determination of conventional protein kinase c isoforms ... of conventional protein kinase c...
TRANSCRIPT

Vision Research 49 (2009) 315–321
Contents lists available at ScienceDirect
Vision Research
journal homepage: www.elsevier .com/locate /v isres
Determination of conventional protein kinase C isoforms involved in highintraocular pressure-induced retinal ischemic preconditioning of rats
Jingwen Ding a,1, Ning Ding a,1, Ningli Wang a,*, Qingjun Lu a, Ningning Lu a, Diya Yang a, Xiangning Bu b,Song Han b, Junfa Li b,*
a Beijing Tongren Eye Center, Capital Medical University Affiliated Beijing Tongren Hospital, Vision Science Laboratory, School of Ophthalmology, Beijing 100730, Chinab Department of Neurobiology and Beijing Institute for Neuroscience, Capital Medical University, #10 You An Men Wai Xi Tou Tiao, Beijing 100069, China
a r t i c l e i n f o
Article history:Received 29 August 2008Received in revised form 13 October 2008
Keywords:Conventional protein kinase C (cPKC)High intraocular pressureRetinal ischemic preconditioningMembrane translocationProtein expression
0042-6989/$ - see front matter � 2008 Elsevier Ltd. Adoi:10.1016/j.visres.2008.10.018
* Corresponding authors. Fax: +86 10 5826 9920 (NLi).
E-mail addresses: [email protected] (N. Wang), ju1 These authors contributed equally to this work,
a b s t r a c t
Evidence indicates that conventional protein kinase C (cPKC) plays a pivotal role in the development ofretinal ischemic preconditioning (IPC). In this study, the effect of high intraocular pressure (IOP)-inducedretinal IPC on cPKC isoform-specific membrane translocation and protein expression were observed. Wefound that cPKCc membrane translocation increased significantly at the early stage (20 min-1 h), whilethe protein expression levels of cPKCa and c were markedly elevated in the delayed retinal IPC(12–168 h) of rats. The increased protein expressions of cPKCa at 72 h and cPKCc at 24 h after IPC werefurther confirmed by immunofluorescence staining. In addition, we found that cPKCc co-localized withretinal ganglion cell (RGC)-specific marker, neurofilaments heavy chain (NF-H) by using doubleimmunofluorescence labeling. These results suggest that increased cPKCc membrane translocation andup-regulated protein expressions of cPKCa and c are involved in the development of high IOP-inducedrat retinal IPC.
� 2008 Elsevier Ltd. All rights reserved.
1. Introduction
Retinal ischemia is clinically considered as one of the majorcauses of visual impairment and blindness. It involves ongoingand irreversible loss of specific classes of neurons in the retina,preferentially retinal ganglion cells (RGCs) (Osborne et al., 2004;Quigley, 1999). Effective strategies for protecting the retina againstischemic injury still remain elusive. Ischemic preconditioning (IPC)refers to the phenomenon whereby a brief period of ischemia pro-tects a tissue against subsequent severe ischemic insult. It has beendemonstrated in myocardium, brain, skeletal muscle, and more re-cently in retina. Roth et al. (Roth et al., 1998) first reported that ret-inal IPC provides complete protection against retinal ischemicinjury in rats. The protective effect of IPC is of great interest andextensive studies in vitro and in vivo have provided accumulateddata on its potential mechanisms. Protein kinase C (PKC) has beenimplicated as a key mediator of the IPC events observed in variousanimals and organs (Bright & Mochly-Rosen, 2005; Kuno et al.,2007; Li et al., 2005; Raval, Dave, Mochly-Rosen, Sick, & Perez-Pin-zon, 2003; Um, Matthews, Song, & Mun, 2005).
ll rights reserved.
. Wang), +86 10 8391 1491 (J.
[email protected] (J. Li).
The PKC family of serine/threonine protein kinases is known toparticipate in signaling pathways mediating critical intracellularevents, including proliferation, differentiation, tumourigenesis,apoptosis, and synaptic transmission (Kanashiro & Khalil, 1998).So far, at least ten PKC isoforms have been identified in mammalsand classified into three subgroups based on their molecular struc-tures and sensitivity to activators: conventional PKCs (cPKCa, bI,bII and c), requiring both Ca2+ and diacylglycerol for activation; no-vel PKCs (nPKCd, e, g and h); and atypical PKCs (aPKCk and n) (Bar-nett, Madgwick, & Takemoto, 2007; Nishizuka, 1984). PKCs arepresent abundantly in neural tissues including retina (Wood,McCord, & Osborne, 1997), and their activations are generallymarked by translocation from the cytosolic fraction to the particu-late or membrane-related fraction (Kurkinen, Busto, Goldsteins,Koistinaho, & Perez-Pinzon, 2001; Niu et al., 2005).
As part of the central nervous system (CNS), the retina sharesmany functional and structural characteristics with the brain.However, one striking difference between them is the relativeresistance of the retina to an ischemic insult (Osborne et al.,2004). Our previous studies have suggested that cPKCbII, c andnPKCe were involved in cerebral hypoxic preconditioning (HPC)(Li et al., 2005, 2006; Niu et al., 2005). In this study, we determinedwhich cPKC isoforms might participate in the retinal IPC by observ-ing the changes in cPKC isoform-specific membrane translocationas well as their protein expression levels in the retina of rat follow-ing high intraocular pressure (IOP)-induced retinal ischemia.

316 J. Ding et al. / Vision Research 49 (2009) 315–321
2. Materials and methods
2.1. Animals
All procedures conducted in this study conformed to the guide-lines of Association for Research in Vision and Ophthalmology(ARVO) Statement for the Use of Animals in Research and were ap-proved by the Animal Care and Use Committee of Capital MedicalUniversity. Specific pathogen-free adult (12–16 weeks of age) maleWistar rats (weighing 250–280 g, purchased from ExperimentalAnimal Center of Chinese Academy of Medical Sciences, China)were housed in temperature-controlled rooms with 12 h light/darkcycles where they received food and water ad libitum.
2.2. High IOP-induced retinal ischemia
Our previously published procedures to generate the retinalischemia model in rats (Wei et al., 2007) were followed with minormodifications. Briefly, rats were initially anesthetized with pento-barbital sodium (50 mg/kg, i.p.) and the body temperature wasmaintained with warming lamp and heating pad. After pupil dila-tion and corneal analgesia occurred, anterior chamber of the righteye was cannulated with a 27-gauge needle connected to an ele-vated saline reservoir. The IOP was raised to 130 mm Hg for only5 min and reperfusion was allowed for a period of 10 min up to168 h. Retinal ischemia was confirmed by the blanching of the irisand retinal circulation. Sham-operated eyes were treated in thesimilar way but without elevating the reservoir. To prevent infec-tion, antibiotic ointment was applied topically after the procedure.If there existed lens damaging, excessive hemorrhaging, or otherocular complications, the rats were discarded and immediatelyeuthanized.
2.3. Subcellular fractionations and whole tissue homogenatepreparations
According to previous studies (Li et al., 2005; Niu et al., 2005),the liquid nitrogen-frozen retinas were immediately placed into80 ll ice-cold, freshly prepared homogenization buffer A (50 mMTris–Cl, pH 7.5, 2 mM DTT, 2 mM EDTA, 2 mM EGTA, 5 mg/ml eachof leupeptin, aprotinin, pepstatin A and chymostatin, 50 mM potas-sium fluoride, 50 lM okadaic acid, 5 mM sodium pyrophosphate)and homogenized. The homogenate was centrifuged at 30,000gfor 30 min, and the supernatants were collected as cytosolic frac-tions. The resulting pellets were further solubilized in 80 llhomogenization buffer B (buffer A containing 0.5% Nonidet P-40)on ice before being sonicated and centrifuged at 30,000g for30 min again. The supernatants were taken as particulate fractions.Both fractions were prepared for membrane translocation analysis.To determine cPKC isoform-specific protein expression levels, fro-zen retinas were homogenized in 150 ll homogenization buffer C(buffer A containing 2% SDS) on ice and sonicated to extract thewhole protein. All procedures were performed at 4 �C, and proteinconcentration was determined by the BCA protein assay (Thermo,Rockford, IL). After normalization, samples were boiled for 5 minand stored at �70 �C until use.
2.4. Western blot analysis
As described previously (Li et al., 2005; Wei et al., 2007), equalamounts of retinal protein (20 lg from either cytosolic or particu-late fractions, and 40 lg from whole homogenates) per lane wereloaded on 10% SDS-PAGE gels which contained the broad-rangePrestained Protein Marker (NEB, Ipswich, MA). The gels were elec-trophoresed, and then transferred onto polyvinylidene difluoride(PVDF) membrane (GE Healthcare, UK) at 4 �C. After several rinses
with TTBS (20 mM Tris–Cl, pH 7.5, 0.15 M NaCl and 0.05% Tween-20), the transferred PVDF membrane was blocked with 10% non-fatmilk in TTBS for 1 h and incubated with the primary rabbit poly-clonal antibodies against cPKCa, bI, bII and c (Santa Cruz Biotech-nology Inc., CA) at a 1:1000 dilution for 3 h. To verify equal loadingof protein, the blots were stripped by incubating the membranesfor 45 min in stripping buffer containing 62.5 mM Tris–Cl (pH6.7), 2% SDS and 100 mM 2-mercaptoethanol at 55 �C and reprobedwith primary monoclonal antibody against b-actin (Sigma-AldrichCompany, St. Louise, MO). The horseradish peroxidase-conjugatedgoat anti-rabbit or anti-mouse IgG (Stressgen, Canada) were usedas second antibodies at a 1:3000 dilution for 1 h incubation. Fol-lowing incubation with the primary and secondary antibodies,the Enhanced Chemiluminescence (ECL) kit (GE Healthcare, UK)was employed to detect the signals.
2.5. Immunostaining analysis
As previous report (Wei et al., 2007), rats were anesthetizedwith pentobarbital sodium (50 mg/kg, i.p.) and perfused transcar-dially with 0.1 M phosphate- buffered saline (PBS, pH 7.4) followedby 4% paraformaldehyde. After enucleation and removal of theanterior segment, the eyecup was post-fixed with the same fixativefor at least 4 h at 4 �C, and subsequently washed in PBS. The sam-ples were then cryoprotected in a series of sucrose in PBS (10%, 20%and 30%), embedded in Tissue-Tek OCT compound (Sakura Finetec,Torrance, CA), and vertically cryosectioned at 16-lm thicknesswith a cryostat (Leica CM1850, Germany). Cryosections that cutthrough the optic nerve head were collected on coated slides,pre-incubated in PBS containing 5% bovine serum albumin (BSA)and 0.5% Triton X-100 for 1 h at room temperature (RT), and thensubsequently incubated in one of primary rabbit polyclonalantibodies against cPKCa and c or a combination of two primaryantibodies such as anti-cPKCc and mouse monoclonal anti-neuro-filaments heavy chain (NF-H, Sigma-Aldrich, St. Louis, MO) at a1:200 dilution overnight at 4 �C. Following rinses in PBS, sectionswere incubated for 2 h in secondary antibodies or a mixture fordouble immunolabeling. Secondary antibodies were either fluores-cein isothiocyanate (FITC)-labeled goat anti-rabbit (green color) orrhodamine-labeled goat anti-mouse IgG (red color, Jackson Immu-noResearch, West Grove, PA) at a dilution of 1:400. Control sec-tions were conducted the same way, but primary antibodieswere omitted. The sections were then cover-slipped in mountingmedium containing DAPI (Vector Laboratories, Burlingame, CA)to visualize cell nuclei (blue color). Fluorescent sections wereviewed by confocal laser-scanning microscope (Leica TCS SP2,Germany).
2.6. Statistical analysis
Quantitative analysis for immunoblotting was done after scan-ning of the X-ray film with Quantitative-one software (Gel Doc2000 imaging system, Bio-Rad, Hercules, CA). To quantitate mem-brane translocation, the ratio of cPKC isoform (band density in par-ticulate/band densities in both particulate and cytosolic fractions)of naïve control group was normalized to 100%, and IPC andsham-operated groups were expressed as percentages of naïvecontrol group. For the quantitative analysis of cPKC protein expres-sion, the optical density of each band corresponding to cPKC iso-forms (from whole tissue homogenate) was normalized to that ofb-actin. The protein expression ratio in naïve control group wasregarded as 100%, and data from IPC and sham-operated groupswere also expressed as percentages of naïve control group. Thepresented values were expressed as mean ± SE. from at least sixindependent experiments. Statistical analysis was conducted byone-way analysis of variance followed by all pairwise multiple
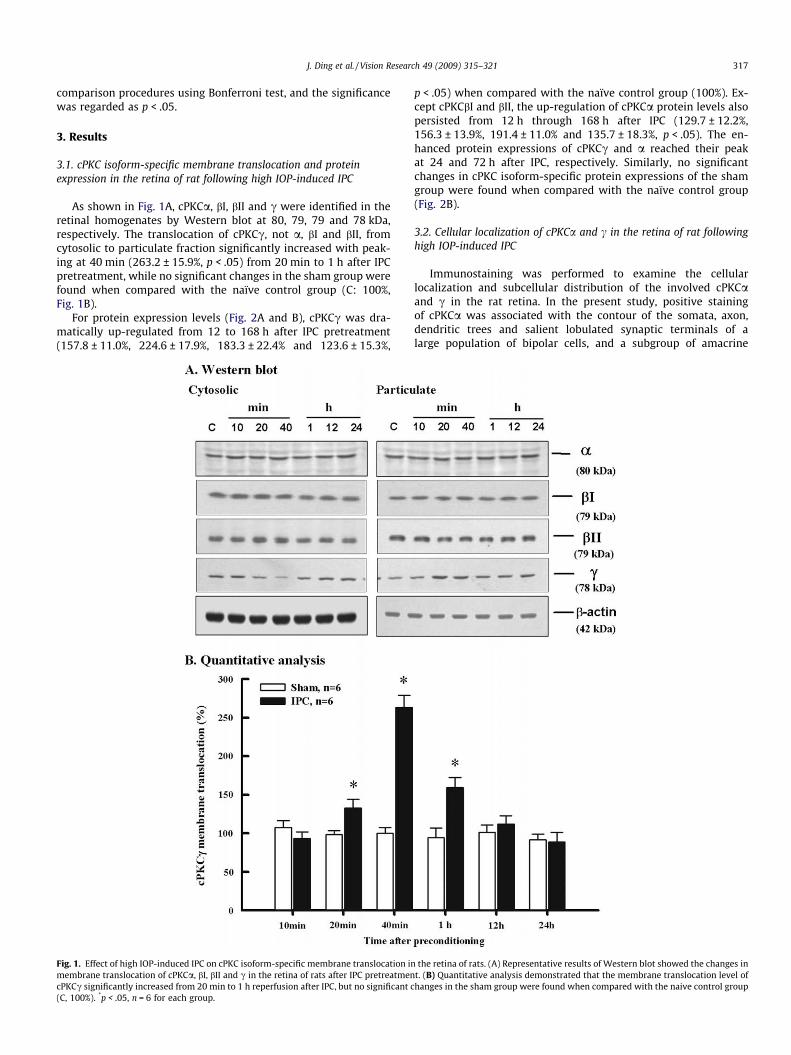
J. Ding et al. / Vision Research 49 (2009) 315–321 317
comparison procedures using Bonferroni test, and the significancewas regarded as p < .05.
3. Results
3.1. cPKC isoform-specific membrane translocation and proteinexpression in the retina of rat following high IOP-induced IPC
As shown in Fig. 1A, cPKCa, bI, bII and c were identified in theretinal homogenates by Western blot at 80, 79, 79 and 78 kDa,respectively. The translocation of cPKCc, not a, bI and bII, fromcytosolic to particulate fraction significantly increased with peak-ing at 40 min (263.2 ± 15.9%, p < .05) from 20 min to 1 h after IPCpretreatment, while no significant changes in the sham group werefound when compared with the naïve control group (C: 100%,Fig. 1B).
For protein expression levels (Fig. 2A and B), cPKCc was dra-matically up-regulated from 12 to 168 h after IPC pretreatment(157.8 ± 11.0%, 224.6 ± 17.9%, 183.3 ± 22.4% and 123.6 ± 15.3%,
Fig. 1. Effect of high IOP-induced IPC on cPKC isoform-specific membrane translocation inmembrane translocation of cPKCa, bI, bII and c in the retina of rats after IPC pretreatmencPKCc significantly increased from 20 min to 1 h reperfusion after IPC, but no significant c(C, 100%). *p < .05, n = 6 for each group.
p < .05) when compared with the naïve control group (100%). Ex-cept cPKCbI and bII, the up-regulation of cPKCa protein levels alsopersisted from 12 h through 168 h after IPC (129.7 ± 12.2%,156.3 ± 13.9%, 191.4 ± 11.0% and 135.7 ± 18.3%, p < .05). The en-hanced protein expressions of cPKCc and a reached their peakat 24 and 72 h after IPC, respectively. Similarly, no significantchanges in cPKC isoform-specific protein expressions of the shamgroup were found when compared with the naïve control group(Fig. 2B).
3.2. Cellular localization of cPKCa and c in the retina of rat followinghigh IOP-induced IPC
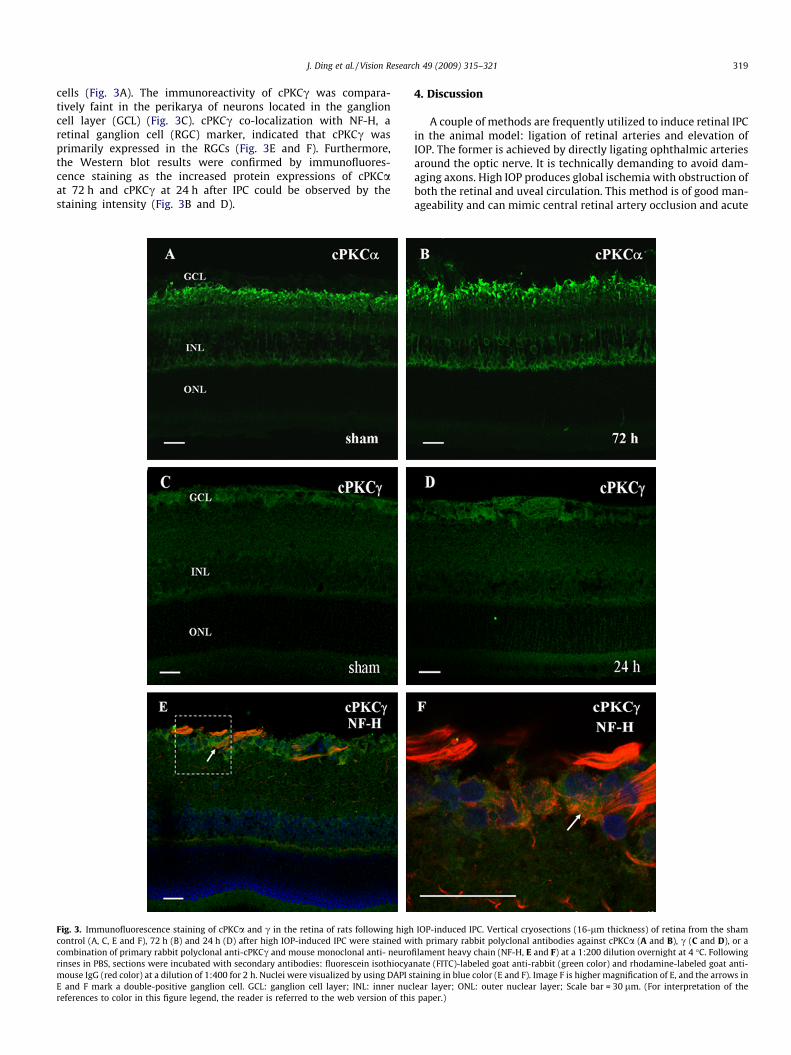
Immunostaining was performed to examine the cellularlocalization and subcellular distribution of the involved cPKCaand c in the rat retina. In the present study, positive stainingof cPKCa was associated with the contour of the somata, axon,dendritic trees and salient lobulated synaptic terminals of alarge population of bipolar cells, and a subgroup of amacrine
the retina of rats. (A) Representative results of Western blot showed the changes int. (B) Quantitative analysis demonstrated that the membrane translocation level ofhanges in the sham group were found when compared with the naive control group
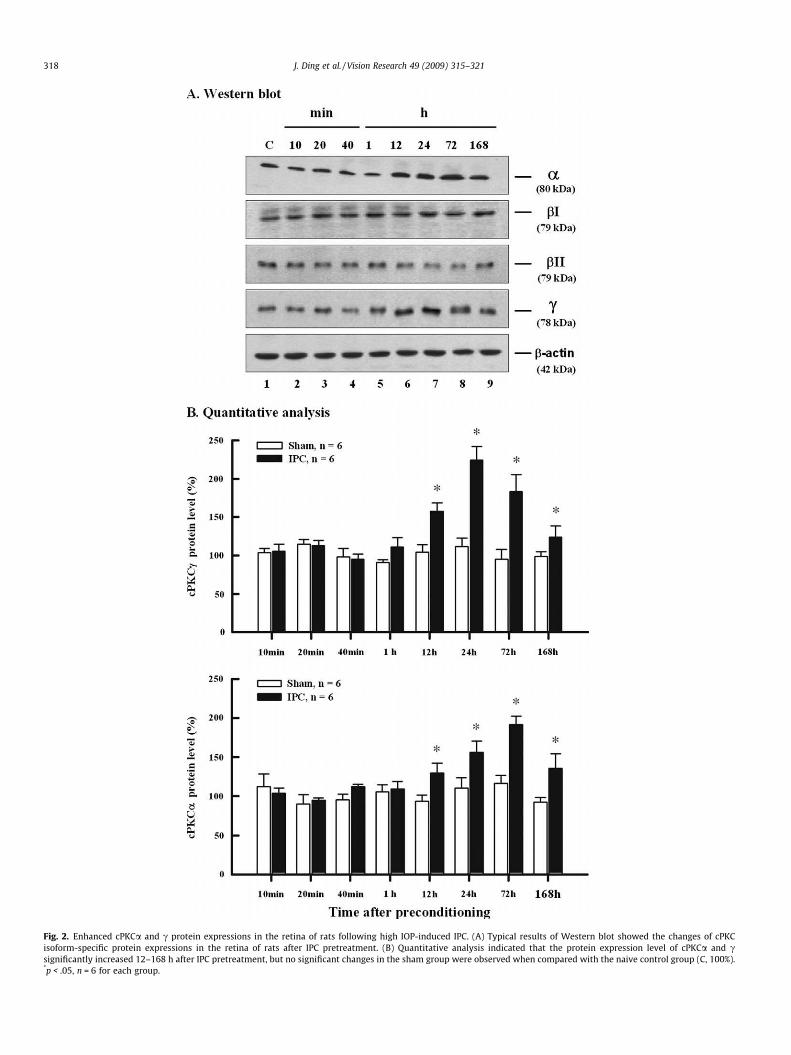
Fig. 2. Enhanced cPKCa and c protein expressions in the retina of rats following high IOP-induced IPC. (A) Typical results of Western blot showed the changes of cPKCisoform-specific protein expressions in the retina of rats after IPC pretreatment. (B) Quantitative analysis indicated that the protein expression level of cPKCa and csignificantly increased 12–168 h after IPC pretreatment, but no significant changes in the sham group were observed when compared with the naive control group (C, 100%).*p < .05, n = 6 for each group.
318 J. Ding et al. / Vision Research 49 (2009) 315–321
J. Ding et al. / Vision Research 49 (2009) 315–321 319
cells (Fig. 3A). The immunoreactivity of cPKCc was compara-tively faint in the perikarya of neurons located in the ganglioncell layer (GCL) (Fig. 3C). cPKCc co-localization with NF-H, aretinal ganglion cell (RGC) marker, indicated that cPKCc wasprimarily expressed in the RGCs (Fig. 3E and F). Furthermore,the Western blot results were confirmed by immunofluores-cence staining as the increased protein expressions of cPKCaat 72 h and cPKCc at 24 h after IPC could be observed by thestaining intensity (Fig. 3B and D).
Fig. 3. Immunofluorescence staining of cPKCa and c in the retina of rats following highcontrol (A, C, E and F), 72 h (B) and 24 h (D) after high IOP-induced IPC were stained wcombination of primary rabbit polyclonal anti-cPKCc and mouse monoclonal anti- neurorinses in PBS, sections were incubated with secondary antibodies: fluorescein isothiocyamouse IgG (red color) at a dilution of 1:400 for 2 h. Nuclei were visualized by using DAPI sE and F mark a double-positive ganglion cell. GCL: ganglion cell layer; INL: inner nucreferences to color in this figure legend, the reader is referred to the web version of thi
4. Discussion
A couple of methods are frequently utilized to induce retinal IPCin the animal model: ligation of retinal arteries and elevation ofIOP. The former is achieved by directly ligating ophthalmic arteriesaround the optic nerve. It is technically demanding to avoid dam-aging axons. High IOP produces global ischemia with obstruction ofboth the retinal and uveal circulation. This method is of good man-ageability and can mimic central retinal artery occlusion and acute
IOP-induced IPC. Vertical cryosections (16-lm thickness) of retina from the shamith primary rabbit polyclonal antibodies against cPKCa (A and B), c (C and D), or afilament heavy chain (NF-H, E and F) at a 1:200 dilution overnight at 4 �C. Followingnate (FITC)-labeled goat anti-rabbit (green color) and rhodamine-labeled goat anti-taining in blue color (E and F). Image F is higher magnification of E, and the arrows inlear layer; ONL: outer nuclear layer; Scale bar = 30 lm. (For interpretation of thes paper.)

320 J. Ding et al. / Vision Research 49 (2009) 315–321
angle closure glaucoma. According to previous studies, a transient(5 min) pretreatment by raising IOP to 130 mmHg does not resultin histological and functional lesion to the retina (Sakamoto,Kuwagata, Nakahara, & Ishii, 2001; Sakamoto et al., 2004).
It has been described that IPC is a biphasic phenomenon in theheart and brain (Das & Das, 2008; Stenzel-Poore, Stevens, King, &Simon, 2007). The two phenotypes differ in the duration andinduction mechanism. The early IPC is initiated and wanes rapidlywithin a few hours, which relates to changes in ion channel perme-abilities, protein phosphorylation and other post-translationalmodifications; the delayed IPC otherwise develops slowly and lastsfor several days, which requires gene activation and de novo pro-tein synthesis (Gidday, 2006). Roth et al. (Roth et al., 1998) re-ported that early preconditioning does not occur in the retina.The window of opportunity between the IPC and subsequent ische-mia is 1–168 h and a 24 or 72 h separation protects the retina func-tionally and histologically, with a greater degree than any knownneuroprotective agent. However, Sakamoto et al. (Sakamotoet al., 2004) later demonstrated that preconditioning 20–40 minbefore sustained ischemia can completely prevent the retinalischemic damage. These conflicting reports may stem from varyingischemic models and intensities of insult. Further investigationshave provided data to suggest that retinal IPC occurs by mecha-nisms similar to other parts of the CNS. Brief ischemia activatesthe adenosine A1 and A2a receptors, which is followed by an in-crease in expression or activation of PKC, or an opening of ATP-sen-sitive K+ channels (Roth, 2004; Roth, Dreixler, Shaikh, Lee, &Bindokas, 2006; Sakamoto et al., 2001). Activated PKC phosphory-lates unknown effector proteins and triggers signal cascades lead-ing to neuroprotection. Nonspecific PKC inhibitors can attenuatethe protection effect in both the early and delayed IPC (Li, Yang,Rosenbaum, & Roth, 2000; Sakamoto et al., 2004). However, differ-ent PKC isoforms complicate the situation and the question regard-ing PKC isoform-specific role in the event is far from answered. Wethereby assessed alterations in the activation and protein level ofcPKC isoforms following IPC stimuli, and found that IPC pretreat-ment stimulates early membrane translocation of cPKCc, and laterelevates protein expression of cPKCa and c in the retina of rats.
Activation of cPKCs is related to cell signaling pathwaysinvolving changes in cytoplasmic Ca2+ levels and transmitter re-lease in neurons of CNS. cPKCa is considered as a good molecularmarker for rod bipolar cells in the vertebrate retina. Ischemia canactivate and downregulate cPKCa in the rabbit retina (Osborne,Wood, & Muller, 1995), whereas IPC is assumed not to affectcPKCa (Casson, Wood, Melena, Chidlow, & Osborne, 2003). How-ever, we found that the expression level of cPKCa in rat retina didsignificantly increase 12–168 h after IPC. cPKCa is prominentlyexpressed in the synaptic terminals of the bipolar cells, whichseems to suggest that cPKCa might fulfill its protective functionby modulating synaptic contacts with RGCs. cPKCbI and bII aregenerated by alternative splicing from a single gene (Kawakami,Kawakami, & Kitaura, 2002). They are mostly expressed in conebipolar cells, amacrine cells and RGCs in the retina. We recentlyreported that both of these isoforms are activated in neuroblas-toma cells after hypoxic exposure (Li et al., 2006). There was nosubcellular translocation or altered expression of cPKCbI and bIIobserved in the IPC retina. cPKCc is primarily expressed in theneurons of brain and spinal cord (Saito & Shirai, 2002). The iso-form may thus play a pivotal CNS-specific role in mediating re-sponses to ischemic insult. It is also found in the eye tissuesincluding retina and lens epithelium and cortex (Correia, Duarte,Faro, Pires, & Carvalho, 2003; Saleh, Takemoto, Zoukhri, & Takem-oto, 2001). The absence of cPKCc in the rat retina has beenproved by immunohistochemistry, Western blot, and in situhybridization (Kosaka, Suzuki, Morii, & Nomura, 1998; Osborne,Barnett, Morris, & Huang, 1992). In our study, nevertheless, cPKCc
was actually detected by Western blot and immunostainingmethods. Moreover, double-labeled staining showed cPKCc pre-dominantly resided in the RGCs. In order to exclude the nonspe-cific effect, we used hippocampus and lung as positive andnegative controls respectively (data not shown). The results cor-roborated cPKCc presence in the rat retina. The discrepancymay be explained by the different antibodies or procedures.Although Western blot analysis confirmed early membrane trans-location of cPKCc, we did not observe any subcellular redistribu-tion 40 min after IPC by immunofluoresence staining. Lack ofcorrelation between the two methods was probably owing to lim-itations of the cryosection staining and low levels of antigen pres-ent in the retina. Taken together, the striking increases inmembrane translocation and expression level of cPKCc demon-strate its possible protective effect on RGCs against ischemic in-jury. As cPKCc has been implicated in regenerating neuritis andaxons (Okajima, Mizoguchi, Tamai, Hirasawa, & Ide, 1995; Tsaiet al., 2007), cPKCc may exert its protective effect by promotingaxonal growth after the IPC insult.
In conclusion, we provide new data concerning cPKC isoform-specific membrane translocation and protein expression in theIPC retina of rat. The development of retinal IPC is accompaniedby increased cPKCc membrane translocation in the RGCs at theearly stage, and enhanced protein expression levels of cPKCa andc at the delayed stage. Studies are underway to identify the protec-tive role of cPKCa and c in the retina against severe ischemic inju-ries, and their downstream targets. Elucidation of the molecularmechanisms of IPC may lead to the findings in novel strategiesfor ischemic retinal diseases.
Acknowledgments
This work was supported by the following Grants: National Nat-ural Science Foundation of China (30571991, 30670782 and30871219), Beijing Natural Science Foundation (7043067 and5072008), Key Scientific Developing Program of Beijing MunicipalCommission of Education (KZ200810025012), Funding Project forAcademic Human Resources Development in Institutions of HigherLearning Under the Jurisdiction of Beijing Municipality (Li J) andChina 973 Program (2006CB504100).
References
Barnett, M. E., Madgwick, D. K., & Takemoto, D. J. (2007). Protein kinase C as a stresssensor. Cell Signalling, 19, 1820–1829.
Bright, R., & Mochly-Rosen, D. (2005). The role of protein kinase C in cerebralischemic and reperfusion injury. Stroke, 36, 2781–2790.
Casson, R. J., Wood, J. P., Melena, J., Chidlow, G., & Osborne, N. N. (2003). The effect ofischemic preconditioning on light-induced photoreceptor injury. InvestigativeOphthalmology & Visual Science, 44, 1348–1354.
Correia, S. S., Duarte, C. B., Faro, C. J., Pires, E. V., & Carvalho, A. L. (2003). Proteinkinase C gamma associates directly with the GluR4 alpha-amino-3-hydroxy-5-methyl-4-isoxazole propionate receptor subunit. Effect on receptorphosphorylation. The Journal of Biological Chemistry, 278, 6307–6313.
Das, M., & Das, D. K. (2008). Molecular mechanism of preconditioning. IUBMB Life,60, 199–203.
Gidday, J. M. (2006). Cerebral preconditioning and ischaemic tolerance. NatureReviews. Neuroscience, 7, 437–448.
Kanashiro, C. A., & Khalil, R. A. (1998). Signal transduction by protein kinase C inmammalian cells. Clinical and Experimental Pharmacology & Physiology, 25,974–985.
Kawakami, T., Kawakami, Y., & Kitaura, J. (2002). Protein kinase C beta (PKC beta):Normal functions and diseases. Journal of Biochemistry, 132, 677–682.
Kosaka, J., Suzuki, A., Morii, E., & Nomura, S. (1998). Differential localization andexpression of alpha and beta isoenzymes of protein kinase C in the rat retina.Journal of Neuroscience Research, 54, 655–663.
Kuno, A., Critz, S. D., Cui, L., Solodushko, V., Yang, X. M., Krahn, T., et al. (2007).Protein kinase C protects preconditioned rabbit hearts by increasing sensitivityof adenosine A2b-dependent signaling during early reperfusion. Journal ofMolecular and Cellular Cardiology, 43, 262–271.
Kurkinen, K., Busto, R., Goldsteins, G., Koistinaho, J., & Perez-Pinzon, M. A. (2001).Isoform-specific membrane translocation of protein kinase C after ischemicpreconditioning. Neurochemical Research, 26, 1139–1144.

J. Ding et al. / Vision Research 49 (2009) 315–321 321
Li, J., Niu, C., Han, S., Zu, P., Li, H., Xu, Q., et al. (2005). Identification of protein kinaseC isoforms involved in cerebral hypoxic preconditioning of mice. Brain Research,1060, 62–72.
Li, J., Qu, Y., Zu, P., Han, S., Gao, G., Xu, Q., et al. (2006). Increased isoform-specificmembrane translocation of conventional and novel protein kinase C in humanneuroblastoma SH-SY5Y cells following prolonged hypoxia. Brain Research,1093, 25–32.
Li, B., Yang, C., Rosenbaum, D. M., & Roth, S. (2000). Signal transduction mechanismsinvolved in ischemic preconditioning in the rat retina in vivo. Experimental EyeResearch, 70, 755–765.
Nishizuka, Y. (1984). The role of protein kinase C in cell surface signal transductionand tumour promotion. Nature, 308, 693–698.
Niu, C., Li, J., Cui, X., Han, S., Zu, P., Li, H., et al. (2005). Changes in cPKC isoform-specific membrane translocation and protein expression in the brain of hypoxicpreconditioned mice. Neuroscience Letters, 384, 1–6.
Okajima, S., Mizoguchi, A., Tamai, K., Hirasawa, Y., & Ide, C. (1995). Distribution ofprotein kinase C (alpha, beta, gamma subtypes) in normal nerve fibers and inregenerating growth cones of the rat peripheral nervous system. Neuroscience,66, 645–654.
Osborne, N. N., Barnett, N. L., Morris, N. J., & Huang, F. L. (1992). The occurrence ofthree isoenzymes of protein kinase C (alpha, beta and gamma) in retinas ofdifferent species. Brain Research, 570, 161–166.
Osborne, N. N., Casson, R. J., Wood, J. P., Chidlow, G., Graham, M., & Melena, J. (2004).Retinal ischemia: Mechanisms of damage and potential therapeutic strategies.Progress in Retinal and Eye Research, 23, 91–147.
Osborne, N. N., Wood, J., & Muller, A. (1995). The influence of experimentalischaemia on protein kinase C and the GABAergic system in the rabbit retina.Neuropharmacology, 34, 1279–1288.
Quigley, H. A. (1999). Neuronal death in glaucoma. Progress in Retinal and EyeResearch, 18, 39–57.
Raval, A. P., Dave, K. R., Mochly-Rosen, D., Sick, T. J., & Perez-Pinzon, M. A. (2003).Epsilon PKC is required for the induction of tolerance by ischemic and NMDA-mediated preconditioning in the organotypic hippocampal slice. The Journal ofNeuroscience, 23, 384–391.
Roth, S. (2004). Endogenous neuroprotection in the retina. Brain Research Bulletin,62, 461–466.
Roth, S., Dreixler, J. C., Shaikh, A. R., Lee, K. H., & Bindokas, V. (2006). Mitochondrialpotassium ATP channels and retinal ischemic preconditioning. InvestigativeOphthalmology & Visual Science, 47, 2114–2124.
Roth, S., Li, B., Rosenbaum, P. S., Gupta, H., Goldstein, I. M., Maxwell, K. M.,et al. (1998). Preconditioning provides complete protection against retinalischemic injury in rats. Investigative Ophthalmology & Visual Science, 39,777–785.
Saito, N., & Shirai, Y. (2002). Protein kinase C gamma (PKC gamma): Function ofneuron specific isotype. Journal of Biochemistry, 132, 683–687.
Sakamoto, K., Kuwagata, M., Nakahara, T., & Ishii, K. (2001). Late preconditioning inrat retina: Involvement of adenosine and ATP-sensitive K(+) channel. EuropeanJournal of Pharmacology, 418, 89–93.
Sakamoto, K., Yonoki, Y., Kuwagata, M., Saito, M., Nakahara, T., & Ishii, K. (2004).Histological protection against ischemia-reperfusion injury by early ischemicpreconditioning in rat retina. Brain Research, 1015, 154–160.
Saleh, S. M., Takemoto, L. J., Zoukhri, D., & Takemoto, D. J. (2001). PKC-gammaphosphorylation of connexin 46 in the lens cortex. Molecular Vision, 7,240–246.
Stenzel-Poore, M. P., Stevens, S. L., King, J. S., & Simon, R. P. (2007). Preconditioningreprograms the response to ischemic injury and primes the emergence ofunique endogenous neuroprotective phenotypes: A speculative synthesis.Stroke, 38, 680–685.
Tsai, S. Y., Yang, L. Y., Wu, C. H., Chang, S. F., Hsu, C. Y., Wei, C. P., et al. (2007). Injury-induced Janus kinase/protein kinase C-dependent phosphorylation of growth-associated protein 43 and signal transducer and activator of transcription 3 forneurite growth in dorsal root ganglion. Journal of Neuroscience Research, 85,321–331.
Um, J. W., Matthews, J. B., Song, J. C., & Mun, E. C. (2005). Role of protein kinase C inintestinal ischemic preconditioning. The Journal of Surgical Research, 124,289–296.
Wei, Y., Wang, N., Lu, Q., Zhang, N., Zheng, D., & Li, J. (2007). Enhanced proteinexpressions of sortilin and p75NTR in retina of rat following elevatedintraocular pressure-induced retinal ischemia. Neuroscience Letters, 429,169–174.
Wood, J. P., McCord, R. J., & Osborne, N. N. (1997). Retinal protein kinase C.Neurochemistry International, 30, 119–136.