
Mol. Nutr. Food Res. 61, 6, 2017, 1600659 (1 of 19) 1600659DOI 10.1002/mnfr.201600659
REVIEW
Phytochemicals enhance antioxidant enzyme expression
to protect against NSAID-induced oxidative damage of
the gastrointestinal mucosa
Yu-Ting Cheng, Chi-Cheng Lu and Gow-Chin Yen
Department of Food Science and Biotechnology, National Chung Hsing University, Taichung, Taiwan
Received: July 29, 2016Revised: November 15, 2016
Accepted: November 17, 2016
The gastrointestinal (GI) mucosa provides the first protective barrier for digested food andxenobiotics, which are easily attacked by toxic substances. Nonsteroidal anti-inflammatorydrugs, including aspirin, diclofenac, indomethacin, and ketoprofen, are widely used in clinicalmedicine, but these drugs may cause oxidative stress, leading to GI damage such as ulcers.Lansoprazol, omeprazole, and other clinical drugs are widely used to treat duodenal and gas-tric ulcers and have been shown to have multiple biological functions, such as antioxidantactivity and the ability to upregulate antioxidant enzymes in vivo. Therefore, the reduction ofoxidative stress may be an effective curative strategy for preventing and treating nonsteroidalanti-inflammatory drug induced ulcers of the GI mucosa. Phytochemicals, such as dietary phe-nolic compounds, phenolic acids, flavan-3-ols, flavonols, flavonoids, gingerols, carotenes, andorganosulfur, are common antioxidants in fruits, vegetables, and beverages. A large amountof evidence has demonstrated that natural phytochemicals possess bioactivity and potentialhealth benefits, such as antioxidant, anti-inflammatory, and antibacterial benefits, and they canprevent digestive disease processes. In this review, we summarize the literature on phytochem-icals with biological effects, such as angiogenic, antioxidant, antiapoptotic, anti-inflammatory,and antiulceration effects, and their related mechanisms are also discussed.
Keywords:
Antioxidant enzymes / NSAIDs / Oxidative stress / Phytochemicals / Ulcer
1 Introduction: prevalence
Peptic ulcers induced by mucosal damage are a prevalentdigestive disease in the United States, and approximately500,000 individuals with gastrointestinal (GI) ulcers are diag-nosed each year [1–4]. The GI mucosa provides the first pro-tective barrier against xenobiotics, but it is easily damaged bytoxic substances. Serious ulceration may lead to GI bleedingor even perforation. The pathophysiology of peptic ulcers is of-
Correspondence: Dr. Gow-Chin YenE-mail: [email protected]
Abbreviations: COX, cyclooxygenase; EGCG, (−)-epigallocatech-in-3-gallate; GI, gastrointestinal; GSH, glutathione; GSSG, ox-idized GSH; GPx, glutathione peroxidase; GR, glutathione re-ductase; HO-1, heme oxygenase-1; HSP70, heat shock protein-70; iNOS, inducible nitric oxide synthase; LPO, lipid peroxida-tion; MDA, malondialdehyde; MPO, myeloperoxidase; Nrf2, nu-clear factor erythroid 2 related factor 2; NSAID, nonsteroidal anti-inflammatory drug; PCA, protocatechuic acid; PG, prostaglandin;RCS, reactive carbonyl species; ROS, reactive oxygen species;SOD, superoxide dismutase; TGF, transforming growth factor
ten depicted as an imbalance between aggressive and protec-tive factors (Fig. 1). Common endogenous aggressive factorsinclude bile salts, gastric acid, and pepsin, whereas exoge-nous aggressive factors include the consumption of alco-hol, infection by Helicobacter pylori, and nonsteroidal anti-inflammatory drugs (NSAIDs). Relevant protective factorsinclude mucosal blood flow, mucosal cellular proliferationand restitution, and prostaglandin (PG) [4–7].
NSAIDs, including aspirin, diclofenac, indomethacin,naproxen, and ketoprofen, are all available to control acuteand chronic pain from rheumatoid arthritis and to alleviateswelling as anti-inflammatory medications. However, theiruse can cause oxidative damage to the GI mucosa in the clini-cal stage [5]. It has been reported that intestinal diseases, suchas ulcers, are related to lipid peroxidation (LPO) and oxidativedamage in the mucosa [8]. It is important to reduce the levelsof reactive oxygen species (ROS) in GI ulcers because ROS cancause oxidative damage to biological macromolecules such aslipids, DNA, and proteins.
Colour online: See the article online to view Fig. 1 in colour.
C© 2016 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.mnf-journal.com

1600659 (2 of 19) Y.-T. Cheng et al. Mol. Nutr. Food Res. 61, 6, 2017, 1600659
Figure 1. The defensive and aggres-sive factors related to peptic ulcer dis-eases. The pathophysiology of a gas-trointestinal ulcer is often depictedas an imbalance between aggressivefactors and protective factors. Com-mon endogenous aggressive factorsinclude bile salts, gastric acid, andpepsin, whereas exogenous aggres-sive factors include the consumption ofalcohol, Helicobacter pylori infection,and NSAIDs. The relevant protectivefactors include prostaglandin (PG), mu-cosal blood flow, mucosal cellular pro-liferation, and restitution.
The appearance of LPO and oxidative damage in the GImucosa is correlated with intestinal diseases, chronic intesti-nal inflammation, or peptic ulcers [9–11]. Phytochemicalspossess multiple biofunctional activities, including scaveng-ing of ROS, trapping of reactive carbonyl species (RCS), andchelation of metal ions [12]. The dietary intake of natural an-tioxidants could decrease the risk of oxidative stress relateddiseases in living systems. Hence, the use of oxygen rad-ical scavengers and antioxidant supplements from naturalfoods might be an effective and potential therapeutic strat-egy for preventing tissue damage under oxidative stress. Inthis review, we summarize and characterize the beneficialhealth effects of recent experimental and clinical studies de-scribing the major antioxidant compounds of vegetables and
fruits that upregulate antioxidant enzymes to protect againstNSAID-induced oxidative damage of the GI mucosa.
2 Relationship between NSAID-inducedoxidative injury of the GI mucosa andrelated effects such as apoptosis,angiogenesis, antioxidant enzymes,and inflammation
Peptic ulcers are a multifactorial and complex dis-ease involved in duodenal and gastric ulcers. Althoughthere have been medical advances, the managementof peptic ulcers and their complications remain a
Figure 2. Mechanisms of NSAID-induced gastrointestinal ulcers.There are several mechanisms forNSAID-induced gastrointestinal ul-cers, including the upregulation ofoxidative stress, cyclooxygenase-2(COX-2), and apoptosis, and thedownregulation of cyclooxygenase-1 (COX-1), prostaglandin (PG),endothelin-converting enzyme-1 (ECE-1), angiogenesis, mucosal cellularproliferation, and restitution.
C© 2016 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.mnf-journal.com

Mol. Nutr. Food Res. 61, 6, 2017, 1600659 (3 of 19) 1600659
challenge, including high morbidity and death ratesfor the disease [13, 14]. NSAIDs are widely used tolessen inflammation, fevers, and pain in clinical medicineand can be divided into non-cyclooxygenase (non-COX)selective (COX-1 and COX-2) inhibitors and COX-2selective inhibitors [15]. Epidemiologic studies indicate thatpatients with non-COX selective NSAIDs have a higher riskof ulcers in the GI tract [6, 13, 15]. The major non-COX se-lective inhibitors include aspirin, diclofenac, indomethacin,piroxicam, naproxen, and ketoprofen [5, 16, 17].
Nonselective NSAID induction of peptic ulcers is a mul-tifaceted process that includes the generation of ROS, induc-ing inflammatory molecules of the COX-2, cytosolic phos-pholipases A2, leukotriene B4, 5-lipoxygenase, PGE2, TNF-�,IL-1�, and IL-6 production [18–21], increasing LPO, xanthineoxidase, and DNA damage (8-OHdG production) [3, 22], andthe inhibition of antioxidant enzymes including glutathioneperoxidase (GPx), glutathione reductase (GR), glutathioneS-transferase, catalase, superoxide dismutase (SOD), andheme oxygenase-1 (HO-1) as well as glutathione (GSH)/ oxi-dized GSH (GSSG) ratio in vivo [23,24]. Moreover, ROS mayalso cause oxidative damage to biological macromoleculesand react with proteins, lipids, and mitochondrial DNA, lead-ing to cells death by affecting various apoptotic pathways inthe GI tract [3,5,22]. Furthermore, taking NSAIDs also down-regulates angiogenesis, inhibits mucosal cellular restitution,and promotes extracellular matrix degradation [25, 26], as il-lustrated in Fig. 2 and Table 1. Thus, it is important to regulatethe levels of ROS in the GI tract. In this study, we summarizethe literature on multi-NSAIDs with varying degrees of ul-cerogenic activity associated with different biological effectsincluding oxidative stress, inflammation, and GI protection.
2.1 The role of oxidative stress and antioxidant
enzymes in ulceration
NSAID-induced LPO and oxidative stress might also causeulcers in the GI mucosa [27]. Some reports have indicatedthat oral administration of NSAIDs causes GI oxidative injurythrough increased lactate dehydrogenase leakage, mucosalLPO (malondialdehyde, MDA), DNA damage, and decreasedgastric mucus secretion in vivo [3, 27]. Thus, upregulationof antioxidant enzymes, such as GPx, GR, SOD, and HO-1, might be a major mechanism of action against oxidativestress associated GI ulcers.
The selenium-dependent enzyme GPx is thought to actas a barrier against hydroperoxide attack [28]. Brigelius-Flohe[29] indicated that GI isoenzyme (GI-GPx) is the most re-lated to a classical enzyme and might provide a barrieragainst hydroperoxides from the metabolism of xenobiotics.GR is responsible for maintaining a balance of reduced GSHand GSSG. The activation of GR plays an important rolein increasing the concentration of GSH, which maintainsthe redox status of the organism [30]. Thus, the ratio ofGSH/GSSG is an index for oxidative stress. The levels of
the total sulfhydryl groups are important in maintaining an-tioxidation or detoxification of ROS and intracellular redoxstatus [31]. Cheng et al. [3] reported that ketoprofen not onlyincreased the levels of LPO and ROS but also decreased thelevels of intracellular antioxidants in vitro, such as GPx, GR,and total sulfhydryl groups. In addition, HO-1 and SOD areimportant antioxidant enzymes for the elimination of ROSand RCS, and they can act as metal ion chelators in the hu-man body [32–34]. HO-1 protects GI cells against oxidativedamage through the breakdown of heme to biliverdin andcarbon monoxide [32] and suppresses inflammation and tis-sue damage after ROS induction in the intestine and stom-ach [33]. Some reports also indicate that administration ofNSAIDs such as indomethacin, diclofenac, piroxicam, andketoprofen reduced the levels of GPx, GR, SOD, catalase ac-tivities, and the GSH and GSH/GSSG ratio in the GI mucosa[3,19,35]. These updated data confirm the well-established di-verse benefits of pharmacological actions and might supportthe standpoint for nutraceutical applications.
2.2 Inhibition of PGs by COX in ulceration
Inflammatory diseases such as asthma, rheumatoid arthri-tis, and hepatitis are a major cause of morbidity in humans.Reports on chronic inflammation indicate that it can lead tothe development of cancer as well as cardiovascular and neu-rodegenerative diseases [36–38]. The rate of conversion ofarachidonic acid into prostanoids depends on the availabilityof two COX enzymes, COX-1 and COX-2 [39]. COX-1 con-verts arachidonic acid into PGs. PGs are potent vasodilatorsthat control almost all aspects of the gastric mucosal defenseand healing. Inhibition of COX-1 and PG synthesis has beenshown to cause serious GI damage [15]. It has been previ-ously reported that the GI toxicity of NSAIDs in rats resultsfrom a combination of the inhibition of COX-1 and the stim-ulation of COX-2 production in gastric mucosal lesions [40].Furthermore, increased COX-2 gene expression is commonlyassociated with inflammatory responses [41].
2.3 NSAIDs induce apoptosis in the GI mucosa
GI ulcer formation is a multifaceted process that includesROS generation, extracellular matrix degradation, and mito-chondrial damage. Mitochondria play a crucial role in ROShomeostasis and cell survival [42]. Musumba et al. [5] alsoreported that NSAID-induced ROS might also cause GImucosal cell apoptosis by affecting various signaling cas-cades such as those involving Bax, caspase-8, caspase-9, andcaspase-3 activities. Hence, the downregulation of ROS lev-els may be the most important cytoprotective mechanismagainst NSAID-induced GI tissue/cell apoptosis. Therapiesthat effectively target NSAID-induced apoptosis likely re-quire inhibition of both mitochondria- and caspase-mediatedpathways.
C© 2016 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.mnf-journal.com
1600659 (4 of 19) Y.-T. Cheng et al. Mol. Nutr. Food Res. 61, 6, 2017, 1600659
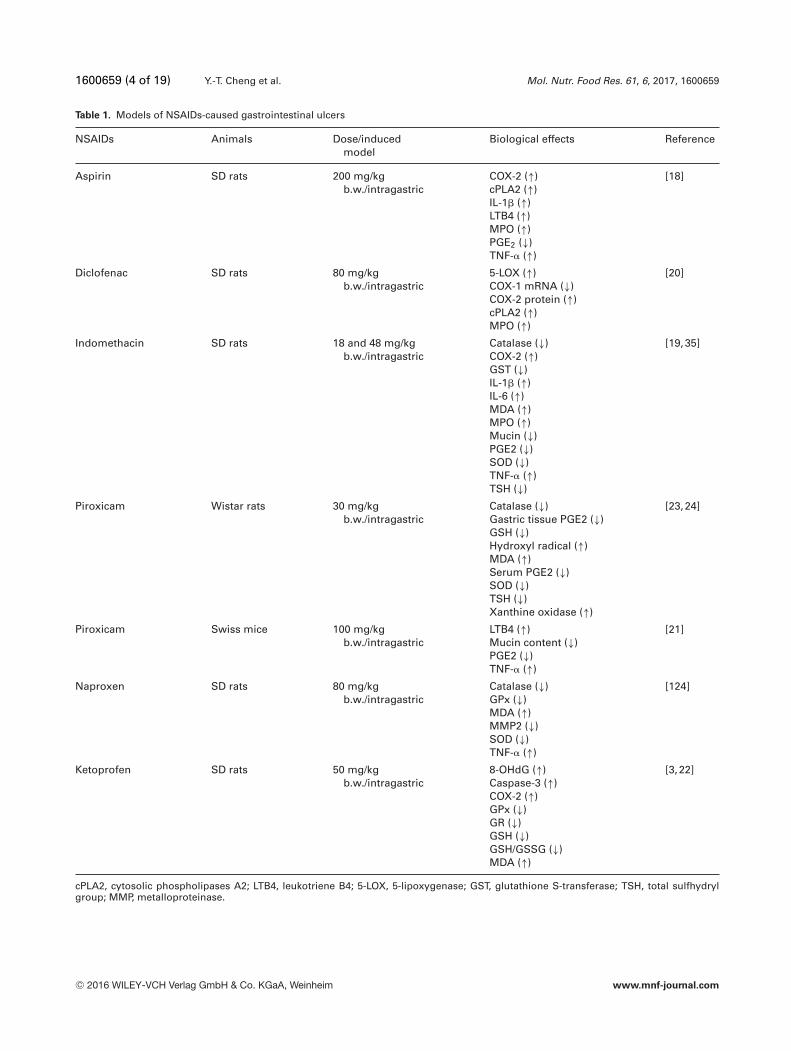
Table 1. Models of NSAIDs-caused gastrointestinal ulcers
NSAIDs Animals Dose/inducedmodel
Biological effects Reference
Aspirin SD rats 200 mg/kgb.w./intragastric
COX-2 (↑)cPLA2 (↑)IL-1� (↑)LTB4 (↑)MPO (↑)PGE2 (↓)TNF-� (↑)
[18]
Diclofenac SD rats 80 mg/kgb.w./intragastric
5-LOX (↑)COX-1 mRNA (↓)COX-2 protein (↑)cPLA2 (↑)MPO (↑)
[20]
Indomethacin SD rats 18 and 48 mg/kgb.w./intragastric
Catalase (↓)COX-2 (↑)GST (↓)IL-1� (↑)IL-6 (↑)MDA (↑)MPO (↑)Mucin (↓)PGE2 (↓)SOD (↓)TNF-� (↑)TSH (↓)
[19,35]
Piroxicam Wistar rats 30 mg/kgb.w./intragastric
Catalase (↓)Gastric tissue PGE2 (↓)GSH (↓)Hydroxyl radical (↑)MDA (↑)Serum PGE2 (↓)SOD (↓)TSH (↓)Xanthine oxidase (↑)
[23,24]
Piroxicam Swiss mice 100 mg/kgb.w./intragastric
LTB4 (↑)Mucin content (↓)PGE2 (↓)TNF-� (↑)
[21]
Naproxen SD rats 80 mg/kgb.w./intragastric
Catalase (↓)GPx (↓)MDA (↑)MMP2 (↓)SOD (↓)TNF-� (↑)
[124]
Ketoprofen SD rats 50 mg/kgb.w./intragastric
8-OHdG (↑)Caspase-3 (↑)COX-2 (↑)GPx (↓)GR (↓)GSH (↓)GSH/GSSG (↓)MDA (↑)
[3,22]
cPLA2, cytosolic phospholipases A2; LTB4, leukotriene B4; 5-LOX, 5-lipoxygenase; GST, glutathione S-transferase; TSH, total sulfhydrylgroup; MMP, metalloproteinase.
C© 2016 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.mnf-journal.com

Mol. Nutr. Food Res. 61, 6, 2017, 1600659 (5 of 19) 1600659
2.4 Role of angiogenesis in GI mucosal repair
Another method for protecting against NSAID-induced dam-age of the mucosa may be involved in the upregulation of neo-vascularization in the GI tract [25,26,43]. Neovascularization,which is the generation of new capillaries, is tightly controlledand normally occurs during embryonic and postembryonicdevelopment, the reproductive cycle, and wound repair [43].Reports indicate that against NSAID-induced damage of pep-tic ulcers may involve the upregulation of neovasculariza-tion in the GI mucosa [25, 26]. An effective approach for theremedy of stomach ulcers might be through enhancement ofstomach epithelial cell migration [44]. Angiogenic growth fac-tors, such as fibroblast growth factors , transforming growthfactor-alpha (TGF-�), trefoil factors, hepatocyte growth factor,basic fibroblast growth factor, platelet-derived growth factor,TGF-�, and vascular endothelial growth are the most impor-tant factors in GI mucosal reconstruction, mucosal defense,and ulcer healing [43–45]. Deng et al. [43] indicated that anincrease in angiostatin or endostatin (an endogenous angio-genesis inhibitor) plays a pathologic role in impaired anddelayed healing of intestinal mucosal ulcers. These resultssuggest that angiogenesis serves as a barrier against NSAID-induced GI mucosal damage and can help mucosal repair.
3 Gastroprotective effects ofphytochemicals and their underlyingmechanisms
NSAIDs, including aspirin, diclofenac, indomethacin,naproxen, and ketoprofen, are all available to control theacute and chronic pain of rheumatoid arthritis, and theiranti-inflammatory effects alleviate swelling; however, theiruse could cause oxidative injury of the GI mucosa in clinicalmedicine [6,41]. Reports indicate that NSAID-induced pepticulcers are associated with oxidative stress and apoptosis. Theappearance of LPO and oxidative damage in the GI mucosa iscorrelated with intestinal diseases, chronic intestinal inflam-mation, or ulcers [8]. These results suggest that GI ulcersinduced by NSAIDs result from the induction of oxidativestress and/or the reduction of antioxidant enzymes.
Nuclear factor erythroid 2 related factor 2 (Nrf2), which isa vital transcription factor, binds to an antioxidant responseelement in the promoter regions of genes encoding sev-eral antioxidant and detoxifying enzymes and related stress-responsive proteins [46]. Nrf2 elicits rescue pathways thatprotect cells against oxidative damage, abnormal immune re-sponses, abnormal inflammation, apoptosis, and carcinogen-esis [47, 48]. Under normal conditions, Nrf2 is inactive andis bound in the cytosol by Kelch-like ECH-associated protein1 (Keap1). Stimuli of the cytoplasmic Keap1 protein, includ-ing antioxidants, oxidants, chemopreventive agents, and elec-trophilic agents, that possess the ability to modify thiol groupsand release Nrf2 from Keap1 allow Nrf2 to translocate intothe nucleus and bind to the antioxidant response element.
Nrf2 regulates antioxidant and phase II detoxification en-zymes such as NAD(P)H:quinone acceptor oxidoreductase 1,SOD, catalase, GPx, GR, and HO-1 [49].
GPx, HO-1, and SOD are important antioxidant enzymesfor the elimination of ROS and RCS, and they can act as metalion chelators in the human body [32–34]. The GI isoenzyme-GPx is the main gastric enzyme, which serves as a barrieragainst hydroperoxide [29]. HO-1 protects GI cells against ox-idative damage through the breakdown of heme to biliverdinand carbon monoxide [32] and suppresses inflammation andtissue damage by ROS induction in the stomach and intes-tine [33]. Hence, the upregulation of antioxidant enzymesby Nrf2 may be related to the prevention of NSAID-inducedGI ulcers. Phytochemicals, such as sulforaphane, catechin,and caffeic acid, effectively prevent the NSAID-induced in-jury of intestinal epithelial cells by upregulating Nrf2/HO-1signaling pathways [3,22,50]. Hence, the upregulation of an-tioxidant enzymes by Nrf2 may be related to the preventionof NSAID-induced GI ulcers.
Recent reports have suggested that cytosolic Nrf2 becomesunstable without DJ-1 (also referred to as PARK7), and the re-sponses of Nrf2-regulated downstream antioxidant enzymesare attenuated [51, 52]. Early-onset Parkinson’s disease andneurodegenerative disorders have been linked to pathogenicmutations in DJ-1 [53]. Taira et al. [54] indicated that upreg-ulation of DJ-1 prevented neuroblastoma cell death due tooxidative stress in Parkinson’s disease. Previous reports indi-cated that catechin effectively prevented ketoprofen-inducedoxidative damage by the upregulation of HO-1 in Int-407 cells,which might play a key role in the prevention of NSAID-induced mucosal damage [3]. Cheng et al. [22] also showedthat caffeic acid effectively prevented ketoprofen-induced ox-idative damage to the GI cells by inducing the expressionof antioxidant enzymes and through the DJ-1/Nrf2-mediatedprocess. These results indicate that increasing antioxidant en-zyme expression by Nrf2 might represent a potential curativeapproach to protect patients against oxidative stress relateddevelopment of GI mucosal ulcers. Importantly, it is docu-mented that dietary foods may be effective against GI ulcersby the upregulation of antioxidant enzymes and the down-regulation of oxidative stress [55].
Aspirin and other NSAIDs, such as indomethacin anddiclofenac, are commonly used for treating pain and in-flammation [56, 57]. Leung et al. [56] indicated that aspirinwith caspase inhibitors causes gastric mucosal cell deaththrough a caspase-independent pathway that is indicativeof apoptosis-like programmed cell death. Liu et al. [58]also showed that indomethacin induced oxidative damageto the GI mucosa by downregulating heat shock protein-70 (HSP70) and Bax protein expression instead of sup-pressing the Bcl-2 level in gastric tissue. Epidemiologicstudies indicate that 75 mg/day body weight (b.w.) of di-clofenac treatment increased DNA damage (8-OHdG pro-duction) and decreased HSP70 and caspase-3 expression inthe gastric mucosa of humans [58]. It was suggested thatthe gastroprotective effects of natural antioxidants were due
C© 2016 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.mnf-journal.com

1600659 (6 of 19) Y.-T. Cheng et al. Mol. Nutr. Food Res. 61, 6, 2017, 1600659
to the improvement of antioxidative status, activation ofCOX-mediated PGE2 synthesis, and downregulation of theBax level, as well as upregulation of Bcl-2 and HSP70 pro-teins [58]. Proton pump inhibitors, including lansoprazole,are widely used to treat duodenal and gastric ulcers [59].Some studies have indicated that lansoprazole can reversethe effects of NSAIDs (diclofenac, indomethacin, ketopro-fen, and piroxicam) on the mucosal content of LPO prod-ucts (MDA) and myeloperoxidase (MPO) via upregulationof antioxidant enzymes [59–61]. In addition, the heat shockresponse exhibits defense machinery, and its associated reg-ulatory signals or heat shock proteins are involved in GI pro-tection. HO-1 is known as HSP32 and can be stimulatedduring pathological conditions and oxidative stress [62, 63].Recent evidence indicates that HO-1 breaks down heme tobiliverdin and carbon monoxide, has antioxidant effects, andcan mitigate inflammation, control cell growth, and elimi-nate cell death [32]. The upregulation of HO-1 expressionreduces inflammation and tissue damage caused by ROS inthe GI tract [41]. Megias et al. [64] indicated that HO-1 mod-ulated PGE2 production in osteoarthritic chondrocytes. PGscan maintain GI mucosal integrity, control most gastric mu-cosal defenses, and resolve inflammation [25,65]. Hence, up-regulation of antioxidant enzymes, especially HO-1, shouldbe an effective prevention or curative intervention strategyagainst oxidative stress induced GI damage leading to an ulcerdisease.
A recently published report indicates that dairy prod-ucts with certain probiotics, such as Lactobacillus and Bifi-dobacterium, exhibit prevention against NSAIDs, alcohol, andH. pylori induced GI ulcers and possess antioxidant, anti-apoptotic, anti-inflammatory, and antibacterial activity [66].Moreover, probiotics can accelerate mucosa repair and help tomaintain mucosal health via induction of angiogenic growthfactors by H. pylori [66]. Similarly, a large amount of evi-dence has demonstrated that phytochemicals possess bioac-tivity and potential health benefits. Some reports indicatedthat dietary phenolic compounds (curcumin, mangiferin,and �-myrcene), phenolic acids (caffeic acid and ellagicacid), flavan-3-ols (catechin and (−)-epigallocatechin-3-gallate[EGCG]), flavonols (quercetin and morin), flavonoids (resver-atrol and pogostone), gingerols (6-gingerol and 6-shogaol),carotenes (lycopene and �-carotene), and organosulfur(s-allyl cysteine) possess multiple bioactivity and potentialhealth benefits, such as antioxidant, anti-inflammatory, andantibacterial benefits, and they can prevent digestive diseaseprocesses [35,47,67–70]. In addition, some of the most bene-ficial phytochemicals also appear to increase antioxidant en-zymes, mucosal defense, and the capacity to heal ulcers viaupregulation of the Nrf2 signaling pathway, which may aidmucosal repair [3,22,46,47,71–75]. These results suggest thatNrf2 expression induced by phytochemicals/pharmacologicalmodulators may be a novel means of increasing GI protec-tion. In this review, we summarize the actions of phyto-chemicals and their related mechanisms are also discussed inTable 2.
3.1 Phenolic compounds (curcumin, mangiferin, and
�-myrcene)
3.1.1 Curcumin
Curcumin is a dietary pigment in turmeric (popularly called“curry powder”) and is widely used as a spice and color-ing agent in food [76]. Curcumin possesses multiple healthbenefits and might be a highly efficient antioxidant forthe GI tract [77, 78]. He et al. [79] indicated that orallyadministrated (p.o.) with 20 mg/kg b.w. curcumin for 7days downregulated the acetylation of histone H3 at thesite of the H+ and K+–ATPase gene promoters inducedby water immersion-restraint stress for 4 h. Thong-Ngamet al. [67] indicated that oral administration of curcumin at200 mg/kg b.w. for 30 min decreased the oxidative dam-age to the gastric mucosa, the inflammatory cytokine TNF-�,and leukocyte adhesion of ICAM-1 that were induced by in-domethacin (150 mg/kg b.w. for 8 h, p.o.) in rats. Theseresults suggest that curcumin has preventive and therapeu-tic effects on gastric ulcers by the modulation of gastric acidsecretions and through its antioxidant and anti-inflammatorycapabilities.
3.1.2 Mangiferin
Mangiferin is a natural phenolic glucosylxanthone fromMangifera indica L. (Anacardiaceae) [46, 80]. Reports indi-cate that mangiferin might have highly efficient antioxidant,antimicrobial, antidiabetes, anti-inflammatory, antitumori-genic, and antiulcer activities [80, 81]. Sahoo et al. [81] in-dicated that mangiferin (10 �g/mL) can inhibit TNF-inducedLPO and increase catalase activity, which thereby inhib-ited apoptosis in U937 and HepG2 cell lines. Mangiferin(30 mg/kg b.w. for 3 days, p.o.) has proven to have antigastricacid secretory and antioxidant gastroprotective effects againstabsolute ethanol (96%, 0.2 mL for 30 min) or indomethacin(30 mg/kg b.w. for 8 h) induced gastric ulcers [2, 46]. Mech-anistic studies revealed that mangiferin (10 mg/kg b.w. for3 days, p.o.) stimulation increased PPAR�, nuclear Nrf2, andtotal HO-1 protein expression but decreased the inductionof nuclear NF-�B in the gastric mucosa of rats treated withischemia/reperfusion [46]. These results imply a protectiveeffect of mangiferin on GI ulcers.
3.1.3 �-Myrcene
�-Myrcene (7-methyl-3-methylene-1,6-octadiene) is a com-mon antioxidant found in lemongrass, hops, and verbena andis widely used in cosmetics [82]. Moraes et al. [82] indicatedthat Citrus aurantium and its major compound, �-myrcene,provided significant gastric protection against absolute al-cohol and indomethacin-induced mucosal oxidative injury.Bonamin et al. [83] showed that intragastrically administrated
C© 2016 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.mnf-journal.com

Mol. Nutr. Food Res. 61, 6, 2017, 1600659 (7 of 19) 1600659Ta
ble
2.
Th
ep
rote
ctiv
eef
fect
so
fp
hyto
chem
ical
sag
ain
stri
skfa
cto
rsin
du
ced
gast
roin
test
inal
ulc
erm
od
els
and
thei
ru
nd
erly
ing
mec
han
ism
s
Str
uct
ure
Trea
tmen
tM
ech
anis
mR
efer
ence
Ph
en
olic
co
mp
ou
nd
s
Cu
rcu
min
SD
rats
wer
eo
rally
adm
inis
trat
edw
ith
curc
um
inat
200
mg
/kg
b.w
.fo
r30
min
,fo
llow
edb
ytr
eatm
ent
wit
hin
do
met
hac
in(1
50m
g/k
gb
.w.f
or
8h
,p
.o.)
.
Cu
rcu
min
cou
ldd
ecre
ase
the
oxid
ativ
ed
amag
eo
fga
stri
cm
uco
sab
yd
ecre
asin
gth
ein
flam
mat
ory
cyto
kin
esT
NF-
�an
dle
uko
cyte
adh
esio
no
fIC
AM
-1in
du
ced
by
ind
om
eth
acin
.
[67]
SD
rats
wer
eo
rally
adm
inis
trat
edw
ith
curc
um
inat
20m
g/k
gb
.w.f
or
7d
ays,
follo
wed
by
trea
tmen
tw
ith
wat
erim
mer
sio
n-r
estr
ain
tst
ress
(WIR
S)
for
4h
.
Cu
rcu
min
do
wn
reg
ula
ted
the
acet
ylat
ion
of
his
ton
eH
3at
the
site
of
the
H+
and
K+ -
AT
Pase
gen
ep
rom
ote
rs.
[79]
Man
gif
erin
Sw
iss
alb
ino
mic
eo
rW
ista
rra
tsw
ere
ora
llyad
min
istr
ated
wit
hm
ang
ifer
inat
3,10
,an
d30
mg
/kg
b.w
.fo
r3
day
s,fo
llow
edb
yis
chem
ia/r
eper
fusi
on
(I/R
for
4h
),ab
solu
teet
han
ol(
0.2
mL
for
30m
in),
or
ind
om
eth
acin
(30
mg
/kg
b.w
.fo
r8
h),
resp
ecti
vely
,to
ind
uce
ulc
er.
Man
gif
erin
stim
ula
tio
nin
crea
sed
PPA
R�,n
ucl
ear
Nrf
2,an
dto
talH
O-1
pro
tein
exp
ress
ion
bu
td
ecre
ased
the
ind
uct
ion
of
nu
clea
rN
F-�B
.
[2,4
6]
�-M
yrce
ne
Sw
iss
alb
ino
mic
ew
ere
ora
llyad
min
istr
ated
wit
hes
sen
tial
oil
fro
mfr
uit
pee
lso
fC
itru
sau
ran
tiu
m(1
.43%
myr
cen
e)fo
r0.
5–1
h,f
ollo
wed
by
trea
tmen
tw
ith
abso
lute
alco
ho
l(fo
r1
h,i
.g.)
and
ind
om
eth
acin
(100
mg
/kg
b.w
.fo
r5
h,i
.g.)
.
Ess
enti
alo
ilfr
om
Cit
rus
aura
nti
um
pro
vid
edef
fect
ive
99%
gast
ric
pro
tect
ion
by
mo
du
lati
on
of
PG
syn
thes
is.
[82]
Wis
tar
rats
wer
eo
rally
adm
inis
trat
edw
ith
�-m
yrce
ne
at7.
5m
g/k
gb
.w.f
or
30m
in,f
ollo
wed
by
trea
tmen
tw
ith
ind
om
eth
acin
(50
mg
/kg
b.w
.fo
r6
h,
i.g.)
.
�-M
yrce
ne
cou
ldd
ecre
ase
ind
om
eth
acin
(50
mg
/kg
b.w
.)in
du
ced
-oxi
dat
ive
dam
age
toth
ega
stri
cm
uco
saan
dp
reve
nt
the
dep
leti
on
of
GP
x,G
R,G
SH
,an
dN
Ole
vels
,an
dsu
lfhy
dry
lgro
up
s.
[83]
Ph
en
olic
acid
s
Caf
feic
acid
Hu
man
inte
stin
alIn
t-40
7ce
llsw
ere
trea
ted
wit
hca
ffei
cac
idat
50�
Mfo
r1–
4h
.S
Dra
tsw
aso
rally
adm
inis
trat
edw
ith
caff
eic
acid
at12
0m
g/k
gb
.w.f
or
21d
ays,
follo
wed
by
trea
tmen
tw
ith
keto
pro
fen
(50
mg
/kg
b.w
.fo
r1
day
,i.g
.).
Up
reg
ula
tio
no
fG
Px,
GR
,an
d�-G
CS
anti
oxid
ant
enzy
me
mR
NA
exp
ress
ion
by
DJ-
1/N
rf2
sig
nal
ing
pat
hw
ayin
Int-
407
cells
.D
ow
nre
gu
lati
on
of
MD
A,L
DH
leak
age,
CO
X-2
,NO
,an
d8-
OH
dG
inS
Dra
tsin
du
ced
by
keto
pro
fen
.
[22]
C© 2016 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.mnf-journal.com

1600659 (8 of 19) Y.-T. Cheng et al. Mol. Nutr. Food Res. 61, 6, 2017, 1600659Ta
ble
2.
Co
nti
nu
ed
Str
uct
ure
Trea
tmen
tM
ech
anis
mR
efer
ence
Ella
gic
acid
Sw
iss
alb
ino
mic
ew
ere
ora
llyad
min
istr
ated
wit
hin
do
met
hac
in(1
8m
g/k
gb
.w,s
ing
led
ose
),fo
llow
edb
ytr
eatm
ent
wit
hel
lag
icac
idat
7m
g/k
gb
.w.f
or
3d
ays.
Ella
gic
acid
cou
ldin
hib
itlip
idp
erox
idat
ion
,TN
F-�
,an
dIL
-1�
exp
ress
ion
and
incr
ease
the
exp
ress
ion
of
IL-4
,EG
F,an
dH
GF
inth
ega
stri
cti
ssu
ein
du
ced
by
ind
om
eth
acin
.
[91]
Bal
b/C
mic
ew
ere
trea
ted
wit
h5%
dex
tran
sulf
ate
sod
ium
and
2%el
lag
icac
id(i
nd
rin
kin
gw
ater
)fo
r7
day
s.
Ella
gic
acid
sho
wed
sig
nifi
can
tp
rote
ctio
no
fth
eco
lon
icm
uco
sain
dex
tran
sulf
ate
sod
ium
ind
uce
du
lcer
ativ
eco
litis
inm
ice
and
red
uce
dth
eg
ross
mu
cosa
ldam
age
by
do
wn
reg
ula
tin
gC
OX
-2,i
NO
S,
and
thei
rre
late
dsi
gn
alin
gp
ath
way
s,su
chas
p38
MA
PK
,N
F-�B
,an
dth
esi
gn
altr
ansd
uce
rS
TAT
3w
asal
sob
lock
ed.
[94]
Fla
van
-3-o
l
Cat
ech
inH
um
anin
test
inal
Int-
407
cells
wer
etr
eate
dw
ith
caff
eic
acid
at10
0�
Mfo
r1–
4h
.S
Dra
tsw
ere
ora
llyad
min
istr
ated
wit
hca
tech
inat
35m
g/k
gb
.w.f
or
21d
ays,
follo
wed
by
trea
tmen
tw
ith
keto
pro
fen
(50
mg
/kg
b.w
.fo
r1
day
,i.g
.).
Up
reg
ula
tio
no
fG
Px,
GR
,an
dH
O-1
anti
oxid
ant
enzy
me
by
Nrf
2in
Int-
407
cells
.D
ow
nre
gu
lati
on
of
MD
A,L
DH
leak
age,
and
8-O
Hd
Gin
SD
rats
ind
uce
db
yke
top
rofe
n.
[3]
BA
LB/c
mic
ew
ere
trea
ted
wit
h0.
5m
LH
.p
ylo
rib
acte
rial
susp
ensi
on
(1011
bac
teri
a/L,
i.g.f
or
7d
ays)
,fo
llow
edb
ytr
eatm
ent
wit
hca
tech
inat
320
�g
/0.5
mL
for
7–14
day
s(p
.o.)
.
Cat
ech
ins
hav
esh
ow
np
ote
nti
alan
ti-H
.pyl
ori
acti
vati
on
and
red
uce
du
lcer
atio
nb
yd
ow
nre
gu
lati
ng
the
infl
amm
aso
me/
casp
ase-
1p
ath
way
inm
ice.
[97]
SD
rats
wer
eo
rally
adm
inis
trat
edw
ith
cate
chin
at10
0m
g/k
gb
.w.f
or
30m
in,
follo
wed
by
trea
tmen
tw
ith
1m
LH
Cl-
eth
ano
lso
luti
on
for
1h
(i.g
.).
Cat
ech
insi
gn
ifica
ntl
yin
crea
sed
the
mu
cus
con
ten
tin
SD
rats
.[9
5]
SD
rats
wer
etr
eate
dis
chem
ia/r
eper
fusi
on
(I/R
)fo
r1
h,
follo
wed
by
trea
tmen
tw
ith
cate
chin
at50
mg
/kg
b.w
.fo
r3
day
s(p
.o.)
.
Cat
ech
insi
gn
ifica
ntl
yin
crea
sed
the
leve
lof
cata
lase
and
dec
reas
edth
eox
idat
ive
stre
ssin
rats
that
was
ind
uce
db
yp
ylo
rus
ligat
ure
.
[98]
Fou
rca
tech
ins,
(+)-
cate
chin
,(–
)-ep
icat
ech
in,(
–)-e
pic
atec
hin
galla
te,
and
(–)-
epig
allo
cate
chin
wer
ed
isso
lved
ind
imet
hyls
ulf
oxid
e,an
dth
ein
hib
itio
no
fga
stri
cen
zym
eac
tive
sw
ere
anal
yzed
by
H+
and
K+ -
AT
Pase
,wh
ich
wer
ep
rep
ared
fro
mh
og
sst
om
ach
.
Cat
ech
in(1
70�
M),
epic
atec
hin
(47
�M
),E
CG
(93
�M
),an
dEG
C(0
.11
�M
)co
uld
inh
ibit
50%
acti
vity
of
gast
ric
H+
and
K+ -
AT
Pase
.
[101
]
C© 2016 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.mnf-journal.com
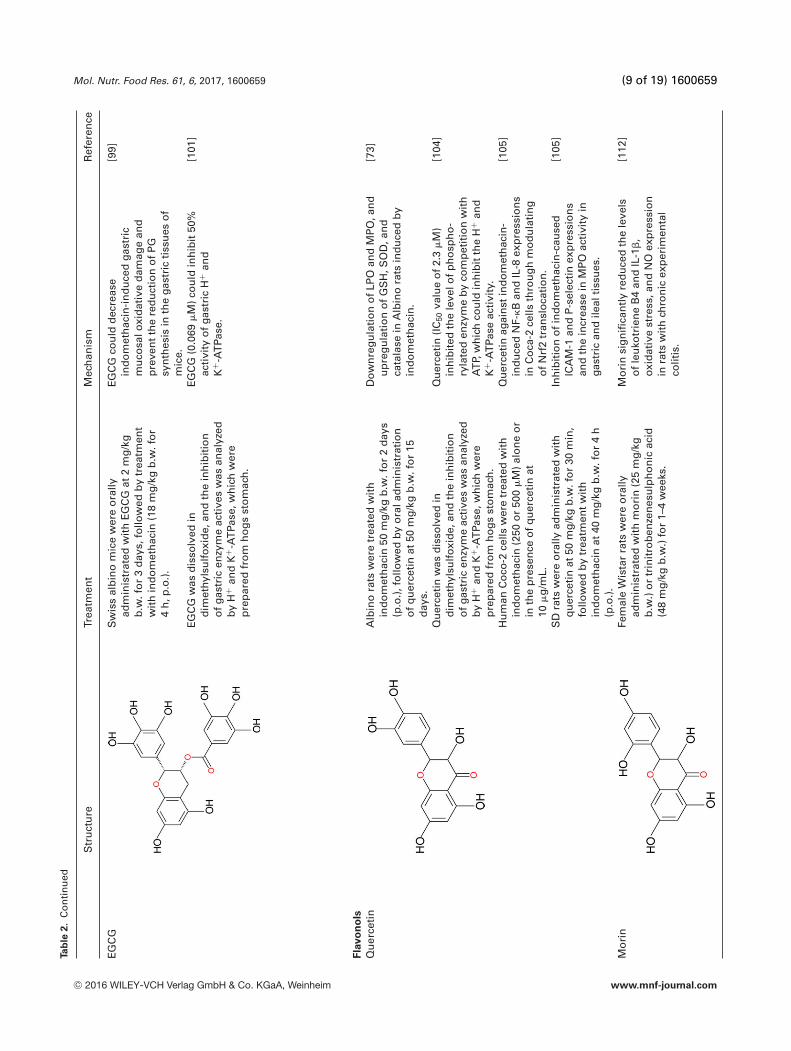
Mol. Nutr. Food Res. 61, 6, 2017, 1600659 (9 of 19) 1600659
Ta
ble
2.
Co
nti
nu
ed
Str
uct
ure
Trea
tmen
tM
ech
anis
mR
efer
ence
EGC
GS
wis
sal
bin
om
ice
wer
eo
rally
adm
inis
trat
edw
ith
EGC
Gat
2m
g/k
gb
.w.f
or
3d
ays,
follo
wed
by
trea
tmen
tw
ith
ind
om
eth
acin
(18
mg
/kg
b.w
.fo
r4
h,p
.o.)
.
EGC
Gco
uld
dec
reas
ein
do
met
hac
in-i
nd
uce
dga
stri
cm
uco
salo
xid
ativ
ed
amag
ean
dp
reve
nt
the
red
uct
ion
of
PG
syn
thes
isin
the
gast
ric
tiss
ues
of
mic
e.
[99]
EGC
Gw
asd
isso
lved
ind
imet
hyls
ulf
oxid
e,an
dth
ein
hib
itio
no
fga
stri
cen
zym
eac
tive
sw
asan
alyz
edb
yH
+an
dK
+ -A
TPa
se,w
hic
hw
ere
pre
par
edfr
om
ho
gs
sto
mac
h.
EGC
G(0
.069
�M
)co
uld
inh
ibit
50%
acti
vity
of
gast
ric
H+
and
K+ -
AT
Pase
.
[101
]
Fla
vo
no
ls
Qu
erce
tin
Alb
ino
rats
wer
etr
eate
dw
ith
ind
om
eth
acin
50m
g/k
gb
.w.f
or
2d
ays
(p.o
.),f
ollo
wed
by
ora
lad
min
istr
atio
no
fq
uer
ceti
nat
50m
g/k
gb
.w.f
or
15d
ays.
Do
wn
reg
ula
tio
no
fLP
Oan
dM
PO
,an
du
pre
gu
lati
on
of
GS
H,S
OD
,an
dca
tala
sein
Alb
ino
rats
ind
uce
db
yin
do
met
hac
in.
[73]
Qu
erce
tin
was
dis
solv
edin
dim
ethy
lsu
lfox
ide,
and
the
inh
ibit
ion
of
gast
ric
enzy
me
acti
ves
was
anal
yzed
by
H+
and
K+ -
AT
Pase
,wh
ich
wer
ep
rep
ared
fro
mh
og
sst
om
ach
.
Qu
erce
tin
(IC
50va
lue
of
2.3
�M
)in
hib
ited
the
leve
lof
ph
osp
ho
-ry
late
den
zym
eb
yco
mp
etit
ion
wit
hA
TP,
wh
ich
cou
ldin
hib
itth
eH
+an
dK
+ -A
TPa
seac
tivi
ty.
[104
]
Hu
man
Co
co-2
cells
wer
etr
eate
dw
ith
ind
om
eth
acin
(250
or
500
�M
)al
on
eo
rin
the
pre
sen
ceo
fq
uer
ceti
nat
10�
g/m
L.
Qu
erce
tin
agai
nst
ind
om
eth
acin
-in
du
ced
NF-
�B
and
IL-8
exp
ress
ion
sin
Co
ca-2
cells
thro
ug
hm
od
ula
tin
go
fN
rf2
tran
slo
cati
on
.
[105
]
SD
rats
wer
eo
rally
adm
inis
trat
edw
ith
qu
erce
tin
at50
mg
/kg
b.w
.fo
r30
min
,fo
llow
edb
ytr
eatm
ent
wit
hin
do
met
hac
inat
40m
g/k
gb
.w.f
or
4h
(p.o
.).
Inh
ibit
ion
of
ind
om
eth
acin
-cau
sed
ICA
M-1
and
P-s
elec
tin
exp
ress
ion
san
dth
ein
crea
sein
MP
Oac
tivi
tyin
gast
ric
and
ileal
tiss
ues
.
[105
]
Mo
rin
Fem
ale
Wis
tar
rats
wer
eo
rally
adm
inis
trat
edw
ith
mo
rin
(25
mg
/kg
b.w
.)o
rtr
init
rob
enze
nes
ulp
ho
nic
acid
(48
mg
/kg
b.w
.)fo
r1–
4w
eeks
.
Mo
rin
sig
nifi
can
tly
red
uce
dth
ele
vels
of
leu
kotr
ien
eB
4an
dIL
-1�
,ox
idat
ive
stre
ss,a
nd
NO
exp
ress
ion
inra
tsw
ith
chro
nic
exp
erim
enta
lco
litis
.
[112
]
C© 2016 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.mnf-journal.com

1600659 (10 of 19) Y.-T. Cheng et al. Mol. Nutr. Food Res. 61, 6, 2017, 1600659
Ta
ble
2.
Co
nti
nu
ed
Str
uct
ure
Trea
tmen
tM
ech
anis
mR
efer
ence
SD
rats
wer
etr
eate
dw
ith
mo
rin
at50
mg
/kg
b.w
.fo
r30
min
(p.o
.),f
ollo
wed
by
trea
tmen
tw
ith
ind
om
eth
acin
at48
mg
/kg
b.w
.fo
r4
h(p
.o.)
.
Mo
rin
pro
tect
sag
ain
stin
do
met
hac
inin
du
ced
-gas
tric
mu
cosa
dam
age
by
do
wn
reg
ula
tio
no
fM
PO
,NF-
�B
,T
NF-
�,M
CP
-1,i
NO
S,I
CA
M-1
,IL-
6,ca
spas
e-3,
and
up
reg
ula
tio
no
fP
GE
2,S
OD
,GS
Tin
the
gast
ric
mu
cosa
.
[35]
Fla
vo
no
ids
Res
vera
tro
lS
Dra
tsw
ere
trea
ted
wit
hre
sver
atro
l(1
00�
mo
l/Lfo
r15
min
,p.o
.),f
ollo
wed
ulc
erin
du
ced
by
isch
emia
for
2h
,an
dre
per
fusi
on
for
4h
.
Res
vera
tro
lco
uld
incr
ease
Na+
-K+ -
AT
Pase
and
Ca2+
-AT
Pase
acti
vity
,an
dth
ele
vels
of
Bcl
-2an
dS
OD
exp
ress
ion
,bu
tit
do
wn
reg
ula
ted
MD
Aan
dLD
Hle
vels
,cas
pas
e-3
acti
vity
,an
dB
axex
pre
ssio
nin
the
gast
ric
mu
cosa
ind
uce
db
yis
chem
ia/r
eper
fusi
on
.
[115
]
Sw
iss
alb
ino
mic
ew
ere
trea
ted
wit
hre
sver
atro
lat2
mg
/kg
b.w
.fo
r4–
7d
ays
(p.o
.),f
ollo
wed
by
trea
tmen
tw
ith
ind
om
eth
acin
(18
mg
/kg
b.w
.,si
ng
leo
rald
ose
).
Trea
tmen
tw
ith
resv
erat
rolc
ou
ldin
hib
iteN
OS
exp
ress
ion
inth
ega
stri
cti
ssu
ein
du
ced
by
ind
om
eth
acin
.
[71]
Ku
nm
ing
mic
ew
ere
ora
llyin
ocu
late
d10
8
CFU
H.p
ylo
rifo
r1
wee
k,fo
llow
edtr
eate
dw
ith
resv
erat
rola
t10
0m
g/k
gb
.w.f
or
6w
eeks
(p.o
.).
Trea
tmen
tw
ith
resv
erat
rolc
ou
ldag
ain
stH
.pyl
ori
ind
uce
dIL
-8,i
NO
S,
and
NF-
�B
exp
ress
ion
by
acti
vati
on
of
the
Nrf
2/H
O-1
pat
hw
ay.
[75]
Pog
ost
on
eS
Dra
tsw
ere
trea
ted
wit
hp
og
ost
on
eat
40m
g/k
gb
.w.f
or
7d
ays
(i.g
.),f
ollo
wed
by
trea
tmen
tw
ith
ind
om
eth
acin
(50
mg
/kg
b.w
.,fo
r1
h,i
.g.)
.
Th
ep
rom
oti
on
of
CO
X-m
edia
ted
PG
E2
and
cellu
lar
anti
oxid
ant
mec
han
ism
by
incr
easi
ng
gast
ric
SO
D,C
AT,
and
GS
Hle
vels
asw
ell
asd
ecre
asin
glip
idp
erox
idat
ion
and
mu
cosa
lap
op
tosi
so
fH
SP
70ac
tiva
tio
naf
ter
ind
uce
db
yin
do
met
hac
in.
[72]
Gin
gero
ls
6-G
ing
ero
lW
ista
rra
tsw
ere
ora
lad
min
istr
atio
no
f6-
gin
ger
ol(
2m
g/k
gb
.w.)
wit
has
pir
in(2
00m
g/k
gb
.w)
for
4h
.
Trea
tmen
to
f6-
gin
ger
olc
ou
ldre
du
ceth
ein
cid
ence
of
mu
cosa
lesi
on
san
dle
vels
of
infl
amm
ato
rycy
toki
ne,
such
asiN
OS
,TN
F-�
,an
dIL
-1�
inth
ega
stri
cm
uco
sa.
[69]
C© 2016 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.mnf-journal.com

Mol. Nutr. Food Res. 61, 6, 2017, 1600659 (11 of 19) 1600659
Ta
ble
2.
Co
nti
nu
ed
Str
uct
ure
Trea
tmen
tM
ech
anis
mR
efer
ence
6-S
ho
gao
lW
ista
rra
tsw
ere
ora
lad
min
istr
atio
no
f6-
sho
gao
l(1
mg
/kg
b.w
.)w
ith
asp
irin
(200
mg
/kg
b.w
)fo
r4
h.
6-S
ho
gao
lco
uld
inh
ibit
iNO
S,T
NF-
�,
and
IL-1
�ex
pre
ssio
nin
the
gast
ric
mu
cosa
ind
uce
db
yas
pir
in(2
00m
g/k
gb
.w.)
.
[69]
Caro
ten
es
Lyco
pen
eW
ista
rra
tsw
ere
trea
ted
wit
hly
cop
ene
(100
mg
/kg
b.w
.fo
r5
min
,p.o
.),
follo
wed
by
trea
tmen
tw
ith
ind
om
eth
acin
(25
mg
/kg
b.w
.fo
r6
h,
p.o
.).
Trea
tmen
to
fly
cop
ene
pri
or
toth
ead
min
istr
atio
no
fin
do
met
hac
in(2
5m
g/k
gb
.w.)
red
uce
dth
ein
cid
ence
of
mu
cosa
lesi
on
san
dle
vels
of
oxid
ativ
est
ress
,su
chas
MD
Aan
dM
PO
inga
stri
cm
uco
sal.
Lyco
pen
eco
uld
enh
ance
SO
D,c
atal
ase,
and
GS
Hex
pre
ssio
nin
the
gast
ric
tiss
ue
ind
uce
db
yin
do
met
hac
in.
[70]
�-C
aro
ten
eW
ista
rra
tsw
ere
trea
ted
wit
h�
-car
ote
ne
(30
or
60m
g/k
gb
.w.f
or
5d
ays,
i.p),
follo
wed
ulc
erin
du
ced
by
isch
emia
for
30m
in,a
nd
rep
erfu
sio
nfo
r3
h.
Pret
reat
men
tw
ith
�-c
aro
ten
ep
rote
cted
the
gast
ric
mu
cosa
agai
nst
isch
emia
-rep
erfu
sio
nin
jury
by
red
uci
ng
IL-1
�,T
NF-
�,a
nd
TG
F-�
inra
ts.
[123
]
Org
an
osu
lfu
r
S-A
llyl
cyst
ein
eC
57B
L/6
mic
ew
ere
trea
ted
wit
hs-
ally
lcy
stei
ne
(30
mg
/kg
b.w
.fo
r1
h,i
.g.)
,fo
llow
edb
ytr
eatm
ent
wit
hin
do
met
hac
in(2
0m
g/k
gb
.w.f
or
24h
,i.g
.).
Dec
reas
edga
stri
cle
sio
ns
ind
uce
db
yin
do
met
hac
in.A
dec
reas
ein
mac
rop
hag
ein
filt
rati
on
;in
crea
sein
the
mu
cus
secr
etio
nan
dH
O-1
anti
oxid
ant
enzy
mes
exp
ress
ion
.
[68]
SD
rats
wer
eo
rally
adm
inis
trat
edw
ith
PM
K-S
005
(asy
nth
etic
s-al
lyl-
cyst
ein
e)at
5m
g/k
gb
.w.f
or
1h
,fo
llow
edb
ytr
eatm
ent
wit
has
pir
in(2
00m
g/k
gb
.w.
for
4h
,i.g
.),d
iclo
fen
ac(8
0m
g/k
gb
.w.
for
4h
,i.g
.),o
rin
do
met
hac
in(4
0m
g/k
gb
.w.f
or
4h
,i.g
.).
Asy
nth
etic
s-al
lyl-
L-cy
stei
ne
cou
ldag
ain
stN
SA
IDs
(asp
irin
,dic
lofe
nac
,an
din
do
met
hac
in)
ind
uce
dac
ute
gast
ric
dam
age
by
do
wn
reg
ula
tin
gp
roin
flam
mat
ory
cyto
kin
es,s
uch
ascP
LA2,
CO
X-2
,an
dLT
B4
exp
ress
ion
,an
din
crea
sin
gth
esy
nth
esis
of
mu
cus
inra
ts.
[18]
CA
T,ca
tala
se;
cPLA
2,cy
toso
licp
ho
sph
olip
ases
A2;
LDH
,la
ctat
ed
ehyd
rog
enas
e;LT
B4,
leu
kotr
ien
eB
4;G
ST,
glu
tath
ion
eS
-tra
nsf
eras
e;H
GF,
hep
ato
cyte
gro
wth
fact
or;
EGC
,(−
)-ep
igal
loca
tech
in;E
CG
,(−)
-ep
icat
ech
in-3
-gal
late
.
C© 2016 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.mnf-journal.com

1600659 (12 of 19) Y.-T. Cheng et al. Mol. Nutr. Food Res. 61, 6, 2017, 1600659
(i.g.) with 7.5 mg/kg b.w. �-myrcene for 30 min could reduceindomethacin-induced (50 mg/kg b.w. for 6 h, i.g.) oxidativedamage to the gastric mucosa and prevent the depletion ofGPx, GR, GSH, NO, and sulfhydryl groups.
3.2 Phenolic acids (caffeic acid and ellagic acid)
3.2.1 Caffeic acid
Caffeic acid and protocatechuic acid (PCA) are common phy-tochemical compounds found in fruits, vegetables, grains,and traditional Chinese herbs that possess many beneficialeffects, including anticancer, antioxidant, antihypertension,and anti-inflammatory effects [84–87]. Vitaglione et al. [85]indicated that in humans, PCA is primarily produced as ametabolite of anthocyanins. The report indicated that the fateof anthocyanins in humans was that less than 1% of intakewas absorbed. Oral administration of PCA has shown highbioavailability and bioefficacy in rats [85], which might mod-erately decrease the oxidative stress in the GI tract. However,this hypothesis may require more evidence. Similarly, caffeicacid was distributed to the plasma in an intact form becauseof its easy absorption in the small intestinal mucosa [88, 89].Therefore, it is more suitable for investigating GI protection.Cheng et al. [22] reported that caffeic acid at 50 �M for 1–4 hstrongly induced GPx activity in Int-407 cells, which sug-gests that caffeic acid is important in preventing GI oxida-tive damage. Caffeic acid also attenuated excessive COX-2protein expression and NO generation in lipopolysaccharide-induced RAW 264.7 cells, which suggests that caffeic acidprevents a form of inflammation-associated GI damage [90].Moreover, the pretreatment of caffeic acid at 120 mg/kg b.w.for 21 days (p.o.) effectively inhibited ketoprofen-induced(50 mg/kg b.w. for 1 day, p.o.) oxidative damage of theGI mucosa by inducing the expression of antioxidant en-zymes and modulating the DJ-1/Nrf2 pathway [22]. There-fore, caffeic acid is a potential curative compound that canbe used for treating GI mucosal injuries caused by oxidativestress.
3.2.2 Ellagic acid
Ellagic acid (2,3,7,8-tetrahydroxy[1] benzopyrano[5,4,3-cde][1]benzopyran-5,10-dione) is one of the naturally occurringpolyphenols found in cranberries, Indian gooseberries,pecans, pomegranates, raspberries, strawberries, walnuts,and other plant foods and is found mainly in the form ofellagitannins [91]. Some reports have indicated the antipro-liferative and antioxidant activities of ellagic acid in vitro andin vivo [92]. Ellagic acid has been shown to exert a potentscavenging effect on both hydroxy anions and superoxideanions. Moreover, the rats were orally administrated withellagic acid (7 mg/kg b.w., p.o.) for 3 days could inhibitLPO, TNF-�, and IL-1� expression, and increased the ex-
pression of IL-4, EGF, and hepatocyte growth factor in gas-tric tissue induced by indomethacin (18 mg/kg b.w., singledose, p.o.) [91, 93]. Moreover, diets supplemented with 2%ellagic acid for 7 days showed significant protection of thecolonic mucosa in dextran sulfate sodium induced (1 and5% DSS) ulcerative colitis in mice and reduced the grossmucosal damage by downregulating COX-2, inducible ni-tric oxide synthase (iNOS), and their related signaling path-ways, such as p38 MAPK, NF-�B, and the signal transducerand activator of transcription 3 (STAT3) was also blocked[94].
3.3 Flavan-3-ol (catechin and EGCG)
Catechin derivatives, such as EGCG, (−)-epigallocatechin,(−)-epicatechin-3-gallate, and (−)-epicatechin, have beenwidely investigated and provide a potential therapeutic ap-proach for oxidative stress associated GI ulcer diseases [3,95].EGCG and catechins are stable under acidic conditions andmay be more suitable for studying GI protection [96]. More-over, these catechins at 320 �g/0.5 mL for 7 days (p.o.) haveshown potential anti-H. pylori activation and reduced ulcer-ation by downregulating the inflammasome/caspase-1 path-way in mice [97]. Jung et al. [95] indicated that pretreatmentwith catechin at 100 mg/kg b.w. for 30 min followed by HCl-ethanol for 1 h significantly increased the mucus content inrats. Rao et al. [98] demonstrated that catechin at 50 mg/kgb.w. for 3 days significantly increased the level of catalase anddecreased the oxidative stress in rats that was induced by py-lorus ligature. Cheng et al. [3] showed that supplementationwith catechin (100 �M for 1–4 h) appeared to increase theantioxidant capacity of Int-407 cells, not only through its ownnatural antioxidant activity, but also by stimulating the expres-sion of antioxidants and detoxifying enzymes such as HO-1[3]. Pretreatment of SD rats with catechin (35 mg/kg b.w. for21 days) could decrease ketoprofen-induced (50 mg/kg b.w.for 1 day) gastric mucosal oxidative damage and could pre-vent the reduction of GPx, GRd antioxidant enzymes, and theGSH/GSSG ratio in the intestinal mucosa [3]. Similarly, oraladministration of EGCG (2 mg/kg b.w. for 3 days) could de-crease indomethacin-induced (18 mg/kg b.w. for 4 h) gastricmucosal oxidative damage and prevented the reduction of PGsynthesis in the gastric tissues of mice [99]. The administra-tion of EGCG at 40 and 80 mg/kg/day for 6 days reducedthe formation of cholesterol gallstones in a dose-dependentmanner in a C57BL/6 mouse model by downregulating theactivity of NF-�B and upregulating the activity of PPAR�
[100]. Furthermore, catechin (170 �M), epicatechin (47�M), (−)-epicatechin-3-gallate (93 �M), (−)-epigallocatechin(0.11 �M), and EGCG (0.069 �M) could inhibit 50% activ-ity of gastric H+, K+-ATPase, and EGCG and showed themost inhibitory activity in vitro [101]. These findings suggestthat the antisecretory and antiulcerogenic effects of catechinsare due to their inhibitory activity on gastric H+ and K+−
ATPase.
C© 2016 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.mnf-journal.com

Mol. Nutr. Food Res. 61, 6, 2017, 1600659 (13 of 19) 1600659
3.4 Flavonols (quercetin and morin)
3.4.1 Quercetin
Quercetin (3,5,7,3′,4′-pentahydroxy flavone) is a flavonewidely found in fruits and vegetables that has health-promoting and disease-preventing activity [13]. The dailyintake of this phenolic compound is approximately 5–40mg/day in a healthy diet [102]. Quercetin is one of the mostpotent scavengers of free radicals and is more active thanthe well-known antioxidant vitamins C and E. Its antioxi-dant and anti-inflammatory activities are contributing fac-tors to its therapeutic efficacy for peptic ulcers [102, 103].Abourehab et al. [73] indicated that the oral administration ofquercetin at 50 mg/kg b.w for 15 days significantly downreg-ulated the levels of LPO and MPO but upregulated the levelsof GSH, SOD, and catalase in Albino rats treated with in-domethacin (30 mg/kg b.w. for 2 days, p.o.). Murakami et al.[104] indicated that quercetin (IC50 value of 2.3 �M) inhib-ited the levels of phosphorylated enzyme by competition withATP, which could inhibit the H+ and K+–ATPase activity invitro. Carrasco-Pozo et al. [105] demonstrated that quercetin(10 �g/mL) against indomethacin (250 or 500 �M) inducedinflammation of NF-�B and IL-8 expressions in Coca-2cells through modulating of Nrf2 translocation. Moreover,quercetin (50 mg/kg b.w. for 30 min, p.o.) could inhibit in-domethacin (40 mg/kg b.w. for 4 h, p.o.)-induced ICAM-1and P-selectin expressions and MPO activity in gastric andileal tissue [105].
3.4.2 Morin
Morin (2-(2,4-dihydroxyphenyl)-3,5,7-trihydroxy-4H-1-benzopyran-4-one) is found in guava (Psidium guajava), almond(Prunus dulcis), old fustic (Maclura tinctoria), and osage or-ange (Maclura pomifera) [106, 107]. It possesses many ben-eficial characteristics and might be a highly efficient an-tioxidant, anti-inflammatory, and xanthine oxidase inhibitoryagent [108–111]. Galvez et al. [112] indicated that oral ad-ministration of morin (25 mg/kg b.w.) for 4 weeks signif-icantly reduced the levels of leukotriene B4 and IL-1�, ox-idative stress, and NO expression in rats with chronic ex-perimental colitis. Sinha et al. [35] also indicated that ratpretreatment with morin (50 mg/kg b.w. for 30 min, p.o.)protected against indomethacin-induced (48 mg/kg b.w. for4 h) gastric mucosal damage by downregulating MPO, NF-�B,TNF-�, MCP-1, iNOS, ICAM-1, IL-6, and caspase-3, as wellas upregulating PGE2, SOD, and glutathione S-transferasein the gastric mucosa. These findings suggest that morinhas a beneficial anti-inflammatory effect on the intestinalmucosa.
3.5 Flavonoids (resveratrol and pogostone)
3.5.1 Resveratrol
Resveratrol (3,5,4′-trihydroxystilbene), which belongs to thegroup of stilbenes, is a natural polyphenol originally iso-lated from white hellebore and is present in berries, grapes,and peanuts [113, 114]. There are various natural resveratrol-related analogs in plants. The compound 3,5,4′-trimethoxy-trans-stilbene (MR-3) is methoxylated instead of hydroxy-lated at positions 3, 5, and 4′ in resveratrol. The compound3,5,3′,4′,5′-pentamethoxystilbene (MR-5), a methoxy deriva-tive of resveratrol, can be synthesized by artificial methods[114]. Shen et al. [115] indicated that oral administration ofresveratrol at 100 �mol/L for 15 min could increase Na+-K+-ATPase and Ca2+-ATPase activity and the levels of Bcl-2 andSOD expression, but it downregulated MDA and lactate dehy-drogenase levels, caspase-3 activity, and Bax expression in thegastric mucosa induced by ischemia/reperfusion. Dey et al.[71] indicated that rat treatment with resveratrol (2 mg/kgb.w. for 4–7 days, p.o.) inhibited eNOS expression in the gas-tric tissue induced by indomethacin (18 mg/kg b.w., singleoral dose). Zhang et al. [75] also indicated that mice treatedwith resveratrol (100 mg/kg b.w. for 6 weeks) could againstH. pylori (108 CFU)-induced IL-8, iNOS, and NF-�B expres-sions through activation of the Nrf2/HO-1 pathway in gastrictissue.
3.5.2 Pogostone
Pogostemonis Herba is widely used to treat GI diseases withtraditional Chinese herbs, and the major constituent of pogo-stone indicates the occurrence of multiple biological activ-ities such as antioxidant and anti-inflammatory activities[116, 117]. Some reports also indicate that the oral adminis-tration of pogostone was highly absorbed and bioavailable invivo. The mechanisms of pogostone are potentially associatedwith the increase in COX-mediated PGE2 and cellular antiox-idant mechanisms by increasing gastric SOD, catalase, andGSH levels as well as decreasing LPO and mucosal apoptosisand upregulation of HSP70 activation in vivo [72]. Moreover,Chen et al. showed that pogostone (40 mg/kg b.w. for 7 day,i.g.) exerted a gastroprotective effect against indomethacin-induced (50 mg/kg b.w. for 1 h, i.g.) gastric injury in SD rats[72]. These findings suggest that the upregulation of HSPmight work against oxidative stress associated GI ulcers.
3.6 Gingerols (6-gingerol and 6-shogaol)
Ginger (Zingiber officinale), a natural dietary rhizome, iswidely used as a flavoring agent, as a traditional medic-inal herb, and is shown to have multibiological func-tions [118, 119]. Shogaols are dehydrated products of thestructurally related gingerols and are rich in fresh and
C© 2016 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.mnf-journal.com
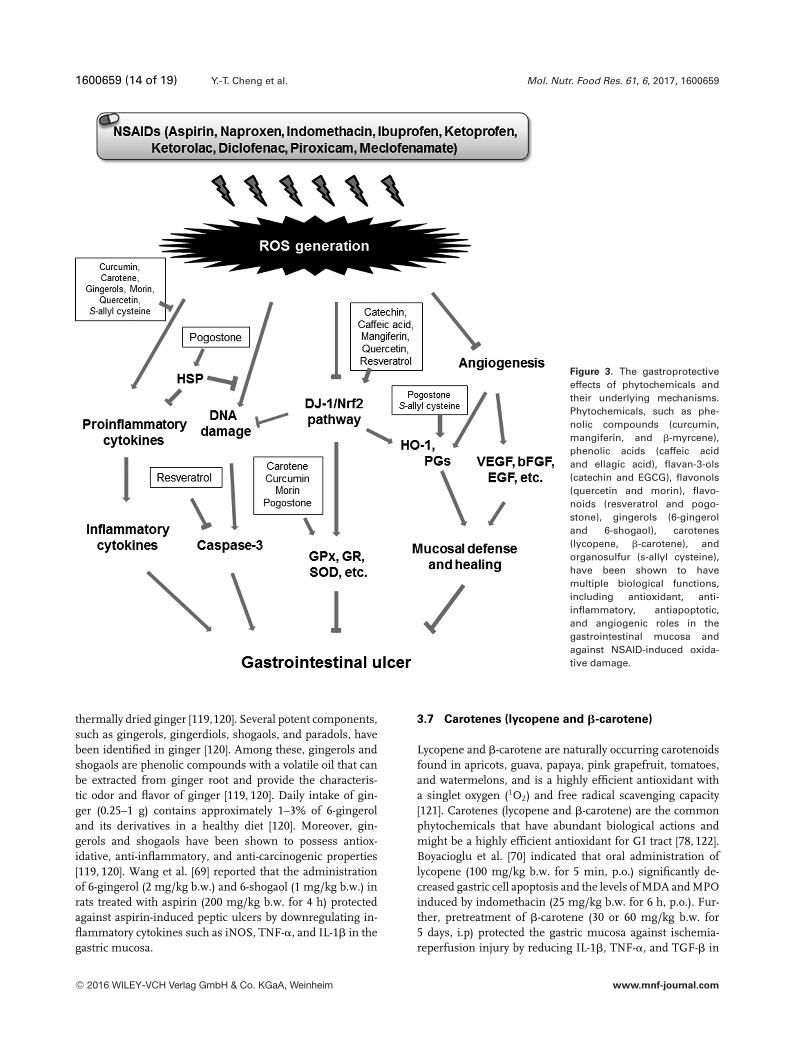
1600659 (14 of 19) Y.-T. Cheng et al. Mol. Nutr. Food Res. 61, 6, 2017, 1600659
Figure 3. The gastroprotectiveeffects of phytochemicals andtheir underlying mechanisms.Phytochemicals, such as phe-nolic compounds (curcumin,mangiferin, and �-myrcene),phenolic acids (caffeic acidand ellagic acid), flavan-3-ols(catechin and EGCG), flavonols(quercetin and morin), flavo-noids (resveratrol and pogo-stone), gingerols (6-gingeroland 6-shogaol), carotenes(lycopene, �-carotene), andorganosulfur (s-allyl cysteine),have been shown to havemultiple biological functions,including antioxidant, anti-inflammatory, antiapoptotic,and angiogenic roles in thegastrointestinal mucosa andagainst NSAID-induced oxida-tive damage.
thermally dried ginger [119,120]. Several potent components,such as gingerols, gingerdiols, shogaols, and paradols, havebeen identified in ginger [120]. Among these, gingerols andshogaols are phenolic compounds with a volatile oil that canbe extracted from ginger root and provide the characteris-tic odor and flavor of ginger [119, 120]. Daily intake of gin-ger (0.25–1 g) contains approximately 1–3% of 6-gingeroland its derivatives in a healthy diet [120]. Moreover, gin-gerols and shogaols have been shown to possess antiox-idative, anti-inflammatory, and anti-carcinogenic properties[119, 120]. Wang et al. [69] reported that the administrationof 6-gingerol (2 mg/kg b.w.) and 6-shogaol (1 mg/kg b.w.) inrats treated with aspirin (200 mg/kg b.w. for 4 h) protectedagainst aspirin-induced peptic ulcers by downregulating in-flammatory cytokines such as iNOS, TNF-�, and IL-1� in thegastric mucosa.
3.7 Carotenes (lycopene and �-carotene)
Lycopene and �-carotene are naturally occurring carotenoidsfound in apricots, guava, papaya, pink grapefruit, tomatoes,and watermelons, and is a highly efficient antioxidant witha singlet oxygen (1O2) and free radical scavenging capacity[121]. Carotenes (lycopene and �-carotene) are the commonphytochemicals that have abundant biological actions andmight be a highly efficient antioxidant for GI tract [78, 122].Boyacioglu et al. [70] indicated that oral administration oflycopene (100 mg/kg b.w. for 5 min, p.o.) significantly de-creased gastric cell apoptosis and the levels of MDA and MPOinduced by indomethacin (25 mg/kg b.w. for 6 h, p.o.). Fur-ther, pretreatment of �-carotene (30 or 60 mg/kg b.w. for5 days, i.p) protected the gastric mucosa against ischemia-reperfusion injury by reducing IL-1�, TNF-�, and TGF-� in
C© 2016 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.mnf-journal.com

Mol. Nutr. Food Res. 61, 6, 2017, 1600659 (15 of 19) 1600659
rats [123]. These findings suggest that protective roles of ly-copene and carotene are performed by regulating antioxidantand anti-inflammatory effects in the gastric mucosa.
3.8 Organosulfur (s-allyl cysteine)
S-Allyl cysteine enriched in garlic is one of the naturalorganosulfuric compounds that are beneficial to humanhealth. Recently, s-allyl cysteine has been reported to pos-sess antioxidant and anti-inflammatory effects in vitro andin vivo. Park et al. [68] indicated that s-allyl cysteine (30mg/kg b.w. for 1 h, i.g.) alleviated indomethacin (20 mg/kgb.w. for 24 h) induced gastric mucosal damage by inhibi-tion of COX-2 and upregulation of HO-1 expression. More-over, s-allyl cysteine decreased macrophage infiltration butincreased the mucus secretion and total antioxidant activityin C57BL/6 mice [68]. Choi et al. [18] also indicated that oraladministration of PMK-S005 (a synthetic s-allyl-L-cysteine) at5 mg/kg b.w. for 1 h against NSAIDs, such as aspirin (200mg/kg b.w. for 4 h, i.g.), diclofenac (80 mg/kg b.w. for 4 h,i.g.), and indomethacin (40 mg/kg b.w. for 4 h, i.g.), inducedacute gastric damage by downregulating proinflammatory cy-tokines (cytosolic phospholipases A2, COX-2, and leukotrieneB4) expression, and increasing the synthesis of mucus inrats.
4 Conclusions
Epidemiological reports have shown that increased dietary in-take of phytochemicals are beneficial for GI health. In this re-view, we summarized the protective effects of phytochemicalsand their underlying mechanisms against NSAID-inducedGI ulcers following oxidative stress. The current review con-firms that dietary phytochemicals possess protective effectsand curative potential for peptic ulcers by improving angio-genesis, cytoprotection and re-epithelialization, upregulatingtissue growth factors and PGs, suppressing oxidative damageof the mucosa, increasing endogenous mucosal defensiveagents, and blocking oxidative stress associated gastroduo-denal inflammation and ulceration, as illustrated in Fig. 3.Notably, some of the most beneficial phytochemicals, suchas catechins, caffeic acid, mangiferin, pogostone, quercetin,and resveratrol appear to antiulcer properties through upreg-ulation of Nrf2 signaling pathways, which might play a keyrole to control oxidative stress and prevent cell death for GItract. These results suggest the Nrf2-mediated antioxidant en-zymes expression by phytochemicals/pharmacological mod-ulators, which may be a novel means of increasing GI pro-tection. These updated data confirm the well-established di-verse beneficial biological actions of these phytochemicalsand might support their therapeutic use, as illustrated inTable 2.
This research work was supported in part by the Ministry ofEducation, Taiwan under the ATU plan.
The authors have declared no conflict of interest.
5 References
[1] Smith, J. W., Mathis, T., Benns, M. V., Franklin, G. A. et al.,Socioeconomic disparities in the operative managementof peptic ulcer disease. Surgery 2013, 154, 672–678; discus-sion 678–679.
[2] Carvalho, A. C., Guedes, M. M., de Souza, A. L., Trevisan,M. T. et al., Gastroprotective effect of mangiferin, a xan-thonoid from Mangifera indica, against gastric injury in-duced by ethanol and indomethacin in rodents. Planta Med.2007, 73, 1372–1376.
[3] Cheng, Y. T., Wu, C. H., Ho, C. Y., Yen, G. C., Catechin protectsagainst ketoprofen-induced oxidative damage of the gastricmucosa by up-regulating Nrf2 in vitro and in vivo. J. Nutr.Biochem. 2013, 24, 475–483.
[4] Mota, K. S., Dias, G. E., Pinto, M. E., Luiz-Ferreira, A. et al.,Flavonoids with gastroprotective activity. Molecules 2009,14, 979–1012.
[5] Musumba, C., Pritchard, D. M., Pirmohamed, M., Review ar-ticle: cellular and molecular mechanisms of NSAID-inducedpeptic ulcers. Aliment. Pharmacol. Ther. 2009, 30, 517–531.
[6] Rainsford, K. D., Profile and mechanisms of gastrointesti-nal and other side effects of nonsteroidal anti-inflammatorydrugs (NSAIDs). Am. J. Med. 1999, 107, 27S–35S; discus-sion 35S–36S.
[7] Slomiany, B. L., Piotrowski, J., Slomiany, A., Omepra-zole fails to suppress up-regulation of gastric mucosalendothelin-converting enzyme-1 by Helicobacter pylorilipopolysaccharide. J. Physiol. Pharmacol. 2000, 51, 421–431.
[8] van der Vliet, A., Bast, A., Role of reactive oxygen speciesin intestinal diseases. Free. Radic. Biol. Med. 1992, 12, 499–513.
[9] Kuo, B., Bhasin, M., Jacquart, J., Scult, M. A. et al., Genomicand clinical effects associated with a relaxation responsemind-body intervention in patients with irritable bowel syn-drome and inflammatory bowel disease. PLoS One 2015,10, e0123861.
[10] Suleyman, H., Albayrak, A., Bilici, M., Cadirci, E. et al.,Different mechanisms in formation and prevention ofindomethacin-induced gastric ulcers. Inflammation 2010,33, 224–234.
[11] Indo, H. P., Matsui, H., Chen, J., Zhu, H. et al., Manganese su-peroxide dismutase promotes interaction of actin, S100A4and Talin, and enhances rat gastric tumor cell invasion. J.Clin. Biochem. Nutr. 2015, 57, 13–20.
[12] Saija, A., Uccella, N., Olive biophenols functional effects onhuman wellbeing. Trends Food Sci. 2001, 11, 357–363.
[13] Farzaei, M. H., Abdollahi, M., Rahimi, R., Role of dietarypolyphenols in the management of peptic ulcer. World J.Gastroenterol. 2015, 21, 6499–6517.
C© 2016 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.mnf-journal.com

1600659 (16 of 19) Y.-T. Cheng et al. Mol. Nutr. Food Res. 61, 6, 2017, 1600659
[14] Melcarne, L., Garcia-Iglesias, P., Calvet, X., Managementof NSAID-associated peptic ulcer disease. Expert Rev. Gas-troenterol. Hepatol. 2016, 10, 723–733.
[15] Marjoribanks, J., Ayeleke, R. O., Farquhar, C., Proctor, M.,Nonsteroidal anti-inflammatory drugs for dysmenorrhoea.Cochrane Database Syst. Rev. 2015, 7, CD001751.
[16] Holland, S., Silberstein, S. D., Freitag, F., Dodick, D. W. et al.,Evidence-based guideline update: NSAIDs and other com-plementary treatments for episodic migraine prevention inadults: report of the Quality Standards Subcommittee ofthe American Academy of Neurology and the AmericanHeadache Society. Neurology 2012, 78, 1346–1353.
[17] Falcao Hde, S., Maia, G. L., Bonamin, F., Kushima, H. et al.,Gastroprotective mechanisms of the chloroform and ethylacetate phases of Praxelis clematidea (Griseb.) R.M.King &H.Robinson (Asteraceae). J. Nat. Med. 2013, 67, 480–491.
[18] Choi, Y. J., Kim, N., Lee, J. Y., Nam, R. H. et al., Protectiveeffects of garlic extract, PMK-S005, against nonsteroidalanti-inflammatory drugs-induced acute gastric damage inrats. Dig. Dis. Sci. 2014, 59, 2927–2934.
[19] Chatterjee, S., Chatterjee, A., Roy, S., Bera, B. et al., L-Theanine healed NSAID-induced gastric ulcer by modulat-ing pro/antioxidant balance in gastric ulcer margin. J. Nat.Med. 2014, 68, 699–708.
[20] Lim, J. H., Kim, J. H., Kim, N., Lee, B. H. et al., Gastro-protective effect of Cochinchina momordica seed extract innonsteroidal anti-inflammatory drug-induced acute gastricdamage in a rat model. Gut Liver 2014, 8, 49–57.
[21] Shimoyama, A. T., Santin, J. R., Machado, I. D., de Oliveirae Silva, A. M. et al., Antiulcerogenic activity of chlorogenicacid in different models of gastric ulcer. Naunyn Schmiede-bergs Arch. Pharmacol. 2013, 386, 5–14.
[22] Cheng, Y. T., Ho, C. Y., Jhang, J. J., Lu, C. C. et al., DJ-1plays an important role in caffeic acid-mediated protectionof the gastrointestinal mucosa against ketoprofen-inducedoxidative damage. J. Nutr. Biochem. 2014, 25, 1045–1057.
[23] Firdaus, S. B., Ghosh, D., Chattyopadhyay, A., Jana, J. et al.,A combination of aqueous curry (Murraya koenigii) leafextract and melatonin protects against piroxicam inducedgastric ulcer in male albino rats: involvement of antioxidantmechanism(s). J. Pharm. Res. 2014, 8, 428–436.
[24] Halabi, M. F., Shakir, R. M., Bardi, D. A., Al-Wajeeh, N. S.et al., Gastroprotective activity of ethyl-4-[(3,5-di-tert-butyl-2-hydroxybenzylidene) amino]benzoate against ethanol-induced gastric mucosal ulcer in rats. PLoS One 2014, 9,e95908.
[25] Tarnawski, A. S., Ahluwalia, A., Molecular mechanisms ofepithelial regeneration and neovascularization during heal-ing of gastric and esophageal ulcers. Curr. Med. Chem.2012, 19, 16–27.
[26] Shibuya, A., Onda, K., Kawahara, H., Uchiyama, Y. et al.,Sofalcone, a gastric mucosa protective agent, increasesvascular endothelial growth factor via the Nrf2-heme-oxygenase-1 dependent pathway in gastric epithelial cells.Biochem. Biophys. Res. Commun. 2010, 398, 581–584.
[27] Stancova, V., Zikova, A., Svobodova, Z., Kloas, W., Effects ofthe non-steroidal anti-inflammatory drug(NSAID) naproxen
on gene expression of antioxidant enzymes in zebrafish(Danio rerio). Environ. Toxicol. Pharmacol. 2015, 40, 343–348.
[28] Iloki-Assanga, S. B., Lewis-Lujan, L. M., Fernandez-Angulo,D., Gil-Salido, A. A. et al., Retino-protective effect of Bu-cida buceras against oxidative stress induced by H2O2 inhuman retinal pigment epithelial cells line. BMC Comple-ment. Altern. Med. 2015, 15, 254.
[29] Brigelius-Flohe, R., Tissue-specific functions of individualglutathione peroxidases. Free. Radic. Biol. Med. 1999, 27,951–965.
[30] Banning, A., Deubel, S., Kluth, D., Zhou, Z. et al., The GI-GPxgene is a target for Nrf2. Mol. Cell Biol. 2005, 25, 4914–4923.
[31] Bansal, A. K., Bilaspuri, G. S., Oxidative stress alters mem-brane sulfhydryl status, lipid and phospholipid contents ofcrossbred cattle bull spermatozoa. Anim. Reprod. Sci. 2008,104, 398–404.
[32] Kirkby, K. A., Adin, C. A., Products of heme oxygenase andtheir potential therapeutic applications. Am. J. Physiol. Re-nal. Physiol. 2006, 290, F563–F571.
[33] Guo, X., Shin, V. Y., Cho, C. H., Modulation of heme oxy-genase in tissue injury and its implication in protectionagainst gastrointestinal diseases. Life Sci. 2001, 69, 3113–3119.
[34] Husain, K., Mejia, J., Lalla, J., Kazim, S., Dose response ofalcohol-induced changes in BP, nitric oxide and antioxidantsin rat plasma. Pharmacol. Res. 2005, 51, 337–343.
[35] Sinha, K., Sadhukhan, P., Saha, S., Pal, P. B. et al., Morin pro-tects gastric mucosa from nonsteroidal anti-inflammatorydrug, indomethacin induced inflammatory damage andapoptosis by modulating NF-kappaB pathway. Biochim.Biophys. Acta 2015, 1850, 769–783.
[36] Duvillard, L., Ortega-Deballon, P., Bourredjem, A., Scherrer,M. L. et al., A case-control study of pre-operative levels ofserum neutrophil gelatinase-associated lipocalin and otherpotential inflammatory markers in colorectal cancer. BMCCancer 2014, 14, 912.
[37] Singh, R. G., Rathore, S. S., Wani, I. A., Agrawal, A. et al.,Effects of Salacia oblonga on cardiovascular risk factors inchronic kidney disease patients: a prospective study. SaudiJ. Kidney Dis. Transpl. 2015, 26, 61–66.
[38] Sadhu, A., Upadhyay, P., Agrawal, A., Ilango, K. et al., Man-agement of cognitive determinants in senile dementia ofAlzheimer’s type: therapeutic potential of a novel poly-herbal drug product. Clin. Drug Investig. 2014, 34, 857–869.
[39] Hawkey, C. J., COX-1 and COX-2 inhibitors. Best Pract. Res.Clin. Gastroenterol. 2001, 15, 801–820.
[40] Adhikary, B., Yadav, S. K., Bandyopadhyay, S. K., Chat-topadhyay, S., Role of the COX-independent pathways inthe ulcer-healing action of epigallocatechin gallate. FoodFunct. 2011, 2, 338–347.
[41] Liu, S., Mizu, H., Yamauchi, H., Photoinflammatoryresponses to UV-irradiated ketoprofen mediated bythe induction of ROS generation, enhancement ofcyclooxygenase-2 expression, and regulation of multiplesignaling pathways. Free Radic. Biol. Med. 2011, 48, 772–780.
C© 2016 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.mnf-journal.com

Mol. Nutr. Food Res. 61, 6, 2017, 1600659 (17 of 19) 1600659
[42] Chakraborty, S., Stalin, S., Das, N., Choudhury, S. T. et al.,The use of nano-quercetin to arrest mitochondrial damageand MMP-9 upregulation during prevention of gastric in-flammation induced by ethanol in rat. Biomaterials 2012,33, 2991–3001.
[43] Deng, X., Xiong, X., Khomenko, T., Sandor, Z. et al., Inap-propriate angiogenic response as a novel mechanism ofduodenal ulceration and impaired healing. Dig. Dis. Sci.2011, 56, 2792–2801.
[44] Taupin, D., Podolsky, D. K., Trefoil factors: initiators ofmucosal healing. Nat. Rev. Mol. Cell Biol. 2003, 4, 721–732.
[45] Amagase, K., Inaba, A., Senta, T., Ishikawa, Y. et al., Gastriculcerogenic and healing impairment effects of risedronate,a nitrogen-containing bisphosphonate in rats. Comparisonwith alendronate and minodronate. J. Physiol. Pharmacol.2012, 62, 609–618.
[46] Mahmoud-Awny, M., Attia, A. S., Abd-Ellah, M. F., El-Abhar, H. S., Mangiferin mitigates gastric ulcer in ischemia/reperfused rats: involvement of PPAR-gamma, NF-kappaBand Nrf2/HO-1 signaling pathways. PLoS One 2015, 10,e0132497.
[47] Surh, Y. J., Cancer chemoprevention with dietary phyto-chemicals. Nat. Rev. Cancer 2003, 3, 768–780.
[48] Ho, C. Y., Cheng, Y. T., Chau, C. F., Yen, G. C., Effect of dial-lyl sulfide on in vitro and in vivo Nrf2-mediated pulmonicantioxidant enzyme expression via activation ERK/p38 sig-naling pathway. J. Agric. Food Chem. 2012, 60, 100–107.
[49] Ci, X., Lv, H., Wang, L., Wang, X. et al., The antioxidative po-tential of farrerol occurs via the activation of Nrf2 mediatedHO-1 signaling in RAW 264.7 cells. Chem. Biol. Interact.2015, 239, 192–199.
[50] Yeh, C. T., Chiu, H. F., Yen, G. C., Protective effect of sul-foraphane on indomethacin-induced cytotoxicity via hemeoxygenase-1 expression in human intestinal Int 407 cells.Mol. Nutr. Food Res. 2009, 53, 1166–1176.
[51] van Horssen, J., Drexhage, J. A., Flor, T., Gerritsen, W. et al.,Nrf2 and DJ1 are consistently upregulated in inflammatorymultiple sclerosis lesions. Free Radic. Biol. Med. 2010, 49,1283–1289.
[52] Lee, E., Yin, Z., Sidoryk-Wegrzynowicz, M., Jiang, H.et al., 15-Deoxy-delta12,14-prostaglandin J(2) modulatesmanganese-induced activation of the NF-kappaB, Nrf2, andPI3K pathways in astrocytes. Free Radic. Biol. Med. 2012,52, 1067–1074.
[53] Jaramillo-Gomez, J., Nino, A., Arboleda, H., Arboleda,G., Overexpression of DJ-1 protects against C2-ceramide-induced neuronal death through activation of the PI3K/AKTpathway and inhibition of autophagy. Neurosci. Lett. 2015,603, 71–76.
[54] Taira, T., Saito, Y., Niki, T., Iguchi-Ariga, S. M. et al., DJ-1 hasa role in antioxidative stress to prevent cell death. EMBORep. 2004, 5, 213–218.
[55] Odabasoglu, F., Halici, Z., Cakir, A., Halici, M. et al., Benefi-cial effects of vegetable oils (corn, olive and sunflower oils)and alpha-tocopherol on anti-inflammatory and gastroin-
testinal profiles of indomethacin in rats. Eur. J. Pharmacol.2008, 591, 300–306.
[56] Leung, A. M., Redlak, M. J., Miller, T. A., Aspirin-inducedmucosal cell death in human gastric cells: role of a caspase-independent mechanism. Dig. Dis. Sci. 2009, 54, 28–35.
[57] Ramirez-Alcantara, V., LoGuidice, A., Boelsterli, U. A., Pro-tection from diclofenac-induced small intestinal injury bythe JNK inhibitor SP600125 in a mouse model of NSAID-associated enteropathy. Am. J. Physiol. Gastrointest. LiverPhysiol. 2009, 297, G990–G998.
[58] Liu, Y. H., Zhang, Z. B., Zheng, Y. F., Chen, H. M. et al.,Gastroprotective effect of andrographolide sodium bisul-fite against indomethacin-induced gastric ulceration in rats.Int. Immunopharmacol. 2015, 26, 384–391.
[59] Chandranath, S. I., Bastaki, S. M., Singh, J., A compara-tive study on the activity of lansoprazole, omeprazole andPD-136450 on acidified ethanol- and indomethacin-inducedgastric lesions in the rat. Clin. Exp. Pharmacol. Physiol.2002, 29, 173–180.
[60] Fornai, M., Natale, G., Colucci, R., Tuccori, M. et al.,Mechanisms of protection by pantoprazole against NSAID-induced gastric mucosal damage. Naunyn SchmiedebergsArch. Pharmacol. 2005, 372, 79–87.
[61] Blandizzi, C., Fornai, M., Colucci, R., Natale, G. et al., Lan-soprazole prevents experimental gastric injury induced bynon-steroidal anti-inflammatory drugs through a reduc-tion of mucosal oxidative damage. World J. Gastroenterol.2005, 11, 4052–4060.
[62] Sikora, A., Grzesiuk, E., Heat shock response in gastroin-testinal tract. J. Physiol. Pharmacol. 2007, 58(Suppl 3), 43–62.
[63] Yoda, Y., Amagase, K., Kato, S., Tokioka, S. et al., Preventionby lansoprazole, a proton pump inhibitor, of indomethacin -induced small intestinal ulceration in rats through inductionof heme oxygenase-1. J. Physiol. Pharmacol. 2010, 61, 287–294.
[64] Megias, J., Guillen, M. I., Clerigues, V., Rojo, A. I.et al., Heme oxygenase-1 induction modulates microsomalprostaglandin E synthase-1 expression and prostaglandinE(2) production in osteoarthritic chondrocytes. Biochem.Pharmacol. 2009, 77, 1806–1813.
[65] Wallace, J. L., Prostaglandins, NSAIDs, and gastric mucosalprotection: why doesn’t the stomach digest itself? Physiol.Rev. 2008, 88, 1547–1565.
[66] Khoder, G., Al-Menhali, A. A., Al-Yassir, F., Karam, S. M.,Potential role of probiotics in the management of gastriculcer. Exp. Ther. Med. 2016, 12, 3–17.
[67] Thong-Ngam, D., Choochuai, S., Patumraj, S., Chayanu-patkul, M. et al., Curcumin prevents indomethacin-inducedgastropathy in rats. World J. Gastroenterol. 2012, 18, 1479–1484.
[68] Park, J. M., Han, Y. M., Kangwan, N., Lee, S. Y. et al.,S-allyl cysteine alleviates nonsteroidal anti-inflammatorydrug-induced gastric mucosal damages by increasingcyclooxygenase-2 inhibition, heme oxygenase-1 induction,and histone deacetylation inhibition. J. Gastroenterol. Hep-atol. 2014, 29(Suppl 4), 80–92.
C© 2016 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.mnf-journal.com

1600659 (18 of 19) Y.-T. Cheng et al. Mol. Nutr. Food Res. 61, 6, 2017, 1600659
[69] Wang, Z., Hasegawa, J., Wang, X., Matsuda, A. et al., Protec-tive effects of ginger against aspirin-induced gastric ulcersin rats. Yonago Acta Med. 2011, 54, 11–19.
[70] Boyacioglu, M., Kum, C., Sekkin, S., Yalinkilinc, H. S. et al.,The effects of lycopene on DNA damage and oxidativestress on indomethacin-induced gastric ulcer in rats. Clin.Nutr. 2016, 35, 428–435.
[71] Dey, A., Guha, P., Chattopadhyay, S., Bandyopadhyay, S. K.,Biphasic activity of resveratrol on indomethacin-inducedgastric ulcers. Biochem. Biophys. Res. Commun. 2009, 381,90–95.
[72] Chen, X. Y., Chen, H. M., Liu, Y. H., Zhang, Z. B. et al., Thegastroprotective effect of pogostone from PogostemonisHerba against indomethacin-induced gastric ulcer in rats.Exp. Biol. Med. (Maywood) 2016, 241, 193–204.
[73] Abourehab, M. A., Khaled, K. A., Sarhan, H. A., Ahmed, O.A., Evaluation of combined famotidine with quercetin forthe treatment of peptic ulcer: in vivo animal study. DrugDes. Devel. Ther. 2015, 9, 2159–2169.
[74] Sun, C. Y., Xu, L. Q., Zhang, Z. B., Chen, C. H. et al., Protectiveeffects of pogostone against LPS-induced acute lung injuryin mice via regulation of Keap1-Nrf2/NF-kappaB signalingpathways. Int. Immunopharmacol. 2016, 32, 55–61.
[75] Zhang, X., Jiang, A., Qi, B., Ma, Z. et al., Resveratrol Protectsagainst Helicobacter pylori-associated gastritis by combat-ing oxidative stress. Int. J. Mol. Sci. 2015, 16, 27757–27769.
[76] Weng, C. J., Yen, G. C., Chemopreventive effects of dietaryphytochemicals against cancer invasion and metastasis:phenolic acids, monophenol, polyphenol, and their deriva-tives. Cancer Treat. Rev. 2012, 38, 76–87.
[77] Ghosh, S., Banerjee, S., Sil, P. C., The beneficial role of cur-cumin on inflammation, diabetes and neurodegenerativedisease: a recent update. Food. Chem. Toxicol. 2015, 83,111–124.
[78] Kim, M. J., Kim, H., Anticancer effect of lycopene in gastriccarcinogenesis. J. Cancer Prev. 2015, 20, 92–96.
[79] He, P., Zhou, R., Hu, G., Liu, Z. et al., Curcumin-induced his-tone acetylation inhibition improves stress-induced gastriculcer disease in rats. Mol. Med. Rep. 2015, 11, 1911–1916.
[80] Rajendran, P., Rengarajan, T., Nishigaki, I., Ekambaram, G.et al., Potent chemopreventive effect of mangiferin on lungcarcinogenesis in experimental Swiss albino mice. J. Can-cer Res. Ther. 2015, 10, 1033–1039.
[81] Sahoo, B. K., Zaidi, A. H., Gupta, P., Mokhamatam, R. B.et al., A natural xanthone increases catalase activity butdecreases NF-kappa B and lipid peroxidation in U-937 andHepG2 cell lines. Eur. J. Pharmacol. 2015, 764, 520–528.
[82] Moraes, T. M., Kushima, H., Moleiro, F. C., Santos, R. C.et al., Effects of limonene and essential oil from Citrus au-rantium on gastric mucosa: role of prostaglandins and gas-tric mucus secretion. Chem. Biol. Interact. 2009, 180, 499–505.
[83] Bonamin, F., Moraes, T. M., Dos Santos, R. C., Kushima, H.et al., The effect of a minor constituent of essential oil fromCitrus aurantium: the role of beta-myrcene in preventingpeptic ulcer disease. Chem. Biol. Interact. 2014, 212, 11–19.
[84] Jiang, R. W., Lau, K. M., Hon, P. M., Mak, T. C. et al., Chem-istry and biological activities of caffeic acid derivatives fromSalvia miltiorrhiza. Curr. Med. Chem. 2005, 12, 237–246.
[85] Vitaglione, P., Donnarumma, G., Napolitano, A., Galvano, F.et al., Protocatechuic acid is the major human metaboliteof cyanidin-glucosides. J. Nutr. 2007, 137, 2043–2048.
[86] Mehrotra, A., Shanbhag, R., Chamallamudi, M. R., Singh,V. P. et al., Ameliorative effect of caffeic acid against in-flammatory pain in rodents. Eur. J. Pharmacol. 2011, 666,80–86.
[87] Kono, Y., Kobayashi, K., Tagawa, S., Adachi, K. et al., An-tioxidant activity of polyphenolics in diets. Rate constantsof reactions of chlorogenic acid and caffeic acid with re-active species of oxygen and nitrogen. Biochim. Biophys.Acta 1997, 1335, 335–342.
[88] Ye, Z., Hong, C. O., Lee, K., Hostetter, J. et al., Plasma caffeicacid is associated with statistical clustering of the anticoliticefficacy of caffeic acid in dextran sulfate sodium-treatedmice. J. Nutr. 2011, 141, 1989–1995.
[89] Yeh, C. T., Huang, W. H., Yen, G. C., Antihypertensive effectsof Hsian-tsao and its active compound in spontaneouslyhypertensive rats. J. Nutr. Biochem. 2009, 20, 866–875.
[90] Wang, B. S., Yen, G. C., Chang, L. W., Yen, W. J. et al., Protec-tive effects of burdock (Arctium lappa Linne) on oxidation oflow-density lipoprotein and oxidative stress in RAW 264.7macrophages. Food Chem. 2007, 101, 729–738.
[91] Chatterjee, A., Chatterjee, S., Das, S., Saha, A. et al., Ellagicacid facilitates indomethacin-induced gastric ulcer heal-ing via COX-2 up-regulation. Acta Biochim. Biophys. Sin.(Shanghai) 2012, 44, 565–576.
[92] Solon, S., Lopes, L., Teixeira de Sousa, P., Jr., Schmeda-Hirschmann, G., Free radical scavenging activity of Lafoen-sia pacari. J. Ethnopharmacol. 2000, 72, 173–178.
[93] Iino, T., Nakahara, K., Miki, W., Kiso, Y. et al., Less damagingeffect of whisky in rat stomachs in comparison with pureethanol. Role of ellagic acid, the nonalcoholic component.Digestion 2001, 64, 214–221.
[94] Marin, M., Maria Giner, R., Rios, J. L., Recio, M. C., Intestinalanti-inflammatory activity of ellagic acid in the acute andchronic dextrane sulfate sodium models of mice colitis. J.Ethnopharmacol. 2013, 150, 925–934.
[95] Jung, J., Bae, K. H., Jeong, C. S., Anti-Helicobacter pyloriand antiulcerogenic activities of the root cortex of Paeoniasuffruticosa. Biol. Pharm. Bull. 2013, 36, 1535–1539.
[96] Jang, J. H., Park, Y. D., Ahn, H. K., Kim, S. J. et al., Analysisof green tea compounds and their stability in dentifrices ofdifferent pH levels. Chem. Pharm. Bull. (Tokyo) 2014, 62,328–335.
[97] Yang, J. C., Yang, H. C., Shun, C. T., Wang, T. H. et al., Cate-chins and sialic acid attenuate Helicobacter pylori-triggeredepithelial caspase-1 activity and eradicate Helicobacter py-lori infection. Evid. Based Complement Alternat. Med. 2013,2013, 248585.
[98] Rao Ch, V., Vijayakumar, M., Protective effect of (+)-catechinagainst gastric mucosal injury induced by ischaemia-reperfusion in rats. J. Pharm. Pharmacol. 2007, 59, 1103–1107.
C© 2016 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.mnf-journal.com

Mol. Nutr. Food Res. 61, 6, 2017, 1600659 (19 of 19) 1600659
[99] Adhikary, B., Yadav, S. K., Bandyopadhyay, S. K., Chat-topadhyay, S., Epigallocatechin gallate accelerates healingof indomethacin-induced stomach ulcers in mice. Pharma-col. Rep. 2011, 63, 527–536.
[100] Shan, D., Fang, Y., Ye, Y., Liu, J., EGCG reducing the sus-ceptibility to cholesterol gallstone formation through theregulation of inflammation. Biomed. Pharmacother. 2008,62, 677–683.
[101] Murakami, S., Muramatsu, M., Otomo, S., Gastric H+, K(+)-ATPase inhibition by catechins. J. Pharm. Pharmacol. 1992,44, 926–928.
[102] Russo, M., Spagnuolo, C., Tedesco, I., Bilotto, S. et al.,The flavonoid quercetin in disease prevention and therapy:facts and fancies. Biochem. Pharmacol. 2012, 83, 6–15.
[103] Butkovic, V., Klasinc, L., Bors, W., Kinetic study of flavonoidreactions with stable radicals. J. Agric. Food Chem. 2004,52, 2816–2820.
[104] Murakami, S., Muramatsu, M., Otomo, S., Inhibition of gas-tric H+, K(+)-ATPase by quercetin. J. Enzyme Inhib. 1992,5, 293–298.
[105] Carrasco-Pozo, C., Castillo, R. L., Beltran, C., Miranda, A.et al., Molecular mechanisms of gastrointestinal protectionby quercetin against indomethacin-induced damage: roleof NF-kappaB and Nrf2. J. Nutr. Biochem. 2016, 27, 289–298.
[106] Wijeratne, S. S., Abou-Zaid, M. M., Shahidi, F., Antioxidantpolyphenols in almond and its coproducts. J. Agric. FoodChem. 2006, 54, 312–318.
[107] Aggarwal, B. B., Shishodia, S., Molecular targets of dietaryagents for prevention and therapy of cancer. Biochem. Phar-macol. 2006, 71, 1397–1421.
[108] Xie, M. X., Long, M., Liu, Y., Qin, C. et al., Characterization ofthe interaction between human serum albumin and morin.Biochim. Biophys. Acta 2006, 1760, 1184–1191.
[109] Hanasaki, Y., Ogawa, S., Fukui, S., The correlation betweenactive oxygens scavenging and antioxidative effects offlavonoids. Free Radic. Biol. Med. 1994, 16, 845–850.
[110] Fang, S. H., Hou, Y. C., Chang, W. C., Hsiu, S. L. et al.,Morin sulfates/glucuronides exert anti-inflammatory activ-ity on activated macrophages and decreased the incidenceof septic shock. Life Sci. 2003, 74, 743–756.
[111] Yu, Z., Fong, W. P., Cheng, C. H., The dual actions of morin(3,5,7,2′,4′-pentahydroxyflavone) as a hypouricemic agent:uricosuric effect and xanthine oxidase inhibitory activity. J.Pharmacol. Exp. Ther. 2006, 316, 169–175.
[112] Galvez, J., Coelho, G., Crespo, M. E., Cruz, T. et al., Intestinalanti-inflammatory activity of morin on chronic experimen-
tal colitis in the rat. Aliment. Pharmacol. Ther. 2001, 15,2027–2039.
[113] Baur, J. A., Sinclair, D. A., Therapeutic potential of resver-atrol: the in vivo evidence. Nat. Rev. Drug Discov. 2006, 5,493–506.
[114] Weng, C. J., Wu, C. F., Huang, H. W., Wu, C. H. et al.,Evaluation of anti-invasion effect of resveratrol and relatedmethoxy analogues on human hepatocarcinoma cells. J.Agric. Food Chem. 2010, 58, 2886–2894.
[115] Shen, M., Wu, R. X., Zhao, L., Li, J. et al., Resveratrol at-tenuates ischemia/reperfusion injury in neonatal cardiomy-ocytes and its underlying mechanism. PLoS One 2012, 7,e51223.
[116] Li, Y., Su, Z., Lin, S., Li, C. et al., Characterisation of themetabolism of pogostone in vitro and in vivo using liq-uid chromatography with mass spectrometry. Phytochem.Anal. 2014, 25, 97–105.
[117] Verma, S., Kumar, V. L., Attenuation of gastric mucosaldamage by artesunate in rat: modulation of oxidative stressand NFkappaB mediated signaling. Chem. Biol. Interact.2016, 257, 46–53.
[118] Lantz, R. C., Chen, G. J., Sarihan, M., Solyom, A. M. et al.,The effect of extracts from ginger rhizome on inflammatorymediator production. Phytomedicine 2007, 14, 123–128.
[119] Weng, C. J., Chou, C. P., Ho, C. T., Yen, G. C., Molecular mech-anism inhibiting human hepatocarcinoma cell invasion by6-shogaol and 6-gingerol. Mol. Nutr. Food Res. 2012, 56,1304–1314.
[120] O’Hara, M., Kiefer, D., Farrell, K., Kemper, K., A review of 12commonly used medicinal herbs. Arch. Fam. Med. 1998, 7,523–536.
[121] Luxsuwong, D., Indranupakorn, R., Wongtrakul, P., Prepa-ration of vesicles entrapped lycopene extract. J. Oleo Sci.2014, 63, 645–652.
[122] Jung, S., Wu, K., Giovannucci, E., Spiegelman, D. et al.,Carotenoid intake and risk of colorectal adenomas in a co-hort of male health professionals. Cancer Causes Control2013, 24, 705–717.
[123] Seyyed Ali, M., Neisi, N., Darbor, M., Hassanpour, M. et al.,�-Carotene protects the gastric mucosa against ischemia-reperfusion injury in rats. Eur. Cytokine Netw. 2012, 23,173–178.
[124] Kim, S. J., Lee, H. J., Kim, B. S., Lee, D. et al., An-tiulcer activity of anthocyanins from Rubus coreanusvia association with regulation of the activity of matrixmetalloproteinase-2. J. Agric. Food Chem. 2011, 59, 11786–11793.
C© 2016 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.mnf-journal.com

本文献由“学霸图书馆-文献云下载”收集自网络,仅供学习交流使用。
学霸图书馆(www.xuebalib.com)是一个“整合众多图书馆数据库资源,
提供一站式文献检索和下载服务”的24 小时在线不限IP
图书馆。
图书馆致力于便利、促进学习与科研,提供最强文献下载服务。
图书馆导航:
图书馆首页 文献云下载 图书馆入口 外文数据库大全 疑难文献辅助工具







