differentpatternsoflectinbindingandcellsurfacesialylation...
TRANSCRIPT

(CANCER RESEARCH 43, 5138-5144, November 1983]
Different Patterns of Lectin Binding and Cell Surface SialylationDetected on Related High- and Low-Metastatic Tumor Lines
Peter Altevogt,1 Mina Fogel, Rachanee Cheingsong-Popov, Jim Dennis, Peter Robinson, andVolker Schirrmacher2
Institut fur Immunologie und Genetik, Deutsches Krebsforschungszentrum, 6900 Heidelberg, Federal Republic of Germany [P. A., M. F., R. C-P. P. R., V. S.¡,andThe Hospital for Sick Children, Toronto, Ontario, Canada [J. D.J
ABSTRACT
We have analyzed cell surface-bound carbohydrates in two
different model systems for metastasis composed of closelyrelated tumor cell lines with differing metastatic potential. Thefirst system studied was that of the DBA/2-derived T-lymphoma
lines (Eb/ESb) and some recently established sublines of ESbwith altered metastatic behavior (ESb-M and ESb-MR). Thesecond system consisted of the highly metastatic MDAY-D2cells, a wheat germ agglutinin-resistant low metastatic subline
MDW40, and two metastatic revertants from the latter. The cellswere stained with fluorescein isothiocyanate-conjugated lectinsand analyzed by flow cytofluorography. All low-metastatic tumor
lines expressed receptor sites for the lectins soybean agglutinin(SBA) and Vicia villosa (VV). The metastatic lines had the respective lectin binding sites blocked by sialic acid (SA). A goodcorrelation was found within the cell lineages Eb —»ESb —»ESb-M —ESb-MR and MDAY-D2 -»MDW40 -»MDW40M1 between
reactivity of SBA and VV and metastatic potential.The amount of neuraminidase-accessible SA was similar for
all cell types (except MDW40) indicating differences in the positioning of SA. For high-metastatic ESb cells, the Sialylation of
SBA and VV receptor sites was paralleled by a relative decreaseof SA associated with receptor sites for peanut agglutinin. Low-
metastatic Eb cells, in contrast, had their peanut agglutininreceptor sites sialylated but expressed asialylated SBA and VVreceptor sites. Eb cells were also found to have 2-fold higheractivities in galactose-specific sialyltransferases. It is proposed
that the differences in positioning of SA on the cell surfaceleading to masking or unmasking of terminal sugars could influence the metastatic potential of tumor cells.
INTRODUCTION
Alterations in cell surface carbohydrates have been found toseverely affect the metastatic potential of experimental tumors.For example, tumor cell variants selected in the presence oftoxic concentrations of certain plant lectins appeared to havesignificantly changed metastatic behavior (11, 28, 34). Suchselection procedures can induce changes in specific surfacecarbohydrates (11, 12), glycosyltransferase activities (12, 15,23), and glycoprotein content (22). One could theoretically envisage at least 3 steps in the metastatic cascade that involve cellsurface carbohydrate interactions: (a) in the release of tumorcells from the primary tumor mass due to altered homotypicadhesion phenomena; (b) in the mechanism of blood transpor-
1To whom requests for reprints should be addressed. Recipient of a grant from
Meyenburg-Stiftung.* Supported by the Deutsche Forschungsgemeinschaft through Sonderfor-
schungsbereich 136.Received September 2, 1982; accepted August 8, 1983.
tation of metastatic tumor cells by heterotypic cell interactions;and (c) in the arrest in organs by specific interactions with thetarget tissue.
Such important involvements of carbohydrates in metastasisformation would imply that tumor cells with different metastaticpotential possess distinct qualitative or quantitative differencesin their glycosylation of membrane glycoconjugates. A correlationbetween the quantity of cell surface-exposed SA3 and metastatic
capacity was recently found by Yogeeswaran (36) for a varietyof different tumor cell lines. Of particular importance seemed tobe SA residues in association with subterminal sugar moietieswhich could be oxidized by galactose oxidase and subsequentlyreduced with NaBH4 (36).
We have further investigated this topic with cell lines of 2independent tumor model systems for metastasis. Differences inglycosylation and the role of SA were studied by using a varietyof FITC-conjugated lectins with different sugar specificities and
flow cytofluorographic analysis. The first tumor system was theDBA/2 T-lymphoma Eb and its spontaneous high-metastatic
variant ESb (30). It was found that acceptor sites of SBA andVV were covered by SA on all high-metastatic ESb sublines butnot on parental type Eb cells or on low-metastatic variants from
ESb cells. Similar differences were noted in a second tumorsystem (9, 11, 18, 19). From these results, we suggest that theSialylation of particular carbohydrate residues on tumor cellswhich are recognized by certain lectins may facilitate their metastatic spread possibly by masking specific cellular adhesionsites.
MATERIALS AND METHODS
Cells. The tumor cell Eb (Heidelberg subline of L5178Y) and itsmetastatic variant ESb were obtained from Professor P. Alexander(London, England). The etiology of the cell lines has been described (30).Both cell lines were passaged as ascites in syngeneic DBA/2 mice(Bomholtgaard, Denmark) or grown in suspension in Roswell Park Memorial Institute tissue culture Medium 1640 supplemented with penicillin(100 units/ml), streptomycin (100 /jg/ml), glutamine (5 HIM), 4-(2-hydrox-yethyl)-1-piperazineethanesulfonic acid (50 mM), 2-mercaptoethanol (5 x10~5 M), and 5 or 10% fetal bovine serum. ESb-M is a plastic-adherent
variant of ESb with highly decreased metastatic capacity (14). MDAY-
D2 is a metastatic tumor cell line of DBA/2 and was established byKerbel ef al. (18,19). A WGAR cell line MDW40, with altered tumorigenic-
ity and metastatic capacity (9), and 2 metastatic and wheat germagglutinin-sensitive revertants thereof (MDW40M1 and MDW40M4) have
been described previously (11 ).Staining of Tumor Cells with FITC-conjugated Lectins and Cyto
fluorographic Analysis. FITC-conjugated lectins (1 mg/ml) were obtained from Medac (Hamburg, Federal Republic of Germany). Fluoro-
3The abbreviations used are: SA, sialic acid; FITC, fluorescein isothiocyanate;SBA, soybean agglutinin; VV, Vicia vil/osa: WGA", Wheat germ agglutinin-resistant;HP, Helix oomatia, GalNAc, N-acetylgalactosamine; PNA, peanut agglutinin; o-Gal.D-galactose; HBP, hepatocyte-binding protein; GlcNAc, N-acetylglucosamine.
5138 CANCER RESEARCH VOL. 43
on May 8, 2018. © 1983 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

chrome/protein ratios (495/280 nm) were between 1.3 and 1.8. Cells(106) were incubated with 50 n\ of 1/20 diluted FITC-lectins in the absence
or presence of 0.2 M competitive sugars on ice for 45 min. The reactionwas carried out in 0.9% NaCI solution supplemented with 1 HIM calciumand magnesium salts. Cells were washed and analyzed by flow cytofluo-rography using an ORTHO 50-H cell sorter. The argon laser was operatedat 488 nm, and 3 x 104 cells were analyzed using settings to exclude
nonviable cells according to scatter characteristics. Gains were kept atidentical settings for each FITC-lectin analyzed.
Neuraminidase Treatment of Cells. Cells were incubated in Dulbec-co's phosphate-buffered saline at 20 to 40 x 106/ml for 45 min at 37°in
the presence of 2.5 units of neuraminidase of Vibrio cholerae (Behring-
werke, Marburg, Federal Republic of Germany) and were then washed3 times and analyzed for lectin binding as described above.
SA Determination. Fifteen to 20 x 106 cells/ml were treated with 2units of neuraminidase for 45 to 60 min at 37°.Neuraminidase-releasable
SA of the supernatants was determined by the thiobarbiturate methodof Warren (35).
Determination of Glycosyltransferase Activities. Cell homogenatesfrom Eb and ESb were prepared according to the method of Chatterjeeef al. (8). One g of packed cells was resuspended in 3 ml of 0.25 Msucrose and 0.1 mw CaCI2 adjusted to pH 8.0 with 1 M Tris buffer andhomogenized with a glass homogenizer. The homogenate was spun at1500 rpm for 10 min. The supernatant was collected and stored at -70°.
Asialofetuin and asialoagalactofetuin were prepared from fetuin (GradeIII; Sigma Chemical Co., Munich, Federal Republic of Germany) accordingto standard procedures (32, 33). Ovalbumin (crystallized 3 times) waspurchased from Miles Laboratories, and RNase A was from Sigma.Assays for sialyltransferase (5), galactosyltransferase (6), fucosyltrans-ferase (7), and N-acetylglucosaminyltransferase (5) were performed essentially as described in the literature. [14C]CMP SA, [14C]UDP galactose,[I4C]GDP-fucose, and [14C]UDP-W-acetyl-D-glucosamine were obtained
from NEN (Dreieich, Federal Republic of Germany).
RESULTS
Staining of Eb and ESb Tumor Cells with FITC-conjugatedLectins. FITC-conjugated lectins with different sugar specificities
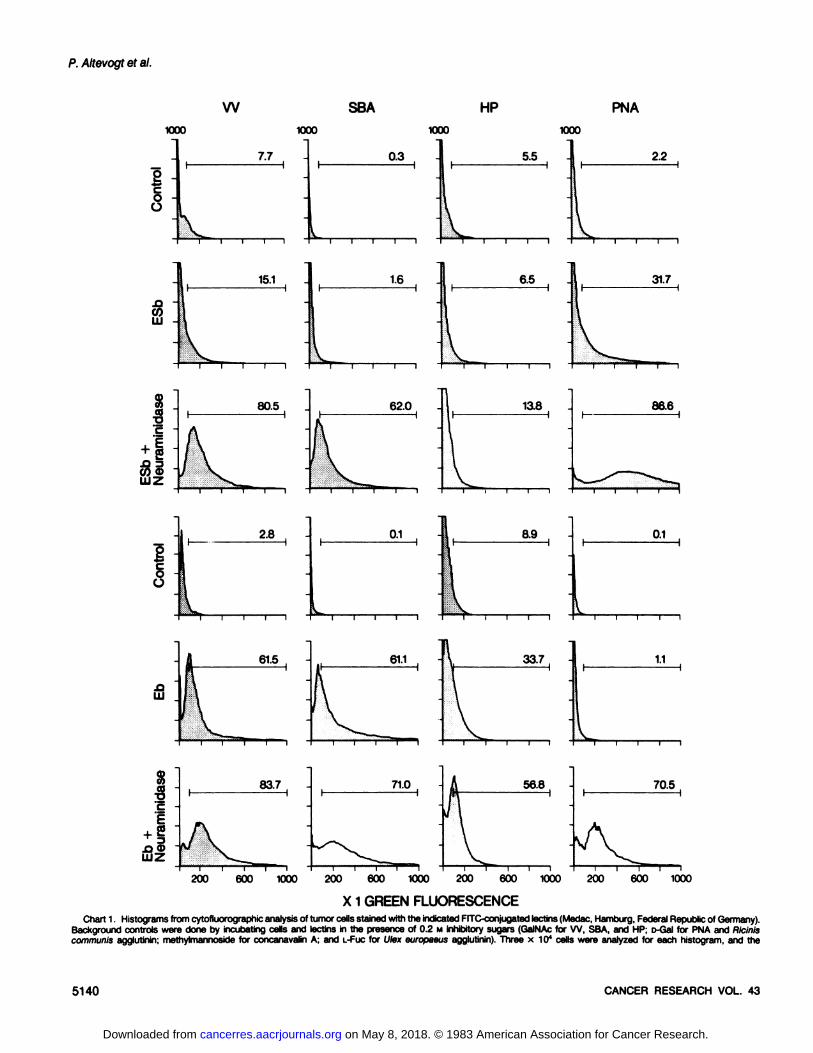
(Table 1) were used to study the expression of correspondingbinding sites on the standard Eb and ESb tumor cells. Althoughcloned sublines of both tumor cell populations were available,we intentionally studied here the noncloned populations to getan impression of the overall differences. As negative controls,we used lectin binding values in the presence of 0.2 M inhibitorysugars. The results obtained for ESb and Eb cells with or withoutneuraminidase treatment are shown in Chart 1. The lectins VV,SBA, and HP stained the metastatic ESb tumor cells only weakly(close to background controls), while the parental line Eb wasable to bind these lectins (Chart 1). The failure of these lectinsto bind to ESb cells was apparently due to a masking of receptorsites by SA, since treatment of the cells with neuraminidaseuncovered these sites and led to staining of ESb cells to a similardegree as that for Eb cells (Chart 1). There was, however,virtually no staining observed with HP, even when neuraminidasetreatment was performed. This is in agreement with the previousobservation that T130, the major HP binding glycoprotein (2), is
Cell Surface Glycosylation and Metastatic Potential
absent on ESb tumor cells but present on Eb cells (1).Staining of Eb cells with SBA, VV, and HP was only slightly
increased after neuraminidase treatment, indicating that thesereceptor sites were mostly unsialylated and accessible for thelectins (Chart 1).
ESb cells showed weak reactivity with PNA, while Eb cellswere negative as judged by the histograms in Chart 1. Afterneuraminidase treatment, both cell lines were stained by PNA,but the ESb cells still showed stronger reactivity as seen by thepercentage of stained cells and by the fluorescence intensity. Incontrast, Ricinus communis agglutinin was able to bind to bothEb and ESb cells, possibly via unblocked terminal o-galactose
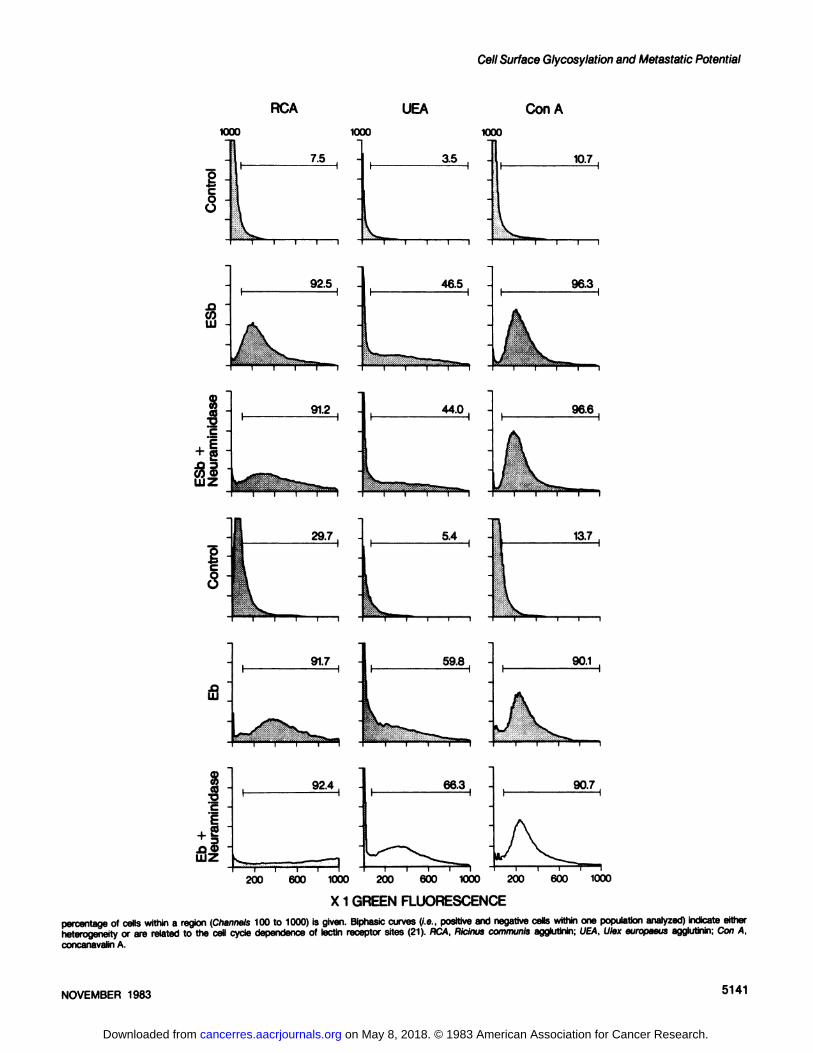
residues. This binding was enhanced after treatment with neuraminidase and most strongly for Eb cells, indicating a higherdegree of o-galactose sialylation (Chart 1S). No major differencesin lectin binding properties of Eb and ESb cells were noted for u-fucose- or o-mannose-detecting lectins [see results in Chart 16with Ulex europaeus (L-fucose) or concanavalin A (o-mannose)].
The results taken together indicate obvious differences between the low- and high-metastatic cells, Eb and ESb, in the
expression of receptor sites for lectins with specificity for GalNAcresidues (VV, SBA, and HP), which are expressed on Eb but notESb cells. On the other hand, receptor sites for PNA, which hasspecificity for D-Gal-1-3-D-GalNAc configurations, were better
expressed on ESb than on Eb cells. Removal of SA by neuraminidase treatment led to the uncovering of binding sites forSBA and VV by ESb cells and of terminal o-Gal-1-3-o-GalNAc
groups on Eb cells detectable by the respective specific lectins.We have recently isolated a plastic-adherent variant line from
the high-metastatic ESb cell (ESb-M) which was found to have
lost its metastatic potential while still being tumorigenic in normalsyngeneic hosts (14). The variant retained most of its ESb-
derived antigenic and biochemical characteristics, but showedincreased expression of binding sites for the lectins VV and SBA,very similar to the characteristics of the low-metastatic parental
line Eb. While such sites were masked by SA on metastatic ESbcells, they became unmasked on ESb-M cells. Furthermore,metastatic revertants (ESb-MR) from ESb-M cells isolated fromthe spleen (Esb-MSP) or the brain (ESb-MBR) of tumor-bearing
mice were again found to have the respective lectin binding sitesmasked by SA. There was, thus, within the lineage Eb —»ESb—»ESb-M —»ESb-MR, a correlation between the phenomenon
described here, i.e., exposure or masking of SBA receptor sitesand overall metastatic capacity (Table 2).
Staining of MDAY-D2 Tumor Cells and Its Wheat Germ
Agglutinin Selected Mutants with VV and SBA. In order toevaluate whether the relation between exposure or blocking ofSBA receptor sites and metastatic capacity was a peculiarity ofthe Eb/ESb tumor system or was perhaps of more generalsignificance, we investigated another tumor system consistingof high- and low-metastatic related tumor lines. The parentalhigh-metastatic tumor line MDAY-D2, the nonmetastatic WGAR
mutant, and 2 metastatic revenant lines were compared for their
Table 1
Sugar specificities of lectins used in this study
HPWGlycine max (SBA)Ulex europeusRicinus communisArachis hypogea (PNA)Conavalia enziformis (concanavalin A)
D-GalNAc-a-1 -»D-GalNAc-«-1—3-D-Gal-/3-1-»D-GalNAc-a-1 -»3-o-Gal-/3-1 -> 3-D-GlcNAcL-Fuc-a-1 —2-D-Gal-tf-1 —4-D-GlcNAc-/3-1 -D-Gal-/3-l-»D-Gal-/3-1 -> 3-D-GalNAcD-Man-a-1 -»
NOVEMBER 1983 5139
on May 8, 2018. © 1983 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
P. Altevogt et al.
W SBA HP
XX» XX» 1000 1000
l8 J
•4-*
o -ü
7.7 0.3 5.5
PNA
2.2
Sì -coLU -
15.1 16 6.5 31.7
8to•o'
&Å“LUZ
80.5 62'° 13.8
2.8 0.1 8.9 0.1
61.5 61.1 33.7 1.1
a'c -
S
83.7 71.0 56.8 70.5
200 600 1000 200 600 1000 200 600 1000 200 600 1000
X 1 GREEN FLUORESCENCEChart 1. Histograms from cytofluorographic analysis of tumor cells stained with the indicated FITC-conjugated lectins (Medac, Hamburg, Federal Republic of Germany).
Background controls were done by incubating cells and lectins in the presence of 0.2 M inhibitory sugars (GalNAc for VV. SBA, and HP; D-Gal for PNA and Riciniscommunis agglutinin; methylmannoside for concanavalin A; and L-Fuc for Ulex europaeus agglutinin). Three x 104 cells were analyzed for each histogram, and the
5140 CANCER RESEARCH VOL. 43
on May 8, 2018. © 1983 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
on May 8, 2018. © 1983 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

P. Altevogt et al.
Table 2Binding of FITC-conjugated lectins W and SBA to ESb-derived tumor cell lines
Cells were stained and analyzed by flow cytofluorography as described in the legend to Chart 1.
CellsESbESb-M
ESb-MSPESb-MBRControls
in presence of sugarMetastatic
capacityHigh
LowHighHigh%
of cellsbindingVV24.6
74.928.7
38.714.0
(0.2 M GalNAc)SBA30.2
63.433.9
44.29.0
(0.2 M GalNAc)Concanavalin
A*93.189.4
93.787.21
1.0 (0.2 M n-methylmannoside)
" Binding of concanavalin A to internal sugar moieties was more uniform and could be inhibited in the presence of «-methylmannoside.
Table 3Binding of FITC-conjugated lectins VV and SBA to MDAY-D2-derived tumor cell lines
Cells were stained and analyzed by flow cytofluorography as described in the legend to Chart 1.
CellsMDAY-D2
MDW40MDW40M1MDW40M4Metastatic
capacityHigh
LowHighHighIn
thepresence of
sugar1.8
17.76.4
16.2%
of cells binding Wneuraminidase
treatment2.886.5
11.327.6+94.3
94.775.090.5In
the presene«of sugar0.5
5.61.52.1%
of cells bindingSBAneuraminidasetreatment»1.8
80.25.83.1+81.1
86.682.584.7
Table 4Neuraminidase-releasable SA of murine tumor cell lines used in this study
CellsMDAY-D2MDW40MDW40M1MDW40M4EbESbESb-MMetastaticcapacityHighLowHighHighLowHighLownmolSA/mg cellprotein1.91±0.35a(5)"0.59
±0.13(4)1.89 ±0.06(2)1.69
±0.12(3)1.08
+ 0.19(2)1.42±0.10(3)1.45±0.01 (2)
" Mean ±S.D.6 Numbers in parentheses, number of experiments performed.
binding properties to the lectins VV and SBA. The results,presented as the percentage of positive cells, are shown in Table3. The parental metastatic line MDAY-D2 and the wheat germagglutinin-sensitive revenants, MDW40M1 and MDW40M4,
could not be stained significantly above the background controlswith either of the 2 lectins. They were, however, readily stainedif the cells were pretreated with neuraminidase. In contrast, thenonmetastatic WGAR MDW40 cells stained brightly without neur
aminidase treatment.Determination of SA Content of the Tumor Cell Lines. The
results presented indicated similar differences in VV and SBAlectin staining between high- and low-metastatic tumor lines of
2 different tumor systems. Since the differences found could beaffected by neuraminidase pretreatment, we determined the totalamount of neuraminidase-releasable SA for all of the lines studied. As shown in Table 4, cells from the MDAY-D2 lineageshowed a 3-fold reduction of SA for the nonmetastatic MDW40
line, while the revenants showed values similar to those of theparental line. In the case of the Eb, ESb, and ESb-M cells, thevalues for neuraminidase-accessible SA differed only slightly. At
least in the Eb/ESb system, there was no correlation betweenmetastatic potential and net amount of neuraminidase-releasable
SA.Determination of Glycosyltransferase Activities of Eb and
ESb Tumor Cells. Altered levels of glycosyltransferase activitiesof ESb tumor cells could possibly account for the noted differences in glycosylation. Therefore, we determined some of these
enzyme activities using standard procedures (5-8) (Table 5).Table 5 shows that A/-acetylglucosaminyltransferase and sialyl-transferase activities were, respectively, 5- or 2-fold higher in Eb
than in ESb tumor cells. Activities of fucosyltransferase A and Band galactosyltransferases were only slightly different.
These data clearly document changes in glycosyltransferaseactivities between Eb and ESb cells.
DISCUSSION
Changes in the metastatic behavior of defined tumor cell lineshave been observed to be associated with changes in cell surfaceglycosylation (9,13). An important role of surface carbohydrateswas also supported by the effect of tunicamycin on metastaticarrest properties (16). In this particular context, SA has receivedincreased attention recently (36).
In the present study, we have analyzed cell surface carbohydrates by means of lectin binding in 2 different tumor systemsconsisting of various tumor sublines differing greatly in metastaticpotential. The most consistent differences in lectin binding properties between the pairs of high- and low-metastatic tumor linesinvestigated (i.e., Eb versus ESb; ESb versus ESb-M; ESb-Mversus ESb-MR; MDAY-D2 versus MDW40; and MDW40 versusMDW40M1) were found with the D-GalNAc-specific lectins, VV
and SBA. Both lectins were found to bind directly only to thelow-metastatic tumor lines, while the high-metastatic lines re
quired neuraminidase pretreatment in order to expose the respective binding sites.
In a recent study, Kaladas ef al. (17) have investigated the finespecificity of the interaction of VV lectin to carbohydrates.The purified lectin agglutinated type A human erythrocytes preferentially. Inhibition assays with various monosaccharides, gly-cosides, and oligosaccharides indicated that the VV lectin isspecific for terminal, nonreducing «-linked D-GalNAc (17) residues. Among the disaccharides tested, o-GalNAc-«-1-3-D-Galwas most active in inhibition, and the importance of the «-1-3-
linkage was indicated (17). In contrast to VV, SBA is known toreact strongly with o-galactose as well as with o-GalNAc (25)
and has no anomeric specificity (17). The question of which
5142 CANCER RESEARCH VOL. 43
on May 8, 2018. © 1983 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

Cell Surface Glycosylation and Metastatic Potential
Table 5
Glycosyltransferase activities of the Eb and ESb tumor cells
Enzyme activities (cpm/mg protein)
Analyzed enzymeactivitiesSialyltransferaseGalactosyltransferaseFucosyltransterase
AFucosyltransferase
B/v-Acetylglucosaminyltransfer-ase-1N-Acetylglucosaminyltransfer-ase-2SubstrateCMP
(["Cjsialicacid)CMP(["CJsialicacid)UDP
(Ì14C]galactose)UDP(['4C)galactose)GDP([14C]fucose)GDP(r14C]fucose)GDP(['4C)fucose)GDP(|'4C]fucose)UDP([14C)NAcGlc)UDP
(['4C]NAcGlc)UDP([14C]NAcGlc)UDP
(["CJNAcGIc)AcceptorAsialofetuinOAsialoagalactofetuin—Asialofetuin—Asialoagalactofetuin—Ovalbumin—RNase
A—Eb61
,400 ±618a433
±157,753±85650
±1825,220±2111000
±2117,950±30450
±1020,925±159320
±405,000±18375
±38ESb32,957
±87466±2810,992
±178775±2020,260±1201
,400 ±2513,316±82550
±124,175±39.1,125
±32550
±13ESb/Eb0.541.410.800.740.200.23a Mean ±S.D.6 —,assays performed in absence of exogenous acceptor;
acceptor.the values obtained include background plus possible contribution of endogenous
tumor cell surface structures were involved in the binding of VVand SBA has to be discussed in light of these specificity data.Terminal GalNAc residues, which could act as potential bindingsites for SBA and VV, have not been observed to be sialylatedin any glycolipid or W-glycosidically linked protein carbohydrateside chain. 0-Glycosidically linked mucin-type carbohydrate side
chains have been observed to be sialylated on galactose residuesor on the innermost GalNAc moiety. Biochemical studies arepresently being performed on the tumor cell surface carbohydrates with serve as receptor sites for VV and SBA, on theirlinkage type, and on the nature of the respective glycoconju-
gate(s).Interestingly, it was shown by Kimura and Wigzell (20) that
cytotoxic mouse T-lymphocytes display a cell surface glycopro-
tein, T145, to which VV has an exquisite specificity. A similarglycoprotein could be detected after neuraminidase treatmentand selective labeling by the galactose oxidase/NaBH4 techniqueon some cloned ESb tumor cells (1). The T145 glycoproteinamong putative others might be a candidate for the interactionof VV with neuraminidase-treated ESb tumor cells.
In the 2 tumor systems studied, the binding of VV and SBAcorrelated with the metastatic capacity of the tumor sublines. Inthe B16 melanoma system, however, which, in contrast toMDAY-D2 and Eb/ESb cells, is plastic-adherent, binding of iodi-
nated SBA did not correlate with the metastatic capacity of thecells (27). Therefore, it will be interesting to extend lectin bindingstudies to other tumor cells with differences in metastatic potential and adhesion properties in order to evaluate a possiblegeneral significance of our findings.
Little is known at present about the possible physiological roleof the carbohydrate side chains which serve as specific lectinreceptor sites for VV or SBA. It is conceivable that such sitesmight mediate cellular adhesion to certain substrates, such asfibronectin, laminin, or collagen. In support of this assumption,we have recently found that the low-metastatic variant MDW40,
which showed a much better expression of receptor sites for VVand SBA than did the metastatic parental line MDAY-D2 (Table
3), also showed a much better attachment to basement membrane collagen type IV and fibronectin than to laminin. Furthermore, neuraminidase treatment of the metastatic MDAY-D2 tu
mor cells selectively enhanced their attachment to collagen IVand fibronectin (10).
In the Eb/ESb/ESb-M tumor system, the total amount of
neuraminidase-accessible cell surface SA was quite similar.
Therefore, the difference in sialylation of the receptor sites forVV and SBA in this tumor system was most likely due todifferences in the exact positioning of SA on the various glyco-conjugates of the cell surface. In the MDAY-D2 tumor system,the low-metastatic lectin-resistant variant MDW40 had onlyabout one-third of the amount of neuraminidase-accessible SA
of the corresponding metastatic lines (i.e., the parental line and2 metastatic revertants). In this system, therefore, increase inmetastatic capacity was associated both with an increase in thetotal amount of cell surface sialylation and in the site-specific
sialylation of SBA and VV receptor sites. From these and otherfindings (36), it seems that the receptor sites for SBA or VV ontumor cells can have a negative impact on their metastaticcapacity.
How could the differences between Eb and ESb cells inexpression of specific lectin binding sites be explained? Relevantto this context may be our recent observations on hepatocyte-
tumor cell interactions in vitro, where we found that only ESbtumor cells and not Eb cells were able to form spontaneousrosettes with the hepatocytes (29). Eb cells required treatmentwith neuraminidase in order to exhibit similar rosette formation(29). A detailed molecular analysis of this interaction allowed theidentification of a lectin-like protein on hepatocytes (HBP), whichseemed to recognize terminal Gal-GalNac residues and to besimilar to PNA in specificity." As can be seen from Chart M3, this
lectin, in contrast to VV or SBA, reacted more strongly with ESbthan with Eb cells. Thus, it appears as if the transition from thelow-metastatic Eb cells to their high-metastatic variant was
associated with a change in the sialylation of specific bindingsites. While Eb cells expressed binding sites for VV and SBAand had the binding sites for HBP and PNA masked by sialylation,ESb cells had the binding sites for VV and SBA sialylated andinstead expressed binding sites for HBP and PNA. A differencein the positioning of terminal SA on high- and low-metastatic
tumor cells could thus have a dual effect: a blocking of certainbinding sites and an unblocking of others. This could have apronounced effect on the biological behavior of the respectivecancer cells (10,14, 29) and could lead to altered charge distribution patterns of metastatic versus nonmetastatic cells as was
4 R. Cheingsong-Popov, P. Robinson, P. Altevogt, and V. Schirrmacher. A mousehepatocyte carbohydrate-specific receptor and its interaction with metastasizingtumor cells. Int. J. Cancer, in press, 1983.
NOVEMBER 1983 5143
on May 8, 2018. © 1983 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

P. Altevogt et al.
shown ¡nthe B16 melanoma system (26).A further important question is the possible role of glycosi-
dases and glycosyltransferases in the modulation of expressionof carbohydrate binding sites. A proper interpretation of theresults on the glycosyltransferase activities of Eb and ESb cells(Table 4) depends on the carbohydrate side chain structure ofthe acceptor proteins used in these assays. This seems ofparticular importance, because it appears that there exist glycosyltransferases for the same sugar with different substratefine specificity (3, 4, 24). Fetuin, asialofetuin, and asialogalacto-
fetuin were used as acceptor proteins for the determination ofsialyl-, galactosyl-, and fucosyltransferase activities. It is known
from earlier studies by Spiro (32, 33) on the structure of thecarbohydrate units of fetuin that the O-glycosidically linked oneshave a terminal sequence of SA-o-Gal-GalNAc-(SA)-Ser(Thr),while the A/-glycosidically linked ones have a terminal sequenceof SA-D-Gal-GlcNAc4-Man3-GlcNAc-GlcNAc-Asn. Accordingly,asialofetuin expresses numerous terminal galactosyl-residues
and thus can be assumed not to test for all sialyltransferasesbut perhaps predominantly for galactosyl-specific ones. The
finding of a higher sialyltransferase activity in Eb cells (possiblyspecific for galactosyl residues) could thus account for the higherdegree of sialylation of terminal ß-galactose structures as rec
ognized by HBP or PNA. This is also in agreement with theobservation that Ricinus commuais agglutinin receptor sites onEb cells are more sensitive to neuraminidase treatment than arethose on ESb cells (Chart 10). Similarly, the higher activity ofgalactosyltransferase in ESb-type cells could contribute to a
better expression of HBP or PNA binding sites on these cells.The strongly reduced activities of NAc-glucosaminyltransferases
in ESb cells could perhaps indicate a less complete and complexbiosynthesis of carbohydrate side chains as compared to Eb-
type cells, but further experimentation is required before such aconclusion might be drawn.
Based on the results presented here and from previous workon glycosylation differences between Eb and ESb cells (1, 31 ),we propose: (a) that cell surface carbohydrate changes takeplace during tumor progression; (o) that these changes are notrandom but show certain patterns detectable with lectins; and(c) that glycosyltransferases and, in particular, sialyltransferasesplay an important role in these changes, leading to the maskingof some binding sites and the exposure of others.
REFERENCES
1. Altevogt, P., Kumick, J. T., Kimura, A. K., Bosstet, «.,and Schirrmacher, V.Different expression of Lyt differentiation antigens and cell surface glycopro-teins by a murine T lymphoma line and its highly metastatic variant. EUT.J.Immunol., 12: 300-307, 1982.
2. Axelsson, B., Kimura, A., Hammarstrom, S., Wigzell, H., Nilsson, K., andMellstedt, H. Helix pomatia A hemagglutinin: selectivity of binding to lymphocyte surface glycoproteins on T cells and certain B cells. Eur. J. Immunol., 8:757-764,1978.
3. Bartholomew. B. A., Jourdian, G. W., and Roseman. S. The sialic acids: XV.Transfer of sialic acid to glycoproteins by a sialyltransferase from colostrum.J. Biol. Chem., 248: 5751-5762,1973.
4. Carlson, D. M., Jourdian, G. W., and Roseman, S. The sialic acids: XIV.Synthesis of sialyl-lactose by a sialyltransferase from rat mammary gland. J.Biol. Chem., 248: 5742-5750,1973.
5. Chatterjee, S. K. Glycosyltransferases in metastasizing and non-metastasizingrat mammary tumors and the release of these enzymes in the host sera. Eur.J. Cancer, 75: 1351-1356, 1979.
6. Chatterjee, S. K., and Kim, U. Galactosyltransferase activity in metastasizingand non-metastasizing rat mammary carcinoma and its possible relationshipwith tumor cell surface antigen shedding. J. Nati. Cancer Inst., 58: 273-280,1977.
7. Chatterjee, S. K., and Kim, U. Fucosyltransferase activity in metastasizing andnon-metastasizing rat mammary carcinomas. J. Nati. Cancer Inst., 67: 151-
162, 1978.8. Chatterjee, S. K., Kim, U., and Bielat, K. Plasma membrane-associated en
zymes of mammary tumors as the biochemical indicators of metastasizingcapacity. Analysis of enriched plasma membrane preparations. Br. J. Cancer,33: 15-22. 1976.
9. Dennis, J., Donaghue, T. P., Florian, H., and Kerbel, R. S. Apparent reversionof stable in vitro genetic markers detected in tumor cells from spontaneousmétastases.Nature (Lond.), 292. 242-245, 1981.
10. Dennis, J., Waller, C., Timpl, R., and Schirrmacher, V. Surface sialic acidreduces attachment of metastatic tumour cells to collagen and fibronectin.Nature (Lond.), 300: 274-276, 1982.
11. Dennis, J. W., Donaghue, T. P., and Kerbel, R. S. Membrane-associatedalterations detected in poorly tumorigenic lectin-resistant variant sublines of ahighly malignant and metastatic murine tumor. J. Nati. Cancer Inst., 66: 129-139, 1981.
12. Finne, J., Burger, M. M., and Prieels, J-P. Enzymatic basis for lectin-resistant
phenotype: increase in a fucosyltransferase in mouse melanoma cells. J. CellBiol., 92: 277-282,1982.
13. Finne, J., Tao, T. W., and Burger, M. M. Carbohydrate changes in glycoproteinsof a poorly metastasizing wheat germ agglutinin-resistant melanoma clone.Cancer Res., 40. 2580-2587, 1980.
14. Fogel, M., Altevogt, P., and Schirrmacher, V. Metastatic potential severelyaltered by changes in tumor cell adhesiveness and cell surface sialylation. J.Exp. Med., 757: 371-376, 1983.
15. Gottlieb, C., Baenzinger, J., and Kornfeld, S. Deficient uridine diphosphate-N-acetylglucosamine: glycoprotein W-acetylglucosaminyltransferase activity in a
clone of Chinese hamster ovary cells with altered surface glycoproteins. J.Biol. Chem., 250: 3303-3309,1974.
16. Irimura, T., Gonzales, R., and Nicolson, G. L. Effect of tunicamycin on B16metastatic melanoma cell surface glycoproteins and blood-borne arrest andsurvival properties. Cancer Res., 47: 3411-3418,1981.
17. Kaladas, P. M., Kabat, E. A., Kimura, A. K., and Ersson, B. The specificity ofthe combining site of the lectin from Vicia villosa seeds which reacts withcytotoxic T lymphoblasts. Mol. Immunol.. 73: 969-977,1981.
18. Kerbel, R. S., Florian, H., Man, M. S., Dennis, J., and McKencie, l. F. C.Carcmogenicity of tumor cell populations: origin of a putative H-2 isoantigenicloss variant tumor. J. Nati. Cancer Inst., 64:1221-1230,1980.
19. Kerbel, R. S., Twiddy, R. R., and Robertson, D. M. Induction of a tumor withgreatly increased metastatic growth potential by injection of cells from a lowmetastatic H-2 heterozygous tumor cell line into an H-2 incompatible parentalstrain. Int. J. Cancer, 22: 583-594, 1978.
20. Kimura, A. K., and Wigzell, H. Cell surface glycoproteins of murine cytotoxic Tlymphocytes. I. T145, a new cell surface glycoprotein selectively expressedon Ly 1-2* cytotoxic T lymphocytes. J. Exp. Med., 747: 1418-1434,1978.
21. Mannino, R. J., and Burger, M. M. Growth inhibition of animal cells bysuccinated concanavalin A. Nature (Lond.), 256: 19-22, 1975.
22. Meager, A., Ungkitchanukit, A., Nairn, R., and Hughes, R. C. Ricin resistancein baby hamster kidney cells. Nature (Lond.), 257:137-139,1975.
23. Narasimhan, S., Stanley, P., and Schachter, H. Control of glycoprotein biosynthesis: lectin-resistant mutant containing only one of two distinct A/-acetylglu-cosaminyltransferase activities present in wild type Chinese hamster ovarycells. J. Biol. Chem., 252. 3926-3933, 1977.
24. Paulson, J. C., Rearick, J. I., and Hill, R. L. Purification of a sialyltransferasefrom bovine colostrum by affinity chromatography on CDP-agarose. J. Biol.Chem., 252: 2363-2371, 1977.
25. Pereira, M. E. A., Kabat, F. A., and Sharon, N. Immunochemical studies on thespecificity of soybean agglutinin. Carbohydr. Res., 37: 89-102, 1974.
26. Raz, A., Bucana, C., McLellan, W., and Fidler, I. J. Distribution of membraneanionic sites on B16 melanoma variants with differing lung colonizing potential.Nature (Lond.), 284: 363-364,1980.
27. Raz, A., McLellan, W., Hart, I. R., Bucana, C. D., Hoyer, L. C., Sela, B. A.,Dragsten, P., and Fidler, I. J. Cell surface properties of B16 melanoma variantswith differing metastatic potential. Cancer Res., 40. 1645-1651, 1980.
28. Reading, C. L., Betoni, P. N., and Nicolson, G. L. Selection and in vivoproperties of lectin-attachment variants on malignant lymphosarcoma cell lines.J. Nati. Cancer Inst., 64: 1241-1249, 1980.
29. Schirrmacher, V., Cheingsong-Popov, R., and Amheiter, H. Hepatocyte-tumorcell interaction in vitro. I. Conditions for rosette formation and inhibition by antiH-2 antibody. J. Exp. Med., 757. 984-989,1980.
30. Schirrmacher, V., Fogel, M., Russmann, E., Bosslet, K., Altevogt, P., andBeck, L. Antigenic variation in cancer metastasis: immune escape versusimmune control. Cancer Met. Rev., 7: 241-274, 1982.
31. Schwartz, R. Ph.D. Dissertation, University of Braunschweig, 1982.32. Spiro, R. G. Periodate oxidation of glycoprotein fetuin. J. Biol. Chem., 239:
567-573, 1964.33. Spiro, R. G. Analysis of sugars found in glycoproteins. Methods Enzymol., 8:
3-26, 1966.34. Tao, T. W., and Burger, M. M. Non-metastasizing variants selected from
metastasizing melanoma cells. Nature (Lond.), 270: 437-438,1977.35. Warren, L. The thiobarbituric acid assay of sialic acids. J. Biol. Chem., 243:
1971-1979,1959.36. Yogeeswaran. G., and Salk, P. L. Metastatic potential is positively correlated
with cell surface sialylation of cultured murine tumor cell lines. Science (Wash.D.C.), 272:1514-1516,1981.
5144 CANCER RESEARCH VOL. 43
on May 8, 2018. © 1983 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

1983;43:5138-5144. Cancer Res Peter Altevogt, Mina Fogel, Rachanee Cheingsong-Popov, et al. Detected on Related High- and Low-Metastatic Tumor LinesDifferent Patterns of Lectin Binding and Cell Surface Sialylation
Updated version
http://cancerres.aacrjournals.org/content/43/11/5138
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/43/11/5138To request permission to re-use all or part of this article, use this link
on May 8, 2018. © 1983 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from