
REVIEW ARTICLE
A historical perspective on protein crystallization from1840 to the present dayRichard Gieg�e
Institut de Biologie Mol�eculaire et Cellulaire, Universit�e de Strasourg et CNRS, France
Keywords
automation; crystal growth; high-throughput;
macromolecular assemblies; methods of
crystallization; microgravity; nucleation;
nucleic acids; protein crystallization; virus
Correspondence
R. Gieg�e, Architecture et R�eactivit�e de
l’ARN, Universit�e de Strasbourg, CNRS,
IBMC, 15 rue Ren�e Descartes, F-67084,
Strasbourg, France
Fax: +33 (0)3 88 60 22 18
Tel: +33 (0)3 88 41 70 58
E-mail: [email protected]
Note
The term ‘Protein’ is often taken as the
generic name for a biological
macromolecule or a macromolecular
assembly. ‘Protein solubility’ can have two
distinct meanings, either the amount of
protein that can be dissolved in a solvent or
the protein concentration in a solution in
equilibrium with a phase containing its
crystalline form. ‘Crystallants’ (often
referred as ‘precipitants’) and ‘nucleants’
are the chemical or physical factors that
promote nucleation and/or crystallization.
(Received 12 July 2013, revised 30 August
2013, accepted 27 September 2013)
doi:10.1111/febs.12580
Protein crystallization has been known since 1840 and can prove to be
straightforward but, in most cases, it constitutes a real bottleneck. This stim-
ulated the birth of the biocrystallogenesis field with both ‘practical’ and
‘basic’ science aims. In the early years of biochemistry, crystallization was a
tool for the preparation of biological substances. Today, biocrystallogenesis
aims to provide efficient methods for crystal fabrication and a means to opti-
mize crystal quality for X-ray crystallography. The historical development of
crystallization methods for structural biology occurred first in conjunction
with that of biochemical and genetic methods for macromolecule production,
then with the development of structure determination methodologies and,
recently, with routine access to synchrotron X-ray sources. Previously, the
identification of conditions that sustain crystal growth occurred mostly
empirically but, in recent decades, this has moved progressively towards
more rationality as a result of a deeper understanding of the physical chemis-
try of protein crystal growth and the use of idea-driven screening and high-
throughput procedures. Protein and nucleic acid engineering procedures to
facilitate crystallization, as well as crystallization methods in gelled-media or
by counter-diffusion, represent recent important achievements, although the
underlying concepts are old. The new nanotechnologies have brought a sig-
nificant improvement in the practice of protein crystallization. Today, the
increasing number of crystal structures deposited in the Protein Data Bank
could mean that crystallization is no longer a bottleneck. This is not the case,
however, because structural biology projects always become more challeng-
ing and thereby require adapted methods to enable the growth of the appro-
priate crystals, notably macromolecular assemblages.
Introduction
The art of crystallization dates back to antiquity and,
for a long time, primarily comprised the growth of salt
crystals by evaporation procedures. Protein crystalliza-
tion is much more recent and appeared in the first half
Abbreviations
2D, two-dimensional; aaRS, aminoacyl-tRNA synthetase (e.g. AspRS for aspartyl-tRNA synthetase, PheRS for phenylalanyl-tRNA synthetase,
etc.); AFM, atomic force microscopy; DLS, dynamic light scattering; HEW, hen egg-white; ICCBM, International Conference on the
Crystallization of Biological Macromolecules; IEF, isoelectric focusing; PAGE, polyacrylamide gel electrophoresis; PDB, Protein Data Bank;
SANS, small-angle neutron scattering; SAXS, small-angle X-ray scattering; TEW, turkey egg-white; TMV, tobacco mosaic virus.
6456 FEBS Journal 280 (2013) 6456–6497 ª 2013 FEBS

of 19th Century, with an initial publication in 1840 on
the observation of crystallites in blood preparations [1],
which in fact were haemoglobin crystals. Over the years,
the diversity of crystallized proteins has expanded,
although crystallization often occurred by chance and
using empirical procedures. For approximately one cen-
tury, crystallization was used as a means of protein
purification and characterization by biochemists and
physiologists. The situation changed when X-ray crys-
tallography entered biology in 1934 after the first X-ray
photograph of a protein crystal was taken [2]. Improve-
ments in crystallization procedures and the fabrication
of crystals suitable for structure determination arose in
parallel with advances in X-ray crystallographic meth-
ods and the ambition of structural biologists who were
seeking to image the macromolecular components of
living organisms. This became possible as a result of
interdisciplinary efforts merging biochemistry/molecu-
lar biology, chemistry, physics and engineering, which
gradually transformed the field of protein crystallization
into a scientific discipline of its own. I have named this
discipline ‘crystallogenesis’ [3], where the aim is to
understand and control crystal growth and quality; note
that a German version, ‘Krystallogenese’, was already
proposed in the 19th Century by different individuals,
such as Preyer [4]. The literature on biocrystallogenesis
is manifold. The present review restricts itself to a few
introductory references on historical [3,5–7] as well as
on methodological and physicochemical [8–16] aspectsand to a selection of most significant research articles
and focused reviews. More citations on facts listed in
Tables and Figures are provided in Data S1 to S10.
Additional bibliographic sources, particularly books,
reviews and International Conference on the Crystalli-
zation of Biological Macromolecules (ICCBM) Pro-
ceedings, are given in Data S11.
The present review is divided into three sections
describing how biocrystallogenesis emerged and became
a mature field, as well as how it became seminal for mod-
ern structural biology. They cover: (a) the period of phys-
iological and colloidal chemistry before the birth of
protein X-ray crystallography; (b) the early years of
structural biology when conventional methods of protein
crystallization were established; and (c) the years of more
recent technologies and structural genomics. The conclu-
sion outlines perspectives and sketches a few applications
beyond the field of structural biology (e.g. in medicine).
The time of physiology and chemistry(1840–1934)
In the 19th and early 20th Centuries, knowledge on
proteins was elusive and the name ‘protein’ (coined by
Berzelius in 1838) was not of universal use in biology
and chemistry. Terms such as ‘Proteid/Eiweissk€orper’
substances, ‘albumineous’ material or ‘colloids’ were
often employed for these mysterious substances. How-
ever, during this period, a few visionary physiologists,
chemists and physico-chemists established the corner-
stones of modern biology, notably structural biology,
when they worked out protocols leading to the pro-
duction of crystalline proteins. The basic methods of
protein crystallization were established and the essen-
tial physico-chemical properties of proteins discovered.
Crystallinity of haemoglobin and plant globulins
In 1840, Friedrich Ludwig H€unefeld published a book
entitled Der Chemismus in der thierischen Organisation
(Chemical Properties in the Animal Organization) in
which he reported (p. 160 and 161) how he acciden-
tally discovered the formation of crystalline material in
samples of earthworm blood held under two glass
slides and occasionally observed small plate-like crys-
tals in desiccated swine or human blood samples [1].
These were crystals of ‘haemoglobin’, a name coined
1864 by Felix Hoppe-Seyler for the ‘colorant substance
of blood’ [17]. In the following years, and likely even
before H€unefeld, many scientists observed haemo-
globin crystals when examining various animal tissues
or animal blood (e.g. Julius Budge, Otto Funke,
Albert von K€olliker, Karl G Lehmann, Franz Leydig
and Karl Reichert) but except Funke did not investi-
gate further the properties of these crystals [4].
In 1855, Theodor Hartig discovered a second family
of crystalline proteids in the gluten flour ‘Klebermehl’
from the Bertholletia excelsa Brazil nut [18]. Soon,
‘crystalloids’, as they were named, of globulins were
described by several authors in extracts of other plant
seeds (e.g. from Avena, Camelia, Crocus, Croton and
Ricinus), notably by Heinrich Ritthausen [19] and
mostly by Thomas B. Osborne [20] who knew and
extended Ritthausen’s work. By 1889, when Osborne
started his thorough biochemical work on plant globu-
lins, his main interest was to prepare pure specimens
of globulins by employing all of the available methods
at the time (particularly crystallization) to ensure
homogeneity of the preparations. As a result, he
obtained crystals of several globulins (two examples
are provided in Fig. 1A) and assigned them specific
designations; for example, ‘excelsin’ for the globulin
from Brazil nut (an allergen presently known as the
Ber e 2 protein) [21] and ‘edestin’ or ‘avenalin’ for
those from hemp seeds or oat kernels [20]. In 1907,
Osborne published a monograph summarizing his
investigations (revised in 1924) in which he described
FEBS Journal 280 (2013) 6456–6497 ª 2013 FEBS 6457
R. Gieg�e Protein crystallization for structural biology

procedures to obtain crystals that are based, amongst
others, on protein extractions from warm salt solutions
(40–60 °C) followed by slow cooling to room tempera-
ture; for further details, see the online version of the
original 1924 publication [22].
Deliberate protein crystallizations in the 19th and
early 20th Centuries
After the seminal findings by H€unefeld and Hartig,
many other physiological chemists and botanists tried
to deliberately produce crystals of haemoglobin and
plant globulins using more controlled protocols [6].
Thus, in 1851, Funke described how to grow human
haemoglobin crystals by successively diluting red blood
cells with solvents such as pure water, alcohol or ether,
followed by slow evaporation of the solvent from the
protein solution [23]. This was the first use of organic
solvents in protein crystallization. In 1871, the English-
born physiologist William T. Preyer, Professor at Uni-
versity of Jena, published a book entitled Die Blutk-
rystalle (The Crystals of Blood), reviewing the features
of haemoglobin crystals from ~ 50 species of mammals,
birds, reptiles and fishes [4]. Franz Hofmeister entered
the theater of crystal science in 1890 when he crystal-
lized hen egg-white (HEW) albumin [24].
The interest in haemoglobin crystals did not decline
in the 20th Century and was first highlighted in 1909
when the physiologist Edward T. Reichert, together
with the mineralogist Amos P. Brown, published an
impressive treatise on the preparation, physiology and
geometrical characterization of haemoglobin crystals
from several hundreds animals, including extinct
species such as the Tasmanian wolf (Thylacyanus cy-
nocephalus) [25] (Fig. 1B,C). The crystallization of
other proteins was also actively pursued in the first
half of the 20th Century. As was common practice in
chemistry, crystallization became a powerful step in
purification protocols. Examples are the crystalliza-
tion of animal and plant globulins (e.g. various serum
albumins and canavalin from jack beans), the crystal-
lization of a plant lectin (concanavalin A), the crys-
A
B
C
Fig. 1. Animal haemoglobins and plant
globulins, comprising the first animal and
vegetable proteins that were crystallized.
(A) Crystals of B. excelsa exelsin from the
Brazil nut (left) and of Avena sativa
avenalin from oat kernels (right) [20]. (B,
C) Haemoglobin crystals of the Tasmanian
wolf: (B) photographs of a-oxyhaemoglobin
showing groups of plates in parallel
growth (left) and of b-oxyhaemoglobin
showing small dodecahedral crystals
(right); (C) schematized drawing of the
above crystals emphasizing their prismatic
(left) and dodecahedral (right) habits [25] .
6458 FEBS Journal 280 (2013) 6456–6497 ª 2013 FEBS
Protein crystallization for structural biology R. Gieg�e

tallization of several enzymes (carboxypeptidase, cata-
lase, chymotrypsin, ribonuclease, pepsin, trypsin, ure-
ase, etc.), the crystallization of the diphtheria toxin,
and the crystallization of the polypetidic hormone
insulin [6]. All of the investigators noted the impor-
tance of salts, organic solvents, pH and/or tempera-
ture for crystallization. Progressively, they took
advantage of the new ideas of Hofmeister on salt
effects (especially the ‘salting in’ and ‘salting out’
phenomena) to reach supersaturation and discovered
the crucial role of metal ions in protein crystalliza-
tion, notably for insulin crystallization where Zn2+
ions are indispensable [26].
Precursors that impacted upon the field of
protein crystallization
The biographical notes of pioneers and the highlights
of their achievements in crystal science are summarized
in Table 1. Besides H€unefeld, Funke and Hartig who
opened the field, the inspired physico-chemical contri-
butions of Hofmeister and Ostwald deserve particular
attention, although they were of indirect influence on
early crystallization investigations. Hofmeister was the
first individual to systematically study the effects of
salts on protein stability and solubility [27]. He is the
father of what is presently known as the Hofmeister
lyotropic salt series, which ranks the relative influence
of ions on the physical behaviour of proteins [28].
These salt effects (with NH4+ having the strongest
effect with respect to decreasing solubility) turned out
to be critical for understanding protein crystallization
[29,30]. Ostwald established the rules for time depen-
dent phase changes in chemical mixtures (solid–liquidtransitions) and discovered the phenomenon of ripen-
ing [31] that has found recent applications in macro-
molecular crystallization [32].
Reichert and Brown aimed to correlate the classifi-
cation of animal species with their evolution on the
basis of the morphology of their haemoglobin crystals
[25]. Today, this appears naive but, by 1909, the idea
underlying their work was in some way visionary
because, in present biology, evolution is accounted for
by protein sequences and three-dimensional structures.
In a more crystallographic perspective, they were the
first individuals to thoroughly describe polymorphism
in protein crystals, which is now amply demonstrated.
The motivation of James B. Sumner was different.
In 1917, when he was at Cornell University and had
heavy teaching obligations, he decided to accomplish
something of real importance during his spare time.
This was the risky project of purifying an enzyme.
Fortunately, using urease from the jack bean, he opted
for a good experimental model. Two years later, he
obtained crystals of the lectin concanavalin, which is
abundant in the jack bean. It took him an additional
7 years to find the appropriate recipe to prepare crys-
talline urease. The clue to success was the extraction
of the enzyme from the protein bulk with 30% alcohol
[33]. However, his 1926 paper, in which he reported
that solutions of dissolved urease crystals possess ‘to
an extraordinary degree the ability to decompose urea
into ammonium carbonate’ [33] generated skepticism
and his conclusion was rejected by the renowned Ger-
man organic chemist Willst€atter (1915 Nobel Prize in
Chemistry), who was convinced that the catalytic
activity of enzymes is a result of organic compounds
copurified or adsorbed on carrier proteins [5]. This
forced Sumner to provide stronger arguments and
stimulated John H. Northrop to study the crystalliza-
tion of swine pepsin for which he had strong biochem-
ical evidence of its protein nature [34]. Despite
intensive efforts, no putative catalytic entity could be
separated from either urease or pepsin. The contro-
versy was resolved when Northrop developed better
quantitative tools to purify, characterize and crystallize
proteins, and thereby generalized the concept of cata-
lytic proteins to pepsin, trypsin and chymotrypsin [35].
By 1937, Sumner closed the debate with a decisive
publication on catalase from beef liver showing that its
catalytic activity requires both the protein and an iron
porphyrin group [36]. Both Sumner and Northrop
received the Nobel Prize in Chemistry in 1946 for these
biochemistry-focused contributions [5]. They shared
the Nobel Prize with Wendel M. Stanley, who was the
first to have prepared a crystalline virus [37], although
he did not immediately realize the implications of his
finding as he was on a quest to prepare protein con-
stituents of tobacco mosaic virus (TMV) [38].
More influential from the viewpoint of crystal science
was Arda A. Green with her seminal papers on the phys-
ical chemistry of proteins completed in a continuation of
the early observations of Hofmeister on the solubility of
horse carboxy- and oxyhaemoglobin as a function of the
concentration of various salts, pH and temperature
[39,40]. Accordingly, she deduced an empirical relation-
ship between protein solubility and ionic strength
log S = b – Ksl
where S is solubility and l is ionic strength, Ks is the
salting-out constant considered to be independent of
pH and temeperature, and b is a protein-, pH- and
temperature-dependent constant). Interestingly, she
noted decreasing values of Ks correlated with the rank-
ing of the salts in the Hofmeister series. Arda
A. Green was active in many other domains of protein
FEBS Journal 280 (2013) 6456–6497 ª 2013 FEBS 6459
R. Gieg�e Protein crystallization for structural biology

Table 1. Early pioneers that impacted upon the emerging science of crystallogenesis. For references, see text and Data S1.
Pioneers Highlights Biographical notes
Friedrich L.
H€unefeld
1840: First observation of crystals in blood
samples (haemoglobin)
German MD and chemist, b.1799 M€uncheberg – †1882
Greifswald. Was active at Greifswald University (Professor of
Chemistry and Mineralogy). Was with Berzelius in 1827
Otto Funke 1851: First deliberate crystallization of
haemoglobin (Blutfarbstoffes) by evaporation
German MD and chemist, b.1828 Chemnitz – †1879 Freiburg.
Was active at Leipzig, then Freiburg University (Professor of
Medicine, then Physiological Chemistry)
Theodor Hartig 1855: Crystalline particles from extracts of the
Brazil nut (a storage protein known as
B. excelsa excelsin)
German forestry biologist and botanist, b.1805 Dillenburg – †1880
Brauschweig. Was active at the German Forestry Organization
Franz Hofmeister 1888: Different salts can be placed in a regular
order with respect to their salting-out effect
on proteins (ranking now known as the
‘Hofmeister series’ or ‘lyotropic series’)
1890: First crystals of HEW albumin
Bohemian-German MD, physiologist, chemist and pharmacologist,
b.1850 Prague – †1922 W€urzburg. Worked at Prague University
until 1896 (Professor of Pharmacology), succeeded Hoppe-Seyler
1896 at Strasbourg University, left for W€urzburg in 1919
Wilhelm Ostwalda 1897: Phenomenon of ripening describing the
change of an inhomogeneous structure over
time (called Ostwald ripening). Applies to
proteins, where large crystals can grow at the
expense of small ones
Baltic-German physical chemist and philosopher, b.1853 Riga –
†1932 Grossbothen. Educated in Tartu; worked at Riga
(1881–87) then Leipzig University (Professor of Chemistry and
Philosophy). Nobel Prize in Chemistry in 1909
Edward T. Reichert
and
Amos P. Brown
1909: Publication of an impressive opus on the
solubility, crystallization and crystal
characterization (shape, angles, etc.) of
haemoglobins from ~ 100 mammalian species
and a few Batrachia, birds, fishes and reptiles
American MD from the Medical Department of Pennsylviana
University, b.1855 – †1931. Educated in Berlin, Leipzig and
Geneva; worked mainly at University of Pennsylvania (Professor
of Physiology)
American mineralogist, b.1864 Germantown – †1918 Philadelphia.
Was head Professor of Department of Mineralogy and Geology,
University of Pennsylvania, Philadelphia, PA
James B. Sumner 1919: Crystals of Canavalis ensiformis
concanavalin A & B (jack bean)
1926: First crystallization of an enzyme, urease
from jack bean. Despite skepticism he claimed
that the crystalline enzyme is a protein
American chemist and biochemist, b.1887 Canton, MA – †1955
Buffalo, NY. Graduated from Harvard University; most research
at Cornell University, Ithaca, NY (Professor of Biochemistry).
Nobel Prize in Chemistry in 1946
John H. Northrop 1930: Pepsin in crystalline form. Northrop was
visionary in realizing that a crystalline form of a
protein is not in itself a criterion of purity
American biochemist, b.1891 Yonkers, NY – †1987
Wickenburg, AZ. Main work at Rockfeller Institute in New York,
NY, and Princeton, NJ. Nobel Prize in Chemistry in 1946
1931–33: Crystallization of trypsin and
chymotrypsin
Wendel M. Stanley 1930–40: Use of chemical methods, including
crystallization, for isolation of active substances
from viruses that are harmful to plants. In 1935,
isolated tobacco mosaic virus in crystalline form
American biochemist and virologist, b.1891 Ridgeville, IN – †1987
Salamanca, Spain. Main work at the Rockfeller Institute in
Princeton, NJ; after 1948 at University of California, Berkeley, CA
(Professor of Biochemistry). Nobel Prize in Chemistry in 1946
Arda A. Green 1931–32: Seminal papers on the solubility of
horse haemoglobin as a function of pH, ionic
strength and temperature
American protein chemist and biochemist, b.1899 Prospect, PA –
†1958 Baltimore. Many prominent scientists worked under
A. Green (e.g. Krebs, 1992 Nobel Prize) or were associated with
her (e.g. the Cori’s, 1947 Nobel Prize). Posthumous Garvan
Medal awarded to notable women chemists
1956: Crystallization of luciferase (her last
contribution)
John D. Bernal
and
1934: First X-ray diffraction pattern of a protein
crystal (pepsin)
British crystallographer, b.1901 Nenagh, Ireland – †1971 London.
Mentor of D. Hodgkin at Cambridge University (Professor of
Physics); 1937: moved to Birkbeck College, London (Professor of
Crystallography)
Dorothy (Crowfoot)
Hodgkin
British chemist and protein crystallographer, b.1910 Cairo – †1994
Ilmington. Educated in Oxford; was in Cambridge with J. Bernal
and held a post at Sommerville College, Oxford, until 1977.
Nobel Prize in Chemistry in 1964
aHis son Wolfgang Ostwald (1883–1943) was the initiator of colloid chemistry and biochemistry.
6460 FEBS Journal 280 (2013) 6456–6497 ª 2013 FEBS
Protein crystallization for structural biology R. Gieg�e

science while working with the most famous American
biochemists and this explains why her work on protein
solubility did not receive the recognition that it
deserves, although it did influence the two Cori’s and
Krebs, and all three were awarded a Nobel Prize, and
later impacted decisively upon the whole field of pro-
tein crystallization.
Considerations on protein crystallization in the
epoch of physiology and chemistry
In this epoch, crystallization was a tool for protein puri-
fication and was instrumental to demonstrate that the
catalytic activity of enzyme resides within the protein
itself. In the 19th Century, most crystalline proteins
were of plant origin and only a few animal proteins
(haemoglobins and albumins) were characterized as
pure substances. Crystals were obtained in the micro-
scale range by desiccation/evaporation procedures of
crude biological materials, mainly from extracts treated
by water, alcohol, hot acetic acid or salts to solubilize
their ‘albumineous’ entities. Scaling-up procedures rep-
resented a challenge that was first tackled by Preyer
with haemoglobin [4] and pursued by the biochemists in
the early 20th Century, who significantly enriched the
repertoire of crystalline proteins. All of these proteins
were easily available and had rather robust structures, a
feature not known at the time. In retrospect, one can
wonder why the early investigators were not intrigued
by the fact that proteins considered as colloidal sub-
stances with an elusive structure can be crystallized.
Being physiologists and biochemists, it is fortunate that
they were not refrained by the rules of classical crystal-
lography, which claim that crystals are formed by
strictly identical entities, although, today, it is well
established that macromolecular crystals can encompass
proteins with disordered domains.
The paradigm change in the field occurred in 1934
when John Desmond Bernal and Dorothy Crowfoot
(Hodgkin), two prominent figures in British science,
reported the first diffraction pattern of a protein crys-
tal [2]. This closed the epoch of chemistry and physiol-
ogy in biocrystallogenesis and marked the beginning of
structural biology.
The birth of biocrystallogenesis as ascience (1934–1990)
Growing crystals was not the major concern for the
pioneers of structural biology who were busy establish-
ing methods for structure determination. They used
proteins available in large amounts and easy to crystal-
lize with the bulk methods worked out by the biochem-
ists (see above). Once the first protein structures were
solved in the 1950/60s (Table 2), researchers became
more ambitious and enrolled in objective-focused pro-
Table 2. Landmarks inmacromolecule crystallization leading to three-
dimensional (3D) structures. For references, see text andData S2
Macromolecular
class
Subclass
Example of
crystallized entities
Name, origin, (year)a
3D structure
(year)a and
PDB codeb
Proteins
Globins Haemoglobin,
human (1840)
(1963) – 4hhb
Phytoglobulins Excelsin, Brazil nut
(1855)
(2007) – 2lvfc
Enzymes HEW lysozyme (1890) (1965) – 1lyz
Urease, jack been (1926) (2012) –4h9m
Pepsin, pig (1929) (1990) – 1pep
Catalase, beef liver (1937) (1985) – 7cat
Hormones Insulin, rabbit (1926) (1969) – 4insd
Toxins Erabutoxin, sea snake (1971) (1989) – 5ebx
Antibodies Intact IgG, human (1969) (1973) – 7fab
Membrane
proteins
Porin, Escherichia coli (1980) (1995) – 1opf
Sweet tasting
proteins
Thaumatin, Thaumatococcus
daniellii (1975)
(2002) – 1kwne
Nucleic acids
tRNAs tRNAPhe, Saccharomyces
cerevisiae (1968)
(1974) – 1tn2
DNA fragments Synthetic DNA
duplexes (1988)
(1989) – 2d13f
Supramolecular
assemblies
Viruses Plant virus, TMV (1935) (1986) – 1vtmg
Enzyme:RNA
complexes
AspRS:tRNAAsp,
S. cerevisiae (1980)
(1991) – 1asy
Membrane
embedded
assemblies
Photosynthetic
reaction center
Rhodopseudomonas
viridis (1982)
(1986) – 1prc
Protein:DNA
complexes
Nucleosome,
Xenopus laevis (1984)
(1998) – 1aoi
Ribosomes 70S,
Bacillus stearothermophilus
(1980)
(2001) – 1giyh
a Prime publication(s) of crystallization or structure.b PBD codes can correspond to a refined structure deposited after
the prime publication.c Crystal structure not yet in PDB, although NMR structure has
been solved.d PBD codes can also correspond to a structure of different taxo-
nomic origin than the first crystallized entity.e Structure at 1.2 �A resolution solved from crystals grown under
microgravity in gel with data collected at room temperature.f Example of a structure of an A-DNA decamer at 2 �A resolution.g Fibre-diffraction structure at 3.5 �A resolution.h First X-ray structure of a ribosome (i.e. that of Thermus thermo-
philus) at moderate resolution (5.5 �A).
FEBS Journal 280 (2013) 6456–6497 ª 2013 FEBS 6461
R. Gieg�e Protein crystallization for structural biology

jects. The supply of interesting proteins became a limit-
ing factor and the fabrication of crystals turned out to
be a major bottleneck. Studies aiming to understand
the functioning of enzymes and the mechanisms of
protein synthesis were probably the first emblematic
objective-focused projects that stimulated worldwide
interdisciplinary efforts to overcome this bottleneck
(e.g. for understanding tRNA biology) [7]). Further-
more, deciphering protein synthesis enlarged the prob-
lem of protein crystallization to nucleic acids [41] and
nucleoprotein complexes [42]. Crystallization of mem-
brane proteins was the other great challenge [43].
Widening and exploring the crystallization
parameter space
Crystallization processes are multiparametric phenom-
ena and therefore the primordial duty of experimenters
is to properly choose the parameters leading to best
crystal growth. In the field of protein crystallization,
early investigators were not always aware of this fact
and often obtained crystals by chance, although it was
soon noted that some factors were of importance, such
as the solubility of the protein, the type of salts used to
induce supersaturation, the temperature, the need for
metal ions, and the source and amount of the protein.
Nevertheless, many crystallographers considered pro-
tein crystallization as an art where magic skills are essen-
tial for success. This idea remained popular for some
time, especially because the amount of material avail-
able for crystallization purposes was often limited. This
prevented systematic studies aiming to understand the
global or specific effects brought by the known parame-
ters affecting protein crystallization [10] (Table 3).
When projects became more ambitious, the poor suc-
cess rate in crystallization attempts led a few pioneers
to develop methods better adapted to the requirements
of nascent structural biology. The aim was to produce
the rather large crystals needed at the time for diffrac-
tion measurements with limited amounts of protein
material [44] and, importantly, to enable an exploration
of the huge crystallization parameter space (Table 3).
Handling the diversity of parameters then became
another motivation to devise new crystallization proce-
dures. Thus, in the 1980s, ~ 90 different crystallants
were tested, with ammonium sulfate and poly(ethylene
glycol) 6000 ranking at the first places [9].
From conventional and forgotten methods to
project-driven approaches
Batch and dialysis methods were commonly employed
to obtain protein crystals for X-ray crystallography
(Table 4). In conventional batch methods, supersatu-
rated protein solutions containing all the required
ingredients are left undisturbed in sealed vessels. How-
ever, the success of crystallization, notably in terms of
number and size of grown crystals, is dependent on
the level of supersaturation at time zero, which should
be chosen and tuned appropriately. Accordingly, con-
ditions can easily be varied by temperature changes or
the addition of small aliquots of chemicals in the
experimental vessels. Alternatively, sealed crystalliza-
tion chambers can be opened to allow concentration
changes by evaporation. An attractive variation of the
conventional batch method is a sequential extraction
procedure by ammonium sulfate, which applies tem-
perature gradients on protein solutions at high ionic
strength [45]. It was validated with several proteins
and employed for crystallizing E. coli MetRS [46].
Similarly, in dialysis methods, modification or
exchange of the solutions in which the dialysis bags
are immersed allows tuning of the experimental condi-
tions. However, the main drawback of both methods
is the large volume of samples (in the millilitre range)
and, consequently, the large amounts of material
(> 10 mg) required for each assay.
The advent of molecular biology and the first suc-
cesses of X-ray crystallography stimulated biologists
and crystallographers to embark on ambitious pro-
jects. This was a driving force to devise adapted crys-
tallization methods. A initial breakthrough with an
immediate impact on structural biology came in 1968
with the invention of user-friendly vapour-diffusion
methods; first, in a sitting drop version for the crystal-
lization of tRNAs [47] that rapidly evolved in a num-
ber of variants, notably hanging and sandwiched
drops displayed in various experimental arrangements.
The method is based on the equilibration of a drop
with the protein to be crystallized and all ingredients
for crystallization against a reservoir containing the
crystallant at a higher concentration than in the drop.
Equilibration proceeds by diffusion of the volatile spe-
cies (e.g. water in most cases, although it can be
organic solvents or ammonia always present in ammo-
nium sulfate) until the vapour pressure of the drop
equals that of the reservoir, which is accompanied by
a volume decrease in the drop and an increase of the
protein concentration that can enter in the supersatu-
rated phase during which crystallization can occur.
The method can operate in a reverse regime if the ini-
tial concentration of the crystallant in the reservoir is
lower than that in the drop. In that case, water
exchange occurs from the reservoir to the drop. The
reverse vapour-diffusion method was discovered fortu-
itously in the course of an attempt to gently dissolve a
6462 FEBS Journal 280 (2013) 6456–6497 ª 2013 FEBS
Protein crystallization for structural biology R. Gieg�e

Table 3. Parameters affecting protein crystallization. For references, see text and Data S3.
Main parametersa Commentsa
Chemical and biochemical
Macromolecule Can be considered as the most important parameter
Purity Purity and homogeneity essential but not absolute prerequisites
Concentration Mostly in the range 5–20 mg�mL�1 (but examples at 1 mg�mL�1 or > 60 mg�mL�1)
Deliberate modification Chemical modification of amino acids, fragmentation into structural domains
Crystallants > 40 Single compounds and ~ 40 associations of two or more compounds
Saltsb 22 Compounds, with ammonium sulfate at rank 1
Organic moleculesb 13 Compounds, with 2-methyl-2,4-pentanediol at rank 1
Polymers 10 Families, notably poly(ethylene glycol) (first use in 1976), Jeffaminesc,d(1992),
poloxamersc,e(2009), miscelleanous polymersc(2010), polysaccharidesc,f (2011) with
poly(ethylene glycol) 6000 at rank 1
Ionic liquids Imidazolium-based compounds (first use in 2007) after a precursory finding in 1999 on
the properties of ethylammonium nitrate
Buffer and pHb High success rate in the pH 6–8 range and near pI of proteins
Supersaturationb Controls nucleation (number of crystals)
Ligands Modify macromolecules properties (importance of stoichiometry)
Additives Metal ionsb; other ions; miscellaneous small compounds (in mM range)
Detergents > 50 Potentially useful detergents for membrane proteins; can be useful for ‘soluble’ proteins
Physical
Purity Beneficial effects of conformational purity; solid impurities (dust particles)
Temperatureb Tested in the range 4–60 °C; temperature-dependent solubility; temperature fluctuations
Time Minutes to years for nucleation; can modify properties of macromolecules
Pressure (up to 220 MPa) Affects solubility and nucleation (first tested in 1990)
Magnetic field (up to 10 T) Diminishes convection, can orient crystals (first tested in 1997)
Electric field (up to 10 kV�cm�1) Affects nucleation rate (first tested in 1999)
Gravitational fieldg
Earth gravity (1 g) Importance of convection & sedimentation at 1 g
Microgravity (10�6 g range) In space shuttles, stations or satellites; frequent g-variations during flights
Hypergravity (> 1000 g) In ultracentrifuges (from 1000 to 40000 g) (first tested in 1936)
Flow and motion Hampers or enhances crystal growth (combined effects of convection and diffusion)
Minimized In gelled or viscous media (microgravity mimicry), first tested in 1954
Enhanced/amplified By deliberate stirring (first explicitely tested in 2002)
Vibrations and soundsc Mostly uncontrolled; also deliberate sonocrystallization proceduresc
Laser irradiationc Triggers nucleation by cavitation effect (first tested in 2006)
Geometry of set-ups Influences crystallization kinetics (equilibration) (see text)
Volume and geometry of samples Affects physico-chemical properties of sample media (see text)
Contact surfacesc Heterogeneous nucleation and deliberate epitaxy)
Biological
Macromolecule Can be considered as the most important parameter
State Homogeneity; purity; presence of natural contaminants
Origin Extremophiles versus mesophiles and difficulty with Eukarya
In vivo modification Modification of amino acids/nucleotides; enzymatic fragmentation
Genetic variants Crystallizability affected by mutations (e.g. disruption of crystal contacts; conformational
changes in the protein)
a For details, see text and crystallization databases (e.g. http://xpdb.nist.gov:8060/BMCD4/index.faces). Most of the crystallization parame-
ters were known in 1990.b Only a few crystallization parameters were explicitly characterized before 1934.c Note the few additional parameters that were identified after 1990.d Jeffamines are polyetheramines based on either propylene oxide (PO), ethylene oxide (EO) or mixed PO/EO backbones with terminal
amino groups.e Poloxamers are amphiphilic non-ionic multiblock polymers.f Polysaccharides include alginic acids, chitosans, pectins and dextrins.g The gravity force is g (on Earth, the standard acceleration due to g is 9.81 m.s�2).
FEBS Journal 280 (2013) 6456–6497 ª 2013 FEBS 6463
R. Gieg�e Protein crystallization for structural biology

tRNAAsp precipitate in a drop (provoked by spermine)
by adjunction of water in the reservoir that was fol-
lowed by the appearance of a new crystal form of this
tRNA [48]. Vapour-diffusion was followed in the early
1970s by improved dialysis methods with the invention
of new dialysis arrangements (e.g. Cambridge buttons,
Lagerkvist cells, capillaries), initially promoted by pro-
jects on aminoacyl-tRNA synthetase (aaRS) crystalli-
zation [49,50]. At the same time, the free interface
diffusion method became popular [51] and generated
related methods, such as a liquid-bridge variant [52]
and a hybrid method combining dialysis and diffusion
in capillaries. The latter method is attractive because it
allows a decrease in the number of crystal nuclei and
an increase in crystal size by temperature pulses [53].
Of practical interest was the miniaturization of the
batch method to the microlitre level. In that case,
experiments are conducted in sitting drops under oil to
prevent evaporation and to keep volume constant [54].
A common characteristic of these methods is a signifi-
cant reduction of the volume of individual assays that
decreased by ~ 100-fold (from the millilitre- down to
the 2–50-lL range), thereby allowing a more extensive
screening of the parameter space with limited amounts
of macromolecules.
Two methods that were forgotten for a long time
and that have recently been rediscovered are worth
mentioning at this point. The first is protein crystalli-
zation by centrifugation. This was already used in
1936 to crystallize the coat protein of TMV [55] and
its physical basis was investigated in some depth in the
1970/80s with the growth of catalase crystals of vari-
ous sources in a preparative ultracentrifuge at Institute
of Crystallography in Moscow [56,57]. Even though
the original Russian publications were translated into
English, they were overlooked by most western scien-
tists, despite the fact that the centrifuge-grown crystals
led to the first structure of a catalase solved in collabo-
ration with the Rossmann laboratory [58]. The method
was rejuvenated and miniaturized in 1992 with the
crystallization of the Trichoderma resei aspartic pro-
teinase [59] and was thoroughly reinvestigated in 2008
with the crystallization of a panel of model proteins
and a RNA plant virus at hypergravity levels between
1000 and 22000 g [60]. The underlying idea of the
method is to create by centrifugation a gradient of
protein concentration in the crystallization vessel that
encompasses a supersaturated region favourable for
nucleation. A similar rationale underlies a hybrid dial-
ysis method where an increase of protein concentration
Table 4. Early crystallization methods and their variants with examples of deliberate crystallizations for X-ray crystallography. For
crystallization data, see http://xpdb.nist.gov:8060/BMCD4/index.faces. For references, see text and Data S4.
Method Remarks, Cell type (sample volume) Early applications, Year (macromolecule)
Batch methods
Conventional Vials (mL range) 1971 (sea snake erabutoxin)
Jakoby variant Applicable to protein samples of ≥ 4 mg 1971 (proteolyzed E. coli MetRS)
Microbatch Drops under oil (≤ 2 lL) 1990 (e.g. lysozyme)
Dialysis
Conventional Dialysis bags (> 1 mL) 1959 (e.g. yeast cytochrome b2)
Microdiffusion Zeppezauer cells (≤ 100 lL) 1968 (e.g. lysozyme)
Heavy-walled capillary cells (≤ 100 lL) 1970 (e.g. aldolase)
Meso and micro methods Lagerkvist cells (~ 50 lL) 1972 (S. cerevisiae LysRS)
Cambridge cells (4–350 lL) 1973 (B. stearothermophilus TyrRS)
Microcaps (< 50 lL) 1985 (E. coli enterotoxin)
Double-dialysis Cambridge buttons 1989 (Staphylococcus aureus delta toxin)
Interface diffusion
Conventional free interface
method
Pasteur pipettes and other types of glass tubes
(diameter < 6 mm)
1972 (validated with several proteins,
e.g. cytochromes)
Liquid-bridge variant Droplets of protein sample and mother liquor
connected by a liquid-bridge
1974 (Chlorobium limicola bacteriochlorophyll-
protein)
Hybrid diffusion/dialysis
method
Capillaries submitted to temperature pulses 1975 (Lactobacillus casei thymidylate synthetase)
Vapour-diffusion
Sitting drop Plates/trays with 6–100 drops (2–40 lL) 1968 (S. cerevisiae tRNAPhe)
Hanging drop Tissue culture plates with 24 wells (2–20 lL) 1971 (carp albumins)
Sandwiched drop Drops between two glass plates 1994 (bacterial cytochrome C-552)
Capillary apparatus Sample in a capillary (≤ 1 lL) 1988 (ribosome)
6464 FEBS Journal 280 (2013) 6456–6497 ª 2013 FEBS
Protein crystallization for structural biology R. Gieg�e

occurs by electrophoresis [61]. The second forgotten
method, first published in 1954 [62], is protein crystal-
lization in a gelled medium where convection is
reduced. This diffusion-dependent method, validated
by the crystallization of human serum albumin (dimer
form) in gelatin, was rediscovered 34 years later [63] in
the frame of microgravity projects. Other methods
only marginally exploited are crystal growth under lev-
itation [64] and at high pressure [65]. Interestingly, in
all of these atypical procedures, the parameter ‘diffu-
sion’ and hence its counterpart ‘convection’ are on the
forefront (see implications below).
Attempts to control and understand the
crystallization process of biomacromolecules
Before the first interdisciplinary conference on protein
crystallization in 1986 (ICCBM1), attempts to under-
stand the physico-chemical basis of protein crystal
growth were extremely scarce [66]. For example,
Schlichtkrull concluded that, after initial nucleation,
subsequent nucleations occur mainly on the surface of
existing beef insulin crystals [67] and Bunn distin-
guished between amorphous and crystalline material
when measuring the solubility of calf rennin [68]. The
situation changed radically when physicists outside of
biology entered the game and tried to adapt the theo-
retical background of small molecule crystal growth to
the protein field [69–71]. This trend was also fostered
by the first protein crystallizations in microgravity [72–74]. As a result, data accumulated rapidly and signifi-
cant information was obtained with model proteins
(lysozyme, canavalin, concanavalin A) on precrystalli-
zation [75–77], nucleation [78–83], growth [78,79,81,84]
and cessation of growth [79,80,83]. For example,
nucleation rate and final lysozyme crystal size were
found to depend upon the rate at which critical super-
saturation is approached [81].
The establishment and exploration of phase dia-
grams represented important trends (Fig. 2A). Initial
investigations were conducted on nucleic acid crystals
grown by the vapour-diffusion micromethod; first of
yeast tRNAPhe [85] and, subsequently, of DNA frag-
ments [86]. The combined effects of Mg2+ and sper-
mine concentrations on crystal quality were explored
and, in the case of tRNA, this allowed the identifica-
tion of a crystal polymorph diffracting at high resolu-
tion [85]. In the protein field, initial investigations were
conducted on lysozyme by material-consuming batch
methods (1–80 mg of protein per measurement), show-
ing the rate-limiting attachment of protein molecules
on growing crystals, the preferential growth of large
crystals from moderately saturated protein solutions
[87] and temperature-dependent solubility accompanied
by negative or positive crystallization heats for tetrago-
nal or orthorhombic polymorphs [66]. Because high
amounts of material were needed refrained to explore
phase diagrams, this encouraged the development of
user-friendly micromethods based on vapour-diffusion
in 10-lL drops and of sensitive microassays for mea-
surement of protein concentration. This allowed sys-
tematic studies with Arthrobacter glucose isomerase,
jack bean concanavalin A and HEW lysozyme. Thus,
glucose isomerase crystallization was found to be pH-
A B
Fig. 2. Phase diagrams. (A) Theoretical 2D phase diagram showing how macromolecule concentration relates to crystallant concentration;
the diagram is multidimensional and can encompass additional dimensions covering physical parameters (see list provided in Table 3). The
diagram shows how the solubility curve separates the undersaturated region with the three zones of the supersaturated region and also
how parameters vary in a crystallization assay (from the undersaturated region toward the nucleation zone following trajectory a, until a
nucleus forms in b that will grow following trajectory c until the crystal/solute equilibrium is reached in d) [14]. (B) Part of the phase diagram
of concanavalin A from jack bean showing the solubility of the lectin as a function of ammonium sulfate concentration and temperature [82].
FEBS Journal 280 (2013) 6456–6497 ª 2013 FEBS 6465
R. Gieg�e Protein crystallization for structural biology

dependent over pH range 5.5–6.5 [88]. With concanava-
lin A, solubility decreased when salt concentration
increased in accordance with the empirical Green’s law
(see above) and increased with temperature (Fig. 2B).
Moreover, crystal morphology was found to be temper-
ature dependent [82]. Importantly, as found for HEW
lysozyme, the main effects of salts on protein solubility
were a result of anions ranked in the reverse order of
the Hofmeister series (SCN� > NO3� > Cl� > cit-
rate2� > acetate� ~ H2PO4� > SO4
2�) [29]. On the
other hand, the discovery of the peculiar effects of
ammonium sulfate at high concentration was
unexpected and was beneficial for the crystallization of
the yeast AspRS:tRNAAsp complex [42,89,90].
In summary, many different crystal forms were
observed when exploring the parameter space of
A B C D
E F G H
I J K L
M N O P
Fig. 3. A gallery of crystals illustrating shape and habit variability, as well as growth pathologies, as observed under different experimental
conditions. (A–H) Crystals of model proteins: (A–D) Diversity of lysozyme crystals grown with NaCl as the crystallant [from HEW: (A)
microcrystalline precipitate, (B) twinned embedded crystals, (C) classical tetragonal habit obtained at high pressure (50 MPa); from TEW: (D)
hexagonal prisms obtained in agarose gel under 75 MPa pressure (length increased and width diminished)]. (E–G) Example of three habits
of jack bean concanavalin A crystals found in a phase diagram screening solubility as a function of ammonium sulfate concentration (0.4–
2.0 M), pH (5.0–7.0) and temperature (4–40 °C): (E) the typical form grown under almost all conditions, (F) round-shaped crystals grown
especially at 12 °C and (G) small crystals growing out of the fracture of a large crystal by 2D nucleation, as occasionally observed in 10 lL
sitting drops. (H) Tetragonal bipyramidal crystals of T. daniellii thaumatin grown in free interface diffusion reactors after 10 days of
microgravity at 20 °C with 1.6 M Na tartrate as the crystallant [USML-2 (United States Microgravity Laboratory-2) mission in October 1995;
note the increased number of smaller crystals at the crystallant entrance of the crystallization chamber at the right side and the gravity
vector from right to left]. (I–P) Crystals of key partners in translation: (I) An orthorhombic yeast tRNAAsp crystal that was useful for structure
determination. (J–L) Crystals of yeast and T. thermophilus AspRS: (J) tetragonal crystals from the yeast enzyme showing growth defects
together with brush-like spherulitic needle bunches and (K, L) gorgeous crystals of the bacterial enzyme from T. thermophilus grown (K)
under microgravity or (L) on earth from a microcrystalline precipitate by Ostwald ripening. (M) Crystals of yeast initiator tRNAMet with
growth pathology not suitable for X-ray analysis. (N–P) Crystalline diversity in yeast tRNAAsp:AspRS complex crystals grown in the presence
of a high concentration of ammonium sulfate, showing a great sensitivity of enzyme purity and RNA/protein stoichiometry: (N) spherulite-like
bodies observed with heterogeneous AspRS and a slight stoichiometric excess of tRNA (spherulites are circular bodies composed of thin
crystalline and divergent needles/fibres), (O) polymorphism in the same crystallization drop showing cubic and orthorhombic crystal habits
and (P) orthorhombic P212121 polymorphs diffracting up to 2.7 �A. For references, see text and S5.
6466 FEBS Journal 280 (2013) 6456–6497 ª 2013 FEBS
Protein crystallization for structural biology R. Gieg�e

crystallization. A few examples from the author’s lab-
oratory are shown in Fig. 3. The important
conclusion to emphasize at this point was the absence
of a positive correlation between apparent perfection
of crystal habits and high diffraction quality.
From another viewpoint, the breakthroughs brought
by light microscopy, electron microscopy [and later by
atomic force microscopy (AFM), see below] and
dynamic light scattering (DLS), either to visualize and
quantify protein crystal growth processes or as a tool
for crystallization diagnostics, were important. Thus,
monodispersity of protein solutions under precrystalli-
zation conditions, as monitored by DLS, was shown to
be a good indicator of crystallizability [75–77]. Also of
fundamental importance were investigations on lyso-
zyme crystallization that monitored the size and shape
distribution of small aggregates appearing during pre-
nucleation and kinetic features characterizing the
growth and cessation of growth phases [79]. These were
concluded later for non-uniform growth over time
accompanied by imperfections on fast-growing faces
[80] and growth by lattice defects at low supersatura-
tion and two-dimensional (2D) nucleation at high
supersaturation [83]. On the other hand, the time-
dependent pH changes that can occur in vapour-diffu-
sion set-ups [91] and the dramatic variations in water
equilibration rates when varying temperature and ini-
tial drop volume [92] confirmed the importance of
kinetic effects in protein crystallization.
Towards better and optimized crystallization
strategies
The initial efforts towards rationality in protein crystal
growth and the many observations gathered during
empirical practice of crystallization in the 1970s and
1980s led to new concepts (notably on purity) and tech-
nologies for apprehending protein crystal growth, to
the search for optimization strategies, and to proposals
regarding improved crystallization strategies that were
developed in the 1990s (see below). The fact that many
proteins remained recalcitrant to crystallize also stimu-
lated work on the physical chemistry of protein crystal-
lization and the search of biology-based strategies.
A reasonable assumption made by investigators
working with proteins recalcitrant to crystallize was to
conjecture that evolution has shaped more stable pro-
teins in organisms adapted to live under extreme con-
ditions. The idea was validated with a thermophilic
TyrRS [50] that yielded better crystals than the meso-
philic counterparts. The same is true for thermophilic
ribosomes [93]. Rationalization of the concept of
purity was another accomplishment. It was based on
personal observations (e.g. the presence of microheter-
ogeneities in tRNA and protein samples) [94] and data
from literature (e.g. beneficial effects of purification on
both crystal growth and crystal quality) [95–97]. Alto-
gether, this led to a refined definition of what is really
protein purity, namely chemical and conformational
homogeneity, an absence of protein and small mole-
cule contaminants, and stability over time. Consider-
ations about purity gave also a refined view on the
nature and importance of impurities (isoforms or
denatured/aggregated versions of the protein of inter-
est, foreign protein material, small molecule contami-
nants) in protein samples that could affect
crystallization. Striking examples concern contami-
nants present in poly(ethylene glycol), especially phos-
phate or sulfate anions [accounting for the growth of
Eco elongation factor polymorphs depending on the
brand of poly(ethylene glycol) used] [98] or aldehydes
and peroxides that were shown to affect the crystalliza-
tion of rabbit muscle phosphoglucomutase [99]. In this
context, a crystallization method combining purifica-
tion and protein conditioning in crystallization media
[100] is worth mentioning.
Accordingly, it was conjectured that the intrinsic
flexibility of peptides and many proteins would be det-
rimental to their crystallization. A remedy would be to
stabilize the unstable structures with other macromo-
lecular partners. The idea was validated with the crys-
tallization of antibody:antigen complexes, with initial
proof-of-concept experiments using lysozyme as the
antigen [101]. The relative ease to prepare monoclonal
antibodies permitted a rapid generalization of the
strategy with, for example, the crystallization of neur-
aminidase from influenza virus [102] or of the human
angiotensin II peptide [103] in complex with specific
Fab fragments derived from monoclonal antibodies.
Today, cocrystallization strategies have many applica-
tions in structural biology (see below).
Among thermodynamic parameters, temperature
and time [104] were identified first as being important
for protein crystallization. Both affect protein confor-
mation and, consequently, solubility, as well as crystal
habits (Fig. 3) and growth mechanisms. Similar effects
are brought about by pressure [65] and pH changes
[104]. On the other hand, nonreproducibility remained
a major drawback and pointed to the primordial role
of the geometry and size of set-ups (both crystalliza-
tion chambers/drops and reservoirs) that affect equili-
bration kinetics and modulate the balance between
convective and diffusive mass transport during crystal
growth, as well as the extent of crystal floating or
sedimenting in the mother liquor. Furthermore, exper-
imental evidence indicated heterogeneous and epitaxial
FEBS Journal 280 (2013) 6456–6497 ª 2013 FEBS 6467
R. Gieg�e Protein crystallization for structural biology

nucleation brought about by contact of proteins with
solid surfaces [105], with such phenomena even occur-
ring on the surface of growing crystals (Fig. 3G). The
fact that diffusion is favoured under microgravity
(and convection disfavoured), as well as the expecta-
tion of better crystals when grown in this environ-
ment, was the main justification of crystallization
programs in weightlessness. Initial experiments show-
ing growth of larger lysozyme crystals [72] were the
start of a race for the access to microgravity [106],
which generated both controversial debate [107] and a
search for an alternate means to favour diffusive mass
transport on earth. This line of thinking was first sug-
gested in 1988 by Robert and Lefaucheux, who grew
lysozyme and porcine trypsin crystals in gelled media
[63], and was largely exploited in the 1990s with stud-
ies of protein crystallization by counter-diffusion or
under magnetic- and electric-fields (Table 3; see also
below). On the other hand, seeding procedures were
recognized as practical means to optimize crystalliza-
tion as soon as initial crystalline material becomes
available. They have been shown to trigger new nucle-
ations or to enlarge the size of crystals [108].
Because of the impressive number of parameters
affecting protein crystal growth and crystal quality,
which largely exceed that involved in small molecule
crystal growth (Table 2), it became rapidly evident
that identifying the appropriate crystallization condi-
tions could not occur by systematic screening of the
parameter space. The need to understand the hierarchy
of parameters and their relationships became essential.
This was not an easy task because this hierarchy is
dependent on the class of macromolecules. Emblematic
examples are the detergents essential for membrane
protein crystallization [43,109] but not required for sol-
uble proteins, although they can have beneficial effects
[110], and the polyamines that are only essential for
tRNA crystallization [111]. To overcome difficulties,
statistical methods were invented. The first comprised
an incomplete factorial method validated with B. ste-
arothermophilus TrpRS that aimed to find correlations
between crystallization parameters and crudely esti-
mated crystal quality [112,113]. It was followed in the
1990s by sparse-matrix methods (see below). In paral-
lel, robotic systems were proposed to facilitate the
practice of crystallization and to achieve better repro-
ducibility [54,114].
Summary before entering the era of structural
genomics
The cooperation between biologists and physicists in
the 1980s with respect to crystal growth, as illustrated
by the first ICCBM Conferences, provided insights
into the mechanistic aspects of protein crystal growth.
On the other hand, crystallization was no longer
restricted to isolated proteins and now also concerned
protein assemblies, nucleic acids and nucleic acid:pro-
tein complexes. Highlights were the miniaturization of
conventional batch and dialysis crystallization and also
the invention of vapour-diffusion methods. Vapour-
diffusion methods were rapidly adopted by structural
biologists because of their versatility, although draw-
backs were soon intuitively recognized. They rely on
the fluctuating geometry of the crystallizing drops and
the dynamic nature of the vapour-diffusion process
leading to a decrease of protein concentration and a
concomittant increase of impurities in the crystallizing
media, accompanied by an enhanced poisoning of the
growing crystals, as first suggested by Wayne Ander-
son [115]. Because physico-chemical conditions in crys-
tallization drops are not well defined, this might
explain the large number of irreproducible results.
Batch methods that are more static and easier to
implement remained popular, especially in their minia-
turized versions under oil. Despite the remaining poor
understanding of many aspects of protein crystalliza-
tion, the sound theoretical basis that emerged in the
period between 1934 and 1990 opened new routes for
more rational and efficient biocrystallogenesis, which
were successfully explored in the era of structural
genomics.
Crystallogenesis in the era oftechnologies and structural genomics(1990–2013)
Crystallogenesis always benefited from the interplay
between science and technology. This trend became
especially prevalent after 1990 when the new biotech-
nologies provided tools for the preparation of any
type of protein or nucleic acid present in nature and,
when new instrumentations and robotic systems
became accessible, for more efficient crystal growth
and faster crystal analysis [116]. The initial fundamen-
tal work with HEW lysozyme and a few other model
proteins was pursued, and extended to membrane
proteins and a large panel of other proteins and mac-
romolecular assemblies of high biological value. In
parallel, ideas originating from fundamental work
were translated into applications useful for the growth
of crystals for structural biology. Altogether, this led
to a paradigm change with deep impact upon the
field. New strategies for qualitative and quantitative
evaluation of the different steps of the crystal growth
process were proposed and specific crystallizability
6468 FEBS Journal 280 (2013) 6456–6497 ª 2013 FEBS
Protein crystallization for structural biology R. Gieg�e

features were discovered. The need for large crystals
declined with the easy access to second-generation
synchrotron X-ray sources, a trend that even applies
for modern neutron crystallography. Automation pro-
gressively became essential in crystallogenesis and,
recently, the nascent nanotechnologies found many
applications in structural biology. Last but not least,
the biocrystallogenesis field received support from
Space Agencies that fostered microgravity research
and were particularly interested in protein crystalliza-
tion. Altogether, during the period between 1990 and
2013, the field benefited from dramatic advances in
analytical and gene technologies and was nourished
by a constant interplay between fundamental and
practical focused research. For simplicity, these two
aspects are discussed separately.
Fundamental crystallogenesis
The effects exerted by physical and chemical parame-
ters on macromolecular crystal growth and many
related questions have been investigated in depth by a
variety of approaches [117,118]. Exploration of param-
eter-space in phase diagrams was first on the forefront
for the selection of parameters leading to protein crys-
tallization. Imaging growth processes, scrutinizing
crystal anatomies and, important from the viewpoint
of structural biology, comparing X-ray structures
solved from crystals grown under different conditions,
represented other challenges. In the late 1990s, investi-
gations on atypical physical, chemical and method-
related parameters that might affect crystallization
became more prevalent and led to alternative crystalli-
zation methods. This was first the case for micrograv-
ity and related factors and, more recently, for light,
ionic liquids, new additives, the volume of crystallizing
samples and the geometry of set-ups. Other goals were
to control nucleation and to uncouple it from growth.
Solubility and phase diagrams
Because of the multiparametric nature of the crystalli-
zation process, protein phase diagrams are multidi-
mensional and therefore can only be partly explored.
Their landscapes represent the solubility behaviour of
proteins under crystallization conditions and can be
considered as footprints characterizing individual pro-
teins or group of proteins. Thus, the proper handling
of these parameters could be used to initiate and con-
trol crystallization. Based on empirical rules derived
from Arda A. Green’s work [39,40] and precursory
theoretical thoughts on protein solubility, it was
expected that some general rules could govern protein
solubility and thus predict crystallization. However, as
a result of a poor understanding of the crystallization
process, only qualitative rules could be expected at the
time. Thus, the pH-dependent solubility of proteins,
with a minimum at the isoelectric point (pI), where the
average charge is zero, is accounted for by the zwitter-
ionic nature of proteins. Similarly, the salt-dependent
solubility relies on the ionic interactions that salts can
make with proteins. This occurs especially at high
ionic strength as reflected by salting-out (i.e. decrease
in solubility when the salt concentration increases) and
at the less frequent opposite and poorly understood
salting-in (i.e. increase in solubility) phenomena.
Understanding how the many other factors listed in
Table 3 affect protein crystallization remained essen-
tially unknown, notably the gravity-related factors
convection and diffusion, which are well explored for
the crystal growth of conventional molecules but not
for that of proteins [69,119].
To obtain insight into these unexplored issues, sys-
tematic studies were initiated in the early 1990s, first
on the effects of Hofmeister salt concentrations, pH
and temperature on protein solubility and crystal
growth, and later on those of a variety of additional
parameters, either chemical [organic crystallants such
as poly(ethylene glycol)] or physical (pressure, convec-
tion, diffusion, light, etc.). Experiments were con-
ducted not only with the standard models, but also
with proteins of interest for structural biology. Thus,
besides exploring crystallizability of HEW lysozyme
[120–123], partial phase diagrams were established,
amongst others, for a collagenase [124], two membrane
proteins (bovine cytochrome c, Rhodobacter sphaero-
ides photoreaction centre) [125,126], a carboxypepti-
dase [127], S. cerevisiae AspRS [128] and even plant
viruses [129,130]. Thus, with lysozyme and whatever
the pH, increasing pressure resulted in greater numbers
of crystals, as well as a transition from the initial
tetragonal to the orthorhombic polymorph [122]. On
the other hand, huge temperature effects on solubility
were observed with most investigated proteins
[131,132]. In the majority of cases, a normal tempera-
ture-dependence was observed (increase of solubility
with temperature, as for lysozyme, trypsin and insulin)
but retrograde solubilities were suggested as well
(decreased solubility, as for catalase and glucose
isomerase) [131]. The situation was paradoxical with
T. daniellii thaumatin known to crystallize with Na
tartrate because the temperature-dependence of its sol-
ubility depends on the chirality of the tartrate ion (i.e.
normal with L-tartrate and retrograde with D-tartrate)
[133]. This observation is of importance because it rec-
onciles the contradictory results obtained with crystals
FEBS Journal 280 (2013) 6456–6497 ª 2013 FEBS 6469
R. Gieg�e Protein crystallization for structural biology

grown from solutions of racemic Na D,L-tartrate.
Other dramatic temperature effects were observed with
S. cerevisiae tRNAPhe, notably a transition between
three different growth mechanisms within a narrow
range of only 5 °C as seen on AFM images [134].
Regarding the effects of Hofmeister salts on solubility,
these differ globally for acidic and basic proteins and,
in the case of individual proteins, they depend on the
acidic (pH < pI) and basic (pH > pI) state of the pro-
tein [135], as well as the kosmotropic and chaotropic
nature of the salts (making strong or weak, respec-
tively, water interactions in the solvent shell around
the protein).
In the case of the poly(ethylene glycol), often associ-
ated with salts, the situation becomes more complex
because liquid–liquid phase separations are frequently
observed, with consequences on protein solubility
[136,137]. Thus, with the extremelly soluble Aspergil-
lus flavus urate oxidase, a poly(ethylene glycol)-
induced depletion potential in the protein solution
could be demonstrated by small-angle X-ray scattering
(SAXS) measurements and validated by theory [137].
It was also shown that the liquid–liquid phase separa-
tion precedes and slows down crystallization [138].
Globally, poly(ethylene glycol) modifies phase dia-
grams and favours the attractive intermolecular inter-
actions needed for crystallization. This offers the
possibility to control crystallization by varying the size
and concentration of the poly(ethylene glycol) in crys-
tallization media.
Of practical interest were light scattering studies
[both SAXS and small-angle neutron scattering
(SANS)] (Table 5) that established a correlation
between the second virial coefficient B22 and solubility
[139,140]. This coefficient characterizes the nature and
the strength of the interactions between protein parti-
cles in solution and thus provides essential information
on crystallizability. If B22 is positive, the overall intrec-
tions are repulsive. By contrast, if B22 is negative, the
interactions are globally attractive, which favours crys-
Table 5. Diagnostic tools for protein homogeneity, crystallizability and crystal quality. For references, see text and Data S7.
Tool Type of information (year of early inputs)
AFM Growth mechanisms (1992); growth pathologies (1992)
Calorimetry Thermodynamics of crystal growth; stabilization of proteins by additives (1996)
DLS Screening homogeneity protein homogeneity under precrysrallization conditions (1978); detection of
nucleation (1978)
Electron microscopy toolsa Visualization of lattice defects and 2D nucleation (1990); in situ detection of crystalline phase in biological
samples (2002); sample-quality analysis of membrane proteins (2003)
Fluorescence spectroscopiesb Detection of salts in crystals (1997), of protein aggregation in solution (2009)
Interferometryc Quantitative mapping of solution properties (solute concentration, convection, etc.) around growing crystals
(1993)
Informatic predictionsd Incomplete factorial and sampling methods (1979); database screening (2003)
Sequence-based crystallizability prediction (2006); nucleation prediction (2012)
pI (2004)
Mass spectrometry Content of macromolecules in crystals and detection of bound or contaminating small molecules within
crystals (2000)
Optical light microscopiese Crystal habit (1840); protein crystal detection in crystallization media with precipitates (2010, 2012);
measure of growth velocities on individual elementary steps (2012)
PAGE and IEF Sequence size homogeneity (1982); crystallization screening (2001)
Raman microscopy Quality control of crystals with derivatized proteins (2008)
SANS Time resolved diagnostic of the crystallization process (2008); protein fate in precrystallization (1994) and
supersaturated solutions (1995)
SAXS Crystallization screening (1995), following crystallization process (1998); detection of crystallization artefacts
(2010)
Surface plasmon resonance For identifying compounds that bind to target proteins (2012)
Thermophoresis Search of macromolecule solubility on a thermal gradient device (1998) and crystallization screening (1999)
X-ray topography Visualizing crystal perfection (1996)
a Classical and scanning electron microscopy, transmission electron microscopy, etc.b X-ray, UV and correlation fluorescence spectroscopies, etc.c Mach–Zehnder, Michelson and dual polarization interferometry, etc.d Experimental and virtual bioinformatic predictions, etc.e Classical light microscopy and advanced methods, such as laser confocal differential interference contrast miscroscopy, second-order non-
linear optical imaging of chiral crystals and ultrahigh resolution optical coherence tomography.
6470 FEBS Journal 280 (2013) 6456–6497 ª 2013 FEBS
Protein crystallization for structural biology R. Gieg�e

tallization, a conclusion that received theoretical sup-
port [141]. From the viewpoint of phase diagrams, the
existence of a metastable liquid–liquid immiscibility
region was predicted in which small liquid droplets
with a high protein concentration form before nucle-
ation proceeds. This region corresponds to the ‘crystal-
lization window’ (–8 9 10�4 < B22 < �2 9
10�4 mL�mol�1�g�2), as proposed by George and Wil-
son [139]. A refinement of this concept proposes a
‘kinetic crystallization window’, independent of the
shape and conformation of the protein [142]. It is
characterized by a kinetic coefficient, fc, defined as the
ratio between the diffusion rate of the protein in solu-
tion and its surface integration rate (based on the
kinetics of protein surface self-assembly at the air/
water interface as evaluated by surface tension mea-
surements). Formation of single crystals is kinetically
favoured in the range 1 < fc < 8 where both diffusion
and integration rates are comparable. This criterion
has been succesfuly verified for several proteins [142].
Nucleation and growth
In the 1990s, the focus was to crystallize recalcitrant
proteins and to enhance quality of crystals not suitable
for structural work. This necessitated fundamental
research and was influenced by space-crystallization
programmes. Indeed, theory claimed that a number of
gravity-dependent phenomena that prevent or perturb
crystal growth on earth are minimized in weightless-
ness (e.g. sedimentation, mass transfer, concentration
gradients and convective currents). The logical conse-
quence is an enhanced quality of space-grown protein
crystals. The expectation received support from the
early space-crystallization experiments, thereby justify-
ing ground-based research aiming to obtain deeper
insight into the mechanisms of protein crystallization
and to optimize the forthcoming microgravity mis-
sions. This also stimulated new research lines aiming
to simulate microgravity conditions on earth and to
develop alternative methods of crystallization (see
below) (Table 6). The main results are summarized
below.
Although it was known that nucleation occurs at
much higher supersaturation than growth and that,
once a nucleus is formed, growth follows spontane-
ously, little was known in the 1990s about its exact
mechanism in the protein world, except that it should
depend exponentially on supersaturation and should
occur preferentially on solid surfaces [70]. The reality
of heterogeneous nucleation of proteins induced by
substances, such as contaminating dust or other solid/
colloidal particles, was rapidly confirmed by experi-
ments [105]. It took more time to unravel the nucle-
ation process itself because two decades of intensive
work were needed [143–146] before a comprehensive
two-step mechanism emerged [147]. One reason for
this is that researchers applied classical nucleation the-
ory to solution crystallization without taking into
account differences between theoretical predictions and
experimental results [148]. Thus, according to the two-
step model, crystalline protein nuclei appear inside
pre-existing metastable clusters, which consist of dense
liquid and are suspended in the solution. Such small-
size nuclei have been visualized by AFM [149]. At high
supersaturation, the nuclei are generated in the spinod-
al regime where the nucleation barrier is negligible.
The solution–crystal spinodal helps to clarify the role
of heterogeneous substrates in nucleation and the
selection of crystalline polymorphs. These ideas pro-
vide powerful tools for the control of the nucleation
process by varying the solution thermodynamic param-
eters [147]. It is essential to note that this two-step
model worked out for proteins appears to apply for all
crystallization processes occurring in nature and indus-
try [148]. This new nucleation scenario could explain
specific effects observed with poly(ethylene glycol)
where liquid–liquid phase separations are often
observed, as well as with various substances or solid
supports known as heterologous crystallization nucle-
ants [105,150]. In the particular case of human hair,
which can act as a heterologous nucleant, it was
shown by confocal fluorescent microscopy that the
protein is concentrated on the nucleating surface, with
a substantial accumulation of protein on the sharp
edges of the hairs cuticles [151].
Controlling nucleation has practical applications. A
simple solution consists of removing uncontrolled solid
nucleants by filtration [152]. This can also be achieved
by counter-diffusion methods [153] or by application
of an electric field [154,155]. Another simple solution
is to eliminate poor quality crystals appearing after
nucleation by manual selection [156]. An alternative
possibility would be to stimulate protein nucleation by
temperature or ultrasounds, as demonstrated for small
molecules [157]. This was achieved in the 1990s for
temperature [158] and, more recently, for ultrasound
waves [159] (Table 6).
Growth of protein crystals highly depends on
supersaturation and on the presence of impurities in
the solute. It is also sensitive to crystal-dependent
parameters, such as structure and defects of crystal
faces, as well as on the bonds established between
growth units. At low supersaturation, crystals pre-
dominantly grow by screw dislocations propagating in
a helical path around lattice defects. At higher super-
FEBS Journal 280 (2013) 6456–6497 ª 2013 FEBS 6471
R. Gieg�e Protein crystallization for structural biology

Table 6. New advanced and old rejuvenated methods of protein crystallization. For references, see text and Data S7.
Method Comments on instrumentation and outputs
Proof-of-
concept (year)
Containerless Electrostatic levitation method (air/liquid contacts) for vapour-diffusion; nice crystals 1990
Batch method with floating drops (5–100 lL) under two oil layers
(liquid/liquid contacts); fewer crystals
1990
Epitaxy Many recent advances: epitaxial growth can occur in vapour-diffusion
set-ups, on minerals, on lipid or protein layers, on etched surfaces
1988
Flow-cell Crystallization under well-defined conditions (e.g. for either quiescent
or forced convection), growth under constant protein concentration)
1986
Hybrid methods combining
Microgravity and gels X-ray topographs indicate more ordered thaumatin crystals than the earth control 1999
Gel and oil Can be operated automatically in microbatch technology; improves the
gel acupuncture method; reduces growth rate
2002
Magnetic field and
levitation
Observation of new phenomena for crystallization and dissolution processes 2008
Microgravity and
counter-diffusion
High quality crystals of a lectin grown in Gel-Tube R crystallization kit
flown in Russian Service Module and crystals of several proteins grown
in the dedicated Granada box operated in ESA FOTON mission
2008
Gel and laser pulses Enhancement of nucleation at very low supersaturation 2013
Induced nucleation by
Continuous light Crystallization by Xe-lamp irradiation or by photon pressure produced by a
continuous wave laser
2006
Laser light pulses Cavitation effects essential for induction of nucleation; allowed
crystallization of many proteins, including membrane proteins;
nucleation can be induced at very low supersaturation at gel-solution interfaces
2003
Natural or modified
surfaces
Modified glass or mica surfaces, porous surfaces, organic fibres, etc. 2000
Langmuir–Blodgett
technology
Use of template protein film for growth of microcrystals 2002
Temperature First conducted in a thermonucleator (with local supersaturation control);
adapted for vapour-diffusion, batch and multiwell microbatch with T-gradient
1992
Ultrasound Nucleation of lysozyme after short ultrasonic irradiation (100 kHz and
100 W); reduction of metastable zone and crystal growth at lower supersaturation
2006
Microgravity Batch, dialysis, vapour-diffusion, free interface diffusion in advanced
protein crystallization facility (APCF) and protein crystallization diagnostic
facility (PCDF), counter-diffusion in Granada box; convection
minimized but frequent perturbations by g-jitters
1984
Microgravity features (e.g. reduced convection and favored diffusion, crystal orientation), simulated in/by
‘Ceiling’ geometry A ‘seed’ crystal attached to the top of a growth cell continues to grow in a
diffusion-limited regime; sedimenting microcrystals do not perturb the growing crystal
2009
Counter-diffusion e.g. Granada box; first known as gel acupuncture method; at present generalized use of
capillaries, works with membrane proteins
1993
Electric field Adapted to microbatch or vapour-diffusion; control of nucleation rate and
better quality of HEW lysozyme crystals
1999
Gelled media Classical devices; mass transport restricted to diffusion 1954
High pressure Batch reactors; control of solubility and crystallization 1990
Hypergravity Operated in batch vessel in ultracentrifuge; metastable starting conditions become
supersatureted during centrifugation
1936
Magnetic field Latest advances in superconducting magnets that provide quasi-microgravity conditions:
improvement of crystal quality (resolution, B-factor) observed
1997
Microfluidic Free interface diffusion, nanobatch, counter-diffusion, formulation chips; variety of chips available
(e.g. for visual crystal inspection, initial X-ray screening and high-throughput data collection)
2002
Levitation Crystals obtained under ultrasonic levitation grow at higher rates are fewer and have better
shape and larger size
2012
Reverse vapour-diffusion Operated in any classical vapour-diffusion system; requires gentle drop volume increase by
vapour-diffusion to dissolve protein precipitates (rediscovered in 1995)
1977
Stirring/vibration/flow Rotatory shaker or mechanical vibrator; improvement of resolution and mosaicity of crystals
grown in stirring mode
2002
6472 FEBS Journal 280 (2013) 6456–6497 ª 2013 FEBS
Protein crystallization for structural biology R. Gieg�e
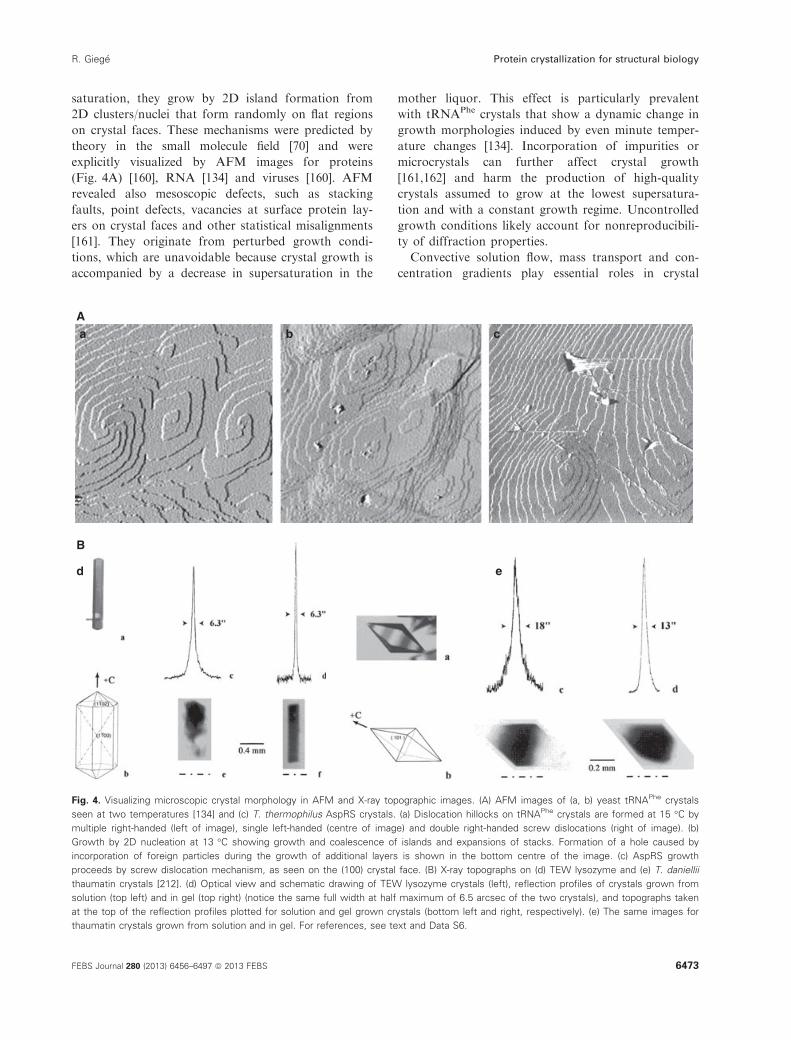
saturation, they grow by 2D island formation from
2D clusters/nuclei that form randomly on flat regions
on crystal faces. These mechanisms were predicted by
theory in the small molecule field [70] and were
explicitly visualized by AFM images for proteins
(Fig. 4A) [160], RNA [134] and viruses [160]. AFM
revealed also mesoscopic defects, such as stacking
faults, point defects, vacancies at surface protein lay-
ers on crystal faces and other statistical misalignments
[161]. They originate from perturbed growth condi-
tions, which are unavoidable because crystal growth is
accompanied by a decrease in supersaturation in the
mother liquor. This effect is particularly prevalent
with tRNAPhe crystals that show a dynamic change in
growth morphologies induced by even minute temper-
ature changes [134]. Incorporation of impurities or
microcrystals can further affect crystal growth
[161,162] and harm the production of high-quality
crystals assumed to grow at the lowest supersatura-
tion and with a constant growth regime. Uncontrolled
growth conditions likely account for nonreproducibili-
ty of diffraction properties.
Convective solution flow, mass transport and con-
centration gradients play essential roles in crystal
Aa b c
d e
B
Fig. 4. Visualizing microscopic crystal morphology in AFM and X-ray topographic images. (A) AFM images of (a, b) yeast tRNAPhe crystals
seen at two temperatures [134] and (c) T. thermophilus AspRS crystals. (a) Dislocation hillocks on tRNAPhe crystals are formed at 15 °C by
multiple right-handed (left of image), single left-handed (centre of image) and double right-handed screw dislocations (right of image). (b)
Growth by 2D nucleation at 13 °C showing growth and coalescence of islands and expansions of stacks. Formation of a hole caused by
incorporation of foreign particles during the growth of additional layers is shown in the bottom centre of the image. (c) AspRS growth
proceeds by screw dislocation mechanism, as seen on the (100) crystal face. (B) X-ray topographs on (d) TEW lysozyme and (e) T. daniellii
thaumatin crystals [212]. (d) Optical view and schematic drawing of TEW lysozyme crystals (left), reflection profiles of crystals grown from
solution (top left) and in gel (top right) (notice the same full width at half maximum of 6.5 arcsec of the two crystals), and topographs taken
at the top of the reflection profiles plotted for solution and gel grown crystals (bottom left and right, respectively). (e) The same images for
thaumatin crystals grown from solution and in gel. For references, see text and Data S6.
FEBS Journal 280 (2013) 6456–6497 ª 2013 FEBS 6473
R. Gieg�e Protein crystallization for structural biology

growth. According to theory, crystal quality is usually
better under diffusion-limited growth where a depleted
zone of the solution surrounds the growing crystal
[70,157]. However, because of convective fluid
motions, the depletion zone is hardly maintained
around a crystal on earth, which might explain why
protein crystals should be of higher quality when
grown under microgravity conditions. To explore these
issues, interferometric studies were undertaken under
earth-gravity and reduced space-gravity. The first data
on protein crystals were obtained in 1993 [163,164]
and were followed by a series of investigations using
Michelson or Mach–Zehnder interferometry that quan-
titatively characterized concentration gradients, deple-
tion zones and diffusion boundary layers around
growing and dissolving protein crystals [161,165–168].From all these studies, it was possible to propose
kinetic models of growth and to realize that quasi-sta-
ble depletion zones form around growing crystals in
space and, consequently, that best conditions for crys-
tal growth occur under microgravity and that vapour-
diffusion geometry does not provide spatially stable
crystal position or fluid conditions for optimized
growth under a diffusive regime. This last conclusion
is in line with other observations made about difficul-
ties encountered in vapour-diffusion methods, as a
result of drop size and shape, geometry of the crystalli-
zation set-up and associated evaporation kinetics that
all control the output of crystallization trials [92,169–171], and accounts for the nonreproducibility likely
explained by uncontrolled physics inside droplets.
A few comments about Ostwald ripening and mem-
brane protein crystallization are worth noting. Ripen-
ing concerns the fate of precipitates occuring at a high
supersaturation that occasionally transform into large
crystals. In the macromolecular field, the phenomenon
was first explicitly described in 1996 for Thermus ther-
mophilus AspRS [32] (Fig. 3), although it has been
occasionally seen by many protein crystal growers. A
recent study shed some light on the mechanism. Using
a combination of DLS, optical microscopy and micro-
fluidics, it could be shown that a dense amorphous
phase constituted by precrystalline protein clusters dis-
plays classical Ostwald ripening growth kinetics but
deviates from this trend after nucleation of the crystal
phase. It was concluded that this behaviour arises
from a metastable relationship between the clusters
and the ordered solid phase [172].
Regarding the mechanism underlying membrane
protein crystallization, although it likely follows gen-
eral rules demonstrated for soluble proteins, it presents
specific features as a result of the intricate interaction
networks created under crystallization conditions by
the detergents, amphiphiles, crystallants and hydro-
phobic membrane proteins. Thus, using SANS, it was
shown that optimization of micelle size and shape for
crystallization requires specific combinations of deter-
gent, amphiphile and crystallant [173]. It was also
shown that poly(ethylene glycol), often included in
crystallization media for membrane proteins, favours
solution conditions where the stability of the liquid
phase changes from stable to unstable [174]. A great
breakthrough came in 1996 when Ehud Landau and
Jurg Rosenbusch replaced the micellar crystallization
media with lipidic cubic phases [175]. These are gel-like
lipid–water systems comprising lipidic compartments
interpenetrated by aqueous channnels that were dis-
covered in the 1960s by Vittorio Luzzati [176]. Recent
data indicate that nucleation of bacteriorhodopsin
crystals occurs in such media following local rear-
rangement of the highly-curved lipidic cubic phase into
a lamellar structure mimicking the native membrane in
which the crystals will grow in a constrained environ-
ment surrounded by lamellar structures [177]. This
mechanism leads to an absence of dislocations and the
generation of new crystalline layers at numerous loca-
tions, as well as to voids and block boundaries. The
characteristic macroscopic lengthscale of these defects
suggests that the crystals grow by attachment of single
molecules to the nuclei [177]. At present, the in cubo
method is widely used [178] and applications for solu-
ble proteins are expected. Recently, the method was
extended to other mesophases in the lipid (monoolein)/
water diagrams and led to a user-friendly fast screen-
ing technology [179].
Microgravity and related issues
A further step towards understanding protein crystalli-
zation consisted of an evaluation of the parameters
governing mass transport and dynamic flow during the
process. Viscosity and gravity are the major parameter
accounting for convection/diffusion and buoyancy-
induced phenomena. Their effects are well known with
respect to the crystal growth of conventional molecules
but were only thoroughly studied in the case of pro-
teins after the first protein crystallization in micrograv-
ity [72] and the claim that reduced convection under
such conditions should favour crystal quality [106]. It
took approximately two decades to convincingly vali-
date the expectation [180]. Overall, space-grown crys-
tals grow larger, and have more regular external
morphology and better internal order with reduced
mosaic spread [181,182], although contradictory results
have been reported [14]. Resolution is sometimes over-
whelmingly improved, as for space-grown paralbumin
6474 FEBS Journal 280 (2013) 6456–6497 ª 2013 FEBS
Protein crystallization for structural biology R. Gieg�e

crystals that diffract at 0.9 �A, whereas the earth con-
trols are not suitable for diffraction analysis [183]. In a
few cases, when real ground controls were available,
space-grown crystals gave more accurate structures
(e.g. obtained with better defined initial electron den-
sity maps) [180,184,185].
Access to space is not easy and, already in 1988, a first
solution to simulate microgravity effects was proposed
consisting of crystallization in gelled media [63]. This
possibility was followed by proposals advertising pro-
tein crystallization by counter-diffusion [186], under
magnetic [187] or electric [188] fields, and, most promis-
ingly, under microfluidic conditions [189] (Table 6).
Preferential orientations of crystals were observed under
a magnetic field [190] and numerical predictions
revealed the damping of convection by magnetization
[191]. It was also realized that some atypical methods
could reproduce any potential beneficial features of the
microgravity environment, such as crystallization in
containerless systems [192], under hypergravity or at
high pressure.
In all of these methods, mass transport and fluid
movements are affected, as accounted by the dimen-
sionless Grashof GrN number [193], a classical predic-
tor in fluid mechanics, which, in the present case,
evaluates how buoyancy and viscosity forces affect pro-
teins in their liquid crystallization media according to:
GrN� buoyancy forces/viscosity forces�L3 a Dc g m�2
where L is the characteristic length of the system in
which a protein is immersed (e.g. the diameter of a
sphere in which the protein can move), a Dc is a den-
sity gradient dependent on the concentration of the
protein, g is the gravity force, and m is the viscosity of
the fluid. It can be easily seen that the same GrN value
can characterize both microgravity and earth condi-
tions provided that the low gravity force (g) in space is
balanced by adapted geometrical characteristics of the
crystallization device (L) and viscosity forces (m) on
earth. This typically occurs in gelled media and under
counter-diffusion and microfluidic conditions.
A posteriori, the usefulness of these methods for
structural biology is demonstrated by the increasing
number of Protein Data Bank (PDB) entries of struc-
tures solved with crystals grown by these atypical pro-
cedures. In a few proof-of-concept cases, it was shown
that the quality of the X-ray structures solved from
diffraction data originating from crystals grown under
conditions simulating microgravity conditions are
improved. For thaumatin, the crystals grown in aga-
rose gel diffracted to a previously unachieved resolu-
tion and yielded a structure at 1.2 �A resolution
computed from diffraction data collected at room
temperature [194]. In the case of magnetic field, a
comparison of HEW lysozyme structures of 0-T and
10-T crystals revealed only limited overall structural
changes but demonstrated significant fluctuations at a
few residues, improvement in crystal perfection and
increased diffracted intensities leading to a higher res-
olution [195]. Interestingly, for earth- and space-grown
HEW lysozyme crystals grown in the advanced pro-
tein crystallization facility apparatus, counter-diffusion
crystallization even improved the resolution of the
tetragonal crystals from 1.40 to 0.94 �A [196].
The dimensionless Reynold ReN number [193] quan-
tifies the relative importance of inertial and viscosity
forces in fluid dynamics according to:
ReN � inertial forces/viscosity forces � L v/m
where L is the characteristic length of the system, v is
the mean velocity of the protein and m is the kinematic
viscosity of the crystallization medium (m = l/q; wherel is the dynamic viscosity and q is the density of the
fluid). Evaluation of Reynold numbers was used to
find optimal stirring conditions for HEW lysozyme
crystallization [197]. Note that the stirring crystalliza-
tion method is widely used in the small molecule field
for growing high-quality crystals and, in the present
case, it was shown that intermittent flow and low ReNvalues contribute to the growth of a smaller number
of larger crystals [197]. Finally, crystallization of HEW
lysozyme was also analyzed in quiescent and forced-
convection environments [198].
Other theories and simulations predict that shear
flow could enhance or, conversely, suppress the nucle-
ation of crystals from solution. These ideas were tested
in droplets held on a hydrophobic substrate in an
enclosed environment and in a quasi-uniform constant
electric field that induces a rotational flow with a max-
imum rate at the droplet top [199]. The likely mecha-
nism of the rotational flow involves adsorption of the
protein and amphiphilic buffer molecules on the air–water interface and their redistribution in the electric
field, leading to non-uniform surface tension of the
droplet and surface tension-driven flow.
Thermodynamic considerations on protein crystallization
Although thermodynamic approaches are appropriate
to describe solubility, phase separation and crystal
growth processes, they were only scarcely used in the
field of protein crystallization. A first phenomenologi-
cal approach in 1996 with HEW lysozyme found
agreement between values of crystallization enthalpies
determined by calorimetry and by analysis of van’t
Hoff solubility plots [200]. It was followed by a few
FEBS Journal 280 (2013) 6456–6497 ª 2013 FEBS 6475
R. Gieg�e Protein crystallization for structural biology

other studies [201]. However, a fruitful paradigm
change occurred when researchers tried to understand
the enthalpic and entropic contributions to the Gibbs
free energy of crystallization (DG°cryst = DH°cryst –TDS°cryst) from the viewpoint of chemistry; in other
words, taking into account the contribution during
crystallization of intermolecular bond formation
between protein and solvent. Thermodynamic data
were gathered for several proteins (apoferritin, haemo-
globin C, insulin, lysozyme) and showed that their
crystallization is dominated by entropic phenomena
[202–204]. Thus, the solvent structure, together with
the trapping and release of water molecules, is essential
in the crystallization of these proteins. This implies
structural rearrangements in protein and solvent, mim-
icking by some aspects that which occurs in macromo-
lecular recognition phenomena. These facts have
important consequences for protein crystallization
because, by engineering DS°cryst, it becomes possible to
find thermodynamically favoured crystallization condi-
tions. The idea was exploited under two versions:
either by protein surface engineering to favour inter-
molecular interactions [205] or by calorometry-based
selection of additives for their propensity to stabilize
protein structures in crystallization solutions [206]. A
few proof-of-concept cases of crystal structures of pro-
tein variants showing modified crystal packing contacts
provided strong support for these approaches [205]
(for applications, see below).
Anatomy and quality of protein crystals
Optical microscopy images show a variety of crystal
habits, with some exhibiting perfect shape and symme-
try. However, at higher resolution, as seen by AFM,
crystal faces are not flat but have rough surfaces with
growth-dependent morphologies comprising frequent
imperfections and level differences reaching up to
1000 �A and even more. This raises the important
question of the impact that growth conditions and
growth-induced defects can have on the internal order
of crystals, as ultimately reflected by diffraction prop-
erties. X-ray topography is an appropriate technique
to answer such concerns. It was used for the first time
with protein crystals in 1996 [207,208]. The method
informs about the spread of mosaic blocks, and detects
imperfections and variations in the internal order
within a crystal [209–211]. Typical X-ray topographs
obtained from TEW lysozyme and T. daniellii thauma-
tin crystals grown under two different growth condi-
tions are shown in Fig. 4B. They clearly show,
especially for TEW lysozyme, more homogeneous
images for the gel-grown crystals, demonstrating that
the gel improves crystal quality [212]. Over the years,
the technique has been refined and applied to an
increasing number of proteins. The most recent studies
have characterized individual domains in HEW lyso-
zyme crystals (with homogeneous diffraction qualities)
[213], as well as the presence of loop and curve shaped
dislocations in such crystals [214]. Information on the
internal structure of protein crystals may be useful to
aid in the improvement of crystal growth techniques
[213] and may guide femtosecond laser processing of
gel-grown crystals for diffraction data collection on
the most perfect crystal domain [215].
Summary and main conclusions
The science of biocrystallogenesis has made consider-
able progress in the period between 1990 and 2013, in
great part through the combined efforts of biochem-
ists, biophysicists, protein crystallographers and scien-
tists from the small molecule crystal growth
community. Thus, as anticipated, it was convincingly
demonstrated that the general rules of crystal growth
apply to the protein field. In the case of nucleation, a
novel two-step mechanism was proposed by Peter Ve-
kilov and coworkers that could be generalized for all
crystallization processes, as reported by experts of the
crystal science of conventional molecules [148]. Alto-
gether, a better understanding of the physical chemis-
try of proteins in the different zones of phase
diagrams (Fig. 2A) emerged and, in the case of mem-
brane proteins, understanding how lipidic cubic phases
sustain their crystallization was an important achieve-
ment. From the standpoint of structural biology, it
was realized that crystal growth under diffusive condi-
tions enhances the quality of protein crystals, which is
reflected by the better quality of the crystallographic
models of macromolecules. To reach these conclusions,
a panel of analytical and diagnostic tools (listed with
their characteristic features in Table 5) were of opera-
tional importance. These tools were adapted to the
specific requirements of protein crystallization, in par-
ticular for measurements on microsamples (down to
the microlitre-scale for DLS) [216] or on small and
fragile crystals. The fact that some of these tools are
used by the practitioners of crystallization in structural
biology laboratories is rewarding, especially with
respect to DLS presently being widely used as a diag-
nostic tool regarding protein quality and crystallizabili-
ty [217,218].
Other offspring of the interplay between crystal sci-
ence and technology were proposals followed by vali-
dations of new crystallization methods and an update
of more conventional methods (Table 6). Most of the
6476 FEBS Journal 280 (2013) 6456–6497 ª 2013 FEBS
Protein crystallization for structural biology R. Gieg�e

novelties exploit atypical parameters (well known for
the crystallization of conventional molecules but not
yet explicitely assayed with proteins, such as tempera-
ture, pressure, stirring) or are based on emerging new
technologies (e.g. the femtosecond laser and nanotech-
nologies). Here also, it is satisfying that practitioners
became progressively convinced of the usefulness of
several of the new crystallization methods. This is
especially the case for counter-diffusion, and gel and
microfluidic based-methods (partly inspired by the
microgravity programmes that created so many con-
troversies among structural biologists), and is reflected
by the increasing number of structures in the PDB
solved from crystals obtained by these methods.
Practical crystallogenesis
This section discusses the changes in the practice of
protein crystallization in the period between 1990 and
2013 and shows how the knowledge gained from basic
research has benefited structural genomics. Different
topics developed synergically (e.g. purity, screening,
structure engineering, high-throughput and automa-
tion, nanotechnology-based methods, optimization)
but, for simplicity, they are discussed separately.
Taken together, a series of strategies for facilitating
and/or enhancing protein crystallization could be
defined (Table 7) and were succesfully employed.
Predicting likelihood of crystallization
Identifying the crystallization conditions of a protein
target can be challenging and explains why research-
ers have tried to relate sample properties with crystall-
izability. This was achieved by exploring the vast
ensemble of data available on macromolular struc-
tures and crystallization features. Although predicting
exact crystallization condition remains a dream,
important guidelines for practitioners originated from
these studies. Thus, predictors of crystallizability were
proposed, with the most emblematic being the second
virial coefficient B22 characterizing undersaturated
protein solutions [217,218]. Also of potential utility is
the kinetic coefficient fc, a predictor reflecting compe-
tition between protein volume transport and protein
surface integration within single crystals or amor-
phous aggregates [142]. Other predictors of crystalliza-
tion likelihood are based on sequence features and
intrinsic physico-chemical properties of the target
proteins (pI, melting temperature, hydrophobicity,
flexibility, etc.) [219–222] or on an analysis of experi-
mentally characterized phase diagrams [223]. For
example, analysis of crystallization data in the PDB
Table 7. A large and diversified panel of crystallization strategies.
For references, see text and Data S1.
Early quotation and year Strategy
Proof-of-
concept
(year)
1971: Creating more compact/less flexible structures
Limited proteolysis 1971
Removal of floppy protein extensions or
fragmentation in structural modules
1994
Chemically synthesized RNA domains 1995
1973: Protein as a variable
Various methods using homologous
proteins with potential better
crystallizability (e.g. from thermophiles,
etc.); screening alternate intrinsic protein
characteristics
1973
1981: Optimization
By seeding procedures 1981
By automation 1990
By controlled drop size variations 2001
By controlled temperature variations 2005
By solubility screening 2005
By advanced DLS methods (e.g. aggregate
size, drop volume., etc.)
2008
By Thermofluor method (estimation of
protein thermal stability)
2011
1983: Cocrystallization with chaperones for soluble and membrane
protein crystallization
Antibody-assisted (antibody fragments
selected by hybridoma or phage display)
1983
Ankyrin-assisted (ankyrins selected by
ribosome display)
2004
1988: Robotics/automation
Use of laboratory robotics to help
crystallization
1987/8
First dedicated system for microbatch
crystallization
1990
Automation of all procedures, in particular
for high-throughput crystallogenesis
2000
1991: Sparse-matrix sampling
A plethora of commercially available
crystallization screens
1991
1991: Protein engineering
Site-directed mutagenesis for modifying
structural properties (e.g. stability)
1991/2
Surface entropy reduction mutagenesis 2001
Site-directed mutagenesis for modifying
physical properties (e.g. solubility)
2012
1992: Uncoupling nucleation and growth
Attempts to control nucleation by a variety
of novel methods
1992
1994: Cocrystallization with chaperones for RNA crystallization
With designed protein modules 1994
With designed RNA module 1998
With antibodies 2011
1998: Heterologous cocrytallization
Easier crystallization if partners originate
from different organisms
1991
For RNA structure determination 1998
FEBS Journal 280 (2013) 6456–6497 ª 2013 FEBS 6477
R. Gieg�e Protein crystallization for structural biology

revealed a significant relationship between the calcu-
lated pI of successfully crystallized proteins and the
reported pH at which they were crystallized, thus pro-
viding information for the optimal choice of range
and distribution of the pH sampling in crystallization
trials [220]. Another analysis of the PDB indicated
that protein crystals favour some space groups over
others and suggested that symmetric proteins, such as
homodimers, would crystallize more readily on aver-
age than asymmetric monomeric proteins [224]. This
idea was validated experimentally and led to the crys-
tallization of bacteriophage T4 lysozyme after creating
by mutagenesis an artificial homodimer [225]. A
recent attractive tool for crystallization prediction
combines experimentally characterized physico-chemi-
cal features and sequence-derived data from target
proteins [226]. Note that most criteria of crystalliz-
ability are correlated with the fact that the target
should have an enhanced structural stability, as amply
confirmed by many successful crystallization projects
based on this idea (see below).
Biotechnological tools for macromolecule purification
and crystallization purposes
It is common sense, although not always taken into
consideration, that the macromolecule itself is an
important parameter, if not the most important one,
for crystallization, as explicitly discussed for proteins
[227] and nucleic acids [228]. This emphasizes the
importance of purification and macromolecule modifi-
cation procedures in crystallogenesis. In the protein
field, advances towards efficient protein expression
and purification for crystallography are well covered
in the literature [229–231]. However, although DNA
recombinant methods present many advantages, a few
drawbacks detrimental to crystallization should be
noted, such as uncontrolled overexpression leading to
inclusion bodies (particles containing protein aggre-
gates), precipitated and/or denatured proteins, proteo-
lytic degradations, incomplete post-translational
modifications, and so on. Solving these problems can
be time-consuming and costly. Lowering the overex-
pression level represents a possible remedy that
decreases the amount of inclusion bodies. The alter-
nate technology that eliminates most of these draw-
backs is cell-free in vitro protein synthesis. Presently,
only a few crystallography groups have employed this
technology to prepare soluble proteins [232,233] and,
recently, membrane proteins [234]. In the case of
RNAs as well, specific drawbacks have to be over-
come for the preparation of homogeneous samples for
crystallization. They rely on the structural and confor-
mational diversity of RNA molecules and their suscep-
tibility to enzymatic or chemical hydrolytic cleavages.
Although pure RNA samples can be prepared from
crude biological material, enzymatic and chemical syn-
thesis are presently favoured and in case of large
RNAs, enzymatic synthesis using T7 RNA polymerase
is the only possible technology [235]. Note that, for
some RNAs, annealing methods are required to
assume conformational homogeneity [236].
Many methods were used for engineering protein or
nucleic acid variants with enhanced structural stability
favouring crystallization [14]. Limited proteolysis is
probably the simplest one, as already employed in the
1970s [46] and recently rejuvenated in a version where
trace amounts of protease are added/seeded in situ to
crystallization assays [237,238]. Other methods that
aim to refine physico-chemical properties of proteins
were used to specifically favour packing contacts. They
consist of changing surface residues on the targets,
either by DNA recombinant technology [205,239,240],
or by chemical modification [241], in particular by
reductive methylation of lysine residues [242]. These
new methods are based on important precursory
observations, such as the change of a single amino
acid that created a packing contact enabling the crys-
tallization of a human ferritin [243], the application of
the concept of entropy-driven crystal growth of pro-
teins [205], or the idea that intermolecular contacts can
favour or disfavour crystallization and therefore
should be created or eliminated. Also of great poten-
tial is the DARPin technology based on the natural
ankyrin repeat-protein fold with randomized surface
residues allowing specific binding to virtually any tar-
get protein [244,245], thus allowing chaperone-assisted
crystallization.
Producing stable homogenous samples of membrane
proteins for crystallization is particularly challenging
and, as for soluble proteins, screening large numbers
of target proteins is common practice. A new strategy
has recently been proposed that involves the use of
green fluorescent protein fusion constructs and screen-
ing procedures based on expression level, detergent
solubilization yield and homogeneity, as determined by
high-throughput and automated chromatographies
[246]. Notably, antibody-assisted crystallization, intro-
duced by 1983 for soluble proteins [247], applies also
to membrane proteins [248].
Screening crystallization parameters
The idea of using condition screens for the crystalli-
zation of proteins was proposed in 1991 with the
sparse-matrix method [249]. In its original version,
6478 FEBS Journal 280 (2013) 6456–6497 ª 2013 FEBS
Protein crystallization for structural biology R. Gieg�e

the method used a set of 50 conditions statistically
chosen in a crystallization database to screen the
crystallization of a target protein. After validation of
the method, a rapid release of new screens was
observed, as illustrated by screens of general use
based, for example, on alternate polymeric crystal-
lants [250] and screens specifically designed, for exam-
ple, for RNAs [235,251], protein assemblies [252] or
membrane proteins [253,254]. Today, a large panoply
of crystallization kits is available, either for initial
screening or for optimization [255]. However, many
screens are redundant and making a good choice can
be delicate, especially for challenging projects when
the amount of macromolecular entities is limited and
the number of required trials before success is large.
A new database for the comparison of crystallization
screens could be useful for a rational choice of the
adequate screen [256].
Because the parameter-space for crystallization is
quasi-unlimited, there was always a quest to find new
compounds that sustain or improve protein crystalli-
zation. This quest was pursued in the period 1990–2013 and led to the identification of several classes of
new crystallants, such as Jeffamines, ionic liquids,
poloxamers, polysaccharides and other polymers com-
mercially available, as well as of new detergents.
Among them (Table 3), ionic liquids are particularly
appealing because of the many potential interactions
that they may establish with proteins. Thus, in a pre-
cursory work on lysozyme crystallization published in
1999, it was suggested that the liquid organic salt
ethylammonium nitrate could be of interest for pro-
tein crystallography [257]. It took several years and
more systematic crystallization studies, however,
before the concept could be firmly established [258–260]. On the other hand, the catalogue of additives
that can be of potential use in crystallization trials
constantly enlarges [255], as well as the possible buf-
fers and salt combinations. This creates a huge com-
binatorial diversity of crystallization conditions that
will even augment if the parameter temperature is
included in the screens. Several condition-screening
strategies aiming to restrict the number of trials either
consist of the use of mixes of properly chosen crystal-
lants and/or additives [255] or optimization of the
choice of additives or the buffer formulation by calo-
rimetric approaches [206,261]. Also of practical inter-
est are the positive effects on crystallization of
heterogeneous nucleants introduced on purpose in
crystallization experiments, particularly fragments of
hairs [151], which have their efficacy enhanced when
included in sparse-matrix or high-throughput screens
[262,263].
It should be noted that most compounds within the
screens (except salts) were found empirically and that
their mechanisms of action are not well understood,
especially for the small additive molecules. This is not
satisfactory and does not facilitate the design of effi-
cient new screens. A few recent dedicated studies have
provided some answers with respect to this issue. A
first case study investigated the thermodynamic effects
of acetone on insulin crystallization and concluded
that acetone displaces water molecules on the surface
of the insulin molecules [204]. Another additive widely
used in protein and nucleic acid crystallization, 2-
methyl-2,4-pentanediol, was found bound to proteins
in many crystal structures. Similarly, it could be con-
cluded that binding is accompanied by the displace-
ment of water molecules and promotes stabilization of
the protein molecules, thereby enhancing crystallizabil-
ity [264]. Interestingly, the calorimetric approaches,
discussed above, arrived at the same conclusion
[206,261]. This is in line with the working hypothesis
tested by Alex McPherson, according to which addi-
tives promote crystallization by enhancing intermolec-
ular contacts between proteins or by removing such
contacts between proteins or solvent [241].
Purity and impurities
Purity was a hot topic all along the history of biocrys-
tallogenesis [94] and its versatile importance is con-
stantly emphasized by new publications [265–268]. Forexample, commercial HEW lysozyme used in nucle-
ation studies was shown to contain significant popula-
tions of large pre-assembled lysozyme clusters that
result in a deterioration of the quality of macroscopic
crystals [265]. At the other extreme, lipidic cubic
phase-based crystallizations appear to be more robust
than crystallizations conducted in more classical deter-
gent environments because up to 50% of impurities
are tolerated in the case of the R. sphaeroides photo-
synthetic reaction centre crystals grown in cubic phases
[267]. Similarly, it was shown that highly contaminated
samples of a recombinant Eco protein yielded repro-
ducibly crystals diffracting at high resolution [266].
Given such data, it is understandable that bulk crystal-
lization from impure protein batches remains an open
issue [268].
From the standpoint of applications, it would be
important to understand how impurities can exert
either detrimental or beneficial effects on crystal
growth. When studying the surface morphology of
Bence-Jones protein crystals, it was shown that impuri-
ties adsorbed on the crystalline surface form an impu-
rity adsorption layer that prevents further growth of
FEBS Journal 280 (2013) 6456–6497 ª 2013 FEBS 6479
R. Gieg�e Protein crystallization for structural biology

the crystal: by growth–dissolution–growth cycles,
impurities can be removed and growth can resume
[269]. In another study, the role of the rate of super-
saturation was highlighted. Thus, when impurity
adsorption on crystal surface is delayed, crystal growth
is enhanced and a ‘purifying’ effect takes place. By
contrast, when impurity desorption is delayed, crystal
‘poisoning’ occurs [270]. This would imply that vibra-
tions, stirring or forced flow during crystallization
[271] could protect from detrimental impurity effects.
Automation and high-throughput
Robotic crystallization systems are efficient, tireless
and accurate, and can carry out experiments using
drop samples of very small volume (1 lL in most
cases, nanolitres in some). They can perform enor-
mous numbers of trials using remarkably small
amounts of biological sample. Many of the robotic
systems reproduce procedures currently used for man-
ual experiments, such as sitting and hanging drops.
They are affordable and well implemented in academic
laboratories [272]. In recent years, and as boosted by
the large Structural Genomics Consortia and Plat-
forms, entire integrated systems have been developed
to accelerate all steps of the crystallization process.
Besides automation of the crystallization trials and
their monitoring, screening of recombinant protein
expression [273], protein purification for crystallization
[274], protein stability [275], image analysis [276], seed-
ing [277] and other optimization procedures [278],
ligand soaking [279], crystal harvesting [280], and crys-
tal mounting [281] have also been automated. More-
over, integrated systems have been installed near to
synchrotron sources enabling in situ diffraction analy-
ses [282]. In summary, automated crystallization by
sparse-matrix methods and screening techniques to
optimize protein homogeneity and crystal quality
improved dramatically and revolutionized the crystal-
logenesis field in the last decade.
It was noted, already one decade ago, that high-
throughput screening of crystallization conditions
does not necessarily produce reproducible results
when carried out in different laboratories, demon-
strating that some important features before crystalli-
zation trials are not under control [283]. This
explains the recent efforts aiming to automatize and
standardize the preparation and handling of samples.
It would also be timely to share worldwide the huge
amount of data generated by the automated high-
throughput crystallization systems with the objective
of extracting useful predictive information. Being
aware of this need, a group a structural biologists
and bioinformaticians convened to develop a crystalli-
zation ontology [284].
Towards nanocrystallogenesis
Scaling-down of crystallization methods was a contin-
uing goal both for practical and theoretical reasons,
aiming at low sample consumption and especially for
the provision of growth conditions favouring crystal
quality. The challenge was to invent miniaturized crys-
tallization devices based either on conventional meth-
ods (batch, vapour-diffusion, etc.) or on alternative
methods that were shown to favour crystal quality
(counter-diffusion, under stirring, etc.) [197,285,286]. A
breakthrough was the demonstration in 2002 of the
feasibility of growing protein crystals in volumes as
small as 1 nL [287]. The same year saw also the entry
of the microfluidic technology in the protein crystalli-
zation field [189]. At the same time, synchrotron tech-
nologies made significant advances (see below) and
offered the possibility of collecting diffraction data on
small crystals [288].
The first microfluidic chip on the market was
based on the free interface diffusion technique [189].
It consists of a complex integrated fluidic circuit
including two networks of channels: one for liquid
handling and a second serving as actuation valves.
The chip was dedicated to high-throughput screening
and was designed to test 48 crystallization conditions
with < 10 lL of sample solution in total. This chip
was modified for the establishment of precipitation
diagrams useful for crystallization screening [289] and
for fine tuning supersaturation in combining free
interface diffusion with vapour-diffusion [290]. A
great step in miniaturization was the possibility to
generate complex mixtures of reagents in 5-nL reac-
tors [289].
Batch crystallization was implemented in a microflui-
dic system in 2003 [291]. In this case, the chip design
was extremely simple and consisted of inlets for pro-
tein, buffer and crystallant solutions, and a microfluidic
channel in which 10-nL droplets are prepared by mix-
ing these solutions in various ratios. This device allows
formulation of thousands of nanodrops, which are car-
ried by a flow of inert oil. The nanodrops can be stored
on the chip and the crystals appearing therein can be
easily analyzed by X-ray diffraction [292]. Based on the
nanodrop approach, a more complex system was
designed for basic research purposes. It is able to for-
mulate droplets and to flow them to storage chambers
where they can be concentrated or diluted by water
permeation through the chamber walls. This PhaseChip
was designed to establish phase diagrams with total
6480 FEBS Journal 280 (2013) 6456–6497 ª 2013 FEBS
Protein crystallization for structural biology R. Gieg�e

control over supersaturation, nucleation and growth
kinetics in each individual drop [293]. This technology
evolved for measuring nucleation rates [294] and for
manipulating temperature and concentrations in phase
diagrams [295].
Subsequently, counter-diffusion features were suc-
cessfully reproduced in microchannels with the produc-
tion of crystalline material ranging from single crystals
to larger monocrystals along the supersaturation gradi-
ent. When made of the appropriate polymer material,
these counter-diffusion chips allow direct on-chip char-
acterization of the crystals by X-ray diffraction, with-
out any further (and potentially deleterious) sample
handling [296,297].
Another microfluidic technology designated ‘Micro-
capillary Protein Crystallization System’ enables nano-
litre-volume screening of crystallization conditions and
in situ X-ray diffraction studies [298]. The latest
released method is based on controlled evaporation in
the microfluidic device [299]. The many advantages of
the microfluidic chips explain why the microfluidic
technology has become a popular and affordable tool
for various applications, such as condition screening,
optimization, X-ray analysis and basic crystallogenesis
research.
Laser technologies have also been miniaturized and
new laser-based tools for crystal processing have
recently been validated with HEW lysozyme crystals
grown in semi-solid agarose gel and generalized for
other crystals. Processing is carried out using a focused
femtosecond laser, enabling the preparation of small
well cut crystal fragments that are not damaged by the
laser irradiation and are suitable for X-ray analysis
[215]. Such protein microcrystals can be handled by
micromethods [281] and can be used for X-ray studies
by synchrotron microbeam technology [288].
Optimizing protein crystallization methods and crystals
Fabrication of protein crystals suitable for diffraction
studies almost always requires optimization of the ini-
tial crystallization conditions. Seeding is probably the
oldest optimization procedure, as already practiced in
the 1980s [108], and has subsequently been constantly
improved. Seeding techniques (either homogeneous or
heterogeneous cross-seeding with seeds originating
from a different protein) fall into two categories that
employ either macroseeds [108] or microcrystals as
seeds [300,301]. In both cases, the solution to be
seeded should be only slightly supersaturated so that
controlled growth can occur. Several microseeding
methods have been employed, such as streak-seeding
developed by Enrico Stura in the 1990s [300], and
recently automated [302], as well as a microseed
matrix screening method [303], as also automated
[304]. Most recent developments concern, for example,
the adaptation of seeding methods to nanocrystalliza-
tion [305] and the preparation of single microseeds by
femtosecond laser ablation [306].
Most steps and variables in the crystallization pro-
cess can be optimized [307,308] (Table 7). For exam-
ple, this concerns the choice of the crystallization
method. Thus, changing from standard methods to the
counter-diffusion technique improved the crystal of the
core complex of a hydrophobic plant photosystem
[309]. It also concerns the choice of the best tempera-
ture and pH when screening, as well as the size and
volume of the crystallizing samples. Accordingly, tem-
perature cycling [310] and pH optimization [311] strat-
egies have been proposed that were shown to increase
the possibility of obtaining crystals. Another optimiza-
tion technology keeps the crystallization solution meta-
stable during the growth process by controlled
temperature variation of the crystallization solution
[312]. Similarly, it was observed that ultrasound can
optimize nucleation by decreasing the energy barrier
for crystal formation [313]. Furthermore, Thermofluor-
based high-throughput screening methods can be
employed to optimize protein sample homogeneity,
stability and solubility [275].
Optimization also concerns crystal quality, post-
crystallization treatment for enhancing diffraction
quality [314] and crystal size. For a long time, the pro-
duction of crystals of a sufficient size and quality
proved to be a bottleneck in structural investigations.
Although techniques for screening crystals have
improved dramatically, the methods for obtaining
large crystals have progressed more slowly. Despite
many structures were solved from small crystals with
synchrotron radiation, it is far easier to solve and
refine structures when robust data are recorded from
larger crystals. In an effort to improve the size of crys-
tals, a strategy for a small-scale batch method has
been developed, which, in many cases, yields far larger
crystals than attainable by vapour-diffusion [315].
Large crystals are required for neutron crystallography
and, for that purpose, the crystal growth technique
based on temperature variations is particularly appro-
priate [312]. It has been applied to grow high-quality
large crystals of several proteins of interest, which, in
the case of A. flavus urate oxidase, yielded neutron dif-
fraction data in the range 1.9–2.5 �A [316].
Another paradigm change occurred with the advent
of sophisticated X-ray optics, ultrasensitive detectors
and microbeams at new-generation synchrotron
sources [288]. Similar to microfluidic systems, this will
FEBS Journal 280 (2013) 6456–6497 ª 2013 FEBS 6481
R. Gieg�e Protein crystallization for structural biology

revolutionize the practice of structural biology, with
the consequence that large crystals are no longer a pre-
requisite in X-ray crystallography. Thus, crystals as
small as 20 lm3, corresponding to not more than
2 9 108 unit cells, can yield usable diffraction data
[288]. The same trend, although less extreme, occurred
in neutron crystallography. In that case, a crystal of
0.15 mm3 of perdeuterated human aldose reductase
yielded a structure at 2.2 �A resolution [317]. Impor-
tantly, from the viewpoint of structural biology, smal-
ler crystals are potentially of enhanced quality (see
‘Fundamental crystallogenesis’).
An overall picture of crystallization strategies and their
outputs for biology
Although much remains unclear, the ever deeper
knowledge on crystallization has generated more
rational strategies to produce protein crystals and to
improve their diffraction quality. These strategies are
diverse (Table 7) and have contributed to solving
many bottlenecks in crystallization projects. They illus-
trate how the field of biocrystallogenesis has evolved
in the last 50 years from mainly empirical methods to
sophisticated trial-and-error strategies, as well as to
idea- and basic science-driven methods that slowly
infiltrate structural biology laboratories. Their number
(and the accompanying crystallization methods)
(Table 6) augmented progressively from 1971 until
2013, with a significant boost in the last decade.
Early strategies were based on understanding and
modifying global structural properties of proteins in
view of efficient crystallization; in other words, they
considered the protein, as such, as a parameter affect-
ing crystallization. Thus, simplified and more compact
architectures obtained by proteolysis or genetic engi-
neering, or stabilized by the addition of different types
of structural chaperones, such as antibodies, ankyrins
or macromolecular natural or designed ligands,
showed enhanced crystallizability. This applies to all
types of proteins, including membrane proteins, as
well as RNAs. In that case, the chaperone can be a
general RNA module [318,319] or a protein [320–322].Interestingly, this allowed the opportunity to crystal-
lize biologically significant RNA:protein complexes
[320,322–324].Sparse-matrix sampling combined with robotics
(introduced in the 1990s) played an essential role in
allowing quicker experiments and providing better
reproducibility. Strategies for controlling the physical
chemistry of crystallization were also of prime impor-
tance. They concern uncoupling nucleation and growth
and procedures for optimizing crystallization. As an
example, the screening space of crystallization in
vapour-diffusion methods can be reduced by control-
ling water equilibration, protein solubility and drop
preparation [325]. On the other hand, macromolecular
engineering employed to modify physical properties of
proteins that affect solubility or favour crystal packing
allowed many difficult crystallization problems to be
solved.
In case of difficulties in crystallizing an essential
protein from a given organism, switching to another
organism or, in a more systematic way, screening
orthologues is one remedy [326]. This strategy has
already been employed for proteins [50] and the ribo-
some [93], considering the relative ease of crystallizing
macromolecules from extremophiles, and has been
generalized in a screening procedure of orthologues
[327]. Strategies to optimize crystallization can take
advantage of the large panoply of available crystalliza-
tion methods (Table 6). To date, this potential has
only been partly explored, if not ignored, by practitio-
ners of crystallization, especially hybrid methods com-
bining, for example, crystallization in gel and laser
pulses to induce nucleation or methods based on stir-
ring or vibrations (although vibrations have likely
induced many uncontrolled crystallizations in the
past). Similarly, new devices allowing the growth of
protein crystals in gradient magnetic fields [328] or
assisting with protein crystallization electrochemically
[329] await more thorough testing by practitioners.
Note also an alternative approach of crystallization,
orthogonal to current approaches, developed by Alex
McPherson and colleagues, with the objective of dou-
bling current success rates [330]. It is based on the
hypothesis that many conventional small molecules,
including new crystallants, might establish stabilizing,
intermolecular and noncovalent cross-links in crystals.
Summarizing, it is rewarding to note that the
crytallization toolbox of diagnostic tools, methods and
strategies at the disposal of structural biologists
(Tables 5–7) led to the structure determination of a
variety of important proteins and macromolecular
assemblies (Table 8). Note that many of these successes
are based on recently developed crystallization strate-
gies, such as the femtosecond laser technique [331], mi-
crofluidics [332], the crystallization of complexes with
specific cross-links [333] or hybrid methods [334].
Protein crystals outside crystallography and structural
biology
Although not the subject of the present review, it is
important to note a few applications where knowledge
of protein crystal growth was crucial. This is, for
6482 FEBS Journal 280 (2013) 6456–6497 ª 2013 FEBS
Protein crystallization for structural biology R. Gieg�e

example, the case with respect to the design of protein-
based biosensors [335]. It also concerns crystallization
for protein purification [336] or safe protein storage
for pharmacological formulations (e.g. the emblematic
example of crystalline insulin) [337]. In this context, it
is worth noting that nature uses this strategy under
certain circumstances to protect macromolecules
against degradation, as is the case of ribosome crystals
found in hibernating animals [338]. More generally,
protein crystals, spherulite-like aggregates and fibres
have been found in vivo in many organisms [339,340],
including in the human body where they are often
associated with severe pathologies, such as Alzheimer’s
and Parkinson’s diseases [340]. Preventing or inhibiting
their formation could therefore have therapeutic appli-
cations. This idea is being explored with respect to the
rational inhibition of amyloid fibril formation [341].
On the other hand, it is known that antibodies can be
raised against protein or small molecule crystals [342].
This opens the possibility of medical applications, such
as for the diagnostic of crystal-based diseases (gout,
Alzheimer’s disease, etc.).
The status in 2013 and perspectives
In 2013, biocrystallogenesis is a mature science based
on strong interdisciplinarity between biology, physics,
chemistry and associated technologies. Today, the
physics and chemistry of protein crystallization are
globally known, although some aspects remain elusive,
Table 8. Examples of emblematic crystallizations based on fundamental or practical advances in protein crystal growth, that led to structure
determination. For references, see text and Data S10.
Year Biomacromolecular particle Crystallization strategy
1968 S. cerevisiae and E. coli pure native tRNA species Conventional method and/or first use of vapour-diffusion (organic solvents as
crystallants)
1980 S. cerevisiae AspRS:tRNAAsp complex Ammonium sulfate as crystallant (most salts disrupt protein:RNA complexes)
1980 B. stearothermophilus ribosome (large subunit) Homologous crystallization (crystallizability of ribosome from thermophiles
better than from mesophiles)
1982 R. viridis photosynthetic reaction centre (first
crystallzation of a membrane embedded assembly)
Ammonium sulfate (crystallant), N,N-dimethyl dodecylamine N-oxide
(detergent) and heptane-1,2,3-triol (additive)
1983 Lysozyme in complex with a monoclonal anti-
lysozyme antibody
Cocrystallization with antibodies
1991 Human ferritin Engineering crystal contacts by analogy with homologous rat ferritin
1994 S. cerevisiae RNA polymerase II Epitaxial growth on 2D crystals on positively-charged lipid layers
1994 Beef mitochondrial cytochrome b-c1 complex Crystals grown in agarose gel
1998 Group II intron domain 5–6 and hepatitis delta virus
ribozyme RNA constructs
Cocrystallization with designed RNA motif
1999 Complex between Eco tRNACys and
Thermus aquaticus elongation factor
Heterologous cocrystallization with partners from two organisms (access to
targeted structure)
2001 Human RhoGDI (cytosolic regulator of GTPases) Protein surface entropy reduction
2002 T. daniellii thaumatin Hybrid method combining microgravity and gel (data collected up to 1.2 �A
resolution at room temperature)
2004 Human aldose reductase Fine biochemistry; crystallization with cofactor and inhibitor (crystals
diffracting at ultrahigh 0.66 �A resolution)
2004 Maltose binding protein and two eukaryotic kinases Cocrystallization with an ankyrin repeat protein
2006 Complex of tRNAGlu and MnmA (an enzyme that
synthesizes 2-thioU at the wobble position of
certain tRNAs)
Femtosecond laser technique
2006 R. viridis photosynthetic reaction centre (proof-of-
concept experiment)
Droplet-based microfluidic batch (at present ≥ 14 solved structures in PDB)
2006 Human glutamate carboxypeptidase II (a large
glycosylated)
Fine biochemistry; heterologous overexpression
2011 Complex of human gankyrin and C-terminal domain
of S6 proteasomal protein
Crystallization of a specific photo cross-linked complex (via incorporation of a
photophore by genetic code expansion)
2012 Thermococcus thioreducens pyrophosphatase Counter-diffusion for neutron crystallography (Hughes RC, Coates C,
Garcia-Ruiz J-M, Blakely M & Ng JD)
2013 Human epidermal growth factor receptor (an apo
cancer-associated mutant)
Hybrid method: microgravity and counter-diffusion
(in JAXA Crystallization Box)
2013 Decameric bacterial SelA:tRNASec ring structure with
heterologous tRNA
Choice of the best bacterial orthologue (Aquifex aealicus) and heterologous
cocrystallization
FEBS Journal 280 (2013) 6456–6497 ª 2013 FEBS 6483
R. Gieg�e Protein crystallization for structural biology

such as an understanding of the growth of undesirable
protein spherulites in crystallization trials, as appar-
ently favoured by heterogeneous nucleation [343]. It is
expected that the current studies on model proteins
will contibute to finding methods preventing their for-
mation [340,343]. Predicting the likelihood of crystalli-
zation as well has made progress, although
uncertainties still remain in crystallization experiments.
Thus, despite a solid fundamental background, many
protein crystals continue to be obtained by trial-and-
error strategies. However, rational-based crystallization
methods, such as counter-diffusion, as well as crystalli-
zation in gels or nanocrystallogenesis, are slowly being
adopted by crystal growers, and a few others await
more systematic testing, such as stirring methods.
On the other hand, in 2013, the PDB contains
~ 82 000 macromolecular structures solved by X-ray
crystallography (but only 64 structures determined
and/or refined using neutron diffraction data), repre-
senting a large panel of proteins originating from
throughout the tree of life. This could mean that the
bottleneck of crystallization is solved such that, in the
future, protein crystallization will be straightforward.
However, this is not true for three main reasons. First,
the majority of these structures correspond to soluble
proteins and there is a dramatic lack of membrane
protein structures, which are predicted to represent
approximately half of the proteome. Second, the pres-
ently solved RNA structures represent only 3% of the
total and the crystallography of lipids is quasi-inexis-
tent. An increasing awareness of the importance of
RNA and lipids in biology requires a much better
knowledge of their structures. Third, biologists are
becoming more and more ambitious and want to know
ever more intricate and larger macromolecular struc-
tures and assemblies; they especially want to compre-
hend the plasticity and dynamics of proteins and are
even more ambitious regarding macromolecular
machines. In addition, there will always be a need for
structures solved at high and ultrahigh resolution.
Given this situation, one can anticipate further devel-
opments in the crystallogenesis of membrane proteins
[109] and lipids [344], RNAs either free or in complex
with proteins [345] and glycoproteins [346]. Improving
crystallization methods and their application to ambi-
tious biological problems will continue to be at the
forefront of research (e.g. the gel method) [347,348].
This also concerns crystallization on solid nanotem-
plates [349] and other advanced nanocrystallogenesis
methods [350]. Studying crystal polymorphs should
also be pursued and could enable better access to the
structural plasticity of macromolecules, and also erase
possible artefacts resulting from packing effects [351].
From a more global perspective, concepts of mac-
romolecular crowding and macromolecular confine-
ment both in vitro and in vivo [352], should enter
the field of biocrystallogenesis. Thus, one could
question the actual physico-chemical properties of
concentrated protein solutions in nanodrops and
crystallizability (enhanced or inhibited) in crowded
media. Being able to answer such questions could
foster applications for more controlled protein crys-
tallization and, importantly, shed light on in vivo
protein crystallizations and their relation with
pathologies. Moreover, in vivo-grown crystals could
be usable for the emerging technology of free-elec-
tron laser-based serial femtosecond crystallography
[339]. In conclusion, an exciting future is expected
and it is anticipated that the interplay between sci-
ence and technology will continue in the science of
biocrystallogenesis [7].
Acknowledgements
This text is based on lectures given at the FEBS Prac-
tical Courses on ‘Advanced methods in macromolecu-
lar crystallization’, held in Nove Hrady (Czech
Republic) in 2004–2012. It is written to acknowledge
FEBS with respect to its support for the field of bio-
crystallogenesis, which started in 1987 with a FEBS
Lecture Course: namely ICCBM2, in Bischenberg
(France). Warm thanks are extended to Ivana Kuta
Smatanova, Pavlina Rezacova and Rolf Hilgenfeld,
the organizers of the Nove Hrady Courses, to all my
students and coworkers from Strasbourg, past and
present, and to all my colleagues from the biocrystallo-
genesis and structural biology communities for the
exchange of ideas and knowledge over the last
40 years. During all of this period, the support of
CNRS and the University of Strasbourg was essential.
References
1 H€unefeld FL (1840) Der Chemismus in der
thierischen Organisation. Physiologisch-chemische
Untersuchungen der materiellen Ver€anderungen, oder
des Bildungslebens im thierischen Organismus;
insbesondere des Blutbildungsprocesses, der Natur
der Blut K€orperchen und ihrer Kernchen. Ein
Beitrag zur Physiologie und Heilmittellehre.
Brockhaus, Leipzig.
2 Bernal JD & Crowfoot D (1934) X-ray photographs of
crystalline pepsin. Nature 133, 794–795.
3 Gieg�e R & Ducruix A (1992) An introduction to the
crystallogenesis of biological macromolecules. In
Crystallization of Nucleic Acids and Proteins: A
6484 FEBS Journal 280 (2013) 6456–6497 ª 2013 FEBS
Protein crystallization for structural biology R. Gieg�e

Practical Approach (Ducruix A & Gieg�e R, eds), pp.
1–18. IRL Press, Oxford.
4 Preyer WT (1871) Die Blutkrystalle. Mauke’s Verlag,
Jena.
5 Dounce AL & Allen PT (1988) Fifty years later:
recollections of the early days of protein
crystallization. Trends Biochem Sci 13, 317–320.
6 McPherson A (1991) A brief history of protein crystal
growth. J Cryst Growth 110, 1–10.
7 Gieg�e R (2013) Crystallogenesis at the heart of the
interplay between science and technology in the quest
to comprehend tRNA biology. Cryst Growth Des 13,
405–414.
8 McPherson A (1982) Preparation and Analysis of
Protein Crystals. Wiley and Sons, New York.
9 Gilliland GL & Davies DR (1984) Protein
crystallization: the growth of large-scale single crystals.
Methods Enzymol 104, 370–381.
10 McPherson A (1990) Current approaches to
macromolecular crystallization. Eur J Biochem 189,
1–23.
11 Gieg�e R, Drenth J, Ducruix A, McPherson A &
Saenger W (1995) Crystallogenesis of biological
macromolecules. Biological, microgravity, and other
physico-chemical aspects. Prog Crystal Growth Charact
30, 237–281.
12 Chernov AA (2003) Protein crystals and their growth.
J Struct Biol 142, 3–21.
13 Gieg�e R & Sauter C (2010) Biocrystallography: past,
present, future. HFSP J 4, 109–121.
14 Sauter C, Lorber B, McPherson A & Gieg�e R (2012)
Crystallization. General methods. In International
Tables for Crystallography (Arnold E, Himmel D &
Rossmann M, eds), pp. 99–121. John Wiley and Sons,
Chichester.
15 Russo Krauss I, Merlino A, Vergara A & Sica F
(2013) An overview of biological macromolecules
crystal growth. Int J Mol Sci 14, 11643–11691.
16 Wlodawer A, Minor W, Dauter Z & Jaskolski M
(2013) Protein crystallography for aspiring
crystallographers, or how to avoid pitfalls and traps in
macromolecular structure determination. FEBS J 280,
5705–5736.
17 Hoppe-Seyler F (1864) Ueber die optische und
chemische Eigenschaften des Blutfarbstoffs. Virchows
Archiv Path Anat und Physiol 29, 233–257.
18 Hartig T (1855) Ueber das Klebermehl. Botanische
Zeitung 13, 881.
19 Ritthausen H (1872) Die Eiweissk€orper der
Getreidearten, H€ulsenfr€uchte und €Olsamen.
Beitr€age zur Physiologie der Samen der
Culturgew€achse, der Nahrungs- und Futtermilch.
Cohen & Sohn, Bonn.
20 Osborne TB (1892) Crystallized vegetable proteids.
Amer Chem J 14, 662–689.
21 Guo F, Jin T, Howard A & Zhang YZ (2007)
Purification, crystallization and initial crystallographic
characterization of brazil-nut allergen Ber e 2. Acta
Cryst F 63, 976–979.
22 Osborne TB (2007) The vegetable proteins. In
Monographs on Biochemistry. 2nd edn. (Thomas B
Osborne ed.). pp. xiii+154. London: Longmans Green
and Co. 1924. J Soc Chem Industry 43, 413–452.
23 Funke O (1851) €Uber das Milzvenenblut. Z Rat
Medicin 1, 172–218.
24 Hofmeister F (1890) €Uber die Darstellung von
krystallisirten Eieralbumin und die
Krystalallisierbarkeit von colloider Stoffe. Hoppe-
Seylers Z Physiol Chem 14, 165.
25 Reichert ET & Brown AP (1909) The Differentiation
and Specificity of Corresponding Proteins and Other
Vital Substances in Relation to Biological
Classification and Evolution: The Crystallization of
Hemoglobins. Carnegie Institution, Washington DC.
26 Abel JJ (1926) Crystalline Insulin. Proc Natl Acad Sci
USA 12, 132–136.
27 Hofmeister F (1888) 11. Zur Lehre von der Wirkung
der Salze. Arch exp Path Pharm 24, 247–260.
28 Zhang Y & Cremer PS (2006) Interactions between
macromolecules and ions: the Hofmeister series. Curr
Opin Chem Biol 10, 658–663.
29 Ri�es-Kautt M & Ducruix A (1989) Relative
effectiveness of various ions on the solubility and
crystal growth of lysozyme. J Biol Chem 264,
745–748.
30 Collins KD (2004) Ions from the Hofmeister series and
osmolytes: effects on proteins in solution and in the
crystallization process. Methods 34, 300–311.
31 Ostwald W (1897) Studien €uber die Bildung und
Umwandlung fester K€orper. Z physic Chemie 22,
289–330.
32 Ng J, Lorber B, Witz J, Th�eobald-Dietrich A, Kern D
& Gieg�e R (1996) The crystallization of
macromolecules from precipitates: evidence for
Ostwald ripening. J Cryst Growth 168, 50–62.
33 Sumner JB (1926) The isolation and crystallization of
the enzyme urease. J Biol Chem 69, 435–441.
34 Northrop JH (1929) Crystalline pepsin. Science 69,
580.
35 Northrop JH (1930) Crystalline pepsin: II. General
properties and experimental methods. J Gen Physiol
13, 767–780.
36 Sumner JB & Dounce AL (1937) Crystalline catalase.
Science 85, 366–367.
37 Stanley W (1935) Isolation of a crystalline protein
possessing the properties of tobacco-mosaic virus.
Science 81, 644–645.
38 Kay LE (1986) W. M. Stanley’s crystallization of
the tobacco mosaic virus, 1930–1940. ISIS 77,
450–472.
FEBS Journal 280 (2013) 6456–6497 ª 2013 FEBS 6485
R. Gieg�e Protein crystallization for structural biology

39 Green AA (1931) Studies in the physical chemistry of
the proteins. VIII. The solubility of hemoglobin in
concentrated salt. A study of the salting out of
proteins. J Biol Chem 93, 495–516.
40 Green AA (1932) Studies in the physical chemistry of
the proteins: X. The solubility of hemoglobin in
solutions of chlorides and sulfates of varying
concentration. J Biol Chem 95, 47–66.
41 Kim S-H & Rich A (1968) Single crystals of transfer
RNA: an X-ray diffraction study. Science 162,
1381–1384.
42 Gieg�e R, Lorber B, Ebel J-P, Moras D & Thierry J-C
(1980) Crystallization of the complex formed between
yeast aspartyl tRNA and its specific aminoacyl tRNA
synthetase. CR S�eances Acad Sci S�er D Sci Nat 291,
393–396.
43 Michel H (1983) Crystallization of membrane proteins.
Trends Biochem Sci 8, 56–59.
44 Zeppezauer M (1971) Formation of large crystals.
Methods Enzymol 22, 235–266.
45 Jakoby WB (1968) A technique for the crystallization
of proteins. Anal Biochem 26, 295–298.
46 Waller J-P, Risler J, Monteilhet C & Zelwer C (1971)
Crystallisation of trypsin-modified methionyl-tRNA
synthetase from Escherichia coli. FEBS Lett 16,
186–188.
47 Hampel A, Labanauskas M, Connors PG, Kirkegard
L, RajBhandary UL, Sigler PB & Bock RM (1968)
Single crystals of transfer RNA from
formylmethionine and phenylalanine transfer RNA’s.
Science 162, 1384–1387.
48 Gieg�e R, Moras D & Thierry J-C (1977) Yeast transfer
RNAAsp: a new high resolution X-ray diffracting
crystal form of a transfer RNA. J Mol Biol 115, 91–96.
49 Lagerkvist U, Rymo L, Lindquist O & Andersson E
(1972) Appendix: Some properties of crystals of lysine
transfer ribonucleic acid ligase from yeast. J Biol Chem
247, 3897–3899.
50 Reid BR, Koch GLE, Boulanger Y, Hartley BS &
Blow D (1973) Crystallization and preliminary X-ray
diffraction studies on tyrosyl transfer RNA synthetase
from Bacillus stearothermophilus. J Mol Biol 80,
199–201.
51 Salemme FR (1972) A free interface diffusion
technique for crystallization of proteins for X-ray
crystallography. Arch Biochem Biophys 151, 533–540.
52 Fenna RE, Matthews BW, Olson JM & Shaw EK
(1974) Structure of a bacteriochlorophyll-protein from
the green photosynthetic bacterium Chlorobium
limicola: Crystallographic evidence for a trimer.
J Mol Biol 84, 231–240.
53 Koeppe RE II, Stroud RM, Pena VA & Santi DV
(1975) A pulsed diffusion technique for the growth of
protein crystals for X-ray diffraction. J Mol Biol 98,
155–160.
54 Chayen NE, Shaw Stewart PD, Maeder DL & Blow
DM (1990) An automated system for microbatch
protein crystallisation and screening. J Appl Cryst 23,
297–302.
55 Wyckoff RW & Corey RB (1936) The ultracentrifugal
crystallization of tobacco mosaic virus protein. Science
84, 513.
56 Karpukhina SY, Barynin VV & Lobanova GM (1975)
Crystallization of catalase in the ultracentrifuge. Sov
Phys Crystallogr 20, 417–418.
57 Barynin VV & Melik-Adamyan VR (1982) The
mechanism of crystallization of proteins in an
ultracentrifuge. Sov Phys Crystallogr 27, 588–591.
58 Vainshtein BK, Melik-Adamyan WR, Barynin VV,
Vagin AA, Grebenko AI, Borisov VV, Bartels KS,
Fita I & Rossmann MG (1986) Three-dimensional
structure of catalase from Penicillium vitale at 2.0 �A
resolution. J Mol Biol 188, 49–61.
59 Pitts JE (1992) Crystallization by centrifugation.
Nature 355, 117.
60 Lorber B (2008) Virus and protein crystallization
under hypergravity. Cryst Growth Des 8, 2964–
2969.
61 Chin C-C, Dence JB & Warren JC (1976)
Crystallization of human placental estradiol 17b-
dehydrogenase. A new method for crystallizing labile
enzymes. J Biol Chem 251, 3700–3706.
62 Low BW & Richards FM (1954) Measurements of the
density, composition and related unit cell dimensions
of some prootein crystals. J Am Chem Soc 76,
2511–2518.
63 Robert M-C & Lefaucheux F (1988) Crystal growth in
gels: principle and applications. J Cryst Growth 90,
358–367.
64 Rhim W-K & Chung SK (1990) Isolation of
crystallizing droplets by electrostatic levitation.
Methods: A Companion to Methods Enzym 1, 118–127.
65 Visuri K, Kaipainen E, Kivim€aki J, Niemi H, Leissla
M & Palosaari S (1990) A new method for protein
crystallization using high pressure. Biotechnology 8,
547–549.
66 Howard SB, Twigg PJ, Baird JK & Meehan EJ (1988)
The solubility of hen egg-white lysozyme. J Cryst
Growth 90, 94–104.
67 Schlichtkrull J (1957) Insulin crystals. V. The
nucleation and growth of insulin crystals. Acat Chem
Scand 11, 439–460.
68 Bunn CW, Moews PC & Baumber ME (1971) The
crystallography of calf rennin (chymosin). Proc R Soc
Lond B Biol Sci 178, 245–258.
69 Rosenberger F (1986) Inorganic and protein crystal
growth – similarities and differences. J Cryst Growth
76, 618–636.
70 Boistelle R & Astier J-P (1988) Crystallization
mechanisms in solution. J Cryst Growth 90, 14–30.
6486 FEBS Journal 280 (2013) 6456–6497 ª 2013 FEBS
Protein crystallization for structural biology R. Gieg�e

71 Feigelson RS (1988) The relevance of small molecule
crystal growth theories and techniques to the growth
of biological macromolecules. J Cryst Growth 90, 1–13.
72 Littke W & John C (1984) Protein single crystal
growth under microgravity. Science 225, 203–204.
73 DeLucas LJ, Suddath FL, Snyder RS, Naumann R,
Broom MB, Pusey M, Yost V, Herren B, Carter D,
Nelson B et al. (1986) Preliminary investigations of
protein crystal growth using the space shuttle. J Cryst
Growth 76, 681–693.
74 Erdmann VA, Lippmann C, Betzel C, Dauter Z,
Wilson K, Hilgenfeld R, Hoven J, Liesum A, Saenger
W, M€uller-Fahrnow A et al. (1989) Crystallization of
proteins under microgravity. FEBS Lett 259,
194–198.
75 Baldwin ET, Crumly KV & Carter CW (1986)
Practical, rapid screening of protein crystallization
conditions by dynamic light scattering. Biophys J 49,
47–49.
76 Mikol V, Hirsch E & Gieg�e R (1990) Diagnostic of
precipitant for biomacromolecule crystallization by
quasi-elastic light-scattering. J Mol Biol 213, 187–195.
77 Kadima W, McPherson A, Dunn MF & Jurnak FA
(1990) Characterization of precrystallization
aggregation of canavalin by dynamic light scattering.
Biophys J 57, 125–132.
78 Heidner E (1978) Protein crystallizations. The
functional dependence of the nucleation rate on the
protein concentration and solubility. J Cryst Growth
44, 139–144.
79 Kam Z, Shore HB & Feher G (1978) On the
crystallization of proteins. J Mol Biol 123, 539–555.
80 Durbin SD & Feher G (1986) Crystal growth studies
of lysozyme as a model of protein crystallization.
J Cryst Growth 76, 583–592.
81 Gernert KM, Smith R & Carter DC (1988) A simple
apparatus for controlling nucleation and size in
protein crystal growth. Anal Biochem 168, 141–147.
82 Mikol V & Gieg�e R (1989) Phase diagram of a
crystalline protein: determination of the solubility of
concanavalin A by a microquantitation assay. J Cryst
Growth 97, 324–332.
83 Durbin SD & Feher G (1990) Studies of crystal
growth mechanisms by electron microscopy. J Mol
Biol 212, 763–774.
84 Pusey ML, Snyder RS & Naumann R (1986) Protein
crystal growth. Growth kinetics for tetragonal
lysozyme crystals. J Biol Chem 261, 6524–6529.
85 Ladner JA, Finch JT, Klug A & Clark BFC (1972)
High-resolution X-ray diffraction studies on a pure
species of transfer RNA. J Mol Biol 72, 99–101.
86 Malinina L, Makhaldiani VV, Tereshko V, Zarytova
VF & Ivanova Z (1987) Phase diagrams for DNA
crystallization systems. J Biomol Struct Dyn 5,
405–433.
87 Ataka M & Asai M (1988) Systematic studies on the
crystallization of lysozyme. Determination and use of
phase diagrams. J Cryst Growth 90, 86–93.
88 Chayen NE, Akins J, Campbell-Smith S & Blow DM
(1988) Solubility of glucose isomerase in ammonium
sulphate solutions. J Cryst Growth 90, 112–116.
89 Gieg�e R, Lorber B, Ebel J-P, Moras D, Thierry J-C,
Jacrot B & Zacca€ı G (1982) Formation of a
catalytically active complex between tRNAAsp and
aspartyl-tRNA synthetase from yeast in high
concentrations of ammonium sulphate. Biochimie 64,
357–362.
90 Lorber B, Gieg�e R, Ebel J-P, Berthet C, Thierry J-C
& Moras D (1983) Crystallization of a tRNA-
aminoacyl-tRNA synthetase complex.
Characterization and first crystallographic data. J Biol
Chem 258, 8429–8435.
91 Mikol V, Rodeau J-L & Gieg�e R (1989) Changes of
pH during biomacromolecule crystallization by vapor
diffusion using ammonium sulfate as the precipitant.
J Appl Cryst 22, 155–161.
92 Mikol V, Rodeau J-L & Gieg�e R (1990) Experimental
determination of water equilibration rates in the
hanging drop method of protein crystallization. Anal
Biochem 186, 332–339.
93 Yonath A, M€ussig J, Tesche B, Lorenz S, Erdmann
VA & Wittmann H (1980) Crystallization of the large
ribosomal subunits from Bacillus stearothermophilus.
Biochem Int 1, 428–435.
94 Gieg�e R, Dock A-C, Kern D, Lorber B, Thierry J-C &
Moras D (1986) The role of purification in the
crystallization of proteins and nucleic acids. J Cryst
Growth 76, 554–561.
95 Bott RR, Navia MA & Smith JL (1982) Improving
the quality of protein crystals through purification
by isoelectric focusing. J Biol Chem 257,
9883–9886.
96 Spangler BD & Westbrook EM (1989) Crystallization
of isoelectrically homogeneous cholera toxin.
Biochemistry 28, 1333–1340.
97 Van der Laan JM, Swarte MBA, Groendijk H, Hol
WGJ & Drenth J (1989) The influence of purification
and protein heterogeneity on the crystallization of
p-hydroxybenzoate hydroxylase. Eur J Biochem 179,
715–724.
98 Jurnak F (1985) Induction of elongation factor Tu-
GDP crystal polymorphism by polyethylene glycol
contaminants. J Mol Biol 185, 215–217.
99 Ray WJJ & Bracker CE (1986) Polyethylene glycol:
catalytic effect on the crystallization of
phosphoglucomutase at high salt concentration.
J Cryst Growth 76, 562–576.
100 Gulewicz K, Adamiak D & Sprinzl M (1985) A new
approach to the crystallization of proteins. FEBS Lett
189, 179–182.
FEBS Journal 280 (2013) 6456–6497 ª 2013 FEBS 6487
R. Gieg�e Protein crystallization for structural biology

101 Mariuzza RA, Poljak RJ, Mihaesco C & Mihaesco E
(1983) Crystals of the human heavy chain disease
protein Riv and human Fc fragment are isomorphous:
further evidence for conformational flexibility in the
hinge region of immunoglobulins. J Mol Biol 165,
559–561.
102 Laver WG, Webster RG & Colman PM (1987)
Crystals of antibodies complexed with influenza virus
neuraminidase show isosteric binding of antibody to
wild-type and variant antigens. Virology 156, 181–184.
103 Garcia KC, Ronco P, Verroust PJ & Amzel LM
(1989) Crystallization and preliminary X-ray
diffraction data of an anti-angiotensin II Fab and
of the peptide-Fab complex. J Biol Chem 264,
20463–20466.
104 McPherson A (1985) Crystallization of proteins by
variation of pH or temperature. Methods Enzymol 114,
125–127.
105 McPherson A & Shlichta P (1988) Heterogeneous and
epitaxial nucleation of protein crystals on mineral
surfaces. Science 239, 385–387.
106 DeLucas LJ & Bugg CE (1987) New directions in
protein crystal growth. Trends Biotechnol 5, 188–193.
107 Leberman R (1985) Crystals in space. Science 230,
373–374.
108 Thaller C, Weaver LH, Eichele G, Wilson E, Karlsson
R & Jansonius JN (1981) Repeated seeding technique
for growing large single crystals of proteins. J Mol
Biol 147, 465–469.
109 Michel H (2012) Crystallization of membrane proteins.
In International Tables for Crystallography (Arnold E,
Himmel D & Rossmann M, eds), pp. 122–128. John
Wiley and Sons, Chicheter.
110 McPherson A, Koszelak S, Axelrod H, Day J,
Williams R, McGrath M, Robinson L & Cascio D
(1986) An experiment regarding crystallization of
soluble proteins in the presence of b-octyl-glucoside.J Biol Chem 261, 1969–1975.
111 Dock A-C, Lorber B, Moras D, Pixa G, Thierry J-C
& Gieg�e R (1984) Crystallization of transfer
ribonucleic acids. Biochimie 66, 179–201.
112 Carter CW & Carter CW Jr (1979) Protein
crystallization using incomplete factorial experiments.
J Biol Chem 254, 12219–12223.
113 Carter CW Jr, Baldwin ET & Frick L (1988)
Statistical design of experiments for protein crystal
growth and the use of a precrystallization assay.
J Cryst Growth 90, 60–73.
114 Cox MJ & Weber PC (1987) Experiments with
automated protein crystallization. J Appl Cryst 20,
366–373.
115 Anderson WF, Boodhoo A & Mol CF (1988) The
importance of purity in the crystallization of DNA
binding immunoglobulin Fab fragments. J Cryst
Growth 90, 153–159.
116 Arnold E, Himmel DM & Rossmann M (2012)
Crystallography of biological macromolecules. In
International Tables for Crystallography, 2nd edn.
John Wiley and Sons, Chichester.
117 Chayen NE, Boggon TJ, Cassetta A, Deacon A,
Gleichmann T, Habash J, Harrop SJ, Helliwell JR,
Nieh YP, Peterson MR et al. (1996) Trends and
challenges in experimental macromolecular
crystallography. Q Rev Biophys 29, 227–278.
118 Wiencek JM (1999) New strategies for protein crystal
growth. Annu Rev Biomed Eng 1, 505–534.
119 Chernov AA (1993) Present-day understanding of
crystal growth from aqueous solutions. Prog Crystal
Growth Charact Mat 26, 121–151.
120 Ri�es-Kautt M & Ducruix A (1991) Crystallization of
basic proteins by ion pairing. J Cryst Growth 110,
20–25.
121 Retailleau P, Ri�es-Kautt M & Ducruix A (1997) No
salting-in of lysozyme chloride observed at low ionic
strength over a large range of pH. Biophys J 73,
2156–2163.
122 Kadri A, Damak M, Lorber B, Gieg�e R & Jenner G
(2003) Pressure vs pH phase diagrams of two
lysozymes crystallized in agarose gels. High Press Res
23, 485–491.
123 Zhang Y & Cremer PS (2009) The inverse and direct
Hofmeister series for lysozyme. Proc Natl Acad Sci
USA 106, 15249–15253.
124 Carbonnaux C, Ri�es-Kautt M & Ducruix A (1995)
Relative effectiveness of various anions on the
solubility of acidic Hypoderma lineatum collagenase at
pH 7.2. Protein Sci 4, 2123–2128.
125 Ataka M, Shinzawa-Itoh K & Yoshikawa S (1992)
Phase diagram of a crystalline membrane protein,
bovine heart cytochrome c oxidase, in the salting-in
region. J Cryst Growth 122, 60–65.
126 Odhara T, Ataka M & Katsura T (1994) Phase
diagram determination to elucidate the crystal growth
of the photoreaction center from Rhodobacter
sphaeroides. Acta Cryst D 50, 639–642.
127 Saridakis EEG, Shaw Stewart PD, Lloyd LL & Blow
DM (1994) Phase diagram and dilution experiments in
the crystallization of carboxypeptidase G2. Acta Cryst
D 50, 293–297.
128 Sauter C, Lorber B, Kern D, Cavarelli J, Moras D &
Gieg�e R (1999) Crystallogenesis studies on aspartyl-
tRNA synthetase: use of phase diagram to improve
crystal quality. Acta Cryst D 55, 149–156.
129 Lorber B & Witz J (2008) An investigation of the
crystallogenesis of an icosahedral RNA plant virus
with solubility phase diagrams. Cryst Growth Des 8,
1522–1529.
130 Schellenberger P, Demangeat G, Lemaire O,
Ritzenthaler C, Bergdoll M, Olieric V, Sauter C &
Lorber B (2011) Strategies for the crystallization of
6488 FEBS Journal 280 (2013) 6456–6497 ª 2013 FEBS
Protein crystallization for structural biology R. Gieg�e

viruses: using phase diagrams and gels to produce 3D
crystals of Grapevine fanleaf virus. J Struct Biol 174,
344–351.
131 Christopher GK, Phipps AG & Gray RJ (1998)
Temperature-dependent solubility of selected proteins.
J Cryst Growth 191, 820–826.
132 Guilloteau JP, Rieskautt MM & Ducruix AF (1992)
Variation of lysozyme solubility as a function of
temperature in the presence of organic and inorganic
salts. J Cryst Growth 122, 223–230.
133 Asherie N, Ginsberg C, Blass S, Greenbaum A &
Knafo S (2008) Solubility of thaumatin. Cryst Growth
Des 8, 1815–1817.
134 Ng JD, Kuznetsov YG, Malkin AJ, Keith G, Gieg�e R
& McPherson A (1997) Visualization of RNA crystal
growth by atomic force microscopy. Nucleic Acids Res
25, 2582–2588.
135 Ri�es-Kautt M & Ducruix A (1997) Interferences
drawn from physico-chemical studies of
crystallogenesis and precrystalline state. Methods
Enzymol 276, 23–59.
136 Annunziata O, Asherie N, Lomakin A, Pande J, Ogun
O & Benedek GB (2002) Effect of polyethylene glycol
on the liquid–liquid phase transition in aqueous
protein solutions. Proc Natl Acad Sci USA 99,
14165–14170.
137 Vivar�es D, Belloni L, Tardieu A & Bonnet�e F (2002)
Catching the PEG-induced attractive interaction
between proteins. Eur Phys J E 9, 15–25.
138 Vivar�es D & Bonnet�e F (2004) Liquid–liquid phase
separations in urate oxidase/PEG mixtures:
characterization and implications for protein
crystallization. J Phys Chem B 108, 6498–6507.
139 George A & Wilson WW (1994) Predicting protein
crystallization from a dilute-solution property.
Acta Cryst D 50, 361–365.
140 Bonnet�e F, Finet S & Tardieu A (1999) Second virial
coefficient: variations with lysozyme crystallization
conditions. J Cryst Growth 196, 403–414.
141 Haas C, Drenth J & Wilson WW (1999) Relation
between the solubility of proteins in aqueous solutions
and the second virial coefficient of the solution.
J Phys Chem B 103, 2808–2811.
142 Jia YW & Liu XY (2005) Prediction of protein
crystallization based on interfacial and diffusion
kinetics. Applied Phys Lett 87, 103902.
143 Rosenberger F, Vekilov PG, Muschol M & Thomas
BR (1996) Nucleation and crystallization of globular
proteins – what do we know and what is missing.
J Cryst Growth 168, 1–27.
144 Nanev CN & Tsekova D (2000) Heterogeneous
nucleation of hen-egg-white lysozyme – molecular
approach. Cryst Res Technol 35, 189–195.
145 Galkin O & Vekilov PG (2000) Control of protein
crystal nucleation around the metastable liquid–liquid
phase boundary. Proc Natl Acad Sci USA 97,
6277–6281.
146 Penkova A, Chayen N, Saridakis E & Nanev CN
(2002) Nucleation of protein crystals in a wide
continuous supersaturation gradient. Acta Cryst D 58,
1606–1610.
147 Vekilov PG (2010) Nucleation. Cryst Growth Des 10,
5007–5019.
148 Erdemir D, Lee AY & Myerson AS (2009) Nucleation
of crystals from solution: classical and two-step
models. Acc Chem Res 42, 621–629.
149 Yau S-T & Vekilov PG (2000) Quasi-planar nucleus
structure in apoferritin crystallization. Nature 406,
494–497.
150 Saridakis E, Khurshid S, Govada L, Phan Q, Hawkins
D, Crichlow GV, Lolis E, Reddy SM & Chayen NE
(2011) Protein crystallization facilitated by molecularly
imprinted polymers. Proc Nat Acad Sci USA 108,
11081–11086.
151 Georgieva DG, Kuil ME, Oosterkamp TH,
Zandbergen HW & Abrahams JP (2007)
Heterogeneous nucleation of three-dimensional protein
nanocrystals. Acta Cryst D 63, 564–570.
152 Blow DM, Chayen NE, Lloyd LF & Saridakis E
(1994) Control of nucleation of protein crystals.
Protein Sci 3, 1638–1643.
153 Bolanos-Garcia VM (2005) The tight control of
nucleation in counterdiffusive flow regimes by
hyperfluorinated oils. J Cryst Growth 283, 215–221.
154 Hammadi Z, Astier JP, Morin R & Veesler S (2009)
Spatial and temporal control of nucleation by
localized DC electric field. Cryst Growth Des 9,
3346–3347.
155 Koizumi H, Fujiwara K & Uda S (2010) Role of the
electric double layer in controlling the nucleation rate
for tetragonal hen egg white lysozyme crystals by
application of an external electric field. Cryst Growth
Des 10, 2591–2595.
156 Han Q & Lin S-X (1996) A microcrystal selection
technique in protein crystallization. J Cryst Growth
168, 181–184.
157 Chernov AA (1984) . Modern Crystallography, III
Crystal Growth Vol. 36. Springer, Berlin.
158 DeMattei RC & Feigelson RS (1992) Controlling
nucleation in protein solutions. J Cryst Growth 122,
21–30.
159 Kakinouchi K, Adachi H, Matsumura H, Inoue T,
Murakami S, Mori Y, Koga Y, Takano K &
Kanaya S (2006) Effect of ultrasonic irradiation on
protein crystallization. J Cryst Growth 292,
437–440.
160 Malkin AJ, Kuznetsov YG, Land TA, DeYoreo JJ
& McPherson A (1995) Mechanisms of growth for
protein and virus crystals. Nat Struct Biol 2,
956–959.
FEBS Journal 280 (2013) 6456–6497 ª 2013 FEBS 6489
R. Gieg�e Protein crystallization for structural biology

161 McPherson A, Malkin AJ, Kuznetsov YG, Koszelak
S, Wells M, Jenkins G, Howard J & Lawson G (1999)
The effects of microgravity on protein crystallization:
evidence for concentration gradients around growing
crystals. J Cryst Growth 196, 572–586.
162 Nakada T, Sazaki G, Miyashita S, Durbin SD &
Komatsu H (1999) Direct AFM observations of
impurity effects on a lysozyme crystal. J Cryst Growth
196, 503–510.
163 Komatsu H, Miyashita S & Suzuki Y (1993)
Interferometric observation of the interfacial
concentration gradient layers around a lysozyme
crystal. Jpn J Appl Phys 32, L1855–L1857.
164 Vekilov PG, Ataka M & Katsura T (1993) Laser
Michelson interferometry investigation of protein
crystal growth. J Cryst Growth 130, 317–320.
165 Kuznetsov YG, Malkin AJ, Greenwood A &
McPherson A (1995) Interferometric studies of
growth kinetics and surface morphology in
macromolecular crystal growth: canavalin, thaumatin,
and turnip yellow mosaic virus. J Struct Biol 114,
184–196.
166 Boggon TJ, Chayen NE, Snell EH, Dong J,
Lautenschlager P, Potthast L, Siddons DP, Stojanoff
V, Gordon E, Thompson AW et al. (1998) Protein
crystal movements and fluid flows during
microgravity growth. Phil Trans R Soc Lond A 356,
1045–1061.
167 Miyashita S, Kurihara K, Hasegawa K, Sazaki G,
Nakada T, Durbin SD, Komatsu H, Fujishima M &
Koyama M (1998) Concentration field around a
growing crystal under microgravity. J Jpn Soc
Microgravity Appl 15, 577–581.
168 Ot�alora F, Garcia-Ruiz JM, Carotenuto L,
Castagnolo D, Novella ML & Chernov AA (2002)
Lysozyme crystal growth kinetics in microgravity.
Acta Cryst D 58, 1681–1689.
169 Luft JR, Albright DT, Baird JK & DeTitta GT
(1996) The rate of water equilibration in vapor-
diffusion crystallization. Dependence on the distance
from the droplet to the reservoir. Acta Cryst D 52,
1098–1106.
170 Baird JK (1999) Theory of protein crystal nucleation
and growth controlled by solvent evaporation. J Cryst
Growth 204, 553–562.
171 Martins PM, Rocha F & Damas AM (2008)
Understanding water equilibration fundamentals as a
step for rational protein crystallization. PLoS ONE 3,
e1998.
172 Streets AM & Quake SR (2010) Ostwald ripening of
clusters during protein crystallization. Phys Rev Lett
104, 178102.
173 Thiyagarajan P & Tiede DM (1994) Detergent micelle
structure and micelle-micelle interactions determined
by small-angle neutron-scattering under solution
conditions used for membrane-protein crystallization.
J Phys Chem 98, 10343–10351.
174 Tanaka S, Ataka M, Onuma K & Kubota T (2003)
Rationalization of membrane protein crystallization
with polyethylene glycol using a simple depletion
model. Biophys J 84, 3299–3306.
175 Landau EM & Rosenbusch JP (1996) Lipidic cubic
phases: a novel concept for the crystallization of
membrane proteins. Proc Natl Acad Sci USA 93,
14532–14535.
176 Luzzati V, Tardieu A, Gulik-Krzywicki T, Rivas E &
Reiss-Husson F (1968) Structure of the cubic phases
of lipid-water systems. Nature 220, 485–488.
177 Qutub Y, Reviakine I, Maxwell C, Navarro J, Landau
EM & Vekilov PG (2004) Crystallization of
transmembrane proteins in cubo: mechanisms of
crystal growth and defect formation. J Mol Biol 343,
1243–1254.
178 Caffrey M (2011) Crystallizing membrane proteins for
structure–function studies using lipidic mesophases.
Biochem Soc Trans 39, 725–732.
179 Kubicek J, Schlesinger R, Baeken C, Buldt G, Schafer
F & Labahn J (2012) Controlled in meso phase
crystallization – a method for the structural
investigation of membrane proteins. PLoS ONE 7,
e35458.
180 Judge RA, Snell EH & van der Woerd MJ (2005)
Extracting trends from two decades of microgravity
macromolecular crystallization history. Acta Cryst D
61, 763–771.
181 Snell EH, Weisgerber S, Helliwell JR, Weckert E,
H€olzer K & Schroer K (1995) Improvements in
lysozyme protein crystal perfection through
microgravity growth. Acta Cryst D 51, 1099–1102.
182 Ng JD, Lorber B, Gieg�e R, Koszelak S, Day J,
Greenwood A & McPherson A (1997) Comparative
analysis of thaumatin crystals grown on earth and in
microgravity. Acta Cryst D 53, 724–733.
183 Declercq J-P, Evrard C, Carter DC, Wright BS,
Etienne G & Parello J (1999) A crystal of a typical
EF-hand protein grown under microgravity diffracts
X-rays beyond 0.9 �A resolution. J Cryst Growth 196,
595–601.
184 Carter DC, Lim K, Ho JX, Wright BS, Twigg PD,
Miller TY, Chapman J, Keeling K, Ruble J, Vekilov
PG et al. (1999) Lower dimer impurity incorporation
may result in higher perfection of HEWL crystal
grown in lg – a case study. J Cryst Growth 196, 623–
637.
185 Ng JD (2002) Space-grown protein crystals are more
useful for structure determination. Ann N Y Acad Sci
974, 598–609.
186 Garcia-Ruiz JM & Moreno A (1994) Investigations on
protein crystal growth by the gel acupuncture method.
Acta Cryst D 50, 484–490.
6490 FEBS Journal 280 (2013) 6456–6497 ª 2013 FEBS
Protein crystallization for structural biology R. Gieg�e

187 Wakayama NI, Yin DC, Harata K, Kiyoshi T,
Fujiwara M & Tanimoto Y (2006) Macromolecular
crystallization in microgravity generated by a
superconducting magnet. Ann N Y Aca Sci 1077, 184–
193.
188 Taleb M, Didierjean C, Jelsch C, Mangeot JP,
Capelle B & Aubry A (1999) Crystallization of
proteins under an external electric field. J Cryst
Growth 200, 575–582.
189 Hansen CL, Skordalakes E, Berger JM & Quake SR
(2002) A robust and scalable microfluidic metering
method that allows protein crystal growth by free
interface diffusion. Proc Natl Acad Sci USA 99,
16531–16536.
190 Sazaki G (2009) Crystal quality enhancement by
magnetic fields. Prog Biophys Mol Biol 101, 45–55.
191 Qi J, Wakayama NI & Ataka M (2001) Magnetic
suppression of convection in protein crystal growth
processes. J Cryst Growth 232, 132–137.
192 Lorber B & Gieg�e R (1996) Containerless protein
crystallization in floating drops: application to crystal
growth monitoring under reduced nucleation
conditions. J Cryst Growth 168, 204–215.
193 Squires TM & Quake SR (2005) Microfluidics: fluid
physics at the nanoliter scale. Rev Mod Phys 77,
977–1026.
194 Sauter C, Lorber B & Gieg�e R (2002) Towards atomic
resolution with crystals grown in gel: the case of
thaumatin seen at room temperature. Proteins Struct
Funct Genet 48, 146–150.
195 Saijo S, Yamada Y, Sato T, Tanaka N, Matsui T,
Sazaki G, Nakajima K & Matsuura Y (2005)
Structural consequences of hen egg-white lysozyme
orthorhombic crystal growth in a high magnetic field:v
of X-ray diffraction intensity, conformational energy
searching and quantitative analysis of B factors and
mosaicity. Acta Cryst D 61, 207–217.
196 Sauter C, Ot�alora F, Gavira JA, Vidal O, Gieg�e R &
Garcia-Ruiz JM (2001) Structure of tetragonal hen egg
white lysozyme at 0.94 �A from crystals grown by the
counter-diffusion method. Acta Cryst D 57,
1119–1126.
197 Maki S, Murai R, Yoshikawa HY, Kitatani T,
Nakata S, Kawahara H, Hasenaka H, Kobayashi A,
Okada S, Sugiyama S et al. (2008) Protein
crystallization in a 100 nl solution with new stirring
equipment. J Synchrotron Radiat 15, 269–272.
198 Grant ML & Saville DA (1995) Long-term studies on
tetragonal lysozyme crystals grown in quiescent and
forced-convection environments. J Cryst Growth 153,
42–54.
199 Penkova A, Pan WC, Hodjaoglu F & Vekilov PG
(2006) Nucleation of protein crystals under the
influence of solution shear flow. Ann N Y Acad Sci
1077, 214–231.
200 Schall CA, Arnold E & Wiencek JM (1996) Enthalpy
of crystallization of hen egg-white lysozyme. J Cryst
Growth 165, 293–298.
201 Zielenkiewicz W (2008) Towards protein
crystallization – some thermodynamic studies.
J Therm Anal Calorim 92, 105–108.
202 Vekilov PG, Feeling-Taylor AR, Yau ST & Petsev D
(2002) Solvent entropy contribution to the free energy
of protein crystallization. Acta Cryst D 58, 1611–1616.
203 Vekilov PG, Feeling-Taylor AR, Petsev DN, Galkin
O, Nagel RL & Hirsch RE (2002) Intermolecular
interactions, nucleation, and thermodynamics of
crystallization of hemoglobin C. Biophys J 83, 1147–
1156.
204 Bergeron L, Filobelo LF, Galkin O & Vekilov PG
(2003) Thermodynamics of the hydrophobicity in
crystallization of insulin. Biophys J 85,
3935–3942.
205 Derewenda ZS & Vekilov PG (2006) Entropy and
surface engineering in protein crystallization. Acta
Cryst D 62, 116–124.
206 Yeh JI & Beale SI (2007) Calorimetric approaches to
characterizing effects of additives on protein
crystallization. Cryst Growth Des 7, 2134–2139.
207 Izumi K, Sawamura S & Ataka M (1996) X-ray
topography of lysozyme crystals. J Cryst Growth 168,
106–111.
208 Stojanoff V, Snell EF, Siddons DP & Helliwell JR
(1996) An old technique with a new application: X-ray
topography of protein crystals. Synchrotron Radiat
News 9, 25–26.
209 Nave C (1998) A description of imperfections in
protein crystals. Acta Cryst D 54, 848–853.
210 Caylor CL, Dobrianov I, Lemay SG, Kimmer C,
Kriminski S, Finkelstein KD, Zipfel W, Webb WW,
Thomas BR, Chernov AA et al. (1999)
Macromolecular impurities and disorder in protein
crystals. Proteins 36, 270–281.
211 Capelle B, Epelboin Y, Hartwig J, Moraleda AB,
Otalora F & Stojanoff V (2004) Characterization of
dislocations in protein crystals by means of
synchrotron double-crystal topography. J Appl Cryst
37, 67–71.
212 Lorber B, Sauter C, Ng JD, Zhu D-W, Gieg�e R, Vidal
O, Robert M-C & Capelle B (1999) Characterization
of protein and virus crystals by quasi-planar wave X-
ray topography: a comparison between crystals grown
in solution and in agarose gel. J Cryst Growth 204,
357–368.
213 Lovelace JJ & Borgstahl GEO (2010) The use of
orthogonal projection to visualize mosaic domains
from topographic data collected on protein crystals.
J Appl Cryst 43, 907–912.
214 Sawaura T, Fujii D, Shen M, Yamamoto Y, Wako
K, Kojima K & Tachibana M (2011)
FEBS Journal 280 (2013) 6456–6497 ª 2013 FEBS 6491
R. Gieg�e Protein crystallization for structural biology

Characterization of dislocations in monoclinic hen
egg-white lysozyme crystals by synchrotron
monochromatic-beam X-ray topography. J Cryst
Growth 318, 1071–1074.
215 Hasenaka H, Sugiyama S, Hirose M, Shimizu N,
Kitatani T, Takahashi Y, Adachi H, Takano K,
Murakami S, Inoue T et al. (2009) Femtosecond laser
processing of protein crystals grown in agarose gel. J
Cryst Growth 312, 73–78.
216 Dierks K, Meyer A, Einspahr H & Betzel C (2008)
Dynamic light scattering in protein crystallization
droplets: adaptations for analysis and optimization of
crystallization processes. Cryst Growth Des 8, 1628–
1634.
217 Wilson WW (2003) Light scattering as a diagnostic for
protein crystal growth – a practical approach. J Struct
Biol 142, 56–65.
218 Borgstahl GE (2007) How to use dynamic light
scattering to improve the likelihood of growing
macromolecular crystals. Methods Mol Biol 363, 109–
129.
219 Sanchez-Puig N, Sauter C, Lorber B, Gieg�e R &
Moreno A (2012) Predicting protein crystallizability
and nucleation. Protein Pept Lett 19, 725–731.
220 Kantardjieff KA & Rupp B (2004) Protein isoelectric
point as a predictor for increased crystallization
screening efficiency. Bioinformatics 20, 2162–2168.
221 Dupeux F, Rower M, Seroul G, Blot D & Marquez
JA (2011) A thermal stability assay can help to
estimate the crystallization likelihood of biological
samples. Acta Cryst D 67, 915–919.
222 Mizianty MJ & Kurgan LA (2012) CRYSpred:
accurate sequence-based protein crystallization
propensity prediction using sequence-derived structural
characteristics. Protein Pept Lett 19, 40–49.
223 Zhu DW, Garneau A, Mazumdar M, Zhou M, Xu GJ
& Lin S-X (2006) Attempts to rationalize protein
crystallization using relative crystallizability. J Struct
Biol 154, 297–302.
224 Wukovitz SW & Yeates TO (1995) Why protein
crystals favour some space-groups over others. Nat
Struct Biol 2, 1062–1067.
225 Banatao DR, Cascio D, Crowley CS, Fleissner MR,
Tienson HL & Yeates TO (2006) An approach to
crystallizing proteins by synthetic symmetrization.
Proc Natl Acad Sci USA 103, 16230–16235.
226 Zucker FH, Stewart C, Rosa JD, Kim J, Zhang L,
Xiao L, Ross J, Napuli AJ, Mueller N, Castaneda LJ
et al. (2010) Prediction of protein crystallization
outcome using a hybrid method. J Struct Biol 171, 64–
73.
227 Dale GE, Oefner C & D’Arcy A (2003) The protein as
a variable in protein crystallization. J Struct Biol 142,
88–97.
228 Dock-Bregeon A-C, Moras D & Gieg�e R (1999)
Nucleic acids and their complexes. In Crystallization
of Nucleic Acids and Proteins A Practical Approach,
2nd edn. (Ducruix A & Gieg�e R, eds), pp. 209–243.
IRL Press, Oxford.
229 Banci L, Bertini I, Cusack S, de Jong RN, Heinemann
U, Jones EY, Kozielski F, Maskos K, Messerschmidt
A, Owens R et al. (2006) First steps towards effective
methods in exploiting high-throughput technologies
for the determination of human protein structures of
high biomedical value. Acta Cryst D 62, 1208–1217.
230 Kim Y, Bigelow L, Borovilos M, Dementieva I,
Duggan E, Eschenfeldt W, Hatzos C, Joachimiak G,
Li H, Maltseva N et al. (2008) High-throughput
protein purification for X-ray crystallography
and NMR. Adv Protein Chem Struct Biol 75,
85–105.
231 Hughes SH & Stock AM (2012) Preparing
recombinant proteins for X-ray crystallography. In
International Tables for Crystallography (Arnold E,
Himmel D & Rossmann M, eds), pp. 75–91. John
Wiley and Sons, Chicheter.
232 Kigawa T, Yamaguchi-Nunokawa E, Kodama K,
Matsuda T, Yabuki T, Matsuda N, Ishitani R, Nureki
O & Yokoyama S (2002) Selenomethionine
incorporation into a protein by cell-free synthesis.
J Struct Funct Genomics 2, 29–35.
233 Yokoyama S (2003) Protein expression systems for
structural genomics and proteomics. Curr Opin Chem
Biol 7, 39–43.
234 Jaehme M & Michel H (2013) Evaluation of cell-free
protein synthesis for the crystallization of membrane
proteins – a case study on a member of the glutamate
transporter family from Staphylothermus marinus.
FEBS J 280, 1112–1125.
235 Holbrook SR, Holbrook EL & Walukiewicz HE
(2001) Crystallization of RNA. Cell Mol Life Sci 58,
234–243.
236 Lietzke SE, Barnes CL & Kundrot CE (1995)
Crystallization and structure determination of RNA.
Curr Opin Struct Biol 5, 645–649.
237 Wernimont A & Edwards A (2009) In situ proteolysis
to generate crystals for structure determination: an
update. PLoS ONE 4, e5094.
238 Huang J, Gong Y, Huang D, Haire L, Liu J & Peng
Y (2012) ‘Seeding’ with protease to optimize protein
crystallization conditions in in situ proteolysis. Acta
Cryst F 68, 606–609.
239 D’Arcy A, Stihle M, Kostrewa D & Dale G (1999)
Crystal engineering: a case study using the 24 kDa
fragment of the DNA gyrase B subunit from
Escherichia coli. Acta Cryst D 55, 1623–1625.
240 Gallagher DT, Smith NN, Kim SK, Robinson H &
Reddy PT (2009) Protein crystal engineering of
6492 FEBS Journal 280 (2013) 6456–6497 ª 2013 FEBS
Protein crystallization for structural biology R. Gieg�e

YpAC-IV using a strategy of excess charge reduction.
Cryst Growth Des 9, 3570–3574.
241 McPherson A, Nguyen C, Cudney R & Larson SB
(2011) The role of small molecule additives and
chemical modification in protein crystallization. Cryst
Growth Des 11, 1469–1474.
242 Kim Y, Quartey P, Li H, Volkart L, Hatzos C, Chang
C, Nocek B, Cuff M, Osipiuk J, Tan K et al. (2008)
Large-scale evaluation of protein reductive
methylation for improving protein crystallization. Nat
Methods 5, 853–854.
243 Lawson DM, Artymiuk PJ, Yewdall SJ, Smith JMA,
Livingstone JC, Treffry A, Luzzago A, Levi S, Arosio
P, Cesarini G et al. (1991) Solving the structure of
human H ferritin by genetically engineering
intermolecular crystal contacts. Nature 349, 541–544.
244 Binz HK, Amstutz P, Kohl A, Stumpp MT, Briand C,
Forrer P, Gr€utter MG & Pluckthun A (2004) High-
affinity binders selected from designed ankyrin repeat
protein libraries. Nat Biotechnol 22, 575–582.
245 Sennhauser G & Gr€utter MG (2008) Chaperone-
assisted crystallography with DARPins. Structure 16,
1443–1453.
246 Low C, Moberg P, Quistgaard EM, Hedren M,
Guettou F, Frauenfeld J, Haneskog L & Nordlund P
(2013) High-throughput analytical gel filtration
screening of integral membrane proteins for structural
studies. Biochim Biophys Acta 1830, 3497–3508.
247 Mariuzza RA, Jankovic DL, Boulot G, Amit AG,
Saludjian P, Le Guern A, Mazie JC & Poljak RJ
(1983) Preliminary crystallographic study of the
complex between the Fab fragment of a monoclonal
anti-lysozyme antibody and its antigen. J Mol Biol
170, 1055–1058.
248 Hunte C & Michel H (2002) Crystallisation of
membrane proteins mediated by antibody fragments.
Curr Opin Struct Biol 12, 503–508.
249 Jancarik J & Kim S-H (1991) Sparse matrix sampling;
a screening method for crystallization of proteins.
J Appl Cryst 24, 409–411.
250 Grimm C, Chari A, Reuter K & Fischer U (2010) A
crystallization screen based on alternative polymeric
precipitants. Acta Cryst D 66, 685–697.
251 Golden BL, Podell ER, Gooding AR & Cech TR
(1997) Crystals by design: a strategy for crystallization
of a ribozyme derived from the Tetrahymena group I
intron. J Mol Biol 270, 711–723.
252 Radaev S & Sun PD (2002) Crystallization of protein–protein complexes. J Appl Cryst 35, 674–676.
253 Newstead S, Ferrandon S & Iwata S (2008)
Rationalizing alpha-helical membrane protein
crystallization. Protein Sci 17, 466–472.
254 Parker JL & Newstead S (2012) Current trends in
alpha-helical membrane protein crystallization: an
update. Protein Sci 21, 1358–1365.
255 McPherson A & Cudney B (2006) Searching for silver
bullets: an alternative strategy for crystallizing
macromolecules. J Struct Biol 156, 387–406.
256 Hedderich T, Marcia M, Kopke J & Michel H (2011)
PICKScreens, a new database for the comparison of
crystallization screens for biological macromolecules.
Cryst Growth Des 11, 488–491.
257 Garlitz JA, Summers CA, Flowers RA 2nd &
Borgstahl GE (1999) Ethylammonium nitrate: a
protein crystallization reagent. Acta Cryst D 55, 2037–
2038.
258 Pusey ML, Paley MS, Turner MB & Rogers RD
(2007) Protein crystallization using room temperature
ionic liquids. Cryst Growth Des 7, 787–793.
259 Judge RA, Takahashi S, Longenecker KL, Fry EH,
Abad-Zapatero C & Chiu ML (2009) The effect of
ionic liquids on protein crystallization and
X-ray diffraction studies. Cryst Growth Des 9, 3463–
3469.
260 Chen XW, Ji YP & Wang JH (2010) Improvement on
the crystallization of lysozyme in the presence of
hydrophilic ionic liquid. Analyst 135, 2241–2248.
261 Santos SP, Bandeiras TM, Pinto AF, Teixeira M,
Carrondo MA & Romao CV (2012) Thermofluor-
based optimization strategy for the stabilization and
crystallization of Campylobacter jejuni
desulforubrerythrin. Protein Expres Purif 81, 193–200.
262 Thakur AS, Robin G, Guncar G, Saunders NF,
Newman J, Martin JL & Kobe B (2007) Improved
success of sparse matrix protein crystallization
screening with heterogeneous nucleating agents. PLoS
ONE 2, e1091.
263 Nederlof I, Hosseini R, Georgieva D, Luo JH, Li DF
& Abrahams JP (2011) A straightforward and robust
method for introducing human hair as a nucleant into
high throughput crystallization trials. Cryst Growth
Des 11, 1170–1176.
264 Anand K, Pal D & Hilgenfeld R (2002) An overview
on 2-methyl-2,4-pentanediol in crystallization and in
crystals of biological macromolecules. Acta Cryst D
58, 1722–1728.
265 Parmar AS, Gottschall PE & Muschol M (2007) Pre-
assembled clusters distort crystal nucleation kinetics in
supersaturated lysozyme solutions. Biophys Chem 129,
224–234.
266 Lohkamp B & Dobritzsch D (2008) A mixture of
fortunes: the curious determination of the structure of
Escherichia coli BL21 Gab protein. Acta Cryst D 64,
407–415.
267 Kors CA, Wallace E, Davies DR, Li L, Laible PD &
Nollert P (2009) Effects of impurities on membrane-
protein crystallization in different systems. Acta Cryst
D 65, 1062–1073.
268 Giffard M, Ferte N, Ragot F, El Hajji M, Castro B &
Bonnet�e F (2011) Urate oxidase purification by
FEBS Journal 280 (2013) 6456–6497 ª 2013 FEBS 6493
R. Gieg�e Protein crystallization for structural biology

salting-in crystallization: towards an alternative to
chromatography. PLoS ONE 6, e19013.
269 Plomp M, McPherson A & Malkin AJ (2003) Repair
of impurity-poisoned protein crystal surfaces. Proteins
50, 486–495.
270 Ferreira C, Rocha FA, Damas AM & Martins PM
(2012) Running away from thermodynamics to
promote or inhibit crystal growth. Cryst Growth Des
12, 40–43.
271 Adachi H, Takano K, Yoshimura M, Mori Y &
Sasaki T (2002) Promotion of large protein crystal
growth with stirring solution. Jpn J Appl Phys 41,
L1025–L1027.
272 Newman J, Egan D, Walter TS, Meged R, Berry I,
Ben Jelloul M, Sussman JL, Stuart DI & Perrakis A
(2005) Towards rationalization of crystallization
screening for small- to medium-sized academic
laboratories: the PACT/JCSG+ strategy. Acta Cryst D
61, 1426–1431.
273 Busso D, Stierle M, Thierry J-C & Moras D (2008)
Automated recombinant protein expression screening
in Escherichia coli. Methods Mol Biol 426, 175–186.
274 Kim Y, Babnigg G, Jedrzejczak R, Eschenfeldt WH,
Li H, Maltseva N, Hatzos-Skintges C, Gu M,
Makowska-Grzyska M, Wu R et al. (2011) High-
throughput protein purification and quality assessment
for crystallization. Methods 55, 12–28.
275 Ericsson UB, Hallberg BM, Detitta GT, Dekker N &
Nordlund P (2006) Thermofluor-based high-
throughput stability optimization of proteins for
structural studies. Anal Biochem 357, 289–298.
276 Vallotton P, Sun CM, Lovell D, Fazio VJ & Newman
J (2010) DroplIT, an improved image analysis method
for droplet identification in high-throughput
crystallization trials. J Appl Cryst 43, 1548–1552.
277 Khurshid S, Haire LF & Chayen NE (2010)
Automated seeding for the optimization of crystal
quality. J Appl Cryst 43, 752–756.
278 Chayen NE (2007) Optimization techniques for
automation and high throughput. Methods Mol Biol
363, 175–190.
279 Sugahara M, Shimizu K, Asada Y, Fukunishi H,
Kodera H, Fujii T, Osada E, Kasazaki T, Sawada T,
Chikusa H et al. (2010) Autolabo: an automated
system for ligand-soaking experiments with protein
crystals. J Appl Cryst 43, 940–944.
280 Viola R, Carman P, Walsh J, Miller E, Benning M,
Frankel D, McPherson A, Cudney B & Rupp B (2007)
Operator-assisted harvesting of protein crystals using a
universal micromanipulation robot. J Appl Cryst 40,
539–545.
281 Makino M, Wada I, Mizuno N, Hirata K, Shimizu N,
Hikima T, Yamamoto M & Kumasaka T (2012) Fine-
needle capillary mounting for protein microcrystals. J
Appl Cryst 45, 785–788.
282 Bingel-Erlenmeyer R, Olieric V, Grimshaw JPA,
Gabadinho J, Wang X, Ebner SG, Isenegger A,
Schneider R, Schneider J, Glettig W et al. (2011) SLS
crystallization platform at beamline X06DA – a fully
automated pipeline enabling in situ X-ray diffraction
screening. Cryst Growth Des 11, 916–923.
283 Page R & Stevens RC (2004) Crystallization data
mining in structural genomics: using positive and
negative results to optimize protein crystallization
screens. Methods 34, 373–389.
284 Newman J, Bolton EE, Muller-Dieckmann J, Fazio VJ,
Gallagher DT, Lovell D, Luft JR, Peat TS, Ratcliffe D,
Sayle RA et al. (2012) On the need for an international
effort to capture, share and use crystallization screening
data. Acta Cryst F 68, 253–258.
285 Mueller U, Nyarsik L, Horn M, Rauth H, Przewieslik
T, Saenger W, Lehrach H & Eickhoff H (2001)
Development of a technology for automation and
miniaturization of protein crystallization. J Biotech 85,
7–14.
286 Ng JD, Stevens RC & Kuhn P (2008) Protein
crystallization in restricted geometry: advancing old
ideas for modern times in structural proteomics.
Methods Mol Biol 426, 363–376.
287 Kuil ME, Bodenstaff ER, Hoedemaker FJ &
Abrahams JP (2002) Protein nano-crystallogenesis.
Enz & Microb Technol 30, 262–265.
288 Schneider TR (2008) Synchrotron radiation:
micrometer-sized X-ray beams as fine tools for
macromolecular crystallography. HFSP J 2, 302–306.
289 Hansen CL, Sommer MO & Quake SR (2004)
Systematic investigation of protein phase behavior
with a microfluidic formulator. Proc Natl Acad Sci
USA 101, 14431–14436.
290 Hansen CL, Classen S, Berger JM & Quake SR (2006)
A microfluidic device for kinetic optimization of
protein crystallization and in situ structure
determination. J Am Chem Soc 128, 3142–3143.
291 Zheng B, Roach LS & Ismagilov RF (2003) Screening
of protein crystallization conditions on a microfluidic
chip using nanoliter-size droplets. J Am Chem Soc
125, 11170–11171.
292 Yadav MK, Gerdts CJ, Sanishvili R, Smith WW,
Roach LS, Ismagilov RF, Kuhn P & Stevens RC
(2005) In situ data collection and structure refinement
from microcapillary protein crystallization. J Appl
Cryst 38, 900–905.
293 Shim JU, Cristobal G, Link DR, Thorsen T, Jia Y,
Piattelli K & Fraden S (2007) Control and
measurement of the phase behavior of aqueous
solutions using microfluidics. J Am Chem Soc 129,
8825–8835.
294 Selimovic S, Jia Y & Fraden S (2009) Measuring the
nucleation rate of lysozyme using microfluidics. Cryst
Growth Des 9, 1806–1810.
6494 FEBS Journal 280 (2013) 6456–6497 ª 2013 FEBS
Protein crystallization for structural biology R. Gieg�e

295 Selimovic S, Gobeaux F & Fraden S (2010) Mapping
and manipulating temperature-concentration phase
diagrams using microfluidics. Lab Chip 10, 1696.
296 Ng JD, Clark PJ, Stevens RC & Kuhn P (2008) In situ
X-ray analysis of protein crystals in low-birefringent
and X-ray transmissive plastic microchannels. Acta
Cryst D 64, 189–197.
297 Dhouib K, Khan Malek C, Pfleging W, Gauthier-
Manuel B, Duffait R, Thuillier G, Ferrigno R,
Jacquamet L, Ohana J, Ferrer JL et al. (2009)
Microfluidic chips for the crystallization of
biomacromolecules by counter-diffusion and on-chip
crystal X-ray analysis. Lab Chip 9, 1412–1421.
298 Gerdts CJ, Elliott M, Lovell S, Mixon MB, Napuli
AJ, Staker BL, Nollert P & Stewart L (2008) The
plug-based nanovolume Microcapillary Protein
Crystallization System (MPCS). Acta Cryst D 64,
1116–1122.
299 Yu Y, Wang X, Oberthur D, Meyer A, Perbandt M,
Duan L & Kang Q (2012) Design and application of a
microfluidic device for protein crystallization using an
evaporation-based crystallization technique. J Appl
Cryst 45, 53–60.
300 Stura E (1999) Seeding techniques. In Crystallization
of Nucleic Acids and Proteins: A Practical Approach,
2nd edn. (Ducruix A & Gieg�e R, eds), pp. 177–208.
IRL Press, Oxford.
301 Bergfors T (2003) Seeds to crystals. J Struct Biol 142,
66–76.
302 Georgiev A, Vorobiev S, Edstrom W, Song T, Laine
A, Hunt J & Allen P (2006) Automated streak-seeding
with micromachined silicon tools. Acta Cryst D 62,
1039–1045.
303 Ireton GC & Stoddard BL (2004) Microseed matrix
screening to improve crystals of yeast cytosine
deaminase. Acta Cryst D 60, 601–605.
304 D’Arcy A, Villard F & Marsh M (2007) An
automated microseed matrix-screening method for
protein crystallization. Acta Cryst D 63, 550–554.
305 Walter TS, Mancini EJ, Kadlec J, Graham SC,
Assenberg R, Ren J, Sainsbury S, Owens RJ, Stuart
DI, Grimes JM et al. (2008) Semi-automated
microseeding of nanolitre crystallization experiments.
Acta Cryst F 64, 14–18.
306 Yoshikawa HY, Hosokawa Y, Murai R, Sazaki G,
Kitatani T, Adachi H, Inoue T, Matsumura H,
Takano K, Murakami S et al. (2012) Spatially precise,
soft microseeding of single protein crystals by
femtosecond laser ablation. Cryst Growth Des 12,
4334–4339.
307 DeLucas LJ, Hamrick D, Cosenza L, Nagy L,
McCombs D, Bray T, Chait A, Stoops B, Belgovskiy
A, Wilson WW et al. (2005) Protein crystallization:
virtual screening and optimization. Prog Biophys Mol
Biol 88, 285–309.
308 Iino H, Naitow H, Nakamura Y, Nakagawa N, Agari
Y, Kanagawa M, Ebihara A, Shinkai A, Sugahara M,
Miyano M et al. (2008) Crystallization screening test
for the whole-cell project on Thermus thermophilus
HB8. Acta Cryst F 64, 487–491.
309 Kuta Smatanova I, Gavira JA, Rezacova P, Vacha F
& Garcia-Ruiz JM (2006) New techniques for
membrane protein crystallization tested on
photosystem II core complex of Pisum sativum.
Photosynth Res 90, 255–259.
310 Zhang CY, Yin DC, Lu QQ, Guo YZ, Guo WH,
Wang XK, Li HS, Lu HM & Ye YJ (2008) Cycling
temperature strategy: a method to improve the
efficiency of crystallization condition screening of
proteins. Cryst Growth Des 8, 4227–4232.
311 Meged R, Dym O & Sussman JL (2008) High
throughput pH optimization of protein crystallization.
Methods Mol Biol 426, 411–418.
312 Budayova-Spano M, Dauvergne F, Audiffren M,
Bactivelane T & Cusack S (2007) A methodology and
an instrument for the temperature-controlled
optimization of crystal growth. Acta Cryst D 63,
339–347.
313 Crespo R, Martins PM, Gales L, Rocha F & Damas
AM (2010) Potential use of ultrasound to promote
protein crystallization. J Appl Cryst 43, 1419–1425.
314 Newman J (2006) A review of techniques for
maximizing diffraction from a protein crystal in stilla.
Acta Cryst D 62, 27–31.
315 Rayment I (2002) Small-scale batch crystallization of
proteins revisited: an underutilized way to grow large
protein crystals. Structure 10, 147–151.
316 Oksanen E, Blakeley MP, Bonnet�e F, Dauvergne MT,
Dauvergne F & Budayova-Spano M (2009) Large
crystal growth by thermal control allows
combined X-ray and neutron crystallographic studies
to elucidate the protonation states in Aspergillus
flavus urate oxidase. J Royal Soc Interface 6,
S599–S610.
317 Hazemann I, Dauvergne MT, Blakeley MP, Meilleur
F, Haertlein M, Van Dorsselaer A, Mitschler A,
Myles DA & Podjarny A (2005) High-resolution
neutron protein crystallography with radically small
crystal volumes: application of perdeuteration to
human aldose reductase. Acta Cryst D 61,
1413–1417.
318 Ferr�e-D’Amar�e AR, Zhou K & Doudna JA (1998) A
general module for RNA crystallization. J Mol Biol
279, 621–631.
319 Ravindran PP, Heroux A & Ye JD (2011)
Improvement of the crystallizability and expression of
an RNA crystallization chaperone. J Biochem 150,
535–543.
320 Ke A & Doudna JA (2004) Crystallization of RNA
and RNA–protein complexes. Methods 34, 408–414.
FEBS Journal 280 (2013) 6456–6497 ª 2013 FEBS 6495
R. Gieg�e Protein crystallization for structural biology

321 Ferr�e-D’Amar�e AR (2010) Use of the spliceosomal
protein U1A to facilitate crystallization and structure
determination of complex RNAs. Methods 52, 159–167.
322 Ye JD, Tereshk V, Frederiksen JK, Koide A,
Fellouse FA, Sidhu SS, Koide S, Kossiakoff AA &
Piccirilli JA (2007) Chaperone-assisted RNA
crystallography: selection and co-crystallization of a
Fab-Delta C209 P4-P6 complex. J Biomol Struct Dyn
24, 688–689.
323 Oubridge C, Ito N, Teo C-H, Fearnley I & Nagai K
(1995) Crystallization of RNA–protein complexes. II.
The application of protein engineering for
crystallization of the UIA protein–RNA complex. J
Mol Biol 249, 409–423.
324 Gieg�e R, Touz�e E, Lorber B, Th�eobald-Dietrich A &
Sauter C (2008) Crystallogenesis trends of free and
liganded aminoacyl-tRNA synthetases. Cryst Growth
Des 8, 4297–4306.
325 Martins P, P€essoa J, Sarkany Z, Rocha F & Damas A
(2008) Rationalizing protein crystallization screenings
through water equilibration theory and protein
solubility data. Cryst Growth Des 8, 4233–4243.
326 Itoh Y, Brocker MJ, Sekine S, Hammond G, Suetsugu
S, S€oll D & Yokoyama S (2013) Decameric
SelA*tRNASec ring structure reveals mechanism of
bacterial selenocysteine formation. Science 340, 75–78.
327 Grimm C, Klebe G, Ficner R & Reuter K (2000)
Screening orthologs as an important variable in
crystallization: preliminary X-ray diffraction studies of
the tRNA-modifying enzyme S-adenosyl-methionine:
tRNA ribosyl transferase isomerase. Acta Cryst D 56,
484–488.
328 Poodt PWG, Heijna MCR, Christianen PCM, van
Enckevort WJP, de Grip WJ, Tsukamoto K, Maan JC
& Vlieg E (2006) Using gradient magnetic fields to
suppress convection during crystal growth. Cryst
Growth Des 6, 2275–2280.
329 Espinoza-Montero PI, Moreno-Narvaez ME,
Frontana-Uribe BA, Stojanoff V & Moreno A (2013)
Investigations on the use of graphite electrodes using a
Hull-type growth cell for ellectrochemically assisted
protein crystallization. Cryst Growth Des 13, 590–598.
330 Larson SB, Day JS, Nguyen C, Cudney R &
McPherson A (2008) Progress in the development of
an alternatuve approach to macromolecule
crystallization. Cryst Growth Des 8, 3038–3052.
331 Numata T, Ikeuchi Y, Fukai S, Adachi H,
Matsumura H, Takano K, Murakami S, Inoue T,
Mori Y, Sasaki T et al. (2006) Crystallization and
preliminary X-ray analysis of the tRNA thiolation
enzyme MnmA from Escherichia coli complexed with
tRNAGlu. Acta Cryst F 62, 368–371.
332 Li L, Mustafi D, Fu Q, Tereshko V, Chen DL, Tice
JD & Ismagilov RF (2006) Nanoliter microfluidic
hybrid method for simultaneous screening and
optimization validated with crystallization of
membrane proteins. Proc Natl Acad Sci USA 103,
19243–19248.
333 Sato S, Mimasu S, Sato A, Hino N, Sakamoto K,
Umehara T & Yokoyama S (2011) Crystallographic
study of a site-specifically cross-linked protein complex
with a genetically incorporated photoreactive amino
acid. Biochemistry 50, 250–257.
334 Yoshikawa S, Kukimoto-Niino M, Parker L, Handa
N, Terada T, Fujimoto T, Terazawa Y, Wakiyama M,
Sato M, Sano S et al. (2013) Structural basis for the
altered drug sensitivities of non-small cell lung cancer-
associated mutants of human epidermal growth factor
receptor. Oncogene 32, 27–38.
335 Acosta F, Eid D, Marin-Garcia L, Frontana-Uribe
BA & Moreno A (2007) From cytochrome c crystals
to a solid-state electron-transfer device. Cryst Growth
Des 7, 2187–2191.
336 Judge RA, Johns MR & White ET (1995) Protein
purification by bulk crystallization: the recovery of
ovalbumin. Biotech and Bioengin 48, 316–323.
337 Brader ML, Sukumar M, Pekar AH, McClellan DS,
Chance RE, Flora DB, Cox AL, Irwin L & Myers SR
(2002) Hybrid insulin cocrystals for controlled release
delivery. Nat Biotechnol 20, 800–804.
338 Yonath A (2010) Polar bears, antibiotics, and the
evolving ribosome (Nobel Lecture). Angew Chem Int
Ed Engl 49, 4341–4354.
339 Koopmann R, Cupelli K, Redecke L, Nass K,
Deponte DP, White TA, Stellato F, Rehders D, Liang
M, Andreasson J, et al. (2012) In vivo protein
crystallization opens new routes in structural biology.
Nat Methods 9, 259–262.
340 Domike KR&Donald AM (2009) Kinetics of spherulite
formation and growth: salt and protein concentration
dependence on proteins beta-lactoglobulin and insulin.
Int J Biol Macromol 44, 301–310.
341 Martins PM (2013) True and apparent inhibition of
amyloid fibril formation. Prion 7, 136–139.
342 Kam M, Perl-Treves D, Sfez R & Addadi L (1994)
Specificity in the recognition of crystals by antibodies.
J Mol Recognit 7, 257–264.
343 Heijna MCR, Theelen MJ, van Enckevort WJP &
Vlieg E (2007) Spherulitic growth of hen egg-white
lysozyme crystals. J Phys Chem B 111, 1567–1573.
344 Kulkarni CV (2012) Lipid crystallization: from self-
assembly to hierarchical and biological ordering.
Nanoscale 4, 5779–5791.
345 Perederina A & Krasilnikov AS (2012) Crystallization
of RNA–protein complexes: from synthesis and
purification of individual components to crystals.
Methods Mol Biol 905, 123–143.
346 Mesters JR & Hilgenfeld R (2007) Protein
glycosylation, sweet to crystal growth? Cryst Growth
Des 7, 2251–2253.
6496 FEBS Journal 280 (2013) 6456–6497 ª 2013 FEBS
Protein crystallization for structural biology R. Gieg�e

347 Sugiyama S, Shimizu N, Sazaki G, Hirose M,
Takahashi Y, Maruyama M, Matsumura H, Adachi
H, Takano K, Murakami S et al. (2013) A novel
approach for protein crystallization by a synthetic
hydrogel with thermoreversible gelation polymer.
Cryst Growth Des 13, 1899–1904.
348 Gangloff M, Moreno A & Gay NJ (2013) Liesegang-
like patterns of Toll crystals grown in gel. J Appl
Crystallogr 46, 337–345.
349 Shah UV, Allenby MC, Williams DR & Heng JYY
(2012) Crystallization of proteins at ultralow
supersaturations using novel three-dimensional
nanotemplates. Cryst Growth Des 12, 1772–1777.
350 Villasenor AG, Wong A, Shao A, Garg A, Donohue
TJ, Kuglstatter A & Harris SF (2012) Nanolitre-scale
crystallization using acoustic liquid-transfer
technology. Acta Cryst D 68, 893–900.
351 Bonnefond L, Schellenberger P, Basquin J, Demangeat
G, Ritzenthaler C, Chenevert R, Balg C, Frugier M,
Rudinger-Thirion J, Gieg�e R et al. (2011) Exploiting
protein engineering and crystal polymorphism for
successful X-ray structure determination. Cryst Growth
Des 11, 4334–4343.
352 Minton AP (2001) The influence of macromolecular
crowding and macromolecular confinement on
biochemical reactions in physiological media. J Biol
Chem 276, 10577–10580.
Supporting information
Additional supporting information may be found in
the online version of this article at the publisher’s web
site:Data S1 to S10. Complete bibliography for Table 1–8
and Figs 1–4 : references [353–488] are supplemental.
Data S11. Contains additional bibliography and com-
ments on ‘Books, historical accounts & reviews on
crystal growth’, on ‘ICCBM proceedings’, and on
‘Specialized reviews & research articles’.
FEBS Journal 280 (2013) 6456–6497 ª 2013 FEBS 6497
R. Gieg�e Protein crystallization for structural biology







