does predator interference cause alternative stable states in multispecies communities?
TRANSCRIPT

Theoretical Population Biology 82 (2012) 170–176
Contents lists available at SciVerse ScienceDirect
Theoretical Population Biology
journal homepage: www.elsevier.com/locate/tpb
Does predator interference cause alternative stable states inmultispecies communities?Jianfeng Feng a,∗, Vasilis Dakos b, Egbert H. van Nes b
a Key Laboratory of Pollution Processes and Environmental Criteria at Ministry of Education, College of Environmental Science and Engineering, Nankai University,Tianjin 300071, Chinab Department of Aquatic Ecology and Water Quality Management, Wageningen University, P.O. Box 47, 6700 AA, Wageningen, The Netherlands
a r t i c l e i n f o
Article history:Received 12 December 2011Available online 23 June 2012
Keywords:Predator–prey modelCompetition modelBeddington–DeAngelis functional responseResilienceCommunity shiftsCritical transitions
a b s t r a c t
Whereas it is well known that simple ecological mechanisms may promote stability in simplespecies models, their consequences for stability and resilience in multispecies communities are largelyunexplored. Here,we studied the effect of predator interference on the occurrence of alternative attractorsand complex dynamics in randomly constructed multispecies predator–prey communities. We studiedthree types of interference: random interference (‘‘asymmetric’’), random interference but symmetricalbetween pairs of predators (‘‘symmetric’’), and interference among only the same species (‘‘conspecific’’).In all cases predator interference increased the average number of alternative attractors, whereas atthe same time it reduced the emergence of oscillatory or chaotic dynamics. Our findings demonstratea contrasting effect of predator interference on the stability of a community: on the one hand it reducescycles and chaos in the dynamics, on the other hand predator interference increases the likelihood thatcommunities may undergo critical transitions between multiple stable states.
© 2012 Elsevier Inc. All rights reserved.
1. Introduction
Theoretical and empirical studies suggest that alternativeattractors may occur in different ecological systems ranging frommarine systems (Knowlton, 2004; Mumby et al., 2007; Petraitiset al., 2009), to coastal environments (Heffernan, 2008; Van deKoppel et al., 2001), shallow lakes (Scheffer et al., 1993; Schefferand van Nes, 2007), and terrestrial ecosystems (Chartier andRostagno, 2006; Janssen et al., 2008; Kefi et al., 2007). The existenceof alternative attractors implies that ecosystems run the risk ofshifting from one state to another. Such critical transitions haveimportant consequences for ecosystem management: becauserestoration of an ecosystem to its initial state is often difficult oreven impossible (Folke et al., 2004; Scheffer and Carpenter, 2003),critical transitions are usually associated with permanent losses ofecosystems services.
Alternative attractors often emerge in the presence of a destabi-lizing positive feedback combined with negative feedbacks (Schef-fer et al., 2009). For example, strong interspecific competition is amechanism thatmay produce alternative attractorswhen linked to
∗ Corresponding author.E-mail addresses: [email protected] (J. Feng), [email protected]
(V. Dakos), [email protected] (E.H. van Nes).
0040-5809/$ – see front matter© 2012 Elsevier Inc. All rights reserved.doi:10.1016/j.tpb.2012.06.003
a positive feedback (Knowlton, 1992). A dominant species in a com-munity will prevent the invasion of a competitor as long as inter-specific competition is stronger than conspecific competition. In-deed, the classical two-species Lotka–Volterra competition model(Lotka, 1925; Volterra, 1926) can yield two stable point-attractors:each represents the dominance of either competitor and the ex-clusion of the other. The outcome of the competition depends onstrength of competition as well as on the initial densities of bothcompetitors.
While many studies have focused on multiple alternative at-tractors (or alternative stable states) in models with few species,little is known about alternative attractors in multispecies com-munities (Feng et al., 2006; Van Nes and Scheffer, 2004). A rea-sonable hypothesis is that the conditions that lead to alternativeattractors in simple models should also create alternative attrac-tors in complex multispecies settings. It appears, however, thatthis is not necessarily the case. For instance, Van Nes and Schef-fer (2004) showed that alternative attractors occur in multispeciescompetition models even if interspecific competition is smallerthan intraspecific competition, contrary to theoretical expecta-tions based on simple two-species models. The implications ofsuch discrepancies are fundamental when it comes to estimat-ing the resilience of multispecies communities. Thus, scaling up tomultispecies models in order to investigate the occurrence of al-ternative attractors presents a research field worth exploring.
The implications of predator interference for the occurrenceof alternative attractors have not been thoroughly investigated.

J. Feng et al. / Theoretical Population Biology 82 (2012) 170–176 171
Predator interference refers to situations where a predation rate issuppressed in the presence of other predators (Beddington, 1975;DeAngelis et al., 1975). It is a widely observed phenomenon inpredator–prey and host–parasitoid systems (Arditi and Akcakaya,1990) ranging from hares to fish and insects (Skalski and Gilliam,2001). In simple two or three species predator–prey models(Arditi et al., 2004; Rall et al., 2008; Van Voorn et al., 2008;Zhang et al., 2007), predator interference can stabilize populationdynamics, in the sense that it reduces the parameter spacewhere unstable attractors give rise to limit cycles. Rall et al.(2008) studied whether predator interference tended to reduceoscillations in complex food web models or not. They showed thatspecies’ survival, species’ richness and their average coefficient ofvariation changed with predator interference and demonstratedthat predator interference will stabilize the dynamics of complexfood web models in contrast to the destabilizing effect ofenrichment in classic food web models. These results imply thatpredator interference enhances the stability of species dynamics(in terms of reducing the occurrence of unstable attractors), but it isunclear whether it may also have implications for the resilience ofa community in terms of its likelihood of shifting to an alternativeattractor. It seems reasonable to assume that this likelihoodincreases if there are more alternative stable states possible (VanNes and Scheffer, 2004).
Here we study the effect of predator interference on the oc-currence of alternative attractors in predator–prey communities.We first explore how predator interference can lead to alterna-tive stable states in a simple model of one prey with two interfer-ing predators. We then extend the analysis to a model of multiplecompeting prey species with multiple predators. In particular, weuse a Beddington–DeAngelis functional response to model preda-tor interference to investigate its effects on the dynamics and theemergence of alternative attractors in randomly constructed mul-tispecies communities.
2. Model and methods
2.1. A simple model with two interfering predators and one prey
We explored whether predator interference can lead toalternative states in a lowdimensional systemwith two interferingpredators P1 and P2 consuming one prey species C . The modelfollows the classical Rosenzweig–MacArthur predator–preymodel(Dakos et al., 2009; Rosenzweig and MacArthur, 1963; Van Nesand Scheffer, 2004; Vandermeer, 1993), which we extended witha Beddington–DeAngelis functional response to include predatorinterference (Beddington, 1975; DeAngelis et al., 1975):
dCdt
= rC1 −
CK
− gP1
CC + H + c1P1 + kP2
− gP2C
C + H + c2P2 + kP1dP1dt
= egP1C
C + H + c1P1 + kP2− mP1
dP2dt
= egP2C
C + H + c2P2 + kP1− mP2.
(1)
The prey species grows logistically with a maximum growthrate r and carrying capacity K . Parameter e is the assimilationefficiency of predators P1,2 and m is the death rate of predatorsP1,2. Predators consumeprey according to a Beddington–DeAngelisfunctional response (Beddington, 1975; DeAngelis et al., 1975)withmaximumgrazing rate g and fixedhalf saturationH . kdenotesinterference between predator species (interspecific), whereasc1 and c2 describe interferences within predator species P1, P2(conspecific).
For simplicity, we assumed that all parameters were the samefor both predators, including interspecific interference k. Lastly,we assumed that prey species C was at its stable (non-cyclic)equilibrium (dC/dt = 0, C = C∗), and we performed a stabilityanalysis of the following model:
dP1dt
= P1
eg
C∗
C∗ + H + c1P1 + kP2− m
dP2dt
= P2
eg
C∗
C∗ + H + c2P2 + kP1− m
.
(2)
Although the assumption of stable non-cyclic equilibrium isvalid only for a limited part of parameter space, it serves ourpurpose to illustrate the effect of predator interference on theoccurrence of alternative states. More details can be found in theElectronic Supplementary Material (ESM) 1.
2.2. A multispecies predator–prey model with predator interference
We studied the effects of predator interference on theoccurrence of alternative states by extending the three speciespredator–prey model with predator interference to a multispeciespredator–prey community, following earlier contributions (Dakoset al., 2009; Rosenzweig and MacArthur, 1963; Van Nes andScheffer, 2004; Vandermeer, 1993):
dCi
dt= riCi
1 −
j(αijCj)
Ki
−
k
gkPkSikCi
j(SjkCj) + b
n
(BknPn) + Hk
+ u
dPkdt
= ekgkPk
jSjkCj
j(SjkCj) + b
n
(BknPn) + Hk− mkPk + u
(3)
in which Ci and Pk are now the biomasses of prey i and predatorspecies k respectively. The prey species grow logistically witha maximum growth rate ri and a carrying capacity Ki. Thecompetition coefficient between prey species i and j is written asαij. Sik is the preference of predator Pk for prey Ci. It indicates theavailability of prey i to predator k (Pelletier, 2000). Parameter ek isthe assimilation efficiency of predator Pk andmk is the death rate ofpredator Pk. Factor u introduces a small immigration factor whichis used to prevent unrealistically low species biomass values andheteroclinic cycles (Van Nes and Scheffer, 2004).
Predators consume prey with maximum grazing rate gk andfixed half saturation Hk. Interference between predator species kand n is included by factor Bkn whereas the parameter b scales theoverall strength of predator interference.
2.3. Model parameterization
Wemodeled 10 competing prey and 6 predator species. Defaultparameters representative for plankton communities (Dakos et al.,2009; Scheffer et al., 1997) are summarized in Table 1. Interspecificcompetition terms (αij) were drawn from a uniform distributionbetween 0.5 and 1.5, while intraspecific competition terms (αij)were set by definition to 1. Prey preferences S were also drawnfrom a uniform distribution between 0 and 1 to create a matrixof generalists with average selectivity of 0.5. Carrying capacityK was equal for all competing prey species (i.e. Ki = K ). Allpredator-interference coefficients Bkn were drawn from a uniform

172 J. Feng et al. / Theoretical Population Biology 82 (2012) 170–176
Fig. 1. Nullclines of the two interfering predators model feeding on a common constant prey. Similar to the classical Lotka–Volterra competition model, it is the relativestrength of interspecific and conspecific interference that determines whether stable coexistence is possible or whether only one of the two predators will prevail.(a) If interspecific interference is weaker than conspecific (k < c1,2), stable coexistence is possible. (b) When interspecific interference becomes greater than conspecificinterference (k > c1,2), alternative attractors can emerge.
Table 1Parameter values or ranges used in the model simulations.
Symbol Interpretation Range Units
ri Maximum specific growth rate of prey 0.5 d−1
mk Mortality rate of predators 0.15 d−1
αij Prey competition coefficient 0.5–1.5 –gk Maximum grazing rate of predators 0.4 d−1
Sik Preference coefficient of predators 0–1 –Hk Half-saturation constant of predators 0.6 mg L−1
ek Assimilation efficiency 0.7 –u Immigration rate 10−7 mgL−1d−1
Ki Carrying capacity of prey species 2, 5, 10 mg L−1
b Strength of predator interference 0, 0.05, 0.5 –
Table 2Overview of structure and strength settings of predator interference used in thestudy.
Type of predator interference Structure Bkn Strength b
No interference Not applicable. 0Symmetrical interference Bkn = Bnk 0.05 (weak)Asymmetrical interference Bkn = Bnk 0.05 (weak)Conspecific interference Bkn = 0, n = k 0.05 (weak)Symmetrical interference Bkn = Bnk 0.5 (strong)Asymmetrical interference Bkn = Bnk 0.5 (strong)Conspecific interference Bkn = 0, n = k 0.5 (strong)
distribution between 0 and 1. (More details about the parametersampling values in ESM 2.)
We investigated the effect of both structure and strengthof predator interference on the occurrence and dynamics ofalternative attractors (Table 2). We assumed three different cases:(a) symmetric predator interference,Bkn = Bnk (predator species Pnand Pk interfered equally with each other), (b) random asymmetricpredator interference, Bkn = Bnk (predator species Pn and Pkinterfered randomly with each other), and (c) interference onlyamong conspecifics, Bkn = 0, n = k (predators interfered onlywith individuals of the same species). We also tested for differentaverage strengths of predator interference (b = 0, 0.05, or 0.5), aswell as different levels of carrying capacity (K = 2, 5, and 10).
2.4. Model analysis
We started simulations with random initial conditions for all16 species (between 0 and 10 mg/L). To ensure that the systemreached an attractor, we first simulated our model communitiesfor 400 years (146,000 days). These transients were discardedand we continued the simulation for another 40 years. Wecategorized the dynamics of the 40 year long generated timeseries automatically into point attractors, simple cyclic attractors,
complex cyclic attractors, and chaotic attractors. Point attractorswere solutions without oscillations. Simple cyclic attractors weredefined as limit cycles with a period of one. Complex cyclicattractorswere limit cycles with a period of more than one. Chaoticattractors were identified by a positive Lyapunov exponent. TheLyapunov exponent quantifies the rate of exponential divergence(or convergence) of nearby trajectories (Strogatz, 1994). A positiveLyapunov exponent indicates chaos, and its magnitude is ameasure of the system’s sensitivity to initial conditions.
We searched for alternative attractors by performing 100simulations with different initial conditions for each parametersetting. For every simulation, the selectivity coefficients matrixS, competition coefficients matrix A and predator interferencecoefficients matrix B were randomly sampled from a uniformdistribution. We identified alternative point attractors basedon the species with maximum biomass, i.e. point attractorswith different species of maximum biomass were consideredalternative attractors. We identified alternative simple cyclicattractors by checking if species with maximum biomasses werethe same. In the case of alternative complex cyclic attractorswe first checked if the periods were the same. If that was thecase, we checked for differences in the species with maximumbiomass. As we could not differentiate between alternative chaoticattractors, we assumed that all chaotic solutions belonged to thesame attractor.
All simulations were implemented in MATLAB using thesoftware package GRIND (freely available at http://www.dow.wau.nl/aew/grind).
3. Results
3.1. The mechanistic origin of alternative attractors due to predatorinterference in a three-species model
When both types of interference were present, stable coex-istence of both predators was feasible, but only when predatorinterspecific interference was weaker than predator conspecificinterference (k < c1,2) (Fig. 1(a), see also ESM 1). When interspe-cific interference exceeded conspecific interference (k > c1,2) al-ternative attractors emerged, and only one of the two predatorsdominated (Fig. 1(b)). The transition between the two alternativeattractors (from dominance to extinction of the dominant preda-tor) occurred through a transcritical bifurcation either through adecrease in interspecific interference k of the dominant predator(Fig. 2(a)) or through an increase in conspecific interference c ofthe dominant predator (Fig. 2(b)). These results resemble those ofoutcompetition in the classical Lotka–Volterra model, and show
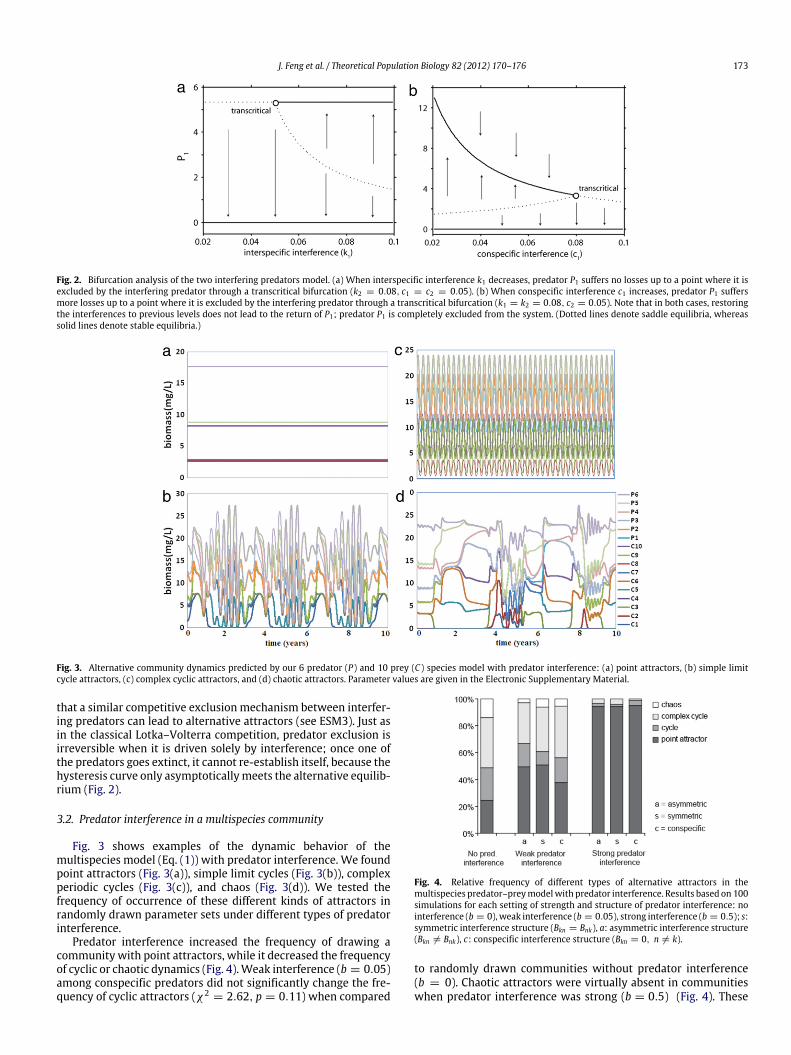
J. Feng et al. / Theoretical Population Biology 82 (2012) 170–176 173
Fig. 2. Bifurcation analysis of the two interfering predators model. (a) When interspecific interference k1 decreases, predator P1 suffers no losses up to a point where it isexcluded by the interfering predator through a transcritical bifurcation (k2 = 0.08, c1 = c2 = 0.05). (b) When conspecific interference c1 increases, predator P1 suffersmore losses up to a point where it is excluded by the interfering predator through a transcritical bifurcation (k1 = k2 = 0.08, c2 = 0.05). Note that in both cases, restoringthe interferences to previous levels does not lead to the return of P1; predator P1 is completely excluded from the system. (Dotted lines denote saddle equilibria, whereassolid lines denote stable equilibria.)
Fig. 3. Alternative community dynamics predicted by our 6 predator (P) and 10 prey (C) species model with predator interference: (a) point attractors, (b) simple limitcycle attractors, (c) complex cyclic attractors, and (d) chaotic attractors. Parameter values are given in the Electronic Supplementary Material.
that a similar competitive exclusion mechanism between interfer-ing predators can lead to alternative attractors (see ESM3). Just asin the classical Lotka–Volterra competition, predator exclusion isirreversible when it is driven solely by interference; once one ofthe predators goes extinct, it cannot re-establish itself, because thehysteresis curve only asymptoticallymeets the alternative equilib-rium (Fig. 2).
3.2. Predator interference in a multispecies community
Fig. 3 shows examples of the dynamic behavior of themultispecies model (Eq. (1)) with predator interference. We foundpoint attractors (Fig. 3(a)), simple limit cycles (Fig. 3(b)), complexperiodic cycles (Fig. 3(c)), and chaos (Fig. 3(d)). We tested thefrequency of occurrence of these different kinds of attractors inrandomly drawn parameter sets under different types of predatorinterference.
Predator interference increased the frequency of drawing acommunity with point attractors, while it decreased the frequencyof cyclic or chaotic dynamics (Fig. 4).Weak interference (b = 0.05)among conspecific predators did not significantly change the fre-quency of cyclic attractors (χ2
= 2.62, p = 0.11) when compared
Fig. 4. Relative frequency of different types of alternative attractors in themultispecies predator–preymodelwith predator interference. Results based on 100simulations for each setting of strength and structure of predator interference: nointerference (b = 0),weak interference (b = 0.05), strong interference (b = 0.5); s:symmetric interference structure (Bkn = Bnk), a: asymmetric interference structure(Bkn = Bnk), c: conspecific interference structure (Bkn = 0, n = k).
to randomly drawn communities without predator interference(b = 0). Chaotic attractors were virtually absent in communitieswhen predator interference was strong (b = 0.5) (Fig. 4). These

174 J. Feng et al. / Theoretical Population Biology 82 (2012) 170–176
Fig. 5. Relative frequency of number of attractors in the multispecies predator–prey model with predator interference. Results based on 100 simulations for each setting ofstrength and structure of predator interference: (a)–(c): no interference (b = 0), (d)–(f): weak interference (b = 0.05), (g)–(i): strong interference (b = 0.5); upper panels:symmetric interference (Bkn = Bnk), middle panels: asymmetric interference (Bkn = Bnk), lower panels: conspecific interference (Bkn = 0, n = k).
results are in accordance with the stabilizing effect of predatorinterference found in two species population cycles (Arditi et al.,2004; DeAngelis et al., 1975). Interestingly, the structure of preda-tor interference had little effect on the occurrence of complex dy-namics except in the case of weak conspecific interference, whichseemed to slightly favor complex dynamics (Fig. 4).
Predator interference not only increased the stability in the dy-namics of our randomly drawn multispecies communities, but italso increased both the emergence (Fig. 4) and the total num-ber of alternative attractors (Fig. 5). In the absence of preda-tor interference, alternative attractors occurred in 30% of thesimulations. In the presence of predator interference this fre-quency was significantly higher (on average 66%, χ2
= 12.56,p < 0.01). In particular, symmetric predator interference had thestrongest effect on the emergence and the number of different al-ternative attractors (n = 7) (Fig. 5(d), (g)). Conspecific predator in-terference had the weakest effect on the emergence of alternativeattractors, even though its effect on the emergence of complex dy-namics was the strongest (Fig. 5(f), (i)).
Our results appear to be dependent on the level of productivity(K ) (Table 3). Generally, a decrease in productivity increased thefrequency of stable solutions (chaotic and cyclic dynamics werereduced) from 57% on the high productivity level (K = 10) to 100%on the low productivity level (K = 2). Decreasing productivityalso led to an increase in the occurrence of alternative attractorsfrom 53% on the high productivity level (K = 10) to 82% on thelow productivity level (K = 2). At low productivity (K = 2) thereare all stable point solutions and strong predator interferencehad little effect on the number of alternative attractors comparedwith no predator interference (χ2
= 3.63, p = 0.06). However, atintermediate and high levels of productivity (K = 5, 10), strongpredator interference caused a significant rise in the frequencyof alternative attractors in the communities (χ2
= 19.66,p < 0.001, K = 5; χ2
= 64.58, p < 0.001, K = 10) and at thesame time stabilized the system with more stable point attractors(χ2
= 21.81, p < 0.001, K = 5; χ2= 95.94, p < 0.001, K = 10)
compared with no predator interference.
4. Discussion
Our main finding is that predator interference reduces thefrequency of finding cycles and chaos in randomly drawnmultispecies communities, while it increases the frequency of
alternative attractors. This result is very robust to the type ofpredator interference modeled; it made little difference whetheronly conspecifics or different species were interfering (Fig. 4).However, the type of interference mattered when looking at thenumber of alternative attractors, as symmetrical interference ledto the highest number of alternative attractors (Fig. 5).
4.1. Why do we get more stable dynamics?
Our results are in line with previous studies that show thatpredator interference has a stabilizing effect on the dynamicsof models with a few species (Rall et al., 2008; Van Voornet al., 2008; Zhang et al., 2007) as well as on the dynamicsof multispecies communities (Rall et al., 2008). This effect caneasily be explained as follows. When predator density increases,increased competition amongpredators causes predation to be lessefficient. Prey species can thus maintain a relatively high biomassand predator–prey cycles are dampened. This also explains whythe structure of predator interference has little influence on theoverall stability of the dynamics: in all cases we get less efficientpredation if the total biomass of predators is high. It is thus notthe structure (determined by the interference matrix Bkn) but thestrength (determined by b) of predator interference that influencesthe stability of predator–prey dynamics.
4.2. Why do we get more alternative attractors?
Competition is a well-studied mechanism that causes multipleattractors in ecosystems (Gilpin and Case, 1976; Van Nes andScheffer, 2004). In terms of a positive feedback it can be understoodthat once a species has reached a sufficient biomass, it hasan advantage in terms of growth and drives its competitorto extinction. It, thus, follows that while predator interferencereduces predator–prey cycles there are more alternative stablestates (Van Nes and Scheffer, 2004). Indeed, under stationaryconditions, higher predator interference leaves more space forcompetition among prey species to drive dynamics, and allows theemergence ofmultiple attractors through outcompetition betweenprey species.
However, a careful look at the results of the multispeciescommunities shows that an increase in the strength of predatorinterference does not always lead to an increase in the frequency

J. Feng et al. / Theoretical Population Biology 82 (2012) 170–176 175
Table 3Occurrence of different numbers of alternative attractors and different types of attractors in communities with asymmetrical predator interference under differentproductivities K .
Number ofalternativeattractors
K = 10 K = 5 K = 2
Predator interference Predator interference Predator interferenceNo(b = 0) (%)
Weak(b = 0.05) (%)
Strong(b = 0.5) (%)
No(b = 0) (%)
Weak(b = 0.05) (%)
Strong(b = 0.5) (%)
No(b = 0) (%)
Weak(b = 0.05) (%)
Strong(b = 0.5) (%)
1 70 59 13 42 27 13 22 21 112 26 30 24 48 49 25 48 33 323 4 10 32 8 21 38 20 31 334 0 1 23 2 3 19 10 13 185 0 0 8 0 0 5 0 2 6
Type ofattractors
Point 25 51 94 62 75 91 100 100 100Simple cycle 24 10 2 13 8 5 0 0 0Complex cycle 37 33 4 23 18 4 0 0 0Chaos 14 6 0 1 0 0 0 0 0
of occurrence of alternative attractors. The analysis of themodel with two interfering predators feeding on a single preyshows that one should only expect a higher frequency ofalternative attractors when interspecific interference is strongerthan conspecific interference. In our multispecies simulations,however, we assumed similar strengths for both types ofinterference. What is then the mechanism that may boost theoccurrence of alternative attractors due to ‘‘interfering exclusion’’in multispecies communities? The answer is not only in thestrength, but also in the structure of predator interference.
When interference is symmetric, we clearly get more alter-native attractors in randomly drawn communities. Symmetricinterspecific interference maximizes the chance of interspecificinterference to be stronger than conspecific interference amongpairs of predators. When one predator becomes dominant, it re-duces considerably the effective grazing rate of its symmetricallyinteracting predators; this can potentially lead to outcompeti-tion of the invading predator species by the dominant predator.Van Nes and Scheffer (2004) found a similar effect for symmet-ric competition. Even when interspecific competition was weakerthan intraspecific competition, symmetry in the competition ma-trix between species led to competitive exclusion. We show thateven for a three-species Lotka–Volterra model alternative equi-libria are possible when almost all competition coefficients aresmaller than one (see ESM 3). Interestingly, when predators inter-fere only among their conspecifics, we get only a small increase inthe occurrence of alternative attractors. This result is in full agree-ment with the results from themodel of two interfering predators.Therein, alternative attractors can only emerge in the presence ofboth conspecific and interspecific interference. In the multispeciescommunities where only conspecific interference was present, in-terference among predators had no effect; the increase in alterna-tive attractors was only due to competition between prey species.
4.3. What are the ecological consequences of these findings?
Perhaps the most interesting conclusion from our study is thatmultiplicity of attractors in multispecies communities may bemore common than expected. Competitive interaction betweenmultiple prey species is already known as a mechanism that leadsto alternative states (Van Nes and Scheffer, 2004). On top of that,our results show that interspecific predator interference enhancesthe occurrence of multiple attractors even further. This can beexplained due to a mechanism of ‘‘interfering exclusion’’ basedon our simple model of two interfering predators on a singleprey. Our multispecies simulations showed that when these two
mechanisms act together, both the frequency of occurrence andthe number of alternative attractors increasemost strongly (Fig. 5).Van Nes and Scheffer (2004) showed that environmental changein multispecies systems could lead to abrupt unexpected shiftsbetween alternative states. It seems reasonable to expect that thisrisk will increase if there are more alternative attractors.
Nevertheless, the effect of predator interference on the risk ofabrupt shifts seems to depend on the type of community and levelof productivity. We found that predator interference has minimaleffects when it is weak or when it occurs among conspecifics. In-stead, its maximum impact is demonstrated in cases with strongand symmetric interference between different species. Strong in-terference is to be expected in populations with territorial and an-tagonistic behavior, such as mammal predators and insects. Suchcommunities aremore likely to have alternative stable states com-pared to communities of organisms in which it is physically im-possible to achieve strong predator interference, like herbivorouszooplankton or parasites (Arditi et al., 2004). In this case, alterna-tive attractors may be driven more by competition among prey.
Generally, an increase in carrying capacity destabilized thesystem and increased the frequency of chaotic and cyclic dynamicssolutions (Roy and Chattopadhyay, 2007) and at the same timeled to an increase in the occurrence of alternative attractors.In environments of low productivity, predator interference alsodoesn’t really enhance the chance of alternative attractors orstabilize the dynamics of multispecies communities. The reasonis simply because under such conditions the dynamics arealready mostly stable (see many publications on the ‘‘paradox ofenrichment’’ of Rosenzweig (1971), reviewed recently by Roy andChattopadhyay, 2007) and the occurrence of alternative attractorswill be mostly bottom-up driven by competition among prey. Butthings are different in environmentswith high productivity; strongpredator interference caused very significant rise in the occurrenceof alternative attractors in the communities and at the same timestabilized the dynamics of multispecies communities with morestable point attractors compared to no predator interference.
In conclusion, our results suggest opposing effects of predatorinteractions on the stability of amultispecies community. Predatorinterference, on the one hand, decreases the occurrence of non-equilibrium dynamics, whereas on the other hand, it increases theoccurrence of alternative attractors. Such contrasting effects areimportant for the study of multispecies communities. Networkstudies have been focusing on mechanisms that promote thestability in terms of dynamics of complex food web models(Rall et al., 2008), but have not considered the issue of multiplestates as an extra dimension of stability. Apparently, network

176 J. Feng et al. / Theoretical Population Biology 82 (2012) 170–176
structures that allow for stable (non-oscillatory) dynamics butincrease the occurrence of multiple attractors, are examples oflocally stable systems in a global unstable landscape where shiftsfrom one state to another may even be catastrophic.
Acknowledgments
We thank the two anonymous reviewers whose commentshelped improve the manuscript. We also thank Andrea Downingfor improving our text and Lin Zhu for the simulation analysisand comments. VD and EvN are supported by a EuropeanUnion Advanced Grant (ERC) awarded to Marten Scheffer. JF issupported by the National Natural Science Foundation of China(No. 41006065), National Major Project of Science and TechnologyMinistry of China (No. 2012ZX07501003) and FundamentalResearch Funds for the Central Universities.
Appendix. Supplementary data
Supplementary material related to this article can be foundonline at http://dx.doi.org/10.1016/j.tpb.2012.06.003.
References
Arditi, R., Akcakaya, H.R., 1990. Underestimation of mutual interference ofpredators. Oecologia 83, 358–361.
Arditi, R., et al., 2004. Does mutual interference always stabilize predator–preydynamics? a comparison ofmodels. Comptes Rendus Biologies 327, 1037–1057.
Beddington, J.R., 1975. Mutual interference between parasites or predators and itseffects on searching efficiency. Journal of Animal Ecology 51, 597–624.
Chartier, M.P., Rostagno, C.M., 2006. Soil erosion thresholds and alternative statesin northeastern patagonian rangelands. Rangeland Ecology & Management 59,616–624.
Dakos, V., et al., 2009. Interannual variability in species composition explained asseasonally entrained chaos. Proceedings of the Royal Society Biological SciencesSeries B 276, 2871–2880.
DeAngelis, D.L., et al., 1975. A model for trophic interaction. Ecology 56, 881–892.Feng, J., et al., 2006. Alternative attractors in marine ecosystems: a comparative
analysis of fishing effects. Ecological Modelling 195, 377–384.Folke, C., et al., 2004. Regime shifts, resilience, and biodiversity in ecosystem
management. Annual Review of Ecology, Evolution, and Systematics 35,557–581.
Gilpin, M.E., Case, T.J., 1976. Multiple domains of attraction in competitioncommunities. Nature 261, 40–42.
Heffernan, J.B., 2008. Wetlands as an alternative stable state in desert streams.Ecology 89, 1261–1271.
Janssen, R.H.H., et al., 2008. Microscale vegetation-soil feedback boosts hysteresisin a regional vegetation-climate system. Global Change Biology 14, 1104–1112.
Kefi, S., et al., 2007. Spatial vegetation patterns and imminent desertification inmediterranean arid ecosystems. Nature 449, 213–217.
Knowlton, N., 1992. Thresholds and multiple stable states in coral reef communitydynamics. American Zoologist 32, 674–682.
Knowlton, N., 2004. Multiple ‘‘stable’’ states and the conservation of marineecosystems. Progress in Oceanography 60, 387–396.
Lotka, A.J., 1925. Elements of Physical Biology. Dover, NewYork, (Reprinted in 1956).Mumby, P.J., et al., 2007. Thresholds and the resilience of caribbean coral reefs.
Nature 450, 98–101.Pelletier, J.D., 2000. Are large complex ecosystems more unstable? a theoretical
reassessment with predator switching. Mathematical Biosciences 163, 91–96.Petraitis, P.S., et al., 2009. Experimental confirmation of multiple community states
in a marine ecosystem. Oecologia 161, 139–148.Rall, B.C., et al., 2008. Food-web connectance and predator interference dampen the
paradox of enrichment. Oikos 117, 202–213.Rosenzweig, M.L., 1971. Paradox of enrichment: destabilization of exploitation
ecosystems in ecological time. Science 171, 385–387.Rosenzweig, M.L., MacArthur, R.H., 1963. Graphical representation and stability
conditions of predator–prey interactions. American Naturalist 97, 209–223.Roy, S., Chattopadhyay, J., 2007. The stability of ecosystems: a brief overview of the
paradox of enrichment. Journal of Biosciences 32, 421–428.Scheffer, M., Carpenter, S.R., 2003. Catastrophic regime shifts in ecosystems: linking
theory to observation. Trends in Ecology & Evolution 18, 648–656.Scheffer, M., et al., 1993. Alternative equilibria in shallow lakes. Trends in Ecology
& Evolution 8, 275–279.Scheffer,M., et al., 1997. On the dominance of filamentous cyanobacteria in shallow,
turbid lakes. Ecology 78, 272–282.Scheffer, M., et al., 2009. Early warnings of critical transitions. Nature 461, 53–59.Scheffer, M., van Nes, E.H., 2007. Shallow lakes theory revisited: various alternative
regimes driven by climate, nutrients, depth and lake size. Hydrobiologia 584,455–466.
Skalski, G.T., Gilliam, J.F., 2001. Functional responses with predator interference:viable alternatives to the Holling type II model. Ecology 82, 3083–3092.
Strogatz, S.H., 1994. Nonlinear Dynamics and Chaos—With Applications to Physics,Biology, Chemistry and Engineering. Addison-Wesley Publishing Company,Reading.
Van de Koppel, J., et al., 2001. Domultiple stable states occur in natural ecosystems?evidence from tidal flats. Ecology 82, 3449–3461.
Vandermeer, J., 1993. Loose coupling of predator–prey cycles-entrainment, chaos,and intermittency in the classic McArthur consumer-resource equations.American Naturalist 141, 687–716.
Van Nes, E.H., Scheffer, M., 2004. Large species shifts triggered by small forces.American Naturalist 164, 255–266.
Van Voorn, G.A.K., et al., 2008. Stabilization due to predator interference:comparison of different analysis approaches. Mathematical Biosciences andEngineering 5, 567–583.
Volterra, V., 1926. Fluctuations in the abundance of a species consideredmathematically. Nature 118, 558–560.
Zhang, F., et al., 2007. Influence of consumermutual interference on the stabilizationof a three-level trophic system. Population Ecology 49, 155–163.