diploma misa dinarska voluharica koncna - core.ac.uk · biro, m.: dolo čanje starosti in...
TRANSCRIPT

UNIVERZA V MARIBORU
FAKULTETA ZA NARAVOSLOVJE IN MATEMATIKO
Oddelek za biologijo
DIPLOMSKO DELO
Miša BIRO
Maribor, 2008


UNIVERZA V MARIBORU
FAKULTETA ZA NARAVOSLOVJE IN MATEMATIKO
Oddelek za biologijo
Miša BIRO
Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici, Dinaromys bogdanovi (Martino in Martino, 1922)
(Rodentia, Mammalia)
DIPLOMSKO DELO
Mentor: izr. prof. dr. Franc JANŽEKOVIČ
Somentor: red. prof. dr. Boris KRYŠTUFEK
Maribor, 2008

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
II
UNIVERZA V MARIBORU
FAKULTETA ZA NARAVOSLOVJE IN MATEMATIKO
IZJAVA
Podpisana Miša BIRO, rojena 4. 6. 1982, študentka Fakultete za
naravoslovje in matematiko Univerze v Mariboru, smer biologija in
kemija, izjavljam, da je diplomsko delo z naslovom Določanje starosti in
morfometrija meljaka m1 pri dinarski voluharici, Dinaromys bogdanovi,
(Martino in Martino, 1922) (Rodentia, Mammalia), pri mentorju izr. prof.
dr. Francu JANŽEKOVIČU in somentorju red. prof. dr. Borisu
KRYŠTUFKU, avtorsko delo. V diplomskem delu so uporabljeni viri in
literatura korektno navedeni; teksti niso prepisani brez navedbe avtorja.
___________________________
(podpis študenta-ke)
Maribor, 7. 5. 2008

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
III
BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski
voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia,
Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za
naravoslovje in matematiko, Oddelek za biologijo, 2008.
IZVLEČEK
Starost dinarske voluharice, Dinaromys bogdanovi, je bila ocenjena na
osnovi višine korenine prvega spodnjega meljaka. V analizo je bilo
zajetih 188 osebkov, nabranih na 36 različnih geografskih območjih
Dinaridov in združenih v tri regije (klade): zahodna, centralna in južna
regija. Največ osebkov je zastopanih iz centralne regije (118), iz
zahodne regije je zastopanih 31 osebkov in 24 iz južne regije. Analiza
meritev je pokazala, da sekundarni spolni dimorfizem ni prisoten.
Umrljivost samcev in samic je skozi vso življenje bolj ali manj enako. V
prvem starostnem obdobju imajo večjo sposobnost preživetja samice, v
tretjem (od 24 do 36 mesecev) pa samci. Največja pričakovana
življenjska doba je pri samicah v prvem starostnem obdobju do 12
mesecev (1,35 let); prav tako pri samcih, vendar je nekoliko nižja (0, 89
let).
Ključne besede: dinarska voluharica, spodnji meljak, višina korenine,
določanje starosti, starostna struktura, populacijska struktura.

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
IV
BIRO, M.: Age determination and morphometry of the first lower molar in
martino's vole, Dinaromys bogdanovi (Martino in Martino, 1922)
(Rodentia, Mammalia). Graduation Thesis. University of Maribor, Faculty
of nautral science and mathematics, Department of Biology, 2008.
ABSTRACT
Age of Martino's vole, Dinaromys bogdanovi, has been estimated on the
basis of the root height of the first lower molar. The research included
188 specimen collected from 36 different localities in Dinaric Alps
grouped into three regions: the Western, Central and Southern. Most
specimen were from the Central region (118), 31 were from the Western
region and 24 from the Southern region. The results showed no
secondary sexual dimorphism. Mortality is through whole life more or
less equal. Females from the first age group show greater life
expectancy than males of the same age, though males of the age 24 to
36 months (third age group) live longer than females. The greatest life
expectancy of females for the first age group is up to 12 months
(expected age 1,35 years), as well as in males only something lower
(0,89 years).
Descriptors: Martino's vole, lower molar, rooth length, age
determination, age structure, population structure.

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
V
KAZALO VSEBINE
IZJAVA.................................................................................................... II
IZVLEČEK ............................................................................................. III
ABSTRACT............................................................................................ IV
KAZALO VSEBINE .................................................................................V
KAZALO SLIK.......................................................................................VII
KAZALO TABEL ..................................................................................VIII
KAZALO GRAFOV................................................................................. IX
1 UVOD .................................................................................................. 1
1.1 Dinarska voluharica (Dinaromys bogdanovi) ............................ 1
1.1.1 Taksonomija dinarske voluharice............................................ 2
1.1.2 Opis dinarske voluharice......................................................... 3
1.1.3 Filogeografija in paleontologija ............................................... 5
1.1.4 Habitat in razširjenost ............................................................. 6
1.1.5 Prehranjevanje........................................................................ 8
1.1.6 Razmnoževanje ...................................................................... 9
1.2 Namen in delovne hipoteze......................................................... 10
1.2.1 Hipoteze................................................................................ 10
2 MATERIALI IN METODE ................................................................... 11
2.1 Material ....................................................................................... 11
2.2 Metode ........................................................................................ 15
2.2.1 Statistične analizne metode ................................................ 18
2.2.2 Sestavljanje življenjske preglednice.................................... 19
3 REZULTATI ....................................................................................... 21
3.1 Opisna statistika .......................................................................... 21
3.2 Sekundarni spolni dimorfizem ..................................................... 26

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
VI
3.3 Starostna struktura ...................................................................... 28
3.4 Starostna in spolna struktura po geografskih regijah................... 29
3.5 Življenjska preglednica in krivulja preživetja................................ 30
4 DISKUSIJA ........................................................................................ 33
5 POVZETEK........................................................................................ 36
6 ZAHVALA .......................................................................................... 37
7 LITERATURA .................................................................................... 38
8 PRILOGE........................................................................................... 41

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
VII
KAZALO SLIK
Slika 1: Dinarska voluharica, Dinaromys bogdanovi, endemit dinarskega
krasa (Fotografija: Alenka Kryštufek)................................................. 1
Slika 2: Zgornji (levo) in spodnji (desno) niz meljakov pri dinarski
voluharici, Dinaromys bogdanovi (legenda: PP – prednja pentlja, ZZ
– zadnja zanka, PZ – prednja zanka, ZP – zadnja pentlja, Tx –
trikotnik, Mx – zgornji meljaki, Mx – spodnji meljaki) (Kryštufek, 1987).
.......................................................................................................... 4
Slika 3: Gorovje Korab (vir:
http://en.wikipedia.org/wiki/Image:Panair_Korab.jpg pridobljeno 7. 5.
2008). ................................................................................................ 8
Slika 4: Lokalitete ulovljenih dinarskih voluharic, Dinaromys bogdanovi,
ki so bile vključene v analizo. Zaporedne številke prikazujejo
posamezne lokalitete, barve pa posamezne regije (Rumež, 2006). 12
Slika 5: Leva spodnja čeljustnica s sekalcem in tremi meljaki dinarske
voluharice, Dinaromys bogdanovi, meljak m1 je z označenimi
spremenljivkami............................................................................... 17
Slika 6: Spodnja čeljustnica dinarske voluharice, pri kateri zobna
korenina prvega meljaka še ni razvita. ............................................ 22
Slika 7: Spodnja čeljustnica dinarske voluharice, kjer je razvita zobna
korenina. Višina korenine prvega meljaka je skoraj na polovici
celotne višine zoba. ......................................................................... 22
Slika 8: Spodnja čeljustnica dinarske voluharice, kjer predstavlja višina
korenine prvega meljaka skoraj celo višino zoba. ........................... 23

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
VIII
KAZALO TABEL
Tabela 1: Skupen seznam lokalitet in število dinarskih voluharic,
Dinaromys bogdanovi, vključenih v analizo. .................................... 13
Tabela 2: Opisna statistika meritev 188 vzorcev dinarske voluharice. . 24
Tabela 3: Opisna statistika za zahodno klado. ..................................... 24
Tabela 4: Opisna statistika za centralno klado. .................................... 25
Tabela 5: Opisna statistika za južno klado. .......................................... 25
Tabela 6: Rezultati enosmerne analize variance testiranja sekundarnega
spolnega dimorfizma pri dinarski voluharici, Dinaromys bogdanovi, za
prvi starostni razred. ........................................................................ 26
Tabela 7: Rezultati enosmerne analize variance testiranja sekundarnega
spolnega dimorfizma pri dinarski voluharici, Dinaromys bogdanovi, za
drugi starostni razred....................................................................... 27
Tabela 8: Rezultati enosmerne analize variance testiranja
sekundarnega spolnega dimorfizma pri dinarski voluharici,
Dinaromys bogdanovi, za tretji starostni razred............................... 27
Tabela 9: Koledarska leta izražena kot delež molarnega indeksa. ....... 28
Tabela 10: Velikost vzorcev glede na koledarsko leto. ......................... 28
Tabela 11: Starostna in spolna struktura populacij. .............................. 29
Tabela 12: Življenjska preglednica za samice in samce dinarske
voluharice Dinaromys bogdanovi..................................................... 31
Tabela 13: Osnovi podatki o dinarski voluharici in meritve. .................. 41

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
IX
KAZALO GRAFOV
Graf 1: Krivulja preživetja za samce in samice dinarske voluharice......32

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
1
1 UVOD
1.1 Dinarska voluharica (Dinaromys bogdanovi)
Glodavci so najštevilčnejša skupina sesalcev (predstavljajo več kot 40%
vseh sesalcev). So uspešna in prilagodljiva skupina. Ena izmed
poddružin glodavcev so voluharice (Arvicolinae), med katere uvrščamo
tudi dinarsko voluharico. Dinarsko voluharico so do leta 1922 poznali le
kot fosilno vrsto, tega leta pa sta Vladimir in Evgenija Martino našla živi
primerek. Tedaj seveda še nista vedela, da sta našla prvi (zabeležen)
živi primer tega rodu, kar kaže dejstvo, da sta novo vrsto prvotno opisala
kot Microtus (Chionomys) bogdanovi. Šele nadaljnja proučevanja so
pokazala, da gre za dinarsko voluharico, endemičnega in reliktnega
glodavca dinarskega krasa. Leta 1955 so jo uvrstili v rod Dinaromys. V
literaturi jo lahko zasledimo pod več imeni; najpogosteje kot dinarska
voluharica, pa tudi kot reliktna voluharica in Martinojeva voluharica.
Slika 1: Dinarska voluharica, Dinaromys bogdanovi, endemit dinarskega krasa
(Fotografija: Alenka Kryštufek).

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
2
1.1.1 Taksonomija dinarske voluharice
Za glodavce je diagnostično zobovje. Zobna formula voluharic je:
i1/1 c0/0 p0/0 m3/3 = 16, kar pomeni, da je v vsaki čeljustnici po en
sekalec, podočnikov ni, prav tako ni predmeljakov, meljaki pa so trije.
Torej je med sekalci in meljaki diastema.
Za nekatere voluharice je značilno, da imajo hipsodontne kočnike, ki
stalno rastejo. Takšni kočniki so se razvili pri tistih glodavcih, ki se
hranijo z najmanj hranljivim rastlinjem in se jim zato zobje hitro
obrabljajo. Pri dinarski voluharici pa se je pojavila druga posebnost,
namreč meljaki starejših osebkov razvijejo korenino, kar velja pri
voluharicah za primitiven znak.
Sistematska ureditev družine miši, voluharice in hrčki ni stabilna, na kar
kažejo tudi večkratni poskusi uvrstitve dinarske voluharice v sistem.
Različni avtorji so jo uvrstili pod različnimi imeni v različne rodove oz.
skupine. Tako jo je Sergej I. Ognjev leta 1984 prvi uvrstil pod imenom
Dolomys bogdanovi, skupaj z rodovoma Clethrionomys in
Prometheomys. Hooper in Hart (1962) ter Gromov in Poljakov (1977) so
rod uvrstili v pleme Clethrionomyni (Kolarič, 1999). Kretzoi je rod
Dinaromys najprej uvrstil v pleme Ondatrini, zaradi podobnosti z
rodovoma Ondatra in Neofiber, kasneje pa ga je prestavil v pleme
Pliomyni (Kretzoi, 1969), kot edinega recentnega predstavnika.

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
3
Uvrstitev v sistem, povzeta po Kretzoiu (1969):
(deblo): Chordata (strunarji)
(poddeblo): Vertebrata (vretenčarji)
(razred): Mammalia (sesalci)
ordo (red): Rodentia (glodavci)
familia (družina): Muridae (miši)
subfamilia (poddružina): Microtinae
tribus (pleme): Pliomyni Kretzoi, 1969
subtribus (podpleme): Pliomyi Kretzoi, 1969
genus (rod): Dinaromys Kretzoi, 1955
species (vrsta): bogdanovi (Martino in Martino, 1922)
1.1.2 Opis dinarske voluharice
Dinarska voluharica je srednje velika žival, skupaj z glavo meri od 99 do
152 mm. Dlake na hrbtu so pri odraslih živali rjavkaste, pri mladih pa
modrikaste barve. Trebušna stran je sivo-bela. Kožuh je mehak, gost in
dolgodlak, najdaljše dlake na hrbtu so dolge 17–18 mm. Pozimi je kožuh
gostejši in temnejši kot poleti, kar je posledica črnih konic dlak. Slednje
se tekom zime obrabijo in kožuh posvetli. Menjava dlake se prične poleti
in traja čez vso poletje in jesen. Dlake na nogah so bele barve; noht je
razvit le na palcu, na ostalih prstih pa so ostri kremplji, ki so dolgi od 3
do 3,3 mm. Za voluharico ima relativno dolg rep, od 60–75 mm. Rep je
dvobarven, dorzalno je siv, ventralno in ob straneh pa bel. Konice dlak
na repu so bele, dolge 4–6 mm. Ušesa so relativno velika, od 16, 8–19,
7 mm, na notranji in zunanji strani so pokrita s kratkimi dlačicami in malo
štrlijo od kožuha. Smrček je dokaj dolg, na koncu smrčka je območje
srsti, ki je veliko in gosto. Najdaljša srst zgornje ustne je dolga do 60
mm, povprečno število srsti je 62,8.

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
4
Sekalci dinarske voluharice so relativno majhni; meljaki imajo navadno 2
pentlji in razvijejo tekom postnatalnega razvoja korenino. Sprednja krona
m1 je enostavna, prva zunanja sinklinala na m3 je navadno zelo ploska,
tretja zunanja zanka na m3 je jasna in pogosto sodeluje z zadnjim
lobusom zoba.
Slika 2: Zgornji (levo) in spodnji (desno) niz meljakov pri dinarski voluharici,
Dinaromys bogdanovi (legenda: PP – prednja pentlja, ZZ – zadnja zanka, PZ –
prednja zanka, ZP – zadnja pentlja, Tx – trikotnik, Mx – zgornji meljaki, Mx –
spodnji meljaki) (Kryštufek, 1987).

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
5
1.1.3 Filogeografija in paleontologija
Dinarska voluharica je edina preživela vrsta tega rodu in poseljuje gore
zahodnega Balkana (Petrov, 1992). V pleistocenu so med glacialnima
cikloma nastale klimatske spremembe, ki so povzročile spremembe oz.
premike pri vseh evropskih taksonih. Zaradi omejenosti primernih
habitatov v takšnih razmerah se je njeno območje v poznem pleistocenu
zožilo, posledica tega je najmanjša razširjenost med evropskimi sesalci.
Večina filogeografskih študij je pokazala, da sta južna Evropa in Balkan
v tem času delovala kot glacialno zatočišče.
Dinarsko voluharico obravnavajo kot paleoendemita zahodnega
Balkana.
Po zunanjosti so dinarsko voluharico najprej uvrstili v rod Chionomys.
Podrobnejše raziskave so pokazale, da meljaki pri dinarski voluharici
razvijejo korenino, kar je kazalo na bolj unikaten položaj med evropskimi
sesalci (Hinton, 1926). Paleontološki dokazi kažejo, da je bila
razširjenost monotipskega rodu Dinaromys vedno majhna in verjetno
izključno omejena na zahodni in južni Balkan. Iz fosilnih najdb iz
pliocena so poznane le tri vrste Dinaromys, od katerih sta dve izumrli.
Omenjeno kaže na nizko stopnjo evolucije.
Todorović (1956) je razdelil dinarsko voluharico v dve glavni starostni
liniji, osnovani na morfologiji prvega meljaka in kasnejši študiji
prekrivanja dveh vrst. Elektroforetske raziskave frekvenc alelov na 28
lokusih so pokazale visok nivo genetske delitve dveh morfotipov –
poddružin miši Muridae, ter da je rod Dinaromys sestrski takson
Clethrionimys – klade Microtus (Gill in sod., 1986).
Prostorska genetska struktura dinarske voluharice je bila raziskana z
uporabo 555 bp sekvence mitohondrijskega gena citokroma b. Študija
temelji na 63 primerkih iz 18 različnih lokacij celotnega območja, ki ga
vrsta poseljuje. Med 47 halotipi sta bili prepoznani 2 kladi, prva obsega
vzorce 4 geografskih območij jugovzhodno od reke Neretve na

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
6
Hrvaškem ter zahodni Bosni in Hercegovini, to je tako imenovana
zahodna klada, druga pa vse ostale vzorce. Večja klada je bila nadalje
ločena v dve podskupini, poskusno ločeni z reko Drim. Prva zajema
preostale vzorce iz Bosne in Hercegovine, Črne gore in Kosova, tako
imenovana centralna klada, druga, tako imenovana južna klada, pa
zajema vzorce iz Makedonije. Med slednjima filoskupinama ni bilo
deljenih haplotipov. Pomembna je razlika v nukleotidih, ki je bila največja
v zahodni kladi (π = 0,016), najmanjša pa v južni kladi (π = 0,0062). Na
podlagi pridobljenih podatkov je bila postavljena hipoteza, da se je pred
približno 1 milijonom let zahodna klada začela širiti južneje, preko reke
Neretve. Tako bi naj kasneje z alopatrijo nastala centralna klada. Pred
približno 0,3 milijoni let bi se naj začela širiti južneje tudi centralna klada,
preko reke Drim ter se postopoma ločevala v sedanjo južno klado, prav
tako z alopatrijo. Omenjeno je podprto tudi s paleontološkimi dokazi
(Kryštufek in sod., 2007).
1.1.4 Habitat in razširjenost
Habitat dinarske voluharice je unikaten, sestavljen iz kamenja, v obliki
izoliranih gora, osamelih skal ali popolnoma kamnitega terena. Živi v
razpokah in votlinah apnenčastih skal, kot jih nudi predvsem Kras. Našli
so jo tako v majhnih luknjah, kot pri vhodih v večje jame. Raje ima
apnenec, vendar so jo zasledili tudi na tleh druge sestave.
J. Martino predpostavlja, da je premer dostopov v brloge v skalnatih
razpokah velik 8 do 12 cm. To bi lahko bil tudi razlog, zakaj Martinu ni
uspelo odkopati oz. razkriti gnezda dinarske voluharice.
Primerni bivalni prostori so pogosto v veliki meri izolirani, tako da je
kontakt s sosednjimi populacijami težko predstavljiv. Zasledili so le
manjšo, krajevno zamenjavo prostorov, in sicer spomladi. Zaradi taljenja
snega in dežja postanejo skalnate razpoke mokre, zato jih živali
zapustijo, vendar jih čez poletje spet poiščejo (Martino, 1948). Naključno

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
7
so našli dinarsko voluharico v pastirskih kočah in drugih hišah, na primer
postavljenih opazovalnicah na vrhovih Bjelašnice (Bolkay, 1927).
Dinarsko voluharico so našli na 10 m nadmorske višine (Tvrtković, 1994)
in vse do 2200 m, v glavnem pa živijo na nadmorski višini 1400 m. Na
nižjih legah živijo predvsem na območju Krasa, in sicer na pečinah,
orientiranih k Jadranu, na višjih nadmorskih višinah pa se biotop približa
severni in vzhodni arealni meji.
Populacije so majhne in široko razpršene prav zaradi točkaste narave
njenega habitata. Mnogo lokalnih populacij smatrajo za visoko ogrožene
in nekatere so, kljub rasti zavesti o njihovi ohranitvi (Mitchell-Jones in
sod., 1999), verjetno že izumrle (Tvrtković, 1994). Namreč vsi trije
kriteriji 7-delnega modela redkosti po Rabinowitz in sod. (1986) so
značilni za dinarsko voluharico, najbolj pa so izraženi pri zahodni kladi.
Večji del areala dinarske voluharice obdaja Jadran, le manjši del
Egejsko in Črno morje. V pleistocenu se je razprostiral od severne Italije
do vzhodne Grčije, na kar kažejo fosilna nahajališča (Grotte di Veia
(Bartolomei in Broglio 1975), Bosco Chiesanouva (Bartolomei 1977),
Malo I., San Vito di Leguzzano (Bartolomei 1970), Colli Barici
(Bartolomei 1964), Ilirska Bistrica (Bartolomei 1970), Varkiza I, Atene
(Weerd 1973)). V Sloveniji so, poleg okolice Ilirske Bistrice, našli fosile
tudi v okolici Logatca in črnokalske prelomnice (ustno Kryštufek, 1997;
povzeto po Kolarič, 1999). Današnji življenjski prostor sega od
severnega Velebita in Male Kapele na Hrvaškem, preko Bosne in
Hercegovine, Črne gore, Kosova do Galičice v Makedoniji (Petrov in
Todorović, 1982). Južna meja je neznana, zelo verjetno sega v Albanijo
in morda celo v severozahodno Grčijo.

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
8
Slika 3: Gorovje Korab (vir: http://en.wikipedia.org/wiki/Image:Panair_Korab.jpg
pridobljeno 7. 5. 2008).
1.1.5 Prehranjevanje
Odvisno od nadmorske višine se spreminja tudi vegetacija: od makije,
preko listnatih gozdov do iglastih gozdov. To vpliva na prehrano dinarske
voluharice, ki je izrazit rastlinojed. Hrani se predvsem z zelenjem, ki
raste v bližini njihovih bivališč. Med letoma 1946 in 1948 je v mesečnih
intervalih J. Martino izvedel ulov dinarske voluharice v osrednji Bosni.
Poleg lovišč v gorovju Trebević je našel Martino dele rastlin Festuca
ovina, Agropyrum repens, Veronica chamaedrys, Cirsium sp., Scabiosa
leucophylla, Cephalaria sp., Helianthenum vulgare, Potentilla sp.,
Galium sp., Thalictrum sp., Asarum europaeum, Aremonia
agrimoniodes, Arabis alpina, Convolvulus arvensis, Ajuga reptans in
Hylocomium proliferum.
Čeprav se je zdelo, da je Martino v tem času zbral bogat material, so
poročila le v obliki naključnih opazovanj. Tako je zapisano, da je pred
spomladjo vsebina želodca dinarske voluharice vonjala po lesu iglavcev,
maja in junija pa so v skalnih razpokah našli vršičke jelke. Dinarsko
voluharico je prav tako opazoval M. Milenković, in sicer konec maja v
gorovju Orjen, v coni gozda. Poročal je, da je dinarska voluharica čez
dan večkrat splezala na skalo, odtrgala mlade bukove liste in vejice ter
jih hitro odnesla v svoje skrivališče, poleg pa je nabrala še zelenje. Na

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
9
podlagi navedenega ter ostankov delov rastlin pred bivalnim prostorom
dinarske voluharice je J. Martino (1948) sklepal, da le-ta dela zimske
zaloge. Kljub pomanjkljivim dokazom iz zimskih obdobij, so nekateri
znanstveniki mnenja, da je dinarska voluharica aktivna skozi vso leto. Po
pričevanjih J. Martina (1948) in Gligića (1959) je dinarska voluharica
predvsem nočna žival, vendar govorijo številna opazovanja tudi o dnevni
aktivnost, med drugim zgoraj omenjeno opazovanje M. Milenkovića.
1.1.6 Razmnoževanje
Tekom dvoletnih opazovanj dinarske voluharice v gorovju Trebević je J.
Martino (1948) zabeležil v enem letu dve paritveni obdobji: prvo v
mesecu marcu, drugo v mesecu juniju. Kolarič (1999) ugotavlja, da bi naj
imela še tretje paritveno obdobje konec meseca septembra, saj so bile
nekatere dinarske voluharice, ki so jih ulovili oktobra, breje. Brejost traja
približno 1 mesec, kar je za voluharico relativno dolgo. Povprečno število
embrijev je 2, redko 3. Prvi mladiči so bili ulovljeni v juniju.
Temu logično sledi populacijska dinamika – na začetku poletja
dominirajo odrasli. V obdobju od avgusta do sredine septembra je število
mladičev in odraslih bolj ali manj izenačeno, oktobra pa prevladujejo
mladiči. Voluharice živijo približno 2 leti. Petrov in Todorović (1982) sta iz
razpoložljivih podatkov o dinarski voluharici ugotovila, da dosežejo
nekatere tudi tretje leto starosti, en samec pa bi naj bil celo v četrtem
letu starosti.

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
10
1.2 Namen in delovne hipoteze
Meljaki dinarske voluharice razvijejo med postnatalnim razvojem
korenino, kar omogoča uporabo dolžine korenine za določitev absolutne
starosti osebkov.
V diplomski nalogi želim predstaviti morfometrijo zoba, natančneje
prvega spodnjega meljak tekom postnatalnega razvoja dinarske
voluharice. Na podlagi dolžine korenine meljaka m1 ovrednotiti starost
posameznega osebka, starostno strukturo populacij iz geografskih
vzorcev ter sekundarni spolni dimorfizem.
Cilji
• Določitev prisotnosti sekundarnega spolnega dimorfizma.
• Predstavitev starostne strukture na podlagi velikosti korenin m1.
1.2.1 Hipoteze
• Predpostavljam, da pri razvoju zoba dinarske voluharice spolni
dimorfizem ni prisoten.
• Predpostavljam, da je mogoče na osnovi razvoja m1 ovrednotiti
starostno strukturo populacij dinarske voluharice.

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
11
2 MATERIALI IN METODE
2.1 Material
V statistično analizo sem vključila 188 vzorcev čeljusti dinarske
voluharice, Dinaromys bogdanovi, ki so imele ohranjen prvi spodnji levi
meljak (m1). Večina materiala je last Prirodoslovnega muzeja Slovenije
(PMS) – zbirka obsega 167 osebkov, nekaj jih je last Prirodoslovno-
matematične fakultete sveučilišča (PMF) iz Zagreba (zbirka obsega 4
osebke), ostali pa so last Zemeljskega muzeja Bosne in Hercegovine
(ZMS) iz Sarajeva (zbirka obsega 17 osebkov). Živali so bile ulovljene na
različnih lokalitetah, ki so uvrščene v tri geografsko ločene regije:
zahodno (W), centralno (C) in južno regijo (S) – mejo predstavljata reki
Neretva in Drim, ter so prikazane na sliki 2.
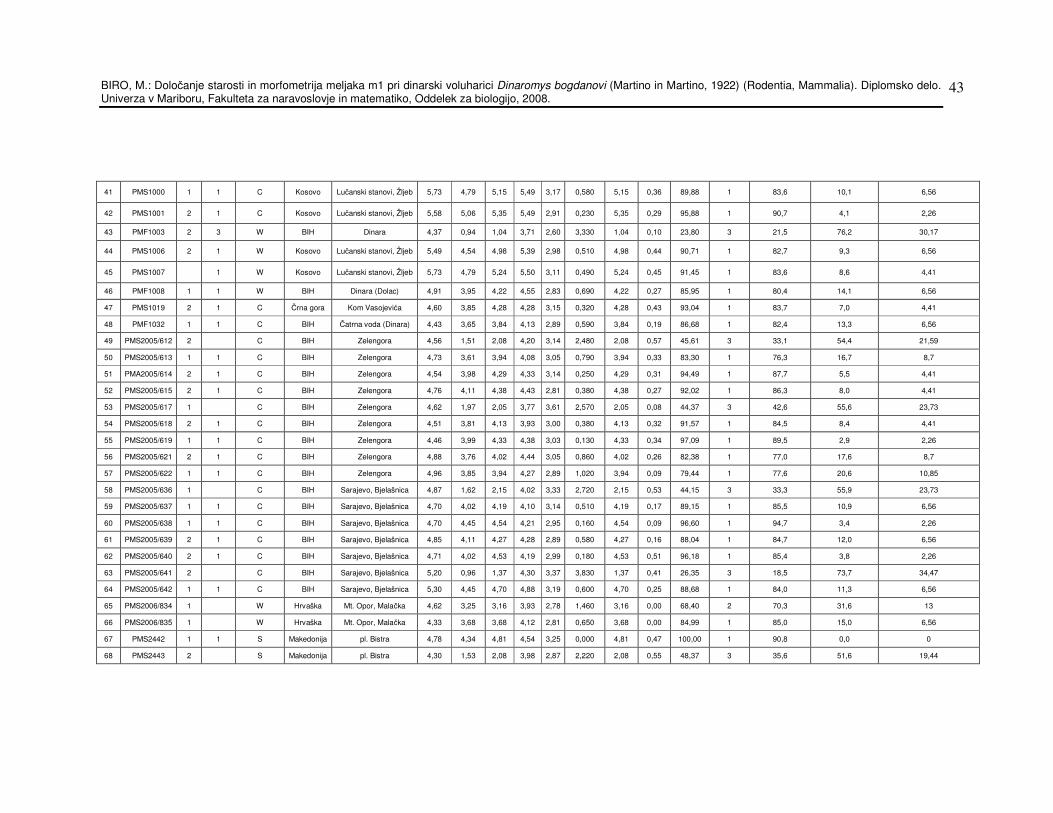
Inventarne številke osebkov in lokalitete so navedeni v prilogi 1, tabela
št.13. Tabela 1 pa prikazuje skupen seznam lokalitet in število osebkov
ulovljenih oz. najdenih na posamezni lokaciji.

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
12
Slika 4: Lokalitete ulovljenih dinarskih voluharic, Dinaromys bogdanovi, ki so
bile vključene v analizo. Zaporedne številke prikazujejo posamezne lokalitete,
barve pa posamezne regije (Rumež, 2006).

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
13
Tabela 1: Skupen seznam lokalitet in število dinarskih voluharic, Dinaromys
bogdanovi, vključenih v analizo.
Zap. št. Geografska regija Lokaliteta Število osebkov
1 S Korab 2
2 S Jablanica 3
3 S Bistra 14
4 S Stogovo 3
5 S Galičica 5
6 S Jakupica 2
7 S Šar planina 6
8 C Bjelašnica 11
9 C Trebević 10
10 C Gacko 1
11 C Bjelasica 28
12 C Zelengora 28
13 C Žljeb 7
14 C Kom 3
15 C Vilusi 2
16 C Durmitor 7
17 C Prenj 3
18 C Mostar 1
19 C Ljubinje 3
20 C Lovčen 2
21 C Sutorman 1
22 C Orjen 12
23 C Čapljina 1
24 C Maglić 1
25 C Orebić 1
26 C Čemerno 3
27 C Lebršnik 3
28 C Vrelo 1
29 C Čatrna voda 1

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
14
30 W Dinara 2
31 W Šatorsko jezero 2
32 W Split (Malačka) 5
33 W Cincar 10
34 W Paklenica 1
35 W Poklečani 1
36 W Opor 2

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
15
2.2 Metode
Poznamo različne metode za določanje relativne ali absolutne starosti
živali. Razdelimo jih lahko v 4 skupine: spremembe v velikosti in masi,
degeneracija struktur, določanje absolutne starosti s pomočjo rastnih linij
(rastni krogi, obroči) in ocenitev relativne starosti s pomočjo rastnih
sprememb. Kot je znano, se zobovje pri sesalcih tekom življenja
spreminja. Pri dinarski voluharici meljaki razvijejo korenino. S starostjo
osebka korenina raste, torej je višina korenine vedno večja. Tako lahko
na podlagi meritve višine korenine določimo starost osebkov.
Za določanje starosti sem uporabila rentgenske posnetke prvega
spodnjega meljaka leve čeljustnice.
Najprej sem pripravila material za rentgensko slikanje. Na kartončke
velikosti približno 15 cm krat 10 cm, sem z lepilnim trakom prilepila
spodnje čeljustnice vseh 189 vzorcev, obrnjene z lingvalno stranjo
navzgor. Sproti sem k posamezni čeljusti pripisala inventarno številko,
kot je bilo zabeleženo na oznaki embalaže, v kateri so bile hranjene
čeljusti. Na kartončke sem ustrezno dopisala območje, kjer je bil material
nabran, da sem se kasneje lažje orientirala. Na enak način smo kasneje
označili tudi rentgenske posnetke.
Material, pritrjen na kartončkih, smo slikali z mamografom PLANMED
Sophie. Mamograf je zaradi nizkih doz (doza je določena energija
rentgenskega sevanja, ki jo prejme posamezno tkivo ali organ)
primernejši za slikanje drobnega kostnega tkiva od klasičnega rentgena.
Daje tudi bolj natančne slike kostnega in zobnega tkiva, kar je za
merjenje spremenljivk na m1 zelo pomembno.

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
16
Za slikanje smo uporabili mamografsko folijo MAMORAY screens in film
AGFA DETAIL 18x24 cm. Tehnika slikanja je bila mehka tehnika slikanja
– mamografska tehnika z nizko napetostjo. Razdalja med fokusom in
filmom je bila 60 cm. Ta razdalja je pomembna, saj je slika tem boj
realna, čim bolj približujemo objekt. Slika objekta je bila povečana za
faktor 1,8. Pri tej povečavi ter napetosti 21 kV in količini žarkov 35 mAs
smo dobili najbolj jasne in najprimernejše slike za nadaljnjo obdelavo.
Na enega izmed kartončkov s pritrjenimi čeljustmi dinarskih voluharic
smo položili tudi objekt z znanim premerom (izravnano sponko). To nam
je kasneje pomagalo pri preračunavanju dobljenih meritev v programu
TPS iz »pikslov« v milimetre.
Ves material smo slikali na radiološkem oddelku Univerzitetnega
kliničnega centra Maribor.
Rentgenske posnetke čeljusti sem skenirala v digitalno obliko, meritve
pa izvedla s pomočjo paketa računalniških programov TPS. Po
naslednjem izračunu sem preračunala meritve iz »pikslov« v milimetre:
a×0,85x = ×1,8
17,9,
pri čemer je x meritev preračunana v milimetre; a je posamezna meritev
m1; 0,85 je meritev uporabljenega objekta z znanim premerom (sponke)
v milimetrih; 17,9 je izmerjena višina sponke (premer) v pikslih, 1,8 pa je
faktor povečave objekta pri slikanju z mamografom.

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
17
V nadaljevanju sem izračunala še višino korenine spodnjega meljaka in
višino vratu zoba. S pomočjo slednjih pa indekse, in sicer molarni,
skleninski in koreninski indeks. Molarni indeks je parameter, ki izraža
višino krone kot delež višine zoba. Skleninski indeks izraža višino
sklenine (meritev 2, slika 4) kot delež višine zoba. Koreninski indeks
izraža višino korenine kot delež višine zoba.
Nato sem meritve statistično obdelala s pomočjo programa SPSS 13.0.
Pri 189 slikah čeljusti dinarskih voluharic sem izvedla 5 meritev (slika 4)
na 1. spodnjem meljaku (m1), in sicer:
• višina zoba iz anteriorne strani (1),
• višina zareze (2),
• višina krone (3),
• višina m1 iz posteriorne strani (4) ter
• dolžina zoba (5) – vertikalno.
Slika 5: Leva spodnja čeljustnica s sekalcem in tremi meljaki dinarske
voluharice, Dinaromys bogdanovi, meljak m1 je z označenimi spremenljivkami.

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
18
2.2.1 Statistične analizne metode
Za statistično obdelavo podatkov sem uporabila naslednje matematične
operacije in izraze.
Aritmetična sredina
Aritmetična sredina je matematično izračunana vrednost normalne
distribucije numeričnih podatkov, ki se nahaja na sredini celotne
distribucije. Dobimo je, če vsoto vrednosti opažanj delimo s številom
opažanj (n), npr. 1, 2, 3, 4, 5 → x = 3). Je najbolj natančna srednja
vrednost v primerih, ko imamo simetrične in homogene porazdelitve.
Minimum
Minimum je najmanjša vrednost.
Maksimum
Maksimum je vrednost, ki je ne presega nobena druga vrednost,
največja vrednost.
Standardna deviacija
Je kvadratni koren variance. Uporabljamo jo kot merilo za variabilnost
posameznih vrednosti okrog srednje vrednosti ( 2s s= ).
ANOVA oz. F-test (enosmerna analiza variance)
ANOVA lahko imenujemo tudi f-test oz enosmerna analiza variance. S
pomočjo tega testa ugotovimo ali se varianci dveh vzorcev razlikujeta
statistično značilno ali le naključno (Sokal in Rohlf, 1995).
S pomočjo F-testa sem preverila razlike med tremi geografskimi vzorci in
razlike med spoloma znotraj posamezne geografske skupine. Postavila
sem dve hipotezi:

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
19
• ničelna hipoteza, ki pravi: če je stopnja tveganja večja od 5 %
oz. p > 0,05, se varianci dveh vzorcev razlikujeta naključno. Torej
lahko rečemo, da med primerjanimi vzorci ni razlik.
• alternativna hipoteza, ki pravi: pri dani stopnji tveganja, ki je
manjša od 5 % oz. p < 0,05 se vzorca razlikujeta statistično
značilno, kar pomeni, da med vzorci obstajajo razlike.
V nadaljevanju sem na podlagi višine korenine s pomočjo tabele 34 v
Kolaričevi diplomski nalogi (1999) ocenila starost posameznih osebkov
ter jih uvrstila v starostne razrede. (glej prilogo 1).
2.2.2 Sestavljanje življenjske preglednice
Življenjska preglednica daje popolno sliko o umrljivosti za določeno
vrsto. Ko sem ocenila starost 188 osebkov, sem jih razvrstila v razrede.
V prvem razredu se nahajajo osebki brez korenine. V ostalih razredih so
zajeti osebki z razvito korenino, njihova starost je eno, dve, tri ali štiri
leta. Število živali v enem razredu je dalo parameter nx, iz tega pa sem
lahko izračunala druge parametre dx, lx, qx, Lx in ex, kjer je:

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
20
dx – število poginulih osebkov v starostnem obdobju, računano na 1000
rojenih osebkov;
lx – število preživelih v začetku starostnega obdobja, računano od 1000
rojenih;
qx – umrljivost na 1000 živih ob začetku starostnega obdobja
xx
x
dq = x1000
l
Lx – število živih osebkov, ki je povprečno preživelo starost v starostnem
razredu
x x-1x
l + lL =
2
ex – pričakovana življenjska doba glede na doseženo starost (v letih)
∞
∑ xx
xx
Le =
l

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
21
3 REZULTATI
3.1 Opisna statistika
V analizo morfometrije m1 sem vključila živali iz 36 lokalitet (tabela 1), ki
so združene v 3 geografske regije: zahodno, centralno in južno regijo.
Za 188 vzorcev čeljusti dinarske voluharice Dinaromys bogdanovi, ki so
imele ohranjen prvi spodnji meljak, sem naredila 5 meritev (slika št. 4), ki
so prikazane v tabeli 13 (priloga 1).
Legenda za starost (povzeto po metodologiji Kolariča, 1999):
1. osebki, mlajši od 4-ih mesecev (1),
2. osebki, stari od 4 do 12 mesecev (2),
3. osebki, stari od 12 do 24 mesecev (3),
4. osebki, stari od 24 do 36 mesecev (4) in
5. osebki, stari nad 48 mesecev (A).
Legenda za spol:
1 – samec,
2 – samica.
Rezultat slikanja z mamografom so elektronski posnetki spodnjih
čeljustnic dinarske voluharice. Na spodnjih slikah je postopno prikazan
razvoj korenine. Absolutna višina zoba ostaja bolj ali manj
nespremenjena, saj se zobna krona tekom staranja zmanjšuje na račun
rasti korenine, pa tudi zaradi obrabe zoba, kot posledice prehranjevanja.

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
22
Slika 6: Spodnja čeljustnica dinarske voluharice, pri kateri zobna korenina
prvega meljaka še ni razvita.
Slika 7: Spodnja čeljustnica dinarske voluharice, kjer je razvita zobna korenina.
Višina korenine prvega meljaka je skoraj na polovici celotne višine zoba.

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
23
Slika 8: Spodnja čeljustnica dinarske voluharice, kjer predstavlja višina
korenine prvega meljaka skoraj celo višino zoba.
Analiza združenih vzorcev prikazuje minimalno in maksimalno meritev
višine zoba iz anteriorne strani, višino zareze, višino krone, višino m1 iz
posteriorne strani in dolžino zoba – vertikalno, kar prikazuje tabela 2.
Prav tako so razvidni parametri: višina korenine, višina krone ter vratu.
Število meritev ni povsod enako. Do razhajanj je prišlo, ker določenih
parametrov nisem mogla izmeriti, predvsem zaradi slabše vidnih
posnetkov.

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
24
Tabela 2: Opisna statistika meritev 188 vzorcev dinarske voluharice.
Valid N Mean Minimum Maximum Std.Dev.
m1 188 4,83 4,20 5,98 0,33
m2 183 3,10 0,69 5,06 1,12
m3 183 3,44 0,79 5,35 1,05
m4 187 4,36 3,68 5,50 0,34
m5 187 3,18 2,40 3,79 0,27
Ko 183 1,40 0,00 4,07 1,04
Kr 183 3,44 0,79 5,35 1,05
vrat 182 0,36 0,00 1,60 0,27
molarni indeks 183 71,07 16,25 100,00 21,34
skleninski indeks 183 64,21 14,20 99,35 22,91
koreninski indeks 183 28,95 0,00 83,75 21,31
Statistično analizo sem naredila posebej tudi za vsako klado.
Tabela 3: Opisna statistika za zahodno klado.
Valid N Mean Minimum Maximum Std.Dev.
m1 25 4,77 4,28 5,73 0,38
m2 25 3,36 0,94 4,79 1,06
m3 25 3,64 1,04 5,24 1,05
m4 25 4,42 3,70 5,50 0,45
m5 25 2,99 2,60 3,69 0,26
Ko 25 1,13 0,00 3,33 0,95
Kr 25 3,64 1,04 5,24 1,05
vrat 25 0,28 0,00 0,68 0,19
molarni indeks 25 76,07 23,80 100,00 20,53
starostni razred 25 1,52 1,00 3,00 0,77
skleninski indeks 25 70,40 21,51 95,32 21,22
koreninski indeks 25 23,92 0,00 76,20 20,53
BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
25
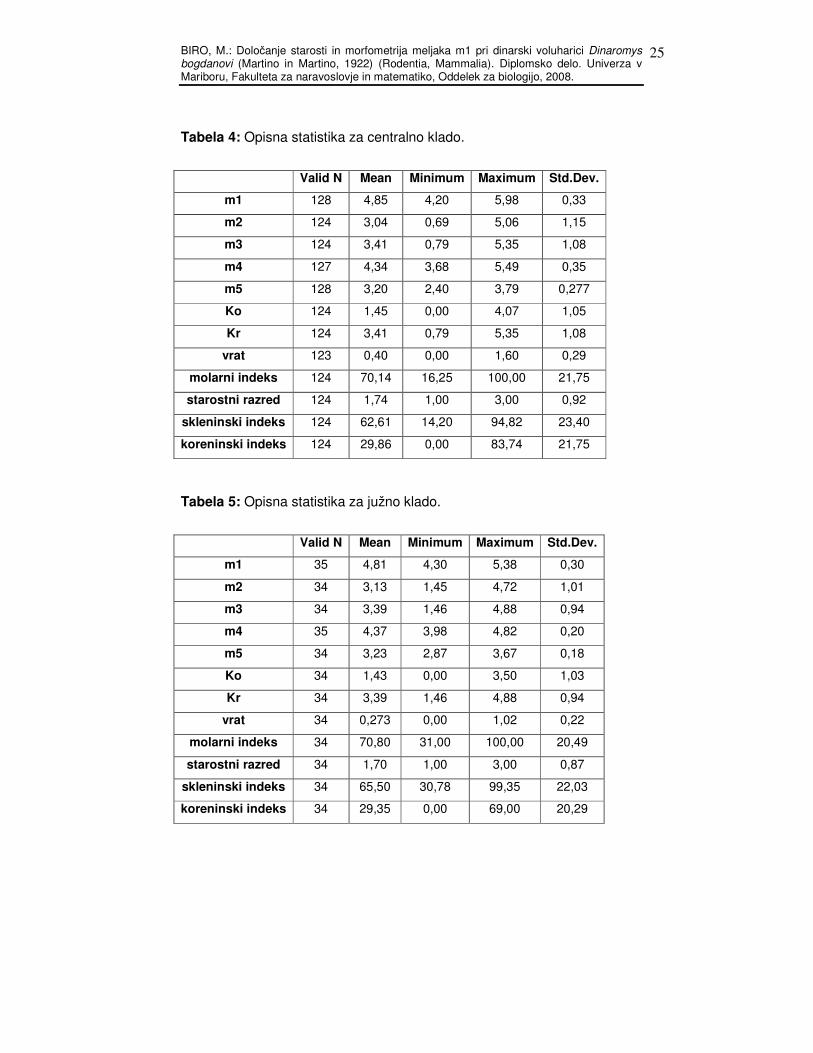
Tabela 4: Opisna statistika za centralno klado.
Valid N Mean Minimum Maximum Std.Dev.
m1 128 4,85 4,20 5,98 0,33
m2 124 3,04 0,69 5,06 1,15
m3 124 3,41 0,79 5,35 1,08
m4 127 4,34 3,68 5,49 0,35
m5 128 3,20 2,40 3,79 0,277
Ko 124 1,45 0,00 4,07 1,05
Kr 124 3,41 0,79 5,35 1,08
vrat 123 0,40 0,00 1,60 0,29
molarni indeks 124 70,14 16,25 100,00 21,75
starostni razred 124 1,74 1,00 3,00 0,92
skleninski indeks 124 62,61 14,20 94,82 23,40
koreninski indeks 124 29,86 0,00 83,74 21,75
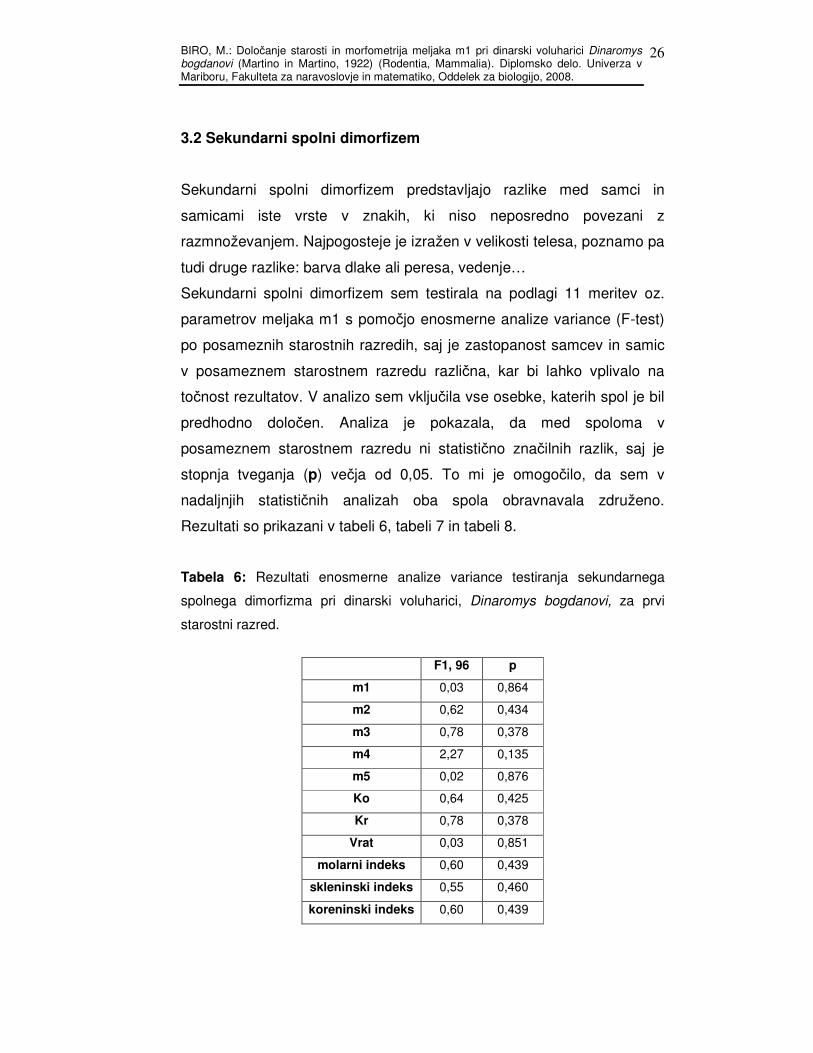
Tabela 5: Opisna statistika za južno klado.
Valid N Mean Minimum Maximum Std.Dev.
m1 35 4,81 4,30 5,38 0,30
m2 34 3,13 1,45 4,72 1,01
m3 34 3,39 1,46 4,88 0,94
m4 35 4,37 3,98 4,82 0,20
m5 34 3,23 2,87 3,67 0,18
Ko 34 1,43 0,00 3,50 1,03
Kr 34 3,39 1,46 4,88 0,94
vrat 34 0,273 0,00 1,02 0,22
molarni indeks 34 70,80 31,00 100,00 20,49
starostni razred 34 1,70 1,00 3,00 0,87
skleninski indeks 34 65,50 30,78 99,35 22,03
koreninski indeks 34 29,35 0,00 69,00 20,29
BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
26
3.2 Sekundarni spolni dimorfizem
Sekundarni spolni dimorfizem predstavljajo razlike med samci in
samicami iste vrste v znakih, ki niso neposredno povezani z
razmnoževanjem. Najpogosteje je izražen v velikosti telesa, poznamo pa
tudi druge razlike: barva dlake ali peresa, vedenje…
Sekundarni spolni dimorfizem sem testirala na podlagi 11 meritev oz.
parametrov meljaka m1 s pomočjo enosmerne analize variance (F-test)
po posameznih starostnih razredih, saj je zastopanost samcev in samic
v posameznem starostnem razredu različna, kar bi lahko vplivalo na
točnost rezultatov. V analizo sem vključila vse osebke, katerih spol je bil
predhodno določen. Analiza je pokazala, da med spoloma v
posameznem starostnem razredu ni statistično značilnih razlik, saj je
stopnja tveganja (p) večja od 0,05. To mi je omogočilo, da sem v
nadaljnjih statističnih analizah oba spola obravnavala združeno.
Rezultati so prikazani v tabeli 6, tabeli 7 in tabeli 8.
Tabela 6: Rezultati enosmerne analize variance testiranja sekundarnega
spolnega dimorfizma pri dinarski voluharici, Dinaromys bogdanovi, za prvi
starostni razred.
F1, 96 p
m1 0,03 0,864
m2 0,62 0,434
m3 0,78 0,378
m4 2,27 0,135
m5 0,02 0,876
Ko 0,64 0,425
Kr 0,78 0,378
Vrat 0,03 0,851
molarni indeks 0,60 0,439
skleninski indeks 0,55 0,460
koreninski indeks 0,60 0,439

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
27
Tabela 7: Rezultati enosmerne analize variance testiranja sekundarnega
spolnega dimorfizma pri dinarski voluharici, Dinaromys bogdanovi, za drugi
starostni razred.
F1, 20 p
m1 2,52 0,128
m2 0,01 0,905
m3 1,01 0,327
m4 2,05 0,167
m5 1,46 0,241
Ko 0,80 0,381
Kr 1,01 0,327
Vrat 0,79 0,383
molarni indeks 0,02 0,877
skleninski indeks 0,70 0,414
koreninski indeks 0,02 0,877
Tabela 8: Rezultati enosmerne analize variance testiranja sekundarnega
spolnega dimorfizma pri dinarski voluharici, Dinaromys bogdanovi, za tretji
starostni razred.
F1, 51 p
m1 0,75 0,389
m2 0,51 0,478
m3 0,61 0,440
m4 0,34 0,559
m5 0,19 0,665
Ko 0,05 0,831
Kr 0,61 0,440
Vrat 0,38 0,539
molarni indeks 0,38 0,538
skleninski indeks 0,33 0,565
koreninski indeks 0,38 0,538

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
28
3.3 Starostna struktura
Na podlagi meritev meljaka m1 sem izrazila višino krone kot delež višine
zoba, kar je dalo molarni indeks (MI). 188 obravnavanih osebkov sem
razvrstila v 3 starostne razrede po koledarskih letih. Razporeditev temelji
na tabeli 12, tabeli 13 in tabeli 14 v delu Kolarič (1999). Glede na razpon
molarnega indeksa lahko ocenimo približno starost osebka.
Tabela 9: Koledarska leta izražena kot delež molarnega indeksa.
Koledarska leta Obdobje Razpon molarnega
indeksa (%)
1. leto rojstvo do prva zima 100–69,8
2. leto prva zima do druga zima 69,7–56,26
3. leto druga zima in naprej 56,25 in manj
V tabeli 10 sem izrazila podatke o velikosti vzorca glede na koledarsko
leto. Rezultati kažejo podatke le za 183 osebkov. To je posledica
manjkajočih oz. nemerljivih parametrov.
Tabela 10: Velikost vzorcev glede na koledarsko leto.
Koledarsko leto N
1 107
2 23
3 53
skupaj 183

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
29
3.4 Starostna in spolna struktura po geografskih regijah
188 živali, zajetih v analizo, je bilo zbranih na 35 lokalitetah (tabela 3), ki
so združene v 3 geografske regije: zahodno (W), centralno (C) in južno
regijo (S). Zanimala me je zastopanost osebkov v posamezni regiji po
spolu in starostnem razredu. Rezultati so prikazani v tabeli 11. kot je
razvidno iz tabele, je skupno število osebkov zajetih v analipri preostalih
15 osebkih je manjkal podatek za spol.
Največ primerkov izvira iz centralne regije – 118. V prvem starostnem
obdobju prevladujejo samci, v drugem in tretjem pa je več samic.
Iz južne klade je zastopanih 31 primerkov. Tudi pri južni kladi v prvem
starostnem obdobju prevladujejo samci, v drugem in tretjem pa samice.
Nekoliko drugače je pri zahodni kladi, kjer je zastopanih 24 osebkov, v
vseh treh starostnih obdobjih pa prevladujejo samice.
Tabela 11: Starostna in spolna struktura populacij.
Geografska regija spol starostni
razred 1
starostni razred
2
starostni razred
3
Skupaj vrsta
S samci 11 1 1 13 S samice 5 5 8 18 Skupaj 16 6 9 31 C samci 43 4 13 60 C samice 24 7 27 58 Skupaj 67 11 40 118 W samci 6 2 0 8 W samice 9 3 4 16 Skupaj 15 5 4 24 Skupaj kolone 98 22 53 173

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
30
3.5 Življenjska preglednica in krivulja preživetja
Na podlagi višine korenine sem ocenila starost osebkov, pomagala sem
si s tabelo 16 v delu Kolarič 1999, in jih nato razdelila v 5 razredov. Na
podlagi starosti osebkov sem nato naredila življenjsko preglednico
(tabela 12), ki je dala vpogled v starostno strukturo populacije glede na
spol. Starost osebkov je ocenjena na podlagi višine korenine. V prvem
razredu so zajeti osebki do starosti 12 mesecev, v drugem osebki od 12
do 24 mesecev, v tretjem od 24 do 36 mesecev, v četrtem starostnem
razredu pa od 36 do 48 mesecev.
V posameznem starostnem razredu je zastopanih več samic kot
samcev. Krivulji na grafu 1 kažeta, da je umrljivost samcev in samic
skozi vso leto bolj ali manj enaka. Najmanjša stopnja umrljivosti je pri
obeh spolih v prvem starostnem obdobju. S starostjo se pri obeh spolih
veča, nekoliko bolj pri samicah. Samice imajo v prvem starostnem
obdobju največjo stopnjo verjetnosti, da bodo dosegle starost dveh let
(ex=1,35), prav tako samci, le da je vrednost nekoliko nižja (ex=0,89).
Analiza razpoložljivega materiala je pokazala, da v materialu ni bilo
prisotne nobene samice, ki bi bila v četrtem starostnem obdobju,
medtem ko je pri samcih en dosegel četrto starostno obdobje.

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
31
Tabela 12: Življenjska preglednica za samice in samce dinarske voluharice
Dinaromys bogdanovi (nx – število živali v razredu; dx – število poginulih
osebkov v starostnem obdobju, računano od začetnih 1000 rojenih osebkov; lx
– število preživelih v začetku starostnega obdobja, računano od 1000 rojenih;
qx – umrljivost na 1000 živih ob začetku starostnega obdobja; Lx – število živih
osebkov, ki je povprečno preživelo v starostnem razredu; ex – pričakovana
življenjska doba glede na doseženo starost (v letih).
samice samci
razred Starost v
mesecih
nx dx lx qx Lx ex nx dx lx qx Lx ex
1 0–12 35 380 1000 380 810 1,35 56 700 1000 700 650 0,89
2 12–24 36 392 620 632 424 0,87 18 225 300 750 188 0,79
3 24–36 21 228 228 1000 114 0,50 5 62,5 75 833 43,75 0,67
4 36–48 0 0 0 0 0 0 1 12,5 12,5 1000 6,25 0,50
5 > 48 0 0 0 0 0 0 0 0 0 0 0 0
∑ 92 1000 1348 80 1000 888

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
32
-
Graf 1: Krivulja preživetja za samce in samice dinarske voluharice.
0
100
200
300
400
500
600
700
800
900
1 2 3 4 5
starostni razred
št. ži
vih o
sebko
v
samice samci

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
33
4 DISKUSIJA
Raziskav in objav o dinarski voluharici, Dinaromys bogdanovi, je do
danes zelo malo, saj so jo do leta 1922 poznali le v fosilni obliki. Od
odkritja živih primerkov do danes so različni avtorji (Gill in sod., 1982)
večinoma obravnavali taksonomsko problematiko, ki pa še vedno ni
dorečena. Todorović (1956) dopušča možnost, da obstajata znotraj vrste
Dinaromys bogdanovi dve glavni starostni liniji, torej dve dobro ločljivi
podvrsti. Teza je osnovana na morfologiji prvega meljaka in kasnejši
študiji prekrivanja dveh vrst. Potrdile so jo tudi elektroforetske raziskave
frekvenc alelov na 28 lokusih pri poddružinah Muridae in Microtus, ki so
pokazale visok nivo genetske delitve dveh morfotipov, ter da je rod
Dinaromys sestrski takson Clethrionomys – klade Microtus (Gill in sod.,
1986). Novejše raziskave (Kryštufek, 2007) so bile narejene na osnovi
genetske strukture dinarske voluharice, in sicer z uporabo 555 bp
sekvence mitohondrijskega gena citokroma b. Študija temelji na 63
primerkih iz 18 različnih lokacij celotnega območja, ki ga vrsta poseljuje.
Med 47 halotipi sta bili prepoznani 2 kladi, podprti s 6, 64 % sekvenčno
divergenco. Prva klada, ti. zahodna klada obsega vzorce 4 geografskih
območij jugovzhodno od reke Neretve na Hrvaškem ter zahodni Bosni in
Hercegovini, druga (večja) pa vse ostale vzorce. Večja klada se nadalje
loči v dve podskupini, podprti z 2, 9 % sekvenčno divergenco. Kladi
ločuje reka Drim. Prva zajema preostale vzorce iz Bosne in
Hercegovine, Črne gore in Kosova - centralna klada, druga, ti. južna
klada, pa zajema vzorce iz Makedonije. Med slednjima filoskupinama ni
bilo deljenih halotipov. Pomembna razlika je razlika v nukleotidih, ki je
bila največja v zahodni kladi (π = 0,016), najmanjša pa v južni kladi (π =
0,0062). Na podlagi pridobljenih podatkov je bila postavljena hipoteza,
da se je pred približno 1 milijonom let zahodna klada začela širiti južneje,
preko reke Neretve, tako bi naj kasneje z alopatrijo nastala centralna
klada. Pred približno 0,3 milijoni let bi se naj začela širiti južneje tudi

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
34
centralna klada, preko reke Drim, ter se postopoma ločevala v sedanjo
južno klado, prav tako z alopatrijo. Omenjeno je podprto tudi s
paleontološkimi dokazi, saj so ostanki dinarske voluharice iz zgodnjega
pleistocena najdeni le na tem območju. Medtem pa so iz območja južne
regije znani le fosilni ostanki iz kasnejšega časa (Kryštufek in sod.,
2007).
Na osnovi omenjenih ugotovitev sta me zanimali spolna in starostna
struktura po posameznih starostnih razredih in geografskih regijah
(kladah). Ker ostajajo očitne razlike med tremi, zgoraj omenjenimi
kladami, sem opravila analize ločeno za posamezno klado.
Kot je znano, je za glodavce diagnostično zobovje, ki se tekom življenja
spreminja. Pri nekaterih stalno raste, pri drugih pa mlečne zobe
zamenjajo stalni. Posebnost zob dinarske voluharice je, da tekom
postnatalnega razvoja razvijejo korenino. Aboslutna višina zoba ostaja
bolj ali manj enaka, saj se zob zaradi načina prehranjevanja (rastline)
obrablja, višina korenine zoba pa se s starostjo veča. Na osnovi
omenjenih dejstev in ugotovitev sem postavila hipotezo, da lahko na
podlagi morfometričnih lastnosti meljaka dinarske voluharice določim
starost osebka za posamezno klado na podlagi višine korenine zoba.
Meritve sem opravljala na prvem spodnjem meljaku (m1).
Na podlagi opravljenih meritev sem najprej ugotavljala prisotnost
sekundarnega spolnega dimorfizma za posamezni starostni razred.
Rezultati analize so pokazali, da v velikosti posameznih parametrov med
samci in samicami ni statistično značilnih razlik. Stopnja tveganja je pri
vseh višja od 0,05. Podobno ugotavlja tudi Kolarič (1999). Druge
raziskave (Rumež, 2006; Klemen, 2007), in sicer na področju
morfometrične variabilnosti pri dinarski voluharici prav tako ne kažejo
razlik med spoloma v nereprodukcijskih znakih.
Za ocenjevanje starosti sem uporabila višino korenine, ne pa molarnega
indeksa, saj vsebuje slednji nekatere neželene lastnosti. Sestavljata ga

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
35
dva parametra: višina krone in višina zoba, katerih velikost ni odvisna
samo od fizioloških dejavnikov, ampak tudi od zunanjih, saj vpliva tip
hrane na obrabo žvekalne površine. Tako sem želela z izražanjem višine
krone kot deležem višine celotnega zoba odstraniti vpliv višine zoba.
Kadar imamo opravka z alometričnimi odnosi med spremenljivkami, ne
moremo učinkovito odstraniti vpliva velikosti z enostavnim parametrom
(indeksom) (Packard in Boardman, 1987), torej je takšen pristop lahko
zavajajoč. Iz tega razloga sem za ocenjevanje starosti raje uporabila
absolutno višino korenine. Rezultati analize so pokazali, da pri centralni
in južni kladi v prvem starostnem razredu prevladujejo samci, v drugem
in tretjem starostnem obdobju pa samice. Pri zahodni kladi prevladujejo
samice v vseh treh starostnih obdobjih.
Umrljivost samcev in samic skozi vso leto bolj ali manj enaka Večje
razlike so v prvem letu starosti, v obdobju do 12 mesecev, ko imajo
samice večjo sposobnost preživetja. V drugem obdobju (od 12 do 24
mesecev) med vrednostma koeficienta umrljivosti (pri samicah: qx=620,
pri samcih: qx= 750) ni tako očitne razlike kot v prvem starostnem
razredu. Od 24 meseca naprej pa imajo večjo sposobnost preživetja
samci. Število živih osebkov, ki je povprečno preživelo v posameznem
starostnem razredu, je večje pri samicah kot pri samcih, so pa dosegli
samci višje starosti kot samice. Podobne rezultate je dobil tudi Kolarič
(1999).

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
36
5 POVZETEK
Namen moje naloge je bila določiti prisotnosti sekundarnega spolnega
dimorfizma in predstaviti starostno strukturo na podlagi velikosti korenin
m1 pri dinarski voluharici, Dinaromys bogdanovi. Meljaki dinarske
voluharice razvijejo tekom postnatalnega razvoja korenino, kar omogoča
uporabo dolžine korenine kot indikator absolutne starosti. Analiza je
temeljila na 188 vzorcih, vendar so bili pri nekaterih osebkih podatki
pomanjkljivi. Uporabljen material bil zbran na 36 različnih geografskih
lokacijah in razdeljen v tri klade (zahodno, centralno in južno klado). Na
podlagi meritev meljaka m1 sem izrazila višino krone kot delež višine
zoba, kar je dalo molarni indeks (MI). Dobljene rezultate sem razvrstila v
tri starostne razrede po koledarskih letih. Glede na razpon molarnega
indeksa lahko določimo približno starost osebka. Rezultati analize za
spolni dimorfizem so pokazali, da pri dinarski voluharici sekundarni
spolni dimorfizem ni prisoten. Analiza starostne in spolne strukture po
geografskih regijah pa je pokazala, da pri centralni in južni kladi v prvem
starostnem razredu prevladujejo samci, v drugem in tretjem starostnem
obdobju pa samice, medtem ko pri zahodni kladi v vseh treh starostnih
obdobjih prevladujejo samice. Pri samicah je tudi večja stopnja
verjetnosti, da bodo dosegle starost dveh let. Sicer pa je umrljivost pri
obeh spolih skozi vse življenje enakomerna.

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
37
6 ZAHVALA
Zahvaliti se želim vsem, ki so kakor koli prispevali k mojemu uspešno
zaključenemu študiju.
Prva zahvala velja mojemu mentorju, izr. prof. dr. Francu Janžekoviču,
za odlično mentorstvo, pomoč in koristne nasvete pri izdelavi diplomske
naloge.
Prav tako se zahvaljujem somentorju red. prof. dr. Borisu Kryštufku za
posojo materiala in idejno vodstvo.
Zahvaljujem se tudi prim. Jožefu Mateli, dr. med., predstojniku
radiološkega oddelka Univerzitetnega kliničnega centra Maribor, ki mi je
omogočil slikanje materiala ter ge. Ireni Hercog, dipl. inž. radiologije, ki
mi je pomagala pri rentgenskem slikanju materiala.
Posebna zahvala gre mojim staršem in bratu, ker so verjeli vame in me
razumeli, mi ves čas študija stali ob strani in s svojo dobro voljo ter
moralno podporo doprinesli k nastanku tega dela.
Vsem najlepša hvala

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
38
7 LITERATURA
Bolkay, St. J. (1924). Catalogue of the Mammals occuring in Bosnia-
Hercegovina and Preserved in the Land-Museum Sarajevo. Biol.
Hung., I, str. 4-5.
Bratina, T. (2003). Primer uporabe SPSS. Maribor: Pedagoška fakulteta.
Bratina, T. (2006). Slovarček statističnih metod s primeri. Maribor:
Pedagoška fakulteta.
Cardini, A., Tongiorgi, P. (2003). Yellow-bellied marmots (Marmota
flaviventris) »in the shape space«(Rodentia, Scuridea): sexual
dimorphism, growth and allometry of the mandible. Zoomorphology
122: 11-23.
Cardini, A., O'Higgins, P. (2005). Post-natal onogeny of the mandible
and ventral cranium in Marmota species (Rodentia, Scuridae):
allometry and phylogeny. Zoomorphology 124: 189-203.
Đulić, B., Savić, I., Soldatović, B. (1971). The chromosomes of two rodent
species Dolomys bogdanovi (V. and E. Martino 1922) and Glis glis
(Linnaeus 1766) (Mammalia, Rodentia). Caryologia, 24, str.299-305.
Gill, A., Petrov, B., Živković, S., Rimsa, D. 1986. Biochemical
comparisons in Yugoslavian rodents of the families Arvicolidae in
Muridae. Z. f. Säugetierkunde, 52, 4, s. 247-256.
Gligić, M. O. 1059. Prilog poznavanju roda Dolomys Nehring na
centralnim planinama NR Bosne i Hercegovine. Glasnik
prirodnjačkog muzeja. Beograd, Serija B, Knjiga 14, s. 265-297.

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
39
http://en.wikipedia.org/wiki/Image:Panair_Korab.jpg
Klemen, M. (2007). Variabilnost zgornjih meljakov dinarske voluharice,
Dinaromys bogdanovi (Martino in Martino, 1922). Diplomsko delo,
Maribor: Fakulteta za naravoslovje in matematiko, Oddelek za
biologijo.
Kolarič, K. (1999). Določanje starosti in starostna struktura pri dinarski
voluharici Dinaromys bogdanovi (Rodentia, Mammalia). Diplomsko
delo, Ljubljana: Biotehniška fakulteta, Oddelek za biologijo.
Kryštufek, B. (1991). Sesalci Slovenije. Prirodoslovni muzej Slovenije.
Ljubljana. Str. 131-132.
Kryštufek, B. (1987). Taksonomska diferenciacija, zoogeografija in
ekologija voluharic (Arvicolidae, Rodentia, Mammalia) v Sloveniji.
Doktorska disertacija, Ljubljana.
Kryštufek, B., Kolarič, K. in Paunović, M. (2000). Age determinatio and
age structure in Martino's vole Dinaromys bogdanovi. Mammalia 64,
str. 361-370.
Kryštufek, B., Bužan, E., Hutchinson, W. F., in Hanfling, B. (2007).
Phylogeography of the rare Balkan endemic Martino's vole,
Dinaromys bogdanovi, reveals strong differentiation within the
westwrn Balkan Peninsula. Mol. Ecol., 2007, vol. 16, iss. 6, str. 1221-
1232.
Martino, J. (1948). Prilog poznavanju biologije recentnih voluharica rodu
Dolomys. Godišnjak Biol. Instituta (Sarajevo), 1, str. 83-88.

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
40
Martino, J. (1948). Prilog poznavanju recentnih voluharica rodu
Dolomys. Godišnjak Biol. Instituta (Sarajevo), 1, str. 71-78.
Packard, G. C., Boardman, T. J., (1987). The misuse of ratios to scale
physiological data that vary allometrically with body size. New
directions in ecological physiology. Cambridge, Cambridge University
Press, s. 216-239.
Petrov, B., Todorović, M. (1982). Dinaromys bogdanovi (V. et E. Martino,
1922) – Bergmaus. Handbuch der Säugetiere Europas, 2, I.
Akademische Verlagsgesellschaft, Wiesbaden, str. 193-208.
Rohlf, J. (2006) tpsDig, Version 2.10. SUNY at Stony Brook.
Rumež, S. (2006). Geografska variabilnost dimenzij zgornjig meljakov pri
dinarski voluharici Dinaromys bogdanovi. Diplomsko delo, Maribor:
Pedagoška fakulteta, Oddelek za biologijo.

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
41
8 PRILOGE
Priloga 1
Tabela 13: Osnovi podatki o dinarski voluharici in meritve.
zap. št. inv. št. spol
starost (v
letih)
geografska regija država lokaliteta m1 m2 m3 m4 m5 korenina krona vrat
molarni indeks
starostni razred
skleninski indeks
koreninski indeks
starost v mesecih (ocenjena starost)
1 PMS162 2 S Makedonija Korab, Kobilino polje, Crna Čuka 5,02 1,73 2,16 4,19 3,38 2,860 2,16 0,43 43,03 3 34,5 57,0 25,88
2 PMS163 2 S Makedonija Korab, Kobilino polje, Crna Čuka 4,79 2,40 2,58 4,10 3,41 2,210 2,58 0,18 53,86 3 50,1 46,1 19,44
3 PMS191 1 S Makedonija Jablanica, Podgorci, Podgoriško jezero 4,96 3,76 3,93 4,36 3,15 1,030 3,93 0,17 79,23 1 75,8 20,8 10,85
4 PMS192 1 S Makedonija Jablanica, Podgorci, Podgoriško jezero 5,15 4,26 4,24 4,71 3,16 0,910 4,24 0,00 82,33 1 82,7 17,7 8,7
5 PMS193 2 S Makedonija Jablanica, Podgorci, Podgoriško jezero 5,38 1,75 1,88 4,43 3,21 3,500 1,88 0,13 34,94 3 32,5 65,1 32,32
6 PMS197 1 S Makedonija Bistra 4,71 1,45 1,46 4,26 3,56 3,250 1,46 0,01 31,00 3 30,8 69,0 30,17
7 PMS198 1 1 S Makedonija Bistra 4,79 3,68 3,85 4,36 3,17 0,940 3,85 0,17 80,38 1 76,8 19,6 8,7
8 PMS200 1 S Makedonija Stogovo, Ajdar Ćesmana 4,46 2,99 3,13 4,23 3,25 1,330 3,13 0,14 70,18 1 67,0 29,8 13
9 PMS202 2 S Makedonija Stogovo, Ajdar Ćesmana 5,18 2,20 2,35 4,41 3,37 2,830 2,35 0,15 45,37 3 42,5 54,6 25,88
10 PMS203 2 S Makedonija Stogovo, Ajdar Ćesmana 4,73 3,67 3,72 4,38 3,18 1,010 3,72 0,05 78,65 1 77,6 21,4 10,85
11 ZMS234 C BIH Bjelašnica 5,34 2,84 3,37 4,36 2,83 1,970 3,37 0,53 63,11 2 53,2 36,9 17,29
12 ZMS235 C BIH Bjelašnica 4,73 2,65 3,43 3,97 3,15 1,300 3,43 0,78 72,52 1 56,0 27,5 13
13 ZMS236 1 C BIH Bjelašnica 4,40 3,47 3,80 4,20 3,06 0,600 3,80 0,33 86,36 1 78,9 13,6 6,56
14 ZMS237 1 C BIH Bjelašnica 4,60 3,69 3,98 4,47 2,76 0,620 3,98 0,29 86,52 1 80,2 13,5 6,56
15 ZMS258 1 2 C BIH Trebević 4,52 2,11 2,45 3,88 3,21 2,070 2,45 0,34 54,20 3 46,7 45,8 19,44
16 ZMS260 1 4 C BIH Trebević 4,22 3,72 3,10 88,2
17 ZMS261 2 1 C BIH Trebević 4,44 4,21 4,34 4,30 2,79 0,100 4,34 0,13 97,75 1 94,8 2,3 2,26
18 ZMS262 1 2 C BIH Trebević 4,79 1,87 2,41 4,17 3,48 2,380 2,41 0,54 50,31 3 39,0 49,7 21,59

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
42
19 ZMS263 2 C BIH Trebević 4,57 2,19 2,72 3,68 3,13 1,850 2,72 0,53 59,52 2 47,9 40,5 17,29
20 ZMS265 2 C BIH Trebević 4,40 1,56 1,90 3,86 3,21 2,500 1,90 0,34 43,18 3 35,5 56,8 23,73
21 ZMS266 2 2 C BIH Trebević 4,23 1,12 2,00 3,76 3,05 2,230 2,00 0,88 47,28 3 26,5 52,7 19,44
22 ZMS267 1 1 C BIH Trebević 4,54 4,10 4,27 4,11 3,08 0,270 4,27 0,17 94,05 1 90,3 5,9 4,41
23 ZMS268 1 1 C BIH Trebević 4,40 3,77 4,33 4,24 2,81 0,070 4,33 0,56 98,41 1 85,7 1,6 2,26
24 ZMS269 1 1 C BIH Trebević 4,46 3,94 3,93 4,11 2,87 0,530 3,93 0,00 88,12 1 88,3 11,9 6,56
25 ZMS358 2 1 C BIH Lebršnik 5,04 3,95 4,27 4,71 3,60 0,770 4,27 0,32 84,72 1 78,4 15,3 8,7
26 ZMS359 1 1 C BIH Lebršnik 5,36 3,00 4,26 4,68 3,59 1,100 4,26 1,26 79,48 1 56,0 20,5 10,85
27 ZMS361 C BIH Lebršnik 5,73 4,38 4,97 4,80 3,68 0,760 4,97 0,59 86,74 1 76,4 13,3 6,56
28 PMS486 1 1 C Črna gora Hotel Jelovica,
Jelovica, Lubnice, Bjelasica
4,92 4,38 4,53 4,69 3,06 0,390 4,53 0,15 92,07 1 89,0 7,9 4,41
29 PMS487 2 2 C Črna gora Zekova glava, Bjelasica 5,29 2,73 3,32 4,81 3,45 1,970 3,32 0,59 62,76 2 51,6 37,2 17,29
30 PMS488 2 3 C Črna gora Zekova glava, Bjelasica 5,43 1,92 2,35 4,68 2,95 3,080 2,35 0,43 43,28 3 35,4 56,7 28,03
31 PMS489 1 4 C Črna gora Zekova glava, Bjelasica 4,74 0,84 1,22 4,35 2,95 3,520 1,22 0,38 25,74 3 17,7 74,3 32,32
32 PMS491 2 3 C Črna gora Hotel Jelovica,
Jelovica, Lubnice, Bjelasica
4,97 1,73 2,64 4,04 3,10 2,330 2,64 0,91 53,12 3 34,8 46,9 21,59
33 PMS492 2 2 C Črna gora Hotel Jelovica,
Jelovica, Lubnice, Bjelasica
5,00 2,20 2,70 4,67 3,18 2,300 2,70 0,50 54,00 3 44,0 46,0 21,59
34 PMS493 1 1 C Črna gora Hotel Jelovica,
Jelovica, Lubnice, Bjelasica
4,98 3,91 4,40 4,62 3,27 0,580 4,40 0,49 88,35 1 78,5 11,6 6,56
35 PMS494 1 1 C Črna gora Hotel Jelovica,
Jelovica, Lubnice, Bjelasica
4,73 4,19 4,60 4,66 3,29 0,130 4,60 0,41 97,25 1 88,6 2,7 2,26
36 PMS990 2 1 S Makedonija Galičica 4,68 4,34 4,30 4,55 3,03 0,380 4,30 0,00 91,88 1 92,7 8,1 4,41
37 PMS991 2 S Makedonija Galičica 4,54 2,25 2,82 4,13 3,44 1,720 2,82 0,57 62,11 2 49,6 37,9 15,14
38 PMS992 2 S Makedonija pl. Bistra 4,75 1,72 2,74 4,27 3,15 2,010 2,74 1,02 57,68 2 36,2 42,3 19,44
39 PMS998 2 2 C Kosovo Lučanski stanovi, Žljeb 5,30 2,57 3,08 4,45 3,77 2,220 3,08 0,51 58,11 2 48,5 41,9 19,44
40 PMS999 1 1 C Kosovo Lučanski stanovi, Žljeb 5,60 4,91 4,98 5,14 3,11 0,620 4,98 0,07 88,93 1 87,7 11,1 6,56
BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
43
41 PMS1000 1 1 C Kosovo Lučanski stanovi, Žljeb 5,73 4,79 5,15 5,49 3,17 0,580 5,15 0,36 89,88 1 83,6 10,1 6,56
42 PMS1001 2 1 C Kosovo Lučanski stanovi, Žljeb 5,58 5,06 5,35 5,49 2,91 0,230 5,35 0,29 95,88 1 90,7 4,1 2,26
43 PMF1003 2 3 W BIH Dinara 4,37 0,94 1,04 3,71 2,60 3,330 1,04 0,10 23,80 3 21,5 76,2 30,17
44 PMS1006 2 1 W Kosovo Lučanski stanovi, Žljeb 5,49 4,54 4,98 5,39 2,98 0,510 4,98 0,44 90,71 1 82,7 9,3 6,56
45 PMS1007 1 W Kosovo Lučanski stanovi, Žljeb 5,73 4,79 5,24 5,50 3,11 0,490 5,24 0,45 91,45 1 83,6 8,6 4,41
46 PMF1008 1 1 W BIH Dinara (Dolac) 4,91 3,95 4,22 4,55 2,83 0,690 4,22 0,27 85,95 1 80,4 14,1 6,56
47 PMS1019 2 1 C Črna gora Kom Vasojevića 4,60 3,85 4,28 4,28 3,15 0,320 4,28 0,43 93,04 1 83,7 7,0 4,41
48 PMF1032 1 1 C BIH Čatrna voda (Dinara) 4,43 3,65 3,84 4,13 2,89 0,590 3,84 0,19 86,68 1 82,4 13,3 6,56
49 PMS2005/612 2 C BIH Zelengora 4,56 1,51 2,08 4,20 3,14 2,480 2,08 0,57 45,61 3 33,1 54,4 21,59
50 PMS2005/613 1 1 C BIH Zelengora 4,73 3,61 3,94 4,08 3,05 0,790 3,94 0,33 83,30 1 76,3 16,7 8,7
51 PMA2005/614 2 1 C BIH Zelengora 4,54 3,98 4,29 4,33 3,14 0,250 4,29 0,31 94,49 1 87,7 5,5 4,41
52 PMS2005/615 2 1 C BIH Zelengora 4,76 4,11 4,38 4,43 2,81 0,380 4,38 0,27 92,02 1 86,3 8,0 4,41
53 PMS2005/617 1 C BIH Zelengora 4,62 1,97 2,05 3,77 3,61 2,570 2,05 0,08 44,37 3 42,6 55,6 23,73
54 PMS2005/618 2 1 C BIH Zelengora 4,51 3,81 4,13 3,93 3,00 0,380 4,13 0,32 91,57 1 84,5 8,4 4,41
55 PMS2005/619 1 1 C BIH Zelengora 4,46 3,99 4,33 4,38 3,03 0,130 4,33 0,34 97,09 1 89,5 2,9 2,26
56 PMS2005/621 2 1 C BIH Zelengora 4,88 3,76 4,02 4,44 3,05 0,860 4,02 0,26 82,38 1 77,0 17,6 8,7
57 PMS2005/622 1 1 C BIH Zelengora 4,96 3,85 3,94 4,27 2,89 1,020 3,94 0,09 79,44 1 77,6 20,6 10,85
58 PMS2005/636 1 C BIH Sarajevo, Bjelašnica 4,87 1,62 2,15 4,02 3,33 2,720 2,15 0,53 44,15 3 33,3 55,9 23,73
59 PMS2005/637 1 1 C BIH Sarajevo, Bjelašnica 4,70 4,02 4,19 4,10 3,14 0,510 4,19 0,17 89,15 1 85,5 10,9 6,56
60 PMS2005/638 1 1 C BIH Sarajevo, Bjelašnica 4,70 4,45 4,54 4,21 2,95 0,160 4,54 0,09 96,60 1 94,7 3,4 2,26
61 PMS2005/639 2 1 C BIH Sarajevo, Bjelašnica 4,85 4,11 4,27 4,28 2,89 0,580 4,27 0,16 88,04 1 84,7 12,0 6,56
62 PMS2005/640 2 1 C BIH Sarajevo, Bjelašnica 4,71 4,02 4,53 4,19 2,99 0,180 4,53 0,51 96,18 1 85,4 3,8 2,26
63 PMS2005/641 2 C BIH Sarajevo, Bjelašnica 5,20 0,96 1,37 4,30 3,37 3,830 1,37 0,41 26,35 3 18,5 73,7 34,47
64 PMS2005/642 1 1 C BIH Sarajevo, Bjelašnica 5,30 4,45 4,70 4,88 3,19 0,600 4,70 0,25 88,68 1 84,0 11,3 6,56
65 PMS2006/834 1 W Hrvaška Mt. Opor, Malačka 4,62 3,25 3,16 3,93 2,78 1,460 3,16 0,00 68,40 2 70,3 31,6 13
66 PMS2006/835 1 W Hrvaška Mt. Opor, Malačka 4,33 3,68 3,68 4,12 2,81 0,650 3,68 0,00 84,99 1 85,0 15,0 6,56
67 PMS2442 1 1 S Makedonija pl. Bistra 4,78 4,34 4,81 4,54 3,25 0,000 4,81 0,47 100,00 1 90,8 0,0 0
68 PMS2443 2 S Makedonija pl. Bistra 4,30 1,53 2,08 3,98 2,87 2,220 2,08 0,55 48,37 3 35,6 51,6 19,44

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
44
69 PMS2444 1 1 S Makedonija pl. Bistra 4,74 4,13 4,62 4,53 3,16 0,120 4,62 0,49 97,47 1 87,1 2,5 2,26
70 PMS2501 1 2 W Hrvaška Split, Malačka 4,81 2,59 2,83 4,37 2,87 1,980 2,83 0,24 58,84 2 53,8 41,2 17,29
71 PMS2502 2 2 W Hrvaška Split, Malačka 4,71 2,91 3,42 4,10 2,92 1,290 3,42 0,51 72,61 1 61,8 27,4 13
72 PMS2503 1 1 W Hrvaška Split, Malačka 4,34 3,63 3,78 3,97 2,76 0,560 3,78 0,15 87,10 1 83,6 12,9 6,56
73 PMS2504 2 3 W Hrvaška Split, Malačka 4,47 1,55 1,63 3,70 3,10 2,840 1,63 0,08 36,47 3 34,7 63,5 25,88
74 PMS2505 1 1 W Hrvaška Split, Malačka 4,44 3,94 4,03 4,37 2,68 0,410 4,03 0,09 90,77 1 88,7 9,2 4,41
75 PMS2577 2 2 W BIH Cincar, Ćejića ravne 4,88 2,31 2,84 4,45 3,35 2,040 2,84 0,53 58,20 2 47,3 41,8 19,44
76 PMS2578 2 2 W BIH Cincar, Ćejića ravne 5,11 2,10 2,59 4,36 3,45 2,520 2,59 0,49 50,68 3 41,1 49,3 23,73
77 PMS2579 2 1 W BIH Cincar, Ćejića ravne 4,82 3,95 4,15 4,44 3,06 0,670 4,15 0,20 86,10 1 82,0 13,9 6,56
78 PMS2580 2 1 W BIH Cincar, Ćejića ravne 4,98 4,03 4,28 4,45 3,27 0,700 4,28 0,25 85,94 1 80,9 14,1 6,56
79 PMS2581 2 1 W BIH Cincar, Ćejića ravne 4,82 4,05 4,47 4,45 3,00 0,350 4,47 0,42 92,74 1 84,0 7,3 4,41
80 PMS2582 2 1 W BIH Cincar, Ćejića ravne 4,48 4,04 4,53 4,36 2,80 0,000 4,53 0,49 100,00 1 90,2 0,0 0
81 PMS2583 2 1 W BIH Cincar, Ćejića ravne 4,60 4,22 4,33 4,54 3,69 0,270 4,33 0,11 94,13 1 91,7 5,9 4,41
82 PMS2584 2 1 W BIH Cincar, Ćejića ravne 4,70 4,48 4,56 4,78 2,73 0,140 4,56 0,08 97,02 1 95,3 3,0 2,26
83 PMS2585 1 1 W BIH Cincar, Ćejića ravne 4,56 3,91 4,23 4,39 2,95 0,330 4,23 0,32 92,76 1 85,7 7,2 4,41
84 PMS2591 2 2 W BIH Cincar, Ćejića ravne 5,35 2,37 2,83 4,97 2,89 2,520 2,83 0,46 52,90 3 44,3 47,1 23,73
85 PMS2882 1 C Črna gora Vilusi, Obodina 4,79 3,84 3,92 4,30 3,02 0,870 3,92 0,08 81,84 1 80,2 18,2 8,7
86 PMS3422 2 C Črna gora Durmitor, Žabljak, Crepuljna poljana 4,70 1,97 2,35 4,33 3,70 2,350 2,35 0,38 50,00 3 41,9 50,0 21,59
87 PMS3423 2 C Črna gora Durmitor, Žabljak, Crepuljna poljana 4,51 1,21 1,64 3,77 3,11 2,870 1,64 0,43 36,36 3 26,8 63,6 25,88
88 PMS3424 2 C Črna gora Durmitor, Žabljak, Crepuljna poljana
4,92 1,38 4,28 3,03 3,540 1,38 28,05 3 72,0 32,32
89 PMS3425 1 C Črna gora Durmitor, Žabljak, Crepuljna poljana 4,86 0,69 0,79 4,62 2,95 4,070 0,79 0,10 16,26 3 14,2 83,7 36,63
90 PMS3426 1 C Črna gora Durmitor, Žabljak, Crepuljna poljana 5,05 2,15 2,93 4,11 3,49 2,120 2,93 0,78 58,02 2 42,6 42,0 19,44
91 PMS3707 2 2 W BIH Šator, Šatorsko jezero 4,35 2,27 2,55 3,84 2,75 1,800 2,55 0,28 58,62 2 52,2 41,4 17,29
92 PMS3733 1 1 W BIH Šator, Šatorsko jezero 4,97 4,29 4,38 4,54 3,16 0,590 4,38 0,09 88,13 1 86,3 11,9 6,56
93 PMS4214 1 1 C BIH Zelengora, Orlovačko
jezero 5,04 4,02 4,02 4,55 3,22 1,020 4,02 0,00 79,76 1 79,8 20,2
94 PMS4215 1 1 C BIH Zelengora, Orlovačko jezero 4,85 3,37 4,31 4,23 2,88 0,540 4,31 0,94 88,87 1 69,5 11,1 6,56

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
45
95 PMS4229 2 1 C BIH Zelengora, pod Orlovcem 4,97 3,97 4,44 4,63 2,95 0,530 4,44 0,47 89,34 1 79,9 10,7 6,56
96 PMS4230 1 3 C BIH Zelengora, pod Orlovcem 4,71 1,46 1,54 3,93 3,08 3,170 1,54 0,08 32,70 3 31,0 67,3 28,03
97 PMS4231 2 4 C BIH Zelengora, pod Orlovcem 4,74 3,95 2,56
98 PMS4232 1 2 C BIH Zelengora, pod
Orlovcem 4,75 1,61 2,33 4,55 3,37 2,420 2,33 0,72 49,05 3 33,9 50,9 21,59
99 PMS4233 1 1 C BIH Zelengora, pod Orlovcem 4,90 4,51 4,70 4,38 3,79 0,200 4,70 0,19 95,92 1 92,0 4,1 2,26
100 PMS4234 1 1 C BIH Zelengora, pod
Orlovcem 4,87 3,93 4,03 4,30 2,87 0,840 4,03 0,10 82,75 1 80,7 17,2 8,7
101 PMS5128 2 2 C BIH Prenj, Poljana 5,10 1,38 1,73 4,37 3,51 3,370 1,73 0,35 33,92 3 27,1 66,1 30,17
102 PMS5129 1 1 C BIH Prenj, Poljana 4,61 4,15 4,24 4,30 2,73 0,370 4,24 0,09 91,97 1 90,0 8,0 4,41
103 PMS5130 1 1 C BIH Prenj, Poljana 4,20 2,75 3,86 3,78 2,40 0,340 3,86 1,11 91,90 1 65,5 8,1 4,41
104 PMS5239 2 1 W BIH Paklenica, Velebit 5,04 4,36 4,62 4,96 3,12 0,420 4,62 0,26 91,67 1 86,5 8,3 4,41
105 PMS5680 2 1 C BIH Mostar, Podveležje 4,76 4,06 4,30 4,53 2,78 0,460 4,30 0,24 90,34 1 85,3 9,7 4,41
106 PMS6473 2 2 C BIH Zelengora, pod Orlovcem 5,38 2,57 3,68 4,56 3,46 1,700 3,68 1,11 68,40 2 47,8 31,6 15,14
107 PMS6474 1 2 C BIH Zelengora, pod Orlovcem 4,56 2,08 2,49 3,89 3,35 2,070 2,49 0,41 54,61 3 45,6 45,4 19,44
108 PMS6475 2 2 C BIH Zelengora, pod Orlovcem 5,23 2,32 2,87 4,54 3,70 2,360 2,87 0,55 54,88 3 44,4 45,1 21,59
109 PMS6476 2 2 C BIH Zelengora, pod Orlovcem 5,37 1,81 2,41 4,66 3,67 2,960 2,41 0,60 44,88 3 33,7 55,1 25,88
110 PMS6477 2 3 C BIH Zelengora, pod Orlovcem 4,56 0,92 1,45 3,79 3,15 3,110 1,45 0,53 31,80 3 20,2 68,2 28,03
111 PMS7539 1 2 C BIH Čemerno 5,14 3,28 3,71 4,24 3,20 1,430 3,71 0,43 72,18 1 63,8 27,8 13
112 PMS7542 1 1 C BIH Zelengora, pod
Orlovcem 4,87 4,31 4,59 4,59 3,22 0,280 4,59 0,28 94,25 1 88,5 5,7 4,41
113 PMS7543 2 1 C BIH Zelengora, pod Orlovcem
4,96 3,42 3,76 4,79 3,14 1,200 3,76 0,34 75,81 1 69,0 24,2 10,85
114 PMS7544 1 1 C BIH Zelengora, pod Orlovcem 5,05 4,20 4,44 4,54 2,94 0,610 4,44 0,24 87,92 1 83,2 12,1 6,56
115 PMS7545 1 1 C BIH Zelengora, pod Orlovcem 5,06 4,08 4,41 4,79 3,13 0,650 4,41 0,33 87,15 1 80,6 12,8 6,56
116 PMS7556 1 1 C BIH Čemerno 4,53 4,02 3,95 4,23 2,94 0,580 3,95 0,00 87,20 1 88,7 12,8 6,56
117 PMS7557 2 2 C BIH Čemerno 4,74 3,64 3,77 4,11 3,22 0,970 3,77 0,13 79,54 1 76,8 20,5 8,7
118 PMS7558 2 2 C BIH Zelengora, pod
Orlovcem 5,16 3,33 3,60 4,66 3,23 1,560 3,60 0,27 69,77 2 64,5 30,2 15,14

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
46
119 PMS7559 2 1 C BIH Zelengora, pod Orlovcem 4,84 4,37 5,05 4,71 3,11 0,000 5,05 0,68 100,00 1 90,3 0,0 0
120 PMS7608 2 2 C BIH Ljubinje 4,75 1,28 1,55 4,27 3,08 3,200 1,55 0,27 32,63 3 26,9 67,4 28,03
121 PMS7609 2 1 C BIH Ljubinje 4,39 2,24 3,10 3,95 2,95 1,290 3,10 0,86 70,62 1 51,0 29,4 13
122 PMS7610 1 2 C BIH Ljubinje 4,51 2,06 2,58 3,85 3,19 1,930 2,58 0,52 57,21 2 45,7 42,8 17,29
123 PMS7806 2 2 C Črna gora Lovčen, Jezero
Lovčen 4,98 1,73 3,33 4,44 3,41 1,650 3,33 1,60 66,87 2 34,7 33,1 15,14
124 PMS7827 2 2 C Črna gora Lovčen, Jezerski vrh, Čista strana 4,66 2,20 2,51 4,42 3,02 2,150 2,51 0,31 53,86 3 47,2 46,1 19,44
125 PMS7834 2 2 C Črna gora Sutorman, Čukuljeri 4,83 2,00 2,76 4,19 3,33 2,070 2,76 0,76 57,14 2 41,4 42,9 19,44
126 PMS8035 1 S Makedonija Bistra Mts., Careva Češma 4,81 2,61 2,85 4,47 3,37 1,960 2,85 0,24 59,25 2 54,3 40,7 17,29
127 PMS8036 2 S Makedonija Bistra Mts., Careva
Češma 5,10 2,75 2,92 4,28 3,19 2,180 2,92 0,17 57,25 2 53,9 42,7 19,44
128 PMS8038 1 1 S Makedonija Bistra Mts., Careva Češma 4,58 4,26 3,88 4,19 3,06 0,700 3,88 0,00 84,72 1 93,0 15,3 6,56
129 PMS9210 2 S Makedonija Bistra Mts., Brzovec 4,98 2,39 2,67 4,45 3,41 2,310 2,67 0,28 53,61 3 48,0 46,4 21,59
130 PMS9217 2 S Makedonija Jakupica, Gorno Begovo 4,98 2,58 3,08 4,27 3,46 1,900 3,08 0,50 61,85 2 51,8 38,2 17,29
131 PMS9218 2 S Makedonija Jakupica, Gorno Begovo 5,13 1,63 1,97 4,32 3,67 3,160 1,97 0,34 38,40 3 31,8 61,6 28,03
132 PMS9533 2 S Makedonija Bistra Mts., Carevec 5,25 2,95 3,24 4,58 3,40 2,010 3,24 0,29 61,71 2 56,2 38,3 19,44
133 PMS9534 1 1 S Makedonija Bistra Mts., Carevec 4,76 3,55 3,89 4,57 3,35 0,870 3,89 0,34 81,72 1 74,6 18,3 8,7
134 PMS9895 1 2 C Črna gora Orjen Mt., sedlo 4,62 1,80 2,91 4,11 3,36 1,710 2,91 1,11 62,99 2 39,0 37,0 15,14
135 PMS9896 2 1 C Črna gora Orjen, sedlo 5,05 3,67 3,64 4,48 3,44 1,410 3,64 0,00 72,08 1 72,7 27,9 13
136 PMS9897 2 3 C Črna gora Orjen, sedlo 4,55 1,38 1,73 4,18 2,68 2,820 1,73 0,35 38,02 3 30,3 62,0 25,88
137 PMS9898 2 4 C Črna gora Orjen, sedlo 4,70 4,29 2,94
138 PMS9899 2 1 C Črna gora Orjen, sedlo 4,70 3,59 3,77 4,28 3,32 0,930 3,77 0,18 80,21 1 76,4 19,8 8,7
139 PMS9900 2 1 C Črna gora Orjen, sedlo 4,73 4,13 4,32 4,43 3,35 0,410 4,32 0,19 91,33 1 87,3 8,7 4,41
140 PMS9901 1 2 C Črna gora Orjen, sedlo 4,41 1,90 3,20 4,13 3,33 1,210 3,20 1,30 72,56 1 43,1 27,4 10,85
141 PMS9902 1 1 C Črna gora Orjen, sedlo 4,74 3,95 4,11 4,36 3,43 0,630 4,11 0,16 86,71 1 83,3 13,3 6,56
142 PMS9903 1 1 C Črna gora Orjen, sedlo 4,62 3,16 3,68 3,94 3,41 0,940 3,68 0,52 79,65 1 68,4 20,3 8,7
143 PMS9905 1 1 C Črna gora Orjen, sedlo 5,06 3,86 4,28 4,63 3,38 0,780 4,28 0,42 84,58 1 76,3 15,4 8,7
144 PMS10409 2 1 S Makedonija Šar planina, Jelak 4,91 4,72 4,88 4,70 3,24 0,030 4,88 0,16 99,39 1 96,1 0,6 2,26
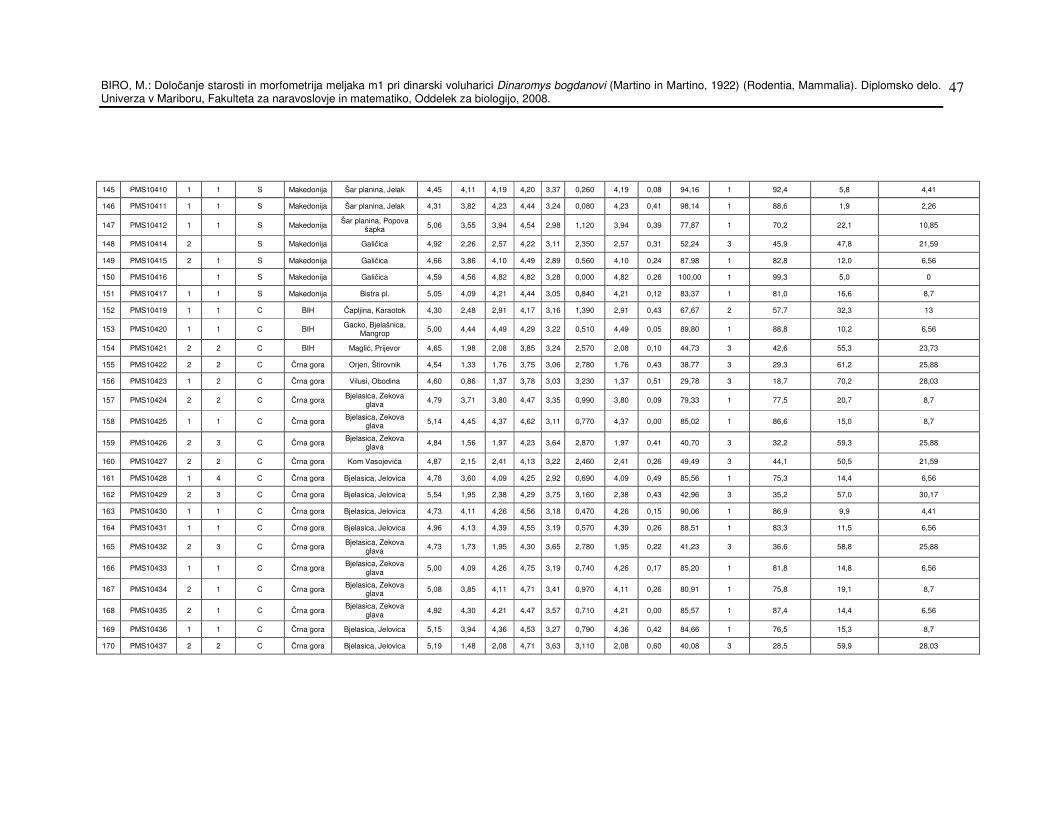
BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
47
145 PMS10410 1 1 S Makedonija Šar planina, Jelak 4,45 4,11 4,19 4,20 3,37 0,260 4,19 0,08 94,16 1 92,4 5,8 4,41
146 PMS10411 1 1 S Makedonija Šar planina, Jelak 4,31 3,82 4,23 4,44 3,24 0,080 4,23 0,41 98,14 1 88,6 1,9 2,26
147 PMS10412 1 1 S Makedonija Šar planina, Popova
šapka 5,06 3,55 3,94 4,54 2,98 1,120 3,94 0,39 77,87 1 70,2 22,1 10,85
148 PMS10414 2 S Makedonija Galičica 4,92 2,26 2,57 4,22 3,11 2,350 2,57 0,31 52,24 3 45,9 47,8 21,59
149 PMS10415 2 1 S Makedonija Galičica 4,66 3,86 4,10 4,49 2,89 0,560 4,10 0,24 87,98 1 82,8 12,0 6,56
150 PMS10416 1 S Makedonija Galičica 4,59 4,56 4,82 4,82 3,28 0,000 4,82 0,26 100,00 1 99,3 5,0 0
151 PMS10417 1 1 S Makedonija Bistra pl. 5,05 4,09 4,21 4,44 3,05 0,840 4,21 0,12 83,37 1 81,0 16,6 8,7
152 PMS10419 1 1 C BIH Čapljina, Karaotok 4,30 2,48 2,91 4,17 3,16 1,390 2,91 0,43 67,67 2 57,7 32,3 13
153 PMS10420 1 1 C BIH Gacko, Bjelašnica, Mangrop 5,00 4,44 4,49 4,29 3,22 0,510 4,49 0,05 89,80 1 88,8 10,2 6,56
154 PMS10421 2 2 C BIH Maglić, Prijevor 4,65 1,98 2,08 3,85 3,24 2,570 2,08 0,10 44,73 3 42,6 55,3 23,73
155 PMS10422 2 2 C Črna gora Orjen, Štirovnik 4,54 1,33 1,76 3,75 3,06 2,780 1,76 0,43 38,77 3 29,3 61,2 25,88
156 PMS10423 1 2 C Črna gora Vilusi, Obodina 4,60 0,86 1,37 3,78 3,03 3,230 1,37 0,51 29,78 3 18,7 70,2 28,03
157 PMS10424 2 2 C Črna gora Bjelasica, Zekova glava 4,79 3,71 3,80 4,47 3,35 0,990 3,80 0,09 79,33 1 77,5 20,7 8,7
158 PMS10425 1 1 C Črna gora Bjelasica, Zekova glava 5,14 4,45 4,37 4,62 3,11 0,770 4,37 0,00 85,02 1 86,6 15,0 8,7
159 PMS10426 2 3 C Črna gora Bjelasica, Zekova
glava 4,84 1,56 1,97 4,23 3,64 2,870 1,97 0,41 40,70 3 32,2 59,3 25,88
160 PMS10427 2 2 C Črna gora Kom Vasojevića 4,87 2,15 2,41 4,13 3,22 2,460 2,41 0,26 49,49 3 44,1 50,5 21,59
161 PMS10428 1 4 C Črna gora Bjelasica, Jelovica 4,78 3,60 4,09 4,25 2,92 0,690 4,09 0,49 85,56 1 75,3 14,4 6,56
162 PMS10429 2 3 C Črna gora Bjelasica, Jelovica 5,54 1,95 2,38 4,29 3,75 3,160 2,38 0,43 42,96 3 35,2 57,0 30,17
163 PMS10430 1 1 C Črna gora Bjelasica, Jelovica 4,73 4,11 4,26 4,56 3,18 0,470 4,26 0,15 90,06 1 86,9 9,9 4,41
164 PMS10431 1 1 C Črna gora Bjelasica, Jelovica 4,96 4,13 4,39 4,55 3,19 0,570 4,39 0,26 88,51 1 83,3 11,5 6,56
165 PMS10432 2 3 C Črna gora Bjelasica, Zekova glava 4,73 1,73 1,95 4,30 3,65 2,780 1,95 0,22 41,23 3 36,6 58,8 25,88
166 PMS10433 1 1 C Črna gora Bjelasica, Zekova
glava 5,00 4,09 4,26 4,75 3,19 0,740 4,26 0,17 85,20 1 81,8 14,8 6,56
167 PMS10434 2 1 C Črna gora Bjelasica, Zekova glava 5,08 3,85 4,11 4,71 3,41 0,970 4,11 0,26 80,91 1 75,8 19,1 8,7
168 PMS10435 2 1 C Črna gora Bjelasica, Zekova
glava 4,92 4,30 4,21 4,47 3,57 0,710 4,21 0,00 85,57 1 87,4 14,4 6,56
169 PMS10436 1 1 C Črna gora Bjelasica, Jelovica 5,15 3,94 4,36 4,53 3,27 0,790 4,36 0,42 84,66 1 76,5 15,3 8,7
170 PMS10437 2 2 C Črna gora Bjelasica, Jelovica 5,19 1,48 2,08 4,71 3,63 3,110 2,08 0,60 40,08 3 28,5 59,9 28,03

BIRO, M.: Določanje starosti in morfometrija meljaka m1 pri dinarski voluharici Dinaromys bogdanovi (Martino in Martino, 1922) (Rodentia, Mammalia). Diplomsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2008.
48
171 PMS10438 2 1 C Črna gora Bjelasica, Jelovica 4,90 4,27 4,50 4,82 3,24 0,400 4,50 0,23 91,84 1 87,1 8,2 4,41
172 PMS10439 2 2 C Črna gora Bjelasica, Jelovica 4,88 1,80 2,31 3,85 3,61 2,570 2,31 0,51 47,34 3 36,9 52,7 23,73
173 PMS10440 1 2 C Črna gora Kom Vasojevića 4,94 2,16 2,74 4,15 3,40 2,200 2,74 0,58 55,47 3 43,7 44,5 19,44
174 PMS10441 2 2 C Črna gora Durmitor, Dobri Do 4,87 1,60 1,78 4,11 3,60 3,090 1,78 0,18 36,55 3 32,9 63,4 28,03
175 PMS10442 1 S Makedonija Bistra planina 4,70 4,04
176 PMS10443 2 1 S Makedonija Šar planina, Jelak 4,79 4,01 4,09 4,30 3,10 0,700 4,09 0,08 85,39 1 83,7 14,6 6,56
177 PMS10444 1 4 C Črna gora Orjen 4,40 3,72 3,18
178 PMS10445 1 3 C Črna gora Bjelasica, Zekova
glava 4,38 1,39 1,73 3,78 3,52 2,650 1,73 0,34 39,50 3 31,7 60,5 23,73
179 PMS10446 1 1 C Črna gora Bjelasica, Zekova glava 4,71 3,64 3,71 4,30 2,87 1,000 3,71 0,07 78,77 1 77,3 21,2 10,85
180 PMS10447 1 1 C Črna gora Bjelasica, Jelovica 5,25 3,97 4,23 4,54 3,11 1,020 4,23 0,26 80,57 1 75,6 19,4 10,85
181 PMS10448 2 1 C Črna gora Bjelasica, Jelovica 5,21 3,82 4,64 4,71 3,25 0,570 4,64 0,82 89,06 1 73,3 10,9 6,56
182 PMS10449 1 1 C Črna gora Bjelasica, Jelovica 5,09 4,46 4,53 4,87 3,55 0,560 4,53 0,07 89,00 1 87,6 11,0 6,56
183 PMS10450 1 1 C Črna gora Durmitor, Dobri Do 5,02 3,87 4,39 4,54 3,52 0,630 4,39 0,52 87,45 1 77,1 12,5 6,56
184 PMS10451 2 2 W BIH Poklečani, Rakitno 4,28 1,97 2,65 4,27 3,10 1,630 2,65 0,68 61,92 2 46,0 38,1 15,14
185 PMS10453 S Makedonija Šar planina 4,33 2,65 3,19 4,13 3,05 1,140 3,19 0,54 73,67 1 61,2 26,3 10,85
186 PMS10455 2 2 C Kosovo Vrelo Belog Drima, Radovačka pećina 5,14 2,41 2,87 4,58 3,63 2,270 2,87 0,46 55,84 3 46,9 44,2 21,59
187 PMS10456 1 2 C Kosovo Žljeb, Lučanski stanovi 5,98 2,82 3,16 5,13 3,71 2,820 3,16 0,34 52,84 3 47,2 47,2 25,88
188 PMF13556 1 2 C Hrvaška Orebić (Sv. Ilija) 4,62 2,83 3,25 3,95 2,90 1,370 3,25 0,42 70,35 1 61,3 29,7 13