dimerization of signalling modules of the evgas and bvgas ... · dimerization of signalling modules...
TRANSCRIPT

Dimerization of signalling modules of the EvgAS and BvgASphosphorelay systems
Anne-Laure Perraud a, Karsten Rippe b, Marcus Bantsche¡ c, Michael Glocker c,Magnus Lucassen d, Kirsten Jung d, Walter Sebald e, Verena Weiss f , Roy Gross a;*
a Lehrstuhl fu«r Mikrobiologie, Biozentrum, Universita«t Wu«rzburg, Am Hubland, D-97074 Wu«rzburg, Germanyb Deutsches Krebsforschungszentrum, Abteilung Biophysik der Makromoleku«le, D-69115 Heidelberg, Germany
c Fakulta«t fu«r Chemie, Universita«t Konstanz, D-78457 Constanz, Germanyd Abteilung Mikrobiologie, Universita«t Osnabru«ck, D-46069 Osnabru«ck, Germany
e Lehrstuhl fu«r Physiologische Chemie II, Biozentrum, Universita«t Wu«rzburg, D-97074 Wu«rzburg, Germanyf Fakulta«t fu«r Biologie, Universita«t Konstanz, D-78434 Constanz, Germany
Received 13 December 1999; received in revised form 28 February 2000; accepted 9 March 2000
Abstract
Biophysical and biochemical properties of signalling proteins or domains derived from the unorthodox EvgAS and BvgAStwo-component phosphorelay systems of Escherichia coli and Bordetella pertussis were investigated. Oligomerization of theeffector proteins EvgA and BvgA and of truncated EvgS and BvgS derived signalling proteins containing the receiver andhistidine containing phosphotransfer (HPt) domains or comprising only the HPt domains were characterized by native gelelectrophoresis, gel permeation experiments and analytical ultracentrifugation. The results obtained by the different methodsare consistent with non-phosphorylated EvgA and BvgA proteins being dimers in solution with a dissociation constantsignificantly below 1 WM. In contrast, all sensor derived domains of EvgS and BvgS were observed to be monomers in vitro.No indications for a phosphorylation induced stimulation of oligomerization of the C-terminal histidine kinase domainscould be detected. In agreement with these data, surface plasmon resonance studies revealed a 2:1 stoichiometry in theinteraction of EvgA with the immobilized EvgS HPt domain and an affinity constant of 1.24U106 M31. ß 2000 ElsevierScience B.V. All rights reserved.
Keywords: Analytical ultracentrifugation; Surface plasmon resonance; Dimerization; Phosphorelay; BvgAS; EvgAS; Mass spectrometry
1. Introduction
Two-component systems are found in eubacteria,archaea, several lower eukaryotic organisms andplants [1^3]. Signals are perceived by a sensory his-
tidine kinase, which, after autophosphorylation at ahistidine residue in the conserved transmitter do-main, serves as a phosphodonor for phosphorylationof further signalling domain(s). In classical two-com-ponent systems, a one-step phosphotransfer occursfrom the phosphohistidine in the transmitter to anaspartic acid residue in the receiver domain of thee¡ector protein. Phosphorylation of the e¡ector pro-tein has regulatory consequences for its biologicalactivity. For instance, many two-component e¡ector
0167-4838 / 00 / $ ^ see front matter ß 2000 Elsevier Science B.V. All rights reserved.PII: S 0 1 6 7 - 4 8 3 8 ( 0 0 ) 0 0 0 5 2 - 2
* Corresponding author. Fax: +49-931-888-4402;http://www.biozentrum.uni-wuerzburg.de/microbiology/ ;E-mail : [email protected]
BBAPRO 36134 11-5-00
Biochimica et Biophysica Acta 1478 (2000) 341^354www.elsevier.com/locate/bba

proteins are transcription factors, which become ac-tivated by phosphorylation [4]. Instead of the phos-phohistidine presented by the histidine kinases, com-pounds such as acetyl phosphate may be used asphosphodonors for the receiver domains of the e¡ec-tor proteins in vitro and in some cases possibly alsoin vivo [5,6]. This demonstrates that the receiver do-mains of the e¡ector proteins harbor the enzymaticactivity for their phosphorylation. On the basis ofsequence similarities and di¡erences in their domainstructures, the e¡ector proteins can be classi¢ed intoseveral subfamilies [4].
In `unorthodox' two-component systems, a multi-step phosphorelay occurs between alternating histi-dine and aspartate residues (His-Asp-His-Asp) beforethe e¡ector proteins are activated [2]. These systemscontain additional signalling modules intercalatedbetween the transmitter domain of the sensor proteinand the receiver domain of the e¡ector protein.These additional domains consist of a second `inter-mediate' receiver domain and the so-called histidinecontaining phosphotransfer (HPt) domain [7]. Theintermediate receiver domains appear to be highlyrelated to the receiver domains found in the e¡ectorproteins. The various phosphorelay domains are ei-ther present as independent proteins or combined invarious combinations in complex multidomain pro-teins [3]. For example, in the case of the Bvg phos-phorelay system of Bordetella pertussis, the etiologi-cal agent of whooping cough, and of the highlyrelated Evg phosphorelay system of Escherichia colithree domains, the transmitter, the intermediate re-ceiver and the HPt domain, are fused in the singlemultidomain sensor proteins BvgS and EvgS, respec-tively [8^10]. The Bvg system of B. pertussis is knownto be the master regulator of virulence controllingthe expression of several toxins and adhesins [11].Nothing is known so far about the function of theEvg system in E. coli. However, in agreement withthe extensive sequence conservation between the twosystems, both appear to perceive the same environ-mental signals which include sulfate ions and temper-ature [8,9].
The structural consequences of phosphorylationfor the signal transduction proteins are not wellunderstood. Oligomerization of signalling domainsappears to be an important feature during signaltransduction by two-component systems. For exam-
ple, for several e¡ector proteins it has been reportedthat phosphorylation induces dimerization and/or co-operative binding to the respective target sites on theDNA [12^16]. Similarly, histidine kinases are able todimerize and autophosphorylation of the histidinekinases in fact may occur between two homodimericprotein partners [17]. Genetic and biochemical evi-dence indicated that the BvgA and BvgS proteinsof B. pertussis, the etiological agent of whoopingcough, also have dimerizing properties [9,18^20].
In the present report we attempted to gain moreinsight in the molecular basis of signal transductionin the Bvg and Evg phosphorelay systems of B. per-tussis and E. coli. We investigated the dimerizationand autophosphorylation properties of several sig-nalling domains derived from the unorthodox histi-dine kinases EvgS and BvgS and of their cognatee¡ector proteins EvgA and BvgA.
2. Materials and methods
2.1. Bacterial strains, plasmids and growth conditions
The bacterial strains and plasmids used in thisstudy are listed in Table 1. E. coli DH5K was usedas a host for gene cloning and E. coli M15 andSF120 for overexpression of proteins as describedpreviously [21,22]. The bacteria were grown eitherin Luria^Bertani (LB) liquid broth or on LB-agarplates. When necessary antibiotics were added tothe following ¢nal concentrations: ampicillin, 100Wg/ml; kanamycin, 50 Wg/ml.
2.2. Overexpression and puri¢cation oftwo-component proteins
The proteins analyzed are shown in Fig. 1. Forpuri¢cation the genes encoding the response regula-tors EvgA and BvgA were cloned in the expressionvector pCYTEXP1 [22] resulting in plasmids pCYT-EvgA and pCYT-BvgA [23]. The native proteinswere then overproduced as described [23]. The openreading frames encoding the sensor derived domainsEvgS-RO, EvgS-O, BvgS-RO and BvgS-O werecloned in the pQE30 and pQE31 vectors (Qiagen)creating an N-terminal His-tag and resulting in plas-mids pQE-EvgS-RO, pQE-EvgS-O, pQE-BvgS-RO
BBAPRO 36134 11-5-00
A.-L. Perraud et al. / Biochimica et Biophysica Acta 1478 (2000) 341^354342

and pQE-BvgS-O, respectively (Table 1). The pro-teins were overexpressed in E. coli M15 and puri¢edby a¤nity chromatography according to the manu-facturer's protocol (Qiagen). The detailed puri¢ca-tion protocols of the various proteins were describedrecently [23].
2.3. Biotinylation of EvgS-O and EvgS-RO
The two proteins EvgS-O and EvgS-RO were bio-tinylated using N-[biotinyl-6-amino]-capronate-N-succinimidylester (Fluka, Buchs, Switzerland) ac-cording to manufacturers instructions. A stock solu-tion of biotinylation reagent (0.5 Wg/Wl) in dimethyl-formamide was freshly prepared prior to reaction.Proteins (0.3 Wg/Wl) were dissolved in 50 mMNaHCO3 (pH 8.5). The modi¢cation reactions wereperformed at di¡erent protein/reagent ratios (1:2 and1:5, respectively) for 30 min at 37³C and stopped bythe addition of glacial acetic acid. Biotinylation sitesin the EvgS-RO and EvgS-O proteins were identi¢edby mass spectrometric peptide mapping as describedbelow.
2.4. Tryptic on-target digestion of biotinylatedEvgS-O
A protein solution (2 Wl) containing biotinylatedEvgS-O was spotted on the MALDI-MS target. Pro-teolytic on-target digestion was performed afterevaporation of the solvent. A freshly prepared tryp-sin solution (2 Wl ; 0.01 Wg/Wl ; Sigma, Deisenhofen,
Germany), dissolved in 50 mM NH4HCO3, pH 8,was added, resulting in an enzyme/substrate ratioof 1:100. The target was placed in a humid atmos-phere and was incubated at 37³C for 3 h. Subse-quently, the solvent was evaporated within 30 minat room temperature. The on-target digestion prod-ucts were analyzed by MALDI-MS peptide mappingas described below.
2.5. Avidin Sepharose a¤nity isolation of biotinylatedEvgS-O peptide
The biotinylated amino acid residue of EvgS-Owas identi¢ed using the avidin^biotin chemistry com-bined with MALDI-MS as described previously[24,25]. In brief, the biotinylated EvgS-O was enzy-matically digested in NH4HCO3 bu¡er, pH 7.8 for3 h at 37³C using endoprotease Asp-N (BoehringerMannheim, Mannheim, Germany). The enzyme/sub-strate ratio was 1:200. Avidin (Sigma, Deisenhofen,Germany) was coupled to NHS-activated Sepharoseusing standard protocols as suggested by the supplier(Amersham Pharmacia Biotech, Freiburg, Germany).Three Wl of avidin coupled Sepharose suspension inPBS bu¡er (30 mM sodium phosphate, 3 M NaCl,pH 7.2) was added to 5 Wl of the digestion mixture.The suspension was vortexed for approximately10 min, centrifuged to settle the Sepharose and thesupernatant was discarded. Several washing stepswere carried out with PBS (1U50 Wl) followed bydeionized water (2U100 Wl) and ¢nally the pelletwas resuspended in 3 Wl of a solvent mixture consist-
Table 1Strains and plasmids
Bacterial strains and plasmids Relevant features Source
E. coli DH5K strain for high e¤ciency transformation GibcoE. coli M15 strain for cloning and overproducing His-tagged proteins QiagenE. coli SF120 strain for overexpression of proteins [21]pQE30 and pQE31 expression vectors for N-terminal His-tag cloning QiagenpCYTEXP1 expression vector [22]pQE-BvgS-RO derivative of pQE30 expressing the His-tagged BvgS receiver and output (HPt)
domains[23]
pQE-BvgS-O derivative of pQE30 expressing the His-tagged BvgS output (HPt) domain [23]pQE-EvgS-RO derivative of pQE30 expressing the His-tagged EvgS receiver and HPt domains [23]pQE-EvgS-O derivative of pQE30 expressing the His-tagged EvgS HPt domain [23]pCYT-BvgA derivative of pCYTEXP1 expressing BvgA [23]pCYT-EvgA derivative of pCYTEXP1 expressing EvgA [23]
BBAPRO 36134 11-5-00
A.-L. Perraud et al. / Biochimica et Biophysica Acta 1478 (2000) 341^354 343

ing of 0.1% aqueous tri£uoroacetic acid (TFA) and40% CH3CN (v/v). This suspension was subjected tomass spectrometric analysis.
2.6. Mass spectrometry
MALDI-MS analyses were carried out with aBruker Bi£ex linear time-of-£ight spectrometer(Bruker-Franzen, Bremen, Germany). For molecularweight determination of the Bvg/Evg proteins 1 Wl ofsample solution was mixed with 0.8 Wl of a saturatedsolution of K-cyano-4-hydroxy cinnamic acid(HCCA) dissolved in CH3CN/0.1% TFA (2:1, v/v)directly on the target. After evaporation of the sol-vent, the matrix/analyte mixture was recrystallizedfrom 1.5 Wl CH3CN/0.1% TFA (2:1, v/v) prior todata acquisition.
In the case of avidin Sepharose a¤nity separatedsamples a thin layer of matrix (saturated HCCA so-lution in acetone) was ¢rst placed on the target.After evaporation of the solvent, 1 Wl of Sepharoseslurry was added and allowed to dry. Finally, 0.4 Wlof a saturated solution of HCCA dissolved inCH3CN/0.1% TFA (2:1, v/v) was added. The driedSepharose was blown o¡ by applying a stream ofnitrogen gas.
Nano-ESI-MS spectra were recorded on a VestecA 201 single quadrupole mass spectrometer (PerSep-tive Biosystems, Framingham, MA, USA), as re-ported elsewhere [26,27]. Volumes of 0.5^2 Wl wereloaded by dipping the capillaries into the correspond-ing sample solutions.
2.7. Native polyacrylamide gradient gels and gelpermeation experiments
Native polyacrylamide gradient gels were preparedas described previously [28]. The polyacrylamide gra-dient ranged from 4 to 28% and was combined witha sucrose gradient ranging from 0 to 20%. Electro-phoresis was carried out for 12 h at 100^150 V. Asmolecular weight markers the puri¢ed proteins of theMW-GF-70 Kit (Sigma) were used which includealdolase (158 kDa), L-lactoglobulin (36.6 kDa), an-hydrase (29 kDa) and cytochrome c (12.6 kDa). Thegel permeation experiments were carried out accord-ing to the manufacturer's protocols using a Superdex75 HR-10 FPLC column (Pharmacia) in NH4HCO3
bu¡er at pH 8.5. For molecular weight calibration ofthe column the proteins of the MW-GH-70 Kit (Sig-ma) were used once more.
2.8. Chemical synthesis of acetyl [32P]phosphate andphosphorylation experiments
For the synthesis of acetyl [32P]phosphate the pro-tocol of Stadtman [29] was modi¢ed as follows [30] :0.19 ml of pyridine, 0.3 ml of 0.33 M K2HPO4 and0.1 ml of carrier-free [32P]orthophosphate (speci¢cactivity 10 mCi ml31 ; Amersham Buchler) werestirred on ice in a 16U100 Pyrex tube. Then 22 Wlacetic anhydride was added slowly and incubated onice. LiOH (4 N) was added to adjust to pH 7. Fi-nally, acetyl [32P]phosphate was precipitated with4.5 ml of 100% ethanol for 1 h on ice. The precipi-tated acetyl [32P]phosphate was collected by centrifu-gation, washed twice with 5 ml cold ethanol anddried over KOH and CaCl2. Acetyl [32P]phosphatewas solved in 180 Wl of 50 mM Tris^HCl, pH 7.5, 5%glycerol, 0.1 mM EDTA and 1 mM dithiothreitoland used immediately for the phosphorylation assays.
For in vitro phosphorylation with acetyl[32P]phosphate the proteins were incubated in50 mM Tris^HCl, pH 7.5, 20 mM MgCl2 and300 mM KCl with 20 mM acetyl [32P]phosphate at30³C. The reaction was stopped using SDS^PAGEsample bu¡er [31] and subjected to SDS^PAGE. Invitro phosphorylation of proteins with [33P]QATPwas performed as described recently [23]. Thephosphorylated proteins were detected using a phos-phoimager (Molecular Dynamics).
2.9. Analytical sedimentation equilibriumultracentrifugation (AUC)
Sedimentation equilibrium experiments were car-ried out on a Beckman Instruments Optima XL-Aanalytical ultracentrifuge with absorbance optics.Protein solutions of A280 = 0.1^0.2 (correspondingto concentrations of 10^20 WM in terms of proteinmonomer) in a bu¡er containing 50 mM Tris^HCl,pH 7.5, 50 mM KCl and 4% glycerol (HK bu¡er)were analyzed at 10 000 and 25 000 rpm at 4³C withan An60Ti rotor. For measurements of the responseregulator proteins BvgA and EvgA the bu¡er wassupplemented with 5 mM MgCl2 (RR bu¡er). The
BBAPRO 36134 11-5-00
A.-L. Perraud et al. / Biochimica et Biophysica Acta 1478 (2000) 341^354344

e¡ect of chemical phosphorylation of truncatedforms of BvgS and EvgS was studied by the additionof 10 mM acetyl phosphate and 10 mM MgCl2. Cal-culation of the partial speci¢c volume from the ami-no acid composition of the proteins and analysis ofthe data by one exponential ¢t was conducted asdescribed recently [32]. The density b of both theHK and RR bu¡er was measured to be 1.017 gml31 at 4³C with a DMA 5000 density meter (AntonPaar, Graz, Austria).
2.10. Biosensor experiment
All surface plasmon resonance (SPR) experimentswere carried out in a BIAcore 2000 system (Pharma-cia Biosensor) at 25³C with a £ow rate of 10 Wlmin31 in HBS bu¡er consisting of 10 mM HEPES,pH 7.4, 150 mM NaCl, 3.4 mM EDTA, 0.005% sur-factant P20 with a data collection rate of 2.5 s31. Toanalyze the interaction of EvgA with the HPt do-main of EvgS, the biotinylated derivatives of theEvgS-RO and EvgS-O proteins containing eitherthe EvgS receiver and the HPt domain or only theHPt domain, respectively, were coupled to BIAcorestreptavidin sensor chips (Pharmacia) at a density ofabout 1000 resonance units (RU) as described re-cently [33]. The sensorgrams were generated andevaluated using the BIAevaluation 2.1 software.
3. Results
3.1. Dimerization properties of e¡ector proteins andisolated signalling domains
The e¡ector proteins EvgA and BvgA and the pre-
viously described His-tagged signalling domainsEvgS-RO, EvgS-O, BvgS-RO and BvgS-O [9,23](Fig. 1) were isolated in mg quantities and subjectedto biophysical and biochemical characterizationmethods including native gradient gel electrophore-sis, gel permeation chromatography (GPC) andAUC. First, all protein samples were checked forpurity and homogeneity by SDS^PAGE (data notshown) and mass spectrometric molecular mass de-terminations. The determined molecular masses of allproteins precisely matched the calculated massesfrom the DNA-translated amino acid sequences (Ta-ble 2). The mass spectrometric results, thus, provedthe integrity of all proteins and constructs and con-¢rmed the absence of high molecular mass contami-nants in the protein samples.
Providing initial evidence for the dimeric status ofEvgA, on native gradient gels the 22.69 kDa proteinmigrated at a position of about 46 kDa (Fig. 2, top).Under denaturing conditions the EvgA protein mi-grated at a position corresponding to its monomericform, but a band at the position of the dimer could
Table 2MALDI-MS molecular weight determinations of e¡ector pro-teins and signalling modules
Protein Mr MW (experimental)a
BvgA 22 952 22 949BvgS-RO 30 892 30 915BvgS-O 17 279 17 275EvgA 22 690 22 693EvgS-RO 31 115 31 120EvgS-O 16 127 16 126aCon¢rmed by ESI-MS.
Fig. 1. Schematic presentation of the architecture of the pro-teins of the BvgAS and EvgAS two-component systems andtheir derivatives used in this study. At the top the domainstructure and the phosphorelay within the wild type systems isdepicted schematically. At the bottom the domain compositionof the histidine kinase derived proteins BvgS-RO, BvgS-O,EvgS-RO and EvgS-O and of the e¡ector proteins BvgA andEvgA is shown, respectively. The box in front of the histidinekinase derived proteins indicates the N-terminal His-tag.
BBAPRO 36134 11-5-00
A.-L. Perraud et al. / Biochimica et Biophysica Acta 1478 (2000) 341^354 345
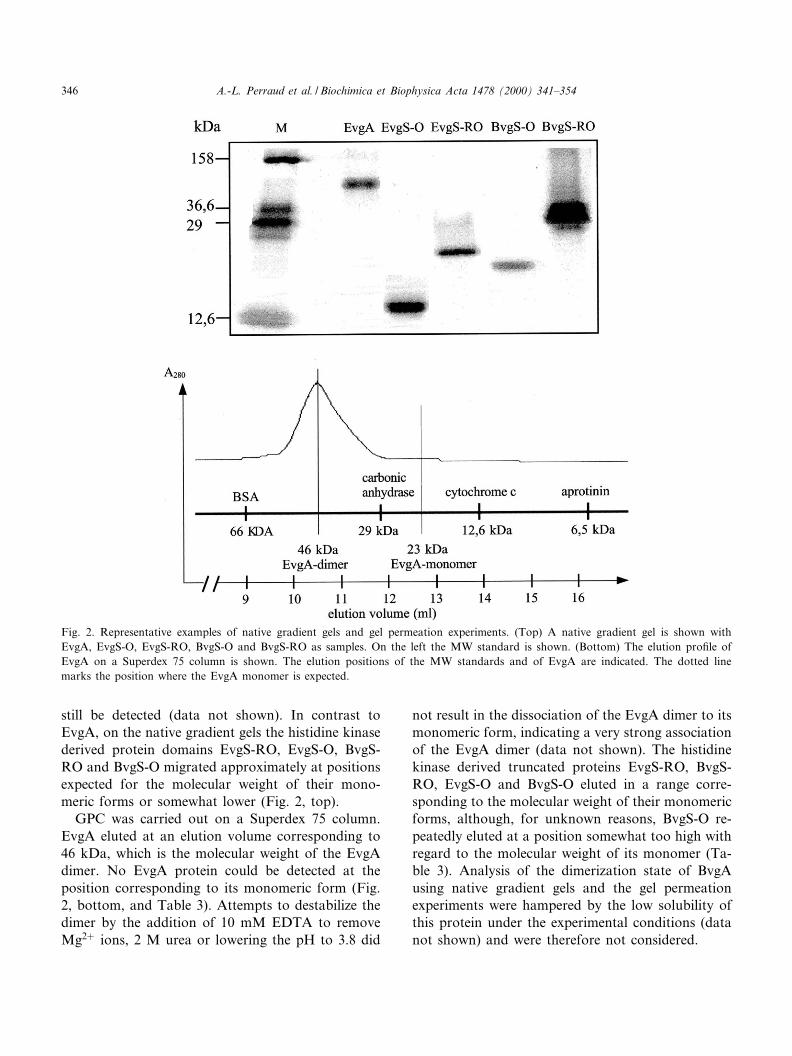
still be detected (data not shown). In contrast toEvgA, on the native gradient gels the histidine kinasederived protein domains EvgS-RO, EvgS-O, BvgS-RO and BvgS-O migrated approximately at positionsexpected for the molecular weight of their mono-meric forms or somewhat lower (Fig. 2, top).
GPC was carried out on a Superdex 75 column.EvgA eluted at an elution volume corresponding to46 kDa, which is the molecular weight of the EvgAdimer. No EvgA protein could be detected at theposition corresponding to its monomeric form (Fig.2, bottom, and Table 3). Attempts to destabilize thedimer by the addition of 10 mM EDTA to removeMg2� ions, 2 M urea or lowering the pH to 3.8 did
not result in the dissociation of the EvgA dimer to itsmonomeric form, indicating a very strong associationof the EvgA dimer (data not shown). The histidinekinase derived truncated proteins EvgS-RO, BvgS-RO, EvgS-O and BvgS-O eluted in a range corre-sponding to the molecular weight of their monomericforms, although, for unknown reasons, BvgS-O re-peatedly eluted at a position somewhat too high withregard to the molecular weight of its monomer (Ta-ble 3). Analysis of the dimerization state of BvgAusing native gradient gels and the gel permeationexperiments were hampered by the low solubility ofthis protein under the experimental conditions (datanot shown) and were therefore not considered.
Fig. 2. Representative examples of native gradient gels and gel permeation experiments. (Top) A native gradient gel is shown withEvgA, EvgS-O, EvgS-RO, BvgS-O and BvgS-RO as samples. On the left the MW standard is shown. (Bottom) The elution pro¢le ofEvgA on a Superdex 75 column is shown. The elution positions of the MW standards and of EvgA are indicated. The dotted linemarks the position where the EvgA monomer is expected.
BBAPRO 36134 11-5-00
A.-L. Perraud et al. / Biochimica et Biophysica Acta 1478 (2000) 341^354346
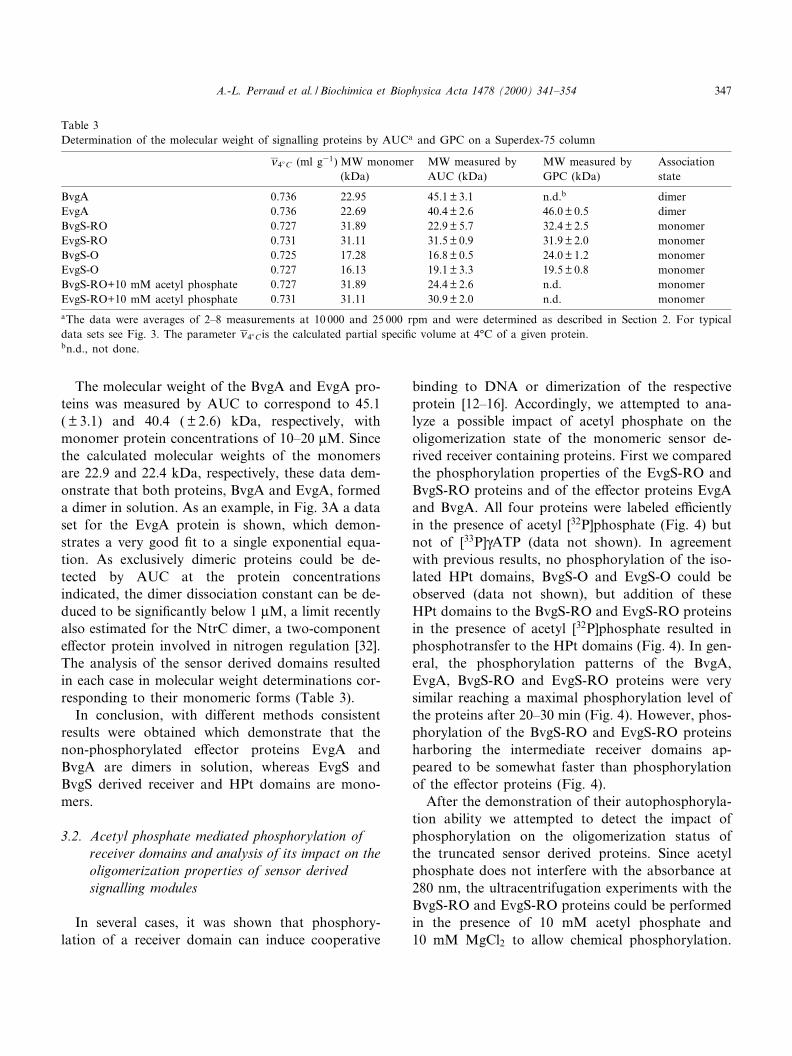
The molecular weight of the BvgA and EvgA pro-teins was measured by AUC to correspond to 45.1( þ 3.1) and 40.4 ( þ 2.6) kDa, respectively, withmonomer protein concentrations of 10^20 WM. Sincethe calculated molecular weights of the monomersare 22.9 and 22.4 kDa, respectively, these data dem-onstrate that both proteins, BvgA and EvgA, formeda dimer in solution. As an example, in Fig. 3A a dataset for the EvgA protein is shown, which demon-strates a very good ¢t to a single exponential equa-tion. As exclusively dimeric proteins could be de-tected by AUC at the protein concentrationsindicated, the dimer dissociation constant can be de-duced to be signi¢cantly below 1 WM, a limit recentlyalso estimated for the NtrC dimer, a two-componente¡ector protein involved in nitrogen regulation [32].The analysis of the sensor derived domains resultedin each case in molecular weight determinations cor-responding to their monomeric forms (Table 3).
In conclusion, with di¡erent methods consistentresults were obtained which demonstrate that thenon-phosphorylated e¡ector proteins EvgA andBvgA are dimers in solution, whereas EvgS andBvgS derived receiver and HPt domains are mono-mers.
3.2. Acetyl phosphate mediated phosphorylation ofreceiver domains and analysis of its impact on theoligomerization properties of sensor derivedsignalling modules
In several cases, it was shown that phosphory-lation of a receiver domain can induce cooperative
binding to DNA or dimerization of the respectiveprotein [12^16]. Accordingly, we attempted to ana-lyze a possible impact of acetyl phosphate on theoligomerization state of the monomeric sensor de-rived receiver containing proteins. First we comparedthe phosphorylation properties of the EvgS-RO andBvgS-RO proteins and of the e¡ector proteins EvgAand BvgA. All four proteins were labeled e¤cientlyin the presence of acetyl [32P]phosphate (Fig. 4) butnot of [33P]QATP (data not shown). In agreementwith previous results, no phosphorylation of the iso-lated HPt domains, BvgS-O and EvgS-O could beobserved (data not shown), but addition of theseHPt domains to the BvgS-RO and EvgS-RO proteinsin the presence of acetyl [32P]phosphate resulted inphosphotransfer to the HPt domains (Fig. 4). In gen-eral, the phosphorylation patterns of the BvgA,EvgA, BvgS-RO and EvgS-RO proteins were verysimilar reaching a maximal phosphorylation level ofthe proteins after 20^30 min (Fig. 4). However, phos-phorylation of the BvgS-RO and EvgS-RO proteinsharboring the intermediate receiver domains ap-peared to be somewhat faster than phosphorylationof the e¡ector proteins (Fig. 4).
After the demonstration of their autophosphoryla-tion ability we attempted to detect the impact ofphosphorylation on the oligomerization status ofthe truncated sensor derived proteins. Since acetylphosphate does not interfere with the absorbance at280 nm, the ultracentrifugation experiments with theBvgS-RO and EvgS-RO proteins could be performedin the presence of 10 mM acetyl phosphate and10 mM MgCl2 to allow chemical phosphorylation.
Table 3Determination of the molecular weight of signalling proteins by AUCa and GPC on a Superdex-75 column
X 4�C (ml g31) MW monomer(kDa)
MW measured byAUC (kDa)
MW measured byGPC (kDa)
Associationstate
BvgA 0.736 22.95 45.1 þ 3.1 n.d.b dimerEvgA 0.736 22.69 40.4 þ 2.6 46.0 þ 0.5 dimerBvgS-RO 0.727 31.89 22.9 þ 5.7 32.4 þ 2.5 monomerEvgS-RO 0.731 31.11 31.5 þ 0.9 31.9 þ 2.0 monomerBvgS-O 0.725 17.28 16.8 þ 0.5 24.0 þ 1.2 monomerEvgS-O 0.727 16.13 19.1 þ 3.3 19.5 þ 0.8 monomerBvgS-RO+10 mM acetyl phosphate 0.727 31.89 24.4 þ 2.6 n.d. monomerEvgS-RO+10 mM acetyl phosphate 0.731 31.11 30.9 þ 2.0 n.d. monomeraThe data were averages of 2^8 measurements at 10 000 and 25 000 rpm and were determined as described in Section 2. For typicaldata sets see Fig. 3. The parameter X 4�C is the calculated partial speci¢c volume at 4³C of a given protein.bn.d., not done.
BBAPRO 36134 11-5-00
A.-L. Perraud et al. / Biochimica et Biophysica Acta 1478 (2000) 341^354 347
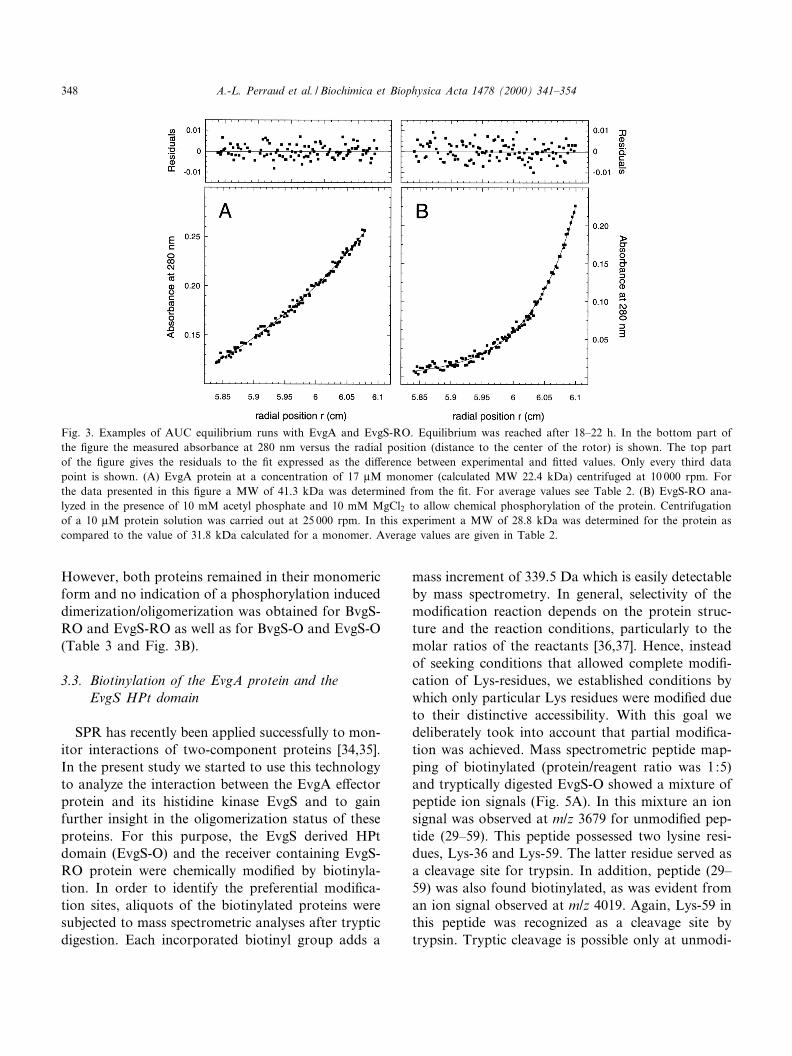
However, both proteins remained in their monomericform and no indication of a phosphorylation induceddimerization/oligomerization was obtained for BvgS-RO and EvgS-RO as well as for BvgS-O and EvgS-O(Table 3 and Fig. 3B).
3.3. Biotinylation of the EvgA protein and theEvgS HPt domain
SPR has recently been applied successfully to mon-itor interactions of two-component proteins [34,35].In the present study we started to use this technologyto analyze the interaction between the EvgA e¡ectorprotein and its histidine kinase EvgS and to gainfurther insight in the oligomerization status of theseproteins. For this purpose, the EvgS derived HPtdomain (EvgS-O) and the receiver containing EvgS-RO protein were chemically modi¢ed by biotinyla-tion. In order to identify the preferential modi¢ca-tion sites, aliquots of the biotinylated proteins weresubjected to mass spectrometric analyses after trypticdigestion. Each incorporated biotinyl group adds a
mass increment of 339.5 Da which is easily detectableby mass spectrometry. In general, selectivity of themodi¢cation reaction depends on the protein struc-ture and the reaction conditions, particularly to themolar ratios of the reactants [36,37]. Hence, insteadof seeking conditions that allowed complete modi¢-cation of Lys-residues, we established conditions bywhich only particular Lys residues were modi¢ed dueto their distinctive accessibility. With this goal wedeliberately took into account that partial modi¢ca-tion was achieved. Mass spectrometric peptide map-ping of biotinylated (protein/reagent ratio was 1:5)and tryptically digested EvgS-O showed a mixture ofpeptide ion signals (Fig. 5A). In this mixture an ionsignal was observed at m/z 3679 for unmodi¢ed pep-tide (29^59). This peptide possessed two lysine resi-dues, Lys-36 and Lys-59. The latter residue served asa cleavage site for trypsin. In addition, peptide (29^59) was also found biotinylated, as was evident froman ion signal observed at m/z 4019. Again, Lys-59 inthis peptide was recognized as a cleavage site bytrypsin. Tryptic cleavage is possible only at unmodi-
Fig. 3. Examples of AUC equilibrium runs with EvgA and EvgS-RO. Equilibrium was reached after 18^22 h. In the bottom part ofthe ¢gure the measured absorbance at 280 nm versus the radial position (distance to the center of the rotor) is shown. The top partof the ¢gure gives the residuals to the ¢t expressed as the di¡erence between experimental and ¢tted values. Only every third datapoint is shown. (A) EvgA protein at a concentration of 17 WM monomer (calculated MW 22.4 kDa) centrifuged at 10 000 rpm. Forthe data presented in this ¢gure a MW of 41.3 kDa was determined from the ¢t. For average values see Table 2. (B) EvgS-RO ana-lyzed in the presence of 10 mM acetyl phosphate and 10 mM MgCl2 to allow chemical phosphorylation of the protein. Centrifugationof a 10 WM protein solution was carried out at 25 000 rpm. In this experiment a MW of 28.8 kDa was determined for the protein ascompared to the value of 31.8 kDa calculated for a monomer. Average values are given in Table 2.
BBAPRO 36134 11-5-00
A.-L. Perraud et al. / Biochimica et Biophysica Acta 1478 (2000) 341^354348
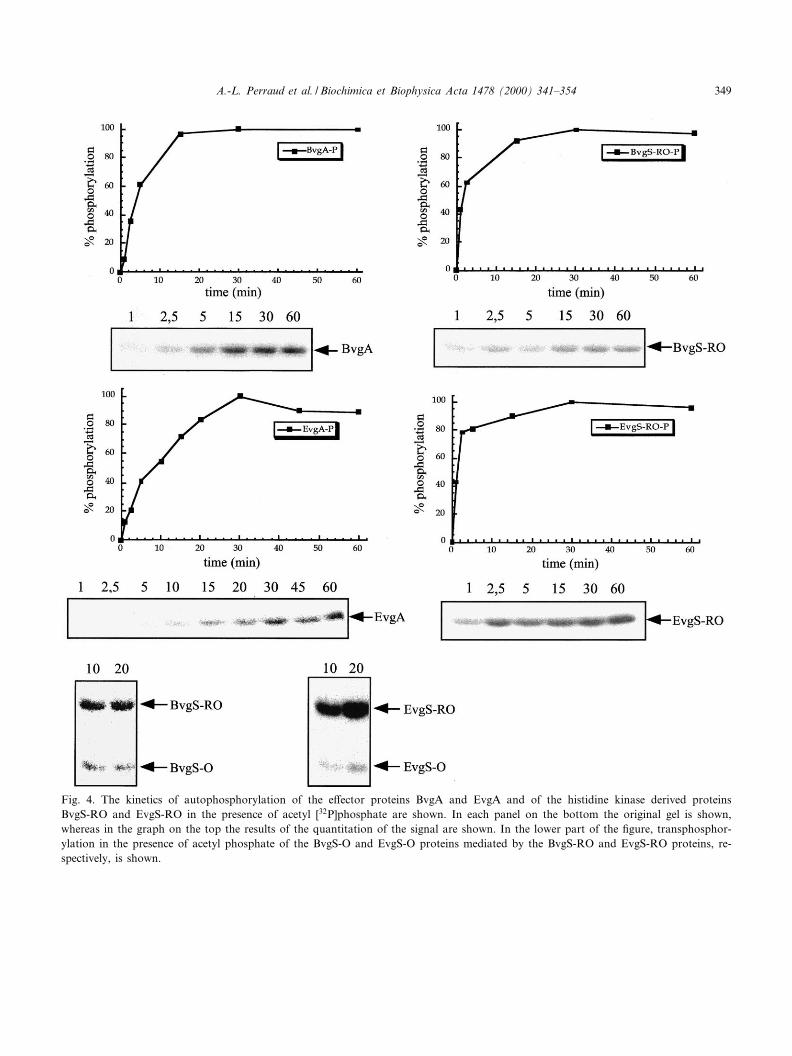
Fig. 4. The kinetics of autophosphorylation of the e¡ector proteins BvgA and EvgA and of the histidine kinase derived proteinsBvgS-RO and EvgS-RO in the presence of acetyl [32P]phosphate are shown. In each panel on the bottom the original gel is shown,whereas in the graph on the top the results of the quantitation of the signal are shown. In the lower part of the ¢gure, transphosphor-ylation in the presence of acetyl phosphate of the BvgS-O and EvgS-O proteins mediated by the BvgS-RO and EvgS-RO proteins, re-spectively, is shown.
BBAPRO 36134 11-5-00
A.-L. Perraud et al. / Biochimica et Biophysica Acta 1478 (2000) 341^354 349

¢ed lysine residues. Hence, mass spectrometry andthe enzymatic cleavage speci¢city located the pre-dominant biotinylation site at Lys-36. Lysine residueLys-36 in the HPt domain (EvgS-O) corresponds toLys-1092 in the EvgS wild type sequence.
Biotinylation at Lys-36 was independently provenby using mass spectrometry together with speci¢cbiotin a¤nity isolation of proteolytic peptides afterdigestion of biotinylated EvgS-O (protein/reagent ra-
tio was 1:2). After digestion with Asp-N protease thepeptide mixture was incubated with Sepharosecoupled avidin. As avidin exhibits a very strongbinding a¤nity to biotin, only biotinylated peptidesremained in the Sepharose precipitate whereas allother peptides were removed in the supernatant byextensive washing. After transferring the Sepharoseavidin/biotinylated peptide complex to the MALDI-MS target and after addition of the matrix, spectrawere recorded in which the only observed compo-nents were ions from singly and multiply chargedavidin and from biotinylated EvgS-O peptide (33^59) (Fig. 5B). Similar experiments were carried outwith the biotinylated EvgS-RO protein and showedthat this signalling module was modi¢ed preferen-tially within its receiver domain at the conserved ly-sine residue which corresponds to K1059 in wild typeEvgS (data not shown).
3.4. Interaction between EvgA and the EvgS HPtdomain
To characterize the interaction of the solubleEvgA protein with the immobilized histidine kinasederived protein domains using the BIAcore biosensorsystem, increasing EvgA concentrations were addedto the sensor chip ranging from 0.11 to 11 WM. Fig. 6shows the SPR signals expressed in arbitrary RUobtained at di¡erent EvgA concentrations with theimmobilized EvgS-O protein. The a¤nity constant ofthe EvgS-O/EvgA interaction was estimated to be1.24U106 ( þ 1.3U104) M31 demonstrating a rela-tively weak but speci¢c interaction of these proteins.Evidence for the stoichiometry of the interactionpartners could be deduced from the SPR signals, asthe relative amount of the two protein partners canbe determined considering the ratio of the SPR signaland the molecular weight of the respective proteins.The respective values were found to be 0.065 and0.038 for EvgS-O and EvgA, respectively, indicatinga 1:2 stoichiometry. The same a¤nity constant andstoichiometry of the interacting partners were ob-tained with the immobilized EvgS-RO protein (datanot shown). This stoichiometry is in agreement withthe results reported above indicating that EvgAforms dimers in solution. The e¡ect of phosphory-lation of the EvgS-RO protein on its interaction withEvgA could not be further analyzed, because, as
Fig. 5. Mass spectrometric identi¢cation of the biotinylationsite in EvgS-O. (A) The MALDI-spectrum (range: m/z 2500^4500) obtained from biotinylated and tryptically digested EvgS-O shows a mixture of peptide ion signals. The peptide ion sig-nal that contains the biotinylated residue K-36 is indicated. (B)MALDI-MS analysis of biotinylated EvgS-O peptide obtainedby digestion with endoproteinase Asp-N and subsequent a¤nityisolation using avidin Sepharose. After extensive washing a sin-gle biotinylated EvgS-O peptide remained and was detected to-gether with ion signals belonging to avidin. A indicates avidin;A* represents a deglycosylated form of avidin. Numbers givem/z values. Numbers in parentheses refer to amino acid posi-tions as in the EvgS-O sequence. HCCA was used as a matrix.
BBAPRO 36134 11-5-00
A.-L. Perraud et al. / Biochimica et Biophysica Acta 1478 (2000) 341^354350

pointed out above, biotinylation of the EvgS-ROprotein occurred at the crucial lysine residue withinthe receiver domain. The interaction of the BvgS-Oand BvgA proteins could not be analyzed in thissystem due to solubility problems with the BvgAprotein under the given experimental conditions(data not shown).
4. Discussion
Phosphorylation of the e¡ector proteins by theircognate histidine kinases is the crucial event whichregulates their biological activity [4]. Although thestructural consequences of phosphorylation on thee¡ector proteins are not well understood, in severalcases it has been observed that phosphorylation in-duces dimerization and/or cooperative binding toDNA. [12^16]. Thus, e¡ector proteins engaged inthe activation of transcription are at least in partactivated by increasing their a¤nity to their respec-tive target sites. Dimerization/oligomerization of sig-nalling domains may therefore be an important fea-ture involved in the phosphorylation dependentregulation of the biological activity of the e¡ectorproteins. In the present study we analyzed the oligo-merization state of the non-phosphorylated BvgAand EvgA e¡ector proteins. As the multi-domain
BvgS and EvgS histidine kinases are integral mem-brane proteins and as such di¤cult to study, we an-alyzed several truncated derivatives lacking the trans-membrane domains of the two proteins. Previouslywe have shown that the puri¢ed response regulatorsand also the truncated proteins derived from the his-tidine kinases retain important activities such asphosphorylation properties, DNA binding activityor the capability of transcomplementation in vivo[9,18,23]. As indicated by mass spectrometry the pu-ri¢ed EvgA and BvgA proteins were not phosphor-ylated at the time of their analysis (Table 2). Thelack of phosphorylation corresponds to the factthat, in the case of BvgA, the cognate histidine ki-nase BvgS is not present in the overexpressing E. colistrain, whereas in the case of EvgA its overexpres-sion was achieved under growth conditions whichinactivated the EvgS histidine kinase (addition ofMgSO4 to the medium) [8].
4.1. Oligomerization state of signalling proteinsderived from the BvgAS and EvgAS systems
Consistently, the AUC experiments, the native gra-dient gels and gel permeation experiments demon-strated that the non-phosphorylated e¡ector proteinsEvgA and BvgA are dimers in solution. These resultsare in agreement with preliminary data publishedpreviously which also indicated the occurrence ofdimeric forms of BvgA in vivo [9,19,20]. Due to ex-tensive sequence similarities the EvgA and BvgA pro-teins are classi¢ed in the so-called NarL subfamily ofe¡ector proteins. Although previously considered tobe a monomer in solution which dimerizes uponphosphorylation, the recent comparison of severaldi¡erent crystal structures of the closely relatedNarL e¡ector protein indicated that also NarL mayhave the tendency to form homodimers in the non-phosphorylated form [38,39]. In the future, it will beinteresting to analyze the oligomerization state ofthese proteins after phosphorylation and/or afterbinding of DNA. In contrast to the e¡ector proteins,no indication for a dimeric state of the non-phosphorylated histidine kinase derived domainsBvgS-RO, BvgS-O, EvgS-RO and EvgS-O could befound.
To analyze whether phosphorylation may induceoligomerization of the histidine kinase derived pro-
Fig. 6. Graphic representation of the RU detected with EvgS-Oimmobilized on the BIAcore sensor chip after addition of in-creasing concentrations of EvgA.
BBAPRO 36134 11-5-00
A.-L. Perraud et al. / Biochimica et Biophysica Acta 1478 (2000) 341^354 351

teins, we performed ultracentrifugation experimentsin the presence of acetyl phosphate. This was possi-ble, because, as discussed below, the histidine kinasederived intermediate receiver domains are able toautophosphorylate in vitro in the presence of acetylphosphate (Fig. 4). Under the experimental condi-tions of ultracentrifugation acetyl phosphate is ex-pected to be stable. Hence, di¡erences in the sedi-mentation properties of the proteins in its absenceor presence should be detectable. However, no indi-cation for a phosphorylation dependent dimerizationof these domains could be observed (Fig. 3B). There-fore, it is likely that these protein domains do notoligomerize upon phosphorylation.
4.2. Phosphorylation of receiver domain containingproteins
In the case of the receiver domains of e¡ector pro-teins it was shown that they harbor the enzymaticactivity for phosphorylation and, accordingly, areable to phosphorylate in the presence of compoundssuch as acetyl phosphate. Here we show, that theintermediate receiver domains derived from the un-orthodox histidine kinases EvgS and BvgS are alsoable to phosphorylate using 32P-labelled acetyl phos-phate as a substrate. As shown in Fig. 4, the phos-phorylation properties of the e¡ector proteins and ofthe proteins containing the intermediate receiver do-mains are very similar. Note, however, that the in-terpretation of the phosphorylation patterns musttake into account that the visible phosphorylationsignals of the EvgS-RO and BvgS-RO proteins deriveexclusively from the phosphohistidine of their HPtdomains. It had already been reported that the phos-phoaspartates of the EvgS and BvgS receiver do-mains are very unstable and can not be visualizeddirectly after SDS^PAGE [23,40]. The HPt domainsrequire phosphorylation of the intermediate receiverdomains prior to their phosphorylation. Therefore,the phosphorylation signals detected with the EvgS-RO and BvgS-RO proteins re£ect the phosphory-lation of their receiver domains only indirectly (Fig.4) [9,40]. One other intermediate receiver domain ofa multi-domain histidine kinase, DokA of Dictyoste-lium discoideum, has recently been shown to be ableto autophosphorylate [41]. It is likely that, similar tothe receiver domains of e¡ector proteins, the inter-
mediate receiver domains present in unorthodox his-tidine kinases generally harbor the enzymatic activityfor the phosphotransfer reaction.
4.3. Characterization of the EvgA^EvgS interactionby SPR
The biosensor experiments allowed preliminarycharacterization of the interaction of EvgA with theEvgS HPt domain (Fig. 6). The a¤nity constant wascalculated to be about 1.24U106 M31, which indi-cates a relatively weak interaction of the two proteinsas compared to results obtained with the CheA/CheY and VanS/VanR two-component systems[34,42]. It is possible that this relatively low a¤nitymay be the consequence of a lack of phosphorylationof the histidine kinase derived protein domains inour biosensor experiments. In fact, an impact ofphosphorylation on the stability of the complex be-tween the histidine kinase and the e¡ector proteinwas recently demonstrated for the CheA/CheY pro-teins [34] and, accordingly, will be analyzed in thefuture with the Evg and Bvg proteins. Alternatively,it is possible that an e¤cient interaction betweenEvgA and EvgS requires an EvgS dimer. All histidinekinases analyzed so far including BvgS are able toform homodimers. Homodimerization is mediated bythe transmitter domains of the histidine kinases andappears to be independent of the phosphorylationstate of the proteins [9,28,43,44]. As shown here,the EvgS-RO and EvgS-O proteins lacking theEvgS transmitter domain are monomers, a confor-mation which may not allow a high a¤nity interac-tion of the proteins.
Recently, Utsumi et al. attempted to characterizethe oligomerization state of EvgA using SPR. Forthis purpose they coupled EvgA to the sensor chipand added soluble non-phosphorylated or phosphor-ylated EvgA. However, as no resonance signal couldbe detected, it was concluded that EvgA is a mono-mer independent of its phosphorylation state [35]. Inmarked contrast, AUC, native gradient gels and thegel permeation experiments described here consis-tently demonstrate that non-phosphorylated EvgAis a dimer in solution. Moreover, the biosensor ex-periments revealed that EvgA interacts as a dimerwith the EvgS HPt module coupled to the sensorchip. There are several possible explanations for the
BBAPRO 36134 11-5-00
A.-L. Perraud et al. / Biochimica et Biophysica Acta 1478 (2000) 341^354352

di¡erent results. We show that EvgA is a dimer insolution and as such may not interact anymore withEvgA protein coupled to the sensor chip. Also, asshown here for the EvgS-RO protein, chemical mod-i¢cation of the protein required for its immobiliza-tion on the sensor chip can signi¢cantly interferewith important functional parts of a protein. In thecase of the EvgS-RO protein a strictly conserved ly-sine residue important for the structural integrity ofthe receiver domain and for phosphorylation inducedconformational changes is the main target of bioti-nylation (Fig. 5). As the structure of the chemicallymodi¢ed EvgA proteins was not characterized in theprevious work, it is possible that the correspondinglysine residue within its receiver domain was also apreferred target for biotinylation of EvgA and maytherefore interfere strongly with the structure of theimmobilized protein and with its biophysical proper-ties.
Acknowledgements
We thank Dagmar Beier for critical reading of themanuscript. This work was supported by grantGr1243/2-4 from the Deutsche Forschungsgemein-schaft (Priority program `Regulatory Networks inBacteria') and by the Fonds der Chemischen Indus-trie.
References
[1] W.F. Loomis, A. Kuspa, G. Shaulsky, Curr. Opin. Micro-biol. 1 (1997) 643^648.
[2] J.L. Appleby, J.S. Parkinson, R.B. Bourret, Cell 86 (1996)845^848.
[3] A.-L. Perraud, V. Weiss, R. Gross, Trends Microbiol. 7(1999) 115^120.
[4] J.B. Stock, A.M. Stock, J.M. Mottonen, Nature 344 (1990)395^400.
[5] W.R. McCleary, J.B. Stock, A.J. Ninfa, J. Bacteriol. 175(1993) 2793^2798.
[6] J. Feng, M.R. Atkinson, W.R. McCleary, J.B. Stock, B.L.Wanner, A.J. Ninfa, J. Bacteriol. 174 (1992) 6061^6070.
[7] K. Ishige, S. Nagasawa, S. Tokishita, T. Mizuno, EMBO J.13 (1994) 5195^5202.
[8] R. Utsumi, S. Katayama, M. Taniguchi, T. Horie, M. Ikeda,S. Igaki, H. Nakagawa, A. Miwa, H. Tanabe, M. Noda,Gene 140 (1994) 73^77.
[9] D. Beier, B. Schwarz, T.M. Fuchs, R. Gross, J. Mol. Biol.248 (1995) 596^610.
[10] M.A. Uhl, J.F. Miller, EMBO J. 15 (1996) 1028^1036.[11] B. Arico, J.F. Miller, C. Roy, S. Stibitz, D. Monack, S.
Falkow, R. Gross, R. Rappuoli, Proc. Natl. Acad. Sci.USA 86 (1989) 6671^6675.
[12] U. Fiedler, V. Weiss, EMBO J. 14 (1995) 3696^3705.[13] K.J. Huang, C.Y. Lan, M.M. Igo, Proc. Natl. Acad. Sci.
USA 94 (1997) 2828^2832.[14] V. Weiss, F. Claverie-Martin, B. Magasanik, Proc. Natl.
Acad. Sci. USA 89 (1992) 5088^5092.[15] J. Li, S. Kustu, S. Stewart, J. Mol. Biol. 241 (1994) 150^165.[16] W.R. McCleary, Mol. Microbiol. 20 (1996) 1155^1163.[17] J.B. Stock, Science 274 (1996) 370^371.[18] D. Beier, H. Deppisch, R. Gross, Mol. Gen. Genet. 252
(1996) 169^176.[19] V. Scarlato, A. Prugnola, B. Arico, R. Rappuoli, Proc. Natl.
Acad. Sci. USA 87 (1990) 6753^6757.[20] P.E. Boucher, F.D. Menozzi, C. Locht, J. Mol. Biol. 241
(1994) 363^377.[21] F. Baneyx, G. Georgiou, J. Bacteriol. 173 (1991) 2696^
2703.[22] T.N. Belev, M. Singh, J.E.G. McCarthy, Plasmid 26 (1991)
147^150.[23] A.-L. Perraud, B. Kimmel, V. Weiss, R. Gross, Mol. Micro-
biol. 27 (1998) 875^887.[24] D.C. Schriemer, L. Li, Anal. Chem. 68 (1996) 3382^3387.[25] D.C. Schriemer, T. Yalcin, L. Li, Anal. Chem. 70 (1998)
1569^1575.[26] T. Fligge, K. Bruns, M. Przybylski, J. Chromatogr. B Bio-
med. Sci. Appl. 706 (1998) 91^100.[27] M. Bantsche¡, V. Weiss, M.O. Glocker, Biochemistry 38
(1999) 11012^11020.[28] R. Heermann, K. Altendorf, K. Jung, Biochim. Biophys.
Acta 1415 (1998) 114^124.[29] E.R. Stadtman, Methods Enzymol. 3 (1957) 228^231.[30] S. Bouche, E. Klauck, D. Fischer, M. Lucassen, K. Jung, R.
Hengge-Aronis, Mol. Microbiol. 27 (1998) 787^795.[31] U.K. Laemmli, Nature 227 (1970) 680^685.[32] K. Rippe, N. Mu«cke, A. Schulz, J. Mol. Biol. 278 (1998)
915^933.[33] B.J. Shen, T. Hage, W. Sebald, Eur. J. Biochem. 240 (1996)
252^261.[34] S.C. Schuster, R.V. Swanson, L.A. Alex, R.B. Bourret, M.I.
Simon, Nature 365 (1993) 343^347.[35] H. Tanabe, T. Masuda, Y. Yamasaki, A. Katoh, S. Yoshio-
ka, R. Utsumi, Biosci. Biotechnol. Biochem. 62 (1998) 78^82.
[36] M.O. Glocker, C. Borchers, W. Fiedler, D. Suckau, M. Przy-bylski, Bioconjug. Chem. 5 (1994) 583^590.
[37] M. Kalkum, M. Przybylski, M.O. Glocker, Bioconjug.Chem. 9 (1998) 226^235.
[38] I. Baikalov, I. Schro«der, M. Kaczor-Grzeskowiak, K. Grze-skowiak, R.P. Gunsalus, R.E. Dickerson, Biochemistry 35(1996) 11053^11061.
[39] I. Baikalov, I. Schro«der, M. Kaczor-Grzeskowiak, D. Cas-
BBAPRO 36134 11-5-00
A.-L. Perraud et al. / Biochimica et Biophysica Acta 1478 (2000) 341^354 353

cio, R.P. Gunsalus, R.E. Dickerson, Biochemistry 37 (1998)3665^3676.
[40] M.A. Uhl, J.F. Miller, J. Biol. Chem. 271 (1996) 33176^33180.
[41] S.S. Schuster, A.A. Noegel, F. Oehme, G. Gerisch, M.I.Simon, EMBO J. 15 (1996) 3880^3889.
[42] S.L. Fisher, S.K. Kim, B.L. Wanner, C.T. Walsh, Biochem-istry 35 (1996) 4732^4740.
[43] Y. Hidaka, H. Park, M. Inouye, FEBS Lett. 400 (1997) 238^242.
[44] S.Q. Pan, T. Charles, S. Jin, Z.L. Wu, E.W. Nester, Proc.Natl. Acad. Sci. USA 90 (1993) 9939^9943.
BBAPRO 36134 11-5-00
A.-L. Perraud et al. / Biochimica et Biophysica Acta 1478 (2000) 341^354354