
Lo scheletro idrostatico.
Lo scheletro idrostatico rappresenta l'apparato di sostegno
più semplice descritto negli animali: lo si ritrova negli
Cnidari (anemoni di mare e meduse), negli Anellidi
(lombrichi) e in altri invertebrati.
Il suo nome deriva dal fatto che consiste in una cavità
corporea colma di un liquido incomprimibile (l'acqua),
circondata da muscoli. Quando un gruppo di muscoli
opportunamente orientati si contrae, viene generata una
compressione sulla cavità contenente il liquido. Una
porzione della parete della cavità dello scheletro idrostatico
si estroflette dal versante opposto rispetto al sito di
compressione. Un esempio di tale meccanismo si può
osservare in un anemone di mare, il quale dispone di un
idroscheletro contenente acqua marina. Per estendere il
proprio corpo e le estroflessioni tentacolari, l'animale
chiude l'apertura orale e quindi contrae la guaina
muscolare circolare che circonda la propria cavità corporea,
provocando, con la pressione del liquido, l'effetto di
estensione. In condizioni di pericolo l'anemone può ottenere
l'effetto contrario, utilizzando muscoli disposti
longitudinalmente intorno allo scheletro idrostatico e
parallelamente rispetto all'asse maggiore dei tentacoli.
In certi gruppi animali questo tipo di scheletro risulta
adattato a svolgere una funzione motoria rispetto al
substrato, come accade nel caso dei lombrichi. In tali
animali la cavità corporea è ripartita in diversi segmenti
modulari, mentre la sua parete possiede un duplice strato
muscolare: uno, interno, presenta fibre ad andamento
longitudinale; l'altro, esterno, ad andamento circolare
(▶figura 1).
Ogni segmento corporeo dispone di un comparto chiuso
ripieno di liquido sul quale si esercita la compressione dei
muscoli: quando si contraggono quelli circolari il singolo
segmento tende a farsi più lungo e più sottile; se invece si
contraggono quelli ad andamento longitudinale si verifica
un accorciamento del segmento, che tende a rigonfiarsi.
Figura 1. II ruolo dello scheletro idrostatico nella locomozione
(A) Lo scheletro idrostatico di un lombrico è costituito da cavità contenenti liquido, isolate l'una dall'altra mediante setti, (B) La contrazione dei muscoli disposti circolarmente (alla periferia) determina l'allungamento dei comparti contenenti il liquido e dei segmenti rispettivi, mentre quella dei muscoli disposti longitudinalmente (più internamente) ne riduce la lunghezza. Il movimento dell'animale su un dato substrato risulta dall'alternanza dei processi di allungamento e di quelli di accorciamento: il lombrico avanza con un estremo che prima si assottiglia e quindi incrementa il diametro. Le «setole» (o chete) consentono l'ancoraggio al substrato e impediscono il movimento retrogrado dei segmenti corporei, quando questi sono interessati dalle onde di contrazione.
Il funzionamento alternato dei due rivestimenti muscolari
determina onde di compressione e di dilatazione,che
interessano il corpo dell'animale. I segmenti brevi ed
espansi funzionano come dispositivi di presa sul substrato
grazie anche all'ancoraggio garantito da specializzazioni
tegumentali (chete) simili in aspetto a setole. In questo
modo l'animale si muove abbastanza celermente tanto
negli spostamenti in superficie quanto in quelli
sotterranei.
Un interessante adattamento dello scheletro idrostatico
consente ad alcuni molluschi cefalopodi (polpi e calamari) di
muoversi mediante un dispositivo a reazione. I muscoli
disposti intorno alla cavità contenente il liquido si
contraggono e ne provocano l'espulsione, determinando
il movimento dell'animale in senso opposto.
1. Il sistema scheletrico.
In assenza di un dispositivo rigido sul quale applicarsi, un muscolo può solamente contrarsi o rilasciarsi, cioè cambiare
forma, ma non può eseguire alcun lavoro. Tra gli animali si descrivono sistemi scheletrici di vario tipo, in grado di
tradurre in lavoro la contrazione muscolare e di rendere possibili movimenti coordinati e opportunamente diretti. Oltre
a concorrere a tali funzioni cinetiche, le strutture scheletriche degli animali svolgono anche un ruolo di protezione nei
confronti dei danni di tipo meccanico che l'ambiente esterno può arrecare. In alcuni gruppi, non necessariamente
primitivi, questa protezione esterna coinvolge l'intero corpo: è noto che molti vertebrati, persino alcuni mammiferi,
dispongono di una vistosa corazzatura esterna. Nei vertebrati, tuttavia, la funzione protettiva delle strutture scheletriche
è riservata prevalentemente agli organi di senso e al sistema nervoso centrale.
Le fondamentali varietà dei dispositivi scheletrici degli animali si riducono a tre: lo scheletro idrostatico, l'esoscheletro e
l'endoscheletro. La prima parte di questo capitolo riguarda lo scheletro dal punto di vista generale e comparativo mentre
la seconda parte tratta in particolare lo scheletro dell'uomo.

L’esoscheletro.
Si definisce in tal modo una complessa struttura
sclerotizzata (cioè notevolmente indurita), disposta sulla
superficie corporea e in grado di dare attacco ai muscoli lo-
calizzati internamente. Tale rivestimento è costituito da
segmenti reciprocamente articolati e dunque in grado di
muoversi l'uno rispetto all'altro, per opera dei muscoli su
essi applicati. L'esempio più semplice di esoscheletro è
rappresentato dalla conchiglia di un Mollusco, la quale può
essere costituita da due parti speculari e simmetriche
definite valve (ad esempio quelle delle vongole) oppure da
una struttura unica (ad esempio il guscio delle chiocciole).
Alcune forme marine (a conchiglia bivalve o unica) hanno un
esoscheletro costituito da proteine rinforzate da cristalli di
carbonato di calcio; il prodotto complessivo risulta simile a
un materiale calcareo per consistenza. La conchiglia può
raggiungere spessori e dimensioni considerevoli,
conferendo dunque una protezione integrale nei confronti
dei predatori. Diversamente, l'esoscheletro delle chiocciole
terrestri è privo di sali minerali e risulta estremamente
fragile. Le dimensioni delle conchiglie dei molluschi
aumentano nel tempo, assecondando lo sviluppo
dell'animale, e tale evoluzione dimensionale è
documentata dalla presenza di caratteristici anelli di
accrescimento. Occorre tener presente che il corpo dei
molluschi dispone anche di uno scheletro interno
(idrostatico), il quale viene utilizzato per la locomozione,
mentre quello esterno svolge un ruolo protettivo. Alcuni
pettinidi (i noti bivalvi dotati di una conchiglia ondulata
secondo linee radiali) riescono a spostarsi, sia pure in modo
non direzionale, impiegando un peculiare meccanismo a
reazione, espellendo cioè rapidamente l'acqua mediante
rapidi e ripetuti movimenti di apertura e di chiusura delle
valve.
L'esoscheletro più complesso si rinviene negli Artropodi
(nei crostacei e negli insetti per esempio) dove si descrive
un rivestimento completo e articolato, appendici comprese.
Ogni elemento dell'esoscheletro viene prodotto da una
definita area di epitelio tegumentale; il suo materiale
costitutivo viene definito cuticola e di regola è a sua volta
rivestito da uno strato ceroso. Il piano cuticolare subisce un
processo di sclerotizzazione che lo rende molto rigido ma
che non coinvolge le zone di articolazione tra le diverse
placche dell'esoscheletro, in modo tale che la flessibilità sia
comunque garantita. Il rivestimento comprende uno strato
interno, l'endocuticola, che è relativamente spesso e
rappresenta il principale costituente dell'esoscheletro, e
un piano esterno, l'epicuticola, sul quale viene deposta la
cera per proteggere l'animale dai rischi della
disidratazione. L'endocuticola rappresenta la parte
dell'endoscheletro più caratteristica, composta da proteine
e da un polisaccaride contenente azoto; nei crostacei marini
sono inoltre presenti sali di calcio insolubili. Lo spessore
dell'esoscheletro cuticolare risulta variabile a seconda dei
diversi gruppi di artropodi e, nel singolo animale, a
seconda delle diverse porzioni del corpo; in questo modo il
rivestimento risulta adeguato a molteplici esigenze. I
muscoli si inseriscono sulla superficie interna degli elementi
cuticolari e ne consentono il movimento in corrispondenza
delle articolazioni (▶figura 2).
Figura 2. L'esoscheletro degli insetti Come nei vertebrati, ma partendo da una posizione inversa, i muscoli si applicano ai segmenti scheletrici e ne consentono i movimenti reciproci in corrispondenza delle articolazioni.
Il ruolo preminente dell'esoscheletro, quello che ne ha
decretato il successo evolutivo, corrisponde alla protezione
che garantisce alle porzioni molli del corpo; esso è tuttavia
esposto ai rischi di abrasione o di rotture.
Il maggior limite dell'esoscheletro degli artropodi è dovuto alla
sua incapacità di accrescersi, e per tale motivo lo sviluppo
dimensionale dell'animale comporta la necessità di una muta,
la perdita cioè del vecchio esoscheletro e la formazione di
uno nuovo e più ampio. La muta espone l'animale a una
serie di rischi, poiché il nuovo esoscheletro richiede un
certo tempo prima di sclerotizzarsi. In questo periodo il
corpo rimane scarsamente protetto, ed è inoltre incapace di
muoversi celermente, poiché i muscoli, non potendo
disporre di stabili punti di attacco, non sono in grado di
sviluppare un'adeguata tensione. I granchi che hanno da
poco compiuto la muta e hanno un carapace tenero
vengono considerati un piatto molto appetibile.
Molti vertebrati possedevano e altri possiedono un efficiente
scheletro superficiale: i pesci ossei attuali dispongono di
scaglie ossee che derivano dagli estesi tavolati cutanei
presenti nei loro progenitori arcaici.
Tra i rettili ricordiamo i cheloni (le tartarughe e le forme
affini) e i loricati (i coccodrilli e gli alligatori), che presentano
uno spesso piano osseo al di sotto dello strato corneo
della pelle.
Anche l'armadillo, tra i mammiferi, dispone di una
corazzatura ossea cutanea, per molti versi simile a quella
dei cheloni.
In tutti questi casi parliamo di scheletro superficiale,
evitando l'attributo esterno; occorre infatti precisare
che l'esoscheletro dei vertebrati è contenuto nello
spessore del derma (lo specifico connettivo cutaneo)
e quindi è localizzato al di sotto dell'epidermide e
non al di sopra dì questa.
L’endoscheletro dei vertebrati.
Si ritiene di solito che l'endoscheletro o più
correntemente l'apparato scheletrico dei vertebrati
consista in una serie di strutture ossee. In realtà
esistono altri due tipi di tessuto di sostegno che sono
di complemento o addirittura alternativi all'osso: il
tessuto cordale e la cartilagine, i quali formano
rispettivamente la corda dorsale e segmenti scheletrici
più o meno complessi. Dal punto di vista dello
sviluppo, sia il tessuto cordale che quello cartilagineo
danno origine a strutture per lo più effimere, che nei
vertebrati superiori possono essere soppiantate in
misura variabile da formazioni ossee. Tuttavia, almeno
in una fase della vita dell'organismo, la corda e la
cartilagine sono presenti e funzionanti. Residui
cordali più o meno consistenti sì rinvengono tra le
vertebre degli adulti, e porzioni scheletriche
cartilaginee permangono ancora nello scheletro dei
vertebrati, là dove la robustezza deve coniugarsi con la
flessibilità e con l'elasticità. Esiste inoltre un'intera
classe di vertebrati acquatici, i condroitti o pesci
cartilaginei (comprendenti squali e razze), il cui
endoscheletro è totalmente cartilagineo, per una
probabile riduzione secondaria. Vediamo ora in
dettaglio alcune caratteristiche strutturali delle
formazioni che costituiscono l'endoscheletro dei
vertebrati.
La corda dorsale.
La corda dorsale rappresenta una struttura assile di
sostegno posta ventralmente rispetto al sistema
nervoso centrale e dorsalmente al canale alimentare,
secondo una posizione che viene acquisita nel corso
delle fasi avanzate della gastrulazione.
Nei vertebrati è di regola presente solo in fasi
precoci dello sviluppo, negli embrioni e,
eventualmente, nelle larve; inoltre la sua estensione è
limitata in questo gruppo al tronco. Nei vertebrati
primitivi la corda dorsale rappresenta ancora la
fondamentale struttura di sostegno del tronco, e su
essa prendono attacco i muscoli segmentali che
promuovono il nuoto (▶figura 3).
. Figura 3. Lo scheletro assile della lampreda. La corda dorsale di questo primitivo vertebrato è costituita da cellule notevolmente idratate e da robuste guaine connettivali esterne. Si tratta di un organo assile sufficientemente rigido da fornire un efficace attacco ai muscoli segmentali.
Da un punto di vista strutturale, la corda appare
come un organo cilindrico costituito da una polpa di
voluminose cellule poliedriche con ampie superfici a
mutuo contatto; tali elementi sono separati,
mediante un piano di elementi più piccoli, da una
guaina di rivestimento, fibrosa ed elastica. Le
prestazioni meccaniche della corda sono strettamente
correlate alle caratteristiche funzionali e strutturali
delle cellule della polpa, che presentano un notevole
stato di imbibizione, mantenuto contro gradiente, e
quindi con spesa di energia. Questo turgore, al quale
si oppone la guaina periferica, determina le
contemporanee caratteristiche di rigidità e
flessibilità; tali qualità meccaniche fanno della corda
una struttura scheletrica adeguata per l'attacco dei
deboli muscoli di organismi acquatici in grado di
nuotare solo con mediocre velocità. La sostituzione
della corda da parte di segmenti scheletrici (le
vertebre), rappresenta nei vertebrati adulti un
fenomeno generale; tuttavia permangono ancora
gruppi di pesci ossei arcaici con estesi residui
cordali, e persino nell'uomo rudimenti di tale organo
si ritrovano tra vertebra e vertebra. Nell'anfiosso (un
cordato affine ai vertebrati) la corda dorsale presenta
una struttura peculiare, essendo costituita da
particolari cellule muscolari. Queste subiscono una
stimolazione nervosa continua e sono costantemente
in contrazione (o in tono), una condizione che
conferisce rigidità a tale corda muscolare.
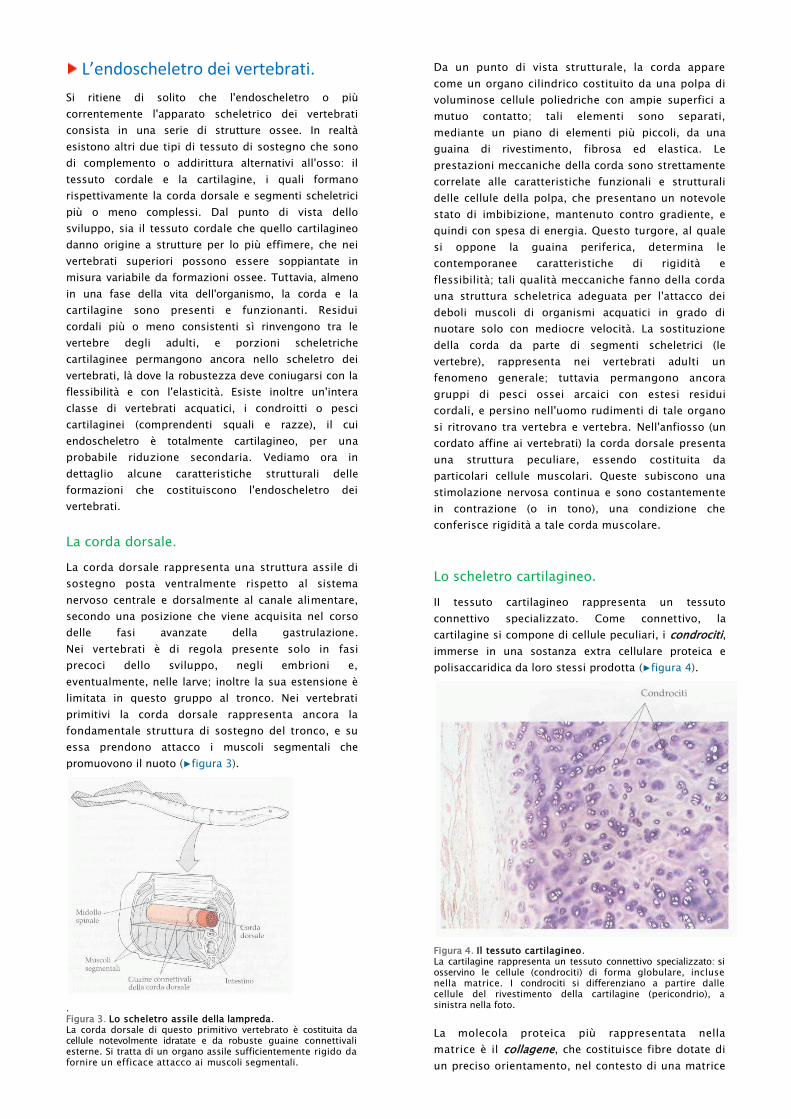
Lo scheletro cartilagineo.
II tessuto cartilagineo rappresenta un tessuto
connettivo specializzato. Come connettivo, la
cartilagine si compone di cellule peculiari, i condrociti,
immerse in una sostanza extra cellulare proteica e
polisaccaridica da loro stessi prodotta (▶figura 4).
Figura 4. Il tessuto cartilagineo. La cartilagine rappresenta un tessuto connettivo specializzato: si osservino le cellule (condrociti) di forma globulare, incluse nella matrice. I condrociti si differenziano a partire dalle cellule del rivestimento della cartilagine (pericondrio), a sinistra nella foto.
La molecola proteica più rappresentata nella
matrice è il collagene, che costituisce fibre dotate di
un preciso orientamento, nel contesto di una matrice

con prerogative di gel. Queste caratteristiche
strutturali sono alla base delle prestazioni
meccaniche della cartilagine, che consistono in una
buona resistenza alle sollecitazioni meccaniche e, in
particolare, nella flessibilità e nella notevole resilienza;
quest'ultimo termine definisce la capacità di
sopportare urti improvvisi senza subire fratture.
Nello scheletro dei vertebrati adulti la cartilagine è
dunque presente in particolari distretti corporei che
sono soggetti a notevoli sollecitazioni meccaniche,
fondamentalmente in corrispondenza delle
articolazioni interossee, dove tuttavia non si
descrivono veri elementi scheletrici cartilaginei, ma
semplici strutture di rivestimento. Altrove, come
nella laringe e nella trachea, il tessuto cartilagineo
forma invece strutture a morfologia definita,
interpretabili come segmenti scheletrici autonomi.
Come abbiamo già detto, la cartilagine costituisce lo
scheletro dell'embrione sino a quando essa non viene
sostituita dal tessuto osseo; come vedremo tra poco,
esiste un'intera categoria di elementi ossei dello
scheletro caratterizzati proprio da questa
derivazione.
Lo scheletro osseo.
II tessuto osseo corrisponde fondamentalmente a un
connettivo dotato di una sostanza extracellulare
ancora più specializzata di quella che caratterizza la
cartilagine. Tale sostanza contiene non solo fibre di
collagene, ma anche cristalli costituiti prevalentemente
da fosfato di calcio, i quali sono insolubili e con-
feriscono all'osso la caratteristica rigidità e la
notevole robustezza. Per la sua componente salina,
l'osso rappresenta la banca del calcio dell'intero
organismo, poiché la quantità degli ioni di tale
elemento fissata nella sua matrice intercellulare
risulta in equilibrio con quella presente in soluzione
nel liquido interstiziale e nel sangue. Tale equilibrio è
garantito dall'azione antagonista di due ormoni; la
calcitonina e il paratormone. In condizioni normali il
prelievo del calcio non è mai comunque tale da
compromettere la consistenza e l'integrità dell'osso.
Il ruolo regolativo svolto dall'osso nei confronti del
contenuto del calcio ematico introduce il concetto
della natura dinamica delle strutture scheletriche, le
quali sono soggette a un continuo rimo del lamento.
Due linee cellulari specifiche partecipano al turnover
dell'osso: gli osteoblasti e gli osteoclasti. I primi
depositano la matrice extracellulare sul segmento
scheletrico (prevalentemente sulla sua superficie)
restando imprigionati nel loro stesso prodotto. In
questa situazione cessa la deposizione della matrice
ossea, mentre gli osteoblasti sopravvivono accolti in
lacune e assumono il nome di osteociti. Nonostante la
notevole quantità di matrice interposta tra queste
cellule, che di norma appaiono ben distanziate, esse
rimangono in connessione reciproca tramite sottili
prolungamenti citoplasmatici contenuti in canalicoli
ossei. Sembra che questi rapporti tra gli osteociti
svolgano un ruolo determinante ai fini della
regolazione del turnover metabolico dell'osso. Gli
osteoclasti, che rappresentano le cellule coinvolte nel
riassorbimento del tessuto osseo, appartengono pro-
babilmente alla stessa linea cellulare dei macrofagi. Si
tratta di elementi plurinucleati in grado di creare, nel
contesto della matrice ossea, spazi di erosione e
tunnel, all'interno dei quali penetrano, al loro
seguito, gli osteoblasti, per deporre nuovo materiale
osseo (▶figura 5).
Figura 5. Il turnover del tessuto osseo. Sopra: L’osso viene continuamente rimodellato per azione degli osteoclasti, che hanno il compito di demolirlo, e degli osteoblasti, che invece depongono nuova matrice ossea; questi ultimi rimangono intrappolati nella sostanza prodotta dalla propria attività, trasformandosi in osteociti.
Non è ancora ben noto come questa cooperazione sia
coordinata ma si ritiene che un ruolo notevole spetti
alle sollecitazioni meccaniche che agiscono sui diversi
distretti dello scheletro. Gli astronauti che
permangono lungamente in assenza di gravita sof-
frono di processi di decalcificazione, mentre gli atleti
presentano un notevole irrobustimento delle ossa
degli arti sollecitati. Questi fenomeni si verificano
anche nel corso dei processi riparativi, nel caso di
fratture degli arti inferiori. Le ossa dell'arto ingessato
si assottigliano, mentre quelle della gamba sana, che
sopportano un maggior peso, si irrobustiscono. La
mandibola degli anziani che hanno perso i denti
subisce sollecitazioni diverse rispetto alle condizioni
normali e dunque viene notevolmente rimodellata
(▶figura 6).
Figura 6. Gli effetti della perdita dei denti sulla mandibola.Sotto: In seguito alla caduta dei denti vengono a mancare le forze di compressione che agiscono sulla mandibola; ne consegue un processo di riassorbimento del tessuto osseo che modifica la forma di questo elemento scheletrico, L'immagine mostra due mandibole umane di individui di diversa età.

Le diverse varietà di tessuto osseo.
Le ossa vengono divise in due categorie, sulla base
delle diverse modalità di formazione. Le ossa di
rivestimento si sviluppano direttamente nello
spessore di un tessuto connettivo membranoso
localizzato al di sotto dell'apparato cutaneo, mentre
le ossa di sostituzione derivano secondariamente da
modelli cartilaginei. Esempi di ossa derivate da
materiale membranoso sono gli elementi scheletrici
della volta del cranio dei vertebrati e dell'uomo,
mentre le ossa degli arti si sviluppano da modelli
preformati in cartilagine, di aspetto simile ai
segmenti scheletrici definitivi. Anche la base del
cranio è comunque costituita da parti ossee derivate
da cartilagine. L'ossificazione dei modelli cartilaginei
può essere un processo complicato; nelle ossa lunghe
del braccio e della gamba, per esempio, si descrivono
tre centri di ossificazione: uno centrale, nel fusto del
segmento scheletrico (la diafisi) e due localizzate in
corrispondenza dell'una e dell'altra estremità o
epifisi del segmento stesso (▶figura 7). Il processo di
ossificazione si accompagna a quello di allungamento
per tutto il tempo in cui permane un anello di
cartilagine interposto tra la diafisi e ciascuna epifisi
(metafisi). Tale struttura anulare, contenente elementi
cellulari in proliferazione, viene definita cartilagine di
coniugazione (o di accrescimento).
Figura 7. L’ossificazione e la crescita di un osso lungo. Le ossa lunghe compaiono nell'embrione come modelli cartilaginei miniaturizzati , simili per morfologia ai corrispondenti segmenti scheletrici definitivi. L'ossificazione inizia a partire dalla dialisi e secondariamente compare anche in corrispondenza delle due epifisi. Il tessuto cartilagineo che permane ai contini tra i tre centri di ossificazione (cartilagini di accrescimento) garantisce l'allungamento dell'osso, sino a che non sarà coinvolto esso stesso dal processo di trasformazione.
Anche nelle ossa di rivestimento del cranio i processi
di accrescimento continuano parallelamente
all'ossificazione; l'arresto della crescita ossea in
questo caso avviene al momento in cui i margini delle
aree di ossificazione limitrofe si incontrano. Le
fontanelle, riconoscibili al tatto sulla calotta cranica
dei bambini, rappresentano zone in corrispondenza
delle quali non è ancora giunta l'ossificazione. Dal
punto di vista strutturale l'osso può essere compatto
o spugnoso: nel primo caso, come dice l'attributo
stesso, l'architettura interna risulta molto robusta; nel
secondo caso la struttura si presenta complessamente
trabecolata e caratterizzata dalla presenza di
numerose cavità, le quali tuttavia non compromettono
la robustezza di tale varietà ossea. La struttura
complessiva di un segmento scheletrico osseo è
correlata alla posizione che esso occupa
nell'organismo e al ruolo che svolge, ma in ogni caso
componenti compatte e componenti spugnose
coesistono.
Per esempio, la diafisi di un osso lungo di un arto
appare conformata come un cilindro cavo contenente
il midollo, ma le estremità presentano una struttura
spugnosa (▶figura 8).
Figura 8. Architettura interna dell’osso. Le ossa sottoposte a intense sollecitazioni meccaniche dispongono di norma di regioni a struttura compatta e di altre a struttura spugnosa. L'osso compatto si ritrova nella diafisi delle ossa lunghe, che appaiono conformate come un cilindro cavo, mentre la componente spugnosa è presente nelle epifisi.
Nonostante la sua leggerezza, l'osso spugnoso risulta
molto solido, poiché la rete tridimensionale interna
costituisce una travatura disposta secondo le linee di
forza che vi agiscono e può sopportare compressioni
elevate. La diafisi, conformata come un tubo dalla
spessa parete costituita da osso compatto, può
opporsi efficacemente alle compressioni e alle
sollecitazioni che comportino una curvatura. La
tecnologia umana e la natura utilizzano ampiamente
strutture tubulari cave come elementi costruttivi,
leggeri ma solidi. In un cilindro pieno che si curva, un
versante è sottoposto a uno stiramento mentre
l'altro risulta compresso. Ambedue gli effetti
determinano una risposta in opposizione alla forza che
causa la curvatura. Un cilindro cavo di pari diametro
presenta la medesima risposta alla curvatura,
nonostante sia meno pesante (▶figura 9).
Figura 9. Caratteristiche meccaniche dell’osso. Un cilindro pieno (in alto) e un tubo cavo di pari diametro (in basso) presentano lo stesso comportamento in risposta alle forze di tensione e a quelle di compressione; la struttura cava risulta ovviamente più leggera.

Dunque la porzione interna di un cilindro pieno non
concorre in alcun modo alla resistenza nei confronti
delle sollecitazioni che tendano a piegarlo. L'osso
compatto dei mammiferi è formato da unità
strutturali chiamate sistemi di Havers od osteoni.
Ogni osteone consiste di un insieme di sottili cilindri
ossei concentrici e di diametro variabile, tra i quali
sono scavate le lacune ossee contenenti gli osteociti.
AI centro del complesso dei cilindri ossei c'è un
canale contenente vasi sanguigni. Gli osteociti di un
osteone si mantengono in reciproca connessione, ma
tali rapporti non coinvolgono le cellule di osteoni
diversi, a causa della presenza di linee cementanti.
Queste delimitano ogni sistema di Havers e di tatto
arrestano il percorso dei processi citoplasmatici degli
osteociti alla periferia dell'osteone. Le linee
cementanti svolgono un ruolo fondamentale dì
protezione nel caso di traumi, poiché interrompono
le linee di frattura.
L’organizzazione dell’endoscheletro dei
vertebrati.
Nella maggior parte dei vertebrati lo scheletro interno
presenta un piano organizzativo comune illustrato
dalla (▶figura 10), riferita a un pesce osseo e a un
mammifero. Secondo uno schema generalizzato, nei
vertebrati troviamo uno scheletro assile e uno
scheletro appendicolare.
La prima porzione si compone di un cranio e di una
colonna vertebrale, ambedue coinvolti in funzioni
protettive e motorie.
Il cranio protegge organi di senso ed encefalo ed è
implicato tanto nei movimenti della mascella e della
mandibola (quando presenti), quanto nei movimenti
respiratori dell'apparato branchiale o dei suoi
derivati.
La colonna vertebrale offre una protezione al midollo
spinale e da attacco ai muscoli del tronco, i quali
possono partecipare in modo più o meno deter-
minante ai movimenti dell'organismo.
Nella maggioranza dei vertebrati lo scheletro assile è
completato da una gabbia toracica costituita dalle
coste, articolate con la colonna vertebrale; nei
vertebrati terrestri o tetrapodi la gabbia toracica è
completata dallo sterno sul quale converge un certo
numero di coste.
La porzione appendicolare comprende le pinne pari,
nei pesci ossei e in quelli cartilaginei, o gli arti nei
tetrapodi. Le prime svolgono un ruolo
prevalentemente direzionale o stabilizzante rispetto
al nuoto, promosso dai muscoli della porzione
caudale del tronco. I secondi di regola sono
responsabili dei movimenti sul terreno, tranne i casi
di riduzione o di scomparsa secondarie degli arti,
come avviene nei serpenti. Molti tetrapodi hanno
conseguito una postura bipede, specializzando gli
arti posteriori ( in tal caso definibili come inferiori)
per la deambulazione sul terreno, e quelli anteriori
per il volo, la prensione e/o la manipolazione. Lo
scheletro appendicolare risulta connesso più o meno
direttamente con quello assile. Nei pesci tale
connessione coinvolge esclusivamente la coppia delle
pinne anteriori, mentre quelle posteriori sono
semplicemente applicate alla muscolatura. Nei
tetrapodi, in cui ambedue le coppie dì arti sono
collegate in qualche modo alla colonna vertebrale,
quella posteriore mantiene un rapporto più stabile con
lo scheletro assile. In tutti i vertebrati, queste
connessioni sono garantite da particolari complessi
scheletrici definiti cinture o cingoli. Parlando dei
vertebrati terrestri, il cingolo anteriore è noto come
cintura toracica o scapolare, quello posteriore viene
definito cintura pelvica. Una caratteristica dello
scheletro dei tetrapodi consiste nel fatto che la
porzione dorsolaterale della cintura pelvica si
connette sui lati del corpo con una o più vertebre
definite sacrali. Questo processo di sacralizzazione
garantisce una connessione efficiente tra gli arti
posteriori, che sono quelli più impegnati nella
deambulazione, e la colonna vertebrale. La cintura
scapolare non si connette direttamente con la colonna
vertebrale ma si appoggia semplicemente sulla gabbia
toracica e può prendere eventualmente rapporto con
lo sterno.
Figura 10. L'endoscheletro dei vertebrati. L’endoscheletro dei vertebrati è riconducibile a un comune piano organizzativo Sono posti a confronto lo scheletro di un pesce osseo evoluto (la perca) di un mammifero (il ratto). Si osservi come nella colonna vertebrale del ratto sia riconoscibile una ripartizione topografica più complessa. Nel pesce sono presenti componenti scheletriche esclusive dei vertebrati acquatici: le pinne impari (dorsali, caudali e anale) mentre, tra le pinne pari, le posteriori (pelviche) si sono portate secondariamente a livello delle anteriori (pettorali). Nel ratto, infine, la gabbia costale si chiude ventralmente in corrispondenza dello sterno.

Lo scheletro assile.
2. Il sistema scheletrico dell’uomo.
In termini generali, l'architettura dello scheletro umano (▶figura 11) corrisponde pienamente a quella di un vertebrato
terrestre, con le uniche specializzazioni correlate alla posizione bipede e alla presenza di mani prensili. Ambedue queste
caratteristiche rappresentano comunque specializzazioni non eccezionali e sono presenti anche negli altri primati più evoluti,
in particolare nelle grandi scimmie (quali il gorilla, lo scimpanzé e l'orango). In questa breve trattazione dell'osteologia
umana saranno pertanto utilizzati i criteri di suddivisione dello scheletro già enunciati a proposito dei vertebrati, individuando
una sezione assile e una appendicolare. Saranno inoltre descritte le cinture, che svolgono una funzione di raccordo tra asse
scheletrico e ossa degli arti; tale connessione non deve essere intesa in senso puramente statico, poiché le cinture forniscono
un adeguato supporto per l'attacco di muscoli coinvolti direttamente nei movimenti degli arti. Il criterio del confronto con lo
schema fondamentale dei vertebrati, sarà sempre presente, poiché esso rappresenta, come paradigma generale, uno strumento
adeguato per comprendere la grande varietà degli adattamenti presenti nello scheletro umano.
Figura 11. Lo scheletro dell’uomo nei suoi principali componenti.

Lo scheletro assile.
L'asse scheletrico dell'uomo presenta la tradizionale
suddivisione in colonna vertebrale e cranio, ai quali
si aggiunge tradizionalmente lo scheletro del torace,
cioè il complesso dello sterno e delle coste. La
funzione ancestrale dello scheletro assile primitivo
(sostanzialmente la corda e/o la colonna vertebrale) è
stata verosimilmente quella di dare attacco ai muscoli
del tronco che consentivano il movimento
nell'ambiente acquatico dei primitivi vertebrati
pisciformi, e contemporaneamente di offrire una
protezione al sistema nervoso centrale. I rapporti tra
le strutture scheletriche e quelle nervose hanno
guidato anche le tappe successive dell'evoluzione
dello scheletro assile. portando alla comparsa di una
porzione cefalica specializzata, il cranio. Nella regione
della testa compaiono infatti precocemente organi di
senso essenziali per l'animale e, secondariamente,
l'encefalo. Nel corso della filogenesi sì presentano
dunque ben presto le esigenze di garantire la
protezione a strutture così "nobili” e si organizza un
cranio neurale o neurocranio. La regione cefalica
dell'animale è inoltre impegnata nei processi
dell'alimentazione e della respirazione; tali funzioni
richiedono strutture mobili e sostegni scheletrici, che
nel loro insieme costituiscono il cranio viscerale o
splancnocranio. La stessa colonna vertebrale, che
svolge un essenziale ruolo di supporto meccanico,
protegge con porzioni specializzate dorsali il
midollo spinale. Seguendo un criterio topografico
cefalo-caudale prenderemo ora in considerazione il
cranio, la colonna vertebrale e la gabbia toracica.
Il cranio.
Nel cranio dei mammiferi e dell'uomo sia la consueta
ripartizione in ossa di sostituzione e in ossa di
rivestimento che quella in porzioni neurali (e
sensoriali) e in porzioni viscerali (o splancniche) si
dimostrano troppo rigide per dare una descrizione
esauriente della complessità di tale struttura
scheletrica, poiché nel corso dello sviluppo si
verificano fenomeni di fusione che alterano
notevolmente questo schema generale. Per una
visione complessiva del cranio umano possiamo
riferirci alle (▶figura 12, 13, 14, 15) che mostrano
rispettivamente: la visione frontale, la visione laterale,
la visione superiore e la visione basale (vista
internamente ed esternamente).
Comunque, per comodità di studio, il cranio può essere
può essere suddiviso in due porzioni:
il neurocranio che protegge l’encefalo
lo splancnocranio che costituisce lo scheletro della
faccia.
NEUROCRANIO.
La porzione del cranio che riveste dorsalmente e
lateralmente l'encefalo è costituita in senso antero-
posteriore dal frontale, dai due parietali, dalle grandi ali
dello sfenoide, dalle squame dei due temporali e da
quella unica dell'occipitale. Tranne le grandi ali dello
sfenoide, si tratta sempre di ossa (o di porzioni di
ossa) di rivestimento, cioè derivate da connettivo
membranoso. La base del cranio, che accoglie la faccia
ventrale dell'encefalo, comprende una porzione
anteriore di rivestimento, corrispondente al frontale,
e poi una serie di ossa preformate in cartilagine.
Tali ossa sono: l'etmoide, lo sfenoide, le parti petrose
dei due temporali (ognuna contenente l'orecchio
interno del proprio lato) e la parte basilare
dell'occipitale.
SPLANCNOCRANIO
E’ la porzione che costituisce lo scheletro della faccia
e comprende i nasali, i lacrimali i mascellari, gli
zigomatici, il vomere, i palatini, la conca nasale
inferiore, e la mandibola, che è l’unico osso articolato
al resto del cranio tramite un’articolazione mobile.
Tutte le altre ossa del cranio sono articolate tramite
articolazioni fisse dette suture.
Figura 12. Visione frontale del cranio.
Figura 13. Visione laterale del cranio.
Figura 14. Visione superiore del cranio.
Figura 15. Base del cranio.
A sinistra la visione interna, a destra la visione esterna.
Tutte le ossa della faccia derivano da connettivo
membranoso e sono dunque di rivestimento.
Compreso tra i due mascellari troviamo il palato
osseo, costituito dagli stessi mascellari e dai palatini.
Tra il palato e la base del cranio è presente il vomere,
che assieme all'etmoide sovrastante, separa la cavità
nasale destra dalla sinistra. Al di sotto del palato
osseo, lo scheletro viscerale è completato dalla
mandibola.
Esistono altre strutture scheletriche derivate dal
primitivo scheletro cartilagineo viscerale
dell'embrione: il martello, l'incudine, la staffa (ossa
pari), l'osso ioide e gran parte delle cartilagini
laringee. 1 primi tre elementi costituiscono la catena
degli ossicini dell'orecchio medio; essi rappresentano
residui dell'ancestrale arco orale cartilagineo degli
antichi vertebrati (il martello e l'incudine) e della
porzione dorsale dell'arco ioideo (la staffa), il quale
nei pesci ossei e in quelli cartilaginei è intercalato tra
lo scheletro orale e lo scheletro branchiale. L'osso
ioide è situato al confine tra il pavimento della bocca
e la faccia ventrale del collo e da attacco a muscoli
della lingua, del pavimento della cavità orale e del
collo stesso; si compone di materiale derivato dalla
porzione ventrale dell'arco ioideo insieme a
contributi del primitivo scheletro branchiale. I residui
degli altri archi branchiali costituiscono gli elementi
cartilaginei dello scheletro della laringe, la quale
rappresenta il primo tratto retrorale delle vie aeree e
pertanto deve essere sempre pervia.
La colonna vertebrale.
Le vertebre umane sono tutte olospondile, cioè
dotate di un corpo completo e robusto (di forma
cilindrica), dal quale si dipartono alcune formazioni
accessorie; queste sono localizzate dorsalmente e
lateralmente, e consistono in un arco neurale
(dorsale) e in due processi trasversi (laterali). L'arco
si prolunga in un processo spinoso e delimita con il
corpo vertebrale il foro omonimo, occupato dal
midollo spinale. 1 processi trasversi si proiettano sui
due lati a partire dalla zona di raccordo tra arco e
corpo, dove sono presenti evidenti incisure. Nella
giustapposizione tra vertebra e vertebra, le incisure
determinano la formazione di una duplice serie di
fori vertebrali, nei quali passano i nervi spinali. Altre
formazioni caratteristiche delle vertebre
corrispondono ai processi articolari che regolano i
loro rapporti e che concorrono a determinarne
l'entità dei movimenti reciproci. Nella colonna
vertebrale dell'uomo si riconosce la ripartizione
topografica tipica dei mammiferi, correlata a una
precisa suddivisione del tronco (▶figura 16).
Figura 16. La colonna vertebrale. A sinistra la visione anteriore, al centro la visione laterale e a destra quella posteriore.
Si descrivono quindi 5 regioni:
7 vertebre cervicali per la regione del collo;
12 vertebre toraciche per l'omonima regione;
5 vertebre lombari per la regione addominale;
5 vertebre sacrali
3-5 vertebre coccigee,

REGIONE CERVICALE:
La prima e la seconda vertebra cervicale sono molto
specializzate e vengono definite rispettivamente
atlante ed epistrofeo. La prima ha il corpo ridotto a
una semiluna, che con l'arco neurale costituisce una
sorta di anello. L'atlante si articola con l'osso
occipitale del cranio consentendo alla testa di oscillare
in senso antero-posteriore rispetto alla colonna
vertebrale. Esso si articola inoltre con un voluminoso
processo (dente) dell'epistrofeo; potendo ruotare
intorno a questo da lato a lato, coinvolge nel suo
movimento lo stesso cranio. In tutte le vertebre
cervicali i processi trasversi sono fusi con rudimenti
costali, delimitando su ambo ì lati un foro
trasversario, attraverso il quale passano strutture
nervose simpatiche e vasi.
REGIONE TORACICA:
Le vertebre del tratto toracico sono connesse con le
coste e dunque concorrono alla chiusura dorsale
della gabbia toracica. Esse sono relativamente
voluminose e tendono a incrementare le proprie
dimensioni procedendo verso il basso; questo
aumento dimensionale riguarda l'intera colonna
(▶figura 16) e deve essere messo in relazione con
l'aumento progressivo del carico che si verifica
scendendo lungo l'asse vertebrale. Sui processi
trasversi delle vertebre toraciche si applica
un'estremità della costa definita tubercolo, mentre un
altro estremo costale (il capitello) prende connessione
con una piccola faccia articolare scavata tra due corpi
vertebrali consecutivi. In corrispondenza
dell'articolazione tra coste e vertebre si individua un
foro trasversario mediocremente definito.
REGIONE LOMBARE:
Nelle vertebre lombari i processi trasversi hanno
incorporato, come avviene nella regione cervicale, i
rudimenti delle coste corrispondenti, e si presentano
come formazioni ampiamente espanse sui due lati.
REGIONE SACRALE:
L'osso sacro rappresenta il risultato di una fusione
che ha coinvolto cinque vertebre, ma due di queste
sono state cooptate secondariamente, nel processo di
sacralizzazione. Ricordiamo che con questo termine si
intende il rapporto che si stabilisce nei vertebrati
terrestri tra la cintura pelvica e la colonna vertebrale.
L'origine segmentale di questa ampia struttura ossea
è ben evidente sia sulla faccia ventrale che su quella
dorsale.
REGIONE COCCIGEA O CAUDALE:
Nell'uomo anche le poche vertebre caudali subiscono
un processo di fusione reciproca, formando una
struttura rudimentale definita coccige. Una formazione
omologa (il pigostilo) si ritrova anche negli uccelli,
mentre negli anfibi degli anuri (che sono privi di coda)
le vertebre caudali si fondono reciproca mente a dare
uno stiletto osseo (l'urostilo).
Poiché, come abbiamo detto, il diametro dei corpi
vertebrali tende ad aumentare in senso cefalo-
caudale, la colonna umana appare conformata come
un tronco di cono. Le possibilità di rotazione, o
comunque dì spostamento, di una vertebra rispetto
alle due adiacenti sono limitate (per evitare danni al
midollo spinale), e tuttavia questo asse scheletrico
risulta adeguatamente flessibile. La colonna vertebrale
umana presenta alcune curvature di natura fisiologica
e altre che rappresentano malformazioni. Tra le
prime, che sono antero-posteriori, se ne descrivono a
concavità anteriore (cifosi) o posteriore (lordosi).
Dall'alto in basso si susseguono: una lordosi
cervicale, una cifosi toracica, una lordosi lombare e
una cifosi sacrococcigea. Le curvature non fisiologiche
si definiscono scoliosi e sono curvature perlaterali (si
sviluppano a destra o a sinistra rispetto all'asse della
colonna vertebrale).
Lo scheletro del torace.
La gabbia toracica costituisce una protezione per
organi interni di vitale importanza come cuore,
polmoni, esofago, trachea, bronchi, grossi vasi, ecc.
Essa è formata posteriormente dai corpi vertebrali
delle vertebre toraciche, anteriormente da un osso
piatto impari e mediano: lo sterno e lateralmente da
dodici paia di ossa lunghe e appiattite: le costole.
Figura 17. La gabbia toracica. In evidenza lo sterno costituito da tre parti e le coste (7 vere, 3 false e 2 fluttuanti); nell'immagine vengono inoltre messi in risalto il livello dell'articolazione del manubrio sternale con la clavicola sinistra e l'angolo diedro formato dal manubrio stesso e dal corpo. Come nelle figure successive, le linee in colore indicano le inserzioni di specifici muscoli.
Le coste.
Nell'uomo alle dodici vertebre toraciche sono annesse
altrettante coppie di coste, segmenti arcuati derivati
da abbozzi cartilaginei, che si estendono
dorsalmente, lateralmente e, in misura variabile,
ventralmente (▶figura 17). Sono composte da un
segmento dorsale (vertebrale), che è ossificato, e da
un segmento ventrale che è invece cartilagineo.

Le prime sette paia di coste sono dette coste vere o
sternali perché raggiungono lo sterno tramite la
porzione costituita da cartilagine.
Le seguenti tre paia sono dette coste false o asternali
perché contraggono un rapporto indiretto con lo
sterno, in quanto si connettono tramite il proprio
segmento cartilagineo a quello delle precedenti.
Le ultime due sono dette coste libere o fluttuanti
perché non si connettono in nessun modo allo sterno.
Esse sono provviste di una modestissima porzione
cartilaginea, hanno uno sviluppo laterale e ventrale
ridotto (soprattutto la dodicesima).
Lo sterno.
Tutti i vertebrati terrestri presentano una struttura
scheletrica impari ventrale, lo sterno, più o meno
ossificato, che risulta spesso connessa con la cintura
scapolare e, a partire dai rettili, anche con numerose
coste. Secondo un piano strutturale che si rinviene
anche nei rettili, lo sterno umano si compone di tre
parti: un manubrio superiore, un corpo intermedio e
un processo xifoideo inferiore (▶figura 17). Gli uccelli
dispongono di uno sterno molto specializzato, di
forma carenata, sul quale si applicano i muscoli
pettorali propulsori del volo. Manubrio, corpo e
processo xifoideo permangono nettamente distinti
da piani di separazione; tale discontinuità è resa
ancor più evidente dal fatto che la seconda e la
settima costa raggiungono rispettivamente lo sterno
proprio al limite tra manubrio e corpo e tra corpo e
processo xifoideo. Nei giovani anche il corpo presenta
una struttura segmentale, poiché i moduli
componenti (gli sternebri) si saldano non prima del
venticinquesimo anno.
Le cinture.
Secondo lo schema che si ritrova in tutti i vertebrati
terrestri, tranne le forme che hanno perso
secondariamente gli arti, l'uomo presenta due
strutture, definite cinture, che servono da raccordo
tra l'osso prossimale di ogni arto (cioè quello più
vicino al corpo) e lo scheletro assile. Delle due cinture
quella superiore, scapolare, non ha rapporti diretti
con la colonna vertebrale, ma può connettersi
ventralmente con lo sterno (come avviene nella nostra
specie), e in tal caso risulta solidale rispetto alla
gabbia costale. In molti mammiferi, in assenza di
clavicole, viene a mancare ogni connessione, anche
indiretta, tra il cingolo scapolare e lo scheletro assile.
La cintura pelvica si connette direttamente con la
regione sacrale della colonna vertebrale, e dunque
conferisce stabilità agli arti inferiori soggetti alle
sollecitazioni meccaniche della deambulazione
bipede.
La cintura scapolare
Nell'uomo il raccordo tra l'arto superiori e lo scheletro
assile è garantito da una coppia di ossa presenti su
ogni lato del corpo: la scapola e la clavicola (▶figura
18).
Figura 18. La cintura scapolare. L'immagine mostra la scapola destra dalla faccia dorsale (a) e la clavicola dello stesso lato vista superiormente (b). Oltre alle inserzioni muscolari, la figura (a) mostra i tratti caratterizzanti della scapola: il solco che accoglie un vaso arterioso, la faccetta articolari per la clavicola e l'impronta dovuta a una parte (conoide) del legamento teso Ira il processo coracoideo e la clavicola. La scapola è derivata da materiale cartilagineo e
risulta adagiata dorso-lateralmente sullo scheletro
toracico. Si tratta dì un osso piuttosto appiattito, con
un'ampia superficie di forma triangolare. Essa è
dotata di un'estesa porzione articolare in
corrispondenza del vertice laterale, la cavità
glenoidea, la quale accoglie la testa dell'omero, l'osso
del braccio. La superficie dorsale della scapola è
percorsa superiormente da un rilievo (la spina) che la
interessa dal margine laterale a quello mediale.
L'estremo laterale della spina viene definito acromion
ed è in rapporto articolare con un estremo della
clavicola. La scapola presenta inoltre un processo
coracoideo, omologo a un elemento scheletrico
indipendente, definito coracoide, assente in tutti i
mammiferi evoluti e piuttosto diffuso negli altri
vertebrati, compresi i mammiferi monotremi. Il
secondo elemento della cintura scapolare umana è
rappresentato dalla clavicola, un osso sagomato a S
che deriva da una matrice membranosa ed è spesso
assente nei vertebrati terrestri, inclusi molti

mammiferi. Le due clavicole sono caratterizzate da un
orientamento quasi trasversale rispetto all'asse della
colonna vertebrale e da una posizione
sottocutanea. Raggiungono l'acromion della scapola
omolaterale (cioè dello stesso lato) con il proprio
estremo acromiale e, sul lato opposto, si connettono,
tramite l'estremo sternale, con il manubrio dello
sterno.
La cintura pelvica
In tutti i vertebrati terrestri tre elementi ossei
preformati in cartilagine si uniscono su ogni lato delle
vertebre sacrali e formano, unendosi con i processi
trasversi di queste, un cingolo scheletrico definito
cintura pelvica. Là dove si uniscono reciprocamente,
le tre ossa delimitano un'ampia cavità che accoglie il
femore, definita acetabolo in quanto ricorda per forma
la tazza in cui i Romani raccoglievano l'aceto. Se si
individuano tre raggi sulla superficie circolare
dell'acetabolo, in modo da ricavare una sorta di Y
rovesciata, si rileva che ognuno dei tre raggi
corrisponde all'asse di un elemento scheletrico di
questa cintura: l'ileo, il pube e l'ischio. Nell'uomo le
tre ossa presentano una posizione peculiare: l'ileo,
che si connette con l'osso sacro, è localizzato su-
periormente, il pube inferiormente e ventralmente e
l'ischio inferiormente e dorsalmente. Nella nostra
specie i tre elementi ossei sono in realtà fusi in
un'unica formazione scheletrica per lato, definita osso
dell'anca (▶figura 19).
Figura 19. L’osso dell’anca. I tre segmenti scheletrici che costituiscono l'anca umana sinistra sono evidenti in questa immagine: l'ileo, l'ischio e il pube. La faccia esterna (o glutea) dell'ileo è divisa in quattro aree da tre linee glutee. Si osservi che il bordo della cavità articolare dell'acetabolo è interrotto in basso in corrispondenza dell'incisura acetabolare.
La sua porzione dorsale iliaca è piuttosto ampia,
mentre al di sotto dell'acetabolo, pube e ischio sono
relativamente sottili e delimitano un'ampia finestra
definita foro otturato; in più, i due elementi pubici si
connettono ventralmente sul piano di simmetria,
realizzando una connessione fibrosa e cartilaginea
definita sinfisi pubica. L'insieme delle due ossa
dell'anca e del sacro delimita una cavità rota come
pelvi o bacino, le cui dimensioni (diametri e assi)
presentano un notevole interesse per l'ostetricia, per
la medicina legale e per l'antropologia (▶figura 20).
Figura 20. La pelvi femminile. La figura mostra i fondamentali diametri di questa struttura: quello antero-posteriore E-F, quello trasverso C-D e quelli o-bliqui A-B e A'-B'.
Lo scheletro appendicolare.
Figura 21. Lo schema generale dell’arto generalizzato di un tetrapode primitivo.
Si confronti con la figura 11 e si rilevino le omologie con gli
arti umani (tra parentesi i nomi delle ossa dell'arto posteriore).
Lo scheletro degli arti umani può essere descritto
secondo lo schema generalizzato di tutti i vertebrati
terrestri. Può sembrare in qualche misura
sorprendente che le ossa del nostro braccio, quelle
dell'avambraccio e quelle della mano ricordino molto
da vicino l'arto anteriore di un tritone. Questa è
tuttavia la realtà, e, se mai, la somiglianza dimostra
ulteriormente come la nostra evoluzione abbia
premiato altre caratteristiche anatomo-funzionali (le
capacità integrative del sistema nervoso centrale per
esempio) piuttosto che quelle legate al movimento
nell'ambiente. La (▶figura 21) illustra lo schema
dell'arto generalizzato di un primitivo vertebrato
terrestre e riporta le corrispondenze rispetto
all'arto superiore e a quello inferiore dell'uomo.
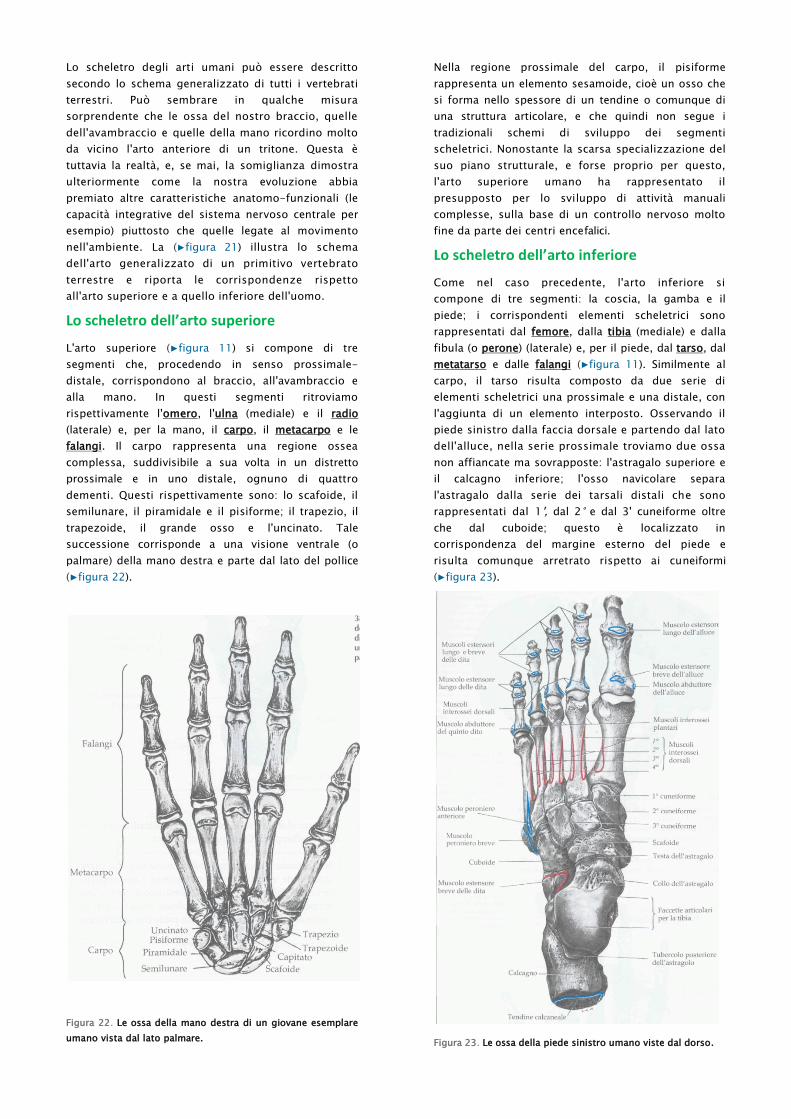
Lo scheletro dell’arto superiore
L'arto superiore (▶figura 11) si compone di tre
segmenti che, procedendo in senso prossimale-
distale, corrispondono al braccio, all'avambraccio e
alla mano. In questi segmenti ritroviamo
rispettivamente l'omero, l'ulna (mediale) e il radio
(laterale) e, per la mano, il carpo, il metacarpo e le
falangi. Il carpo rappresenta una regione ossea
complessa, suddivisibile a sua volta in un distretto
prossimale e in uno distale, ognuno di quattro
dementi. Questi rispettivamente sono: lo scafoide, il
semilunare, il piramidale e il pisiforme; il trapezio, il
trapezoide, il grande osso e l'uncinato. Tale
successione corrisponde a una visione ventrale (o
palmare) della mano destra e parte dal lato del pollice
(▶figura 22).
Figura 22. Le ossa della mano destra di un giovane esemplare
umano vista dal lato palmare.
Nella regione prossimale del carpo, il pisiforme
rappresenta un elemento sesamoide, cioè un osso che
si forma nello spessore di un tendine o comunque di
una struttura articolare, e che quindi non segue i
tradizionali schemi di sviluppo dei segmenti
scheletrici. Nonostante la scarsa specializzazione del
suo piano strutturale, e forse proprio per questo,
l'arto superiore umano ha rappresentato il
presupposto per lo sviluppo di attività manuali
complesse, sulla base di un controllo nervoso molto
fine da parte dei centri encefalici.
Lo scheletro dell’arto inferiore
Come nel caso precedente, l'arto inferiore si
compone di tre segmenti: la coscia, la gamba e il
piede; i corrispondenti elementi scheletrici sono
rappresentati dal femore, dalla tibia (mediale) e dalla
fibula (o perone) (laterale) e, per il piede, dal tarso, dal
metatarso e dalle falangi (▶figura 11). Similmente al
carpo, il tarso risulta composto da due serie di
elementi scheletrici una prossimale e una distale, con
l'aggiunta di un elemento interposto. Osservando il
piede sinistro dalla faccia dorsale e partendo dal lato
dell'alluce, nella serie prossimale troviamo due ossa
non affiancate ma sovrapposte: l'astragalo superiore e
il calcagno inferiore; l'osso navicolare separa
l'astragalo dalla serie dei tarsali distali che sono
rappresentati dal 1', dal 2° e dal 3' cuneiforme oltre
che dal cuboide; questo è localizzato in
corrispondenza del margine esterno del piede e
risulta comunque arretrato rispetto ai cuneiformi
(▶figura 23).
Figura 23. Le ossa della piede sinistro umano viste dal dorso.

Le caratteristiche più notevoli dell'arto inferiore
consistono nella posizione del suo asse, che risulta
parasagittale, cioè parallela al piano di simmetria del
corpo, e nella plantigradia. L'assetto parasagittale
degli arti si realizza compiutamente nel corso della
transizione dai rettili ai mammiferi e coinvolge, sia
pure in misura minore, anche la coppia degli arti
anteriori. Si tratta di un adattamento che consente una
maggiore celerità negli spostamenti, rispetto a quanto
si verifica con arti disposti su un piano ortogonale
rispetto a quello di simmetria. La posizione
parasagittale, riferita al femore umano, prevede la
presenza di una testa, accolta nell'acetabolo e
raccordata al corpo dell'osso (diafisi) tramite un collo,
che forma un angolo ottuso con asse diafisario.
L'assetto plantigrado, cioè il sostegno del corpo
garantito dall'intera serie delle ossa podali dal tarso
alle falangi rappresenta un carattere di primitività.
Tra i mammiferi più evoluti dal punto di vista della
deambulazione, i carnivori poggiano sulle dita (sono
cioè digitigradi), mentre gli unguligradì (i bovini, gli
ovini e i cavalli, per esempio) utilizzano a tal scopo
unghie specializzate definite zoccoli, e da ciò deriva il
loro nome sistematico. Si può pensare che l'assetto
plantigrado sia correlato alla posizione eretta, poiché
offre la più ampia superficie di appoggio possibile agli
arti impegnati a sorreggere il corpo. Tuttavia si
conoscono vertebrati evoluti, quali gli uccelli e alcuni
rettili giganteschi, oggi estinti, i quali hanno
conseguito il bipedalismo poggiando sulle sole dita.
Occorre ancora una volta accettare l'idea che, almeno
dal punto di vista degli adattamenti al movimento
sul terreno, la nostra specie occupa una posizione
mediocre nella serie evolutiva.







