die Änderung der chemischen resistenz pflanzlicher plasmen mit dem entwicklungszustand
TRANSCRIPT

Die ~nderung der ehemischen Resistenz pflanzlicher Plasmen mit dem Entwieklungszustand
Woll
Richard Biebl und Waldtraut Rossi-Pillhofer
Aus dem Pflanzenphysiolog~is&en Institut der Universit~it Wien
Mit 9 Textabbildungen
(Eiugelangt am 12. M~rz 1954)
Einleitung
Die Erforschung der Resistenzeigenschaften der Pflanze gegenfiber chemischen Einfliissen z~ihlt heute mit zu den meist untersuchten Fragen der Pflanzenphysiologie. Mit dem Bekanntwerden der Wirkung kleinster Stoffmengen, seien es die fiir die Ern~ihrung wichtigen Spurenelemente (Lit. b. S c h a r r e r 1944, Bibliography 1948, 1951, 1953 u. a.) oder die fiir das Wachstum ent.scheidenden Wuchsstoffe (Lit. b. L i n s e r 195l, F r o h b e r g e r 1951, B l a c k m a n n 1952 u. a.), ergab sich die Notwendigkeit, die optimalen Mengen gegen grSl~ere, sch~idigende Do,sen abzugrenzen. Dabei wurde vor allem die Wirkung der dutch den Boden, dutch Bespriihen oder durch In- jek/ionen verabreichten Stoffgaben auf versehiedenste physiologische Leistungen, wie Wachstum, Atmung, Assimilation, Chlorophyllbildung usw., untersucht.
Die Zellphysiologie beniitzte ihrerseits schon lange die verschiedene Resistenz pflanzlicher Protoplaste gegen ~iul~ere Einwirkungen, darunter auch gegen chemische Agenzien, fiir Fragen tier proioplasmatischen Ana- tomie ( W e b e r 1929, R e u t e r 1949) oder der vergleichenden Protoplasma- tik (H 5 f l e r 1932, 1939). hn Bereich 5kologisch orientierter Plasmaforschung zeigte es sich, daft zwi~schen Standortfaktoren und Resistenz gegen diese enge Beziehungen bestehen und daft den Pflanzen eines und desselben Siandortes oft gemeinsame Ziige in ihrer plasmatischen Resistenz zukommen (B ieb l 1939). Vergleicht man jedoeh die Resistenz der Pflanzen eines be- stimmten Standortes gegen Stoffe oder Einfliisse, die an diesem keine Rolle spielen, so hat die Gemeinsamkeit des plasmatischen Verhaltens ein Ende Protoplasma, Bd. XLIV2 9

114 R. Biebl und Waldtraut Rossi-Pillhofer
(Biebl 1952a, b). Die , , iSkologische t { e s i s f e n z " gegen bestimrnte Um- weltfak~oren ist also ftir Pflanzen eines Biofops vielfaeh iihnlieh, die ~m i e h t u m w e l t b e z o g e n e n I l e s i s f e n z e n " k~innen hingegen rectlt versehieden sein (Biebl 1949). Lefztgenannte sind daher besonders geeignet, versehie- dene pflanzliche Plasmasorfen zu kennzeiehnen (Bi eb I 1947 a, 1950 a).
Erfahrungen der letzten Dezennien lehrfen, dal3 die Protoplasmen ver- sehiedener Pflanzen nieht allein untereinander versehiedenartig sind, son- dern daft sie aueh alters- und jahreszeitlieh bedingte Veriinderungen er- fahren kgnnen. Das gilt sowohI fiir die Permeabilif?it (K r as s i n s k y 1929, M a r k l u n d 1956, HiSf ler 1957, H o f m e i s t e r 1938, L e n k 1955 u. a.) wie ftir den osmotisehen Weft ( L i d f o r s s 1907, U r s p r u n g und B l u m 1911. S t e i n e r 1959, P i s e k 1950 u. a.) oder die Viskositiit und die K:~ilferesistenz (Weber und H o h e n e g g e r 1923, K e s s l e r 1935, K e s s l e r und R u h l a n d I958 u. a.). Zus.ammenfassende Darstellungen der Nnderungen der Plasma- eigensehaften beim Altern pflanzlieher Zellen gaben P a e e h (1940) und F i s c h e r (1950). Die GriStle der Sehwankungen ist bet den versehiedenen Pflanzen nieht gleieh.
Siehf man die ,,nieht umwelfbezogenen konsfitutionellen Resisfenzen" als Ansdruck eines gegebenen Plasmazusfandes, so ist zu erwarien, daI~. aneh diese -- in unserem Falle die ehemisehen Resistenzen -- eniwi&]ungs- bedingte Yeriinderungen erfahren werden.
Seholl ill der ersten Untersuehung tiler die Pl.asmaresistenz gegen Zink, Bor und Mangan (Biebl 1947 a) kom~fe an einigen Moo,sen gezeigt werden, daft die Resistenz bet der einen Art wiihrend des ganzen Jahres gleieh bleibt, bet ether anderen hingegen weehself. Es wurde daher in bezug ant den in Frage siehenden St.off zwisehen , , r e s i s t e n z k o n s t a n t e n ' " und , , r e s i s t e n z v e r i i n d e r l i e h e n '~ Plasmen unfersehieden.
Yorliegende Untersuehung betrifft die u d e r ehe- m i s e h e n R e s i s t e n z der Blatf- oder Stengelepidermen verschiedener Bliitenpflanzen im L a u f e i h r e r E n t w i e k l u n g . Beobaehtet wurde die Wirkung yon Borsiiure und "eon Zink-, Mangan- und zum Tell aueh yon Chromsulfat. Um den jahreszeitliehen Einflu~ yore Einflufl des Alters unfer- seheiden zu kiSnnen, wurden an ether Reihe yon Objekten zur gleiehen Jahreszeit versehieden alte Individuen bzw. Organe untersuehf. Die znr Untersuehung besfimmten Kulturpflanzen wurden zu diesem Zweek mehr- reals hintereinander angebaut. Die Mitteilungen stiifzen sieh im wesent- lichen ant die "~on W. P~ossi in ihrer Dissertation (Wien 1951) nieder- gelegten Beobaehtnngen. Es set daher an dieser Stelle yon einer vollst~in- digen Wiedergabe der Protokolle und der Einzelbeobaehfungen abgesehen.
Methodik
Epidermissehnitte de,- Versuehspflanzen wurden in weithalsigem 30cm ,~ fassenden Fl~isehehen tiber 48 Sfunden in die abgestuft konzentrierten L~i- sungen eingelegt und ansehliel~end mikroskopiseh ant ihren Lebenszustand

itnderung d. diem. Resistenz pflanzl. Plasmen 1nit dem Entwickhmg'szustand 115
untersucht. Die Einwirkungsdauer yon 48 Stunden hat sich in den vora~l- gegangenen Arbeiten als hinreichend lange erwiesen. Nach dieser Zeit blei- ben die erreichten Sch~idigungen meist ann~ihernd konstant. Ein weiteres t2bergreifen auf niedrigere Konzentrafionen geht dann nut mehr alhnhhlieh oder oft fiberhaupt nicht mehr vor sich. Als Lebensreaktion diente Plasmo- lyse in 0,5--1,0tool KC1. Kontrollschnitte wurden gleich lange in doppelt destilliertem Wasser gehalten.
lm allgemeinen wurden yon Borshure (H3BO~), ZnSO~, MnSO4 und Cr2(SO,) 5%ige LiSsungen in doppelt dest. H~O hergestellt und aus die- sen 1, 0,1, 0,01, 0,001 und 0,0001%ige Verdiinnungen bereitet. Um die Prozentangaben mit solchen in Mol-Werten vergleichen zu kiinnen, seie~ die Mol-Werte der l%igen Liisungen bzw. die Prozentwerte einmolarer Ltisungen unserer Versuchsstoffe angegeben:
1% H~BO~ ~ 0,16 Mol, 1 Mol - 6,2% 1% Z n S O 4 . 7 H 2 0 ---- 0,0_54 ,, 1 ,, ~ 28,7 % :l% Crz(SO~)2.6H:O =0,020 ,, 1 ,, -=50,0% 1% MnSO~.4H.~O =0,044 ,, 1 ,, ~22,5%
Jedes Objekt wurde im Verlauf seiner Entwicklung monatlich ein- bis zweimal untersucht. In den graphischen Darstellungen sind die jeweils nodt vitalen Grenzkonzentrationen angegeben.
1. R e s i s t e n z w i l d w a c h s e n d e r , w i n t e r g r i i n e r P f l a n z e n in d e r Ze i t yon M~irz J949 bis A p r i l 1950
Um die Beobachtungen auch fiber den Winter fortffihren zu kSnnen, wurden zu dieser ersten Versuchsreihe Pflanzen mit wintergriinen Bl~ittern gewhhlt, und zwar Anemone hepatica, Cyclamen europaeum und Hedera helix. Untersucht wurde die Epidermis der Blattunterseite und des Blatt- stieles, bet Hedera yon Sonnen- und Schattenbl~ittern. In jedem Versuch wurden vier Schnitte beobachtet.
Der Zeitpunkt der Blattentfaltung fiillt bet den drei Pflanzen nicht in denselben Monat. Bei Anemone hepatica beginnt die Beblhtterung durch- schnittlich im April und bleibt auf kurze Zeit beschr~inkt, w/ihrend die Bl~itter des Vorjahres allmghlich absterben. Bet Cyclamen europaeum finden sich besonders im Juli junge Bl~itter, w~ihrend die alien Bliitter, die den Winter fiberdauert haben, his Oktober erhalten bleiben. Hedera helix schliefilich setzt mit der Neubebl~itterung im allgemeinen im April ein, wobei die Bl~itter h~iufig im selben ~[ahr nicht mehr ihre voile GrSl~e erreichen. Man kann bet Hedera bis zu einem gewissen Grad aus dem Aus- sehen der Bl~itter auf ihr Alter schliel~en. Sie werden grS~er, dunkler grfin, ledriger und verlieren den feinen, an der Unterseite ganz junger BlOtter vorhandenen Flaum. Die Epidermen der Unterseite iiberwinternder Bliitter sind meist anthokyanhgltig. Sonnen-und Schattenbl~itter unterscheiden sich
9*

116 R. Biebl und Waid t rau t Rossi-Pil lhofer
z u d e m w e s e n t l i e h d u r e h i h r e G e s t a l t . D i e S o n n e n b l i i t t e r s ind o f t f a s t
e i f S r m i g , g rS f l e r u n d h e l l e r g r t in .
a) Anemone hepatica
T a b . I g i b t e in B e i s p i e l d e r P r o t o k o l l f i i h r u n g u n d z e i g t g l e i e h z e i t i g d a s
u m e r e i n a n d e r m e i s t s e h r g u t i i b e r e i n s t i m m e n d e R e s i s t e n z v e r h a l t e n d e r
v i e r P a r a l l e l s e h n i t t e . F e s t g e h a l t e n s i n d in d i e s e m B e i s p i e l d i e R e s i s t e n z -
Tab. 1. Anemone hepatica, R e s i s t e n z v e r s c h i e d e n a l t e r B l ~ i t t e r ge- g e n B o r s f i u r e , Zn-, C r - u n d M n - s u l f a t i n d e r Z e i t y o n M f i r z b i s J u l i . (4 = alles lebend, 3 = mehr als die H/i l f te lebend, 2 - ungef~ihr die H~lf te leben& J = einige Zellen lebend, 0 = alles tot. In den Kurvenda r s t e l lungen wurde ein Sdmi t t als , , lebend" bezeiehnet, w e a n alle oder zumindes t fast alle Zellen lebend waren.)
% H3BO;~ ZnSOa Cr~(SOd~ MnSO,~ % MnSO.,
17. 3. 49, Blair vom Vor jahr 5 0000 0000 0000 4444 20 4444 1 0000 0000 0000 4444 15 4444 0,1 4444 0000 0000 4443 10 4443 0,01 4444 0000 0000 4444 8 4444 0,001 4444 4444 0000 4444 5 4435 0,0001 4444 4444 3322 4444 4 4444
3. 4. 49, Blat t vom Vor jahr 5 0000 221t 0000 4442 20 0000 1 4322 1111 0000 4444 15 3522 0,1 3322 0000 0000 4444 10 4444 0,01 4433 3222 0000 4444 8 4331 0,001 4444 4444 0000 4444 5 2211 0,000l 4444 4444 0000 4444 4 2211
4. 5. 49. Blat t vom u 3 0000 3211 0000 4422 20 1100 l 3511 2100 0000 4444 15 1100 0,1 4443 2100 0000 4444 10 3322 0,01 4444 1100 0000 4322 8 3211 0,001 4444 4352 2000 4452 5 3522 0,0001 4444 4422 2100 4420 4 4444
4. 5. 49, F r f ih jahrsb la t t 1949 5 0000 0000 0000 0000 1 4444 0000 0000 0000 0,1 4444 0000 0000 0000 0,01 4444 0000 0000 0000 0,001 4444 0000 0000 0000 0,0001 4444 4000 2200 4443
6. 7. 49, Blat t yon 1949 0000 0000 0000 0000
l 0000 0000 0000 0000 0,I 4444 0000 0000 0000 0,01 4444 4200 0000 0000 0,001 4444 4444 0000 4444
0,0001 4444 4444 0000 4444
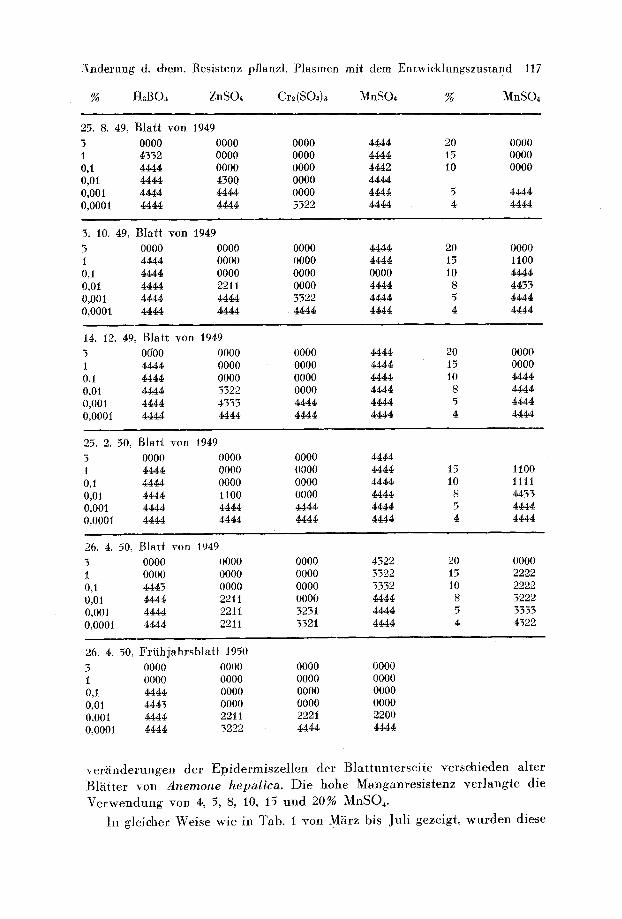
-~inderung d. chem. Res is tenz pflanzl . P l a s m e n mi t dem E n t w i c k l u n g s z u s t a n d 117
% H~BOa ZnSO~ Cr~(SO~)~ MnSO~ % MnSO~
25. 8. 49, Bla i r yon 1949 5 0000 0000 0000 4444 20 0000 1 4552 0000 0000 4444 15 0000 0,I 4444 0000 0000 4442 i0 0000 0,01 4444 4500 0000 4444 0,001 4444 4444 0000 4444 5 4444 0o0001 4444 4444 3522 4444 4 4444
3. 10. 49, Bla i r yon 1949 3 0000 0000 0000 4444 20 0000 1 4444 0000 0000 4444 15 1100 0,1 4444 0000 0000 0000 10 4444 0,01 4444 2211 0000 4444 8 4455 0,001 4444 4444 3522 4444 5 4444 O,O00t 4444 4444 4444 4444 4 4444
14. 12. 49, B l a t t yon 1949 3 0if00 0000 0000 4444 20 0000 1 4444 0000 0000 4444 15 0000 0,1 4444 0000 0000 4444 10 4444 0,0l 4444 3322 0000 4444 8 4444 0,001 4444 4353 4444 4444 5 4444 0,0001 4444 4444 4444 4444 4 4444
25. 2. 50, B la t t yon 1949 3 0000 0000 0000 4444 1 4444 0000 0000 4444 0,1 4444 0000 0000 4444 0,0l 4444 1100 0000 4444 0,001 4444 4444 4444 4444 0,0001 4444 4444 4444 4444
15 1100 10 1111
S 4453 5 4444 4 4444
26. 4. 50, Bla i r yon t949 3 0000 0000 0000 4522 20 0000 1 0000 0000 0000 5522 15 2222 0,1 4445 0000 0000 5332 10 2222 0,01 4444 2211 0000 4444 S 5222 0,001 4444 2211 5251 4444 5 5555 0,0001 4444 2211 5521 4444 4 4322
26. 4. 50, F r f i h j a h r s b l a t t 1950 3 0000 0000 0000 0000 1 0000 0000 0000 0000 0,1 4444 0000 0000 0000 0,01 4445 0000 0000 0000 0,001 4444 2211 2221 2200 0,0001 4444 5222 4444 4444
v e r ~ i n d e r u n g e n d e r E p i d e r m i s z e l l e n d e r B l a t t u n t e r s e i t e v e r s c h i e d e n a l t e r
Bl~ i f l e r y o n Anemone hepatica. D i e h o h e M a n g a n r e s i s t e n z v e r l a n g i e d i e
V e r w e n d u n g y o n 4, 5, 8, 10, 15 u n d 2 0 % M n S O , .
I n g l e i c h e r W e i s e w i e i n T a b . i y o n M~irz b i s J u l i g e z e i g t , w u r d e n d i e s e
t18 ll. Biebl und Waldtraut Rossi-Pillhofer
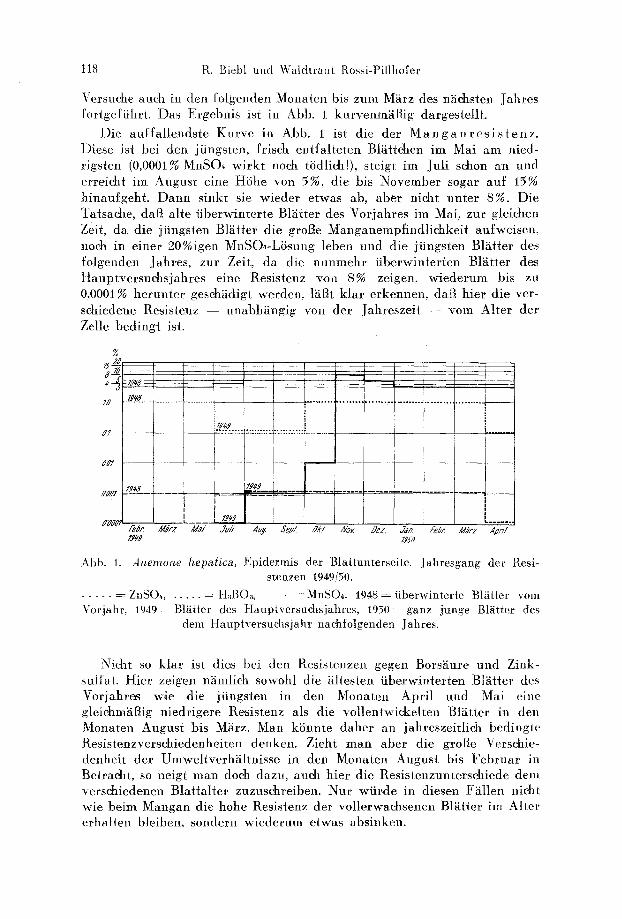
Versuehe audl in den folg'enden Monaten bis zum M~irz des niidrsten Jahres forigefiihrt. Das Ergebnis ist in Abb. I kurvenmii[~ig dargestellt.
Die auffa l lendste Kurve in Abb. 1 ist die der M a n g a n r e s i s t e n z . Diese ist bei den jiingsten, frisch entfa l te ten Bliittehen im Mai am nied- rigsten (0,0001% MnSO'~ wirkt noeh tSdlieh!), steigt im Juli sehon an und erreieht im August eine HShe yon 5%, die his November sogar auf 15% hinaufgeht. Dann sinkt sie wieder etwas ab, aber nieht unter 8%. Die Tatsaehe, da~ alte i iberwinterte Bliitter des Vorjahres im Mai, zur gleiehen Zeit, da die jiingsten Bliitter die grofte Manganempfindliehkeit aufweisen, noeh in einer 20%igen MnSO~-LSsung leben und die jiingsten Bliitter des folgenden ]ahres, zur Zeit, da die nunmehr f iberwinterten Bltitter des Hauptversuehsjahres eine Resistenz rot1 8% zeigen, wiederum his zu 0,0001% herunter gesehiidigt werden, lh[ft klar erkennen, dal~ bier die ver- schiedene Resistenz - - unabhiingig yon der Jahreszeit - - Yore Alter der Zelle bedingt ist.
%
$~ --I-- r ~ :72.<7 - - - - -
I i
oool i! 000~ ~h/: Mdrz
I ...... I
!zdcy i t"::::-::::~
I
I, r ,~al' dub' Au~ ,fer Ok~ Ho~ Bez.
I I
q
] 1 t i
J d~/z /-e~r. MJrz /~SD'
I I
t [ //dr//
Abb. l. Anemone hepatica, Epidermis der Blattunterseite. Jahresgang der Resi- stenzen 1949i50.
. . . . . . ZnSO~, . . . . . - H:~BO:~, -- MnSO~. 1948 = fiberwinterte Bl~itter yore Voriahr, 1949- B1/itter des Hauptversuchsjahres, i950 = ganz junge Bl/itter des
dem Hauptversu&sjahr nachfolgenden Jahres.
Nicht so klar ist dies bei den Resistenzen gegen Borsiiure und Zink- suJfat. Hier zeigen niimlich sowohl die ~iltesten i iberwinterten Bliitter des Vorjahres wie die jiingsten in den Monaten Apri l und Mai eine gleiehmiil3ig niedrigere ttesistenz als die vollentwickelten Bliitter in den Monaten August bi,s M~irz. Mail kSnnte daher an jahreszeitlieh bedingte Resistenzversehiedenheiten denken. Zieht man aber die gro[te Versdlie- denheit der Umweltverhiil tnisse in den Monaten August his Feb rua r in Betraeht, so neigt man d oeh dazu, aueh bier die Resistenzuntersehiede dem verschiedenen Blat tal ter zuzusehreiben. Nur wtirde in diesen Fal len nieht wie beim Mangan die hohe Resistenz der vollerwaehsenen Bliitter im Alter erhalten bleiben, sondern wiederum etwas absinken.
_-r3;nderung d. &em. Resistenz pflanzl. Plasmen mit dem Entwieklungszustand 119
b) Cyclamen europaeum
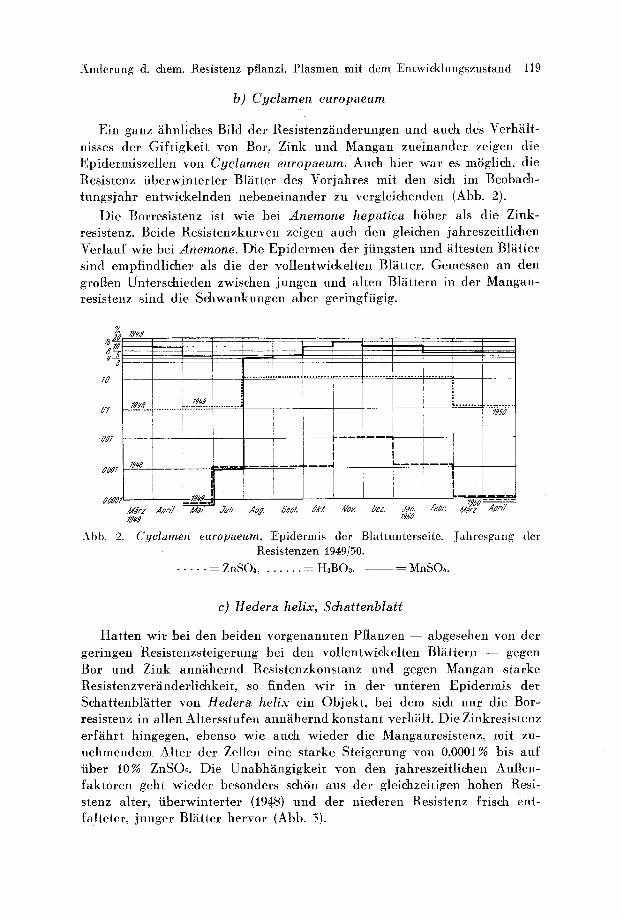
Ein ganz ~ihnliehes Bild der Resistenz~inderungen und aueh des VerhMt- nisses der Giftigkeit yon Bor, Zink und Mangan zueinander zeigen die Epidermiszellen yon Cyclamen europaeum. Auch hier war es m6glich, die Resi stenz tiberwjnterter Bl~itter des u mit den sieh im Beobach- tungsjahr entwickelnden nebeneinander zu vergleichenden (Abb. 2).
Die Borresistenz ist wie bet Anemone hepatica htiher als die Zink- resistenz. Beide Resistenzkurven zeigen aueh den gleichen jahreszeitlichen Verlauf wie bet Anemone. Die Epidermen der jiingsten und hltesten BlOtter sind empfindlicher als die der vollentwickelten Bl~itter. Gemessen an den grofien Unterschieden zwischen jungen und alten Bl~ittern in der Mangan- resistenz sind die Sehwankungen aber geringfiJgig.
IY
0~
ON
0~07
%@8
L
Abb. 2.
i i ! !
I I �9 i .......... [ I
i , !
I
78~
J. , i
I
' !
' ...7. I ' ! ! ! . . . . . ~ ,
i t I , i J i
i i i I , i I I t ~
I#s
Cyclamen europaeum, Epidermis der Blattunterseite. Jahresgang der Resistenzen 1949150.
. . . . . . ZnSOa . . . . . . . . H~BO3, -- MnSO~.
r o
A#i'z .4#n7 lg~#
c) Hedera helix, Schattenblatt
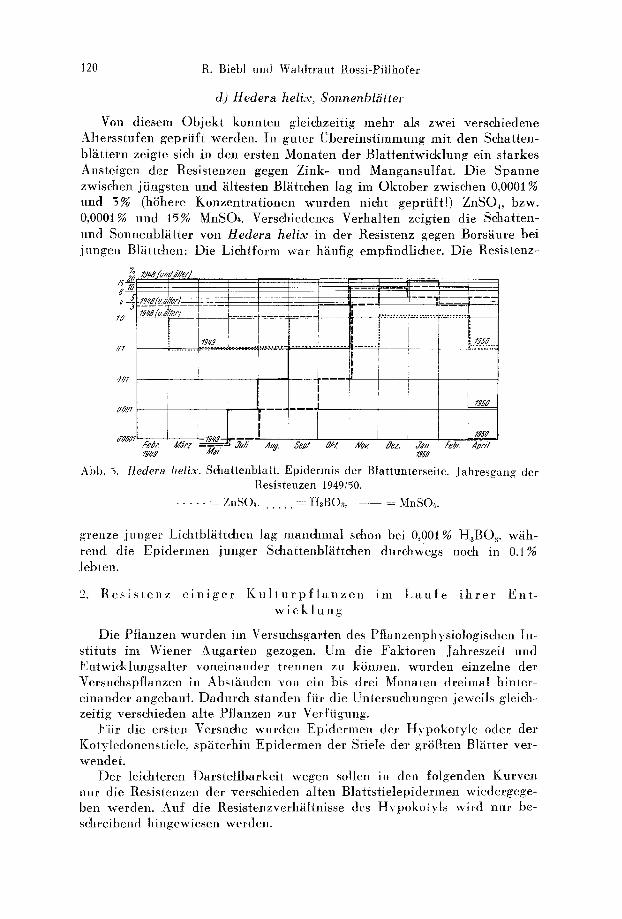
Hatten wir bet den beiden vorgenannten Pflanzen - - abgesehen v o n d e r geringen Resistenzsteigerung bet den vollentwickelten Bliittern - - gegen Bor und Zink amfiihernd Resistenzkonstanz und gegen Mangan starke Resistenzveriinderliehkeit, so linden wir in der unteren Epidermis der Schattenbliitter yon Hedera helix ein Objekf, bet dem sieh nur die Bor- resistenz in allen Altersstufen anniihernd konstant verh~ilt. Die Zinkresistenz erfiihrt hingegen, ebenso wie auch wieder die Manganresistenz, mit zu- nehmendem Alter der Zellen eine starke Steigerung yon 0,0001% bis auf tiber 10% ZnSO~. Die Unabhiingigkeit yon den jahreszeitliehen Aul~en- faktoren geht wieder besonders sehiin aus der gleiehzeitigen hohen Resi- stenz alter, iiberwinterter (194,8) und der niederen Resistenz friscll ent- falteter, junger Bl~itter hervor (Abb. 5).
120 R. Biebl und Waldtraut Rossi-Pillhofer
d) Hedera helix, SonnenbI~tter
Von diesem Objekt konnten gleichzeitig mehr a|s zwei versdfiedene Altersstufen gepriift werden. In guter t~bereinsfimmung mit den Schatten- bliittern zeigte sich in den ersten Monaten der Blaflentwicklung ein starkes Ansteigen der Itesistenzen gegen Zink- und Mangansulfat. Die Spanne zwischen jiingsten und iiltesten Bl~ittchen lag im Oktober zwischen 0,0001% und 3% (hbhere Konzentrationen wurden nicht gepriift!) ZnSO~, bzw. 0,0001% und 15% MnSO< Ver~schiedenes Verhalten zeigten die Schatten- und Sonnenbliitter yon Hedera helix in der Resistenz gegen Borsiiure bei .iungen Bl~ittchen: Die Lichfforln war h~iufig empfindlicher. Die Resistenz-
~o :~ndgTler}
10
0o7
;01
00g/
0~00"/
I r 1 - , 7 Q / 7 5 ~ 7 ) - - i - - - ]
i I
I i i
i I
i y..._= :ca,: bld:z --~eT" 3al:" Ave /.9#$
I I i
Ti J
I
Sep/ 0/'/. AIo~
} " I i
! . . . . . . . . . . I? : :',s3: i i V ".'.J2.V..
i
i :YSd i i- :.gYo
Dez. 3e~ i-e#,: ,4or:~ 7550
Abb. 5. Hedera helix. Sehattenblatt. Epidermis der Blattunterseite. Jahresgang der Resistenzen 194:9./50.
. . . . . . . ZnSO4. =: HaB()a. = MnSO,~.
grenze junger Lichtbliittehen lag manehmal sehon bei 0,001% HaBOa, wiih- rend die Epidermen junger Sehattenbliittehen durehwegs noeh in 0,1% Jeblen.
2. R e s i s t e n z e i n i g e r K u l t u r p f l a n z e n im L a u f e i h r e r Ent - w i e k l u n g
Die Pilanzen wurden im Versuehsgar/en des Pflanzenphysiologisehen In- stituf,s im Wiener Augarten gezogen. Um die Faktoren Jahreszeit und Entwieklungsalter voneinander trennen zu k6mmn, wurden einzelne der Versuehspflanzen in Abstiinden yon ein his drei Monaten dreimal hinter- einauder angebaut. Dadureh standen fiir die Untersuehungen jeweils gleieh- zeitig versehieden alte Pflanzen zur Verfiigung.
Fiir die ersten Versuehe wurden Epidermen cler Hypokotyle oder der Kotyledonenstiele. sp~iterhin Epidermen der Stiele der griSlY.ten Bl~itier ver- wende t.
Der leieh/eren Darstellbarkeit wegen sollen in den folgenden Kurven nut die Resistenzen der versehieden alten Blattstielepidermen wiedergege- ben werden. Auf die Resisfenzverhiiltnisse des Hypokotyls wird nur be- s&reibend hingewiesen werden.

;S, nderung d. chem. Resistenz pflanz]. Plasmen mit dem Entwicklungszustand 121
a) Raphanus satious L. f. albus D. C., Miinchner Bierrettich, mei[~
Die Resistenzen der Blattstielepidermen gegen Bors~iure, Zink- und Mangansulfat (Abb. 4) iinderten sich unter dem Einflult des Alters und zum Teil wohl auch unter dem der Jahreszeit. Im allgemeinen stieg die Resisteuz der Epidermiszellen yon Hypokotyl, Kotyledonenstiel und Blattstiel mit dem Alter der Pflanzen an. Die Resistenzen der Hypokotyle gegen Mangan lagen wesentlich, die der Kotyledonenstiele um ein geringes hSher als die junger Blatts/iele.
Die Resistenz gegen Mangansulfat lag scho~ bei jungeu Blattstielepi- dermen im Vergleich zu anderen Pflanzen hoch. Die Lebeusgrenze glcich
.4z/~q8#21.- 4~ s I~r
- 7 - - - -
1~7 - - l - -
1 i
Y7
I oo~ Z ~ d ~
oooo~ ,.Tun/ Jl/// Av~, ,5"e#t. Ok/,
,4z/58a,~F Z~ 3.19~,4 ,4u333,~I.' .~ ~ lYg, o
.... J. i. r i
I
I
J �9 ,f/vz ,geol. OA'/, N o ~ Ok/. A/o~
Abb. 4. Raphanus satious L. [. albus, D. C., Miinchner Bierrettich, weifl. Blatt- stielepidermis. Resistenz~inderungen im Laufe der Entwicklung.
- - - -- ZnSO~, �9 . . . . . H.~BO3, -- MnSO,,
junger Blattstielepidermen lag, unabh~ingig ~on der Jahreszeit, in allen drei Versuchen (August, September, November) bet 0,01% und stieg bis zu einem bestimmten Enfwicklungszustand auf 3 % an. Dieser Zustand war bet der ersten Aussaat im September, bet der zweiten im November erreicht. Eine im ]uni untersuchte alte Pflanze hatte kurz vor dem Bltihen glei&- falls ihre Resistenzgrenze bet 3%.
Bemerkenswert ist, daft bet Pflanzen mittleren Alters Konzentrationen yon 0,1 und 1% MnSO'~ hiiufig schiidlicher wirkten als 3%ige LiJsungen, w/ihrend die Epidermen alter Pflanzen in allen diesen Konzeutrationen am Leben bleiben.
Fiir Zinksulfat ergab sich folgendes Bild der Resistenz: Hypokotyle hatten nut im Juni ihre Lebensgrenze bet 0,0001% ZnSO~, im Juli, Sep- tember und Oktober aber bet 0,001% und im November sogar erst bet 0,01%. Parallel dazu lag die Lebensgrenze der Blattstielepidermen im August bei 0,001% und im November fiir die verschieden alten BlOtter aller drei Versuche einheitlich bet 0,0I%. Es scheiuen demnach bier engere Be- ziehungen zwischen Resistenz und Jahreszeit zu bestehen als zwischen Re- sistenz und Alter.

122 R. Biebl und Waldtraut Rossi-Pillhofer
In der Resistenz gegen Bors~iure liet]en sich keine besonderen Beziehun- gen zu Jahreszeit oder Alter feststellen. Die Resistenz war in den ver- schiedenen Altersstufen ann~ihernd konstant. Die Lebensgrenzen lagen bet 0.1 oder 1% H~BO~.
- 7C 8~
/0
07
001
0~0:
0~00/
b) Brassica napa oar. Rapa (L.) Thellung, Wei[}e Riibe, Turnips
Es wurden zwei Aussaaten ant 4. Juni und am 24. August gemacht und ab Juli bzw. ab Mitte September laufend untersueht. Die Resistenzen der Blattstielepidermen gegen Borsiiure, Zink- und Mangansulfat iinderten sidl
bier wiederum deutlich mit Auss28L- qs 7YCY
__~:.:.:...
4r
~,
A~$sJJ:~8 7J#Y
l...i ....... .........
iii
i i I
i ' I
f ' I I
zv: Se:,~ :,:/. J e m " Ju~" 4/oz Sept OM Noz
Abb. 5. Brassica napa oar. Rapa, weifle Riibe. Blattstielepidermis. Resistenzen im Laufe der Ent-
wicklung. . . . . . . ZnSO~, . . . . . . H.~BOa, MnSO~,
MnSO~.
dem Alter der Bliitter, uu- abh~ingig yon der Jahres- zeit (Abb. 5).
Die Lebensgrenze fiir Zinksulfat lag bet den jiingsten Bliittern im Juli und Oktober bet 0,001% und stieg bet der Aussaat im Juli bis November auf 0,l%, bet der Aussaat im Oktober bis November auf 0,01% an.
Die Resistenz gegen Mangansulfat zeigt gleiche Abh~ingigkeit vom Alter und Unabh~ingigkeit yon der Jahreszeit. Die jiing- sten Blattchen der ersten
Aussaat haben im lull ihre Lebensgrenze in 0,1%, die im August bereits auf 8% hinaufsdmellt. Diese hohe Resistenz bleibt bis November mehr oder weniger gleichlnhl}ig erhalten. Die jiingsten Blhtter der zweiten Aussaat haben im September wieder eine niedere Resistenz (0,01%), die aber gleich- falls innerhalb eines Monates auf 5 und im n~ichsten Monat auf 4% MnSO~ ansteigt. Wie beim Rettich finden wir auch hier gelegentlich bet 0,1--1% MnSO4 eine grofie Anzahl abgestorbener Zellem w~ihrend il~ 5% MnSO~ wiederuin alle Zellen leben.
Die Resistenzgrenze gegen Borshure lag im allgemeinen in jeder Alters- stufe bet 1%, nut beim jungen Blattstiel der zweiten Aussaat sch.on bet 0.1%. Hier stieg sie erst sp~iter auf 1% an.
c) Brassica oleracea oar. 8ongylodes, Kohlrabi, blauer Speck
Die Resistenzen gegen Zink- and Mangansulfat stiegen wiederum mit dem Alter, unabh~ingig yon der ]ahreszeit, stark an, die gegen Borshure lag" hingegen bet allen untersuchten Blattstielepidermen, wie auch bei den Hypokotylen, unabhhngig yon Alter und Jahreszeit gleichm~tfiig bet 1% (Abb. 6).

Xnderung d. chem. Resistenz pflanz !. Plasmen mit dem Entwieklungszustand 123
d) Daucus carota, M6hre, gelber Riese, Champion
7y
d7
dY7
OOdl
d ~Yd;,
Die Resistenzverschiedenheiten junger und alter Bl~itter sind bet der MShre geringer als bet den bisher besprochenen Objekten. Die Wider- standsfhhigkeif gegen Mangan- und Zinksulfat nimmt aber gleichfalls mit dem Alter etwas zu. Gegen Bors~iure sank die Resistenz im August und September yon i % auf 0,1%, um Anfang Oktober wieder auf 1% Bors~iure anzusteigen (Abb. 7).
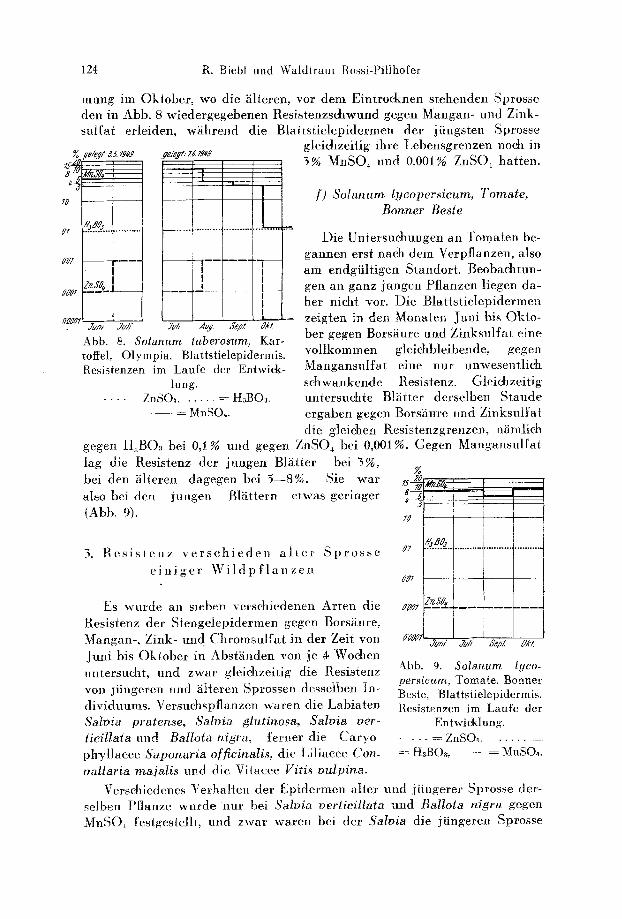
e) Solanum tuberosum, Kartoffel, Olympia
Die ersten Untersu- chungen wurden an Kartoffelpflanzen durch- gefiihrt, die im Mat im ungedtingten Boden ge-
lO
07
O01
OOOl
00007
dussygl .~d lY~ I ' !
I
; E I
I I
~i~ssas! Zr 81,.~zr
Yedd d,~d #az
I 31ii71" YU/l' AaJ. SepA Okt. #av.
Abb. 6. Brassica oleracea var. gongylodes, Kohlrabi. Blattstielerpidermis. Resistenzen im Laufe der Ent-
wicklung. . . . . . . ZnSO,~, . . . . . . HaBO3, -- MnSO,,.
legt wurden. Diese Pflanzen waren schon bei der ersten Untersuehung roll entwid<elt und konnten im August nicht mehr verwendet werden.
Eine zweiie Serie wurde im .&.i3sd, e/.- ~d Lr162
! I
iz~ so~_ ~___ !
~u&~eel Z8 d If'4.] -+
! I
!
3uiz' 3u// Au~ ~r 0~l #or 0~t iVav
ADD. 7. Daucus carota, MShre. Blattstielepi- dermis. Resistenzen im Laufe der Entwicklung.
. . . . . . ZnSO4 . . . . . . . H3BOa, -- MnSO~.
Juni gelegt und im Jugend- stadium (Scm hoeh) erstmals gepriift. Bei der letzten Unter- suchung im Oktober waren diese Pflanzen schon kurz vor dem Abtrocknen (Abb. 8).
Die hohe, Manganresistenz (3--10%) junger und roll ent- wiekelter Bl~itter steht in f)bereinstimmung mit friihe- re n Beobaehtungen (B i e b 1 1950 b). Sie fhllt im Gegensatz zu den bisher besproehenen Objekten bet den ~iltesten Bl'~ttern im Oktober yon 3,0 auf 0,i % ab. Gegen Bors~iure
zeigen demgegeniiber die Epidermen der Blattstiele w~ihrend der gesamten Versuchsdauer unver~indert Resistenzkonstanz. Die Zinkresistenz beginut auf 0,01% und sinkt in den alten Bl~ittern im Oktober his auf 0,0001% ab.
Ein Unterschied verschieden alter Bl~itter ein und derselben Kartoffel- staude ist kaum merkbar, mit Ausnahme bet der letzten Resistenzbestim-
124 R. Biebl und Waldtraut Rossi-Pillhofer
%
1 0
0~
0~7
a~Q]
Oagg;
mung im Oktober, wo die iilteren, vor dem Eintroeknen stehenden Sprosse den in Abb. 8 wiedergegebenen t/esistenzsehwund gegen Mangan- und Zink- sutfat erleiden, wiihrend die Blat ts t ielepidermen der jtingsten Sprosse
gleiehzeifig ihre Lebensgrenzen no& in 3% MnSO~ und 0,001% ZnSO~ bat ten.
$ee/efl 3 S 7-oe~ o oae/egZ" 7s 13~2
~S6-
I z~s4_]
,]uo/ ]a// ]c,/~ Au~ ~fe/JA OA'f
Abb. 8. SoIanum tuberosum, Kar- toffel, Olympia. Blattstielepidermis. Resistenzen im Laufe der Entwiek-
lung. . . . . . ZnSO~ . . . . . . . HaBOa,
-- MnSO~.
D Solarium lycopersicum, Tomate, Banner Beste
Die Untersuehungen an "Fomaten be- gannen erst naeh dem Verpflanzen, Also am endgiiltigen Standoff. Beobaehtun- geI1 an ganz jungen Pflanzen liegen da- her nicht ~or. Die Blat ts t ie lepidermen zeigten in den Monaten Juni his Okto- her gegen Bors~iure und Zinksulfat eine vollkommen gleiehbteibende, gegen Mangansulfa t eine nur unwesentlich sehwankende Resistenz. Gleiehzeitig untersuchte Bliitter derselben Staude ergaben gegen Borsiiure und Zinksulfat die gleiehen Resistenzgrenzen, niimlieh
gegen H:~BO.~ bei 0,I% und gegen ZnSO~ bei 0,001%. Gegen Mangansulfai lag die Resistenz der jungen Bl~itter bei 3 %, g bei den iilteren dagegen bei 5--8%. Sie war l s ~ : ~ 4 / Also bei deft jungen Bliittern etwas geringer s,~ I (Abb. 9). 7~ '
3. t l e s i s t e n z v e r s c h i e d e n a l t e r S p r o s s c e i n i g e r W i l d p f l a n z e n
Es wurde an sieben versdi iedenen Arten die Resistenz der Stengelepidermen gegen Bors~iure, Mangan-, l i nk - mid Chromsulfa t in der Zeit yon Juni bis Oktober in Abstiinden yon je 4 Woehen untersucht, und zwar gleiehzeitig die Resistenz yon jiingeren und ii]teren Sprossen desselben In- dividumns. Versuehspflanzen waren die Labiaten Salvia pratense, Salvia glutinosa, Salvia oer-
~07 '. . . . . . . . . . .
g07
0"OOO7 i 3um" JJ/ ~qe?A Ok/.
Abb. 9. Solarium lyco- persicurn, Tomate, Bonnet Besie, Blattsiielepidermis. Resistenzen im Laufe der
Entwicklung. ticillata und Ballota nigra, ferner die Caryo . . . . . . . ZnSO,~ . . . . . . . . ph3;llacee Saponaria officinalis, die Liliacee Con- == HaBO.~, - . . . . . . MnSO~. oatlaria majatis und die Vitacee Vitis oulpina.
Verschiedenes Verhallen der Epidermen alter und jiingerer Sprosse der- selben Ptlanze wurde nu t bei Salvia oerticillata und Ballota nigra gegen MnSO~ festgestellt, und zwar waren bei der Salvia die jiingeren Sprosse

Anderung d. chem. Resistenz pflanzl. Plasmen mit dem Entwicklungszustand 125
manganresis tenter als die /ilteren, bet Ballota hingegen die ji ingeren manganempfindlicher.
Tab. 2 gibt eine tJbersicht des Schwankungsbereiches der Resistenz gegen die vier verwendeten Stoffe in den Monaten Juni bis Oktober 1%9.
Tab. 2. S c h w a n k u n g s b e r e i c h de r R e s i s t e n z e i n i g e r W i l d p f l a n z e n in den M o n a t e n J u n i bis O k t o b e r (bei Vitis oulpina a ) = a l t e r Trieb,
b) = junge Triebspitze).
Voll erwachsene Pflanzen
(Stengelepidermen) H3B02
Salvia pratensis Salvia glutinosa Salvia verticillata Ballota nigra Saponaria officinalis Convallaria ma]alis Vitis vulpina
1--1% 0,1--0,1% 0,1--0,1%
1--0,1--1% 1--0,1--1%
1--0,1% 0,1--0,1%
Schwankungsbereich tier Resistenz in den Monaten Juni--Oktober
ZnS04
0,001--0,01% 0,001--0,01% 0,001--0,001% 0,001--0,001% 0,001--0,001% 0,01 --0,01% 0,001--0,001%
MnSQ
4--20% 8--15%
0,01--15% 3--20%
10~20% 8--10%
a) 0,01--1--0,01% b) 0,01--3--0,01%
Cr2(S0~)3
0,0001--0,001% 0,001 --0,001% 0,001 --0,0001% 0,001 --0,0001% 0,001 --0,001% 0,001 --0,001% 0,001 --0,001%
Die R e s i s t e n z g e g e n B o r s ~ u r e ist im Verlauf der Beobachtungs- monate konstant bet Saloia pratense, Saloia glutinosa, Saloia oerticillata und Vitis oulpina. Bet Ballota nigra l inden wir ein leichtes Absinken der Resistenz in den Monaten Juli bis September, bet Saponaria in den Monaten Juli und August. Dann steigt die Resistenz wieder auf den Ausgangswert. Bet Conoallaria (Blattscheiden) schliefilich sehen wir ab Ende Juli ein blei- bendes Absinken yon 1,0 auf 0,1% H~BO3.
Die R e s i s t e n z g e g e n Z i n k , s u l f a t ist in den Monaten Juni bis Oktober konstant bet SalDia oerticillata, Ballota nigra, Saponaria offici- halts, Conoallaria majalis und Vitis oulpina. Bet Saloia pratensis und Saloia ghttinosa steigt sie im Juli von 0,001 auf 0,01% an, sinkt aber b e t Saloia pratensis in den n~ichsten Monaten wieder auf 0,001% ab. Bemer- kenswert ist die verhaltnism~ifiig hohe, konstante Zinkresistenz (0,01% ZnS OJ der Ep idermen der Blattscheiden yon Conoallaria majalis, die yon anderen Pflanzen nu t selten oder nu t vori ibergehend erreieht wird. Gleich- zeitig durchgefi ihrte Best immungen an ~iufieren, inneren und chlorophyll- freien inneren Blattscheiden yon Conoallaria majalis ergaben gleiche Resi- stenzgrenzen.
Auffa l lend sind hingegen die grol~en R e s is t e 1l z s c h w a n k u n g e u g e g e n M a n g a n s u l f a t . hn allgemeinen ist die Resistenz sehr grolq. Vita]e Plasmolysen yon Saloia glutinosa und Saloia oerticillata in 15%, yon Saponaria o[ficinalis in 15 und 20% sowie Plasmolysen, die allerdings zum Tode fiihren, in 15 und 20% MnSO4 bet Conoallaria majalis zeigen --- hhn- lich wie dies auch fiir die sehr manganresis tenten Moose gilt (B i e b 1 1949) - - , dal~ nicht e twa die Zellmembran dem Mangansulfa t den Durchtr i t t ver- wehrt, sondern daft die LSsung tatshchlich den Protoplas ten umspiilt. Wie welt diese hohen Manganresistenzen etwa auf ether erschwerten Wegsam-

126 R. Biebl und Waldtraut Rossi-Pillhofer
keit eines besonders widerstandsfhhigen Plasraaleramas und damit einer Verhinderung einer Intrabilifiit (vgl. H S f l e r 1951) beruht, wurde noch nicht untersucht.
Wahrend die meisten dieser Wildpflanzen in jungen und alten Sprofi- epidermen schon ira Juni eine hohe Manganresistenz yon 5 bis 8% MnSO~ zeigten, die in den nachsten Monarch noch his auf 10--20% anstieg, fielen Saloia oerticillata und Vitis ouipina durch zeitweise weitaus grSfiere Manganerapfindlichkeit auf. Saloia oerticillata hatte ira Juni eine Lebens- grenze bet nur 0,01% MnSO,, die sich allerdin~s im Juli auf 1%, ira Septem- ber auf 8 % und ira Oktober auf 15 % erhSh~e. Es ist dies ein Ansteigen, wie es ira vorigen hhnlich z. B. bet Hedera helix und Cyclamen europaeum zu sehen war. Bet Vitis oulpina lag die Resistenzgrenze ira Juni g'leichfalls bet 0,01% MnSO~, stieg ira Juli bet einera dunkelroten alten Trieb ant ~ %, bet einer jungen heIlen Triebspitze auf 3% an, sank abet schon ira September bet beiden wieder auf 0,01% ab.
4. Die R e s i s t e n z v e r s e h i e d e n e r E p i d e r m e n d e r s e l b e n P f l a n z e (Cyclamen persicnm und Cyclamen enropaeum)
Fiir Perlneabilit~itsstudien wurden schon raehrfach verscbiedene Gewebe und ZelIsorten derselben Pflanze herangezog'en ( H S f l e r und S t i e g l e r 1930, H u r c h 1935, H S f l e r 1936, 1937, Ur l 1951, 1952), zu Resi.stenzunCer- suchungen wurde ,d,ies hingegen erst selten getan (R e u t e r 1941, B i e b 1 1950 a).
Bet Cyclamen persicum wurde an verschieden alten Bl~ittern rait roter und griiner Unterseite das Verhalten der unteren und oberen Blattepider- men sowie der Stengelepidermis geprtift. Zura Vergleich wurde Cyclamen e,.tropaeum herangezogen, und zwar eine Waldpflanze rait roter Blattunter- seite und eine Topfpflanze rait griiner Unterseite.
Die Resistenz wurde wiederum gepriift gegen Bors~iure, Zink-, Mangan-, Chromsulfat und aul~erdem gegen Aluminiumchlorid (A1CI~),
Die Resistenzgrenzen der Blattober- und Blattnnterseite yon Cyclamen persicum waren bet den im J~inner und Februar dnrchgefiihrten Unter- suchungen gleich in Zinksulfat (0,001%) und in Mangansulfat (fiber 5%) sowohl bet ~ilteren und jiingeren Bl~ittern wie auch bet solchen rait roter und mit grfiner Blattunterseite. Gegen B ors~iure und Aluminiuraddorid er- wies sich die rote wie auch die griine Blattunterseite empfindlicher als die entsprechenden Blattoberseiten. Letztere lebten noch in 1% Borsgnre gegen- iiber 0,1% bet Blattunterseiten yon Bl~ittern verschiedenen Alters. In Alu- miniumchlorid lebten die Epidermen der Oberseiten in 1% LSsung, die der Blattunterseiten hingegen nut in 0,1%, wobei die griinen Unterseiten der glteren Blgtfer etwas resistenter waren. Die Resistenz der Blattunterseite gegeniiber Chromsulfat war bet Blhttern rait toter Unterseite geringer (0,0001%) als die der Blattoberseite und der griinen Unterseite ~ilterer Bl~itter (0,001%).
Ein V e r g 1 e i e h d e r Resistenzen yon Cyclamen persicum und Cyclamen europaeum ergab ffir die Blattoberseite gleiche Lebensgrenzen in Bors~iure

5_nderung d. &era. Resistenz pflanzl. Plasmen mit dem Entwieklungszustand 127
(1%), Chromsulfat (0,001%) und Mangansulfat (tiber 3%), ftir die Unter- seite gleiche in Zinksulfat (0,001%) und Mangansulfat (fiber 3%) und an- ni~hernd gleiche in Chromsulfat (0,0005%). Hingegen erwies sich die Blatt- oberseite yon Cyclamen europaeum in Zinksulfatl~Jsungen (0,01%) um eine Konzentrationsstufe resistenter als die Blattoberseite yon Cyclamen per- sicum (0,001%) und gegen Aluminiumchlorid mit 0,05% um eine Konzen- trationsstufe empfindlicher als die yon Cyclamen persicum (0,5%). Die Blattunterseite war hingegen bet Cyclamen europaeum resistenter gegen Borshure (1%) als bet Cyclamen persicum (0,1%) und mn ein wesentliches empfindlicher gegen Aluminiumchlorid (0,005%) als bet Cyclamen persicum (0,1 -0,5%).
Der Vergleich der Resistenzen der Epidermen unterseits toter und grtiner Bliitter yon Cyclamen europaeum wurde im Februar vorgenommen. Dazu wurde die Waldpflanze mit roter Blattunterseite unter dem Schnee hervor- geholt, wiihrend die griinbl~ittrige Pflanze schon ein halbes Jahr in Topf- kuliur gehalten worden war. Die Resistenzgrenzen waren ftir a|le unter- suchten Substanzen fast viJllig gleich. Nur gegen Aluminiumchlorid war die grtine Unterseite der Topfpflanzen (0,01%) etwas empfindlicher als die rote Unterseite der Waldpflanzen (0,1%). Auf Grund dieses trotz so ver- schiedener Aufienbedingungen iihnlichen Verhaltens der Cyclameu euro- paeum-Pflanzen dar t man wohl folgern, da~ auch bet den Unterschieden zwischen der Glashauspflanze CYclamen persicum und der Freilandptlanze Cyclamen europaeum die Aufienfaktoren keinen wesentlichen Einflult aus- geiibt haben, sondern dal~ die gefundenen Resistenzverschiedenheiten auf spezifische Plasmaunterschiede hinweisen.
Anzumerken ist noch eine Erscheinung, die auch in friiheren Versuchen gelegentlich zu beobachten war, daft n~imlich niedrigere Konzentrationen manchmal schiidlicher wirkten als hbhere, vor allem auf anthokyanhaltige Epidermen der Blattunterseite. Dies frat bet L~sungen ,on MangansulfaL Aluminiulnchlorid und Borsiiure e~wa bet 0,001%, bet Zinksulfat bet 0,0001% auf. Etwas hbhere Konzentrationen bewirkten dann keine Schhdigung und ersi in noch stiirkeren LiJsungen, bet Mangansulfat z. B. erst tiber 5%, kam es zum allgememen Absferben der Schnitte. Auf diese Erscheinung soil in der Besprechung der Versuche noch zurtickgekommen werden.
Besprechung der Versuehe
Resistenzversuche in abgestuften LiJsungen yon Schwermetallsalzen (Zn, Mn, Cr, V) oder yon Bors~iure haben gezeigt, daft die Protoplasmen ver- scbiedener Pflanzen charakteristische Resis/enzunterschiede besitzen, die in ihrer Kombination geeignet sind, verschiedene pflanzliche Plasmen zu kennzeichnen (Biebl 1947--1950). Einzelbeobachtungen, die auch in einer neueren Untersuchung iiber die Resistenz pflanzlicher Plasmen gegen 2, 4-D (Biebl 1953) ihre Bestiitigung fanden, wiesen ferner darauf him dalt die Resistenz pflanzlicher Zellen gegentiber dem einen oder anderen einwirken- den Stoff sich im Laufe des Jahres hndern kann. Die Untersuchung soleher

128 R. Biebl und Waldtraut Rossi-Pillhofer
chemischer Resistenzhnderungen im Verlaufe der jahreszeitlichen Entwick- lung war Kernpunkt vorliegender Arbeit.
Um den Einflufi der jahreszeitlich verschiedenen Umweltfaktoren yore Alter der untersuchten B!at t -oder Stengelepidermen trennen zu kSnnen, wurden jeweils gleichzeitig verschieden alte Organe untersucht. Dies war entweder dadurch mSg'lich, da[] an Pflanzen ,nit langlebigen, iiberwintern- den Blhttern junge und alte Bl~itter nebeneinander auf ihr Resistenz- verhalten g'eprtift wurden (Anemone hepatica, C ljclamen europaeum, Hedera helix), da[~ die Versuchspflanzen mehrmals im Jahr frisch angebaut wurden, so daf~ zum gleichen Zeitpunkt verschieden alte Pflanzen zur Ver- fiigung standen (Rettich, Weif}e Rtibe, Kohlrabi, MShre~ Kartoffel, Tomate) oder dalt schlie~lich an ein und demselben einj~ihrigen Individuum die Epidermen jiingerer und ~ilterer Sprosse verg'lichen wurden.
Alle diese verschiedenen Untersuchungen erbrachten die gemeinsame Tatsache, daf t d ie m e i s t e n P f l a n z e n im Laufe des Jahres zumindes~ gegeniiber dem einen oder anderen Stoff i h r e c h e m i s c h e R e s i s t e n z ~indern, w oran Alter des Gewebes und j ahreszeitliche Ein{liisse in wech- selnd starkem Ausmal~. beteiligt zu sein scheinen. In den meisten F~illen ist in Abh~ngigkeit yore Alter eine Resistenzsteigerung yon jiingsten Sta- dien bis zur Vollentwicklung zu beobachten. Besonders trifft dies fiir die Resistenz gegen Mangan- und Zinksulfat zu. In Ausnahmefhllen nimmt die Resistenz alter, iiberwinterter Bl~itter wieder ab. Da dann die niedere Resi- stenz in den alten Organen mit der noch niederen der jungen Organe zeit- Itch zusammenfhllt, kSnnte hier auch an einen jahreszeitlichen Einflu~ ge- dacht werden. Und auch in solchen F~illen ist ein Einflul3 der klimatisdlen Verh~iltnisse der Jahreszeit in Betracht zu ziehen, wo Pflanzen, die all- gemein eine Resistenzzunahme yon jung zu alt zeigen, bet einem sp~te- ren Anbau im Jahr schon mit einer hSheren Resistenzstufe beginnen als die friiher angebauten (Rettich, Kohlrabi). Relativ h~iufig finder sieh Resi- stenzkonstanz in allen Altersstufen gegen Bors~ure (z. B. Kohlrabi, Kar- toffel, Tomate, Sal~ia-Arten). Eine seltene Resistenzkon,stanz nicht nut gegen Bors~iure, sondern auch gegen Z i n g und Mangansulfat zeigten in den Beobachtungsmonaten .]uni his Oktober die Sprol3epidermen yon Solanum lycopersicum, und zwar in gleicher HShe in den jiingsten wie in den :~iltesten Seifensprossen. Hier sind Alter und Jahreszeit ohne EinflulL
Y e r s c h i e d e n e E p i d e r m e n yon Cyclamen persicum bzw. yon Cy- clameu europaeum (Blatt ober- und -unterseite, Stengel, Bla~tstiel) zeigen fast gleiche Resistenzeigenschaften. Nut gegen Bors~iure und A1CI~ erwiesen sich sowohl die roten wie auch die griinen Blattunterseiten yon C.yclamen persicum empfindlicher als die entsprechenden Blattoberseiten. Bet unter- seits roten und griinen Bl~ittern yon Cyclamen europaeum hingegen fehlten auch diese Unterschiede.
Auch ~iltere Beobachtungen ergaben h~iufig ein gleiches chemisches Resi- stenzverhalten verschiedener Zellsorten einer gegebenen Pflanze. So be- sitzen griine und gelbliche Mesophyllzellen panaschierter Arbutilon Thomp- sonii-Bl'fitter gleiche Resistenz gegen Borshure, Zink-. Mangan- und Vana-

Xnderung d. chem. Resistenz pflanzl. Plasmen mit dem Entwicklungszustand 129
dylsulfat (B ieb l 1950 c). Au& die Blattsfielepidermen yon fiinf verschie- denen Sorten yon Solanum tuberosum und die Epidermen der Dunkeltriebe yon vier verschiedenen Kartoffelsorfen zeigfen gegen die genannten Stoffe gleichartige Resistenzgrenzen ( B i e b l 1949). Demgegeniiber bot Atropa Belladonna in VOSO~-L5sung ein Beispiel fiir eine gr6flere Empfindlich- keit der Epidermis der Blatfoberseite gegeniiber jener der Blattunterseite und des Blattstiels (Bieb 1 1950 a). Subepidermiszellen zeigten sieh bei ver- schiedenen Pflanzen resistenter als die Epidermen. Ein auffallend gegen- siitzliches Verhalten der Resistenz gegen Bors~iure und Zinksulfa* war schliefllich an den Bliittern des Laubmooses Mnium rostratum zwischen den Blattrand- und Mitielrippenzellen einerseits und den Blattfl~ichenzellen andererseits zu beobachten ( B i e b l 1947b).
[3ber die U r s a c h e n der alters- oder jahreszeitlich bedingten chemischen Resisfenzhnderungen wie iiberhaupt tier Resistenzun~erschiede zwischen verschiedenen Plasmasorten lassen sich bisher noch keinerlei gesicherte An- gaben machen. Ebensowenig ist es m5glich, bestimm~e Beziehungen zwi- sehen chemischem Resistenzverhalten und anderen im zellphysiologischen Versuch faflbaren plasmafischen Ver~inderungen (osmotischer Wert, Per- meabiliSi~, u u sw.) im Verlaufe des Alterns (vgl. P a e c h 1940, F i s c h e r 1950) aufzustellen.
H5 f l e t (1951) schreibt in einer Untersuchung fiber die Plasmolyse mit Natriunxkarbonat: ,,Wenn eine Substanz binnen gleieher Zeif die eine Zelle tStet, die andere am Leben l~iflf, so kann dies zweierlei Ursache haben: das Plasmalemma ist bei den einen Zellen fiir die betreffenden Smffe durch- liis sig, bei anderen nicht, d. h. die Infrabilitht isf hoch bzw. niedrig. Oder die S~offe trefen in beiden F~illen ein, aber die einen Zytoplasmen sind gegen sie unempfindlich, die anderen werden getStet." Diese zwei M5glichkeiten k5nnten durchaus auch fr unsere Resistenzversuche zu~reffen. Danach l@e der Schwerpunkt der Resistenz entweder im Plasmalemlna oder die ver- sehiedene Widerstandsfiihigkeit kiime dem gesamten Plasma zu.
Betrachten wir daraufhin die chemischen Resistenzen unserer Versuchs- pflanzen, so dtirfen wit uns vielleich~ in allen F~illen, in denen die Resi- sfenzgrenzen bei niederen Konzentrationen liegen, yon vornherein fiir den zweiten Fall enfscheiden. Es isf kaum anzunehmen bzw. nichf wahrschein- lich, daft z. B. bei einer letalen Konzentration yon 0,1%, bei welcher die LSsung also zweifellos ins Plasma eingedrungen isf und dieses ge~5tet hal, das Plasmalemma einer Konzenfration yon 0,01% den Eintritt verwehrf hat.
Anders liegt der Fall bei jenen Zellen, die z. B. sogar dutch 15--20%ige (0,66--0,88 Mol) hypertonische MangansulfailSsungen nicht gesch~idigf wer- den und zwei oder mehr Tage (bei einigen Laub- und Lebermoosen his zu acht Tagen beobachtef! B i e b 1 t9~9) in diesen hodlkonzenfrierten LSsungen plasmolysiert liegen kSnnen, ohne dadurch get5tet zu werden. Diese F~ille hhneln sehr den Beobachtungen H 5 f 1 e r s in seiner oben angefiihrten Unfer- suchung, wo manche Desmidiaceen-Zellen Plasmolyse in NazCO3 tagelang ertragen, w~ihrend Gewebszellen yon Anthophyten dutch hypertonische Protoplasma, Bd. XLIV/2 ] 0

130 R. Biebl und WaIdtraut Rossi-Pillhofer
Sodalbsungen entweder sofort gettitet werden odes bald naeh et.folgter Plasmolyse Plasmaaufquellungen erfahren und dann ganz odes his auf dell Tonoplasten absterben. HiSf le r nimmt ftir diese Sonderfglle hoher Resi- stenz gegen Na~CO3 eine ungleiehe Durehlgssigkeit bzw. Sehutzwirkung des '~iul3eren Plasmagrenzsehiehte als hi3ehstwahrseheinlidle Ursache an.
In diesem Zusammenhang kommt l~un den Beobaehtungen fiber die letale Wirkung maneher Substanzen in niedrigeren Konzenfrationen, die in hiihe- sen Konzentrationen dann wieder unseh~idlieh erscheinen und erst bei nod~ st~rkerer Konzentration zum Tod fiihren, besondere Beachtung zu. Wit haben z. B. beim Rettieh und b ei des Weil~en Riibe darauf hingewiesen. Dort fanden sieh h~iufig in 0,1 und 1% MnSO, eine grof~e Anzahl yon ab- gestorbenen Zellen, wiihrend in 5% MnSO, wiederum alle Zellen lebten. Solehe Beobachtungen wurden aueh sehon yon anderen Autoren gemaeht. [ l j i n (1935) erhielt z. B. an Zellen yon Reseda lutea naeh 48stiindigem Auf- enthalt in NaC1 (ebenso in KC1) bei Verwendung vers&iedener Konzen- lrationen folgende Ergebnisse: In 0,5 und 0,8 Mol NaC1 waren al!e Zellen tot (Beginn des Absterbens sehon nach 5 Stunden), in 1.2 Mol lebte ein Tell des Zellen, in 1,8--5,0 Mol aber lebten alle Zetlen. Von 27 yon II.~in unter- suehten Pflanzen verhielten sieh 21 naeh diesem Schema. Die iibrigen starben mit zunehmender Konzentration des Lbsung ab. In unseren Versuchen, wo diese Erseheinung hauptsiiehlieh in MnSO,-Lbsungsreihen zu beobaehten war, war allerdings die zmlehmende Sehadenwirkung mit Ansteigen des Konzentration die Regel und das Absterben in den verdiinnten LSsungen bei gleiehzeitiger Widerstandsfiihigkeit in den konzentrierten die Aus- nahme. Manches seheint audl darauf hinzuweisen, daft diese beiden Ersehei- nungen unter bestimmten, n.och nicht niiher bekannten ~uI~.eren und inneren Umst~inden bei des gleiehen Pflanze auftreten kSnnem flier kSnnten wit also vielleieht annehmen, daft es ab einer bestimmten Konzentration des MnSO~ zu eilmr ,,Abdiehtung" .des Plasmalemmas kommt, wiihrend schwh- ehere Lbsungen noeh leiehter ins Binnenplasma eintreten und dieses sehiidigen k~innen. Betont sei, da|~. es sieh bei unseren Versuehen um MnSO~- Konzentrationen handelte, die noeh weir unter dem plasmolyfisehen Grenz- west lagen. Nut in einzelnen Fiillen, wie z. B. bei den Laub- und Leber- moosen, lagen die letzten letalen Grenzen so hoeh, dal~ die sd~idliehen Konzentrationen bereits in den Bereieh des Hypertonie fielen.
Dal~ sieh Protoplaste maneher Pflanzen, iihnlieh wie in unseren hoeh- konzentrierten Mangansulfatli3sungen, aueh in anderen h~cpertonisehen SehwermetallsalzliSsungen, wie Kupfer- odes Zinksulfat, lange im plasmo- lysiertem Zustand erhalten ki3nnen, gibt aueh P r i n g s h e i m (1924~) an. Des Verfasser teilt mit, da~ er z. B. fast alle Zellen des Laubmooses Muium cuspidatum noeh nach 3 Tagen Plasmolyse in 18--20% Zinksulfat dur& Ubertragung in Wasser wieder deplasmol,,.,sieren und dann mit KNO:, neuerlieh plasmolysieren konnte und dal3 in mit ZnSO~ (14,t%) plasmo- 1ysierten Zellen yon Allium, Rhoeo, Elodea deusa mad Elodea cauadeusi.s naeh 10 Minuten his 5 Stunden no& oft kriiftige PlasmastrSmung zu sehen war. Es ist naeh unseren Erfalarungen wohl als sidles anzunehmen, da[~ be-

Anderung d. chem. Resistenz pflanzl. Plasmen mit dem Entwicklungszustand 131
deutend SCilW~ichere, hypotonische LSsungen innerhalb yon 48 Stunden den Zelltod herbeigefiihrt h~itten. Die htichste an unseren Versuchsobjekten im abgestuflen Reihenversuch beobachtete Zinkresistenz zeigten vollentwickelfe Efeublhfter im Dezember und J~inner mit einer Lebensgrenze bet 10% ZnSO4. Aber auch dies entspricht erst einer Konzentrafion von 0,34 Mol, ist also noeh hypotonisch. Die durchschnittlichen Lebensgrenzen bet den bisher untersuchten Pflanzenepidermen liegen aber viel tiefer, etwa bet 0,01% (~=0,0034Mol) und darunter. Auch Bl~ttehen yon Mnium punctatum und ]/lnium rostratum starben innerhalb yon 48 Stunden in 1 bzw. 0,01% ZnSO, ab (Bieb 1 1947). Hypertonisehe Zinksulfatltisungen wurden nicht gepriift. Solche Versuche whren nachzutragen.
K a h o (1933), ein Schiller L e p e s c h k i n s , ist der Ansicht, daft die Schwermetallsalze bet der Beriihrung mit der Pflanzenzelle auf der Plasma- oberIl~iche eine Schutzsehicht (irreversibel koagullierte Oberfl~ichenschieht) erzeugen, die das Eindringen des Salzes ant einige Zeit verhindert nnd die griJfiere Menge des Plasmas vor mo.mentaner AbtiStung schiitzt. Auch P r i n g s h e i m wies auf diese MiSglichkeit hin.
Nach S c h i n d l e r (1944) wirken Nickel- und Kobaltsalze ,,erh[irtend" auf die Protoplastenoberfliiche. Diese Ver~inderung des Plasmalemmas ~iul]ert sich sehon beim ersten Plasmolyseeintritt dutch fiidiges Ausziehen des Zytoplasmas, also ether versfiirkten Bildung yon Heehtschen Fhden. S c h i n d l e r hebt auch hervor, dal] sich die Salze je nach dem Anion in ihrer Wirkung g~inzlich verschieden verhalten.
Der Annahme HiS f i e f s , daft in bestimmten F~]len die Resistenzunter- SC|liede verschiedener Plasmen ant Verschiedenheiten ihrer PlasmaleInmen zuriickgehen, dart somit erg~nzend hinzugefiigt werden, dal~ die verschie- dene ,,Dichte ~ des Plasmalemmas nicht yon vornherein gegeben sein muff, sondern sieh unter Umst~inden erst bei Einwirkung hiSherer Salzkonzentra- tionen ausbitden kann. In diesen F~illen kiJianten daher niedrigere Konzen- trationen st~irker sch~digen als hiShere. Neu ist, daI~ solche Ver~nderungen des Plasma]emmas nicht al|ein am plasmolytisch abgehobcnen Protoplasten, sondern auch schon im hypotonischen Bereich auftreten kiJnnen.
Auch die Beobachtung yon K a m i y a (1939), daft 1--3 Wochen in Wasser liegende Schnitte der Aufienepidermis yon Zwiebelsehuppen (Allium cepa) yon der Mitre her abzusterben beginnen, so da[~ schliel]lich nur am Rand des Sehnittes ein Ring yon lebenden Zellen erhalten bleibt, l~_[~t sich mit der Vorstel]ung ether sekund~iren Ver:~inderung der :,iu~eren Plasmagrenz- "schiehte vereinen. Daftir wiirden auch die p]asmolytischen Beobachtungen HiSf l e r s (1934) sprechen, wonach an Zwiebelepidermisschnitten bet Plas- molyse mit 0,6 Mol KCI eine Zonenbildung auftritt: eine Randzone mif wechselnden ,pathologischen" Plasmolyseformen, dann eine Zone mit star- ker Kappenplasmolyse und schlie[~lich ein Mittelfeld mit ,,normaler" Konvexplasmolyse.
Unsere Beobachtungen haben welter gezeigt, dal~ die im Yerlauf des Jahres festgestellten Resistenz~nderungen entweder in gleicher Weise alle verwendeten Teststoffe oder abet nnr den einen odor anderen yon ihneI~
10"

132 R. Biebl und Waldtraut Rossi-Pi]lhofer
betreffen kthlnen. So sehen wit z. B. bet Hedera helix die Mangan- und Zinkresistenz yon einer Lebensgrenze bet 0,0001% in jugendliehen Bliitterl~ tin Mat his auf 3--10% in vollentwiekelten Bl~itfern tin November und Dezember ansteigen, whhrend bet Cyclamen europaeum Init gleiehfalls gleich grol]er Zink- und Manganempfindlichkeit tin Mat und ]uli (Lebensgrenze bet 0,0001%) die Resistenz gegen MnSO~ im November bis 10%, die gegen ZnSO~ abet nur auf 0,001% ansteigt. Es miissen sieh also die fiir die Resi- stenz verantwortliehen Zust~inde tin Zytoplasina oder tin Ptasinaleinina im Laufe der Entwicklmlg in elner fiir die beiden Sehwermetallsalze verschie- denen Wei.se ge~indert haben. Die sieh aus Untersuehungen yon L i e b e n und J e s s e r (1936, zit. in S e h i n d l e r 194~) ergebende Tatsaebe, da~ z.B. Nickel-, Koba]t- oder Kupfereiwei~-Verbindungen trotz mancher iiuI~erer Nhnlichkeiten in ihrein feineren cheinischen Aufbau grundlegende Unter- schiede aufweisen, kann vielleieht die untereinander oft gro~e Versehieden- heir in der Mangan-, Zink- oder Chroinresistenz versfiindlich machen.
Wit Iniissen uns einstweilen Init dein Ergebnis beseheiden: ]in Verlauf der jahreszeitlichen Entwicklung bzw. des Alterns erfiihrt das Plasma der Ineisten Pflanzen Yeriinderungen, die eine ganz bedeutende "S~nderung seiner Resistenz gegeniiber verschiedenen cheinisehen Substanzen (H,~BO:, ZnSO~, MnSO~, Cr2SO4 und nach friiheren Untersuehungen u 2,4-D) zur Folge haben kanI1. Wesentlleh ist dabei, da~ diese Resistenzhnderungen gegeniiber den untersuehten Stoffen nieht gleiehlaufend oder gleieh stark zu sein branehen. Nur wenige Plasinen erweiseu sieh wiihrend des ganzen Jahres gegen einen oder gegen einige dieser Stoffe resistenzkonstant. Ob die den Resistenz~inderungen zugrunde liegenden plasmatisehen Ver~inde- rungen tin Binnenplasina oder in besonderen F~illen vielleieht vorwiegend tin Plasinalemlna erfolgen und weleher Art sic sind, wissen wit noeh ni&t.
Zusammeniassung
1. Epidermisschnitte yon Bl~ittern, Blattstielen oder Stengeln der Ver- suetlspflanzen wurden in konzentrafionsabgestufte LSsungsreihen yon H~B(),~ (Bors~iure), ZnSO~', MnSO~ und Cr~(SO+)~ (0,0001--1 bzw. 20%) durdt ~8 Stunden eingelegt und dann auf ihren Lebenszustand untersueht. Die eben no& vitale Grenzkonzentration wurde als Resistenzgrenze bezeiehnet.
2. Uin den jahreszeitliehen Uinwelteinftufl voin Einflul] des Entwick- lungsalters unterseheiden zu ktinnen, wurden an einer Reihe yon Objekten zur gleiehen Jahreszeit versehieden alte Individuen bzw. versehieden al[e Stengel oder Bliitter untersueht.
5. Von Anemone hepatica, Cyclamen europaeum und Hedera helix konnten nebeneinander jmlge, in Entfaltung begriffene Bliittchen und alte, 4iberwinterte Bliitter untersueht werden. Blattepiderinen aller drei Objekte zeigen gegen MnSO~ Init zunehinendena Alter eine starke Steigerung der Manganresistenz yon 0,0001% his auf 10% und anniihernde Resistenzkon- stanz bzw. nut eine nail der Entwieklung leieht ansteigende, im Alter abet wieder etwas absinkende Resistenz gegen Borsiiure. Die Resistenz gegen

Xnderung d. dram. Resistenz pflanzl. Plasmen mit dem Entwieklungszustand 133
ZnSO4 nimmt bet Anemone hepatica und Cyclamen europaeum nur etwas, bei Hedera helix hingegen gleich stark wie gegen MnSO4 zu.
4. Von den untersuchten Kulturpflanzen erwiesen sich die Cruciferen (Rettieh, Weif~e Riibe und Kohlrabi) besonders gegen MnSQ und ZnSO,~ als Stark resistenzverinderlich, Daucus carota und Solanum tuberosum zeig- ten geringe Resistenzgnderungen, und die Tomate, Solanum lycopersicum, war in ihrer Hauptentwicklungszeit yon Juni bis Oktober gegen B, Zn und Mn nahezu resistenzkonstant.
5. In den Monaten Juni bis Oktober zeigten die Epidermen verschieden alter Sprosse yon sieben Wildpflanzen (SalDia pratensis, Saloia glutinosa, Saloia r, erticillata, Ballota nigra, Saponaria o[ficinalis, ConDallaria majalis und Vitis Dulpina) gleichfalls altersbedingte Resistenzverschiebungen, wo- bet wiederum die Manganresistenz die grSlten Unterschiede aufwies.
6. Die Resistenz verschiedener Epidermen (Blattober- und -unterseite, Blattstiel, Stengel) von Cyclamen europaeum und Cyclamen persicum zeigte keine gro[~.en Unterschiede. Lediglich die Blattunterseite yon Cycla- men persicum erwies sich gegeniiber der Blattoberseite gegen Bors~iure und gegen Atuminiumchlorid als etwas empfindlicher.
7. Im wesentlichen sind die Resistenz~inderungen "yore Alterszustand der Zellen abhingig, zum Tell scheinen aber auch jahreszeitliche Aufienein- fliisse yon Bedeutung zu seth.
8. Die Frage, ob das Plasmalemma oder das gesamte Zytoplasma fiir die Hiihe der Resistenz der Zelle malgebend ist, wird diskutiert. Es scheint so zu sein, dal~ im allgemeinen die Empfindlichkeit des Zytoplasmas die Hiihe der Resistenz bestimmt, dal~ abet in F~illen besonders hoher Resistenz Ver- ~inderungen im Plasmalemma (Abdichtung und damit geringere lntrabilitit) das Absterben der Zellen verhindern kiJnnen.
9. Nur wenige Plasmen erweisen sich w~ihrend des ganzen Jahres gegen einen oder gegen einige der angewandten Teststofte vtillig resistenzkonstant. Die Tatsache, daft aber die Resistenzverinderungen im Laufe der Ent- wicklung nicht alle Teststoffe in gleicher Weise treften, liiftt annehmen, dal~ die ihnen zugrunde liegenden plasmatischen Verinderungen fiir die ver- sehiedenen chemischen Einwirkungen nicht yon gleicher Bedeutung sind. Worin diese plasmatischen Ver~inderun,gen bestehen, steht noch vtillig often.
L i t e r a t u r
B i b l i o g r a p h y of the literature of the Minor Elements and their relation to plant and animal nutrition. 4. ed., Vol. I 1948, Yol. II 1951, u III 1953, Chil. Nitr. Edue. Bur. Inc., New York.
Biebl, R., 1939: Protoplasmatisehe Okologie der Meeresalgen. Ber. dtsdl. Bot. Ges. 57 (78).
- - 1947 a: Die Resistenz gegen Zink, Bor und Mangan als Mittel znr Kennzeiehnung versehiedener pflanzlieher Plasmasorten. Sitzber. Akad. Wiss. Wien, math.-nat. KI., Abt. I, 155, 145.
- - 1947 b: t3ber die gegens~itzliche Wirkung der Spurenelemente Zink und Bor auf die Blattzellen VOl~ Mnium rostratum. Ost. Bot. Z. 94, 61.

134 R. Biebl und Wald t rau t Rossi-Pillhoier
1949: Vergleiehende chemis&e Resistenzstudien an pflanzlieheu Plasmen. Proto- plasma 39, 1.
- - 1 9 5 0 a : (Yber die Resistenz pflanzli&er Plasmen gegen Vanadium. Proto- plasma 39, 251. /950 b: Zellphysiologische Untersuchungen an Gemiisepflanzen. Biol. gen. 19, 236. 1950e: Zellphysiologisdm Beobachtungen an panasdl ier ten Abutilon-Pflanzen. ()st. Bot. Z. 97, 168.
- - 1952 a: tlesisfenz der Meeresalgen gegen siehtbares Lieht und gegen kurzwell ige UV-Strahlen. Protoplasma 41, 353. 1952 b: Ecological and non-environmental consti tutional resistance of the proto- plasm of marine algae. Journ. Mar. Biol. Ass. Un. King& 3L 307.
- 1953: Resistenz pflanzticher Plasmen gegen 2,4-D. Protoplasms 42, 193. B l a c k m a n n , G. E., 1952: Studies in the Principles of Phytotoxie i ty I., Journ. Exp.
Bot. 3, 1. F i s c h e r , H., 1950: l~ber protoplasmatisdle Veriinderungen beim Altern yon
pflanzlichen Zellen. Sammelreferat. Protoplasma 39, 661. F r o h b e r g e r , E., 1951: Zur Wirkung der 2 4-Didflorphenoxvessigs~ure auf Ge-
webe und Stoffwechsel der Pflanze. Htifehen-Briefe, VeriSff. d. , ,Bayer"-Pflan- zensehutzabt. Leverkusen 4, 236.
H t i f l e r , K., 1952: Vergleichende Protoplasmatik. Bet. dtsdl. Bot. Ges. 50, (53). - - 1934: Kappenplasmolyse und Salzpermeabil i tat . Z. f. wiss. Mikr. 31, 70. - - 1936: Permeabiliti i tsuntersehiede in versehiedenen Geweben ether Pflanze and
ihre vermutlichen Ursachen. Mikroehemie (Moliseh-Festschrift) 224. 1937: Spezifis&e Permeabilit~itsreihen verschiedener Zelisorten derselben Pflanze. Ber. dtseh. Bot. Ges. 55 (155).
- 1939: Nekroseformen pflanzlieher Zellen. Ber. dts&. Bot. Ges. 56, 45~. - - 1951: Plasmolyse mit Natr iumkarbonat . Protoplasms 40, 426. - - u n d H. S t i e g t e r , 1930: Permeabilit~itsverteilung in versdliedenen Geweben
der Pflanze. Protoplasma 9, 469. H o f m e i s t e r , L., 1958: Versehiedene Permeationsreihen bet einer und derselben
Zellsorte yon Ranunculus repefzs. Jb. wiss. Bot. 86, 401. H u t c h , H., 1955: Beitr~ige zur Kenntnis der Permeabili t / i tsverteilung in den ~er-
sehiedenen Geweben des Blattes. Beth. bot. Zentrbl. 50. I l j i n , W. S., 1935: Das Absterben der Pflanzenzellen in reinen und balaneicrten
SalzliSsungen. Protoplasms 24, 409. K a h o , It., 1953: Das Verhalten der Pflanzenzellen gegen Sdtwermetallsalze.
Plants 18, 664~ K a m i y a, N., 1939: Zytomorphologische Plasmolysestudien an Allium-Epidermen.
Protoplasma .32, 373. K e s s l e r , W., 1935: Crber die inneren Ursa&en der K/ilteresistenz der [)flanzc.
Planta 24, 51% und W. R u h l a n d , 1958: Weitere Untersuchungen tiber die inneren Ursadlen der K~lteresistenz. Plants 28, 159.
K r a s s i n s k y , N., ~929: f3ber jahreszeitliche Ver~inderung'el~ der Pemaeabilit/it de:s Protoplasmas. Protoplasma 9, 622.
L e n k , J., 1953: Uber die Plasmapermeabil i t / i t ether Spirogyra in verschiedenen Entwieklungsstadien nnd zu verschiedenen Jahreszeiten. Sitzber. d. ()st. Akad. Wiss., math.-nat. K1., Abt. 1, 162, 235.
l ~ i d f o r s s , B., 1907: Die wintergriine Flora. Eine biolog'ische Untersudmng'. L u , d s Univ. Arsskr., N. F., 2 II. Nr. 13,

Xnderung d. chem. Resistenz pflanzl. Plasmen mit dem Entwieklungszustand 135
L i n s e r, H., 1951: Unkrautbek~impfung auf hormonaler Basis. Die Bodenkultur 5, 19I.
M a r k l u n d, G., 1936: Vergleiehende Permeabili t / i tsstndien an ptlanzlichen Proto- plasten. Aeta bot. Fenn. 18, 1.
P a e c h , K., 1940: Ver~nderungen des Plasmas w/ihrend des Alterns pflanzlicher Zellen. Planta 31, 293.
P is e k, A., 1950: Frosth~irte und Zusammensetzung des Zellsaftes bei Rhododendron ferrugineum, Pinus cerebra und Picea excelsa. Protoplasma .39, 129.
P r i n g s h e i m , E., 1924: t3ber Plasmolyse dutch Schwermetallsalze. Beih. hot. Zentrbl. 4I, I. Abt., 4.
R e u t e r , L., 1941: l~ber die Salzresistenz der Epidermiszellen des Blattes yon Pisum satioum. Protoplasma 35, 530.
- 1949: Protoplasmatische Pflanzenanatomie. Sammelref. Phyton 1, 229. R o s s i , W., 195t: Ober die chemisehe Resistenz ptlanzlicber Plasmen und ihre Yer-
'anderung mit dem Entwieklungszustand. Diss. Univ. Wien (unveriiff.). S c h a r r e r, K., t944: Bio&emie der Spurenelemente. Berlin, Parey. S e h i n d l e r , H., 1949: Protoplasmatod dureh Sehwermetaltsalze I. Protoplasma 38,
225. S t e i n e r , M., 1939: Die Zusamlnensetzung des Zellsaftes bei htiheren Pflanzen in
ihrer i~kologischen Bedeutung. Erg. Biol. 17, 151. Ur l , W., 1951: Permeabilit~itsverteilung in den Zellen des Stengels yon Taraxaeum
officinale und anderer kraut iger Pflanzen. Protoplasma 40, 475. - - 1952: Untersehiede der Plasmapermeabilit~it in den Gewebesehidlien krant iger
Stengel. Physiologia P lan tarum 5, 155. U r s p r u n g , A., und B. B l u m , t911: 1. Verteilung des osmotischen Wertes in der
Pflanze, 2. Einflul~ der Aui3enbedingungen auf den osmotisehen Wert, 5. Uber die periodisehen Sehwankungen des osmotisehen Wertes. Bet. dtseh. Bot. Ges. 34. 88.
W e b er, Ft. , 1929: Protoplasmatisehe Pflanzenanatomie. Protoplasma 8, 291. - - und H. H o h e n e g ge r, 1923: Reversible Viskosit/itserhShung des Protoplasmas
bei K/ilte. Bet. dts&. Bot. Ges. 41, 198.