dinâmica da vegetação ripária em riachos de mata atlântica ... · diretamente relacionados com...
TRANSCRIPT

UNIVERSIDADE FEDERAL DE SANTA CATARINA
CENTRO DE CIÊNCIAS BIOLÓGICAS
DEPARTAMENTO DE ECOLOGIA E ZOOLOGIA
PROGRAMA DE PÓS-GRADUAÇÃO EM BIOLOGIA VEGETAL
Leonardo Kleba Lisboa
Dinâmica da vegetação ripária em riachos de Mata Atlântica
subtropical: composição da matéria orgânica alóctone e
interação com invertebrados aquáticos
Dissertação apresentada ao Programa de
Pós-Graduação em Biologia Vegetal da
Universidade Federal de Santa Catarina
como parte dos requisitos para obtenção do
título de Mestre em Biologia Vegetal
Orientador: Mauricio Mello Petrucio
Florianópolis
2012

À VIDA
"Toda interpretação feita por um cientista é
uma hipótese, e todas as hipóteses são
tentativas. Elas devem sempre ser testadas e
sempre revisadas quando insatisfatórias.
Portanto, uma mudança na mente de um
cientista, particularmente na mente de um
grande cientista, não é um sinal de fraqueza,
mas sim uma evidência da atenção contínua
para o respectivo problema e da habilidade
de testar a hipótese de novo, e de novo..."
Ernst Mayr
“O que me preocupa não é o grito dos
maus, mas sim o silêncio dos bons.”
Martin Luther King

Agradecimentos
Primeiramente gostaria de agradecer aos meus pais, Armando Lisboa e Teresa
Kleba, pelo incondicional apoio aos estudos, por acreditarem em mim e apoiarem
minhas decisões sempre e por incentivarem a busca pela qualidade do trabalho
acadêmico como professores universitários. Dedico este trabalho a eles!
Agradeço também aos meus irmãos, João e Cristiane, exemplos de caráter e
atitudes no trabalho e na vida, a quem sempre segui e busquei me espelhar como irmão
mais novo. Também aos tios, primos, vós e toda família pelo suporte conferido e
confiança gerada.
Agradeço ao Programa de Pós-Graduação em Biologia Vegetal (PPGBVE) da
Universidade Federal de Santa Catarina, e à Coordenação de Aperfeiçoamento de
Pessoal de Nível Superior (CAPES) pela bolsa de estudos concedida.
Agradeço ao professor Mauricio Mello Petrucio, por ter me aceitado como aluno
de mestrado e acreditado no meu potencial, pelas orientações, suporte fornecido e
trabalho agradável. Também ao professor José Francisco Gonçalves Júnior, da UNB,
pelo apoio, correções pertinentes, agradável recepção em Brasília e ideias para
metodologias experimentais, análises e referências da presente dissertação.
Agradeço à pós-doc Ana Emília e à doutoranda Áurea Lemes pelo
imprescindível apoio nas coletas, nas análises estatísticas, identificação dos
macroinvertebrados e companheirismo. A todos os companheiros do Laboratório de
Ecologia de Águas Continentais que ajudaram nas coletas direta ou indiretamente,
análises de água, e foram essenciais para descontração do ambiente de trabalho: Jéssica,
Denise, Natália, Mônica, Rafael, Guto, Eduardo... Agradeço ao Cássio do PPGBVE
pela identificação das espécies vegetais, auxílio em campo e ensinamentos botânicos.
Agradeço ao LAPAD (UFSC) pelo suporte fornecido essencial para realização
das coletas, através de equipamentos e pessoal de campo, principalmente ao Pedrão,
Mauricio, Ronaldo, Cézinha e Davi. Também ao Parque Municipal da Lagoa do Peri
por aceitar a pesquisa e disponibilizar equipamento e pessoal para realização das
coletas, em nome de Danilo, Mauro, Rafael, Vado, entre outros.
Ao Departamento de Ecologia e Zoologia (ECZ) da UFSC, pelo apoio fornecido
pelos funcionários Carla, Walter, Félix, Rogério entre outros. Também ao pessoal do
DMSG da UFSC pela disponibilização do transporte.

Aos colegas dos programas de Pós-Graduação em Biologia Vegetal e em
Ecologia pelo companheirismo, trocas de ideias e festas frequentadas. À secretária
Verinha pela agilidade e momentos de conversa sobre a família, e ao secretário Roberto.
Agradeço aos amigos de sempre pelos momentos de abstração à vida acadêmica,
das rodas de samba às empurradas de Kombi: Henrique, Pietro, Lucas, Muraro, Vitor,
Mateus, Bernardo, Aita, entre tantos outros...

5
Sumário
Resumo ......................................................................................................................... 6
Abstract ........................................................................................................................ 7
Lista de Tabelas ............................................................................................................ 8
Lista de Figuras............................................................................................................. 9
Introdução Geral ......................................................................................................... 11
Material e Métodos ..................................................................................................... 17
Área de estudo ......................................................................................................... 17
Mata Atlântica ..................................................................................................... 17
Bacia Hidrográfica da Lagoa do Peri .................................................................... 18
Delineamento amostral ............................................................................................ 20
Experimento de dinâmica da vegetação ripária ..................................................... 20
Interação entre a vegetação ripária e invertebrados aquáticos ............................... 22
Análise estatística dos dados .................................................................................... 23
Capítulo 1 ................................................................................................................... 26
Dinâmica e composição da matéria orgânica alóctone em um riacho de Mata
Atlântica subtropical
Capítulo 2 ................................................................................................................... 54
Relação entre a estrutura da comunidade de invertebrados aquáticos com a
composição e biomassa de detritos foliares em riachos preservado e impactado
Considerações Finais................................................................................................... 84
Referências Gerais ...................................................................................................... 86

6
Resumo
O aporte de detritos vegetais alóctones é de suma importância para a dinâmica
energética de riachos florestados de cabeceira, influenciando na estruturação das
comunidades aquáticas. Padrões de aporte e composição desse material estão
diretamente relacionados com o tipo e a integridade da vegetação ripária, e são
influenciados pelo clima e geomorfologia local. Poucos estudos sobre o tema têm sido
feitos em riachos de Mata Atlântica, e são ainda mais raros em ambiente subtropical. O
objetivo da presente dissertação foi avaliar padrões temporais de aporte e composição
da matéria orgânica (MO) vegetal alóctone em um riacho de Floresta Ombrófila Densa
preservada (capítulo 1), e determinar a influência da composição e biomassa dos
detritos foliares do estoque bêntico na estruturação espaço-temporal das comunidades
de invertebrados aquáticos associados em riachos com diferentes graus de integridade
da vegetação ripária (capítulo 2). Foram realizados experimentos entre agosto de 2010 a
agosto de 2011 no riacho Cachoeira Grande (preservado), e até junho de 2011 no riacho
Ribeirão Grande (impactado), ambos localizados na mesma bacia, coberta por Floresta
Ombrófila Densa subtropical. O experimento de dinâmica da vegetação ripária
evidenciou uma participação muito maior do aporte lateral, em detrimento ao vertical,
na incorporação direta de MO ao riacho, superando também os valores do aporte
terrestre e estoque bêntico. Houve grande variação temporal na quantidade e na
composição do detrito vegetal ao longo do ano, influenciada principalmente pela
precipitação e padrões fenológicos. As principais espécies na dinâmica da MO do riacho
foram Schizolobium parahyba, Ficus eximia, Virola bicuhyba, Ficus adhatodifolia,
Cupania vernalis e Tetrorchidium rubrivenium. Para comunidade de invertebrados
aquáticos, a riqueza de espécies do detrito, assim como temperatura da água , biomassa
de galhos e condutividade foram responsáveis pela variação espacial entre os riachos. A
variação temporal na distribuição da comunidade em ambos ocorreu em função da
temperatura da água e precipitação, mas apenas para o preservado a quantidade de
folhas foi importante. A variação no riacho impactado foi mais pronunciada. Os
resultados descrevem um padrão de aporte diferente dos encontrados na literatura, e
demonstram a importância da integridade da vegetação ripária no fornecimento de
subsídios e estabilidade da comunidade de invertebrados aquáticos em riachos de
cabeceira. Assim, se evidencia a importância da integridade das zonas ripárias para os
sistemas aquáticos, enfatizando a necessidade de conservação e recuperação dessas
áreas.
Palavras chave: serapilheira, composição do detrito foliar, mata ciliar

7
Abstract
Input of allochthonous leaf litter is extremely important for energetic dynamics
of forested headwater streams, influencing the aquatic communities’ structures. Patterns
of input and composition of this material are directly related to riparian vegetation type
and integrity, and respond to local climate and geomorphology. Little attention to this
issue has been paid for Atlantic Forest streams, and studies are even rarer in subtropical
environments. The objectives of the present dissertation were to evaluate temporal
patterns of input and composition of vegetable allochthonous organic matter (OM) in a
preserved Atlantic Rain Forest stream (chapter 1), and determine the influence of
benthic leaf litter biomass and composition on aquatic invertebrate communities’
temporal-space structure in streams with different riparian vegetation integrity states
(chapter 2). Experiments were undertaken between August/2010 and August/2011 for
Cachoeira Grande (preserved) stream, and until June/2011 in Ribeirão Grande
(impacted) stream, both situated in the same watershed covered by subtropical Atlantic
Rain Forest vegetation. Riparian vegetation dynamics experiment showed a higher
contribution of lateral input, when compared with vertical input, in direct OM entry into
the stream, exceeding values of terrestrial input and benthic stocks. There was a
temporal variation in quantity and composition of vegetable litter during the year,
controlled mainly by precipitation and phenological factors. The main species of OM
dynamics were Schizolobium parahyba, Ficus eximia, Virola bicuhyba, Ficus
adhatodifolia, Cupania vernalis e Tetrorchidium rubrivenium. For aquatic invertebrate
community, leaf litter species richness, as well as water temperature, branch biomass
and conductivity were responsible for spatial variations between the streams. Temporal
variation in the distributions of communities of both streams occurred in function of
water temperature and precipitation, but just for the preserved one leaf biomass was
important. Variation in the impacted stream was more pronounced. The results describe
an input pattern distinct from those found in literature, and demonstrate the importance
of riparian vegetation integrity on providing subsidies and stability of aquatic
invertebrate community in headwater streams. Thus, the importance of riparian zone
integrity is shown for aquatic systems, which highlights the necessity for conservation
and restoration of these areas.
Key-words: litterfall, leaf litter composition, allochthonous input

8
Lista de Tabelas
Capítulo 1
Tabela 1. Valores de correlação de Pearson entre a biomassa vegetal incorporada das
categorias de aporte e principais espécies com as variáveis ambientais mensuradas no
rio Cachoeira Grande entre ago/10 e ago/11. Valores em vermelho significativos ( p <
0,01) ............................................................................................................................... 35
Tabela 2. Resumo de dados do aporte médio de MO (g.m-2
.mês-1
) vegetal em riachos e
rios de ambientes tropicais e temperados. AV = aporte vertical; AL = aporte lateral; AT
= aporte terrestre; EB = estoque bêntico; % F = porcentagem de folhas. * dados do
presente artigo. **foram feitos experimentos de 48h, o que acaba por não demonstrar a
variabilidade representativa de um mês. ........................................................................ 41
Apêndice. Médias mensais (g.m-2
.mês-1
± erro padrão) do aporte de MO vegetal por
espécie nos compartimentos vertical (n = 90), lateral (n = 10), terrestre (n = 10) assim
como do estoque bêntico (n = 15), mensurados no rio Cachoeira Grande entre agosto de
2010 e agosto de 2011. ............................................................................................. 50-53
Capítulo 2
Tabela 1. Principais famílias de invertebrados aquáticos em valores de abundância
média (Abu méd), percentual de contribuição da abundância total (Contrib %) e
contribuição acumulativa (Cum %) observados entre agosto de 2010 e junho de 2011
nos riachos preservado (Cachoeira Grande) e impactado (Ribeirão Grande). As famílias
foram escolhidas a partir da análise de SIMPER, desconsiderando-se aquelas que juntas
representaram menos de 10% da abundância da comunidade. ...................................... 64
Apêndice. Composição das folhas do estoque bêntico nos riachos preservado (CG) e
impactado (ST) mensuradas de agosto de 2010 a junho de 2011, especificando-se as
médias mensais (g.m-2
) e a porcentagem de cada táxon no total mensurado. * representa
espécies exóticas. ..................................................................................................... 81-83

9
Lista de Figuras
Introdução Geral
Figura 1. Localização geográfica da Bacia Hidrográfica da Lagoa do Peri na cidade de
Florianópolis, SC, Brasil, e seus principais tributários, os rios Cachoeira Grande e
Ribeirão Grande. ............................................................................................................ 20
Figura 2. Desenho esquemático do delineamento amostral realizado no capítulo 1 –
experimento de dinâmica da vegetação ripária – mostrando os coletores verticais,
laterais e terrestres nos 5 pontos amostrais em um trecho de 100 m do riacho. ............ 22
Capítulo 1
Figura 1. Média e erro-padrão da biomassa de MO vegetal dos diferentes
compartimentos para o riacho Cachoeira Grande entre ago/2010 e ago/2011. ............. 36
Figura 2. Percentual de folhas, galhos, frutos e restos vegetais na formação dos
diferentes compartimentos de aporte de detritos para o riacho Cachoeira Grande
medidos entre ago/2010 a ago/2011. ............................................................................. 36
Figura 3. Variação mensal do aporte de biomassa vegetal seca dos diferentes
compartimentos estudados no riacho Cachoeira Grande entre ago/2010 e ago/2011.
Barras representam o erro padrão. ................................................................................. 37
Figura 4. Porcentagem das cinco principais famílias botânicas no aporte de MO vegetal
ao riacho Cachoeira Grande no período de ago/2010 a ago/2011. ................................ 38
Figura 5. Contribuição média e erro padrão da biomassa foliar das 6 principais espécies
vegetais nos 4 compartimentos do riacho Cachoeira Grande entre ago/2010 e ago/2011.
........................................................................................................................................ 38
Figura 6. Médias e erros-padrão das principais espécies do aporte vertical (a), aporte
lateral (b), aporte terrestre (c) e estoque bêntico (d) mensurados entre agosto de 2010 e
agosto de 2011 no riacho Cachoeira Grande. ................................................................ 40
Capítulo 2
Figura 1. Valores médios de precipitação acumulada (PA), pH, oxigênio dissolvido
(OD), condutividade elétrica (CE) e temperaturas (Temp) da água e do ar medidos entre
agosto de 2010 e junho de 2011 nos riachos preservado (CG) e impactado (ST) ......... 63

10
Figura 2. Curva de acumulação de espécies baseada no índice de “MaoTau” utilizando
o número de famílias de invertebrados aquáticos encontradas por amostragens, nos
ambientes preservado (círculos) e impactado (triângulos), amostrados entre agosto de
2010 a junho de 2011 na Bacia Hidrográfica da Lagoa do Peri, SC. Pontilhados
representam intervalo de confiança de 95%. ................................................................. 65
Figura 3. Ordenação de escalonamento multidimensional (MDS) obtida pela
similaridade de Bray Curtis dos dados de abundância da comunidade de invertebrados
aquáticos transformada por log(x+1) dos riachos preservado (CG) e impactado (ST),
coletados entre agosto de 2010 e junho de 2011 na Bacia Hidrográfica da Lagoa do Peri,
SC. ................................................................................................................................. 66
Figura 4. Ordenação de escalonamento multidimensional (MDS) dos dados de
composição específica dos detritos foliares feita através da similaridade de distância
Euclidiana da matriz de presença/ausência de espécies vegetais dos riachos preservado
(CG) e impactado (ST) coletados entre agosto de 2010 e junho de 2011 na Bacia
Hidrográfica da Lagoa do Peri, SC. ............................................................................... 66
Figura 5. Quantidade de detritos vegetais do estoque bêntico representados em
biomassa total de recursos (MO, matéria orgânica), e riqueza de espécies vegetais do
folhiço coletado, observadas entre agosto de 2010 e junho de 2011 nos riachos
preservado (CG) e impactado (ST). Barras quadriculadas representam a quantidade de
biomassa de folhas referentes a espécies exóticas – encontradas apenas no rio ST. ..... 67
Figura 6. Análise de Correspondência Canônica (CCA) entre as matrizes de abundância
da comunidade de invertebrados aquáticos (transformada em log(x)) e das variáveis
ambientais mensuradas entre agosto de 2010 e junho de 2011 nos rios Cachoeira Grande
(CG, preservado, círculos cinzas) e Ribeirão Grande (ST, impactado, círculos brancos).
Triângulos representam a ordenação das famílias de invertebrados. Consideraram-se só
as variáveis ambientais significativas (Monte Carlo, p < 0,05); Condut = condutividade
da água, Temp = temperatura da água (°C), Riqueza = número de espécies vegetais do
detrito, Galhos = biomassa de galhos. O eixo 1 representou 25,7% da variação da
comunidade, e o eixo 2 representou 9,2% dessa variação. Correspondências entre as
variáveis bióticas e abióticas foram significativas (Monte Carlo, p < 0,05) ................. 68
Figura 7. Abundância média mensal das principais famílias de invertebrados aquáticos
nos riachos preservado (CG) e impactado (ST) medidas entre agosto de 2010 e junho de
2011. Para o gráfico foram utilizadas apenas famílias que em algum mês apresentaram
abundância maior que 10 % da comunidade. ................................................................ 69

11
Introdução Geral
Zonas ripárias podem ser entendidas como um mosaico surpreendentemente
diverso de acidentes geográficos, comunidades e ambientes compreendidos dentro de
uma paisagem maior, e funcionam como estrutura básica conceitual a fim de se
compreender a organização, diversidade e dinâmica das comunidades associadas aos
sistemas fluviais (Naiman & Décamps, 1997). Estendendo esse conceito no tempo e
espaço, essas zonas podem ser vistas em termos de padrões espaço-temporais levando
em consideração a relação entre processos hidrológicos e geomorfológicos, sucessão de
plantas terrestres e processos ecossistêmicos aquáticos (Gregory et al., 1991).
Especificamente, essas áreas estão localizadas ao longo de rios e no entrono de
nascentes, lagos e reservatórios (Rodrigues & Nave, 2000). O delineamento preciso de
uma zona ripária se mostra muitas vezes difícil de ser determinado. Segundo Naiman &
Décamps (1997), sua área abrange o canal do riacho entre o nível mais alto e o mais
baixo de água, e aquela porção da paisagem terrestre que vai desde a marca de água
mais alta até onde a vegetação é influenciada por cheias e o solo tem alta capacidade de
reter água. Já segundo Gregory et al. (1991), a zona ripária se estende horizontalmente
além dos limites das cheias, e verticalmente compreendendo o dossel da vegetação
adjacente.
As formações florestais localizadas nestas áreas recebem diversas
denominações: vegetação ripária, mata ciliar, mata de galeria, florestas ribeirinhas, entre
muitas outras como bem discutido por Rodrigues (2009). A despeito de denominações
generalistas, fica claro que essas formações não se constituem como um tipo
vegetacional único, pois mesmo possuindo estrutura e função ecossistêmicas similares,
elas diferem fundamentalmente em composição taxonômica conforme o domínio
morfoclimático e fitogeográfico do País (Ab’Sáber, 2009). A vegetação ripária ocupa
uma das áreas mais dinâmicas de uma paisagem, sendo que a distribuição e composição
dessa comunidade refletem tanto influências do ambiente fluvial na forma de cheias e
vazantes, quanto influências das áreas terrestres adjacentes como fogo, vento, doenças e
herbivoria (Gregory et al., 1991). Além disso, essas formações vegetais apresentam um
elevado grau de complexidade estrutural e heterogeneidade ambiental devido
principalmente às variações edáficas, topográficas e às características hidrológicas da
bacia e do curso d’água (Rodrigues & Shepherd, 2009).

12
As florestas ripárias atuam na manutenção da integridade de uma bacia
hidrográfica pela ação direta que têm em uma série de processos importantes para
estabilidade da microbacia, para manutenção da qualidade e quantidade de água
presente, assim como para manutenção da saúde ecológica dos próprios ecossistemas
aquáticos (Lima & Zakia, 2009). Elas também são chamadas de zona tampão,
justamente pelo fato de amortecerem os impactos provenientes das áreas que circundam
o ecossistema aquático (Ferraz, 2001), representando um importante link de transição
aquático-terrestre (Wallace et a.l, 1997; Naiman & Décamps, 1997). Além disso, esses
ambientes têm grande importância ambiental e social, sendo inclusive considerados
áreas de preservação permanente (APPs) pela legislação ambiental brasileira segundo o
Código Florestal (Lei n° 4777/65), que limita sua ocupação e proíbe sua supressão.
Dentre as muitas funções ecológicas importantes, está o funcionamento da vegetação
ripária como corredores ecológicos para fauna e flora silvestres, sua atuação como filtro
natural de sedimento e água, no controle de erosão das margens, da incidência luminosa
e um papel chave na ciclagem de nutrientes de riachos florestais (Gregory et al., 1991;
Naiman & Décamps, 1997; Lima & Zakia, 2009).
Esse papel chave da vegetação ripária na ciclagem de nutrientes está diretamente
relacionado com a dinâmica de energia dos ecossistemas aquáticos, principalmente de
rios sombreados de pequenas ordens, que são sistemas predominantemente
heterotróficos (Cummins et al, 1973; Vannote et al., 1980). Estudos vêm demonstrando
que riachos de cabeceira têm o suprimento de energia dependente da incorporação da
matéria orgânica (MO) produzida nos sistemas terrestres adjacentes, onde muitos dos
organismos aquáticos utilizam essa MO vegetal (Cummins & Klug, 1979; Vannote et
al., 1980; Naiman & Décamps, 1997; Wallace et al., 1997; Graça, 2001; Tank et al.,
2010), e inclusive animal (Kawaguchi et al., 2003) como recurso nutricional. Segundo
Wallace et al. (1997), a supressão deste aporte alóctone pode afetar diretamente a teia
dos detritívoros reduzindo drasticamente a comunidade de invertebrados aquáticos de
um rio.
Vannote et al. (1980) observaram que as comunidades de produtores e
consumidores características de um determinado trecho de rio são estabelecidas em
harmonia com a dinâmica das condições físicas do canal. Os autores evidenciaram a
importância da geomorfologia do rio no estabelecimento da relação produção/respiração
dentro d’água, e propuseram o “Conceito do Rio Contínuo” (RCC). Esse conceito
propõe que, de acordo com a ordem do rio, se tem uma mudança na importância do

13
aporte vegetal alóctone para o suprimento energético do sistema, e consequentemente há
uma mudança conjunta na predominância dos grupos tróficos das comunidades
presentes, sendo que o aumento da ordem do rio coincide com a diminuição da
importância do aporte de MO terrestre.
Na proposta do RCC, riachos de cabeceira e sua vegetação ripária associada
influenciam fortemente o fluxo de materiais da nascente para o ecossistema à jusante,
onde o material vegetal produzido nas cabeceiras afeta as cadeias alimentares de
diversos organismos ao longo do curso do rio e nas regiões de borda (Vannote et al.,
1980; Begon et al., 2007; Binckley et al., 2010). Apesar da grande aceitação do RCC,
existem outras propostas de funcionamento da dinâmica de ecossistemas lóticos que
levam em conta especificidades hidro-geológicas e climáticas, como o Conceito da
Série Descontínua, o Conceito do Pulso de Inundações e o Conceito do Corredor
Hiporreico (Poole, 2002). A ecologia de paisagem também tem mostrado a importância
em se considerar um rio como um mosaico de paisagens e manchas heterogêneas, onde
padrões espaciais locais assim como padrões temporais influenciam os processos
ecológicos, enfatizando a importância de se considerar o contexto, geomorfologia,
estrutura e metaestrutura na determinação da dinâmica e funções de um ecossistema
lótico (Poole, 2002; Wiens, 2002).
A MO de ambientes lóticos provém de duas principais fontes: (1) autóctone,
produzida pelos organismos fotossintetizantes dentro do próprio rio; e (2) alóctone, de
origem terrestre proveniente da vegetação do entorno (Dobson & Frid, 1998). Ecólogos
de rios têm classificado a MO através de seu tamanho em: particulada grossa (MOPG;
>1 mm), particulada fina (MOPF; 0,45 µm - 1 mm) e matéria orgânica dissolvida
(MOD, <0,45 µm) (Tank et al., 2010). Evidências sugerem que a MOD, representada na
forma de carbono orgânico dissolvido (COD), é um grande estoque de matéria orgânica
nos ecossistemas de águas correntes, e provém predominantemente dos solos e de folhas
da vegetação ripária (Tank et al., 2010).
A MOPG alóctone, por sua vez pode atingir os cursos de água através de
movimentos verticais, que seria a queda da serapilheira diretamente sobre o curso de
água, ou movimentos laterais, que consiste no carreamento da MO depositada no
entorno para dentro do rio (Benfield, 1997; Webster & Meyer, 1997). Dados na
literatura indicam uma participação muito maior (80-90%) do aporte vertical quando
comparado com o lateral (Conners & Naiman, 1984; Benson & Person, 1993; Benfield,
1997). Porém, há estudos em sistemas Neotropicais controversos, demonstrando não

14
haver diferença entre as duas fontes (Gonçalves et al., 2006), ou mesmo que o aporte
lateral é maior (França et al., 2009). É válido considerar também, que a maioria dos
estudos sobre o aporte da serapilheira em rios não consideram o aporte lateral (Abelho,
2001). A MOPG pode conter folhas, flores, sementes, galhos, cascas, frutos e outras
partes vegetais (Beinfield, 1997), sendo constituída principalmente por folhas em ambos
ambientes temperados e tropicais, que podem representar de 41 a 98% da MO presente
(Abelho & Graça, 1998; Benson, & Pearson, 1993; França et al., 2009).
Um fator determinante da importância e quantidade dessas influências é o
padrão do fluxo e vazão da água, onde os períodos de vazão máxima e consequentes
inundações têm o potencial de carrear uma grande quantidade de nutrientes para dentro
do ecossistema (Townsend, 1996; Acuña et al., 2007). A estrutura da vegetação ripária e
o grau de preservação da mesma também são fatores determinantes deste processo.
Comparando o aporte de MO proveniente da zona ripária entre um rio de vegetação
preservada com outro de vegetação em regeneração, por exemplo, Carvalho & Uieda
(2010) encontraram uma quantidade do aporte lateral 10 vezes maior no rio preservado.
Outra problemática é a substituição de espécies nativas por exóticas, que pode afetar a
qualidade dos detritos que entram no ecossistema aquático assim como o ciclo de
nutrientes e a estrutura da comunidade do rio (Alonso et al., 2010). Comparando
plantações de eucalipto com florestas nativas em zonas ripárias na Espanha, Pozo et al.
(1997) observaram uma drástica redução na qualidade do aporte de MO aos rios com
vegetação da espécie exótica, com uma redução de 67% do conteúdo de nitrogênio e
65% do conteúdo de fósforo nas folhas incorporadas ao ambiente aquático.
A queda de serapilheira tem forte influência sazonal em ambientes temperados,
onde o grande aporte desse material se dá principalmente no outono, que marca a época
característica de abscisão foliar (Benfield, 1997; Pozo et al., 1997). A queda de
serapilheira em ambientes tropicais pode também ser sazonal, principalmente quando
existe a presença de estações bem definidas. No Cerrado, por exemplo, o maior aporte
de MO vegetal se dá justamente no fim da estação seca (Gonçalves et al., 2006;
Carvalho & Uieda, 2010). Diferentemente, Rezende & Mazzoni (2005) encontraram o
pico de aporte de serapilheira na estação úmida trabalhando em um rio de Mata
Atlântica. Como esclarece Abelho (2001), é provável que o clima regional associado às
características decíduas da floresta sejam os fatores determinantes desse padrão sazonal
de queda de serapilheira.

15
A composição do material alóctone dentro do rio afeta diretamente a estrutura da
comunidade e a dieta da fauna aquática, e consequentemente, o ecossistema aquático
como um todo (Carvalho & Uieda, 2010). Dentro do ecossistema aquático as folhas são
decompostas por uma combinação de fatores físicos, químicos e biológicos, incluindo
as etapas de lixiviação, que seria a dissolução de compostos solúveis em água, o
condicionamento, que é representado pela colonização por micro-organismos, e a
fragmentação pela abrasão física e por invertebrados (Cummins, 1974; Kirby et al.;
1983; Gessner et al., 1999).
Estudos na literatura também diferem sobre o valor de taxas de decomposição da
MO entre rios temperados e tropicais, e a importância da participação dos
macroinvertebrados aquáticos nesse processo, sugerindo que invertebrados
fragmentadores tendem a ser menos importantes em rios tropicais por causa da
colonização e processamento mais rápidos dos microorganismos devido às temperaturas
mais altas (Abelho, 2001; Gonçalves et al., 2004; Moretti et al., 2007; Carvalho &
Uieda, 2010). Avaliando a colonização de detritos foliares por macroinvertebrados
dentro de rios tropicais, Moretti et al. (2007) encontraram poucos fragmentadores, e
Gonçalves et al. (2007) assim como Rezende et al. (2010) não encontraram sequer
organismos desse grupo, concluindo que a decomposição se dá principalmente por
processos microbiológicos e físicos. Em contrapartida, Landeiro et al. (2010)
observaram uma significativa participação de invertebrados fragmentadores no processo
de decomposição de folhas em um rio tropical na Floresta Amazônica, o que pode
indicar uma diferença nos padrões deste processo entre os biomas Cerrado e Floresta
Amazônica. Para Mata Atlântica, o componente microbiano é provavelmente o mais
importante na decomposição (Moulton & Magalhães, 2003), e a participação de macro-
consumidores onívoros, como camarões e caranguejos, parece ser mais importante que
insetos e outros grupos de fragmentadores especialistas nesse processo (Moulton et al.,
2010).
É importante lembrar que a composição química e fatores físicos são variáveis
intrinsecamente ligadas à taxa de decaimento da massa das folhas e seu consumo por
detritívoros, e consequentemente à colonização pela comunidade de invertebrados
aquáticos. Particularmente a composição de nutrientes como o nitrogênio, compostos
estruturais como lignina e celulose, e as defesas químicas como os polifenóis, assim
como fatores físicos como o revestimento por cutícula e a dureza, são os fatores internos
determinantes da taxa de decomposição (Graça & Zimmer, 2005).

16
O processo de decomposição da MO alóctone, além de liberar nutrientes e MOD
para o ambiente, também produz CO2 e CH4, gases causadores do efeito estufa (Prast &
Pinho, 2008). É importante ressaltar que os ecossistemas de água doce representam um
componente relevante no balanço continental desses gases (Batsviken et al., 2011) e,
apesar da pequena área que ocupam, podem afetar o balanço regional do carbono em
uma bacia (Cole et al., 2007). Por serem subsidiados pelo aporte alóctone terrestre,
ecossistemas aquáticos continentais funcionam tanto como fontes de emissão quanto
como sumidouros de CO2 atmosférico (Cole et al., 2007). De acordo com Cole et al.
(2007), de todo aporte de carbono que as águas interiores recebem, 40% retorna para
atmosfera como CO2, 12% é sequestrado nos sedimentos, e por volta dos 48% restantes
são transportados para os oceanos.
Está claro que diferentes biomas podem ter padrões de comportamento e
dinâmica das formações florestais muito distintos em virtude de condições
geomorfológicas e climáticas específicas. Segundo Naiman & Décamps (1997),
Webster & Meyer (1997) e Tank et al. (2010), a estrutura e a composição da vegetação
ripária parecem ser os principais fatores determinantes da entrada de material alóctone
em riachos, onde a fenologia particular de cada espécie ripária irá determinar
ultimamente os padrões de input de MO ao rio. Nesse sentido, Binckley et al. (2010)
observaram que o modo como a vegetação ripária é manejada influencia diretamente o
sedimento, quantidade de detritos e a quantidade de presas disponíveis aos
consumidores de topo em um rio, alertando que previsões das consequências de um
manejo dessa vegetação requerem um conhecimento de ambos fatores locais e
regionais, e enfatizando a importância da ecorregião e do tempo na dinâmica dos
nutrientes no sistema aquático. Como enfatizam Carvalho & Uieda (2010), mudanças na
composição da vegetação ripária afetam a estrutura e os processos nos rios na medida
em que se altera a qualidade e a quantidade dos detritos que são incorporados na cadeia
alimentar.
Dentro disso, estudos específicos por região fitogeográfica se fazem necessários
para determinarmos padrões de dinâmica da vegetação ripária a nível inter-regional,
avaliarmos espécies-chave da vegetação na manutenção da integridade dos ecossistemas
e fornecermos subsídios para conservação e restauração das cadeias tróficas de sistemas
fluviais, assim como para cálculos de balanço do carbono incluindo a produção alóctone
nos sistemas hídricos continentais. Sendo assim, o objetivo da presente dissertação foi
avaliar o padrão temporal de aporte e a composição da MO vegetal alóctone em um

17
riacho de Floresta Ombrófila Densa preservada (capítulo 1), e determinar a influência
da composição e biomassa dos detritos foliares do estoque bêntico na estruturação
espaço-temporal das comunidades de invertebrados aquáticos associados a riachos com
diferentes graus de integridade da vegetação ripária (capítulo 2).
Material e Métodos
Área de estudo
Mata Atlântica
A Mata Atlântica é a floresta mais rica do mundo em diversidade de árvores, tem
proporcionalmente maior diversidade biológica quando comparada com a Floresta
Amazônica, e representa o segundo bioma com o maior risco de extinção do mundo
(Campanili & Prochnow, 2006). Hoje ela se encontra reduzida a apenas 7,2% de sua
área original, e foi considerada pela International Union for Conservation of Nature
(IUCN) como um hot-spot de biodiversidade mundial por ter sua grande diversidade
biológica e alto grau de endemismo sob perigo imediato de extinções de espécies e
destruição de habitats (Primack & Rodrigues, 2001).
Inserida no domínio da Mata Atlântica está a formação da Floresta Ombrófila
Densa, ou Mata Pluvial da Encosta Atlântica. Segundo Klein (1984), no sul do Brasil
essa formação se estende desde o Vale do Ribeira (SP) até a Lagoa dos Barros (RS),
ocupando planícies quaternárias do pleistoceno e as encostas orientais da Serra do Mar e
ramificações da Serra Geral. De acordo com Campanili & Prochnow (2006) a Floresta
Oombrófila Densa tem característica perenifólia, com dossel de até 15 m e árvores
emergentes até 40 m, densa vegetação arbustiva de samambaias arborescentes,
bromélias, palmeiras e abundantes trepadeiras. Na região sul predominam espécies das
famílias das mirtáceas, leguminosas, rutáceas, lauráceas, meliáceas, apocináceas,
palmeiras, entre outras (Wettstein, 1970).
No sul do Brasil, grande parte da floresta é dominada por Ocotea catharinensis
Mez (canela preta), o que gera uma uniformidade fitofisionômica não igualada por
nenhuma outra floresta latifoliada brasileira (Klein, 1980). Outras arbóreas frequentes
são Sloanea guianensis (Aubl.) Benth. (laranjeira-do-mato), Copaifera trapezifolia

18
Hayne (pau-óleo), Cryptocarya aschersoniana Mez (canela-fogo ou canela-pururuca),
Aspidosperma olivaceum Müll.Arg. (peroba-vermelha), Guapira opposita (Vell.) Reitz
(maria-mole), Matayba guianensis Aubl. (camboatá-branco), Ocotea odorifera (Vell.)
Rohwer (canela-sassafrás), Virola bicuhyba (Schott ex Spreng.) Warb. (bicuíba),
Hieronyma alchorneoides Allemão (licurana), Calyptranthes strigipes O.Berg
(guamirim-chorão), Inga sellowiana Benth. (ingá-mirim), Marlierea silvatica (O.Berg)
Kiaersk. (guamirim-chorão), Myrcia pubipetala Miq. (guamirim-araçá), Buchenavia
kleinii Exell (garajuva) entre outras (Klein, 1984).
No estrato médio há uma grande dominância de Euterpe edulis Mart.
(palmiteiro), onde também se encontram Garcinia gardneriana (Planch. & Triana)
Zappi (bacupari), Marlierea eugeniopsoides (Kausel & D.Legrand) D.Legrand
(guamirim-branco), Ocotea teleiandra (Meisn.) Mez (canela-pimenta), Sorocea
bonplandii (Baill.) W.C.Burger et al. (soroca), Actinostemon concolor (Spreng.)
Müll.Arg. (pau-rainha), Maytenus alaternoides Reissek. (coração-de-bugre) e Pera
glabrata (Schott) Poepp. ex Baill. (seca-ligeiro) (Klein, 1984).
Bacia Hidrográfica da Lagoa do Peri
O município de Florianópolis está localizado em zona subtropical, e, segundo a
classificação climática de Köeppen, tem clima do tipo Cfa, que representa a ausência de
estação seca, com chuvas distribuídas uniformemente durante o ano e verões rigorosos
(Nascimento, 2002). A vegetação da Ilha de Santa Catarina pode ser apresentada sob
três aspectos distintos: Áreas de Formação Pioneiras, de constituição herbáceo-
arbustiva, raramente arbórea e solos pedologicamente instáveis; Vegetação Secundária,
caracterizada por sucessivos estágios de regeneração que tendem a reconstituir a
vegetação original, ocorrendo após devastação ou abandono do terreno cultivado; e
Floresta Primária, restringida apenas a pequenos focos no alto das encostas do morros
do Ribeirão e da Costa da Lagoa (Neto & Klein, 1991).
A Bacia Hidrográfica da Lagoa do Peri possui 20,3 Km2 (CECCA, 1996), e tem
os mesmos limites do Parque Municipal da Lagoa do Peri, criado pela Lei Municipal n°
1.828/81, situado ao sul da parte insular de Florianópolis (Prefeitura Municipal de
Florianópolis, 2009) (Figura 1). A região é de extrema importância para a cidade na
medida em que na Lagoa do Peri funciona um sistema de captação de água com vazão

19
média captada de 200 l/s (CASAN, 2009), que abastece cinco distritos do município:
Pântano do Sul, Campeche, Ribeirão da Ilha, Lagoa da Conceição e Barra da Lagoa
(GGE-UFSC, 2006).
Aproximadamente 75% da bacia é constituída pelo complexo cristalino pré-
cambriano, com relevo acidentado representado na forma de altos morros como o Morro
da Chapada (440 m), da Tapera (371 m), da Boa Vista (350 m) e do Peri (320 m)
(Santos et al., 1989). A maior parte desses terrenos cristalinos é coberta por vegetação
de Floresta Ombrófila Densa em estádios de regeneração avançado. Segundo Caruso
(1990), as regiões noroeste e sudeste da Lagoa do Peri possuem um dos estádios mais
desenvolvidos de vegetação secundária na Ilha de Santa Catarina, e algumas dessas
áreas apresentam aspecto fisionômico muito semelhante à floresta primária.
A parte leste da bacia é constituída geologicamente por terrenos de sedimentação
quaternária, que separam a lagoa do Oceano Atlântico através de uma estreita faixa de
250 m de largura (Santos et al., 1989). Essa região é coberta principalmente por
vegetação litorânea característica de restinga, com algumas áreas ocupadas por casas e
pequenos trechos de plantação de Eucalipto.
Dentre os dois principais tributários da bacia, o rio Cachoeira Grande drena uma
área de 1,66 km2, tem suas nascentes situadas na cota de 280 m de altitude e percorre
1,17 km até desaguar na Lagoa do Peri. Já o rio Ribeirão Grande drena uma área de 6,98
km2, tem suas nascentes por volta de 285 m de altitude e percorre 4,6 km até desaguar
na lagoa (Santos et al., 1989) (Figura 1).

20
Figura 1. Localização geográfica da Bacia Hidrográfica da Lagoa do Peri na cidade de
Florianópolis, SC, Brasil, e seus principais tributários, os rios Cachoeira Grande e Ribeirão
Grande.
Delineamento amostral
A presente dissertação foi subdividida em dois capítulos que representaram
experimentos distintos, porém complementares. Para o primeiro capítulo, foi realizado
um experimento sobre dinâmica da vegetação ripária a fim de se estimar o padrão
temporal e a composição do aporte de detritos vegetais alóctones no rio Cachoeira
Grande ao longo de um ano. Já no segundo capítulo, foi analisada a relação da biomassa
e riqueza do estoque bêntico de detritos vegetais depositados no leito dos rios Cachoeira
Grande e Ribeirão Grande com a variação espacial e temporal da estrutura das
comunidades de invertebrados aquáticos dos dois ambientes durante onze meses.
Experimento de dinâmica da vegetação ripária
Para avaliar a dinâmica de incorporação do material vegetal alóctone no
ecossistema lótico, o experimento foi realizado em um trecho de 100 m do rio
Ribeirão Grande
Cachoeira Grande

21
Cachoeira Grande, representando um sistema lótico de 3ª ordem, com densa cobertura
de dossel e vegetação ripária em estádio médio–avançado de regeneração. Esse trecho
do rio foi dividido em cinco pontos amostrais separados a uma distância média de 20 m.
Em cada ponto amostral a quantificação do aporte de MO vegetal alóctone foi realizada
em quatro frentes distintas, que correspondem aos quatro compartimentos diferentes:
Aporte vertical: para mensuração da MO vegetal alóctone que cai diretamente
sobre o curso de água, três fileiras foram penduradas sobre o rio a uma altura de 2 m da
lâmina de água. Cada fileira continha seis coletores, representados por baldes de 26 cm
de diâmetro com pequenos furos no fundo para impedir o acúmulo de água.
Aporte lateral: o aporte de MO que escoa do solo para o riacho foi mensurado
através de coletores com 0,5 m de comprimento e 0,2 m de altura (0,1 m2), com
profundidade de 30 cm e abertura de malha de 5 mm. Os coletores foram armados no
limite entre a água e o solo da margem do rio, de modo a capturar toda MO que se
movimenta do solo para água, um na margem direita e outro na margem esquerda de
cada ponto.
Aporte terrestre: considerado como um estoque potencial de MO que pode
chegar ao leito do rio, o aporte terrestre foi mensurado através de coletores de 1 m2
armados dentro da vegetação ripária, com profundidade de 10 cm e 5 mm de malha. Em
cada ponto de coleta foi armado um coletor na margem direita e outro na margem
esquerda do rio a uma distância de 10 m do curso d’água.
Estoque bêntico: a MO vegetal depositada no leito do rio foi mensurada com
auxílio de um coletor tipo Surber, tamanho 0,3 x 0,3 m e abertura de malha de 0,25 mm.
Em cada ponto de coleta foram realizadas aleatoriamente três amostras feitas em regiões
do rio onde se encontrava depósito de folhiço.
Coletas foram feitas a cada trinta dias durante treze meses, compreendendo o
período de agosto de 2010 a agosto de 2011. Todo material coletado foi seco em estufa
a 60ºC até a estabilização do peso, separado qualitativamente em folhas, galhos, partes
reprodutivas e miscelânea, e pesado em balança analítica para determinação do peso
seco. As folhas foram identificadas até o menor grupo taxonômico possível com auxílio
de bibliografia adequada (Sobral et al., 2006), auxílio de especialista e através de
comparações morfológicas com exsicatas previamente identificadas provenientes de
outro projeto que fez a fitossociologia da área de estudo. Pluviosidade, intensidade e
direção de ventos e temperatura média do ar de cada mês estudado foram obtidas na

22
estação meteorológica da Empresa de Pesquisa Agropecuária e Extensão Rural de Santa
Catarina (EPAGRI).
Figura 2. Desenho esquemático do delineamento amostral realizado no capítulo 1 –
experimento de dinâmica da vegetação ripária – mostrando os coletores verticais, laterais e terrestres dispostos nos 5 pontos amostrais em um trecho de 100 m do riacho.
Interação entre a vegetação ripária e invertebrados aquáticos
O experimento foi realizado nos dois principais tributários da Bacia Hidrográfica
da Lagoa do Peri. Os riachos foram classificados como “preservado” e “impactado” de
acordo com o grau de preservação da vegetação ripária. O rio Cachoeira Grande
representou o ambiente preservado por se encontrar em local de ótimo estado de
conservação, com densa vegetação ripária nativa e forte cobertura do dossel. Já o rio
Ribeirão Grande foi considerado como ambiente impactado, pois atravessa áreas de
pequenas propriedades rurais, e teve parte de sua vegetação ripária transformada em
pasto e em plantação de espécies exóticas, havendo também a presença de algumas
espécies nativas esparsas ao longo do curso de água.
Foram escolhidos cinco pontos de coleta em cada riacho, separados por uma
distância de 20 m entre si. Os detritos vegetais depositados no leito do riacho (estoque

23
bêntico) foram coletados com auxílio de um amostrador de Surber, tamanho 0,3 x 0,3 m
e abertura de malha de 0,25 mm. Em cada ponto de coleta foram realizadas três
amostragens onde se encontrava depósito de folhiço, totalizando 15 amostras mensais
por riacho. As coletas foram realizadas ao longo de onze meses, compreendendo o
período de agosto de 2010 a junho de 2011.
Todo material coletado foi levado a laboratório e lavado em peneiras de 0,25
mm para retenção de macroinvertebrados aquáticos. Após a lavagem, o material vegetal
foi seco em estufa a 60ºC até a estabilização do peso, e então determinado o peso seco
através de balança analítica de precisão 0,01 g. Os detritos vegetais foram separados
qualitativamente entre folhas, galhos, partes reprodutivas e miscelânea. As folhas foram
identificadas até o menor grupo taxonômico possível com bibliografia adequada (Sobral
et al., 2006) e com auxílio de observações externas realizadas por especialista. Os
animais retidos na peneira foram triados, acondicionados em frascos com álcool 70% e
posteriormente identificados até o menor nível taxonômico possível utilizando-se
chaves de identificação disponíveis (Fernández & Domínguez, 2001; Mugnai et al.,
2010) e auxílio de especialistas.
Simultaneamente às coletas de estoque bêntico nos riachos, foram medidas in
situ variáveis ambientais da água como oxigênio dissolvido (mg.l-1
), pH, temperatura
(°C) e condutividade (μS.cm-1
) com a sonda específica WTW-Multi350i. A velocidade
da água (m.s-1
) foi obtida através do método do flutuador, com cinco réplicas por ponto,
a profundidade (m) medida com metro em 10 réplicas por ponto, e a temperatura do ar
medida com termômetro. Dados de precipitação referentes ao município foram
fornecidos pela Empresa de Pesquisa Agropecuária e Extensão Rural de Santa Catarina
(EPAGRI).
Análise estatística dos dados
Para o primeiro capítulo, cálculos dos percentuais da participação qualitativa do
aporte e da contribuição por compartimento foram feitos a partir da matriz de dados
brutos com o total de peso seco (g) de MO vegetal observado no período de estudo. O
percentual da contribuição específica dos táxons e das famílias foi calculado a partir do
peso seco total das folhas. A partir da matriz de dados brutos foi feita a padronização da
área de cada coletor para 1 m2, assim como a padronização de todos os meses coletados

24
para períodos de 30 dias. Com a matriz de dados padronizada, calculou-se a média
mensal em gramas de peso seco (g.m-2
.mês-1
) dos diferentes compartimentos, assim
como das espécies e táxons. A fim de se verificarem variações na quantidade de
biomassa vegetal aportada entre os pontos amostrais, entre os compartimentos de
aporte, assim como entre os meses, foram realizadas ANOVAs one-way a partir da
matriz de biomassa padronizada (g.m-2
.mês-1
). Para verificar a influência das variáveis
ambientais no padrão de aporte vegetal ao rio, foi feita uma análise de correlação de
Pearson com a matriz padronizada de biomassa e as relativas médias mensais das
variáveis mensuradas. As análises estatísticas foram realizadas com auxílio do programa
Statistica 7.
Já para o segundo capítulo, variações dos parâmetros ambientais entre os pontos
amostrais e entre os meses foram verificadas a partir do teste de Kruskal-Wallis, feito
com a matriz bruta de amostragem de dados para cada rio em separado. A fim de se
verificar variações das variáveis da água entre os dois ambientes estudados foi realizado
o teste de Mann-Whitney, também com a matriz de dados abióticos brutos.
A riqueza da comunidade aquática e o esforço amostral foram avaliados através
da curva de acumulação de espécies referente a todo período de estudo, calculada a
partir da matriz bruta de abundância de indivíduos com auxílio do programa EstimateS
8.2.0, e plotada utilizando-se o método de Mao Tau. Variações espaciais na estrutura da
comunidade entre os dois rios foram verificadas através da análise de escalonamento
multidimensional (MDS), obtida a partir da matriz de similaridade de Bray Curtis dos
dados médios de abundância de todo período transformados em log (x+1).
Similaridades foram testadas estatisticamente através de uma análise de similaridade
(ANOSIM), tendo como fatores as duas áreas. Quando diferenças significativas foram
detectadas, a análise de porcentagem de similaridade (SIMPER) foi utilizada para
identificar os táxons responsáveis pela diferença observada nas comunidades (Clarke &
Warwick, 2001). A análise SIMPER não é um teste estatístico, mas uma análise
exploratória que gera um ranking de porcentagem de contribuição dos táxons mais
importantes para determinar a diferença entre os grupos de amostras, i.e., neste caso,
riachos com diferente complexidade da vegetação ripária (Clarke & Warwick, 2001).
A variação espacial da composição dos detritos foliares foi analisada através de
um MDS, calculado a partir das matrizes de distância Euclidiana dos dados de presença
e ausência das espécies vegetais dos dois riachos. Similaridades entre os riachos foram
testadas através de uma ANOSIM, tendo como fatores as duas áreas.

25
Para verificar a influência das variáveis ambientais na variação espacial da
comunidade, foi feita uma Análise de Correspondência Canônica (CCA), calculada
através da matriz de médias mensais de abundância da comunidade, transformada em
log (x+1), e da matriz de variáveis ambientais, analisando-se conjuntamente os dois
ambientes estudados. As variáveis foram incluídas na análise através de seleção manual,
excluindo-se aquelas não significativas (Monte Carlo, p > 0,05) na análise, e
representando-se as amostragens por ambiente e por mês de coleta. A significância foi
verificada com o teste de Monte Carlo (p < 0,05). A CCA foi feita com auxílio do
programa Canoco for Windows.
Variações temporais da comunidade e da composição dos detritos foliares foram
analisadas em separado para cada riacho, e testadas a partir de ANOSIMs utilizando-se
as mesmas matrizes do MDS, e considerando como fatores os meses estudados. A fim
de se evidenciar os principais fatores ambientais responsáveis pela variação temporal da
comunidade, foi feita uma análise de BIOENV para cada riacho em separado
relacionando-se a matriz de similaridade de Bray Curtis das médias de abundância
transformada (log x + 1), com a matriz de distância Euclidiana dos dados abióticos
transformados (log x + 1) e padronizados. Para análises multivariadas, além dos
parâmetros físico-químicos da água e variáveis meteorológicas, foram considerados
fatores ambientais as categorias do estoque bêntico vegetal como “folhas”, “galhos”,
“reprodutivos”, “miscelânea” e “total de matéria orgânica” medidas em biomassa média
(g), e “riqueza” de espécies na composição do detrito. Todas as análises de MDS,
ANOSIM, SIMPER e BIOENV foram feitas com auxilio do programa estatístico Primer
6.

26
Capítulo 1
Dinâmica e composição da matéria orgânica alóctone em um riacho de
Mata Atlântica subtropical
Composition and dynamics of allochthonous organic matter in a
subtropical Atlantic Rain Forest stream
Leonardo Kleba Lisboa1,3
, Aurea Luiza Lemes da Silva2,4
, Ana Emilia Siegloch3,
José Francisco Gonçalves Júnior2 e Mauricio Mello Petrucio
1,3
1Programa de Pós-graduação em Biologia Vegetal, Universidade Federal de
Santa Catarina - Trindade, Florianópolis, CEP 88010-970, Santa Catarina, Brasil
E-mail: [email protected]
2Laboratório de Limnologia, Departamento de Ecologia, Universidade de
Brasília, Darcy Ribeiro, Brasília- DF - CEP 70910-900. E-mail: [email protected]
3Laboratório de Ecologia de Águas Continentais, Universidade Federal de Santa
Catarina - Trindade, Florianópolis, CEP 88010-970, Santa Catarina, Brasil
E-mail: [email protected]
4Programa de Pós-Graduação em Ecologia, Universidade de Brasília, Darcy
Ribeiro, Brasília – DF – CEP 70910-900

27
Resumo
O aporte de matéria orgânica (MO) vegetal alóctone tem grande importância no
fluxo de energia de riachos de cabeceira, e a maneira com que esse material entra no
ecossistema têm sido bastante estudada, particularmente em ambientes temperados.
Estudos desse tipo em Mata Atlântica subtropical são raros. O presente estudo teve
como objetivo quantificar e qualificar o aporte de MO da vegetação ripária em um
riacho de 3ª ordem de Mata Atlântica subtropical. O estudo se baseou nas hipóteses de
que (1) o aporte vertical é maior que o lateral, e (2) não há variação temporal no aporte
de detritos do riacho. Os aportes vertical, lateral, terrestre e o estoque bêntico foram
mensurados a cada 30 dias durante treze meses. O material coletado foi qualificado em
folhas, galhos, partes reprodutivas, e restos, e as folhas identificadas. O aporte lateral foi
o que mais contribuiu com a incorporação direta de MO ao riacho, com média de 175,5
g.m-2
.mês-1
, contra apenas 32,4 g.m-2
.mês-1
observado no aporte vertical. O estoque
bêntico superou em termos de biomassa o aporte do estoque potencial terrestre. As
folhas representaram 57,5 % de todo material coletado, sendo predominantes no aporte
vertical, lateral, terrestre e estoque bêntico. Um total de 122 táxons foram identificados,
separados em 45 famílias. Myrtaceae, Lauraceae, Annonaceae e Euphorbiaceae foram
as famílias mais ricas, porém, Fabaceae e Moraceae foram as famílias que mais
contribuíram com o aporte de folhas. Seis espécies representaram 52,9 % do aporte total
de folhas para o riacho, sendo Schizolobium parahyba a principal com 25 % desse total.
A quantidade média de MO mensurada ao longo do período de estudo variou
significativamente entre os compartimentos assim como entre os meses. A partir dos
resultados foram rejeitadas as duas hipóteses: o aporte lateral foi muito superior ao
vertical, e houve variação temporal nos padrões de aporte de detritos ao riacho,
principalmente regulada através da precipitação.
Palavras chave: aporte vegetal, serapilheira, vegetação ripária

28
Abstract
Allochthonous organic matter (OM) input has great importance in headwater
stream energy flows, and the way these materials enter the ecosystem has been long
studied, particularly in temperate environments. Such studies are rare in subtropical
Atlantic Rain Forest. The objective of this study was to quantify and identify the OM
input of riparian vegetation in an Atlantic Rain Forest 3rd
order stream. The study had
two hypotheses: (1) vertical input is higher than lateral input, and (2) there is no
temporal variation in litter input to the stream. The vertical and lateral inputs, and the
terrestrial and benthic stocks were measured along 13 months. All collected material
was quantified as leaves, branches, reproductive and others, and the leaves were
identified into species. Lateral input was the highest compartment in direct litter input to
the stream, with mean of 175.5 g.m-2
.month-1
, compared to 32.4 g.m-2
.month-1
observed
in vertical input. Benthic stocks exceeded in terms of biomass the terrestrial stocks.
Leaves represented 57.5 % of all collected material, prevailing in vertical, lateral,
terrestrial and benthic stocks inputs. A total of 122 taxa were identified, classified into
45 families. Myrtaceae, Lauraceae, Annonaceae and Euphorbiaceae were the richest
families, although Fabaceae and Moraceae were the most important in leaves inputs. Six
species represented 52.9 % of total leaves input; Schizolobium parahyba was the main
species with 25 % of this amount. The mean value of OM measured in the study period
varied significantly between the compartments as well as between the months. Given
the results, the two hypotheses were rejected. Lateral input surpassed greatly the vertical
input, and temporal variation was observed in litter input patterns to the stream. Organic
matter dynamics in the stream was principally driven by precipitation.
Key-words: organic input, litterfall, riparian vegetation

29
Introdução
Estudos sobre o balanço de matéria orgânica (MO) em riachos (quantificação do
input e output de energia) auxiliam na compreensão da importância relativa dos recursos
alimentares como ferramentas para avaliação da integridade e comparação dos
ecossistemas (Tank et al., 2010). Para maioria dos riachos florestais, o entendimento de
tal balanço energético está intimamente ligado à dinâmica de MO proveniente da
vegetação ripária, que exerce um papel chave na dinâmica energética destes ambientes
(Gregory et al., 1991; Naiman & Décamps, 1997; Lima & Zakia, 2009). Por serem
sombreados, riachos de cabeceira são na maioria das vezes heterotróficos (Cummins et
al, 1973; Vannote et al., 1980), e suas comunidades dependem da incorporação alóctone
de MO vegetal (Cummins & Klug, 1979; Wallace et al., 1997) e inclusive animal
(Kawaguchi et al., 2003) provenientes da zona ripária. Assim, esta região representa
uma importante ligação aquático-terrestre essencial para manutenção da produtividade
de pequenos riachos florestais (Wallace et al., 1997; Naiman & Décamps, 1997;
Wallace et al., 1999; Magana, 2001).
Diversos fatores influenciam na maneira com que se dá a incorporação de
energia alóctone em riachos. A nível global, a dinâmica da matéria orgânica nesses
sistemas é controlada primariamente pelo clima através de seu efeito na vegetação
terrestre (Webster & Meyer, 1997). Segundo Wantzen et al. (2008), variações temporais
no aporte alóctone podem ocorrer de acordo com a fenologia da vegetação, com os
mecanismos de retenção na zona de transição aquático-terrestre, e com o clima local. A
estrutura e composição da vegetação ripária determina padrões de queda de serapilheira
(Tank et al., 2010), e a composição específica dos detritos varia de acordo com a
vegetação, estrutura física (Conners & Naiman, 1984) e biodiversidade do ecossistema
ripário (Wantzen et al., 2008). Apesar de certas generalizações, a quantidade e o tipo de
detrito, assim como a variação espacial e temporal do aporte de MO é extremamente
variável (Tank et al., 2010).
A precipitação também se mostra muito importante para o entendimento da
incorporação da MO alóctone. Períodos de intensas chuvas influenciam drasticamente o
aporte de detritos através das consequentes inundações, aumento da vazão, e
carreamento de detritos, mostrando serem eventos determinantes da dinâmica de
matéria orgânica em riachos (Acuña et al., 2007). Segundo Benfield et al. (1997), existe
uma correlação direta entre o aporte de detritos vegetais para os sistemas e a

30
precipitação anual das localidades onde eles se encontram. Não obstante, a vazão de
água afeta diretamente a capacidade de retenção de um rio, e pode influenciar
fortemente os estoques de matéria orgânica presentes no ecossistema aquático (Pozo et
al., 1997a; Acuña et al., 2007).
Outro fator relacionado a esse processo é o tamanho do corpo d’água. Estudos
demonstram que o aporte de serapilheira por unidade de área da superfície de um rio
decai exponencialmente com o aumento de sua ordem (Conners & Naiman, 1984;
Benfield, 1997; Richardson, 2005). Conners & Naiman (1984) demonstraram
experimentalmente que riachos de 1ª a 2ª ordem chegam a ter mais de 80% da produção
alóctone, enquanto rios de 5ª a 6ª ordem mais de 80% autóctone, indo de encontro aos
processos descritos e bem fundamentados no Conceito do Rio Contínuo (RCC)
(Vannote et al., 1980).
O aporte, processamento e exportação da matéria orgânica em sistemas lóticos
são fortemente influenciados por perturbações, sejam elas naturais ou antrópicas
(Richardson, 2005). Mudanças no uso da terra reduzem a área recoberta com vegetação
e alteram a quantidade, composição e retenção do aporte de detritos em rios e riachos
(Kominoski & Rosemond, 2012). Estudos indicam que a quantidade de matéria
orgânica aportada em rios reduz drasticamente de áreas de vegetação preservada para
áreas abertas por influência antrópica (Afonso et al., 2000; Magana, 2001; Carvalho &
Uieda, 2010; Kiffney & Richardson, 2010). A urbanização reduz drasticamente a
disponibilidade desse material através da diminuição significativa do input, e do
aumento da frequência e magnitude das cheias que acabam por controlar ultimamente os
padrões de retenção de matéria orgânica nos sistemas (Imberger et al., 2011). Além
disso, espécies exóticas também podem reduzir a quantidade e diversidade de detritos e
consequentemente diminuir o input de nutrientes para riachos (Pozo et al., 1997b).
A composição do material alóctone dentro do rio afeta diretamente a estrutura da
comunidade e a dieta da fauna aquática, e consequentemente, o ecossistema aquático
como um todo (Carvalho & Uieda, 2010). A matéria orgânica particulada grossa
(MOPG) vegetal pode atingir os cursos de água através de movimentos verticais, que
seria a queda da serapilheira diretamente sobre o curso de água, ou movimentos laterais,
que consiste no carreamento da matéria orgânica depositada no entorno para dentro do
rio (Benfield, 1997). Dados na literatura indicam uma participação muito maior (80-
90%) do aporte vertical quando comparado com o lateral (Conners & Naiman; 1984;
Benson & Person, 1993; Benfield, 1997). Esse material é representado por folhas,

31
flores, sementes, galhos, cascas, frutos e outras partes vegetais (Beinfield, 1997), sendo
constituído principalmente por folhas, que podem representar de 41 a 98% da matéria
orgânica presente (Abelho & Graça, 1998; Benson & Pearson, 1993; França et al.,
2009). Dentro do ecossistema aquático as folhas são decompostas por uma combinação
de fatores físicos, químicos e biológicos, incluindo as etapas de lixiviação,
condicionamento, e fragmentação pela abrasão física e por invertebrados (Cummins,
1974; Kirby et al.; 1983; Gessner et al., 1999). O processo de decomposição da MO
alóctone, além de liberar nutrientes e matéria orgânica dissolvida (MOD) para o
ambiente, também produz CO2 e CH4, gases causadores do efeito estufa (Prast & Pinho,
2008).
Comparando-se sistemas temperados com tropicais, observa-se que os
temperados têm um período de grande queda de folhas no outono e inverno, e
consequente maior incorporação de detritos aos rios (Fisher & Likens, 1973; Benfield,
1997; Pozo et al., 1997a). Os rios tropicais que possuem duas estações climáticas
marcantes (chuvosa e seca), apresentam um aporte de detritos muito maior no final da
estação seca (Gonçalves et al., 2006; Selva et al., 2007; França et al., 2009; Carvalho &
Uieda, 2010). Está claro que diferentes biomas podem ter padrões próprios na dinâmica
das formações florestais devido às condições fenológicas, geomorfológicas e climáticas
específicas. No entanto, o funcionamento dos ecossistemas aquáticos ainda não está
bem descrito devido à escassez de estudos de dinâmica da matéria orgânica em riachos
de regiões subtropicais.
Diante da discussão observada em literatura e baseado no fato de que no clima
subtropical não há uma estação seca marcante, este estudo propõem duas hipóteses a
serem testadas: (1) o aporte vertical é a principal via de entrada de material alóctone, e
(2) que não há variação temporal no aporte de detritos vegetais ao rio. O objetivo deste
trabalho foi determinar a composição e dinâmica da matéria orgânica vegetal alóctone
em um riacho de cabeceira de Mata Atlântica subtropical.
Métodos
Área de estudo
A pesquisa foi realizada no riacho Cachoeira Grande, um riacho de 3ª ordem
segundo a classificação de Strahler (1957), localizado na Bacia Hidrográfica da Lagoa
do Peri (27° 44’ S e 48° 31’ W), situada ao sul da parte insular de Florianópolis, região

32
sul do Brasil. O riacho tem suas nascentes situadas na cota de 280 m, se estende por
1,17 km até desaguar na Lagoa do Peri, drena uma área de 1,66 km², e apresenta uma
declividade que varia entre 0,26 e 0,24 m.m-1
nos cursos superior e médio, até 0,1 m.m-1
no baixo curso (Santos et al., 1989). A área está localizada em zona subtropical, que
segundo a classificação climática de Köppen tem clima do tipo Cfa, com ausência de
estação seca, chuvas distribuídas uniformemente durante o ano e verões rigorosos
(Nascimento, 2002). A bacia hidrográfica tem 20,3 km2 e está inteiramente
compreendida em uma unidade de conservação, o Parque Municipal da Lagoa do Peri,
criado em 1981. A região é de extrema importância para a cidade na medida em que na
lagoa funciona um sistema de captação de água com vazão média captada de 200 L.s-1
.
A região da área de estudo é recoberta por Floresta Ombrófila Densa em estádio
avançado de regeneração, possuindo um dos fragmentos vegetais mais desenvolvidos da
Ilha de Santa Catarina (Caruso, 1990), podendo inclusive abrigar focos de floresta
primária restritos ao alto das encostas nas regiões de nascentes (Neto & Klein, 1991).
Isso faz com que a área tenha grande relevância para conservação, visto que a Mata
Atlântica se encontra reduzida a apenas 7,2% de sua área original, e foi considerada pela
International Union for Conservation of Nature (IUCN) como um hot-spot de
biodiversidade mundial (Myers et al., 2000). De acordo com Campanili & Prochnow
(2006), a Floresta Ombrófila Densa tem característica perenifólia, com dossel de até 15
m e árvores emergentes até 40 m, densa vegetação arbustiva de samambaias
arborescentes, bromélias, palmeiras e abundantes trepadeiras. No sul do Brasil
predominam espécies das famílias Myrtaceae, Fabaceae, Rutaceae, Lauraceae,
Meliaceae, Apocynaceae, Palmae entre outras (Wettstein, 1970).
Delineamento amostral e metodologia de campo
O experimento para se avaliar a dinâmica de material orgânico (MO) vegetal no
ambiente lótico foi realizado em um trecho de 100 m do riacho Cachoeira Grande.
Foram escolhidos cinco pontos amostrais neste trecho separados a uma distância média
de 20 m, e em cada ponto amostral a quantificação do aporte de MO vegetal alóctone
foi realizada em quatro compartimentos do ecossistema:
Aporte vertical: para mensuração da MO vegetal alóctone que cai diretamente
sobre o curso de água, três cordas foram penduradas sobre o riacho a uma altura de 2 m
da água. Cada corda continha seis coletores, representados por baldes de 26 cm de
diâmetro (0,053 m2) com pequenos furos no fundo para impedir o acúmulo de água.

33
Aporte lateral: o aporte de MO que escoa do solo para o riacho foi mensurado
através de coletores com 0,5 m de comprimento e 0,2 m de altura (0,1 m2), com
profundidade de 30 cm e abertura de malha de 5 mm. Os coletores foram armados no
limite entre a água e o solo da margem do riacho, de modo a capturar a MO que se
movimenta do solo para água, um na margem direita e outro na margem esquerda de
cada ponto.
Aporte terrestre: considerado como um estoque potencial de MO que pode
chegar ao leito do rio, o aporte terrestre foi mensurado através de coletores de 1 m2
armados dentro da vegetação ripária, com profundidade de 10 cm e 5 mm de malha. Em
cada ponto de coleta foi armado um coletor na margem direita e outro na margem
esquerda do rio a uma distância de 10 m do curso d’água.
Estoque bêntico: a MO vegetal depositada no leito do riacho foi mensurada com
auxílio de um coletor tipo Surber, tamanho 0,32 x 0,32 m (0,1024 m2) e abertura de
malha de 0,25 mm. Em cada ponto de coleta foram realizadas aleatoriamente três
amostras feitas em regiões onde se encontrava depósito de folhiço.
Coletas do material retido nos coletores e do estoque bêntico foram feitas a cada
trinta dias durante treze meses, compreendendo o período de agosto de 2010 a agosto de
2011. Todo material coletado foi seco em estufa a 60ºC por no mínimo três dias até a
estabilização do peso, separado qualitativamente entre folhas, galhos e partes
reprodutivas, e pesado em balança analítica para determinação do peso seco. As folhas
foram identificadas até o menor grupo taxonômico possível com auxílio de bibliografia
adequada (Sobral et al., 2006), e através de comparações morfológicas com exsicatas
previamente identificadas provenientes de outra pesquisa do mesmo projeto que fez a
fitossociologia da área de estudo. Dados de pluviosidade, intensidade e direção de
ventos e temperatura média do ar ao longo do período de estudo foram fornecidos pela
Empresa de Pesquisa Agropecuária e Extensão Rural de Santa Catarina (EPAGRI). Para
análises foram consideradas as médias de temperatura, de ventos e o acumulado de
precipitação dos 30 dias antecedentes às coletas. Os valores de fotoperíodo considerado
foram os previsíveis para o 15º dia de cada mês na latitude 27°.
Cálculos dos percentuais da participação qualitativa do aporte e da contribuição
por compartimento foram feitos a partir da matriz de dados brutos com peso seco (g)
total de MO vegetal observado no período de estudo. O percentual da contribuição
específica dos táxons e das famílias foi calculado a partir do peso seco total das folhas.
A partir da matriz de dados brutos foi feita a padronização da área de cada coletor para

34
1 m2, assim como a padronização de todos os meses coletados para períodos de 30 dias.
Com a matriz de dados padronizada, calculou-se a média mensal em gramas de peso
seco (g.m-2
.mês-1
) dos diferentes compartimentos, assim como das espécies e táxons.
Tratamento estatístico
A fim de se verificarem variações na quantidade de biomassa vegetal aportada
entre os pontos amostrais, entre os compartimentos de aporte, assim como entre os
meses, foram realizadas ANOVAs one-way a partir da matriz de biomassa padronizada,
utilizando-se como variável dependente a biomassa total vegetal, e variável
independente, separadamente, os pontos, meses e compartimentos. Para verificar a
influência das variáveis ambientais no padrão de aporte vegetal foi feita uma análise de
correlação de Pearson com a matriz padronizada de biomassa e as relativas médias
mensais das variáveis mensuradas. Os dados para análises foram checados para
normalidade (Shapiro-Wilk) e homogeneidade de variâncias (Levene’s) (Zar, 1996). As
análises estatísticas foram realizadas com auxílio do programa Statistica 7.
Resultados
Dinâmica x fatores climáticos
Relacionando a incorporação de biomassa vegetal no riacho com as variáveis
ambientais, pode-se constatar uma correlação direta entre a pluviosidade com o aporte
total de MO (Pearson, r = 0,59; p < 0,05), assim como com o aporte lateral (Pearson, r =
0,43; p < 0,05) (Tabela 1). Os aportes vertical e terrestre, assim como o estoque bêntico,
não estiveram relacionados a nenhuma variável mensurada. Para biomassa de folhas em
específico, considerando-se as seis principais espécies, as médias de temperatura e o
fotoperíodo foram os principais fatores relacionados à queda de folhas. A incorporação
vegetal de Schizolobium parahyba (Vell.) Blake esteve negativamente relacionada às
médias de temperaturas máximas e mínimas do ar (Pearson, r = -0,22 e r = -0,3; p <
0,05) e ao fotoperíodo (Pearson, r = -0,2; p < 0,05), mesmo comportamento observado
na espécie Ficus eximia Schott (Pearson, r = -0,19, r = -0,22 e r = -0,18,
respectivamente; p < 0,05), o que indica que ambas as espécies tem um padrão de queda
foliar acentuado no inverno. Já as espécies Cupania vernalis Cambess. e Virola
bicuhyba (Schott ex Spreng.) Warb. tiveram um padrão de queda foliar relacionado

35
positivamente às médias de temperaturas do ar e ao fotoperíodo (Tabela 1), ou seja, um
padrão de queda foliar acentuada no verão.
Tabela 1. Valores de correlação de Pearson entre a biomassa vegetal incorporada das categorias
de aporte e principais espécies com as variáveis ambientais mensuradas no rio Cachoeira Grande entre ago/10 e ago/11. Representados apenas valores significativos ( p < 0,05)
Dinâmica de MO
A quantidade média de MO mensurada ao longo do período de estudo variou
significativamente entre os compartimentos (ANOVA, F3,625 = 23,18; p < 0,05), apesar
de não ter sido observada variação entre os pontos amostrais (ANOVA, F4,624 = 1,8; p >
0,05). O aporte lateral foi o compartimento que mais contribuiu com a incorporação
direta de MO vegetal para o riacho, com uma média mensal de 175,5 g.m-2
.mês-1
,
superando largamente o aporte vertical, que apresentou uma média de 32,4 g.m-2
.mês-1
.
O estoque bêntico apresentou uma média de 115,4 g.m-2
.mês-1
, superando o aporte do
estoque potencial terrestre de apenas 69,1 g.m-2
.mês-1
(Fig. 1).
Na composição qualitativa do input vegetal observado, as folhas representaram a
maior parte do aporte, com 57,5 % de todo material coletado no período de estudo.
Galhos representaram 25,2 %, partes reprodutivas apenas 5,1 % e restos 12 % desse
total. As folhas formaram a principal parte da MO em todos os compartimentos,
representando 77,8 % do aporte vertical, 58,5 % do aporte terrestre, 43,6 % do aporte
lateral e 45,6 % do estoque bêntico (Fig. 2).
pluviosidade vento temp. mín temp. máx fotoperíodo
MO total 0,59 0,27
vertical
terrestre
lateral 0,43
bêntico
miscelânia 0,21 0,15
galhos 0,17
Cupania vernalis 0,25 0,25 0,27
Ficus adhatodifolia 0,13 0,13
Ficus eximia -0,19 -0,23 -0,18
Schizolobium parahyba -0,22 -0,3 -0,2
Tetrorchidium rubrivenium -0,02
Virola bicuhyba 0,19 0,18 0,21
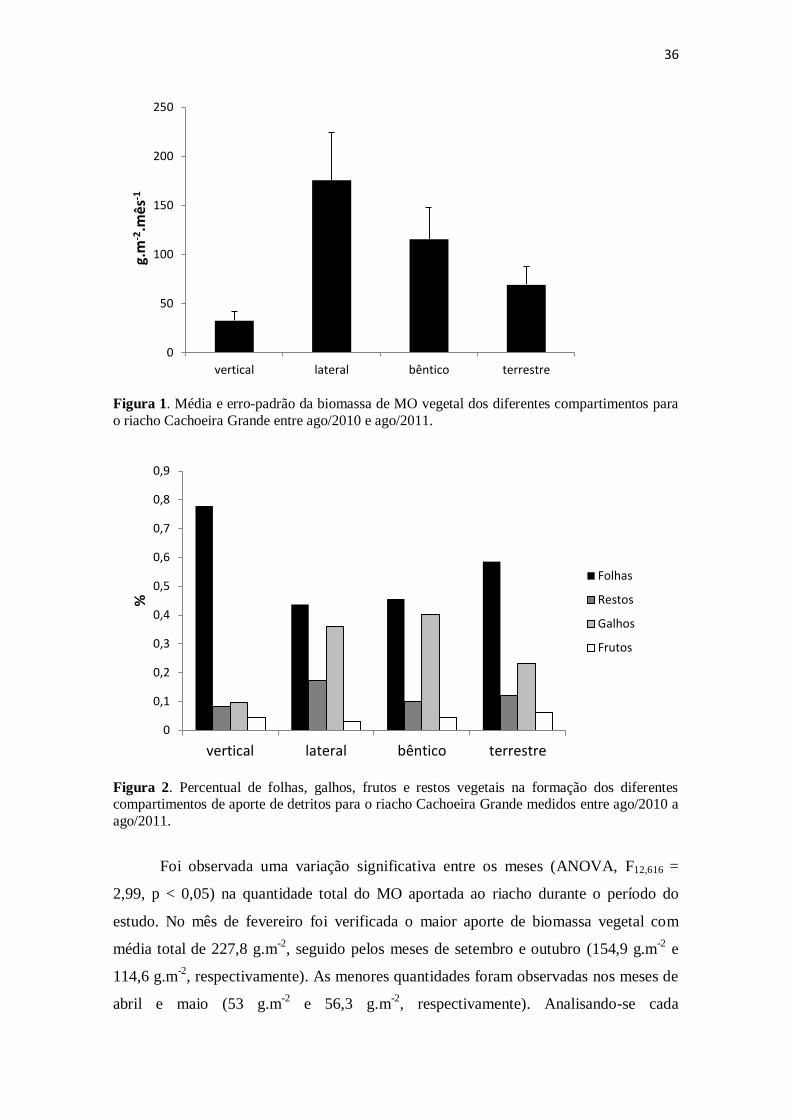
36
Figura 1. Média e erro-padrão da biomassa de MO vegetal dos diferentes compartimentos para
o riacho Cachoeira Grande entre ago/2010 e ago/2011.
Figura 2. Percentual de folhas, galhos, frutos e restos vegetais na formação dos diferentes compartimentos de aporte de detritos para o riacho Cachoeira Grande medidos entre ago/2010 a
ago/2011.
Foi observada uma variação significativa entre os meses (ANOVA, F12,616 =
2,99, p < 0,05) na quantidade total do MO aportada ao riacho durante o período do
estudo. No mês de fevereiro foi verificada o maior aporte de biomassa vegetal com
média total de 227,8 g.m-2
, seguido pelos meses de setembro e outubro (154,9 g.m-2
e
114,6 g.m-2
, respectivamente). As menores quantidades foram observadas nos meses de
abril e maio (53 g.m-2
e 56,3 g.m-2
, respectivamente). Analisando-se cada
0
50
100
150
200
250
vertical lateral bêntico terrestre
g.m
-2.m
ês-1
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
vertical lateral bêntico terrestre
%
Folhas
Restos
Galhos
Frutos

37
compartimento em separado ao longo do ano, pode-se observar que foram mais
constantes os valores dos aportes vertical (variou de 19,8 g.m-2
em ago/2010 a 57,8 g.m-
2 em ago/2011) e terrestre (variou de 42,2 g.m
-2 em março a 136,9 g.m
-2 em ago/2011).
A variação temporal foi mais marcante no aporte lateral (variou de 697,5 g.m-2
em
fevereiro, a 51,2 g.m-2
em ago/2010) e no estoque bêntico (variou de 392,7 g.m-2
em
setembro a 42,5 g.m-2
em ago/2011). (Fig. 3).
Figura 3. Variação mensal do aporte de matéria orgânica (MO) vegetal seca nos diferentes
compartimentos estudados no riacho Cachoeira Grande entre ago/2010 e ago/2011. Barras
representam o erro padrão.
Composição de espécies
As folhas dos aportes foram identificadas em 122 taxa de traqueófitas
subdivididos em 45 famílias botânicas (Apêndice). As famílias com o maior número de
taxa foram Myrtaceae, Lauraceae, Annonaceae, Euphorbiaceae e Sapindaceae, com 12,
10, 8, 8 e 7 taxa, respectivamente. As cinco principais famílias em termos de
incorporação de biomassa foliar ao rio foram responsáveis por 71,9% do aporte total.
Fabaceae foi a família mais representativa contando com 30% desse total, seguida de
Moraceae (17,4%), Euphorbiaceae (11,6%), Sapindaceae (7,5%) e Myristicaceae (5,3%)
(Fig. 4).
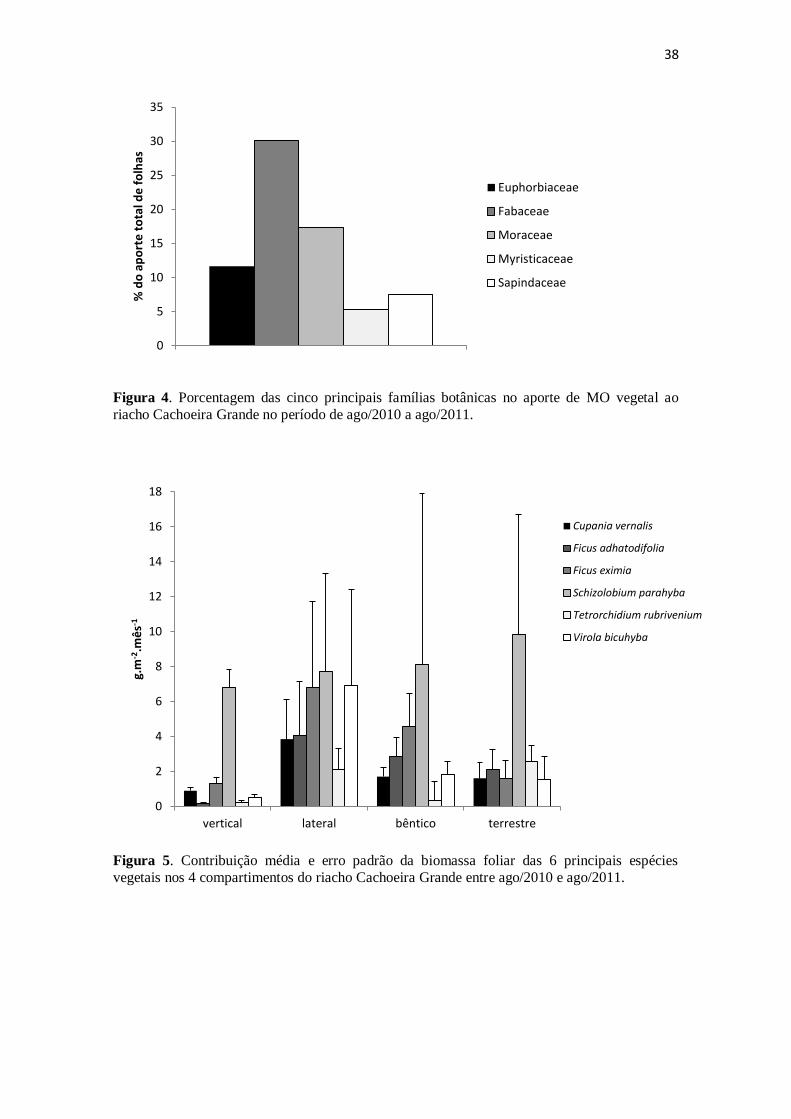
38
Figura 4. Porcentagem das cinco principais famílias botânicas no aporte de MO vegetal ao
riacho Cachoeira Grande no período de ago/2010 a ago/2011.
Figura 5. Contribuição média e erro padrão da biomassa foliar das 6 principais espécies
vegetais nos 4 compartimentos do riacho Cachoeira Grande entre ago/2010 e ago/2011.
0
5
10
15
20
25
30
35
% d
o a
po
rte
to
tal d
e f
olh
as
Euphorbiaceae
Fabaceae
Moraceae
Myristicaceae
Sapindaceae
0
2
4
6
8
10
12
14
16
18
vertical lateral bêntico terrestre
g.m
-2.m
ês-1
Cupania vernalis
Ficus adhatodifolia
Ficus eximia
Schizolobium parahyba
Tetrorchidium rubrivenium
Virola bicuhyba

39
A principal espécie ripária na dinâmica de MO do rio foi S. parahyba, que
representou mais de 25% da biomassa total de folhas nos compartimentos. Essa espécie
apresentou uma média de aporte de 8,1 g.m-2
.mês-1
, sendo predominante em todos os
quatro compartimentos estudados. Outras espécies foram representativas, como: F.
eximia (3,5 g.m-2
.mês-1
), V. bicuhyba (2,7 g.m-2
.mês-1
), Ficus adhatodifolia Schott ex
Spreng. (2,3 g.m-2
.mês-1
), C. vernalis (2 g.m-2
.mês-1
) e Tetrorchidium rubrivenium
Poepp. (1,3 g.m-2
.mês-1
) (Fig. 5). Estas espécies foram responsáveis por 52,9% da
biomassa total de folhas encontradas, e representativas em todos os compartimentos na
dinâmica de energia do riacho.
As principais espécies variaram quanto a representatividade da biomassa total
nos compartimentos e quanto aos padrões temporais de aporte. Somente S. parahyba e
F. eximia estiveram entre as quatro espécies mais representativas de todos
compartimentos. S. parahyba apresentou picos de queda de folhas nos meses de agosto
e setembro de 2010, e novamente entre maio e agosto de 2011 para o aporte vertical
(Fig. 6, a) e o terrestre (Fig. 6, c). No estoque bêntico, essa espécie mostrou um
acúmulo máximo em setembro de 2010 que superou largamente todas outras medições
feitas para o compartimento (Fig. 6, d). A espécie F. eximia apresentou picos de queda
em setembro e outubro de 2010 e entre junho a agosto de 2011 para os aportes vertical e
terrestre, padrão similar ao observado para o aporte lateral e o estoque bêntico (Fig. 6).
C. vernalis mostrou maiores taxas de aporte tanto vertical quanto lateral entre outubro
de 2010 e fevereiro de 2011. F. adhatodifolia apresentou maior aporte de biomassa
foliar entre novembro e dezembro de 2010 no aporte terrestre (Fig. 6, c), e máximos de
acumulação em agosto e novembro de 2010 no estoque bêntico (Fig. 6, d). A espécie V.
bicuhyba apresentou um aumento na biomassa incorporada entre novembro e dezembro
de 2010 para o aporte vertical (Fig. 6, a), e entre outubro de 2010 e janeiro de 2011 para
o aporte lateral (Fig. 6, b). T. rubrivenium apresentou picos de incorporação de
biomassa foliar entre setembro e novembro de 2010 no aporte terrestre (Fig. 6, c).
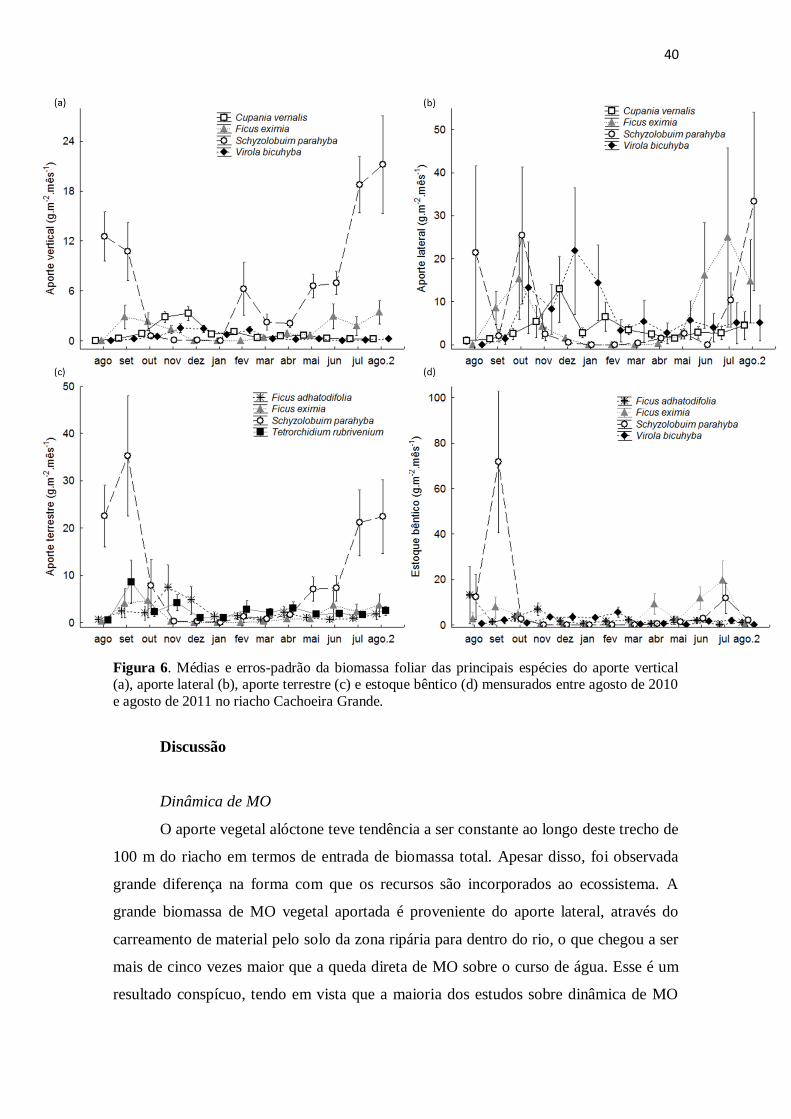
40
Figura 6. Médias e erros-padrão da biomassa foliar das principais espécies do aporte vertical (a), aporte lateral (b), aporte terrestre (c) e estoque bêntico (d) mensurados entre agosto de 2010
e agosto de 2011 no riacho Cachoeira Grande.
Discussão
Dinâmica de MO
O aporte vegetal alóctone teve tendência a ser constante ao longo deste trecho de
100 m do riacho em termos de entrada de biomassa total. Apesar disso, foi observada
grande diferença na forma com que os recursos são incorporados ao ecossistema. A
grande biomassa de MO vegetal aportada é proveniente do aporte lateral, através do
carreamento de material pelo solo da zona ripária para dentro do rio, o que chegou a ser
mais de cinco vezes maior que a queda direta de MO sobre o curso de água. Esse é um
resultado conspícuo, tendo em vista que a maioria dos estudos sobre dinâmica de MO

41
em riachos demonstram uma maior contribuição do aporte vertical quando comparado
com o lateral, tanto para ambientes tropicais (Afonso et al., 2000; Gonçalves et al.,
2006; Selva et al., 2007; Carvalho & Uieda, 2010) como para ambientes temperados
(Fisher & Likens, 1979; Stewart & Davis, 1990; Pozo et al., 1997b; Webster et al.,
1997; Acunña et al., 2007) (Tabela 2). Apenas França et al. (2009) observaram uma
maior participação do aporte lateral comparando-se com o aporte vertical em um riacho
de transição Cerrado/Mata Atlântica, porém, os valores entre o aporte vertical e lateral
foram próximos, e a média de entrada de biomassa lateralmente foi cinco vezes menor
que o observado neste estudo. Essas raras observações de estudos com maior
participação do aporte lateral provavelmente estão relacionadas ao fato de existirem
poucas pesquisas desse gênero em riachos de Mata Atlântica, assim como à falta de
dados como topografia e declividade da área de drenagem dos ecossistemas estudados.
Como evidenciaram Selva et al. (2007) e Wantzen et al. (2008), a declividade da
margem assim como características do solo e superfície de escoamento estão
intimamente relacionadas ao transporte lateral de biomassa vegetal em riachos.
Tabela 2. Resumo de dados do aporte médio de MO (g.m-2
.mês-1
) vegetal em riachos e rios de
ambientes tropicais e temperados. AV = aporte vertical; AL = aporte lateral; AT = aporte terrestre; EB = estoque bêntico; % F = porcentagem de folhas. * dados do presente artigo.
**foram feitos experimentos de 48h, o que acaba por não demonstrar a variabilidade
representativa de um mês.
A grande incorporação lateral observada esteve fortemente relacionada com
períodos de intensa precipitação, que acabaram por criar pequenos fluxos laterais
Rio Clima (bioma) Conservação Ordem AV AL AT EB % F Referência
Cachoeira Grande, SC (BR) Subtropical (Mata Atlântica) preservado 3ª 32 175 69 115 57 Lisboa et al., 2012*
Doné, MG (BR) Tropical (Cerrado / Mata Atlântica) preservado 3ª 28 31 59 138 77,5 Franca et al., 2009
Itaúna, SP (BR) Tropical (Cerrado / Mata Atlântica) preservado 3ª 59 35 94 - 73 Afonso et al., 2000
área aberta 3ª 7 10 17 - 79 Afonso et al., 2000
Ribeirão da Quinta, SP (BR) Tropical (Cerrado / Mata Atlântica) preservado 3ª 102 64 166 - 71 Carvalho & Uieda, 2010
área aberta 3ª 0 6 6 - 71 Carvalho & Uieda, 2010
Indaiá, MG (BR) Tropical (Cerrado) preservado 3ª 24 15 39 316 >50 Gonçalves et al., 2006
Juruena, MT (BR) Tropical (Amazônia) preservado 1ª 98 34 - - 50 Selva et al., 2007
vários (BR) Tropical (Amazônia) preservado 1-3ª 58 12 - - - McClain & Richey, 1996
Andorinha, RJ (BR) Tropical (Mata Atlântica) preservado 3ª 9** - - - - Rezende & Mazzoni, 2005
Nojoro (KE) Tropical preservado - 96 - 175 168 40 Magana, 2001
área aberta - 42 - 86 91 35 Magana, 2001
Fuirosos (ES) Temperado (Mediterrâneo) preservado 3ª 55 16 71 - 65 Acuña et al., 2007
Window (SA) Temperado (Fynbos / Mediterrâneo) preservado 1ª - - 35 41 62 Stewart & Davis, 1990
Saiderrey (ES) Temperado (Floresta Decídua) preservado 1ª 32 9 41 18 77 Pozo et al., 1997b
Jerguerón (ES) Temperado (Eucalipto) preservado 1ª 24 2 26 37 92 Pozo et al., 1997b
Bear Brook (US) Temperado (Conifera) preservado 2ª 46 - 55 95 66 Fisher & Likens, 1973
Hugh Withe Creek (US) Temperado (Floresta Decídua) preservado 2ª 42 8 - 494 82 Webster et al., 1997

42
intermitentes facilitando o carreamento do material vegetal acumulado no solo da zona
ripária para dentro do riacho. Segundo Acuña et al. (2007), a precipitação é um dos
principais fatores que controlam a dinâmica da MO de riachos através dos distúrbios
causados por suas consequentes inundações. Outras pesquisas em riachos tropicais de
Mata Atlântica também evidenciaram uma relação direta entre o aporte total de MO
com períodos chuvosos (Rezende & Mazzoni, 2005; Nin, 2007). Dentro disso, a Mata
Atlântica se diferencia daqueles ambientes tropicais que apresentam sazonalidade
fortemente marcada em estações chuvosa e seca. Pesquisas vêm demonstrando que é na
estação seca, mais especificamente no fim deste período, onde ocorre a maior parte do
aporte de biomassa vegetal em riachos tropicais tanto do Cerrado (Gonçalves et al.,
2006; França et al., 2009; Carvalho & Uieda, 2010), como da Floresta Amazônica
(Selva et al., 2007). Esse padrão está diretamente relacionado ao estresse hídrico
causado por um período de pouca pluviosidade, que força a abscisão foliar de muitas
das espécies de árvores presentes (Campanili & Prochnow 2006). Além disso, a alta
taxa de acúmulo de material na estação seca também influencia o balanço do transporte
final da MO vegetal no riacho (Gonçalves et al., 2006). Entretanto, essa sazonalidade
em ambientes tropicais é diferente e muitas vezes menos marcada que o comportamento
típico de riachos em ambientes temperados (Wantzen et al., 2008), cuja maior parte da
produção de MO se dá no outono (Benfield, 1997; Pozo et al., 1997a), e esse período
pode representar até 90 % da queda de folhas total anual e cerca de 54 % do aporte total
de MO (Fisher & Likens, 1973).
No presente estudo, apesar da variação nas taxas de aporte mensais, não foi
observado um padrão sazonal na incorporação direta de MO para o riacho. Essa
variação se deu principalmente por causa de um evento de cheias causado por altas
taxas de precipitação observadas no mês de fevereiro, que apresentou um pico máximo
de aporte lateral, e acabou por representar um outlier se comparado com os outros
meses. Por ser uma floresta de copa sempre-verde perenifólia (Campanili & Prochnow
2006), para Mata Atlântica propriamente dita é esperada uma queda constante de
serapilheira entre os meses, e, desconsiderando-se eventos de inundações, a entrada de
material alóctone nos riachos ocorre quase que constantemente ao longo do ano (Nin,
2007).
Composição da MO

43
A composição específica de folhas variou ao longo do ano, apesar do caráter
perenifólio da vegetação. A principal espécie para dinâmica da MO do riacho em termos
de biomassa incorporada, S. parahyba, apresentou uma queda de folhas bastante
acentuada no inverno, com pico máximo nos meses de julho e agosto de 2011. Com o
começo da primavera a queda de folhas da espécie teve uma redução brusca, chegando a
zerar no mês de janeiro, e apresentou novamente um pequeno pico no mês de fevereiro
(verão). A espécie F. eximia também apresentou uma queda de folhas acentuada nos
meses de inverno, e uma redução brusca no verão chegando a zerar em janeiro. Essa
diminuição do aporte de duas das principais espécies nos meses mais quentes parece ter
sido compensada por um aumento na queda de folhas de outras espécies, principalmente
C. vernalis, V. bichyba e F. adhatodifolia, que apresentaram picos de aporte de folhas
nos meses quentes.
É provável que a grande diversidade de plantas contribuindo com incorporação
de folhas para o ecossistema esteja influenciando na dinâmica final de MO de maneira a
resultar em um padrão de aporte vertical total constante ao longo do ano, tendo em vista
os diferentes comportamentos ecológicos e fenológicos de cada espécie. Esse é um
padrão que foi observado também por Gonçalves et al. (2006), onde existem diferenças
na contribuição específica porque as espécies de plantas perdem as folhas em épocas
diferentes do ano. Em ecossistemas temperados, cuja sazonalidade é bem marcada, a
vegetação ripária é normalmente pobre em diversidade. Na Europa central, por
exemplo, é comum não se achar mais que dez espécies contribuindo com o aporte
vegetal de determinado riacho (Wantzen et al., 2008). Dessa forma, os mais de 122
táxons de plantas encontrados aqui superam largamente o que se encontra normalmente
em ambientes temperados, inclusive supera outras observações para ambientes tropicais
(França et al., 2009). Esse fato era de certa forma esperado, na medida em que a Mata
Atlântica é uma das florestas mais ricas do mundo em diversidade de árvores
(Campanili & Prochnow 2006).
As folhas formaram a principal fração do aporte total de MO para o riacho,
assim como o observado em outros estudos relacionados, tanto de ecossistemas
tropicais como temperados (Tabela 2). Certas espécies de árvores contribuem com a
maioria desse total, e acabam por ser determinantes e reguladoras dessa ligação
aquático-terrestre presente nesses sistemas. De acordo com o observado aqui, pode-se
sugerir algumas espécies-chave na dinâmica de matéria orgânica de riachos de Mata
Atlântica subtropical em estádio médio-avançado de regeneração, cujas presenças em

44
matas ripárias se faz de extrema importância tanto pela incorporação de energia como
para manutenção da saúde ecológica dos próprios ecossistemas aquáticos (Lima e Zakia,
2009). São elas as espécies S. parahyba, F. eximia, V. bicuhyba, F. adhatodifolia, C.
vernalis e T. rubrivenium.
Considerando-se os resultados encontrados, foram rejeitadas as duas hipóteses
formuladas inicialmente. Ao contrário do que é normalmente encontrado na literatura, o
aporte lateral no riacho em questão foi muito superior ao aporte vertical, e teve uma
forte ligação com a precipitação acumulada, provavelmente associada à declividade do
terreno. Houve variação temporal no aporte de detritos ao rio, mas, diferentemente dos
padrões de Cerrado e Floresta Amazônica, essa temporalidade não é baseada em
estações bem definidas de “chuva” e “seca”, mas sim relacionada diretamente com
períodos pontuais de intensa precipitação distribuídos imprevisivelmente ao longo do
ano. É evidente a falta de estudos sobre dinâmica e composição do aporte da vegetação
ripária em riachos de Mata Atlântica, sobretudo em clima subtropical, sendo que mais
estudos são necessários para comparações relativas a padrões gerais de aporte e
identificação de espécies-chave da dinâmica de MO nas zonas ripárias.
Agradecimentos
Os autores gostariam de agradecer à Coordenação de Aperfeiçoamento de
Pessoal de Nível Superior (CAPES) pela bolsa de estudos concedida durante o período
de estudo. Agradecemos também ao suporte estrutural fornecido através da
Universidade Federal de Santa Catarina (UFSC), do Laboratório de Biologia e Cultivo
de Peixes de Água Doce (LAPAD) da UFSC, do Parque Municipal da Lagoa do Peri e
do departamento de Ecologia e Zoologia (ECZ) da UFSC. Agradecemos também à
EPAGRI pelos dados fornecidos. Agradecemos por último a toda equipe do Laboratório
de Ecologia de Águas Continentais (LIMNOS) da UFSC.
Referências capítulo 1
Abelho, M. & Graça, M.A.S. (1998). Litter in a first-order stream of a temperate
deciduous forest (Margaraça Forest, central Portugal). Hydrobiologia, 386, 147–152.

45
Acuña, V., Giorgi, A., Muñoz, I., Sabater, F. & Sabater, S. (2007).
Meteorological and riparian influences on organic matter dynamics in a forested
Mediterranean stream. Journal of the North American Benthological Society, 26(1):54-
69.
Afonso, A.A.O., Henry, R. & Rodella, R.C.S.M. (2000). Allochthonous matter
input in two different stretches of a headstream (Itatinga, São Paulo, Brazil). Brazilian
Archives of Biology and Technology, 43, 335-343.
Benfield, E.F. (1997). Comparison of litterfall input streams. Journal of the
North American Benthological Society, 16,104-108.
Benson, L.J. & Pearson, R.G. (1993). Litter inputs to a tropical Australian
rainforest stream. Australian Journal of Ecology, 18, 377–383.
Campanili, M. & Prochnow, M. (orgs) (2006). Mata Atlântica – uma rede pela
floresta. RMA, Brasília: BR.
Caruso, M.M.L. (1990). O desmatamento na Ilha de Santa Catarina de 1500 aos
dias atuais. Ed. da UFSC, Florianópolis, BR.
Carvalho, E.M. & Uieda, V.S. (2010). Input of litter in deforested and forested
areas of a tropical headstream. Brazilian Journal of Biology, 70, 283-288.
Conners, M.E. & Naiman, R.J. (1984). Particulate allochthonous inputs:
relationships with stream size in an undisturbed watershed. Canadian Journal of
Fisheries and Aquatic Sciences, 41, 1473-1484.
Cummins, K.W., Petersen, R.C., Howard, F.O., Wuycheck, J.C. & Holt, V.I.
(1973). The utilization of leaf litter by stream detritivores. Ecology, 54 (2), 336-345.
Cummins, K.W. (1974). Structure and function of stream ecosystems.
BioScience, 24, 631-641.

46
Cummins, K.W. & Klug, M.J. (1979). Feeding ecology of stream invertebrates.
Annual Review of Ecology, Evolution and Systematics, 10, 147-172.
Fisher, S.G. & Likens, G. E. (1973). Energy flow in Bear Brook, New
Hampshire: an integrative approach to stream ecosystem metabolism. Ecological
Monographs, 43, 421-439.
França, J.S., Gregorio, R.S., De Paula, J.D., Gonçalves Jr., J.F.; Ferreira, F.A. &
Callisto, M. (2009). Composition and dynamics of allochthonous organic matter inputs
and benthic stock in a Brasilian stream. Marine and Freshwater Research, 60, 990-998.
Gessner, M.O., Chauvet E. & Dobson M. (1999). A perspective on leaf litter
breakdown in streams. Oikos, 85, 377-384.
Gonçalves Jr., J.F., França, J.S. & Callisto, M. (2006). Dynamics of
allochthonous organic matter in a tropical Brazilian headstream. Brazilian Archives of
Biology and Technology, 49, 967-973.
Gregory, S.V., Swanson, F.J., Mckee, W.A. & Cummins, K.W. (1991). An
ecosystem perspective of riparian zones – focus on links between land and water.
BioScience, 41, 540–551.
Imberger, S.J., Thompson, R.M. & Grace, M.R. (2011). Urban catchment
hydrology overwhelms reach scale effects of riparian vegetation on organic matter
dynamics. Freshwater Biology, doi:10.1111/j.1365-2427.2011.02575.x.
Kawaguchi, Y., Taniguchi, Y. & Nakano, S. (2003). Terrestrial invertebrate
inputs determine the local abundance of stream fishes in a forested stream. Ecology,
84(3), 701–708.
Kiffney, P.M. & Richardson, J.S. (2010). Organic matter inputs into headwater
streams of southwestern British Columbia as a function of riparian reserves and time
since harvesting. Forest Ecology and Management, 260, 1931–1942.

47
Kirby, J.M., Webster, J.R. & Benfield, E.F. (1983). The role of shredders in
detrital dynamics of permanent and temporary streams. In: Fontaine, T.D & Bartell,
S.M. Dynamics of lotic ecosystems. Michgan, Ann Arbor Science, 494p.
Kominoski J.S. & Rosemond, A.D. (2012). Conservation from the bottom up:
forecasting effects of global change on dynamics of organic matter and management
needs for river networks. Freshwater Science, 2012, 31(1), 51–68.
Lima, W.P. & Zakia, M.J.B. (2009). Hidrologia de matas ciliares. In: Rodrigues,
R.R. & Leitão-Filho, H.F.L. (eds.). Matas ciliares: conservação e recuperação. São
Paulo, Edusp, 2ª ed. 2ª reimp. 319p.
Magana, A.E.M. (2001). Litter input from riparian vegetation to streams: a case
study of the Njoro River, Kenya. Hydrobiologia 458, 141–149.
Myers, N., Mittermeier, R.A., Mittermeier C.G., Fonseca G.A.B. & Kents, J.
(2000). Biodiversity hotspots for conservation priorities. Nature, 403, 853-845.
Naiman, R.J. & Décamps, H. (1997). The ecology of interfaces: Riparian zones.
Annual Review of Ecology, Evolution, and Systematics, 28, 621-658.
Nascimento, R. (2002). Atlas Ambiental de Florianópolis. Instituto Larus,
Florianópolis, BR.
Neto, A.B.C. & Klein, R.M. (1991). Mapeamento temático do município de
Florianópolis: Vegetação - síntese temática. IPUF, Florianópolis, BR.
Nin, C.S., Ruppenthal, E.L. & Rodrigues, G.G. (2007). Vegetação ripária e suas
funções ecológicas referentes à produção de folhiço em cursos de água, São Francisco
de Paula, RS. Revista Brasileira de Biociências, 5, 861-863.
Pozo, J., González, E., Díez, J.R., & Elósegui, A. (1997a). Leaf-litter budgets in
two contrasting forested streams. Limnetica, 13(2), 77-84.

48
Pozo, J., González, E., Díez, J.R., Molinero, J. & Elósegui, A. (1997b). Inputs of
particulate organic matter to streams with different riparian vegetation. Journal of the
Notrh America Benthological Society, 16, 602-611.
Prast, A.E. & Pinho, L. (2008). Ciclo do carbono em ecossistemas aquáticos
continentais brasileiros. Oecologia Brasiliensis, 12(1), 03-05.
Rezende, C.F. & Mazzoni, R. (2005). Seasonal variation in the input of
allochthonous matter in an Atlantic Rain Forest stream, Ilha Grande-RJ. Acta
Limnologica Brasiliensia, 17, 167-175.
Richardson, J.S., Bilby R.E. & Bondar, C.A. (2005). Organic matter dynamics in
small streams of the Pacific Northwest. Journal of the American Water Resources
Association, 41(4), 921-934.
Santos, G.F., Silva, J.T.N., Mendoça, M. & Veado, R.W. ad-V. (1989). Análise
Ambiental da Lagoa do Peri. Geosul, 8, 101-123.
Selva, E.C., Couto, E.G., Johnson, M.S. & Lehmann, J. (2007). Litterfall
production and fluvial export in headwater catchments of the southern Amazon. Journal
of Tropical Ecology, 23, 329–335.
Sobral, M., Jarenkow, J.A., Brack, P., Irgang, B., Larocca, J. & Rodrigues, R.s.s.
(2006). Flora arbórea e arborescente do Rio Grande do Sul, Brasil. São Carlos,
RiMA/Novo Ambiente.
Stewart, B.A. & Davis, B.R. (1990). Allochthonous input and retention in a
small mountain stream, South Africa. Hydrobiologia, 202, 135-146.
Strahler, A.N. (1957). Quantitative Analysis of Watershed Geomorphology.
Transactions, American Geophysical Union 38, 913-920.

49
Tank, J.L., Rosi-Marshall, E.J., Griffiths, N.A., Entrekin, S.A. & Stephen, M.L.
(2010). A review of allochthonous organic matter dynamics and metabolism in streams.
Journal of the North American Benthological Society, 29, 118–146.
Vannote, R.L., Minshall, G.W., Cummins, K.W., Sedell, J.R. & Cushing, C.E.
(1980).The river continuum concept. Canadian Journal of Fisheries and Aquatic
Sciences. 37, 130–137.
Wallace, J.B., Eggert, S.L., Mayer, J.L. & Webster, J.R. (1997). Multiple trophic
levels of a forest stream linked to terrestrial litter inputs. Science, 277,102-104.
Wallace, J.B., Eggert, S.L., Mayer, J.L. & Webster, J.R. (1999). Effects of
resource limitation on a detrital-based ecosystem. Ecological Monographs, 69, 409–
442.
Wantzen, K.M, Yule, C.M., Mathooko, J.M. & Pringle, C.M. (2008). Organic
Matter Processing in Tropical Streams. In: Dudgeon, D. & Cressa, C. (Org). Tropical
Stream Ecology. Elsevier, 2008, p. 44-64.
Webster, J.R. & J.L. Meyer. (1997). Organic matter budgets for streams: a
synthesis. Journal of the North American Benthological Society,16, 141–161.
Webster, J.R., Meyer, J.L., Wallace, J.B. & Benfield E.F. (1997). Organic matter
dynamics in Hugh White Creek, Coweeta Hydrologic Laboratory, North Carolina,
USA. Journal of the North American Benthological Society,16, 74-78.
Wettstein, R.R.V. (1970). Plantas do Brasil: aspectos da vegetação do sul do
Brasil. , Edgar Blücher, São Paulo, BR.
Zar, J.H. (1996). Biostatistical Analysis. 3rd ed., Prentice-Hall: NewYork.
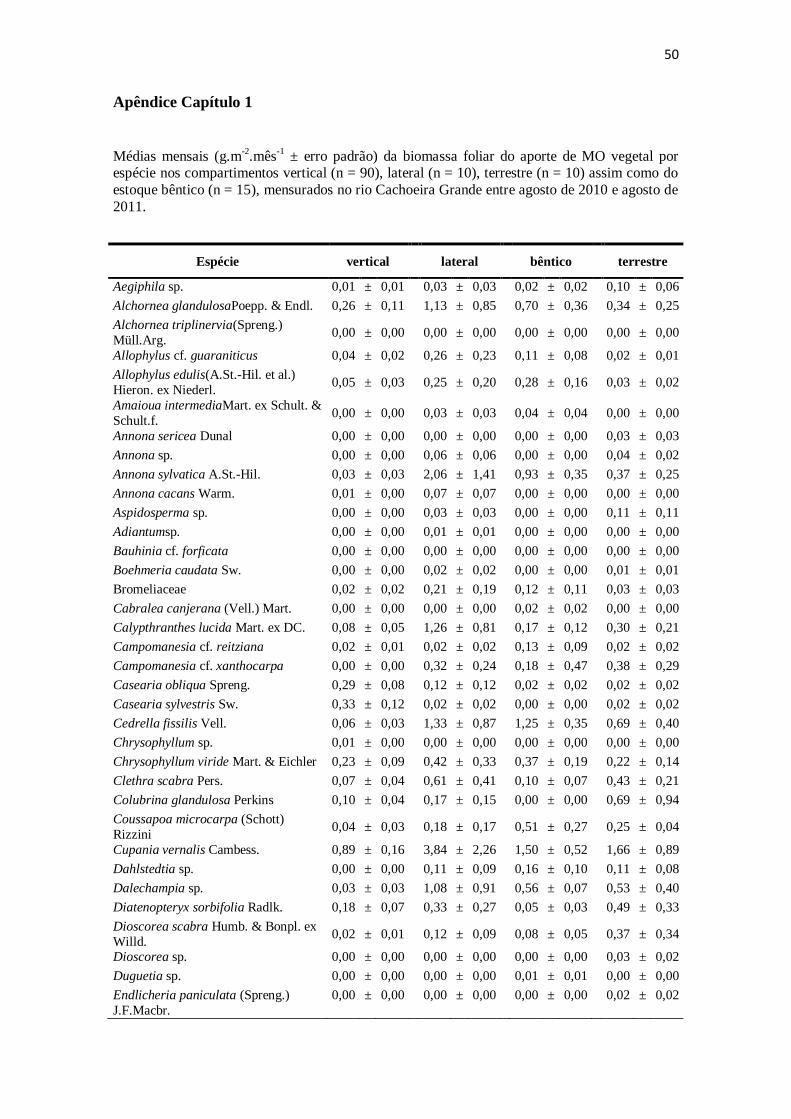
50
Apêndice Capítulo 1
Médias mensais (g.m-2
.mês-1
± erro padrão) da biomassa foliar do aporte de MO vegetal por espécie nos compartimentos vertical (n = 90), lateral (n = 10), terrestre (n = 10) assim como do
estoque bêntico (n = 15), mensurados no rio Cachoeira Grande entre agosto de 2010 e agosto de
2011.
Espécie vertical lateral bêntico terrestre
Aegiphila sp. 0,01 ± 0,01 0,03 ± 0,03 0,02 ± 0,02 0,10 ± 0,06
Alchornea glandulosaPoepp. & Endl. 0,26 ± 0,11 1,13 ± 0,85 0,70 ± 0,36 0,34 ± 0,25
Alchornea triplinervia(Spreng.)
Müll.Arg. 0,00 ± 0,00 0,00 ± 0,00 0,00 ± 0,00 0,00 ± 0,00
Allophylus cf. guaraniticus 0,04 ± 0,02 0,26 ± 0,23 0,11 ± 0,08 0,02 ± 0,01
Allophylus edulis(A.St.-Hil. et al.)
Hieron. ex Niederl. 0,05 ± 0,03 0,25 ± 0,20 0,28 ± 0,16 0,03 ± 0,02
Amaioua intermediaMart. ex Schult. &
Schult.f. 0,00 ± 0,00 0,03 ± 0,03 0,04 ± 0,04 0,00 ± 0,00
Annona sericea Dunal 0,00 ± 0,00 0,00 ± 0,00 0,00 ± 0,00 0,03 ± 0,03
Annona sp. 0,00 ± 0,00 0,06 ± 0,06 0,00 ± 0,00 0,04 ± 0,02
Annona sylvatica A.St.-Hil. 0,03 ± 0,03 2,06 ± 1,41 0,93 ± 0,35 0,37 ± 0,25
Annona cacans Warm. 0,01 ± 0,00 0,07 ± 0,07 0,00 ± 0,00 0,00 ± 0,00
Aspidosperma sp. 0,00 ± 0,00 0,03 ± 0,03 0,00 ± 0,00 0,11 ± 0,11
Adiantumsp. 0,00 ± 0,00 0,01 ± 0,01 0,00 ± 0,00 0,00 ± 0,00
Bauhinia cf. forficata 0,00 ± 0,00 0,00 ± 0,00 0,00 ± 0,00 0,00 ± 0,00
Boehmeria caudata Sw. 0,00 ± 0,00 0,02 ± 0,02 0,00 ± 0,00 0,01 ± 0,01
Bromeliaceae 0,02 ± 0,02 0,21 ± 0,19 0,12 ± 0,11 0,03 ± 0,03
Cabralea canjerana (Vell.) Mart. 0,00 ± 0,00 0,00 ± 0,00 0,02 ± 0,02 0,00 ± 0,00
Calypthranthes lucida Mart. ex DC. 0,08 ± 0,05 1,26 ± 0,81 0,17 ± 0,12 0,30 ± 0,21
Campomanesia cf. reitziana 0,02 ± 0,01 0,02 ± 0,02 0,13 ± 0,09 0,02 ± 0,02
Campomanesia cf. xanthocarpa 0,00 ± 0,00 0,32 ± 0,24 0,18 ± 0,47 0,38 ± 0,29
Casearia obliqua Spreng. 0,29 ± 0,08 0,12 ± 0,12 0,02 ± 0,02 0,02 ± 0,02
Casearia sylvestris Sw. 0,33 ± 0,12 0,02 ± 0,02 0,00 ± 0,00 0,02 ± 0,02
Cedrella fissilis Vell. 0,06 ± 0,03 1,33 ± 0,87 1,25 ± 0,35 0,69 ± 0,40
Chrysophyllum sp. 0,01 ± 0,00 0,00 ± 0,00 0,00 ± 0,00 0,00 ± 0,00
Chrysophyllum viride Mart. & Eichler 0,23 ± 0,09 0,42 ± 0,33 0,37 ± 0,19 0,22 ± 0,14
Clethra scabra Pers. 0,07 ± 0,04 0,61 ± 0,41 0,10 ± 0,07 0,43 ± 0,21
Colubrina glandulosa Perkins 0,10 ± 0,04 0,17 ± 0,15 0,00 ± 0,00 0,69 ± 0,94
Coussapoa microcarpa (Schott)
Rizzini 0,04 ± 0,03 0,18 ± 0,17 0,51 ± 0,27 0,25 ± 0,04
Cupania vernalis Cambess. 0,89 ± 0,16 3,84 ± 2,26 1,50 ± 0,52 1,66 ± 0,89
Dahlstedtia sp. 0,00 ± 0,00 0,11 ± 0,09 0,16 ± 0,10 0,11 ± 0,08
Dalechampia sp. 0,03 ± 0,03 1,08 ± 0,91 0,56 ± 0,07 0,53 ± 0,40
Diatenopteryx sorbifolia Radlk. 0,18 ± 0,07 0,33 ± 0,27 0,05 ± 0,03 0,49 ± 0,33
Dioscorea scabra Humb. & Bonpl. ex
Willd. 0,02 ± 0,01 0,12 ± 0,09 0,08 ± 0,05 0,37 ± 0,34
Dioscorea sp. 0,00 ± 0,00 0,00 ± 0,00 0,00 ± 0,00 0,03 ± 0,02
Duguetia sp. 0,00 ± 0,00 0,00 ± 0,00 0,01 ± 0,01 0,00 ± 0,00
Endlicheria paniculata (Spreng.)
J.F.Macbr.
0,00 ± 0,00 0,00 ± 0,00 0,00 ± 0,00 0,02 ± 0,02

51
Erythroxylum argentinum O.E.Schulz 0,14 ± 0,04 0,11 ± 0,10 0,00 ± 0,00 1,17 ± 0,74
Erythroxylum sp.1 0,02 ± 0,02 0,02 ± 0,02 0,07 ± 0,06 0,06 ± 0,05
Erythroxylum sp.2 0,01 ± 0,01 0,08 ± 0,08 0,04 ± 0,04 0,37 ± 0,31
Eugenia bacopari D.Legrand 0,00 ± 0,00 0,02 ± 0,02 0,00 ± 0,00 0,00 ± 0,00
Eugenia stigmatosa DC. 0,00 ± 0,00 0,00 ± 0,00 0,04 ± 0,02 0,00 ± 0,00
Ficus adhatodifolia Schott ex Spreng. 0,15 ± 0,07 4,07 ± 3,05 1,70 ± 1,10 1,84 ± 1,20
Ficus cestrifolia Schott ex Spreng. 0,89 ± 0,18 6,80 ± 4,94 3,76 ± 1,88 1,25 ± 1,01
Ficus eximia Schott 1,28 ± 0,38 0,57 ± 0,50 1,22 ± 0,49 0,61 ± 0,28
Guatteria australis A.St.-Hil. 0,00 ± 0,00 0,74 ± 0,44 0,30 ± 0,21 0,08 ± 0,06
Guatteria sp. 0,01 ± 0,00 0,00 ± 0,00 0,00 ± 0,00 0,00 ± 0,00
Handroanthus heptaphyllus (Vell.)
Mattos 0,14 ± 0,05 0,93 ± 0,93 0,48 ± 0,28 0,06 ± 0,06
Hieronyma alchorneoides Allemão 0,01 ± 0,01 0,00 ± 0,00 0,02 ± 0,02 0,00 ± 0,00
Hirtella hebeclada Moric. ex DC. 0,04 ± 0,03 0,09 ± 0,09 0,00 ± 0,00 0,03 ± 0,03
Ilex cf. brevicuspis 0,08 ± 0,03 0,03 ± 0,03 0,01 ± 0,01 0,03 ± 0,03
Ilex sp. 0,00 ± 0,00 0,12 ± 0,11 0,02 ± 0,01 0,03 ± 0,02
Inga cf. vera 0,04 ± 0,03 0,03 ± 0,03 0,00 ± 0,00 0,07 ± 0,07
Inga marginata Willd. 0,49 ± 0,14 0,02 ± 0,02 0,06 ± 0,05 0,00 ± 0,00
Lauraceae 0,03 ± 0,01 0,13 ± 0,06 0,01 ± 0,01 0,02 ± 0,02
Leandra sp. 0,01 ± 0,01 0,01 ± 0,01 0,00 ± 0,00 0,01 ± 0,01
Luehea divaricata Mart. & Zucc. 0,05 ± 0,02 0,00 ± 0,00 0,02 ± 0,08 0,02 ± 0,01
Machaerium stipitatum (DC.) Vogel 0,16 ± 0,05 0,00 ± 0,00 0,00 ± 0,00 0,01 ± 0,01
Maclura tinctoria (L.) D.Don ex
Steud. 0,07 ± 0,04 0,00 ± 0,00 0,00 ± 0,00 0,00 ± 0,00
Magnolia ovata (A.St.-Hil.) Spreng. 0,01 ± 0,01 2,91 ± 2,78 0,90 ± 0,67 0,19 ± 0,19
Marliera sp. 0,00 ± 0,00 0,07 ± 0,05 0,04 ± 0,03 0,06 ± 0,06
Matayba elaeagnoides Radlk. 0,00 ± 0,00 0,07 ± 0,07 0,00 ± 0,00 0,01 ± 0,01
Matayba guianensis Aubl. 0,12 ± 0,05 0,20 ± 0,20 0,03 ± 0,03 0,19 ± 0,12
Miconia cabucu Hoehne 0,00 ± 0,00 0,03 ± 0,03 0,00 ± 0,00 0,09 ± 0,09
Miconia cf. cinerascens 0,00 ± 0,00 0,00 ± 0,00 0,02 ± 0,01 0,00 ± 0,00
Miconia cinnamomifolia (DC.) Naudin 0,00 ± 0,00 0,32 ± 0,24 0,06 ± 0,06 0,38 ± 0,38
Microgramma cf. vacinifolia 0,06 ± 0,04 0,01 ± 0,01 0,00 ± 0,00 0,00 ± 0,00
Mikania campanulata Gardner 0,01 ± 0,01 0,00 ± 0,00 0,00 ± 0,00 0,05 ± 0,05
Mikania sp. 0,01 ± 0,01 0,15 ± 0,15 0,80 ± 0,41 0,02 ± 0,02
Molinedia cf. calodonta 0,00 ± 0,00 0,08 ± 0,06 0,32 ± 0,20 0,01 ± 0,01
Mollinedia schottiana (Spreng.)
Perkins 0,00 ± 0,00 0,42 ± 0,42 0,13 ± 0,08 0,24 ± 0,18
Myrcia sp. 0,00 ± 0,00 0,30 ± 0,27 0,24 ± 0,13 0,16 ± 0,12
Myrsine guianensis (Aubl.) Kuntze 0,00 ± 0,00 0,00 ± 0,00 0,00 ± 0,00 0,00 ± 0,00
Myrsine sp. 0,00 ± 0,00 0,05 ± 0,04 0,00 ± 0,00 0,01 ± 0,01
Myrtaceae 3 0,03 ± 0,02 0,01 ± 0,01 0,00 ± 0,00 0,02 ± 0,02
Myrtaceae 4 0,05 ± 0,03 0,02 ± 0,02 0,00 ± 0,00 0,03 ± 0,03
Myrtaceae 5 0,00 ± 0,00 0,08 ± 0,08 0,00 ± 0,00 0,02 ± 0,02
Myrtaceae 6 0,00 ± 0,00 1,66 ± 1,63 0,24 ± 0,17 0,11 ± 0,11
Nectandra lanceolata Nees 0,03 ± 0,02 0,02 ± 0,02 0,00 ± 0,00 0,00 ± 0,00
Nectandra oppositifolia Ness 0,00 ± 0,00 0,00 ± 0,00 0,00 ± 0,00 0,00 ± 0,00
Ocotea cf. pulchella 0,01 ± 0,01 0,00 ± 0,00 0,02 ± 0,02 0,01 ± 0,01
Ocotea cf. silvestris 0,00 ± 0,00 0,88 ± 0,79 0,73 ± 0,33 0,53 ± 0,27
Ocotea mandioccana A.Quinet 0,17 ± 0,07 0,05 ± 0,05 0,05 ± 0,05 0,03 ± 0,02

52
Ocotea sp.1 0,00 ± 0,00 0,02 ± 0,02 0,00 ± 0,00 0,00 ± 0,00
Ocotea sp.2 0,00 ± 0,00 0,01 ± 0,01 0,00 ± 0,00 0,01 ± 0,01
Ouratea parviflora (A.DC.) Baill. 0,00 ± 0,00 0,00 ± 0,00 0,00 ± 0,00 0,02 ± 0,02
Passiflora sp. 0,00 ± 0,00 0,13 ± 0,08 0,00 ± 0,00 0,01 ± 0,01
Pausandra morisiana (Casar.) Radlk. 0,37 ± 0,14 0,30 ± 0,26 0,07 ± 0,01 0,02 ± 0,01
Pouteria salicifolia (Spreng.) Radlk. 0,00 ± 0,00 0,47 ± 0,26 0,14 ± 0,07 0,23 ± 0,10
Pera glabrata (Schott) Poepp. ex
Baill. 0,15 ± 0,07 0,02 ± 0,02 0,03 ± 0,03 0,00 ± 0,00
Persea sp. 0,01 ± 0,01 0,20 ± 0,17 0,05 ± 0,05 0,04 ± 0,03
Piper aduncum L. 0,00 ± 0,00 0,15 ± 0,11 0,02 ± 0,02 0,11 ± 0,07
Piptocarpha cf. tomentosa 0,00 ± 0,00 0,26 ± 0,22 0,00 ± 0,00 0,10 ± 0,07
Poaceae 0,00 ± 0,00 0,02 ± 0,02 0,00 ± 0,00 0,00 ± 0,00
Posoqueria latifolia (Rudge) Schult. 0,06 ± 0,03 0,00 ± 0,00 0,00 ± 0,00 0,21 ± 0,13
Prunus myrtifolia (L.) Urb. 0,01 ± 0,01 0,00 ± 0,00 0,00 ± 0,00 0,02 ± 0,02
Psidium cattleianum Sabine 0,00 ± 0,00 0,00 ± 0,00 0,01 ± 0,01 0,01 ± 0,01
Roupala brasiliensis (Klotzsch) 0,02 ± 0,01 0,91 ± 0,43 0,33 ± 0,29 0,49 ± 0,21
Rubiaceae 0,00 ± 0,00 0,03 ± 0,02 0,00 ± 0,00 0,02 ± 0,02
Rudgea sp.1 0,00 ± 0,00 0,33 ± 0,27 0,05 ± 0,03 0,49 ± 0,33
Rudgea sp.2 0,00 ± 0,00 0,00 ± 0,00 0,05 ± 0,05 0,00 ± 0,00
Sapium glandulosum (L.) Morong 0,01 ± 0,01 0,00 ± 0,00 0,00 ± 0,00 0,01 ± 0,01
Schizolobium parahyba (Vell.) Blake 6,79 ± 1,04 7,69 ± 5,61 1,65 ± 9,80 11,0 ± 6,85
Sebastiana sp.2 0,01 ± 0,01 0,00 ± 0,00 0,07 ± 0,06 0,03 ± 0,03
Sebastiania sp.1 0,17 ± 0,06 0,03 ± 0,03 0,00 ± 0,00 0,01 ± 0,01
Serjania sp. 0,00 ± 0,00 0,02 ± 0,02 0,05 ± 0,04 0,01 ± 0,01
Sloanea guianensis (Aubl.) Benth. 0,00 ± 0,00 0,19 ± 0,16 0,33 ± 0,20 0,09 ± 0,07
Smilax sp. 0,00 ± 0,00 0,00 ± 0,00 0,04 ± 0,04 0,02 ± 0,02
Solanacea 0,00 ± 0,00 0,84 ± 0,73 0,04 ± 0,04 0,40 ± 0,18
Solanum cf. mauritianum 0,10 ± 0,05 0,08 ± 0,08 0,08 ± 0,07 0,05 ± 0,02
Solanum sp. 0,00 ± 0,00 0,00 ± 0,00 0,00 ± 0,16 0,00 ± 0,00
Sorocea bonplandii (Baill.)
W.C.Burger et al. 0,00 ± 0,00 0,04 ± 0,04 0,00 ± 0,00 0,00 ± 0,00
Strychnus trinervis (Vell.) Mart. 0,01 ± 0,01 0,08 ± 0,08 0,08 ± 0,07 0,05 ± 0,02
Symplocos sp. 0,00 ± 0,00 0,10 ± 0,09 0,07 ± 0,05 0,03 ± 0,03
Tetrorchidium rubrivenium Poepp. 0,23 ± 0,10 2,08 ± 1,22 0,26 ± 1,08 2,03 ± 0,89
Tillandsia sp. 0,00 ± 0,00 1,68 ± 1,20 1,21 ± 0,52 0,66 ± 0,42
Trema micrantha (L.) Blume 0,08 ± 0,04 0,39 ± 0,32 0,15 ± 0,08 0,22 ± 0,13
Trichilia cf. clausseni 0,03 ± 0,02 0,67 ± 0,58 0,18 ± 0,13 0,46 ± 0,34
Vernonanthura discolor (Spreng.)
H.Rob. 0,00 ± 0,00 0,00 ± 0,00 0,00 ± 0,00 0,00 ± 0,00
Virola bicuhyba (Schott ex Spreng.)
Warb. 0,51 ± 0,17 6,93 ± 5,49 1,65 ± 0,74 1,49 ± 1,34
Vriesea sp. 0,01 ± 0,01 0,00 ± 0,00 0,00 ± 0,00 0,00 ± 0,00
Xylopia brasiliensis Spreng. 0,00 ± 0,00 0,00 ± 0,00 0,00 ± 0,00 0,08 ± 0,07
Xylosma sp. 0,01 ± 0,01 0,00 ± 0,00 0,00 ± 0,00 0,00 ± 0,00
Zanthoxylum sp. 0,00 ± 0,00 0,19 ± 0,19 0,00 ± 0,00 0,06 ± 0,05
Indeterminada 1 0,06 ± 0,04 0,04 ± 0,03 0,00 ± 0,00 0,02 ± 0,01
Indeterminada 2 0,02 ± 0,01 0,00 ± 0,00 0,00 ± 0,00 0,00 ± 0,00
Indeterminada 3 0,00 ± 0,00 0,68 ± 0,65 0,49 ± 0,25 0,18 ± 0,15
indeterminada 4 0,20 ± 0,08 0,10 ± 0,10 0,04 ± 0,04 0,00 ± 0,00
Indeterminada 5 0,00 ± 0,00 0,01 ± 0,01 0,05 ± 0,04 0,03 ± 0,02
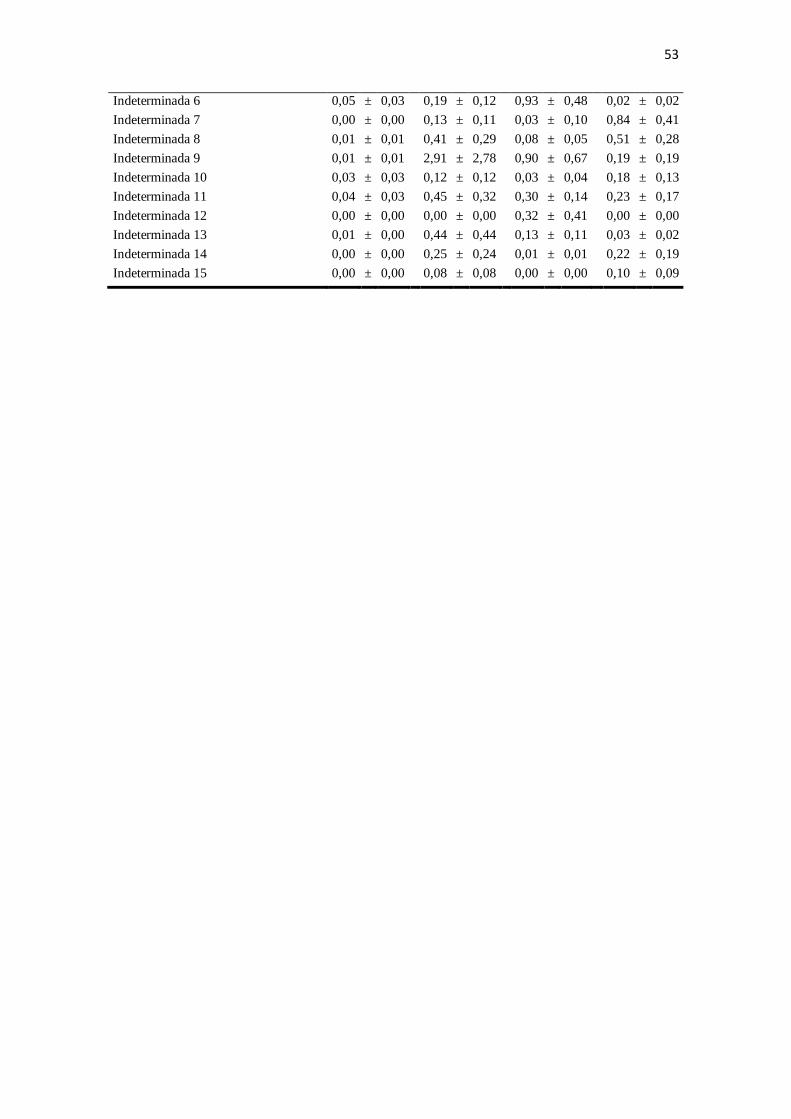
53
Indeterminada 6 0,05 ± 0,03 0,19 ± 0,12 0,93 ± 0,48 0,02 ± 0,02
Indeterminada 7 0,00 ± 0,00 0,13 ± 0,11 0,03 ± 0,10 0,84 ± 0,41
Indeterminada 8 0,01 ± 0,01 0,41 ± 0,29 0,08 ± 0,05 0,51 ± 0,28
Indeterminada 9 0,01 ± 0,01 2,91 ± 2,78 0,90 ± 0,67 0,19 ± 0,19
Indeterminada 10 0,03 ± 0,03 0,12 ± 0,12 0,03 ± 0,04 0,18 ± 0,13
Indeterminada 11 0,04 ± 0,03 0,45 ± 0,32 0,30 ± 0,14 0,23 ± 0,17
Indeterminada 12 0,00 ± 0,00 0,00 ± 0,00 0,32 ± 0,41 0,00 ± 0,00
Indeterminada 13 0,01 ± 0,00 0,44 ± 0,44 0,13 ± 0,11 0,03 ± 0,02
Indeterminada 14 0,00 ± 0,00 0,25 ± 0,24 0,01 ± 0,01 0,22 ± 0,19
Indeterminada 15 0,00 ± 0,00 0,08 ± 0,08 0,00 ± 0,00 0,10 ± 0,09

54
Capítulo 2
Relação entre a estrutura da comunidade de invertebrados
aquáticos com a composição e biomassa de detritos foliares em riachos
preservado e impactado
Relationship of aquatic invertebrate community structure to the
composition and biomass of leaf litter in preserved and impacted
streams
Leonardo Kleba Lisboa1,3
, Aurea Luiza Lemes da Silva2,3
, Ana Emília Siegloch3,
e Mauricio Mello Petrucio1,3
1Programa de Pós-graduação em Biologia Vegetal, Universidade Federal de
Santa Catarina - Trindade, Florianópolis, CEP 88010-970, Santa Catarina, Brasil
Email: [email protected]
2Programa de Pós-graduação em Ecologia, Universidade de Brasília, Darcy
Ribeiro, Brasília- DF - CEP 70910-900.
3Laboratório de Ecologia de Águas Continentais, Universidade Federal de Santa
Catarina - Trindade, Florianópolis, CEP 88010-970, Santa Catarina, Brasil

55
Resumo
Detritos vegetais alóctones depositados no estoque bêntico formam um dos
principais recursos energéticos de riachos florestados, mas pouca atenção tem sido dada
para a composição de folhas em riachos tropicais e sua relação com a comunidade de
invertebrados aquáticos. O presente trabalho avaliou a relação entre a estrutura da
comunidade de invertebrados aquáticos e a composição e biomassa de folhas do estoque
bêntico em dois riachos de Mata Atlântica com diferentes graus de conservação da
vegetação ripária ao longo de um ano. O estudo se baseou nas hipóteses de que (1) a
comunidade de invertebrados varia espacialmente de acordo com a composição e
quantidade dos recursos, e (2) as comunidades variam temporalmente em função da
variabilidade dos recursos. O riacho preservado apresentou maior riqueza tanto na
comunidade de invertebrados como na composição das folhas do estoque bêntico.
Chironomidae e Simuliidae foram as famílias mais abundantes nos dois ambientes. A
estrutura das comunidades e a composição do detrito foram significativamente
diferentes entre os dois riachos, e em ambas variaram temporalmente. A variação
espacial da comunidade de invertebrados foi influenciada pela riqueza de espécies
vegetais e quantidade de recursos, além de temperatura da água e condutividade.
Variações temporais foram influenciadas principalmente pela temperatura da água e
precipitação nos dois riachos, sendo que a biomassa de folhas foi uma variável
importante apenas para o riacho preservado. A variação temporal na comunidade do
riacho impactado foi mais pronunciada, provavelmente relacionada à menor integridade
da vegetação ripária. Sugerimos ainda, que a riqueza de espécies do detrito influenciou
a variação espacial das comunidades, mas parece não influenciar variações temporais. A
primeira hipótese foi acatada, e a segunda apenas em parte para o riacho preservado.
Palavras chave: variação espacial e temporal, estoque bêntico, Mata Atlântica,
integridade da vegetação ripária

56
Abstract
Allochtonous leaf litter in benthic stock is one of the main energetic resources in
forested streams, but little attention has been paid to leaf composition in tropical
streams and its relation with the aquatic invertebrate community. The present study
evaluated the relation between aquatic invertebrate community structure to the benthic
leaf litter composition and biomass in two Atlantic Rain Forest streams with different
riparian vegetation integrity states during a year. We hypothesized that (1) the
invertebrate community changes spatially according to the quantity and composition of
resources, and (2) communities change temporally in function of resource variability.
The preserved stream showed higher richness in invertebrate community and in benthic
leaf litter composition. Chironomidae and Simuliidae were the dominant families in
both streams. Community structure and leaf composition were significantly different
between the streams, and both showed temporal variation. Spatial invertebrate
communities variation was influenced by leaf species richness and resources quantity,
and also by water temperature and conductivity. Temporal variations were influenced
mainly by water temperature and precipitation in both streams, but just for the preserved
one the biomass of leaves was also important. Temporal variation was more pronounced
in the impacted stream, possibly related to the worse riparian vegetation integrity state.
We suggest that leaf litter richness influences community spatial variation, but seems to
not influence temporal variations. The first hypothesis was accepted, and the second
accepted just in part for the preserved stream.
Key-words: Spatio-temporal variation, benthic stock, Atlantic Forest, riparian
vegetation integrity

57
Introdução
Detritos vegetais alóctones representam uma das principais fontes de energia
para riachos de cabeceira, e têm papel crucial no suporte e estruturação das
comunidades aquáticas (Cummins et al, 1973; Cummins & Klug, 1979; Vannote et al.,
1980; Wallace et al., 1997). Ao entrarem nos riachos, esses detritos são agregados de
acordo com mecanismos de retenção do ecossistema (Richardson et al., 2005), e
acumulados de acordo com o regime do fluxo de água (Acuña et al., 2007) e taxas de
incorporação e processamento da matéria orgânica (Gonçalves. et al., 2006). A
composição do detrito alóctone está diretamente relacionada à composição da vegetação
ripária (Swan & Palmer, 2006; Kominoski et al., 2011), que por sua vez irá determinar a
qualidade e quantidade de subsídios de detritos que entram nos ecossistemas
(Kominoski et al., 2012). Assim, a vegetação ripária tem influência direta na
disponibilidade alimentar aos invertebrados aquáticos (Ardón & Pringle, 2008;
Kominoski et al., 2012), fornecendo folhas de diferentes qualidades que acabam por
representar diferentes tipos de recursos para insetos e micro consumidores (Ardón &
Pringle, 2008). Além disso, as comunidades aquáticas também utilizam indiretamente o
material vegetal depositado no leito de riachos (Carvalho & Uieda, 2010), que podem
servir como locais de refúgio e forrageio (Taniguchi & Tokeshi, 2004).
Estudos demonstram que a estrutura da comunidade de invertebrados aquáticos é
influenciada pela abundância de recursos (Burdett & Watts, 2009), tempo de exposição
dos detritos (Abelho, 2008), diversidade do substrato (Boyero, 2003) e heterogeneidade
do habitat (Taniguchi & Tokeshi, 2004; Brown, 2007). Segundo Swan & Palmer
(2006), misturas de folhas apresentam fatores tanto positivos (nutrientes disponíveis)
como negativos (compostos secundários), que direcionam a preferência e performance
de alimentação dos consumidores. Muitos invertebrados, principalmente pertencentes ao
grupo trófico dos fragmentadores (ex: Calamoceratidae, Trichoptera), obtêm energia e
nutrientes necessários colonizando e ingerindo preferencialmente folhas de maior
qualidade nutricional (Bastian et al., 2007; Janke & Trivinho-Strixino, 2007; Abelho,
2008; Graça & Cressa, 2010), em especial aquelas já previamente condicionadas pela
colonização por microrganismos (Bastian et al., 2007; Graça & Cressa, 2010).
De acordo com Wantzen et al. (2008), a diminuição da diversidade de plantas
vasculares em latitudes mais elevadas faz com que a composição do aporte de matéria
orgânica alóctone em riachos tenha variações substanciais entre ambientes temperados e

58
tropicais. Assim, a riqueza na composição de folhas do estoque bêntico tende a ser
muito maior em ambientes preservados tropicais (Wantzen et al., 2008; Boyero et al.,
2011), fazendo com que invertebrados neotropicais colonizem uma gama de espécies de
folhas muito maior (Bastian et al., 2007). Apesar disso, a diversidade de fragmentadores
é maior em ambientes temperados (Boyero et al., 2011), pois a qualidade dos detritos
tende a ser maior (Graça & Cressa, 2010), e a participação de microrganismos no
processo de decomposição menor (Abelho et al., 2005). Além disso, a dinâmica da
vegetação ripária e consequentemente padrões de acumulação de detritos vegetais no
leito de riachos são extremamente variáveis entre ambientes tropicais e temperados,
principalmente em função do clima e fenologia da vegetação (Webster & Meyer, 1997;
Wantzen et al., 2008). Em latitudes elevadas, as poucas espécies presentes tendem a
perder as folhas no outono, ocasionando grande incorporação de biomassa vegetal no
ambiente aquático em um mesmo período (Fisher & Likens, 1973; Pozo et al., 1997).
Em muitos ambientes tropicais, entretanto, a distribuição dos detritos vegetais tende a
ser fragmentada e imprevisível durante o ano (Bastian et al., 2007). Estudos sobre a
composição de espécies dos detritos foliares depositados no leito de riachos são
virtualmente inexistentes para sistemas lóticos tropicais (Bastian et al., 2007). O
consumo de detritos foliares por invertebrados aquáticos de pequenos riachos tem sido
bastante estudado, mas muitos estudos têm sido feitos experimentalmente com pacotes
de folhas de espécies abundantes do local (Yanoviak, 1999; Janke e Trivinho-Strixino,
2007; Burdett & Watts, 2009; Neiff et al., 2009), não contemplando a diversidade da
composição do material do estoque bêntico, e pouca atenção vem sendo dada para a
influência da diversidade de folhas nesse processo (Bastian et al., 2007).
A composição e retenção do aporte de detritos em rios e riachos também é
fortemente influenciada por mudanças no uso da terra (Kominoski & Rosemond, 2012),
sendo que a quantidade de matéria orgânica diminui drasticamente de áreas com
vegetação preservada para áreas abertas por influência antrópica (Afonso et al., 2000;
Carvalho & Uieda, 2010; Kiffney & Richardson, 2010). Neste contexto, Áreas de
Preservação Permanente (APPs) conseguem manter o fluxo energético em riachos de
cabeceira através da preservação da vegetação ripária (Kiffney & Richardson, 2010).
Como mudanças na composição dessa vegetação podem afetar os ecossistemas
aquáticos através de alterações na dinâmica de matéria orgânica (Kominoski &
Rosemond, 2012), diferentes graus de preservação da vegetação tendem a propiciar
mosaicos de folhiço com comunidades de invertebrados dissimilares, possivelmente

59
influenciada pela variação na quantidade e composição do detrito vegetal acumulado.
Nesse sentido, diferentes estágios sucessionais da vegetação ripária parecem influenciar
a distribuição dos invertebrados aquáticos (Bastian et al., 2007).
Partindo do princípio que a comunidade de invertebrados aquáticos está
diretamente associada às características dos recursos, e de que existe variação temporal
no aporte desses recursos, este estudo se baseou nas seguintes hipóteses: (1) há variação
na estrutura da comunidade de invertebrados entre riachos de vegetação ripária
preservada e impactada em função de diferenças na composição e quantidade do detrito
no estoque bêntico, e (2) a distribuição dos invertebrados aquáticos varia temporalmente
de acordo com a variabilidade na riqueza e biomassa dos recursos do estoque bêntico.
Material e métodos
Área de estudo
O município de Florianópolis está localizado em zona subtropical, e segundo a
classificação climática de Köeppen, o clima é do tipo Cfa, com ausência de estação
seca, chuvas distribuídas uniformemente durante o ano e verões rigorosos (Nascimento,
2002). A Bacia Hidrográfica da Lagoa do Peri (27° 43' 30” S e 48° 32' 18” W) possui
20,3 km2, e tem os mesmos limites do Parque Municipal da Lagoa do Peri, criado pela
Lei Municipal n° 1.828/81, situado ao sul da parte insular de Florianópolis (CECCA,
1996). Aproximadamente 75% da bacia é constituída pelo complexo cristalino pré-
cambriano, com relevo acidentado representado na forma de altos morros como o Morro
da Chapada (440 m), da Tapera (371 m), da Boa Vista (350 m) e do Peri (320 m)
(Santos et al., 1989). A maior parte desses terrenos cristalinos, situados no norte, oeste e
sul da bacia, são cobertos por densa vegetação de Mata Atlântica (Floresta Ombrófila
Densa) em estádios de regeneração avançado. A parte leste da bacia é constituída
geologicamente por terrenos de sedimentação quaternária, que separam a lagoa do
Oceano Atlântico através de uma estreita faixa de 250 m de largura. Essa região é
coberta principalmente por vegetação litorânea característica de restinga e de praia, com
algumas áreas ocupadas por casas e pequenos trechos de plantação de Eucalipto.
Representando um dos dois principais tributários da bacia hidrográfica, o rio
Cachoeira Grande drena uma área de 1,66 km2, tem suas nascentes situadas na cota de
280 m de altitude e percorre 1,17 km até desaguar na Lagoa do Peri (Santos et al.,
1989). Esse rio possui densa vegetação ripária justamente por se estender através de um

60
dos estádios mais desenvolvidos de vegetação secundária da ilha, podendo até mesmo
abrigar áreas de floresta primária (Caruso, 1990). O experimento foi realizado perto da
foz do rio, em um trecho de 3ª ordem, com densa cobertura de dossel. Em virtude disso,
esse ambiente foi considerado como preservado.
Já o rio Ribeirão Grande drena uma área de 6,98 km2, tem suas nascentes por
volta de 285 m de altitude e percorre 4,6 km até desaguar na lagoa (Santos et al., 1989).
Esse rio atravessa uma área de pequenas propriedades rurais, cujos agricultores
transformaram parte da vegetação ripária com o plantio de espécies nativas (Manihot
esculenta) e exóticas (Saccharum sp., Musa sp. e Bambusa sp.), e em determinados
trechos com a supressão para pecuária. No trecho estudado deste rio, também de 3ª
ordem, também ocorrem espécies nativas remanescentes na vegetação, sendo que estas
não são representativas e têm abundância muito menor que as espécies exóticas. Para o
presente estudo esse ambiente foi considerado como impactado.
Metodologia experimental
O experimento foi realizado nos dois principais tributários da Bacia Hidrográfica
da Lagoa do Peri, os quais foram classificados como “preservado” e “impactado” de
acordo com o grau de preservação da vegetação ripária. Foram escolhidos cinco pontos
de coleta em cada rio, separados por uma distância média de 20 m entre si. Os detritos
vegetais depositados no leito do rio (estoque bêntico) foram coletados com auxílio de
um amostrador tipo Surber, tamanho 0,3 x 0,3 m e abertura de malha de 0,25 mm. Em
cada ponto de coleta foram realizadas três amostragens aleatórias onde se encontrava
depósito de folhiço, totalizando 15 amostras mensais por rio. As coletas foram
realizadas ao longo de onze meses, compreendendo o período de agosto de 2010 a junho
de 2011.
Todas as amostras foram levadas a laboratório e lavadas em peneiras de 0,25
mm para retenção de invertebrados aquáticos. Após a lavagem, o material vegetal foi
seco em estufa a 60ºC até a estabilização do peso, e então determinado o peso seco
através de balança analítica de precisão 0,01 g. Os detritos vegetais foram separados
qualitativamente entre folhas, galhos e partes reprodutivas. As folhas foram
identificadas até o menor grupo taxonômico possível com bibliografia adequada (Sobral
et al., 2006) e com auxílio de especialista. Os animais retidos na peneira foram triados,
acondicionados em frascos com álcool 70% e posteriormente identificados até o menor

61
nível taxonômico possível utilizando-se chaves de identificação disponíveis (Fernández
& Domínguez, 2001; Mugnai et al., 2010) e ajuda de especialistas.
Simultaneamente às coletas de estoque bêntico foram medidas in situ variáveis
ambientais da água como oxigênio dissolvido (mg.l-1
), temperatura (°C), condutividade
(μS.cm-1
) e pH com auxílio de sonda específica WTW-Multi350i. A velocidade da água
(m.s-1
) foi obtida através do método do flutuador com cinco réplicas por ponto,
profundidade (m) medida com trena em dez réplicas por ponto, e a temperatura do ar
medida com termômetro. Dados de precipitação referentes ao município foram
fornecidos pela Empresa de Pesquisa Agropecuária e Extensão Rural de Santa Catarina
(EPAGRI).
Análise dos dados
A fim de se verificar a existência de diferenças entre os dois rios nos parâmetros
da água, foi realizado o teste de Mann-Whitney com as matrizes de dados abióticos
brutos. Para verificar a existência de variação espacial entre os pontos amostrais dentro
de cada rio, e de variação temporal entre os meses quanto aos parâmetros ambientais,
foi realizado o teste de Kruskal-Wallis com os parâmetros individuais brutos para cada
ambiente. Foram utilizados testes não-paramétricos pois as variáveis da água não se
encaixaram na distribuição normal de acordo com o teste de Shapiro-Wilk. Os testes
acima citados foram realizados com auxílio do programa estatístico Statistica 7.
A riqueza da comunidade aquática e o esforço amostral foram avaliados através
da curva de acumulação de espécies referente a todo período de estudo, calculada a
partir da matriz bruta de abundância de indivíduos com auxílio do programa EstimateS
8.2.0, e plotada utilizando-se o método de Mao Tau. Variações na estrutura da
comunidade entre os dois rios foram verificadas através da análise de escalonamento
multidimensional (MDS), obtida a partir da matriz de similaridade de Bray Curtis das
médias de abundância de todo período transformados em log (x+1). Similaridades entre
as comunidades foram testadas estatisticamente através de uma análise de similaridade
(ANOSIM), tendo como fatores as duas áreas. Quando diferenças significativas foram
detectadas (a partir da ANOSIM), a análise de porcentagem de similaridade (SIMPER)
foi utilizada para identificar os táxons responsáveis pela diferença observada nas
comunidades (Clarke & Warwick, 2001). A análise SIMPER não é um teste estatístico,
mas uma análise exploratória que gera um ranking de porcentagem de contribuição dos

62
táxons mais importantes para determinar a diferença entre os grupos de amostras, i.e.,
neste caso, riachos com diferente complexidade da vegetação ripária.
A variação espacial da composição dos detritos foliares foi analisada através de
um MDS, calculado a partir das matrizes de distância Euclidiana dos dados de presença
e ausência das espécies vegetais dos dois riachos. Similaridades entre os riachos foram
testadas através de uma ANOSIM, tendo como fatores as duas áreas.
Para verificar a influência das variáveis ambientais na estrutura da comunidade
entre as duas áreas, foi feita uma Análise de Correspondência Canônica (CCA),
calculada através das médias mensais de abundância e das médias mensais de variáveis
ambientais transformadas em log (x+1), analisando-se conjuntamente os dois ambientes
estudados. As variáveis foram incluídas na análise através de seleção manual,
excluindo-se aquelas não significativas (Monte Carlo, p > 0,05), representando-se as
amostragens por ambiente e por mês de coleta. A significância foi verificada com o
teste de Monte Carlo (p < 0,05). A CCA foi feita com auxílio do programa Canoco for
Windows 4.5.
Variações temporais da comunidade e da composição dos detritos foliares foram
analisadas em separado para cada riacho, e testadas a partir de ANOSIMs utilizando-se
as mesmas matrizes do MDS, e considerando como fatores os meses. A fim de se
evidenciar os principais fatores ambientais responsáveis pela variação temporal da
comunidade, foi feita uma análise de BIOENV para cada riacho em separado
relacionando-se a matriz de similaridade de Bray Curtis das médias de abundância
transformada (log x + 1), com a matriz de distância Euclidiana dos dados abióticos
transformados (log x + 1) e padronizados. Para análises multivariadas, além dos
parâmetros físico-químicos da água e variáveis meteorológicas, foram considerados
como fatores ambientais as categorias do estoque bêntico vegetal como “folhas”,
“galhos”, “reprodutivos”, “miscelânea” e “total de matéria orgânica” medidas em
biomassa média (g), e “riqueza” de espécies na composição do detrito. Todas as análises
de MDS, ANOSIM, SIMPER e BIOENV foram feitas com auxilio do programa
estatístico Primer 6 (Clarke & Gorley, 2006).
Resultados
Variação espacial
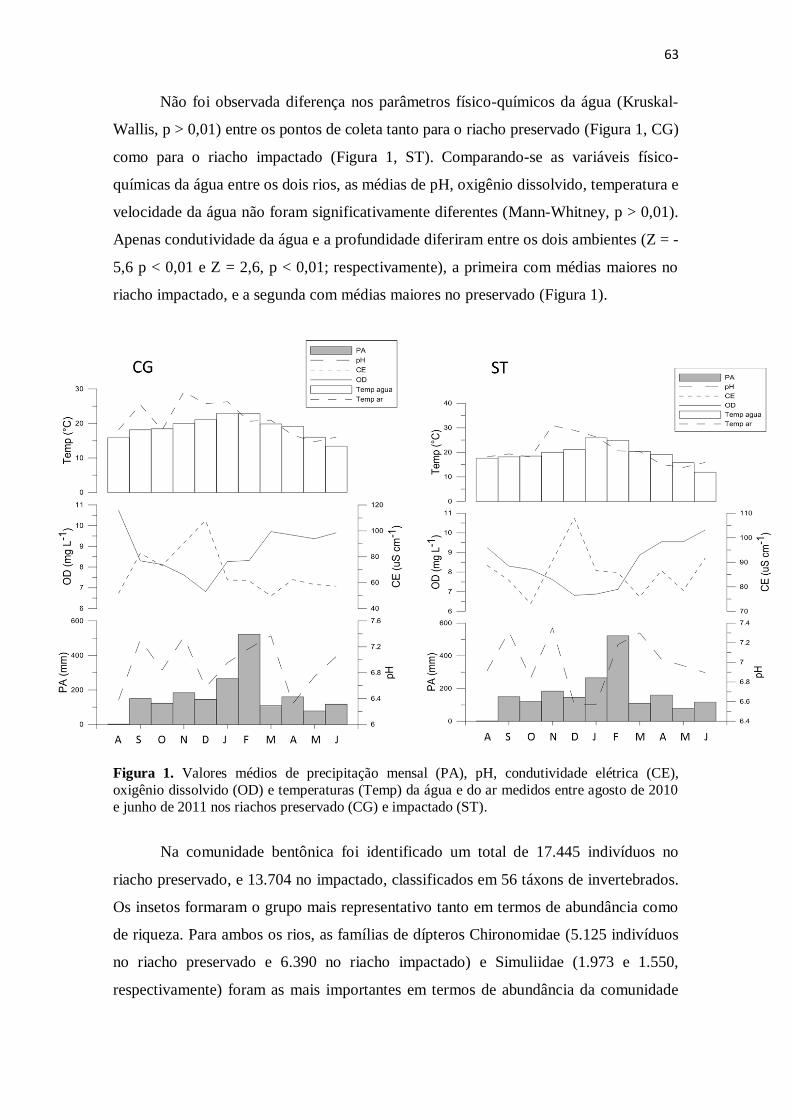
63
Não foi observada diferença nos parâmetros físico-químicos da água (Kruskal-
Wallis, p > 0,01) entre os pontos de coleta tanto para o riacho preservado (Figura 1, CG)
como para o riacho impactado (Figura 1, ST). Comparando-se as variáveis físico-
químicas da água entre os dois rios, as médias de pH, oxigênio dissolvido, temperatura e
velocidade da água não foram significativamente diferentes (Mann-Whitney, p > 0,01).
Apenas condutividade da água e a profundidade diferiram entre os dois ambientes (Z = -
5,6 p < 0,01 e Z = 2,6, p < 0,01; respectivamente), a primeira com médias maiores no
riacho impactado, e a segunda com médias maiores no preservado (Figura 1).
Figura 1. Valores médios de precipitação mensal (PA), pH, condutividade elétrica (CE),
oxigênio dissolvido (OD) e temperaturas (Temp) da água e do ar medidos entre agosto de 2010
e junho de 2011 nos riachos preservado (CG) e impactado (ST).
Na comunidade bentônica foi identificado um total de 17.445 indivíduos no
riacho preservado, e 13.704 no impactado, classificados em 56 táxons de invertebrados.
Os insetos formaram o grupo mais representativo tanto em termos de abundância como
de riqueza. Para ambos os rios, as famílias de dípteros Chironomidae (5.125 indivíduos
no riacho preservado e 6.390 no riacho impactado) e Simuliidae (1.973 e 1.550,
respectivamente) foram as mais importantes em termos de abundância da comunidade
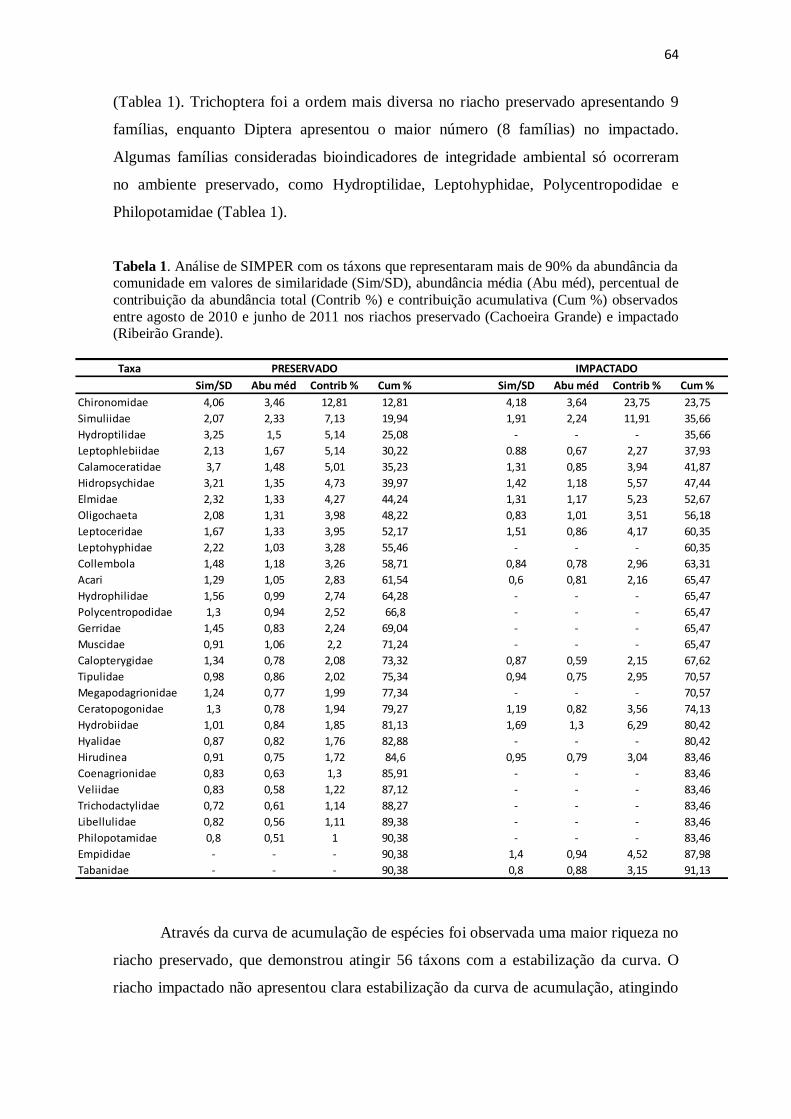
64
(Tablea 1). Trichoptera foi a ordem mais diversa no riacho preservado apresentando 9
famílias, enquanto Diptera apresentou o maior número (8 famílias) no impactado.
Algumas famílias consideradas bioindicadores de integridade ambiental só ocorreram
no ambiente preservado, como Hydroptilidae, Leptohyphidae, Polycentropodidae e
Philopotamidae (Tablea 1).
Tabela 1. Análise de SIMPER com os táxons que representaram mais de 90% da abundância da comunidade em valores de similaridade (Sim/SD), abundância média (Abu méd), percentual de
contribuição da abundância total (Contrib %) e contribuição acumulativa (Cum %) observados
entre agosto de 2010 e junho de 2011 nos riachos preservado (Cachoeira Grande) e impactado (Ribeirão Grande).
Através da curva de acumulação de espécies foi observada uma maior riqueza no
riacho preservado, que demonstrou atingir 56 táxons com a estabilização da curva. O
riacho impactado não apresentou clara estabilização da curva de acumulação, atingindo
Taxa
Sim/SD Abu méd Contrib % Cum % Sim/SD Abu méd Contrib % Cum %
Chironomidae 4,06 3,46 12,81 12,81 4,18 3,64 23,75 23,75
Simuliidae 2,07 2,33 7,13 19,94 1,91 2,24 11,91 35,66
Hydroptilidae 3,25 1,5 5,14 25,08 - - - 35,66
Leptophlebiidae 2,13 1,67 5,14 30,22 0.88 0,67 2,27 37,93
Calamoceratidae 3,7 1,48 5,01 35,23 1,31 0,85 3,94 41,87
Hidropsychidae 3,21 1,35 4,73 39,97 1,42 1,18 5,57 47,44
Elmidae 2,32 1,33 4,27 44,24 1,31 1,17 5,23 52,67
Oligochaeta 2,08 1,31 3,98 48,22 0,83 1,01 3,51 56,18
Leptoceridae 1,67 1,33 3,95 52,17 1,51 0,86 4,17 60,35
Leptohyphidae 2,22 1,03 3,28 55,46 - - - 60,35
Collembola 1,48 1,18 3,26 58,71 0,84 0,78 2,96 63,31
Acari 1,29 1,05 2,83 61,54 0,6 0,81 2,16 65,47
Hydrophilidae 1,56 0,99 2,74 64,28 - - - 65,47
Polycentropodidae 1,3 0,94 2,52 66,8 - - - 65,47
Gerridae 1,45 0,83 2,24 69,04 - - - 65,47
Muscidae 0,91 1,06 2,2 71,24 - - - 65,47
Calopterygidae 1,34 0,78 2,08 73,32 0,87 0,59 2,15 67,62
Tipulidae 0,98 0,86 2,02 75,34 0,94 0,75 2,95 70,57
Megapodagrionidae 1,24 0,77 1,99 77,34 - - - 70,57
Ceratopogonidae 1,3 0,78 1,94 79,27 1,19 0,82 3,56 74,13
Hydrobiidae 1,01 0,84 1,85 81,13 1,69 1,3 6,29 80,42
Hyalidae 0,87 0,82 1,76 82,88 - - - 80,42
Hirudinea 0,91 0,75 1,72 84,6 0,95 0,79 3,04 83,46
Coenagrionidae 0,83 0,63 1,3 85,91 - - - 83,46
Veliidae 0,83 0,58 1,22 87,12 - - - 83,46
Trichodactylidae 0,72 0,61 1,14 88,27 - - - 83,46
Libellulidae 0,82 0,56 1,11 89,38 - - - 83,46
Philopotamidae 0,8 0,51 1 90,38 - - - 83,46
Empididae - - - 90,38 1,4 0,94 4,52 87,98
Tabanidae - - - 90,38 0,8 0,88 3,15 91,13
PRESERVADO IMPACTADO

65
44 táxons de invertebrados (Figura 2). Os ambientes preservado e impactado
demonstraram significativa diferenciação quanto à estrutura da comunidade de
invertebrados aquáticos (ANOSIM, R = 0,69, p < 0,001). A ordenação de
escalonamento multidimensional dos dados de abundância demonstrou claramente essa
separação dos ambientes (Figura 3), com um valor de “stress” relativamente baixo, de
0,18, indicando que o MDS foi uma representação bidimensional aceitável das
distâncias multivariadas entre os dados da matriz de similaridade (Benstead & Huryn,
2011).
Figura 2. Curva de acumulação de
espécies baseada no índice de
“MaoTau”, utilizando o número de famílias de invertebrados aquáticos
por amostragens, nos riachos
preservado (círculos) e impactado (triângulos), amostradas entre
agosto de 2010 a junho de 2011 na
Bacia Hidrográfica da Lagoa do Peri, SC. Pontilhados representam
intervalo de confiança de 95%.
Comparando a composição taxonômica foliar do estoque bêntico entre os dois
riachos, foi observada uma diferenciação significativa (ANOSIM, R = 0,6, p < 0,001),
representada através da ordenação do MDS (Figura 4). Além da composição, essa
diferenciação também se deu na riqueza e biomassa vegetal (Figura 5). A presença de
espécies exóticas foi observada apenas no rio impactado, e, apesar de representarem
uma fração pequena da quantidade total de matéria orgânica, formaram uma fração
significativa da biomassa total de folhas presentes no estoque bêntico (Figura 5).

66
Figura 3. Ordenação de escalonamento multidimensional (MDS) obtida pela matriz de
similaridade de Bray Curtis dos dados de abundância da comunidade de invertebrados aquáticos, transformada por log(x+1), dos riachos preservado (CG) e impactado (ST), coletados
mensalmente entre agosto de 2010 e junho de 2011 na Bacia Hidrográfica da Lagoa do Peri, SC.
Figura 4. Ordenação de escalonamento multidimensional (MDS) dos dados de composição
específica dos detritos foliares feita através da similaridade de distância Euclidiana da matriz de presença/ausência de espécies vegetais dos riachos preservado (CG) e impactado (ST) coletados
entre agosto de 2010 e junho de 2011 na Bacia Hidrográfica da Lagoa do Peri, SC.

67
Figura 5. Quantidade de detritos vegetais do estoque bêntico
representados em biomassa total de
recursos (MO, matéria orgânica), e
riqueza de espécies vegetais do folhiço coletado, observadas entre
agosto de 2010 e junho de 2011
nos riachos preservado (CG) e impactado (ST). Barras
quadriculadas representam a
quantidade de biomassa de folhas referentes a espécies exóticas –
encontradas apenas no rio ST.
De acordo com a CCA, as principais variáveis ambientais responsáveis por
diferenças na estruturação das comunidades de invertebrados entre os riachos foram
riqueza de espécies do estoque bêntico, biomassa de galhos e a condutividade elétrica,
enquanto a temperatura parece explicar diferenças entre os meses, tendo em vista que
esteve positivamente relacionada com os meses quentes de ambos os riachos (Figura 6).
Apesar da explicação dos eixos da CCA na estruturação da comunidade ter sido
relativamente baixa (eixo 1 = 25,7%; eixo 2 = 9,2%), a análise mostrou significativa
correspondência (Monte Carlo, p < 0,05) entre as variáveis ambientais e a estrutura da
comunidade. A partir da CCA pode-se observar primeiramente uma separação entre os
dois ambientes através do eixo 1. Os vetores de riqueza de espécies do detrito e de
biomassa de galhos foram positivamente relacionados às amostragens do riacho
preservado, enquanto que a temperatura da água se mostrou positivamente relacionada
aos meses quentes do riacho impactado, assim como a condutividade mais relacionada
às amostragens desse ambiente (Figura 6).
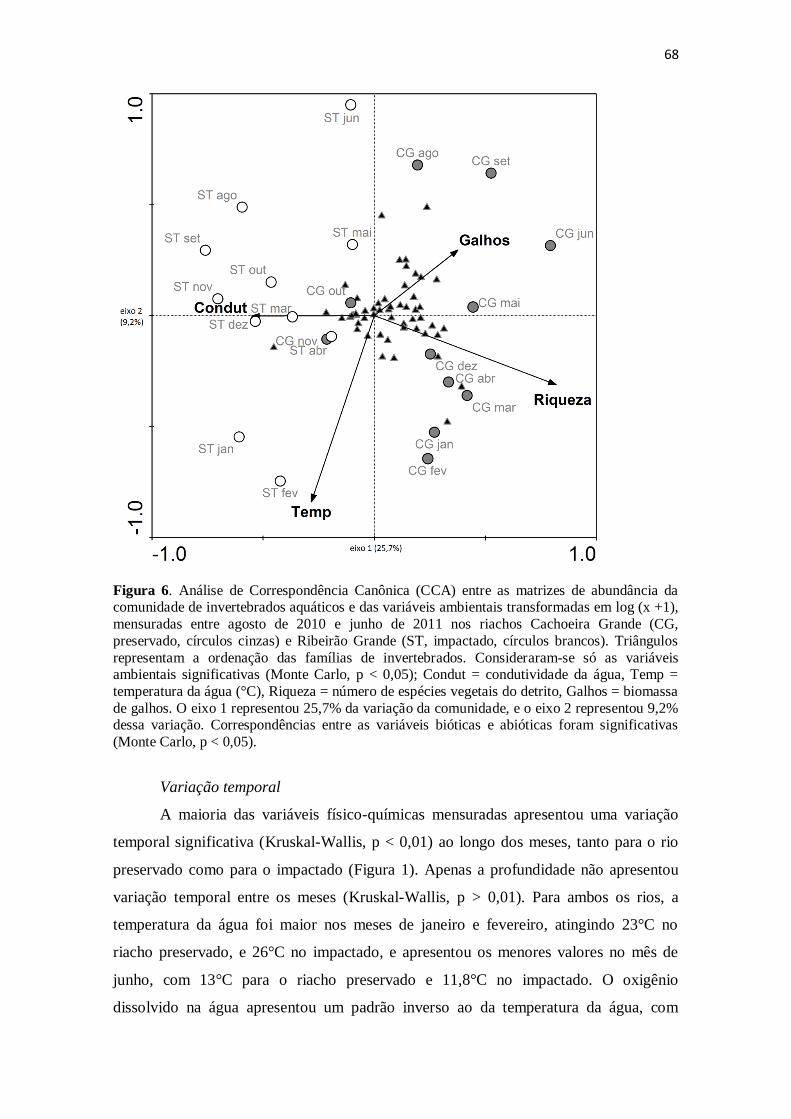
68
Figura 6. Análise de Correspondência Canônica (CCA) entre as matrizes de abundância da comunidade de invertebrados aquáticos e das variáveis ambientais transformadas em log (x +1),
mensuradas entre agosto de 2010 e junho de 2011 nos riachos Cachoeira Grande (CG,
preservado, círculos cinzas) e Ribeirão Grande (ST, impactado, círculos brancos). Triângulos
representam a ordenação das famílias de invertebrados. Consideraram-se só as variáveis ambientais significativas (Monte Carlo, p < 0,05); Condut = condutividade da água, Temp =
temperatura da água (°C), Riqueza = número de espécies vegetais do detrito, Galhos = biomassa
de galhos. O eixo 1 representou 25,7% da variação da comunidade, e o eixo 2 representou 9,2% dessa variação. Correspondências entre as variáveis bióticas e abióticas foram significativas
(Monte Carlo, p < 0,05).
Variação temporal
A maioria das variáveis físico-químicas mensuradas apresentou uma variação
temporal significativa (Kruskal-Wallis, p < 0,01) ao longo dos meses, tanto para o rio
preservado como para o impactado (Figura 1). Apenas a profundidade não apresentou
variação temporal entre os meses (Kruskal-Wallis, p > 0,01). Para ambos os rios, a
temperatura da água foi maior nos meses de janeiro e fevereiro, atingindo 23°C no
riacho preservado, e 26°C no impactado, e apresentou os menores valores no mês de
junho, com 13°C para o riacho preservado e 11,8°C no impactado. O oxigênio
dissolvido na água apresentou um padrão inverso ao da temperatura da água, com
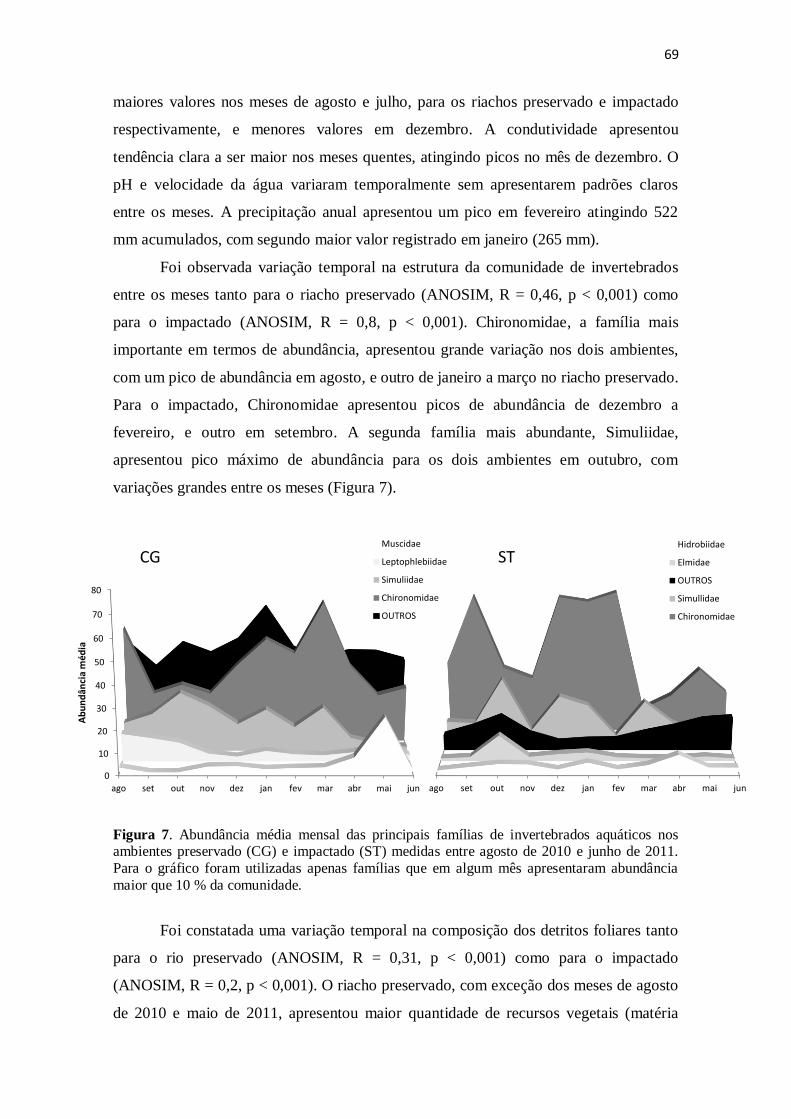
69
maiores valores nos meses de agosto e julho, para os riachos preservado e impactado
respectivamente, e menores valores em dezembro. A condutividade apresentou
tendência clara a ser maior nos meses quentes, atingindo picos no mês de dezembro. O
pH e velocidade da água variaram temporalmente sem apresentarem padrões claros
entre os meses. A precipitação anual apresentou um pico em fevereiro atingindo 522
mm acumulados, com segundo maior valor registrado em janeiro (265 mm).
Foi observada variação temporal na estrutura da comunidade de invertebrados
entre os meses tanto para o riacho preservado (ANOSIM, R = 0,46, p < 0,001) como
para o impactado (ANOSIM, R = 0,8, p < 0,001). Chironomidae, a família mais
importante em termos de abundância, apresentou grande variação nos dois ambientes,
com um pico de abundância em agosto, e outro de janeiro a março no riacho preservado.
Para o impactado, Chironomidae apresentou picos de abundância de dezembro a
fevereiro, e outro em setembro. A segunda família mais abundante, Simuliidae,
apresentou pico máximo de abundância para os dois ambientes em outubro, com
variações grandes entre os meses (Figura 7).
Figura 7. Abundância média mensal das principais famílias de invertebrados aquáticos nos ambientes preservado (CG) e impactado (ST) medidas entre agosto de 2010 e junho de 2011.
Para o gráfico foram utilizadas apenas famílias que em algum mês apresentaram abundância
maior que 10 % da comunidade.
Foi constatada uma variação temporal na composição dos detritos foliares tanto
para o rio preservado (ANOSIM, R = 0,31, p < 0,001) como para o impactado
(ANOSIM, R = 0,2, p < 0,001). O riacho preservado, com exceção dos meses de agosto
de 2010 e maio de 2011, apresentou maior quantidade de recursos vegetais (matéria
0
10
20
30
40
50
60
70
80
ago set out nov dez jan fev mar abr mai jun
Ab
un
dân
cia
méd
ia
Muscidae
Leptophlebiidae
Simuliidae
Chironomidae
OUTROS
CG
ago set out nov dez jan fev mar abr mai jun
Hidrobiidae
Elmidae
OUTROS
Simullidae
Chironomidae
ST

70
orgânica) disponíveis se comparado com o impactado, além de apresentar uma maior
riqueza média de espécies na composição de folhas do estoque bêntico (Figura 5). O
riacho preservado apresentou um total de 93 espécies identificadas ao longo do período
de estudo, contra apenas 62 espécies no impactado (apêndice).
A partir da CCA, pode se observar secundariamente uma separação temporal
através do eixo 2, com a agregação entre os meses quentes de dezembro, janeiro e
fevereiro na parte inferior relacionados positivamente à temperatura da água, em
oposição aos meses frios de junho, agosto e setembro na parte superior (Figura 6). A
análise de BIOENV mostrou uma correlação significativa (Rho = 0,529; p < 0,01) entre
as variáveis ambientais de temperatura da água, velocidade da corrente, precipitação e
biomassa de folhas com a matriz temporal da estrutura da comunidade do riacho
preservado. Para o riacho impactado, a BIOENV demonstrou que as variáveis de
condutividade, temperatura da água e a precipitação foram as mais relacionadas (Rho =
0,556; p < 0,01) à matriz temporal da comunidade.
Discussão
Variação espacial
A evidente diferenciação na estrutura da comunidade de invertebrados e na
composição dos detritos foliares do estoque bêntico entre os riachos preservado e
impactado, aliada à correlação significativa da mesma com riqueza e biomassa do
detrito, sugere que tais variáveis foram preditoras importantes da variação espacial da
comunidade de invertebrados na área de estudo. Corroborando com Afonso et al.
(2000), o riacho com vegetação ripária preservada apresentou maior quantidade e
riqueza de recursos. Estudos mostram que uma maior diversidade de detritos foliares
suporta uma comunidade de invertebrados colonizadores mais diversa (Boyero, 2003).
Ainda, Swan & Palmer (2006) observaram que a mistura de folhas influencia
diretamente o crescimento de consumidores, afetando taxas de decomposição e
consumo e consequentemente a comunidade como um todo. A partir disso, os
resultados apresentados na CCA nos levam a crer que ambientes com vegetação ripária
em bom estado de conservação têm a estrutura da comunidade de invertebrados
aquáticos primariamente influenciada pela riqueza de folhas e quantidade de recursos
disponíveis no estoque bêntico, enquanto que a perda de espécies e redução na

71
quantidade de detritos podem fazer com que a comunidade seja regulada por fatores
ambientais como temperatura e condutividade da água.
Para abundância de grupos funcionais específicos (detritívoros), a palatabilidade
das folhas se mostra mais importante que a diversidade de recursos (Janke & Trivinho-
Strixino, 2007; Abelho, 2008; Shah, 2010). Como evidenciou Bastian et al. (2007),
fragmentadores de rios tropicais colonizam uma ampla gama de espécies, priorizando a
qualidade das mesmas em detrimento à composição específica. Como a qualidade das
folhas muda de acordo com as espécies, a diferenciação na composição dos detritos
foliares entre os dois ambientes tratados aqui tende a representar detritos de diferentes
qualidades, influenciando na estruturação das comunidades de invertebrados. Outro
fator que aponta para influência das características do recurso na diferenciação espacial
das comunidades, se dá devido ao fato de que as variáveis ambientais pouco diferiram
entre os dois ambientes.
Os dois riachos apresentaram uma homogeneidade nas características da água
dos trechos estudados, e se mostraram bastante similares quanto a esses parâmetros
físico-químicos, com diferenças observadas apenas na condutividade e profundidade. A
condutividade apresentou valores maiores no rio impactado, provavelmente por se tratar
de um ambiente com alguma influência antrópica, cujas pequenas propriedades rurais
devem ter participação na incorporação pontual de efluentes na água e consequente
aumento de íons. Concentrações de oxigênio e valores de pH encontrados aqui não
foram limitantes para a comunidade de invertebrados aquáticos.
A alta abundância, principalmente da família Chironomidae, provavelmente
esteve associada a altos níveis de abundância de recurso, corroborando com o
encontrado por Burdett & Watts (2009). Essa família é frequentemente dominante nos
sistemas lóticos de Mata Atlântica (Moulton & Magalhães, 2003; Milesi et al., 2009;
Oliveira & Nessimian, 2010; Rosa et al., 2011). Apesar do predomínio de
Chironomidae nos dois ambientes, pode-se observar maior dominância dessa família no
ambiente impactado, corroborando com o encontrado por Moulton & Magalhães
(2003). Como esclarece Roque et al. (2007), a família Chironomidae inclui a maioria
dos grupos funcionais de invertebrados e é capaz de colonizar inúmeros habitats. Nesse
sentido, a maior riqueza encontrada na comunidade do riacho preservado, assim como a
presença de táxons bioindicadores sensíveis (Bispo & Oliveira, 2007), provavelmente
estão relacionadas às características de substrato, já que um substrato mais diverso
promove a ausência de táxons dominantes em uma comunidade (Boyero, 2003; Shah,

72
2010). Em pequenos riachos de Mata atlântica, a riqueza de grupos sensíveis tende a
aumentar com a disponibilidade de habitats, estando diretamente relacionada à
integridade da vegetação ripária (Milesi et al., 2009). Já para segunda família mais
abundante, Simuliidae, a quantidade de recursos parece ser menos importante que
fatores estruturais (tamanho do rio, largura, velocidade da água, descarga, estabilidade
do substrato e regime de chuvas) para estruturação da comunidade em nascentes
Neotropicais (Grillet & Barrera, 1997).
Variação temporal
A variação temporal da comunidade de invertebrados foi influenciada por
variáveis distintas em cada ambiente. Os resultados sugerem que a precipitação e a
temperatura da água têm forte influência na variação temporal da comunidade para
ambos ambientes. De fato, a distribuição temporal de insetos aquáticos em riachos
subtropicais é influenciada por variações na temperatura da água (Yan & Li, 2007;
Tejerina & Malizia, 2012), e períodos de intensa precipitação alteram drasticamente a
dinâmica de riachos (Acuña et al., 2007). Para o riacho preservado, porém, a quantidade
de folhas também foi responsável por parte dessa variação, o que não ocorreu para o
ambiente impactado, que por sua vez mostrou influência significativa da condutividade.
Isso sugere que a comunidade de invertebrados aquáticos de ambientes com vegetação
ripária preservada varia temporalmente, entre outros fatores, em conjunto com a
quantidade de folhas presentes no estoque bêntico. É sabido que a abundância de
recursos alimentares e de refúgio é um dos principais fatores determinantes da
estruturação da comunidade (Burdett & Watts, 2009), e que riachos preservados de
Mata Atlântica subtropical têm grande variação temporal na quantidade e composição
do detrito do estoque bêntico ao longo do ano, resultado em parte da forte influência da
precipitação na incorporação desse material ao ecossistema (Lisboa et al., 2012 –
Capítulo 1). Já a comunidade de invertebrados do riacho impactado variou
temporalmente devido a fatores climáticos (temperatura, precipitação e condutividade).
De fato, a precipitação teve uma variação temporal muito grande no presente
estudo, marcada com picos de chuva imprevisíveis. Os valores de temperatura da água
tiveram padrões esperados para ambientes subtropicais, com variação temporal
condizente com as tendências de meses quentes no verão e frios no inverno
(Hennemann & Petrucio, 2011). A amplitude de variação da temperatura da água
também se mostrou maior no rio impactado, provavelmente devido à supressão de parte

73
de sua vegetação ripária e consequente maior exposição à incidência solar. Assim, a
principal diferença dos parâmetros ambientais entre os dois riachos se dá nas
características de integridade das matas ciliares.
A variação temporal mais pronunciada na comunidade de invertebrados do rio
impactado possivelmente se deu devido ao menor número de espécies vegetais no
detrito do estoque bêntico. É sabido que a heterogeneidade espacial do habitat reduz a
variabilidade temporal na comunidade de insetos de riachos (Brown, 2003). Apesar da
maioria dos estudos com heterogeneidade em riachos utilizarem como critérios
diferentes categorias de substrato, sugerimos aqui que as mais de 93 espécies vegetais
encontradas no estoque bêntico do riacho preservado representam maior
heterogeneidade no substrato, relativa às variações morfológicas e possivelmente
nutricionais das folhas. Dessa forma, os dados apresentados aqui corroboram com
Boyero (2003), onde a diversidade do substrato está positivamente relacionada à
abundância e riqueza da comunidade de invertebrados de um rio. É claro que outros
fatores são determinantes na estruturação e distribuição temporal dos invertebrados
aquáticos em riachos, como o ciclo de vida dos táxons (Johnson et al., 2012), a
complexidade do habitat (Taniguchi & Tokeshi, 2004) e mesmo a heterogeneidade
espacial ligada à conectividade (Neiff et al. 2009).
Em resumo, a estrutura da comunidade de invertebrados aquáticos variou de
acordo com diferentes graus de integridade da vegetação ripária, principalmente através
da influência da composição e quantidade do detrito foliar depositado no estoque
bêntico, e da proteção conferida pela vegetação ripária à variação de temperatura da
água. Assim, a hipótese de que a estrutura da comunidade de invertebrados difere entre
riachos preservado e impactado em função de diferenças na composição, riqueza e
quantidade do detrito no estoque bêntico foi corroborada. Enquanto que a estrutura da
comunidade variou ao longo do tempo de forma diferente ao se considerar sistemas com
distintos graus de integridade da vegetação ripária. No riacho preservado, a estrutura da
comunidade variou temporalmente em função da variabilidade na quantidade de detritos
foliares no estoque bêntico, e de variáveis ambientais como temperatura e precipitação.
No riacho impactado, a variação temporal da comunidade se deu em decorrência da
maior instabilidade ocasionada pela retirada da vegetação ripária, que acentuou o efeito
das chuvas e da temperatura sobre a comunidade. Além disso, a comunidade de
invertebrados aquáticos variou temporalmente de forma mais acentuada no riacho de
vegetação impactada. Esse estudo sugere experimentações específicas em campo sobre

74
influencia da riqueza de folhas do estoque bêntico na comunidade de invertebrados,
assim como análises da qualidade do detrito para um melhor detalhamento e discussão
aprofundada.
Agradecimentos
À bolsa de estudos concedida pela Coordenação de Aperfeiçoamento de Pessoal
de Nível Superior (CAPES) que possibilitou o desenvolvimento da pesquisa.
Gostaríamos de agradecer em especial ao professor José Francisco Gonçalves Júnior
pelas considerações pertinentes e críticas construtivas ao manuscrito. Ao suporte
estrutural fornecido através da Universidade Federal de Santa Catarina (UFSC), do
Laboratório de Biologia e Cultivo de Peixes de Água Doce (LAPAD) da UFSC, do
Parque Municipal da Lagoa do Peri e do departamento de Ecologia e Zoologia (ECZ) da
UFSC. À EPAGRI pelos dados fornecidos.
Referências capítulo 2
Abelho, M., Cressa, C. & Graça, M. A. S., 2005. Microbial biomass, respiration,
and decomposition of Hura crepitans L. (Euphorbiaceae) leaves in a tropical stream.
Biotropica 37: 397-402.
Abelho, M., 2008. Effects of Leaf Litter Species on Macroinvertebrate
Colonization during Decomposition in a Portuguese Stream. International Review of
Hydrobiology 93: 358-372.
Acuña, V., Giorgi, A., Muñoz, I., Sabater, F. & Sabater, S., 2007.
Meteorological and riparian influences on organic matter dynamics in a forested
Mediterranean stream. Journal of the North American Benthological Society 26:54-69.
Afonso, A. A. O., Henry, R. & Rodella, R. C. S. M., 2000. Allochthonous matter
input in two different stretches of a headstream (Itatinga, São Paulo, Brazil). Brazilian
Archives of Biology and Technology 43: 335-343.

75
Ardón, M. & Pringle, C. M., 2008. Do secondary compounds inhibit microbial-
and insect-mediated leaf breakdown in a tropical rainforest stream, Costa Rica?
Oecologia 155: 311-323.
Bastian, M., Boyero, L., Jackes, B. R. & Pearson, R. G., 2007. Leaf litter
diversity and shredder preferences in an Australian tropical rain-forest stream. Journal
of Tropical Ecology 23: 219–229.
Benstead, J. P. & Huryn, A. D., 2011. Extreme seasonality of litter breakdown in
an arctic spring-fed stream is driven by shredder phenology, not temperature.
Freshwater Biology, doi:10.1111/j.1365-2427.2011.02635.x
Bispo, P. C. & Oliveira, L. G., 2007. Diversity and structure of Ephemeroptera,
Plecoptera and Trichoptera (Insecta) assemblages from riffles in mountain streams of
Central Brazil. Revista Brasileira de Zoologia 24: 283–293.
Boyero, L., 2003. The quantification of local substrate heterogeneity in streams
and its significance for macroinvertebrate assemblages. Hydrobiologia 449: 161-168.
Boyero, L., Perason, R. G., Dudgeon, D., Graça, M. A. S., Gessner, M. O. et al.,
2011. Global distribution of a key trophic guild contrasts with common latitudinal
diversity patterns. Ecology 92: 1839–1848.
Brown, B. L., 2003. Spatial heterogeneity reduces temporal variability in stream
insect communities. Ecology Letters 6: 316-325.
Brown, B. L., 2007. Habitat heterogeneity and disturbance influence patterns of
community temporal variability in a small temperate stream. Hydrobiologia 586: 93-
106.
Burdett, A. S. & Watts, R. J., 2009. Modifying living space: an experimental
study of the influences of vegetation on aquatic invertebrate community structure.
Hydrobiologia 618: 161-173.

76
Caruso, M. M. L., 1990. O desmatamento na Ilha de Santa Catarina de 1500
aos dias atuais. Ed. da UFSC, Florianópolis, BR. 158p.
Carvalho, E. M. & Uieda, V. S., 2010. Input of litter in deforested and forested
areas of a tropical headstream. Brazilian Journal of Biology 70, 283-288.
CECCA/FNMA, 1996. Uma cidade numa ilha – relatório sobre os problemas
sócio-ambientais da Ilha de Santa Catarina. Ed. Insular, Florianópolis. 248p.
Clarke K. R. & Gorley R. N., 2006. Primer v6: user manual/tutorial. PRIMER-
E, Plymouth.
Clarke K. R. & Warwick., R. M., 2001. Change in marine communities: an
approach to statistical analysis and interpretation. Plymouth, PRIMER-E, UK, 2nd ed.,
172p.
Cummins, K. W., Petersen, R. C., Howard, F. O., Wuycheck, J. C. & Holt, V. I.,
1973. The utilization of leaf litter by stream detritivores. Ecology 54: 336-345.
Cummins, K. W. & Klug, M. J., 1979. Feeding ecology of stream invertebrates.
Annual Review of Ecology, Evolution and Systematics 10: 147-172.
Fernández, H. R. & Dominguéz, E., 2001. Guia para determinación de los
artropodos bentônicos Sudamericanos. UNT, Tucumán: 282 p.
Fisher, S. G. & Likens, G. E., 1973. Energy flow in Bear Brook, New
Hampshire: an integrative approach to stream ecosystem metabolism. Ecological
Monographs, 43: 421-439.
Gonçalves Jr., J. F., França, J. S. & Callisto, M., 2006. Dynamics of
allochthonous organic matter in a tropical Brazilian headstream. Brazilian Archives of
Biology and Technology 49: 967-973.

77
Graça, M. A. S. & Cressa, M., 2010. Leaf quality of some tropical and temperate
tree species as food resource for stream shredders. International Review of
Hydrobiology 95: 27-41.
Grillet, M. A. & Barrera, R., 1997. Spatial and temporal abundance, substrate
partitioning and species co-occurrence in a guild of Neotropical blackflies (Diptera:
Simuliidae). Hydrobiologia 345: 197–208.
Hennemann, M. C. & Petrucio, M. M., 2011. Spatial and temporal dynamic of
trophic relevant parameters in a subtropical coastal lagoon in Brazil. Environmental
Monitoring Assessment, DOI 10.1007/s10661-010-1833-5.
Janke, H. & Trivinho-Strixino, S., 2007. Colonization of leaf litter by aquatic
macroinvertebrates: a study in a low order tropical stream. Acta Limnologica
Brasiliensia 19: 109-115.
Johnson, R. C., Carreiro, M. M., Jin, H-S & Jack, J. D., 2012. Within-year
temporal variation and life-cycle seasonality affect stream macroinvertebrate
community structure and biotic metrics. Ecological Indicators 13: 206-214.
Kiffney, P. M. & Richardson, J. S., 2010. Organic matter inputs into headwater
streams of southwestern British Columbia as a function of riparian reserves and time
since harvesting. Forest Ecology and Management 260: 1931–1942.
Kominoski, J. S. & Rosemond, A. D., 2012. Conservation from the bottom up:
forecasting effects of global change on dynamics of organic matter and management
needs for river networks. Freshwater Science 31: 51–68.
Kominoski, J. S., Larrañaga, S. & Richardson, J. S., 2012. Invertebrate feeding
and emergence timing vary among streams along a gradient of riparian forest
composition. Freshwater Biology : doi:10.1111/j.1365-2427.2012.02740.x.

78
Kominoski J. S., Marczak L. B. & Richardson J. S., 2011. Riparian forest
composition affects stream litter decomposition despite similar microbial and
invertebrate communities. Ecology 92: 151–159.
Lisboa, L. K., da Silva, A. L. L., Siegloch, A. E. et al., 2012. Composition and
dynamics of allochtonous organic matter in a subtropical Atlantic Rain Forest stream.
Em submissão.
Milesi, S. V., Biasi, C., Restello, R. M. & Hepp, L. U., 2009. Distribution of
benthic macroinvertebrates in Subtropi cal streams (Rio Grande do Sul, Brazil). Acta
Limnologica Brasiliensia 21: 419-429.
Moulton, T. P. & Magalhães, S. A. P., 2003. Responses of leaf processing to
impacts in streams in Atlantic Rain Forest, Rio de Janeiro, Brazil – a test of the
biodiversity-ecosystem functioning relationship?. Brazilian Journal of Biology. 63: 87-
95.
Mugnai, R., Nessimian, J. L. & Baptista, D. F., 2010. Manual de identificação
de macroinvertebrados aquáticos do Estado do Rio de Janeiro. Technical Books Rio de
Janeiro: 176 p.
Nascimento, R. 2002. Atlas Ambiental de Florianópolis. Instituto Larus,
Florianópolis. 35p.
Neiff, A. P. de., Galassi, M. E. & Fransceschini, M. C., 2009. Invertebrate
assemblages associated with leaf litter in three floodplain wetlands of the Paraná River.
Wetlands, 29: 896-906.
Oliveira, A. L. H. de & Nessimian, J. L., 2010. Spatial distribution and
functional feeding groups of aquatic insect communities in Serra da Bocaina streams,
southeastern Brazil. Acta Limnologica Brasiliensia 22: 424-441.
Pozo, J., González, E., Díez, J. R., & Elósegui, A., 1997. Leaf-litter budgets in
two contrasting forested streams. Limnetica, 13(2): 77-84.

79
Richardson, J. S., Bilby R. E. & Bondar, C. A., 2005. Organic matter dynamics
in small streams of the Pacific Northwest. Journal of the American Water Resources
Association 41: 921-934.
Roque, F. O., Trivinho-Strixino, S., Milan, L. & Leite, J. G., 2007. Chironomid
species richness in low-order streams in the Brazilian Atlantic Forest: a first
approximation through a Bayesian approach. Journal of the North American
Benthological Society 26: 221-231.
Rosa, B. F. J. V., Silva, M. V. D. da, Oliveira, V. C. de, Martins, R. T. & Alves,
R. da G., 2011. Macroinvertebrates associated with bryophyta in a first-order Atlantic
Forest stream. Zoologia 28: 351–356.
Santos, G. F., Silva, J. T. N., Mendoça, M. & Veado, R. W. ad-V., 1989. Análise
Ambiental da Lagoa do Peri. Geosul 8: 101-123.
Shah, V., 2010. Colonization of Leaf Litter by Benthic Macroinvertebrates in
Redwood Creek of Mt.Tamalpais State Park, California. Spring, 1-21.
Sobral, M., Jarenkow, J. A., Brack, P., Irgang, B., Larocca, J. & Rodrigues, R.
S., 2006. Flora arbórea e arborescente do Rio Grande do Sul, Brasil. RiMA/Novo
Ambiente, São Carlos. 350p.
Swan, C. M. & Palmer, M. A., 2006. Composition of speciose leaf litter alters
stream detritivore growth, feeding activity and leaf breakdown. Oecologia 147: 469–
478.
Taniguchi, H. & Tokeshi, M., 2004. Effects of habitat complexity on benthic
assemblages in a variable environment. Freshwater Biology 49: 1164–1178.
Tejerina, E. G. & Malizia, A., 2012. Chironomidae (Diptera) larvae assemblages
differ along an altitudinal gradient and temporal periods in a subtropical montane stream
in Northwest Argentina. Hydrobiologia, 686: 41-54.

80
Vannote, R. L., Minshall, G. W., Cummins, K. W., Sedell, J. R. & Cushing, C.
E., 1980. The river continuum concept. Canadian Journal of Fisheries and Aquatic
Sciences 37: 130–137.
Yan, Y. & Li, X., 2007. Production dynamics and life cycle of dominant
chironomids (Diptera, Chironomidae) in a subtropical stream in China: Adaptation to
variable flow conditions in summer and autumn. Chinese Journal of Oceanology and
Limnology, 25: 330-342.
Yanoviak, S. P., 1999. Effects of leaf litter species on macroinvertebrate
community properties and mosquito yield in Neotropical tree hole microcosms.
Oecologia 120: 147-155.
Wallace, J. B., Eggert, S. L., Mayer, J. L. & Webster, J. R., 1997. Multiple
trophic levels of a forest stream linked to terrestrial litter inputs. Science 277:102-104.
Wantzen, K. M, Yule, C. M., Mathooko, J. M. & Pringle, C. M., 2008. Organic
Matter Processing in Tropical Streams. In: Dudgeon, D. & Cressa, C. (Org). Tropical
Stream Ecology. Elsevier, p. 44-64.
Webster, J.R. & J.L. Meyer. (1997). Organic matter budgets for streams: a
synthesis. Journal of the North American Benthological Society 16: 141–161.

81
Apêndice capítulo 2
Composição das folhas do estoque bêntico nos rios preservado (CG) e impactado (ST)
mensuradas de agosto de 2010 a junho de 2011, especificando-se as médias mensais (g.m-2
) e a
porcentagem de cada táxon no total mensurado. * representa espécies exóticas.
Família Espécie
CG ST
média % média %
Annonaceae Annona sylvatica A.St.-Hil. 1,10 2,49 0,23 0,69
Annona sericea Dunal 0,00 0,01 - -
Guatteria australis A.St.-Hil. 0,01 0,02 - -
Guatteria sp. 0,02 0,05 - -
Aquifoliaceae Ilex cf. brevicuspis 0,01 0,02 0,00 0,00
Asteraceae Mikania campanulata Gardner 0,02 0,04 0,02 0,05
Piptocarpha cf. tomentosa - - 1,33 3,98
Vernonia discolor (Spreng.) H.Rob. - - 0,38 1,15
Bignoniacceae Handroanthus heptaphyllus (Vell.) Mattos 0,64 1,44 - -
Bromeliaceae Bromeliaceae 0,08 0,19 0,02 0,07
Cannabaceae Trema micrantha (L.) Blume 0,07 0,16 0,01 0,03
Chrysobalanaceae Hirtella hebeclada Moric. ex DC. 0,07 0,16 0,03 0,08
Clethraceae Clethra scabra Pers. 0,22 0,50 - -
Dilleniaceae Davila cf. rugosa 0,10 0,22 0,15 0,46
Dioscoreaceae Dioscorea scabra Humb. & Bonpl. ex Willd. 0,07 0,15 0,05 0,14
Elaeocarpaceae Sloanea guianensis (Aubl.) Benth. 0,08 0,19 - -
Erythroxylaceae Erythroxylum argentinum O.E.Schulz 0,08 0,19 - -
Erythroxylum sp.1 0,08 0,19 - -
Erythroxylum sp.2 0,05 0,10 - -
Erythroxylum sp.3 0,06 0,13 - -
Euphorbiaceae Alchornea glandulosa Poepp. & Endl. 1,67 3,76 4,81 14,41
Alchornea triplinervia (Spreng.) Müll.Arg. 0,01 0,01 - -
Dalechampia sp. 0,07 0,17 0,01 0,02
Pausandra morisiana (Casar.) Radlk. 0,28 0,63 0,01 0,04
Sapium glandulosum (L.) Morong 0,01 0,02 - -
Sebastiania sp. 0,40 0,89 0,03 0,10
Tetrorchidium rubrivenium Poepp. 0,36 0,82 0,17 0,51
Fabaceae Bauhinia cf. forficata - - 0,12 0,36
Dahlstedtia sp. 0,21 0,47 - -
Inga cf. vera 0,08 0,18 - -
Inga marginata Willd. 1,12 2,52 - -
Machaerium stipitatum (DC.) Vogel 0,04 0,09 - -
Schyzolobium parabyba (Vell.) Blake 8,38 18,93 - -
Lamiaceae Aegephila sp. 0,02 0,04 - -
Lauraceae Lauraceae 0,18 0,41 0,06 0,18
Nectandra lanceolata Ness 0,95 2,15 0,14 0,43
Nectandra opositifolia Ness 0,33 0,74 0,03 0,10
Ocotea cf. silvestris 0,14 0,32 1,99 5,95
Ocotea mandioccana A.Quinet 0,26 0,59 0,03 0,08
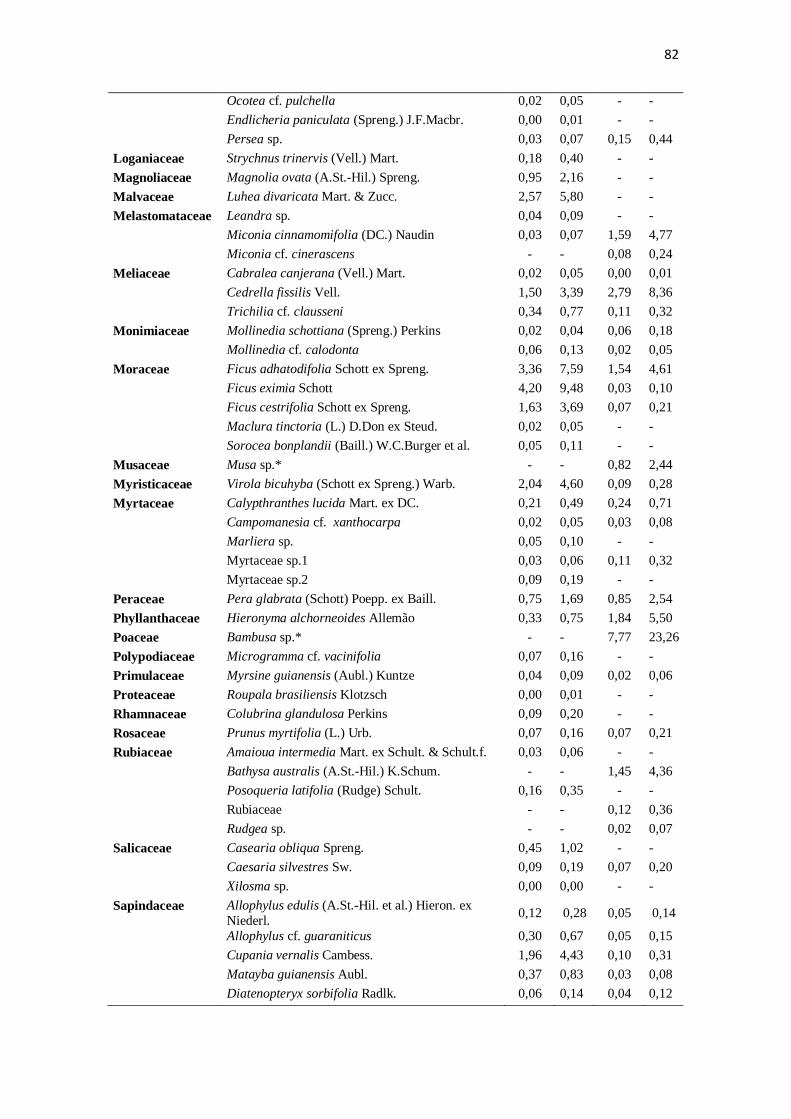
82
Ocotea cf. pulchella 0,02 0,05 - -
Endlicheria paniculata (Spreng.) J.F.Macbr. 0,00 0,01 - -
Persea sp. 0,03 0,07 0,15 0,44
Loganiaceae Strychnus trinervis (Vell.) Mart. 0,18 0,40 - -
Magnoliaceae Magnolia ovata (A.St.-Hil.) Spreng. 0,95 2,16 - -
Malvaceae Luhea divaricata Mart. & Zucc. 2,57 5,80 - -
Melastomataceae Leandra sp. 0,04 0,09 - -
Miconia cinnamomifolia (DC.) Naudin 0,03 0,07 1,59 4,77
Miconia cf. cinerascens - - 0,08 0,24
Meliaceae Cabralea canjerana (Vell.) Mart. 0,02 0,05 0,00 0,01
Cedrella fissilis Vell. 1,50 3,39 2,79 8,36
Trichilia cf. clausseni 0,34 0,77 0,11 0,32
Monimiaceae Mollinedia schottiana (Spreng.) Perkins 0,02 0,04 0,06 0,18
Mollinedia cf. calodonta 0,06 0,13 0,02 0,05
Moraceae Ficus adhatodifolia Schott ex Spreng. 3,36 7,59 1,54 4,61
Ficus eximia Schott 4,20 9,48 0,03 0,10
Ficus cestrifolia Schott ex Spreng. 1,63 3,69 0,07 0,21
Maclura tinctoria (L.) D.Don ex Steud. 0,02 0,05 - -
Sorocea bonplandii (Baill.) W.C.Burger et al. 0,05 0,11 - -
Musaceae Musa sp.* - - 0,82 2,44
Myristicaceae Virola bicuhyba (Schott ex Spreng.) Warb. 2,04 4,60 0,09 0,28
Myrtaceae Calypthranthes lucida Mart. ex DC. 0,21 0,49 0,24 0,71
Campomanesia cf. xanthocarpa 0,02 0,05 0,03 0,08
Marliera sp. 0,05 0,10 - -
Myrtaceae sp.1 0,03 0,06 0,11 0,32
Myrtaceae sp.2 0,09 0,19 - -
Peraceae Pera glabrata (Schott) Poepp. ex Baill. 0,75 1,69 0,85 2,54
Phyllanthaceae Hieronyma alchorneoides Allemão 0,33 0,75 1,84 5,50
Poaceae Bambusa sp.* - - 7,77 23,26
Polypodiaceae Microgramma cf. vacinifolia 0,07 0,16 - -
Primulaceae Myrsine guianensis (Aubl.) Kuntze 0,04 0,09 0,02 0,06
Proteaceae Roupala brasiliensis Klotzsch 0,00 0,01 - -
Rhamnaceae Colubrina glandulosa Perkins 0,09 0,20 - -
Rosaceae Prunus myrtifolia (L.) Urb. 0,07 0,16 0,07 0,21
Rubiaceae Amaioua intermedia Mart. ex Schult. & Schult.f. 0,03 0,06 - -
Bathysa australis (A.St.-Hil.) K.Schum. - - 1,45 4,36
Posoqueria latifolia (Rudge) Schult. 0,16 0,35 - -
Rubiaceae - - 0,12 0,36
Rudgea sp. - - 0,02 0,07
Salicaceae Casearia obliqua Spreng. 0,45 1,02 - -
Caesaria silvestres Sw. 0,09 0,19 0,07 0,20
Xilosma sp. 0,00 0,00 - -
Sapindaceae Allophylus edulis (A.St.-Hil. et al.) Hieron. ex
Niederl. 0,12 0,28 0,05 0,14
Allophylus cf. guaraniticus 0,30 0,67 0,05 0,15
Cupania vernalis Cambess. 1,96 4,43 0,10 0,31
Matayba guianensis Aubl. 0,37 0,83 0,03 0,08
Diatenopteryx sorbifolia Radlk. 0,06 0,14 0,04 0,12
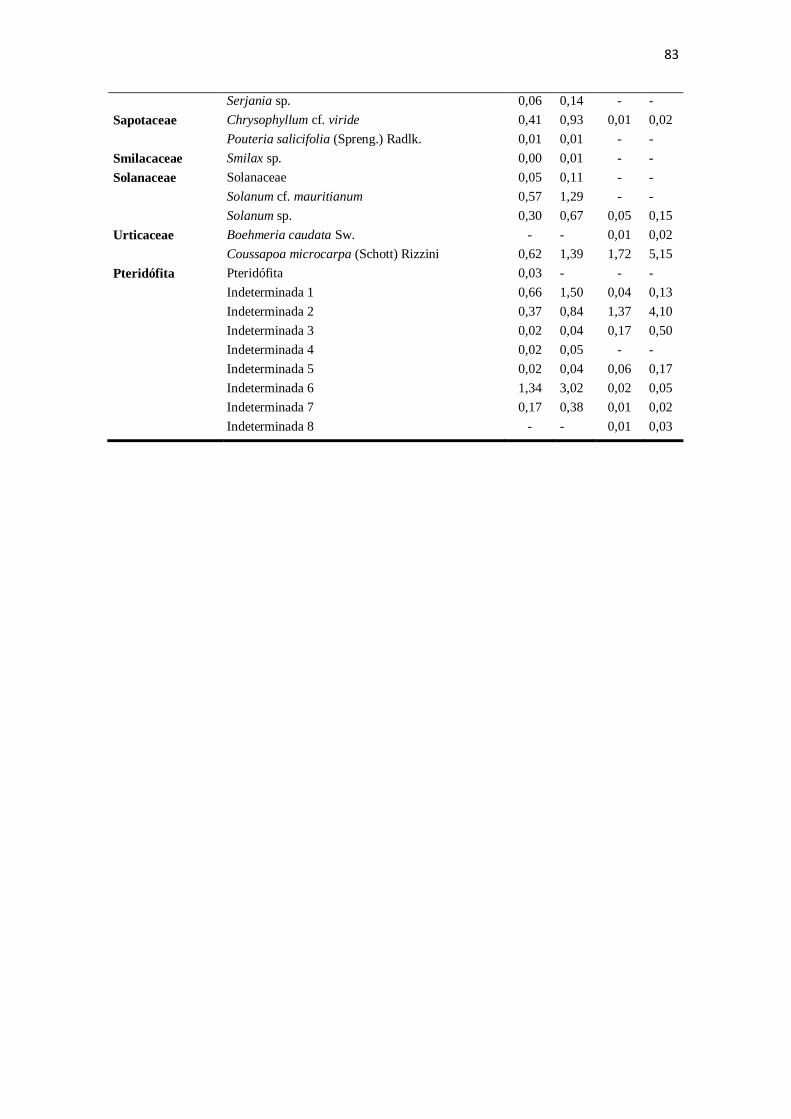
83
Serjania sp. 0,06 0,14 - -
Sapotaceae Chrysophyllum cf. viride 0,41 0,93 0,01 0,02
Pouteria salicifolia (Spreng.) Radlk. 0,01 0,01 - -
Smilacaceae Smilax sp. 0,00 0,01 - -
Solanaceae Solanaceae 0,05 0,11 - -
Solanum cf. mauritianum 0,57 1,29 - -
Solanum sp. 0,30 0,67 0,05 0,15
Urticaceae Boehmeria caudata Sw. - - 0,01 0,02
Coussapoa microcarpa (Schott) Rizzini 0,62 1,39 1,72 5,15
Pteridófita Pteridófita 0,03 - - -
Indeterminada 1 0,66 1,50 0,04 0,13
Indeterminada 2 0,37 0,84 1,37 4,10
Indeterminada 3 0,02 0,04 0,17 0,50
Indeterminada 4 0,02 0,05 - -
Indeterminada 5 0,02 0,04 0,06 0,17
Indeterminada 6 1,34 3,02 0,02 0,05
Indeterminada 7 0,17 0,38 0,01 0,02
Indeterminada 8 - - 0,01 0,03

84
Considerações Finais
A dinâmica da vegetação ripária exerce uma forte influência sobre os
ecossistemas de riachos florestados de cabeceira, sendo essencial na incorporação de
recursos alimentares, que também funcionam como locais de forrageio e refúgio para
fauna de invertebrados aquáticos. Desse modo, ela tem um papel determinante na
estruturação das comunidades aquáticas e na manutenção da integridade funcional
desses ecossistemas. Entretanto, há uma carência de estudos abordando esses assuntos
em riachos de Mata Atlântica, principalmente em ambientes subtropicais. Em se
tratando de um bioma considerado um Hot-Spot de biodiversidade, é necessário um
aprofundamento e melhor descrição dessas relações para o melhor entendimento da
funcionalidade desses sistemas, assim como auxiliar em esforços de conservação e
recuperação desse ambiente.
Os resultados deste trabalho demostram que a dinâmica da matéria orgânica
(MO) em riachos de cabeceira de Mata Atlântica subtropical é controlada
primariamente por períodos de intensa precipitação, que ocasionam picos de aporte de
detritos para os sistemas lóticos através do carreamento lateral. Por ser tratar de uma
formação sem sazonalidade bem marcada, a Mata Atlântica propriamente dita (Floresta
Ombrófila Densa) não apresenta períodos definidos de estação “chuvosa” e “seca”,
como em sistemas tropicais do Cerrado e Floresta Amazônica, fazendo com que a
dinâmica da MO fique sujeita à imprevisibilidade das chuvas.
No riacho Cachoeira Grande, foi observada uma taxa de aporte lateral maior que
todas as outras encontradas em literatura. Duas hipóteses foram levantadas para explicar
tal fato. Primeiro, existe uma real carência de estudos desse tipo em riachos de Mata
Atlântica, que podem ter padrões de dinâmica da MO distintos da maioria dos biomas já
estudados, assim como muitos estudos não consideram o grau de declividade da
margem e a geomorfologia do leito dos ambientes. Segundo, o período estudado
compreendeu um mês de fevereiro atípico, com 522 mm de chuva acumulados, e
consequentemente um pico de aporte lateral que acabou aumentando significativamente
a média mensal desse compartimento. Os resultados também sugerem possíveis
espécies-chave na dinâmica da MO de riachos de Mata Atlântica propriamente dita para
ambientes subtropicais, como Schizolobium parahyba, Ficus eximia, Virola bicuhyba,
Ficus adhatodifolia, Cupania vernalis e Tetrorchidium rubrivenium.

85
Quando comparamos dois riachos com diferentes graus de integridade da
vegetação ripária, constatamos que a riqueza na comunidade de invertebrados aquáticos,
e na composição de folhas do estoque bêntico é significativamente maior em riachos de
vegetação preservada. A integridade da vegetação ripária foi fundamental para
estruturação das comunidades através da diversidade, quantidade e composição de
detritos incorporados, assim como através da proteção fornecida contra variações na
temperatura da água dos ecossistemas. De fato, o riacho impactado apresentou maior
variação na temperatura de água ao longo do ano, e maior variação temporal na
distribuição da comunidade de invertebrados.
Através das lacunas observadas aqui, e para uma discussão mais aprofundada do
assunto, sugerimos estudos que congreguem a comunidade de invertebrados aquáticos e
a riqueza de folhas na composição do detrito em experimentações em campo. Além
disso, se faz necessário um melhor detalhamento sobre a relação entre qualidade e
riqueza de espécies do detrito para ambientes tropicais, assim como estudos sobre
preferência alimentar a partir de análises de isótopos estáveis de carbono.
Medições de matéria orgânica dissolvida (MOD) também se fazem necessárias
na medida em que esse recurso representa o principal estoque de MO em águas
correntes (Fisher & Likens, 1973) e contribui com uma porção significativa do aporte
total em riachos (Webster & Meyer, 1997), derivado principalmente do solo da
vegetação ripária (Tank et al., 2010). Apesar dos avanços no entendimento dos
processos ecológicos com matéria orgânica em riachos, muitas das diferenças entre
ambientes e dúvidas refletem omissões de importantes componentes na dinâmica de
MO, especialmente medidas acuradas da área do leito do rio, respiração heterotrófica,
estoque bêntico de MOPF e aporte de MOD através de água subterrânea (Webster e
Meyer, 1997).
Impactos antrópicos vêm causando a degradação das zonas ripárias ao redor do
mundo, alterando desse modo a dinâmica de aporte de folhas para ecossistemas de
riachos (Wallace et al., 1997). Não obstante, segundo Kominosky & Rosemond, (2012),
as mudanças globais vão influenciar a dinâmica de MO dos ecossistemas de rios e
riachos alterando a quantidade e qualidade de MO, fenologia da vegetação ripária, e as
taxas de produção heterotróficas e autotróficas. Manter e restaurar esse aporte de
detritos ripários se torna essencial para conservação e restauração das cadeias tróficas de
riachos (Wallace et al., 1997).

86
Referências Gerais
Ab’Saber, A.N. (2009). O suporte ecológico das florestas beiradeiras (ciliares).
In: Rodrigues, R.R. e Leitão-Filho, H.F. (eds.) Pp 15-25. Matas ciliares: conservação e
recuperação. São Paulo: EDUSP, 2ª ed. 2ª reimp. 320p.
Abelho, M. & Graça, M.A.S. (1998). Litter in a first-order stream of a temperate
deciduous forest (Margaraça Forest, central Portugal). Hydrobiologia, 386, 147–152.
Abelho, M. (2001). From litterfal to breakdown in streams: a review. The
Scientific World, 1, 656-680.
Acuña, V., Giorgi, A., Muñoz, I., Sabater, F. & Sabater, S. (2007).
Meteorological and riparian influences on organic matter dynamics in a forested
Mediterranean stream. Journal of the North American Benthological Society, 26(1):54-
69.
Alonso, A., González-Muñoz, N. & Castro-Díez, P. (2010). Comparison of leaf
decomposition and macroinvertebrate colonization between exotic and native trees in a
freshwater ecosystem. Ecological Research 25: 647-653.
Batsviken, D, Tranvik, L.J., Downing, J.A., Crill, P.M. & Prast, A.E. (2011).
Freshwater Methane Emissions Offset the Continental Carbon Sink. Science, 331, 50.
Begon, M., Townsend, C.R., Harper, J.L. (2007). Ecologia: de indivíduos a
ecossistemas. Porto Alegre, Artmed, 752p.
Benfield, E.F. (1997).Comparison of litterfall input streams. Journal of the
North American Benthological Society, 16, 104-108.
Benson, L.J. & Pearson, R.G. (1993). Litter inputs to a tropical Australian
rainforest stream. Australian Journal of Ecology, 18, 377–383.

87
Binckley, C.A., Wipfli, M.S., Medhurst, R.B., Polivka, K., Hessburg, P., Salter,
R.B. & Kill, J.Y. (2010). Ecoregion and land-use influence invertebrates and detritus
transport from headwater streams. Freshwater Biology 55, 1205-1218.
Campanili, M. & Prochnow, M. (orgs) (2006). Mata Atlântica – uma rede pela
floresta. RMA, Brasília: BR.
CASAN (2009) In: http://www.casan.com.br/index.php?sys=370 acesso em
05/11/2009, 21h50min.
Caruso, M.M.L. (1990). O desmatamento na Ilha de Santa Catarina de 1500 aos
dias atuais. Ed. da UFSC, Florianópolis, BR.
Carvalho, E.M. & Uieda, V.S. (2010). Input of litter in deforested and forested
areas of a tropical headstream. Brazilian Journal of Biology, 70, n 2.
CECA/FNMA (1996). Uma cidade numa ilha – relatório sobre os problemas
sócio-ambientais da Ilha de Santa Catarina. Florianópolis, Ed. Insular, 248p.
Clarke K.R. & Warwick., R.M. (2001). Change in marine communities: an
approach to statistical analysis and interpretation. Plymouth, PRIMER-E, UK, 2nd ed.,
172p.
Cole, J.J., Prairie, Y.T, Caraco, N.F., Mcdowell, W.H., Tranvik, L.J., Striegl,
R.G., Duarte, C.M., Downing J.A., Middelburg, J.J. & Melack, J. (2007). Plumbing the
global carbon cycle: integrating inland waters into the terrestrial carbon budget.
Ecosystems, 10, 171–184.
Conners, M.E. & Naiman, R.J. (1984). Particulate allochthonous inputs:
relationships with stream size in an undisturbed watershed. Canadian Journal of
Fisheries and Aquatic Sciences, 41, 1473-1484.
Cummins, K.W., Petersen, R.C., Howard, F.O., Wuycheck, J.C. & Holt, V.I.
(1973) The utilization of leaf litter by stream detritivores. Ecology, 54 (2), 336-345.

88
Cummins, K.W. (1974). Structure and function of stream ecosystems.
BioScience, 24, 631-641.
Cummins, K.W. & Klug, M.J. (1979). Feeding ecology of streams invertebrates.
Annual Review of Ecology, Evolution and Systematics, 10, 147-172.
Dobson, M. & Frid, C. (1998). Ecology of Aquatic Systems. Longman, Essex,
232p.
Fernández, H.R. & Dominguéz, E. (2001). Guia para determinación de los
artropodos bentônicos Sudamericanos. UNT, Tucumán: 282 p.
Ferraz, D.K. (2001). O papel da vegetação na margem de ecossistemas
aquáticos. In: Primack, R. B. & Rodrigues, E. (eds.) Biologia da conservação.
Londrina, 328p.
Fisher, S.G. & Likens, G. E. (1973). Energy flow in Bear Brook, New
Hampshire: an integrative approach to stream ecosystem metabolism. Ecological
Monographs, 43, 421-439.
França, J.S., Gregorio, R.S., De Paula, J.D., Gonçalves Jr., J.F.; Ferreira, F.A. &
Callisto, M. (2009). Composition and dynamics of allochthonous organic matter inputs
and benthic stock in a Brasilian stream. Marine and Freshwater Research 60, 990-998.
Gessner, M.O., Chauvet E. & Dobson M. (1999). A perspective on leaf litter
breakdown in streams. Oikos, 85, 377-384.
GGE (GRUPO GESTÃO DO ESPAÇO – UFSC). Atlas ambiental municipal,
Florianópolis, SC, Brasil. Coord. ORTH, D. M. Projeto Funcitec, UFSC, 2006.
Gonçalves Jr., J.F., Santos, A.M. & Esteves, F.A. (2004). The influence of the
chemical composition of Typha domingensis and Nymphaea ampla detritus on
invertebrate colonization during decomposition in a Brazilian coastal lagoon.
Hydrobiologia, 527, 125-137.

89
Gonçalves Jr., J.F., França, J.S. & Callisto, M. (2006). Dynamics of
allochthonous organic matter in a tropical brazilian headstream. Brazilian Archives of
Biology and Technology, 49, 967-973.
Graça, M.A.S. (2001). The role of invertebrates on leaf litter decomposition in
streams – a review. International Review of Hydrobiology, 86: 383-393.
Graça, M.A.S. & Zimmer, M. (2005). Leaf toughness. In: Graça, M.A.S.,
Barlocher, F. & Gessner, M.O. Methods to Study Litter Decomposition: A Practical
Guide. Springer, Netherlands, 342p.
Gregory, S.V., Swanson, F.J., Mckee, W.A. & Cummins, K.W. (1991). An
ecosystem perspective of riparian zones – focus on links between land and water.
BioScience, 41, 540–551.
Kawaguchi, Y., Taniguchi, Y. & Nakano, S. (2003). Terrestrial invertebrate
inputs determine the local abundance of stream fishes in a forested stream. Ecology,
84(3), 701–708.
Kirby, J.M., Webster, J.R. & Benfield, E.F. (1983). The role of shredders in
detrital dynamics of permanent and temporary streams. In: Fontaine, T.D & Bartell,
S.M. Dynamics of lotic ecosystems. Michgan, Ann Arbor Science, 494p.
Klein, E.M. (1980). Ecologia da flora e vegetação do Vale do Itajaí. Sellowia –
Anais Botânicos do Herbárieo “Barbosa Rodrigues”, 32, 165-389.
Klein, R.M. (1984). Aspectos dinâmicos da vegetação do sul do Brasil. Sellowia
– Anais Botânicos do Herbário “Barbosa Rodrigues”, 36, 5-54.
Kominoski J.S. & Rosemond, A.D. (2012). Conservation from the bottom up:
forecasting effects of global change on dynamics of organic matter and management
needs for river networks. Freshwater Science, 2012, 31(1), 51–68.

90
Landeiro, V.L., Hamanda, N., Godoy, B.S. & Melo, A.S. (2010). Effects of litter
patch area on macroinvertebrate assemblage structure and leaf breakdown in Central
Amazonian streams. Hydrobiologia, 649, 355-363.
Lima, W.P. & Zakia, M.J.B. (2009). Hidrologia de matas ciliares. In: Rodrigues,
R.R. & Leitão-Filho, H.F.L. (eds.). Matas ciliares: conservação e recuperação. São
Paulo, Edusp, 2ª ed. 2ª reimp. 319p.
Moretti, M.S., Gonçalves Jr., J.F., Ligeiro, R. & Callisto, M. (2007).
Invertebrates colonization on native tree leaves in a neotropical stream (Brazil).
International Review of Hydrobiology, 92, 199-210.
Moulton, T.P. & Magalhães, S.A.P. (2003). Responses of leaf processing to
impacts in streams in Atlantic Rain Forest, Rio de Janeiro, Brazil – a test of the
biodiversity-ecosystem functioning relationship?. Brazilian Journal of Biology. 63: 87-
95.
Moulton, T.P., Magalhães-Fraga, S.A.P., Brito, E.F. & Barbosa, F.A. (2010).
Macroconsumers are more important than specialist macroinvertebrate shredders in leaf
processing in urban forest streams of Rio de Janeiro, Brazil. Hydrobiologia, 638: 55-66.
Mugnai, R., Nessimian, J.L. & Baptista, D.F. (2010). Manual de identificação de
macroinvertebrados aquáticos do Estado do Rio de Janeiro. Technical Books Rio de
Janeiro: 176 p.
Naiman, R.J. & Décamps, H. (1997). The ecology of interfaces: Riparian zones.
Annual Review of Ecology, Evolution, and Systematics, 28, 621-658.
Nascimento, R. (2002). Atlas Ambiental de Florianópolis. Instituto Larus,
Florianópolis, BR.
Neto, A.B.C. & Klein, R.M. (1991). Mapeamento temático do município de
Florianópolis: Vegetação - síntese temática. IPUF, Florianópolis, BR.

91
PMF (2009). In: http://www.pmf.sc.gov.br/floram/nova/parques/lagoa_peri.htm
acesso em 05/11/2009, 21h18min.
Primack, R.B. & Rodrigues, E. (2001). Biologia da conservação. Efraim
Rodrigues, Londrina, BR.
Poole, G. C. (2002). Fluvial landscape ecology: adressing uniqueness within the
river discontinuum. Freshwater Biology 47: 641-660.
Pozo, J., González, E., Díez, J.R., Molinero, J. & Elósegui, A. (1997). Inputs of
particulate organic matter to streams with different riparian vegetation. Journal of the
Notrh America Benthological Society, 16, 602-611.
Prast, A.E. & Pinho, L. (2008). Ciclo do carbono em ecossistemas aquáticos
continentais brasileiros. Oecologia Brasiliensis, 12, 03-05.
Primack, R.B. & Rodrigues, E. (2001). Biologia da conservação. Londrina,
328p.
Rezende, C.F. & Mazzoni, R. (2005). Seasonal variation in the input of
allochthonous matter in an Atlantic Rain Forest stream, Ilha Grande-RJ. Acta
Limnologica Brasiliensia, 17, 167-175.
Rezende, R.S.; Gonçalves Jr. J.F. & Petrucio, M.M. (2010). Leaf breakdown and
invertebrate colonization of Eucalyptus grandis (Myrtaceae) and Hirtella glandulosa
(Chrysobalanaceae) in two Neotropical lakes. Acta Limnologica Brasiliensia, 22, 23-34.
Rodrigues, R.R. (2009). Uma discussão nomenclatural dos formações ciliares.
In: Rodrigues, R.R. e Leitão-Filho, H.F. (eds.), Pp 91-99. Matas ciliares: conservação e
recuperação. São Paulo: EDUSP, 2ª ed. 2ª reimp. 320p.
Rodrigues, R.R. & Nave, A.G. (2000). Heterogeneidade florística das matas
ciliares. In: Rodrigues, R.R. e Leitão Filho, H.F.L. (eds.), Pp. 45-71. Matas ciliares:
conservação e recuperação. São Paulo: EDUSP. 320p.

92
Rodrigues, R.R. & Shepherd, G.J. (2009). Fatores condicionantes da vegetação
ciliar. In: Rodrigues, R.R. e Leitão-Filho, H.F. (eds.), Pp 101-107. Matas ciliares:
conservação e recuperação. São Paulo: EDUSP, 2ª ed. 2ª reimp. 320p.
Santos, G.F., Silva, J.T.N., Mendoça, M. & Veado, R.W. ad-V. (1989). Análise
Ambiental da Lagoa do Peri. Geosul, 8, 101-123.
Sobral, M., Jarenkow, J.A., Brack, P., Irgang, B., Larocca, J. & Rodrigues, R.s.s.
(2006). Flora arbórea e arborescente do Rio Grande do Sul, Brasil. São Carlos,
RiMA/Novo Ambiente.
Tank, JL, Rosi-Marshall, EJ., Griffiths, na., Entrekin, SA. & Stephen, ML.
(2010). A review of allochthonous organic matter dynamics and metabolism in streams.
Journal of the North American Benthological Society, 29(1), 118–146.
Townsend, C.R. (1996). Concepts in river ecology: pattern and process in the
catchment hieraechy. Archiv fur Hidrobiologie, 113, 3-21.
Vannote, R.L., Minshall, G.W., Cummins, K.W., Sedell, J.R. & Cushing, C.E.
(1980).The river continuum concept. Canadian Journal of Fisheries and Aquatic
Sciences. 37, 130–137.
Wallace, J.B., Eggert, S.L., Mayer, J.L. & Webster, J.R. (1997). Multiple trophic
levels of a forest stream linked to terrestrial litter inputs. Science, 277,102-104.
Webster, J.R. & J.L. Meyer. (1997). Organic matter budgets for streams: a
synthesis. Journal of the North American Benthological Society, 16, 141–161.
Wettstein, R.R.V. (1970). Plantas do Brasil: aspectos da vegetação do sul do
Brasil. São Paulo, Edgar Blücher, 126p.
Wiens, J.A (2002). Riverine landscape: taking landscape ecology into the water.
Freshwater Biology, 47, 501-515.

93