
1
RN927C, a site-specific Trop-2 antibody-drug-conjugate (ADC) with enhanced stability, is
highly efficacious in preclinical solid tumor models
Pavel Strop1,5,*, Thomas-Toan Tran1,6,*, Magdalena Dorywalska1, Kathy Delaria1,8, Russell
Dushin2, Oi Kwan Wong1, Wei-Hsien Ho1, Dahui Zhou2, Aidong Wu3, Eugenia Kraynov3, Laura
Aschenbrenner4, Bora Han4,9, Christopher J. O’Donnell2, Jaume Pons1,7, Arvind Rajpal1,5, Dave
L. Shelton1 and Shu-Hui Liu1
1Oncology-Rinat R&D, Pfizer Inc., 230 E Grand Ave, South San Francisco, CA 94080 USA
2Worldwide Medicinal Chemistry, Pfizer Inc., 445 Eastern Point Rd, Groton, CT 06340 USA
3Pharmacokinetics, Dynamics & Metabolism, 4Drug Safety R&D, Pfizer Inc., 10646 Science
Center Dr, San Diego, CA 92121 USA
5Current Address: Bristol-Myers Squibb, 700 Bay Rd Ste A, Redwood City, CA 94063 USA
6Current Address: NGM Biopharmaceuticals, Inc. 630 Gateway Blvd, South San Francisco, CA
94080 USA
7Current Address: Alexo Therapeutics, 951 Gateway Blvd Ste 201, South San Francisco, CA
94080 USA
8Current Address: Grifols Diagnostic Solutions, 6455 Christie Ave B-334C, Emeryville, CA
9Current Address: Adheren Inc. 5858 Horton St Ste 255, Emeryville, CA 94608 USA
*Pavel Strop and Thomas-Toan Tran contributed equally to this article
Corresponding author: Shu-Hui Liu, Email: [email protected] TEL: 650-615-7462
Running title: Site-specific Trop-2 ADC with enhanced stability
Key Words: Trop-2, ADC, site-specific
The authors disclose no potential conflicts of interest
on May 14, 2018. © 2016 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2016; DOI: 10.1158/1535-7163.MCT-16-0431

2
Abstract
Trop-2, also known as TACSTD2, EGP-1, GA733-1 and M1S1, is frequently expressed
on a variety of human carcinomas and its expression is often associated with poor prognosis of
the diseases. However, it is also present on the epithelium of several normal tissues. A
comprehensively designed Trop-2-targeting ADC, balancing both efficacy and toxicity, is
therefore necessary to achieve clinical utility. To this end, we developed a cleavable Trop-2-
ADC (RN927C) using a site-specific transglutaminase-mediated conjugation method and a
proprietary MTI (microtubule inhibitor) linker-payload, PF-06380101. Robust in vitro
cytotoxicity of RN927C was observed on a panel of Trop-2 expressing tumor cell lines, with IC50
generally in the subnanomolar range. As expected for an MTI-containing ADC, RN927C readily
induced mitotic arrest of treated cells in vitro and in vivo, followed by subsequent cell death.
The in vivo efficacy of RN927C was tested in multiple cell line and patient-derived xenograft
tumor models including pancreatic, lung, ovarian and triple negative breast tumor types. Single
dose administration of RN927C at 0.75 - 3 mg/kg was generally sufficient to induce sustained
regression of Trop-2 expressing tumors and showed superior efficacy over standard treatment
with paclitaxel or gemcitabine. Administration of RN927C in non-human primate toxicity
studies resulted in target mediated effects in skin and oral mucosa, consistent with Trop-2
expression in these epithelial tissues with minimal, non-dose limiting off-target toxicities. Based
on the combined efficacy and safety results, RN927C is postulated to have a favorable
therapeutic index for treatment of solid tumors.
Introduction
on May 14, 2018. © 2016 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2016; DOI: 10.1158/1535-7163.MCT-16-0431

3
Antibody-drug conjugates (ADCs) were developed to improve the therapeutic indices of
cytotoxic anti-cancer agents. The approach makes use of an immunoconjugate in which a
cytotoxic agent is chemically or enzymatically linked to an antibody that selectively binds to an
internalizing tumor-associated antigen (1-3). This strategy allows specific delivery of the
cytotoxic agent to the tumor site while minimizing the exposure to normal tissues. Trop-2
(Trophoblast cell-surface antigen 2), also referred to as M1S1, GA733-1 (gastric antigen 733-1),
EGP-1 (epithelial glycoprotein-1), or TACSTD2 (tumor-associated calcium signal transducer 2),
is a type I transmembrane cell surface glycoprotein originally identified in human placental
trophoblasts (4) and subsequently found to be highly expressed in most human carcinomas.
Although the biological role of Trop-2 is still under investigation (5), various studies have shown
that overexpression of Trop-2 correlates with increased cancer growth (6, 7), tumor
aggressiveness, metastasis, and poor prognosis in various human carcinomas (8-14). Studies
have also shown that Trop-2 contributes to tumor pathogenesis at least in part by activating the
ERK1/2 MAPK pathway which has important implications in cancer cell proliferation,
migration, invasion, and survival (15). Regulated proteolysis of Trop-2 is suggested to drive
epithelial hyperplasia and stem cell self-renewal via β-catenin signaling (16). Other studies,
however, have indicated that loss of Trop-2 can also contribute to tumorigenesis depending on
the cell type and context (17-19).
Immunohistochemistry (IHC) analysis on human tumor samples has demonstrated
expression of Trop-2 protein in multiple tumor types including pancreatic (8), ovarian (9),
endometrial (10), non-small cell lung (11), colon (12), gastric (13) and oral cancers (14). The
association of Trop-2 expression with advanced disease and/or clinical outcome in multiple
tumor types, and the relatively restricted expression in normal adult tissues, underscore the
on May 14, 2018. © 2016 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2016; DOI: 10.1158/1535-7163.MCT-16-0431

4
potential benefit of targeting Trop-2 to fill an unmet need in cancer treatment. An anti-Trop-2
SN38 conjugate, hRS7-CL2A-SN-38 (IMMU-132), consisting of a humanized Trop-2 antibody
conjugated to SN38, the active component of irinotecan, has been shown to be efficacious in
several epithelial cancer cell line xenograft models (20-22). IMMU-132 (sacituzumab
govitecan) is currently being tested in clinical trials for solid tumors and has reported
encouraging therapeutic activity in patients with difficult to treat cancers (23). The safety profile
of IMMU-132 is similar to irinotecan with neutropenia and diarrhea being the most commonly
observed side effects (23).
ADCs synthesized by conjugating to reduced cysteine sulfhydryl groups or lysine side
chain amines yield heterogeneous conjugates that have been associated with aggregation,
increased toxicity and inconsistent pharmacokinetics (24-26). One approach that can potentially
overcome these limitations is to employ site-specific conjugation of antibodies and many
experimental approaches have been developed including conjugation through engineered
cysteine residues, non-natural amino acids, antibody carbohydrates, aldehyde tags, and utilizing
enzymes such as sortases and phosphopantetheinyl transferases (2, 27-36). A site-specifically
conjugated anti-CD33 ADC, SGN-CD33A, has shown encouraging activities in targeting drug-
resistant acute myeloid leukemia (AML) (37). We have previously disclosed the unique qualities
of conjugating antibodies through an enzymatic method employing microbial transglutaminase
(30). Here we describe the design and utility of a new Trop-2-ADC, RN927C, comprised of a
humanized anti-Trop-2 hIgG1 antibody, conjugated with the AcLys-VC-PABC-PF-06380101
linker payload at the C-terminus of the antibody heavy chain via an enzymatic process. This
conjugate contains a valine-citruline cleavable linker and PF-06380101 (38), which is a potent
anti-mitotic agent that inhibits tubulin polymerization. Upon binding to the extracellular portion
on May 14, 2018. © 2016 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2016; DOI: 10.1158/1535-7163.MCT-16-0431

5
of Trop-2 on the cell surface, RN927C is internalized and traffics to lysosomes. After being
processed by the lysosomal proteases, the payload PF-06380101 is released and induces cell
cycle arrest resulting in cell death. Indeed, RN927C showed potent in vitro cell killing activity
in tumor cell lines with wide range of Trop-2 expression levels. In vivo testing of RN927C in
cell line and PDX models representing several solid tumor types confirmed robust activity often
inducing tumor regression with a single dose. The AcLys-VC-PABC linker in RN927C is
shown to be stable in the bloodstream from preclinical mouse PK studies and is expected to
provide an enhanced off-target safety profile (30, 39). In addition, our site-specific conjugation
technology produces highly homogeneous conjugates (30, 40) and has the potential to minimize
unwanted PK behavior and toxicity often associated with highly loaded ADCs (24, 27).
Materials and Methods
Generation of RN927C
The antibody portion of RN927C is derived from immunization of Balb/c mice using
recombinant Trop-2 extracellular domain fused with human Fc. The mouse antibody was then
humanized by CDR grafting/affinity maturation and cloned into human IgG1/k constant domains
to create the parental antibody. The C-terminal lysine on the heavy chain of the humanized anti-
Trop-2 antibody was replaced with the LLQGA tag using standard molecular biology techniques.
The DNA encoding this antibody was cloned into an in-house expression plasmid, transiently
expressed in HEK293 cells, and then purified with Protein-A MabSelect SuRe columns (GE
Healthcare). For AcLys-VC-PABC-PF-06380101 conjugation, the antibody concentration was
adjusted to 5 mg/mL in buffer containing 25 mM Tris-HCl, pH 8.0, 150 mM sodium chloride.
Linker-payload (40) was added in a 10-fold molar excess over antibody, the conjugation reaction
on May 14, 2018. © 2016 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2016; DOI: 10.1158/1535-7163.MCT-16-0431

6
was initiated by addition of 2% (w/v) bacterial transglutaminase (Ajinomoto Activa TI, Japan)
and incubated with gentle shaking at 37°C for 16 hours. The reaction mixture was adjusted to
0.75 M ammonium sulfate, 25 mM potassium phosphate, pH 7 (Buffer A), and the material was
applied to a Butyl HP column (Butyl HP, GE Healthcare), washed with 5 column volumes of
Buffer A, and eluted with a linear gradient over 20 column volumes into 25 mM potassium
phosphate, pH 7. Fractions containing the conjugate were pooled, dialyzed into 1x PBS,
concentrated using a 10 kDa Amicon Ultra centrifugal filter unit (Millipore), and 0.2 µm sterile
filtered. The final product DAR (Drug-Antibody-Ratio) was 2.0 as determined by HIC and mass
spectrometry analysis.
Cell lines and Reagents
A431, Fadu, OVCAR3, BxPC3, Calu-3, NCI-H292, NCI-H1650, HCC-827, MDA-MB-468,
Colo205, SKBR3 and SW620 were all purchased from American Type Culture Collection
(ATCC) in 2008. Cell lines were tested for Trop-2 expression using immunofluorescence and
FACS. No further authentication was performed. Gemcitabine (GEMZAR) was purchased from
Eli Lily and company. Paclitaxel was purchased from Sigma-Aldrich. Receptor quantitation
was performed using Quantum Simply Cellular kits (Bangs Laboratories, Inc.) according to
manufacturer’s instructions.
Cytotoxicity assays
Human tumor cells from ATCC were cultured in DMEM + 10% FBS (fetal bovine serum)
(A431, Fadu, OVCAR3, SKBR3, MDA-MB-468 and SW620) or RPMI + 10% FBS (BxPC3,
NCI-H1650, NCI-H292, Calu-3, HCC-827 and Colo205). Cells were seeded at 2000 cells/well
(A431, Fadu, MDA-MB-468, SW620, BxPC3, NCI-H1650, NCI-H292, Calu-3, HCC-827 and
on May 14, 2018. © 2016 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2016; DOI: 10.1158/1535-7163.MCT-16-0431

7
Colo205) or 3000 cells/well (OVCAR3 and SKBR3) in growth media on white 96 well plates on
day 0. RN927C or negative control ADC were added to each well on day 1 in the dilution range
of 0.0006 - 40 μg/mL final concentration (equivalent to 0.004 - 267 nM). Free PF-06380101(38)
was tested in the final concentration range of 0.0006-40 nM. CelltiterGlo viability assays
(Promega) were performed 4 days after treatment. Luminescence signals were detected on the
M5 plate reader (Molecular Devices). All the readings were normalized to % of value derived
from control untreated wells. IC50s were calculated by logistic non-linear regression (GraphPad
Prism 6.03) and reported as the concentration of antibody (nM) that reduced cell viability by
50%.
Internalization assay
HCC-827 cells were cultured until nearly confluent, harvested and re-suspended in 5 ml of cold
binding buffer (DMEM + 10% FBS + 10 mM HEPES) at 1.6 million cells/ml. Primary antibody
(RN927C or parental Ab) was labeled with Dylight 800 antibody labeling kit per manufacturer’s
protocol (Thermo Scientific). The labeled antibody was then added to the cells to a final
concentration of 2 μg/ml and incubated at 4oC for 1 hour. To initiate internalization, cells were
washed and re-suspended in 5 ml pre-warmed binding buffer supplemented with 2 μg/ml
unlabeled parental Ab and incubated in a 37oC water bath. At each duplicate time point, 0.25 ml
sample was removed and internalization was stopped by the addition of 4 volumes of quench
buffer (cold 1 x PBS + 0.2% sodium azide) and placed on ice. Subsequently, cells were treated
with 1x trypsin-EDTA (Mediatech) supplemented with 5 mg/ml papain (Sigma-Aldrich) and
incubated in a 37oC water bath for 25 min to remove un-internalized antibodies on the surface.
After wash cells were re-suspended in 200 μl of fix buffer (1xPBS + 1.5% paraformaldehyde +
on May 14, 2018. © 2016 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2016; DOI: 10.1158/1535-7163.MCT-16-0431

8
1:4000 Draq5 (Biostatus) and centrifuged onto a 96-well poly-D-lysine coated plate (BD
biosciences). For time point zero, a second set of samples was collected without the
trypsin/papain step to measure the maximal amount of surface-bound fluorescence signals. The
fluorescence signals were acquired by Odyssey CLx Infrared Imaging System (LI-COR
Biotechnology) and analyzed using Odyssey software version 3.0.30. For each time point,
relative fluorescence signal, Fr(t), was determined from the fluorescence antibody signal divided
by the Draq5 DNA signal. Subsequently, the normalized amount of internalized antibody at each
time, Y(t), was calculated from Fr(t) divided by Fmax, the maximum surface-bound fluorescence
signal at time zero, and plotted as a function of time. The resulting graph was fitted to the
equation Y(t) = (Bmax * t)/(T½ + t) using GraphPad Prism 6.03 where t is time, Bmax is the
maximal amount of internalized antibody and T½ is the time at which half maximum
internalization occurs.
Immunofluorescence
Trop-2 expression: BxPC3 and SW620 cells were seeded on 12 mm coverslips put into 24 well
plates in growth media until 70-80% confluent. Mouse Ab from which RN927C was derived
was used at 2 μg /mL in RPMI + 10 % FBS + 20 mM HEPES and incubated at 4°C for 1 hour.
The cells were washed twice with 1 x PBS and then fixed in 4% formaldehyde solution for 10
minutes at room temperature. Afterwards the cells were washed 2x with PBS and then incubated
in 400 μL non-permeabilizing blocking buffer (1 x PBS + 5 % Donkey serum) for 1 hour. Trop-2
expression was then detected with Cy3-conjugated donkey anti-mouse 2nd Ab (1:300, Jackson
ImmunoResearch Laboratories). Colocalization with lysosomal marker: cells were grown on
coverslips as described previously. After removing the growth media, 10 μg/mL of RN927C in
on May 14, 2018. © 2016 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2016; DOI: 10.1158/1535-7163.MCT-16-0431

9
RPMI + 10 % FBS + 20 mM HEPES were added to the cells and incubated at 4°C for 1 hour.
The antibody solution was then removed and the cells were washed once with 1 x PBS. 0.5 mL
growth media was then added and the cells were moved to 37°C incubator for 2 hours. The cells
were washed twice with 1 x PBS, fixed and then incubated in 400 μL blocking buffer (1 x PBS +
0.3 % TX-100 + 5 % Donkey serum) for 1 hour. Lysosomal compartments were labeled with
anti-LAMP2 (Abcam) diluted 1:100 in 300 μL blocking buffer for 1 hour at room temperature
followed by 2nd Ab detection (1:300 for Cy3-donkey anti-mouse and 1:200 for Dylight488 goat
anti-human, both from Jackson ImmunoResearch Laboratories). The coverslips were mounted
on slides with a drop of Vectashield with DAPI (Vector Laboratories) to stain the nuclei. Images
were observed and acquired by a scanning confocal microscope LAS AF from Leica.
Phosphorylated histone H3 staining: cells were grown on chamber slides (Thermo Scientific)
until 50-70% confluent and then treated overnight with 0.1, 1 and 10 μg/mL of RN927C. Anti-
phospho-histone H3 rabbit antibody (Cell Signaling Technology) was used at 1:1600 dilution for
staining and detection was done using Alex488-F(ab’)2 donkey anti-rabbit IgG (Jackson
ImmnuoResearch Laboratories) as 2nd Ab. Images were observed and acquired by a Nicon
Eclipse E800 microscope (Nicon Instruments).
In vivo studies:
All animal studies were conducted in an AAALAC accredited facility under IACUC
(Institutional Animal Care and Use Committee) approved protocols. Three animals were used at
each time point in the PK study and five animals per cohort were used in all the efficacy studies.
For BxPC3 xenograft model used in both PK and efficacy studies, 2 million cells were implanted
subcutaneously into 5-6 weeks old female CB17/SCID mice (The Jackson Laboratory) until the
on May 14, 2018. © 2016 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2016; DOI: 10.1158/1535-7163.MCT-16-0431

10
tumor sizes reached 250-300 mm3 before treatment started. For SW620 model, 3 million tumor
cells were implanted subcutaneously into 5-7 weeks old female NCr nu/nu mice (Taconic) until
the tumor sizes reached ~300 mm3. Pan 0123 (pancreatic ductal carcinoma from peritoneal
biopsy), Pan 0135 (pancreatic adenosquamous carcinoma) and Pan0146 (metastatic pancreatic
carcinoma from liver biopsy), and LG0476 (Squamous non-small cell lung carcinoma) PDX
models were acquired from The Jackson Laboratory, CTG-1017 (triple negative breast cancer)
model was acquired from Champion Oncology, and the Ova196756 (ovarian adenocarcinoma
metastasized to colon) model was established in house from surgical tumor specimen propagated
first in NSG mice (The Jackson Laboratory). For these models 1-2 mm3 of tumor fragments
were implanted subcutaneously into the lateral flanks of female CB17/SCID mice from Taconic.
Animals were randomized by tumor sizes once they reached ~300 mm3 or otherwise indicated,
and RN927C and other reagents were administered through bolus tail vein injection. Tumor
volume was calculated with the following formula: Tumor volume = (length x width 2) / 2.
Animals were humanely sacrificed before their tumor volumes reached 2000 mm3. All tumor
models except SW620 were tested positive for Trop-2 expression by immunohistochemistry
(data not shown).
Pharmacokinetic analysis:
Serum samples were analyzed using an enzyme-linked immunosorbent assays (ELISA)
developed on GyroLab@ immunoassay platform. For the total antibody assay, RN927C or
unconjugated Ab were captured using biotinylated polyclonal goat anti-human IgG (H+L)
antibody (Southern Biotech), and detected with a polyclonal goat anti-human IgG (H+L)
(Bethyl) labeled with Alexa Fluor 647. For the ADC assay, RN927C was captured with
biotinylated polyclonal goat anti-human IgG (H+L) antibody (Southern Biotech). The detection
on May 14, 2018. © 2016 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2016; DOI: 10.1158/1535-7163.MCT-16-0431

11
of the captured RN927C was done with a polyclonal antibody generated in-house that recognizes
PF-06380101. The instrument response was used to construct a standard curve and calculate
concentration of study samples and QCs. For both antibody and ADC assays, the lower limit of
quantitation was 50 ng/mL. Frozen tumor samples were thawed on ice and homogenized
(FastPrep®-24 tissue homogenizer) in ice-cold lysis buffer (Invitrogen). The resulting
homogenate was centrifuged at 12,000 rpm for 20 minutes at 4 °C to separate the supernatant
containing RN927C. The samples were analyzed for total antibody and ADC using the same
analytical procedures as for serum. Pharmacokinetic data analysis was performed by the non-
compartmental method using Phoenix software v. 6.3 (Pharsight, Prinseton, NJ).
Immunohistochemistry: Formalin-fixed, paraffin embedded tumor sections were processed and
stained with anti-phospho-H3 (Ser10) rabbit pAb (#9701, Cell Signaling Technology) according
to manufacturer’s instruction followed by detection with EnVision HRP-labeled polymer anti-
rabbit 2nd Ab (DAKO).
Results
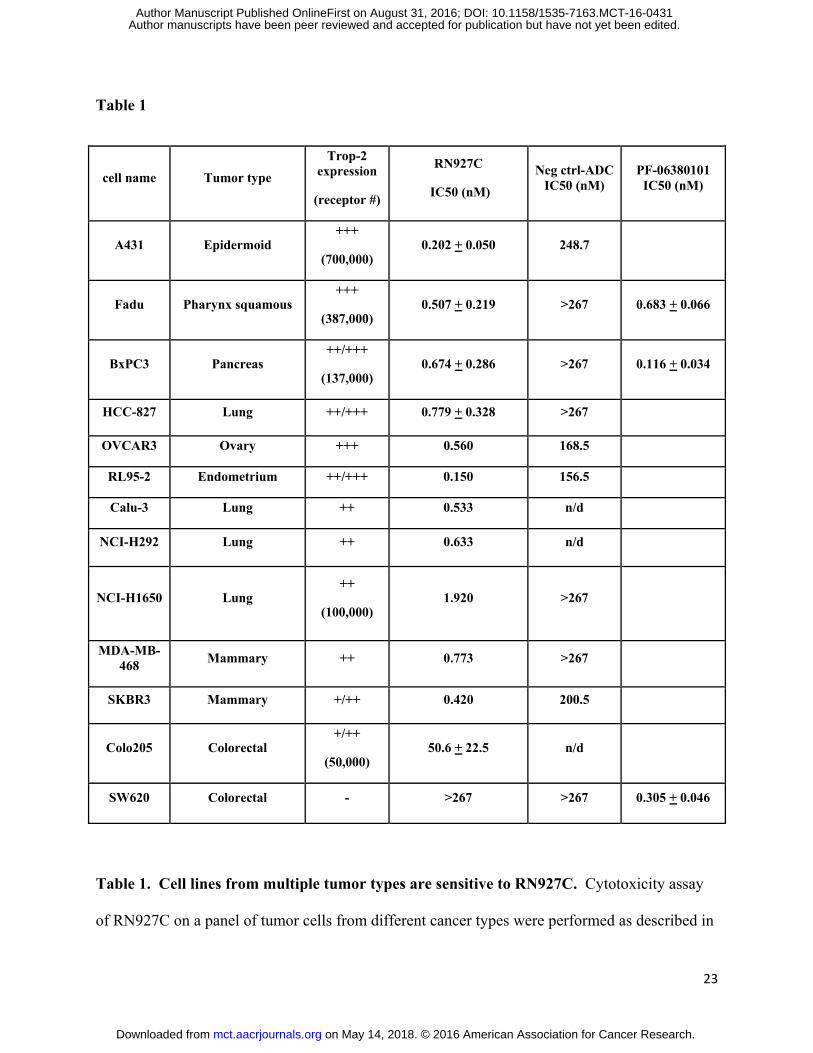
RN927C is a homogeneous ADC compound
The chemical composition of RN927C is shown in Figure 1A. RN927C is generated by
conjugating AcLys-VC-PABC-PF-06380101 (38, 40) to the transglutaminase tag (LLQGA)
located at the C-terminus of the antibody heavy chain (see Materials and Methods). The AcLys
linker was chosen to improve the stability of the conjugate in circulation (40). The conjugation
reaction yielded a product with DAR (Drug-Antibody-Ratio) of 1.9 and after a single HIC
purification step a homogeneous conjugate with DAR 2.0 was achieved (Fig. 1B). The process
on May 14, 2018. © 2016 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2016; DOI: 10.1158/1535-7163.MCT-16-0431

12
is readily scalable, and high degree of reproducibility was observed among multiple gram scale
production lots as measured by cytotoxicity on Trop-2-expressing BxPC3 cells (Fig. 1C).
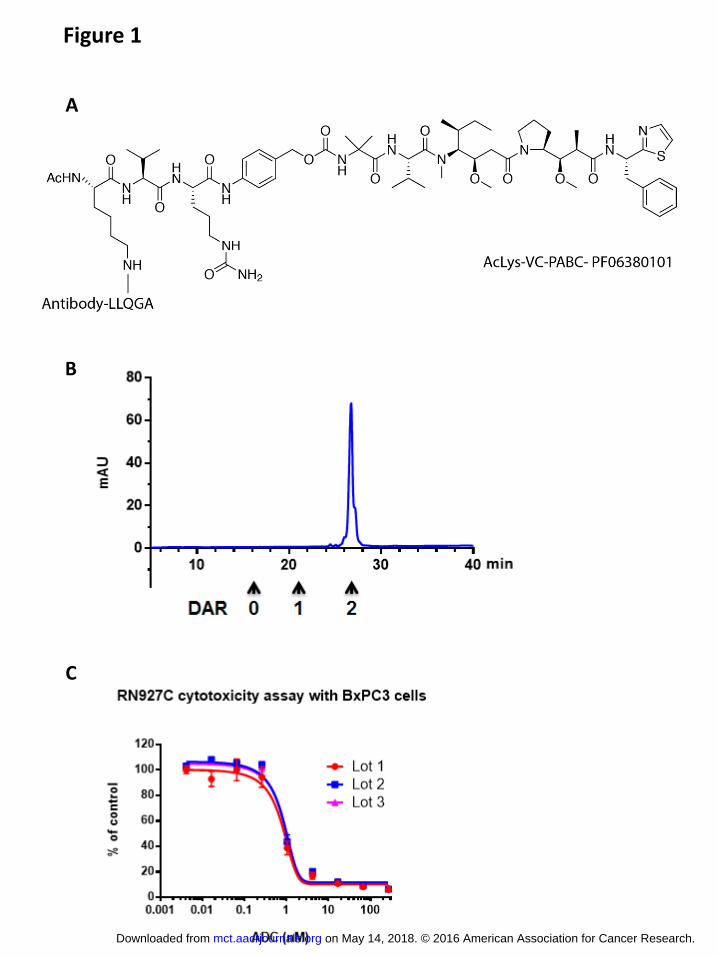
RN927C internalizes efficiently and traffics to the lysosomal compartment
The uptake of antibody drug conjugates into the target cells and its subsequent release,
usually by lysosomal proteases, is important for cytotoxic drug delivery. The internalization
kinetics of RN927C was measured and compared to the parental unconjugated Ab in Trop-2
expression NSCLC cell line HCC-827. Both RN927C and the unconjugated parental Ab
internalized efficiently, with t1/2 at 23.4 and 31.6 minutes, respectively (Fig. 2A). The
localization of RN927C was further examined by confocal microscopy, and significant amount
of RN927C was found to co-localize with the lysosomal marker LAMP-2 after two hour
incubation (Fig. 2B).
RN927C induces mitotic arrest in Trop-2-expressing cells
The presumed mechanism through which RN927C elicits cytotoxicity is by intracellular
release of the PF-06380101 payload, which results in the disruption of microtubule
polymerization leading to mitotic arrest, apoptosis and cell death. To confirm this mechanism,
Trop-2 positive BxPC3 and Trop-2 negative SW620 cells (Fig. 3A) were incubated with various
concentrations of RN927C overnight and processed for immunofluorescence (Fig. 3B and C).
Cells in mitotic phase were indicated by positive staining of phosphorylated histone H3 (41).
RN927C induced a dramatic increase of positive phosphorylated histone H3 staining in BxPC3
cells treated with RN927C at the lowest experimental concentration of 0.1 μg/mL (Fig. 3B),
indicating many treated cells were trapped in the mitotic phase. Cells started to detach from the
slides after overnight incubation of RN927C at 10 μg/mL, resulting in loss of cells. In contrast,
on May 14, 2018. © 2016 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2016; DOI: 10.1158/1535-7163.MCT-16-0431

13
the untreated cells display the typical low percentage of mitotic cells (Fig. 3B). RN927C
incubation did not show any increase of phosphorylated histone H3 staining on Trop-2 negative
SW620 cells at 10 μg/mL (Fig. 3C), indicating that the activity of RN927C to induce mitotic
arrest in these cells is dependent on Trop-2 expression. RN927C - induced increase in
phosphorylated H3 staining is also observed in vivo on a pancreatic PDX model Pan0146, with
the peak staining observed 3-5 days after RN927C dosing (Supplementary Fig. S1).
RN927C exerts potent in vitro tumor cell killing activity
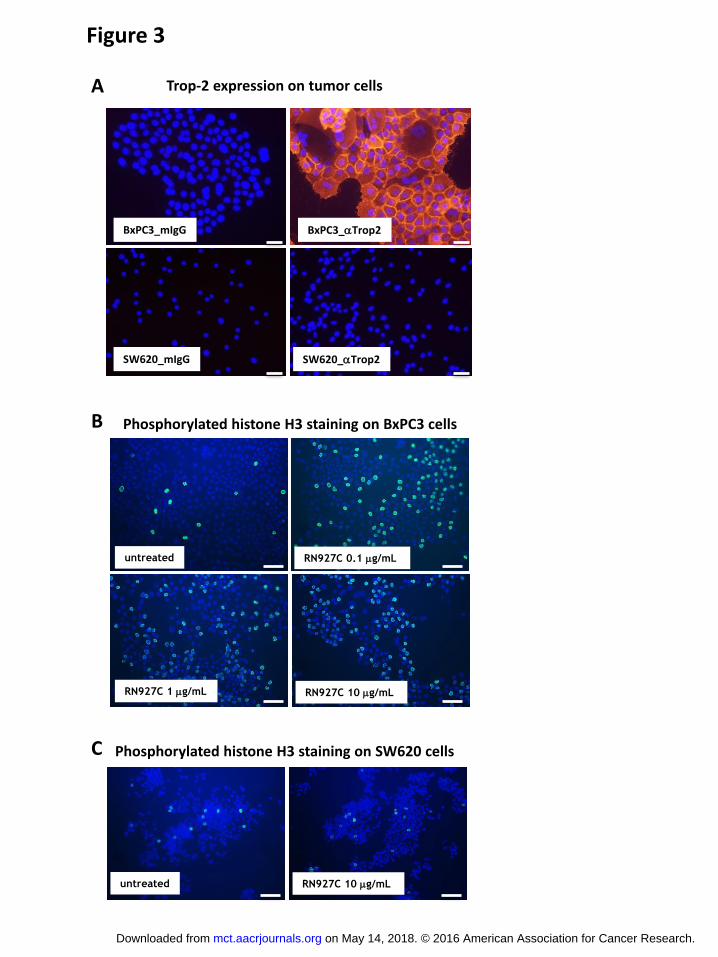
Next we examined the in vitro cytotoxicity of RN927C on a variety of tumor cell lines.
Tumor cell lines from multiple tumor types including skin, pancreas, lung, head and neck, breast,
ovary and colon were exposed to RN927C and negative control conjugate at concentrations
ranging from 0.04 to 267 nM for 4 days. Most cell lines were readily killed by RN927C with
IC50 below 1 nM. Negative control conjugate was not active against any of the cell lines, and
RN927C did not show any cytotoxicity toward Trop-2 negative SW620 cells (Table 1). The lack
of killing of Trop-2 negative SW620 cells is not due to intrinsic insensitivity to the PF-06380101
as free payload is active against SW620 cells with IC50 of 0.305 nM, comparable of that of the
Trop-2 expressing BxPC3 cells at 0.116 nM or Fadu cells at 0.683 nM (Table 1). These results
indicated that RN927C is efficacious in killing cells derived from multiple tumor types and the
killing requires Trop-2 expression.
RN927C linker is stable in vivo
To evaluate the pharmacokinetic characteristics of RN927C in vivo, single 1.5 mg/kg dose of
RN927C was injected i.v. into BxPC3 tumor-bearing animals with tumor sizes ~250mm3. Both
blood and tumor samples were collected at the following time points (0, 0.083, 2, 6, 24, 72, 120,
on May 14, 2018. © 2016 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2016; DOI: 10.1158/1535-7163.MCT-16-0431

14
168, 240 and 336 hours). Total antibody (including both conjugated and unconjugated
antibodies) and ADC concentration were measured as described in the Materials and Methods.
Note that the immunoassay-based measurement cannot distinguish between ADC of DAR1 and
DAR2 therefore certain degree of payload loss can still occur and not be shown by the assay.
ADC and total antibody concentration in serum were similar throughout the time course,
indicating only a slow release of the payload in circulation (Fig. 4A). The serum AUC0-336 for
total Ab is 2488 (μg*hr/mL) and for ADC is 2154 (μg*hr/mL), suggesting that most of the Ab
remaining in circulation still contains payload PF-06380101. There is a slightly faster loss of
payload within the tumors, possibly due to increased internalization and processing of RN927C
occurring in Trop-2 positive tumor cells (Fig. 4B). AUC0-336 percentage of ADC from the total
Ab population in serum is 87% (Fig. 4A), showing good serum linker stability of RN927C in
vivo.
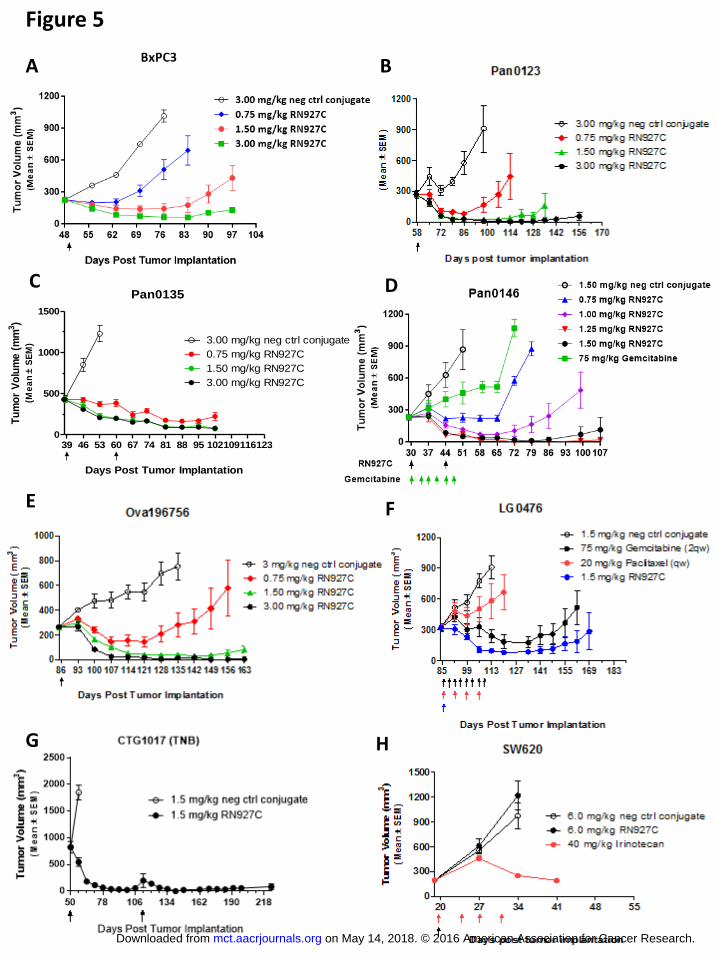
RN927C is highly efficacious in multiple tumor xenograft models
The in vivo efficacy of RN927C was tested on multiple tumor models. In a panel of
pancreatic tumor xenograft models including cell line BxPC3 and PDX models Pan0123,
Pan0135 and Pan0146, one or two doses (every two or three weeks) of RN927C treatment
resulted in sustained tumor growth inhibition/regression at doses between 0.75 to 1.5 mg/kg (Fig.
5A-D). A negative control conjugate consisting of a non-binding antibody conjugated in the
same manner as RN927C showed no effect on these tumor models at 3 mg/kg, the highest dose
tested in these studies. Additionally, in the Pan0146 model, two doses of RN927C at ≥ 0.75
mg/kg were more efficacious than gemcitabine treatment given at 75 mg/kg twice weekly for 6
doses (Fig. 5D).
on May 14, 2018. © 2016 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2016; DOI: 10.1158/1535-7163.MCT-16-0431

15
RN927C is also highly efficacious in other solid tumor types tested (Fig. 5E-G). Robust
anti-tumor activity was observed in an ovarian PDX model Ova196756 with single dose
treatment of RN927C at doses > 1.50 mg/kg resulting in long-term tumor regression and tumor
eradication (Fig. 5E). In a lung PDX model LG0476, 1.5 mg/kg single injection of RN927C is
more efficacious in inducing tumor regression than gemcitabine treatment given at 75 mg/kg
twice weekly for 8 doses. Four weekly doses of paclitaxel at 20 mg/kg also only achieved partial
tumor growth inhibition in the same study (Fig. 5F).
In a TNB (triple negative) breast cancer PDX model CTG-1017, single injection of
RN927C induces tumor regression of large tumors (~830 mm3) for more than 60 days. Tumors
that re-grew were treated again 63 days after the first RN927C injection and tumor regression
was again achieved for long duration (Fig. 5G). The anti-tumor effect of RN927C is Trop-2-
dependent as an SW620 colon cancer model devoid of Trop-2 expression showed no response to
RN927C compared to the control conjugate, indicating target expression is required for efficacy
(Fig. 5H). On the other hand standard of care irinotecan resulted in significant tumor growth
inhibition in SW620 model after 4 doses treatment at 40 mg/kg (Fig. 5H).
Exploratory toxicology
The non-clinical safety profile of RN927C has been characterized in rats (up to 30
mg/kg) and non-human primates (up to 6 mg/kg) in repeat-dose studies (two doses; on day 1 and
day 15). The key safety signals observed in cynomolgus monkeys were found in tissues that
express Trop-2. Reversible findings included increased mitoses and single cell necrosis in
multiple epithelial tissues in the skin, injection sites, upper alimentary canal (oral mucosa and
esophagus) and vagina. These were likely related to the activity of the PF-06380101 payload in
on May 14, 2018. © 2016 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2016; DOI: 10.1158/1535-7163.MCT-16-0431

16
the Trop-2 expressing tissues. On the other hand, in rats, organ toxicities were more pronounced
in the hematopoietic system and lymphoid tissues, presumably due to the ability to test higher
doses in rats due to lack of cross-reactivity of RN927C to tissues that express rat Trop-2.
Discussion
In this paper we described a novel anti-Trop-2 ADC, RN927C, composed of a humanized
anti-Trop-2 antibody with a site-specifically conjugated novel MTI-payload, PF-06380101 (30,
38, 40), resulting in improved ADC stability and durable anti-tumor responses after a single dose
of therapy. Trop-2 as a tumor antigen has the advantage of high prevalence in many solid tumor
types. It also internalizes efficiently with t1/2 ~ 30 min (Fig. 2), generally considered a
prerequisite for efficient ADC delivery into the cells. We and others have found Trop-2 to be
highly expressed on multiple epithelial tumor types including pancreatic, breast, ovarian,
NSCLC, prostate, gastric and oral cancers (5) . However, it is also expressed on a number of
normal epithelial tissues such as skin and oral mucosa. For example, human keratinocytes were
found to express Trop-2 at ~ 50,000 copies / cell based on receptor quantitation (data not shown).
In general, it is determined that there is 2-10 fold increase of Trop-2 expression in tumors (++ to
+++) compared to target-positive normal tissues (+ to ++). The design of an anti-Trop-2 ADC
thus required a careful balance between efficacy and safety. Several factors were taken into
consideration: first different antibody affinities were explored and a carrier Ab of medium
affinity to Trop-2 (KD ~ 14 nM at 37 degree) was chosen to favor binding of higher Trop-2
expressing tumor tissue over normal tissues. In the cytotoxicity assays this medium affinity Ab
was similarly active to Abs with 10 fold higher affinity on cell lines with high Trop-2 expression
on May 14, 2018. © 2016 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2016; DOI: 10.1158/1535-7163.MCT-16-0431

17
(+++), but showed lower killing activity in low Trop-2 expressing cells (+) that have similar
target levels as normal tissues (data not shown). Secondly, use of a transglutaminase-mediated
site-specific conjugation technology to enable production of a nearly homogeneous Trop-2
ADCs carrying two payloads (DAR2) (30). The majority of ADCs that have been used clinically
have been manufactured through cysteine disulfide bond or lysine based conjugations, producing
mixtures of ADC species with various DARs with an average around 4. By decreasing the
payload to antibody ratio, making a homogeneous ADC, and eliminating high DAR species we
aim to reduce the toxicity to lower Trop-2 expressing tissue. The generally high Trop-2
expression on tumor cells and efficient internalization of the ADC should compensate for the
reduced drug loading (Fig. 2). Indeed, most of the tumor cell lines we tested are sensitive to
RN927C with IC50 in the sub-nanomolar range (Table 1). RN927C is highly efficacious in vivo,
with tumor regression generally achieved with single injection of RN927C at a dose of > 1.5
mg/kg with some sensitive models responding to a dose of 0.75 mg/kg (Fig. 5). Efficacy is seen
in pancreatic, ovarian, lung and breast cancer models and is Trop-2 expression dependent. We
also found that RN927C is generally more efficacious than standard of care in these models. For
instance, in the pancreatic cancer PDX model Pan0146, RN927C treatment at >1 mg/kg Q2W for
two doses induces tumor regression, while multiple twice a week gemcitabine dosing only
resulted in tumor growth inhibition (Fig. 5D). Favorable comparison is also observed in NSCLC
PDX model LG0476 where a single dose of RN927C treatment outperforms multiple doses of
gemcitabine or paclitaxel (Fig. 5F).
Many ADCs in the clinic are limited by the off-target toxicity, most notably
hematological toxicity and neurotoxicity. Off-target toxicities can arise from both non-target-
specific uptake or premature release of payloads. Loss of payloads from ADCs in circulation can
on May 14, 2018. © 2016 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2016; DOI: 10.1158/1535-7163.MCT-16-0431

18
result in reduced efficacy and a toxicity profile resembling that of the free payload. Several
ADC programs utilizing a cleavable linker and MMAE payload reported neutropenia and
peripheral neuropathy as dose-limiting toxicities (42). IMMU-132, an ADC targeting Trop-2
with payload SN38 (active metabolite of irinotecan) also concluded their safety profile to be
similar to irinotecan (hematological and gastrointestinal toxicity), with no/little toxicities related
to Trop-2 expressing normal tissues (23). Additionally, rapid loss of payloads likely results in
accumulation of conjugate-free antibodies, as antibody half-lives are usually longer than the
intact ADCs. Cardillo et al., reported in a mouse PK study that the AUCs for the intact IMMU-
132 versus the carrier antibody hRS7, were 1516 (h*μg/mL) and 13112 (h*μg/mL), respectively
(21). The persistent presence of unconjugated antibody could have complicated effects to the
ADC efficacy. While some antibodies can elicit anti-tumor effect and possibly aid to the
response such as in the case of T-DM1 for HER2 positive breast cancer, the unconjugated
antibody is also a potential competitor for target binding and could negatively ADC efficacy. In
fact, we have observed a strong inhibitory effect from unconjugated Trop-2 antibodies on the in
vivo efficacy of RN927C, and this effect is most profound within the first three days
(Supplementary Fig. S2).
To reduce the off-target toxicities resulted from de-conjugation of free payloads or linker
cleavage observed with ADCs made with conventional cysteine disulfide bond or lysine based
conjugations, we utilized stable isopeptide linkage and incorporated an AcLys-VC-PABC linker
that was shown to be more stable in circulation (30, 39, 40). We have indeed observed
improved stability of our Trop-2 ADC in vivo, evidenced by the small difference between ADC
and total Ab pharmacokinetic profiles (Fig. 4). Preclinical exploratory safety studies of RN927C
conducted in non-human primates with doses up to 6 mg/kg for two doses showed mostly on
on May 14, 2018. © 2016 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2016; DOI: 10.1158/1535-7163.MCT-16-0431

19
target epithelial toxicities (rash and mucositis) that were fully recoverable, consistent with Trop-
2 expression on skin and oral mucosa. Notably there were no adverse hematological findings
observed in monkeys, indicating reduction of off-target toxicity presumably due to the stable
nature of our linker-payload. With a more stable compound we have observed a potential
correlation between Trop-2 expression and efficacy. Trop-2- negative tumor models such as
SW620 typically being insensitive to RN927C treatment, and tumors with high and
homogeneous Trop-2 expression responding well with sustained efficacy (Fig. 5). It is likely
that a companion diagnostic test for patient selection based on Trop-2 expression level would
enhance clinical activity of a Trop-2 targeting ADCs. In summary, we have developed a
homogeneous site-specific Trop-2 ADC with enhanced stability, and the preclinical efficacy and
safety data support clinical testing of RN927C in multiple solid tumor types.
Acknowledgements
The authors would like to thank Jessica Yu and Jeanette Dilley for their hybridoma work, Kevin
Lindquist, Christine Bee, and Yasmina Abdiche for Biosensor analysis; Bryant Chau, Colleen
Brown, Ishita Barman and Michael Chin for protein and antibody production; Jyothirmayee
Kudaravalli, Ratika Chopra, and Jing-Tyan Ma for antibody characterization and technical
assistance; Victor Lui and Santiago Farias for Mass Spec analytical support, Rachel DeVay for
confocal microscopy assistance, and Birte Nolting for technical development in scale up. The
authors wish to acknowledge the contributions of Andreas Maderna and Matthew Doroski for
their discovery of the PF-06380101 payload, and Michael Green and Ramalakshmi
Chandrasekaran for preparation of key intermediates used in the synthesis of AcLys-VC-PF-
06380101.
on May 14, 2018. © 2016 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2016; DOI: 10.1158/1535-7163.MCT-16-0431

20
References:
1. Sievers EL, Senter PD. Antibody-drug conjugates in cancer therapy. Annual review of medicine. 2013;64:15-29. 2. Panowksi S, Bhakta S, Raab H, Polakis P, Junutula JR. Site-specific antibody drug conjugates for cancer therapy. mAbs. 2014;6:34-45. 3. Polakis P. Antibody Drug Conjugates for Cancer Therapy. Pharmacological reviews. 2016;68:3-19. 4. Lipinski M, Parks DR, Rouse RV, Herzenberg LA. Human trophoblast cell-surface antigens defined by monoclonal antibodies. Proceedings of the National Academy of Sciences of the United States of America. 1981;78:5147-50. 5. Shvartsur A, Bonavida B. Trop2 and its overexpression in cancers: regulation and clinical/therapeutic implications. Genes & cancer. 2015;6:84-105. 6. Wang J, Day R, Dong Y, Weintraub SJ, Michel L. Identification of Trop-2 as an oncogene and an attractive therapeutic target in colon cancers. Molecular cancer therapeutics. 2008;7:280-5. 7. Trerotola M, Jernigan DL, Liu Q, Siddiqui J, Fatatis A, Languino LR. Trop-2 promotes prostate cancer metastasis by modulating beta(1) integrin functions. Cancer research. 2013;73:3155-67. 8. Fong D, Moser P, Krammel C, Gostner JM, Margreiter R, Mitterer M, et al. High expression of TROP2 correlates with poor prognosis in pancreatic cancer. British journal of cancer. 2008;99:1290-5. 9. Bignotti E, Todeschini P, Calza S, Falchetti M, Ravanini M, Tassi RA, et al. Trop-2 overexpression as an independent marker for poor overall survival in ovarian carcinoma patients. European journal of cancer. 2010;46:944-53. 10. Bignotti E, Zanotti L, Calza S, Falchetti M, Lonardi S, Ravaggi A, et al. Trop-2 protein overexpression is an independent marker for predicting disease recurrence in endometrioid endometrial carcinoma. BMC clinical pathology. 2012;12:22. 11. Kobayashi H, Minami Y, Anami Y, Kondou Y, Iijima T, Kano J, et al. Expression of the GA733 gene family and its relationship to prognosis in pulmonary adenocarcinoma. Virchows Archiv : an international journal of pathology. 2010;457:69-76. 12. Ohmachi T, Tanaka F, Mimori K, Inoue H, Yanaga K, Mori M. Clinical significance of TROP2 expression in colorectal cancer. Clin Cancer Res. 2006;12:3057-63. 13. Muhlmann G, Spizzo G, Gostner J, Zitt M, Maier H, Moser P, et al. TROP2 expression as prognostic marker for gastric carcinoma. Journal of clinical pathology. 2009;62:152-8. 14. Fong D, Spizzo G, Gostner JM, Gastl G, Moser P, Krammel C, et al. TROP2: a novel prognostic marker in squamous cell carcinoma of the oral cavity. Modern pathology : an official journal of the United States and Canadian Academy of Pathology, Inc. 2008;21:186-91. 15. Cubas R, Zhang S, Li M, Chen C, Yao Q. Trop2 expression contributes to tumor pathogenesis by activating the ERK MAPK pathway. Molecular cancer. 2010;9:253. 16. Stoyanova T, Goldstein AS, Cai H, Drake JM, Huang J, Witte ON. Regulated proteolysis of Trop2 drives epithelial hyperplasia and stem cell self-renewal via beta-catenin signaling. Genes & development. 2012;26:2271-85. 17. Wang J, Zhang K, Grabowska D, Li A, Dong Y, Day R, et al. Loss of Trop2 promotes carcinogenesis and features of epithelial to mesenchymal transition in squamous cell carcinoma. Molecular cancer research : MCR. 2011;9:1686-95. 18. Lin JC, Wu YY, Wu JY, Lin TC, Wu CT, Chang YL, et al. TROP2 is epigenetically inactivated and modulates IGF-1R signalling in lung adenocarcinoma. EMBO molecular medicine. 2012;4:472-85.
on May 14, 2018. © 2016 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2016; DOI: 10.1158/1535-7163.MCT-16-0431

21
19. Zhang K, Jones L, Lim S, Maher CA, Adkins D, Lewis J, et al. Loss of Trop2 causes ErbB3 activation through a neuregulin-1-dependent mechanism in the mesenchymal subtype of HNSCC. Oncotarget. 2014;5:9281-94. 20. Cardillo TM, Govindan SV, Sharkey RM, Trisal P, Goldenberg DM. Humanized anti-Trop-2 IgG-SN-38 conjugate for effective treatment of diverse epithelial cancers: preclinical studies in human cancer xenograft models and monkeys. Clin Cancer Res. 2011;17:3157-69. 21. Cardillo TM, Govindan SV, Sharkey RM, Trisal P, Arrojo R, Liu D, et al. Sacituzumab Govitecan (IMMU-132), an Anti-Trop-2/SN-38 Antibody-Drug Conjugate: Characterization and Efficacy in Pancreatic, Gastric, and Other Cancers. Bioconjugate chemistry. 2015;26:919-31. 22. Goldenberg DM, Cardillo TM, Govindan SV, Rossi EA, Sharkey RM. Trop-2 is a novel target for solid cancer therapy with sacituzumab govitecan (IMMU-132), an antibody-drug conjugate (ADC). Oncotarget. 2015;6:22496-512. 23. Starodub AN, Ocean AJ, Shah MA, Guarino MJ, Picozzi VJ, Jr., Vahdat LT, et al. First-in-Human Trial of a Novel Anti-Trop-2 Antibody-SN-38 Conjugate, Sacituzumab Govitecan, for the Treatment of Diverse Metastatic Solid Tumors. Clin Cancer Res. 2015;21:3870-8. 24. Hamblett KJ, Senter PD, Chace DF, Sun MM, Lenox J, Cerveny CG, et al. Effects of drug loading on the antitumor activity of a monoclonal antibody drug conjugate. Clin Cancer Res. 2004;10:7063-70. 25. Wang L, Amphlett G, Blattler WA, Lambert JM, Zhang W. Structural characterization of the maytansinoid-monoclonal antibody immunoconjugate, huN901-DM1, by mass spectrometry. Protein science : a publication of the Protein Society. 2005;14:2436-46. 26. Boswell CA, Mundo EE, Zhang C, Bumbaca D, Valle NR, Kozak KR, et al. Impact of drug conjugation on pharmacokinetics and tissue distribution of anti-STEAP1 antibody-drug conjugates in rats. Bioconjugate chemistry. 2011;22:1994-2004. 27. Junutula JR, Raab H, Clark S, Bhakta S, Leipold DD, Weir S, et al. Site-specific conjugation of a cytotoxic drug to an antibody improves the therapeutic index. Nature biotechnology. 2008;26:925-32. 28. Shen BQ, Xu K, Liu L, Raab H, Bhakta S, Kenrick M, et al. Conjugation site modulates the in vivo stability and therapeutic activity of antibody-drug conjugates. Nature biotechnology. 2012;30:184-9. 29. Axup JY, Bajjuri KM, Ritland M, Hutchins BM, Kim CH, Kazane SA, et al. Synthesis of site-specific antibody-drug conjugates using unnatural amino acids. Proceedings of the National Academy of Sciences of the United States of America. 2012;109:16101-6. 30. Strop P, Liu SH, Dorywalska M, Delaria K, Dushin RG, Tran TT, et al. Location matters: site of conjugation modulates stability and pharmacokinetics of antibody drug conjugates. Chemistry & biology. 2013;20:161-7. 31. Okeley NM, Toki BE, Zhang X, Jeffrey SC, Burke PJ, Alley SC, et al. Metabolic engineering of monoclonal antibody carbohydrates for antibody-drug conjugation. Bioconjugate chemistry. 2013;24:1650-5. 32. Zimmerman ES, Heibeck TH, Gill A, Li X, Murray CJ, Madlansacay MR, et al. Production of site-specific antibody-drug conjugates using optimized non-natural amino acids in a cell-free expression system. Bioconjugate chemistry. 2014;25:351-61. 33. Drake PM, Albers AE, Baker J, Banas S, Barfield RM, Bhat AS, et al. Aldehyde tag coupled with HIPS chemistry enables the production of ADCs conjugated site-specifically to different antibody regions with distinct in vivo efficacy and PK outcomes. Bioconjugate chemistry. 2014;25:1331-41. 34. Beerli RR, Hell T, Merkel AS, Grawunder U. Sortase Enzyme-Mediated Generation of Site-Specifically Conjugated Antibody Drug Conjugates with High In Vitro and In Vivo Potency. PloS one. 2015;10:e0131177. 35. Grunewald J, Klock HE, Cellitti SE, Bursulaya B, McMullan D, Jones DH, et al. Efficient Preparation of Site-Specific Antibody-Drug Conjugates Using Phosphopantetheinyl Transferases. Bioconjugate chemistry. 2015;26:2554-62.
on May 14, 2018. © 2016 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2016; DOI: 10.1158/1535-7163.MCT-16-0431

22
36. Lyon RP, Bovee TD, Doronina SO, Burke PJ, Hunter JH, Neff-LaFord HD, et al. Reducing hydrophobicity of homogeneous antibody-drug conjugates improves pharmacokinetics and therapeutic index. Nature biotechnology. 2015;33:733-5. 37. Kung Sutherland MS, Walter RB, Jeffrey SC, Burke PJ, Yu C, Kostner H, et al. SGN-CD33A: a novel CD33-targeting antibody-drug conjugate using a pyrrolobenzodiazepine dimer is active in models of drug-resistant AML. Blood. 2013;122:1455-63. 38. Maderna A, Doroski M, Subramanyam C, Porte A, Leverett CA, Vetelino BC, et al. Discovery of cytotoxic dolastatin 10 analogues with N-terminal modifications. Journal of medicinal chemistry. 2014;57:10527-43. 39. Dorywalska M, Strop P, Melton-Witt JA, Hasa-Moreno A, Farias SE, Galindo Casas M, et al. Effect of attachment site on stability of cleavable antibody drug conjugates. Bioconjugate chemistry. 2015;26:650-9. 40. Dorywalska M, Dushin R, Moine L, Farias SE, Zhou D, Navaratnam T, et al. Molecular basis of valine-citrulline-PABC linker instability in site-specific ADCs and its mitigation by linker design. Molecular cancer therapeutics. 2016. 41. Goto H, Tomono Y, Ajiro K, Kosako H, Fujita M, Sakurai M, et al. Identification of a novel phosphorylation site on histone H3 coupled with mitotic chromosome condensation. The Journal of biological chemistry. 1999;274:25543-9. 42. Burris H. Antibody drug conjugates: promise beyond breast cancer. SOWG plenary session, San Francisco. 2014.
on May 14, 2018. © 2016 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2016; DOI: 10.1158/1535-7163.MCT-16-0431
23
Table 1
cell name Tumor type Trop-2
expression
(receptor #)
RN927C
IC50 (nM) Neg ctrl-ADC
IC50 (nM) PF-06380101
IC50 (nM)
A431 Epidermoid +++
(700,000) 0.202 + 0.050 248.7
Fadu Pharynx squamous +++
(387,000) 0.507 + 0.219 >267 0.683 + 0.066
BxPC3 Pancreas ++/+++
(137,000) 0.674 + 0.286 >267 0.116 + 0.034
HCC-827 Lung ++/+++ 0.779 + 0.328 >267
OVCAR3 Ovary +++ 0.560 168.5
RL95-2 Endometrium ++/+++ 0.150 156.5
Calu-3 Lung ++ 0.533 n/d
NCI-H292 Lung ++ 0.633 n/d
NCI-H1650 Lung ++
(100,000) 1.920 >267
MDA-MB-468 Mammary ++ 0.773 >267
SKBR3 Mammary +/++ 0.420 200.5
Colo205 Colorectal +/++
(50,000) 50.6 + 22.5 n/d
SW620 Colorectal - >267 >267 0.305 + 0.046
Table 1. Cell lines from multiple tumor types are sensitive to RN927C. Cytotoxicity assay
of RN927C on a panel of tumor cells from different cancer types were performed as described in
on May 14, 2018. © 2016 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2016; DOI: 10.1158/1535-7163.MCT-16-0431

24
Material and Methods. Trop-2 expression (number of + symbols) levels were empirically
assigned based on staining intensity of RN927C parent Ab by FACS (data not shown), and
corresponded to the following receptor copy number ranges (+: < 10-50,000; ++: 50-100,000;
+++: > 100,000). For A431, Fadu, BxPC3, NCI-H1650 and Colo205 cells the actual receptor
numbers were listed in parentheses. For A431, Fadu, BxPC3, HCC-827, and Colo205 cells the
inhibitory concentration of 50% (IC50) was calculated by logistic nonlinear regression and is
reported as the mean± stdev in nM of antibody concentration from multiple experiments. For
SW620 no average can be calculated as most IC50 values from various experiments exceeded the
top concentration of 267 nM. Only single experimental value was listed for OVCAR3, RL95-2,
Calu-3, NCI-H292, NCI-H1650, MDA-MB-468, and SKBR3 cells. Free payload PF-06380101
was tested on tumor cell lines BxPC3, SW620, and Fadu tumor cells. Note that Trop-2 negative
cell SW620 is sensitive to free payload killing.
on May 14, 2018. © 2016 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2016; DOI: 10.1158/1535-7163.MCT-16-0431

25
Figure Legends
Figure 1. Structure, homogeneity and manufacturing reproducibility of RN927C. A. The
chemical structure of the RN927C linker payload including the engineered transglutaminase tag
LLQGA. B. The HIC (hydrophobic interaction chromatography) profile of RN927C after
purification. RN927C was conjugated and purified as described in Material and Methods. The
resulting conjugate appeared as a single peak with payload loading equivalent of DAR 2 on the
HIC column. C. Cytotoxicity assay of three batches of RN927C showed reproducible killing
activity of Trop-2 expressing BxPC3 cells.
Figure 2. Internalization kinetics and intracellular trafficking of RN927C. A. The
internalization kinetics of parental unconjugated Ab and RN927C on Trop-2 expressing NSCLC
HCC-827 cells were measured as described in Material and Methods in duplicates. The t1/2
(time at which half maximum internalization occurs) was determined to be 31.6 minutes for the
unconjugated parent Ab and 23.4 minutes for RN927C. B. The internalized RN927C is shown to
be co-localized with lysosomal marker LAMP-2. RN927C (green) was seen on the cell surface
with confocal microscopy after one hour incubation at 4 degree (0 hr), while LAMP2 marked the
lysosomes (red). The nuclei were stained with DAPI (blue). After 2 hr incubation at 37 degree,
significant amount of RN927C colocalized with LAMP-2 (yellow). Scale bar, 10 μm.
Figure 3. Mitotic arrest induced by RN927C in Trop2 expressing cells. A. BxPC3 and
SW620 cells were stained with mouse parental Ab for RN927C and then detected with Cy3-
conjugated donkey anti-mouse 2nd Ab (red). BxPC3 expressed high level of Trop-2 on the cell
surface while SW620 showed no staining. Scale bar, 20 μm. B. Trop-2 positive BxPC3 cells
were seeded on tissue culture slides and treated with RN927C overnight at concentration
on May 14, 2018. © 2016 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2016; DOI: 10.1158/1535-7163.MCT-16-0431

26
indicated. The cells were then washed and fixed and stained with anti-phosphorylated histone
H3 (pH3) Ab and detected by Alexa488-conjugated donkey anti-rabbit 2nd Ab (green). In
control untreated cells (control) only a few cells were pH3 positive (green, compared to nuclear
staining by DAPI in blue), reflecting a normal low number of cells in mitosis in unsynchronized
population. Treatment of RN927C for as low as 0.1 μg/mL drastically increases the percentage
of pH3 positive cells after overnight incubation, indicating that many cells in the stage of mitotic
arrest. Scale bar, 50 μm. C. In contrast, Trop-2 negative SW620 cells were insensitive to
RN927C treatment at 10 μg/mL (right panel) and the pH3 staining signal did not change
compare to control (untreated). Scale bar, 50 μm.
Figure 4. RN927C is relatively stable in tumor bearing mice. 1.5 mg/kg of RN927C was
administered i.v. to BxPC3 tumor bearing mice and the concentrations of total Ab and ADC
were determined at time points indicated in both the serum samples (A) and tumor specimens (B)
as described in Material and Methods. ADC exposure (AUC0-336) was 87% to total Ab in the
serum (A) and 73% in the tumor (B).
Figure 5. RN927C induces tumor regression in multiple tumor models. A. Single dose
treatment of RN927C at 0.75, 1.5 and 3.0 mg/kg on pancreatic cell line BxPC3 xenograft model.
Tumor regression was observed at ≥1.5 mg/kg. B. Single dose treatment of RN927C at 0.75, 1.5,
and 3.0 mg/kg in pancreatic PDX Pan0123 xenograft model. Tumor regression was observed at
all doses. C. RN927C was administered to Pancreatic PDX model Pan0135 at 0.75, 1.5, and
3.0 mg/kg every two weeks for two treatments. Prolong regression was observed in all doses.
D. RN927C was given to pancreatic PDX model Pan0146 at 0.75 – 1.50 every two weeks for
two doses. Regression was seen at ≥ 1.0 mg/kg. Contrarily Gemcitabine treatment (75 mg/kg)
on May 14, 2018. © 2016 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2016; DOI: 10.1158/1535-7163.MCT-16-0431

27
at twice weekly dosing for 6 doses resulted in only partial growth inhibition. E. Single dose
treatment of RN927C at 0.75, 1.5, and 3.0 mg/kg in ovarian Ova196756 PDX xenograft model.
Tumor regression was observed at all doses but more persistent at doses ≥1.5 mg/kg. F. Single
dose treatment of RN927C at 1.5 mg/kg resulted in sustained tumor regression in Lung LG0476
PDX xenograft model. Negative control conjugate at 1.5mg/kg showed no effect. Gemcitabine
at 75 mg/kg twice weekly dosing for 8 doses induces tumor regression but tumor re-growth
occurred sooner than the RN927C treated group. Paclitaxel at weekly dosing of 20 mg/kg for 4
doses resulted in partial tumor growth inhibition. G. In TNB (triple negative breast) cancer
PDX model CTG-1017, single injection of RN927C induces tumor regression of large tumors
(~830 mm3) for more than 60 days. Tumors that re-grew were treated again 63 days after the
first RN927C injection and tumor regression was again achieved for long duration. H. Single
dose treatment of RN927C at 6.0 mg/kg has no anti-tumor effect in Trop-2 negative colon cancer
cell line SW620 xenograft model. Irinotecan at 40 mg/kg twice weekly dosing of 4 doses
resulted in significant tumor growth inhibition. All tumor models continued to grow when
treated with non-binding negative control conjugates at the highest doses in all studies.
on May 14, 2018. © 2016 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2016; DOI: 10.1158/1535-7163.MCT-16-0431
A
C
Figure 1
B
on May 14, 2018. © 2016 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2016; DOI: 10.1158/1535-7163.MCT-16-0431
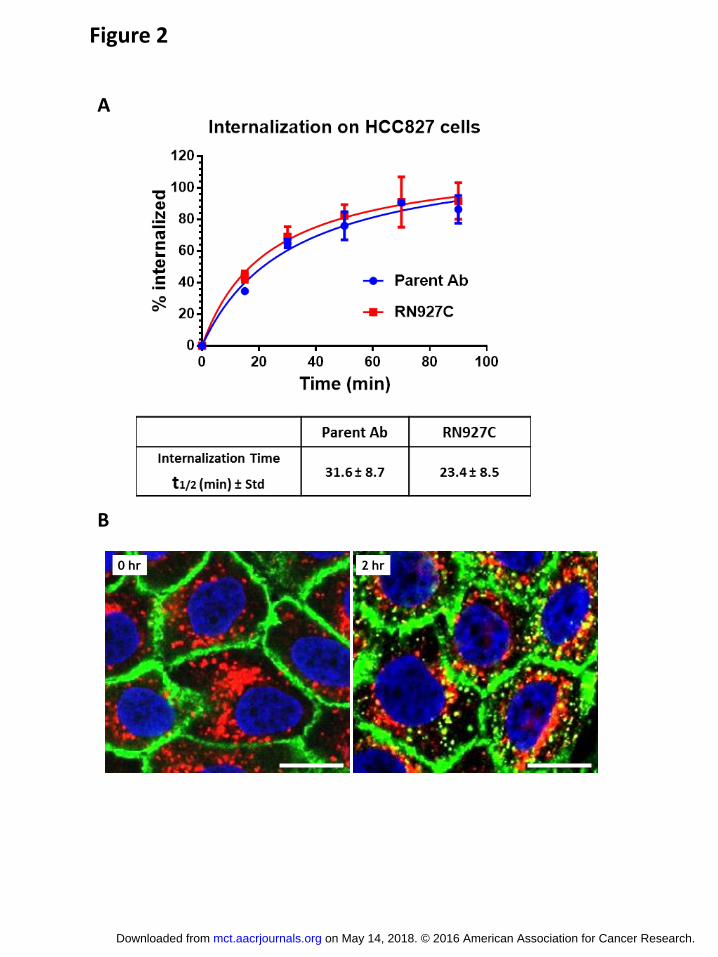
Figure 2
0 hr 2 hr
A
B
on May 14, 2018. © 2016 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2016; DOI: 10.1158/1535-7163.MCT-16-0431
BxPC3_mIgG
SW620_mIgG
BxPC3_aTrop2
SW620_aTrop2
A
B
C
Figure 3
untreated
RN927C 1 mg/mL RN927C 10 mg/mL
RN927C 0.1 mg/mL
untreated RN927C 10 mg/mL
Trop-2 expression on tumor cells
Phosphorylated histone H3 staining on BxPC3 cells
Phosphorylated histone H3 staining on SW620 cells
on May 14, 2018. © 2016 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2016; DOI: 10.1158/1535-7163.MCT-16-0431

A
B
Figure 4
on May 14, 2018. © 2016 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2016; DOI: 10.1158/1535-7163.MCT-16-0431
39 46 53 60 67 74 81 88 95 1021091161230
500
1000
1500
3.00 mg/kg neg ctrl conjugate
0.75 mg/kg RN927C
1.50 mg/kg RN927C
3.00 mg/kg RN927C
Pan0135
Days Post Tumor Implantation
Tu
mo
r V
olu
me (
mm
3)
(Me
an
SE
M)
Figure 5
A B
C D
G H
E F
on May 14, 2018. © 2016 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2016; DOI: 10.1158/1535-7163.MCT-16-0431

Published OnlineFirst August 31, 2016.Mol Cancer Ther Pavel Strop, Thomas-Toan Tran, Magdalena Dorywalska, et al. tumor modelswith enhanced stability, is highly efficacious in preclinical solid RN927C, a site-specific Trop-2 antibody-drug-conjugate (ADC)
Updated version
10.1158/1535-7163.MCT-16-0431doi:
Access the most recent version of this article at:
Material
Supplementary
http://mct.aacrjournals.org/content/suppl/2016/08/27/1535-7163.MCT-16-0431.DC1
Access the most recent supplemental material at:
Manuscript
Authoredited. Author manuscripts have been peer reviewed and accepted for publication but have not yet been
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://mct.aacrjournals.org/content/early/2016/08/27/1535-7163.MCT-16-0431To request permission to re-use all or part of this article, use this link
on May 14, 2018. © 2016 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on August 31, 2016; DOI: 10.1158/1535-7163.MCT-16-0431







