
Con@. Biochem. Physiol., 1972, Vol. 42A, pp. 1049 to 1053. Pergamon Press. Printed in Great Britain
ASPECTS OF NITROGEN EXCRETION DURING AESTIVATION IN THE AMPHIBIOUS SNAIL,
PILA GLOBOSA-I. EVIDENCE FOR NEUROENDOCRINE INVOLVEMENT IN THE PATTERN OF EXCRETION
NAYEEMUNNISA
Department of Zoology, Bangalore University, Bangalore, S. India
(Recked 19 October 1971)
Abstract-l. The ammonia, urea and uric acid contents of the mantle were determined in active and aestivated snails.
2. Ammonia decreases significantly (P> 0.001) and urea and uric acid increase on aestivation (percentage increase is 90 and P> 0.05 respectively). The adaptive significance of these changes is explained.
3. Neuroendocrine influence on the pattern of nitrogen excretion was studied by injecting extracts from pleuropedal and cerebral ganglia of active and aestivated snails into the foot muscle of the aestivated and active group.
4. On injection of the ganglionic extract from the active snail, aestivated animals showed a significant increase in ammonia (P > O-02) and a decrease in urea and uric acid of the mantle (percentage change is 7 and 53 respectively).
5. Active pila injected with the extract from the aestivated animal showed a significant decrease in ammonia (P < 0.001) and a significant increase in uric acid (P>O*OS).
INTRODUCTION
ASPECTS of nitrogen excretion in pulmonates have been extensively studied (Baldwin & Needham, 1934; Baldwin, 1935; Needham, 1935). The available information has been recently reviewed by Florkin (1966) and Potts (1968). Jezewska et al. (1963) reported that in the snail Helix pomuticz 90 per cent of the nitrogen is excreted in the form of purines. Duerr (1967) determined the content of uric acid in pulmonates and prosobranches. Speeg & Campbell (1968a) pre- sented information on aestivating snails and purine biosynthesis. Recently, Speeg & Campbell (1968a, b, 1969) h ave studied problems of nitrogen excretion in pulmonates in relation to enzymes involved in the metabolism of nitrogenous substances.
The amphibious snail, Pila globosu, is known to aestivate during drought and also in summer (Reddy, 1963, 1965). There are several reports concerning the metabolic as well as the biochemical changes in these snails during aestivation (Reddy, 1963, 1965). P. globosa resorts to uricotelism during aestivation (Sexana, 1955). Reddy & Swami (1963) have reported that protein and purine metabolism is affected in this snail on aestivation leading to the accumulation of urea and uric
1049

1050 NAYEEMUNNISA
acid in various tissues. Mostly the mantle is known to accumulate uric acid and urea. But there is no information on the mechanisms of regulation (nervous or hormonal) of excretion in molluscs (Potts, 1968). The results of the present study indicate neurohumoral regulation of the pattern of nitrogen excretion in the amphibious snail, P. globosa, on aestivation.
MATERIALS AND METHODS
The common amphibious snail, Pila gZobosa, were purchased and stocked in laboratory aquaria. The animals were divided into two batches. One batch, consisting of about two dozen animals, was left in the aquarium tank maintained at the laboratory temperature (26 k 3°C). These animals were the normal active pila (controls). The other batch was buried in sand at a temperature of 35 k 2°C in a temperature bath for 4 months for the purpose of aestivation. Active animals were fed with pieces of cabbage leaves at regular intervals.
The whole mantle from individuals was utilized for the analysis. To study the neuro- hormonal control of the nitrogen excretory pattern, normal and aestivated pila were injected with the aqueous extract (O-5 ml) of the pleuropedal and cerebral ganglia of aestivated and normal animals respectively. Injections were given in the early hours of the day (between 8 a.m. and 9 a.m.), into the muscle of the foot. Injected animals were sacrificed 2i hr after injection, and the mantle was made use of for analysis. Active and aestivated animals injected with ganglionic extracts from active and aestivated animals respectively constituted the controls. These animals were also similarly sacrificed 2) hr after injection and used for analysis. Thus the total mantle mass from active, aestivated, active injected (with normal and aestivated extracts) and aestivated injected (with aestivated and normal extracts) sets were assayed for the determination of ammonia, urea and uric acid.
The mantle was weighed and homogenized in 21 ml of distilled water. One ml of the homogenate was utilized for the estimation of uric acid and 10 ml each was used for analyses of ammonia and urea respectively.
Analyses of ammonia and urea contents were made by the method of Vanslyke & Cullen as given in Hawk’s Physiological Chemistry by Oser (1965). The uric acid content was measured by the cyanide-free calorimetric procedure as described in Practical Physiological Chemistry by Hawk et al., (1954).
RESULTS AND DISCUSSION
The results obtained are summarized in Tables 1 and 2. From the results given in Table 1 it is clear that on aestivation ammonia
decreases significantly (I’> 0.001) while uric acid shows a significant increase (P> 0.05). There was a considerable increase in the urea content (percentage increase was 90), but this was not statistically significant.
On injection of the nervous extract from the active animal, aestivated animals showed a significant increase in ammonia (P> O-02) and a decrease in urea (7 per cent) and uric acid (53 per cent). But these changes in urea and uric acid are statistically not significant compared to the control values. Likewise active pila injected with the extract from an aestivated animal showed a significant decrease in the level of ammonia (I’> 0.001). Uric acid increased significantly (P> O-05) and urea content showed a decrease (11 per cent). In all the injected cases men- tioned above t-values are calculated between injected controls and animals injected with the extract from the other set.
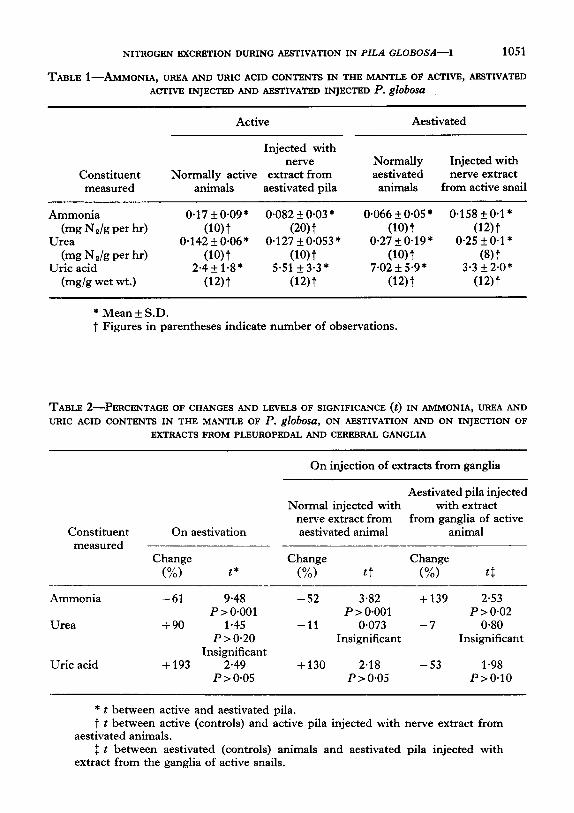
NITROGEN EXCRBTION DURING ABSTIVATION IN PILA GLOBOSA-I 1051
TABLE ~-AMMONIA, UREA AND URIC ACID CONTENTS IN THE MANTLE OF ACTIVE, AESTIVATED ACTIVE INJECTED AND ARSTIVATBD INJECTBD P. gJObOsU
Constituent measured
Active
Injected with nerve
Normally active extract from animals aestivated pila
Aestivated
Normally Injected with aestivated nerve extract
animals from active snail
Ammonia 0.17 *0.09* 0.082 ?I 0.03 * 0.066 + 0.05 * 0.158 * 0*1* (mg Na/g per hr) uwt (20) t (10) t (12) t
Urea 0.142f0.06* 0.127 + 0.053 * 0.27+0.19* 0.25 * 0.1* (mg Na/g per hr) (10) t (10) t wt wt
Uric acid 2*4+ l-8* 5.51 f 3.3 * 7.02 f 5.9* 3.3 + 2*0* (mg/g wet wt.) wt wt wt (12) t
* Mean + S.D. t Figures in parentheses indicate number of observations.
TABLE ~-PERCENTAGE 0~ CHANGES AND LEVELS OF SIGNIFICANCE (t) IN AMMONIA, UREA AND URIC ACID CONTENTS IN THE MANTLE 0F P. globosa, 0N ABSTIVATION AND ON INJECTION OF
E?XRACTS FROM PLEUROPEDAL AND CEREBRAL GANGLIA
On injection of extracts from ganglia
Aestivated pila injected Normal injected with with extract
nerve extract from from ganglia of active Constituent On aestivation aestivated animal animal
measured Change Change Change
(%) t* (%I tt (%I t:
Ammonia -61 9.48 -52 3.82 +139 2.53 P > 0.001 P > 0.001 P>O.O2
Urea +90 1.45 -11 0.073 -7 0.80 P > 0.20 Insignificant Insignificant
Insignificant Uric acid + 193 2.49 +130 2.18 -53 1.98
P>O*OS P>O*OS P>O*lO
* t between active and aestivated pila. t t between active (controls) and active pila injected with nerve extract from
aestivated animals. : t between aestivated (controls) animals and aestivated pila injected with
extract from the ganglia of active snails.

1052 NAYEEMUNNISA
From the results presented above two general conclusions can be drawn, concerning the contents of ammonia, urea and uric acid:
(a) Ammonia decreases on aestivation and urea and uric acid show an increase. (b) On injection of extracts considerable changes occur in the nitrogen-
excretory pattern. Extracts from the ganglia of aestivated animals injected into active animals bring about changes in the amount of ammonia urea and uric acid which closely approximate the trend of these constituents in the naturally aestivated animals. Thus, for example, the ammonia content of active pila decreases on injection of extract from aestivated animals (compared with injected controls). Uric acid increased significantly. Only urea showed a decrease, contrary to the expected increase.
Following the injection of extract from the active animal, aestivated snails showed changes which are comparable with the trend of occurrence of these constituents in the naturally active group. For instance, uric acid content decreased, ammonia showed a significant increase (P> O-02) and a slight decrease in urea was found. Therefore the nature of change of these components has followed the pattern of distribution of ammonia urea and uric acid in active pila.
Thus from the facts presented above it can be said that active forms are pre- dominantly ammonotelic, and uric acid found in them may presumably be due to an incomplete breakdown of uric acid derived from purine metabolism. Likewise the urea reported in active forms may quite likely be formed by the action of arginase on exogenous arginine (Potts, 1968). On aestivation the animal resorts to uricotel- ism as the nitrogen metabolism (protein metabolism) is affected and, as a conse- quence, urea and uric acid accumulate (Reddy, 1963, 1965). The accumulation of urea and uric acid can be interpreted in terms of the non-availability of the medium of excretion (water) due to desiccation.
The changes in the ammonia, urea and uric acid levels of active and aestivated pila, produced by the nervous extracts of aestivated and active animals, clearly indicate the presence of one or more humoral agents in the nerve ganglia which can produce in az%o effects. It can be presumed from the results that the nervous extract from the normally active animals is capable of raising the level of ammonia and decreasing that of uric acid in aestivated animals, in r&o. Similarly it appears that an agent with a depressing effect on the ammonia content and an enhancing effect on the uric acid level is present in the ganglionic extract of the aestivated snail.
Thus the results of the injection experiments show that the direction of change, on injection of extracts, in the nitrogen-excretory pattern is almost similar to changes observed in active (normal) and aestivated animals. It can also be said that ammonia and uric acid are more susceptible to the influences of these extracts while the changes observed in urea are not. Therefore, further studies are now in progress (a) to examine the sources of urea on aestivation and on injection of extracts ; (b) to analyse the blood levels of ammonia, urea and uric acid ; and (c) to investigate the kinetics of certain specific enzymes of nitrogen excretion.

NITROGEN EXCRETION DURING AESTIVATION IN PILA GLOBOSA-I 1053
Acknowledgements-The author is immensely indebted to Professor K. Pampapathi Baa, Professor and Head of the Department of Zoology, Bangalore University, Bangalore, India, for his useful suggestions and valuable criticism and for offering facilities to conduct the study. The author is also thankful to M.Sc. Special Physiology students of Bangalore University, Bangalore, India, of the years 1970-l and 1971-2 for their help in the pursuit of this study.
REFERENCES
BALDWIN E. (1935) Problems of nitrogen metabolism in invertebrates-III. Arginase in the invertebrates, with a new method for its determination. Bi0chem.J. 29, 252-262.
BALDWIN E. & NRRDHAM J. (1934) Problems of nitrogen catabolism in invertebrata-1. The snail (Helix pomatia). Biochem. J. 28, 1372-l 392.
DIJERR F. G. (1967) The uric acid content of several species of prosobranch and pulmonate snails as related to nitrogen excretion. Comp. Biochem. Physiol. 22, 333-340.
FLORKIN M. (1966) Nitrogen metabolism. In Physiology of MolZuscs (Edited by WILBER J. & YONGE C.), Vol. II, pp. 309-351. Academic Press, New York.
HAWK P. B., OSER B. L. & SUMMER~ON W. H. (1954) Practical Physiologica Chemistry. McGraw-Hill, New York.
JFZEWSKA M. M., GORZKOWSKI B. & HELLER J. (1963) Nitrogen compounds in snail Helix pomatia excretion. Acta biochem. Polon. 10, 55-65.
NRRDHAM J. (1935) Problems of nitrogen catabolism in invertebrates-II. Correlation between uricotelic metabolism and habitat in the phylum Mollusca. Biochem. J. 29, 238-251.
OSER B. L. (1965) Hawk’s Physiological Chemistry. McGraw-Hill, New York. POTTS W. T. W. (1968) Aspects of excretion in the molluscs. Symp. 2001. Sot. Lond. 22,
187-192. SEXANA B. B. (1955) Physiology of excretion in the common Indian apple snail, Pila globosa
(Swainson). J. Amim. Morph. Physiol. 2, 87-95. R~GHUPATHIRAMIRRDDY S. (1965) The charged properties and metabolism of animals
exposed to changed environments. Doctoral thesis, Sri Venkateswara University, Tirupathi, A.P., India.
R~GHUPATHIRAMIREDDY S. & SWAMI K. S. (1963) Distribution of uric acid in the soft parts of the amphibious snail, Pila. J. Anim. Morph. Physiol. 10, 2, 154-157.
SPRRG K. V. & CAMPBELL J. W. (1968a) Purine biosynthesis and excretion in Otala (Helix) lactea: an evaluation of the nitrogen excretory potential. Comp. Biochem. Physiol. 26, 579-595.
SPEEG K. V. & CAMPBELL J. W. (1968b) Formation and volatilization of ammonia gas by terrestrial snails. Am.J. Physiol. 214, 1392-1402.
SPRRG K. V. & CAMPBELL J. W. (1969) Arginine and urea metabolism in terrestrial snails. Am.J. Physiol. 216, 1003-1012.
Key Word Index--Ammonia; urea; uric acid; mantle; Pila globosa; pattern of nitrogen excretion; neurohormonal control; humoral agent; nervous extract; aestivation; injection of ganglionic extracts.







