dithiol disulphide exchange in redox regulation of...
TRANSCRIPT

R
De
DJa
b
c
d
e
a
ARRAA
KRDDFTC
C
h0
Plant Science 255 (2017) 1–11
Contents lists available at ScienceDirect
Plant Science
journa l homepage: www.e lsev ier .com/ locate /p lantsc i
eview article
ithiol disulphide exchange in redox regulation of chloroplastnzymes in response to evolutionary and structural constraints
esirée D. Gütle a,b,c,d,∗,1, Thomas Roret a,b,1, Arnaud Hecker a,b, Ralf Reski c,d,e,ean-Pierre Jacquot a,b,∗
Université de Lorraine, UMR 1136 Interactions Arbres Microorganismes, F-54500 Vandœuvre-lès-Nancy, FranceINRA, UMR 1136 Interactions Arbres Microorganismes, F-54280 Champenoux, FrancePlant Biotechnology, Faculty of Biology, University of Freiburg, Schänzlestr. 1, 79104 Freiburg, GermanySpemann Graduate School of Biology and Medicine (SGBM), University of Freiburg, 79104 Freiburg, GermanyBIOSS − Centre for Biological Signalling Studies, University of Freiburg, Schänzlestr. 18, 79104 Freiburg, Germany
r t i c l e i n f o
rticle history:eceived 13 September 2016eceived in revised form 4 November 2016ccepted 5 November 2016vailable online 8 November 2016
eywords:edox regulationisulphideithiolerredoxinhioredoxinalvin-Benson cycle
a b s t r a c t
Redox regulation of chloroplast enzymes via disulphide reduction is believed to control the rates of CO2
fixation. The study of the thioredoxin reduction pathways and of various target enzymes lead to thefollowing guidelines:
i) Thioredoxin gene content is greatly higher in photosynthetic eukaryotes compared to prokaryotes;ii) Thioredoxin-reducing pathways have expanded in photosynthetic eukaryotes with four different
thioredoxin reductases and the possibility to reduce some thioredoxins via glutaredoxins;iii) Some enzymes that were thought to be strictly linked to photosynthesis ferredoxin-thioredoxin
reductase, phosphoribulokinase, ribulose-1,5-bisphosphate carboxylase/oxygenase, sedoheptulose-1,7-bisphosphatase are present in non-photosynthetic organisms;
iv) Photosynthetic eukaryotes contain a genetic patchwork of sequences borrowed from prokaryotesincluding �–proteobacteria and archaea;
v) The introduction of redox regulatory sequences did not occur at the same place for all targets. Somepossess critical cysteines in cyanobacteria, for others the transition occurred rather at the green algaelevel;
vi) Generally the regulatory sites of the target enzymes are distally located from the catalytic sites. The
cysteine residues are generally not involved in catalysis. Following reduction, molecular movementsopen the active sites and make catalysis possible;vii) The regulatory sequences are located on surface-accessible loops. At least one instance they can be cutout and serve as signal peptides for inducing plant defence.
© 2016 Elsevier Ireland Ltd. All rights reserved.
ontents
1. Principles of redox regulation of chloroplast enzymes via dithiol-disulphide exchange, what are the actors, what are the targets? . . . . . . . . . . . . . . . . .21.1. Diversification of the electron donor systems . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 21.2. The complex TRX gene family of plants . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2
2. Why did redox regulation via dithiol reduction appear? . . . . . . . . . . . . . . . . . .2.1. Oxidative stress increases during evolution of higher plants . . . . . .2.2. Functional modifications of target proteins . . . . . . . . . . . . . . . . . . . . . . . .
∗ Corresponding authors at: Université de Lorraine, UMR 1136 Interactions Arbres MicrE-mail addresses: [email protected] (D.D. Gütle), j2p@univ-lorra
1 Joint first authors.
ttp://dx.doi.org/10.1016/j.plantsci.2016.11.003168-9452/© 2016 Elsevier Ireland Ltd. All rights reserved.
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4
oorganismes, Vandœuvre-lès-Nancy, F-54500, France.ine.fr (J.-P. Jacquot).

2 D.D. Gütle et al. / Plant Science 255 (2017) 1–11
3. When and how did redox regulation via dithiol reduction appear? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 43.1. Occurrence of TRX reductases . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 43.2. Integration of redox sensitive cysteine residues in target protein sequences . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4
4. In response to which structural constraints did redox regulation happen on molecular targets? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 64.1. ATP synthase . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 64.2. NADP-malate dehydrogenase . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 84.3. Glucose 6-phosphate dehydrogenase . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 84.4. Fructose-1,6-bisphosphatase and sedoheptulose-1,7-bisphosphatase . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 84.5. Phosporibulokinase . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 84.6. Glyceraldehyde 3-phosphate dehydrogenase . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 8
5. Do the regulatory sequences have additional roles in signalling? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 96. Potential new avenues of research in the redox field . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 9
Acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .10 . . . . . .
1dt
1
tiepbdugttsembmNsffiwutpsortaepCtaivCtttFpc
References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
. Principles of redox regulation of chloroplast enzymes viaithiol-disulphide exchange, what are the actors, what arehe targets?
.1. Diversification of the electron donor systems
Redox regulation by dithiol-disulphide exchange reactionsogether with the thiol-dependent repair systems (peroxiredox-ns (PRX) and methionine sulfoxide reductases (Msr)) is gettingver more attention in all biological systems, but it is clearly inlants and especially in land plants that these processes haveecome amazingly diversified [1–5]. Ubiquitously present, thiore-oxin proteins (TRX) are central to those reactions. They havenmatched ability to reduce (and oxidize) disulphide bonds on tar-et enzymes/proteins that perform reduction reactions. TRX arehen reduced at the expense of NADPH via an NADPH-dependenthioredoxin reductase (NTR) in bacteria [6]. The situation is quiteimilar in animal/mammalian cells with NADPH as the primarylectron provider and NTR as a signal transmitter. However, as ani-al cells evolved multiple compartments, the reducing systems
ecame more diversified with the existence of both cytosolic anditochondrial systems. In animal cell cytosolic and mitochondrialTR components are modified by a C-terminal extension pos-
essing a selenocysteine. All animal NTRs have similar propertiesorming a homodimer with FAD and a redox active selenosul-de together with a disulphide in each subunit. One complicationith mammalian thioredoxin reductase genes is their capacity to
ndergo differential exon splicing leading to the generation of mul-iple forms of the enzyme in both compartments [7]. Conversely,lants are even more complicated than animal or fungal cells, pos-essing an extra compartment, the chloroplast which performsxygenic photosynthesis. Accordingly, currently four different TRXeduction pathways have been described in plant cells [8]. Two ofhose are similar to those found in animal cells i.e. mitochondrialnd cytosolic NTR-based systems. There is though a major differ-nce; NTRs in cytosolic and mitochondrial land plant are of therokaryotic type, containing the disulphide and FAD but lacking the-terminal extension and the selenocysteine residue. Selenocys-eines are not present in photosynthetic eukaryotes after the greenlgae lineage [9]. The two other TRX reduction systems are locatedn the chloroplast; one of these is called NTRC and it is actually aariant of the prokaryotic NTR with a built-in TRX domain in the-terminus [10–12]. The other is the ferredoxin-thioredoxin sys-em consisting of a redox cascade involving the photosystems, andhe following stromal components: ferredoxin (FDX), ferredoxin-hioredoxin reductase (FTR) and chloroplast-located TRXs [13,14].
TR is a heterodimer with a so called “variable subunit”, with a likelyrotective task [15], and a somewhat larger and more conservedatalytic subunit. The catalytic subunit bears both a [4Fe-4S] centre. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 10
and the catalytic disulphide. Those four reduction pathways in landplants are schematized in Fig. 1, providing also a list of chloroplasttarget enzymes for which molecular structural details are availableand which will be discussed later in more detail in this review.
1.2. The complex TRX gene family of plants
The diversification of the TRX reductions modes is not the onlysignificant difference between plant cells and other organisms. Oneadditional indicator is the number of TRX genes. Strictly speaking,a TRX should bear a WCGPC redox-active site although WCPPC sig-nature can be quite frequent especially in plants, but other moredeviant motifs can also be found [16,17]. Anyway, when consid-ering only the classical (and more numerous) WCGPC containingTRXs, bacterial and animal species contain but only few of those(1–3 sequences on average) [2,18], while land plants have a largecontent of those genes (nearly 20 on average) with an unequal dis-tribution between compartments with most in chloroplasts (f, m, x,y, z isoforms) or cytosol (h isoforms), whilst mitochondria contain hand o isoforms [3,5,19]. The expansion of the diversity of TRX genesin plants is not unique to this antioxidant protein. It is even moreenhanced for its cousin protein glutaredoxin (GRX) with more than40 genes of that category in plant genomes, some of them unique toland plants [20]. The reasons for this remarkable enhanced diver-sity of proteins of the broad redoxin lineage will be discussed ina later section. It is, however, of interest to note that there areconnections between the TRX and GRX systems and at least oneplant TRX is reduced by a GRX system. It can also be palmitoylatedand has the capacity to move from cell to cell [21,22]. Likewise,there are proteins clearly more related phylogenetically to TRXsbut with non-canonical active sites which in reality are reduced byglutathione (GSH) and behave as GRXs [17]. The complex interplaybetween the TRX and GRX systems has been described in a num-ber of recent reviews [3,4,23,24] and recent paper discuss how FTRand NTRC systems can back up one another and, as a consequence,the analysis of mutants of the regulatory cascades needs to be donewith circumspection [25–27].
2. Why did redox regulation via dithiol reduction appear?
As noted above, two indicators reflect the expansion of theredox networks in land plants, i.e. the diversification of the reduc-tion routes and the expansion and diversification of the TRX andGRX genes. Actually, a third criterion can be added to that list: theappearance of enzyme target sequences containing critical disul-
phides needed for ON/OFF regulation in chloroplasts. During thedark phase, plastid enzymes are generally under an oxidized stateand turn out inactive whereas in the light phase, enzymes arerather in reduced form and are active. In contrast, plastid glucose
D.D. Gütle et al. / Plant Science 255 (2017) 1–11 3
F localizN in-thio−
6Ww
2
pasacrtaaesi(staoaroasop
ig. 1. Schematic representation of the reduction pathways found in land plants
ADPH-thioredoxin reductase; TRX − thioredoxin; Fd − ferredoxin; FTR − ferredox glutaredoxin; PS − photosystem. Pink regions of NTRC indicate TRX domain.
-phosphate dehydrogenase (G6PDH), behaves the opposite way.ith this observation in mind, two questions immediately arise:hy and when did redox regulation appear?
.1. Oxidative stress increases during evolution of higher plants
In response to the why, essentially two hypotheses have beenut forward. The first one is based on the coincidence of thedvent of redox regulatory pathways with oxygenic photosynthe-is. It is believed that oxygen began to accumulate on the earthfter the elaboration of oxygenic photosynthesis based on theapacity of organisms able to cleave water molecules with theelease of protons, electrons and molecular oxygen. Oxygenic pho-osynthesis is believed to have been initiated in cyanobacteriand further developed in algae and land plants. In this scenario,s the oxygen content gradually increased on the still primitivearth, various reactive oxygen species (ROS) have been and aretill being produced (singlet oxygen, hydroxyl radicals, superox-de ions, hydrogen peroxide) as well as reactive nitrogen speciesRNS). Several of ROS and/or RNS compounds are used in cellularignal transduction pathways, but when their levels become uncon-rolled they can damage macromolecules, including lipids, nucleiccids and proteins. The proteins are mostly affected at the levelf methionine and cysteine, the two of the most reactive aminocids. As a consequence, all living cells evolved detoxification andepair systems. The detoxification systems include catalase in per-xisomes (destroys H2O2), ascorbate and glutathione peroxidases
nd superoxide dismutases in various sub-compartments (reduceuperoxide ions) but also some very abundant enzymes called PRXr thiol-peroxidases. The PRX are present in nearly all cell com-artments and they can reduce simple peroxides as H2O2 but alsoed in the three sub-compartments mitochondria, chloroplast and cytosol. NTR −redoxin reductase; GR − glutathione reductase; GSH − reduced gluthathione; GRX
more complex ones as lipid hydroperoxides (compounds that cata-lase cannot reduce) [28]. The catalytic cysteine of PRXs becomesoxidized to a sulfenic acid In the catalytic cycle that is regeneratedvia either the TRX or the GRX systems [29].
Methionine sulfoxide reductases that can reduce methioninesulfoxide back to methionine and thus repair damaged proteins arealso essential in the repair systems. As there are two stereoisomersof methionine sulfoxide (R and S), there are accordingly two dif-ferent types of methionine sulfoxide reductases (MsrA and MsrB).MsrA and MsrB, as PRXs, generate a sulfenic acid (R-SOH) moi-ety in their catalytic cycle which is reduced back to a thiol viaeither the TRX or the GRX system [30–32]. As explained before,the TRX and GRX systems of higher plants have the greatest knowndiversity in nature and it is certainly a reasonable hypothesis topropose that TRX and GRX and their regeneration systems are verymuch involved in overcoming oxidative stress in plants contain-ing chloroplasts i.e. with an additional (and very intense) site ofROS generation. Still it is puzzling to observe that cyanobacteria,which are also able to perform oxygenic photosynthesis lack severalcharacteristics elucidated in algae and land plants and in particu-lar they do not show this expansion of the TRX and GRX genesalthough that if one considers cyanobacterial cells as the equiva-lent of a chloroplast, the difference is somewhat reduced [18]. Thisleads to postulate that either cyanobacteria might have lower oxy-gen yields and thus generate reduced levels of ROS or that theypossess physical/biochemical characteristics that limit ROS diffu-sion and damage. There is no easy answer to this hypothesis, as
comparison of the quantum yield of cyanobacteria and green algaevaries depending on the methodology used, and it has been hypoth-esized that their O2 evolving capacities are not greatly different. Butthere might be large differences in the rates of the O2 consuming
4 t Scie
Mat[icdce
2
ahfawrctowptpAslcis
Tmbmtthpopn(ltceTtc
3r
3
ogphtt
prominently in the Benson-Calvin cycle [51,52]. Initially, threeenzymes of the Benson-Calvin cycle (i.e. phosphoribulokinase(PRK), ribulose 1,5-bisphosphate carboxylase/oxygenase (RubisCO)and sedoheptulose-1,7-bisphophatase (SBPase)) were thought to
land colonization
vascular system
monocotsdicots
mosses
green algae
red algae
cyanobacteria
plantae
FBPase, SBPase,G6PDH
NADP-MDH C-term., ATP synth. γ subunit,GADPH G ap B
NADP-MDH N -term.
PRK*, CP12
Arabidopsis tha liana
Oryza sat iva
Physcomitre lla patens
Chlamydomonas reinha rdtii
Cyanidio schyzon mer olae
Synechoc ystis sp. PCC 6803
(loss of GADP H Gap B)
Fig. 2. Evolutionary appearance of the redox sensitive cysteine residues in redox-regulatory PRK (phosphoribulokinase), CP12, FBPase (fructose-1,6-bisphosphatase),SBPase (sedoheptulose-1,7-bisphosphatase), G6PDH (glucose-6-phosphate dehy-drogenase), NADP-MDH (NADP-malate dehydrogenase), ATP synthase � subunit and
D.D. Gütle et al. / Plan
ehler reaction (reduction of molecular oxygen (O2) to superoxidenion (O2
−) by donation of an electron in photosystem I) leading tohe generation of H2O2 after conversion by superoxide dismutase33]. Cyanobacteria are much more sensitive to hydrogen perox-de than green algae. This physiological difference has been used tolean lakes polluted by cyanobacterial blooms [34,35]. There is littleoubt that this is due at least partially to their reduced anti-oxidantapacities. Also to be noted is that cyanobacteria differ notably fromukaryotes in their PRX content [36].
.2. Functional modifications of target proteins
A second hypothesis that has been put forward is limited to couple enzymes located in the chloroplast. This compartmentosts both a fructose-1,6-bisphosphatase (FBPase) able to cleave
ructose-1,6-bisphosphate and to generate fructose-6-phosphatend inorganic phosphate and also a phosphofructokinase (PFK)hich does the reverse reaction at the expense of ATP. It has been
easoned that if both enzymes functioned simultaneously in thisompartment, it would lead to a futile cycle with useless consump-ion of ATP. So one hypothesis put forward was that FBPase is activenly in the light phase (through reduction through the FTR system)hile conversely PFK operates only in the dark phase in this com-
artment [37]. While this argument has some strength it ignoreshe fact that there are also FBPase and PFK enzymes in the cytosol oflants and they are apparently not subject to this type of regulation.lternatively, metabolic channelling and the formation of enzymeuper complexes might be another way that circumvent the prob-em of futile cycles [38]. Another example of a potential futile cycleould be the sequence F6P->G6P->->->R5P->Ru5P) which is foundn the oxidative pentose pathway and in reverse in the Calvin Ben-on cycle (Ru5P->->->->F6P->G6P).
There are very large lists of possible molecular targets of theRX and GRX systems in various sub-cellular plant compartmentsostly from proteomic data [39–42]. In general, the proteins have
een isolated following a similar procedure involving the replace-ent of the second cysteine of TRX (or GRX) into a serine, trapping
he targets as disulphides, releasing them with a reductant andhen identifying them by mass spectrometry analysis of peptides. Itas become immediately noticeable that some, if not many, of theotential targets do not contain two cysteines, sometimes a singlene is found and sometimes none, casting some doubt on the actualhysiological nature of the latter targets. Still reasonable expla-ations have been provided for single cysteine containing targetsreviewed in [4]) that can be modified by glutathionylation, nitrosy-ation and a variety of other ROS-induced modifications. In additiono the proteomics approach, the best biochemically and structurallyharacterized redox-sensitive enzymes have been discovered inarlier studies using more traditional biochemical approaches.hey essentially function in primary carbon metabolism. Still pro-eomics has uncovered that TRXs and GRXs extend their regulationapacities to many other metabolic systems.
. When and how did redox regulation via dithioleduction appear?
.1. Occurrence of TRX reductases
As discussed above, a first event linked to the developmentf redox regulation was the diversification of TRX and GRXenes [18–20]. Obviously, this happened after the transition from
rokaryotes to eukaryotes and, as pointed out before, it mayave evolved to deal with an increase in ROS generation. Whilehe cytosolic and mitochondrial enzymes are of the prokaryoticype (short NTR), the chloroplast NTRC are uniquely linked tonce 255 (2017) 1–11
oxygenic photosynthetic organisms, being present in land plantsbut also in some cyanobacteria [10,43]. NTRCs also occur in non-photosynthetic tissues such as in plant root plastids, but a homologof this enzyme is also present in non-photosynthetic prokaryotessuch as Mycobacterium leprae [11,44]. The occurrence of fusions ofthat type is actually quite common in pathogenic bacteria, as shownfor a PRX-GRX fusion in Neisseria meningitidis [45], or even in a redalga (Gracilaria gracilis) for a GRX-GRX-Msr fusion [46]. Still con-cerning the reductases, FTR has long been thought to be absolutelyrestricted to photosynthetic organisms, but genes with similarsequences occur in genomes of bacteria and archaea. The protein isalso present in Arabidopsis roots as well as in non-photosyntheticplant tissues as wheat endosperm [1,47–49]. Its function seems tobe related to methane metabolism [50] in methanogenic archaea.Interestingly, in some prokaryotic organisms there are fusionsbetween GRX and the catalytic subunit of FTR, further suggestingthat the enzyme might possess additional unknown functions [8].
3.2. Integration of redox sensitive cysteine residues in targetprotein sequences
A good deal of information concerning the ontogenesis of theredox regulatory systems can be derived from the study of thetarget regulatory enzymes. As pointed out earlier, many of thetarget enzymes are involved in carbon metabolism and most
GADPH (glyceraldehyde 3-phosphate dehydrogenase) in the organisms Synechocys-tis sp PCC 6803 (cyanobacteria), Cyanidioschyzon merolae (red algae), Chlamydomonasreinhardtii (green algae), Physcomitrella patens (moss), Oryza sativa and Arabidopsisthaliana. The GADPH Gap B got lost in losses. *One cysteine residue got lost in redalgae.
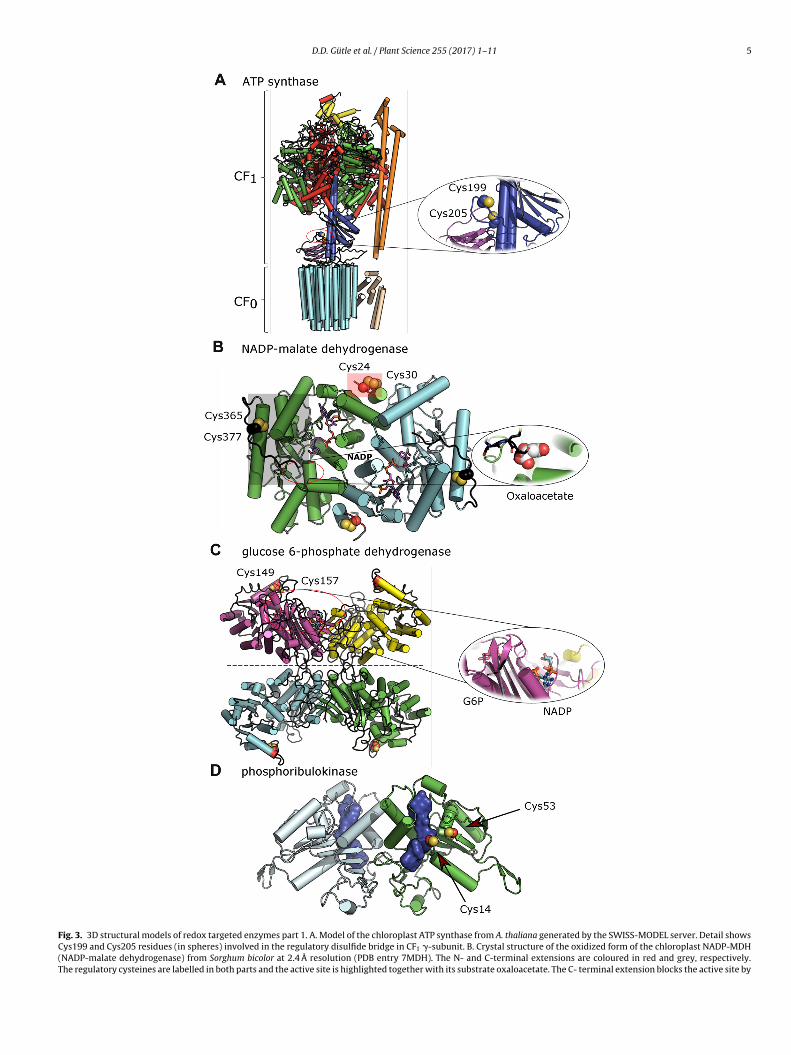
D.D. Gütle et al. / Plant Science 255 (2017) 1–11 5
Fig. 3. 3D structural models of redox targeted enzymes part 1. A. Model of the chloroplast ATP synthase from A. thaliana generated by the SWISS-MODEL server. Detail showsCys199 and Cys205 residues (in spheres) involved in the regulatory disulfide bridge in CF1 �-subunit. B. Crystal structure of the oxidized form of the chloroplast NADP-MDH(NADP-malate dehydrogenase) from Sorghum bicolor at 2.4 Å resolution (PDB entry 7MDH). The N- and C-terminal extensions are coloured in red and grey, respectively.The regulatory cysteines are labelled in both parts and the active site is highlighted together with its substrate oxaloacetate. The C- terminal extension blocks the active site by

6 t Scie
btv[uh(apfMg(1c
sotbtfmsaCeto1
essot
ir−es
tRaa[pprwBgha
oNGrGoblr
D.D. Gütle et al. / Plan
e unique to this cycle. But genome sequencing has shown thathis was not the case, as representatives of those genes occur inarious microorganisms and especially in bacteria and archaea47,53]. In order to evaluate at which point in evolution redox reg-latory cysteines were introduced into the target sequences weave run sequence comparisons that included a cyanobacteriumSynechocystis sp. PCC 6803), a red alga (Cyanidioschyzon merolae),
green alga (Chlamydomonas reinhardtii), a moss (Physcomitrellaatens) and two angiosperms (Arabidopsis thaliana and Oryza sativa)or the following enzymes: NADP-malate dehydrogenase (NADP-
DH), PRK, FBPase, SBPase, the �-subunit of CF1-ATPase, G6PDH,lyceraldehyde-3-phosphate dehydrogenase (GAPDH) and CP12see Supplementary material Fig. S1 in the online version at DOI: 10.016/j.plantsci.2016.11.003). These analyses led to the followingonclusions:
i) Redox regulation has been achieved by single cysteine sub-titution in different routes (G6PDH, PRK, SBPase), or by insertionf redox regulatory modules (FBPase, CF1-ATPase) or even by addi-ions of regulatory extensions either at the N- or C-termini or onoth sides of the sequence (NADP-MDH, GAPDH). Two versions ofhe GAPDH sequence exist: an A4 enzyme, which recruits CP12or its regulation, and an A2B2 form, possessing a built-in C ter-
inal extension that includes the regulatory cysteines in the Bubunit [54]. CP12 is actually a homolog of the extension with thedvantage as serving as a hub for recruiting super complexes of thealvin-Benson cycle. NADP-MDH is a very interesting example ofvolution in progress as the enzyme of C. rheinhardtii lacks the N-erminal regulatory sequence, but already possesses the C-terminalne (Fig. S1 in the online version at DOI: 10.1016/j.plantsci.2016.1.003).
ii) The appearance of redox properties on the regulatorynzymes occurred at different times of evolution. The variousteps at which the regulatory sequences were integrated areynthesized in Fig. 2. It is readily apparent that the integrationf redox properties did occur at different periods of evolu-ion.
iii) The comparison of FBPase and SBPase is very reveal-ng, showing that although the two sequences are clearlyelated, they nevertheless originate from very different ancestors
�-proteobacteria for FBPase and archaea for SBPase [53]. Asxplained below, it also shows that there are several solutions pos-ible to a given structural constraint.
iv) There are at least two examples which indicate that evolu-ion relied on trial and error/success experiments. One of those isubisCO activase for which there is one sequence which includes
regulatory disulphide but does not occur in all angiosperms (it ispparently present in A. thaliana but absent in tobacco and maize)55]. A second example is the redox regulation of C. reinhardtii phos-hoglycerate kinase which relies on two cysteines absent in landlant homologs [56]. Analysis of future evolutionary events willeveal whether these two examples are dead-ends of evolution orhether they will be later more universally adopted. For GAPDH
subunit the situation is actually quite complex as it is present in
reen algae (Characeae and Ostreococcus) [57,58] but the sequenceas apparently been lost in mosses and somehow maintained inngiosperms.ccupying the position of the substrate oxaloacetate (shown in spheres) from the X-ray cryADP molecule is coloured in purple. C. Model of chloroplast G6PDH (glucose 6-phosphate6PDH (45% identity, PDB entry 1QKI). The G6PDH can be observed as a dimer or a dim
egulatory cysteines are on position 149 and 157. Zoom of the active site with the G6P (g6PDH from Leuconostoc mesenteroides (PDB entry 1E7Y). D. Model of the reduced PRK (pf the PRK from Rhodobacter spheroides (24% identity, PDB entry 1A7J). The regulatory clue surface. As indicated elsewhere, the low conservation of the primary sequence and
ower confidence. The overall structures are shown in cartoon representation with �-heliesidues was calculated excluding transit sequences.
nce 255 (2017) 1–11
4. In response to which structural constraints did redoxregulation happen on molecular targets?
As a general rule, the generation of redox regulatory enzymesfrom non-regulatory forms is driven by 3D structural constraintsspecific to each enzyme. The goal is to shield the active site in theinactive form, preventing access of at least one of the substrates.Reduction of the regulatory disulphides is predicted to inducestructural rearrangements that result in an opening of the activesite, allowing catalysis. A very large panel of target enzymes hasbeen provided by proteomics studies and biochemistry. Geneticshas also opened the way to describing new functions in plants, suchas those linked to starch degradation or tetrapyrrole biosynthesis[55,59,60]. Nevertheless, these studies have not yielded structuralmodels that could be useful to our study. We have thus limitedthis section to those enzymes for which there are detailed struc-tures available, or for which we can build credible 3D models usingrelated available structures (same six enzymes as those used forphylogeny) (Figs. 3 and 4 ).
4.1. ATP synthase
The ATP synthase that produces ATP using the proton gradi-ent generated by the photosynthetic electron flow in the light, isa complex assembly of several subunits in which the CF1 extrinsiccatalytic domain is composed of three �- and �-subunits, the redoxregulatory �-subunit, one �- and �-subunits and b/b’-subunits(coloured in red, green, blue, yellow, purple and orange, respec-tively). The CF0 membrane domain is composed of a ring of 14c-subunits and a bundle of �-helices assigned to the �-subunit(coloured in cyan and pale yellow, respectively, only 12 are shownin our model based on mitochondrial F0) (Fig. 3A) [61]. ATP synthaseis mainly regulated by the FTR system [62,63], however, recentstudies postulate a regulation by NTRC especially under low lightconditions [64]. The regulatory site does not occur in cyanobacte-ria and red algae (Fig. S1 in the online version at DOI: 10.1016/j.plantsci.2016.11.003), but it is highly conserved in higher plants.CF0-CF1 ATP synthase is a wonderful model of a rotating enzymeconverting kinetic energy into a high energy molecule and the rotorpart contains the � subunit. Our model clearly shows that the redoxactive site between Cys 199 and Cys 205 is readily accessible forstromal components (e.g. TRX), being situated in a loop distant fromthe adenine nucleotide binding sites situated in the CF1 part of themolecule. How the reduction of the disulphide influences the cat-alytic activity of CF0-CF1 ATP synthase is unknown. Still, one canrather safely postulate that the formation of the disulphide mightbe sufficient to slow down the movement of the rotating part andthus the rate of catalysis. Conversely, its reduction could relax thestrain and improve catalysis. In connection with this possibility, anice artificial example of strain induced by disulphide bridge forma-tion is roGFP where a disulphide engineered between two adjacent
� strands of the � barrel constrains the protein and limits its flu-orescence. Reducing the protein with GSH and GRX releases thatconstraint and this probe can be used to monitor the intra-cellularGSH status [65].stal structure of the NADP-MDH from Aquaspirillum arcticum (PDB entry 1B8U).The dehydrogenase) from A. thaliana obtained by homology modeling from the humaner of dimers (separated by a dashed line) depending on specific conditions, the
lucose 6-phosphate) and NADP molecules obtained from the X-ray structure of thehosphoribulokinase) of A. thaliana. The dimer was obtained by homology modelingysteine residues are highlighted and the active site of each monomer is shown as
the presence of the large insertion between the cysteines result in a model withces and �-strands in cylinders and arrows, respectively. The numbering of cysteine
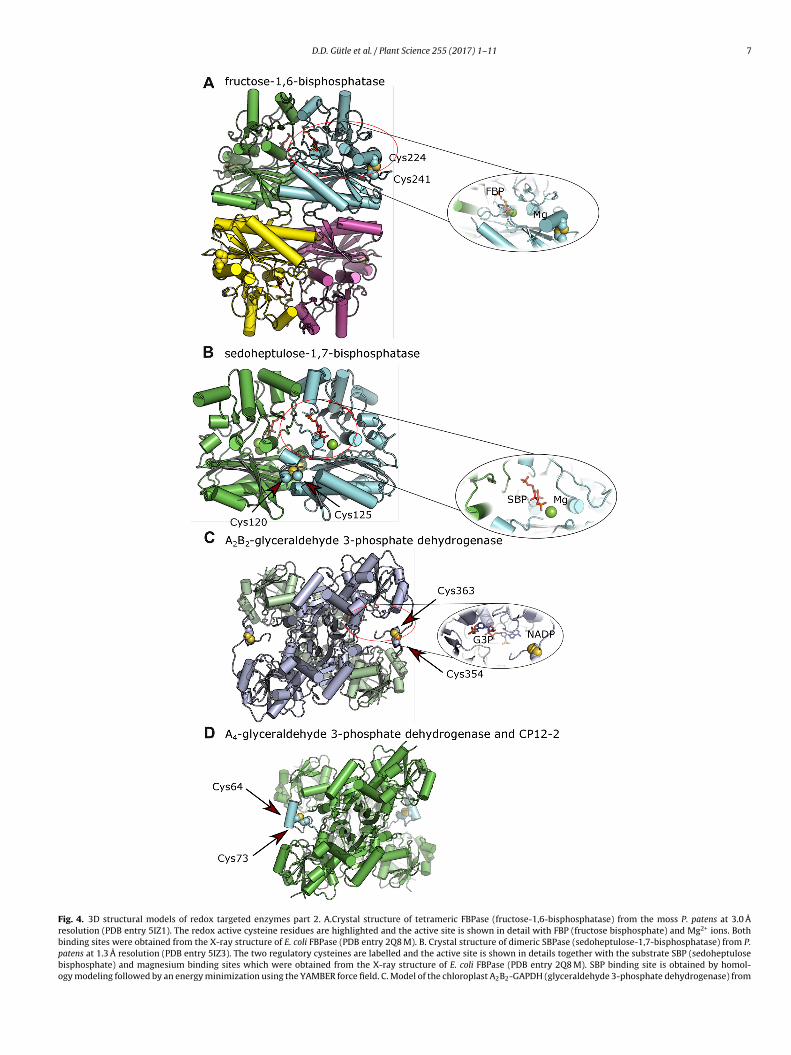
D.D. Gütle et al. / Plant Science 255 (2017) 1–11 7
Fig. 4. 3D structural models of redox targeted enzymes part 2. A.Crystal structure of tetrameric FBPase (fructose-1,6-bisphosphatase) from the moss P. patens at 3.0 Åresolution (PDB entry 5IZ1). The redox active cysteine residues are highlighted and the active site is shown in detail with FBP (fructose bisphosphate) and Mg2+ ions. Bothbinding sites were obtained from the X-ray structure of E. coli FBPase (PDB entry 2Q8 M). B. Crystal structure of dimeric SBPase (sedoheptulose-1,7-bisphosphatase) from P.patens at 1.3 Å resolution (PDB entry 5IZ3). The two regulatory cysteines are labelled and the active site is shown in details together with the substrate SBP (sedoheptulosebisphosphate) and magnesium binding sites which were obtained from the X-ray structure of E. coli FBPase (PDB entry 2Q8 M). SBP binding site is obtained by homol-ogy modeling followed by an energy minimization using the YAMBER force field. C. Model of the chloroplast A2B2-GAPDH (glyceraldehyde 3-phosphate dehydrogenase) from

8 t Scie
4
crtlposfmfTwoaiav
4
rlhhtmbiiumt
4s
lpioTeeutbthcmtsa
Acsprpr
D.D. Gütle et al. / Plan
.2. NADP-malate dehydrogenase
The chloroplast NADP-MDH (Fig. 3B) is an unusual case con-erning redox regulation, as it contains two or more redox cysteineesidue pairs per monomer in vascular plants. The NADP-MDH pro-ein sequence harbours N- and C-terminal sequence extensions inand plants. The enzyme is a head-to-tail-homodimer and the disul-hides are located at the interface of the two subunits (N terminalne) or at the outside corner of each subunit (C-terminal one). Thepacing between the cysteines involved in the disulphides includesour N-terminal and eleven C-terminal residues. Both disulphides
ust be reduced to obtain a fully activated enzyme, but the con-ormational changes occurring upon reduction are likely different.he N-terminal part influences the catalytic rate. The C-terminus,hich mimics the substrate oxaloacetate, fills the active site in the
xidized state, preventing access of oxaloacetate to this part of thective site, and thus catalysis cannot occur. Both extensions occurn algal protein sequences, but only the cysteines in the C-terminusre present and are redox-sensitive and required for enzyme acti-ation [66].
.3. Glucose 6-phosphate dehydrogenase
We have modelled the structure of chloroplast G6PDH of closelyelated sequences (Fig. 3C). This enzyme, which catalyzes the rate-imiting step of the oxidative pentose phosphate pathway, is aomo tetramer and each subunit contains a regulatory disulphide,aving the cysteines separated by seven amino-acids. Here too,he disulphides are surface-exposed at the outside corner of each
onomer and presumably easily accessible for reduction. As notedefore, the redox regulation of G6PDH is unique. The enzyme is
n the oxidized state in the dark and inactive in the reduced staten the light [67–69]. As observed for the other enzymes, the reg-latory disulphide is rather remote from the G6P binding site andolecular reconfiguration is likely to be necessary for modifying
he catalytic properties of the enzyme [67].
.4. Fructose-1,6-bisphosphatase andedoheptulose-1,7-bisphosphatase
FBPase and SBPase are two related enzymes although theyikely originate from different backgrounds: FBPase from �-roteobacteria, SBPase from archaea [53]. At the subunit level there
s extensive homology between the two enzymes, but FBPase isrganized as a tetramer (Fig. 4A) and SBPase (Fig. 4B) as a dimer.he tetrameric FBPase is actually composed of two pairs of dimers,ach pair quite similar to one SBPase molecule, so the homologyxtends at least partially up to the quaternary structure. The reg-latory disulphides of both enzymes are located on the surface ofhe protein, away from the active site, and probably influence theinding of the co-factor Mg2+. Both enzymes share high similari-ies in their catalytic sites, their substrates being very similar (SBPas one additional carbon compared to FBP). The biggest differenceoncerning redox regulation is the position of the disulphides in the
onomers. The redox sensitive disulphide of FBPase is located onhe outer edges of the protein and the cysteines are in an insertioneparated by 16 to 19 amino acids (Fig. S1 in the online versiont DOI: 10.1016/j.plantsci.2016.11.003), whereas for SBPase it is
. thaliana. The tetrameric model was obtained by homology modeling from the photosynontains the regulatory cysteine residues highlighted. The G3P (glyceraldehyde 3-phosphtructure from Geobacillus stearothermophilus (PDB entry 3CM3). D. Crystal structure of trotein CP12-2 (PDB entry 3QV1). The subunits A of the GAPDH tetramer and CP12-2
esidues) are missing in the X-ray structure showing a highly disordered N-terminal partrobably mimics the regulatory disulfide of A2B2-GAPDH. The overall structures are showespectively. The numbering of cysteine residues was calculated excluding transit sequen
nce 255 (2017) 1–11
more hidden between the two monomers and less accessible forthe reductant TRX molecule (the spacing between the cysteines isfour residues). In contrast to earlier work carried out with proteinsisolated directly from plants, the availability of recombinant pro-teins has allowed to show that Trx m can also activate FBPase [70].The more recent comparison of the redox properties of FBPase andSBPase from P. patens has confirmed and extended this observationto SBPase. Nevertheless, Trx f was still found to be the best reductantand reduction was facilitated for FBPase with the more open redoxregulatory sites (those in SBPase are sandwiched between the twosubunits). This observation together with a tighter redox control ofthe SBPase enzyme in vitro leads to the assumption that the loca-tion of redox-sensitive residues differentially impacts the qualityand extent of the redox control [53]. In terms of molecular move-ment following reduction, the comparison of a redox-insensitiveFBPase from pig kidney to the oxidized structure of the chloro-plast pea enzyme suggests that the animal FBPase possesses apseudo-reduced conformation [71]. Comparing the oxidized andpseudo-reduced structures has then lead us to propose that in thepea enzyme, the reduction of the disulphide is likely to induce areversal of the orientation of two �-strands with the shifting of aside chain glutamate that leads to better Mg2+ binding [69].
4.5. Phosporibulokinase
There is no current available structure of plant PRK, but we couldmodel this enzyme from the related structure of its Rhodobactersphaeroides homolog (Fig. 3D). As the sequence homology is low,this model should be considered with caution. In our model, theprotein is a homodimer and the regulatory disulphide, the cysteinesof which are separated by 38 amino acids, is surface-exposed. Notethat these cysteines are also present in the cyanobacterial PRKs, butthe plant sequence has had an insertion of 17 amino acids duringevolution. The bacterial enzyme, on which we have based the modeldoes not contain the regulatory cysteines and is redox insensitive.In the model, the sulfur atom of Cys 14 is close to the binding site ofribulose-5-phosphate. Earlier biochemical studies have proposedthat both regulatory cysteines of PRK were also required for cataly-sis and close to the nucleotide binding site with a more marked rolefor Cys 53 [72,73]. More recent papers have described the redox reg-ulation of PRK in complex and the shape of the molecule via SAXS[74,75].
4.6. Glyceraldehyde 3-phosphate dehydrogenase
The next enzyme of this series is GAPDH, which is presentin super complexes with PRK under oxidized conditions togetherwith the regulatory protein CP12, whereas the activation processrequires the TRX proteins. The photosynthetic GADPH occurs in twodifferent subunits, Gap A and Gap B, which share high sequencesimilarity, but with an extension of the C-terminus at the Gap Bsubunit that harbours the redox regulatory cysteine residues andthe homology with the CP 12 probably evolved from a fusion ofGADPH A subunit with CP12 protein. So far the Gap B subunit is
restricted to higher plants [54] (Fig. S1 in the online version atDOI: 10.1016/j.plantsci.2016.11.003). The enzyme appears in twoconformations, either as a homotetramer A4, without redox sen-sitivity, or as a heterotetramer with two A and two B subunitsthetic GAPDH from Spinacia oleracea (85% identity, PDB entry 2PKQ). Only subunit Bate) and NADP binding sites, shown in detail, were obtained from the GAPDH X-rayhe binary complex of photosynthetic A4-GAPDH with the intrinsically disorderedare coloured in green and cyan, respectively. The first 58 residues in CP12-2 (79. In the folded C-terminal part, Cys73 establishes a disulfide bridge with Cys64 andn in cartoon representation with �-helices and �-strands in cylinders and arrows,ces.

t Scie
(wpsPC[sSgt3t[
5s
stsliFtssso2ltp
lAbhttttiteepDfttaepltaltOusRr
D.D. Gütle et al. / Plan
A2B2). The latter is very sensitive to redox regulation through TRXith an auto-inhibitory process (similar to NADP-MDH) by occu-
ying the active site with the C-terminus extension in the oxidizedtate. The A4 GADPH is kept inactive via interaction with CP12 andRK and after the reductive activation of the two cysteine pairs ofP12 the super complex dissociates and releases active A4 GADPH76,77]. Our structural model of A2B2 GADPH (Fig. 4C) was con-tructed using the A. thaliana sequence and the crystal structure ofpinacia oleracea as a basis. The two binding sites of the substratelyceraldehyde 3-phosphate and NADP were included accordinglyo the structure from Geobacillus stearothermophilus (PDB entryCM3). The crystal structure of the photosynthetic A4 homote-ramer was solved in complex with two CP12 proteins (Fig. 4D)76].
. Do the regulatory sequences have additional roles inignalling?
The structural studies indicate that, as a general rule, the redox-ensitive disulphide bonds of the target enzymes are distal fromhe active site and present in surface-exposed loops. There is noequence similarity whatsoever in these regulatory parts and itooks as if each of the regulatory enzymes was modified dur-ng evolution in response to its own structural constraints. TheBPase/SBPase story [53] indicates that there are several solutionso solve a similar 3D constraint. It is likely, that the ubiquitousurface-exposed position is strategic for making the regulatoryites accessible to TRX. However, this positioning renders theseequences potential targets of endoproteases. There are examplesf endoproteases that are able to cleave before or after 16 of the0 amino acids. We could not find adequate endoproteases in the
iterature for C, N, P and Q, with proline having a strongly nega-ive effect when adjacent to one of the permissive amino acids foreptide hydrolysis.
It is highly puzzling that in cowpea the peptide inceptin was iso-ated and identified as a fragment of the � subunit of the CF0-CF1TP synthase [78]. Inceptin is an eleven amino-acid disulphide-ridged peptide that is released by the plant after attack by insecterbivores such as Spodoptera frugiferda [79]. Inceptin is found inhe oral secretions of the pest and it induces ethylene produc-ion by the plant at concentrations as low as 1 fmol per leaf. Inurn, the plant produces ethylene which triggers the production ofhe defence-related hormones jasmonic acid and salicylic acid. Its thus believed that when the worm starts feeding on the plant,he release of plant peptides acts as a defence signal. Amazinglynough, inceptin has the sequence ICDINGVCVDA, which is nearlyxactly the redox regulatory insertion in the � subunit of the chloro-last CF0-CF1 ATP synthase (see Fig. S1 in the online version atOI: 10.1016/j.plantsci.2016.11.003). The surrounding sequence
rom which inceptin is generated is GE/ICDVNGVCVDA/AE, andhus one can imagine that a combination of a glutamyl endopep-idase (that cuts after E) and of an elastase-like peptidase (can cutfter A) is sufficient to generate this peptide. An interesting hypoth-sis concerning inceptin is that it is probably easily cut out by theroteases because it is surface-exposed, but also possibly stabi-
ized by its disulphide bond thus improving its potential as a signalransmitter. Our structural study indicates that these propertiesre certainly shared by all redox-regulated enzymes. In particu-ar, the regulatory insertion of FBPase could be easily released byhe same combination of a glutamyl endopeptidase and elastase.ther simple combinations of various endoproteases or even the
se of a single endoprotease could easily release the regulatoryequences of SBPase, GAPDH, NADP-MDH, PRK, CP12, G6PDH andubisCO activase. Whether those potential fragments are indeedeleased and could serve as signalling molecules in plant defence,nce 255 (2017) 1–11 9
as demonstrated for inceptin, could be an exciting research area forthe future.
6. Potential new avenues of research in the redox field
A number of problems remain to be elucidated concerning theregulatory enzymes and their interaction with TRXs, but also aboutthe multiplicity of reduction systems. We have pointed out thatthe area of recognition on the target enzymes is enriched in acidicresidues with resulting negatively charged electrostatic surfaces(Fig. S2A–H in the online version at DOI: 10.1016/j.plantsci.2016.11.003).Interestingly, the most efficient TRXs (especially the f type)have a compatible positively charged surface (Fig. S2I in the onlineversion at DOI: 10.1016/j.plantsci.2016.11.003). The interactionwould then be governed at least in part by electrostatic interactions.The active site of TRX is still primarily hydrophobic and these inter-actions should also play an essential role [80]. A number of recentpapers have dealt with TRX specificity although in general they donot address the problem at the molecular structural level [81,82]. Tobetter understand the target enzyme/TRX interaction it is desirableto generate stable heterodimers by replacing backup cysteines byserines and thus generating stable mixed disulphides. This technol-ogy has been used to generate ternary complexes of FDX-FTR-TRX,the structure of which could then be determined by X ray crystal-lography [13]. Those protein-protein interactions are essential forgoverning the reduction of enzymes soluble in the stromal phase ofthe chloroplasts, a location where the biomolecules could be signif-icantly distant from the reductant TRX. The problem is not as crucialin membrane-bound electron transfer chains where the positionsof all components are well defined. It has already been demon-strated that FDX recognizes its protein partners (FTR, FNR) via acombination of charge and hydrophobic recognition [13,83,84].This could possibly serve as an earlier example of what the TRXtarget enzyme interaction could be. Protein-protein interactionscan also be tested using fluorescence methods, isothermal titrationcalorimetry and/or surface plasmon resonance to obtain bindingparameters. The reactivity between a given TRX and its target canalso be described/predicted in terms of redox potentials. In generalTRXs possess disulfides with lower redox potentials than those ofthe targets making the reaction thermodynamically favourable. Anabundant literature dealing with those aspects has already beenreviewed in [1–6].
We have also pointed out to the need for molecular movementsin the redox-regulated enzymes for reaching their active confor-mation. At this point, the only report dealing with that aspect isan NMR study showing modifications of resonances related to theC-terminus of NADP-MDH after reduction [85]. Given the size of theproteins studied, it remains to be seen whether this very excitingapproach could be applied to other enzymes or whether this was alucky draw (which has nevertheless required a lot of perseverancein this case). Another possibility would be to try to solve both oxi-dized and reduced structures of the target enzymes, and possiblyusing anaerobiosis for elucidating the reduced forms.
The origins of the regulatory sequences present as insertionsor additions are also of evolutionary interest. Do they result fromslow evolution processes or are these pieces of DNA borrowedfrom other sequences? This is very difficult to answer because thesequence additions/insertions are short and quite variable. Never-theless, when BLASTing the regulatory sequence of the � subunit ofCF0-CF1 ATPase, one obtains a hit with 50% identity to a seeminglyunrelated chloroplast protein, the 30S ribosomal protein S13 from
Porphyridium purpureum (see below).CF1 gamma ATPase 236 DGKCVDAADDEIFKL 250Conserved residues D KC D DDE+ K+30 S rib protein DTKCKDLQDDEVVKI 56

1 t Scie
tcsmmg
dpIbk
A
Rptm
R
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
0 D.D. Gütle et al. / Plan
This sequence is not totally coincident with the regulatory inser-ion, but it overlaps it partially containing in particular one of theysteines. Of course, given the short length of the sequence, it mighttill be coincidental, but if other examples of that type arise, then itay well be that the genes coding for the redox regulated targetsay also be a composite of sequences borrowed elsewhere in the
enome.Another area that warrants interest is to understand why redun-
ant TRX reduction systems are present in plant cells (five differentossibilities when including the GSH GRX-dependent systems).
n fact, several mutants of NTR A, B and C and FTR have alreadyeen generated, sometimes in combination but so far no completenockout of FTR has been reported.
cknowledgements
This work was supported by a grant from by the French Nationalesearch Agency (ANR) as part of the “Investissements d’Avenir”rogram ANR-11-LABX-0002-01, Lab of Excellence ARBRE, and byhe Excellence Initiative of the German Federal and State Govern-
ents (EXC 294).
eferences
[1] M. Balsera, E. Uberegui, P. Schürmann, B.B. Buchanan, Evolutionarydevelopment of redox regulation in chloroplasts, Antioxid. Redox Signal. 21(2014) 1327–1355.
[2] J. Lu, A. Holmgren, The thioredoxin antioxidant system, Free Radic. Biol. Med.66 (2014) 75–87.
[3] Y. Meyer, C. Belin, V. Delorme-Hinoux, J.-P. Reichheld, C. Riondet, Thioredoxinand glutaredoxin systems in plants: molecular mechanisms, crosstalks, andfunctional significance, Antioxid. Redox Signal. 17 (2012) 1124–1160.
[4] L. Michelet, et al., Redox regulation of the Calvin-Benson cycle: something old,something new, Front. Plant Sci. 4 (2013) 470.
[5] P. Schurmann, J.-P. Jacquot, Plant thioredoxin systems revisited, Annu. Rev.Plant Physiol. Plant Mol. Biol. 51 (2000) 371–400.
[6] J. Kuriyan, et al., Convergent evolution of similar function in two structurallydivergent enzymes, Nature 352 (1991) 172–174.
[7] A.A. Turanov, D. Su, V.N. Gladyshev, Characterization of alternative cytosolicforms and cellular targets of mouse mitochondrial thioredoxin reductase, J.Biol. Chem. 281 (2006) 22953–22963.
[8] J.-P. Jacquot, H. Eklund, N. Rouhier, P. Schürmann, Structural and evolutionaryaspects of thioredoxin reductases in photosynthetic organisms, Trends PlantSci. 14 (2009) 336–343.
[9] S. Dai, M. Saarinen, S. Ramaswamy, Y. Meyer, J.P. Jacquot, H. Eklund, Crystalstructure of Arabidopsis thaliana NADPH dependent thioredoxin reductase at2.5 A resolution, J. Mol. Biol. 264 (1996) 1044–1057.
10] P. Bernal-Bayard, M. Hervás, F.J. Cejudo, J.A. Navarro, Electron transferpathways and dynamics of chloroplast NADPH-dependent thioredoxinreductase C (NTRC), J. Biol. Chem. 287 (2012) 33865–33872.
11] K. Kirchsteiger, J. Ferrández, M.B. Pascual, M. González, F.J. Cejudo, NADPHthioredoxin reductase C is localized in plastids of photosynthetic andnonphotosynthetic tissues and is involved in lateral root formation inArabidopsis, Plant Cell. 24 (2012) 1534–1548.
12] A.J. Serrato, J.M. Pérez-Ruiz, M.C. Spínola, F.J. Cejudo, A Novel NADPHthioredoxin reductase localized in the chloroplast, which deficiency causeshypersensitivity to abiotic stress in Arabidopsis thaliana, J. Biol. Chem. 279(2004) 43821–43827.
13] S. Dai, C. Schwendtmayer, P. Schürmann, S. Ramaswamy, H. Eklund, Redoxsignaling in chloroplasts: cleavage of disulfides by an iron-sulfur cluster,Science 287 (2000) 655–658.
14] S. Dai, et al., Structural snapshots along the reaction pathway offerredoxin-thioredoxin reductase, Nature 448 (2007) 92–96.
15] E. Keryer, V. Collin, D. Lavergne, S. Lemaire, E. Issakidis-Bourguet,Characterization of Arabidopsis mutants for the variable subunit offerredoxin:thioredoxin reductase, Photosynth. Res. 79 (2004) 265–274.
16] N. Coudevylle, A. Thureau, C. Hemmerlin, E. Gelhaye, J.-P. Jacquot, M.-T. Cung,Solution structure of a natural CPPC active site variant, the reduced form ofthioredoxin h1 from poplar, Biochemistry (Mosc.) 44 (2005) 2001–2008.
17] K. Chibani, et al., Atypical thioredoxins in poplar: the glutathione-dependentthioredoxin-like 2.1 supports the activity of target enzymes possessing asingle redox active cysteine, Plant Physiol. 159 (2012) 592–605.
18] M.E. Pérez-Pérez, E. Martín-Figueroa, F.J. Florencio, Photosynthetic regulation
of the cyanobacterium Synechocystis sp. PCC 6803 thioredoxin system andfunctional analysis of TrxB (Trx x) and TrxQ (Trx y) thioredoxins, Mol. Plant. 2(2009) 270–283.19] E. Gelhaye, N. Rouhier, N. Navrot, J.P. Jacquot, The plant thioredoxin system,Cell. Mol. Life Sci. CMLS 62 (2005) 24–35.
[
nce 255 (2017) 1–11
20] N. Rouhier, J. Couturier, J.-P. Jacquot, Genome-wide analysis of plantglutaredoxin systems, J. Exp. Bot. 57 (2006) 1685–1696.
21] C.S. Koh, et al., An atypical catalytic mechanism involving three cysteines ofthioredoxin, J. Biol. Chem. 283 (2008) 23062–23072.
22] L. Meng, J.H. Wong, L.J. Feldman, P.G. Lemaux, B.B. Buchanan, Amembrane-associated thioredoxin required for plant growth moves from cellto cell, suggestive of a role in intercellular communication, Proc. Natl. Acad.Sci. U. S. A. 107 (2010) 3900–3905.
23] Y. Meyer, B.B. Buchanan, F. Vignols, J.-P. Reichheld, Thioredoxins andglutaredoxins: unifying elements in redox biology, Annu. Rev. Genet. 43(2009) 335–367.
24] G. Wittenberg, A. Danon, Disulfide bond formation in chloroplasts: formationof disulfide bonds in signaling chloroplast proteins, Plant Sci. 175 (2008)459–466.
25] K. Yoshida, T. Hisabori, Two distinct redox cascades cooperatively regulatechloroplast functions and sustain plant viability, Proc. Natl. Acad. Sci. U. S. A.113 (2016) E3967–3976.
26] L. Nikkanen, J. Toivola, E. Rintamäki, Crosstalk between chloroplastthioredoxin systems in regulation of photosynthesis, Plant Cell Environ. 39(2016) 1691–1705.
27] I. Thormählen, et al., Thioredoxin f1 and NADPH-dependent thioredoxinreductase C have overlapping functions in regulating photosyntheticmetabolism and plant growth in response to varying light conditions, PlantPhysiol. 169 (2015) 1766–1786.
28] K.-J. Dietz, Thiol-based peroxidases and ascorbate peroxidases: why plantsrely on multiple peroxidase systems in the photosynthesizing chloroplast?Mol. Cells. 39 (2016) 20–25.
29] N. Rouhier, S.D. Lemaire, J.-P. Jacquot, The role of glutathione inphotosynthetic organisms: emerging functions for glutaredoxins andglutathionylation, Annu. Rev. Plant Biol. 59 (2008) 143–166.
30] C. Vieira Dos Santos, et al., Specificity of thioredoxins and glutaredoxins aselectron donors to two distinct classes of Arabidopsis plastidial methioninesulfoxide reductases B, FEBS Lett. 581 (2007) 4371–4376.
31] N. Rouhier, Plant glutaredoxins: pivotal players in redox biology andiron-sulphur centre assembly, New Phytol. 186 (2010) 365–372.
32] N. Rouhier, et al., Functional and structural aspects of poplar cytosolic andplastidial type a methionine sulfoxide reductases, J. Biol. Chem. 282 (2007)3367–3378.
33] R.M. Schuurmans, P. van Alphen, J.M. Schuurmans, H.C.P. Matthijs, K.J.Hellingwerf, Comparison of the photosynthetic yield of cyanobacteria andgreen algae: different methods give different answers, PLoS One 10 (2015)e0139061.
34] E.F.J. Weenink, V.M. Luimstra, J.M. Schuurmans, M.J. Van Herk, P.M. Visser,H.C.P. Matthijs, Combatting cyanobacteria with hydrogen peroxide: alaboratory study on the consequences for phytoplankton community anddiversity, Front. Microbiol. 6 (2015) 714.
35] H.C.P. Matthijs, et al., Selective suppression of harmful cyanobacteria in anentire lake with hydrogen peroxide, Water Res. 46 (2012) 1460–1472.
36] K.-J. Dietz, Peroxiredoxins in plants and cyanobacteria, Antioxid. RedoxSignal. 15 (2011) 1129–1159.
37] B. Heuer, M.J. Hansen, L.E. Anderson, Light modulation of phosphofructokinasein pea leaf chloroplasts, Plant Physiol. 69 (1982) 1404–1406.
38] J. Erales, L. Avilan, S. Lebreton, B. Gontero, Exploring CP12 binding proteinsrevealed aldolase as a new partner for thephosphoribulokinase/glyceraldehyde 3-phosphate dehydrogenase/CP12complex-purification and kinetic characterization of this enzyme fromChlamydomonas reinhardtii, FEBS J. 275 (2008) 1248–1259.
39] Y. Balmer, A. Koller, G. del Val, W. Manieri, P. Schürmann, B.B. Buchanan,Proteomics gives insight into the regulatory function of chloroplastthioredoxins, Proc. Natl. Acad. Sci. U. S. A. 100 (2003) 370–375.
40] Y. Balmer, et al., Thioredoxin links redox to the regulation of fundamentalprocesses of plant mitochondria, Proc. Natl. Acad. Sci. U. S. A. 101 (2004)2642–2647.
41] M. Hall, et al., Thioredoxin targets of the plant chloroplast lumen and theirimplications for plastid function, Proteomics 10 (2010) 987–1001.
42] N. Rouhier, et al., Identification of plant glutaredoxin targets, Antioxid. RedoxSignal. 7 (2005) 919–929.
43] M.B. Pascual, A. Mata-Cabana, F.J. Florencio, M. Lindahl, F.J. Cejudo, Acomparative analysis of the NADPH thioredoxin reductase C-2-Cysperoxiredoxin system from plants and cyanobacteria, Plant Physiol. 155(2011) 1806–1816.
44] B. Wieles, J. van Noort, J.W. Drijfhout, R. Offringa, A. Holmgren, T.H. Ottenhoff,Purification and functional analysis of the Mycobacterium lepraethioredoxin/thioredoxin reductase hybrid protein, J. Biol. Chem. 270 (1995)25604–25606.
45] N. Rouhier, J.P. Jacquot, Molecular and catalytic properties of aperoxiredoxin-glutaredoxin hybrid from Neisseria meningitidis, FEBS Lett. 554(2003) 149–153.
46] J. Couturier, F. Vignols, J.-P. Jacquot, N. Rouhier, Glutathione- andglutaredoxin-dependent reduction of methionine sulfoxide reductase A, FEBSLett. 586 (2012) 3894–3899.
47] M. Balsera, et al., Ferredoxin:thioredoxin reductase (FTR) links the regulationof oxygenic photosynthesis to deeply rooted bacteria, Planta 237 (2013)619–635.

t Scie
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
with ferredoxin and flavodoxin, Proteins 72 (2008) 848–862.[85] I. Krimm, A. Goyer, E. Issakidis-Bourguet, M. Miginiac-Maslow, J.M. Lancelin,
Direct NMR observation of the thioredoxin-mediated reduction of thechloroplast NADP-malate dehydrogenase provides a structural basis for therelief of autoinhibition, J. Biol. Chem. 274 (1999) 34539–34542.
D.D. Gütle et al. / Plan
48] Y. Balmer, et al., A complete ferredoxin/thioredoxin system regulatesfundamental processes in amyloplasts, Proc. Natl. Acad. Sci. U. S. A. 103 (2006)2988–2993.
49] C.H. Marchand, H. Vanacker, V. Collin, E. Issakidis-Bourguet, P.L. Maréchal, P.Decottignies, Thioredoxin targets in Arabidopsis roots, Proteomics 10 (2010)2418–2428.
50] D. Susanti, et al., Thioredoxin targets fundamental processes in amethane-producing archaeon, Methanocaldococcus jannaschii, Proc. Natl.Acad. Sci. U. S. A. 111 (2014) 2608–2613.
51] J. Couturier, K. Chibani, J.-P. Jacquot, N. Rouhier, Cysteine-based redoxregulation and signaling in plants, Front. Plant Sci. 4 (2013) 105.
52] P. Schürmann, B.B. Buchanan, The ferredoxin/thioredoxin system of oxygenicphotosynthesis, Antioxid. Redox Signal. 10 (2008) 1235–1274.
53] D.D. Gütle, et al., Chloroplast FBPase and SBPase are thioredoxin-linkedenzymes with similar architecture but different evolutionary histories, Proc.Natl. Acad. Sci. U. S. A. 113 (2016) 6779–6784.
54] S. Fermani, et al., Molecular mechanism of thioredoxin regulation inphotosynthetic A2B2-glyceraldehyde-3-phosphate dehydrogenase, Proc. Natl.Acad. Sci. U. S. A. 104 (2007) 11109–11114.
55] N. Zhang, A.R. Portis, Mechanism of light regulation of Rubisco: a specific rolefor the larger Rubisco activase isoform involving reductive activation bythioredoxin-f, Proc. Natl. Acad. Sci. U. S. A. 96 (1999) 9438–9443.
56] S. Morisse, et al., Thioredoxin-dependent redox regulation of chloroplasticphosphoglycerate kinase from Chlamydomonas reinhardtii, J. Biol. Chem. 289(2014) 30012–30024.
57] J. Petersen, R. Teich, B. Becker, R. Cerff, H. Brinkmann, The GapA/B geneduplication marks the origin of streptophyta (charophytes and land plants),Mol. Biol. Evol. 23 (2006) 1109–1118.
58] S. Robbens, J. Petersen, H. Brinkmann, P. Rouzé, Y.V. de Peer, Uniqueregulation of the Calvin cycle in the ultrasmall green alga ostreococcus, J. Mol.Evol. 64 (2007) 601–604.
59] M.A. Ballicora, J.B. Frueauf, Y. Fu, P. Schürmann, J. Preiss, Activation of thepotato tuber ADP-glucose pyrophosphorylase by thioredoxin, J. Biol. Chem.275 (2000) 1315–1320.
60] A.S. Richter, B. Grimm, Thiol-based redox control of enzymes involved in thetetrapyrrole biosynthesis pathway in plants, Front. Plant Sci. 4 (2013) 371.
61] M. Vollmar, D. Schlieper, M. Winn, C. Büchner, G. Groth, Structure of the c14rotor ring of the proton translocating chloroplast ATP synthase, J. Biol. Chem.284 (2009) 18228–18235.
62] D.W. McKinney, B.B. Buchanan, R.A. Wolosiuk, Association of athioredoxin-like protein with chloroplast coupling factor (CF1), Biochem.Biophys. Res. Commun. 86 (1979) 1178–1184.
63] H. Ueoka-Nakanishi, Y. Nakanishi, H. Konno, K. Motohashi, D. Bald, T. Hisabori,Inverse regulation of rotation of F1-ATPase by the mutation at the regulatoryregion on the � subunit of chloroplast ATP synthase, J. Biol. Chem. 279 (2004)16272–16277.
64] L.R. Carrillo, J.E. Froehlich, J.A. Cruz, L.J. Savage, D.M. Kramer, Multi-levelregulation of the chloroplast ATP synthase: the chloroplast NADPHthioredoxin reductase C (NTRC) is required for redox modulation specificallyunder low irradiance, Plant J. 87 (2016) 654–663.
65] A.J. Meyer, et al., Redox-sensitive GFP in Arabidopsis thaliana is a quantitativebiosensor for the redox potential of the cellular glutathione redox buffer,Plant J. Cell Mol. Biol. 52 (2007) 973–986.
66] S.D. Lemaire, et al., NADP-malate dehydrogenase from unicellular green algaChlamydomonas reinhardtii. A first step toward redox regulation? Plant
Physiol. 137 (2005) 514–521.67] G. Née, M. Aumont-Nicaise, M. Zaffagnini, S. Nessler, M. Valerio-Lepiniec, E.Issakidis-Bourguet, Redox regulation of chloroplastic G6PDH activity bythioredoxin occurs through structural changes modifying substrateaccessibility and cofactor binding, Biochem. J. 457 (2014) 117–125.
nce 255 (2017) 1–11 11
68] M. Cardi, et al., Plastidic P2 glucose-6P dehydrogenase from poplar ismodulated by thioredoxin m-type: distinct roles of cysteine residues in redoxregulation and NADPH inhibition, Plant Sci. 252 (2016) 257–266.
69] I. Wenderoth, R. Scheibe, A. von Schaewen, Identification of the cysteineresidues involved in redox modification of plant plastidicglucose-6-phosphate dehydrogenase, J. Biol. Chem. 272 (1997) 26985–26990.
70] J. López Jaramillo, et al., High-yield expression of pea thioredoxin m andassessment of its efficiency in chloroplast fructose-1,6-bisphosphataseactivation, Plant Physiol. 114 (1997) 1169–1175.
71] M. Chiadmi, Redox signalling in the chloroplast: structure of oxidized peafructose-1,6-bisphosphate phosphatase, EMBO J. 18 (1999) 6809–6815.
72] S. Milanez, R.J. Mural, F.C. Hartman, Roles of cysteinyl residues ofphosphoribulokinase as examined by site-directed mutagenesis, J. Biol. Chem.266 (1991) 10694–10699.
73] M.K. Geck, F.C. Hartman, Kinetic and mutational analyses of the regulation ofphosphoribulokinase by thioredoxins, J. Biol. Chem. 275 (2000) 18034–18039.
74] L. Marri, et al., Prompt and easy activation by specific thioredoxins of Calvincycle enzymes of Arabidopsis thaliana associated in the GAPDH/CP12/PRKsupramolecular complex, Mol. Plant. 2 (2009) 259–269.
75] A. Del Giudice, et al., Unravelling the shape and structural assembly of thephotosynthetic GAPDH-CP12-PRK complex from Arabidopsis thaliana bysmall-angle X-ray scattering analysis, Acta Crystallogr. D Biol. Crystallogr. 71(2015) 2372–2385.
76] S. Fermani, et al., Conformational selection and folding-upon-binding ofintrinsically disordered protein CP12 regulate photosynthetic enzymesassembly, J. Biol. Chem. 287 (2012) 21372–21383.
77] R. Scheibe, N. Wedel, S. Vetter, V. Emmerlich, S.-M. Sauermann, Co-existenceof two regulatory NADP-glyceraldehyde 3-P dehydrogenase complexes inhigher plant chloroplasts, Eur. J. Biochem. FEBS. 269 (2002) 5617–5624.
78] E.A. Schmelz, et al., Fragments of ATP synthase mediate plant perception ofinsect attack, Proc. Natl. Acad. Sci. U. S. A. 103 (2006) 8894–8899.
79] E.A. Schmelz, S. LeClere, M.J. Carroll, H.T. Alborn, P.E.A. Teal, Cowpeachloroplastic ATP synthase is the source of multiple plant defense elicitorsduring insect herbivory, Plant Physiol. 144 (2007) 793–805.
80] A. Holmgren, B.O. Söderberg, H. Eklund, C.I. Brändén, Three-dimensionalstructure of Escherichia coli thioredoxin-S2 to 2.8 A resolution, Proc. Natl.Acad. Sci. U. S. A. 72 (1975) 2305–2309.
81] B. Naranjo, A. Diaz-Espejo, M. Lindahl, F.J. Cejudo, Type-f thioredoxins have arole in the short-term activation of carbon metabolism and their loss affectsgrowth under short-day conditions in Arabidopsis thaliana, J. Exp. Bot. 67(2016) 1951–1964.
82] K. Yoshida, S. Hara, T. Hisabori, Thioredoxin selectivity for thiol-based redoxregulation of target proteins in chloroplasts, J. Biol. Chem. 290 (2015)14278–14288.
83] J.P. Jacquot, M. Stein, A. Suzuki, S. Liottet, G. Sandoz, M. Miginiac-Maslow,Residue Glu-91 of Chlamydomonas reinhardtii ferredoxin is essential forelectron transfer to ferredoxin-thioredoxin reductase, FEBS Lett. 400 (1997)293–296.
84] M. Medina, R. Abagyan, C. Gómez-Moreno, J. Fernandez-Recio, Dockinganalysis of transient complexes: interaction of ferredoxin-NADP+ reductase