
Linköping University Medical Dissertation
No. 1361
The impact of helminth infection in patients with
active tuberculosis
Ebba Abate
Division of Medical Microbiology
Department of Clinical and Experimental Medicine
Faculty of Health Sciences
Linköping University
SE-58185 Linköping, Sweden
Linköping University 2013

© Ebba Abate, 2013
All rights reserved.
Papers I -III are reprinted with permission from the respective publishers.
Illustrations are created by the author and produced by Liu-Tryck.
ISBN 978-91-7519-653-4
ISSN 0345-0082
Printed in Sweden by LiU Tryck
Linköping 2013

The thesis is dedicated to my mother and brother who have left this world too soon

Supervisors
Thomas Schön (Ass. Prof)
Department of Infectious Diseases
Kalmar County Hospital and Linköping
University
Sweden
Olle Stendahl (Prof)
Department of Clinical and Experimental
Medicine
Medical Microbiology
Linköping University
Sweden
Abraham Aseffa (MD, PhD)
Director, Armauer Hansen Research
Institute
Addis Ababa, Ethiopia
Opponent
Zvi Bentwich (Prof)
Head, Center for Infectious Diseases and
AIDS
Ben-Gurion University
Israel
Evaluation Board
Jan Ernerudh (Prof)
Department of Clinical and Experimental
Medicine
AIR/Clinical Immunology
Linköping University
Sweden
Maria Jenmalm (Prof)
Department of Clinical and Experimental
Medicine
AIR/Clinical Immunology
Linköping University
Sweden
Peter Bergman (Ass. Prof)
Department of Laboratory Medicine
Division of Clinical Microbiology
Karolinska Institute
Sweden

Financial support
This work was supported by the Swedish Research Council, the Swedish Heart and Lung
Foundation, King Oscar II Jubilee Foundation, SIDA/SAREC and European and Developing
Countries Clinical Trials Partnership (EDCTP), Swedish Medical Association (SLS), the
Groschinsky Memorial Foundation and the Marianne and Marcus Wallenberg foundation.

Table of Contents
ABSTRACT .................................................................................................................................... 1
SUMMARY IN SWEDISH/SAMMANFATTNING PÅ SVENSKA ........................................... 3
LIST OF ORIGINAL PAPERS ...................................................................................................... 5
ABBREVIATIONS ........................................................................................................................ 7
BACKGROUND .......................................................................................................................... 10
Historical overview and global epidemiology of tuberculosis .................................................. 10
Mycobacterium tuberculosis ..................................................................................................... 10
Tuberculosis .............................................................................................................................. 10
Tuberculosis and HIV/AIDS ..................................................................................................... 11
Host susceptibility to tuberculosis............................................................................................. 12
Pathogenesis and host immune response to tuberculosis .......................................................... 13
The innate immune response to Mycobacterium tuberculosis .............................................. 13
The adaptive immune response during tuberculosis.............................................................. 14
The granuloma ....................................................................................................................... 15
Clinical characteristics of tuberculosis ...................................................................................... 16
Diagnosis of tuberculosis .......................................................................................................... 17
Treatment of tuberculosis .......................................................................................................... 18
Drug resistance in tuberculosis ................................................................................................. 19
Tuberculosis vaccines ............................................................................................................... 20
Classification, global epidemiology and public health impact of helminths ............................ 21
The host immune response to helminths ................................................................................... 22
The role of regulatory T-cells in health and helminth infection ............................................... 22
Helminth co-infection with HIV/AIDS ..................................................................................... 25

Helminths and non-infectious diseases ..................................................................................... 25
Impact of helminths on vaccine efficacy ................................................................................... 26
Helminth co-infection with tuberculosis ................................................................................... 27
AIMS............................................................................................................................................. 29
PATIENTS, MATERIALS AND METHODS ............................................................................. 30
Study setting............................................................................................................................... 30
Study participants (Paper I-V)................................................................................................... 33
Clinical and laboratory patient characteristics .......................................................................... 36
Baseline characteristics and the TB-score ............................................................................. 36
HIV testing and CD4+ cell counts ......................................................................................... 37
IgE, eosinophil cell count and Quantiferon measurement ..................................................... 37
Stool examination .................................................................................................................. 37
Sputum smear examination ................................................................................................... 38
Isolation of peripheral blood mononuclear cells (PBMCs) .................................................. 38
Analysis of regulatory T-cells by flow cytometry ................................................................. 38
Analysis of IFN-γ, IL-5 and IL-10 by ELISPOT .................................................................. 39
Statistics (Paper I-V) ................................................................................................................. 39
Ethical considerations ............................................................................................................... 40
RESULTS AND DISCUSSIONS (Paper I-V) ............................................................................. 42
CONCLUSIONS........................................................................................................................... 64
CONCLUDING REMARKS ........................................................................................................ 65
REFERENCES ............................................................................................................................. 68
ACKNOWLEDGEMENTS .......................................................................................................... 82

1
ABSTRACT
The geographic distribution of helminth infection and tuberculosis (TB) overlap substantially.
Experimental animal models and limited data from humans have shown that intestinal helminths
could subvert the host immune response towards a T-helper 2 (Th2)-type immune response and
an increased regulatory T-cell activity (Tregs). This in turn affects the host's ability to mount an
effective Th1 immune-mediated protection against Mycobacterium tuberculosis. However,
evidence for this hypothesis in the human setting from helminth infected TB patients is limited.
This thesis primarily focuses on the immunological and clinical impact of helminth infection on
pulmonary TB.
The kinetics of the Quantiferon-Gold (QFN) assay, which measures IFN-γ response to TB-
specific antigens in whole blood was assessed and showed a modest decline during TB treatment
to the level observed for healthy blood donors. We further assessed another clinical monitoring
tool, the-TB-score, composed of clinical signs and symptoms of TB, and found an early decline
two weeks after initiation of TB- treatment where a failure of decline correlated with increased
mortality. Overall, the helminth co-infection rate was significantly higher in TB patients
compared to healthy controls. Helminth co-infection was associated to a significantly higher rate
of eosinophilia and IgE-levels in healthy controls and patients with tuberculosis. During the first
weeks of anti-TB treatment, a marked decrease in the rate of helminth infection was observed in
HIV co-infected compared to HIV-negative TB patients. However, helminth co-infection was
more common in HIV negative than HIV positive TB patients. There was no detectable impact
of helminth infection on the clinical presentation of pulmonary tuberculosis. At baseline,
helminth co-infected TB patients showed an increased frequency of Tregs compared to helminth
negative TB patients and healthy controls. This was accompanied by an increased rate of PPD
stimulated IL-5 and spontaneous production of IL-10 by peripheral blood mononuclear cells
among helminth co-infected TB patients. A placebo controlled randomized trial was conducted
in order to test the hypothesis that albendazole treatment of helminth positive TB patients may
improve the clinical response of TB by reducing the immunmodulatory effect of helminthes on
TB immunity. A total of 140 helminth co-infected TB patients were randomized to albendazole
(400 mg per os for three consecutive days) or placebo. No significant difference was observed
between the albendazole and placebo group in terms of the primary outcome (TB score change

2
between baseline and week 8). Among the secondary outcomes, a significant decline of
peripheral eosinophil cells was observed in the albendazole treated group, but no effect on other
outcome variables (changes in chest x-ray findings, IgE level and sputum smear conversion).
Regarding the immunological assessment no significant difference was observed for changes in
Tregs, and PPD-induced production of IFN- γ or IL-5 although a non-significant trend of a
decrease in IL-10 expressing PBMCs were observed in the albendazole group.
Taken together, the burden of helminth infection was higher in TB patients than in a healthy
control group. Helminth co-infection during pulmonary TB in the human setting induces an
immune response characterized by increased IgE production, eosinophilia as well as increased
levels of Tregs and spontaneous IL-10 production. Thus, the immunological impact of helminth
infection on the outcome and risk for developing TB merits further investigation.

3
SUMMARY IN SWEDISH/SAMMANFATTNING PÅ SVENSKA
Det finns en stor geografiskt överlappning mellan utbredningen av maskinfektioner och områden
som är hårt drabbade av tuberkulos (TBC). Data ifrån experimentella djurmodeller och ett
begränsat antal humanstudier har visat att maskinfektioner generellt sett driver värdorganismens
immunförsvar mot ett T-hjälpaparcell-2 (Th-2)-dominerat cytokinsvar samt ger upphov till en
ökat antal regulatoriska T-celler (Tregs). Dessa immunologiska förändringar anses i sin tur
hindra värdorganismen i att frambringa det Th-1-dominerande cytokinsvar som är nödvändigt för
ett effektivt skydd gentemot Mycobacterium tuberculosis, bakterien som orsakar TBC. Det
primära målet för denna avhandling är att undersöka den immunologiska och kliniska betydelsen
av maskinfektion för lungtuberkulos.
Testresultaten för Quantiferon-Gold (QFN), ett helblods-baserat ELISA-test som kvantifierar den
cellulära produktionen av interferon-γ (IFN-γ) efter stimulering med TB-specifika antigen,
monitorerades hos TBC-patienter som genomgick behandling, och visade på en viss minskning
av IFN-γ-produktion under behandling, till nivåer jämförbara med en frisk kontrollgrupp. Vi
utvärderade även TB-score som är ett annat kliniskt hjälpmedel för att monitorera kliniskt
förlopp vid TBC. TB-score bygger på kliniska symptom och tecken, och våra resultat visar en
tydlig minskning två veckor efter påbörjad behandling där utebliven reducering av TB-score
korrelerade med ökad mortalitet.
Överlag fann vi en högre prevalens av maskinfektion i hos patienter med TBC jämfört med en
frisk kontrollgrupp. Hos båda grupperna var maskinfektion associerat med signifikant högre
förekomst av eosinofili samt ökade nivåer av IgE. Under första behandlingsveckorna mot TBC
observerade vi en tydlig minskning av andel HIV-positiva patienter med maskinfektion jämfört
med TBC-patienter utan HIV. Dessutom var maskinfektion vanligare hos HIV-negativa TBC-
patienter än TBC-patienter med HIV.
Vi fann ingen påverkan på den kliniska bilden vid diagnos av TBC i relation till maskinfektion.
Däremot var samtidig maskinfektion och TBC associerat med en högre andel Tregs jämfört med
TBC-patienter utan maskinfektion och friska individer. Likväl fanns en ökad andel IL-5
producerande vita blodkroppar i perfiert blod (PBMCs) efter PPD-stimulering samt en ökad
spontan produktion av IL-10 hos TBC-patienter med samtidig maskinfektion.
I en randomiserad placebo-kontrollerad studie undersökte vi hypotesen att behandling av
maskinfekterade TBC-patienter med albendazol, förbättrar svaret på TBC-behandling genom att
reducera den immunomodulerande effekten av maskinfektionen på värdförsvaret mot TBC.
TBC-patienter (n=140) med samtidig maskinfektion blev randomiserade till albendazol (400 mg
per os under tre dagar) eller placebo. Ingen signifikant skillnad återfanns hos de patienter som
behandlades med albendazol jämfört med placebo vad gäller förändring i TB-score mellan
behandlingens början och vecka 8, förändringar i lungröntgen, IgE nivåer eller

4
sputumkonversion. Däremot gav albendazol en signifikant minskning av eosinofila granulocyter
i perifert blod.
Vad gäller den immunologiska utvärderingen återfanns inga signifikanta skillnader mellan de
båda grupperna vad gäller Tregs och PPD-inducerad produktion av IFN-γ eller IL-5. Däremot
fanns en trend för minskade nivåer av IL-10 producerande PBMCs hos gruppen som
behandlades med albendazol.
Sammanfattningsvis fann vi en högre prevalens av maskinfektion hos TBC-patienter jämfört
med kontrollgruppen, och maskinfektion hos TBC-patienter gav upphov till ett immunsvar
karaktäriserat av ökade IgE-nivåer, eosinofili samt ökade nivåer av Tregs och spontan
produktion av IL-10. Dessa immunologiska förändringar styrker vikten av fortsatta kliniska
studier av maskinfektioners påverkan på läkningsförmåga vid aktiv TBC och risken hos
exponerade att utveckla TBC.

5
LIST OF ORIGINAL PAPERS
This thesis is based on the following publications and manuscripts, referred to in the text by their
Roman numerals:
Paper I
Idh J, Abate E, Westman A, Elias D, Janols H, Gelaw A, Getachew A, Alemu S, Aseffa A,
Britton S, Stendahl O, Schön T. Kinetics of the QuantiFERON-TB Gold In-Tube test during
treatment of patients with sputum smear-positive tuberculosis in relation to initial TST result and
severity of disease. Scand J Infect Dis. 2010 Sep; 42(9):650-7.
Paper II
Janols H, Abate E, Idh J, Senbeto M, Britton S, Alemu S, Aseffa A, Stendahl O, Schön T. Early
treatment response evaluated by a clinical scoring system correlates with the prognosis of
pulmonary tuberculosis patients in Ethiopia: a prospective follow-up study. Scand J Infect Dis.
2012 Nov;44(11):828-34.
Paper III
Abate E, Belayneh M, Gelaw A, Idh J, Getachew A, Alemu S, Diro E, Fikre N, Britton S, Elias
D, Aseffa A, Stendahl O, Schön T. The impact of asymptomatic helminth co-infection in patients
with newly diagnosed tuberculosis in north-west Ethiopia. PLoS One. 2012;7(8):e42901. doi:
10.1371.
Paper IV
Ebba Abate, Jonna Idh, Meseret Belayneh, Assefa Getachew, Shitaye Alemu, Ermias Diro,
Sven Britton, Daniel Elias, Abraham Aseffa, Olle Stendahl, Thomas Schön. Impact of helminth
infection on the clinical presentation of pulmonary tuberculosis. Manuscript.
Paper V
Ebba Abate, Daniel Elias, Assefa Getachew, Shitaye Alemu, Ermias Diro, Sven Britton,
Abraham Aseffa, Olle Stendahl, Thomas Schön. Effects of albendazole treatment on the clinical
outcome and immunological responses in patients with helminth infection and pulmonary
tuberculosis: a randomized clinical trial. Manuscript.

6
Additional work performed during the PhD period outside the scope of this thesis
1. Idh J, Mekonnen M, Abate E, Abraham Assefa, Elias D, Sundqvist T, Stendahl O, Schön T.
Nitric oxide resistance in Mycobacterium tuberculosis is associated with resistance against
first line drugs and poor clinical response during the initial phase of treatment. PLoS One.
2012;7(6):e39891. Epub 2012 Jun 29.
2. Mekonen M, Abate E, Aseffa A, Anagaw B, Elias D, Hailu E, Idh J, Moges F, Wolde-
Amanuel Y, Asrat D, Yamuah L, Britton S, Stendahl O, Schön T. Identification of drug
susceptibility pattern and mycobacterial species in sputum smear positive pulmonary
tuberculosis patients with and without HIV co-infection in north west Ethiopia. Ethiop Med
J. 2010; 48:203-10.
3. Schön T, Idh J, Westman A, Elias D, Abate E, Diro E, Moges F, Kassu A, Ayele
B, Forslund T, Getachew A, Britton S, Stendahl O, Sundqvist T. Effects of a food
supplement rich in arginine in patients with smear positive pulmonary tuberculosis-a
randomized trial. Tuberculosis (Edinb). 2011 Sep;91(5):370-7. doi:
10.1016/j.tube.2011.06.002. Epub 2011 Aug 2.

7
ABBREVIATIONS
Ab Antibody
AFB Acid fast bacilli
Ag Antigen
AHRI Armauer Hansen Research Institute
AIDS Acquired Immune Deficiency Syndrome
APC Antigen presenting cells
ART Anti-retroviral therapy
BCG Bacillus Calmette Guérin
BMI Body mass index
BSL Biosafety level
CC Community control
CD Cluster of differentiation
CFP Culture filtrate protein
CMI Cell mediated immunity
CNS Central nervous system
DC Dendritic cell
DOTS Directly Observed Treatment, Short-course
DSMB Data and safety monitoring board
ELISA Enizyme -linked immuno sorbent assay
ELISPOT Enizyme- linked immunospot assay
ESAT Early secretory antigenic target
FBS Fetal bovine serum
FCS Fetal calf serum
FITC Fluorescein isothiocyanate
FMO Fluorescence Minus One
FMOH Federal Ministry of Health

8
FOXP3 Forehead box P3
GM-CSF Granulocyte macrophage colony stimulating factors
HAART Highly active anti retroviral therapy
HIV Human immunodeficiency virus
HLA Human leukocyte antigen
IFN-γ Interferon-gamma
Ig Immunoglobulin
IGRA Interferon-gamma release assay
IL Interleukin
IUTLD International Union against Tuberculosis and Lung Diseases
MDR-TB Multi-drug-resistant tuberculosis
MHC Major histocompatibility complex
MNL Mediastinal lymphnode
MUAC Mid-upper arm circumference
NKs Natural killer cell
NNRTI Non-nucleoside reverse-transcriptase inhibitors
NO Nitric-oxide
NRTI Nucleoside reverse transcriptase inhibitor
PBMCs Peripheral blood mononuclear cells
PBS Phosphate Buffered Saline
PE Phycoerythrin
PIHCT Providers initiated HIV counseling and testing
PPD Purified protein derivative
QFN QuantiFERON
RIF Rifampicin
RNA Ribonucleic acid
RPO Research and publication office

9
SD Standard deviation
SEA Soluble egg antigen
SFU Spot forming unit
TB Tuberculosis
Th T- helper
TLR
TNF Tumor necrosis factors
Treg Regulatory T-cell
TST Tuberculin skin test
UNAIDS Joint United Nations Programme on HIV/AIDS
WHO World Health Organization
XDR-TB Extensively drug-resistant tuberculosis

10
BACKGROUND
Historical overview and global epidemiology of tuberculosis
Tuberculosis (TB) is an infectious disease caused by bacilli belonging to the Mycobacterium
tuberculosis complex [1]. This complex consists of seven species including Mycobacterium
tuberculosis (M.tuberculosis), M. canetti, M. africanum, M. pinnipedii, M. microti, M. caprae
and M. bovis [2]. Tuberculosis has a long history and coexisted with humans since ancient times
[3]. It was reported that all modern members of the M. tuberculosis complex had a common
African ancestor [4].
Mycobacterium tuberculosis
Mycobacterium tuberculosis is a fastidious, slow growing, lipid-rich, rod shaped bacterium,
which resists decolorization with acid-alcohol. The bacterium has a slow growth rate of 12-16
hours compared most other bacteria, which have a generation time measured in minutes [5]. The
cell wall of M. tuberculosis is rich in lipids which contribute to the acid fastness and
hydrophobicity. The waxy coat also contributes to the resistance to many disinfectants, common
laboratory stains as well as to antibiotics [6].
Tuberculosis
Tuberculosis is predominantly a disease of the lung, with pulmonary TB accounting for about
70% of the cases. Extra-pulmonary disease sites include lymph nodes, bone, and meninges [7].
There are 22 high-burden countries, which account for approximately 80 % of the estimated
number of new TB cases [1]. The consequences of TB on society are immense. Worldwide, it
has been estimated based on tuberculin skin test (TST) positivity, that one person out of three is
infected with M. tuberculosis [8]. In 2012, there were 8.7 million new cases of TB globally, with
1.4 million deaths in the same year, which confirms TB as a serious global health issue [1]
(Figure 1).
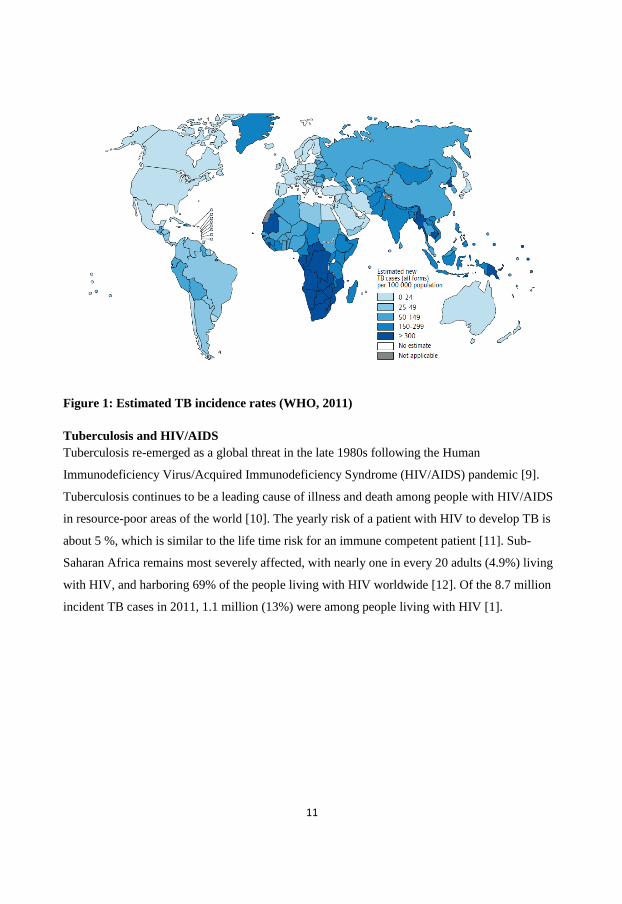
11
Figure 1: Estimated TB incidence rates (WHO, 2011)
Tuberculosis and HIV/AIDS
Tuberculosis re-emerged as a global threat in the late 1980s following the Human
Immunodeficiency Virus/Acquired Immunodeficiency Syndrome (HIV/AIDS) pandemic [9].
Tuberculosis continues to be a leading cause of illness and death among people with HIV/AIDS
in resource-poor areas of the world [10]. The yearly risk of a patient with HIV to develop TB is
about 5 %, which is similar to the life time risk for an immune competent patient [11]. Sub-
Saharan Africa remains most severely affected, with nearly one in every 20 adults (4.9%) living
with HIV, and harboring 69% of the people living with HIV worldwide [12]. Of the 8.7 million
incident TB cases in 2011, 1.1 million (13%) were among people living with HIV [1].

12
Figure 2: Estimated HIV prevalence in new TB cases (WHO, 2011)
Tuberculosis and HIV/AIDS in Ethiopia
Tuberculosis continues to be one of the major public health concerns in Ethiopia fueled by the
expansion of the HIV pandemic since the 1990s. According to the 2012 WHO TB report,
Ethiopia was ranked 7th
among the world’s 22 high burden countries with an estimated incidence
of 258 per 100,000 population, and a TB mortality of 18 per 100,000 individuals. The reported
HIV prevalence in the incident TB cases was 17 per 100,000 individuals [1].
In addition to a high TB burden, Ethiopia has been seriously affected by HIV/AIDS, with an
estimated 1.5 million people living with HIV [12]. The high rate of chronic malnutrition,
poverty, overcrowding, in combination with a high sero-prevalence of HIV infection has created
an environment making TB a very serious health problem in Ethiopia [13].
Host susceptibility to tuberculosis
The host susceptibility to TB and the clinical course of the infection depend on a complex
interplay between host, bacterial as well as environmental factors, such as poverty, malnutrition,
overcrowding, and exposure to other pathogens [14-16]. Several genetic factors have been shown
to influence host susceptibility to TB. Polymorphisms in certain genes encoding for natural-
resistance-associated macrophage protein (NRAMP1), the interleukin-1 (IL-1) gene cluster, the

13
vitamin D receptor and mannose-binding lectin have been associated with susceptibility to TB.
The essential role of T-helper 1 (Th1) associated cytokines such as IFN-γ and IL-12 was
confirmed by the increased susceptibility to mycobacterial infections in patients with mutations
in genes coding for these cytokines [17,18].
Pathogenesis and host immune response to tuberculosis
M. tuberculosis is an air borne pathogen and following infection alveolar macrophages are the
primary target cells for inhaled mycobacteria [19]. If the bacteria are not directly killed by the
innate host response which may be the case in up to 50 % of exposed individuals, the infection is
controlled in part by walling it off from the rest of the uninfected lung in distinct foci known as
granulomas [20]. The immune response against TB plays a fundamental role in the outcome of
M. tuberculosis infection. M. tuberculosis is a pathogen capable of causing both progressive
disease and latent infection [8,21]. After successful control of the primary TB infection, some
bacilli may remain in a non-replicating or slowly replicating dormant state for the rest of the life
of the individual. This infectious state, defined as latent TB infection, is clinically asymptomatic.
Control of M. tuberculosis infection is mainly dependent on the success of the interaction
between innate and adaptive immune response of the host.
The innate immune response to Mycobacterium tuberculosis
Phagocytosis of M. tuberculosis by macrophages represents the first major host-pathogen
interaction in TB [22]. Recognition of M. tuberculosis by phagocytic cells is through surface
receptors including Toll-like receptors (TLRs), complement receptors (CR), mannose receptors,
scavenger receptors, and dendritic cell (DC)-specific intercellular-adhesion-molecule-3-grabbing
non-integrin (DC-SIGN). This recognition causes activation of phagocytic cells and production
of cytokines and pro-inflammatory cytokines such as TNF-α, IL-1-β, IL-6, IL-12 and IFN-γ. In
relation to innate immune effector responses, vitamin D is also involved in the killing of M.
tuberculosis through the production of the peptide cathelicidin [23].
Neutrophils are among the first cells to respond to inflammatory stimuli by migrating to the site
of infection where they kill pathogens by both reactive oxygen species (ROS) and antimicrobial
peptides [24]. The role of neutrophils in host immunity to TB through mechanisms including
apoptosis have been previously shown [25,26]. Other cells of the host innate immune system of
importance for the defense against M. tuberculosis include DCs, natural killer cells (NK) and

14
epithelial cells [27,28]. Innate effector mechanisms against M. tuberculosis include phago-
lysosomal fusion, reactive oxygen and nitrogen intermediates and antimicrobial peptides such as
cathelicidin induced by vitamin-D [29]. Natural killer cells may kill intracellular M. tuberculosis
by the pore-forming perforin, where the anti-mycobacterial factor granulysin binds to the
bacterial cell surface, and disrupts the membrane causing osmotic lysis of the bacteria [30].
However, this mechanism is more pronounced when M. tuberculosis specific CD8+ T-cells are
activated [31]. The major defense mechanisms within the macrophages include low pH, ROS
produced by NADPH oxidase, reactive nitrogen species (RNS) produced by the inducible nitric
oxide synthase (iNOS), iron (Fe2+) scavengers and exporters, proteases and antimicrobial
peptides such as cathelicidin [32]. Apoptosis is an important mechanism for the infected host cell
to limit replication of M. tuberculosis. Apoptosis of phagocytic cells may prevent dissemination
of infection as well as reduce the viability of intracellular mycobacteria [33].
In murine models it has been shown that nitric oxide (NO) is essential for host defense against
M. tuberculosis, but the relative importance of NO has not been fully established in humans
[34,35]. At the site of TB infection in humans, the presence of iNOS and nitrotyrosine in
macrophages has been shown as indicators for NO production [36]. Idh et al., reported low level
NO in both HIV negative and HIV co- infected TB patients compared to blood donors and
house-hold contacts [37]. In a randomized study conducted in Gondar, Ethiopia, supplementation
of peanuts (rich in arginine) in conjunction with anti-TB drugs increased exhaled NO production
and enhanced clinical improvement during anti-TB treatment in HIV-coinfected TB patients
[38].
The adaptive immune response during tuberculosis
Following activation, dendritic cells are recruited to the lung where they transport mycobacterial
antigens to the mediastinal lymphnode (MLN). Within the MLN, antigen presenting cells (APCs)
activate antigen-specific T-cells. The infected macrophages and DCs secrete cytokines including
IL-7, IL-12, IL-15, IL-23 and TNF-α, leading to attraction of more leukocytes to the infection
site [39]. Due to the nature of TB infection, the majority of bacilli reside within an endosome and
M. tuberculosis antigens are presented mostly on major histocompatibility complex (MHC) class
II [40]. However, it has also been shown that M. tuberculosis antigens could be presented by
MHC class I pathways activating CD8+ cytolytic T-cells which kill M. tuberculosis by the
granulysin-perforin system [31]. CD4+ T-cells may differentiate into Th1, Th2, Th17 and

15
regulatory T-cells (Tregs) depending up stimuli. The Th1 response primarily results in increased
IFN-γ production and subsequent activation of macrophages, with killing of intracellular bacteria
by phagolysosome fusion and effector mechanisms of the macrophage such as nitric oxide. In
contrast, the Th2 response produces B- lymphocyte stimulating factors (IL-4, IL-5, IL-10 and IL-
13), which suppress the Th1 response. Th17 cells, stimulated by IL-23, IL-6, IL-21, and low
TGF-β levels, are involved in recruitment of cells of the innate immune system and Th1 cells,
and secrete IL-17. Regulatory T- cells produce anti-inflammatory cytokines such as IL-10 and
TGF-β, which have the capacity to suppress microbicidal mechanisms in the macrophage. The
role of B cells in TB is not clear, but has been recognized as potentially protective [41]. Of
relevance for the Th1/Th2 balance and the effector function of the macrophage, stimulation of
the macrophage with IFN-γ, TNF- or IL-1 will lead to a M1 phenotype, with up-regulation of
iNOS, NO-production and antimicrobial activity. Stimulation with the Th2-related cytokines IL-
4, IL-10 or TGF- will generate a M2 phenotype, with increased arginase activity beneficial for
tissue repair but not for bacterial killing [29].
The granuloma
The granuloma is formed through recruitment of immune cells during the innate response and
consolidated by adaptive immunity, as a result of complex cytokine and chemokine signals. The
formation of a granuloma is a dynamic process that begins shortly after infection and
continuously evolves over time. It can be divided into three distinct phases, the “innate
granuloma,” which is a loose aggregate composed primarily of recruited macrophages and
neutrophils; the “immune granuloma” formed following the emergence of antigen-specific T-
cells; and the “chronic granuloma,” resulting from distinct morphological changes in granuloma
structure [42]. The cellular core of the granuloma contains infected macrophages, and occasional
multi-nucleated giant cells surrounded by dendritic cells, neutrophils, eosinophils, T and B- cells
and fibroblasts [43]. The structure and functions of the granuloma are regulated and controlled
by the complex interplay between cytokines which includes IFN-γ and TNF-α. Regulatory
cytokines including IL-10 and TGF-β may undermine granuloma maintenance [44]. In the case
of an impaired immune response, such as during HIV-infection, the immunological balance is
disturbed, which may lead to tissue damage from necrosis as a result of bacterial dissemination
[45]. It is controversial whether the granuloma may actually provide a shelter for M. tuberculosis

16
(especially during the early stages of infection) or is part of the host protective function during
the later stages of the infection.
Figure 3: Schematic illustration of a TB granuloma: Infected macrophages and dendritic cells
are surrounded by bystander macrophages, neutrophils, and subsets of T -lymphocytes where the
cells are enclosed by a fibrotic cuff (left). M. tuberculosis can be contained within the granuloma
for long periods, but if immune control fails as a result of HIV-infection or other factors, the
bacilli start to replicate, and a necrotic granuloma core develops. The granuloma then ruptures
and M. tuberculosis is dispersed into the airways (right).
Clinical characteristics of tuberculosis
The lungs are the main site of disease for pulmonary TB and are also the primary route of
infection. For pulmonary TB, the universal clinical symptom is cough for more than two weeks,
which is initially dry but in rare cases blood stained sputum could be produced as the disease
progresses [46]. The other most common clinical manifestations after cough include fever,

17
malaise, fatigue, sweating, and weight loss, [46]. Upper lobe infiltrates as well as fibrosis with or
without cavitation are common chest X-ray presentations in active TB [47].
Diagnosis of TB
The laboratory diagnostic methods used for detection and identification of TB include smear
microscopy, culture and molecular techniques involving specific nucleic acid markers [48].
However, culture is still the “gold standard” for the diagnosis of active TB [49]. Smear
microscopy of acid fast-bacilli (AFB) is often the only available diagnostic method in low
resource countries [50]. For the diagnosis of pulmonary TB, at least two consecutive sputum
specimens including one morning sputum should be collected, based on the Directly Observed
Treatment Short-course drug therapy (DOTS) guidelines by the WHO recommendations [51].
Although several methods can be used to visualize acid-fast organisms, the Ziehl-Neelsen
technique is the most widely used method in resource poor countries [52,53].
The culture media used for the isolation of mycobacteria include egg-based and agar-based
media [49,54,55]. All isolates from cultures reported positive for mycobacteria should be
identified to the level of species using either biochemical or molecular methods [56]. Recently, a
nucleic-acid amplification based assay, Xpert MTB/RIF assay (Cepheid-Australia), has been
introduced. The Xpert MTB/RIF assay is a fully automated molecular assay in which real-time
polymerase chain reaction technology is used to simultaneously detect M. tuberculosis and
rifampicin resistance mutations in the rpoB gene [57]. In 2010, WHO endorsed the roll-out of
this test for the investigation of TB suspects, especially in settings with a high prevalence of
HIV-associated disease and MDR-TB [58]. The assay requires minimal laboratory expertise, and
results are available within 2 hours. Even though the assay enables rapid results and is less labor
intensive, it has some drawbacks regarding the high cost of the instrument and cartridges. In
addition, issues related with proper maintenance and services are of relevance for high endemic
rural areas. Additionally, reports of false positive rifampicin resistance results merits further
assessment of the assay [59].
Immunological diagnostic tools
A gold standard for the diagnosis of latent TB is not yet available. The century old tuberculin
skin test (TST) and the more recently developed IFN-γ release assays (IGRAs) are used as
screening tools for latent TB infection [60]. IGRAs are slightly more specific than TST and both

18
tests have a high negative predictive value but a poor positive predictive value (2-3 %) in
estimating the risk for developing active disease [61].
Tuberculin skin test (TST)
The TST has been used for more than 100 years [62,63] and has been widely employed to
measure the prevalence of TB infection in the community as well as to select TST-negative
patients for BCG vaccination. The TST is administered intradermally by injecting 0.1 mL of 5
Tuberculin Units of Purified Protein Derivative (PPD) on the dorsal surface of the forearm. After
48-72 hours, a positive reaction (usually defined as > 10 mm) is indicated by induration [64].
Individuals with prior infection with M. tuberculosis will mount an immune response to
mycobacterial proteins [62]. However, TST is associated with limitations such as low sensitivity
in individuals with impaired cellular immunity as well as false positive results in individuals
previously vaccinated with BCG or exposed to environmental mycobacteria [65,66].
Interferon-γ release assays (IGRAs)
The IGRA assays are based on the principle that T-cells of TB sensitized individuals produce
IFN-γ when they re-encounter M. tuberculosis antigens [67,68]. Recent evaluations showed that
IFN-γ assays that use M. tuberculosis RD1 antigens, such as ESAT6 and CFP10 have advantages
over TST because they are specific to M. tuberculosis and are not affected by BCG or other
Mycobacterium species with a few rare exceptions [69,70]. In contact investigations, the results
of IGRAs had a better correlation with measures of exposure than TST [60,71]. Since 2005,
IGRAs have increasingly been used for the detection of latent TB infection either as replacement
or as adjunct to the tuberculin skin test [72].
Treatment of tuberculosis
The history of TB changed dramatically after the introduction of anti-mycobacterial agents. The
use of anti-TB drugs started in the 1940-ies, through the discovery of streptomycin and para-
aminosalicylic acid [73]. In TB-endemic areas TB-treatment is administered through DOTS
programs, where patients are observed when they take their medication to ensure compliance, as
non-compliance is a major contributor to the development of antibiotic resistance [74].

19
Anti-TB drugs are grouped into first line and second line drugs. First-line anti-TB drugs are
isoniazid, rifampicin, ethambutol and pyrazinamide [75]. Other drugs such as aminoglycosides
(amikacin and kanamycin), fluroquinolones, capreomycin, para-aminosalicylic acid and
thioamides (ethionamide and prothionamide) are used as second-line or alternative agents to treat
TB when the bacilli are resistant to first-line drugs [75]. The current short-course treatment
guideline aims for a complete elimination of active and dormant bacilli and involves two phases.
During the initial phase four drugs (usually isoniazid, rifampicin, pyrazinamide and ethambutol )
are given for two months [76]. The continuation phase in which fewer drugs (usually isoniazid
and rifampicin) are administered for an additional 4 months, targets the killing of any remaining
or dormant bacilli in order to prevent relapse. As a routine, all sputum-positive patients on DOTS
should have repeated sputum specimens examined at the end of the 2nd
and 6th ‘
month, and
treatment outcomes are reported through the WHO guideline as cured, treatment completed,
defaulted, transferred out and treatment failure [58].
Drug resistance in tuberculosis
Multi-drug resistant (MDR) TB is defined by resistance to at least rifampicin (RIF) and isoniazid
whereas extensively drug-resistant (XDR) TB is defined as MDR-TB with additional resistance
to any fluoroquinolone and to at least one of the injectable drugs (capreomycin, kanamycin and
amikacin). The prevalence of MDR and XDR-TB has been increasing with alarming rate [1].
The emergence of drug resistance has posed a major challenge on the control of TB globally.
Drug resistance in TB is the result of spontaneous mutation in combination with selection by
poor programmatic and individual care performance [77]. The effect of anti-TB drug resistance
has been around since the first anti-tuberculosis agent, streptomycin, appeared on the market as a
mono-therapy [78]. The problem of increasing resistance to M. tuberculosis is also of
significance to African countries such as Ethiopia with a high burden of disease. The recent
Ethiopian national estimate of MDR-TB was 1.6% and 11.8% for new and re-treatment patients,
respectively [13]. However, this is likely underestimated considering the limited TB diagnostic
and identification facilities in the country and the need for a regular update of resistance survey
data.

20
Tuberculosis vaccines
The current vaccine against TB, bacille Calmette-Guérin (BCG), is a live vaccine derived from
an attenuated strain of M. bovis. It is an inexpensive vaccine that has been applied since decades
and has a long-established safety profile. Studies have shown that BCG gives good protection
against disseminated TB in young children [79]. Natural exposure to environmental
mycobacteria [80,81], nutritional and socio-economic diversity, genetic differences and co-
infection with other endemic infections such as helminths have been suggested as responsible for
the low protection in adults [14,82-84].
Thus, there is a great need to develop a vaccine that gives better protection against TB.
However, the development of new vaccines against pulmonary TB has proved to be challenging
[84]. In the last ten years more than ten vaccine candidates have been developed [82] .The
approach to develop a TB vaccine includes a replacement vaccine for BCG or a booster vaccine,
a post-exposure vaccine or a therapeutic vaccine. A newly developed booster vaccine containing
two of the M. tuberculosis antigens secreted in the acute phase of infection (Ag85B and ESAT-
6), delayed and reduced clinical disease in cynomolgus macaques exposed to M. tuberculosis
[85]. Another promising approach is the recombinant live vaccine, VPM1002, which is a
recombinant BCG strain expressing listeriolysin of Listeria monocytogenes. The rationale behind
this strategy is the assumption of improved release of BCG-derived antigens into the cytosol and
increased apoptosis of host cells in vitro [86]. However, it appears unlikely to have a protective
vaccine on the market in the coming decade [11]. The very recent phase 2b randomized placebo
controlled clinical trial with MVA85A conducted in South Africa on infants previously
vaccinated with BCG, did not show any protective results [87].

21
Helminths
Classification, global epidemiology and public health impact
Helminths, a Greek word meaning “worms”, are multicellular organisms characterized by
elongated, flat or round bodies. The definitive classification is based on the external and internal
morphology of egg, larval, and adult stages and helminths are primarily divided into two major
phyla: platyhelminths (includes trematodes and cestodes) and round worms (nematodes) [88].
Humans are infected after ingesting eggs from contaminated soil or food, which are referred as
soil transmitted helminths (Ascaris lumbricoides and Trichuris trichiura), or through active
penetration of the skin by infective larval stages present in contaminated soil (hookworm) [89].
The most prevalent helminth species affecting about one-third of the world’s population living in
resource poor regions are A. lumbricoides, T. trichuira and hookworm (Necator
americanus and Ancylostoma duodenale) [90-92].
Infections by most soil-transmitted helminths are asymptomatic, especially in adults. In children,
soil-transmitted helminth is one of the world’s most important causes of physical and intellectual
growth retardation and malnutrition. Helminths induce tissue reactions, such as granuloma, and
provokes intestinal obstruction or rectal prolaps, especially in children [93]. Studies have also
highlighted the profound effect of soil-transmitted helminth infection on school performance and
attendance and future economic productivity [94,95]. Yet, despite all these consequences,
helminth infection is an area which remains largely neglected. It has been suggested that this
neglect originates from three features: (i), the people most affected are the world’s most
impoverished, particularly those who live on less than US$2 per day; (ii), the infections cause
chronic health problems and have subtle clinical presentation; and (iii) a difficulty of quantifying
the effect of soil-transmitted helminth infections on economic and educational development [89].
However, in the last decade it has become clear from revised estimates that the combined disease
burden of helminth infections might be as great as that of malaria or TB [96]. Furthermore, it has
been described that even asymptomatic helminth infection might increase host susceptibility,
affecting other diseases like HIV/AIDS, TB and malaria [14,97-100].

22
The host immune response to helminths
Helminth parasites are an evolutionarily ancient and diverse group [101], but show species-
independent modes of immune modulation in clinical and experimental studies. Because of such
character they have been described as a “master of immunoregulation” capable of escaping the
host defense system through suppression of both Th1 and Th2 associated host immunity and of
establishing chronic infections [93].
Helminths induce a strong Th2-type immune responses characterized by production of cytokines
such as IL- 4, IL-5, IL-9, and IL-13 and increased levels of circulating IgE antibodies,
eosinophils, and mast cells [93]. CD4+ Th2-lymphocytes are the central player in the helminth-
induced immune response by the production of cytokines and chemokines such as CCR3. It has
been shown that mice depleted of CD4+ T-cells, did not mount a protective immune response
following vaccination with Schistosoma mansoni, and lacked the ability to expel the intestinal
helminths [102,103]. The Th2 dominance in many infections is maintained by IL-10 and TGF-β
mediated suppression of competing Th1 and Th17 cell populations [83]. This has been shown in
a mouse filariasis model using L. sigmodontis where the chronic phase of infection is marked by
T-cell anergy, loss of proliferative responses to parasite antigen challenge, reductions in effector
cytokine levels, and elevated expression of inhibitory immune molecules and cells such as, IL-
10, TGF-β and Tregs.
The role of regulatory T-cells in health and helminth infection
Regulatory T-cells are a sub-population of T-cells, which modulate the immune system in order
to prevent tissue damage from pro-inflammatory responses, maintain tolerance to self-antigens,
and abrogate autoimmune disease [104]. These cells can be divided in to two subsets, natural
Tregs that develop in the thymus, and induced Tregs (Tr1, Th3) that arise from conventional
CD4+ T-cells in the periphery, a process promoted by chronic antigen exposure [105].
Regulatory T-cells express CD25, the α chain of the IL-2 receptor. The quantitative identification
of Tregs requires reliable surface markers that are selectively expressed on Tregs. The
forkhead/winged helix transcription factor (Foxp3) is an important marker to identify these
subsets [106], but it may be transiently expressed on activated CD4+ T-cells. CD127 is another
marker expressed on Tregs which inversely correlates with Foxp3 and is considered an
improvement in the definition of Tregs [107].

23
Expansion of both natural and induced Treg populations has been demonstrated in human
helminth infections including an increased frequency of Foxp3 expressing Tregs [108]. These
cells pose a more potent immune suppression in helminth infected mice than helminth uninfected
mice [109,110]. Similarly, in human filarial and Ascaris infections, the presence of helminths
correlate with both the production of IL-10 and TGF-β [111], and generalized T-cell hypo-
responsiveness [112]. In addition, the observed epidemiological association of increased rate of
helminth infection with less allergic manifestations and autoimmune inflammatory conditions in
humans is linked to an attenuated immune response by Tregs [113-115].
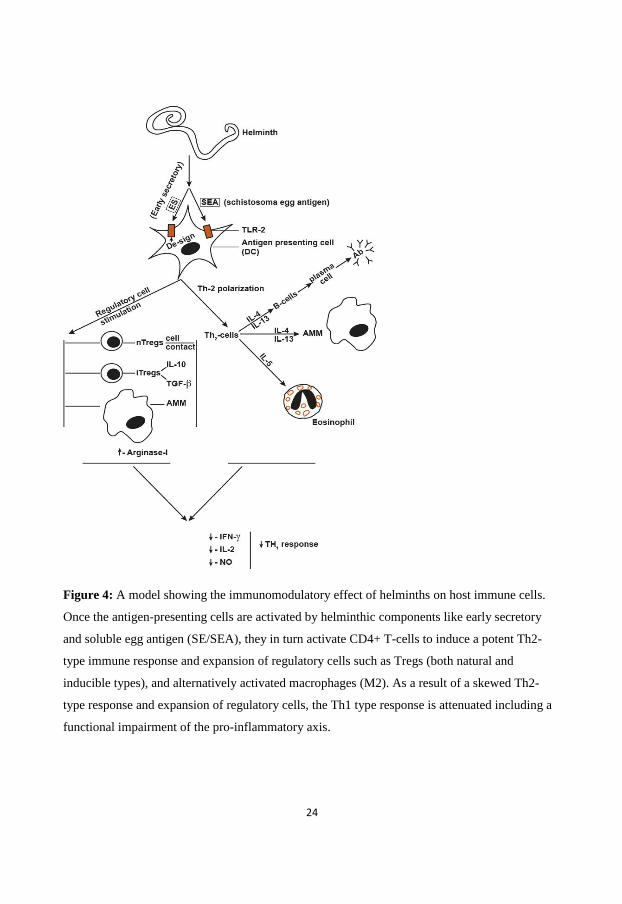
24
Figure 4: A model showing the immunomodulatory effect of helminths on host immune cells.
Once the antigen-presenting cells are activated by helminthic components like early secretory
and soluble egg antigen (SE/SEA), they in turn activate CD4+ T-cells to induce a potent Th2-
type immune response and expansion of regulatory cells such as Tregs (both natural and
inducible types), and alternatively activated macrophages (M2). As a result of a skewed Th2-
type response and expansion of regulatory cells, the Th1 type response is attenuated including a
functional impairment of the pro-inflammatory axis.

25
Helminth co-infection with HIV/AIDS
Following the HIV epidemic, infection with both helminths and HIV-1 is common in resource
poor areas [116]. Epidemiological cross-sectional studies show an association of helminth
infection with HIV [117,118]. However, this was more common with protozoan opportunistic
parasites such as Cryptosporidium parvum. There was a decline in the rate of helminth-HIV
coinfection before as compared to after the era of anti-retroviral therapy in Ethiopia [117,119-
122].
The immunological influences of helminths have been investigated in several human diseases
including HIV and TB. Increased expression of co-receptors of HIV-1 chemokines on T-
lymphocytes and monocytes obtained from helminth infected individuals has been reported
[123,124]. Peripheral blood mononuclear cells (PBMCs) obtained from helminth-infected
individuals show increased susceptibility to HIV-1 infection [125,126]. An increased risk of
mother to child transmission of HIV was observed in pregnant women co-infected with
helminths compared to controls [127]. Furthermore, a decreased CD8+ cytolytic HIV-1-specific
T- cell response, and increased IL-10 production were observed in HIV patients co-infected with
S. mansoni compared to those with HIV-1 infection only [128]. In a placebo controlled trial,
Ascaris co-infected HIV positive patients treated with albendazole showed a significant increase
in peripheral CD4+ T-cells cells and a decreasing trend in HIV-1 RNA level compared to the
placebo group [129].
Helminths and non-infectious diseases
The difference in the prevalence of allergic diseases between urban versus rural and between
economically poor and industrialized countries has launched the “hygiene hypothesis” [130]. An
increased colonization with helminths may lead to a decrease in allergic and autoimmune
diseases possibly by increased activation of anti-inflammatory responses induced by helminths.
Studies on the interactions between helminths and allergic immune responses have shown
variable results, ranging from decreased rate of allergy in helminth infected individuals [131],
increased risk of allergy [132,133] and the lack of any association [134]. Most of the
epidemiological and experimental studies show an important role of helminths in suppressing the
occurrence and outcome of autoimmune diseases [128,135-137]. In mouse models, infection

26
with S. mansoni, inhibited the development of type1 diabetes [128] . Helminths were found to be
protective in several models of inflammatory bowel disease [135,138]. Currently, Trichuris suis
is used to treat patients with ulcerative colitis, Crohn’s disease and multiple sclerosis
[136,137,139].
Impact of helminths on vaccine efficacy
The lack of efficient responsiveness to oral vaccines like cholera, polio and rotavirus in
developing countries initiated the hypothesis that the presence of helminths in the gastrointestinal
tract might have affected efficient uptake of theses vaccines [140]. It was shown that A.
lumbricoides infection impaired the immune responses to oral cholera vaccine by decreasing
seroconversion [140,141]. The negative impact of helminths in the response to vaccines like
BCG has also been shown in Ethiopia. In one study, BCG vaccination improved PPD-specific
IFN-γ production and T-cell proliferation from PBMCs in albendazole-treated healthy subjects
with intestinal helminths compared to controls [142]. It was further shown that BCG-vaccinated
mice with prior S. mansoni infection had significantly higher TB bacillary load as measured by
viable count. In addition, lower levels of IFN-γ and nitric oxide together with increased levels of
IL-4 and IL-5 were observed in mice with prior S. mansoni infection [143].
Other studies demonstrating an attenuated vaccine response in helminth positive individuals are
in agreement with the results observed for BCG vaccination [144-148]. Filarial infections
decreased the responsiveness to tetanus toxoid after tetanus vaccinations [149]. In another study,
a tetanus antitoxin level was measured two months after immunization with tetanus toxoid
vaccine, and only 7.1% of patients infected with onchocerciasis became immunized compared to
44.5% in the control group free from onchocerciasis [150]. Such findings underline the influence
of helminth infections on vaccine efficacy, and thus warranted the need to consider this issue
when designing and testing new vaccines. Indeed, without deworming prior to vaccination, as it
has previously been used empirically in veterinary medicine, the efficacy of vaccines could be
compromised in endemic areas for helminth infection. Therefore, a better understanding of the
interplay between the immune response of the host and the impact of co-infections is needed.

27
Helminth co-infection with tuberculosis
The geographic distributions of helminths and TB overlap substantially, increasing the likelihood
of co-infection with both pathogens [119]. Both helminths and M. tuberculosis use several
independent mechanisms to affect host immune responses and these mechanisms may interact
with important consequences for the immunology and outcome of both infections [14,151,152].
Epidemiological cross-sectional and case-control studies evaluating the prevalence and
association of helminth infection in active TB showed that these infections indeed coexist.
Tristao-SR et al., found that a high intestinal nematode infection rate in pulmonary TB patients
[153]. An increase in the prevalence of helminth co-infection in TB was reported in Gondar,
Ethiopia [121,154]. Moreover, in the same setting an increased rate of helminth infection among
active TB patients compared to healthy community controls was recently shown [119].
Furthermore, significantly higher frequency of intestinal helminths was reported in patients with
the lepromatous and multibacillary form of leprosy than patients with paucibacillary leprosy or
to a control group without leprosy [155]. Even though those epidemiological studies are unable
to address the immunological aspects behind the findings, it is clear from experimental models
and limited data from human infection that immunomodulation induced by helminths may affect
the ability of the host to control infection and disease from M. tuberculosis.
The findings of many of the experimental studies are in favor of the negative impact of helminths
on TB infection and risk of developing TB after exposure. The Th1 immune-mediated protection
against M. tuberculosis is characterized by strong M. tuberculosis-specific Th1 responses and co-
infection with helminths could modulate these immune responses through influence on the
Th1/Th2 balance with increased Th2 dominance as well as increased activity of Tregs [156].
Impairment of Th1 immune response characterized by a decrease in PPD-specific IFN-γ and IL-
17 responses was shown during chronic filarial infection [157]. In this context, it was reported
that peripheral T-cells obtained from individuals with onchocerciasis responded poorly to M.
tuberculosis antigens [158].
On the other hand, in contrast to the hypothesis that chronic helminth infections may increase
susceptibility to and worsen the clinical course of TB infection, the recent results of a mouse
model study could not detect any immunological effects of helminths during TB infection [159].
In this study the histological examinations and quantitative TB cultures demonstrate similar or
reduced mycobacterial burden in helminth co-infected animals compared to those infected with

28
TB only where PPD-specific cellular proliferation and IFN-γ production were not suppressed in
co-infected animals [159]. Additionally, in a cotton rat model with a filarial nematode (S.
litmosoides), it was reported that proliferation and IFN-γ production of spleen cells in response
to PPD were similar between co-infected (M. tuberculosis-S. litmosoides) than those with M.
tuberculosis infection only [159]. A possible explanation for theses divergent findings may lie in
the fact that the impact of helminthiasis on the host response to M. tuberculosis is dependent on
the type of helminth/intensity of infection, timing and the level of exposure to M. tuberculosis
itself.
Concluding remarks on the interaction between helminth infection and tuberculosis
Taken together, the co-existence of helminth infection in areas of high TB incidence may affect
the ability of the host to respond to M. tuberculosis and BCG vaccination. This may be of
importance for the risk of developing TB on exposure as well as for reactivation of latent disease
or the outcome of active TB. Helminth infections have a significant immunomodulatory
influence on the immune system and there is experimental evidence showing that helminth
clearly attenuate the Th1 immune response required for the control of M. tuberculosis infection.
However, whether helminth infection indeed has an impact on the clinical outcome or
immunological responses in human TB remains an open question.

29
AIMS
To assess the initial tuberculin skin test results and kinetics of an interferon-γ release
assay (Quantiferon) during active TB in a high endemic area.
To prospectively investigate whether early changes in a clinical scoring system (TB-
score) can predict treatment outcome in patients with pulmonary TB.
To assess the magnitude and clinical characteristics of helminth infection in TB patients
in comparison with household contacts and community controls and to evaluate its
correlation with eosinophil and IgE responses.
To investigate the immunological response and clinical outcome of helminth co-infection
in patients with active TB before and after albendazole treatment.
30
PATIENTS, MATERIALS AND METHODS
Study setting
The patients in papers I-V were recruited from three health institutions (Gondar University
Teaching Hospital, Gondar Health Center and Debark Hospital) in Ethiopia located in Gondar
town and the surroundings. Gondar is located 750 km from Addis Ababa in North-West
Ethiopia.
Figure 5: With a population of approximately 93 million [160], Ethiopia has the second largest
population in Africa.

31
Figure 6: The majority of the inhabitants practiced Ethiopian Orthodox Christianity
(84.2%)[161]
Figure 7: The Gondar city is nicknamed "The Camelot of Africa" due to the presence of a group
of royal castles (Photograph: Wikipedia, the free encyclopedia).

32
The Gondar University Hospital is a tertiary level teaching and referral hospital with 450 beds
rendering referral health services for over 5 million inhabitants in North-West Ethiopia. The
hospital provides inpatient and outpatient services, including care and treatment for TB and
HIV/AIDS patients. Antiretroviral treatment (ART) for HIV has been available in Gondar
hospital since March 2005. The ART regimen consisted of a combination of nucleoside and non-
nucleoside reverse transcriptase inhibitors (NRTI and NNRTI), according to the national HIV
guidelines [162]. At the end of 2012, there were close to 9470 HIV- positive adults on follow up
in the Gondar ART clinic and about 6000 of these patients were starting ART while the
remaining patients were on pre-ART. In 2010, a TB isolation and treatment ward, with a 28 bed
capacity, was built to improve the hospital TB infection control with a certain focus on MDR-
TB. The need is much greater than the present beds available, but capacity building activities are
underway, including a new hospital construction with 800 beds.
The Gondar Health Center is the largest of the three health centers currently available in Gondar
town. The health center provides primary health care to the population of Gondar town. It has a
TB treatment center (DOTS clinic) where TB patients receive medication after TB diagnosis.
Debark is a small town in northern Ethiopia located 90 kilometers north of Gondar town in the
Semien Gondar Zone of the Amhara Region. Debark is the closest town to the Semien
Mountains National Park. It is the largest settlement in Debark area.
All laboratory work was done in Gondar University Hospital laboratory except for TB culture
which was done in the BSL3 facilities at the Armauer Hansen Research Institute (AHRI), Addis
Ababa, Ethiopia. Optimization for ELISPOT and flow cytometry was done at AHRI and
Linköping University, Sweden while all patient analyses were done in Gondar.

33
Figure 8: Outlook of the University of Gondar Hospital (left) and Debark hospital (right).
Photograph: Ebba Abate
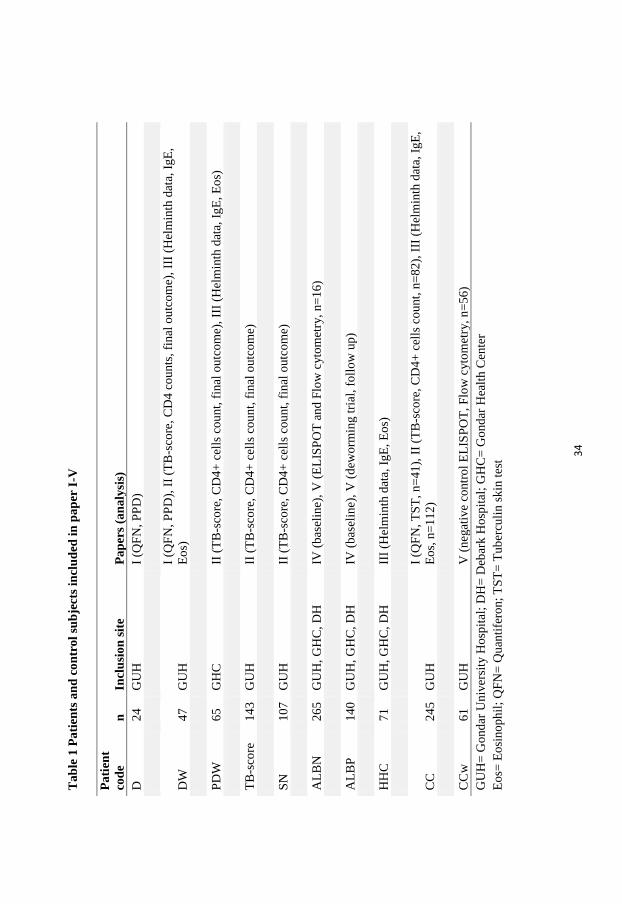
Study participants (Paper I-V)
Inclusion of TB patients and recruitment of blood donors and household contacts to TB patients
took place at the TB treatment clinics (DOTS center) within the Gondar University Hospital,
Ethiopia; Gondar Health Center (DOTS-polyclinic) and Debark Hospital. Only outpatients
fulfilling the inclusion criteria of newly diagnosed smear positive-TB (Paper I, and III) or smear-
positive and smear-negative pulmonary TB patients (Paper II, IV and V), aged between 15 to 60
years, and who provided oral and written informed consent were included. The number and
distribution of participants included in the 5 papers are described in Table 1.
34
T
ab
le 1
Pati
ents
an
d c
on
trol
sub
ject
s in
clu
ded
in
pap
er I
-V
Pa
tien
t
cod
e n
In
clu
sion
sit
e P
ap
ers
(an
aly
sis)
D
24
G
UH
I
(QF
N,
PP
D)
DW
47
G
UH
I (Q
FN
, P
PD
), I
I (T
B-s
core
, C
D4 c
ounts
, fi
nal
ou
tcom
e),
III
(Hel
min
th d
ata,
IgE
,
Eos)
PD
W
65
G
HC
II
(T
B-s
core
, C
D4
+ c
ells
count,
fin
al o
utc
om
e),
III
(Hel
min
th d
ata,
IgE
, E
os)
TB
-sco
re
14
3
GU
H
II (
TB
-sco
re, C
D4
+ c
ells
count,
fin
al o
utc
om
e)
SN
10
7
GU
H
II (
TB
-sco
re, C
D4
+ c
ells
count,
fin
al o
utc
om
e)
AL
BN
26
5
GU
H, G
HC
, D
H
IV (
bas
elin
e), V
(E
LIS
PO
T a
nd F
low
cyto
met
ry,
n=
16)
AL
BP
14
0
GU
H, G
HC
, D
H
IV (
bas
elin
e), V
(d
eworm
ing t
rial
, fo
llo
w u
p)
HH
C
71
G
UH
, G
HC
, D
H
III
(Hel
min
th d
ata,
IgE
, E
os)
CC
24
5
GU
H
I (Q
FN
, T
ST
, n=
41),
II
(TB
-sco
re, C
D4
+ c
ells
co
un
t, n
=82),
III
(H
elm
inth
dat
a, I
gE
,
Eos,
n=
11
2)
CC
w
61
G
UH
V
(n
egat
ive
contr
ol
EL
ISP
OT
, F
low
cyto
met
ry, n
=56)
GU
H=
Gon
dar
Univ
ersi
ty H
osp
ital
; D
H=
Deb
ark H
osp
ital
; G
HC
= G
ondar
Hea
lth C
ente
r
Eos=
Eosi
noph
il;
QF
N=
Qu
anti
fero
n;
TS
T=
Tub
ercu
lin s
kin
tes
t

35
Smear-positive TB was defined as 2 of 3 morning sputum samples positive, or one of 3 positive,
with chest X-ray findings and clinical symptoms, suggestive of active pulmonary TB [13].
Smear- negative pulmonary TB was defined as symptoms suggestive of TB with 3 sputum smear
samples negative for AFB, radiographic abnormalities consistent with pulmonary TB, and a lack
of clinical response to one week of broad-spectrum antibiotic therapy (MOH, 2008). The
exclusion criteria were patients requiring hospital admission, pregnancy, clinical signs or
medical treatment indicating any concomitant chronic or infectious diseases other than TB/HIV.
As control group, healthy community controls were recruited from the blood bank of Gondar
University Hospital. All blood donors had passed pre-donation clinical screening to rule out any
chronic illnesses and previous TB history. Moreover, all controls had a normal chest x-ray
findings, a TB-score ≤ 3 and did not show any clinical signs or symptoms of clinical TB. Patients
were evaluated for laboratory and clinical markers as indicated in the respective papers and as
outlined in Table 1. In addition, household contacts were included (Paper III) based on the
information obtained from the TB patient. A household contact was defined as a person who
lives together (> 6 months) and spends more than 12 hours per day with a TB patient.
Outline of the interventional clinical study on the effect of albendazole in patients co-
infected with helminths and pulmonary TB (Paper V)
The study design of Paper V was a randomized, double-blind follow- up study in which newly
diagnosed pulmonary TB patients with concurrent helminth infection presenting consecutively
from 1st of March 2009 to 15
th of October 2012 at the DOTS clinics at the Teaching and Referral
Hospital of the University of Gondar, the Gondar Health Centre and at Debark Hospital were
randomized to albendazole or placebo groups after oral and written consent. Each patient
received 400 mg X III per os of albendazole or placebo at two weeks after initiation of TB
treatment. The primary outcome was TB-score change at week 8 compared to baseline. The
secondary outcomes were sputum smear conversion after two months, changes in chest x-ray
pattern from baseline to week 12, CD4 cells count, IgE and eosinophil response after 3 months as
well as immunological responses such as changes in Tregs and levels of IFN-γ, IL-5 and IL-10
producing PBMCs after 3 months. Random numbers was generated by a computer and was done
in a block size of eight by the Department of Epidemiology, Addis Continental Institute of Public

36
Health, Ethiopia. The treatment allocation was concealed in individual envelopes opened only
when a patient was enrolled. Albendazole and placebo were identical and numbered by Addis
pharmaceutical company, Ethiopia. Both the investigator and clinical staffs were blinded to the
randomization code and substance. The code was broken after the follow-up visit of the last
patient and when the statistical analysis had been performed.
Clinical and laboratory patient characteristics
Baseline characteristics and determination of the TB-score
The TB-score consists of signs and symptoms which were recorded as previously described
[163]. Each variable of the TB score contributes to one point resulting in a TB-score from 0 to 13
(Table 2). Furthermore, age, sex, the presence of a BCG scars and body mass index (BMI) were
registered for all study participants.
Table 2: Parameters in the TB-score
Cough 1 0
Haemoptysis 1 0
Dyspnoea 1 0
Chest pain 1 0
Night sweating 1 0
Anaemic conjunctiva: 1 0
Tachycardia (>100/min): 1 0
Lung auscultation (pos): 1 0
Temperature >37C 1 0
BMI<18: 1 0
BMI<16: 1 0
MUAC <220: 1 0
MUAC <200: 1 0
TB score sum
BMI: Body Mass Index; MUAC: Middle Upper Arm Circumference

37
HIV testing and determination of CD4+T-cells count
Testing for HIV was done with different diagnostic test methods applied and used in Ethiopia at
different periods based on the national HIV/AIDS guidelines, i.e, (i). Enzygnost Anti-HIV 1/2
Plus (Dade Behring, Germany) and confirmed with Vironistika HIV Uni-Form II Ag/Ab (Biom é
rieux, France) using ELISA, (ii). Rapid HIV test kits which include Determine, Capillus and
Unigold, (iii). Rapid HIV test kits which include KHB, Stat pack and Unigold. CD4+T-cell
counts were analyzed by using a FACSCount machine (BD, San Jose, California, USA) at the
Gondar University Hospital Laboratory which is involved in continuous external and internal
quality control programmes.
Determination of IgE, eosinophil cell count and Quantiferon by ELISA
Serum IgE was determined with a commercial ELISA kit (Immundiagnostik, Germany)
according to the manufacturer’s instruction. The absolute eosinophil count of peripheral blood
was computed in cells/mm3
from the value of total and differential white blood cell counts
obtained using Cell Dyn 1800 (Abbot, USA). The quantiferon (QFN) test was done according to
the manufacturer’s instructions (Cellestis, Australia). All QFN samples were collected in the
morning at 09.00 am.–12.00 am. One milliliter of blood was collected into each of the three QFN
blood collection tubes. At the time of sample collection and prior to incubation, the samples were
mixed thoroughly by shaking the tube 10 times (5 s) to ensure that the entire inner surface of the
antigen coated tube was covered with the blood. The samples were transported and directly
incubated following the shaking procedure at 37 ° C for 18- 22 h after which the supernatant
was frozen at -20 ° C for later analysis.
Stool examination
Stool samples from three consecutive days were collected from each participant and examined
using direct stool microscopy and Kato-Katz techniques [164] by the same technician throughout
the study. The classification into helminth positive or negative was based on the examination of
all three samples together from each patient. If at least one of the samples was positive for
helminth ova or larva the patient was regarded as helminth positive. One in 10 slides were
randomly selected and checked again blindly by a second microscopist for quality control. The
same stool sample collection and examination strategy was used at week 12 in paper V.

38
Sputum smear examination
Acid- fast bacilli (AFB) staining and examination was done at baseline and week 8 on morning
sputum samples from three consecutive days. The AFB load in sputum smears was measured as
previously described by the World Health Organization as: negative (0 AFB/100 oil fields),
scanty (1-9 AFB/100 fields), 1+ (10-99 AFB/100 fields), 2++ (1- 10 AFB/ field) and 3 +++ (> 10
AFB/ field) [165]. Sputum conversion was defined as all three consecutive sputum smears
turning negative for AFB at week 8.
Isolation of peripheral blood mononuclear cells (PBMCs) from whole blood
Heparinized venous blood (25 ml) was collected in DOTS clinics and transported within one
hour to the laboratory where PBMCs were directly isolated. Heparinized blood was layered on a
Lymphoprep density gradient solution (Axis-Shield POC AS, Norway) with a 1:2 proportion of
blood, and then centrifuged at 800g for 30 minutes at 20 ºC. The resulting interphase ring
consisting of a mixture of mononuclear cells was collected and then washed twice with PBS
(Sigma-Aldrich, Munich, Germany) followed by centrifugation at 250g for 10 minutes. Finally,
cells were re-suspended in RPMI-1640 (Sigma-Aldrich) supplemented with 10 % sterile heat-
inactivated FBS (Sigma-Aldrich) and 1% penicillin-streptomycin (Sigma-Aldrich), before
counting in a Bürker chamber. Cells were stored in freezing medium containing of 10% DMSO
and fetal calf serum (FCS) in -80oC freezers until use. Cells were frozen for a maximum of 2
months period. Trypan -blue exclusion dye (Sigma-Aldrich) was used for detection of cell
viability. Only cells from patients with a viability after thawing of above 75% were included.
Analysis of regulatory T-cells by flow cytometry
The isolated PBMCs were stained with fluorochrome-labeled monoclonal mouse anti-human
antibodies CD4-fluorescein-isothiocyanate (FITC) (BD Biosciences, clone RPA-T4), CD25-
Percp Cy5.5 (BD Biosciences, clone M-A251 ),and CD127-Alexa 647 (BD Biosciences, clone
HIL-7R-M21), followed by fixation/permeabilization using cytofix/cytoperm solution (BD
Biosciences) and intracellular staining with monoclonal antibodies to Foxp3- phycoeryhtrin (PE)
(BD Biosciences, clone 236A/E7). Fluorescence minus one (FMO) controls was used for gating
purposes. Cells were included in the analysis if the cell viability was 75% after thawing. Tregs
were defined as the population of cells that were CD4+/CD25hi
/CD127low
/Foxp3+. Flow

39
cytometry data were collected on a FACSCalibur flow cytometer (BD Biosciences) using
CellQuest acquisition software and were then analyzed using Flowjo 7.6.5 (Tree Star, USA).
Analysis of IFN-γ, IL-5 and IL-10 by ELISPOT
Peripheral blood mononuclear cells (250,000/well) were plated onto ester-cellulose-bottomed
plates (PVDF plates, Mabtech, Solna, Sweden) coated with a capture mAb specific for human
IFN-γ (1-D1K), IL-5 (TRFK5) and IL-10 (9D7) at 15µg/ml in PBS. Cells were incubated in a
humidified incubator at 37°C in 5% CO2 for 24 h with PPD-antigen in duplicate using
unstimulated and CD3-stimulated cells as negative and positive controls, respectively. Cells were
then removed by washing five time with PBS-0.5% FCS, and then biotinylated anti-mouse IFN-
γ (7-B6-1), IL-5 (5A10) and IL-10 (12G8) were added at 1 µg/ml in PBS and incubated for 2 h at
room temperature. After washing five times in PBS, streptavidin-alkaline phosphatase (Mabtech)
was added for 1 h at room temperature. After washing with PBS, filtered (0.45µm filter)
BCIP/NPT- plus substrate (Mabtech) was added. Plates were extensively washed using tap
water. The number of spots for each well was counted with automated ELISPOT reader (AID,
Germany), and the results for each stimuli (PPD, negative and positive control) correspond to the
mean of duplicates.
Statistics (Paper I-V)
In Paper I, data are presented as mean + standard deviation (SD). To compare groups, the
Student’s t -test was used for parametric data and the chi-square test for discrete variables. Kappa
was calculated according to Cohen. A multiple regression model was used entering significant
variables using the STATISTICA software package (StatSoft, Tulsa, USA).
In Paper II, it was estimated that to detect a minimal clinically important difference of 1.1 TB-
score points of the TB patients at week 8 compared to the blood donors, 80 blood donors were
needed at a power of 80% (alpha 0.05). We assumed that a sample size of 250 TB patients
(including a 10% loss to follow-up) was needed in order to detect a mortality difference from the
expected 8% to 18% in the patients who did not show a decrease in the TB-score of > 25%
between baseline and week 2, on the assumption of a 10% overall mortality and an estimated
fraction of 20% not reaching a < 25% decrease between baseline and week 2. Data are presented
as the median and inter-quartile range and analyzed using Fisher’s exact test for categorical
variables and Wilcoxon and Mann-Whitney tests for continuous variables. In a multiple logistic

40
regression model setting mortality as the dependent variable (STATISTICA software package;
StatSoft, Tulsa, USA), age, sex, HIV, presence of ART treatment, and CD4+T-cell counts (< 200
cells/mm3) were included in the final analysis.
In Paper III, data are presented as median and inter-quartile range. Significance testing was done
with Mann-Whitney and Wilcoxon tests for continuous data and Fisher’s exact test for discrete
variables. Variables with a p<0.1 in the univariate analysis were entered into a multiple
regression analysis with helminth status as the dependent variable. The regression analysis was
performed using the STATISTICA software (Tusla, USA).
In Paper IV, continuous data are expressed as means with standard deviation (SD). Comparison
between groups was done by Students t-test for continuous data and by chi-square test or
Fisher’s exact test for categorical data. Variables with a p-level <0.1 were entered a multiple
logistic regression model using the STATISTICA software package (StatSoft, Tulsa, USA).
Analysis for the randomised trial (paper V) was by intention to treat. The study code was kept in
a sealed envelope until all data were analyzed. Effects of deworming compared to placebo on
primary and secondary outcomes were evaluated by chi-square test or Fisher´s exact test for
discrete variables and Students t-test for continuous variables. Multiple regression analyses were
performed adjusting for baseline. A p<0.05 was regarded as statistically significant.
The sample size for the randomized trial in paper V was calculated under the following
assumptions. In our previous study [166], newly diagnosed TB patients showed a 75% decline
from baseline TB-score value after two months of anti-TB treatment. Accordingly, the sample
size was calculated under the assumption that 210 helminth positive TB patients would have to
be included giving 80 % power at the 5 % significance level to detect a difference in treatment
response of 15 % including a 5 % drop out rate.
Ethical considerations
Oral and written informed consent was obtained from all study participants. The study has
received ethical clearance from the Ethics Review Board of the University of Gondar
(RPO/55/37/2001; Res/05/530/01/12); and from the Medical Ethics Board at Linköping
University, Sweden (M143-06, T2-08). In addition, approval has been obtained from the federal
Drug Administrative and Control Authority (DACA) (02/6/22/36), Ethiopia for paper V where
an independent data and safety monitoring board (DSMB) regularly reviewed the results and

41
adverse events and adverse reactions. All patients included were treated according to the DOTS
program following the WHO adopted national TB guideline of Ethiopia. Pre-and post-test
counseling was given to all study participants during HIV testing as part of the hospital/health
center routine, provider initiated HIV counseling and testing (PIHCT) programme. Helminth-
positive patients received anti-helminthic treatment free of charge after the completion of the
study at week 12. There was no additional cost to or economic incentive for the patients
participating in the study. Study participants were compensated for their transportation expenses
during follow-up visits.

42
RESULTS AND DISCUSSIONS
The influence of TB treatment and HIV on the tuberculin skin test (TST) and IFN-γ
response to M. tuberculosis specific antigens using T-cell based assays: IGRA (Quantiferon)
and TST
Interferon-γ release assays (IGRA) are based on the cell-mediated immune response and
subsequent measurement of IFN-γ production against a mixture of synthetic peptides simulating
proteins present in M. tuberculosis: the early secretory antigenic target-6 (ESAT-6) and culture
filtrate protein-10 (CFP-10) and in the case of Quantiferon (QFN) also to the antigen TB7.7. The
IGRA tests are more specific than TST in particular in BCG vaccinated individuals as the target
peptides in the IGRA tests are secreted by M. tuberculosis and M. bovis, but are neither present
in the BCG vaccine, nor in most non-tuberculous mycobacteria. Previous studies considering the
impact of anti-TB treatment on the specific IFN-γ response as measured by the QFN assay have
shown controversial results in the case of both latent and active TB treatment. Indeed, some
authors have described that the IFN-γ response to specific mycobacterial antigens decreased or
became negative after TB treatment [70,167-170], while others have reported persistently
positive or even stronger responses during and after anti-TB treatment [66,171-175]. Qualitative
and quantitative changes in the IGRA response may be a candidate biomarker to monitor the
response to TB drug therapy and for the evaluation of interventions. However, little is known
about the kinetics of IGRAs during anti-TB treatment in relation to disease severity in high
endemic areas for TB and HIV. Additionally, only a few studies have used a head-to-head
comparison between TST and an IGRA test.
We conducted a small prospective, head-to-head study (Paper-I) to assess the effect of TB
treatment on T-cell responses as measured by the QFN assay in newly diagnosed active TB
patients in Ethiopia with the aim to use this assay as a prognostic tool for treatment response in
subsequent studies (Paper IV and V). Blood donors were recruited as a healthy community
control group.

43
The proportion of patients positive for QFN positive patients declines during the course of
anti-TB treatment
In paper I, we showed a significant decline in the proportion of QFN positive patients which was
more pronounced in HIV- positive patients following TB treatment (day 0 to month 7). This
finding is in agreement with previous reports [168,176,177]. This has further been supported by
a study assessed the decline of IFN-γ (ESAT-6 and CFP-10) using an in-house ELISPOT
protocol in the Gambias [178]. HIV-positive TB patients showed a higher rate of QFN reversion
(from negative to positive) than HIV-negative TB patients (p<0.05 in that study). The observed
QFN conversion was attributed to recovery of the immune response during anti-retroviral
therapy (ART) and may be dependent on initial CD4+ cells count levels and timing of ART
treatment. The decline in the proportion of QFN positive patients observed in most studies so far,
in particular in HIV-negative TB patients is most likely due to the effect of anti-TB treatment,
which decreases the antigen load and subsequently attenuates the cell mediated immune
activation and IFN-γ production. It was also previously described that the magnitude of the M.
tuberculosis antigen specific IFN-γ T-cell response is proportional to the antigen load of the
infecting organism in human and animal models [179,180].
A high clinical severity score correlated with negative TST results
The correlation of TST and the QFN assay with disease severity, treatment outcome and smear
conversion rate (at 2nd
month) was assessed (paper I). Multiple regression analysis showed a
correlation between an initial negative TST result at baseline and a higher disease severity score
(advanced TB), which was not confounded by sex or HIV. The lack of such correlation in QFN-
negative individuals indicates that the severity of the disease may not affect the performance of
the QFN assay to the same extent. Moreover, this observation underlines the clinical differences
between the two tests during active TB and support the observation that TST reaction may be
suppressed relative to the QFN response in more advanced TB. No correlation was observed
between QFN results and final TB treatment outcome or sputum smear conversion rate at the end
of the intensive phase of TB treatment.

44
The impact of HIV and CD4+ T-cells counts on the performance of the Quantiferon assay
The QFN assay showed a significantly higher rate of positive results in HIV-negative TB
patients compared to HIV-positive TB patients (93.8 % vs. 70.3 %, n=69, p<0.05), whereas no
significant difference was noted when the rate of positive TST results (≥ 10mm) were analyzed
(87.9% vs. 73%). Previous studies showed that HIV co-infection could influence the QFN test
since IFN-γ production might be limited by the reduced number and function of CD4+ T-cells
[65]. A low CD4+ T-cells count (<200 cells/mm3) was significantly associated with TST
negative results (p= 0.03), whereas this association was not present for QFN-negative results. It
has been previously described that HIV-infected patients are more frequently anergic to skin
testing, particularly those with a CD4+ lymphocyte count lower than 100-200 cells/mm3 [181].
The correlation of negative TST result and low CD4+ T-cells count confirms the impact of
immune-suppression (characterized by low CD4+T-cells count) of TB patients on the
performance of TST. This finding is supported by the fact that we observed a significant
correlation of negative TST results with advanced forms of TB in the same cohort. Low CD4+
T-cells counts in some studies have been found to be associated with an increase in both
indeterminate and false-negative QFN results [182,183], but low CD4+ T-cells counts do not
seem to account for all indeterminate and negative results in HIV-positive TB patients [184]. In
agreement with this observation, we could not detect a significant correlation between negative
QFN results and low CD4+ T-cell counts, including patients with CD4+ T-cells counts lower
than 200 cells/mm3 in the HIV-positive/TB group. We found that even patients with very low
CD4+ T-cell counts (30 cells/mm3) may present an IFN-γ response high enough to be detected in
the QFN test, suggesting that the quality and function of the CD4+ T-cells is of great importance.
Possibly, also other cells including CD8+ T-cells could contribute to the increased IFN-γ
production in patients with low levels of CD4+ T-cells as QFN is analyzed in whole blood.
Poor concordance between QFN and TST results
The agreement of the TST and QFN tests was poor in TB patients (Kappa= 0.09) vs. healthy
community controls (kappa=0.59), as described previously [182]. One reason for this difference
may be that the cell mediated immune response to mycobacterial antigens differ in latent versus
active TB. The highest frequency of QFN and TST positive test results was observed in TB
patients with >200 CD4+ T-cells/mm3 compared to TB patients with < 200 CD4+ T-cells/mm
3.

45
This illustrates the influence of the number of CD4+T-cells in the performance of the TST. Thus,
the two tests showed fair agreement in the control group (kappa=0.59) and in this group the
controls in general had higher CD4+ T-cells counts than the HIV-negative TB patients (p=0.01)
in this cohort. Although the BCG vaccine coverage of community controls and TB patients in
our study was low, there was no correlation between discordant results and the presence of a
BCG scar. A limitation of this study was that mycobacterial culture was not done for
confirmation of sputum conversion at the end of the 2nd
month. As in most other follow up
studies, the small sample size was one other limitation of our study as well as a not negligible
loss to follow-up.
We conclude that the average rate of positive QFN tests is significantly reduced from baseline to
the time of TB treatment completion resulting in levels comparable to the background level of
healthy blood donors (51%). The agreement between the TST and QFN tests is poor in TB
patients, including a poor correlation to disease severity. Qualitative conversion of IGRA tests
from positive to negative has a very limited clinical value as a surrogate marker of treatment
efficacy as it does not occur reliably in patients who completed the TB treatment. Further
investigations on T-cell kinetics and functions are needed to understand the significance of
persistent QFN-positive results, which do not correlate to poor treatment responses. The
observation that healthy controls had a relatively high QFN level comparable with the level
observed for TB patients after treatment indicates that this assay likely unsuitable for clinical
monitoring or diagnosis of TB in high endemic areas.
TB clinical scoring (TB-score): relevance to treatment monitoring
The current WHO guidelines recommend sputum smear results for monitoring smear positive
pulmonary TB patients [185,186]. For smear-negative TB patients it is recommended to use
weight and clinical symptoms, which may be crude and in case of the clinical symptoms, a
subjective evaluation tool for clinical studies [187,188]. The proportion of HIV co-infected TB
patients is highest in Africa [1] resulting in a large proportion of smear-negative pulmonary TB
patients, who cannot be monitored using smear conversion and whose treatment effect therefore
is evaluated only by weight gain [58].

46
Clinical scoring systems have been developed and are widely used for many diseases such as the
NYHA score for chronic heart failure [189,190]. Various clinical prediction tools have been
proposed for TB [185,186,191,192], all depending on methods not readily available in high
endemic areas and some only developed for smear positive pulmonary TB patients [192]. Studies
from Guinea-Bissau in west-Africa have proposed a scoring system (TB-score) that can be
applied to both smear positive and negative pulmonary TB patients. High TB-scores correlate
with mortality and this scoring system is suitable for field use in highly endemic areas [163].
However, the score still needs to be evaluated with data from various geographical settings and
TB manifestations as well as for the impact of HIV and CD4+ T-cells counts on its performance.
The purpose of our study (Paper II) was to prospectively investigate the kinetics of the TB-score
during the course of anti-TB treatment and whether early changes in the TB-score results could
predict treatment outcome in Ethiopian TB patients. Another aim was to evaluate the TB-score
for use as a primary clinical outcome marker for subsequent studies described in Paper V. In this
part, 250 active pulmonary TB patients and 82 blood donors (controls) were included.
The TB-score declines during the course of TB treatment and is not influenced by HIV
Pulmonary TB patients were followed at three different time points: at initiation of TB treatment
(week 0), on the 2nd
week and during the completion of the intensive phase of TB treatment (8th
week). A highly significant and rapid decline in the median TB-score value was observed
already after the 2nd
week of TB treatment compared with the base line value (8 points vs. 4
points; p< 0.0001); and the rate further dropped to a lower level at the end of the intensive phase
of anti-TB treatment. The rapid response already at week two coincides with the observational
time point where most patients have clinically responded to anti-TB treatment. The low median
TB-score value observed in TB patients after 2 months of TB treatment was still significantly
higher than the median TB-score value of controls (2 points vs. 0 points; p< 0.0001). The HIV
co-infection rate among TB patients was 53.6% in this study. The TB-score was not influenced
by HIV infection or CD4+ T-cell counts as a similar declining pattern was observed when HIV
negative and HIV positive TB patients were followed from initiation until completion of the 2
month intensive phase of TB treatment. Overall, the progressive decline of the TB-score
observed during the course of TB treatment and the fact that this was not influenced by HIV
infection suggests that TB-score may be an appropriate patient clinical monitoring tool during

47
the course of TB treatment in our Ethiopian study setting as well. So far it has been used in
clinical interventional studies evaluating deworming (paper V) and vitamin D supplementation in
Guinea-Bissau [193].
TB-score correlated with mortality
The TB-score levels correlated with mortality at week 2, moreover, mortality was associated
with a failure to achieve more than a 25% decline in TB-score at week 2. This suggests that
repeated clinical scoring during the initial weeks of TB treatment could be of value to identify
high-risk patients, who might need hospital admission to manage adverse events. A significantly
higher TB-score was observed between TB patients who died vs. patients who successfully
completed the TB treatment at week 0, (8 points vs. 9 points, p< 0.04), week 2, (4 points vs. 7
points, p<0.0001) and week 8, (2 points vs. 5 points, p<0.0038). The highest mortality rate was
observed in HIV positive patients (13/134) and in particular in patients with a low baseline
CD4+ T-cells count. Indeed, HIV infection appears to be the dominant risk factor for mortality
among Ethiopian TB patients, and clinical scoring and close monitoring should be prioritized for
this group.
The TB-score can serve as a simple tool for standardized evaluation of early clinical
improvement in TB patients in high endemic areas. Early monitoring of patients by assessing the
TB-score in the first 2 months of treatment could facilitate the identification of patients at high
risk of a poor clinical outcome. This relatively simple clinical score could provide an objective
tool for healthcare workers to identify high-risk patients. The clinical evaluation can quickly be
carried out without using expensive diagnostic tools and could be suitable for follow-up in
clinical trials [163]. Nonetheless, although the TB-score has attained a similar level of
performance in settings that share epidemiological characteristics [163], further studies are
needed to assess its external validity, its acceptance and implementation by clinicians and its
eventual impact to prospectively predict high- risk TB cases and making decision to hospitalize
pulmonary TB patients. Moreover, the scoring system needs to be developed to improve its
capacity to predict clinical outcomes, especially with regard to HIV co-infection. Furthermore,
the definition of cut-off levels for practical use to pin-point high-risk patients needs to be
explored in future studies, as well as how to implement the scoring algorithm at different levels

48
of the healthcare system. The reproducibility study conducted recently [194] revealed the need
for refinement of the TB-score. It is important to select signs and symptoms showing lack of
reliable inter- and intra-observer variability and the need to refine the TB-score further by
omitting variables with relatively poor reproducibility.
As a limitation of the study, 24% of the TB patients in this cohort did not have survival data at
the end of TB treatment. There might be undetected deaths in the defaulting group (10%). Due to
lack of sputum culture facility, smear-negative TB patients were not confirmed by culture but the
national WHO-based guidelines to define smear negative cases were used instead. The impact of
ART in decreasing mortality in HIV-coinfected TB patients could not be assessed since the study
was not primarily designed to address this issue.
In summary, we showed that early monitoring of patients with TB-score in the first weeks of
treatment could facilitate the identification of patients at high risk of poor clinical outcome. The
TB-score could be a useful clinical monitoring tool during TB treatment. The observation that
the TB-score declines during the course of successful TB treatment and the correlation between
high TB-scores and a poor clinical outcome during TB led us to use this scoring method as a
clinical monitoring tool during the course of TB treatment in Paper V.
TB-Helminth co-infection
In sub-Saharan Africa, there is extensive epidemiological overlap between helminthic infections,
TB, HIV/AIDS and malaria [99,195,196]. There are several reports showing high rate of
helminth co-infection with HIV [127,130,197-199] . Nevertheless, epidemiological reports on
the burden of helminth infection in active pulmonary TB patients are scarce. We therefore aimed
to assess the impact of asymptomatic helminth infection among patients with active pulmonary
TB in Ethiopia which is high endemic for helminth infection and TB. Newly diagnosed TB
patients (n=112), house-hold contacts (n=71) and healthy community controls (n=112) were
included. The burden of helminth infection in the three groups was assessed and correlated with
surrogate markers of Th2-type immune response (IgE levels and eosinophilia). TB patients were
followed and examined for their helminth status at baseline, and in the 2nd
, 8th
and 12th
week
after treatment initiation.

49
High burden of asymptomatic helminth infection among patients with active pulmonary
TB
Twenty nine percent (29%) of the TB patients in paper III had concurrent helminth infection,
which is in line with previous reports showing that the helminth co-infection in active TB
patients was nearly twice as high as their healthy household contacts although this study reported
generally higher rates [154]. A total of seven different helminthic species were identified.
Ascaris lumbricoides was the most common parasite detected followed by hook worm infection
for TB patients, house-hold contacts and healthy individuals. Although still controversial, some
studies have suggested that intestinal parasites could be a risk factor for developing TB upon
exposure due to their potent modulation of the host immune response. Tristao-SR et al. [153]
and Elias, et al. [154] reported a higher rate of intestinal nematodes in patients with pulmonary
TB than control groups, and Diniz et al.[155] observed a strong association between intestinal
nematode infection and multibacillary leprosy. Our findings of a high helminth burden among
TB patients further confirm the impact of chronic helminth infection in TB, but additional follow
up studies with much larger sample sizes are needed to confirm the causal relation between
helminth infection and the risk of developing of active TB.
Helminth infection correlated with eosinophilia and elevated IgE level
Large extracellular pathogens like helminths induce immune mechanisms that are of a Th2-type,
characterized by an elevation of peripheral blood eosinophilia and elevated IgE accompanied by
a profound increase in cytokine responses including IL-4, IL-5, IL-9, IL-13 [200,201]. Helminth
products can drive Th2-type responses even in the presence of Th1 inducers. For example, when
stimulated with Schistosoma mansoni soluble egg antigen (SEA), DCs are able to induce Th2
type responses in the presence of bacterial Th1 stimuli [202].
We assessed the peripheral eosinophil cell count and IgE level among TB patients, household
contacts and healthy community controls in relation with helminth status. In healthy individuals,
eosinophils make up only 2-5% of peripheral white blood cells. However, during active parasitic
helminth infection the proportion of eosinophils in the blood can reach 40% [203]. Together with
high IgE levels and mastocytosis, eosinophilia is considered to be one of the cardinal features of
parasitic helminth infection. It was previously shown that Ethiopians have low levels of
hematological and immunological markers, such as peripheral while blood cells and CD4+ T-

50
cells and an increased immune reactivity compared with Dutch citizens and people from other
African countries [204]. Such variations could be due to differences in genetics, dietary patterns,
sex, age, and altitude. Taking this into consideration, two cut off values were used in our studies
for eosinophilia in this population (>500 cells/mm3 and >300 cells/mm
3). The conventional cut
of value for eosinophilia is 500 cells/mm3 but this may not be appropriate for the Ethiopian
population as previously described [205] . In a multivariate analysis, eosinophilia (adjusted OR:
15.2; 95% CI: 1.4–160.3, p = 0.02) and increased IgE-levels (adjusted OR: 7.6; 95% CI: 1.2–
48.4. p = 0.03) were independently associated with asymptomatic helminth infection, which was
not confounded by sex or HIV status. Significantly lower median eosinophil count and IgE level
was observed among TB patients with concurrent helminth infection compared to helminth
infected household contacts (eosinophil counts: 234 cells/mm3 vs. 600 cells/mm
3, p= 0.005; IgE:
351 IU/L vs. 378 IU/L, p=<0.001), and community controls (Eosinophilia: 234 cells/mm3 vs.
602 cells/mm3, p= <0.001; IgE: 351 IU/L vs. 420 IU/L, <0.001). The elevated IgE level and
eosinophilia observed in our study as a result of helminth infection was in agreement with recent
studies done in Brazil and China [205,206]. Furthermore, helminths have also been described as
strong inducers of IgE in infants [207,208]. It was indicated that specific IgE production
significantly reduces TST reactivity in younger children. The effect of eosinophil degranulation
on the killing of the helminthic parasite, Strongyloides was shown in a previous study [209,210].
Furthermore, eosinophil granule proteins can modify immune responses in L. sigmodontis
infection [211]. Overall, our findings show that peripheral blood eosinophils are highly increased
under the influence of helminths, and that helminths are associated with an isotype switch that
leads to high serum IgE concentration [212].
Helminth infection markedly declined in HIV-positive TB patients during TB treatment
Interestingly, we observed a decline in the rate of helminth infection among HIV-positive TB
patients during anti-TB treatment. All patients were asymptomatic and did not receive any anti-
helminthic treatment during the 12 week follow-up period. There has previously not been any
routine screening programme for helminth infection in the area. There was a marked decline in
the rate of helminth infection among HIV-positive TB patients compared to HIV-negative TB
patients. This pattern was stable among the 77 TB patients included in the analysis, where most
of the helminth positive patients, 86% (66/77) had the same worm status at week 0 compared to

51
the follow up samples at week 2, week 8 and month 3. This observation has to our knowledge
not been previously reported. Since the introduction of highly active antiretroviral therapy
(HAART), mortality and morbidity from a wide variety of opportunistic viral, bacterial, fungal
and parasitic infections have decreased dramatically among HIV-infected individuals in
economically developed countries [213]. It is after such observation that Lucas described the
scenario in his paper entitled “Missing infections in AIDS” [214].
Even though, the target of the protein inhibitors (PIs) is the HIV-1 protease, some parasites could
be an unspecific target for HIV-1 PIs. Protease inhibitors was shown to exhibit an inhibitory
effect against the malaria parasite [215]. Additionally, the results of in-vitro and in-vivo
investigations [213,214,216-218], indicate that HIV-1 PIs have a direct effect on opportunistic
parasites such as Cryptosporidium parvum, in particular aspartic protease inhibitors.
Nonetheless, HIV patients in the current TB cohort did not receive PIs for HIV treatment, a
category which was uncommon in the ART regimen used in Ethiopia during the time of the
study. Thus, the effect of PIs documented previously could not be the reason for the decline of
helminth infection observed in the HIV-coinfected TB patients. Interestingly, it has been
reported recently that drugs used in HIV patients (NNRTI/NRTI and trimethoprim-
sulfamethoxazole) showed an inhibitory effect against the liver stage of Plasmodium parasites
[219]. There was also a report indicating an increased effect of first line drugs used for TB
treatment on a protozoan parasite [220]. The same authors showed the combined effect of
rifampicin, isoniazid, and ethambutol, in treating malaria in a murine model. The findings of the
progressive decline in the rate of helminth infection (presence of eggs/larvae) without anti-
helminthic treatment in asymptomatic helminth co-infected TB- patients merits further study.
The clinical implication of helminth infection on active TB was studied further at base line
(Paper IV) and follow up (Paper V) during TB treatment.
The impact of helminth infection in pulmonary TB
In Paper III, we showed a high burden of helminth infection among TB patients and strong
correlation of eosinophilia and elevated IgE level with helminths. In a larger study cohort of 265
helminth negative TB patients and 140 helminth positive patients with TB, we observed that the
magnitude of helminth infection in patients with active TB was significantly higher than the

52
prevalence in community controls (35% vs. 27%, p= 0.03). Interestingly, the rate of hook worm
infection was significantly higher in the helminth positive community controls compared to
patients with active TB where Ascaris was more prevalent (Table 3a and 3b). Taking the findings
of this and other studies into account, we hypothesized that the skewed immune response caused
by helminths may affect the pathogenesis of tuberculosis including the clinical presentation of
TB. Thus, in paper IV we studied the clinical impact of helminth infection in patients with active
TB at initiation of TB treatment. A total of 377 pulmonary TB patients were included in this
paper. About one third of these patients 81% (96/119) were confirmed as sputum culture positive
for M. tuberculosis and no atypical mycobacterial infection was detected.
Influence of helminth infection on the clinical characteristics of active TB infection
The HIV co-infection rate of TB patients in paper IV was 33% which was lower than the close to
50 % rate previously reported from same area [38,116,119]. A recent report by UN-AIDS
indicated that the rate of new HIV/AIDS cases is declining globally after the advent of HAART
alongside with consistent HIV/AIDS control and prevention measures [12]. This is supported by
the fact that 70% (77/110) of the HIV co-infected TB patients included in the current study had
already started ART. The TB patients in the current cohort had low body mass index ( BMI) (<
18.5 kg/m2
) and mid-upper arm circumference (MUAC) (< 22 cm) indicating malnutrition [221],
although no difference was observed between helminth- positive and negative TB patients
included in this cohort. In addition, no significant difference was observed in the mean TB-score
values between helminth- positive and helminth- negative TB patients. Comparing the individual
parameters of the TB-score, the body temperature as well as the rate of tachycardia was
suppressed in helminth-positive patients suggesting an attenuating effect of the chronic helminth
infection on the Th1 type pro-inflammatory response and increasing Tregs and Th2 activity.
However, following adjustment for HIV in a multivariate model the association with body
temperature and tachycardia was no longer present. Consequently, no major differences in the
clinical presentation of TB in helminth- positive versus helminth-negative patients were
observed at baseline. The immunological impact of helminth co-infection confirmed as an
increased rate of eosinophilia also in this study cohort may be too subtle to induce a detectable
clinical difference in this sample size due to the strong pro-inflammatory background of active
TB, but it may still be of importance for the outcome of disease. Such potential impacts are

53
addressed in the clinical follow up study for the helminth- positive patients in an interventional
trial including deworming (paper V).
Low rate of helminth infection in HIV co-infected TB patients
It has earlier been described that parasitic infections could disturb the balance of anti-HIV
immune responses and contribute to HIV replication [125,222], which could accelerate the
progress of the HIV disease into AIDS [223,224]. The impaired cell-mediated immune response
caused by HIV infection might also lead to a higher susceptibility to parasitic infections. The
high prevalence of opportunistic parasites such as Cryptosporidium parvum and Isospora belli
among HIV-positive patients is well known [116,225], but this correlation is less evident with
helminths [225-229].
The major finding in paper IV among a total of 377 TB patients was a significantly lower rate of
helminth infection in HIV-positive compared to HIV-negative TB patients (24% vs. 36%,
p=0.03). This is in support of our hypothesis from paper III that the HIV treatment (NNRTI and
NRTI) as well as co-trimoxazole prophylaxis given to HIV-positive patients might contribute to
the observed difference. To our knowledge there is no systematic evaluation of helminth
infection or routine deworming in patients initiated on HAART in the study area (Ermias Diro,
personal communication). It has been described earlier that since the introduction of highly
active antiretroviral therapy (HAART) the mortality and morbidity for a wide variety of
opportunistic viral, bacterial, fungal and parasitic infections have decreased dramatically among
HIV-infected patients [213,214]. The potential indirect or direct role of ART on reducing the
rate of helminth infection is supported by the fact that the helminth prevalence in the same area
where we recruited the current cohort and elsewhere in Ethiopia was clearly higher before the
introduction of ART in 2005 [120,122,230]. In a study done in Ethiopia in 2004, a year before
the introduction of ART, it was reported that the prevalence of intestinal helminths was higher in
HIV- positive patients (69%) than healthy controls (57%) [122]. In a similar area, higher
helminth prevalence was reported among HIV positive patients (48%) than healthy controls
(39%) before the introduction of ART in the area [230]. On the other hand, it has been described
earlier that HIV might ‘protect’ against the establishment or survival of a mature nematode
infection, or might lead to under-diagnosis due to reduced fecundity. This has been shown in
relation to schistosomiasis, but has not yet been examined in relation to nematodes [231].

54
Furthermore, apart from the effect of HAART it should also be noted that a change in the health-
related behavior among HIV positive TB patients could contribute for the observed difference as
they received much more health care attention following the HIV diagnosis. Therefore, a well
designed follow up study with larger sample size including a more complete and detailed ART
regimen data, previous treatment history of anti-helminth drugs and hygienic practice among
patients is needed to further explore these findings.
Our sample size as well as the clinical parameters chosen could restrict the possibility to detect
subtle differences in the clinical presentation with regards to helminth infection in the
background of active TB. We used three consecutive stool samples and both direct and Kato-
Katz method in order to optimize the detection rate. Nevertheless, our diagnostic strategy to
assess helminth infections could leave occult helminth-infected patients undetected. Different
helminth species may have a variable effect on the clinical presentation and outcome and apart
from Ascaris lumbricoides this could not be evaluated within the present study due to low
detection rate of other parasites.
Effects of albendazole treatment on the clinical outcome and immunological responses in
patients with helminth infection and pulmonary tuberculosis: a randomized clinical trial
Chronic helminth infections induce important immunomodulatory effects through Tregs,
alternatively activated macrophages (also termed M2 macrophages) which are polarized by anti-
inflammatory responses, and the production of cytokines such as IL-10 and TGF-β known to
down regulate a Th1 type response [152]. It has been demonstrated that the Th1 immune-
mediated protection against M. tuberculosis is characterized by strong M.tuberculosis-specific
Th1 responses and that co-infection with helminths could modulate these immune responses as a
result of an increased Th2 and Treg activity [14,151,156]. The findings of the experimental
studies support the negative immunological impact of helminths on TB infection. Babu et
al.,showed an impairment of Th1 immune responses characterized by a decrease in PPD-specific
IFN-γ and IL-17 production during chronic human filarial infection [157]. This observation was
in agreement with a study conducted in Ethiopia which showed impairment of the Th1 response
in healthy individuals with chronic helminthic infection which was reversed after anti-helminthic
treatment [142]. However, apart from the results from experimental models and a few cross-
sectional clinical studies from the human setting performed so far, there is a lack of clinical

55
studies investigating the clinical and immunological impact of helminth co-infection with TB in
man.
Based on our previous findings, in Paper V we longitudinally assessed the clinical and
immunological impact of helminth infection during active TB infection as an effect of
deworming by albendazole. More specifically, we addressed the hypothesis that asymptomatic
helminth infection may influence the clinical improvement and immunological balance in TB
patients as a result of an effect on Tregs and cytokines associated with the Th1/Th2 balance.
Thus, we designed a randomized placebo-controlled clinical trial where such markers were
assessed before and after anti-helminthic treatment in newly diagnosed pulmonary TB patients.
Baseline clinical and immunological characteristics
A total of 1251 newly diagnosed pulmonary TB patients were screened for eligibility. Of these,
846 TB patients could not be recruited into the study due to a number of exclusion criteria, and
265 were helminth-negative and thus excluded from enrolment. In total, 140 eligible helminth-
positive TB patients were randomized: 72 to the albendazole treatment arm and 68 to the placebo
arm (Figure 9). Despite three years of enrolment of helminth-positive TB patients from three
clinical sites after 2 years of preparation we were still not able to reach our target sample size
(210 helminth positive TB patients).
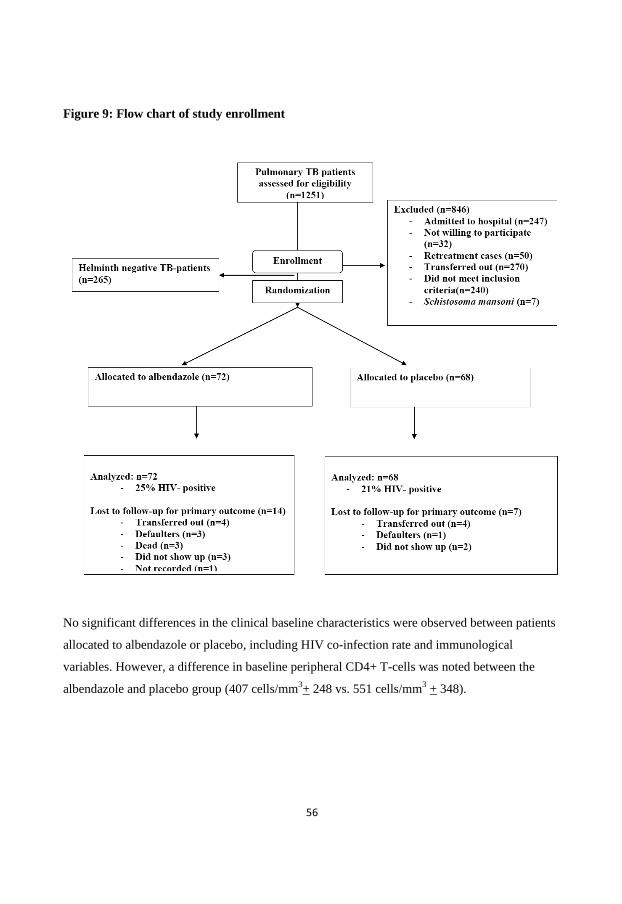
56
Figure 9: Flow chart of study enrollment
No significant differences in the clinical baseline characteristics were observed between patients
allocated to albendazole or placebo, including HIV co-infection rate and immunological
variables. However, a difference in baseline peripheral CD4+ T-cells was noted between the
albendazole and placebo group (407 cells/mm3+ 248 vs. 551 cells/mm
3 + 348).

57
Effect of albendazole treatment on study outcomes
There was a highly significant decrease in helminth infection in the albendazole treated group at
week 12 compared to placebo, 8% vs. 39%, p<0.001. However, a limitation of our study was a
high loss to follow up for helminth evaluation which was equally distributed in the albendazole
(49/72) compared to the placebo group (46/68). In a meta-analysis, albendazole showed
increased efficacy against helminth infections caused by hookworm, Ascaris lumbricoides
compared to mebendazole, and both drugs showed lower efficacy against Trichuris trichiura
[232]. In accordance to the reported efficacy of albendazole, we could observe a tendency for
persistence for trichuris relative to the other helminthes in our trial after week 12. This shows
that anti-helminthic drugs do not have similar potency against helminthic parasites. A similar
result was shown in a deworming trial conducted in HIV patients in Kenya [129], where a
helminth prevalence of 21.6% vs. 40% was observed in the albendazole treated and placebo
groups, respectively, after 12 weeks of deworming. Thus, the efficacy of deworming in our study
was higher at week 12 compared to previous studies although the helminth load and the
composition of the infecting helminth species could have an impact on this difference.
Albendazole treatment did not show any significant difference in the TB-score change
comparing week 8 to the baseline in the albendazole treated group compared with placebo (5.6
±2.87 points vs. 5.9 ±2.54, p=0.59). We further performed a subgroup analysis in a subset of
patients given albendazole treatment (n=30; HIV=21%) and placebo (n=33; HIV=17%) who had
Ascaris infection. No significant difference in the TB-score change from baseline was observed
between the two subgroups (4.89+ 2.08 vs. 5.58 + 4.7, p= 0.45). The failure to detect a clinical
difference with regards to the TB-score might be due to the fact that it is not sensitive enough to
measure subtle clinical changes from helminth infection during active TB disease in our limited
sample size. The effect of anti-TB treatment itself could mask any clinical changes from
helminth infection as a marked decline in TB-score was observed after 8 weeks of anti-TB
treatment in both treatment groups. In general, the TB-score is an appropriate tool for clinical TB
monitoring and could be applicable in clinical trials as has been indicated in Paper II. However,
its usefulness to detect subtle clinical changes due to effects other than anti-TB treatment might
need further refinement.

58
Regarding changes in the secondary outcomes, a significant decline in peripheral blood
eosinophil counts was noted in the albendazole-treated group compared to placebo (week 12:
237 cells/mm3 + 226, n=52 vs. 384 cells/mm
3 + 401, n= 52, p= 0.02). We previously showed a
significant correlation of eosinophilia with helminth infection (Paper III). Thus, even in
asymptomatic helminth infection during TB, albendazole treatment may induce a significant
reversal of immunological changes as measured by a reduction in eosinophil counts. The decline
in the peripheral eosinophil counts could be associated with the reduction in the recruitment and
infiltration of eosinophil granulocytes because of the significantly reduced rate of helminth co-
infection in the albendazole treated TB patients. The impact of such findings for the outcome and
immune response to TB is not known but the response as such is indicative of a role for
deworming in optimizing the immunity against TB. This finding is in agreement with a previous
study conducted in infants with helminth infection in which only a decline in eosinophilia was
noted after anti-helminthic treatment, but not in the levels for IFN-γ, IL-5, IL-10 and IL-13
cytokines [233].
A non-significant increase in the absolute CD4+ T-cell counts change from baseline was
observed in patients given albendazole at evaluation after 3 months, (absolute mean CD4+
change of +107 CD4+ T-cells/mm3 vs. +42 CD4+ T-cells/mm
3; p=0.16). This effect was present
primarily in HIV negative albendazole treated patients compared with the placebo group (+122
CD4+ T-cells/mm3 vs. +42 CD4+ T-cells/mm
3; p=0.17). The impact of increased CD4+ T-cell
counts in patients with TB following albendazole treatment is not clear but may indicate a
restoration of T-cell subsets following clearance of helminth infection. A similar effect of
albendazole treatment on CD4+ T-cells was shown previously in a randomized trial conducted in
Kenya [129] in helminth co-infected HIV patients. A significant increase in CD4+T-cell counts
following albendazole treatment were detected in individuals infected with Ascaris lumbricoides.
In that study, the mean absolute change in the CD4+ T-cell counts in the albendazole treated
group was +109 CD4+ T-cells/mm3 higher than in the placebo arm at 12 weeks of albendazole
treatment. The effect of increased CD4+ T-cells count after albendazole treatment may have
important immunological implications although the composition of the increased CD4+ T-cells
compared to placebo was not investigated. Moreover, the response in the peripheral blood
compared to the site of the disease in terms of host immunity and immune cell expansion may
show discrepancies [234]. The CD4+ T-cells are specialized in providing supporting signals to

59
other cells including macrophages, dendritic cells, B-lymphocytes and CD8+ T -cells through
either secretion of cytokines or direct cell-to-cell interactions. The trend for increased CD4+ T-
cells merit further studies to assess whether albendazole could lead to such increment through
dewomring or if the observed effect is related with the pharmacokinetics of albendazole.
No significant effects as a result of albendazole treatment were detected for other secondary
outcomes than eosinophil counts, including changes in chest x-ray findings, smear conversion
and IgE level. As part of secondary outcome measurement, immunological analysis for Tregs as
well as ELISPOT analysis of IFN-γ, IL-5 and IL-10 producing PBMCs was performed in a
subset of 35 helminth positive TB patients. The HIV co-infection rate for this subset in
albendazole and placebo groups was 50% (6/12) vs.15% (3/20), respectively (p= 0.04). Thus, an
unfortunate balance in the rate of HIV infection was present. Nonetheless, in the baseline
measurements no significant impact of HIV infection was present on the Treg levels. Reports
have shown that the ability of HIV to directly infect Tregs is controversial [235,236]. On the
other hand, it has been reported that exposure of Tregs to HIV selectively promotes their
survival, which is supported by the observation that Tregs are accumulated in the gut or in the
tonsils of HAART-naive HIV positive patients where active viral replication is taking place
[236]. Similarly the kinetics of Tregs lacks a consensus in HIV-positive patients. Increased Tregs
numbers was reported independently of HIV treatment [237], whilst others showed an increase
and a subsequent return to levels comparable to controls [238]. In this study, no significant
difference in Tregs, or IFN-γ, IL-5 or IL-10 response was observed between the two groups.
Sub-group analysis for Tregs, IFN-γ, IL-5 and IL-10 was done for TB patients with Ascaris
infection (n=12 in total), but did not show any significant differences between the albendazole
and the placebo group. This is in agreement with previous studies conducted in HIV patients in
which albendazole treatment of HIV patients with Ascaris infection showed no difference in the
levels of Th1/Th2 cytokines (such as IFN-γ, IL-4, IL-5, IL-13), but where a decline in the IL-10
levels was observed in albendazole treated patients compared to the placebo group [233,239]. In
accordance to these findings, the results from the present study, excluding patients with
persistent helminth infection at week 12 in a multivariate analysis adjusted for baseline level of
IL-10, age, sex, and HIV, showed an increased trend towards significantly lower levels of IL-10
in the albendazole group compared to placebo (p= 0.08). In animal models, an elevated IL-10

60
level was associated with susceptibility to TB, and mice lacking IL-10 were shown to clear M.
bovis BCG infection more efficiently [240]. Additionally, mice over expressing IL-10 were more
susceptible to M. tuberculosis infection, exhibiting an impaired Th1 response as characterized by
decreased numbers of activated T-cells in the peripheral blood and lung tissue [111]. Overall, the
finding of a decrease in IL-10 following deworming compared to placebo support the previous
experimental data.
Regulatory T-cells, IFN-γ, IL-5 and IL-10 are increased in helminth co-infected TB
patients compared to helminth negative TB patients and healthy subjects
There is emerging evidence regarding the influence of Tregs and associated cytokines during
infectious diseases in facilitating pathogen persistence and modulating the host immune response
to infection [241]. The significant increase in Tregs and associated cytokines in TB patients co-
infected with helminths in paper V strengthens this notion. We observed a significant increase in
the frequency of Tregs in helminth co-infected TB patients compared to helminth-negative TB
patients, (2.2+1.5, n=25 vs. 1.2+1.1, n=16; p=0.03) and to helminth-positive community
controls, (1.1+0.99, n=14; p=0.018, (Table 3a and 3b).
The increased expansion of the Treg population in the present study was accompanied by an
increased level of IL-5 and IL-10, among helminth-positive TB patients compared to helminth-
negative TB patients. In animal models, it has been described that T-cells are highly associated
with helminth infection and are in great number in areas prevalent in helminths. Tregs are
common in the gut and most are Foxp3+ Tregs that express CD4. In the colon mucosa, more
than 50% of the Foxp3+ T-cells also produce IL-10. Our finding supports the strong association
of helminths with Tregs and the Th2 associated cytokines IL-5 and IL-10 which have the
capacity to modulate the host immune response. The observed increase in baseline IFN-γ
response in helminth infected TB patients might be associated with induction of a Th1-type
response during the early stages of helminth infection, as previously indicated [242,243]. After
analyzing the distribution IFN-γ responses, it was observed that the increase in the helminth-
positive TB patients is due to the marked increased (more than two standard deviations higher
than the mean) levels of this cytokine in a few patients which were outliers (6/36).
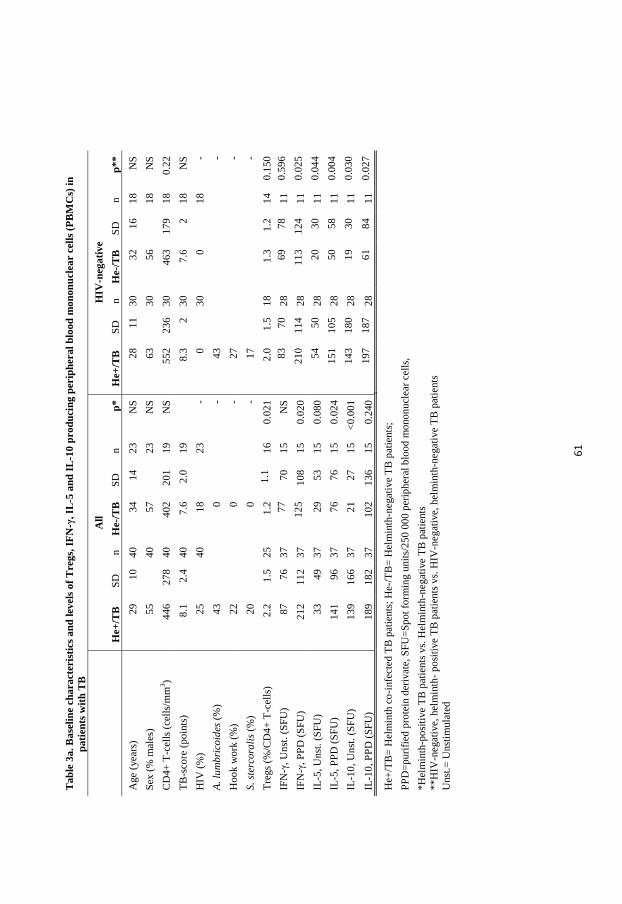
61
T
ab
le 3
a.
Ba
seli
ne
cha
ract
eris
tics
an
d l
evel
s o
f T
reg
s, I
FN
-γ,
IL-5
an
d I
L-1
0 p
rod
uci
ng
per
iph
era
l b
loo
d m
on
on
ucl
ear
cell
s (P
BM
Cs)
in
pa
tien
ts w
ith
TB
A
ll
HIV
-neg
ati
ve
H
e+/T
B
SD
n
H
e-/T
B
SD
n
p
*
He+
/TB
S
D
n
He-
/TB
S
D
n
p*
*
Age
(yea
rs)
29
10
40
34
14
23
NS
2
8
11
30
32
16
18
NS
Sex
(%
mal
es)
55
4
0
57
2
3
NS
6
3
3
0
56
1
8
NS
CD
4+
T-c
ells
(ce
lls/
mm
3)
44
6
27
8
40
40
2
20
1
19
NS
5
52
23
6
30
46
3
17
9
18
0.2
2
TB
-sco
re (
po
ints
) 8
.1
2.4
4
0
7.6
2
.0
19
8
.3
2
30
7.6
2
1
8
NS
HIV
(%
) 2
5
4
0
18
2
3
- 0
30
0
1
8
-
A.
lum
bri
coid
es (
%)
43
0
- 4
3
-
Ho
ok w
ork
(%
) 2
2
0
- 2
7
-
S.
ster
cora
lis
(%)
20
0
- 1
7
-
Tre
gs
(%/C
D4
+ T
-cel
ls)
2.2
1
.5
25
1.2
1
.1
16
0.0
21
2
.0
1.5
1
8
1.3
1
.2
14
0.1
50
IFN
-γ,
Un
st.
(SF
U)
87
76
37
77
70
15
NS
8
3
70
28
69
78
11
0.5
96
IFN
-γ,
PP
D (
SF
U)
21
2
11
2
37
12
5
10
8
15
0.0
20
2
10
11
4
28
11
3
12
4
11
0.0
25
IL-5
, U
nst
. (S
FU
) 3
3
49
37
29
53
15
0.0
80
5
4
50
28
20
30
11
0.0
44
IL-5
, P
PD
(S
FU
) 1
41
96
37
76
76
15
0.0
24
1
51
10
5
28
50
58
11
0.0
04
IL-1
0,
Un
st.
(SF
U)
13
9
16
6
37
21
27
15
<0
.00
1
14
3
18
0
28
19
30
11
0.0
30
IL-1
0,
PP
D (
SF
U)
18
9
18
2
37
10
2
13
6
15
0.2
40
1
97
18
7
28
61
84
11
0.0
27
He+
/TB
= H
elm
inth
co
-infe
cted
TB
pat
ients
; H
e-/
TB
= H
elm
inth
-negat
ive
TB
pat
ients
;
PP
D=
puri
fied
pro
tein
der
ivat
e, S
FU
=S
po
t fo
rmin
g u
nit
s/2
50
00
0 p
erip
her
al b
loo
d m
ono
nu
clea
r ce
lls,
*H
elm
inth
-po
siti
ve
TB
pat
ients
vs.
Hel
min
th-n
egat
ive
TB
pat
ients
**H
IV-n
egat
ive,
hel
min
th-
po
siti
ve
TB
pat
ients
vs.
HIV
-neg
ativ
e, h
elm
inth
-negat
ive
TB
pat
ients
Unst
.= U
nst
imula
ted
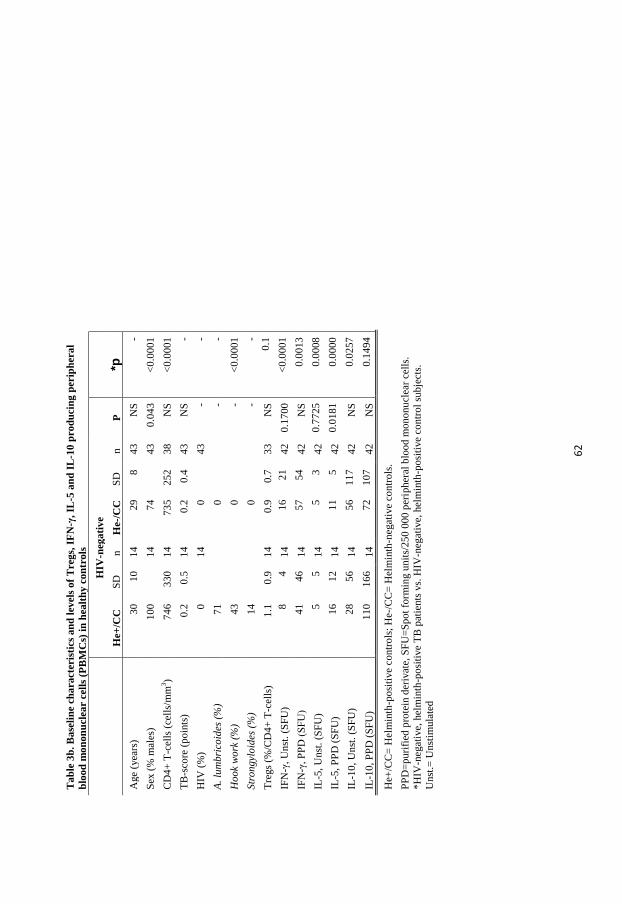
62
T
ab
le 3
b.
Ba
seli
ne
cha
ract
eris
tics
an
d l
evel
s o
f T
reg
s, I
FN
-γ,
IL-5
an
d I
L-1
0 p
rod
uci
ng
per
iph
era
l
blo
od
mo
no
nu
clea
r ce
lls
(PB
MC
s) i
n h
ealt
hy
co
ntr
ols
H
IV-n
ega
tiv
e
H
e+/C
C
SD
n
H
e-/C
C
SD
n
P
*p
Age
(yea
rs)
30
10
14
29
8
43
NS
-
Sex
(%
mal
es)
10
0
1
4
74
4
3
0.0
43
<
0.0
001
CD
4+
T-c
ells
(ce
lls/
mm
3)
74
6
33
0
14
73
5
25
2
38
NS
<
0.0
001
TB
-sco
re (
po
ints
) 0
.2
0.5
1
4
0.2
0
.4
43
NS
-
HIV
(%
) 0
14
0
4
3
- -
A.
lum
bri
coid
es (
%)
71
0
- -
Ho
ok
wo
rk (
%)
43
0
- <
0.0
001
Str
on
gyl
oid
es (
%)
14
0
- -
Tre
gs
(%/C
D4
+ T
-cel
ls)
1.1
0
.9
14
0.9
0
.7
33
NS
0
.1
IFN
-γ,
Un
st.
(SF
U)
8
4
14
16
21
42
0.1
700
<
0.0
001
IFN
-γ,
PP
D (
SF
U)
41
46
14
57
54
42
NS
0
.00
13
IL-5
, U
nst
. (S
FU
)
5
5
14
5
3
42
0.7
725
0
.00
08
IL-5
, P
PD
(S
FU
) 1
6
12
14
11
5
42
0.0
181
0
.00
00
IL-1
0,
Un
st.
(SF
U)
28
56
14
56
11
7
42
NS
0
.02
57
IL-1
0,
PP
D (
SF
U)
11
0
16
6
14
72
10
7
42
NS
0
.14
94
He+
/CC
= H
elm
inth
-po
siti
ve
co
ntr
ols
; H
e-/C
C=
Hel
min
th-n
egat
ive
contr
ols
.
PP
D=
puri
fied
pro
tein
der
ivat
e, S
FU
=S
po
t fo
rmin
g u
nit
s/2
50
00
0 p
erip
her
al b
loo
d m
ono
nu
clea
r ce
lls.
*H
IV-n
egat
ive,
hel
min
th-p
osi
tive
TB
pat
ients
vs.
HIV
-neg
ati
ve,
hel
min
th-p
osi
tive
contr
ol
sub
ject
s.
Unst
.= U
nst
imula
ted

63
We were not able to reach the target sample size in recruitment (n=210). The expansion and
decentralization of anti-TB treatment centres allowed patients to complete the TB treatment in
their vicinity, which made recruitment of the initial calculated sample size in the planned period
unachievable at the study sites. Moreover, consideration of subgroups such as the impact of HIV
infection in immunological and clinical evaluations is problematic and might need even larger
sample sizes or even exclusion in preference of HIV-negative TB patients only. The lack of a
previous study in relation to helminth infection, albendazole treatment and tuberculosis makes
the power calculation challenging. However, we reasoned that the impact of deworming may be
much less pronounced than from the anti-TB drugs. The marked decline in TB-score due to anti-
TB treatment alone in patients allocated to albendazole and placebo groups could mask clinical
effects due to deworming. It is difficult to exclude an undetected low grade helminth infection in
the helminth- negative TB patients and controls, and a possible re-infection after albendazole
treatment.
The major strengths of the study are the combinatory clinical and immunological evaluation.
Compared to previous studies, the sample size for the analysis of Tregs as well as IL-5, IL-10
and IFN-γ was relatively large. Additionally, the approach to use a randomized design to
investigate the effect of albendazole treatment on asymptomatic helminth infection is novel.
In conclusion, albendazole treatment did not affect the clinical outcome of helminth co-infected
TB patients as analyzed by changes in TB-score after two months. Deworming of helminth co-
infected TB patients induced a significant decline in peripheral eosinophil granulocytes, and a
non-significant increase in peripheral CD4+ T-cells after 12 weeks. Tuberculosis patients who
received effective albendazole treatment showed a borderline significant decrease in IL-10 levels
at follow up. In patients with TB, we showed that asymptomatic helminth infection induces an
increased Treg response and increased levels of IL-5 and IL-10 producing PBMCs. Taken
together, a main conclusion of our study is that helminth infection significantly affects the
immunological response to TB. Larger multi-center studies are warranted to further investigate
the effect on deworming on clinical outcome of TB as well as the helminth-induced mechanism
of immune modulation in active TB.

64
CONCLUSIONS
During treatment of TB there is a significant decline in the proportion of QFN positive
patients down to the baseline level similar to a healthy control group.
The agreement between TST and QFN is poor in active TB but satisfactory in community
controls.
Early changes in the TB-score composed of clinical signs and symptoms of TB may be
used to identify patients in need of closer follow up and hospitalization.
More than one third of patients with active pulmonary TB were co-infected with
helminths, which was significantly higher than in healthy controls.
Tuberculosis patients with HIV infection had a significantly lower rate of helminth
infection than HIV-positive TB patients.
Eosinophilia and elevated IgE levels correlated with asymptomatic helminth infection in
patients with active pulmonary TB.
Tuberculosis patients with asymptomatic helminth infection had a significantly increased
frequency of Tregs as well as increased levels of IL-5 and IL-10 PBMCs compared to
helminth-negative TB patients and healthy controls.
Albendazole treatment of helminth-positive TB patients resulted in a marked decline in
the rate of helminth infection after three months compared to placebo whereas it did not
significantly affect the primary outcome (change in TB-score between baseline and 2
months)
Albendazole treatment resulted in a significant decline in eosinophil cells after 3 month
compared to placebo as well as a trend towards a lower rate of IL-10 producing
peripheral mononuclear blood cells after 3 months compared to placebo.

65
CONCLUDING REMARKS
The work presented in this thesis contributes to the understanding of the clinical and
immunological impact of helminth co-infection in active TB. Previously, the clinical and
immunological impact of helminth infection during TB was mainly described in experimental
models or in cells from healthy individuals. The main conclusion of this work is that
asymptomatic helminth infection has an immunological impact in active TB. This has important
implications for the effect of the present BCG and future TB vaccines, but may also be of
importance for the outcome of active TB infection. The randomized clinical trial of albendazole
versus placebo in helminth positive TB patients is the first of its kind and included a thorough
follow-up including immunological assessment. Larger multi-center studies with a sufficient
representation of different helminth species in each treatment arm are important to make sub-
group analysis including considerations of HIV co-infection.
Novel tools for treatment monitoring of TB were evaluated such as the TB-score and
longitudinal QFN measurements. This has significant importance in the clinical care of patients.
It fills a major gap in resource limited settings in particular where there is a deficiency in
objective surrogate markers of clinical outcome in TB which could be monitored longitudinally.
The available WHO recommended strategy of sputum AFB smears and radiology based
assessment is only applicable to a limited subset of TB patients and suffers from limitations in
terms of sensitivity, objectivity and logistics. Our findings evaluated the strength and limitations
of the QFN assay and the TB-score tools for TB treatment monitoring and their relevance in
clinical research. We showed that QFN results are influenced by HIV infection. Furthermore,
after 2 to 6 months of TB treatment, TB patients showed a modest decline in the rate of QFN
positive patients to a level comparable with that recorded for healthy control subjects from the
same area. It was shown that TB-score is a potential marker for early prediction of patients at
high risk of mortality and adverse outcomes. The findings of our helminth-TB co-infection
studies add new clinical and immunological data, which were previously limited. The
immunomodulatory effects of helminth infection have been demonstrated in several
experimental studies. We showed an increased helminth burden in TB patients compared to
healthy controls from the same area. It is not known whether asymptomatic helminth infection
could increase susceptibility to TB disease following exposure. The observed higher burden of

66
helminth co-infection in active TB patients as well as the immunomodulation induced by
helminths, merits further long-term follow-up studies of larger cohorts, including effects of
deworming. Such studies also need to consider other confounders such as HIV co-infection and
nutritional status including vitamin D deficiency. Our immunological findings strongly support
the impact of asymptomatic helminth co-infection on host immunity to active TB. The increase
in Tregs in helminth co-infected TB patients, suggests a significant immunological effect as the
Tregs are known for their immunosuppressive role. Furthermore, the increased levels of the Th1
attenuating cytokine, IL-10, further underlines the Th1 attenuating immunological impact of
helminths on the host immune response during active TB. The hypothesis generated is shown in
Figure 11.
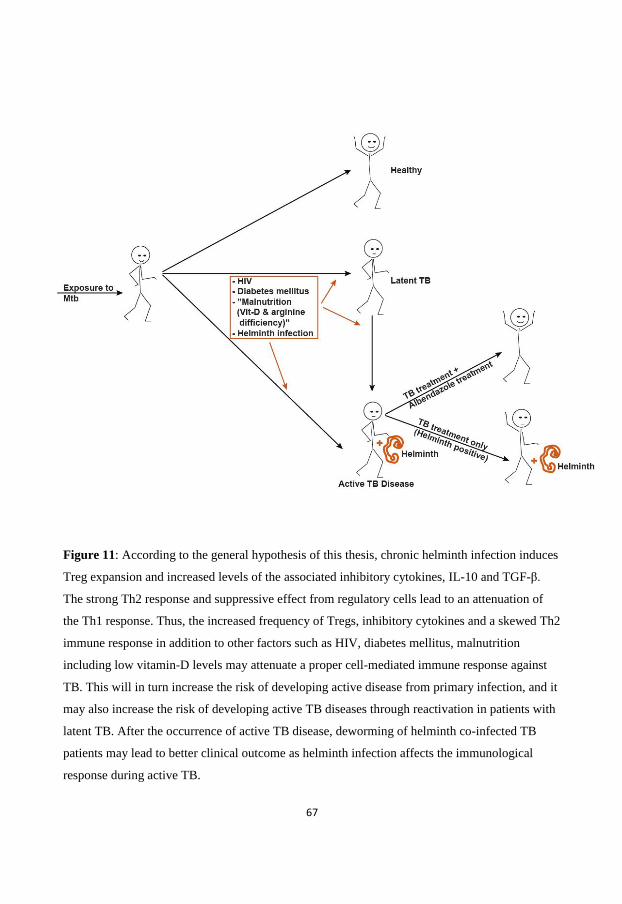
67
Figure 11: According to the general hypothesis of this thesis, chronic helminth infection induces
Treg expansion and increased levels of the associated inhibitory cytokines, IL-10 and TGF-β.
The strong Th2 response and suppressive effect from regulatory cells lead to an attenuation of
the Th1 response. Thus, the increased frequency of Tregs, inhibitory cytokines and a skewed Th2
immune response in addition to other factors such as HIV, diabetes mellitus, malnutrition
including low vitamin-D levels may attenuate a proper cell-mediated immune response against
TB. This will in turn increase the risk of developing active disease from primary infection, and it
may also increase the risk of developing active TB diseases through reactivation in patients with
latent TB. After the occurrence of active TB disease, deworming of helminth co-infected TB
patients may lead to better clinical outcome as helminth infection affects the immunological
response during active TB.

68
REFERENCES
1. WHO (2012) Global Tuberculosis Report. Geneva, Switzerland: WHO. 2. Dye C, Watt CJ, Bleed DM, Hosseini SM, Raviglione MC (2005) Evolution of tuberculosis control and
prospects for reducing tuberculosis incidence, prevalence, and deaths globally. JAMA 293: 2767-2775.
3. Daniel TM (2006) The history of tuberculosis. Respir Med 100: 1862-1870. 4. Gutierrez MC, Brisse S, Brosch R, Fabre M, Omais B, et al. (2005) Ancient origin and gene mosaicism of
the progenitor of Mycobacterium tuberculosis. PLoS Pathog 1: e5. 5. Verma A, Sampla AK, Tyagi JS (1999) Mycobacterium tuberculosis rrn promoters: differential usage
and growth rate-dependent control. J Bacteriol 181: 4326-4333. 6. Palomino JC LS, Ritacco V. (Belgium, 2007, ) Tuberculosis 2007: From basic science to patient care. 1st
edn, BourcillierKamps: 53-680. 7. O'Garra A, Redford PS, McNab FW, Bloom CI, Wilkinson RJ, et al. (2013) The immune response in
tuberculosis. Annu Rev Immunol 31: 475-527. 8. Glaziou P, Falzon D, Floyd K, Raviglione M (2013) Global epidemiology of tuberculosis. Semin Respir
Crit Care Med 34: 3-16. 9. Moore-Gillon JC (2007) Tuberculosis and its future management. Thorax 62: 1019-1021. 10. Ngo AT, Duc NH, Lan NH, Maynart M, Mayaud C, et al. (2007) [Mechanisms and causes of death in
143 Vietnamese HIV-infected patients hospitalized for tuberculosis]. Rev Pneumol Clin 63: 139-146.
11. WHO (2011) Global Tuberculosis control report. Geneva, Switzerland: WHO. 12. UNAIDS (2012) Report on the global AIDS epidemic Joint United Nations Programme on HIV/AIDS
(UNAIDS). 13. Ethiopia FMoH (2008) Tuberculosis, leprosy and TB/HIV prevention and control programme manual.
5-57 p. 14. Borkow G, Weisman Z, Leng Q, Stein M, Kalinkovich A, et al. (2001) Helminths, human
immunodeficiency virus and tuberculosis. Scand J Infect Dis 33: 568-571. 15. Kramnik I, Dietrich WF, Demant P, Bloom BR (2000) Genetic control of resistance to experimental
infection with virulent Mycobacterium tuberculosis. Proc Natl Acad Sci USA 97: 8560-8565. 16. Nardell EA (1993) Environmental control of tuberculosis. Med Clin North Am 77: 1315-1334. 17. Bellamy R, Beyers N, McAdam KP, Ruwende C, Gie R, et al. (2000) Genetic susceptibility to
tuberculosis in Africans: a genome-wide scan. Proc Natl Acad Sci U S A 97: 8005-8009. 18. Wilkinson RJ, Llewelyn M, Toossi Z, Patel P, Pasvol G, et al. (2000) Influence of vitamin D deficiency
and vitamin D receptor polymorphisms on tuberculosis among Gujarati Asians in west London: a case-control study. Lancet 355: 618-621.
19. Behler F, Steinwede K, Balboa L, Ueberberg B, Maus R, et al. (2012) Role of Mincle in alveolar macrophage-dependent innate immunity against mycobacterial infections in mice. J Immunol 189: 3121-3129.
20. Shaler CR, Horvath CN, Jeyanathan M, Xing Z (2013) Within the Enemy's Camp: contribution of the granuloma to the dissemination, persistence and transmission of Mycobacterium tuberculosis. Front Immunol 4: 30.
21. Cooper AM (2009) Cell-mediated immune responses in tuberculosis. Annu Rev Immunol 27: 393-422. 22. Mendez-Samperio P (2012) Immunological mechanisms by which concomitant helminth infections
predispose to the development of human tuberculosis. Korean J Parasitol 50: 281-286. 23. Liu PT, Stenger S, Li H, Wenzel L, Tan BH, Krutzik SR, Ochoa MT, Schauber J, Wu K, Meinken C, Kamen
DL, Wagner M, Bals R, Steinmeyer A, Zügel U, Gallo RL, Eisenberg D, Hewison M, Hollis BW,

69
Adams JS, Bloom BR, Modlin RL. (2006) Toll-like receptor triggering of a vitamin D-mediated human antimicrobial response. Science 311: 1770-1773.
24. Yang CT, Cambier CJ, Davis JM, Hall CJ, Crosier PS, et al. (2012) Neutrophils exert protection in the early tuberculous granuloma by oxidative killing of mycobacteria phagocytosed from infected macrophages. Cell Host Microbe 12: 301-312.
25. Persson A B-JR, Eklund D, Lundström C, Stendahl O. (2009) Induction of apoptosis in human neutrophils by Mycobacterium tuberculosis is dependent on mature bacterial lipoproteins. Microb Pathog 47: 143-150.
26. Persson YA B-JR, Rahman S, Zheng L, Stendahl O. (2008) Mycobacterium tuberculosis-induced apoptotic neutrophils trigger a pro-inflammatory response in macrophages through release of heat shock protein 72, acting in synergy with the bacteria. Microbes Infect 10: 233-240.
27. Fulton SA, Reba SM, Martin TD, Boom WH (2002) Neutrophil-mediated mycobacteriocidal immunity in the lung during Mycobacterium bovis BCG infection in C57BL/6 mice. Infect Immun 70: 5322-5327.
28. Schluger NW (2001) Recent advances in our understanding of human host responses to tuberculosis. Respir Res 2: 157-163.
29. Schön T, Lerm M, Stendahl O. (2013) Shortening the 'short-course' therapy- insights into host immunity may contribute to new treatment strategies for tuberculosis. J Intern Med 273: 368-382.
30. Mueller H, Fae KC, Magdorf K, Ganoza CA, Wahn U, et al. (2011) Granulysin-expressing CD4+ T cells as candidate immune marker for tuberculosis during childhood and adolescence. PLoS One 6: e29367.
31. Stenger S, Mazzaccaro RJ, Uyemura K, Cho S, Barnes PF, Rosat JP, Sette A, Brenner MB, Porcelli SA, Bloom BR, Modlin RL. (1997) Differential effects of cytolytic T cell subsets on intracellular infection. Science 276: 1684-1687.
32. Flannagan RS, Cosío G, Grinstein S. (2009) Antimicrobial mechanisms of phagocytes and bacterial evasion strategies. Nat Rev Microbiol 7: 355-366.
33. Shu CC, Wu MF, Hsu CL, Huang CT, Wang JY, et al. (2013) Apoptosis-associated biomarkers in tuberculosis: promising for diagnosis and prognosis prediction. BMC Infect Dis 13: 45.
34. C. N (2006) Role of iNOS in human host defense. Science 312: 1874-1875. 35. Nathan C, Shiloh MU (2000) Reactive oxygen and nitrogen intermediates in the relationship between
mammalian hosts and microbial pathogens. Proc Natl Acad Sci U S A 97: 8841-8848. 36. Schön T, Elmberger G, Negesse Y, Pando RH, Sundqvist T, Britton S. (2004) Local production of nitric
oxide in patients with tuberculosis. Int J Tuberc Lung Dis 8: 1134-1137. 37. Idh J, Westman A, Elias D, Moges F, Getachew A, Gelaw A, Sundqvist T, Forslund T, Alemu A, Ayele B,
Diro E, Melese E, Wondmikun Y, Britton S, Stendahl O, Schön T. (2008) Nitric oxide production in the exhaled air of patients with pulmonary tuberculosis in relation to HIV co-infection. BMC Infect Dis 8.
38. Schön T, Elias D, Moges F, Melese E, Tessema T, Stendahl O, Britton S, Sundqvist T (2003) Arginine as an adjuvant to chemotherapy improves clinical outcome in active tuberculosis. Eur Respir J 21: 483-488.
39. Dheda K, Schwander SK, Zhu B, van Zyl-Smit RN, Zhang Y (2010) The immunology of tuberculosis: from bench to bedside. Respirology 15: 433-450.
40. Yahagi A, Umemura M, Tamura T, Kariyone A, Begum MD, et al. (2010) Suppressed induction of mycobacterial antigen-specific Th1-type CD4+ T cells in the lung after pulmonary mycobacterial infection. Int Immunol 22: 307-318.
41. Abebe F, Bjune G (2009) The protective role of antibody responses during Mycobacterium tuberculosis infection. Clin Exp Immunol 157: 235-243.

70
42. Shaler CR HC, Jeyanathan M, Xing Z. (2013) Within the Enemy's Camp: contribution of the granuloma to the dissemination, persistence and transmission of Mycobacterium tuberculosis. Front Immunol 4: doi: 10.3389.
43. Flynn JL, Chan J (2001) Immunology of tuberculosis. Annu Rev Immunol 19: 93-129. 44. Cooper AM, Mayer-Barber KD, Sher A (2011) Role of innate cytokines in mycobacterial infection.
Mucosal Immunol 4: 252-260. 45. DG. R (2011) Mycobacterium tuberculosis and the intimate discourse of a chronic infection.
Russell DG. Immunol Rev 240: 252-268. 46. Jeong YJ, Lee KS (2008) Pulmonary tuberculosis: up-to-date imaging and management. AJR Am J
Roentgenol 191: 834-844. 47. Kisembo HN, Boon SD, Davis JL, Okello R, Worodria W, Cattamanchi A, Huang L, Kawooya MG. (2012)
Chest radiographic findings of pulmonary tuberculosis in severely immunocompromised patients with the human immunodeficiency virus. Br J Radiol 85: e130-139.
48. Lawn SD (2013) Diagnosis of pulmonary tuberculosis. Curr Opin Pulm Med 19: 280-288. 49. Leung E, Minion J, Benedetti A, Pai M, Menzies D (2012) Microcolony culture techniques for
tuberculosis diagnosis: a systematic review. Int J Tuberc Lung Dis 16: 16-23, i-iii. 50. Xia H, Song YY, Zhao B, Kam KM, O'Brien RJ, et al. (2013) Multicentre evaluation of Ziehl-Neelsen and
light-emitting diode fluorescence microscopy in China. Int J Tuberc Lung Dis 17: 107-112. 51. Ssengooba W, Kateete DP, Wajja A, Bugumirwa E, Mboowa G, et al. (2012) An Early Morning Sputum
Sample Is Necessary for the Diagnosis of Pulmonary Tuberculosis, Even with More Sensitive Techniques: A Prospective Cohort Study among Adolescent TB-Suspects in Uganda. Tuberc Res Treat 2012: 970203.
52. Somoskovi A, Hotaling JE, Fitzgerald M, O'Donnell D, Parsons LM, et al. (2001) Lessons from a proficiency testing event for acid-fast microscopy. Chest 120: 250-257.
53. Van Deun A, Hamid Salim A, Aung KJ, Hossain MA, Chambugonj N, et al. (2005) Performance of variations of carbolfuchsin staining of sputum smears for AFB under field conditions. Int J Tuberc Lung Dis 9: 1127-1133.
54. Saito H (1998) [Laboratory media for the cultivation of tubercle bacillus]. Kekkaku 73: 329-337. 55. Yamane N, Chilima BZ, Tosaka M, Okazawa Y, Tanno K (1996) [Determination of antimycobacterial
activities of fluoroquinolones against clinical isolates of Mycobacterium tuberculosis: comparative determination with egg-based Ogawa and agar-based Middlebrook 7H10 media]. Kekkaku 71: 453-458.
56. Wu X, Zhang J, Liang J, Lu Y, Li H, et al. (2007) Comparison of three methods for rapid identification of mycobacterial clinical isolates to the species level. J Clin Microbiol 45: 1898-1903.
57. Helb D JM, Story E, Boehme C, Wallace E, Ho K, Kop J, Owens MR, Rodgers R, Banada P, Safi H, Blakemore R, Lan NT, Jones-López EC, Levi M, Burday M, Ayakaka I, Mugerwa RD, McMillan B, Winn-Deen E, Christel L, Dailey P, Perkins MD, Persing DH, Alland D. (2010) Rapid detection of Mycobacterium tuberculosis and rifampin resistance by use of on-demand, near-patient technology. J Clin Microbiol 48: 229-237.
58. WHO (2010) Tuberculosis treatment guideline. Geneva, Switzerland: WHO. 59. Van Rie A, Mellet K, John MA, Scott L, Page-Shipp L, et al. (2012) False-positive rifampicin resistance
on Xpert(R) MTB/RIF: case report and clinical implications. Int J Tuberc Lung Dis 16: 206-208. 60. Arend SM, Thijsen SF, Leyten EM, Bouwman JJ, Franken WP, et al. (2007) Comparison of two
interferon-gamma assays and tuberculin skin test for tracing tuberculosis contacts. Am J Respir Crit Care Med 175: 618-627.

71
61. Rangaka MX WK, Glynn JR, Ling D, Menzies D, Mwansa-Kambafwile J, Fielding K, Wilkinson RJ, Pai M. (2012) Predictive value of interferon-γ release assays for incident active tuberculosis: a systematic review and meta-analysis. Lancet Infect Dis 12: 45-55.
62. Curley C (2003) New guidelines: what to do about an unexpected positive tuberculin skin test. Cleve Clin J Med 70: 49-55.
63. Huebner RE, Schein MF, Bass JB, Jr. (1993) The tuberculin skin test. Clin Infect Dis 17: 968-975. 64. Taylor Z, Nolan CM, Blumberg HM (2005) Controlling tuberculosis in the United States.
Recommendations from the American Thoracic Society, CDC, and the Infectious Diseases Society of America. MMWR Recomm Rep 54: 1-81.
65. Lalvani A (2007) Diagnosing tuberculosis infection in the 21st century: new tools to tackle an old enemy. Chest 131: 1898-1906.
66. Pai M, Zwerling A, Menzies D (2008) Systematic review: T-cell-based assays for the diagnosis of latent tuberculosis infection: an update. Ann Intern Med 149: 177-184.
67. Idh J, Abate E, Westman A, Elias D, Janols H, Gelaw A, Getachew A, Alemu S, Aseffa A, Britton S, Stendahl O,
Schon, T (2010) Kinetics of the QuantiFERON-TB Gold In-Tube test during treatment of patients with sputum smear-positive tuberculosis in relation to initial TST result and severity of disease. Scand J Infect Dis 42: 650-657. 68. Tufariello JM, Chan J, Flynn JL (2003) Latent tuberculosis: mechanisms of host and bacillus that
contribute to persistent infection. Lancet Infect Dis 3: 578-590. 69. Brock I, Munk ME, Kok-Jensen A, Andersen P (2001) Performance of whole blood IFN-gamma test for
tuberculosis diagnosis based on PPD or the specific antigens ESAT-6 and CFP-10. Int J Tuberc Lung Dis 5: 462-467.
70. Lalvani A, Nagvenkar P, Udwadia Z, Pathan AA, Wilkinson KA, et al. (2001) Enumeration of T cells specific for RD1-encoded antigens suggests a high prevalence of latent Mycobacterium tuberculosis infection in healthy urban Indians. J Infect Dis 183: 469-477.
71. Kipfer B, Reichmuth M, Buchler M, Meisels C, Bodmer T (2008) Tuberculosis in a Swiss army training camp: contact investigation using an Interferon gamma release assay. Swiss Med Wkly 138: 267-272.
72. Oxlade O, Pinto M, Trajman A, Menzies D (2013) How Methodologic Differences Affect Results of Economic Analyses: A Systematic Review of Interferon Gamma Release Assays for the Diagnosis of LTBI. PLoS One 8: e56044.
73. Snider GL (1997) Tuberculosis then and now: a personal perspective on the last 50 years. Ann Intern Med 126: 237-243.
74. Bishai JD, Bishai WR, Bishai DM (2010) Heightened vulnerability to MDR-TB epidemics after controlling drug-susceptible TB. PLoS One 5: e12843.
75. Ahmad S, Mokaddas E (2004) Contribution of AGC to ACC and other mutations at codon 315 of the katG gene in isoniazid-resistant Mycobacterium tuberculosis isolates from the Middle East. Int J Antimicrob Agents 23: 473-479.
76. Jordan TS, Davies PD (2010) Clinical tuberculosis and treatment outcomes. Int J Tuberc Lung Dis 14: 683-688.
77. Gillespie SH (2002) Evolution of drug resistance in Mycobacterium tuberculosis: clinical and molecular perspective. Antimicrob Agents Chemother 46: 267-274.
78. Youmans GP, Williston EH, et al. (1946) Increase in resistance of tubercle bacilli to streptomycin; a preliminary report. Proc Staff Meet Mayo Clin 21: 126.
79. Andersen P, Doherty TM (2005) Learning from BCG: Designing a better tuberculosis vaccine. Discov Med 5: 383-387.

72
80. Buddle BM, Skinner MA, Wedlock DN, Collins DM, de Lisle GW (2002) New generation vaccines and delivery systems for control of bovine tuberculosis in cattle and wildlife. Vet Immunol Immunopathol 87: 177-185.
81. de Lisle GW, Wards BJ, Buddle BM, Collins DM (2005) The efficacy of live tuberculosis vaccines after presensitization with Mycobacterium avium. Tuberculosis (Edinb) 85: 73-79.
82. Doherty TM, Andersen P (2005) Vaccines for tuberculosis: novel concepts and recent progress. Clin Microbiol Rev 18: 687-702.
83. Elias D, Britton S, Aseffa A, Engers H, Akuffo H (2008) Poor immunogenicity of BCG in helminth infected population is associated with increased in vitro TGF-beta production. Vaccine 26: 3897-3902.
84. Kaufmann SH (2005) Robert Koch, the Nobel Prize, and the ongoing threat of tuberculosis. N Engl J Med 353: 2423-2426.
85. Lin PL, Dietrich J, Tan E, Abalos RM, Burgos J, et al. (2012) The multistage vaccine H56 boosts the effects of BCG to protect cynomolgus macaques against active tuberculosis and reactivation of latent Mycobacterium tuberculosis infection. J Clin Invest 122: 303-314.
86. Grode L, Ganoza CA, Brohm C, Weiner J, 3rd, Eisele B, et al. (2013) Safety and immunogenicity of the recombinant BCG vaccine VPM1002 in a phase 1 open-label randomized clinical trial. Vaccine 31: 1340-1348.
87. Tameris MD, Hatherill M, Landry BS, Scriba TJ, Snowden MA, et al. (2013) Safety and efficacy of MVA85A, a new tuberculosis vaccine, in infants previously vaccinated with BCG: a randomised, placebo-controlled phase 2b trial. Lancet.
88. Chen J, Chen GW, Wang MZ, Chen HF, Lin MZ, et al. (2012) [Epidemiological characteristics of soil-borne nematodiasis in Xiamen City]. Zhongguo Xue Xi Chong Bing Fang Zhi Za Zhi 24: 109-110.
89. Bethony J, Brooker S, Albonico M, Geiger SM, Loukas A, et al. (2006) Soil-transmitted helminth infections: ascariasis, trichuriasis, and hookworm. Lancet 367: 1521-1532.
90. Brooker S, Kabatereine NB, Smith JL, Mupfasoni D, Mwanje MT, et al. (2009) An updated atlas of human helminth infections: the example of East Africa. Int J Health Geogr 8: 42.
91. de Silva NR, Brooker S, Hotez PJ, Montresor A, Engels D, et al. (2003) Soil-transmitted helminth infections: updating the global picture. Trends Parasitol 19: 547-551.
92. Hotez PJ, Brindley PJ, Bethony JM, King CH, Pearce EJ, et al. (2008) Helminth infections: the great neglected tropical diseases. J Clin Invest 118: 1311-1321.
93. Aranzamendi C, Sofronic-Milosavljevic L, Pinelli E (2013) Helminths: Immunoregulation and Inflammatory Diseases-Which Side Are Trichinella spp. and Toxocara spp. on? J Parasitol Res 2013: 329438.
94. Anderson RM, Truscott JE, Pullan RL, Brooker SJ, Hollingsworth TD (2013) How effective is school-based deworming for the community-wide control of soil-transmitted helminths? PLoS Negl Trop Dis 7: e2027.
95. Brooker S, Miguel EA, Moulin S, Luoba AI, Bundy DA, et al. (2000) Epidemiology of single and multiple species of helminth infections among school children in Busia District, Kenya. East Afr Med J 77: 157-161.
96. Chan MS (1997) The global burden of intestinal nematode infections--fifty years on. Parasitol Today 13: 438-443.
97. Elias D, Mengistu G, Akuffo H, Britton S (2006) Are intestinal helminths risk factors for developing active tuberculosis? Trop Med Int Health 11: 551-558.
98. Fincham JE, Markus MB, Adams VJ (2003) Could control of soil-transmitted helminthic infection influence the HIV/AIDS pandemic. Acta Trop 86: 315-333.

73
99. Hotez PJ, Molyneux DH, Fenwick A, Ottesen E, Ehrlich Sachs S, et al. (2006) Incorporating a rapid-impact package for neglected tropical diseases with programs for HIV/AIDS, tuberculosis, and malaria. PLoS Med 3: e102.
100. Le Hesran JY, Akiana J, Ndiaye el HM, Dia M, Senghor P, et al. (2004) Severe malaria attack is associated with high prevalence of Ascaris lumbricoides infection among children in rural Senegal. Trans R Soc Trop Med Hyg 98: 397-399.
101. Garnier R, Boulinier T, Gandon S (2013) Evolution of the temporal persistence of immune protection. Biol Lett 9: 20130017.
102. Koyasu S, Moro K, Tanabe M, Takeuchi T (2010) Natural helper cells: a new player in the innate immune response against helminth infection. Adv Immunol 108: 21-44.
103. Saenz SA, Noti M, Artis D (2010) Innate immune cell populations function as initiators and effectors in Th2 cytokine responses. Trends Immunol 31: 407-413.
104. Fessler J, Felber A, Duftner C, Dejaco C (2013) Therapeutic Potential of Regulatory T cells in Autoimmune Disorders. BioDrugs.Apr 12
105. Josefowicz SZ, Lu LF, Rudensky AY (2012) Regulatory T cells: mechanisms of differentiation and function. Annu Rev Immunol 30: 531-564.
106. Fontenot JD, Gavin MA, Rudensky AY (2003) Foxp3 programs the development and function of CD4+CD25+ regulatory T cells. Nat Immunol 4: 330-336.
107. Hartigan-O'Connor DJ, Poon C, Sinclair E, McCune JM (2007) Human CD4+ regulatory T cells express lower levels of the IL-7 receptor alpha chain (CD127), allowing consistent identification and sorting of live cells. J Immunol Methods 319: 41-52.
108. Belkaid Y, Rouse BT (2005) Natural regulatory T cells in infectious disease. Nat Immunol 6: 353-360. 109. Finney CA, Taylor MD, Wilson MS, Maizels RM (2007) Expansion and activation of CD4(+)CD25(+)
regulatory T cells in Heligmosomoides polygyrus infection. Eur J Immunol 37: 1874-1886. 110. Rausch S, Huehn J, Kirchhoff D, Rzepecka J, Schnoeller C, et al. (2008) Functional analysis of effector
and regulatory T cells in a parasitic nematode infection. Infect Immun 76: 1908-1919. 111. Turner JD, Jackson JA, Faulkner H, Behnke J, Else KJ, et al. (2008) Intensity of intestinal infection
with multiple worm species is related to regulatory cytokine output and immune hyporesponsiveness. J Infect Dis 197: 1204-1212.
112. Figueiredo CA, Barreto ML, Rodrigues LC, Cooper PJ, Silva NB, et al. (2010) Chronic intestinal helminth infections are associated with immune hyporesponsiveness and induction of a regulatory network. Infect Immun 78: 3160-3167.
113. Elliott DE, Summers RW, Weinstock JV (2007) Helminths as governors of immune-mediated inflammation. Int J Parasitol 37: 457-464.
114. Fallon PG, Mangan NE (2007) Suppression of TH2-type allergic reactions by helminth infection. Nat Rev Immunol 7: 220-230.
115. Fleming J, Fabry Z (2007) The hygiene hypothesis and multiple sclerosis. Ann Neurol 61: 85-89. 116. Karp CL, Auwaerter PG (2007) Coinfection with HIV and tropical infectious diseases. II. Helminthic,
fungal, bacterial, and viral pathogens. Clin Infect Dis 45: 1214-1220. 117. Assefa S, Erko B, Medhin G, Assefa Z, Shimelis T (2009) Intestinal parasitic infections in relation to
HIV/AIDS status, diarrhea and CD4 T-cell count. BMC Infect Dis 9: 155. 118. Saksirisampant W, Prownebon J, Saksirisampant P, Mungthin M, Siripatanapipong S, et al. (2009)
Intestinal parasitic infections: prevalences in HIV/AIDS patients in a Thai AIDS-care centre. Ann Trop Med Parasitol 103: 573-581.
119. Abate E, Belayneh M, Gelaw A, Idh J, Getachew A, et al. (2012) The impact of asymptomatic helminth co-infection in patients with newly diagnosed tuberculosis in north-west ethiopia. PLoS One 7: e42901.

74
120. Hailemariam G, Kassu A, Abebe G, Abate E, Damte D, et al. (2004) Intestinal parasitic infections in HIV/AIDS and HIV seronegative individuals in a teaching hospital, Ethiopia. Jpn J Infect Dis 57: 41-43.
121. Kassu A, Mengistu G, Ayele B, Diro E, Mekonnen F, et al. (2007) HIV and intestinal parasites in adult TB patients in a teaching hospital in Northwest Ethiopia. Trop Doct 37: 222-224.
122. Mariam ZT, Abebe G, Mulu A (2008) Opportunistic and other intestinal parasitic infections in AIDS patients, HIV seropositive healthy carriers and HIV seronegative individuals in southwest Ethiopia. East Afr J Public Health 5: 169-173.
123. Kalinkovich A, Borkow G, Weisman Z, Tsimanis A, Stein M, et al. (2001) Increased CCR5 and CXCR4 expression in Ethiopians living in Israel: environmental and constitutive factors. Clin Immunol 100: 107-117.
124. Kalinkovich A, Weisman Z, Leng Q, Borkow G, Stein M, et al. (1999) Increased CCR5 expression with decreased beta chemokine secretion in Ethiopians: relevance to AIDS in Africa. J Hum Virol 2: 283-289.
125. Gopinath R, Ostrowski M, Justement SJ, Fauci AS, Nutman TB (2000) Filarial infections increase susceptibility to human immunodeficiency virus infection in peripheral blood mononuclear cells in vitro. J Infect Dis 182: 1804-1808.
126. Shapira-Nahor O, Kalinkovich A, Weisman Z, Greenberg Z, Nahmias J, et al. (1998) Increased susceptibility to HIV-1 infection of peripheral blood mononuclear cells from chronically immune-activated individuals. AIDS 12: 1731-1733.
127. Gallagher M, Malhotra I, Mungai PL, Wamachi AN, Kioko JM, et al. (2005) The effects of maternal helminth and malaria infections on mother-to-child HIV transmission. AIDS 19: 1849-1855.
128. McElroy MD, Elrefaei M, Jones N, Ssali F, Mugyenyi P, et al. (2005) Coinfection with Schistosoma mansoni is associated with decreased HIV-specific cytolysis and increased IL-10 production. J Immunol 174: 5119-5123.
129. Walson JL, Otieno PA, Mbuchi M, Richardson BA, Lohman-Payne B, et al. (2008) Albendazole treatment of HIV-1 and helminth co-infection: a randomized, double-blind, placebo-controlled trial. AIDS 22: 1601-1609.
130. Kjetland EF, Ndhlovu PD, Gomo E, Mduluza T, Midzi N, et al. (2006) Association between genital schistosomiasis and HIV in rural Zimbabwean women. AIDS 20: 593-600.
131. van den Biggelaar AH, Rodrigues LC, van Ree R, van der Zee JS, Hoeksma-Kruize YC, et al. (2004) Long-term treatment of intestinal helminths increases mite skin-test reactivity in Gabonese schoolchildren. J Infect Dis 189: 892-900.
132. Kok A, Robinson MJ (1976) Ige, parasites, and allergy. Lancet 2: 633. 133. Lynch NR, Palenque M, Hagel I, DiPrisco MC (1997) Clinical improvement of asthma after
anthelminthic treatment in a tropical situation. Am J Respir Crit Care Med 156: 50-54. 134. Bourke CD, Mutapi F, Nausch N, Photiou DM, Poulsen LK, et al. (2012) Trichuris suis ova therapy for
allergic rhinitis does not affect allergen-specific cytokine responses despite a parasite-specific cytokine response. Clin Exp Allergy 42: 1582-1595.
135. Chu KM, Watermeyer G, Shelly L, Janssen J, May TD, et al. (2013) Childhood Helminth Exposure Is Protective Against Inflammatory Bowel Disease: A Case Control Study in South Africa. Inflamm Bowel Dis 19: 614-620.
136. Hsu SJ, Tseng PH, Chen PJ (2005) Trichuris suis therapy for ulcerative colitis: nonresponsive patients may need anti-helminth therapy. Gastroenterology 129: 768-769; author reply 769.
137. Kuijk LM, Klaver EJ, Kooij G, van der Pol SM, Heijnen P, et al. (2012) Soluble helminth products suppress clinical signs in murine experimental autoimmune encephalomyelitis and differentially modulate human dendritic cell activation. Mol Immunol 51: 210-218.

75
138. Zaccone P, Cooke A (2013) Vaccine against autoimmune disease: can helminths or their products provide a therapy? Curr Opin Immunol.
139. Summers RW, Elliott DE, Urban JF, Jr., Thompson RA, Weinstock JV (2005) Trichuris suis therapy for active ulcerative colitis: a randomized controlled trial. Gastroenterology 128: 825-832.
140. Cooper PJ, Chico M, Sandoval C, Espinel I, Guevara A, et al. (2001) Human infection with Ascaris lumbricoides is associated with suppression of the interleukin-2 response to recombinant cholera toxin B subunit following vaccination with the live oral cholera vaccine CVD 103-HgR. Infect Immun 69: 1574-1580.
141. Cooper PJ, Chico ME, Losonsky G, Sandoval C, Espinel I, et al. (2000) Albendazole treatment of children with ascariasis enhances the vibriocidal antibody response to the live attenuated oral cholera vaccine CVD 103-HgR. J Infect Dis 182: 1199-1206.
142. Elias D, Wolday D, Akuffo H, Petros B, Bronner U, et al. (2001) Effect of deworming on human T cell responses to mycobacterial antigens in helminth-exposed individuals before and after bacille Calmette-Guerin (BCG) vaccination. Clin Exp Immunol 123: 219-225.
143. Elias D, Akuffo H, Pawlowski A, Haile M, Schon T, et al. (2005) Schistosoma mansoni infection reduces the protective efficacy of BCG vaccination against virulent Mycobacterium tuberculosis. Vaccine 23: 1326-1334.
144. Bentwich Z, Kalinkovich A, Weisman Z, Borkow G, Beyers N, et al. (1999) Can eradication of helminthic infections change the face of AIDS and tuberculosis? Immunol Today 20: 485-487.
145. Cooper PJ, Espinel I, Paredes W, Guderian RH, Nutman TB (1998) Impaired tetanus-specific cellular and humoral responses following tetanus vaccination in human onchocerciasis: a possible role for interleukin-10. J Infect Dis 178: 1133-1138.
146. Cooper PJ, Espinel I, Wieseman M, Paredes W, Espinel M, et al. (1999) Human onchocerciasis and tetanus vaccination: impact on the postvaccination antitetanus antibody response. Infect Immun 67: 5951-5957.
147. Malhotra I, Mungai P, Wamachi A, Kioko J, Ouma JH, et al. (1999) Helminth- and Bacillus Calmette-Guerin-induced immunity in children sensitized in utero to filariasis and schistosomiasis. J Immunol 162: 6843-6848.
148. Sabin EA, Araujo MI, Carvalho EM, Pearce EJ (1996) Impairment of tetanus toxoid-specific Th1-like immune responses in humans infected with Schistosoma mansoni. J Infect Dis 173: 269-272.
149. Nookala S, Srinivasan S, Kaliraj P, Narayanan RB, Nutman TB (2004) Impairment of tetanus-specific cellular and humoral responses following tetanus vaccination in human lymphatic filariasis. Infect Immun 72: 2598-2604.
150. Prost A, Schlumberger M, Fayet MT (1983) Response to tetanus immunization in onchocerciasis patients. Ann Trop Med Parasitol 77: 83-85.
151. Elias D, Britton S, Kassu A, Akuffo H (2007) Chronic helminth infections may negatively influence immunity against tuberculosis and other diseases of public health importance. Expert Rev Anti Infect Ther 5: 475-484.
152. van Riet E, Hartgers FC, Yazdanbakhsh M (2007) Chronic helminth infections induce immunomodulation: consequences and mechanisms. Immunobiology 212: 475-490.
153. Tristao-Sa R, Ribeiro-Rodrigues R, Johnson LT, Pereira FE, Dietze R (2002) Intestinal nematodes and pulmonary tuberculosis. Rev Soc Bras Med Trop 35: 533-535.
154. Elias D, Akuffo H, Britton S (2006) Helminthes could influence the outcome of vaccines against TB in the tropics. Parasite Immunol 28: 507-513.
155. Diniz LM, Zandonade E, Dietze R, Pereira FE, Ribeiro-Rodrigues R (2001) Short report: do intestinal nematodes increase the risk for multibacillary leprosy? Am J Trop Med Hyg 65: 852-854.
156. Salgame P (2005) Host innate and Th1 responses and the bacterial factors that control Mycobacterium tuberculosis infection. Curr Opin Immunol 17: 374-380.

76
157. Babu S, Bhat SQ, Kumar NP, Anuradha R, Kumaran P, et al. (2009) Attenuation of toll-like receptor expression and function in latent tuberculosis by coexistent filarial infection with restoration following antifilarial chemotherapy. PLoS Negl Trop Dis 3: e489.
158. Stewart GR, Boussinesq M, Coulson T, Elson L, Nutman T, et al. (1999) Onchocerciasis modulates the immune response to mycobacterial antigens. Clin Exp Immunol 117: 517-523.
159. Hubner MP, Killoran KE, Rajnik M, Wilson S, Yim KC, et al. (2012) Chronic helminth infection does not exacerbate Mycobacterium tuberculosis infection. PLoS Negl Trop Dis 6: e1970.
160. Central Intelligence Agency (CIA) U (2013) The World Fact book. https://www.cia.gov/library/publications/the-world-factbook/geos/et.html.
161. (2007) Census 2007 Tables: Amhara Region. Addis Ababa: Central Statistical Agency, Ethiopia 162. Federal Ministry of Health E (2003) GUIDELINES FOR USE OF ANTIRETROVIRAL DRUGS IN ETHIOPIA.
Ministry of Health Disease Prevention and Control Department In collaboration with HIV/ AIDS Prevention and Control office (HAPCO) and Drug Administration and Control Authority (DACA).
163. Wejse C, Gustafson P, Nielsen J, Gomes VF, Aaby P, et al. (2008) TBscore: Signs and symptoms from tuberculosis patients in a low-resource setting have predictive value and may be used to assess clinical course. Scand J Infect Dis 40: 111-120.
164. Sleigh A, Hoff R, Mott K, Barreto M, de Paiva TM, et al. (1982) Comparison of filtration staining (Bell) and thick smear (Kato) for the detection of quantitation of Schistosoma mansoni eggs in faeces. Trans R Soc Trop Med Hyg 76: 403-406.
165. WHO (2010) Treatment of Tuberculosis Guidelines. Fourth edition ed. http://whqlibdoc.who.int/publications/2010/9789241547833_eng.pdf: WHO.
166. Janols H, Abate E, Idh J, Senbeto M, Britton S, et al. (2012) Early treatment response evaluated by a clinical scoring system correlates with the prognosis of pulmonary tuberculosis patients in Ethiopia: a prospective follow-up study. Scand J Infect Dis 44: 828-834.
167. Aiken AM, Hill PC, Fox A, McAdam KP, Jackson-Sillah D, et al. (2006) Reversion of the ELISPOT test after treatment in Gambian tuberculosis cases. BMC Infect Dis 6: 66.
168. Carrara S, Vincenti D, Petrosillo N, Amicosante M, Girardi E, et al. (2004) Use of a T cell-based assay for monitoring efficacy of antituberculosis therapy. Clin Infect Dis 38: 754-756.
169. Dheda K, Pooran A, Pai M, Miller RF, Lesley K, et al. (2007) Interpretation of Mycobacterium tuberculosis antigen-specific IFN-gamma release assays (T-SPOT.TB) and factors that may modulate test results. J Infect 55: 169-173.
170. Pathan AA, Wilkinson KA, Klenerman P, McShane H, Davidson RN, et al. (2001) Direct ex vivo analysis of antigen-specific IFN-gamma-secreting CD4 T cells in Mycobacterium tuberculosis-infected individuals: associations with clinical disease state and effect of treatment. J Immunol 167: 5217-5225.
171. Al-Attiyah R, Mustafa AS, Abal AT, Madi NM, Andersen P (2003) Restoration of mycobacterial antigen-induced proliferation and interferon-gamma responses in peripheral blood mononuclear cells of tuberculosis patients upon effective chemotherapy. FEMS Immunol Med Microbiol 38: 249-256.
172. Ferrand RA, Bothamley GH, Whelan A, Dockrell HM (2005) Interferon-gamma responses to ESAT-6 in tuberculosis patients early into and after anti-tuberculosis treatment. Int J Tuberc Lung Dis 9: 1034-1039.
173. Ulrichs T, Anding R, Kaufmann SH, Munk ME (2000) Numbers of IFN-gamma-producing cells against ESAT-6 increase in tuberculosis patients during chemotherapy. Int J Tuberc Lung Dis 4: 1181-1183.
174. Vekemans J, Lienhardt C, Sillah JS, Wheeler JG, Lahai GP, et al. (2001) Tuberculosis contacts but not patients have higher gamma interferon responses to ESAT-6 than do community controls in The Gambia. Infect Immun 69: 6554-6557.

77
175. Wu-Hsieh BA, Chen CK, Chang JH, Lai SY, Wu CH, et al. (2001) Long-lived immune response to early secretory antigenic target 6 in individuals who had recovered from tuberculosis. Clin Infect Dis 33: 1336-1340.
176. Chee CB, KhinMar KW, Gan SH, Barkham TM, Koh CK, et al. (2010) Tuberculosis treatment effect on T-cell interferon-gamma responses to Mycobacterium tuberculosis-specific antigens. Eur Respir J 36: 355-361.
177. Dominguez J, De Souza-Galvao M, Ruiz-Manzano J, Latorre I, Prat C, et al. (2009) T-cell responses to the Mycobacterium tuberculosis-specific antigens in active tuberculosis patients at the beginning, during, and after antituberculosis treatment. Diagn Microbiol Infect Dis 63: 43-51.
178. Adetifa IM, Ota MO, Walther B, Hammond AS, Lugos MD, et al. (2010) Decay kinetics of an interferon gamma release assay with anti-tuberculosis therapy in newly diagnosed tuberculosis cases. PLoS One 5.
179. Doherty TM, Demissie A, Olobo J, Wolday D, Britton S, et al. (2002) Immune responses to the Mycobacterium tuberculosis-specific antigen ESAT-6 signal subclinical infection among contacts of tuberculosis patients. J Clin Microbiol 40: 704-706.
180. Vordermeier HM, Chambers MA, Cockle PJ, Whelan AO, Simmons J, et al. (2002) Correlation of ESAT-6-specific gamma interferon production with pathology in cattle following Mycobacterium bovis BCG vaccination against experimental bovine tuberculosis. Infect Immun 70: 3026-3032.
181. Markowitz N, Hansen NI, Wilcosky TC, Hopewell PC, Glassroth J, et al. (1993) Tuberculin and anergy testing in HIV-seropositive and HIV-seronegative persons. Pulmonary Complications of HIV Infection Study Group. Ann Intern Med 119: 185-193.
182. Raby E, Moyo M, Devendra A, Banda J, De Haas P, et al. (2008) The effects of HIV on the sensitivity of a whole blood IFN-gamma release assay in Zambian adults with active tuberculosis. PLoS One 3: e2489.
183. van Zyl-Smit RN, Pai M, Peprah K, Meldau R, Kieck J, et al. (2009) Within-subject variability and boosting of T-cell interferon-gamma responses after tuberculin skin testing. Am J Respir Crit Care Med 180: 49-58.
184. Aabye MG, Ravn P, PrayGod G, Jeremiah K, Mugomela A, et al. (2009) The impact of HIV infection and CD4 cell count on the performance of an interferon gamma release assay in patients with pulmonary tuberculosis. PLoS One 4: e4220.
185. Caetano Mota P, Carvalho A, Valente I, Braga R, Duarte R (2012) Predictors of delayed sputum smear and culture conversion among a Portuguese population with pulmonary tuberculosis. Rev Port Pneumol 18: 72-79.
186. Goodridge A, Cueva C, Lahiff M, Muzanye G, Johnson JL, et al. (2012) Anti-phospholipid antibody levels as biomarker for monitoring tuberculosis treatment response. Tuberculosis (Edinb) 92: 243-247.
187. Enarson DA (1995) The International Union Against Tuberculosis and Lung Disease model National Tuberculosis Programmes. Tuber Lung Dis 76: 95-99.
188. Keeler E, Perkins MD, Small P, Hanson C, Reed S, et al. (2006) Reducing the global burden of tuberculosis: the contribution of improved diagnostics. Nature 444 Suppl 1: 49-57.
189. Knaus WA, Wagner DP, Draper EA, Zimmerman JE, Bergner M, et al. (1991) The APACHE III prognostic system. Risk prediction of hospital mortality for critically ill hospitalized adults. Chest 100: 1619-1636.
190. Lim WS, van der Eerden MM, Laing R, Boersma WG, Karalus N, et al. (2003) Defining community acquired pneumonia severity on presentation to hospital: an international derivation and validation study. Thorax 58: 377-382.

78
191. Chiappini E, Fossi F, Bonsignori F, Sollai S, Galli L, et al. (2012) Utility of interferon-gamma release assay results to monitor anti-tubercular treatment in adults and children. Clin Ther 34: 1041-1048.
192. Horita N, Miyazawa N, Yoshiyama T, Sato T, Yamamoto M, et al. (2013) Development and validation of a tuberculosis prognostic score for smear-positive in-patients in Japan. Int J Tuberc Lung Dis 17: 54-60.
193. Wejse C, Gomes VF, Rabna P, Gustafson P, Aaby P, et al. (2009) Vitamin D as supplementary treatment for tuberculosis: a double-blind, randomized, placebo-controlled trial. Am J Respir Crit Care Med 179: 843-850.
194. Rudolf F, Joaquim LC, Vieira C, Bjerregaard-Andersen M, Andersen A, et al. (2013) The Bandim tuberculosis score: Reliability and comparison with the Karnofsky performance score. Scand J Infect Dis 45: 256-264.
195. Borkow G, Bentwich Z (2004) Chronic immune activation associated with chronic helminthic and human immunodeficiency virus infections: role of hyporesponsiveness and anergy. Clin Microbiol Rev 17: 1012-1030, table of contents.
196. Hotez PJ, Molyneux DH, Stillwaggon E, Bentwich Z, Kumaresan J (2006) Neglected tropical diseases and HIV/AIDS. Lancet 368: 1865-1866.
197. Fontanet AL, Woldemichael T, Sahlu T, van Dam GJ, Messele T, et al. (2000) Epidemiology of HIV and Schistosoma mansoni infections among sugar-estate residents in Ethiopia. Ann Trop Med Parasitol 94: 145-155.
198. Kallestrup P, Zinyama R, Gomo E, Butterworth AE, Mudenge B, et al. (2005) Schistosomiasis and HIV-1 infection in rural Zimbabwe: effect of treatment of schistosomiasis on CD4 cell count and plasma HIV-1 RNA load. J Infect Dis 192: 1956-1961.
199. Modjarrad K, Zulu I, Redden DT, Njobvu L, Freedman DO, et al. (2005) Prevalence and predictors of intestinal helminth infections among human immunodeficiency virus type 1-infected adults in an urban African setting. Am J Trop Med Hyg 73: 777-782.
200. Allen JE, Maizels RM (2011) Diversity and dialogue in immunity to helminths. Nat Rev Immunol 11: 375-388.
201. Al-Qaoud KM, Taubert A, Zahner H, Fleischer B, Hoerauf A (1997) Infection of BALB/c mice with the filarial nematode Litomosoides sigmodontis: role of CD4+ T cells in controlling larval development. Infect Immun 65: 2457-2461.
202. Cervi L, MacDonald AS, Kane C, Dzierszinski F, Pearce EJ (2004) Cutting edge: dendritic cells copulsed with microbial and helminth antigens undergo modified maturation, segregate the antigens to distinct intracellular compartments, and concurrently induce microbe-specific Th1 and helminth-specific Th2 responses. J Immunol 172: 2016-2020.
203. Reimert CM, Fitzsimmons CM, Joseph S, Mwatha JK, Jones FM, et al. (2006) Eosinophil activity in Schistosoma mansoni infections in vivo and in vitro in relation to plasma cytokine profile pre- and posttreatment with praziquantel. Clin Vaccine Immunol 13: 584-593.
204. Tsegaye A MT, Tilahun T, Hailu E, Sahlu T, Doorly R, Fontanet AL, Rinke de Wit TF. (1999) Immunohematological reference ranges for adult Ethiopians. Clin Diagn Lab Immunol 6: 410-414.
205. Vieira Silva CC, Nogueira Ferraz CR, Fornari JV, Sena Barnabe CA (2012) Epidemiological analysis of eosinophilia and elevation of immunoglobulin E as a predictable and relative risk of enteroparasitosis. Rev Cubana Med Trop 64: 22-26.
206. Xie SY, Yuan M, Hu F, Liu YM, Zeng XJ, et al. (2012) [Study on immune status of patients with schistosomiasis japonica in Poyang Lake region I characterization of antigen-specific antibody isotype responses to Schistosoma japonicum]. Zhongguo Xue Xi Chong Bing Fang Zhi Za Zhi 24: 510-513, 521.

79
207. King CL, Malhotra I, Mungai P, Wamachi A, Kioko J, et al. (1998) B cell sensitization to helminthic infection develops in utero in humans. J Immunol 160: 3578-3584.
208. Weil GJ, Hussain R, Kumaraswami V, Tripathy SP, Phillips KS, et al. (1983) Prenatal allergic sensitization to helminth antigens in offspring of parasite-infected mothers. J Clin Invest 71: 1124-1129.
209. Galioto AM, Hess JA, Nolan TJ, Schad GA, Lee JJ, et al. (2006) Role of eosinophils and neutrophils in innate and adaptive protective immunity to larval strongyloides stercoralis in mice. Infect Immun 74: 5730-5738.
210. Herbert DR, Lee JJ, Lee NA, Nolan TJ, Schad GA, et al. (2000) Role of IL-5 in innate and adaptive immunity to larval Strongyloides stercoralis in mice. J Immunol 165: 4544-4551.
211. Specht S, Saeftel M, Arndt M, Endl E, Dubben B, et al. (2006) Lack of eosinophil peroxidase or major basic protein impairs defense against murine filarial infection. Infect Immun 74: 5236-5243.
212. Perona-Wright G MK, Taylor J, Zaph C, Artis D, Pearce EJ, Mohrs M. (2008) Cutting edge: Helminth infection induces IgE in the absence of mu- or delta-chain expression. J Immunol 15;181: 6697-6701.
213. Alfonso Y, Monzote L (2011) HIV Protease Inhibitors: Effect on the Opportunistic Protozoan Parasites. Open Med Chem J 5: 40-50.
214. Lucas SB (1990) Missing infections in AIDS. Trans R Soc Trop Med Hyg 84 Suppl 1: 34-38. 215. Perona-Wright G MK, Taylor J, Zaph C, Artis D, Pearce EJ, Mohrs M. (2013 ) HIV Treatments have
Malaria Gametocyte Killing and Transmission Blocking Activity. J Infect Dis Mar 28. 216. Derouin F, Santillana-Hayat M (2000) Anti-toxoplasma activities of antiretroviral drugs and
interactions with pyrimethamine and sulfadiazine in vitro. Antimicrob Agents Chemother 44: 2575-2577.
217. Hobbs CV, Voza T, Coppi A, Kirmse B, Marsh K, et al. (2009) HIV protease inhibitors inhibit the development of preerythrocytic-stage plasmodium parasites. J Infect Dis 199: 134-141.
218. Morales Gomez MA (2004) [Highly Active AntiRetroviral Therapy and cryptosporidiosis]. Parassitologia 46: 95-99.
219. Hobbs CV, Voza T, De La Vega P, Vanvliet J, Conteh S, et al. (2012) HIV nonnucleoside reverse transcriptase inhibitors and trimethoprim-sulfamethoxazole inhibit plasmodium liver stages. J Infect Dis 206: 1706-1714.
220. Aditya NP, Patankar S, Madhusudhan B (2010) Assessment of in vivo antimalarial activity of rifampicin, isoniazide, and ethambutol combination therapy. Parasitol Res 106: 1481-1484.
221. James WP M-TG, Norgan NG, Bistrian BR, Shetty PS, Ferro-Luzzi A. (1994) The value of arm circumference measurements in assessing chronic energy deficiency in Third World adults. Eur J Clin Nutr 48: 883-894.
222. Newton CR (2005) Interaction between Plasmodium falciparum and human immunodeficiency virus type 1 on the central nervous system of African children. J Neurovirol 11 Suppl 3: 45-51.
223. Karanja DM, Colley DG, Nahlen BL, Ouma JH, Secor WE (1997) Studies on schistosomiasis in western Kenya: I. Evidence for immune-facilitated excretion of schistosome eggs from patients with Schistosoma mansoni and human immunodeficiency virus coinfections. Am J Trop Med Hyg 56: 515-521.
224. Tawill SA, Gallin M, Erttmann KD, Kipp W, Bamuhiiga J, et al. (1996) Impaired antibody responses and loss of reactivity to Onchocerca volvulus antigens by HIV-seropositive onchocerciasis patients. Trans R Soc Trop Med Hyg 90: 85-89.
225. Nielsen NO FH, Magnussen P, Krarup H, Magesa S, Simonsen PE. (2007) Co-infection with subclinical HIV and Wuchereria bancrofti, and the role of malaria and hookworms, in adult Tanzanians: infection intensities, CD4/CD8 counts and cytokine responses. Trans R Soc Trop Med Hyg 101: 602-612.

80
226. Gomez Morales MA AC, Ludovisi A, Rossi P, Scaglia M, Pozio E. (1995) Opportunistic and non-opportunistic parasites in HIV-positive and negative patients with diarrhoea in Tanzania. Trop Med Parasitol 46: 109-114.
227. Li-Guang Tian J-XC, Tian-Ping Wang, Guo-Jin Cheng, Peter Steinmann, Feng-Feng Wang, Yu-Chun Cai, Xiao-Mei Yin, Jian Guo, Li Zhou, Xiao-Nong Zhou (2012) Co-infection of HIV and intestinal parasites in rural area of China. Parasit Vectors 5.
228. Lindo JF DJ, Ager AL, de Gourville EM, Solo-Gabriele H, Klaskala WI, Baum MK, Palmer CJ. (1998) Intestinal parasitic infections in human immunodeficiency virus (HIV)-positive and HIV-negative individuals in San Pedro Sula, Honduras. Am J Trop Med Hyg 58: 431-435.
229. Nielsen NO SP, Magnussen P, Magesa S, Friis H. (2006) Cross-sectional relationship between HIV, lymphatic filariasis and other parasitic infections in adults in coastal northeastern Tanzania. Trans R Soc Trop Med Hyg 100: 543-550.
230. Ebba Abate EM, Mohammed Awol, Afework Kassu (2001) Cryptosporidiosis and Isosporiasis among HIV/AIDS patients in Jimma, Southwest Ethiopia. Journal of Ethiopian Medical Practice 3: 64-69.
231. Brown M MP, Kaleebu P, Elliott AM. (2006) Helminths and HIV infection: epidemiological observations on immunological hypotheses. Parasite Immunol 28: 613-623.
232. Keiser J UJ (2008) Efficacy of current drugs against soil-transmitted helminth infections: systematic review and meta-analysis. JAMA 299: 1937-1948.
233. Wright VJ, Ame SM, Haji HS, Weir RE, Goodman D, et al. (2009) Early exposure of infants to GI nematodes induces Th2 dominant immune responses which are unaffected by periodic anthelminthic treatment. PLoS Negl Trop Dis 3: e433.
234. Brighenti S AJ (2012) Local immune responses in human tuberculosis: learning from the site of infection. J Infect Dis 205: S316-324.
235. Horta A NC, Amorim-Machado P, Coutinho-Teixeira V, Barreira-Silva P, Boavida S, Costa P, Sarmento-Castro R, Castro AG, Correia-Neves M. (2013) Poor immune reconstitution in HIV-infected patients associates with high percentage of regulatory CD4+ T cells. PLoS One 8: e57336.
236. Moreno-Fernandez ME ZW, Blackard JT, Franchini G, Chougnet CA. (2009) Human regulatory T cells are targets for human immunodeficiency Virus (HIV) infection, and their susceptibility differs depending on the HIV type 1 strain. J Virol 83: 12925-12933.
237. Gaardbo JC NS, Vedel SJ, Ersbøll AK, Harritshøj L, Ryder LP, Nielsen JO, Kolte L. (2008) Regulatory T cells in human immunodeficiency virus-infected patients are elevated and independent of immunological and virological status, as well as initiation of highly active anti-retroviral therapy. Clin Exp Immunol 154: 80-86.
238. Presicce P OK, King E, Pratt J, Fichtenbaum CJ, Chougnet CA. (2011) Frequency of circulating regulatory T cells increases during chronic HIV infection and is largely controlled by highly active antiretroviral therapy. PLoS One 6: e28118.
239. Blish CA, Sangare L, Herrin BR, Richardson BA, John-Stewart G, et al. (2010) Changes in plasma cytokines after treatment of ascaris lumbricoides infection in individuals with HIV-1 infection. J Infect Dis 201: 1816-1821.
240. Murray PJ (1999) Defining the requirements for immunological control of mycobacterial infections. Trends Microbiol 7: 366-372.
241. de Almeida AS, Fiske CT, Sterling TR, Kalams SA (2012) Increased frequency of regulatory T cells and T lymphocyte activation in persons with previously treated extrapulmonary tuberculosis. Clin Vaccine Immunol 19: 45-52.
242. du Plessis N, Kleynhans L, Thiart L, van Helden PD, Brombacher F, et al. (2012) Acute helminth infection enhances early macrophage mediated control of mycobacterial infection. Mucosal Immunol.

81
243. Elias D, Akuffo H, Britton S (2005) PPD induced in vitro interferon gamma production is not a reliable correlate of protection against Mycobacterium tuberculosis. Trans R Soc Trop Med Hyg 99: 363-368.

82
ACKNOWLEDGEMENTS
I would like to express my sincere gratitude to everyone who has been involved in completing this work or taken
part in my life in other ways during these years. I would first like to thank all the participants in the studies included
in this thesis. I would especially like to mention the following people:
My supervisor, Thomas Schön for giving me the opportunity to be your student and believing in me. I greatly give
much credit for your continuous effort in refining me. All your enthusiasm to research, encouragement and your
impressive knowledge made it a pleasure working under your guidance. I am always inspired by your endless
energy. Your knowledge is covering the basics and clinical sciences. I always bless our first meeting in Addis. It
grossly changes my carrier and I am now happy to see the results of this choice in life! Thomas, I hope our
collaboration will continue. It would be inappropriate not to mentioning your beloved family. They always surprise
me from the first meeting to this time. They are loving and special. Thank you Maria, Freja, Algot and the lovely
Tyra!
My co-supervisor Olle Stendahl, a man of a big pool of basic and clinical knowledge. I always admire your easy
way of grasping complicated scientific concepts and analysis. You clearly showed me what a true professor is! I
greatly appreciate your critical thinking and the untouchable scope you view things. I will not forget the
encouragement and support you have given me especially in the process to complete the thesis write-up in such
short time. Thank you Olle, for all your scientific and administrative support, without it I would not have managed!
Britt-Inger, I want to extend my appreciation also to you, for your kind personality and hospitality!
My co-supervisor Abraham Aseffa, my appreciation starts more than 15 years back. You are among a handful
Ethiopians who can be perfectly described in every term. Your deep and sharp knowledge, critical thought and
clearness are indescribable. Your kind personality and positive thinking always inspires me. I wish to give you more
honor and working freedom to bring out all your potentials. Thank you Abraham!
Jonna Idh, my Swedish sister “Abelij”. You have a very big place in my PhD journey. We were swimming together
for a couple of years, which enabled me to really get to know you. You are an intelligent, superb and loving sister. I
want to think of many of our good memories and funny moments in a very calm mood after closing this chapter.
Many thanks Jonna for all the unlimited untold support I got from you in all these time. My appreciation to your
family, thank you Nina, for hosting me and the nice time we had in Stockholm! Thank you BG/Maria and
(Rich)Richard- “kirstinaw lene new”.
Daniel Elias, for always believing in me and to our long term true friendship (from the time of “teacher” to “Dany
boy!”). Thank you for bridging me to the excellent Swedish research team. I much appreciated your simple
personality, focused thought and clarity. Share with Hiwot my appreciation!
Sven Britton, for your long way support and intellectual ideas and comments.

83
Mary Esping, for superb administrative service and help with all types of queries, whatever query you receive
wherever from!
My lab and office companions, TB team and other friends in Sweden, Maria Lerm (I admire your strong spirit,
enthusiasm and deep knowledge of TB basics. Thank you for the pulka!), Kalle Magnusson (I like to sit near you
during fika for a relaxing atmosphere you always create), Tommy Sundqvist, I wish to have more time to listen to
your impressive discussions about NO. Danne Eklund/Louise (say hi to the junior in October!), Robert
Blomgran- I like your easy and relaxed personality which give comfort to anyone. Thank you for your help during
optimization of flow assay. Congratulation for the job you did in Brussels), Johanna Raffetseder (you are
extremely helpful and many thanks for the many translations, proof readings and inviting approach, ich danke Ihnen
vielmals!), Clara Braian, You were helpful all the time, thanks for the proof reading and the stroller, which makes
life easy for my kid; Tack somnyket Clara!!), Thommie Karlsson, hope you restore your energy after the
dissertation. I like the few minutes’ talk we usually had in the weekends. My office companions Tony Forslund, for
a very pleasant personality and Maggan Lindroth, it makes me happy hearing your funny talks. You are so relaxing
and create comfort to people surrounding you. Thank you both for the nice working atmosphere we have had in our
office. Elisabet, Lena, Peter (A=A), Elena, Angelika, Henrik, Asia, Anders, Fredrik, Pontus, Magali, Yuhan,
Sadaf, Ramala, Fanny (wish you a fruitful time here and in Peru!). Kajsa Holmgreen, Asa Keita, Olena
Yakymenko. Katarina Tejle and Christina Samuelsson, for excellent technical assistance. The master students
we have supervised over the years Märta, Petra, Sofia, Anna, Caroline and Jonas and all fellow Ph.D students at
Medical Microbiology, old and new.
Peter Söderkvist and Asa from division of Cell Biology; Lars Brudin, Kalmar County Hospital and
Mats Fredrikson, Linkoping for statistical assistance. To Daniel, a friend from Eritrea in Linköping.
To the Gondar research team for your unreserved effort in helping me with my work: Aschalew, Belay Anagaw,
Martha, Tigist Feleke, Lamesgin, Meseret Senbeto, Meseret Atale, Tezera, Abye, Saba, Alemnesh, Kirkim
,Yeshy, Atanaw, Misganaw, Abebech, Bethelhem, Endale Zerfe, Dagne, all staffs in ART and main lab of
Gondar hospital, and the Debark team! All staffs and collegues at the school of Biomedical and Laboratory Sciences
(the seniors-Prof. Moges, Dr. Feleke, Baye, Demekech, Aregawi, and Binyam, Abebe, Meseret,Wubet ,
Gizachew, and to all energetic young academician in the school!) , and to GU/Gondar medical college as a whole,
Drs. Sisay Yifru, Shitaye Alemu, Ermias Diro (Ermy, I appreciate your enthusiasm to research and we should
toast a glass for the nice and fruitful old friendship), Dr. Assefa Getachew for our successful scientific collaboration
and for your unreserved effort in reading all the x-ray films. Oldies but goodies- Buddies: Afework Kassu/Berish,
Gashaw Andarge (“Enkuan maryam marechish Atsedye”!), Dereje Nigusse, Mamo, Kahsay Hiruy, Jemal Ali,
Andargachew Mullu, Belay Tessema, Yeshambel Belihun, Yifokir, for your friendship and all your support! All
JHU staffs in particulalr the Awassa team: Dr Girma Tesfaye,Mekete Reta, Jhony, Edris, Heny. My old true friend
Alex (Cherkos)-wishing a better life over there.
All staffs and students at AHRI/ALERT, especially Martha Zewdie (thank you Marthy for all unreserved support
during optimization of ELISPOT and flow assays, you are really special!), Kidist, Liya, Hiwot, Elena, Markos,

84
Adane, Demis, Alemayehu, Almaz,Wude, Abebe, Menbere, Endalawaw, Haki (you are a superb secretary!),
Jemal, Yonas, Yohannis, Binyam (thank you for handling the finance matters), Drs. Rawligh, Lawrence, Ruth
Likassa and to all other AHRI/ALERT staffs!
A big thank to my amazing life time friends in Ethiopia Zeyede, Amare, Selamina, Yonas, Marthy2,
Endris,Gulilat, Lule, Amha , Etsub and Gecho (US). Kume, a new member but who laid the foundation.
To my entire family, to all your unconditional LOVE and support given to this time, Solome Ebba (my angel who is
the battery of my motor and a source of happiness), my father, Abate Waktola (for all the untold support and
assistance to this end), Helena/Nunish, Derese, Kuyu, Yared, Etaferwa, Kidus, Kidist, KB, Wengel, Barkot,
Gelet, Hailu, Telile, Zewde, Mare, Belayneh, Abiy (“enkuan aterefeh”), Mitaye, and the rest---
Last but certainly not least, thank you Mar/Mesy (what a wonderful wife you are!!!), many thanks for changing my
life and showing the other parts of this world. Thank you Mar for all your patience and unreserved support and
endless Love. I am the luckiest person in this world to have you as a wife!!! I will always Love you!







