dinosaurs in marine strata: evidence from the british ... · dinosaurs in marine strata: evidence...
TRANSCRIPT

III Jornadas Internacionales sobre Paleontología de Dinosaurios y su EntornoSalas de los Infantes, Burgos
Dinosaurs in marine strata: evidence from the BritishJurassic, including a review of the allochthonous vertebrate assemblage from the marine Kimmeridge Clay Formation
(Upper Jurassic) of Great Britain
Dinosaurios en depósitos marinos: evidencias del Jurásico Británico, incluyendo una revi-sión de las asociaciones de vertebrados alóctonos de la Formación marina Kimmerige Clay
D. M. MARTILL, S. EARLAND and D. NAISH
Palaeobiology Research Group, School of Earth and Environmental Sciences, University of Portsmouth, Burnaby
Building, Burnaby Road, Portsmouth, PO1 3QL, UK.
Abstract: Dinosaur remains occur frequently in marine strata including both marginal and deeper basi-
nal facies. Such occurrences are generally considered to represent parautochthonous assembla-ges of dinosaurs that drifted out to sea. All non-avian dinosaurian body plans lack adaptations for a sustained and efficient aquatic existence compared with Mesozoic aquatic reptiles such as plesiosaurs, crocodilians and ichthyosaurs or Recent mammals such as cetaceans, pinnipeds and sirenians. However, this does not exclude some dinosaurs from being at least amphibious. The world’s oceans represent a rich source of nutrients within its surface waters, at depth and in the littoral realm. It therefore seems surprising that non-avian dinosaurs apparently did not exploit marine habitats during the Mesozoic. Some non-avian dinosaurs are known almost exclusively from marine strata (e.g. multiple specimens of Scelidosaurus harrisonii and Lexovisaurus sp.), while some forms (e.g. Baryonyx and Spinosaurus) are proposed to have been piscivorous on the basis of fossil stomach contents and modifications of the skeleton and dentition. Could some dinosaurs therefore have been amphibious and/or marine? This hypothesis is not new and dates back to the earliest studies of dinosaurs (e.g. Leidy, 1858; Owen, 1858). Here we review the oc-currence of dinosaurs in marine Jurassic strata of Great Britain and consider their palaeoecology in terms of their taphonomy and biostratinomy. In addition we present a review of the dinosaur assemblage in the marine Upper Jurassic Kimmeridge Clay Formation.
Martill, D. M., Earland, S. and Naish, D. (2006): Dinosaurs in marine strata: evidence from the British Jurassic, including a review of the allochthonous vertebrate assemblage from the marine Kimmeridge Clay Formation (Upper Jurassic) of Great Britain. En (ColectivoArqueológico-Paleontológico Salense, Ed.): Actas de las III Jornadas sobre Dinosaurios y su Entorno. 1-31. Salas de los Infantes, Burgos, España.
35

III Jornadas Internacionales sobre Paleontología de Dinosaurios y su Entorno36
D. M. Martill, S. Earland and D. Naish
Key words: Dinosauria, Jurassic, Kimmeridge Clay Formation, marine environments.
Resumen:Los restos de dinosaurios aparecen frecuentemente en depósitos marinos incluyendo tanto
facies litorales como facies más profundas. Estos hallazgos se consideran generalmente como asociaciones parautóctonas de dinosaurios que fueron arrastrados hacia el mar. Los cuerpos de todos los dinosaurios no avianos carecen de adaptaciones para mantener una existencia acuática en comparación con reptiles acuáticos Mesozoicos como los plesiosaurios, cocodrilos e ichth-yosaurios. Al igual que con los mamíferos recientes como los cetáceos, pinnípedos y sirénidos. Aunque esto no excluye a algunos dinosaurios de ser al menos anfibios. El mundo de los océanos representa una rica fuente de nutrientes en las aguas de superficie, en profundidad y en la franja litoral. Parece entonces sorprendente que los dinosaurios no avianos aparentemente no hayan explotado los habitats marinos durante el Mesozoico. Algunos dinosaurios no avianos son cono-cidos casi exclusivamente a partir depósitos marinos (e.g. varios especimenes de Scelidosaurus harrisonii y Lexovisaurus sp.), mientras que otras formas (e.g. Baryonyx y Spinosaurus) son propuestas como piscívoras en base al contenido fósil de sus estómagos y a las modificaciones de los esqueletos y la dentición. ¿Pudieron entonces algunos dinosaurios haber sido anfibios y/o marinos?. Esta hipótesis no es nueva, data de los primeros estudios sobre dinosaurios (e.g. Leidy, 1858; Owen, 1858). En este trabajo se revisa la presencia de dinosaurios en depósitos marinos del Jurásico de Gran Bretaña y se considera su paleoecología en términos de su tafonomía y bio-estratinomía. Además se presenta una revisión de la asociación de dinosaurios en la formación marina del Jurásico Superior Kimmeridge Clay Fm.
Palabras clave: Dinosauria, Jurásico, Kimmeridge Clay Formation, ambientes marinos
English Jurassic DinosaurioThere are approximately 23 named dinosaur genera recorded from the English Jurassic (Table
1). Of these, many are nomina dubia, represented by remains that are highly fragmentary. Never-theless, such occurrences are often easily placed within higher clades and represent documenta-ble dinosaurian occurrences. Of these 23 genera only 7 are represented by substantial specimens. Notable occurrences are two near complete skeletons of Scelidosaurus harrisonii (Owen, 1861); the near complete Eustreptospondylus oxoniensis which, despite being discovered before 1870 (Phillips, 1871) remains largely undescribed; partial skeletons of Cetiosaurus oxoniensis Phillips, 1871, Cetiosauriscus stewarti Charig, 1980, Lexovisaurus durobrivensis (Hulke, 1887) and Da-centrurus armatus Owen, 1875 and some substantial skull material (e.g. Megalosaurus hesperis, Proceratosaurus bradleyi). Associated remains, perhaps indicating the pre-collection presence of more substantial remains, include the holotype of Sarcosaurus woodi, associated material of Megalosaurus bucklandii from the Bathonian of Oxfordshire and ‘Ornithopsis’ leedsii from the Kellaways Sand Formation of Peterborough (Martill, 1988).

Salas de los Infantes, Burgos 37
Dinosaurs in marine strata: evidence from the British Jurassic.
Table 1. Facies distribution of dinosaurs in the United Kingdom
Early JurassicSarcosaurus Marine, basinalScelidosaurus Marine, basinal
Also known. A theropod partial limb from Sinemurian of Lyme Regis, Dorset and atheropod tibia from Sinemurian of Isle of Skye, Scotland: both from marine strata.
Mid JurassicEustreptospondylus Marine, basinalIliosuchus Marine, marginalMegalosaurus Marine, marginal + freshwaterMagnosaurus Marine, marginalProceratosaurus Marine, marginalCardiodon Marine, marginalCetiosaurus Marine, marginal + freshwaterCetiosauriscus Marine, basinal‘Ornithopsis’ Marine, basinalCryptosaurus Marine, basinalLexovisaurus Marine, basinal + marginalSarcolestes Marine, basinalCallovosaurus Marine, basinal?Priodontognathus Uncertain
Late JurassicMetriacanthosaurus Marine, basinalCumnoria Marine, basinalDacentrurus Marine, basinalBothriospondylus Marine, basinal‘Cetiosaurus’ Marine, basinalGigantosaurus Marine, basinal‘Megalosaurus’ Marine, basinal
Also known: A new theropod and a possible ornithomimid form the Kimmeridian of Dorset.
Many of these genera are nomina dubia, but the holotype specimens nevertheless represent a dinosaur occurrence. Names in quotes are based on species where designation to that genus is inappropriate and a replacement name is awaited.

III Jornadas Internacionales sobre Paleontología de Dinosaurios y su Entorno38
D. M. Martill, S. Earland and D. Naish
The remainder are highly fragmentary and extremely poorly known. Often, other specimens have been referred to named taxa on no other grounds than having been found in the same strata. Seeley (1869) even assembled type series on the basis of material from different localities.
Of the 23 named genera 21 are known from marine strata, of which 16 genera are recorded from basinal facies occuring in association with open marine fauna such as ammonites and be-lemnites. None of these occurrences are at major hiatus horizons, and thus do not represent remains derived from eroded non-marine strata. Thus a taphonomic or autecological case is re-quired to explain their occurrence in marine strata.
If all British dinosaurian occurrences are included in the analysis then of a total of sixty-six named genera, the percentage (~33%) occurring in marine strata is significantly reduced (a con-sequence of the diverse vertebrate assemblages of the non-marine Wealden Group of southern England and the Triassic red bed sequences of SW England). Of the British Jurassic Sauropoda, 7 genera are recorded from marine strata. For Jurassic Theropoda, 8 genera occur in marine strata while only Megalosaurus is known as a non-marine occurrence. Jurassic ornithopods are also better known from marine occurrences (Callovosaurus, Cumnoria). Stegosaurs, quite well repre-sented in the British fossil record, include two genera discovered in marine strata (Lexovisaurus, Dacentrurus).
British Jurassic ankylosaurs are represented by three genera from marine strata (Scelidosau-rus, Sarcolestes, Cryptosaurus,) whereas the Cretaceous yields only two from non-marine rocks (Hylaeosaurus, Polacanthus) (Pereda Suberbiola and Barrett, 1999). The nodosaurid ankylosaur Priodontognathus cannot be satisfactorily assigned to a formation and may be of Late Jurassic or Early Cretaceous age (Galton, 1980b). Although this British Jurassic dinosaur assemblage re-presents a very restricted small data set when compared with the continental assemblages of Nor-th America, Central Asia and southern South America, it does appear to show some anomalies regarding distribution of dinosaur taxa between marine and non-marine formations. Of particular note is the high diversity of thyreophorans in marine strata with 5 genera (3 ankylosaurs and 2 stegosaurs) documented, whereas only one form definitely comes from non-marine strata.
The Kimmeridge Clay dinosaur assemblage
The Upper Jurassic Kimmeridge Clay Formation of England yields an abundant and often well-preserved vertebrate assemblage (Benton and Spencer, 1995). As well as a diversity of fish, the autochthonous vertebrate assemblage contains a range of marine reptiles, including pliosaurs, plesiosaurs, ichthyosaurs, crocodilians and rare turtles (Brown, 1981; Grange and Benton, 1996;

Salas de los Infantes, Burgos 39
Dinosaurs in marine strata: evidence from the British Jurassic.
Grange, et al., 1996; Noè et al., 2004). A rarer allochthonous vertebrate assemblage includes se-veral named species of dinosaurs and fragmentary pterosaurs (Galton and Powell, 1980; Unwin, 1988). The dinosaur assemblage includes both saurischians and ornithischians, two of which are represented by substantial skeletons (Owen, 1875; Galton and Powell, 1980).
This section provides a historical review of the dinosaur assemblage of the Kimmeridge Clay Formation and comments on their preservation and temporal and geographic distribution. In addition, comparisons are made with other Kimmeridgian dinosaur occurrences.
Geological settingThe onshore Kimmeridge Clay Formation accumulated in a shallow, epeiric marine setting
(Hallam, 1987), between 154 and 145 mya (Gradstein et al., 1994). This was a time of elevated atmospheric carbon dioxide levels and temperatures (Hallam, 1985; Sellwood and Price, 1993; Valdes, 1993) and a global sea level highstand (Hallam, 1992).
Figure 1. Outcrop and subcrop map of the onshore Kimmeridge Clay Formation(Kimmeridgian, Upper Jurassic) of the southern United Kingdom. Dinosaur
localities are marked with black dots. Oxford and Cambridgeshown for geographic reference.

III Jornadas Internacionales sobre Paleontología de Dinosaurios y su Entorno40
D. M. Martill, S. Earland and D. Naish
In England, the Kimmeridge Clay Formation crops out in a narrow strip from Dorset in the south-west to Yorkshire in the north-east (Fig. 1), with some gaps in the outcrop due to overste-pping by Cretaceous sequences. It also crops out in Scotland on the east coast at Eathie and Hel-msdale (Wignall and Pickering, 1993) and on the Hebridean islands of Skye and Mull (Hallam, 1965), but only a few vertebrates have been recorded from these localities (Martill pers. obs) and no dinosaur remains are known. Fossil vertebrates have been reported from more than 60 localities from the English outcrops (Benton and Spencer, 1995), but dinosaur remains are only known from the outcrops between the Wash and Dorset. Lithological and coeval equivalents of the Kimmeridge Clay occur in the Boulonnais and Cap de la Hève areas of northern France (Ty-son, 1996) where both marine and terrestrial vertebrates are reported (Galton and Boiné, 1980; Buffetaut and Cacheleux, 1997).
The stratotype section on the Dorset Coast comprises a series of blocky clays and fissile sha-les, with intermittent oil shales, cementstone bands, and layers rich in septarian nodules (Arkell, 1956). Other facies are developed inland, and in places the Upper Kimmeridge Clay may be represented by sandy facies as at Swindon, Oxford, and Aylesbury (Arkell, 1956; Gallois and Cox, 1994). The base of the formation in Dorset is usually represented by clays with a shallow marine fauna of oysters and brachiopods, with the ammonite Pictonia baylei, and represents the beginning of the Kimmeridgian Stage (Cope, 1980). On the coast the formation is commonly di-vided into a Lower and Upper Kimmeridge Clays, based largely on faunal content. The majority of vertebrates from inland exposures appear to have come from the Lower Kimmeridge Clay, but vertebrates are recorded from much of the sequence on the coast (Etches and Clarke, 1999).
The Kimmeridgian Stage marks a period of widespread marine mudrock deposition in north-west Europe (Wignall, 1990) and the thick sequence of clays and bituminous shales is considered to have been deposited in calm bottom waters, and at times anaerobic conditions may have pre-vailed in a stratified water column: an environment similar to that in the Black Sea today (Aigner, 1980; Raiswell et al., 2001). Most of the sediments are terrestrially derived (Iaonnides et al., 1976), indicating considerable erosion from a nearby landmass. Fossilized wood is common in the formation but other plant macrofossils are rare, suggesting deposition some distance from the palaeoshoreline.
The formation’s maximum thickness of around 140 m is reached at the type section in the Purbeck and Kimmeridge Bay areas of Dorset, decreasing to half this value 32 km westwards around Weymouth, 90-100 m at Swindon and 30-45 m at Oxford (Arkell, 1956; Gallois and Cox, 1994). Despite obvious variations in thickness, a good lithostratigraphic and biostratigraphic correlation of the entire UK onshore Kimmeridge Clay Formation is possible (Cox and Gallois, 1981; Wignall, 1993).

Salas de los Infantes, Burgos 41
Dinosaurs in marine strata: evidence from the British Jurassic.
The Dorset type section of the Kimmeridge Clay Formation can be divided into 13 ammonite biozones, from the Pictonia baylei Zone at the base to the Virgatopavlovia fittoni Zone at the top (Cox and Gallois, 1981; Gallois, 1998). At the type locality the formation can be divided into 63 distinct beds using a combination of palaeontological and lithological criteria (Gallois, 1998).
History of dinosaurs in the Kimmeridge Clay FormationVertebrate remains are relatively abundant in the Kimmeridge Clay Formation: several almost
complete specimens of ichthyosaurs, pliosaurs, crocodilians, and many complete specimens of fish have been discovered over a period of nearly 200 years (Benton and Spencer, 1995; Etches and Clarke, 1999). Dinosaurs are less abundant, but are diverse with sauropods, theropods, orni-thopods, ankylosaurs and stegosaurs reported (Table 2). Several important dinosaur discoveries have been made including the type specimens of the sauropod Bothriospondylus suffossus Owen, 1875, the ornithopod Cumnoria prestwichii (Hulke, 1880) and the stegosaur Dacentrurus arma-tus (Owen, 1875). These specimens were collected from inland exposures in clay pits in the 19th century, with most of these localities now being only of historical importance only.
Sauropoda
Bothriospondylus suffosus Owen, 1875‘Cetiosaurus’ humerocristatus Hulke, 1874Ornithopsis manseli Hulke, 1874Gigantosaurus megalonyx Seeley, 1869.
Theropoda
Megalosaurus insignis Eudes-Deslongchamps, 1870+ undescribed partial skeleton and fragments+ possible ornithomimid elements
Ornithopoda
Cumnoria prestwichii (Hulke, 1880)?Bugenasaura sp.
Thyreophora
Dacentrurus armatus (Owen, 1875)Thyreophora gen., et sp., Indet.
Table 2. Dinosaurs of the Kimmeridge Clay Formation.

III Jornadas Internacionales sobre Paleontología de Dinosaurios y su Entorno42
D. M. Martill, S. Earland and D. Naish
Early collecting of vertebrate remains in clay pits and quarries was often undertaken in a haphazard fashion with little or no biostratinomic data recorded. Bones were usually collected with a ‘rough and ready’ approach, leading to the loss or destruction of some smaller and deli-cate elements. The modern decline of extraction industries and the use of large-scale excavating machinery have severely reduced collecting opportunities for Kimmeridge Clay vertebrates in-land. Consequently, vertebrate remains from inland exposures of the Kimmeridge Clay are now frequently incomplete and damaged (McGowan, 1976).
Coastal exposures have also yielded dinosaur remains, but these are generally less well-pre-served and incomplete, due to prolonged exposure and weathering before collection. This con-trasts with dinosaur material from inland clay pits, which was collected as it was exposed. Ne-vertheless, several specimens have been obtained in the past, including the types of ‘Cetiosaurus’ humerocristatus, Ischyrosaurus manseli and Gigantosaurus megalonyx. As recently as 1984 the most complete theropod from the Kimmeridge Clay was found on the coast and sold to the OUM in 1992. This important specimen (OUM J.3311) is currently under study (Paul Barrett pers. com.).
Kimmeridge Clay dinosaur localities
Dorset coastThe Dorset coast is the type section for the Kimmeridge Clay Formation and the type locality
for the Kimmeridgian Stage (Arkell, 1947, 1956; Cope, 1980). Outcrops occur along three stre-tches of the coast: from St Aldhelm’s Head to Kimmeridge Bay, from Ringstead to Osmington (OS Grid. Ref: SY 764815-735817) and around Portland Bill near Weymouth (OS Grid. Ref: SY 670765). Exposures along the coast are extensive and continual erosion ensures a good supply of fossils (Etches and Clarke, 1999). These localities are now part of the Jurassic Coast World Heri-tage Site. Extensive outcrops of the Kimmeridge Clay on the sea floor to the west of Portland Bill frequently yield vertebrate remains. Local scallop fishermen dredge vertebrate fossils, including dinosaurs from the sea floor and, although many specimens are highly abraded and encrusted with epifauna, many bones in good condition (Powell, 1988; Ensom, pers. com.).
Gillingham, Dorset. (OS Grid. Ref: ST 812 257)An old clay pit near the village of Gillingham in north Dorset, approximately 500 m south of
the railway station on the east side of the B3092, formerly exposed the Kimmeridge Clay. Arkell (1933) claimed that only 25 feet (approximately 8 m) of black and selenitic clays entirely within the Rasenia zones were exposed. Fossils recorded include the dubious ichthyosaur Ophthalmo-saurus pleydelli. Only a single dinosaurian specimen, a right forelimb attributed to the stegosaur Dacentrurus (BMNH R5902), has been noted from this locality (Hoffstetter, 1957).

Salas de los Infantes, Burgos 43
Dinosaurs in marine strata: evidence from the British Jurassic.
Figure 2. Stratigraphic ranges (black bars) of the Kimmeridge Clay exposed at a number of sites in Great Britain, and the approximate hori-zons at which dinosaurs have been collected. Icons grouped with a black dot are stratigraphically approximated. Data compiled from various
sources. In the following list of dinosaur taxa H = holotype specimen, S = syntype specimen. 1, Ornithopsis manseli *H; 2, Dacentrurus vertebra; 3, Ornithopod; 4, Megalosaurid maxilla; 5, ‘Cetiosaurus’ humerocristatus *H; 6, Gigantosaurus megalonyx ungula; 7, Dacentrurus forelimb; 8, Indeterminate thyreophoran scutes; 9, Megalosaurid tibia; 10, Dacentrurus armatus *H; 11, Bothriospondylus suffossus *H; 12,
Dacentrurus armatus (*H D. hastiger); 13, Indet. Theropod vertebra; 14, Cumnoria prestwichii *H; 15, Gigantosaurus megalonyx *S.

III Jornadas Internacionales sobre Paleontología de Dinosaurios y su Entorno44
D. M. Martill, S. Earland and D. Naish
Westbury, Blue Circle cement works, Wilts. (OS Grid. Ref. ST 881 526) An active pit in the Lower Kimmeridge Clay (Eudoxus Zone) that is dug for cement manufac-
ture (Fig. 2). This locality has been the source of a number of important vertebrate discoveries, including turtles, crocodilians and pliosaurs (Grange et al., 1996; Benton and Grange, 1996). Taylor et al. 1993 reported the occurrence of probable thyreophoran osteoderms associated with a pliosaur skull from this locality, but were unable to exclude the possibility that they may be pathological pliosaurian elements.
Wootton Bassett (OS Grid. Ref. SU 070 825)Excellent exposures of the Kimmeridge Clay occurred in the Great Western Railway cutting
(OS Grid. Ref. SU 100-821), but currently fossils are obtained from active mud springs at Woo-tton Fields (OS Grid. Ref. SU 075-804). The only dinosaur remains reported from this locality are two partial dermal spines (BMNH 46320 and 46322) attributed by Lydekker (1888) to the stegosaur Omosaurus hastiger Owen, 1877. These specimens are thought to pertain to the same individual and are now referred to Dacentrurus armatus (Galton, 1985). There is some confusion over the specimen numbers for these spines in Galton (1985).
Rodbourne (OS Grid. Ref: ST 935 835)Delair (1973) reported the occurrence of four probable thyreophoran osteoderms which he
considered similar to those of the Cretaceous ankylosaur Polacanthus, but referred to as undes-cribed ‘stegosaur species A’. He provided no details of the nature of the exposure or the litho-logies but stated erroneously that they came from the Kimmeridge Clay. Rodbourne is on the outcrop of the Callovian to Oxfordian Oxford Clay Formation.
Devizes (OS Grid. Ref: 0161) A number of scrappy remains of Kimmeridge Clay dinosaurs have been collected from
around the small market town of Devizes, including a partial theropod tooth crown referred byLydekker (1888) to Megalosaurus insignis Eudes-Deslongchamps and Lennier vide Lennier, 1870,(Lydekker 1888).
Foxhangers, Wiltshire (OS Grid. Ref: ST 937 615)A single tooth (BMNH 46388) referred to the nomen dubium Megalosaurus insignis Deslon-
gchamps was reported from this locality (Lydekker, 1888, p. 163).
Swindon brick and tile works, Wiltshire (OS Grid. Ref: SU 142 838)A series of old pits around the town of Swindon underwent rapid expansion during the cons-
truction of the railway centre to the north of the town during late Victorian times. Numerous re-mains of marine reptiles were discovered, but a specific account of the discovery of the holotype of Dacentrurus armatus and its extraction is given by Davies (1876).

Salas de los Infantes, Burgos 45
Dinosaurs in marine strata: evidence from the British Jurassic.
Shotover Hill (OS Grid. Ref: SP 565 077)Shotover Hill lies approximately 4 km east of the City of Oxford and comprises an outlier of
Early Cretaceous sandy strata resting unconformably on the Upper Jurassic Portland Sands and Kimmeridge Clay Formation. A number of brickyards on the west side of the hill that excavated the Kimmeridge Clay were highly productive for fossil vertebrates during the late 19th and early 20th centuries (Pocock, 1908). The vertebrate assemblage includes hybodont sharks, chimaeras, actinopterygians, ichthyosaurs, plesiosaurs, pliosaurs and crocodilians. The only dinosaur to be found at Shotover Hill is a theropod caudal vertebra (OUM J 47134).
Cumnor (OS Grid. Ref: SP 475 043)The now disused clay pit at Cumnor west of Oxford is the type locality for the ornithopod
Cumnoria prestwichii described by Prestwich (1879, 1880) and named by Hulke (1880). The sec-tion has been described in detail by Richardson et al. (1946). Pocock (1908) records that a seam of clay laminated with white sand, called the Iguanodon bed, yielded the specimen of Cumnoria prestwichii. He further reports that clays below this level yielded crocodilians, ichthyosaurs and pliosaurs.
Ely (Roswell Pits, OS Grid. Ref: TL 550 815 – 565 815)A number of disused pits around Ely, Cambridgeshire were well known for the remains of
Lower Kimmeridge Clay vertebrates in the 19th century. The history of the pits and their verte-brate palaeontology is reviewed in detail by Benton and Spencer (1995) and Noè et al. (2004). Although the pits yielded a number of marine reptiles, the dinosaurs ‘Cetiosaurus’ humerocrista-tus Hulke, 1874 and some material of the type series of Gigantosaurus megalonyx Seeley, 1869 came from this locality.
Littleport (OS Grid. Ref: TL 567 868)A single large phalanx that has been tentatively referred to Gigantosaurus megalonyx Seeley,
1869 (recorded on a museum label) is all that has been reported from Littleport.
Streatham (OS Grid. Ref: TL 514 745)A single vertebra (CAM. J. 29477) from the Kimmeridge Clay of this locality was part of
Seeley’s, (1869) syntype series for Gigantosaurus megalonyx, and is the same specimen for whi-ch Lydekker (1890, p. 241) erected Pelorosaurus manseli (see below).
Cottenham (OS Grid. Ref: TL 450 675)An indeterminate dinosaur bone (CAM. J. 29478) was discovered at this locality, but no preci-
se details are available. The specimen is believed to be part of Seeley’s (1869) syntype series for Gigantosaurus megalonyx (recorded on museum specimen label).

III Jornadas Internacionales sobre Paleontología de Dinosaurios y su Entorno46
D. M. Martill, S. Earland and D. Naish
Denver, Norfolk (OS Grid. Ref: TF 590 011)Dredging the canalised flood relief channel of the Great Ouse River in 1970 at Denver Sluice
provided material from which was obtained a number of dinosaurian elements, including; a head of a long bone CAM. J.61365, fragment of long bone, CAM. J. 61366, a 1st phalanx of the left manus CAM. J. 61367, a terminal phalanx of the manus CAM. J. 61363, and a metacarpal CAM. J. 61364. This material is presumed to be derived from the Kimmeridge Clay (data is from mu-seum labels).
Systematic palaeontology
Several dinosaur taxa have been erected on the basis of material from the Kimmeridge Clay. Unfortunately, with a few exceptions, most of this material is undiagnostic and consequently many of these taxa can be considered nomina dubia (Upchurch, 1993). The following is a review of the systematic status of the Kimmeridge Clay Formation dinosaurs.
Notes. Synonymy lists include only major name amendments. Trivial name misspellings are excluded for reasons of space.
The following abbreviations are used in the text; BMNH = Natural History Museum, London; CAM = Sedgwick
Museum, Department of Earth Sciences, Cambridge University, Cambridge, UK; OUM = University Museum, Oxford;
DORCM = Dorset County Museum, Dorchester; BRSMG = Bristol City Museum and Art Gallery, Bristol; UCMP =
University of California, Museum of Paleontology, Berkeley, USA.
Dinosauria Owen, 1842Saurischia Seeley, 1888Sauropodomorpha Huene, 1932Sauropoda Marsh, 1878
British sauropod taxonomy is convoluted (McIntosh, 1992; Upchurch, 1993; Upchurch and Martin, 2003; Martill and Naish, 2001). The major diagnostic features occur in the skull, verte-brae and to a lesser degree the feet, which are either not preserved or poorly represented in the Kimmeridge Clay Formation material. However, some specimens can be attributed to specific sauropod clades.
Sauropod remains from the formation have been referred to the genera Bothriospondylus, Cetiosaurus, Ischyrosaurus, Ornithopsis, and Pelorosaurus. McIntosh (1992) suggested it is not possible to confidently assign any of the material to a particular genus, while Upchurch (1993) considered some of the material to be diagnostic.
Neosauropoda Bonaparte, 1986Brachiosauridae Riggs, 1904

Salas de los Infantes, Burgos 47
Dinosaurs in marine strata: evidence from the British Jurassic.
‘Cetiosaurus’ humero-cristatus Hulke, 1874
Synonymy1874 Ceteosaurus humero-cristatus Hulke, 1874 p. 17.1888 Ornithopsis humerocristatus (Hulke, 1874); Lydekker, p. 151.1888 Cetiosaurus humerocristatus Hulke, 1874; Lydekker, p. 151.1890 Pelorosaurus humerocristatus (Hulke, 1874); Lydekker, p. 241.
HolotypeBMNH 44635: Incomplete left humerus that measures 1350 mm in length, possibly 1570 mm when complete (Lydekker, 1988) (Fig. 3).
LocalitySmallmouth Sands, Weymouth, Dorset.
HorizonKimmeridge Clay, probably from the mutabilis Zone (Benton and Spencer, 1995).
Referred materialBMNH R3042: Phalanx. Labelled as Kimmeridge Clay, Swanage. BMNH 49165: Dorsal half of right pubis. Kimmeridge Clay, Weymouth.
DiagnosisAlthough Upchurch (1993) suggested C. humerocristatus to be a nomen dubium, Upchurch
and Martin (2003) considered that the humerus was potentially diagnosable because of a combi-nation of the elongation of the humerus with a distal prolongation of the deltopectoral crest.
DiscussionCetiosaurus was the first sauropod dinosaur to be described (Owen, 1841). Hulke (1874) des-
cribed and figured ‘a very large saurian limb bone adapted for progression on land’ discovered at Smallmouth Sands, Weymouth. Benton and Spencer (1995) state that this bone (BMNH 44635) is a femur of Pelorosaurus humerocristatus (Hulke, 1874), but both Lydekker (1888) and Delair (1959) identified it correctly as a left humerus.
Delair (1959) argued that a sauropod phalanx, BMNH R.3042, is actually from the Wealden Clay, as the Kimmeridge Clay is not known to outcrop at Swanage. However, there are good out-crops of Kimmeridge Clay just 5 km from Swanage between St Aldhelms Head and Chapman’s Pool. Visitors to the area would most likely stay in Swanage, but many Victorians would also take long walks along the coast for the sea air, so it is not impossible that R.3042 did indeed come from the Kimmeridge Clay. Woodward (1895) mentions remains of this taxon from the Brick and

III Jornadas Internacionales sobre Paleontología de Dinosaurios y su Entorno48
D. M. Martill, S. Earland and D. Naish
Tile works North of Swindon, but does not give further details.Lydekker (1888, p. 162) originally referred a partial pubis, BMNH 49165 from the Kimme-
ridge Clay to Ornithopsis humerocristatus, which Delair (1959) later tentatively assigned to Or-nithopsis leedsi, a species from the Callovian of Peterborough (Martill, 1988; Upchurch and Martin, 2003, p. 213-214) though the type of the genus Ornithopsis is from the Wealden of the Isle of Wight (Martill and Naish, 2001).
Ornithopsis manseli Lydekker, 1888
Synonymy1874 Ischyrosaurus Hulke, p. 16.1888 Ornithopsis manseli Hulke MS; Lydekker, p. 152.
Figure 3. Reproduction of Hulke’s (1874) plate of the holotype left humerus of ‘Cetiosaurus’ humerocristatus BMNH 44635. a, caudal aspect; b, cranial aspect; c, distal view; d, cross-sectional shape at break in middle.

Salas de los Infantes, Burgos 49
Dinosaurs in marine strata: evidence from the British Jurassic.
1888 Ischyrosaurus manseli Hulke MS; Lydekker, p. 152.1890 Pelorosaurus manseli (Hulke, 1888); Lydekker, p. 241. 1922 Ornithopsis Manselli (Hulke, 1888); Huene, p. 88
HolotypeBMNH 41626: Left humerus with the deltopectoral crest broken away.
LocalityNear Kimmeridge Bay, possibly Weymouth.
HorizonKimmeridge Clay. No details available.
DiagnosisThis bone is not diagnosable beyond showing some features that suggest it is referable to Bra-
chiosauridae (McIntosh, 1992; Upchurch, 1993).
DiscussionHulke (1869), believing the remains belonged to a crocodile, figured the holotype of Ischyro-
saurus manseli without generic determination, but he referred to the same material in a later work (Hulke, 1874) as the type specimen. The name Ischyrosaurus was preoccupied by Cope (1969), but this is a junior synonym of Ischyrotherium, a name that Cope tried to create when he learned that his Ischyrotherium was not a mammal.
Lydekker (1888) identified the bone as a left humerus and placed it with the sauropod dinosaurs in Ornithopsis manseli commenting that had it been complete it may have agreed closely with ‘Cetiosaurus’ humerocristatus. Delair (1959) stated that ‘there is little doubt that the Weymouth humerus belongs to Ornithopsis’. McIntosh (1992) reinstated the earlier name of Ischyrosaurus manseli Hulke 1874, unaware that the name was preoccupied.
Sauropoda indet.
Bothriospondylus suffossus Owen, 1875
Synonymy1875 Bothriospondylus suffossus Owen, p. 15, pl. III-VI.1888 Bothriospondylus suffosus Owen, 1875, Lydekker, p. 170.1897 Pleurocœlus suffosus (Owen, 1875), Marsh, p. 8.1922 Ornithopsis suffosus (Owen, 1875), Huene, p. 88.

III Jornadas Internacionales sobre Paleontología de Dinosaurios y su Entorno50
D. M. Martill, S. Earland and D. Naish
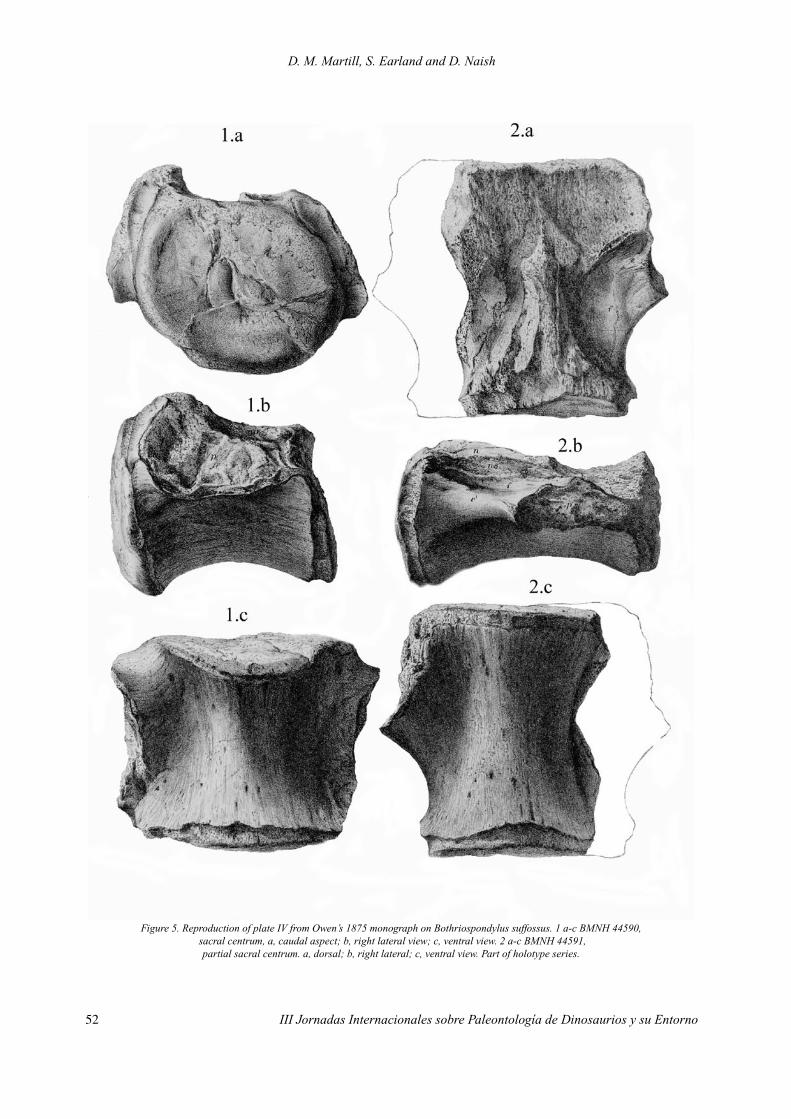
SyntypesBMNH 44589-BMNH 44595: Seven vertebral centra of slightly dubious association (Upchur-
ch, 1993), but all from the same locality. BMNH 44589-91: 3 sacral vertebral centra. BMNH 44592-5: 4 dorsal vertebral centra. (Figs. 4, 5)
LocalitySwindon Brick and Tile Works, Wiltshire. The type material (BMNH 44589-95) was found in
association with a specimen of Pliosaurus brachydeirus Owen, 1841.
HorizonLower Kimmeridge Clay – most vertebrate remains from the Swindon Brick and Tile Works
were recovered from the lower beds, especially the Rasenia and Aulacostephanus zones (Arkell, 1993).
DiscussionOwen (1875) erected four species of Bothriospondylus, all based on vertebrae, but only the
first described species B. suffossus was from the Kimmeridge Clay Formation. The genus Bo-thriospondylus was a wastebasket for various scrappy and indeterminate remains (Naish and Martill, 2001). Owen (1875) recognised B. suffossus as a distinct species on the basis of flattened sacral vertebrae with lateral pleurocoels, dorsal centra gently opisthocoelous, but cranial end almost flat.
Owen’s original description did not list any characters to validate the genus (Upchurch, 1993). Lydekker (1888) included B. suffossus within the Megalosauridae, but later (Lydekker, 1890) noted that this was simply an error. Lydekker (1890, p. 243) suggested that the material might belong to a juvenile or even foetal Pelorosaurus.
Upchurch (1993) suggested possible macronarian affinities for B. suffossus, noting the presen-ce of pleurocoels in the low and long sacral and dorsal centra and the persistence of opisthocoely. However, he concluded that B. suffossus displayed no autapomorphic features. More recently, Salgado and Calvo (1997) suggested that B. suffossus is a basal sauropod or neosauropod and therefore not a macronarian.
Gigantosaurus megalonyx Seeley, 1869
Synonymy1869 Gigantosaurus megalonyx Seeley, p. 64.
SyntypesSeeley assigned several elements to Gigantosaurus megalonyx:

Salas de los Infantes, Burgos 51
Dinosaurs in marine strata: evidence from the British Jurassic.
Figure 4. Reproduction of plate III from Owen’s 1875 monograph on Bothriospondylus suffossus. 1 a-d BMNH 44589, centrum of a sacral vertebra. A, ventral view; b,
caudal aspect; c, right lateral view; d, dorsal aspect. Part of holotype series.
III Jornadas Internacionales sobre Paleontología de Dinosaurios y su Entorno52
D. M. Martill, S. Earland and D. Naish
Figure 5. Reproduction of plate IV from Owen’s 1875 monograph on Bothriospondylus suffossus. 1 a-c BMNH 44590, sacral centrum, a, caudal aspect; b, right lateral view; c, ventral view. 2 a-c BMNH 44591,
partial sacral centrum. a, dorsal; b, right lateral; c, ventral view. Part of holotype series.

Salas de los Infantes, Burgos 53
Dinosaurs in marine strata: evidence from the British Jurassic.
The following are labelled as syntype material:CAM J. 29477: Proximal caudal vertebra, poorly preserved.CAM J. 29478: Middle caudal vertebra (this specimen is referred to Ornithopsis humero-
cristatus on museum label), Cottenham. CAM J.29479: Cast of an ungual phalanx (also referred to Ornithopsis humerocristatus),
Ely.CAM J.29482: Cast of right radius, identified by Seeley as a fibula. Ely.CAM J.29483: Proximal end of the right tibia. CAM J.29481: Osteoderm (no reason to be referred to Sauropoda ((Upchurch, 1993)).
LocalityVaried (see syntypes). Cottenham, Ely
HorizonKimmeridge Clay.
DiagnosisThis taxon cannot be diagnosed and must be considered a nomen dubium.
Other referred materialCAM J.29484: Large phalanx from the Lower Kimmeridge Clay of Littleport, Cambridgeshire.
The following remains were collected as loose, derived material from the Great Ouse flood relief channel, North of Denver, Norfolk by R. Tingay in 1970. They have not been described.
CAM J.61363: Terminal manual phalanx.CAM J.61364: Metacarpal.CAM J.61365: Head of indeterminate limb bone.CAM J.61366: Fragment of indeterminate long bone.CAM J.61367: 1st phalanx, left manus.
DiscussionGigantosaurus megalonyx was erected on the basis of isolated vertebrae and limb bones from
Ely and other nearby sites. All except one bone are undiagnostic and should be considered as Sauropoda incertae sedis. Seeley later admitted that he was aware the series of remains were not only found at different localities, but also at different times (Upchurch, 1993). It is not clear whether all of the remains are the same species, and which of them Seeley regarded as the type

III Jornadas Internacionales sobre Paleontología de Dinosaurios y su Entorno54
D. M. Martill, S. Earland and D. Naish
of the taxon. Upchurch (1993) notes that a caudal vertebra (J29477) could be a proximal caudal vertebra from an animal similar to Diplodocus or Barosaurus. Scrappy though these remains are, they could therefore represent the earliest diplodocid dilpodocoids in the UK.
Damon (1884) also reported G. megalonyx from gritty clay beds (cymodoce Zone, Benton and Spencer, 1995) at Smallmouth Sands, Dorset. Although it is not clear where all of the mate-rial recorded by Damon is now held, Delair (1959) suggested that the remains could include the type humerus of ‘Cetiosaurus’ humerocristatus, held at the BMHM. Lydekker (1888) suggested that Gigantosaurus megalonyx Seeley and material referred to Ornithopsis leedsii Hulke, 1887 may be specifically identical and be referrable to O. humerocristatus (Lydekker, 1888, p. 151) but did not give reasons why. The holotype of O. leedsii comes from the Kellaways Clay (Ca-llovian) while Gigantosaurus megalonyx is from the Kimmeridgian. Historically there has often been confusion over the stratigraphic position of the holotype of O. leedsii. In the original des-cription Hulke (1887) states that the type specimen held in the Leeds Collection comes from the Kimmeridge Clay of Northamptonshire. Martill (1988) noted that the Kimmeridge Clay does not outcrop in that county and that in fact the specimen was found in a well digging on the outcrop of the Kellaways beds and is thus of Callovian age. As O. leedsii is based on dorsal vertebrae and pelvic elements, it cannot be compared with the syntype material of G. megalonyx.
Sauropod conclusionsBothriospondylus suffossus is considered to be a nomen dubium based on limited material of
non-diagnostic value. Ornithopsis humerocristatus and Pelorosaurus humerocristatus are consi-dered to be junior objective synonyms of ‘Cetiosaurus’ humerocristatus, a taxon that is poten-tially diagnosable and may in future merit generic distinction (Upchurch and Martin, 2003). The name Ischyrosaurus manseli was published with no designated type species and the material may be undiagnostic, so this is also considered a nomen dubium. Gigantosaurus megalonyx is based on undiagnostic material collected from different localities at different times, with unclear type material, and is therefore considered a nomen dubium. However, at least some material referred to this taxon may belong to a diplodocid diplodocoid (Upchurch, 1993). Despite so many nomi-na dubia, the remains are clearly sauropod, and some, such as those attributed to ‘Cetiosaurus’ humerocristatus and Ornithopsis manseli, appear to represent the first appearance of Brachiosau-ridae in Europe.
Theropoda Marsh, 1881
Several partial remains and isolated bones from the Kimmeridge Clay have been referred to Theropoda. Some have been referred to Megalosaurus Buckland, 1824 but, like the sauropod Bo-thriospondylus, this genus quickly became a ‘wastebasket’ for theropod remains. Presently, the precise generic position of any British Kimmeridgian theropod remains is at best questionable.

Salas de los Infantes, Burgos 55
Dinosaurs in marine strata: evidence from the British Jurassic.
Tetanurae Gauthier, 1986Carnosauria Huene, 1920?Megalosauridae Huxley, 1869
‘Megalosaurus’ insignis Eudes-Deslongchamps and Lennier vide Lennier, 1870
Synonymy1870 Megalosaurus insignis Eudes-Deslongchamps, vide Lennier, p. 35.
Referred specimenBMNH 46388: Fragment of a single tooth (Lydekker, 1888, p. 163).
LocalityFoxhangers, near Devizes, Wiltshire. The holotype is from the Kimmeridgian of France.
HorizonKimmeridge Clay.
DiagnosisEudes-Deslongchamps (1870) listed the holotype but provided no diagnosis. Isolated thero-
pod teeth are generally considered to be non-diagnostic at generic and specific level, and the only attempt at a diagnosis for M. insignis is Lydekker’s (1888) “Of very large size: the teeth very wide, with the serrations on the distal border usually extending more than two-thirds the entire length of the border, and the serrations very bold.”
Megalosaurus sp.
The following material in the Oxford University Museum collection has been referred to Me-galosauridae by Powell (1987).
OUM J. 47134 Indet. theropod: caudal vertebra. Shotover, Oxford.OUM J. 29886 Indet. theropod: tibia, Swindon.OUM J.13586a,b,c Indet. theropod: three metatarsals.
A worn theropod maxilla fragment containing several teeth (DCM G.10603) dredged from the sea floor to the west of Portland Bill, Dorset, in the 1980’s and probably from the autissiodorensis Zone of the uppermost part of the Lower Kimmeridge Clay (Powell, 1988).

III Jornadas Internacionales sobre Paleontología de Dinosaurios y su Entorno56
D. M. Martill, S. Earland and D. Naish
Galton (1975) reported three indeterminate theropod teeth (UCMP 49612) referred to Mega-losaurus sp. and collected from the Kimmeridge Clay of Weymouth. These comprise two com-plete crowns, each 15 mm high, and a tip of larger tooth. These teeth were figured by Galton (1995) who noted, on the basis of differing denticle densities, that the two complete crowns may represent a different taxon from the larger tooth tip.
DiscussionMegalosaurus was the first dinosaur to be scientifically described (Buckland, 1824), although
the type material from Stonesfield was not given a specific name until Ritgen named it Mega-losaurus bucklandii in 1826, in honour of its original describer. Eudes-Deslongchamps (1870) named Megalosaurus insignis based on an isolated tooth from the Late Jurassic of France, but did not provide a diagnosis. Sauvage (1874) figured an isolated theropod tooth also from the Late Jurassic of France and referred it to M. insignis. However, Megalosaurus insignis is a nomen dubium as isolated teeth are not diagnostic at the specific level.
? Ornithomimidae
Brokenshire and Clarke (1993) reported the occurrence of isolated theropod remains from the Kimmeridge Clay of the shores of the Fleet in Dorset. The material is currently held in the private collection of Mr Adrian J. Brokenshire, specimen AJBC 1662 (Brokenshire and Clarke, 1993, fig. 1 a-c) and comprises a phalanx and ungual phalanx of the pes and has been tentatively referred to Ornithomimidae. The material is from the Cymodoce zone of the Lower Kimmeridge Clay and if the identification as ornithomimid is correct, then this would be the earliest occurren-ce of the group. However, from the figures given it is probably better that the material is consi-dered indeterminate theropod.
Undescribed Kimmeridgian theropod
The remains of a partial theropod skeleton were discovered in the Kimmeridge Clay of Dorset between Chapman’s Pool and Kimmeridge Bay. The remains come from the P. eastlecotten-sis Subzone of the Pectinatus Zone and were purchased by Oxford Univerity museum in 1992 (UOM J3311/1 - ~30). They include cervical, dorsal, sacral and caudal vertebrae, left and right ilia, left and right pubis, left and right ischium, left and right femora, left and right tibiae and other fragments (H. P. Powell pers. com.).
Ornithischia Seeley, 1887Thyreophora Nopsca, 1915Stegosauria Marsh, 1877Stegosauridae Marsh, 1880Dacentrurus Lucas, 1902

Salas de los Infantes, Burgos 57
Dinosaurs in marine strata: evidence from the British Jurassic.
Dacentrurus armatus (Owen, 1875)
Synonymy1875 Omosaurus armatus Owen, p. 46.1877 Omosaurus hastiger Owen, p. 95.1890 Stegosaurus armatus (Owen, 1875); Lydekker, p. 252.1890 Stegosaurus hastiger (Owen, 1877); Lydekker, p. 252.1902 Dacentrurus armatus (Owen, 1875); Lucas, p. 435.1911 Omosaurus lennieri Nopcsa, p.1957 Astrodon pusillus de Lapparent and Zbyszewski, p. 441980 Dacentrurus lenneri (Nopsca, 1911) Galton and Powell, p. 4361980a Osmosaurus lennieri (Nopsca, 1911) Galton, p. 85
HolotypeBMNH 46013: Post-cranial skeleton comprising; 3 cervical vertebrae, 14 dorsal vertebrae,
sacrum, 12 caudal vertebrae, left humerus, radius, ulna, carpus, metacarpus and phalanx, both ischia and ilia, right pubis, femur, tibia, distal end of fibula and calcaneum, left metatarsal IV, right dermal plate from neck region and left dermal tail spine.
LocalitySwindon Brick and Tile Works, Wiltshire.
HorizonKimmeridge Clay; Aulacostephanus or A. mutabilis Zone
Other referable material
BMNH 47329 Centrum of a posterior caudal vertebra; KC, Swindon.BMNH 46320 An incomplete dermal spine; KC of Wootton-Basset, Wilts. (Type speci- men of Omosaurus hastiger).BMNH 46321 Transverse section from the upper extremity of 46320.BMNH 46322 A similar dermal spine, from the same individual as 46320.BMNH 45910 Vertebra from Smallmouth Sands, Weymouth. (G/Ref SY 669764-SY 672771) (Galton, 1985). (Vertebra consists of the neural arch of a mid- caudal vertebra).BMNH R. 5902 Right forelimb from the Gillingham Pottery Company Pit, Dorset.
OUM J1644-45 Dermal spines. Kimmeridge Clay; Swindon, Wilts.

III Jornadas Internacionales sobre Paleontología de Dinosaurios y su Entorno58
D. M. Martill, S. Earland and D. Naish
DiagnosisDorsal vertebrae with massive centra wider than long and with lateral depressions, almost so-
lid dorsal plate to sacrum, deep prepubic process of pubis and long tail spines with sharp lateral and medial edges. This diagnosis, from Galton (1992), is provisional pending a revision of this taxon.
DiscussionOwen (1875) originally named this taxon Omosaurus armatus but Lucas (1902), noting that
the name Omosaurus was preoccupied by a phytosaur, proposed Dacentrurus as a replacement name. This was not noticed by all workers, some of which continued to use Omosaurus for ste-gosaur material well into the 20th century. The type material was found in 1874 at the Swindon Brick and Tile Works. Owen (1875) also described a second species, O. hastiger based on three dermal spines from Wootten Basset (BMNH 46320-22) that Galton (1985, 1992) referred to D. armatus.
A specimen referred to Dacentrurus from the Late Kimmeridgian of Le Havre, France was destroyed during World War 2 (Buffetaut et al., 1991). A stegosaurian femur, probably referable to Dacentrurus, has been reported from Le Havre (Galton and Boiné, 1980), as have other post-cranial remains from the Kimmeridgian of France and Portugal (Galton, 1991).
Ankylosauria , Osborn, 1923
Although no definite ankylosaur remains have been reported from the Kimmeridge Clay a number of osteoderms have been tentatively referred to this group. We review these below.
Material Several fragments of dermal armour have been tentatively referred to Thyreophora including
BRSMG Cc332eu, BRSMG Cc332j, and BRSMG Cc332du: a pair of dermal scutes from oppo-site sides of the animal and a single isolated fragment provisionally identified as a fragment of dermal scute (Taylor et al. 1993). This material comes from the Blue Circle Company Clay Pit, Wiltshire and is from the Aulacostephanus eudoxus Zone (Lower Kimmeridge Clay Formation). In addition to this material four dermal scutes (OUM J1682a-d) were regarded by Galton (1980b) as probably belonging to Ankylosauria. This may be the same material referred to by Delair (1973) from Rodbourne (see above).
DiscussionThe three dermal scutes (BRSMG Cc332eu, j, du) were found in close association with the
skull and mandible of the large pliosaur Pliosaurus brachyspondylus. An explanation proposed for the presence of the scutes is that they were scavenged from a drifting carcass (Taylor et al.,

Salas de los Infantes, Burgos 59
Dinosaurs in marine strata: evidence from the British Jurassic.
1993). One of the paired scutes (BRSMG Cc332eu) was found crushed into the dorsal surface of the palate inside the left orbit of the pliosaur, while the location of the other two scutes was not recorded. Although Taylor et al. (1993) regarded these scutes as belonging to either a stegosaur or ankylosaur, we feel it more likely that they belonged to an ankylosaur. Their morphology is more consistent with an ankylosaurian identity.
Thyreophora indet.
Brokenshire and Clarke (1993, p. 178) reported the occurrence of an osteoderm and a meta-carpal both from a stegosaur. The osteoderm (AJBC4138) was collected from the Pavlovia Zone of the Upper Kimmeridge Clay at Ringstead Bay, Dorset, while the metacarpal (AJBC4137) came from the Cymodoce Zone of the Lower Kimmeridge Clay near Ferrybridge on the shores of the Fleet, Dorset. Although Brokenshire and Clarke (1993) suggested a stegosaurian identi-ty for this material, it cannot certainly be referred to Stegosauria, and we therefore regard it as Thyreophora indet.
Ornithopoda Marsh, 1881Thescelosauridae Gilmore, 1940cf. Bugenasaura Galton, 1995
No certain remains of thescelosaurid dinosaurs have been reported from the Kimmeridge Clay, but a thescelosaurid dentary tooth (UCMP 49611) was said come from the Kimmeridge Clay of Weymouth, Dorset along with three Megalosaurus teeth (Galton, 1975, figs 1D-F, as hypsilo-phodontid: here we regard Thescelosauridae as a taxon distinct from Hypsilophodontidae). The locality data was based on a museum label and Galton (1980) later cast doubt on its reliability, noting that an identical tooth with identical mode of preservation had been collected by Hatcher from the Lance Formation of Wyoming. However, Galton (1999), while also noting doubts about the specimen’s provenance, commented on the strong similarity between UCMP 49611 and the dentition of the North American Maastrichtian thescelosaurid Bugenasaura infernalis Galton, 1995 and therefore proposed that UCMP 49611 be designated cf. Bugenasaura. Although this identification rather supports the specimen not being from the Kimmeridge Clay, it should be noted that phylogenies suggest an origin of Thescelosauridae in or around the Middle Jurassic. (Weishampel et al., 2003).
Iguanodontia Dollo, 1882Camptosauridae Marsh, 1885

III Jornadas Internacionales sobre Paleontología de Dinosaurios y su Entorno60
D. M. Martill, S. Earland and D. Naish
Cumnoria prestwichii (Hulke, 1880)
Synonymy1880 Iguanodon prestwichii Hulke pp. 433-456, pls. 18-20.1888 Cumnoria prestwichii (Hulke, 1880); Seeley, p. 698.1888 Iguanodon prestwichi (Hulke, 1880); Lydekker, p. 196.1890 Camptosaurus prestwichi (Hulke, 1880); Lydekker, p. 258.1890 Cumnoria prestwichi (Hulke, 1880); Lydekker, p. 2581980 Camptosaurus prestwichii (Hulke, 1880); Galton and Powell, p. 438.
HolotypeA partial skeleton (OUM J.3303) comprising an incomplete skull, vertebral column (many
of the vertebrae preserved only as centra), proximal forelimbs and partial shoulder girdle, and incomplete hindlimbs and partial pelvis.
LocalityChawley Brick Pit, Hurst Hill, Near Cumnor, Oxfordshire.
HorizonLower Kimmeridge Clay, Aulacostephanus pseudomutabilis Zone.
DiagnosisSmall intramaxillary cavity, 5 secondary ridges reach middle of tooth crown distal to promi-
nent keel. Cranial end of cervical centra 4 to 9 sub-rhomboidal in outline. Slender long bones and pes, lesser trochanter of femur a sub-equilateral triangle in cross-section (Galton and Powell, 1980). Pending a revision of this taxon and other camptosaurids, it remains to be determined which of these features represent true apomorphies of this taxon.
DiscussionThe holotype material is a well-preserved, almost complete specimen, with detailed accounts
of its discovery and taphonomy (Prestwich, 1879, 1880). Prestwich (1879) originally assigned the remains to the genus Iguanodon, without a specific epithet. Hulke (1880) erected the new species Iguanodon prestwichii to accommodate the specimen. Seeley (1888) however, noted se-veral differences between J.3303 and Iguanodon, and proposed the new genus Cumnoria for the specimen. Lydekker (1889) questioned the validity of the new genus, and instead referred the specimen to Camptosaurus Marsh, 1885 because of its close resemblance to C. leedsi Lydekker, 1889. Galton and Powell (1980) examined OUM J.3303 and agreed with this referral, although they did not list diagnostic features for Camptosaurus seen in the specimen. More recently Nor-man (1998) regarded the specimen as generically distinct.

Salas de los Infantes, Burgos 61
Dinosaurs in marine strata: evidence from the British Jurassic.
This species has also been reported from the Portlandian of Boulogne-Sur-Mer (Sauvage, 1888), although Galton (1980a) and Galton and Powell (1980) noted that this material should be considered generically indeterminate.
Taphonomy of dinosaurs in marine mudrocks
A diverse spectrum of taphonomic modes is encountered for dinosaurs in marine mudrocks such as the Kimmeridge, Oxford and Lias clays. Martill (1988) noted that dinosaur occurrences in the fully marine Oxford Clay Formation (Middle-Upper Jurassic; M. Callovian to L. Oxfor-dian) varied from isolated, but nevertheless well preserved bones, through partial, associated but disarticulated skeletons, to examples that were probably complete skeletons, or nearly so. In the Lias Group of southern England, examples of Scelidosaurus are known in a fully articulated condition with dermal armour in place (Owen, 1861; pers. obs). In at least two examples of Sce-lidosaurus soft tissues are also reported (Martill et al., 2000).
Many isolated bones in the Kimmeridge Clay show encrustations of serpulid annelids and oysters. Such specimens indicate a prolonged period of time on the sea floor prior to burial, but articulated skeletons generally lack encrusting fauna and were probably buried rapidly, perhaps by submergence in soupy substrates (Martill, 1993); a mode of preservation that can result in a high degree of skeletal articulation and the preservation of more resistant soft tissues such as keratin.
In the Kimmeridge Clay of Great Britain dinosaurs are also preserved in a spectrum of tapho-nomic states, though because much of the material was collected in the late 19th century tapho-nomic data is minimal. Probable thyreophoran osteoderms associated with the skull of a pliosaur reported by Taylor et al. (1993) may have been part of a prey item of the pliosaur, but may also have been a result of scavenging on a drifting carcass. The holotype skeleton of Dacentrurus ar-matus (figs. 6-7) shows bones in an articulated condition. Such an individual must have arrived at the sea floor as a near complete carcass with skin and other soft tissues still in place.
Isolated dinosaur bones in museum collections are difficult to assess in taphonomic terms as it is not possible to know if any attempt was made to locate other elements. Isolated bones can occur as falls from drifting carcasses, or as scatter from more complete carcasses that underwent scavenging from larger marine vertebrates or storm events.
There appears to be little difference in taphonomy between different taxa of dinosaurs in ma-rine mudrocks; there being examples of partial, articulated skeletons of ankylosaurs (Scelidosau-rus), stegosaurs (Dacentrurus), ornithopods (Cumnoria), theropods (Eustreptospondylus) and sauropods (Cetiosauriscus), while isolated bones of the same major groups also occur and thus

III Jornadas Internacionales sobre Paleontología de Dinosaurios y su Entorno62
D. M. Martill, S. Earland and D. Naish
the taphonomic factors operating on dinosaurs in marine strata were taxa independent. Indeed, the same taxonomic pathways are encountered in marine reptiles from the same strata (Martill, 1985).
Comparison with other faunas
The dinosaur assemblage of the English Kimmeridge Clay Formation has similarities with the Kimmeridge Clay equivalents of northern France. Galton and Boiné (1980) reported a femur from Kimmeridgian strata of Normandy which they referred to Dacentrurus lennieri rather than to D. armatus. However D. armatus and D. lennieri are now considered conspecific (Galton, 1992).
Buffetaut and Cacheleux (1997) reported the presence of a femur that they attributed to the
ornithopod Dryosaurus from the Kimmeridgian of Cap de la Hève. This taxon has not been re-ported from the English Kimmeridge Clay. In addition to the holotype of Megalosaurus insignis, a number of teeth from the French Kimmeridgian have been referred to Megalosaurus (Buffetaut et. al., 1991). Teeth referred to the indeterminate sauropod Neosodon (Buffetaut, 1995) have in the past been referred to Pelorosaurus but currently are not comparable with any English Kimme-ridge Clay sauropod.
Figure 6. Reproduction of plate XIX from Owen’s 1875 monograph on Dacentrurus armatus. Ventral view of the pelvis. Part of the holotype
specimen BMNH 46013.

Salas de los Infantes, Burgos 63
Dinosaurs in marine strata: evidence from the British Jurassic.
Figure 7. Reproduction of plate XXI from Owen’s 1875 monograph on Dacentrurus armatus. 1. a-b caudal spines. 2. a-d right tibia.
Part of the holotype specimen BMNH 46013.

III Jornadas Internacionales sobre Paleontología de Dinosaurios y su Entorno64
D. M. Martill, S. Earland and D. Naish
Although several dinosaurs are known from the Late Jurassic of Germany and elsewhere in central Europe, including spectacular examples of Archaeopteryx and Compsognathus, no Ki-mmeridgian forms have yet been formally described. In southern Europe Kimmeridgian aged dinosaurs are well known from the Guimarota and other localities in Portugal, with some 14 ge-nera (Dacentrurus, Dracopelta, Trimucrodon, Phyllodon, Hypsilophodon, Dinheirosaurus, Lo-urinhasaurus, Ceratosaurus, Allosaurus, cf. Compsognathus, Aviatyrannis, cf. Richardoestesia, aff. Paronychodon and cf. Archaeopteryx) reported (Antunes and Mateus, 2003). Of these 14, only Dacentrurus appears to be common to both the English and Portuguese Jurassic.
Other dinosaur assemblages elsewhere in the world of Kimmeridgian age include the famous Morrison and Tendaguru formations, but no genera appear to be common to both the Kimmerid-ge Clay Formation and the North American and African assemblages.
Origin of the Kimmeridge Clay dinosaur assemblage
Although some dinosaurs may have lived near water, either coastally or by rivers, most were terrestrial (e.g., Bakker, 1971). Therefore the occurrence of fully terrestrial animals such as dino-saurs in marine basinal facies like as the Kimmeridge Clay can be attributed to direct predation or by accidental or natural death, and then transportation via large river systems.
Accidental drowning may have occurred during catastrophic storm events. Evidence that such events occurred is illustrated by the abundant wood associated with the type of Cumnoria prest-wichii (Woodward, 1895). This suggests wood washed out from forested areas by storm waters or by coastal cliff collapse. Prestwich (quoted in Woodward, 1895) suggests that the presence of the wood “...together with the thinning of this formation as it trends to the south-west, render it probable that land in that direction was not far distant...”. The fairly complete nature of the type specimen of Cumnoria prestwichii is probably due to its being drowned in a flood and swept out to sea, without being disarticulated to a great degree by scavengers or other processes.
The preservation potential of reptiles is determined by their size and lifestyle (Lyman, 1994). The remains of dinosaurs had to be transported into the area of deposition, perhaps drifting for some time before sinking and being buried by the sediment. During this time any number of other animals probably would have scavenged the carcass. The degree of completeness and articulation of the remains is also dependent upon the rate of burial and length of time spent on the sea floor, during which time the hardground ‘islands’ of the remains may have been colonised by epifaunal organisms. Two examples of highly colonised remains include an incomplete ichthyosaur collec-ted in August 2001 from Mepal, Cambridgeshire, and held at the Sedgwick Museum, Cambridge, and also a metriorhynchid skull described and figured by Benton and Grange (1996), held at Oxford University Museum. As a result of these processes, many of the dinosaur remains from the Kimmeridge Clay are fragmentary and encrusted with epifauna.

Salas de los Infantes, Burgos 65
Dinosaurs in marine strata: evidence from the British Jurassic.
Discussion
The dinosaur fauna of the Kimmeridge Clay is surprisingly abundant and diverse. Sauropods, theropods, ornithopods and stegosaurs are all represented, and include six type specimens. This allochthonous input from terrestrial sources must have been highly significant considering the number of terrestrial remains found in the formation. They represent approximately 0.5% of the reptile finds from the major localities of the formation (calculated from data in Benton and Spencer, 1995). Of this small fraction, it appears that saurischian dinosaurs in the English Ki-mmeridgian constitute a little over 50% of the taxic diversity. In terms of specimen abundance (specimens in museums collections), the remains of sauropods constitute approximately 40% (14 accessions) of dinosaur accessions while theropods are a little under 25% (7 accessions). Orni-thishians constitute around 38% (13 accessions), and of these some 11 accession are thyreopho-rans. The occurrence of these terrestrial remains may be due to direct predation or by accidental or natural death. Once in the marine environment the carcasses of dinosaurs would have repre-sented a food source for both pelagic and benthic organisms, providing energy input and also an increase in the diversity of microhabitats on the sea floor. The dinosaur fauna is also significant in terms of palaeobiogeography, with representatives of some families and genera that can be traced from Africa, through Europe to America. However, the true importance of this fauna cannot be realised until the many taxonomic problems are resolved.
Acknowledgements.We are happy to thank the following: Paul Barrett, Sandra Chapman, Mike Dorling, Owen
Jones, Phil Powell, Graham Spiller, George Olshevsky. We are especially grateful to Steve Etches of Kimmeridge, Dorset for sharing his knowledge of the Kimmeridge Clay and to Dr Eric Buffe-taut for refereeing the manuscript.

III Jornadas Internacionales sobre Paleontología de Dinosaurios y su Entorno66
D. M. Martill, S. Earland and D. Naish
References:
Aigner, T. (1980): Biofabric and stratinomy of the lower Kimmeridge Clay (Upper Jurassic, Dorset, England).
Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 159: 324-338. Stuttgart.
Arkell, W. J. (1933): The Jurassic System in Great Britain. Clarendon Press, Oxford. 681 pp.
Arkell, W. J. (1947): The geology of the country around Weymouth, Swanage, Corfe, & Lulworth. Memoir of the
Geological Survey of Great Britain. 386pp.
Arkell, W. J. (1956): Jurassic Geology of the World. Clarendon Press, Oxford. xv + 806 pp. 46pls.
Bakker, R. T. (1971): The ecology of the Brontosaurs. Nature, 229: 172-174.
Benton, M. J. and Grange, D. R. (1996): Kimmeridgian metriorhynchid crocodiles from England. Palaeontology,
39: 497-514.
Benton, M. J. and Spencer, P. S. (1995): Fossil reptiles of Great Britain. Chapman and Hall, London, 345pp.
Brokenshire, A. J. and Clarke, J. B. (1993): Important recently collected dinosaurian remains from the Lower
Kimmeridge Clay at Weymouth. Proceedings of the Dorset Natural History and Archaeological Society,
115: 177-178.
Brown, D. S. (1981): The English Upper Jurassic Plesiosauroidea (Reptilia) and a review of the phylogeny and
classification of the Plesiosauria. Bulletin of the British Museum (Natural History), Geology Series, 35:
253-347.
Buckland, W. (1824): Notice on the Megalosaurus, or great fossil lizard of Stonesfield. Transactions of the Geo-
logical Society of London, 1: 390-396.
Buffetaut, E. (1994): The significance of dinosaur remains in marine sediments: an investigation based on the
French record. Berliner Geowiss. Abh., E: 125-133.
Buffetaut, E. (1995): Dinosaures de France. Éditions BRGM, France, 144 pp.
Buffetaut, E., Cuny, G. and Le Loeuff, J. (1991): French dinosaurs: the best record in Europe? Modern Geology,
16: 17-42.
Buffetaut, E. and Cacheleux, E. (1997): The dinosaur Dryosaurus (Ornithopoda, Dryosauridae) in the Kimme-
ridgian (Late Jurassic) of Normandy: palaeobiogeographical implications. Comptes Rendus Academie des
Sciences, Paris, 324: Series iia, 499-503.
Buckland, W. (1824): Notice on the Megalosaurus, or great fossil lizard of Stonesfield. Transactions of the Geo-
logical Society of London, Series 2, 1: 390-396.
Callomon, J. H. and Cope, J. C. W. (1971): The stratigraphy and ammonite succession of the Oxford and Ki-
mmeridge Clays in the Warlingham borehole. Bulletin of the Geological Society of Great Britain, 36:
147-176.
Charig, A. J. & Milner, A. C. (1997): Baryonyx walkeri, a fish-eating dinosaur from the Wealden of Surrey. Bu-
lletin of the Natural History Museum, Geology Series, 53: 11-70.
Cope, E. D. (1969): Synopsis of the extinct Batrachia, Reptilia, and Aves of the United States. Transactions of
the American Philosophical Society, 14: 1-252.
Cope, J. C. W. (1967): The palaeontology and stratigraphy of the lower part of the Upper Kimmeridge Clay of
Dorset. The Bulletin of the British Museum (Natural History), 15: 1-79.

Salas de los Infantes, Burgos 67
Dinosaurs in marine strata: evidence from the British Jurassic.
Cope, J. C. W. (1974): Upper Kimmeridgian ammonite faunas of the Wash area and a subzonal scheme for the
lower part of the Upper Kimmeridgian. Bulletin of the Geological Survey of Great Britain, 47: 29-37.
Cope, J. C. W. (1978): The ammonite faunas and stratigraphy of the upper part of the Upper Kimmeridge Clay
of Dorset. Palaeontology, 21: 45-56.
Cope, J. C. W. (1980): Kimmeridgian correlation chart. In Cope, J. C. W., Duff, K. L., Parsons, C. F., Torrens, H.
S., Wimbledon, W. A. and Wright, J. K. (eds.). A correlation of Jurassic Rocks in the British Isles. Part 2:
Middle and Upper Jurassic. Geological Society of London, Special Report, 15: 76-85.
Cox, B. M. and Gallois, R. W. (1981): The stratigraphy of the Kimmeridge Clay of the Dorset type area and its
correlation with some other Kimmeridgian sequences. Report of the Institute of Geological Sciences No.
80/4, HMSO, London. 1-45pp.
Damon, R. (1884): Handbook of the Geology of Weymouth, Portland and Coast of Dorsetshire, from Swanage to
Bridport-on-Sea, 2nd Edition. R. F. Damon, Weymouth; E. Stanford, London, 250 pp.
Davies, W. (1876): On the exhumation and development of a large reptile, Omosaurus armatus Owen, from the
Kimmeridge clay of Swindon, Wilts. Geological Magazine, 3 (2): 193-197.
Delair, J. B. (1959): The Mesozoic reptiles of Dorset: Part 2. Proceedings of the Dorset Natural History and
Archaeological Society, 79: 52-90.
Delair, J. B. (1973): The dinosaurs of Wiltshire. Wiltshire Archaeological and Natural History Magazine, 68:
1-7.
Dollo, L. (1882): Premiére note sur les Dinosauriens de Bernissart. Bulletin du Musée Royal d’Histoire Naturelle
de Belgique, 1: 161-180.
Etches, S. and Clarke, J. (1999): Steve Etches’ Kimmeridge Collection Illustrated Catalogue. Published privately
by J. Clarke. Available from 65 Oak Mount Road, Chandlers Ford, Hampshire SO53 2LJ, UK.
Eudes-Deslongchamps, J.-A. (1870): Note sur les reptiles fossiles appartenant à la famille des téléosauriens,
don’t les débris ont été recueillis dans les assises jurassiques de Normandie. Bulletin de Société de Geo-
logique de France (ser. 2), 27: 299-351.
Gallois, R. W. (1998): The stratigraphy of and well-completion reports for the Swanworth Quarry No. 1 and No.
2 and Metherhills No. 1 boreholes (RGCE Project), Dorset, Technical Report WA/97/91. British Geolo-
gical Survey, Exeter. 102 pp.
Gallois, R. W. and Cox, B. M. (1994). The Kimmeridge Clay and underlying strata (Upper Jurassic) at Swindon,
Wiltshire. Proceedings of the Geologists’ Association 105, 99-100.
Galton, P. M. (1975): English hypsilophodontid dinosaurs (Reptilia: Ornithischia). Palaeontology, 18: 741-752.
Galton, P. M. (1980a): European Jurassic ornithopod dinosaurs of the families Hypsilophodontidae and Campto-
sauridae. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 160: 73-95.
Galton, P. M. (1980b): Armored dinosaurs (Ornithischia: Ankylosauria) from the Middle and Upper Jurassic of
England. Geobios, 13: 825-837.
Galton, P. M. (1985): British plated dinosaurs (Ornithischia, Stegosauridae). Journal of Vertebrate Paleontology,
5: 211-254.

III Jornadas Internacionales sobre Paleontología de Dinosaurios y su Entorno68
D. M. Martill, S. Earland and D. Naish
Galton, P. (1991): Postcranial remains of stegosaurian dinosaur Dacentrurus from the Upper Jurassic of France
and Portugal. Geologica et Palaeontologica, 25: 299-327.
Galton, P. (1992): Stegosauria. In: Weishampel, D. B., Dodson, P. and Osmolska, H. (eds). The Dinosauria. Uni-
versity of California, Berkeley. 435-455.
Galton, P. (1995): The species of the basal hypsilophodontid dinosaur Thescelosaurus Gilmore (Ornithischia:
Ornithopod) from the Late Cretaceous of North America. Neues Jahrbuch für Geologie und Paläontolo-
gie, Abhandlungen, 198: 297-311.
Galton, P. M. and Boiné, G. (1980): A stegosaurian dinosaur femur from the Kimmeridgian beds (Upper Jurassic) of
Cap de la Hève, Normandy. Bulletin Trim. Société Geologique de Normandie Amis Muséum, Havre, 67: 40-60.
Galton, P. M. and Powell, H. P. (1980): The ornithischian dinosaur Camptosaurus prestwichii from the Upper
Jurassic of England. Palaeontology, 23: 411-433.
Gauthier, J. (1986): Saurischian monophyly and the origin of birds. Memoirs of the California Academy of
Science, 8: 1-55.
Gradstein, F. M., Agterberg, F. P., Ogg. J. G., Hardenbol, J., Van Veen, P., Thierry, J. and Huang, Z. (1994): A
Mesozoic timescale. Journal of Geophysical Research, 99: 24, 051-24, 074.
Grange, D.R. and Benton, M.J (1996): Kimmeridgian metriorhynchid crocodiles from England. Palaeontology,
39: 497-514.
Grange, D. R., Storrs, G. W., Carpenter, S. and Etches, S. (1996): An important marine vertebrate-bearing lo-
cality from the Lower Kimmeridge Clay (Upper Jurassic) of Westbury, Wiltshire. Proceedings of the
Geologists’ Association, 107: 107-116.
Hallam, A. (1965): Jurassic, Cretaceous and Tertiary sediments. 401-416. In: Craig, G.Y. (ed.). The geology of
Scotland. Oliver and Boyd, Edinburgh, 2 vols., 556 pp.
Hallam, A. (1985): A review of Mesozoic climates. Journal of the Geological Society, London 142, 433-445.
Hallam, A. (1987): Mesozoic marine organic-rich shales. In: Brooks, J. R. V. and Fleet, A. J. (eds.). Marine pe-
troleum source rocks. Special Publication of the Geological Society, London. 26.
Hallam, A. (1992): Jurassic. In: Duff, P. McL. D. and Smith, A. J. (Eds.). Geology of England and Wales. Geo-
logical Society of London, 325-354.
Hoffstetter, R. (1957): Quelques observation sur les stégosaurinés. Bulletin de Muséum d’Histoire Naturel, Paris,
29: 537-547.
Huene, F. von. (1927): Short review of the present knowledge of the Sauropoda. Memoirs of the Queensland
Museum, 8: 121-126.
Huene, F. von. (1932): Die Fossile Reptile-Ordnung Saurischia, ihre Entwicklung und Geschichte. Monogra-
phien zur Geologie und Paläontologie, 4: 361 pp.
Hulke, J. W. (1869): Note on a large saurian humerus from the Kimmeridge Clay of the Dorset coast. Quarterly
Journal of the Geological Society of London, 25: 386-389.
Hulke, J. W. (1874): Note on a very large saurian limb-bone adapted for progression upon land, from the Kimme-
ridge Clay of Weymouth, Dorset. Quarterly Journal of the Geological Society of London, 30: 16-17.

Salas de los Infantes, Burgos 69
Dinosaurs in marine strata: evidence from the British Jurassic.
Hulke, J. W. (1880): Iguanodon prestwichii, a new species from the Kimmeridge Clay, distinguished from I.
mantelli of the Wealden formation in the S.E. of England and the Isle of Wight by differences in the shape
of the vertebral centra, by fewer than 5 sacral vertebrae, by the simpler character of its tooth-serrature,
etc., founded on numerous fossil remains lately discovered at Cumnor, near Oxford. Quarterly Journal of
the Geological Society of London, 36: 433-456.
Hulke, J. W. (1887): Note on some dinosaurian remains in the collection of A. Leeds Esq. of Eyebury, Nor-
thamptonshire. Quarterly Journal of the Geological Society of London, 63: p. 695.
Huxley, T. H. (1870): Further evidence of the Affinity between the Dinosaurian reptiles and birds. Quarterly
Journal of the Geological Society of London, 26: 12-31; SM 3: 465-86.
Iaonnides, N. S., Stavringos, G. N. and Downie, C. (1976): Kimmeridgian microplankton from Clavell’s Hard,
Dorset. Micropaleontology 22, 443-478.
Krebs, B. (1968): Le crocodilien Machimosaurus. Servico Geólogico Portugal, N. S., 14: 21-52.
Lapparent, A. F. and Zbyszewski, G. 1957. Les dinosauriens du Portugal. Services Géologiques du Portugal,
Mémoire 2: Nouvelle Série 1-63 + 36 pls.
Leidy, J. (1858): Hadrosaurus foulkii, a new saurian from the Cretaceous of New Jerzey. Proceedings of the
Academy of Natural Sciences, Philadelphia, 10: 215-218.
Lucas, F. A. (1902): The generic name Omosaurus. Science 16, 435.
Lydekker, R. (1888): Catalogue of the Fossil Reptilia and Amphibia in the British Museum (Natural History).
Part 1. The Orders Ornithosauria, Crocodilia, Dinosauria, Squamata and Proterosauria. British Museum
(Natural History), London, 309 pp.
Lydekker, R. (1889): On the remains and affinities of five genera of Mesozoic reptiles. Quarterly Journal of the
Geological Society of London, 30: 41-59.
Lydekker, R. (1890): Catalogue of the Fossil Reptilia and Amphibia in the British Museum (Natural History).
Part IV. Supplement. British Museum (Natural History), London, 295 pp.
Lyman, R. L. (1994): Vertebrate Taphonomy. Cambridge University Press, 524 pp.
Marsh, O. C. (1877): New order of extinct Reptilia (Stegosauria) from the Jurassic of the Rocky Mountains.
American Journal of Science, Series 3, 14: 513-514.
Marsh, O. C. (1878): Principle characters of American Jurassic dinosaurs. Part 1. American Journal of Science,
16: 411-416.
Marsh, O. C. (1881): Classification of the Dinosauria. American Journal of Science, Series 3, 23: 81-86.
Marsh, O. C. (1885): Names of extinct reptiles. American Journal of Science, Series 3, 29: 169.
Marsh, O. C. (1897): Recent observations on European dinosaurs. Geological Magazine, Decade 4, 5: 6-9.
Martill, D. M. (1988): A review of the terrestrial vertebrate fossils of the Oxford Clay (Callovian-Oxfordian) of
England. Mercian Geologist, 11: 171-190.
Martill, D. M. (1993): Soupy substrates: a medium for the exceptional preservation of ichthyosaurs of the Posi-
donia Shales (Lower Jurassic) of Germany. Kaupia, Darmstädter Beitrage zur Naturgeschiste, 2: 77-97.

III Jornadas Internacionales sobre Paleontología de Dinosaurios y su Entorno70
D. M. Martill, S. Earland and D. Naish
Martill, D. M., Batten, D. J. and Loydell, D. K. (2000): A new specimen of the thyreophoran dinosaur cf. Sceli-
dosaurus with soft tissue preservation. Palaeontology, 43: 549-559.
Martill, D. M. & Naish, D. (2001): Dinosaurs of the Isle of Wight. Palaeontological Association, Field Guides to
fossils. 433 pp.
McIntosh, J. S. (1992): Sauropoda. In: Weishampel, D. B., Dodson, P., and Osmolska, H. (Eds). The Dinosauria.
University of California, Berkeley, pp. 345-401.
McGowan, C. (1976): The description and phylogenetic relationships of a new ichthyosaur genus from the Upper
Jurassic of England. Canadian Journal of Earth Sciences 76: 668-683.
Noè, L. F., Smith, D. T. J. and Walton, D. I. (2004): A new species of Kimmeridgian pliosaur (Reptilia; Saurop-
terygia) and its bearing on the nomenclature of Liopleurodon macromerus. Proceedings of the Geologists’
Association, 115: 13-24.
Nopsca, F. (1915): Die Dinosaurier der Sibenbürgischen Landesteile Ungarns. Mitteilungen Jahrbuch Ungaris-
che Geologische Reichsansalt, 23: 1-26.
Norman, D. B. (1998): On Asian ornithopods (Dinosauria: Ornithischia). 3. A new species of iguanodontid dino-
saur. Zoological Journal of the Linnean Society, 122: 291-348.
Owen, R. (1841): A description of a portion of the skeleton of the Cetiosaurus, a gigantic extinct saurian reptile
occurring in the oolitic formations of different portions of England. Proceedings of the Geological Society
of London 3, 80: 457-462.
Owen R. (1858): Note on the bones of the hindfoot of the Iguanodon, discovered and exhibited by S. H. Beckles.
Quarterly Journal of the Geological Society of London, 14: 174-5.
Owen, R. (1861): British fossil Reptilia of the Liassic formations, Part 1. A monograph on a fossil dinosaur (Sce-
lidosaurus harrisonii Owen) of the Lower Lias. Palaeontographical Society of London [Monographs],
13: 1-14.
Owen, R. (1875-7): Monograph of the fossil Reptilia of the Mesozoic formations. (Pt II) (genera Bothriospon-
dylus, Cetiosaurus, Omosaurus). Palaeontographical Society of London [Monographs], 29: 15-97.
Phillips, J. (1871): Geology of Oxford and the Valley of the Thames. Clarendon Press, Oxford, 529 pp.
Pocock, T. I. (1908): The geology of the country around Oxford. Memoirs of the Geological Survey of England
and Wales. Explanation of Special Oxford Sheet. London. 142 pp.
Powell, H. P. (1988): A megalosaurid dinosaur jawbone from the Kimmeridge Clay of the seabed of West Bay,
Dorset. Proceedings of the Dorset Natural History and Archaeological Society, 109: 105-7.
Prestwich, J. (1879): On the discovery of a species of Iguanodon in the Kimmeridge Clay near Oxford; and a
notice of a very fossiliferous band of the Shotover Sands. Geological Magazine, Decade 2, 6: 193-5.
Prestwich, J. (1880): Notes on the occurrence of a new species of Iguanodon in a brick-pit of the Kimmeridge
Clay at Cumnor Hurst, three miles WSW of Oxford. Quarterly Journal of the Geological Society of Lon-
don, 36: 430-2.
Raiswell, R., Newton, R. and Wignall, P. B. (2001): An indicator of water-column anoxia: Resolution of biofacies
variations in the Kimmeridge Clay (Upper Jurassic, UK). Journal of Sedimentary Research, Section A:
Sedimentary Petrology and Processes, 71 (2): 286 - 294.

Salas de los Infantes, Burgos 71
Dinosaurs in marine strata: evidence from the British Jurassic.
Richardson, L., Arkell, W. J. and Dines, H. G. (1946): Geology of the country around Witney. Memoirs of the
Geological Survey of Great Britain, 150 pp.
Salgado, L. and Calvo, J. O. (1997): Evolution of the titanosaurid sauropods II: the cranial evidence. Ameghi-
niana, 34: 33-48.
Sauvage, H. E. (1874): Mémoire sur les dinosauriens et les crocodiliens des terrains jurassiques de Boulogne-sur-
Mer. Mémoires de Société de Géologique de France (sér. 2), 4: 435-442.
Sauvage, H. E. (1888): Sur les reptiles trouvés dans le Portlandien supérieur de Boulogne-sur-Mer. Bulletin de
Société de Géologique de France (sér. 3), 16: 623-632.
Seeley, H. G. (1869): Index to the Fossil Remains of Aves, Ornithosauria and Reptilia, from the Secondary Sys-
tem of Strata Arranged in the Woodwardian Museum of the University of Cambridge. Deighton, Bell and
Co., Cambridge. 143 pp.
Sellwood, B. W. and Price, G.D. (1993): Sedimentary facies as indicators of Mesozoic palaeoclimate. Philoso-
phical Transactions of the Biological Sciences Royal Society, London, 341: 225-233.
Sereno, P. C. (1986): Phylogeny of the bird-hipped dinosaurs (Order Ornithischia). National Geographic Resear-
ch, 2: 234-256.
Taylor, M. A., Norman, D. B. and Cruickshank, A. R. I. (1993): Remains of an Ornithischian dinosaur in a plio-
saur from the Kimmeridgian of England. Palaeontology, 36: 357-360.
Tyson, R. V. (1996): Sequence stratigraphical interpretation of organic facies variations in marine siliclastic sys-
tems: general principles and application to the onshore Kimmeridge Clay Formation, UK. In: Hesselbro,
S. P. and Parkinson, D. P. (Eds). Sequence Stratigraphy in British geology. Geological Society Special
Publication, 103: 75-93.
Unwin, D. (1988): A new pterosaur from the Kimmeridge Clay of Kimmeridge Bay, Dorset. Proceedings of the
Dorset Natural History and Archaeological Society, 109: 150-153.
Upchurch, P. (1993): The anatomy, phylogeny and systematics of the sauropod dinosaurs. Unpublished PhD
thesis, University of Cambridge, 489 p.
Upchurch, P. (1995): The evolutionary history of sauropod dinosaurs. Philosophical Transactions of the Royal
Society of London, Series B, 349: 365-390.
Upchurch, P. and Martin, J. (2003): The anatomy and taxonomy of Cetiosaurus (Saurischia, Sauropoda) from the
Middle Jurassic of England. Journal of Vertebrate Palaeontology, 23: 208-231.
Valdes, P. J. (1993): Atmospheric general circulation models of the Jurassic. Philosophical Transactions of the
Biological Sciences Royal Society, London, 341: 317-326.
Weishampel, D. B., Jianu, C. –M., Csiki, Z. and Norman, D. B. (2003): Osteology and phylogeny of Zalmoxes
(N. G.), an unusual euornithopod dinosaur from the latest Cretaceous of Romania. Journal of Systematic
Palaeontology, 1: 65-123.
Wignall, P. B. (1990): Benthic palaeoecology of the late Jurassic Kimmeridge Clay of England. Special Papers
in Palaeontology, 43: 74 pp.
Wignall, P. B. (1993): The stratigraphy of the Upper Kimmeridge Clay (late Jurassic) of Golden Hill, Vale of
Pickering, North Yorkshire. Proceedings of the Yorkshire Geological Society, 49: 207-214.

III Jornadas Internacionales sobre Paleontología de Dinosaurios y su Entorno72
D. M. Martill, S. Earland and D. Naish
Wignall, P. B. and Pickering, K. T. (1993): Palaeoecology across a Jurassic fault scar, NE Scotland. Journal of
the Geological Society, London, 150: 323-340.
Woodward, H. B. (1895): The Jurassic rocks of Britain. Volume 5. The Middle and Upper Oolitic Rocks of En-
gland (Yorkshire excepted). Memoirs of the Geological Survey of the United Kingdom. 499 pp.
Zils, W., Werner, C., Moritz, A. and Saanane, C. (1995): Tendaguru, the most famous dinosaur locality of Africa.
Review, Survey and Future Prospects. Documenta Naturae, 97: 1-41.