
IMMU’tiOLOGY TODAY
hysical exercise has always
been prescribed as a panacea
for illness. Its positive effects
are a result of a complex inter-
action of physiological and psychological
effects, including stress relief, improved
cardiopulmonary function and dietary
changes. However, it is becoming clear that
the immune system responds to increased
physical activity and can be given some of
the credit for exercise-related reduction in
illness and mortality.
As early as 1902, the Boston marathon was
used as an experiment in violent (eccentric)
exercise to demonstrate post-exercise leuko-
cytosis’ and. in more recent years, it has
been shown that elite athletes may become
immune suppressed through overtraining’.
On the other hand, regular recreational ex-
ercise is generally understood to be ben-
eficial to health, whereas total inactivity is
detrimental. So, how much exercise is good
for you and how much is too much? Further-
more, if moderate exercise does boost basal
immune parameters, can it help in situations
of extreme immune requirement, for example
in cancer or following infection with human
immunodeficiency virus (HIV)?
Exercise and immune homeostasis In normal individuals, the numbers and
activity of immune cells increase during a
period of acute exercise, after which they de-
crease to a level below normal for a period of
up to six hours. This is true for lymphocytes,
natural killer (NK) cells and lymphokine-
activated killer (LAK) cells, but not for neu-
trophils, which undergo a sustained neutro-
cytosis (reviewed in Ref. 3). This period of
relative immunosuppression following ex-
ercise has led to the ‘open-window’ theory
of post-exercise susceptibility to infection
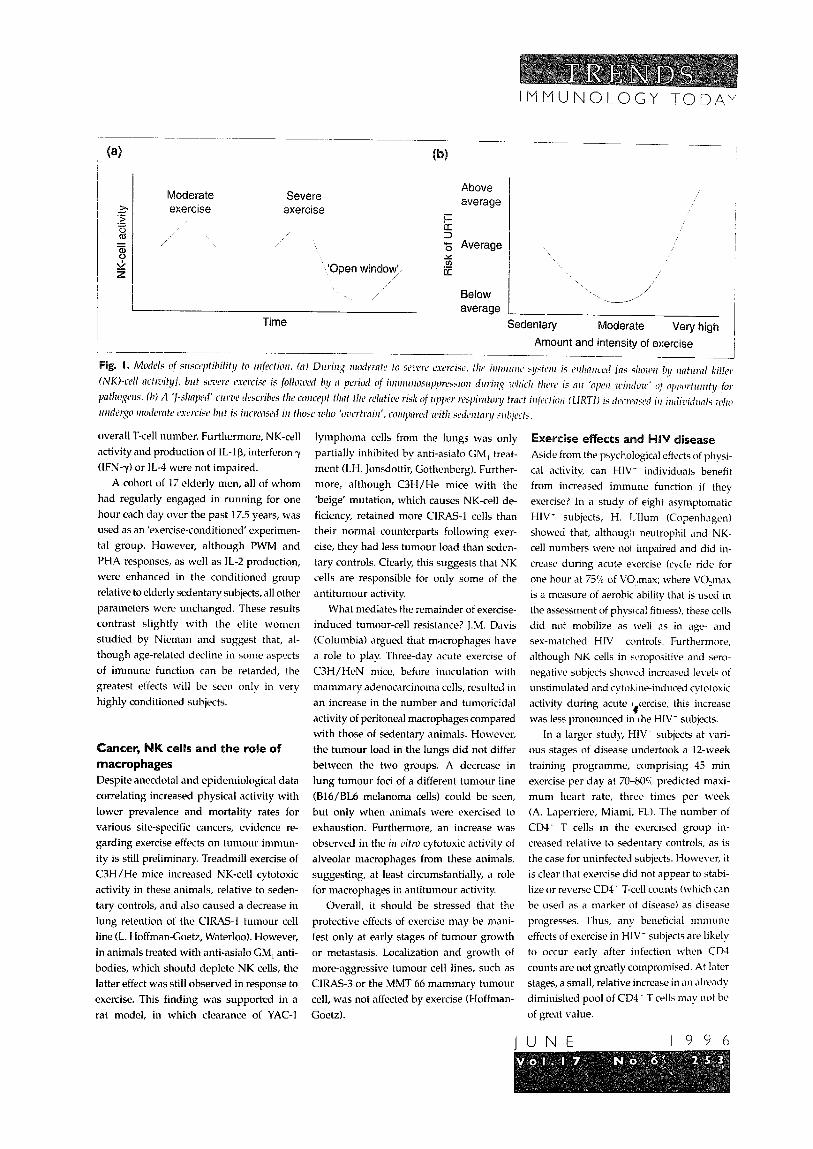
(Fig. la; B.K. Pedersen, Copenhagen). One
explanation for the ‘overtraining effect’ seen
in elite athletes is that the window of oppor-
tunity for pathogens is longer, and the degree
of immunosuppression more pronounced,
following extreme acute or prolonged cxcr-
cise. Certainly, incidence of upper respira-
tory tract infection (URTI) in this group is
greater than in individuals who undergo
regular, moderate exercise (L.T. MacKinnon,
Brisbane; E.M. Peters-Futre, Johannesburg).
Setting a current paradigm, D.C. Nicman
(Boone, NC) proposed a ‘J-shaped’ relation-
ship between regular exercise and immunity
to URTI or other infections. Regular, mod-
erate activity is associated with enhanced
resistance, whereas inactivity or extreme
exercise is linked to decreased resistance
(Fig. lb).
Neutrophils: a last line of defence?
One of the more pronounced features of
physical activity on immune parameters is
the prolonged neutrocytosis following acute
exercise. Since neutrophils comprise nearly
60% of circulating white blood cells and are
an important component of innate immune
responses, what are the implications for
neutrophil function? Overall, human neutro-
phi1 activity as measured by chemilumi-
nescence can be increased by as much as
100% in highly conditioned individuals
(J.A. Smith, Belconnen, Australia; E. Ortega,
Bajadoz, Spain). However, during periods
of exercise, a general pattern is observed in
trained and untrained subjects. Although
not all studies concur, the general view is
that moderate exercise boosts neutrophil
functions, including chcmotaxis, phago-
cytosis and the oxidative burst. During ex-
treme exercise, the picture is quite different:
these functions are chronically suppressed,
with the exception of chemotaxis, which is
not affected.
It is attractive to speculate that, during
the period of immunosuppression of other
immmie components, neutrophils are being
mobilized to compensate for the resulting
deficiencies. The removal of this back-up
system following extreme activity would bc
compatible with the propensity of over-
trained individuals to develop URTI.
Exercise, senescence and immune function The incidence of cancer and respiratory
pathogens both show age-related increases.
Is this a function of immune impairment
and is it avoidable? Nieman studied the
basic mitogenic responses of phytohemag-
glutinin (PHA)-stimulated T cells (these rc-
sponscs are decreased in the aged) in 12
elderly women (average age 73 years) who
were in regular training for long-distance
running events. These ‘elite’ elderly subjects
showed, on average, a T-cell PHA response
that was enhanced by more than 600, as
well as a 64% increase in NK-cell activity
compared with sedentary controls. Sur-
prisingly, however, a 12-week training pro-
gramme of 30 previously sedentary women
resulted in no significant increase in T-cell
or NK-cell function, dcspitc an impressive
13% increase in cardiorespiratory capacity.
A separate analysis in aged men (average
age : $3 years) showed tk?!. in general, they
I-& it .wer circulating numbers ot CD3 and CD8 + r cells compared with younger (average
age 2-1.6 years) men (S. Shinkai, Ehime,
Jnpa~~) T-cell mitogenesis ill Icjponse to
PHA or pokeweed mitogcn (PWM), and in
mixed lymphocyte responses (MLRs), were
also significantly reduced, as was the pro-
duction of interleukin 2 (IL-2). However,
closer analysis suggested that CD45RO
‘memory’ cells and activated CD3’ cells
were elevated as a relative proportion of the
The 2nd Symposium of the International Society of Exercise and Immunology: ‘Practical Applications’ was held at Brussels, Belgium, on 17-18 November 1995.
Moderate Severe exercise exercise
‘Open window/;, /
Time
Above
average
i= 5 5 Average Y 2 eL:
Below average
sedentary Moderate Very high ( Amount and intensity of exercise
I
overall T-cell number. Furthermore, NK-cell
activity and production of IL-lp, interferon y
(IFN-y) or IL-4 were not impaired.
A cohort of 17 elderly men, all of whom
had regularly engaged in running for one
hour each day over the past 17.5 years, was
used as an ‘exercise-conditioned’ experimen-
tal group. However, although PWM and
PHA responses, as well as IL-2 production,
were enhanced in the conditioned group
relative to elderly sedentary subjects, all other
parameters were unchanged. These results
contrast slightly with the elite women
studied by Nieman and suggest that, al-
though age-related decline in some aspects
of immune function can be retarded, the
greatest effects will be seen only in very
highly conditioned subjects.
K cells and the role of
macrophages
Despite anecdotal and epidemiological data
correlating increased physical activity with
lower prevalence and mortality rates for
various site-specific cancers, evidence re-
garding exercise effects on tumour immun-
ity is still preliminary. Treadmill exercise of
C3H/He mice increased NK-cell cytotoxic
activity in these animals, relative to seden-
tary controls, and also caused a decrease in
lung retention of the CIRAS-1 tumour cell
line (L. Hoffman-Goetz, Waterloo). However,
in animals treated with anti-asialo GM, anti-
bodies, which should deplete NK cells, the
latter effect was still observed in response to
exercise. This finding was supported in a
rat model, in which clearance of YAC-1
lymphoma cells from the lungs was only
partially inhibited by anti-asialo GM, treat-
ment (1.H. Jonsdottir, Gothenberg). Further-
more, although C3H/He mice with the
‘beige’ mutation, which causes NK-cell de-
ficiency, retained more CIRAS-1 cells than
their normal counterparts following exer-
cise, they had less tumour load than seden-
tary controls. Clearly, this suggests that NK
cells are responsible for only some of the
antitumour activity.
What mediates the remainder of exercise-
induced tumour-cell resistance? J.M. Davis
(Columbia) argued that macrophages have
a role to play. Three-day acute exercise of
C3H/HeN mice, before inoculation with
mammary adenocarcinoma cells, resulted in
an increase in the number and tumoricidal
activity of peritoneal macrophages compared
with those of sedentary animals. However,
the tumour load in the lungs did not differ
between the two groups. A decrease in
lung tumour foci of a different tumour line
(B16/BL6 melanoma cells) could be seen,
but only when animals were exercised to
exhaustion. Furthermore, an increase was
observed in the irl zrifro cytotoxic activity of
alveolar macrophages from these animals,
suggesting, at least circumstantially, a role
for macrophages in antitumour activity.
Overall, it should be stressed that the
protective effects of exercise may be mani-
fest only at early stages of tumour growth
or metastasis. Localization and growth of
more-aggressive tumour cell lines, Such as
CIRAS-3 or the MMT 66 mammary tumour
cell, was not affected by exercise (Hoffman-
Goetz).
xercise e
Aside from the psychological eftccts of physi-
cal activity, can HIV- individuals benefit
from increased immune function if they
exercise? In a study of eight asymptomatic
HIV- subjects, H. Lllum (Copenhagen)
showed that, although neutrophil and NK-
cell numbers were noi impaired and did in-
crease during acute exercise (cycle ride for
one hour at 75% of VC$max; wrhere VO,max
is a measure of aerobic ability that is used in
the assessment of phvsical fitlless), these cells
did not mobilize as well as in age- and
sex-matched HIV controls. Furthermore,
although NK cells in bcropositive and sero-
negative subjects sholvcd increased levels of
unstimulated and cytoE Ine-induced cytotoxic
activity during acute t -tercise, this increase c
was less pronounced in rhe HIV- subjects.
In a larger study, HIV- subjects at vari-
ous stages of disease undertook a 12-week
training programme, comprising 45 min
exercise per day at 7U-80% predicted maxi-
mum heart rate, three times per week
(A. Laperriere, Miami, FL). The number of
CD4+ T cells in the exercised group in-
creased relative to sedentary controls, as is
the case for uninfected subjects. However, it
is clear that exercise did not appear to stabi-
lize or rel’crse CD4- T-cell counts (which can
be used as a marker of disease) as disease
progresses. Thus, any beneficial immune
effects of exercise in HIV- subjects are likely
to occur early after infection byhen CD-I
counts are not greatly compromised. At later
stages, a small, relative increase in an alread!
diminished pool of CD4 _ T cells mav not be
of great value.
UNE 1996

liiMUN&tiGY TODAY
Overall, HIV* individuals show both im-
paired natural and acquired immunity.
Even though a proportional boost in im-
munity occurs during and after exercise,
any long-term significance of this finding is
a topic of considerable debate. An analysis
of the combined psychological, physical
and immune effects of exercise during HIV
disease is needed to characterize this issue
further.
Concluding remarks Recent work has clearly shown that various
types of exercise may have potentially consid-
erable effects, both positive and negative, on
immune parameters. In order to profit from
the beneficial effects of moderate physical ac-
tivity and to avoid the detrimental effects of
intense exercise, it is important to undersiand
the complex underlying mechanisms. These
certainly involve neuroendocrinological fac-
tors, which are known to have marked effects
on immune cells, and possibly the metabolic
products of muscle exertion.
Studies in the human system have dem-
onstrated that the classical methods of ex-
ercise physiology, in which muscle stress
and cardiorespiratory work-rate act ab a de-
tailed measure of physical capacity, can be
used in the laboratory to generate highly
significant data concerning the interface be-
tween physical activity and immune homeo-
stasis. Furthermore, animal models are now
being developed to provide highly reductive
ifr ivirlo data in support of these fundamental
studies. The critical question is not whether
exercise can influence immune parameters,
but whether these changes can be impressive
clinically.
Rob& Brines is Editor of Immunology Today;
Laurie Hoffman-Goetz (Guetzl.~dcyce~ll.nci.
r7ilz.goz~) is nt the Division of Cnmev Preuentioiz
mrd Cm&o/, Nntionnl Cnrlcer hstifute, Nntioml
Zmtihtes of Health, Bethesda, MD 2089, USA,
otrd nt tk Dept of Hmltk Studies, Llrhcrsify uf
Waterloo, Wnterloo, Chtnrio, Cmndn N2L 3G1;
Bente Klarhd Pedersen is nt fl7e Dcpt of
Illfectiolls Diseases M7722, Riphospitnlet, 20
Tngtwszwj, 2200 Copeulmqerl N, Dcrwwk.
References
1 Larrabee, R.C. (1902) 1. Md. Rm. 7. 76-82 2 Fitzgerald, L. (1988) It~r~r~trr~~~l. %llo!/ 9. 337-339 3 Hoffman-Go&, L. and Pedersen, B.K. (19Y44)
I~ummI. Todq 15, 352-387
urray,
ecrosis is readily de-
tectable in many experi-
mental as well as human
tumors. This is due mainly
to the inability of the vascular supply to
keep up with the nutritional demands of
the rapidly growing tumor (I? Vaupel,
Mainz), as demonstrated by the appearance
of necrosis beyond the diffusion limit of
oxygen. This phenomenon has been ex-
ploited by the development of novel thera-
peutic strategies to treat tumors - collec-
tively known a5 ‘vascular targeting’.
Vascular targeting comprises strategies that
either destroy the pre-existing vasculature
of the tumor or prevent the formation of
new blood vessels, through angiogenesis,
together with the subsequent inhibition
of further tumor growth. In principle,
depriving the tumor parenchyma of nutri-
ents and oxygen will lead to rapid ischemic
necrosis; however, it has become clear that
complete tumor regression is not achieved
50 readily.
The induction of tumor necrosis J. Moore (Manchester) discussed the mecha-
nism of action of photodynamic therapy.
The use of drugs that can be photoactivated
in combination with laser light has shown
great promise in the treatment of canine
prostate cancer, and this therapy is now en-
tering clinical trials. This treatment induces
a rapid hemorrhagic necrosis and its major
target appears to be the endothelium.
However, the effects are more reversible
than tumor necrosis factor (TNF)-induced
necrosis, probably because of the lack of an
immune response.
Many vasoactive chemicals transiently
reduce blood flow in experimental tumors,
without altering the growth of the tumor
(J.C. Murray, Nottingham). Exceptions to
this are flavone-g-acetic acid and 5,6-
dimethylxanthenone acetic acid, which in-
duce a major decrease in blood flow in
experimental tumors, and rapid tumor re-
gression. This activity appears to be retated
to the ability of these agents to induce the
transcription of TNF and interferon y (IFN-y)
in host immune cells. Many conventional
chemotherapeutic agents also induce cyto-
kine production by host cells; in some cases,
this may be due to apoptosis of tumor cells.
Therefore, the initial damaging effect of
*The symposium ‘Tumor Necrosis: Factors and Principles’ was held at Ringberg, Germany, on 29 October-l November 1995.







