
Pharmacological ChaperoneTherapy
Brandon WilcockMarch 27, 2008

Protein Folding and Structure
Wolynes, P.; Luthey-Schulten, Z.; Onuchic, J. Chemistry & Biology 1996, 3, 425-432.http://parasol.tamu.edu/foldingserver/images/landscape.gif
E

Endoplasmic Reticulum-Associated Degradation
Proteasome Ubiquitination
Sec61pComplexRibosome
Molecular Chaperone Recognition
ER
Hsp90
Ali, M.; Roe, S.; Vaughan, C.; Meyer, P.; Panaretou, B.; Piper, P.; Prodromou, C.; Pearl, L. Nature 2006, 440, 1013-1017.

Quality Control
Anelli, T.; Sitia, R. EMBO Journal 2008, 27, 315-327.

PC
ER
MisfoldedProtein
ERAD
ProteasomeDegradation
Transport to Cellular Location
Golgi ApparatusUbiquitination
Pharmacological Chaperone Assisted Rescue
Active-site Chaperone
Specific-site Chaperone

Identification and Characterization
• High-throughput screening
• Rational Design
• Thermal stability
• Mature protein quantification
• Toxicity
Exploring the mechanism
• ERAD and regulatory targets
• Direct interactions
• Specificity

Lysosomal Storage Diseases
Common Symptoms
Neurodegenerative states
anemia
skeletal dysplasia
Approved Treatments
Enzyme replacement therapy
Substrate reduction therapy Cell
ERT
AccumulationSRT

α-Galactosidase A and Fabry Disease
α-Galactosidase A(α-GalA)
Globotriaosylceramide(Gb3)
-GalA
NHOHO
O
O
OHO
HOOH
O
HO
HOOH
OHO
HO
HOOH
OH
NHOHO
O
O
OHO
HOOH
O
HO
HOOH
O
Garman, S.; Garboczi, D.; J. Mol. Biol. 2004, 337, 319-335.

DGJ Enhances R301Q-α-GalA Activity
NH
OHHO
OHHO1-deoxygalactonojirimycin
(DGJ)
Fan, J.; Ishii, S.; Asano, N.; Suzuki, Y. Nat. Med. 1999, 5, 112-115.
4-methylumbelliferyl-α-galactoside
(4-Mu-α-Gal)O
OHHO
HOHO
O
O
O
Me

The Inhibitory Effect
DGJ inhibits R301Q-α-GalA activity at 20 μM
Fan, J.; Ishii, S.; Asano, N.; Suzuki, Y. Nat. Med. 1999, 5, 112-115.
R301Q-α-GalA Activity
ER
Lysosome

ERAD
PC
ER
proteasomedegradation
golgi apparatus, transport
DGJ Increases Mature Protein Levels
Fan, J.; Ishii, S.; Asano, N.; Suzuki, Y. Nat. Med. 1999, 5, 112-115.
Mature α-GalA 46 kD
Immature α-GalA 50 kD
Protein level enhancement is DGJ concentration Dependent

The Stability of R301Q-α-GalA
Fan, J.; Ishii, S.; Asano, N.; Suzuki, Y. Nat. Med. 1999, 5, 112-115.
Rel
ativ
e E
nzym
e A
ctiv
ity (%
)
Rel
ativ
e E
nzym
e A
ctiv
ity (%
)
Time (min) Time (min)
R301Q-α-GalA is unstable at ER pH
pH 5 pH 6 pH 7
DGJ stabilizes R301Q-α-GalA
0.01 μM0.03 μMNone
1 μM[DGJ]

Structure and Inhibitor Strength
NHHO
HO
OHHO
manno-DNJNI
NHHO
OHHO
2-deoxy-DGJ250
Asano, N.; Ishii, S.; Kizu, H.; Ikeda, K.; Yasuda, K.; Kato, A.; Martin, O.; Fan, J. Eur. J. Biochem. 2000, 267, 4179-4186.Ishii, S.; Chang, H.; Kawasaki, K.; Yasuda, K.; Wu, H.; Garman, S.; Fan, J. Biochem. J. 2007, 406, 285-295.

Structure and Inhibitor Strength
N
OHHO
OHHOMe
N-ethyl-DGJ306
N
OHHO
OHHO
OH
N-hydroxyethyl-DGJ520
Asano, N.; Ishii, S.; Kizu, H.; Ikeda, K.; Yasuda, K.; Kato, A.; Martin, O.; Fan, J. Eur. J. Biochem. 2000, 267, 4179-4186.
N-alkylation decreases binding affinity
NH
HOHO
HOOH
-HNJNI
OH
NHHO
HO
OHHO
OH
-manno-HNJ464
NH
HO
HOOH
OHOH
-allo-HNJ4.3
NH
HOHO
OHHO
OH
-galacto-HNJ0.21

Structure and Activity Enhancement
Binding affinity
Cell and ER permeability
Rate of metabolism
Asano, N.; Ishii, S.; Kizu, H.; Ikeda, K.; Yasuda, K.; Kato, A.; Martin, O.; Fan, J. Eur. J. Biochem. 2000, 267, 4179-4186.
Cell ER

R301Q-α-GalA Localization
Yam, G.; Zuber, C.; Roth, J. FASEB J. 2005, 19, 12-18.
Wild Type R301Q DGJ treated R301Q
α-GalA
LAMP1
Overlay
DGJ increases R301Q-α-GalA levels in lysosomes
Yam, G.; Zuber, C.; Roth, J. FASEB J. 2005, 19, 12-18.
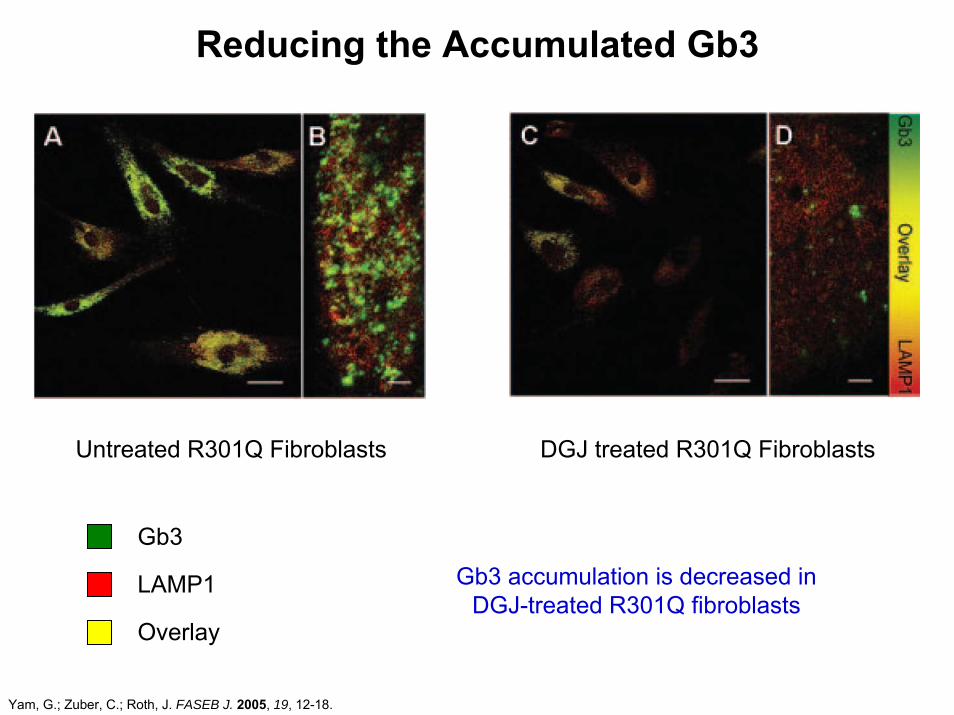
Reducing the Accumulated Gb3
Gb3
LAMP1
Overlay
Untreated R301Q Fibroblasts DGJ treated R301Q Fibroblasts
Gb3 accumulation is decreased in DGJ-treated R301Q fibroblasts

In Vivo Mice Studies
Ishii, S.; Yoshioka, H.; Mannen, K.; Kulkarni, A.; Fan, J. Biochim. Biophys. Acta 2004, 1690, 250-257.
TissueR301Q-α-GalA
Activity Increase
Heart
Kidney
Spleen
Liver
13.8 fold
3.3 fold
3.9 fold
2.6 fold
3 mg/kg DGJ per day
No toxicity observed

Expansion to Other Mutations
Shin, S.; Murray, G.; Kluepfel-Stahl, S.; Cooney, A.; Quirk, J.; Schiffmann, R.; Brady, R.; Kaneski, C. Biochem. Biophys. Res. Comm. 2007, 359, 168-173.
A97V
R301Q
R112H
R112C
A143T
L300P
R356W
G132R
R220X
A143P
30ΔG
Rescued Not Rescued
DGJ treatment success is mutation dependent

Neg. C
ontrolW
ild Type
A20PE59K
E66QM72V
I91TR112H
A97VF113L
P146S
Expansion to Other Mutations
Ishii, S.; Chang, H.; Kawasaki, K.; Yasuda, K.; Wu, H.; Garman, S.; Fan, J. Biochem. J. 2007, 406, 285-295.
DGJ can be therapeutic for several mutants

Application in Cystic Fibrosis
NBD2NBD1
TMD1 TMD2
RD
ATP
P
TMD1 TMD2
NBD2NBD1
RD
PKA ATP
ATP
Cystic fibrosis transmembrane conductance regulator protein (CFTR)
http://www.cystic-fibrosis-symptom.com/cystic_fibrosis_pictures.htm

A Common Mutation ∆F508
Lung infections and inflammation
ΔF508
F508A
F508
90 % of CF patients
Folding defect corrected at 27 °C
Loo, T.; Bartlett, M.; Clarke, D. J. Bioenerg. Biomembr. 2005, 37, 501-507. Lewis, H.; et al. J. Biol. Chem. 2005, 280, 1346-1353.

High-Throughput Screening
YFPYFP
I-
Halide sensitive fluorescent proteins
Cl-
DiSBAC2(3)
Coumarin Coumarin
Voltage sensitive dye
Pedemonte, N.; Lukacs, G.; Du, K.; Caci, E.; Zegarra-Moran, O.; Galietta, L.; Verkman, A.; J. Clin. Invest. 2005, 115, 2564-2571.Van Goor, F.; et al. Am. J. Physiol. Lung. Cell. Mol. Physiol. 2006, 290, 1117-1130.

Potential Pharmacological Chaperones
Lewis, H.; et al. J. Biol. Chem. 2005, 280, 1346-1353.Van Goor, F.; et al. Am. J. Physiol. Lung Cell. Mol. Physiol. 2006, 290, 1117-1130.

CFcor-325 and ∆F508-CFTR
CFcor-325 increases ∆F508-CFTR activity and
mature protein levels
Loo, T.; Bartlett, C.; Clarke, D. Molecular Pharmaceutics 2005, 2, 407-413.
CFcor-325

Further Studies With CFcor-325
Loo, T.; Bartlett, C.; Wang, Y.; Clarke, D. Biochem J. 2006, 395, 537-542.Varga, K.; Goldstein, R.; Jurkuvenaite, A.; Chen, L.; Matalon, S.; Sorscher, E.; Bebok, Z.; Collawn, J. Biochem J. 2008, 410, 555-564.
CFcor-325 extends half-life of ∆F508-CFTR
Treatment is mutation dependent
ControlCFcor-325

P-Glycoprotein and Specificity
Wang, Y.; Bartlett, C.; Loo, T., Clarke, D. Molecular Pharmacology 2006, 70, 297-302.
CFcor-325 inhibits P-Glycoprotein
CFcor-325 Rescues P-Glycoprotein Mutant
P-Glycoprotein Activity
CFpot-532 and corr-2b are more specific for CFTR

Summary
Mutant protein needs to be retained in the ER
Mutant protein must demonstrate some activity
Treatment is mutation specific
Applicable to many genetic diseases characterized by misfolded proteins
High-throughput screening can identify potential pharmacological chaperones
Low impact on other cellular pathways
Limitations
Future
Advantages
More general method for identification

Acknowledgements
Professor Silverman
Chem 575 Class
Professor Burke
Burke Group







