distribution of septohippocampal neurons containing parvalbumin or choline acetyltransferase in the...
TRANSCRIPT

THE JOURNAL OF COMPARATIVE NEUROLOGY 298:362-372 (1990)
Distribution of SeptohippocampalNeurons ContainingParvalbumin or Choline Acety!ltrderase in the Rat Brain
JOZSEF KISS, AMBFUSH J. PATEL, AND TAMAS F. FREUND MRC Collaborative Centre and Division of Neurophysiology and Neuropharmacology,
National Institute for Medical Research, London NW7 1AA (J.K., A.J.P.), and MRC Anatomical Neuropharmacology Unit, Department of Pharmacology,
Oxford (T.F.F.), England
ABSTRACT A combination of retrograde transport of horseradish peroxidase or wheat germ agglutinin-
colloidal gold with either single or double-label imrnunohistochemistry is used to describe the comparative topographic distribution of parvalbumin- and choline acetyltransferase- immunoreactive septal neurons that project to the hippocampal formation of the rat. The morphometric parameters of the retrogradely labelled, parvalbumin-containing neurons were very similar, if not identical, to those neurons of the midline and medial part of the medial septum and the diagonal band regions that had previously been shown to be immunoreactive for y-aminobutyric acid or for glutamate decarboxylase following colchicine treatment. The total number of parvalbumin-immunoreactive and choline acetyltransferase-positive retro- gradely labelled cells was counted at 9 representative levels through the rostrocaudal extension (from 2.4 mm anterior to the level of bregma) of the medial septal-diagonal band complex. In the whole medial septum-vertical limb of the diagonal band region, about 33% of the total retrogradely labelled neurons showed immunoreactivity to parvalbumin, whereas the parvalbu- min-negative cells were mainly choline acetyltransferase-immunopositive. In comparison with the average figure, the proportion of the retrogradely labelled parvalbumin-containing neurons was higher in the middle part (around 1.5 mm anterior to the bregma) than in either the rostra1 or caudal ends. The reverse was true for the distribution of the cholinergic septohippocampal neurons. At the maximum levels the parvalbumin-immunoreactive neurons accounted for more than half of the total retrogradely labelled cells in 4 out of 6 rats. Moreover, within the complexity of the septal neurons, a marked regularity of topographic organisation was observed in the distribution of retrogradely labelled parvalbumin-containing GABAergic and choline acetyltransferase-positive cholinergic neurons as if they were subdivided cytoarchitectonically.
Key words: septohippocampal projections, retrograde tracing, parvalbumin containing GABAergic neurons, cholinergic neurons, double-label immunostaining
In mammals, the septo-hippocampal axis plays an impor- tant role in memory and other cognitive processes (O'Keefe and NadeI, '78; Beatty and Carbone, '80; Low et al., '82; Seifert, '83; Gage et al., '84; Decker, '87; Fischer et al., '87). It has also been suggested that the functional abnormalities in subcortical cholinergic neurons are lesioned conspicu- ously in senescence (Gage et al., '84; Decker, '87; Fischer et al., '87) and in certain neurological disorders including senile dementia of the Alzheimer's type (Kartzman, '83; Roth and Iversen, '86; Decker, '87). This makes the septo- hippocampal pathway an obvious choice for extensive exper- imentation (Raisman, '66; Lewis and Shute, '67; Lewis et al., '67; Lynch et al., '78; Swanson and Cowan, '79; Chandler and Crutcher, '83; Baisden et al., '84; Kohler et
al., '84; Amaral and Kurz, '85; Frotscher and Leranth, '86; LBranth and Frotscher, '87; Freund and Antal, '88; KrnjeviC et a]., '88). In recent years, the endeavour has been tre- mendously helped by the availability of nerve cell specific antibodies and the advancement in immunohistological techniques. It is now well established that f ierents of the medial septal (MS)-diagonal band (DB) travel through the
Accepted April 16, 1990. Address reprint requests to A.J. Patel, MRC Collaborative Centre and
Division of Neurophysiology and Neuropharmacology, National Institute for Medical Research, The Ridgeway, Mill Hill, London NW7 IAA, England.
Dr. T.F. Freund is now at the Institute of Experimental Medicine, Hungarian Academy of Sciences, Budapest, Hungary.
o 1990 WILEY-LISS, INC.

CHARACTERIZATION OF SEPTOHIPPOCAMPAL NEURONS 363
dorsal fornix and fimbria to the hippocampus. In the hippocampal formation, they terminate profusely, espe- cially in the dentate gyrus and CA3 region and to a lesser extent in the CA1 region. These ascending septohippocam- pal projections mainly play a modulatory role in the overall control of the hippocampal electrical activity. When a conditioning pulse is applied to the MS, little or no field potential is recorded in the hippocampus, but it can sharply enhance the response (the population spikes) evoked by other inputs (KrnjeviC et al., '881, most notably by the pathway originating in the entorhinal cortex (Fantie and Goddard, '82; Bilkey and Goddard, '87). The septal input is also essential for the generation of the characteristic theta- rhythm in the hippocampus (Vanderwolf et al., '78; BUZS&, '84).
In recent years the presence of cholinergic and GABAer- gic neurons has been described in the MS-DB area of the adult rat brain (Mesulam et al., '83a; Baisden et al., '84; Kohler et al., '84; Panula et al., '84; Saper, '84; Amaral and Kurz, '85; Wainer et al., '85; Brashear et al., '86; Frotscher and Leranth, '86; Ichikawa and Hirata, '86; Onteniente et al., '86; Sofroniew et al., '87; KrnjeviC et al., '88; Kiss et al., '88, '90). Some of these studies have also provided informa- tion on the cholinergic cell projections from the MS-DB to the hippocampal formation (Lewis et al., '67; Moroni et al., '78; Mesulam et al., '83a; Baisden et al., '84; Amaral and Kurz, '85; Wainer et al., '85; Frotscher and Leranth, '861, but similar information on the MS-DB GABAergic neurons is sparse (Kohler et al., '84). This is because, in comparison with choline acetyltransferase (CUT), the immunoreactiv- ity for either GABA or glutamate decarboxylase (GAD) in this region is very weak and unreliable and can be observed only after colchicine treatment (Kohler et al., '84; Panula et al., '84; Onteniente et al., '86). In recent studies we overcame this problem by using an antibody directed against parvalbumin (PV) to visualise GABAergic neurons, since, in contrast to several other brain areas, a very large proportion of the GABAergic neurons contained PV in the midline and medial parts of the MS and the DB regions (Freund, '89; Kiss et al., '90). In addition, these GABAergic
A a AChE CA CCA C U T co DAB DB DG F FB GABA GAD h hDB HRP MS MS1 MSm MSme PAP PB PV S vDB WGA-gold
Abbreviations
anterior accumbens nucleus acetylcholinesterase anterior commissure corpus callosum choline acetyltransferase optic chiasma diaminobenzidine diagonal band of Broca dentate gyrus fimbria fast blue 7-aminobutyric acid glutamate decarboxylase hilus horizontal limb of the diagonal band horseradish peroxidase medial septum lateral part of the medial septum midline part of the medial septum medial part of the medial septum peroxidase-antiperoxidase phosphate buffer p arv alb u m in subiculum vertical limb of the diagonal band wheat germ agglutinin-colloidal gold
and PV-containing neurons were also shown to project to the hippocampus (Freund, '89). In the present study, using a quantitative retrograde tracing method, we report the relative proportion and the comparative topographic distri- bution of the PV-containing GABAergic and cholinergic MS-DB neurons specifically projecting to the hippocampal formation.
EXPEXUMENTALPROCEDURES Materials
The wheat germ agglutinin-colloidal gold (WGA-gold) conjugate was made and generously supplied by Dr. R.A.J. McIlhinney; antiserum to PV (Baimbridge et al., '82) and anti-ChAT monoclonal antibody (AB8; Mesulam et al., '83a) were kindly provided by Dr. K.G. Baimbridge and Dr. B.H. Wainer, respectively. The silver intensification kit was purchased from Janssen, Wantage. Other materials and chemicals used were obtained as described in previous publications (Freund and Antal, '88; Freund, '89; Kiss et al., '88, '90).
Preparation of the animals Retrograde transport of horseradish peroxidase (HRP,
30% in saline) or WGA-gold conjugate (approximately 17 nm in diameter) was used to determine the location of MS-DB neurons projecting to the hippocampus and dentate gyrus area. Ten Sprague-Dawley male rats aged approxi- mately 12 weeks (250 to 300 g body weight) were used in the present study. Under deep Equithesin anaesthesia (3 ml/kg body weight), a total volume of 200 nl of either HRP (to 4 rats) or WGA-gold conjugate (to 6 rats) was injected into the dorsal hippocampus, at coordinates - 3.2 mm posterior to the bregma, 1.8 mm lateral to the superior sagittal sinus, and 3.3 mm and 2.5 mm (at two sites) vertical below the brain surface. In the case of WGA-gold conjugate, before injection, the stock solution was concentrated by resuspend- ing the sediment, after centrifugation, to one-tenth of the original volume. The retrograde tracers were injected under pressure by a manual micromanipulator at a rate of 5-10 nl per minute and the needle was left in place for an additional 15 minutes to allow the tracer to diffuse away from the needle tip. Animals were killed at 72 to 96 hours after the injection of tracer by transcardial perfusion under deep anaesthesia. The brain areas containing the injection site and septal-diagonal band region of the basal forebrain were dissected and processed further as described by Kiss et al. ('88, '90).
Visualization of retmgmde tracers, and imrnunocytmhemidprocedures
The free-floating, 60-pm-thick serial Vibratome sections were extensively washed (6 times for 30 min) in 0.1 M phosphate buffer (PB), pH 7.4. The sections of the WGA- gold injected experiments were processed for silver intensi- fication (Somogyi and Hodgson, '85). After rinsing in water, the sections were incubated in a 1: 1 (by volumes) mixture of enhancer and initiator component of the silver intensifica- tion kit for 25 minutes. The sections were rinsed in water and then incubated in a 1% sodium thiosulphate solution for 5 minutes, followed by three washes, 10 minutes each, in PB. The sections were processed further for the immuno- cytochemical visualization of ChAT- and PV-containing neurons as described previously (Freund and Antal, '88;

364 J. KISS ET AL.
Kiss et al., ’90). A set of sections was processed for PV immunocytochemistry alone, using the peroxidase-antiper- oxidase (PAP)-diaminobenzidine (DAB) reaction (brown end product). On the other set of sections two-colour, double-label immunocytochemistry was performed to differ- entiate the cholinergic (ChAT-positive) and GABAergic (PV-immunoreactive) neurons containing WGA-gold end product (black granules). The PV immunoreactivity was demonstrated using the PAP-DAB reaction intensified with ammonium nickel sulphate (blue to black end product; Hancock, ’821, whereas the C U T immunoreactivity was demonstrated using the PAP-DAB reaction only (brown end product). In the sections of the HRP-injected experi- ments, the transported HRP was visualized by the nickel- intensified DAB reaction (blue to black granules). These sections were processed further for PV immunocytochemis- try alone using the PAP-DAB reaction (brown end product).
Quantitative studies All sections were prepared for light microscopy and cell
counts were made at x 500 magnification using an ocular grid as described previously by Kiss et al. (’90). The retrogradely transported material in the cells was checked occasionally at x 1,000 magnification. The retrogradely labelled total number of cells, PV-immunopositive neurons, and ChAT-immunoreactive nerve cells were counted at 9 representative levels throughout the rostrocaudal exten- sion ofthe MS-DB region of 6 animals. In carefully matched sections representing the same rostrocaudal level, there was no discrepancy in cell counts obtained on any neuronal population whether they were counted in single- or in double-labelled sections. At different rostrocaudal levels of the MS-DB region, the data on the retrogradely labelled PV- and CUT-immunoreactive neurons were presented sche- matically. Moreover, we have divided the MS-DB complex into the dorsally located MS nucleus and the ventrally located nucleus of the DB; the latter is further subdivided into dorsal and ventral parts according to Harkmark et al. (’75). The dorsal division of the DB is classically defined as the nucleus of the vertical limb of the diagonal band (vDB), whereas at a more caudal level the ventral division extends ventrolaterally to form a separate cell nucleus called the horizontal limb of the diagonal band (hDB) (also see Fig. l a in Kiss et al., ’90). For better visual comprehension, we have given the number of retrogradely labelled PV- and CUT-containing neurons as a percentage of the total retrogradely labelled cells a t different rostrocaudal levels of the MS-vDB region.
RESULTS Description of injection sites
In both the HRP and the WGA-gold conjugate injected rats, no spread of tracers outside the hippocampal forma- tion was observed in any of the 10 animals. Much of the cross-sectional area of the dorsal hippocampal formation was labelled with the heaviest deposition of the tracer in the hilar region and in the regiosuperior involving the area of the pyramidal layer and the stratum oriens (Fig. 1). Since the diffusion of the WGA-gold tracer is very limited, this had restricted the area from which retrograde transport of WGA-gold can take place in comparison with the HRP (Fig. 1).
Generalcommentsandtechnical considerations
In the present study, we have used the term “hippocam- pal formation” or “hippocampal region” or “hippocamp~s” interchangeably; the term includes the hippocampus proper and the dentate gyrus except when they are specified separately. The reaction end products in cells labelled with the retrogradely transported HRP had bluish black gran- ules, whereas WGA-gold conjugate had dark brown-black granules (Fig. 2 ) . Even though we have not carried out a systematic quantitative comparison between the nickel- DAB reaction product for HRP and the end product of silver intensified WGA-gold conjugate, it was apparent that the latter method produced heavy deposition of the reaction product in both PV- and ChAT-immunopositive retro- gradely labelled cells (Fig. 3). In double-immunolabelled experiments, the dark brown-black granulelike WGA-gold reaction end product could be easily distinguished from the bluish coloured reaction product of PAP-nickel-DAB in PV-containing neurons or from the diffuse brownish end product of the cholinergic cells (Fig. 3). In these experi- ments in which sections were processed for PV-immunocy- tochemistry with the PA€’-nickel-DAB and ChAT with the PAP-DAB method, the retrograde end product was found to be accumulated within the cytoplasm and the dendrites of both the bluish-coloured PV-containing and the brownish- coloured ChAT immunopositive neurons (Fig. 3). However, the blue-coloured cytoplasm of heavily retrogradely labelled PV-coptaining cells usually had a slightly dark appearance in the light microscope. The cells that displayed both the diffusely stained immunoreactivity and the reaction end product of the retrograde tracer were therefore defined as cholinergic or PV-containing neurons that project to the
Fig. 1. Location and extent of a typical injection site of (a) horserad- ish peroxidase (HRP) and (b) wheat germ agglutinin-colloidal gold (WGA-gold) in the dorsal hippocampal formation (coordinates - 3.2 mm posterior to the bregma, 1.8 mm lateral to the superior sagittal sinus and 3.3 mm and 2.5 mm vertical below the brain surface). The
diffusion of the WGA-gold tracer is very limited; therefore the area from which retrograde transport can take place is much smaller than that for HRP (if similar volumes were injected). Abbreviations: CA1 and CA3, two areas of pyramidal cell layer; DG, dentate gyrus; F, fimbria; h, hilus; s, subiculum.

CHARACTERIZATION OF SEFTOHIPPOCAMPAL NEURONS 365
hippocampal formation. In the single immunolabel experi- ments, the granular reaction product of the retrogradely transported HRP or WGA-gold conjugate was well distin- guishable from the diffuse staining produced by immunocy- tochemistry (Fig. 2). In these experiments, the cells that contained only HRP or WGA-gold conjugate reaction prod- uct were considered as PV-immunonegative neurons project- ing to the hippocampus (Fig. 2b,d). A great majority of these retrogradely labelled PV-negative cells must be the cholinergic neurons, since more than 90% of the total retrogradely labelled neurons were accounted for by the PV- and the ChAT-immunopositive neurons.
In all divisions of the MS-DB complex, the small round, oval or fusiform and large multipolar cells were retro- gradely labelled after hippocampal injections (Figs. 2, 3). The majority of these neurons were located in the side ipsilateral to the injection; however, in all experiments a few retrogradely labelled cells were also observed in the contralateral side of the MS-DB. The present study was not aimed at examining afferent projections to the septal area, but a few labelled varicose axons on both sides of the MS nucleus were observed in the experiments in which HRP was injected into the hippocampus. For example, an HRP- labelled axon studded with boutons was found to be in contact with a PV-containing cell body and its proximal dendrites located in the contralateral side of the MS nucleus (Fig. 3e). Detailed studies on this interesting observation and on other hippocampal aeren ts are in progress, and these findings will be discussed in a separate publication.
Distribution and morphology of retrogradely labelledparvalbumin-immunoreactive cells The retrogradely labelled PV- and ChAT-immunopositive
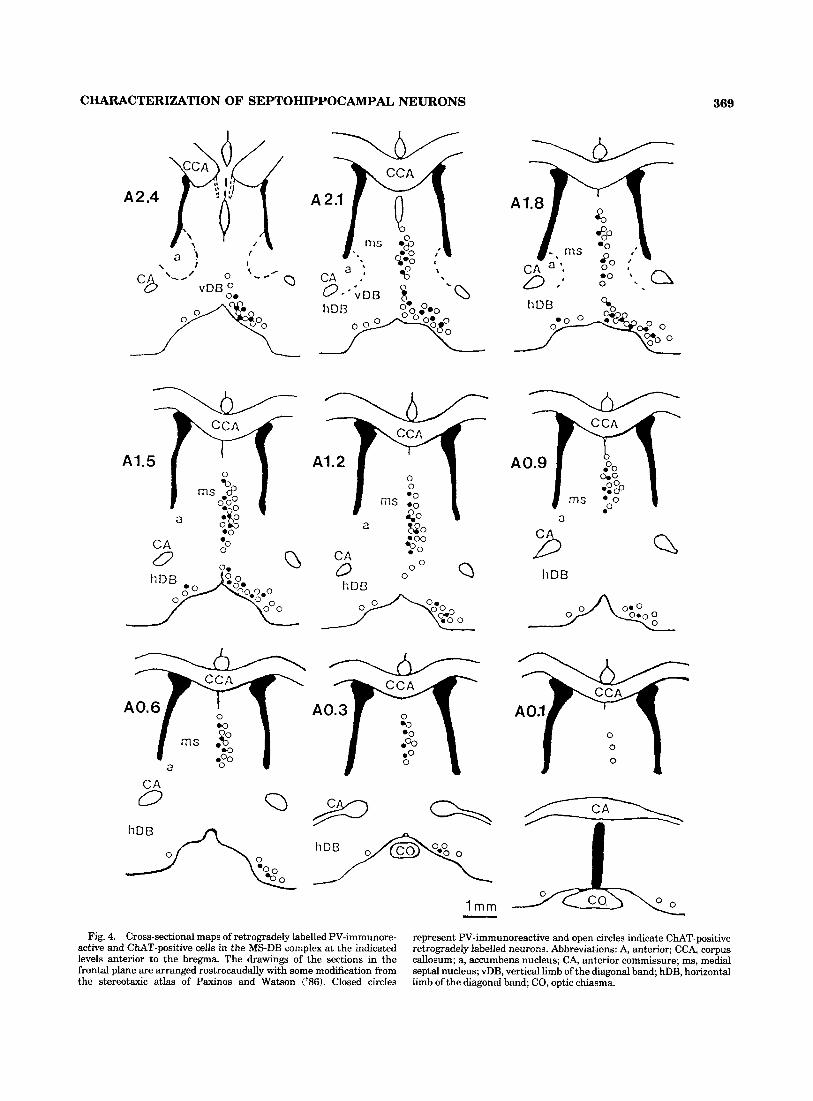
neurons were analysed on the same coronal serial sections starting from the level of the genu of the corpus callosum to the decussation of the anterior commissure (Fig. 4). In agreement with the previous study on the PV- and ChAT- positive cells (Kiss et al., 'go), the retrogradely labelled PV- and ChAT-containing neurons appear to form a continuous band of cells starting rostrally at the level of the anterior hippocampus rudiment and extend caudally as far posteri- orly as the anterior commissure (Fig. 4). In the MS-DB, the retrogradely labelled PV-containing neurons were distrib- uted throughout the region. Occasionally, retrogradely labelled cells with the WGA-gold conjugate were also ob- served in the dorsal and lateral parts of the lateral septal nucleus. These neurons were not immunopositive for either PV or CUT, and in this respect they differed from those present in the MS-DB complex.
At all levels of the MS nucleus examined (Figs. 2-4), the retrogradely labelled PV-containing cells were observed mainly in the midline (MSm) and the medial part (MSme) of the MS (for subdivisions see Fig. l b in Kiss et al., '90). They were mainly small or medium-size cells with an oval or elongated shape, oriented in parallel with the dorsoventral line in the cross-sectioned plane. In the MSme, however, larger multipolar or triangle-shape cells also contained reaction end product of the retrogradely labelled material. In the lateral part of the MS (MSl), a large proportion of neurons were PV-negative; thus it is not surprising to find only rarely retrogradely labelled PV-positive neurons (Figs. 2 4 ) . These retrogradely labelled nerve cells in the MSl were multipolar or triangle shaped and appeared to be very similar to those in the MSme. Even though it was difficult
to rigorously quantitate the mediolateral organization of the retrogradely labelled PV-containing cells, a mediolat- eral gradation was clearly visible in the organization of these neurons in the MS. The reverse was true for the retrogradely labelled PV-negative neurons (in the single immunolabel experiments) or immunopositive for ChAT (in the double-immunolabel experiments). The proportion of these cells in the MSm was very small, but their number increased markedly in the lateral part of the MS.
Retrogradely labelled PV-containing cells in the vDB were generally larger than those in the MS and they usually had a multipolar shape. Topographically these cells, which are scattered amongst the other neurons of the vDB, were more evenly distributed compared to those in the MS (Fig. 3). The PV containing retrogradely labelled cells were present in all parts of the vDB and through the whole rostrocaudal extent of this structure. However, a relatively high density of these retrogradely labelled cells was found in the area following the curvature of the ventral border of the vDB (Fig. 4).
Although in the hDB the total number of retrogradely labelled cells was very high, only a small proportion of these neurons were PV-immunopositive. Within the hDB the distribution of PV-positive neurons in the medial half of the nucleus was higher than in the ventral border of the limb (Fig. 4; also see Fig. 2 in Kiss et al., '90). Retrogradely labelled PV-containing neurons were also found inside the cell group organized in a globularlike pattern (Fig. 2). These PV-containing cells that project to the dorsal hippocampal formation from the hDB were large or medium-size neu- rons usually with a multipolar shape and no prominent orientation (Figs. 2, 4). However, a few bipolar and elon- gated cells could also be found in the hDB.
proportion of parvalbumh-jmmunoreactive retrogradely labelled cells at Merent levels of themedialseptumdiagonalbandregion
A close quantitative examination was carried out on the number of retrogradely labelled cells at 8 matched represen- tative levels of the MS-vDB region in 6 individual experi- ments. The results on the PV- and ChAT-containing neu- rons were expressed as a percentage of the total number of retrogradely labelled cells (Fig. 5). In all experiments, on the rostrocaudal line the values for the percentage of the retrogradely labelled PV-containing cells were low at the rostral end (2.4 mm) but increased progressively to a maximum level in the middle part (between 1.8 mm and 1.2 mm), followed by a gradual decrease toward the caudal end (lower than 0.3 mm) of the MS-vDB region (Fig. 5a), whereas the reverse was true for the ChAT-positive neu- rons (Fig. 5b). On average the number of ChAT-containing cells was always higher than PV-immunopositive neurons at all levels (Fig. 5 ) . However, at the maximum levels, the PV-immunopositive neurons accounted for more than half (between 69% and 52%) of the total retrogradely labelled cells in 4 out of 6 rats; in the other 2 animals the highest proportion of PV-containing retrogradely labelled neurons were 41% and 34% of the total retrogradelly Iabelled cells.
DISCUSSION Septohippocampal cholhergic neurons
The first indications of the types of neurotransmitters involved in the operation of the septohippocampal axis
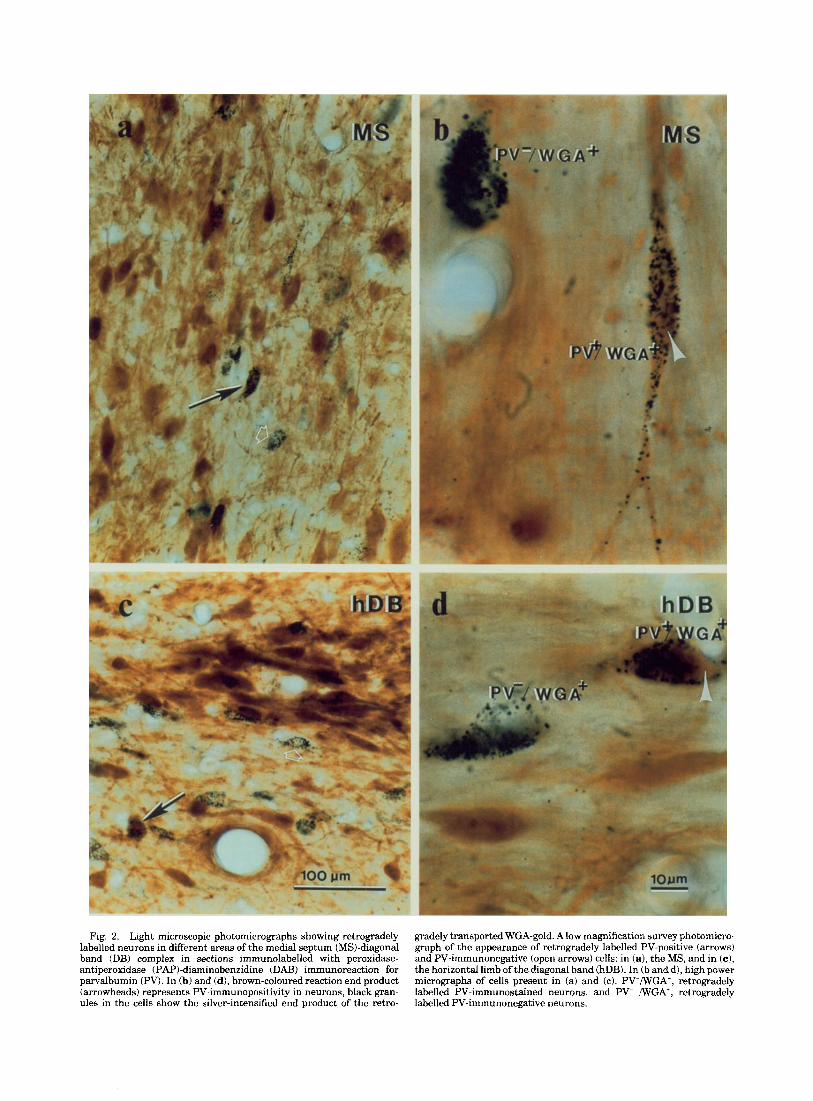
Fig. 2. Light microscopic photomicrographs showing retrogradely gradely transported WGA-gold. A low magnification survey photomicro- labelled neurons in different areas of the medial septum (MS)-diagonal graph of the appearance of retrogradely labelled PV-positive (arrows) band (DB) complex in sections immunolabelled with peroxidase- and PV-immunonegative (open arrows) cells: in (a), the MS, and in (c ) , antiperoxidase (PAP)-diaminobenzidine (DAB) immunoreaction for the horizontal limb of the diagonal band (hDB). In (b and d), high power parvalbumin (PV). In (b) and (d), brown-coloured reaction end product micrographs of cells present in (a) and (c). PV+/WGA+, retrogradely (arrowheads) represents PV-immunopositivity in neurons, black gran- labelled PV-immunostained neurons, and PV- /WGA+, retrogradely ules in the cells show the silver-intensified end product of the retro- labelled PV-immunonegative neurons.

Fig. 3. (a-d) High power photomicrographs showing retrograde labelling and PV- or CUT-immunostaining in neurons of the MS. On the same section, the blue-black-coloured PV-immunoreactive cells (arrowheads) are stained by the PAP-nickel-intensified-DAB reaction, whereas the brown CUT-immunopositive cells are stained by only DAB (thick arrows), using a double-immunolabel procedure. In (c and d), blue-black stained PV-positive and brown coloured ChAT-immuno- stained cells are shown at two different focal planes. Both these cells are retrogradely labelled by WGA-gold. In (c), the CUT-immunopositive
neuron (star) is in focus; in (d) the PV-containing cell (asterisk) is in focus. (e) High power photomicrograph of a section from the MS contralateral to the site of HRP injection into the hippocampal forma- tion. This section is immunostained for PV by the single PAP-DAB immunoreaction and the transported HRP is visualized by nickel intensified-DAB immunostaining. The brown-coloured cell is immuno- positive for PV, an axon with large varicosities (arrows) surrounding the perikaryon and the proximal dendrite exhibits blue to black staining of the transported HRP. Scale bar for all pictures is shown in (c).

368
emerged in the early 1960s. On the basis of electrophysiolog- ical and acetylcholinesterase histochemical findings, it was proposed that the septal afFerents innervating the hippocm- pal formation are cholinergic (Stumpf, '65; Lewis and Shute, '67; Lewis et al., '67). The proposal was supported by later biochemical observations. For example, the destruc- tion of the septal nuclei resulted in a significant decrease in the enzymes responsible for the synthesis and utilization of acetylcholine in the hippocampus (Lewis et al., '67; Mell- gren and Srebro, '73; Fonnum et al., '77; Fonnum and Walaas, '78), and the release or turnover of acetylcholine in the hippocampus was predictably altered by procedures that affect the firing of septal cells in normal animal or in experimental animals pretreated with neurotoxins, which are known to compromise the survival of the septal neurons (Rommelspacher et al., '74; Smith, '74; Dudar, '75; Blaker et al., '83; Galey et al., '85). In the CA1, the CA3 and the dentate gyrus regions of the hippocampal formation, the excitatory, inhibitory, and disinhibitory electrophysiologi- cal responses have been recorded by iontophoretic applica- tion of acetylcholine (Ben-Ari et al., '81; Dodd et al., '81; Cole and Nicoll, '84; KrnjeviC et al., '88). It was also suggested that the periodic excitation of a great number of hippocampal principal neurons by the axons of cholinergic "pacemaker" cells of the septum may be responsible for the theta rhythmic activity in the hippocampus (Bland et al., '80; Buzsaki, '84).
KrnjeviC and colleagues ('88; see also Ben-Ari et al., '81) have suggested that the effect of acetylcholine is modula- tory and is mediated by a combination of postsynaptic depression of potassium currents and disinhibition. The latter effect may be due to a reduction in presynaptic GABA release by hippocampal inhibitory interneurons (Valentino and Dingledine, '81). In contrast, Buzsaki ('84) has argued against the excitatory action of cholinergic septal neurons on the principal cells of the hippocampal formation. He has suggested that septal cells inhibit rather than excite princi- pal neurons in the hippocampal region; the effect may be mediated by direct excitation of the inhibitory interneu- rons. Several morphological studies have attempted to define the cells of origin (Mesulam et al., '83a; Baisden et al., '84; Kohler et al., '84; Amaral and Kurz, '85; Wainer et al., '85; Freund, '89) and the mode of termination of the septohippocampal pathways (Clark, '85; Frotscher and Leranth, '86; Leranth and Frotscher, '87; Freund and Antal, '881, but the picture is still not very clear.
In the present study we have provided precise and comprehensive information on the characterization, the topographical localization, and the semiquantitative estima- tion of the neurons involved in the septohippocampal pathway (Figs. 4, 5). A large proportion of the cholinergic cells projecting to the hippocampal formation are situated in the MS-DB region (also see Amaral and Kurz, '85). On the rostrocaudal axis of the MS-DB, the proportion of projecting cholinergic neurons is much higher at both rostral and caudal ends than in the middle part; at different levels examined the proportion (average value of 6 animals) of the CUT-positive cholinergic cells varied between 53% and 80% of the total retrogradely labelled nerve cells (Fig. 5b). When all retrogradely labelled cells were added to- gether, the cholinergic neurons ranged between 53% and 78% (mean = 65%) of the total projecting neurons in different experiments.
J. KISS ET AL.
Septohippocampal parvalbumjn containing GABAergicneurons
Not all septohippocampal fibres are cholinergic (Lynch et al., '78; Mesulam et al., '83a,b; Amaral and Kurz, '85; Wainer et al., '85). A large proportion of the noncholinergic component appears to utilise GABA as neurotransmitter (Kohler et al., '84; Freund and Antal, '88; Freund, '89). Recently, it has been shown that the GABAergic afferents from the septum terminate mainly on the inhibitory inter- neurons of the hippocampus and the dentate gyrus (Freund and Antal, '88). It has been proposed, therefore, that these GABAergic afferents are responsible for the septal (atro- pine resistent; Fantie and Goddard, '82) facilitation of hippocampal granule cell responses, by inhibiting the inhib- itory interneurons of the hippocampal formation (Fantie and Goddard, '82; Bilkey and Goddard, '87; Freund and Antal, '88). The present findings have extended our under- standing on the noncholinergic neurons of the septum projecting to the hippocampal formation. The sections double labelled for PV- and CUT-positive neurons demon- strated that the majority of retrogradely labelled noncholin- ergic cells showed immunoreactivity for PV. In addition, the topographic distribution, size, and morphological appear- ance of these PV-containing neurons were very similar to those neurons of the midline and medial parts of the MS and the DB regions that had previously been demonstrated to be immunoreactive for GABA or GAD following colchi- cine treatment (Figs. 3-5) (Kohler et al., '84; Panula et al., '84; Brashear et al., '86; Onteniente et al., '86). When retrogradely labelled cells of all levels were added together in different experiments, the PV-positive GABAergic neu- rons ranged between 22% and 47% (mean = 33%) of the total projecting neurons. These figures for the proportion of PV-containing retrogradely labelled septal cells were in good agreement with previous estimations on the noncholin- ergic septohippocampal neurons (Mesulam et al., '83a,b; Baisden et al., '84; Amaral and Kurz, '85; Wainer et al., '85). However, the topographic distribution of the PV- containing neurons was not uniform. On the rostrocaudal axis of the MS-DB, the proportion of the retrogradely labelled PV-containing GABAergic neurons was much higher in the middle part than in either rostral or caudal ends (Fig. 5a). It should be pointed out that even in these defined regions marked heterogeneity does exist, and within each region patches of cells were often seen that would range from mainly PV-positive to almost totally C U T - positive.
Possible functional implications Recently, Griffith and co-workers ('86, '88) have de-
scribed, on the basis of duration of postspike afterhyperpo- larisations, 2 types of neurons in the MS-DB complex using a guinea pig brain slice preparation. The acetylcholines- terase (AChE)-positive neurons (presumably cholinergic) showed slow afterhyperpolarisation and longer duration of evoked action potential. The basal forebrain neurons with the similar electrophysiological characteristics have been observed in living animals (Lamour et al., '84), and they are believed to be responsible for the generation of atropine- sensitive theta-rhythms in the hippocampus (Bland et al., '80; Buzsaki, '84). The second type of neurons (AChE- negative) in the MS-DB was capable of firing very rapidly with fast afterhyperpolarisation. The proportion of these noncholinergic neurons with fast afterhyperpolarisation
CHARACTERIZATION OF SEPTOHIPPOCAMPAL NEURONS 369
A2.4
%'
A l ,
A hDB
$? CA 0 0 O 0 Q
a
D h D B
Q h D B
0.0 0 00 0
1 mm - Fig. 4. Cross-sectional maps of retrogradely labelled PV-immunore-
active and CUT-positive cells in the MS-DB complex at the indicated levels anterior to the bregma. The drawings of the sections in the frontal plane are arranged rostrocaudally with some modification from the stereotaxic atlas of Paxinos and Watson ('86). Closed circles
represent PV-immunoreactive and open circles indicate CUT-positive retrogradely labelled neurons. Abbreviations: A, anterior; CCA, corpus callosum; a, accumbens nucleus; CA, anterior commissure; ms, medial septal nucleus; vDB, vertical limb of the diagonal band; hDB, horizontal limb of the diagonal band; CO, optic chiasma.

370 J. KISS ET AL.
(a) pv-positive
T
2.4 2.1 1.
c
L
1. L .6 0:3
80 -
% 4- 7 0 - 0
0 a,
id C a,
- c
0) 6 0 -
2 a" 5 0 -
I
40 -
2 . 2.1 1.8 1.5 1.2
AP Coordinates anterior from bregrna(mm)
Fig. 5 . Changes in the proportion of retrogradely labelled (a) PV-positive and (b) CUT-immunoreactive neurons in the MS-vDB at 8 indicated levels anterior to the bregma. At each anteroposterior iAP) level the values for retrogradely labelled PV- and CUT-containing
(Griffith, '88) was within the range of our estimates on the proportion of PV-containing GABAergic neurons (Kiss et al., '90) or previously measured noncholinergic neurons (Amaral and Kurz, '85; Wainer et al., '85). The apparent close relationship between the neurons with fast afterhyper- polarisation and the cells with immunoreactivity for PV raises the possibility that functionally different GABAergic neurons in the septum may contain different calcium binding proteins. Indeed, a large number of noncholinergic neurons in the lateral part of the MS and the lateral septum showed immunoreactivity for calbindin (a vitamin D-depen- dent calcium binding protein with 28 KDa molecular weight; Baimbridge et al., '82; Heizmann and Celio, '87; Sloviter, '89), whereas in the MSm, the MSme, and the DB regions only a few scattered neurons were positive for calbindin (unpublished observations). Previous electrophysiological studies have indicated that the cells of the pars dorsalis of the lateral septal nucleus do not project outside the septum (DeFrance, '76). It would, therefore, appear that in the septum calbindin is mainly present in GABAergic interneu- rons, whereas the GABAergic projecting nerve cells, presum- ably with fast afterhyperpolarisation, are endowed with PV. Moreover, it is interesting to note that kindling is shown to produce differential effects on these two types of calcium binding protein containing neurons in the hippocampal formation. In the granule cell of the fascia dentata kindling gradually decreases calbindin (Baimbridge et al., '85), whereas it increases the number of PV-immunoreactive neurons in the CA1 region (Kamphuis et al., '89). In the latter study the presence of PV is found to exert a protective effect against the process that leads to a decrease in the
- . .9
neurons are expressed as a percentage of the total retrogradely labelled cells (100%). Each column is the mean value of 6 experiments with S.E.M. indicated by the bar.
number of GABA-immunoreactive neurons (Kamphuis et al., '89). Also, in the stratum pyramidale of the CA1 region, the fast-spiking interneurons are shown to contain PV, although the actual physiological function of PV in these cells needs to be clarified (Kawaguchi et al., '87).
Concluding remarks In the present study by combining retrograde tracing
with immunohistochemical methods, we have firmly estab- lished the origin and chemical nature of septal neurons projecting to the hippocampal formation. Within the com- plexity of the septal neurons, a marked regularity of topographic organisation was observed in the distribution of retrogradely labelled ChAT-positive cholinergic and PV- containing GABAergic neurons. At present it is not clear: (1) whether the various compartments of the MS-DB complex containing different proportions of GABAergic and cholinergic projection neurons also differ in their input and output characteristics, (2) whether the GABAergic and cholinergic septohippocampal neurons are active during different behaviours generating appropriate patterns of hippocampal electrical activity (i.e., theta activity or sharp waves accompanying low voltage high frequency activity) or they influence the hippocampal circuitry synergistically during the same behaviours but via different mechanisms, and (3) whether the presence of different calcium binding proteins in certain subsets of septal neurons could be related to different functions. Clearly, a well-coordinated anatomical, electrophysiological, pharmacological, and bio- chemical approach to these and other related questions will

CHARACTERIZATION OF SEPTOHIPPOCAMPAL NEURONS 371
aid in our understanding of memory-linked hippocampal processes normally and in patients with learning and memory disorders.
A C K " T S This work was supported in part by Beechams Pharma-
ceuticals. We thank Mrs. K. Somogyi and Mrs. K. Boczko for excellent technical assistance, and Drs. P. Somogyi, C. Clemente, M.S.G. Clark, and F. Brown for helpful discus- sions. We are grateful to Dr. R.A.J. McIlhinney for the gift of a conjugate of WGA and colloidal gold, to Dr. K.G. Baimbridge for the parvalbumin antiserum, and to Dr. B.H. Wainer for the anti-ChAT monoclonal antibody.
Amaral, D.G., and J. Kurz (1985) An analysis of the origins of the cholinergic and noncholinergic septal projections to the hippocampus formation of the rat. J. Comp. Neurol. 240:37-59.
Baimbridge, K.G., J.J. Miller, and C.O. Parkes (1982) Calcium-binding protein distribution in the rat brain. Brain Res. 239:519-525.
Baimbridge, K.G., I. Mody, and J.J. Miller (1985) Reduction of rat hippocam- pal calcium binding protein following commissural, amygdala, septal, perforant path and olfactory bulb kindling. Epilepsia 26t460-465.
Baisden, R.H., M.L. Woodruff, and D.B. Hoover (1984) Cholinergic and noncholinergic septo-hippocampal projections: A double-label horserad- ish peroxidase-acetylcholinesterase study in the rabbit. Brain Res. 290:14&151.
Beatty, W.W., and C.P. Carbone (1980) Septal lesions, intramaze cues and spatial behavior in rats. Physiol. Behav. 24:675-678.
Ben-Ari, Y., K. KrnjeviC, W. Reinhardt, and N. Ropert (1981) Intracellular observations on the disinhibitory action of acetylcholine in hippocampus. Neuroscience 6.2475-2481.
Bilkey, D.K., and G.V. Goddard (1987) Septohippocampal and commissural pathways antagonistically control inhibitory interneurons in the dentate gyrus. Brain Res. 405:320-325.
Blaker, W.D., D.L. Cheney, 0. Gandolfi, and E. Costa (1983) Simultaneous modulation of hippocampal cholinergic activity and extinction by intrasep- tal muscimol. J. Pharmacol. Exp. Ther. 225:361-365.
Bland, B.H., P. Andersen, T. Ganes, and 0. Sveen (1980) Automated analysis of rhythmicity of physiologically identified hippocampal formation neu- rons. Exp. Brain Res. 38:205219.
Brashear, H.R., L. Zaborszky, and L. Heimer (1986) Distributionof GABAer- gic and cholinergic neurons in the rat diagonal band. Neuroscience 17t439-451.
Buzsiki, G. (1984) Feed-forward inhibition in the hippocampal formation. Prog. Neurobiol. 22131-153.
Chandler, J.P., and K.A. Crutcher (1983) The septohippocampal projection in the rat: An electron microscopic horseradish peroxidase study. Neuroscience 10:685496.
Clark, D.J. (1985) Cholinergic innervation of the rat dentate gyms: An immunocytochemical and electron microscopical study. Brain Res. 360: 349354.
Cole, A.E., and R.A. Nicoll(1984) The pharmacology of cholinergic excitatory responses in hippocampal pyramidal cells. Brain Res. 305283-290.
Decker, M.W. (1987) The effects of aging on hippocampal and cortical projections of the forebrain cholinergic systems. Brain Res. Rev. 12:423- 438.
DeFrance, J.F. (1976) A functional analysis of the septal nuclei. Adv. Behav. Biol. 20:185-227.
Dodd, J., R. Dingledine, and J.S. Kelly (1981) The excitatory action of acetylcholine on hippocampal neurons of the guinea pig and rat main- tainedinvitro. Brain Res. 207:109-127.
Dudar, J.D. (1975) The effect of septal nuclei stimulation on the release of acetylcholine from the rabbit hippocampus. Brain Res. 83:123-133.
Fantie, B.D., and G.V. Goddard (1982) Septal modulation of the population spike in the fascia dentata produced by perforant path stimulation in the rat. Brain Res. 252:227-237.
Fischer, W., K. Wictorin, A. Bjorklund, L.R. Williams, S. Varon, and F.H. Gage (1987) Amelioration of cholinergic neuron atrophy and spatial memory impairment in aged rats by nerve growth factor. Nature 329:6548.
Fonnum, F., and I . Walaas (1978) The effect of intrahippocampal kainic acid injections and surgical lesions on neurotransmitters in hippocampus and septum. J. Neurochem. 31:1173-1181.
Fonnum, F., I. Walaas, and E. Iversen (1977) Localization of GABAergic, cholinergic and aminergic structures in the mesolimbic system. J. Neurochem. 29:221-230.
Freund, T.F. (1989) GABAergic septohippocampal neurons containparvalbu- min. Brain Res. 478:375381.
Freund, T.F., and M. Antal(1988) GABA-containing neurons in the septum control inhibitory interneurons in the hippocampus. Nature 336:170- 173.
Frotscher, M., and C. Leranth (1986) The cholinergic innervation of the rat fascia dentata: identification of target structures on granule cells by combining choline acetyltransferase immunocytochemistry and Golgi impregnation. J. Comp. Neurol. 243:58-70.
Gage, F.H., A. Bjorklund, U. Stenevi, S.B. Dunnett, and P.A.T. Kelly (1984) lntrahippocampal septal grafts ameliorate learning impairments in aged rats. Science 225:533-536.
Galey, D., T. Durkin. G. Sifakis, E. Kempf, and R. Jaf€ard (1985) Facilitation of spontaneous and learned spatial behaviours following 6-hydroxydopa- mine lesions of the lateral septum: a chloninergic hypothesis. Brain Res. 340:171-174.
Griffth, W.H. (1988) Membrane properties of cell types within guinea pig basal forebrain nuclei in vitro. J. Neurophysiol. 59:1590-1612.
Griffith, W.H., and R.T. Matthews (1986) Electrophysiology of AChE- positive neurons in basal forebrain slices. Neurosci. Lett. 71t169-174.
Hancock, M.B. (1982) DAB-Nickel substrate for the differential immunoper- oxidase staining of nerve fibres and fibre terminals. J. Histochem. Cytochem. 30:578P.
Harkmark, W., S.I. Mellgren, and B. Srebro (1975) Acetylcholinesterase histochemistry of the septal region in rat and human: distribution of enzyme activity. Brain Res. 95281-289.
Heizmann, C.W., and M.R. Celio (1987) Immunolocalization of parvalbumin. Methods Enzymol. 139:552-570.
Ichikawa, T., andY. Hirata (1986) Organization of choline acetyltransferase- containing structures in the forebrain of the rat. J. Neurosci. 6281-292.
Kamphuis, W., E. Huisman, W.J. Wadman, C.W. Heizmann, andF.H. Lopes de Silva (1989) Kindling induced changes in parvalbumin immunoreac- tivity in rat hippocampus and its relation to long-term decrease in GABA-immunoreactivity. Brain Res. 479:23-34.
Kawaguchi, Y., H. Katsumaru, T. Kosaka, C.W. Heizmann, and K. Hama (1987) Fast spiking cells in rat hippocampus (CA1 region) contain the calcium-binding protein parvalbumin. Brain Res. 416t369-374.
Kartzman, R. (ed) (1983) Biological Aspects of Alzheimer's Disease. Cold Spring Harbor, Ny: Cold Spring Harbor Laboratory.
Kiss, J., J. McGovern, and A.J. Patel (1988) Immunohistochemical localiza- tion of cells containing nerve growth factor receptors in the different regions of the adult rat forebrain. Neuroscience 27:731-748.
Kiss, J., A.J. Patel, K.G. Baimbridge, and T.F. Freund (1990) Topographical localization of neurons containing parvalbumin and choline acetyltrans- ferase in the medial septum-diagonal band region of the rat. Neuro- science 36:61-72.
Kohler, C., V. Chan-Palay, and J-Y. Wu (1984) Septal neurons containing glutamic acid decarboxylase immunoreactivity project to the hippocam- pal region in the rat brain. Anat. Embryol. 169:41-44.
Krnjevic, K., N. Ropert, and J. Casullo (1988) Septohippocampal distribu- tion. Brain Res. 438:182-192.
Lamour, Y., P. Dutar, and A. Jobert (1984) Septohippocampal and other medial septum-diagonal band neurons. electrophysiological and pharma- cological properties. Brain Res. 309:227-239.
Leranth, C., and M. Frotscher (1987) Cholinergic innervation of hippocam- pal GAD- and somatostatin-immunoreactive commissural neurons. J. Comp. Neurol. 26k33-47.
Lewis, P.R., and C.C.D. Shute (1967) The cholinergic limbic system: projections to hippocampal formation, medial cortex nuclei of the ascending cholinergic reticular system and the subfornical organ and supra-optic crest. Brain 90:521-537.
Lewis, P.R., C.C.D. Shute, and A. Silver (1967) Confirmation from choline acetylase analyses of a massive cholinergic innervation to the rat hippocampus. J. Physiol. (Lond.) 191:2 15-224.
Low, W.C., P.R. Lewis, S.T. Bunch, S.B. Dunnett, S.R. Thomas, S.D. Iversen, A. Bjorklund, and U. Stenevi (1982) Function recovery following neural transplantation of embryonic septal nuclei in adult rats with septohippocampal lesions. Nature 300.-260-262.

372 J. KISS ET AL. Roth, M., and L.L. Iversen (eds) (1986) Alzheimer's disease and related
disorders. Br. Med. Bull. 42-116. Saper, C.B. (1984) Organization of cerebral cortical afferent systems in the
rat. 11. Magnocellular basal nucleus. J. Comp. Neurol. 222t313-342. Seifert, W. (ed) (1983) Neurobiology of the Hippocampus. New York
Academic Press. Sloviter, R. (1989) Calcium-binding protein (calbindin-D,,,) and parvalbu-
min immunoqtochemistry: localization in the rat hippocampus with specific reference to the selective vulnerability of hippocampal neurons to seizure activity. J. Comp. Neurol. 280:183-196.
Smith, C.M. (1974) Acetylcholine release from the cholinergic septo- hippocampal pathway. Life Sci. 142159-2166.
Sofroniew, M.V., R.C.A. Pearson, and T.P.S. Powell (1987) The cholinergic nuclei of the basal forebrain of the rat: normal structure, development and experimentally-induced degeneration. Brain Res. 41 lt310-331.
Somogyi, P., and A.J. Hodgson (1985) Antisera to y-aminobutyric acid. 111. Demonstration of GABA in Golgi-impregnated neurons and in conven- tional electron microscopic sections of cat striate cortex. J. Histochem. Cytochem. 33249-257.
Stumpf, C. (1965) Drug action on the electrical activity of the hippocampus. Int. Rev. Neurobiol. 837-138.
Swanson, L.W., and W.M. Cowan (1979) The connections of the septal region in the rat. J. Comp. Neurol. 186;621-656.
Valentino, R.J., and R. Dingledine (1981) Presynaptic inhibitory effect of acetylcholine in the hippocampus. J. Neurosci. 1:784-792.
Vanderwolf, C.H., R. Kramis, and T.E. Robinson (1978) Hippocampal electrical activity during waking behaviour and sleep: analyses using centrally acting drugs. In Functions of the Septo-Hippocampal System. Ciba Foundation Symp. 58. Amsterdam: Elsevier, pp. 199-226.
Wainer, B.H.,A.I. Levey, D.B. Rye, M.-M. Mesulam, andE.J. Mufson (1985) Cholinergic and non-cholinergic septo-hippocampal pathways. Neurosci. Lett. 54:45-52.
Lynch, G., G. Rose, and C. Gall (1978) Anatomical and functional aspects of the septo-hippocampal projections. In Functions of the Septo-Hippocam- pal System. Ciba Foundation Symp. 58. Amsterdam: Elsevier, pp. 5-20.
Mellgren, S.I., and B. Srebro (1973) Changes in acetylcholinesterase and distribution of degeneration fibers in the hippocampal region after septal lesions in the rat. Brain Res. 52:19-36.
Mesulam, M.-M., E.J. Mufson, B.H. Wainer, andA.1. Levey (1983a) Central cholinergic pathways in the rat an overview based on alternative nomenclature (Chl-Ch6). Neuroscience 10: 1185-1201.
Mesulam, M.-M., E.J. Mufson, A.I. Levey, and B.H. Wainer (198313) Cholin- ergic innervation of cortex by the basal forebrain: cytochemistry and cortical connections of the septal area, diagonal band nuclei, nucleus basalis (substantia innominata), and hypothalamus in the rhesus mon- key. J. Comp. Neurol. 214:170-197.
Moroni, F., D. Malthe-Sbenssen, D.L. Cheney, and E. Costa (1978) Modula- tion of acetylcholine turnover in the septal hippocampal pathway hy electrical stimulation and lesioning. Brain Res. 150:333-341.
O'Keefe, J., and L. Nadel (1978) The Hippocampus as a Cognitive Map. Oxford: Clarendon Press.
Onteniente, B., H. Tago, H. Kimura, and T. Maeda (1986) Distribution of 7-aminobutyric acid-immunoreactive neurons in the septal region of the rat brain. J. Comp. Neurol. 248:422430.
Panula, P., A.V. Revuelta, D.L. Cheney, J-Y. Wu, and E. Costa (1984) An immunohistochemical study on the location of GABAergic neurons in rat septum. J. Comp. Neurol. 22259-80.
Paxinos, G., and C. Wat,son (1986) The Rat Brain in Stereotaxic Coordinates, 2nd ed. London: Academic Press.
Raisman, G. (1966) The connexions of the septum. Brain 89:317-348. Rommelspacher, H., A.M. Goldberg, and M.J. Kuhar (1974) Action of
hemicholinium-3 on cholinergic nerve terminals after alteration of neuronal impulse flow. Neuropharmacology 13: 1015-1023.