
Intermediary metabolism
Eva Samcová


Metabolic roles of tissues • Four major tissues play a dominant role in fuel
metabolism : liver, adipose, muscle, and brain.
• These tissues do not function in isolation.
• Communication between tissues is mediated by the nervous systém, by the availability of circulating substrates, and by variation in the levels of plasma hormones.
• The integration of energy metabolism is controlled by the actions of two peptide hormones, insulin, and glucagon (response to changing substrate levels) with catecholamines epinephrine and norepinephrine (response to neural signals).

Liver
Liver lies immediately under the diaphragm. It is supplied
with blood from below through two major vessels: the
hepatic artery (20% of blood) and the hepatic portal vein
which brings the substrates (soluble in water) absorbed
from the intestinal tract including stomach into the blood
and then directly to liver.
Pancreatic vein (insulin, glucagon)
Liver consumes 20 – 30% of total oxygen consumption
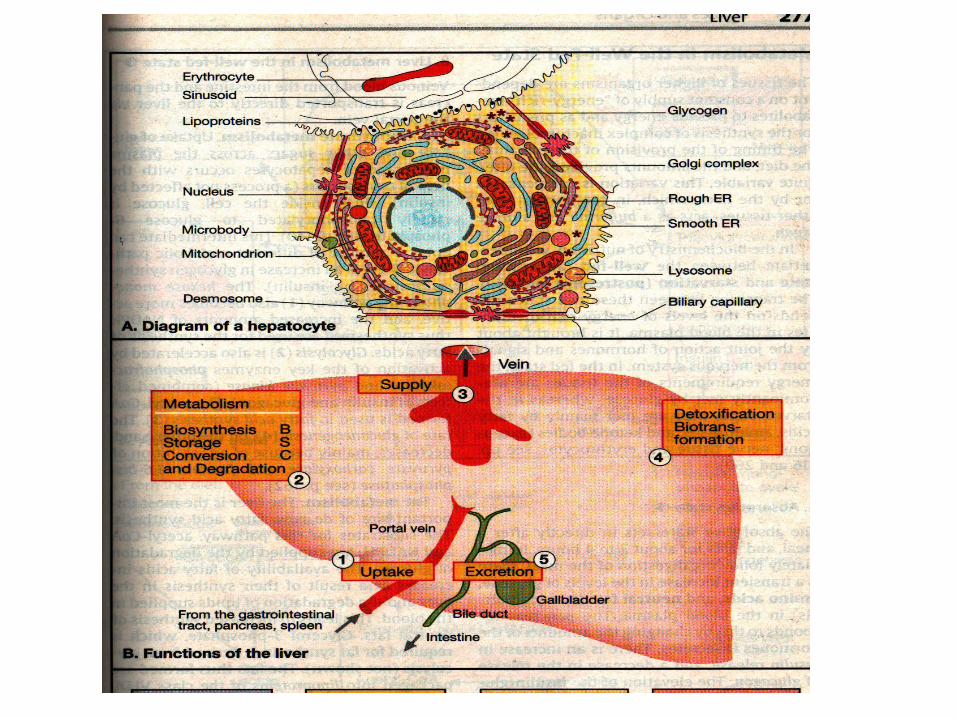

Functions of the liver
The uptake of nutrients delivered from the
digestive tract via portal vein
The synthesis, storage, interconversion and
degradation of metabolite
The regulated supply of energy-rich
intermediates
The detoxification of harmful compounds by
biotransformation
The excretion of substances with the bile;
synthesis and degradation of many blood plasma
constituents

Carbohydrate metabolism in the liver- fed
conditions
Concentration in portal vein after a meal up to 10 mmol/l – GLUT-2 –
type glucose transporter not responsive to insulin, relatively high Km
(rate and direction of movement of glucose through hepatocyte
membrane are determined by concentration inside and outside the cell)
Glucokinase (Km= 12 mmol/l) x hexokinase 0.1 mmol/L
Any increase in glucose concentration against blood conc. leads to
proportional increase in the rate of phosphorylation by glucokinase
.Likewise any decrease in glucose conc. leads to proportional decrease
in the rate of phosphorylation.
Thus liver uses glucose at significant rate only when blood glucose level
is greatly elevated.
The overall result is that when glucose conc. outside the hepatocyte
rises, glucose will be rapidly taken into cells and phosphorylated.

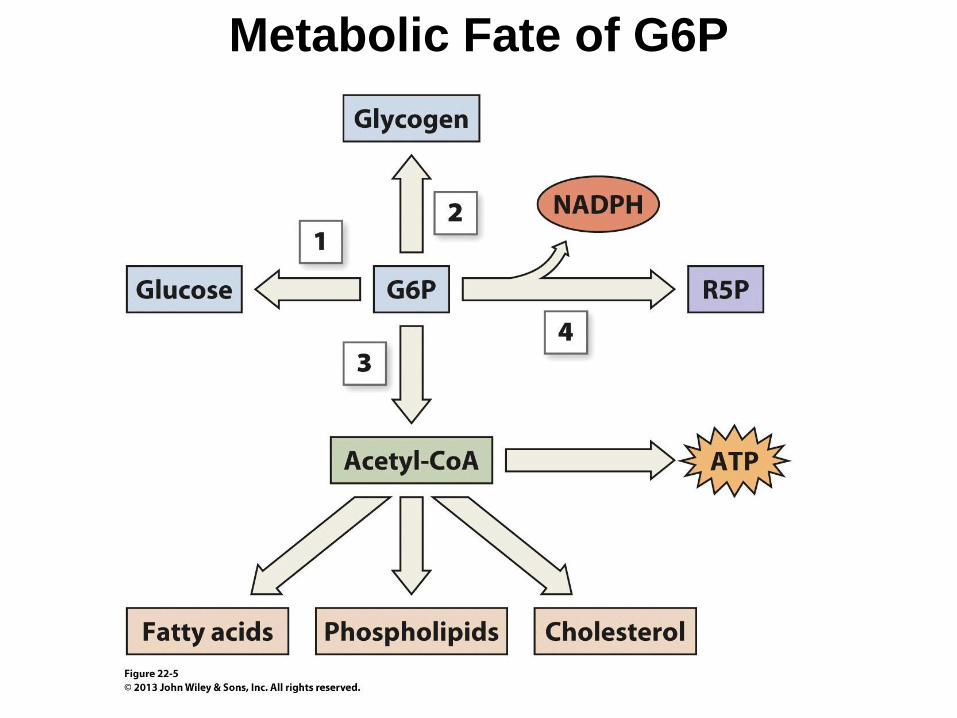
Carbohydrate metabolism in the liver-
fed conditions
The presence of high-Km glucose transporter and
high-Km glucokinase do not enable the hepatocyte
to take up unlimited quantities of glucose as G-6-P
There are specific mechanisms for stimulating the
disposal of Glu-6-P
Glycogen synthesis (activation of glycogen
synthase by insulin and glucose)
Glycolysis metabolizes glucose to pyruvate
TCA, some released after conversion to lactate.
But minor energy source for liver.
Metabolic Fate of G6P

Carbohydrate metabolism –
overnight fasted conditions
Glycogen breakdown (glycogenolysis), controlled by
reciprocal activation of glycogen phosphorylase by
glucagon, adrenalin, noradrenalin, catecholamines.
Glu-1-P produced by glycogenolysis is in equilibrium
with Glu-6-P (enzyme phosphoglucomutase).
Formation of glucose from Glu-6-P is produced by
enzyme Glu-6-phosphatase (membrane ER)
12/2/12

Carbohydrate metabolism in the liver
Synthesis of glucose – gluconeogenesis
Substrates : lactate, alanine, glycerol
Hepatic gluconeogenesis can be also stimulated by
increase in the supply of substrate from other tissue
(after physical exercise-lactate, starvation-glycerol)
and by hormones (glucagon)
Glucose paradox (gluconeogenesis after meal)
The pentose phosphate pathway – alternative fate for
Glu-6-P, conversion to five-carbons sugars (ribose-5-
P for synthesis of nucleic acids)
Formation of NADPH for reductive synthesis

Fat metabolism in the liver
The metabolism of lipids in the liver is closely
linked to metabolism of carbohydrates and
amino acids.
The pathwayof FA oxidation diverges from that
of glycerolipid synthesis when acyl-CoA enters
the mitochondrion for oxidation.
Carnitine-palmitoyl transferase-1 (CPT-1).
Activity of this enzyme is strictly regulated by
means of compound malonyl-CoA (potent
inhibitor). This role of malonyl-CoA provides a
vital link between carbohydrate and fat
metabolism.

Fat metabolism in the liver
The liver converts glucose (Glc) via Acetyl-CoA into
fatty acids (FA) - cytosol. FA and chylomicrons are
used as a sources – neutral fats and phospholipides. In
humans FA synthesis from other molecules (Glc) is
usually small in comparison with dietary fatty acid
intake.
VLDL are formed in smooth ER of hepatocytes.
High concentration of acetyl-CoA (postabsorptive
state, starvation) as a result of β-oxidation of FA in
mitochondrion great amount of ketone bodies :
acetoacetate, 3-hydroxybutyrate and acetone.
12/2/12

Fat metabolism in the liver
Cholesterol has two sources, the diet and de
novo synthesis (in liver significant amount).
Some cholesterol is required for synthesis of bile
acids, some for cell membranes, some is stored
in the form of lipids droplets in esterified form.
The rest in free and esterified form in VLDL (to
supply another tissues)
The liver also degrades lipoprotein complexes
(with cholesterol and cholesterol esters) taking
up from the blood.

Amino acid metabolism in the liver
Our bodies do not continuously accumulate or
lose protein in a net sense. The rate of AA
oxidation in the body must therefore balance the
rate of entry of dietary protein (70-100g per day)
Catabolism of AA occurs predominantly in the
liver with exceptions (branched chain amino
acids in muscles)
AA oxidation provides ½ of the liver´s energy
requirements
It is also the only organ capable of eliminating
the nitrogen from amino acids by urea cycle

Starve-Feed Cycle
• The starve-feed cycle allows a variable fuel and nitrogen consumption to meet a variable metabolic and anabolic demand. Feed refers to intake of meals (variable fuel) after which we store the fuel in the form of glycogen and fat, to meet our metabolic demand while we fast. ATP is energy-transferring agent in this cycle.

Well-Fed State – Amino acids Dietary proteins are hydrolyzed in the intestine (some
of them are used like energy source here : Asp, Asn, Glu, Gln→Ala, Lac, citrul, Pro into the portal blood)
Liver lets most of AA coming from intestine pass through, for synthesis of proteins in peripheral tissue , thanks to high Km .High Km allows to AA to be in excess without catabolism.
Utilization of AA for proteosynthesis (much lower
Km for tRNA-charging enzymes)
Excess of AA can be oxidized to CO2 , water, urea, or metabolites can be used as substrates for lipogenesis

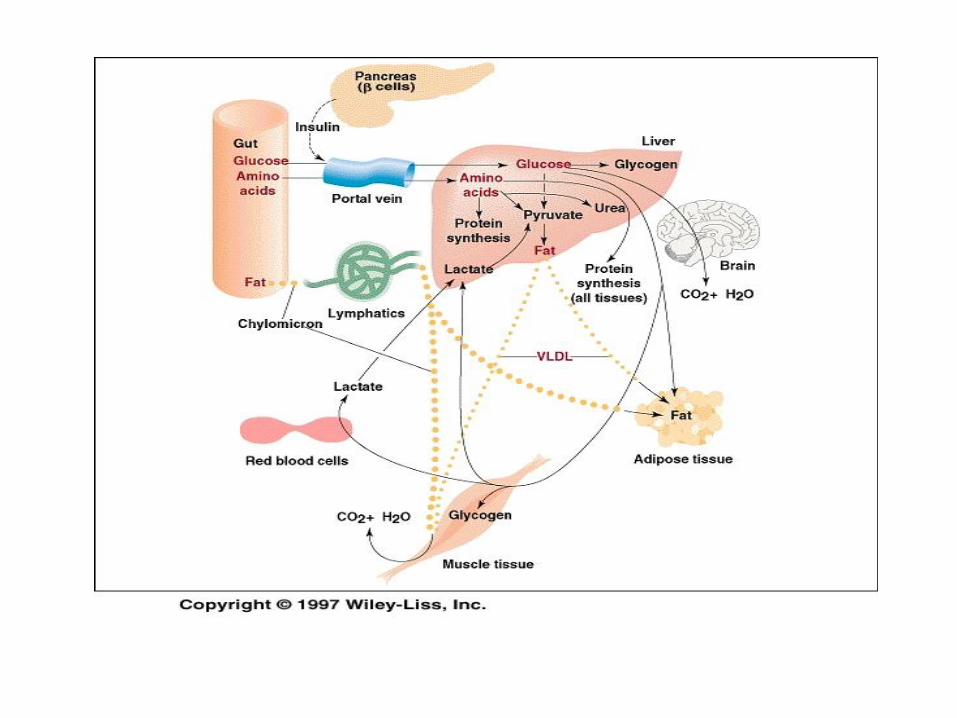
Well-Fed State - glucose
Glucose → glycogen (glycogenesis), pyruvate, lactate
(glycolysis), for pentose phosphate pathway
(NADPH)
Much of glucose from intestine passes through liver
to reach other organs (brain, testis, RBC, renal
medulla, AT)
Number of tissues produce lactate and pyruvate from
circulating glucose, which are taken up by liver , and
fat is formed lipogenesis)
In well-fed state liver does not engage in
gluconeogenesis
Cori cycle is interrupted

Well-βFed State – fat
Glucose, lactate, pyruvate and AA support hepatic lipogenesis.
Fat formed from these substrates is released in the form of VLDL
Chylomicrons, VLDLs circulate in the blood until they meet lipoprotein lipase (near AT), hydrolysis of TAG (FA taken up adipocytes, reesterified with glycerol-3-phosphate to form TAG)
During well-fed state insulin from cells of the pancreas is in high concentration. These cells are very responsive to the influx of glucose and AA in the fed state.
Rate of insulin/glucagon
Well-Fed State

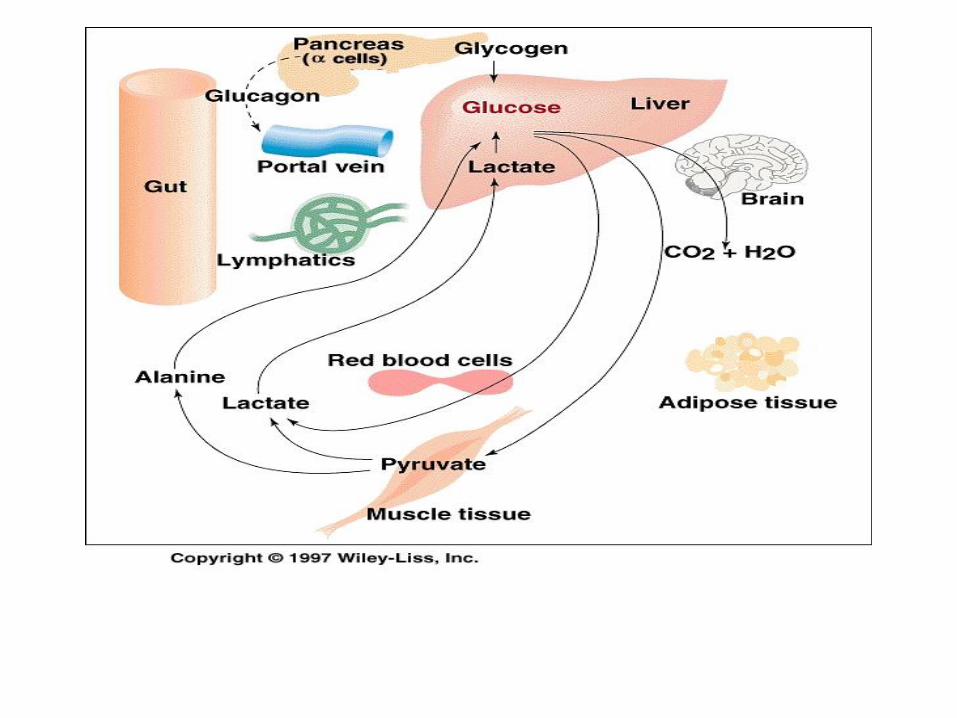
Early fasting state
Hepatic glycogenolysis
Lipogenesis is curtailed
Lactate, pyruvate and AA are diverted into formation
glucose completing Cori cycle (conversion glucose to
lactate, pyruvate in peripheral tissue, they are
substrates for gluconeogenesis in liver)
Alanine cycle, in which carbon and nitrogen return to
the liver in the form of alanine (muscle metabolizes
glucose to alanine, which is coming back to liver as a
substrate for gluconeogenesis)
ing State

• No fuels enters from the gut (duodenum) and little glycogen is left in the liver
• Tissues which require glucose are dependent on hepatic gluconeogenesis
• Cori and Alanine cycles play important role
• FAs can not be used for synthesis of glucose (acetyl-CoA can not be converted to glucose)
• Glycerol (by-product of lipolysis) becomes important substrate for gluconeogenesis
• AA, which are hydrolyzed in skeletal muscle (especially), supply most of the carbon atoms for net glucose synthesis mostly in the for of Ala and Gln

• Most of Gln released from muscle is converted (oxidized) into alanine and NH4
+ by intestinal epithelium and being released into bloodstream (glutaminolysis)
• Gluconeogenesis in the liver fasting is closely connected with to urea cycle (ornithine, carbamoyl phosphate, citrulline). Most AAs can give up the amino nitrogen by transamination with 2-oxo glutarate ….
• AT – lipolysis is activated (low blood insulin), blood level of fatty acids raises and are used by peripheral tissues (heart, muscle, liver – formation of glucose and ketone bodies)

• FA oxidation in liver provide most of ATP needed
for gluconeogenesis
• Acetyl~CoA is mostly converted to ketone bodies (small amount is oxidized completely)
• Ketone bodies and FA are preferred by many tissues over glucose; they can also suppress proteolysis and BCA oxidation in muscle
• Cooperation of tissues : liver synthesizes glucose, muscle and intestinal cells supply the substrate (alanine), and AT supplies the ATP (via FA oxidation in liver) needed for gluconeogenesis
• This cooperation is dependent on levels of hormones (insulin, glucagon, epinephrine)
• Reduction of triiodothyronine – reduces daily basal energy requirements by 25%
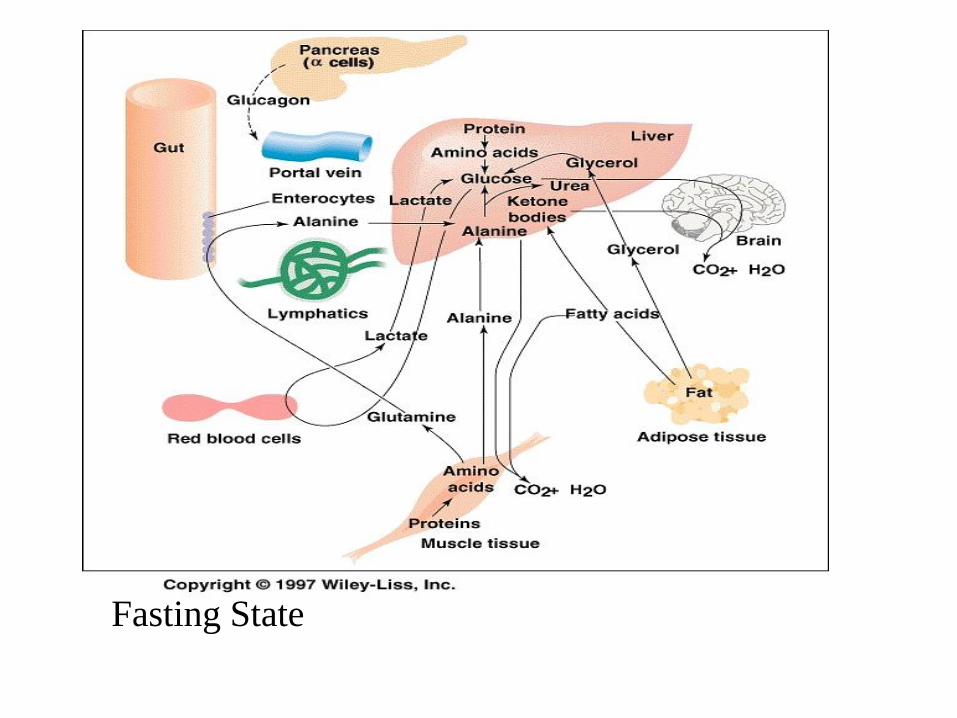
Fasting State

• After meal, fuel is again absorbed from gut
• Fat is metabolized like in well-fed state
• Glucose is poorly extracted by the liver, liver remains in the
gluconeogenic mode for a few hours after refed. Hepatic
gluconeogenesis provides glucose-6-phosphate for
glycogenesis. Rather, glucose is catabolized in peripheral
tissues to lactate which is converted in liver to glycogen
(glucose paradox)and substrates from it are used by liver for
gluconeogenesis and then glycogen.
• After the rate of gluconeogenesis declines, glycolysis becomes
the predominant means of glucose disposal in the liver
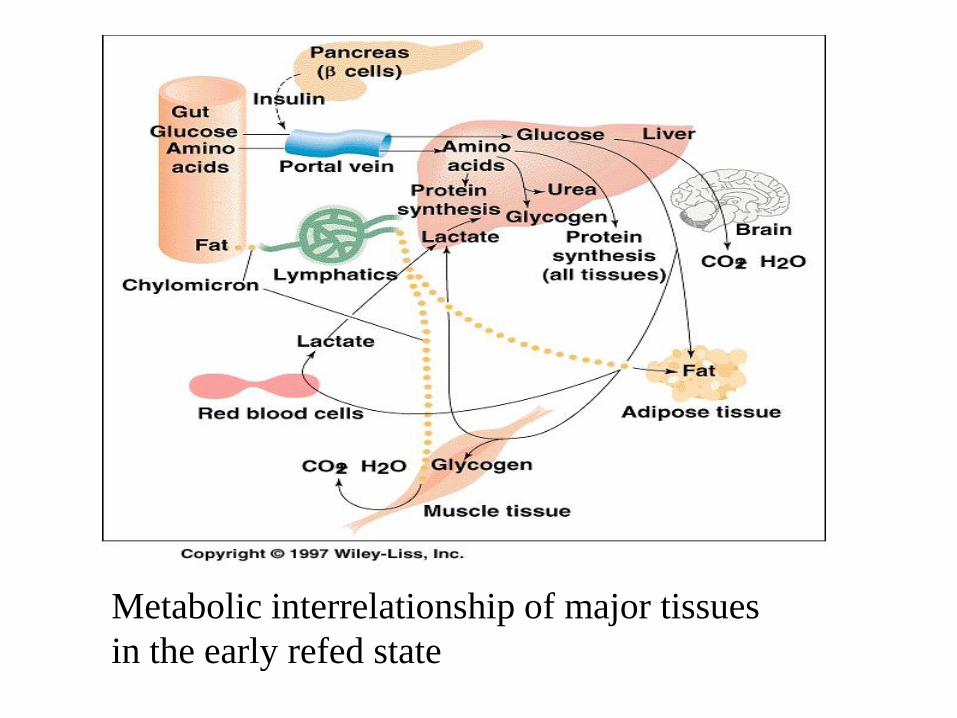
Metabolic interrelationship of major tissues
in the early refed state
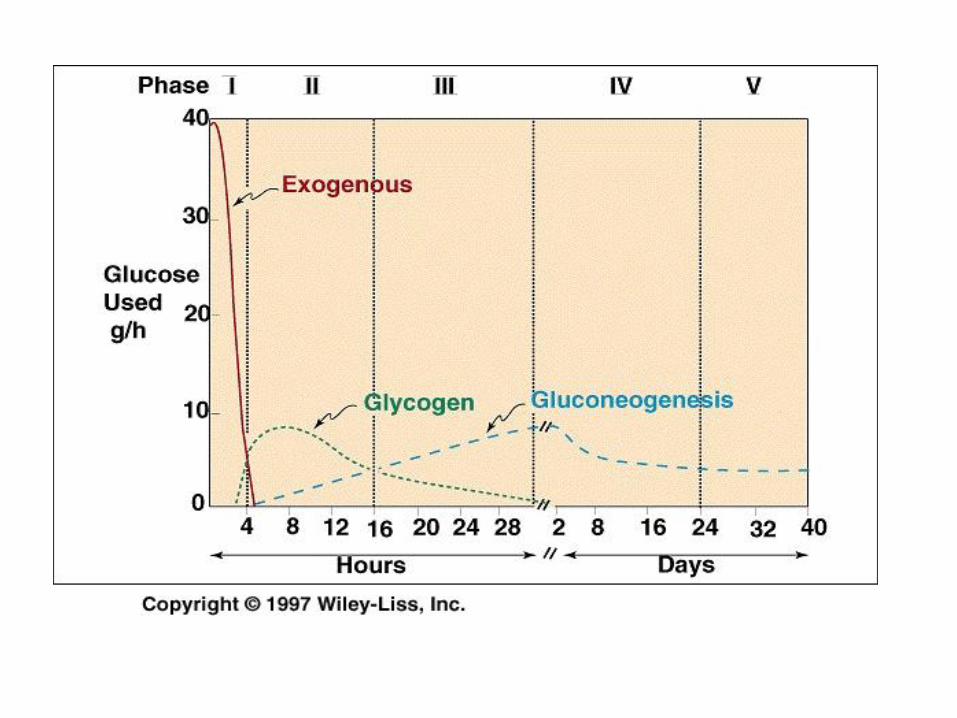
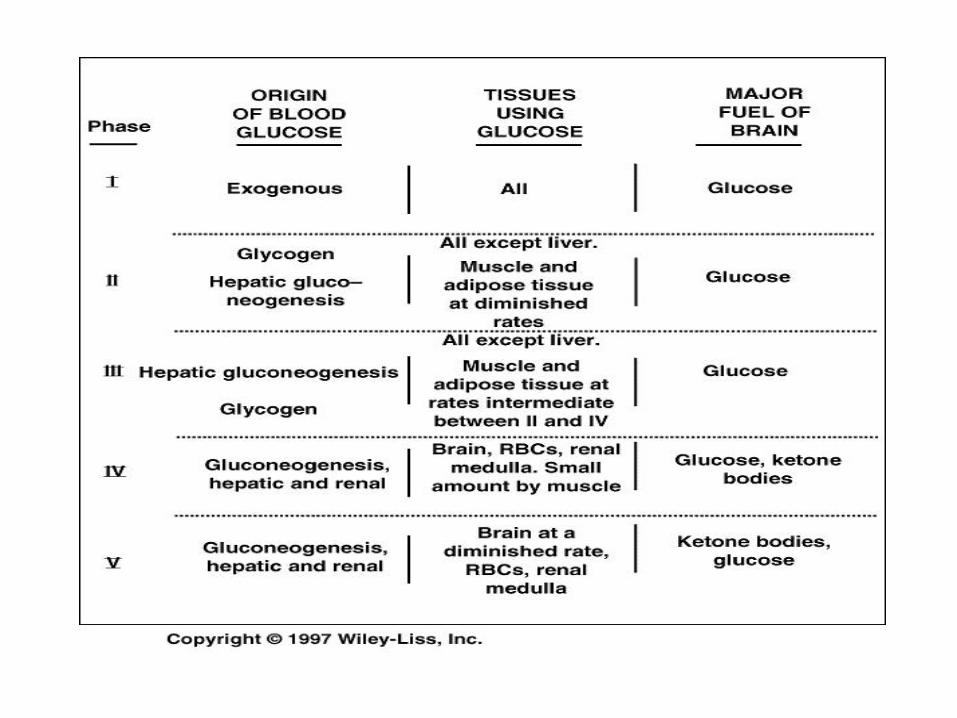

Exercise
Anaerobic exercise : sprinting or weight lifting (very little organ
cooperation), muscle largely relies on its own stored glycogen
and phosphocreatine.
Aerobic exercise : long-distance running
is metabolically more interesting. For moderate exercise, much
of the energy is derived from glycolysis of muscle glycogen -
content of it can be increased by exhaustive exercise that
depletes glycogen, followed by rest and a high-carbohydrate
diet. It is not enough glucose and glycogen for endurance
running – switching to fatty oxidation
The respiratory quotient (the ratio of CO2 exhaled to oxygen
consumed) falls during running-this indicates the progressive
switch from glycogen to fatty acid oxidation during the race.
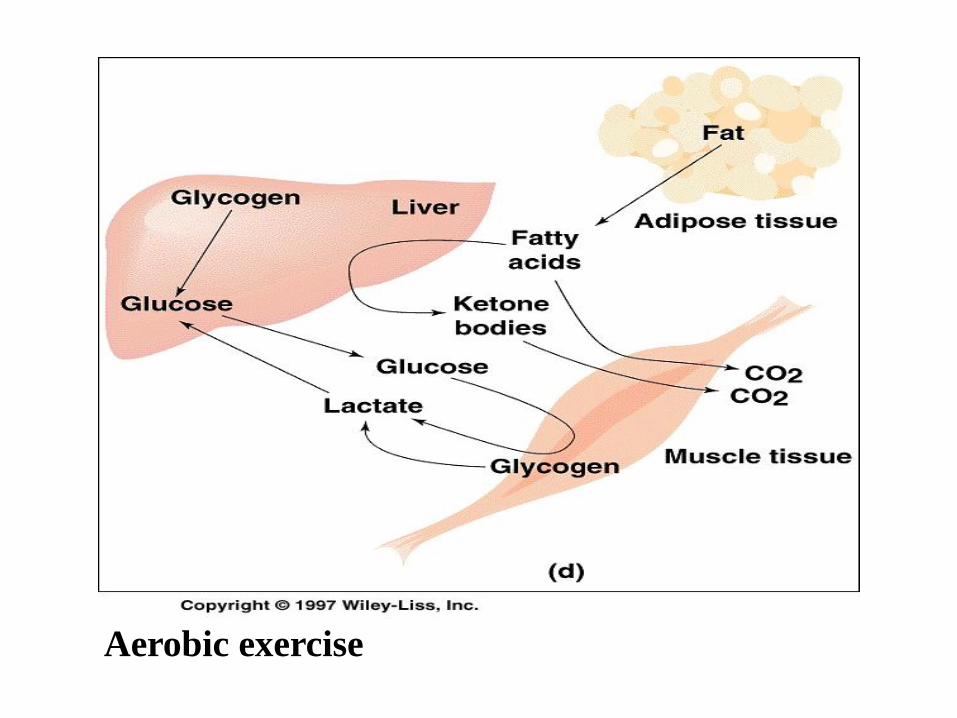
Aerobic exercise

• The liver is primarily responsible for the first two steps of
ethanol catabolism
• Alcohol dehydrogenase
• Aldehyde dehydrogenase
• Liver disposes of NADH generated by this reaction only in
mitochondrial electron transport chain – ethanol generates too
much NADH
• Some enzymes are inhibited by NADH (gluconeogenesis, -
oxidation) and also TCA is inhibited
• The result is fasting hypoglycemia and accumulation of TAG
• Fatty liver , cirhosis
• Redundant acetate is metabolized in peripheral tissue
• Formation of acetaldehyde adducts with proteins in the body
(control of abstinence)
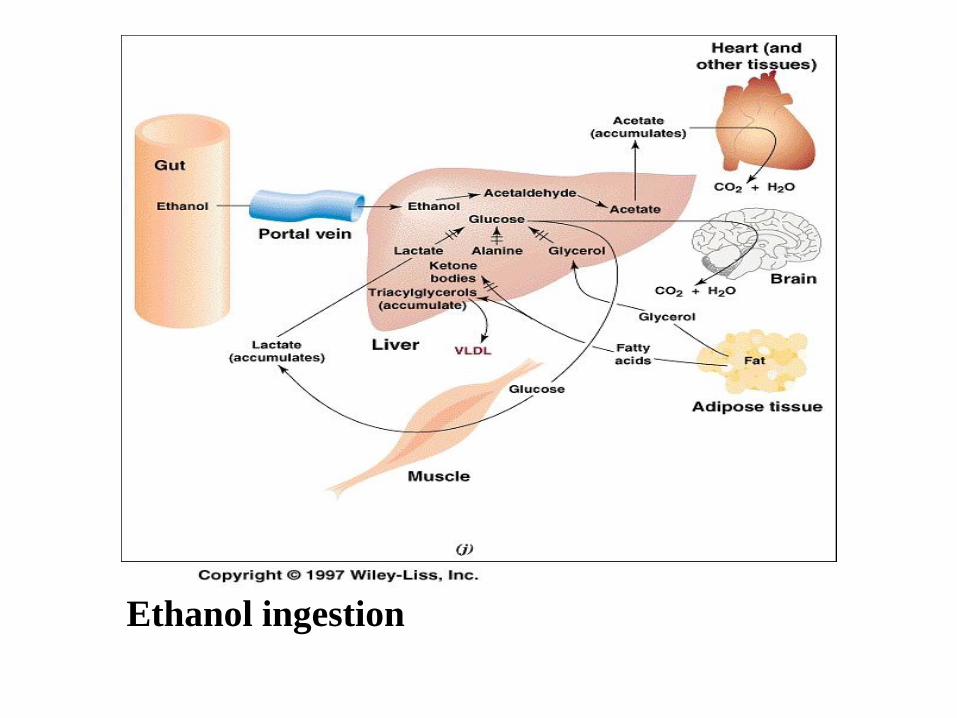
Ethanol ingestion







