determination of quality constituents in the young...
TRANSCRIPT

Food Chemistry 155 (2014) 98–104
Contents lists available at ScienceDirect
Food Chemistry
journal homepage: www.elsevier .com/locate / foodchem
Determination of quality constituents in the young leaves of albino teacultivars
http://dx.doi.org/10.1016/j.foodchem.2014.01.0440308-8146/� 2014 Elsevier Ltd. All rights reserved.
⇑ Corresponding authors. Tel.: +1 306 385 9369; fax: +1 306 385 9482 (M.-J. Gao).Tel./fax: +86 551 65783941 (S. Wei).
E-mail addresses: [email protected] (M.-J. Gao), [email protected](S. Wei).
Lin Feng a, Ming-Jun Gao b,⇑, Ru-Yan Hou a, Xiao-Yi Hu c, Liang Zhang a, Xiao-Chun Wan a, Shu Wei a,⇑a Key Laboratory of Tea Biochemistry and Biotechnology, Anhui Agricultural University, 130 Changjiang Blvd West, Hefei, Anhui 230036, Chinab Agriculture and Agri-Food Canada, Saskatoon Research Centre, 107 Science Place, Saskatoon, SK S7N 0X2, Canadac College of Forestry and Landscape Architecture, Anhui Agricultural University, 130 Changjiang Blvd West, Hefei, Anhui 230036, China
a r t i c l e i n f o
Article history:Received 23 August 2013Received in revised form 15 December 2013Accepted 15 January 2014Available online 23 January 2014
Keywords:Camellia sinensisAlbino teaMetabolic profilingMultivariate analysisTea quality
a b s t r a c t
Albino tea has received increased attention due to its brisk flavour. To identify changes in the key chem-ical constituents conveying important qualities to albino tea, the metabolite profiles of four albinocultivars and one green tea cultivar were analysed. Compared to the green tea control, significantlydecreased contents of chlorophyll (Chl) (p < 0.01), total carotenoids (p < 0.05), caffeine (p < 0.01), andtotal catechins (p < 0.05) were found in albino tea leaves with a few exceptions, whereas increases werenoted in the Chl a/b ratio and the contents of both zeaxanthin and free amino acids, including theanine.Multivariate analysis identified catechins and carotenoids as the most important contributors to the met-abolic profile variance between the albino and green tea cultivars. High levels of amino acids, along withlow levels of chlorophylls, catechins and caffeine, contribute to the qualities of albino tea, which includereduced astringency and bitterness, along with a strong umami taste.
� 2014 Elsevier Ltd. All rights reserved.
1. Introduction
Tea is one of the most widely consumed non-alcoholic bever-ages in the world and imparts marked benefits to human health,such as providing antioxidant activity and reducing the risk of car-diovascular disease and some forms of cancer (Cabrera, Artacho, &Gimenez, 2006). Under certain environmental conditions, such aslow temperature or abnormal light intensity, the albino tea culti-vars generate yellowish to off-white leaves which are deficient inchlorophyll (Du et al., 2006). Compared to normal green cultivars,albino tea germplasm is precious, due to the special flavour, dis-tinct leaf colour and scarcity. Studies have been published for onlya select few of the albino tea cultivars which are commerciallygrown. Chemical analyses and expression profiling of the genesrelated to chlorophyll biosynthesis in these albino cultivars havebegun to reveal the basis of tea leaf albinism at the levels of bio-chemistry and molecular biology (Du et al., 2006, 2008; Wei,Wang, & Zhou, 2012). However, changes in comprehensive meta-bolic profiles of albino tea cultivars and their impact on tea qualityneed to be determined to further improve tea quality.
The quality of processed tea is basically determined by thechemical constituents in tea leaves (Cabrera et al., 2006; Chaturve-dula & Prakash, 2011). The compounds determining tea flavour andastringency include catechins (flavan-3-ols), including (�)-epigal-locatechin gallate (EGCG), (+)-catechin (C), (+)-gallocatechin (GC)and their oxidation products (Chaturvedula & Prakash, 2011; Liang,Ma, Lu, & Wu, 2006; Narukawa, Kimata, Noga, & Watanabe, 2010).In addition, amino acids are the main contributors towards theumami taste of tea infusions (Alcázar et al., 2007). Theanine (c-glut-amyl-L-ethylamide) is the most abundant free amino acid in tea andcan dramatically increase the brothy taste of tea infusions by effec-tively counteracting the astringency and bitterness (Chen, Duan, &Jiang, 2011; Chu, Kobayashi, Juneja, & Yamamoto, 1997). The chlo-rophylls (Chl) and carotenoids, which function as light-harvestingpigments in plants, are the determinants of tea leaf colour.
In order to determine the changes in the key chemical constit-uents affecting albino tea quality, we generated metabolic profilesof four albino cultivars and one green tea cultivar, using highperformance liquid chromatography (HPLC). The data were furtheranalysed by using multivariate analysis approaches to reveal thekey determinants leading to albino tea qualities and metabolic var-iance. Our results suggest that decreased abundances of carote-noids and chlorophylls are accompanied with a correspondingincrease in the abundances of free amino acids (including thea-nine) in albino leaves. These results provide insight into the forma-tion of tea quality-influencing metabolites in albino tea leaves.

L. Feng et al. / Food Chemistry 155 (2014) 98–104 99
2. Materials and methods
2.1. Plant materials
Three-year old plants of Camellia sinensis (L.) cv. ‘Anji baicha’(Ab) (as called White leaf No.1, Li et al., 2011), ‘Huangjinya’ (Hu),‘Tiantai huangcha’ (Th), ‘Yujinxiang’ (Yu) and six-year old plantsof ‘Fuding dabaicha’ (Fd) were grown under the same cultivationmanagement at the experimental tea farm of Anhui AgriculturalUniversity, Hefei, China (latitude: 31.52�N, longitude: 117.17�E).The fully expanded 2nd leaves in current shoots were collectedin the spring when those of albino cultivars ‘Ab’, ‘Hu’, ‘Th’ and‘Yu’ were yellowish, and the ones of normal cultivar ‘Fd’ weregreen (Fig. 1). A leaf sample consisted of pooled leaves (3–4 perplant) collected from four randomly selected tea plants. There werethree biological replicates for each chemical assay. All excised sam-ples were immediately frozen in liquid nitrogen, brought back tothe lab and stored at �80 �C prior to analysis.
2.2. Chemicals and assays
The following chemicals were purchased from Sigma–Aldrich(St. Louis, MO, USA): caffeine (CF), 7-methylxan (7-Mx), (+)-cate-chin (C), (�)-epicatechin (EC), (�)-epicatechin gallate (ECG), (�)-epigallocatechin (EGC), (�)-epigallocatechin gallate (EGCG), (+)-gallocatechin (GC), alanine (Ala), aspartic acid (Asp), arginine(Arg), asparagine (Asn), cysteine (Cys), glutamine (Gln), glutamicacid (Glu), glycine (Gly), histidine (His), isoleucine (Ile), leucine(Leu), lysine (Lys), methionine (Met), phenylalanine (Phe), proline(Pro), serine (Ser), theanine, threonine (Thr), tryptophan (Trp),tyrosine (Tyr) and valine (Val). The following were purchased fromCaroteNature (Lupsingen, Switzerland): b-carotene, b-cryptoxan-thin (b-Cry), lutein, violaxanthin (VX) and zeaxanthin (Zea).
Leaf carotenoids were extracted and analysed according to apreviously published protocol for canola (Yu, Lydiate, Schäfer, &Hannoufa, 2007) with some modifications. Aliquots (5 ll) of theextracts were analysed by HPLC, using the YMC carotenoid columnreverse-phase C30 (4.6 � 250 mm; Waters Ltd., Mississauga, Can-ada) with a column temperature of 35 �C. Peaks were identifiedby comparing their retention times and absorption spectra to thoseof known standards. The carotenoids were quantified, using curvesconstructed from authentic standards. Leaf chlorophylls were ex-tracted and analysed as previously described (Wei et al., 2012)with three independent biological replicates.
Chemical assays of free amino acids and alkaloids in tea leaveswere conducted as previously described (Deng, Wang, Chen, Zhang,& Hu, 2012). Leaf catechins were extracted and abundances weremeasured by a published protocol (Ashihara, Deng, Mullen, &Crozier, 2010), using HPLC with a C18 Column (4.6 � 250 mm;Phenomenex, China). Eluted compounds were identified by com-paring their retention times and absorption spectra with those ofauthentic standards. The compounds were quantified using curvesconstructed from the standards.
Fig. 1. Leaf phenotypes of the 3-year-old albino and 6-year-old normal teacultivars. (A and B) Tiantai huangcha’ (Th); (C and D), ‘Huangjinya’ (Hu) (E and F),‘Yujinxiang’ (Yu); (G and H), ‘Anji baicha’ (Ab); (I and J), Fuding dabaicha’ (Fd).
2.3. Statistical and multivariate analysis
Data were expressed as means ± SD from three independentbiological replicates for all chemical assays. Statistical analysiswas performed by one-way analysis of variance (ANOVA), usingthe software SPSS 17.0 (SPSS Inc., Chicago, IL, USA). The HPLC datamatrix was subjected to multivariate analysis with SIMCA-P 13software (Umetrics AB, Umea, Sweden). Unsupervised principalcomponents analysis (PCA) was run to obtain a general overviewof the variance of metabolites in a data table by forming a lowdimensional model plane; a score plot was then generated to
visualise the relationships among samples in the model plane.Supervised orthogonal partial least-squares discriminant analysis

100 L. Feng et al. / Food Chemistry 155 (2014) 98–104
(OPLS-DA), a more advanced multivariate method for understand-ing the interclass variation, was performed to separate differentvariations among samples in a clearer and more straightforwardway and the corresponding loading plot was used for identifyingvariables important for the class separation (Trygg, Holmes, &Londstedt, 2007). The data points on the OPLS-DA loading plotwere assigned a variable of importance. A range of metaboliteswas selected to represent the variables of importance (VIP) in theprojection based on the OPLS-DA analysis (Ku et al., 2010).
3. Results and discussion
3.1. Phenotypic characterisation of the four albino mutants
In early spring, with the seasonal low temperatures and lowlight intensities, the first flush shoots of all the four albino cultivarswere yellowish. When the shoots reached a stage with three fully
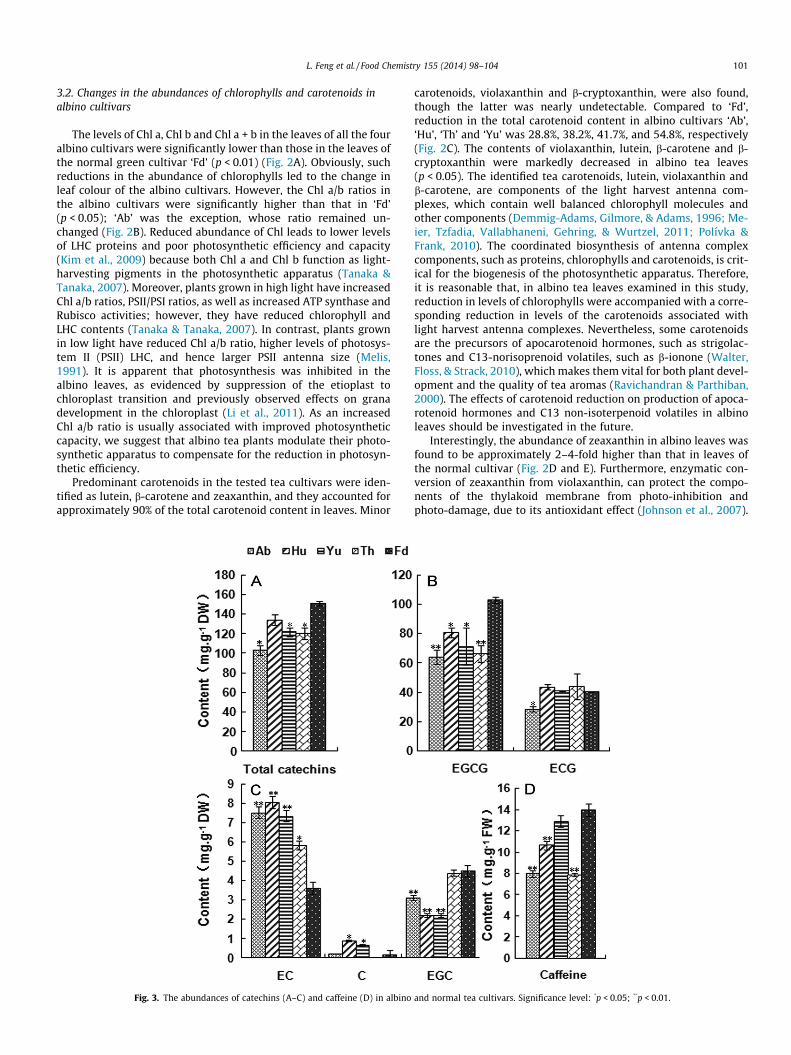
Fig. 2. Changes in the abundances of chlorophylls and predominant carotenoids in albinSignificance level:
⁄p < 0.05;
⁄⁄p < 0.01.
opened leaves in late spring, the leaf phenotypes of the four albinocultivars were further developed, with varying degrees of albinism,while the leaves of the normal green cultivar were dark green, ex-cept for the two slightly yellowish apical leaves (as shown inFig. 1). Interestingly, the leaf colour of ‘Ab’ gradually turned to anormal green colour during the following 2 weeks of leaf growthas the temperature and light intensity increased; this observedchange in ‘Ab’ has been reported previously (Du et al., 2006; Liet al., 2011). In contrast, the leaves of the other albino cultivars,‘Hu’, ‘Th’ and ‘Yu’, generated in spring, remained yellowish underincreased temperatures and light intensities through to autumn,whereas the leaves generated in the previous year changed to anormal green colour (Fig. 1). Such a change in leaf colour of the al-bino cultivars is advantageous, as it allows processing for the un-ique albino tea, using the yellowish or bleached fresh leaves,while maintaining tea plant productivity, due to recovery of leafgreen colour on the older leaves.
o tea leaves. (A) Chlorophyll contents; (B) Chl a/b ratios; (C–E) carotenoid contents.
L. Feng et al. / Food Chemistry 155 (2014) 98–104 101
3.2. Changes in the abundances of chlorophylls and carotenoids inalbino cultivars
The levels of Chl a, Chl b and Chl a + b in the leaves of all the fouralbino cultivars were significantly lower than those in the leaves ofthe normal green cultivar ‘Fd’ (p < 0.01) (Fig. 2A). Obviously, suchreductions in the abundance of chlorophylls led to the change inleaf colour of the albino cultivars. However, the Chl a/b ratios inthe albino cultivars were significantly higher than that in ‘Fd’(p < 0.05); ‘Ab’ was the exception, whose ratio remained un-changed (Fig. 2B). Reduced abundance of Chl leads to lower levelsof LHC proteins and poor photosynthetic efficiency and capacity(Kim et al., 2009) because both Chl a and Chl b function as light-harvesting pigments in the photosynthetic apparatus (Tanaka &Tanaka, 2007). Moreover, plants grown in high light have increasedChl a/b ratios, PSII/PSI ratios, as well as increased ATP synthase andRubisco activities; however, they have reduced chlorophyll andLHC contents (Tanaka & Tanaka, 2007). In contrast, plants grownin low light have reduced Chl a/b ratio, higher levels of photosys-tem II (PSII) LHC, and hence larger PSII antenna size (Melis,1991). It is apparent that photosynthesis was inhibited in thealbino leaves, as evidenced by suppression of the etioplast tochloroplast transition and previously observed effects on granadevelopment in the chloroplast (Li et al., 2011). As an increasedChl a/b ratio is usually associated with improved photosyntheticcapacity, we suggest that albino tea plants modulate their photo-synthetic apparatus to compensate for the reduction in photosyn-thetic efficiency.
Predominant carotenoids in the tested tea cultivars were iden-tified as lutein, b-carotene and zeaxanthin, and they accounted forapproximately 90% of the total carotenoid content in leaves. Minor
Fig. 3. The abundances of catechins (A–C) and caffeine (D) in albino
carotenoids, violaxanthin and b-cryptoxanthin, were also found,though the latter was nearly undetectable. Compared to ‘Fd’,reduction in the total carotenoid content in albino cultivars ‘Ab’,‘Hu’, ‘Th’ and ‘Yu’ was 28.8%, 38.2%, 41.7%, and 54.8%, respectively(Fig. 2C). The contents of violaxanthin, lutein, b-carotene and b-cryptoxanthin were markedly decreased in albino tea leaves(p < 0.05). The identified tea carotenoids, lutein, violaxanthin andb-carotene, are components of the light harvest antenna com-plexes, which contain well balanced chlorophyll molecules andother components (Demmig-Adams, Gilmore, & Adams, 1996; Me-ier, Tzfadia, Vallabhaneni, Gehring, & Wurtzel, 2011; Polívka &Frank, 2010). The coordinated biosynthesis of antenna complexcomponents, such as proteins, chlorophylls and carotenoids, is crit-ical for the biogenesis of the photosynthetic apparatus. Therefore,it is reasonable that, in albino tea leaves examined in this study,reduction in levels of chlorophylls were accompanied with a corre-sponding reduction in levels of the carotenoids associated withlight harvest antenna complexes. Nevertheless, some carotenoidsare the precursors of apocarotenoid hormones, such as strigolac-tones and C13-norisoprenoid volatiles, such as b-ionone (Walter,Floss, & Strack, 2010), which makes them vital for both plant devel-opment and the quality of tea aromas (Ravichandran & Parthiban,2000). The effects of carotenoid reduction on production of apoca-rotenoid hormones and C13 non-isoterpenoid volatiles in albinoleaves should be investigated in the future.
Interestingly, the abundance of zeaxanthin in albino leaves wasfound to be approximately 2–4-fold higher than that in leaves ofthe normal cultivar (Fig. 2D and E). Furthermore, enzymatic con-version of zeaxanthin from violaxanthin, can protect the compo-nents of the thylakoid membrane from photo-inhibition andphoto-damage, due to its antioxidant effect (Johnson et al., 2007).
and normal tea cultivars. Significance level:⁄p < 0.05;
⁄⁄p < 0.01.

102 L. Feng et al. / Food Chemistry 155 (2014) 98–104
Zeaxanthin accumulation can be triggered by adverse environmen-tal conditions, such as excess light, desiccation, and low tempera-ture (Fernández-Marín, Míguez, Becerril, & García-Plazaola, 2011).We speculated that increased levels of zeaxanthin in albino leaveswould be one of the adaptive responses to photooxidative stressfor maintaining photosynthetic efficiency and capacity.
3.3. Changes in the abundances of free amino acids in albino teas
The abundances of free amino acids in each albino cultivar werecompared with those found in the normal cultivar ‘Fd’. A total of17 amino acids were detected in the leaves of all tea plants exam-ined. Theanine was the most abundant free-form amino acidamong the detected compounds, followed by Glu, His and Asp(Supplementary Table 1). The abundances of theanine and Glu inall the tested albino leaves were significantly higher than thosein the leaves of the ‘Fd’ cultivar (p < 0.05). In addition, the concen-trations of serine and total amino acids were markedly increased inthe albino cultivars ‘Hu’ and ‘Yu’ (p < 0.05), while neither changedsignificantly in ‘Ab’ (Supplementary Table 1). High levels of freeamino acids are essential for good quality of green teas (Alcázaret al., 2007; Le Gall, Colquhoun, & Defernez, 2004). Theanine, Ser,and Glu can impart the ‘‘umami’’ or ‘‘brothy’’ taste to teas
Fig. 4. OPLS-DA score plot (A) and corresponding loading plot (B) derived from the integindependent biological replicates. Abbreviations of compounds are outlined in the Mate
(Ekborg-Ott, Taylor, & Armstrong, 1997; Le Gall et al., 2004). Wesuggest that theanine, Glu, and Ser are key components for provid-ing the strong umami flavour of green tea made from albinocultivars.
Theanine is synthesised from Glu and ethylamine by theaninesynthetase (Deng et al., 2012). The Glu is derived from the gluta-mine synthase (GS)/glutamate synthase (GOGAT) cycle, and it isthe direct substrate for theanine biosynthesis. The five-carbonbackbone of Glu also serves in the C5 pathway as the direct precur-sor of 5-aminolevulinic acid (ALA), which is the critical precursorfor chlorophyll synthesis (Tanaka & Tanaka, 2007). It was shownthat increased levels of Glu, resulting from salt stress, led to im-proved theanine biosynthesis in tea seedlings (Deng et al., 2012).Together with our finding in this study that increased theanineand abundant Glu were both present in albino tea leaves, we sug-gest that suppression of chlorophyll biosynthesis in albino leavesmight lead to increased levels of Glu, which are shifted into thetheanine biosynthesis pathway.
3.4. Changes in the abundances of catechins and caffeine in albino teas
The profiles of catechins in albino and normal cultivars werequantitatively analysed using HPLC. The total catechin content
rated HPLC data sets of albino and normal tea cultivars. Data were derived from tworials and methods.

L. Feng et al. / Food Chemistry 155 (2014) 98–104 103
varied from 103 to 150 mg g�1 DW among the different cultivars,with albino cultivars having significantly lower content than thenormal one (p < 0.05; Fig. 3A); our results are in accordance withprevious reports (Du et al., 2006; Wei et al., 2012). The compositionof catechins was also determined. EGCG was the most abundantwhile C was the least abundant in the leaves of all cultivars exam-ined (Fig. 3B and C). Compared to the contents in the normal culti-var, EGCG content in leaves of the four albino cultivars, and EGC inall albino cultivars except ‘Th’, were significantly lower; EC contentin all albino cultivars was significantly higher (p < 0.05; Fig. 3B andC). The abundance of caffeine in albino cultivars ‘Ab’, ‘Hu’, or ‘Th’was significantly lower than that in ‘Fd’ (p < 0.01), but no signifi-cant difference was found in caffeine levels for ‘Yu’ and ‘Fd’. Cate-chins, together with caffeine (1,3,7-trimethylxanthine), are thecompounds determining tea flavour, astringency and overall qual-ity of the tea (Chaturvedula & Prakash, 2011; Liang et al., 2006;Narukawa et al., 2010). Catechins, especially EGCG and EGC, exhi-bit a strong taste of astringency and bitterness; EC provides addi-tional tastes of saltiness and umami (Narukawa et al., 2010). Ourresults suggest that tea made from albino leaves containing re-duced abundances of alkaloids and total catechins, EGCG andEGC might be less astringent and bitter. Reduction of caffeine isgenerally considered beneficial to health (Mohanpuria, Kumar, &Yadav, 2010) whereas reduced catechins might affect antioxidantlevels in the tea.
Fig. 5. Change in the metabolic profiles (p < 0.05 or p < 0.01) of albino tea leaves anphosphoglycerate; ALA, 5-aminolevulinic acid; ch, chloroplast; Chl a, chlorophyll a; Chl bpoly-isoprenoid geranylgeranyl diphosphate; GOGAT, glutamate synthase; GS, glutamimitochondrium; OAA, oxaloacetate; PRPP, phosphoribosyl pyrophosphate; Pchlide, protopyruvate; vac, vacuole.
3.5. Differentiation of cultivars and identification of the metabolitescontributing to cluster formation
Orthogonal partial least-squares discriminant analysis (OPLS-DA) was performed on the metabolite profiles obtained from thefive cultivars to uncover the subtle changes in the occurrenceand abundance of specific compounds. The OPLS-DA score plotshowed a clear separation between the normal cultivar ‘Fd’ andalbino varieties (Fig. 4A). Albino cultivars ‘Ab’, ‘Th’ and ‘Yu’ fellwithin the upper left quadrant, whereas ‘Hu’ fell within the lowerright quadrant, demonstrating that ‘Hu’ has a metabolic profile dis-tinct from the other albino cultivars. The OPLS-DA loading plot as-signed variables of importance (VIP) to each data point. In general,a given metabolite with a smaller influence on cluster formationwill lie closer to the origin. Component P1 in the loading plot isthe principal component and displays the sample variation amongcultivars and P2 is the Y-orthogonal component and models thevariation within a specific cultivar. As shown in Fig. 4B, a broadrange of metabolites, with varying influences on class separation,was established. Carotenoids and EGCG were the most importantcontributors to the observed variance between cultivars, and ami-no acids, with the exception of theanine and methionine, wereamong the least important contributors. A list of top metabolites(VIP > 1) was generated by OPLS-DA (Supplementary Table 2),including catechins, carotenoids, caffeine and amino acids. These
d the network of related metabolic pathways. 2-OG, 2-oxogluatarate; 3-PGA, 3-, chlorophyll b; dKae, dihydrokaempferol; GAP, glyceraldehyde 3-phosphate; GGPP,ne synthetase; hSer, homoserine; lCya, leucocyanidin; lDel, leucodelphinidin; mt,chlorophyllide; PEP, phosphoenolpyruvic acid; Phy-PP, phytyl pyrophosphate; Pyr,

104 L. Feng et al. / Food Chemistry 155 (2014) 98–104
compounds exhibited the greatest contribution to the cluster for-mation within the metabolite profiles. Similar results were ob-tained with respect to the identification of metabolites with themost influence on class separation and the greatest importancefor the differentiation of analysed tea cultivars (SupplementaryFig. 1), based on the principal components analysis (PCA) (Roessneret al., 2001; Trygg et al., 2007).
Taken together, our results, for significant changes in metabolicprofiles of albino tea leaves and the network of related metabolicpathways, are summarised in Fig. 5. The increase in the abundanceof total amino acids (especially theanine and Glu) and decrease inthe levels of catechins and caffeine in the albino leaves contributedto their reduced astringency and bitterness and imparted a strongumami taste. As a high Chl a/b ratio is usually associated with in-creased photosynthetic capacity and increased zeaxanthin isimportant for photoprotection by dissipating excess light energy,our results suggest that albino tea plants can modulate their pho-tosynthetic apparatus to compensate for the changes in photosyn-thetic efficiency and capacity in albino leaves. OPLS-DA analysisrevealed significant differences between the metabolic profiles ofalbino and normal tea cultivars and identified the metabolites pre-dominantly responsible for the observed variance. We speculatethat the biosynthetic inhibition of chlorophylls, carotenoids andcaffeine in albino leaves might lead to a shift in the metabolic net-work towards amino acid biosynthesis.
4. Conclusion
Compared to the normal cultivar, albino tea leaves had a higherchlorophyll a/b ratio and increased abundance of zeaxanthin andfree amino acids, along with lower levels of total carotenoids, cat-echins and caffeine. We suggest that changes in some key metab-olites, such as EGCG, theanine, Glu and caffeine, are responsible forthe decreased astringency and bitterness and enhanced umamitaste of albino tea. Moreover, OPLS-DA identified key determinantsbetween albino and normal control tea cultivars.
Acknowledgements
We thank Dr. Hua Yang and Lin-Long Ma at the Anhui Agricul-tural University for genetics analysis and data processing. Thiswork was funded by the National Science Foundation in China(#31070614), the Research Fund for the Doctoral Program of High-er Education of The ministry of Education (#20123418110002), theProgram for Changjiang Scholars and Innovative Research Team inUniversities (IRT1101), and the ‘‘Twelfth Five-Year’’ National KeyBasic Research and Development Project (973) in China(2012CB722903).
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at http://dx.doi.org/10.1016/j.foodchem.2014.01.044.
References
Alcázar, A., Ballesteros, O., Jurado, J. M., Pablos, F., Martín, M. J., Vilches, J. L., et al.(2007). Differentiation of green, white, black, Oolong and Pu-erh teas accordingto their free amino acids content. Journal of Agricultural and Food Chemistry, 55,5960–5965.
Ashihara, H., Deng, W. W., Mullen, W., & Crozier, A. (2010). Distribution andbiosynthesis of flavan-3-ols in Camellia sinensis seedlings and expression ofgenes encoding biosynthetic enzymes. Phytochemistry, 71, 559–566.
Cabrera, C., Artacho, R., & Gimênez, R. (2006). Beneficial effects of green tea – Areview. Journal of the American College of Nutrition, 25, 79–99.
Chaturvedula, V. S. P., & Prakash, I. (2011). The aroma, taste, color and bioactiveconstituents of tea. Journal of Medicinal Plants Research, 5, 2110–2124.
Chen, Y. L., Duan, J., & Jiang, Y. M. (2011). Production, quality and biological effectsof Oolong Tea (Camellia sinensis). Food Reviews International, 27, 1–15.
Chu, D., Kobayashi, K., Juneja, L. R., & Yamamoto, T. (1997). Theanine-its synthesis,isolation, and physiological activity. In T. Yamamoto, L. R. Juneja, D. Chu, & M.Kim (Eds.), Chemistry and Applications of Green Tea (pp. 129–135). Boca Raton:CRC Press.
Demmig-Adams, B., Gilmore, A. M., & Adams, W. W. (1996). III: Carotenoids 3: Invivo function of carotenoids in higher plants. FASEB Journal, 10, 403–412.
Deng, W. W., Wang, S., Chen, Q., Zhang, Z. Z., & Hu, X. Y. (2012). Effect of salttreatment on theanine biosynthesis in Camellia sinensis seedlings. PlantPhysiology and Biochemistry, 56, 35–40.
Du, Y. Y., Chen, H., Zhong, W. L., Wu, L. Y., Ye, J. H., et al. (2008). Effect of temperatureon accumulation of chlorophylls and leaf ultrastructure of low temperatureinduced albino tea plant. African Journal of Biotechnology, 7, 1881–1885.
Du, Y. Y., Liang, Y. R., Wang, H., Wang, K. R., Lu, J. L., & Zhang, G. H. (2006). A study onthe chemical composition of albino tea cultivars. Journal of Horticultural Scienceand Biotechnology, 81, 809–812.
Ekborg-Ott, K. H., Taylor, A., & Armstrong, D. W. (1997). Varietal differences in thetotal and enantiomeric composition of theanine in tea. Journal of Agriculturaland Food Chemistry, 45, 353–363.
Fernández-Marín, B., Míguez, F., Becerril, J. M., & García-Plazaola, J. I. (2011).Activation of violaxanthin cycle in darkness is a common response to differentabiotic stresses: A case study in Pelvetia canaliculata. BMC Plant Biology, 11,181–185.
Johnson, M. P., Havaux, M., Triantaphylidès, C., Ksas, B., Pascal, A. A., Robert, B., et al.(2007). Elevated zeaxanthin bound to oligomeric LHCII enhances the resistanceof Arabidopsis to photooxidative stress by a lipid-protective, antioxidantmechanism. Journal of Biological Chemistry, 282, 22605–22618.
Kim, E. H., Li, X. P., Razeghifard, R., Anderson, J. M., Niyogi, K. K., Pogson, B. J., et al.(2009). The multiple roles of light-harvesting chlorophyll a/b-proteincomplexes define structure and optimise function of Arabidopsis chloroplasts:A study using two chlorophyll b-less mutants. Biochimica Biophysica Acta, 1787,973–984.
Ku, K. M., Choi, J. N., Kim, J., Kim, J. K., Yoo, L. G., Lee, S. J., et al. (2010). Metabolomicsanalysis reveals the compositional differences of shade grown tea (Camelliasinensis L.). Journal of Agricultural and Food Chemistry, 58, 418–426.
Le Gall, G., Colquhoun, I. J., & Defernez, M. (2004). Metabolite profiling using (1)HNMR spectroscopy for quality assessment of green tea, Camellia sinensis (L.).Journal of Agricultural and Food Chemistry, 52, 692–700.
Li, Q., Huang, J., Liu, S., Li, J., Yang, X., Liu, Y., et al. (2011). Proteomic analysis ofyoung leaves at three developmental stages in an albino tea cultivar. ProteomeScience, 9, 44–49.
Liang, Y. R., Ma, W. Y., Lu, J. L., & Wu, Y. (2006). Comparison of chemical compositionof Ilex latifolia Thumb and Camellia sinensis (L.). Food Chemistry, 75, 339–343.
Meier, S., Tzfadia, O., Vallabhaneni, R., Gehring, C., & Wurtzel, E. T. (2011). Atranscriptional analysis of carotenoid, chlorophyll and plastidial isoprenoidbiosynthesis genes during development and osmotic stress responses inArabidopsis thaliana. BMC System Biology, 5, 77–81.
Melis, A. (1991). Dynamics of photosynthetic membrane composition and function.Biochimica et Biophysica Acta, 1058, 87–106.
Mohanpuria, P., Kumar, V., & Yadav, S. K. (2010). Tea caffeine: Metabolism,functions, and reduction strategies. Food Science and Biotechnology, 19, 275–287.
Narukawa, M., Kimata, H., Noga, C., & Watanabe, T. (2010). Taste characterisation ofgreen tea catechins. International Journal of Food Science & Technology, 45,1579–1585.
Polívka, T., & Frank, H. A. (2010). Molecular factors controlling photosynthetic lightharvesting by carotenoids. Accounts of Chemical Research, 43, 1125–1134.
Ravichandran, R., & Parthiban, R. (2000). Lipid occurrence, distribution anddegradation to flavor volatiles during tea processing. Food Chemistry, 68, 7–13.
Roessner, U., Luedemann, A., Brust, D., Fiehn, O., Linke, T., Willmitzer, L., et al.(2001). Metabolic profiling allows comprehensive phenotype of genetically orenvironmentally modified plant systems. Plant Cell, 13, 11–29.
Tanaka, R., & Tanaka, A. (2007). Tetrapyrrole biosynthesis in higher plants. AnnualReview of Plant Biology, 58, 321–346.
Trygg, J., Holmes, E., & Londstedt, T. (2007). Chemometrics in metabonomics. Journalof Proteome Research, 6, 469–479.
Walter, M. H., Floss, D. S., & Strack, D. (2010). Apocarotenoids: Hormones,mycorrhizal metabolites and aroma volatiles. Planta, 232, 1–17.
Wei, K., Wang, L., & Zhou, Y. J. (2012). Comparison of catechins and purine alkaloidsin albino and normal green tea cultivars (Camellia sinensis L.) by HPLC. FoodChemistry, 130, 720–724.
Yu, B. Y., Lydiate, D. J., Schäfer, U. A., & Hannoufa, A. (2007). Characterisation of a-carotene hydroxylase of Adonis aestivalis and its expression in Arabidopsisthaliana. Planta, 226, 181–192.