
Centro de Ciências Agrárias Depto. de Ciência e Tecnologia de Alimentos
Programa de Mestrado e Doutorado em Ciência de Alimentos
INCIDÊNCIA DE PSE (Pale, Soft,
Exudative) E DFD (Dark, Firm, Dry), AVALIAÇÃO BIOQUÍMICA E ULTRA-
ESTRUTURA DO LOMBO SUÍNO (Longissimus dorsi)
Magali Bernardes Maganhini
Londrina - PR
2007

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

Centro de Ciências Agrárias Depto. de Ciência e Tecnologia de Alimentos
Programa de Mestrado e Doutorado em Ciência de Alimentos
INCIDÊNCIA DE PSE (Pale, Soft, Exudative) E DFD (Dark, Firm, Dry), AVALIAÇÃO
BIOQUÍMICA E ULTRA-ESTRUTURA DO LOMBO SUÍNO (Longissimus dorsi)
Dissertação apresentada ao Programa de Mestrado e Doutorado em Ciência de Alimentos da Universidade Estadual de Londrina, como requisito parcial à obtenção do título de Mestre em Ciência de Alimentos
Mestranda: Magali Bernardes Maganhini
Orientadora: Profa. Dra. Elza Iouko Ida
Co-orientadora: Profª. Drª.Adriana Lourenço Soares
Londrina - PR
2007

DEDICATÓRIA
Aos meus
pais, pelo incentivo, orações e por
tudo para que eu chegasse até aqui.
A minha
irmã, Claudia, pelo apoio e amizade.
Ao meu
esposo Anderson e meu fi lho Caio,
pelo amor, companheirismo e
compreensão.

AGRADECIMENTOS À Profª. Drª Elza Iouko Ida pela sábia e valiosa orientação,
contribuindo para minha formação nos caminhos da ciência e da vida.
À Profª. Drª Adriana Lourenço Soares pela co-orientação e por todos os
momentos de ajuda nunca negados, sugestões e esclarecimentos que
muito enriqueceram este trabalho e minha formação.
Ao Prof. Dr. Massami Shimokomaki pela inestimável colaboração,
sugestões e pelo amor à ciência, mesmo nas horas mais difíceis.
À CAPES/MEC (Coordenação de Aperfeiçoamento de Pessoal de Nível
Superior/Ministério da Educação) pela bolsa concedida.
Ao Prof. Dr. Makoto Matushita do Departamento de Química da
Universidade Estadual de Maringá, que viabil izou a realização da
análise de perfi l de ácidos graxos.
Ao Prof. Dr. Francisco Javier Hernandez Blazquez da Faculdade de
Medicina Veterinária e Zootecnia da Universidade de São Paulo, que
viabil izou a realização da microscopia ótica e eletrônica.
À Seara Alimentos, na pessoa de Paulo Donizett i Guarnieri, pelas
condições oferecidas e amostras cedidas para a realização deste
trabalho.
Ao Programa de Mestrado e Doutorado em Ciência de Alimentos desta
Instituição, pelas condições oferecidas para a realização deste
trabalho.
Aos Docentes do Programa de Mestrado e Doutorado em Ciência de
Alimentos desta Instituição, pelos ensinamentos e atenção dispensada.

v
Aos funcionários do Departamento de Ciência e Tecnologia de
Alimentos desta Instituição, que auxil iaram em todos os momentos nos
laboratórios para execução da parte experimental.
Ao apoio técnico Eduardo Toledo pelo auxíl io e amizade.
Aos estagiários Bruno Mariano, Taísa Saragosa e Valdecir, pela
colaboração no desenvolvimento do trabalho.
Aos colegas do Grupo de Carnes, em especial ao Denis e Cassiana
pela ajuda e amizade.
Aos amigos do Mestrado, pelos momentos que passamos juntos e pela
amizade.
À Deus, pela força em todos os momentos e por tudo que tem realizado
em minha vida.

MAGANHINI, Magali Bernardes. INCIDÊNCIA DE PSE (Pale, Soft, Exsudative) E DFD (Dark, Firm, Dry), AVALIAÇÃO BIOQUÍMICA E ULTRA-ESTRUTURA DO LOMBO SUÍNO (Longissimus dorsi) 93p. 2007. Dissertação de Mestrado em Ciência de Alimentos–Universidade Estadual de Londrina. Londrina-PR
RESUMO O direcionamento da produção de suínos para um aumento na porcentagem de carne magra na carcaça, tem levado à modificações substanciais tanto na composição centesimal como nas características bioquímicas do músculo, como o PSE (Pale, Soft e Exudative), o que signif ica Pálida, Macia e Esxudativa, e DFD (Dark, Firm e Dry) Escura, Firme e Seca. Estas carnes apresentam propriedades funcionais comprometidas, sendo um dos maiores problemas enfrentados pela indústria processadora devido a perdas econômicas. O objetivo deste trabalho foi inves t igar a inc idênc ia de PSE e DFD no lombo su íno (Long iss imus dors i ) , em uma l inha comerc ia l de aba te e ava l ia r as ca rac te r ís t i cas b ioqu ímicas e da u l t ra -es t ru tu ra dos lombos PSE e DFD. Em um frigoríf ico 946 lombos foram analisados quanto ao pH e cor e classif icados como PSE, DFD e Normal. Os lombos PSE, DFD e Normal foram analisados com relação ao valor de R, perda de peso por cozimento (PPC), força de cisalhamento (FC), índice de fragmentação miofibri lar (IFM), l ipídios totais, oxidação l ipídica, perfi l de ácidos graxos e microscopia eletrônica. A incidência de lombos PSE, DFD e Normal foi de 22,83%, 1,06%, 76,11%, respectivamente. Os valores de pH e cor dos lombos suínos PSE, DFD e Normal diferiram signif icativamente (p≤0,05) entre si. O valor de R foi signif icativamente maior (p≤0,05) para os lombos PSE do que DFD. A FC e a PPC não diferiu signif icativamente entre os lombos PSE, DFD e Normal. O IFM foi signif icativamente maior (p≤0,05) para o lombo DFD. A oxidação l ipídica foi 42% maior para o lombo PSE em relação ao normal e DFD. O perfi l de ácidos graxos dos lombos suínos PSE, DFD e Normal não apresentou diferença signif icativa entre si. Finalmente os resultados histológicos apresentaram vários eventos bioquímicos incluindo a contração do sarcômero nos lombos PSE. Ao mesmo tempo foi observada uma seqüência aparente das atividades das proteases endógenas musculares inicialmente pela despolimerização das proteínas miofibri lares próximas e no disco Z e conseqüentemente a fragmentação destes discos e por últ imo o colapso do sarcômero e de algumas estruturas miofibrilares.
Palavras-chave: cor, pH, manejo pré-abate, microscopia eletrônica.

v i i
MAGANHINI, Magali Bernardes . PORK Longissimus dorsi PSE (Pale, Soft, Esudative) AND DFD (Dark, Firm, Dry) OCCURRENCE AND BIOCHEMICAL AND ULTRASTRUCTURAL EVALUATION. 93p. 2007. Master Dissertation in Food Science – Universidade Estadual de Londrina. Londrina-PR
ABSTRACT The aim of pork production in order to have an increase in non fat meat in the carcass has brought about changes not only on proximate chemical composit ion but also within the muscle biochemical characteristics as PSE (Pale, Soft and Exudative) meaning pale, soft and exudative meat and DFD (Dark, Firm and Dry) meaning dark, f irm and dry meat. Both meat types present functional properties impaired and this phenomena represent an actual economical problems to the meat industries. The objective of this work was to investigate the occurrence of pork L. dorsi PSE- and DFD- meat at a commercial plant and to evaluate their biochemical and ultra structural characteristics. Longissimus dorsi samples in number of 946, were analyzed in relation to pH and color thus classif ied as PSE-, DFD- and Normal- meat. They were also analyzed in relation to R value, cooking loss (CL), tenderness measured through shear force (SF) value, miofibri l lar fragmentation index (MFI), total l ipid concentration, l ipid oxidation, fatty acids profi le and ultra structural observation. The occurrence of PSE-, DFD- and normal meat was 22.83, 1.06, 76.11%, respectively . pH and color values differed signif icantly (p≤0.05). R value was signif icantly higher for PSE- in relation to DFD- meat (p≤0.05). Tenderness and CL did not differ signif icantly while MFI was signif icantly higher for DFD meat although this value was not suff icient to show difference in SF. Lipid oxidation was 42.0% higher in PSE- in relation to both normal and DFD- meat samples. Fatty acids profi le did not show signif icantly differences in every samples evaluated. Lastly, histological results showed several biochemical events including sarcomere contraction in PSE meat samples. Also it was noticeable an apparent sequence of muscle endogenous proteases activit ies, f irst ly at the depolimerized myofibri l proteins near and at the Z-disks consequently the fragmentation of the Z-disks and ult imately the collapse of the sarcomere and to some of the myofibri l structures. Keywords: color, pH, pre-slaughtering management, electronic microscopy

v i i i
SUMÁRIO
Página 1 INTRODUÇÃO..... . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 15 2 OBJETIVOS .... . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19 2.1 Objetivos específicos . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19 3 REVISÃO BIBLIOGRÁFICA.... . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 21 3.1 PSE .... . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 21 3.2 DFD . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 31 3.3 Manejo Pré-abate . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 33 3.3.1 Transporte . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 35 3.3.2 Jejum . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 36 3.4 Composição de Ácidos Graxos e Oxidação Lipídica . . . . . . . . . . . . . . . . 38 3.5 Valor de R . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 41 3.6 Ultra-estrutura da Carne . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 43 4 MATERIAL E MÉTODOS ..... . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 46 4.1 Matéria Prima e Classificação dos Lombos em PSE, DFD e Normal . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 46 4.2 Caracterização Bioquímica dos Lombos PSE, DFD e Normal . . . 46 4.2.1 Medida de pH24h . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 47 4.2.2 Medida de Cor . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 47 4.2.3 Classificação das Amostras em PSE e DFD . . . . . . . . . . . . . . . . . . . . . . . . . 48 4.2.4 Medida do Valor de R . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 48 4.2.5 Medida da Perda de Peso por Cozimento (PPC) . . . . . . . . . . . . . . . . . . . 49 4.2.6 Medida da Maciez . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 49 4.2.7 Medida do Índice de Fragmentação Miofibrilar (IFM) . . . . . . . . . . . 50 4.2.7.1 Extração .... . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 50 4.2.7.2 Determinação de Proteína Solúvel ... . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 51 4.2.7.3 Medida do IFM .... . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 52 4.2.8 Determinação de Lipídios Totais . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 52 4.2.9 Medida da Oxidação Lipídica . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 53 4.2.10 Determinação da Composição de Ácidos Graxos . . . . . . . . . . . . . . . 54 4.2.11 Microscopia Eletrônica dos Lombos Suínos . . . . . . . . . . . . . . . . . . . . . . . 55 4.2.12 Análise Estatística . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 57 5 RESULTADOS E DISCUSSÃO ..... . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 58 5.1 INCIDÊNCIA DE PSE E DFD NA CARNE SUÍNA ..... . . . . . . . . . . . . . . . . . . . . 58 5.2 Caracterização Bioquímica e Avaliação da Ultraestrutura dos Lombos PSE, DFD e Normal . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 64 5.2.1 Valores de pH24h dos Lombos Suínos . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 64 5.2.2 Cor dos Lombos Suínos . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 66 5.2.3 Avaliação do Valor de R dos Lombos Suínos . . . . . . . . . . . . . . . . . . . . . . 69 5.2.4 Avaliação da Perda de Peso por Cozimento (PPC) dos Lombos Suínos . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 70 5.2.5 Maciez dos Lombos Suínos . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 72 5.2.6 Índice de Fragmentação Miofibrilar (IFM) dos Lombos Suínos . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 73 5.2.7 Lipídios Totais dos Lombos Suínos . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 74 5.2.8 Oxidação Lipídica dos Lombos Suínos . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 75 5.2.9 Composição de Ácidos Graxos dos Lombos Suínos . . . . . . . . . . . . 77

ix
6 CONCLUSÕES ..... . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 84 7 REFERÊNCIAS BIBLIOGRÁFICAS .... . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 85

LISTA DE QUADROS
Página QUADRO 1 – Classif icação de diferentes raças suínas quanto à
susceptibi l idade ao estresse e predisposição à produção de carne PSE * ... . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 27
QUADRO 2 – Propriedades e características das carnes PSE* ..... . . . . . 30
QUADRO 3 – Propriedades e características das carnes DFD* ..... . . . . . 32

LISTA DE FIGURAS
Página FIGURA 1 - Mecanismo proposto para indução da síndrome PSS
em suínos (Adaptado de MAcLENNAN e PHILLIPS, 1992). .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 24
FIGURA 2 - Parte do eixo da seqüência de aminoácidos no DNA, na qual Arg615 sofreu uma mutação para cisteína (localizado na caixa) e a seqüência normal GCGCTC foi alterada para GTGCTC (FUJII et al., 1991). ... . . . . . . . . . . . 25
FIGURA 3 – Provável relação entre desenvolvimento da oxidação l ipídica e carnes PSE (SOARES et al. , 2005). ... . . . . . . . . . . . . 40
FIGURA 4 – Situação de stress e formação de IMP ..... . . . . . . . . . . . . . . . . . . . . . 42
FIGURA 5 – Ultra-estrutura muscular (JUDGE et al., 1989) ... . . . . . . . . . . . . 44
FIGURA 6 - Estrutura do sarcômero e suas proteínas (SHIMOKOMAKI et al. , 2006) ... . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 44
FIGURA 7 - Histograma de distribuição do valor de L*24h de lombos suínos (n=946) ... . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 59
FIGURA 8 - Histograma de distribuição do valor de pH24h de lombos suínos (n=946) ... . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 59
FIGURA 9 – Incidência de PSE e DFD em lombos suínos (n=946) .... . 61
FIGURA 10 - Lombos suínos classif icados como PSE, DFD e Normal com base no valor de L*24h ... . . . . . . . . . . . . . . . . . . . . . . . . . . . . 61
FIGURA 11 - Correlação entre os valores de L*24h e pH24h de lombos suínos (n=946) (R= -0,46, p ≤ 0,05) ... . . . . . . . . . . . . . . . 63
FIGURA 12 - Correlação entre os valores de b* e L*24h de lombos suínos (n=946) (R= 0,43, p ≤ 0,05) ... . . . . . . . . . . . . . . . . . . . . . . . . . . . . 64
FIGURA 13 - Correlação entre os valores de L*24h e pH24h de lombos suínos (n=36) (R= -0,75, p ≤ 0,05) ... . . . . . . . . . . . . . . . . . 69
FIGURA 14 – Eletromicrografia de uma seção longitudinal do lombo suíno Normal. Aumento 12000 vezes. M = Linha M; Z = disco Z; A = Banda A; I = Banda I. .. . . . . . . . . . . 81
FIGURA 15 – Eletromicrografia de uma seção longitudinal do lombo suíno PSE. Aumento 6000 vezes. ... . . . . . . . . . . . . . . . . . . . . 82
FIGURA 16 - Provável mecanismo para a relação entre estresse pré-abate, alterações bioquímicas e na ultra-estrutura no desenvolvimento da carne suína PSE. 1-

x i i
Despolimerização das proteínas do sarcômero. 2-Atividade das proteases na região do disco Z. 3-Colapso do sarcômero. ... . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 83

LISTA DE TABELAS
Página TABELA 1 - Valores de pH, L* , a* e b* 24 h post mortem dos
lombos suínos PSE, DFD e Normal... . . . . . . . . . . . . . . . . . . . . . . . . . . . . 62
TABELA 2 - Valores de pH24h post mortem dos lombos suínos PSE, DFD e Normal ... . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 65
TABELA 3 - Valores de L* , a*, b* e razão a*/b* 24h post mortem para os lombos suínos PSE, DFD e Normal ... . . . . . . . . . . . . . . . . 68
TABELA 4 – Valor de R dos lombos suínos PSE, e DFD e Normal .... . . 70
TABELA 5 – Perda de peso por cozimento (PPC) dos lombos suínos PSE, DFD e Normal. .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 71
TABELA 6 – Força de cisalhamento (FC) em Newton(N) dos lombos suínos PSE, DFD e Normal in natura e cozido. ... 73
TABELA 7 - Índice de Fragmentação Miofibrilar (IFM) dos lombos suínos PSE, DFD e Normal ... . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 74
TABELA 8 - Lipídios totais dos lombos suínos PSE, DFD e Normal .... 75
TABELA 9 - Oxidação l ipídica dos lombos suínos PSE, DFD e Normal ... . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 76
TABELA 10 – Teor de ácidos graxos dos lombos suínos PSE, DFD e Normal ... . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 78
TABELA 11– Teor de ácidos graxos monoinsaturados, saturados, polinsaturados, ácidos graxos ômega 6 e 3 dos lombos suínos PSE, DFD e Normal... . . . . . . . . . . . . . . . . . . . . . . . . . . . . 79

LISTA DE ABREVIATURAS a* Componente vermelho-verde do Sistema de Cor CIELAB
ABCS Associação Brasileira dos Criadores de Suínos
ABIPECS Associação Brasileira das Indústrias Produtoras e
Exportadoras de Carne Suína
ADP Adenosina Difosfato
ATP Adenosina Trifosfato
b* Componente amarelo-azul do Sistema de Cor CIELAB
CRA Capacidade de Retenção de Água
DFD Escura, Firme e Seca
FC Força de Cisalhamento
HM Hipertermia Maligna
IFM Índice de Fragmentação Miofibri lar
IMP Inosina Monofosfato
L* Luminosidade do Sistema de Cor CIELAB
N Newton
PLA2 Enzima fosfolipase A2
PPC Perda de Peso por Cozimento
PSE Pálida, Macia e Esxudativa
PSS Síndrome do Estresse Suíno
RS Retículo Sarcoplasmático
RYR1 Proteína Rianodina 1
TBA Ácido 2-tiobarbitúrico
TEP 1, 1, 3, 3- tetraetoxipropano

1 INTRODUÇÃO
A carne suína é hoje a forma de proteína animal mais consumida
no mundo e no ano de 1979 ultrapassou a preferência dos
consumidores pela carne bovina (ABCS, 2005).
Em 2006, a produção mundial de carne suína foi de 104,990
milhões de toneladas com crescimento de 2,2%. China, União Européia
e Estados Unidos foram responsáveis por cerca de 81,3% da produção
mundial (ABIPECS, 2007). A produção nacional de carne suína
apresentou índices consideráveis de crescimento nos últ imos 24 anos
com aproximadamente 3,8% ao ano (ABCS, 2005). O Brasil continua na
posição de 4° maior produtor mundial, com 2,825 milhões de toneladas
em 2006. Deste total, 81% foram consumidos no mercado interno, 30%
in natura e 70% industrial izada e os 19% restantes foram destinados
para o mercado internacional (ABIPECS, 2007).
A participação do Brasil no mercado nacional e internacional de
carne suína, e as exigências do consumidor aumentaram. Assim maior
atenção foi dada a carne suína nos diversos segmentos da cadeia
produtiva. Com os avanços tecnológicos nas áreas de genética e
nutrição, o suinocultor passou a desenvolver um animal com menos
gordura, mais carne e com maior eficiência na conversão dos
alimentos. Como resultado deste investimento, surgiu no mercado um
animal mais competit ivo.
Contudo, essas transformações implicaram nas características
físicas e no metabolismo do músculo que influenciaram os aspectos
visuais e sensoriais da carne e a aceitabil idade do produto f inal por

16
parte dos consumidores. Estas característ icas foram determinadas por
um conjunto de fatores que incluíram as condições ambientais
relacionadas ao clima e instalações, técnicas de manejo, nutrição e
genética.
As carnes PSE (Pale, Soft e Exudative) que signif ica Pálida,
Macia e Esxudativa e carnes DFD (Dark, Firm, Dry) Escura, Firme e
Seca, são resultantes de condições anormais das transformações
bioquímicas do músculo com comprometimento de sua qualidade.
Segundo Angerami (2004), as carnes com características de
qualidade comprometidas, resultaram em perdas econômicas que
inviabil izaram as atividades de um frigoríf ico, devido a:
- redução no rendimento de abate e do produto processado;
- desclassif icação de carcaças que l imitam a uti l ização da carne
para a industrial ização;
- alteração na qualidade da carne e de produtos industrial izados
que ocasionaram: comprometimento da cor e da fatiabil idade
dos produtos cárneos cozidos, alterações na formação de cor
em produtos cárneos curados, descoloração e formação de
odores estranhos e redução na vida úti l do produto.
A condição PSE em suínos foi reconhecida e documentada desde
1960, e a condição PSS (Porcine Stress Syndrome) foi reconhecida na
década de 60 (CASSENS, 2000). A PSS foi associada com a PSE e
considerada a causa fundamental do surgimento destas carnes em
suínos. A manifestação da hipertermia maligna (HM), associada a
distúrbios da musculatura esquelética ocorreu quando os animais foram

17
expostos a situações estressantes e a determinados agentes
anestésicos, como o halotano que caracterizou assim, a síndrome do
estresse suíno - PSS (Porcine Stress Syndrome). A relação do PSE
com o gene halotano ou gene rianodina (ryr1), permit iu o processo de
seleção de l inhagens suínas para evitar o desenvolvimento do PSE,
sendo os animais sensíveis ao estresse, os recessivos (nn) e os
heterozigotos (Nn) (FUJII et al., 1991).
A incidência de carne PSE produzida nos EUA, foi de 18% em
1963 e 16% em 1992 (CASSENS, 2000). A indús t r ia f r i go r í f i ca Nor te
Amer icana , t raba lhou com grandes p roporções de carnes PSE
(KAUFFMAN e t a l . , 1992) . Cannon e t a l . (1996) es t imaram que
es tas anomal ias represen ta ram perdas de aprox imadamente 100
mi lhões de dó la res por ano para a economia Nor te Amer icana . Na
Aus t rá l ia , es tas perdas a t ing i ram aprox imadamente U$ 20 mi lhões
por ano .
Fo i re la tado por Murray e Johnson (1998) que cada chuleta de
carne suína PSE perdeu aproximadamente 3 % de água em relação à
carne normal em uma embalagem no varejo, e estimaram que a
condição de carne PSE diminuiu o valor de um suíno em 5 dólares
Canadenses. No Bras i l uma produção de 2 ,7 m i l tone ladas de
ca rne su ína com ocor rênc ia de 15% de an ima is com PSE, fo i
es t imado uma perda de 1 ,5% de água por ca rcaça com um
pre ju ízo de U$ 4 ,5 m i lhões por ano (ODA e t a l . , 2003) .

18
Assim, atenção especial por parte da indústria em monitorar a
qualidade do manejo dos suínos desde a genética até o produto f inal,
são fatores decisivos para garantir a qualidade da carne.
Este trabalho visa investigar a incidência de PSE e DFD no lombo
suíno (Longissimus dorsi), em uma l inha comercial de abate e avaliar
as suas características bioquímicas e da ultraestrutura. No Brasil,
estudos sobre a ocorrência dos fenômenos PSE e DFD em suínos, e a
avaliação da situação da qualidade das carnes e as características
bioquímicas serão úteis para propor procedimentos para melhorá-las e
contribuir para o avanço de conhecimento no desenvolvimento
científ ico e tecnológico das indústrias processadoras de carne.

19
2 OBJETIVOS
Inves t iga r a inc idênc ia de PSE e DFD dos lombos su ínos
(Long iss imus dors i ) em uma l inha comerc ia l de aba te , e ava l ia r as
suas carac te r ís t icas b ioqu ímicas e da u l t ra -es t ru tu ra .
2.1 Objetivos específicos
Es t imar a inc idênc ia de PSE e DFD dos lombos su ínos
(Long iss imus dors i ) em uma l inha comerc ia l de aba te .
Cor re lac ionar os va lo res de pH e co r dos lombos PSE, DFD e
Norma l de su ínos .
Ava l ia r a re lação en t re IMP e ATP (Va lo r de R) dos lombos
su ínos PSE, DFD e Norma l .
Ana l i sa r a mac iez dos lombos su ínos PSE, DFD e Norma l por
me io de med idas de fo rça de c isa lhamento e índ ice de
f ragmentação mio f ib r i l a r .
Es t imar a ox idação l i p íd ica e per f i l de ác idos g raxos dos
lombos su ínos PSE, DFD e Norma l .

20
Avaliar a ultra-estrutura dos lombos suínos (músculo Longissimus
dorsi) PSE, DFD e Normal por meio de microscopia eletrônica.

3 REVISÃO BIBLIOGRÁFICA
3.1 PSE
Uma considerável variação na qualidade da carne suína foi
verif icada nos fr igoríf icos brasileiros, europeus e americanos (CULAU
et al . 1994). As carnes destinadas à industrial ização apresentaram
diversos problemas e foram destacadas as carnes PSE e DFD (TERRA,
1998).
As siglas PSE e DFD referem-se a duas anomalias das
características da carne suína. A abreviatura PSE signif ica Pale, Soft e
Exudative , isto é, carne pálida, macia e esxudativa, devido a perda de
água com facil idade, enquanto que DFD signif ica Dark, Firm e Dry, ou
seja, carne escura, f irme e seca na superfície, devido a retenção de
toda umidade no interior das f ibras musculares (FELÍCIO, 1986).
A carne suína PSE representa o principal problema de qualidade
na indústria, devido à baixa capacidade de retenção de água, textura
f lácida, cor pálida e elevada perda de água no processamento. A carne
PSE foi descrita como indesejável, tanto para os consumidores, como
para a indústria de processamento (BOWKER et al., 1999).
A principal causa do desenvolvimento da condição PSE foi a
decomposição acelerada do glicogênio após o abate, que ocasionou um
valor de pH muscular muito baixo, e geralmente inferior a 5,8. E devido
à temperatura do músculo estar próximo do estado fisiológico (> 38°C),
o decréscimo do pH foi seguido por um processo de desnaturação

22
protéica, que favoreceu o estabelecimento da condição PSE (HONIKEL
& KIM, 1986).
Além dos fatores genéticos, Andrade et al. (1992) descreveram
que havia outros que influenciaram a incidência de carne PSE, tais
como: tempo de transporte dos animais da granja para o fr igoríf ico,
manejo uti l izado no pré abate do frigoríf ico e tempo de descanso dos
animais antes do abate.
As características destas carnes foram provenientes da
manifestação da síndrome PSS (Porcine Stress Syndrome) (CHEAH et
al. 1984) ou HM (Hipertermia maligna) (FUJII et al. 1991). A síndrome
PSS foi desencadeada por fatores de estresse ambientais ou
fisiológicos, como mudança na temperatura ambiente, excitação,
transporte, exercícios, que poderiam levar a morte inesperada dos
animais (CHEAH e CHEAH, 1981b; MITCHELL e HEFFRON, 1982;
CHEAH et al. , 1984). A HM foi descrita como uma miopatia hereditária
e poderia ser desencadeada por indução com halotano (2-bromo-2-
cloro-1, 1,1-tr if luoretano), um anestésico gasoso, ou com relaxantes
musculares como succinilcolina (MITCHELL e HEFRON, 1982; FUJII et
al. ,1991). Os sintomas foram manifestados pela rigidez muscular
generalizada, rápido aumento da temperatura corporal, taquicardia,
hiperventi lação, acidose metabólica severa e elevados níveis de
metabólitos no soro (CHEAH,1981 a ; CHEAH et al.,1984).
O retículo sarcoplasmático (RS) do músculo esquelético de suínos
estoca e l ibera íons cálcio (MITCHELL, 2002), os quais têm importante
papel nas reações bioquímicas celulares e estão associados com a

23
regulação da contração muscular. Portanto, é provável que o aumento
na l iberação dos íons cálcio do retículo sarcoplasmático em animais
com HM, seja o responsável pela série de reações bioquímicas que
conduzem a perda de controle da glicólise com aumento da produção
de ácido lático (CHEAH e CHEAH, 1981b; MITCHELL, 2002). Mac
Lennan e Phil l ips (1992) propuseram o mecanismo de indução da
síndrome em suínos PSS, causado por uma anomalia nos canais
l iberadores de Ca²+ do retículo sarcoplasmático, conforme apresentado
na Figura 1. Este mecanismo propõe que a indução da síndrome PSS
em suínos foi causada por anomalias nos canais l iberadores de Ca²+
localizados no retículo sarcoplasmático (RS). A contração muscular,
gl icólise e função mitocondrial foram reguladas pelas concentrações de
Ca²+ no citoplasma. Em um ciclo normal de contração-relaxamento
(Figura 1A), o Ca²+ foi bombeado para dentro do RS por uma ATPase
para iniciar o relaxamento. Na próxima contração, o Ca²+ foi l iberado
de forma controlada pelos canais do RS. Os metabolismos glicolít ico e
aeróbio foram realizados de forma balanceada, para produzir energia
para a célula. Neste caso, a l iberação de Ca²+ foi regulada pelo ATP,
Mg²+ e pela sua própria concentração. Na condição anormal de PSS
(Figura 1B), os canais l iberadores de Ca²+ não foram sensíveis a
baixas concentrações de Ca²+, mantendo-se abertos e aumentando a
sua concentração no citoplasma. A contração muscular foi mantida,
gerando a rigidez muscular. O metabolismo glicolít ico e aeróbio foi
aumentado, gerando ácido láctico, CO2 e calor, aumentando a tomada

24
de oxigênio. Assim ocorreu injúria nas membranas celulares e
problemas sistêmicos típicos de PSS.
Calor
Ciclo Normal de Contração Muscul ar Ciclo de Contração Muscular HM
Mitocôndria Mitocôndria
Glicogênio Glicogênio
Ácido lá ticoÁcido lático
Fibra muscularFibra muscularActina
ActinaMiosina Miosina
1A 1B FIGURA 1 - Mecanismo proposto para indução da síndrome PSS em
suínos (Adaptado de MAcLENNAN e PHILLIPS, 1992).
Fuji i et al. (1991) descreveram a ocorrência de uma mutação no
gene que codif icou para o receptor rianodina do músculo esquelético,
correlacionada com a HM. Os autores desenvolveram a metodologia em
análise genética, que permitiu diferenciar três genótipos: Hal NN
(normal dominante), HAL Nn (heterozigoto) e Hal nn (sensível
recessivo) e com esta técnica foi possível investigar freqüência do
gene halotano, nas diferentes raças de suínos, e propuseram melhores
estratégias para melhorar o cruzamento.

25
A proteína rianodina tem peso molecular de 350kDa e controla a
l iberação de Ca²+ pelo retículo sarcoplasmático e recebeu a
denominação de rianodina devido à capacidade de l igação a um
alcalóide derivado de planta de mesmo nome (GIANNINI et al ., 1995;
MICKELSON e LOUIS, 1996). O cromossomo 6, responsável pela
codif icação do gene RYR1, que sofreu mutação de uma base
nitrogenada citosina para t imina na posição 1843 da seqüência de
DNA, resultou na alteração do aminoácido 615(Figura 2), onde um
resíduo de arginina foi substituído por um resíduo de cisteína (FUJII et
al. , 1991).
FIGURA 2 - Parte do eixo da seqüência de aminoácidos no DNA, na qual Arg615 sofreu uma mutação para cisteína (localizado na caixa) e a seqüência normal GCGCTC foi alterada para GTGCTC (FUJII et al., 1991).
Esta mutação no gene RYR1 está diretamente relacionada com a
alteração no mecanismo de l iberação de Ca²+ do retículo
sarcoplasmático, embora a natureza e expressão deste defeito ainda
sejam controversias (MICKELSON e LOUIS, 1996). Fuji i et al. (1991)
relataram que Arg 615 está envolvida na l igação de reguladores do
canal de Ca²+, ocasionando a sua abertura. Uma vez aberto, o canal
poderá não responder a Ca²+ e Mg²+, que induzem ao seu fechamento,
conduzindo assim a contratura muscular, hipermetabolismo e

26
hipertermia. Da mesma maneira Mickelson e Louis (1996) descreveram
que em animais sensíveis a HM o aumento das taxas l iberação de Ca²+
do retículo sarcoplasmático e a maior afinidade do canal de Ca²+ para
rianodina, poderiam ser devido a maior permanência do canal no
estado aberto.
O gene halotano, além de determinar a maior predisposição ao
estresse em suínos, também foi responsável pela produção de carcaças
com maior deposição de carne magra, porém relacionado com a
produção de carne PSE (CULAU et al. , 2002).
Embora não seja a única causa de PSE em suínos, o fator
genético provavelmente foi o mais importante. Linhas genéticas de
animais que foram melhorados para o ganho de peso e produção de
carne magra apresentaram ocorrência elevada para a síndrome devido
ao metabolismo energético insuficiente. Entretanto, as empresas de
melhoramento genético reconheceram este problema e procuram
revertê-lo, el iminando o gene halotano, responsável pela PSS (ODA et
al., 2003). O trabalho de melhoria genética visando à qualidade da
carne obrigou a empresa Sadia a identif icar o status (genótipo) para o
gene Hal (halotano), em todas as l inhas, por meio da tecnologia do
teste de DNA. A busca desta qualidade almejada exigiu decisões como:
as l inhas de fêmea obrigatoriamente devem ser l ivres do alelo Halnn.
Isto garantiu animais de abate que no máximo são heterozigotos
(HalNn) e outra parte completamente l ivre do gene Hal. Estas foram as
principais medidas de ordem genética para minimizar a ocorrência da
síndrome PSS nos animais vivos e da carne PSE nas carcaças.

27
Na década de 60, foi reportado que certas raças (Pietrain, Poland
China e Landrace Belga) continham uma grande proporção de animais
propensos a apresentar carne PSE, enquanto que outras raças eram
praticamente l ivres desta anomalia (SELLIER, 1998). Algumas
diferentes raças de suínos foram classif icadas (Quadro 1) conforme sua
susceptibi l idade ao estresse (GREGORY, 1998). Alguns países, como a
Dinamarca, Suíça e Suécia já eliminaram a presença do gene halotano
de suas l inhas selecionadas. Entretanto, somente no f inal da década de
90 algumas empresas internacionais de genética decidiram remover o
gene halotano de suas l inhas de seleção (ROSENVOLD et al. , 2003).
QUADRO 1 – Classif icação de diferentes raças suínas quanto à
susceptibi l idade ao estresse e predisposição à produção
de carne PSE *
Sensível ao estresse
Sensibilidade media ao estresse
Resistente ao estresse
Pietrain Landrace Belga China Polonês Landrace Alemão
Landrace Holandês Landrace Francês Landrace Sueco Landrace Suíço Landrace Dinamarquês Landrace Norueguês Landrace Australiano Landrace Irlandês Yorkshire Holandês Hampshire Americano
Large White Ir landês Large White Australiano Large White Frances Large White Ingles Yorkshire Americano Duroc
*Fonte: GREGORY, 1998
Pommier e Houde (1993) determinaram que 30% das carnes
classif icadas como PSE, com base na cor, foram provenientes de

28
suínos heterozigotos (Nn). Da mesma maneira, os resultados de Cheah
et al. (1995) relataram que os suínos heterozigotos (Nn) apresentaram
propensão para produzir maior proporção de carcaças PSE do que
normais. Culau et al . (1994) identif icaram freqüência de 30,69% de
carcaças PSE no Estado do Rio Grande do Sul, por leitura do pH inicial
no músculo Longissimus dorsi, em 8842 carcaças suínas, entretanto a
identif icação da causa desta freqüência alta de carcaças PSE, não foi
definida, visto que há influência de muitos fatores ambientais e
genéticos.
O teste do halotano em suínos foi uti l izado para detectar a
susceptibi l idade dos animais em desenvolver carnes PSE (CHEAH et
a l . , 1993). Este teste foi baseado na habil idade do suíno de recuperar
ao anestésico halotano. Aqueles animais que se recuperaram
rapidamente foram mais resistentes (NN), enquanto que aqueles que
metabolizaram o halotano lentamente e requereram mais tempo para
recuperar a consciência foram susceptíveis ao estresse ou susceptíveis
ao PSE (nn) (PEARSON e YOUNG, 1989), entretanto, este teste foi
ineficiente para detectar animais heterozigotos (Nn).
Mesmo que existiu correlação signif icativa entre animais
portadores do gene halotano e produção de carne PSE (SILVEIRA,
1996), o genótipo por si só não explicou a ocorrência de carne PSE
(WARRIS, 1995).
A inabil idade da indústria para medir e classif icar objetivamente a
qualidade da carne, foi associada a um aumento da baixa qualidade na
carne suína (KOHLER e FREITAS, 2005). Considerando que as

29
alterações musculares podem se manifestar dentro da primeira hora,
vários parâmetros foram avaliados aos 45 min após o abate para
predizer sobre a qualidade final da carne (WARRISS e BROWN, 1987).
As variáveis que melhor definiram a qualidade da carne suína
foram: pH, cor e algumas propriedades funcionais como capacidade de
retenção de água (CRA). O intervalo de variação dos parâmetros de
pH, CRA, temperatura e cor foram analisados e associados para
qualif icar a carne suína (OURIQUE e NICOLAIEWSKY, 1990).
Joo et al. (2000) descreveram que os fatores mais importantes
para classif icação da qualidade de carne suína foram medidas de pH
após 24 h de abate, cor (L*) e perda de esxudato (drip loss).
Warris e Brown (1987) classif icaram carcaças como PSE quando
apresentaram valores de pH inicial inferior a 5,8 e como DFD, quando
os valores de pH final foram iguais ou superior a 6,0.
A medida da cor através do valor L* (sistema CIELAB) em
associação com pH pode contribuir sobremaneira na caracterização do
PSE (BARBUT, 1997; FLETCHER, 1999; OLIVO et a l . , 2001). A medida
24h após o abate da luminosidade da carne considerada crít ica para
caracterizar PSE está entre 49,0 e 52,0. Valores superiores a esta faixa
implicaram na existência de carne pálida e potencialmente de qualidade
inferior (LARA et al., 2003). A palidez da carne foi diretamente
relacionada com a desnaturação protéica causada pelo baixo pH
(SWATLAND, 1993). A cor observada na superfície das carnes foi
proveniente do resultado da absorção seletiva pela mioglobina,
provocada pela distribuição da luz que emergiu da carne com a

30
diminuição do pH; ocorrência do aumento na birrefringência, com
menos luz sendo transmitida através das fibras e mais luz sendo
dispersa (BENDAL & SWATLAND, 1988; SWATLAND, 1993).
Wirth (1986) descreveu sobre as propriedades e características
da carne PSE e uti l ização na elaboração de produtos cárneos (Quadro
2). Segundo o autor, a carne PSE pode ser destinada até certo l imite
para a elaboração de alguns produtos fermentados e certos t ipos de
emulsionados, porém é inadequada para elaboração de presunto cozido
e outros produtos curados cozidos.
QUADRO 2 – Propriedades e características das carnes PSE*
PROPRIEDADES CARACTERÍSTICAS Capacidade de retenção de água
Maior perda por gotejamento (1 a 4%) em carne fresca; Maior depósito de gelatina e gordura (3 a 5%) em produtos emulsionados; Menor rendimento (3 a 6%) para presunto cozido.
Absorção de ingredientes de cura
Aumento na absorção do sal e alteração na cor curada (esbranquiçada).
Sensoriais Sabor ácido acentuado. Vida úti l Reduzida devido à superfície úmida da
carne fresca. *Fonte: WIRTH, 1986

31
3.2 DFD
Assim como a carne PSE, a carne DFD (dark, f irm, dry) ou dark
cutt ing como é conhecida na terminologia inglesa, foi atribuída a um
mau manejo pré-abate. Os exercícios físicos, transporte,
movimentação, jejum prolongado e contato com suínos estranhos ao
seu ambiente acarretaram o consumo das reservas de glicogênio,
ausência de reversão do ácido lático muscular e lentidão da glicólise
(PARDI et al. , 1993). O pH reduziu l igeiramente nas primeiras horas e
depois se estabil izou, permanecendo ao final em níveis superiores a
6,0 (FELÌCIO, 1986). No caso da carne com anomalia DFD, à falta de
glicogênio já se fez presente na respectiva musculatura antes do abate.
As razões foram quase que exclusivamente de natureza exógenas
(fortes contrações e jejum prolongado) e, por esse motivo, a condição
DFD pode ser prevenida eficientemente pelo manejo pré – abate
adequado (FISCHER, 1987).
Na carne DFD, em decorrência do pH alto, as proteínas
musculares conservaram uma grande capacidade para reter água no
interior das células. Como conseqüência, a superfície de corte do
músculo f icou pegajosa e muito escura. Ainda devido ao pH alto, essa
carne se deteriorou com facil idade (FELÌCIO, 1986).
Na prática, diversas situações podem levar a carne DFD. Em um
estudo onde foram avaliadas condições de transporte, foi relatado que
18% dos suínos transportados num caminhão com piso escorregadio,
sem divisões e sem qualquer mecanismo de venti lação apresentaram a

32
anomalia DFD na carne. Por outro lado, somente 10% dos suínos foram
DFD após o transporte num veículo adequado, com piso não
escorregadio, divisões e mecanismo de venti lação (NIELSEN, 1977).
Wirth (1986) apresentou as características da carne DFD e
l imitações de sua uti l ização na elaboração de produtos cárneos
(Quadro 3). A carne DFD pode ser uti l izada para o processamento de
produtos emulsionados (salsicha tipo Frankfurt) e produtos curados
cozidos (formulados com 60% de carne normal para que seja obtida
uma coloração desejável), não sendo recomendada para o
processamento de produtos fermentados e secos (BARTON-GADE,
1985).
QUADRO 3 – Propriedades e características das carnes DFD*
PROPRIEDADES CARACTERÍSTICAS Capacidade de retenção de água
Menor l iberação de água no tratamento térmico; Vantajoso para salsicha Frankfurt e presunto cozido (produtos mais suculentos e macios).
Absorção de ingredientes de cura
Redução da absorção do sal em porções musculares maiores Pobre desenvolvimento e retenção de cor curada.
Sensoriais Ausência do sabor ácido. Vida úti l Reduzida tanto para a carne fresca
como para produtos industrial izados. *Fonte: WIRTH,1986

33
3.3 Manejo Pré-abate
O termo estresse é uma expressão comum para designar o
conjunto de reações do organismo a agressões de ordem físicas,
psíquicas e outras, capazes de perturbar a homeostase (SIEGEL,
1995).
O manejo pré abate incluiu a mistura de grupos diferentes,
transporte e tempo de repouso na pocilga do abatedouro, práticas que
poderiamm induzir ao estresse e interferir na qualidade da carne
(FERNANDES et al. , 1979).
A intensidade do manejo a que os suínos são submetidos no
carregamento, transporte e chegada no abatedouro foi um dos
principais fatores responsáveis pela ocorrência do estresse pré-abate
(BERTOL,2004).
O estresse pré – abate pode ser dividido em estresse de período
longo, como manejo da granja, misturas de lotes, embarque e
transporte dos animais e o estresse de período curto, incluiu as
condições na pocilga de repouso e a condução dos suínos até o abate.
Esses dois t ipos de estresse não deveriam ser considerados
separadamente, principalmente porque o estresse de período longo
favoreceu ocorrência de carnes DFD, enquanto que o estresse de
período curto favoreceu as carnes PSE (ANGERAMI, 2004).
O embarque, transporte e desembarque dos suínos foram os
fatores pré-abate que mais influenciaram a qualidade da carne.

34
Contribuíram para isso o jejum, movimentação, mistura de lotes,
mudança de local, espaço disponível para cada animal, ruídos,
temperatura e outros eventos estranhos ao animal. Todos contribuiram
para aumentar o estresse, provocar brigas e por conseqüência há
escoriações e prejuízos para a qualidade da carne. Warris (2001)
enfatizou que o mau manejo nos períodos de seleção, embarque,
transporte, desembarque e espera do animal ao abate despertou
condição negativa ao bem-estar, ocasionando quedas no rendimento da
carne. Quando o manejo foi realizado de forma precária ou inadequada,
houve riscos de aumentar a mortalidade em trânsito e a incidência de
danos à carcaça. Além disso, cresceu o número de animais
estressados, gerando maior quantidade de carne PSE e DFD.
Um dos fatores considerado crít ico para o bem estar do animal é
a sua movimentação inadequada. Assim, dependendo da forma de
condução dos suínos, pode-se observar que maus tratos, esforços e
excessiva uti l ização de bastão elétrico, resultaram em perdas
econômicas em função da redução da qualidade obtida. A facil idade de
manejo dos suínos com descargas elétricas, al iadas a falta de
regulamentação, colaboraram com o uso intensivo desse disposit ivo e
conseqüentemente, induziu defeitos na carne, pelo aumento dos níveis
de estresse. Ludtke et al . (2004) relataram que, sem uti l ização do
bastão na movimentação dos suínos, diminuiu os níveis de estresse
suíno, reduzindo em 60% a incidência de carne PSE.
Os métodos de insensibil ização podem influenciar na qualidade
da carne suína. Os suínos insensibil izados eletricamente apresentaram

35
um declínio mais rápido de pH no início do post mortem e diminuição
da capacidade de retenção de água(CRA), quando comparado com os
suínos submetidos à insensibil ização gasosa (CO2), onde o pH em 24h
não foi afetado (CHANONN et al., 2000).
3.3.1 Transporte
Para minimizar os efeitos negativos desses eventos, é
indispensável usar embarcadouros na mesma altura da carroceria do
caminhão, com piso antiderrapante ou rampas com no máximo de 20º
de inclinação, pois os suínos apresentaram dif iculdades em percorrer
descida e de subir quando as inclinações foram superiores a 20º. E a
densidade máxima recomendada foi de 2,5 suínos de 100 kg por m² e o
transporte efetuado durante a noite para aproveitar melhor as horas
mais frescas ou em período de menor temperatura. No transporte, o
motorista deveria dir igir com todo cuidado, evitando freadas bruscas e
outras atitudes que prejudicariam o bem estar dos animais. Todas as
práticas relacionadas com o embarque, transporte e desembarque, se
não conduzidas adequadamente, poderiam contribuir com aumento de
carne PSE nas carcaças (FÁVERO, 2004).
No transporte, a qualidade do veículo, ventilação, densidades e
distâncias das viagens foram importantes para o nível de estresse
induzido nos animais (BARTON-GADE, 1997; WARRIS, 1998 e
FAUCITANO, 2001). A partir do princípio que os animais não
apresentaram totalmente tranqüilos no transporte, viagens mais curtas

36
foram plenamente justif icadas. Portanto, à distância percorrida foi
menos importante do que o tempo de transporte, uma vez que os
eventos que afetaram o tempo (coleta de animais nas diversas granjas,
tráfego lento, engarrafamento, etc) poderiam afetar adversamente no
estresse do animal. Barton-Gade (1985) constataram que a incidência
de PSE foi mais elevada em transportes de curta duração de 30 min e
mais baixa em transportes mais longos de 80 min.
3.3.2 Jejum
O jejum alimentar refere-se ao período em que o animal
permanece na granja com água disponível antes do embarque,
acrescido do tempo de transporte e espera nas instalações de recepção
do abatedouro.
O jejum foi classif icado como o primeiro ponto crít ico do
transporte, e o seu tempo influenciou as outras condições do estresse
no transporte, por ser responsável pelo aumento total das perdas
(ANGERAMI, 2004).Em muitos países, o jejum pré-abate de 12-15 h
consistiu numa prática comum para se reduzir o risco de contaminação
microbiana no abate (WARRIS, 1994). Foi investigado o jejum como
uma maneira de se reduzir as reservas de glicogênio muscular em
suínos no momento do abate, para aumentar o pH em 24h, melhorando
assim, a CRA e a cor (ROSENVOLD et al., 2003).
O uso do jejum pré-abate é uma prática que vem sendo uti l izada
ao longo dos anos, com o objetivo de reduzir as mortes no transporte e

37
l imitar o potencial de contaminação microbiana das carcaças no
processo de evisceração. Com relação às respostas f isiológicas
relacionadas com o manejo, os suínos em jejum submetidos a manejo
de alta ou baixa intensidade apresentaram reduzida temperatura
corporal, quando comparado com suínos alimentados (BERTOL, 2004).
O jejum pode influenciar posit ivamente algumas das
características de qualidade de carne como pH inicial, cor e capacidade
de retenção de água. Foi observado que o jejum pré-abate reduziu a
incidência de carne PSE, porém esta resposta foi mais evidente em
suínos susceptíveis ao estresse. A melhoria da qualidade da carne e a
redução da incidência de PSE em suínos submetidos a jejum, pode
estar parcialmente relacionada com a redução do metabolismo basal e
da temperatura corporal do animal, que resultou em redução da
temperatura da carcaça, e possivelmente uma desaceleração do
metabolismo anaeróbico pós-mortem, já que para ocorrência da
condição PSE foi necessária a condição de elevada temperatura e
baixo pH do músculo(BERTOL, 2004).
O intervalo de jejum ótimo foi considerado entre 16 e 24h
(EIKELENBOOM et al . , 1989). Em se tratando de monogástricos, o
período de repouso, em hipótese alguma, deveria ser inferior a 8h, em
jejum e dieta hídrica (BRASIL, 1995). A etapa de espera no abatedouro
permitiu que os animais recuperassem do estresse no transporte,
favorecendo a recuperação dos níveis de glicogênio. Se mal aplicada,
esta etapa, poderá representar um estresse adicional. Os suínos
abatidos logo após a sua chegada ao abatedouro poderiam produzir até

38
40% de carcaças PSE (EIKELENBOOM e BOLINK, 1991). Entretanto,
longos períodos de jejum implicaram em perda de peso, que variou de
0,12 a 0,20% por hora (WARRIS, 1982).
Períodos de jejum muito prolongados devem ser evitados, pois em
suínos foi observado um aumento na ocorrência de carne DFD pela
diminuição da concentração do glicogênio muscular. O tempo de jejum
muito curto resultou em maior freqüência de carnes PSE pela grande
disponibil idade de glicogênio.
Khöler e Freitas (2005) avaliaram dois tratamentos de 3 e 9 h de
descanso e jejum no frigoríf ico, e indicaram que não houve alteração
de modo signif icativo sobre a qualidade da carne suína, porém com 3h
de descanso obteve-se como redução o estresse, escoriações de pele,
hematomas e fraturas.
3.4 Composição de Ácidos Graxos e Oxidação Lipídica
A oxidação l ipídica é considerada uma das principais causas da
perda de qualidade das carnes, por ser um processo de degradação
que afeta diretamente a aceitabil idade.
A oxidação de l ipídios ou rancidez é a deterioração mais
importante que ocorre nesse tipo de produto e define a vida úti l , na
medida em que há geração de produtos indesejáveis do ponto de vista
sensorial e destruição de vitaminas l ipossolúveis e ácidos graxos
essenciais (OSAWA et al. ,2005).

39
A rancidez inicia-se logo após a morte do animal e afeta a cor,
sabor, textura e valor nutr it ivo das carnes (MORRISSEY et al. , 1998).
Os produtos primários como peróxido e hidroperóxido formados
durante a oxidação l ipídica, são instáveis e se decompõem em produtos
secundários que variam conforme o t ipo de ácido graxo (ARAÚJO,
2004). Como conseqüência compostos são l iberados como alcanos,
alcenos, aldeídos (malonaldeído, hexanal, hidroxihexanal, pentanal),
cetonas, ácidos e álcoois. Em al imentos alguns desses produtos são
voláteis e uti l izados como indicadores da oxidação l ipídica,
principalmente o malonaldeído que é capaz de reagir com o ácido 2-
tiobarbitúrico (TBA) originando um pigmento vermelho que quando
mensurado pode indicar os níveis de rancidez dos produtos
(TARLADGIS et al. , 1964).
O metabolismo de l ipídios pode ser alterado em função dos
fatores estressantes a que são submetidos os animais antes do abate.
Soares (2003) e Soares et al. (2003) constataram que a atividade da
enzima fosfolipase A2 (PLA2) foi aumentada nas carnes de frango com
PSE. A PLA2 é uma enzima l ipolít ica dependente de Ca2+ que catalisou
a hidrólise da posição sn-2 dos glicerofosfolipídios das membranas das
células, l iberando ácido araquidônico (20:4 n-6) e l isofosfolipídos
(MURAKAMI e KUDO, 2002).
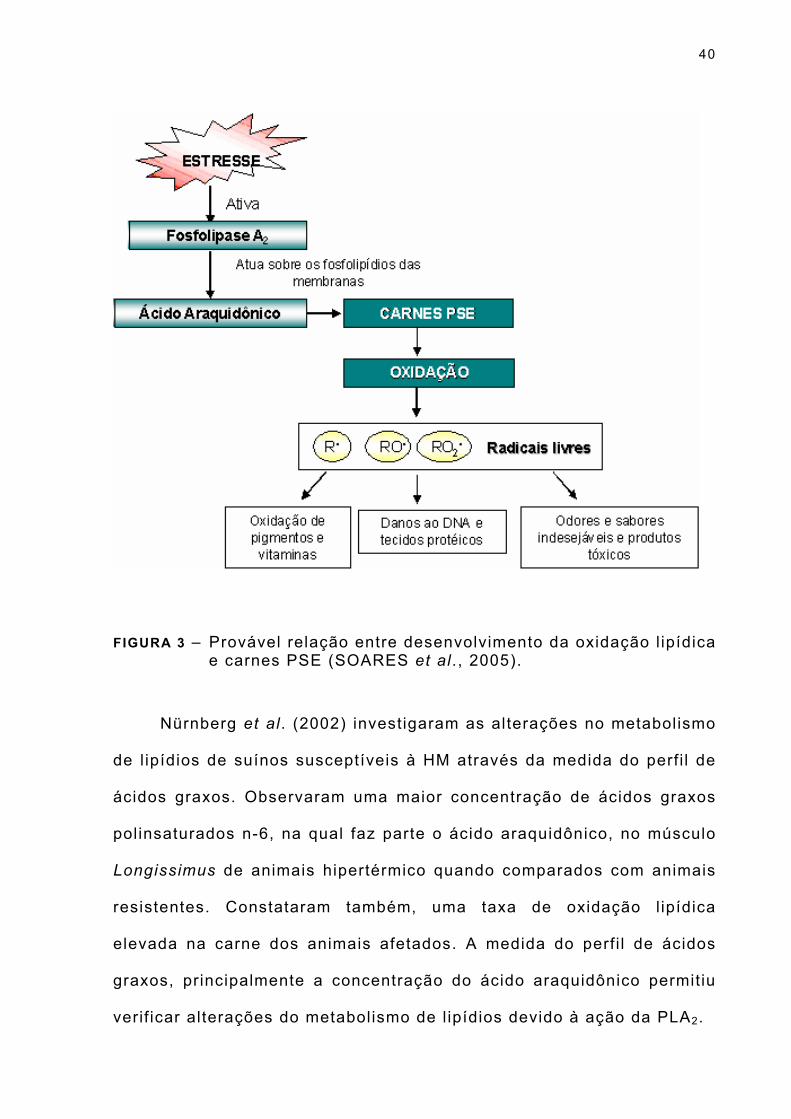
Soares et al. (2005) apresentaram (Figura 3) uma possível
explicação entre a relação do desenvolvimento da oxidação l ipídica e
carnes PSE.
40
FIGURA 3 – Provável relação entre desenvolvimento da oxidação l ipídica e carnes PSE (SOARES et al. , 2005).
Nürnberg et al. (2002) investigaram as alterações no metabolismo
de l ipídios de suínos susceptíveis à HM através da medida do perfi l de
ácidos graxos. Observaram uma maior concentração de ácidos graxos
polinsaturados n-6, na qual faz parte o ácido araquidônico, no músculo
Longissimus de animais hipertérmico quando comparados com animais
resistentes. Constataram também, uma taxa de oxidação l ipídica
elevada na carne dos animais afetados. A medida do perfi l de ácidos
graxos, principalmente a concentração do ácido araquidônico permitiu
verif icar alterações do metabolismo de l ipídios devido à ação da PLA2.

41
3.5 Valor de R
O valor de R representa a razão entre monofosfato de inosina
(IMP) e tr i fosfato de adenosina (ATP) e avalia o consumo de ATP no
momento do abate.
As carnes PSE e DFD apresentaram alterações nos níveis dos
nucleotídeos adenosina e inosina. A depleção de ATP (adenosina
trifosfato) foi a causa do início do r igor mortis, após a morte do animal
o ATP foi rapidamente convertido em ADP (adenosina difosfato) e
subsequentemente em AMP (adenosina monofosfato) e IMP (inosina
monofosfato). Nas carnes PSE e DFD a quebra dos nucleotídeos de
adenosina em IMP ocorreu mais rapidamente do que em carnes
normais conforme proposto por Soares et al. (2007) (Figura 4).
Portanto, músculos PSE e DFD apresentaram baixos níveis de ATP e
altos níveis de IMP, enquanto que elevada concentração de ATP e
baixa de IMP foram observadas para músculos normais (BATLE et al. ,
2000).

42
FIGURA 4 – Situação de stress e formação de IMP Em si tuação de estresse, há maior l iberação de Ca2 + promovendo aceleração
metaból ica do músculo, acarretando em queda brusca do valor de pH,
proporcionando assim carnes PSE. Ao mesmo tempo, acontece a quebra do ATP
em ADP e este f inalmente em IMP e a quant idade destes componentes do
nucleotídeo pode var iar de acordo com as condições de carnes PSE, DFD e normal
(SOARES et al . , 2007).
ATP ADP IMP
Estresse
Anóxia
músculo
Glicólise Anaeróbia
Ácido Lático
pH
Metabolismo Acelerado

43
3.6 Ultra-estrutura da Carne
Existem fatores que influenciam diretamente o r igor mortis e
afetam a ultra-estrutura da carne tais como espécie, genética, manejo
pré- e pós-abate e f isiologia do animal. A f igura 5 apresenta as
características de um músculo com relação a sua ultra-estrutura
conforme Judge et al. (1989). As miofibri las são estruturas que
apresentam um diâmetro de 1 a 2 µm e correm longitudinalmente a
f ibra muscular. São constituídas por uma unidade estrutural
denominada sarcômero, as quais possuem diversas proteínas, onde irá
ocorrer o processo de amaciamento após o abate. Possui um arranjo
ordenado de pontos denominados de miof i lamentos (f i lamentos grossos
e f inos), o qual foi possível identif icar a unidade funcional do músculo
como o sarcômero que é definido como a distância entre dois discos Z.
No sarcômero estão distribuídos os f i lamentos grossos cujo
componente principal é a miosina e os f i lamentos delgados que são
constituídos pelas proteínas actina, tropomiosina, troponinas, beta e
gama actininas. No disco Z estão alocadas outras proteínas como
euactinina, desmina, f i lamina, vimetina e sinemina. Na banda I está a
nebulina e espalhada por todo o sarcômero encontra-se a t i tana,
associadas aos fi lamentos delgados e grossos, respectivamente
(SWARTZ, 1994). A Figura 6 apresenta a estrutura do sarcômero e
suas proteínas, conforme i lustrado por Shimokomaki et al . (2006). A
função destas proteínas musculares é manter a integridade estrutural
das miofibri las (PRICE, 1991). A sua degradação irá causar um

44
enfraquecimento da miofibri la e conseqüentemente o amaciamento da
carne (SHIMOKOMAKI et al. , 2006).
FIGURA 5 – Ultra-estrutura muscular (JUDGE et al., 1989)
FIGURA 6 - Estrutura do sarcômero e suas proteínas (SHIMOKOMAKI
et al., 2006)

45
As mudanças ocorridas no processo de amaciamento foram
resultantes da proteólise, onde proteases localizadas dentro da célula
muscular ou no citosol contribuíram potencialmente para o
amaciamento (JIANG, 1998). O sistema calpaínas atuam sobre certas
proteínas na disco Z, tais como, desmina, f i lamina, nebulina e em
menor extensão a t i tana e requerem íons cálcio (Ca²+) para a sua
atividade. As calpaínas degradam o disco Z, atuando sobre a desmina,
f i lamentos grossos sobre a t itana e f inalmente sobre os f i lamentos
delgados digerindo a nebulina e a troponina T e conduz ao
enfraquecimento do sarcômero (SHIMOKOMAKI et al. , 2006).
No Brasil em 2006 a produção de carne suína foi de 2,825
milhões de toneladas e ocupou a posição de 4º maior produtor mundial.
Desta produção 81% foram consumidos no mercado interno, sendo 30%
in natura e 71% industrial izada, e os 19% restante foram para o
mercado internacional. E considerando que as carnes PSE e DFD
apresentaram grandes perdas e l imitações de uti l ização para indústria
processadora, este trabalho foi desenvolvido para apresentar a
incidência do fenômeno PSE e DFD e contribuir com a sua
caracterização que foi conduzido em uma indústria processadora de
carne suína nacional de grande porte.

4 MATERIAL E MÉTODOS
4.1 Matéria Prima e Classificação dos Lombos em PSE, DFD e Normal
Foram uti l izados como matéria-prima 946 lombos (músculo
Longissimus dorsi), provenientes de suínos da l inhagem Dalland,
machos castrados e fêmeas, de um Frigoríf ico da Região Sul de grande
representatividade no setor que abate aproximadamente 3000 suínos
por dia. O experimento foi realizado no verão em fevereiro de 2006, no
período de uma semana. Os suínos foram abatidos com peso médio de
90kg através do processo convencional com insensibil ização elétrica
(300 V e 1A), sangria (com os animais erguidos), escaldagem em água
a 59,4ºC, evisceração e toalete f inal. As carcaças permaneceram por
24h em câmara fr igoríf ica a 0 ± 1ºC e o músculo Longissimus dorsi foi
removido para as análises de pH e cor para classif icar e investigar a
incidência de carnes PSE, DFD e Normal na carne suína.
4.2 Caracterização Bioquímica dos Lombos PSE, DFD e Normal
Após a classif icação das carnes em PSE, DFD e Normal pela
análise de pH e cor, foram coletados 36 lombos, sendo 12 PSE, 12 DFD
e 12 Normal, codif icados com numeração aleatória de três dígitos e

47
embalados em sacos plásticos e congelados a -18ºC. Em seguida,
foram transportados para o laboratório onde permaneceram congelados
(-18ºC) até a realização das seguintes análises: perda de peso por
cozimento (PPC), maciez, índice de fragmentação miof ibri lar (IFM),
l ipídios totais, oxidação l ipídica e perfi l de ácidos graxos. Para
determinação do Valor de R 5g de amostras foram mantidas em ácido
perclórico 1M e congeladas a -18ºC. Para análise de microscopia os
lombos PSE, DFD e Normal foram coletados no fr igoríf ico, conforme
descrito no item 4.2.11.
4.2.1 Medida de pH24h
As medidas de pH24h foram mensuradas em duplicata, uti l izando
potenciômetro de contato Sentron 1001 diretamente na parte dorsal do
músculo Longissimus dorsi.
4.2.2 Medida de Cor
As medidas de cor foram realizadas tomando-se três pontos
diferentes de leitura para cada amostra. A medida de cor foi analisada
nas mesmas amostras da determinação de pH. Foi uti l izado o
colorímetro Gardner 45/0, com esfera de integração e ângulo de visão
de 10º e i luminante D65. Os valores de luminosidade L*, a*

48
(componente vermelho-verde) b* (componente amarelo-azul) foram
expressos conforme o sistema de cor CIELAB.
4.2.3 Classificação das Amostras em PSE e DFD
O valor de L* foi uti l izado como parâmetro para classif icação dos
lombos em DFD, PSE e Normal conforme recomendações de Warris e
Brown (1987) e Channon et al. (2000). Os lombos com valores de L*24h
> 53 foram classif icados como PSE, com L*24h < 45 como DFD e com
valores intermediários 45 ≤ L*24h ≤ 53 como Normal.
4.2.4 Medida do Valor de R
O valor de R representa a razão entre monofosfato de inosina
(IMP) e tr i fosfato de adenosina (ATP) e avalia o consumo de ATP no
momento do abate e a determinação foi realizada segundo metodologia
de Honikel e Fischer (1977). Neste procedimento foram coletados
aproximadamente 5g de lombo suíno resfriado, colocados em potes
plásticos e adicionado 20 mL de ácido perclórico 1M até realização da
análise.
Uma alíquota de 2 g de amostra l ivre de tecido conjuntivo e
gordura foram homogeneizados com 20 mL de ácido perclórico 1M em
Ultra Turrax por 30 s, na proporção de 1:10 em p/v. A seguir, foi

49
centrifugado por 5 min a 3000g e o sobrenadante foi f i l trado em papel
de f i l tro comum. Uma alíquota de 0,1 mL do sobrenadante foi di luída
em 4,9 mL de tampão fosfato 0,1 M pH 7,0. As leituras foram realizadas
em espectrofotômetro a 250 nm (IMP) e 260 nm (ATP). O tampão
fosfato foi uti l izado como branco. O valor de R foi expresso como a
razão entre absorbância a 250 nm e 260 nm.
4.2.5 Medida da Perda de Peso por Cozimento (PPC)
Para realização da PPC as amostras foram descongeladas sob
refrigeração, cortadas em porções de 2cm e embaladas em saco
plástico hermeticamente fechado. Estas foram pesadas e logo após
foram cozidas em banho-maria a 85ºC até atingir a temperatura interna
de 78ºC. Após o cozimento a água exsudada pela amostra foi
desprezada e pesou-se novamente. O resultado foi expresso em % de
água perdida, considerando o peso inicial e f inal da amostra.
4.2.6 Medida da Maciez
Para análise de textura, as amostras foram descongeladas sob
refrigeração por 24 h. Para medida de maciez das amostras in natura
foram uti l izadas amostras descongeladas. Para determinação da

50
maciez das amostras cozidas, foram uti l izadas as mesmas amostras da
PPC.
A maciez das amostras in natura e cozida foram mensuradas por
meio de força de cisalhamento, uti l izando Texturômetro Universal
TATX-2i, equipado com lâmina Warner Bratzler.
As amostras foram cortadas nas dimensões de 1×1×2cm (altura ×
largura × comprimento), no sentido longitudinal das f ibras musculares.
As amostras foram cortadas com lâmina de Warner Bratzler no sentido
transversal da f ibra muscular. Os resultados obtidos foram expressos
como força máxima em Newton(N) necessária para o corte das
amostras.
4.2.7 Medida do Índice de Fragmentação Miofibrilar (IFM)
A extração, determinação de proteína solúvel e medida do IFM
foram realizadas de acordo com o método descrito por Culler et al.
(1978), conforme segue.
4.2.7.1 Extração
A extração foi conduzida em tripl icata. Amostras de 4g de lombo
congelado l ivre de gordura e tecido conectivo foram colocadas em um
becker de plástico, adicionados 20mL de solução tampão IFM, contendo
100mM KCl, 20mM de tampão fosfato (pH7,0), 1mM de EDTA, 1mM de

51
MgCl2 e 1 mM de azida sódica e homogeneizado por 30s em um
homogeneizador t ipo Ultra Turrax. Após 30s a homogeneização foi
repetida por mais 30s. O homogenato foi transferido para tubos de
centrífuga refrigerada e centrifugados por 15 min a 2ºC e 10000rpm. O
sobrenadante foi f i l trado e reservado. O decantado foi ressuspenso
com 20 mL de tampão de IFM e centrifugado novamente por 15 min a
2ºC e 10000rpm. Os sobrenadantes obtidos foram fi l trados e o
decantado foi resuspenso com 10 mL de tampão IFM e fi l trado
novamente. Assim, os três sobrenadantes foram combinados para
determinação do teor de proteína solúvel.
4.2.7.2 Determinação de Proteína Solúvel
A determinação de proteína solúvel em duplicata foi realizada
conforme o método de Biureto uti l izando uma curva padrão de albumina
sérica bovina nas concentrações de 0; 0,5 ; 1,0; 1,5 ; 2,0 e 2,5 mg/mL.
Em um tubo de ensaio colocou-se 1mL da amostra. Em seguida, foi
adicionado em todos os tubos de ensaio 4 mL de reagente de Biureto.
As absorbâncias foram lidas em espectrofotômetro no comprimento de
onda de 540nm e o teor de proteína solúvel foi calculada para
estabelecer o teor de proteína para medida do IFM.

52
4.2.7.3 Medida do IFM
A medida do IFM em tr ipl icata foi realizada em tubos de vidros
contendo quantidades apropriadas de suspensão, solução tampão e
reagente de Biureto para obter um total de 8 mL de solução com 0,5mg
de proteína/mL. No branco foi adicionado 4 mL de reagente de Biureto
e 4 mL de solução tampão IFM, homogeneizados em vórtex. As
absorbâncias foram lidas em espectrofotômetro no comprimento de
onda de 540nm. O valor do IFM foi expresso como a absorbância
mult ipl icada por 200.
4.2.8 Determinação de Lipídios Totais
A extração de l ipídios foi realizada segundo Bligh e Dyer (1959).
Neste procedimento, 30g de amostra foram pesadas em um Bequer de
250mL e adicionados na seqüência 60mL de clorofórmio e 30mL de
metanol, mais 30mL de clorofórmio e 30mL de água desti lada e agitado
vigorosamente por 10 min. A mistura foi f i l trada à vácuo em um funil de
Buchner com papel de f i ltro Whatman nº1.
O fi l trado foi colocado num funil de separação e adicionado uma
solução de NaCl 0,9% equivalente a 1/5 do volume do fi l trado ou
aproximadamente 30mL. Após a separação espontânea das fases, a
fase inferior contendo clorofórmio e l ipídios foi recolhida no balão de
250mL previamente pesado. Em seguida, foi evaporado no rotavapor
com banho a 34-36oC. Após a evaporação do solvente o balão foi

53
pesado. Os l ipídios totais foram calculados por diferença de peso
inicial e f inal do balão e os resultados expressos em porcentagem (%).
4.2.9 Medida da Oxidação Lipídica
A oxidação l ipídica foi analisada ut i l izando-se o método do ácido
2-tiobarbitúrico (TBA), descrito por Tarladgis et al. (1964) e modif icado
por Crackel et al. (1988). O método consistiu em determinar
espectofotometricamente à formação do complexo de coloração
vermelha formado pela condensação de dois moles do ácido 2-
t iobarbitúrico com um mol de malonaldeído e outras substâncias que
reagirem com o TBA.
Foram util izadas 15g de amostra, em tripl icata, homogeneizada e
acrescida de 98mL de água e 2,5mL de HCL 4 N e duas gotas de
antiespumante ( 8 partes de Span 80 mais 1,3 partes de Tween 20) em
um erlenmeyer de 500mL com algumas pérolas de vidro. Em seguida, a
mistura foi desti lada por 10 min e recolhido cerca de 50mL em um
erlenmeyer. Uma alíquota de 5mL do desti lado foi colocado em um tubo
de ensaio com tampa rosqueável. Foram adicionados 5 mL de TBA e
colocados em banho-maria fervente por 35 min, resfriados a
temperatura ambiente. A leitura foi realizada em espectrofotômetro com
comprimento de onda de 530nm. Uma curva padrão foi elaborada
uti l izando-se solução padrão de 1,1,3,3 tetraetoxipropano (TEP). O
resultado foi expresso em mg de TBARS/kg de amostra.

54
A exatidão do método foi verif icado pela medida de recuperação
do padrão adicionado à amostra comparando com leitura. A % de
recuperação foi expressa como:
( ) 100)()(
% ×++
=amostraabsTEPabs
amostraTEPabsR
4.2.10 Determinação da Composição de Ácidos Graxos
A determinação da composição do teor de ácidos graxos das
amostras PSE, DFD e Normal foi feita conforme o seguinte
procedimento:
A extração de l ipídios foi realizada segundo Bligh e Dyer
(1959), e descrito no item 4.2.8.
A transesterif icação dos l ipídios foi realizada conforme método
5509 da ISO (1978). Aproximadamente 0,200g de l ipídios extraídos,
foram transferidos para um tubo de 10mL com tampa de rosca. Foram
adicionados 2mL de n-heptano e agitado até a completa dissolução.
Foram adicionados 2mL de KOH 2M e agitados vigorosamente até
obtenção de uma solução turva. As fases foram separadas
espontaneamente, e a fase superior contendo n-heptano e ésteres
metíl icos de ácidos graxos foi recolhida num frasco e adicionado o
nitrogênio e fechado hermeticamente. Os frascos foram armazenados
no congelador (-18ºC) até a análise e, realizada por cromatografia
gasosa na Universidade Estadual de Maringá.

55
Os ésteres de ácidos graxos foram separados e quantif icados
uti l izando o cromatógrafo gasoso Variam CP3380, equipado com
detector de ionização de chama e coluna capilar de síl ica fundida CP-
7420, (coluna capilar de dimensão 100 m × 0,25 mm × 0,39 µm). A
temperatura da coluna foi programada uti l izando 165ºC por 18 min,
180ºC (30ºC/min) por 22 min e 240ºC (15ºC/min) por 20 min, uti l izando
uma pressão de 40 psi. As temperaturas do injetor e do detector foram
mantidas a 220ºC e 245ºC, respectivamente. Os fluxos dos gases foram
de 1,2mL/min para o gás de arraste (H2 ); 30mL/min para o gás auxil iar
(N2 ) e 300 mL/min para os gases da chama(H2 e ar sintético),
respectivamente. A razão de divisão da amostra (split) foi de 1/80. A
identif icação dos ácidos graxos foi feita comparando os tempos de
retenção relativos dos picos das amostras com padrões de ésteres
metíl icos de ácidos graxos (Sigma). As áreas de picos foram
determinadas pelo CG-300 integrador (CG Instruments, Brasil). O teor
de ácidos graxos foram expressos como porcentagens de áreas
normalizadas.
4.2.11 Microscopia Eletrônica dos Lombos Suínos
Após a classif icação dos lombos suínos em PSE, DFD e Normal
pela análise de pH e cor, amostras do músculo Longissimus dorsi foram
cortados seguindo o eixo longitudinal da f ibra e f ixados em solução
gelada (4ºC) de glutaraldeído a 2% tamponada com cacodilato de sódio

56
0,14M pH 7,4 contendo sacarose a 180mM. Em seguida, as amostras
foram lavadas em tampão cacodilato 0,1M a pH 7,2, lavadas em ósmio-
cacodilato (um volume de tetróxido de ósmio a 2% em água desti lada
0,1 M a pH 7,2 mais sacarose na concentração de 10,56 g por 100ml) e
permaneceram nesta solução por mais duas horas na temperatura de
4ºC, sofrendo agitação contínua por rotação. Logo após foram lavadas
rapidamente em solução de cloreto de sódio a 0,9% com sacarose na
concentração de 504 g por l itro. Em seguida os fragmentos foram
colocados por 24 h em solução de acetato de uranila a 0,5% em água
desti lada com sacarose na concentração de 10,56 g por 100 ml a 4ºC.
A seguir as peças foram desidratadas em uma série de soluções de
etanol com concentrações crescentes, seguindo-se de duas passagens
em óxido de propileno puro por 20 min cada. Posteriormente realizou-
se a infi l tração com óxido de propileno mais resina em partes iguais por
12 h, seguidas por uma passagem em resina pura por 1 h a 37ºC e
incluídas em resina Spurr (SEM, Hatfiel, Philadelphia, USA) em moldes
de sil icone por 2 dias. Após a polimerização os blocos foram cortados
em ultramicrótomo Porter Blum MT-2, com obtenção de cortes de 0,5
µm de espessura corados com mistura de azul de meti leno 1% e azur II
1% em solução de bórax 1% e observados ao microscópio óptico. Após
a seleção da região de interesse, foram levados ao ultramicrótomo e
obtidos cortes de 70 nm, colhidos em telas de cobre de 200 malhas e
contrastados pelo acetato de uranila a 2% em água desti lada por 1h,
seguindo-se coloração por citrato de chumbo a 0,5% em água desti lada
por 30 minutos após lavagem. O exame dos cortes ultrafinos e as

57
eletron-micrografias foram feitos no microscópio eletrônico JEOL
100CX II do Departamento de Biologia Celular e do Desenvolvimento do
Instituto de Ciências Biomédicas da Universidade de São Paulo.
4.2.12 Análise Estatística
O programa STATISTICA for Windows versão 6.0 foi uti l izado
para as análises dos resultados.
Os lombos suínos foram classif icados com base no valor de L* em
PSE, DFD e Normal e o teste de médias Tukey foi uti l izado para
comparar as diferenças signif icativas entre os três t ipos de lombo com
relação às medidas de pH, cor, valor de R, perda de peso por
cozimento (PPC), maciez, índice de fragmentação miof ibri lar (IFM),
l ipídios totais, oxidação l ipídica e teor ácidos graxos.
O teste de coeficiente de correlação de Pearson foi uti l izado para
avaliar a correlação entre as variáveis pH, L* e b*.

5 RESULTADOS E DISCUSSÃO
5.1 INCIDÊNCIA DE PSE E DFD NA CARNE SUÍNA
Após análise de pH e cor dos lombos suínos (Longissimus dorsi)
as 946 amostras foram classif icadas em PSE, DFD e Normal.
Assim, foi construído o histograma (Figura 7) de distribuição do
valor de L*24h dos lombos suínos. A distribuição da freqüência com
relação ao valor de L*24h de 946 lombos suínos está expressa como
uma curva t ipicamente normal com variação ampla de luminosidade de
muito clara (40), média (50) e muito escura (62).
O histograma de distribuição do pH24h de 946 lombos suínos está
apresentado na Figura 8. Observa-se também uma curva de
distribuição tipicamente normal com valores de pH de 5,3, 5,5 até 6,0.
Assim, presença destas 946 amostras de lombos suínos nos dois
extremos de valor de L*24h associada ao valor de pH24h, sugere o
aparecimento de carnes DFD e PSE.

59
40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60
Valor de L*
0
20
40
60
80
100
120
140
160
180
Freq
üênc
ia
FIGURA 7 - Histograma de distr ibuição do valor de L*24h de lombos suínos (n=946)
5,0 5,1 5,2 5,3 5,4 5,5 5,6 5,7 5,8 5,9 6,0 6,1
pH
0
50
100
150
200
250
300
350
400
Freq
uênc
ia
FIGURA 8 - Histograma de distribuição do valor de pH24h de lombos suínos (n=946)

60
Com base nos valores de L* de 946 lombos suínos, a incidência
de PSE e DFD foi estimada em 22,83% e 1,06%, respectivamente
(Figura 9). Os lombos classif icados como PSE, DFD e Normal estão
i lustrados na Figura 10. Resultados diferentes foram relatados por
Andrade et al . (1993) que encontraram uma freqüência de carcaças
com PSE de 17,2% e DFD de 13,4% na região Oeste do Estado do
Paraná e Culau et al. (1994), identif icaram uma freqüência de carcaças
com 30,69% de PSE no Estado do Rio Grande do Sul. Murray e
Johsons (1998) detectaram no Canadá em lombos suínos com
aproximadamente 13% de PSE. Entretanto, Cassens (2000) relatou nos
EUA que apesar do PSE em suínos já ser identif icado há muito tempo,
esta incidência praticamente não foi alterada, uma vez que em 1963 foi
de 18% e em 1992 cerca de 16%.
Assim, com esta incidência, em 2006 a produção brasileira de
carne suína foi de 33,71 milhões de cabeças, e que cada lombo PSE
perdeu aproximadamente 1,5% de exsudato segundo Oda et a l . , 2003 ,
pode-se estimar o prejuízo econômico de R$ 5,7 milhões. Portanto é
necessário melhorar as condições ambientais de manejo, pré-abate e
conforto animal, realizando um acompanhamento na cadeia produtiva
desses animais com, a f inalidade de se obter um padrão de manejo e
assim estabelecer procedimentos sistemáticos para sua implementação
que levem a diminuição destas perdas.
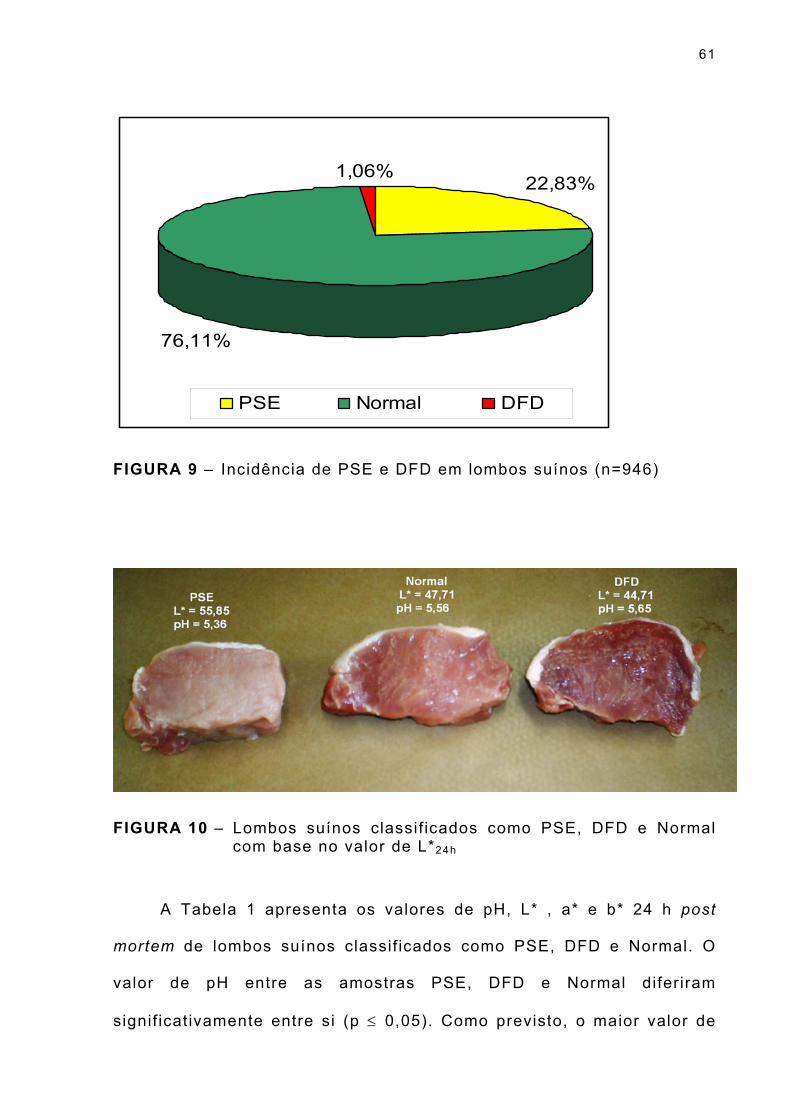
61
22,83%
76,11%
1,06%
PSE Normal DFD
FIGURA 9 – Incidência de PSE e DFD em lombos suínos (n=946)
FIGURA 10 – Lombos suínos classif icados como PSE, DFD e Normal com base no valor de L*24h
A Tabela 1 apresenta os valores de pH, L* , a* e b* 24 h post
mortem de lombos suínos classif icados como PSE, DFD e Normal. O
valor de pH entre as amostras PSE, DFD e Normal diferiram
signif icativamente entre si (p ≤ 0,05). Como previsto, o maior valor de

62
pH foi observado para os lombos DFD e o menor para os PSE.
Resultados similares também foram obtidos por vários pesquisadores
(CHANNON et al. 2000; JENSEN e BARTON-GADE; 1985; LEACH et
al. , 1996). O valor de a*24h (vermelho) não diferiu signif icativamente (p
> 0,05) entre as amostras de lombo com PSE, DFD e Normal, enquanto
que para o componente b*24h (amarelo) o maior valor foi observado
para os lombos PSE, seguidos pelo Normal e DFD.
TABELA 1 - Valores de pH, L* , a* e b* 24 h post mortem dos lombos
suínos PSE, DFD e Normal
pH L* a* b*
PSE (n=216) 5,46c±0,10 54,43a±0,46 1,15a±0,76 8,48a±0,88
DFD(n=10) 5,80a±0,16 43,95c±0,34 1,17a±0,74 6,37c±0,63
Normal (n=720) 5,52b±0,10 50,34b±0,77 1,34a±1,23 7,93b±0,97
Médias seguidas de let ras d i ferentes na mesma co luna d i ferem ent re s i pe lo teste de
Tukey a 5% de probabi l idade
A correlação entre os valores de L*24h e pH24h (Figura 11) e dos
valores de b* e L*24h (Figura 12) dos lombos suínos, músculo
Longissimus dorsi foram investigados. Uma correlação negativa
signif icativa foi encontrada (R= -0,46, p ≤ 0,05) entre o valor de L*24h e
pH24h, indicando que quanto menor o valor de pH maior a
luminosidade, ou seja, o lombo foi mais pálido e vice-versa (Figura 11).
O valor de b*24h correlacionou-se posit ivamente com o valor de L*24h e
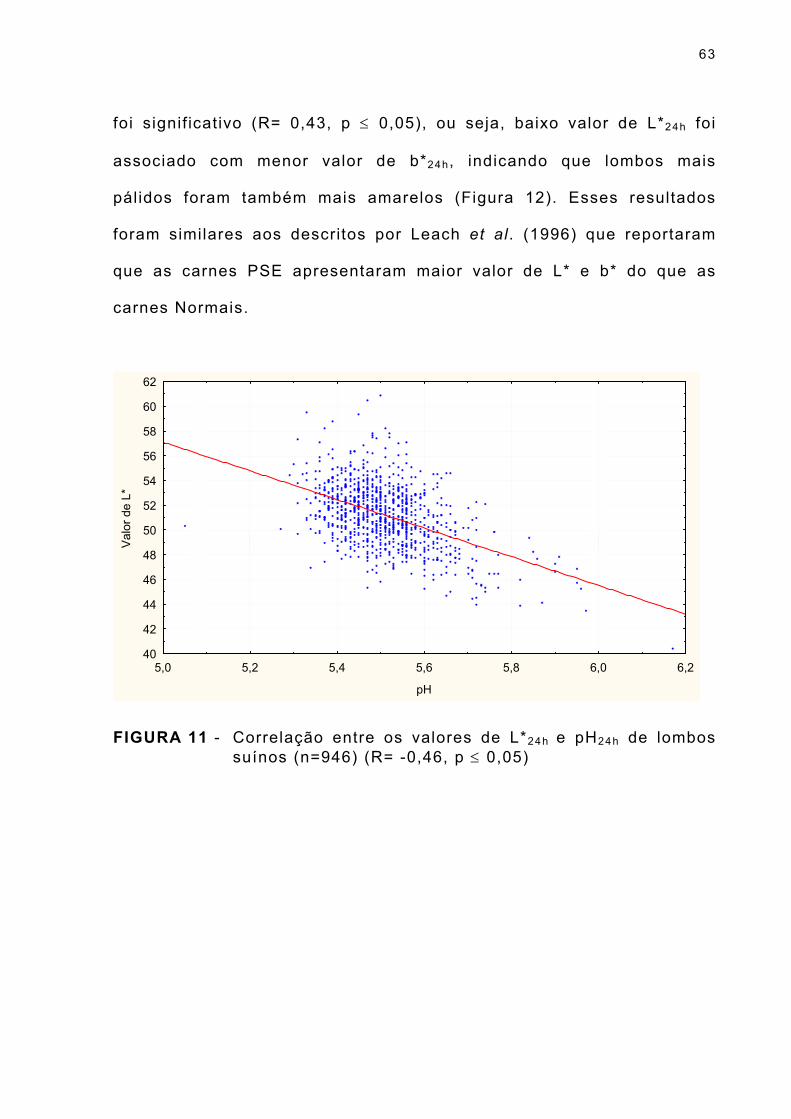
63
foi signif icativo (R= 0,43, p ≤ 0,05), ou seja, baixo valor de L*24h foi
associado com menor valor de b*24h, indicando que lombos mais
pálidos foram também mais amarelos (Figura 12). Esses resultados
foram similares aos descritos por Leach et al. (1996) que reportaram
que as carnes PSE apresentaram maior valor de L* e b* do que as
carnes Normais.
5,0 5,2 5,4 5,6 5,8 6,0 6,2
pH
40
42
44
46
48
50
52
54
56
58
60
62
Valo
r de
L*
FIGURA 11 - Correlação entre os valores de L*24h e pH24h de lombos suínos (n=946) (R= -0,46, p ≤ 0,05)

64
40 42 44 46 48 50 52 54 56 58 60 62
Valor de L*
5
6
7
8
9
10
11Va
lor d
e b*
FIGURA 12 - Correlação entre os valores de b* e L*24h de lombos suínos (n=946) (R= 0,43, p ≤ 0,05)
5.2 Caracterização Bioquímica e Avaliação da Ultraestrutura dos Lombos
PSE, DFD e Normal
5.2.1 Valores de pH24h dos Lombos Suínos
Valores de pH24h post mortem de lombos suínos classif icados
como PSE, DFD e Normal estão apresentados na Tabela 2.
O valor de pH entre amostras PSE e Normal não diferiram
signif icativamente entre si, porém, diferiram com a amostra DFD.
Segundo Honikel e Kim (1986), a principal causa do desenvolvimento

65
da condição PSE foi a decomposição acelerada do glicogênio após o
abate que ocasionou a redução do pH muscular e geralmente inferior a
5,8. Porém, em carnes suínas DFD, Felício (1986) e Pardi et al. (1993)
descreveram que os fatores associados ao pré abate acarretaram o
consumo das reservas de glicogênio, ausência de reversão de ácido
lático muscular e lentidão da glicólise que reduziu o pH l igeiramente
nas primeiras horas e estabil izou ao final em pH superior a 6,0.
Estes resultados foram comparados com outros autores que
verif icaram que os carreadores do gene halotano (Halnn) apresentaram
valores de pH menor (5,36), seguidos pelos heterozigotos (HalNn) com
pH 5,94 e animais não carreadores do gene halotano (HalNN) com pH
6,22(JENSEN e BARTON-GADE, 1985; LEACH et al., 1996; CHANNON
et al., 2000). Assim, evidencia-se nesta investigação, o aparecimento
de carnes tipicamente PSE, DFD e Normal.
TABELA 2 - Valores de pH2 4 h post mortem dos lombos suínos PSE, DFD e Normal
Lombos suínos pH PSE (n=12) 5,62b ± 0,12 DFD (n=12)
Normal (n= 12) 5,96a ± 0,12 5,69b ± 0,10
Médias seguidas de let ras d i ferentes na mesma co luna d i ferem ent re s i pe lo teste de Tukey a 5% de probabi l idade

66
5.2.2 Cor dos Lombos Suínos
Medidas de cor dos lombos suínos PSE, DFD e Normal foram
realizadas com intuito de classif icar e caracterizar o aspecto visual da
carne que estão diretamente relacionados com a aceitação e qualidade.
A Tabela 3 apresenta os valores de L* , a*, b* e razão a*/b*24h post
mortem de lombos suínos classif icados como PSE, DFD e Normal. A
média dos valores de L* para o músculo Longissimus dorsi com PSE,
DFD e Normal foram signif icativamente diferentes (p ≤ 0,05) entre si, e
o maior valor foi encontrado para lombos PSE (L*= 57,88),
intermediários para lombos normais (L*= 49,19) e o menor valor para
lombos DFD (L*= 43,42).
O estudo realizado por Sencic et al. (2002), comprovaram que
animais oriundos de cruzamentos com Pietrain apresentaram maior
produção de carne magra. Entretanto, estes animais apresentaram
deterioração da qualidade da carne, devido a alterações nos valores de
pH e cor.
A variação dos valores de L* do músculo Longissimus dorsi
(Tabela3) indicou a presença de carnes classif icadas como PSE, DFD e
Normal. Estes resultados indicaram que a carne PSE apresentou um
efeito negativo no incremento da luminosidade (L*), contribuindo para
uma coloração mais clara (L*= 57,88), que não é desejável para o
consumidor sob o ponto de vista de aparência da carne.

67
Os suínos carregadores do gene halotano apresentaram
propensão de produção de carnes PSE com valores de L* maior do que
animais l ivres do gene halotano (LEACH et al., 1996 e ANGERAMI,
2004), e no presente estudo estes resultados também foram
observados. Os valores de L* apresentados nesta investigação das
amostras com PSE, DFD e Normal estão de acordo com Fisher et al.
(2000), Leach et al . (1996) e Murray et al. (1989), que confirmaram que
o aumento na disperção da luz foi associado com a carne PSE, que
segundo Fisher et al. (2000) foi devido à ocorrência da desnaturação
protéica.
O valor de a*24h (vermelho) não diferiu signif icativamente (p>
0,05) entre amostras de lombo com PSE, DFD e Normal (Tabela 3).
Entretanto, o componente b*24h (amarelo) não diferiu entre as carnes
PSE e Normal e foram diferentes para carnes DFD. Esses resultados
confirmaram que carnes PSE apresentaram colorações mais
amareladas, que também foram observados na Figura 12.
A razão da medida de cor a*/b* indica a intensidade da coloração
vermelho e amarelo. A razão a*/b* foi signif icativamente menor (p ≤
0,05) para o lombo suíno Longissimus dorsi PSE, quando comparado
com as carnes Normais e DFD (Tabela 3). Desta forma, quanto maior a
razão maior a proporção de oximioglobina e quanto menor, maior a
proporção de metamioglobina. Portanto, o músculo Longissimus dorsi
PSE, apresentou maior teor de metamiolgobina do que as carnes
Normais e DFD. A metamioglobina segundo Cornforth (1994) é a forma
oxidada do pigmento, no qual o átomo central de ferro da mioglobina

68
está no estado férrico (Fe3+) e apresenta a coloração marrom
indesejada (CORNFORTH, 1994).
O melhoramento genético com finalidade de aumentar a produção
de carne magra alterou a taxa de crescimento muscular, devido ao
aumento do número de fibras musculares e da proliferação de células
satélites (ROSENVOLD et al., 2003). Entretanto, este melhoramento
genético resultou em uma carne signif icativamente mais luminosa e
menos vermelha e com conteúdo de pigmento menor em relação às
l inhagens do ano de 1976. Barton-Gade (1990) também observou que o
conteúdo de pigmento do músculo Longissimus dorsi diminuiu entre
início e f inal dos anos 80 na Dinamarca.
TABELA 3 - Valores de L* , a*, b* e razão a*/b* 24h post mortem para os lombos suínos PSE, DFD e Normal
Amostras L* a* b* Razão a*/b*
PSE (n=12)
57,88a±2,54
8,40a±2,48
6,44a±1,70
1,31c±0,15 DFD (n=12)
Normal (n=12) 43,42c±1,32 49,19b±1,45
8,48a±1,88 10,16a±1,45
3,57b±0,82 5,20a±1,23
2,38a±0,19 2,03b±0,39
Médias seguidas de let ras d i ferentes na mesma co luna d i ferem ent re s i pe lo teste de Tukey a 5% de probabi l idade
Uma correlação negativa e signif icativa foi observada (R= -0,75, p
≤ 0,05) entre o valor de L*24h e pH24h do músculo Longissimus dorsi
PSE, DFD e Normal (Figura 13) indicando que quanto menor o pH
maior a luminosidade, ou seja, o lombo apresenta-se mais pálido e
vice-versa.
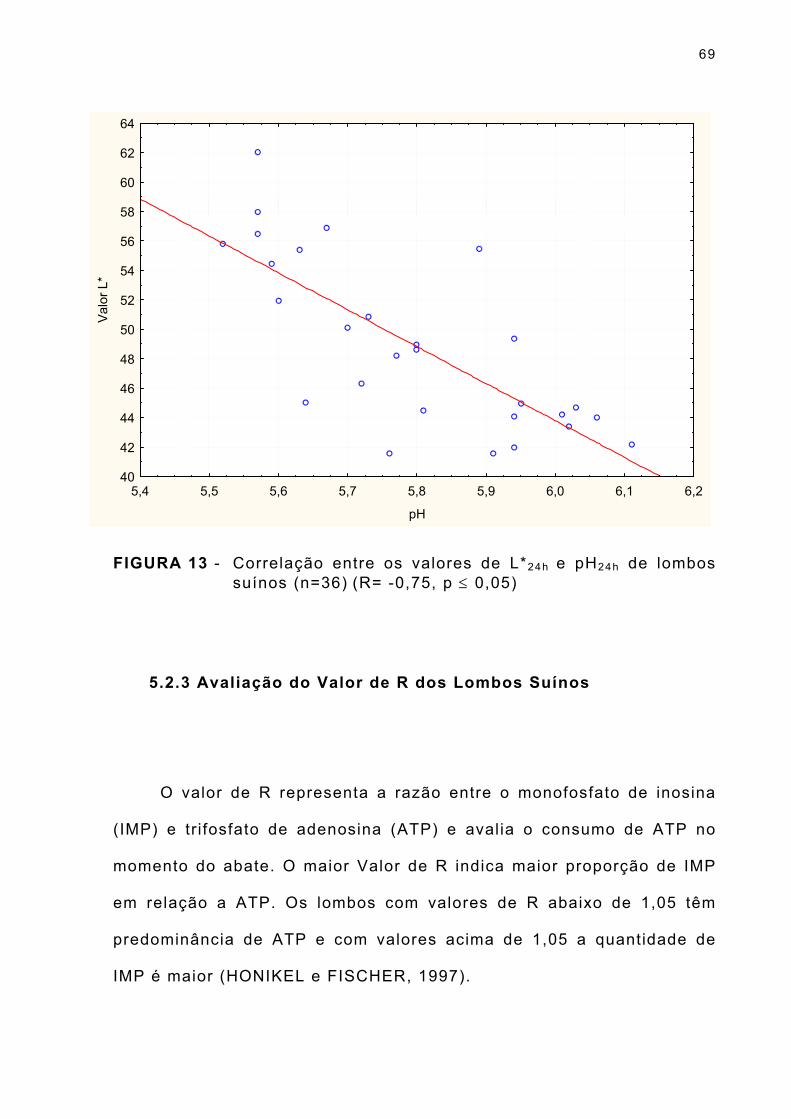
69
5,4 5,5 5,6 5,7 5,8 5,9 6,0 6,1 6,2
pH
40
42
44
46
48
50
52
54
56
58
60
62
64
Valo
r L*
FIGURA 13 - Correlação entre os valores de L*24h e pH24h de lombos suínos (n=36) (R= -0,75, p ≤ 0,05)
5.2.3 Avaliação do Valor de R dos Lombos Suínos
O valor de R representa a razão entre o monofosfato de inosina
(IMP) e tr i fosfato de adenosina (ATP) e avalia o consumo de ATP no
momento do abate. O maior Valor de R indica maior proporção de IMP
em relação a ATP. Os lombos com valores de R abaixo de 1,05 têm
predominância de ATP e com valores acima de 1,05 a quantidade de
IMP é maior (HONIKEL e FISCHER, 1997).

70
A Tabela 4 apresenta os valores de R do músculo Longissimus
dorsi classif icado como PSE, DFD e Normal. As medidas do Valor de R
para o músculo Longissimus dorsi PSE e Normal foram
signif icativamente maior quando comparado com os lombos DFD.
Os resultados obtidos para o Valor de R foram similares aos
encontrados por Batle et al. (2000), que verif icaram valores de R maior
em músculo Longissimus dorsi PSE suíno.
Os lombos PSE e Normal não diferiram signif icativamente,
provavelmente devido a coleta de amostras com r igor já completado.
Para carnes de aves os valores de R acima de 1,10 indicaram o
estabelecimento do r igor (SAMS e MILLS, 1993).
TABELA 4 – Valor de R dos lombos suínos PSE, e DFD e Normal
Lombos suínos Valor de R PSE (n=12) 1,25ª±0,05 DFD (n=12) 1,18b±0,04
NORMAL (n=12) 1,21ª±0,05 Médias seguidas de let ras d i ferentes na mesma co luna d i ferem ent re s i pe lo teste de Tukey a 5% de probabi l idade
5.2.4 Avaliação da Perda de Peso por Cozimento (PPC) dos
Lombos Suínos
A medida da PPC de músculo Longissimus dorsi, classif icadas
como PSE, DFD e Normal está apresentada na Tabela 5. Observa-se
que não houve diferença signif icativa (p> 0,05) para medidas de PPC
entre os lombos PSE, DFD e Normal. Estes resultados estão de acordo
com vários autores (Bridi et al. , 2003, Leach et al. , 1996 e Monin et al .,

71
1999), que não encontraram diferença signif icativa na PPC de animais
susceptíveis ou não ao gene halotano. Entretanto, Fisher et al. (2000) e
Culau et al. (2002) observaram que à medida que aumenta o número de
alelos do gene halotano (n), a PPC foi maior.
Neste trabalho esperava-se que o lombo suíno PSE apresentasse
um maior PPC, devido à desnaturação protéica provocada pela rápida
queda do pH post-mortem conforme observado por Honikel & Kim
(1986). Entretanto, devido o transporte dos lombos congelados para o
laboratório e realização de análises posteriores, provavelmente ocorreu
maior perda de exsudato nas amostras PSE, que não foi considerada, e
não foi possível detectar diferenças signif icativas entre os lombos na
análise de PPC. Assim, medidas de PPC deveriam ser realizadas em
amostras resfriadas logo após a desossa.
TABELA 5 – Perda de peso por cozimento (PPC) dos lombos suínos PSE, DFD e Normal.
Lombos suínos PPC (%)
PSE (n=12) DFD (n=12)
33,51ª±5,22 33,39ª±2,72
NORMAL (n=12) 34,45ª±1,72 Médias seguidas de let ras d i ferentes na mesma co luna d i ferem ent re s i pe lo teste de Tukey a 5% de probabi l idade

72
5.2.5 Maciez dos Lombos Suínos
A textura é descrita pelos consumidores como uma das principais
características da qualidade da carne, e é resultante de uma interação
dinâmica de muitos fatores que podem ser divididos em inerentes
(ante-mortem) e não inerentes (post-mortem) ao animal. Entre os
fatores inerentes, tem-se a genética, f isiologia, al imentação e o manejo
do animal e como fatores não inerentes o resfriamento,
descongelamento e maturação (EMBRAPA, 1999).
Os resultados da medida da Força de Cisalhamento (FC) em
Newton de músculos Longissimus dorsi PSE, DFD e Normal in natura e
cozido estão apresentados na Tabela 6. Não foi encontrada diferença
signif icativa (p> 0,05) entre os lombos PSE, DFD e Normal in natura e
cozido.
Resultados similares foram observados por Bridi et al. (2003),
Leach et al. (1996) e Channon et al. (2000) onde amostras de animais
carreadores ou não do alelo halotano, não apresentaram diferença
signif icativa na FC do músculo Longissimus dorsi. Entretanto, Fisher et
al. (2000) constataram que a carne proveniente de suínos carreadores
do alelo halotano apresentou maior FC, e atribuíram este resultado a
uma maior perda de água no cozimento.
O fato da FC dos lombos PSE e DFD não diferirem
signif icativamente entre si, possivelmente foi devido a PPC das
amostras que também não diferiram.

73
TABELA 6 – Força de cisalhamento (FC) em Newton(N) dos lombos suínos PSE, DFD e Normal in natura e cozido.
Médias seguidas de let ras d i ferentes na mesma co luna d i ferem ent re s i pe lo teste de Tukey a 5% de probabi l idade
5.2.6 Índice de Fragmentação Miofibrilar (IFM) dos Lombos
Suínos
O método de medida de IFM tem sido amplamente adotado para a
degradação do disco Z no músculo post-mortem (KRIESE, 2005). O IFM
é uma medida da média do comprimento das miofibri las e é
signif icativamente relacionada com a maciez, miofibrilas mais curtas
resultam em IFM mais elevado, e assim uma maciez maior (MOLLER et
al. , 1973). O aumento no valor de IFM é devido à proteólise das
proteínas miofibri lares do músculo post-mortem.
Os resultados do índice de fragmentação miofibri lar para o
músculo Longissimus dorsi de suínos classif icados como PSE, DFD e
Normal estão apresentados na Tabela 7. Constata-se que o valor de
IFM do lombo Normal não diferiu entre o lombo PSE ou DFD. Porém o
IFM do lombo PSE e DFD diferiram entre si, com valor maior para o
DFD (83,21).
O maior valor de IFM das carnes DFD, provavelmente foi devido
ao maior valor de pH observado nestas amostras (pH= 5,96, Tabela 2 ).
As calpaínas que atuam sobre certas proteínas do disco Z tais como
Lombos suínos FC (N) amostra in natura FC (N) amostra cozidaPSE (n=12) 16,31ª ± 2,71 36,11ª ±9,06 DFD (n=12) 14,73ª ± 4,44 48,41ª±13,61 NORMAL (n=12) 19,49ª ± 6,86 45,76ª ±13,14

74
desmina, f i lamina, nebulina, e em menor extensão a t i tana requerem
íons cálcio (Ca²+) e apresentam pH ótimo de 6,6-6,8 (SHIMOKOMAKI et
al. , 2006). As carnes DFD apresentaram pH mais próximo do valor
ótimo da atividade da calpaína que favoreceu a degradação das
miofibri las e o amaciamento. Resultados semelhantes foram descritos
por Toldra e Flores (2000) que descreveram maior atividade da
calpaína em lombos suínos DFD.
TABELA 7 - Índice de Fragmentação Miofibri lar (IFM) dos lombos suínos PSE, DFD e Normal
Lombos suínos IFM PSE (n=12) DFD (n=12)
76,04b ±4,63 83,21ª ±7,47
NORMAL (n=12) 79,75ªb ±13,14 Médias seguidas de let ras d i ferentes na mesma co luna d i ferem ent re s i pe lo teste de Tukey a 5% de probabi l idade
5.2.7 Lipídios Totais dos Lombos Suínos
Os l ipídios totais para o músculo Longissimus dorsi de suínos
classif icados como PSE, DFD e Normal estão apresentados na Tabela
8. O teor de l ipídios totais do músculo Longissimus dorsi de suínos
PSE foram signif icativamente diferentes (p ≤ 0,05) dos lombos DFD e
ambos não diferiram do lombo Normal. Destaca-se que os lombos PSE
apresentaram maior teor de l ipídios totais, ou seja, de 4,23%.
Bragagnolo e Rodriguez-Amaya (2002) descreveram que os cortes

75
suínos são 20 a 40% mais magros do que os tradicionais e quando
retira-se a gordura externa são 40% mais magros.
Sinclair e O’dea (1987) descreveram que os novos cortes de
carne suína apresentaram de 2,3 a 3,2 % de l ipídios totais e os cortes
tradicionais de 3,5 a 6,0%. Assim, verif ica-se que os músculos
Longissimus dorsi PSE apresentaram teor de l ipídios totais similares
aos cortes tradicionais, enquanto que o lombo Normal apresentou teor
de l ipídios totais similares aos novos cortes de carne suína. Entretanto,
o teor de l ipídios totais dos lombos com DFD foi inferior aos cortes de
carne suína descrita por Sinclair e O’dea (1987).
TABELA 8 - Lipídios totais dos lombos suínos PSE, DFD e Normal Lombos suínos Lipídios totais(%) PSE (n=12) DFD (n=12)
4,23ª ± 2,01 1,58b ± 0,21
NORMAL (n=12) 3,27ªb ± 0,98 Médias seguidas de le t ras d i ferentes na mesma co luna d i ferem ent re s i pe lo teste de Tukey a 5% de probabi l idade
5.2.8 Oxidação Lipídica dos Lombos Suínos
A medida da oxidação l ipídica foi avaliada por meio de
quantif icação das substâncias reativas ao ácido t iobarbitúrico (TBARS).
Este método apresentou 85,5% de recuperação, indicando uma boa
exatidão. A oxidação l ipídica do músculo Longissimus dorsi de suínos

76
classif icados como PSE, Normal e DFD expressos como mg TBARS/kg
amostra estão apresentados na Tabela 9.
Observa-se que os lombos PSE apresentaram diferença
signif icativa (p ≤ 0,05) na oxidação l ipídica em relação aos lombos DFD
e Normal. E a oxidação l ipídica não diferiu signif icativamente entre os
lombos DFD e Normal.
Os lombos PSE oxidaram 42% a mais do que os lombos normais
e DFD. Uma maior oxidação para os lombos suínos PSE foi esperado
devido o maior teor de l ipídios totais destas amostras (Tabela 8). A
maior oxidação da carne PSE pode ser explicada, possivelmente pelo
fator do estresse que provocou a ativação da enzima fosfolipase A2 e a
sua elevada atividade seria o fator iniciante do surgimento de sintomas
característicos das carnes PSE, ou seja, provenientes de animais que
sofreram estresse térmico no pré-abate (SOARES et al.,2003).
As carnes de suínos susceptíveis a HM, também apresentaram-se
mais oxidadas com aumento no valor de TBARS e maior produção de
pentano e dieno conjugados do que as carnes de suínos resistentes(
DUTHIE et al., 1992 e NÜRNBERG et al., 2002)
TABELA 9 - Oxidação l ipídica dos lombos suínos PSE, DFD e Normal
Lombos suínos
Oxidação lipídica mg TBARS/kg amostra
PSE (n=12) DFD (n=12)
0,050ª ± 0,028 0,019 b ± 0,005
NORMAL (n=12) 0,023 b ± 0,010 Médias seguidas de le t ras d i ferentes na mesma co luna d i ferem ent re s i pe lo teste de Tukey a 5% de probabi l idade

77
O músculo Longissimus dorsi PSE apresentou menor razão a*/b*
(Tabela 3), indicando maior proporção de metamioglobina. Segundo
Rhee et al . (1996) existe uma relação entre a oxidação do pigmento,
com interdependência entre elas, onde a oxidação l ipídica pode
catalisar a oxidação de pigmento e vice-versa. A maior oxidação
l ipídica dos lombos PSE provavelmente conduziu uma oxidação de
mioglobina com maior produção de metamioglobina.
5.2.9 Composição de Ácidos Graxos dos Lombos Suínos
O interesse pela composição dos ácidos graxos da carne suína
PSE, DFD e Normal detém-se principalmente pela verif icação da
concentração dos ácidos graxos polinsaturados n-6, na qual faz parte o
ácido araquidônico. A enzima fosfolipase A2 que apresenta maior
atividade nesses lombos e atuou sobre os fosfolipídios das membranas
celulares, l iberou o ácido araquidônico e l isofosfolipídios (MURAKAMI e
KUDO, 2002). Os resultados da composição de ácidos graxos totais
para o músculo Longissimus dorsi de suínos classif icados como PSE,
DFD e Normal estão apresentados na Tabela 10.
Não foi encontrada diferença signif icativa no teor de todos os
ácidos graxos dos lombos PSE, DFD e Normal. Resultados similares
foram descritos por Tor et al. (2001) com animais portadores de
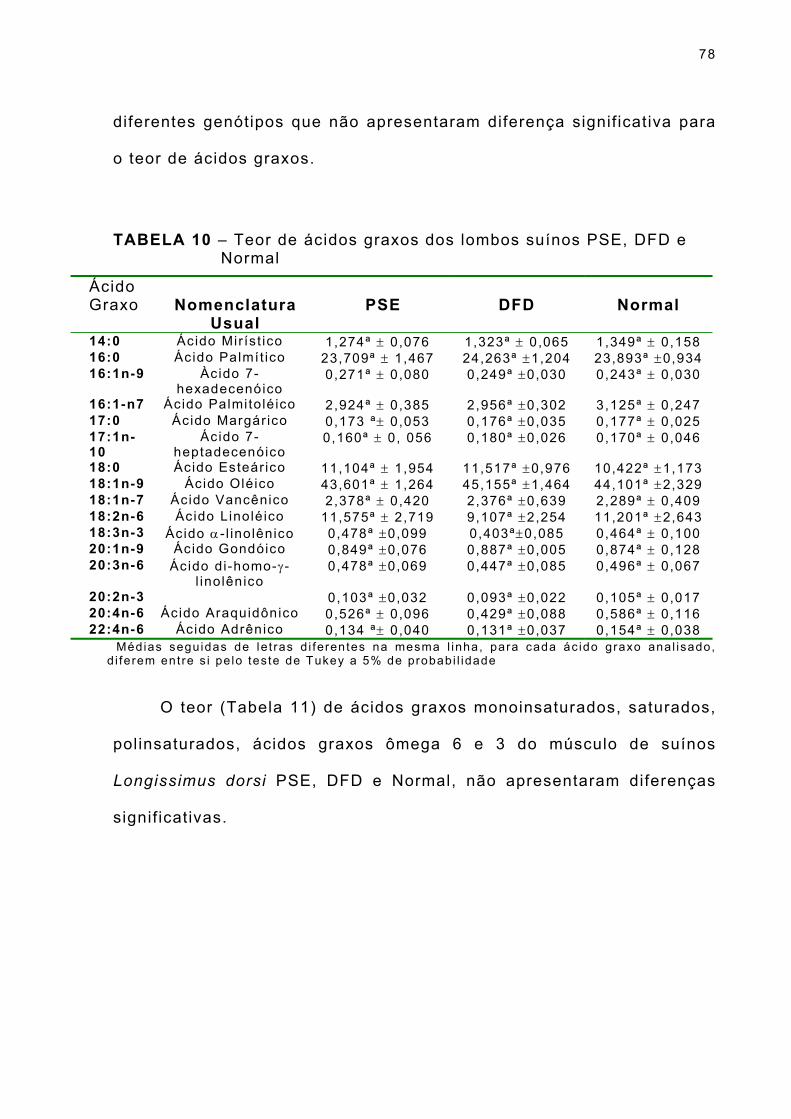
78
diferentes genótipos que não apresentaram diferença signif icativa para
o teor de ácidos graxos.
TABELA 10 – Teor de ácidos graxos dos lombos suínos PSE, DFD e Normal
Ácido Graxo
Nomenclatura
Usual
PSE
DFD
Normal
14:0 Ácido Mir íst ico 1,274ª ± 0,076 1,323ª ± 0,065 1,349ª ± 0,158 16:0 Ácido Palmít ico 23,709ª ± 1,467 24,263ª ±1,204 23,893ª ±0,934 16:1n-9 Àcido 7-
hexadecenóico 0,271ª ± 0,080 0,249ª ±0,030 0,243ª ± 0,030
16:1-n7 Ácido Palmito lé ico 2,924ª ± 0,385 2,956ª ±0,302 3,125ª ± 0,247 17:0 Ácido Margár ico 0,173 ª± 0,053 0,176ª ±0,035 0,177ª ± 0,025 17:1n-10
Ácido 7-heptadecenóico
0,160ª ± 0, 056 0,180ª ±0,026 0,170ª ± 0,046
18:0 Ácido Esteár ico 11,104ª ± 1,954 11,517ª ±0,976 10,422ª ±1,173 18:1n-9 Ácido Oléico 43,601ª ± 1,264 45,155ª ±1,464 44,101ª ±2,329 18:1n-7 Ácido Vancênico 2,378ª ± 0,420 2,376ª ±0,639 2,289ª ± 0,409 18:2n-6 Ácido Linolé ico 11,575ª ± 2,719 9,107ª ±2,254 11,201ª ±2,643 18:3n-3 Ácido α- l inolênico 0,478ª ±0,099 0,403ª±0,085 0,464ª ± 0,100 20:1n-9 Ácido Gondóico 0,849ª ±0,076 0,887ª ±0,005 0,874ª ± 0,128 20:3n-6 Ácido di-homo-γ-
l inolênico 0,478ª ±0,069 0,447ª ±0,085 0,496ª ± 0,067
20:2n-3 0,103ª ±0,032 0,093ª ±0,022 0,105ª ± 0,017 20:4n-6 Ácido Araquidônico 0,526ª ± 0,096 0,429ª ±0,088 0,586ª ± 0,116 22:4n-6 Ácido Adrênico 0,134 ª± 0,040 0,131ª ±0,037 0,154ª ± 0,038
Médias seguidas de le t ras d i ferentes na mesma l inha, para cada ác ido graxo anal isado, d i ferem ent re s i pe lo teste de Tukey a 5% de probabi l idade
O teor (Tabela 11) de ácidos graxos monoinsaturados, saturados,
polinsaturados, ácidos graxos ômega 6 e 3 do músculo de suínos
Longissimus dorsi PSE, DFD e Normal, não apresentaram diferenças
signif icativas.

79
TABELA 11– Teor de ácidos graxos monoinsaturados, saturados, polinsaturados, ácidos graxos ômega 6 e 3 dos lombos suínos PSE, DFD e Normal
PSE DFD Normal AGS* 36,701ª ± 3,313 37,279ª ±2,142 35,842ª ± 2,151
AGMI** 50,184ª ±1,184 51,803ª ±1,816 50,805ª ± 2,378AGPI*** 13,555ª ± 3,122 10,918ª ±2,116 13,357ª ±2,921
n6 12,703ª ± 2,984 10,187ª ±2,501 12,435ª ± 2,786n3 0,683ª ± 0,175 0,539ª ±0,132 0,734ª ± 0,278
AGPI/AGS 0,382ª ± 0,114 0,296ª ±0,084 0,376ª ± 0,094 n6/n3 18,560± 0,708 18,887ª±0,062 16,941± 1,199
Médias seguidas de le t ras d i ferentes na mesma l inha, para cada ác ido graxo anal isado, d i ferem ent re s i pe lo teste de Tukey a 5% de probabi l idade *AGS - ác idos graxos saturados = 14:0 + 16:0 + 17:0 +18:0 **AGMI – ác idos graxos monoinsaturados = 16:1n-9 + 16:1-n7 +17:1n-10 + 18:1n-9 + 18:1n-7 + 20:1n-9 ***AGPI – ác idos graxos po l insaturados = 18:2n-6 + 18:3n-3 + 20:3n-6 + 20:2n-3 + 20:4n-6 + 22:4n-6
A relação entre ômega 6/ ômega 3 para os músculos PSE, DFD
e Normal foi ao redor de 18,13. As dietas ocidentais, segundo
Simopoulus (1994) apresentaram uma relação de consumo de ácidos
graxos ômega 6/ ômega 3 de 20-30:1. Entretanto, segundo Wood et al.
(2003) a relação ideal do consumo desses ácidos graxos ômega 6/
ômega 3 deve ser igual ou menor que 4:1. Os resultados obtidos para a
razão ômega 6/ ômega 3 foram inferiores aos descritos por Bragagnolo
(2001) de 21.
5.2.10 Microscopia Eletrônica dos Lombos Suínos
As f iguras 14 e 15 mostram uma eletromicrografia de seção
longitudinal de f ibras dos lombos suínos Normal e PSE,
respectivamente. Na figura 14 observou uma ultra-estrutura típica do

80
músculo Longissimus dorsi normal de suíno com disco Z definido e
presença de bandas A escuras constituídas pelos f i lamentos grossos,
bandas I claras, f i lamentos finos e l inha M.
Com relação ao lombo suíno DFD na ultra-estrutura foi observado
edema (dados não apresentados).
Na figura 15 observam-se modificações na estrutura do
sarcômero como despolimerização das proteínas do sarcômero (seta 1)
devido às condições fisiológicas do animal durante o estresse térmico a
uma temperatura elevada e baixo valor do pH provocado pelo aumento
da concentração dos ions Ca²+. Nessas condições, houve uma maior
exposição das proteínas disponibil izando-as para a ação das proteases
que provocaram uma desfragmentação do disco Z (seta 2) com o inicio
do surgimento de um colapso na sua ultra-estrutura (seta 3). As
proteases, como as calpaínas degradam as proteínas desmina,
f i lamina, nebulina, troponina T e t itana, sendo que a maioria destas
proteínas estão próximas do disco Z. Pode ser observado até
visualmente que houve encolhimento do sarcômero com diminuição da
banda I. Em uma comparação com as alterações nos f i lés PSE de
frango (Pectoralis major) e de acordo com Guarnieri et al. (2004) que
observou uma hipercontração do sarcômero, o aparecimento de lacunas
ao longo do sarcômero e em alguns casos espessamento e
desfragmentação do disco Z. Aparentemente, as alterações na
ultraestrutura dos frangos foram mais drásticas do que nos suínos.
Uma possível explicação para a não ocorrência da hipercontração do
sarcômero nos suínos está relacionada com a diferença entre o t ipo e

81
tamanho das fibras de ambos os animais. Em lombos suínos ocorre
uma relativa maior proporção das f ibras vermelhas que possuem uma
velocidade de contração mais lenta (JUDGE et al., 1989).
O fenômeno PSE em frangos e suínos ocorreram devido à rápida
glicólise post mortem que levaram a dimiuição do pH enquanto a
carcaça ainda quente promoveu a desnaturação das proteínas. Assim a
Figura 16 apresenta o provável mecanismo para a relação entre
estresse pré-abate, alterações bioquímicas e na ultra-estrutura no
desenvolvimento da carne suína PSE, confirmando a ocorrência do
fenômeno PSE em suínos.
FIGURA 14 – Eletromicrografia de uma seção longitudinal do lombo suíno Normal. Aumento 12000 vezes. M = Linha M; Z = disco Z; A = Banda A; I = Banda I.

82
FIGURA 15 – Eletromicrografia de uma seção longitudinal do lombo suíno PSE. Aumento 6000 vezes.

83
Estresse
Abate
no músculo
Glicólise Anaeróbica
Ácido Lático
pH
MetabolismoAcelerado
DesnaturaçãoProtéica
Ação das Proteases
Colapso daEstrutura
1 2 3
FIGURA 16 - Provável mecanismo para a relação entre estresse pré-abate, alterações bioquímicas e na ultra-estrutura no desenvolvimento da carne suína PSE. 1-Despolimerização das proteínas do sarcômero. 2-Atividade das proteases na região do disco Z. 3-Colapso do sarcômero.

84
6 CONCLUSÕES
A incidência de PSE, DFD e Normal de lombos suínos, músculo
Longissimus dorsi, em uma linha comercial de abate foi de 22,83%,
1,06% e 76,11% respectivamente.
A correlação entre os valores de pH e cor dos lombos suínos
PSE, DFD e Normal foi negativa.
A razão entre IMP e ATP, indicada pelo valor de R, foi
signif icativamente maior (p≤0,05) para os lombos PSE em relação ao
DFD, não diferindo dos normais, com predominância de IMP.
A maciez dos lombos PSE, DFD e Normal não diferiram
signif icativamente entre si pela medida de força de cisalhamento e o
índice de fragmentação miofibri lar foi signif icativamente maior (p≤0,05)
para o lombo DFD.
A oxidação l ipídica dos lombos suínos PSE, foi 42% maior em
relação ao Normal e DFD.
O perfi l de ácidos graxos dos lombos suínos PSE, DFD e Normal
não apresentou diferença signif icativa entre si.
Ocorreu na ultra-estrutura dos lombos suínos PSE a
desorganização do sarcômero com despolimerização das proteínas do
disco Z, ação das proteases e o colapso do sarcômero.

85
7 REFERÊNCIAS BIBLIOGRÁFICAS ABCS – Associação Brasileira dos Criadores de Suínos. Disponível em: http://www.abcs.com.br. Acesso em 19 de junho de 2005.
ABIPECS - Associação Brasileira da Indústria Produtora e Exportadora da Carne Suína. Disponível em: http://www.abipecs.com.br. Acesso em 19 de março de 2007.
ANDRADE JUNIOR, R.; NICOLAIEWSKY, S.; OURIQUE, J. M. R.; CULAU, P. O. V.; BRESSAN, M. C. Análise de alguns fatores determinantes da qualidade da carne suína. Efeito da distância granja-frigoríf ico, tempo de descanso, sexo e peso vivo. In: Congresso Brasileiro de Medicina Veterinária, 22, 1993, Curit iba. Anais.... p.216.
ANGERAMI, C. N. Influência do genótipo, sexo, peso de abate na composição da carcaça e nas características de qualidade da carne suína. 2004. Dissertação (Mestrado)- Esalq. Piracicaba.
ARAÚJO, J. M. A. Química dos alimentos: teoria e prática. 3ed. Viçosa: UFV, 2004.
BARBUT, S. Problem of pale soft exsudative meat in broiler chickens. Britsh Poultry Science, Edinburgh, v. 38, n. 1, p. 355-358, 1997.
BARTON – GADE, P. The effect of pre – slaugther handlig on meat quality of pigs. In: CRANWELL P.D. (Ed.) Manupulating pig production. Melbourne: Frankland, 1997. cap. 6, p. 100 – 123.
BARTON-GADE, P. A. Developments in the pre-slaughter treatment of slaugther animals. In: European Meeting Meat Research Workers, Madri, p. 31-39, 1985.
BARTON–GADE, P. Danish experience in meat quality improvement. In: WORLD COGRESS ON GENETICS APPLIED TO LIVESTOCK PRODUCTION, 4., Edinburgh, 1990. p 511 – 520.
BATLLE, N.; ARISTOY, M.C. e TOLDRÁ, F. Early post mortem detection of exudative pork meat based on nucleotide content. Journal of Food Science, v.65, n.3, 2000.
BENDALL, J. R.; SWATLAND, H. J. A review of the relationships of pH with physical aspects of pork quality. Meat Science, Barking, v. 24, p. 85-126, 1988.
BERTOL, T.M. Estresse pré-abate: conseqüências para a sobrevivência e a qualidade da carne em suínos, 2004. Disponível em <http://www.cnpsa.embrapa.br> . Acesso em: 23/07/2005.

86
BLIGH, E. G.; DYER, W. J. A rapid method of total l ipid extration and purif ication. Canadian Journal of Biochemistry Physiology, v.31, p.911-917, 1959.
BOWKER, B. C.; WYNVEEN, E. J.; GRANT, A. L.; GERRARD, D. E. Effects of electr ical stimulation on early post mortem muscle pH and temperature declines in pigs from different genetic l ine and halothane genotypes. Meat science, v. 53, p. 125-133, 1999.
BRAGAGNOLO, N. Aspectos comparativos entre carnes segundo a composição de ácidos graxos e teor de colesterol. In: 2ª Conferência Internacional Virtual sobre Qualidade de Carne Suína, Concórdia - SC, p. 393-402, 2001.
BRAGAGNOLO, N.; RODRIGUEZ-AMAYA, D. B. Teores de colesterol, l ipídios totais e ácidos graxos em cortes de carne suína. Ciência e Tecnologia de Alimentos, v.22, n. 1. Campinas, 2002.
BRASIL, MAPA – Ministério da Agricultura, Pecuária e Abastecimento. Secretaria de Defesa Agropecuária. Departamento de Inspeção de Produtos de Origem Animal. Normas Técnicas de Instalações e Equipamentos para Abate e Industrial ização de Suínos. Portaria nº 711 de 01 de novembro de 1995. 251 p.
BRIDI, A. M.; RUBENSAM, J. M.; NICOLAIEWSKY, S.; LOPES, R. F. F.; LOBATO, J. F. P. Efeito do genótipo halotano e de diferentes sistemasde produção na qualidade da carne suína. Revista Brasileira de Zootecnia, v.32, n.6, p. 1362-1370,2003.
CANNON, J. E.; MORGAN, J. B.; SCHIMDT, G. R.; TATUM, J. D.; SOFOS, J. N.; SMITH, G. C.; DELMORE, R, J.; WILLIAMS, S. N. Growth and fresh meat quality characteristics of pigs supplemented with vitamin E. Journal of Animal Science, v. 74, p. 98-105, 1996.
CASSENS, R. G. Historical perspectives and current aspects of pork meat quality in the USA. Food Chemistry, v. 69, p. 357-363, 2000.
CHANNON, H. A.; PAYNE, A. M.; WARNER, R. D. Halothane genotype, pre-slaughter handling and stunning method all influence pork quality. Meat Science, v. 56, p. 291-299, 2000.
CHEAH, K. S.; CHEAH, A. M. Mitocondrial calcium transport and calcium-activated phospholipase in porcine malignant hyperthermia. Biochimica et Biophysica Acta, Amesterdam, v. 634, p. 70-84, 1981a.
CHEAH, K. S.; CHEAH, A. M. Skeletal muscle mitochondrial phospholipase A2 and the interaction of mitochondrial and sarcoplasmic reticulum in porcine malignant hyperthermia. Biochimica et Biophysica Acta, Amsterdam, v. 638, p40-49, 1981b.
CHEAH, K. S.; CHEAH, A. M.; CROSLAND, A. R.; CASEY, J. C.; WEBB, A. J. Relathionship between Ca²+ release, sarcoplasmic Ca²+,

87
glycolysis and meat quality in halothane-sensit ive and halothane-insensit ive pigs. Meat Science, Barking, v. 10, n. 2, p. 117-130, 1984.
CHEAH, K. S.; CHEAH, A. M.; KRAUSGRILL, P. I. Variat ions in meat quality in l ive halothane heterozygotes identif ied by biopsy samples of m. Longissimus dorsi . Meat Science, Barking. V. 39, p. 293-300, 1995.
CHEAH, K. S.; CHEAH, A. M.; LAHUCKY, R. ; MOSTO, J.; KOVAC, L. Prediction of meat quality in l ive pigs using stress-susceptible and stress resistant animals. Meat Science, Barking, v.34, p. 179-189,1993.
CORNFORTH, D. Color –its basis and importance. In: PEARSON, A.M.; DUTSON, T.R. Quality attributes and their measurement in meat poultry and fish products. Glasgow: Blackie Academic & Professional, 1994. p.34-78.
CRACKEL, R. L. et al. Some further observations on the TBA test as an index of l ipid oxidation in meats. Food Chemistry, v.28, p.187-196, 1988.
CULAU, P. O. V.; LÓPEZ, J.; RUBENSAM, J. M.; LOPES, R. F. F.; NICOLAIEWSKY, S. Influência do gene halotano sobre a qualidade da carne. Revista Brasileira de Zootecnia, v. 31, p. 1-12. Viçosa, 2002.
CULAU, P. O. V.; OURIQUE, J. M. R.; NICOLAIEWSKY, S.; BRESSAN, M. C. Incidence of PSE in commercial pig carcasses in Rio Grande do Sul state, Brazil. Boletim Técnico da Associação Sul Brasileira das Indústrias de Produtores de Suínos, Porto alegre, 1994.
CULLER, R. D., PARRISH Jr. F.C., SMITH, G. C., CROSS. Relationship of myofibri l fragmentation index to certain chemical, physical, and sensory characteristics of bovine Longissimus muscle. Journal of Food Science, 1978.
DUTHIE, G.G.; WAHLE, K.; HARRIS, C.I.; ARTHUR, J.R. Lipid peroxidation, antioxidant concentrations and fatty acid contents of muscle t issue form malignant hyperthermia-susceptible swine. Archives of Biochemistry and Biophysics, v.296, p.592-296, 1992
EIKELENBOOM, G. & BOLINK, A. H. effects os feed withdrawl before delivery on pork quality and carcass yield. Meat Science . V. 29, p. 25-30,1991.
EIKELENBOOM, G.; BOLINK, A. H. & SYBESMA, W. effect du jeune avant le chargement sur la qualité et le rendement de cacasse des poreso. Qualité de la Viande du Pore. France, 1989.
ELAM, T. E. The world pork industry-rapid change and re-structurinr implications for a global pork market. Elanco Animal halth Iternational Symposium. Indianápolis, Indiana, EUA, 1997.

88
EMBRAPA. Curso Conhecendo a Carne que Você Consome. Campo Grande. Qualidade de Carne Bovina ; Embrapa Gado de Corte, 1999. 25p.
FAUCITANO, L. Causes of skin damage to pig carcasses. Canadian Journal of Animal Science, v. 81, p. 39 – 45, 2001.
FELÍCIO, P. E. O ABC do PSE/DFD. Aliment. Tecnol ., São Paulo, v.2, n. 10, p. 54-57, 1986.
FERNANDES, T. H.; SMITH, W. C.; ARMSTRONG, D. G. The administration of sugar solutions to pigs immediately prior to sleghter. Effect on carcass yield ans some muscle and l iver characteristics. Animal Production , v. 29, p. 213-221, 1979.
FISCHER, K. Quall i tatsabwerchungen bei r indfleisch. Kulumbacher Reihe Band , v. 7, p. 235-268, 1987.
FISHER, P.; MELLETT, F. D.; HOFFMAN, L.C. Halothane genotype and pork quality. 1 Carcass and meat quality characteistics of three halothane genotyps. Meat Science, v. 54, p. 9 – 105, 2000.
FLETCHER, D. L. Broiler breast meat color variation, pH and texture. Poultry Science, v. 78, p. 1323-1327, 1999.
FUJII, I.; OTSU, K.; ZORZATO, F.; DELEON, S.; KHANNA, V. K.; WEILAR, J. E.; O’BRIEN, P. J.; MAC LENNAN, D. H. Identif ication of mutation in porcine ryanodine receptor associated with malignant hyperthermia. Science , Washington, v. 253, p. 448-451, 1991.
GIANNINI, G.; CONTI, A.; MAMMARELLA, S.; SCROBOGNA, M.; SORRENTINO, V. The ryanodine receptor, calcium.I. calcium channel genes are widely and differential ly expressed in murine brain and peripheral t issues. Journal of Cell Biological, New York, v.128, p. 893-904, 1995.
GREGORY, N. G.; GRANDIN, T. Animal welfare and meat science. London. CAB International, p. 286,1998.
GUARNIERI, P. D.; SOARES, A. L.; OLIVO, R.; SCHNEIDER, J.; MACEDO, R. M. G.; IDA, E. I.; SHIMOKOMAKI, M. Preslaughter handling with water shower spray inhibits PSE (Pale, Soft, Exudative) broiler breast meat in a commercial plant. Biochemical and ultrastructural observations. Journal of Food Biochemistry, v.28, n.4, 2004, p.269-277.
HONIKEL, K. O.; KIM, C. J. Causes of the development of PSE pork. Fleischuirts, v. 66, p. 349-353, 1986.
HONIKEL, K.O. & FISCHER, C.A. A rapid method for the detection of PSE and DFD porcine muscle. Journal of food Science, v. 42, n. 7, p. 1663-1676, 1977.

89
INTERNATIONAL ORGANIZATION FOR STANDARDIZATION (ISO), Method ISSO 5509: Animal and vegetable fats and oils- preparation of methyl ésteres of fatty acids. 1978.
JENSEN, P.; BARTON-GADE, P. A. Performance and carcass caracteristics of pigs with know genotypes for halothane susceptibi l i ty. In: COMISSION ON ANIMAL MANEGEMENT AND HEALTH AND COMISSION OF PIG PRODUCTION, Wageningen, 1985. Procedings . Wageningen, 1985, p. 80-93.
JIANG, S.T. Contribution of muscle proteinases to meat tenderization. Proocedings of the National Science Councill, ROC. Part B: Life of Sciences, vol. 22, n. 3, p. 97-107, 1998.
JOO, T. S.; KAUFFMAN, G. R.; WARNER, D. R. ; BORGGAARD, C.; STEVENSON- BARRY, M. J.; LEE, S.; PARK, B. G.; KIM, C. B. Objectively predicting ult imate quality of post-rigor pork musculature: I. Init ial comparison of techniques. Asian Australian Journal of Animal sciences, Suwon, v. 13, n. 1, p. 68-76, 2000.
JUDGE, M. D.; ABERLE, E. D.; FORRESTE, J. C.; HEDRICK, H. B.; MERKEL, R. A. Principles of Meat Science. 2ed. Iowa: Kendall/ Hunt Plublishing Company, 1989. 351p.
KAUFFMAN, G. R.; CASSENS, R. G.; SCHERER, A.; MEEKE, D. L. Variations in pork quality. National Pork Producers Council Publication, p. 56, 1992.
KHÖLER, R. G.; FREITAS, R. J. S. Qualidade da carne suína após dois tempos de descanso no fr igoríf ico. Archives of Veterinary Science, v. 10, n. 1, p. 89-94, 2005.
KRIESE, P. R. Influência da Maturação Na Maciez de Peito de Frango Realizada Em Condições de Processamento Industrial. 2005 Dissertação (Mestrado em Ciência de Alimentos) – Universidade Estadual de Londrina, Londrina, 59p.
LARA, J. A. F.; SOARES, A . L.; IDA, E. I.; OLIVO, R.; SHIMOKOMAKI, M. Carnes PSE em suínos e aves. Uma abordagem comparativa. Boletim SBCTA . V. 37, p. 36-39, 2003.
LEACH, L. M.; ELLIS, M.; SUTTON, D. S. MCKKEIT, F. K.; WILSON, E. R. The growth performance, carcass characteristics, and meat quality of halothane carrier and negative pigs. Journal of Animal Science , v.74, p.934-943, 1996.
LUDTKE, C.B. ; NOGUEIRA, C.E.W..; COSTA, C.S. ; BESSA, L.R. ; ZOCCHE, F. * ; CARRO, S.B.T.; ANTUNEZ, H.C.S.; KUHN, C.R.; MONKS, V.; DORNELLES, G.J.D. Bem-estar no manejo pré-bate e na qualidade da carne eqüina. Disponível em: http://www.multitel.com.uy/congresoselis/suctal/trabajos/147.doc. Acesso em 22/07/2004.

90
MAC LENNAN, D. H.; PHILLIPS, M. S. Malignant hyperthermia. Science, Washington, v. 256, p. 789-794, 1992.
MICKELSON, J. R.; LOUIS, C. F. Malignant hyperthermia: excitation-contraction coupling, Ca²+ release channel, and cell Ca²+ regulation defects. Physiology Review, Balt imore, v. 76, n. 2, p. 537-592, 1996.
MITCHELL, G.; HEFFRON, J. J. A. Porcine stress syndromes. Advances in Food Researche, New York, V. 28, p. 167-230, 1982.
MITCHELL, M.A. Muscle Abnormalit ies: pathophysiological machanisms (Capitulo 3). Disponivel em< http://www.cabipublishing.org/bookshop/redingroom1085/1992374/12374ch03.pdf>. Acesso em 30 de julho de 2005.
MOLLER, A. J., VESTERGAARD, T., WISMER-PEDERSEN j. Myofibri l fragmentation in bovine Longissimus dorsi as an index of tenderness. Journal of Food Science, 38:824, 1973.
MONIN,G.; LARZUL,C.; ROY,P.L.; CULIOLI,J. Effects of the halothane genotype and slaughter weight on texture of pork. Journal Animal Science, v.77, n.2, p. 408-415, 1999
MORRISSEY, P.A.; SHEEHY, P.J.A.; GALVIN, K.; KERRY, J.; BUCKLEY, D.J. Lipid stabil i ty in meat and meat products. Meat Science, v.49, p.73S-86S, 1998.
MURAKAMI, M.; KUDO, I. Phospholipase A2. Journal of Biochemistry, v.131, p.285-292, 2002.
MURRAY, A. C.; JONES, S. D. M.; SATHER,P. The effect of preslaughter feedrestriction and genotype for stress susceptibi l i ty on pork lean quality and composit ion. Canadian Journal of Animal Science, v.49, p.83, 1989.
MURRAY, A.C. and JOHNSON, C.P. 1998. Impact of the halothane gene on muscle quality and pre-slaughter deaths in Western Canadian pigs. Can. Journal Animal Science.78:543-548.
NIELSEN, N. J. The influence of pre-slaugther treatment on meat quality in pigs. Danish Meat Research Institute, 14p., 1977.
NÜRNBERG, K.; KÜCHENMEISTER, U.; JAKSTADT, M.; ENDER, K.; KUHN, G.; NÜRNBERG, G,; GRUNE, T. Compositional changes in muscle of malignant hyperthermia-susceptible pigs due to postmortem alterations in l ipid peroxidation and protein peroxidation. Journal of Food Compositions and Analysis, v. 15, p.283-292, 2002.
ODA, S. H. I.; SCHENEIDER, J.; SOARES, A. L.; BARBOSA, D. M. L; IDA, E. I.; OLIVO, R.; SHIMOKOMAKI, M.Detecção de cor em fi lés de peito de frango. Revista Nacional da Carne , v. 28, p. 30-34, 2003.

91
OLIVO, R.; SOARES, A. L.; IDA, E. I.; SHIMOKOMAKI, M. Dietary vitamin E inhibits poultry PSE and improves meat functional properties. Journal of Food biochemistry, Trumbull, v. 25, n. 4, p. 271-283, 2001.
OSAWA, C. C.; FELÍCIO, P. E.; GONÇALVES, L. A. G. Teste de TBA aplicado a carnes e derivados: métodos tradicionais, modif icados e alternativos. Química Nova, v.28, n.4. São Paulo, 2005.
OURIQUE, M. J.; NICOLAIEWSKY, S. Características físico-químicas e organolépticas e suas relações na avaliação da carne suína. Revista Sociedade Brasileira de zootecnia, Viçosa, v. 19, n. 2, p. 118-125, 1990.
PARDI, M. C.; SANTOS, I.F.; SOUZA, E.R.; PARDI, H.S. Ciência, Higiene e Tecnologia da Carne. V. 1. Niterói: EDUFF, 1993. 586 p.
PEARSON, A. M.; YOUNG, R. B. Muscle and Meat Biochemistry. San Diego: Academic Press, 457p., 1989.
POMMIER, S. A.; HOUDE, A. Effect of the genotype for malignant hyperthermia as determined by a restrict ion endonuclease assay on the quality characteristics of commercial pork loins. Journal of Animal Science, v. 71, 420p., 1993.
PRICE, M. G. In “Advances in Structural Biology”, vol. I., ed. by Melhorta, S. K. J. AI Press, Greenwich, CT, p. 175, 1991.
RHEE, K.S.; ANDERSON, L.M.; SAMS, A.R. Lipid oxidation potential of beef, chicken and pork. Journal of Food Science, v.61, n.1, p.8-12, 1996.
ROSENVOLD, K.; ANDERSEN, H. J. Factors of signif icance for pork quality: a review. Meat Science, v.64, p. 219-237, 2003.
SAMS, A.R.; MILLS, K.A. The effect of feed withdrawal duration on the responsiveness of broiler Pectoralis to r igor mortis acceleration. Polutry Science, v.72, n.9, p.1789-1796, 1993.
SELLIER, P. Genetics of meat and carcass traits.In: Rolhschild, M.; Ruvinsky, A. The genetic of the pig.CAB Internationa l ,p. 436-510, 1998.
SENCIC, D.; ANTUNOVIC, Z.; SPERANDA, M. Meatiness and meat quality in pigs crossed with Pietrain. Stocarstvo, v.56, p.191-196, 2002.
SHEARD, P. R.; ENSER, M.; WOOD, J. D.; NUTE, G. R.; GILL, B. P.; RICHARDISON, R. I. Shelf l ife and quality of pork products with raised n-3 PUFA. Meat Science, v.55, p. 213-221, 2000.
SHIMOKOMAKI, M.; IDA, E.I.; KRIESE, P.R.; SOARES, A.L. Calpaínas e Calpastatinas. In: OLIVO, R.; SHIMOKOMAKI, M,; TERRA, N.N.;

92
FRANCO, B.G.M. Atualidades em Ciência e Tecnologia de Carnes. São Paulo: Varela, 2006. p.185-194.
SIEGEL, H. S. Stress, str ins and resistance. Brit ish Poultry Science, Edinburgh, v. 36, p. 312-329, 1995.
SILVEIRA, E. T. F. Impacto da qualidade na industrialização da carne suína. In: Conferência Internacional Sobre Ciência e Tecnologia de Produção e industrialização de Suínos, 1996, Campinas. Anais.... Campinas: ITAL, p. 99-122, 1996.
SINCLAIR, A. J.; O’DEA, R. The l ipid levels and fatty acid composit ions of the lean portions of pork, chicken and rabbit meats. Food Technologie Austrália, v. 39, p. 232-233, 1987.
SOARES, A. L., SANTOS, T. N., MARCHI, D.OBA, A.,IDA, E. I., SHIMOKOMAKI, M. Similaridade entre os sintomas da síndrome do estresse das aves, PTSS (poultry stress syndrome) e a do suíno PSS (pork stress syndrome) originando as carnes PSE (pale, soft, exudative) de frango. Revista Nacional da Carne, nº 361, março/2007.
SOARES, A., L.; IDA, E. I.; MIYAMOTO, S.; BLAZQUEZ, F. J. H.; OLIVO, R.; PINHEIRO, J.W.; SHIMOKOMAKI, M. Phospholipase A2 activity in poultry PSE, Pale, Soft, Exudative, Journal of Food Biochemistry, v. 27, n. 4, p. 309-319, 2003b.
SOARES, A.L. PSE (Pale, Soft, Exudative) em frangos: implementação de parâmetro de cor e avaliação bioquímica e estrutural do filé (Pectoralis major). 2003. Tese (Doutorado em Ciência de Alimentos) – Universidade Estadual de Londrina, Londrina, 103p.
SOARES, A.L.; MARCHI, D.F.; OBA, A.; IDA, E.I.; SHIMOKOMAKI, M. Carnes PSE (Pale, Soft, Exudative) e DFD (Dark, Firm, Dry) em frangos e rancidez oxidativa. Revista Nacional da Carne, v. 30, n.344, p.138-140, 2005.
SWARTZ, D.R.; LIM, S-S; FASSEL, T.; GREASER, M.L. Mechanism of myofibri l assembly. Annual Reciprocal Meat Conference, Proceedings, v.47, p.141-153,1994.
SWATLAND, H. J. Explaining the P in PSE. Meat Focus Int. , Wall ingford, v.2, n. 8, p. 362-367, 1993.
TARLADGIS, B. G. et al. Chemistry of the 2-thiobarbituric test for determination of oxidative rancidity in foods II . Formation of the TBA-malonaldehyde complex without acid-heat treatment. Journal of Food Science and Agriculture, v.5, p.602-604, 1964.
TERRA, N. N. Apontamentos de tecnologia de carnes. São Leopoldo. Ed UNISINOS, 216p, 1998.

93
TOLDRÁ, F.; FLORES, M. The use of muscle enzymes as predictor of pork meat quality. Food Chemistry, v.69, p.387-395, 2000.
TOR, M.; ESTANY, J. VILLALBA, D.;CUBILO, D.; TIBAU, J.; SOLER,J.; SANCHEZ,A.; NOGUEIRA, J. L. A within-breed comparison of RyR1 pig genotypes for performance, feeding behaviour, and carcass, meat and fat quality traits. Journal of Animal Breeding and Genetics, v.118, p.417-427, 2001.
WARRIS, P. D. Ante mortem handil ing of pigs.In: COLE, D. J. A.; WISEMAN, J.; VRLEY, M. A. Principles of pig science.Loughborough: Nottingham University Press, p. 425-432, 1994.
WARRIS, P. D. New developments in he preslaughter handil ing of pigs. In: Conferência Internacional Ciência e Tecnologia de Produção e industrialização de Suínos 1995, Campinas. Anais.... Campinas: ITAL, p. 81-107, 1995
WARRIS, P. D. The relationship between ph45 and drip in pig muscle. Journal of Food Technology, v. 17, p. 573-578, 1982.
WARRIS, P. D. The welfare of slaughter pigs during transport. Animal Welfare, v.7, p.365-381, 1998.
WARRIS, P. D.; BROWN, S. N. The relationships between init ial pH, reflectance and exudation in pig muscle. Meat Science , barking, v. 20, p. 65-74, 1987.
WARRIS, P.D. Nova Geração. Revista Suinocultura Industrial, n. 150, 2001.
WIRTH, F. Technologie der Verabeitung von flaisc mit abweichender Beschaffenhert.Fleischirwtschaft, v. 65, p. 998-1001, 1986.
WOOD, J. D. Effects of fatty acids on met quality: a review. Meat science, v. 63, p. 21-32, 2003.

Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas

Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo







