
1. Introduction
2. Materials and methods
3. Results
4. Discussion
5. Conclusion
Original Research
Thymus peptides regulate activityof RAW 264.7 macrophage cells:inhibitory analysis and a role ofsignal cascadesSergey M. Lunin†, Olga V. Glushkova, Maxim O. Khrenov,Svetlana B. Parfenyuk, Tatyana V. Novoselova, Evgeny E. Fesenko &Elena G. NovoselovaInstitute of Cell Biophysics of Russian Academy of Sciences, Pushchino, Moscow region, Russia
Objectives: The aim of this study was to reveal T-lymphocyte-independent
mechanisms of thymic peptide-mediated immunomodulation.
Methods: The effects of two thymic peptides---- thymulin and thymopentin were
studied in cultured RAW 264.7 macrophages (lipopolysaccharide-stimulated or
unstimulated) by measuring cytokine production and signal protein levels.
Results: Both peptides increased proinflammatory cytokine secretion by
unstimulated RAW 264.7 macrophages and these effects were blocked by
the NF-kB cascade inhibitor, stress-activated protein kinase (SAPK)/JNK cas-
cade inhibitor and, to a lesser extent, Toll-like 4 receptor activity inhibitor.
In macrophages stimulated by bacterial lipopolysaccharide, peptides alone
did not affect cytokine secretion, but significantly enhanced effects of
each of the inhibitors. Thymopentin increased activation of both NF-kBand SAPK/JNK cascades in unstimulated macrophages, while thymulin
significantly decreased activation of the SAPK/JNK but not NF-kB cascade
in LPS-stimulated macrophages. Thymulin and thymopentin increased pro-
duction of the heat shock protein HSP72 both in LPS-stimulated and
unstimulated cells.
Conclusions: Thymulin and thymopentin are effective anti-inflammatory
modulators with direct actions on innate immune cells; the effects involve
multiple signal cascades, including NF-kB and SAPK/JNK pathways. Since sig-
naling cascades are now considered to be targets for new therapies, thymic
peptides may be prospective modulators of signaling cascades, acting alone
or in combination with other agents.
Keywords: CLI-095, IKK Inhibitor XII, JNK Inhibitor II, SP600125, thymopentin, thymulin
Expert Opin. Ther. Targets (2011) 15(12):1337-1346
1. Introduction
The thymus is considered to be an organ providing T-lymphocyte maturation anddifferentiation. But, along with forming the appropriate microenvironment for thy-mocyte activity, thymic epithelial cells are capable of secreting several peptides intothe circulation. These peptides have been shown to participate in thymocyte differ-entiation and proliferation. However, the extrathymic and systemic functions ofthese peptides are only beginning to be elucidated [1].
Thymulin is a metallopeptide consisting of a nonapeptide (Glu-Ala-Lys-Ser-Gln-Gly-Gly-Ser-Asp), originally named ‘facteur thymique serique’ (FTS), that coupleswith zinc ion. The biological activity of FTS is highly dependent of the presence ofZn2+ [2]. Along with the intrathymic effects on thymocyte maturation, the expression
10.1517/14728222.2011.641952 © 2011 Informa UK, Ltd. ISSN 1472-8222 1337All rights reserved: reproduction in whole or in part not permitted
Exp
ert O
pin.
The
r. T
arge
ts D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Sask
atch
ewan
on
06/2
9/12
For
pers
onal
use
onl
y.

of phenotypic markers, such as CD90, CD3, CD4 andCD8 [3], and extrathymic maturation of T-lymphocytes [4], thy-mulin can also modulate the activity of mature immune cells.Most data indicates that the effects of thymulin on inflamma-tory responses and on cytokine networks are predominantlyinhibitory [5-7].A synthetic pentapeptide, thymopentin (Arg-Lys-Asp-Val-
Tyr), corresponding to residues 32 -- 36 of another thymicpeptide, thymopoietin, reproduces its biological activities [8].Thymopoietin has profound effects on thymocyte differentiationandmaturation. For example, it induces the phenotypic differen-tiation of T precursor cells in vitro and the expression ofCD4 andCD8 cell surfacemarkers on human thymocytes [9]. Now, there isa growing body of data on the immune-balancing role of thymo-poietin or thymopentin in inflammatory diseases. A trendmay beseen toward the immunopotentiating effects of thymopentin inits normal state or upon immunodepressive conditions. Wehave observed previoualy the activation of production of severalcytokines, NO, and heat shock proteins (Hsp70 and Hsp90) inperitoneal macrophages and splenic lymphocytes from healthymice treated with thymopentin injections [10]. In addition, thy-mopentin prevented the lipopolysaccharide (LPS) -inducedphosphorylation of Rel A, an important marker of activation ofthe NF-kB signaling pathway [11].Almost all available data on thymic peptides were either
directly related to T-lymphocytes or were observed in conditionsthat cannot exclude T-lymphocyte effects. Few data are availableon the direct effects of thymic peptides on non-thymus-derivedimmune cells. Thus, Orriger et al. [12] showed no significantincreases in responsiveness to IFN-g in HD-11 macrophage cellscultured with thymulin. Also, thymulin was shown to upregulateNK cell activity, and this effect was considered probably directrather than T-lymphocyte-mediated [13]. Our preliminarydata showed that added in vitro thymulin reduced the peak ofTNF-aproduction inLPS-stimulatedperitonealmacrophages [7].In the present study, we used RAW 264.7 macrophage-like cellsthat seem to be a suitable model to study the T-lymphocyte-independent mechanisms of thymic peptides activity. Theinvestigation of direct effects of thymic peptides on macro-phages will help in discovery of new targets for the therapy ofimmunity-related diseases.Recently, new approaches related to the targeting of intracel-
lular signal transduction have been developed in the therapy ofinflammatory and other disorders. As a result, new inhibitors oragonists of signal pathways have been developed [14-19]. Theseare not only prospective drugs but also useful tools for studyingintracellular machinery. The inhibitors of two signaling path-ways that were shown to be involved in the macrophage cellresponse to bacterial toxins, the NF-kB and stress-activatedprotein kinase (SAPK)/JNK signaling cascades were used inthe experiments. In addition, with regard to the role ofToll-like receptors in the cell response to LPS, inhibitor ofTLR4 signaling was used. So, to reveal the mechanisms of thy-mic peptide activity, the inhibitory analysis was provided usingthree novel inhibitors:
(1) NF-kB cascade inhibitor, named IKK inhibitor XII, acell-permeable compound that acts as a potent ATP-site-targeting inhibitor against kinases IKK-1 and IKK-2 exhib-iting more than 50-fold less potency toward a panel ofmore than 50 other kinases. It has been shown to suppresseffectively the LPS-stimulated secretion of TNF-a, IL-1band IL-6 by human peripheral blood monocytes as wellas TNF-a-induced NF-kB nuclear translocation in humanlung fibroblast cells [14].(2) SAPK/JNK cascade inhibitor SP600125 (also knownas JNK inhibitor II), a potent, cell-permeable, selectiveand reversible inhibitor of JNK-1, JNK-2 and JNK-3.The inhibition is competitive with respect to ATP. Itexhibits over 300-fold greater selectivity for JNK as com-pared with extracellular signal-regulated kinase 1 (ERK1)and p38-2 MAPKs. It inhibits the phosphorylation ofc-Jun and blocks cellular expression of IFN-g , TNF-aand COX-2. It prevents IL-1-induced accumulation ofphospho-Jun and induction of c-Jun transcription [15-17].(3) TLR4 receptor inhibitor CLI-095 (TAK-242), anovel cyclohexene derivative that suppresses TLR4 signal-ing specifically, inhibiting the production of NO andpro-inflammatory cytokines [18]. It acts by blockingthe signaling mediated via the intracellular domain ofTLR4, but not the extracellular domain. It potentlysuppresses both ligand-dependent and -independentsignaling of TLR4 [19].
2. Materials and methods
2.1 Cell culture and reagentsRAW 264.7 cells (mouse leukemic monocyte macrophagecell line), obtained from the American Type Culture Collec-tion (ATCC; catalogue number TIB-71) were cultured inDulbecco’s Modified Eagle’s Medium (DMEM) supplementedwith 10% fetal calf serum (FCS), penicillin G (100 units/ml),gentamycin (100 µg/ml), and streptomycin (100 µg/ml) at37�C in a 5% CO2 atmosphere. Cells were seeded at a conflu-ence of approximately 10% (1 � 106 cells in 100-mm plates)and grown to a confluence of approximately 80%. This proce-dure required the cells to be split every 2 days. In each of fourindependent experiments, the cells from a separate passage(over the range from the third to seventh passages) were pooled,and the cell’s aliquots were treated separately with 1 µg/ml CLI-095 (TLR4 inhibitor, InvivoGen, USA), or 2.2 µg/mlSP600125 (JNK-1, -2, -3 inhibitor, Stressgen, USA), or 1 µg/mlIKK Inhibitor XII (inhibitor of IKK-a, -b and -"-activity,Merck, Germany). In the presence of inhibitors, the cellswere cultured for 1 h (SP600125, IKK Inhibitor XII) or for15 min (CLI-095). Then 5 ng thymopentin/ml (American Pep-tides, USA) or 5 ng thymulin/ml (American Peptides, USA)were added, and cells were cultured for a further 30 min (thebiological activity of thymulin is dependent on Zn2+ ions; sobefore use, serum thymic factor was supplemented with anequimolar quantity of ZnCl2). Finally, the lipolysaccharide
Thymus peptides regulate activity of macrophage cells
1338 Expert Opin. Ther. Targets (2011) 15(12)
Exp
ert O
pin.
The
r. T
arge
ts D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Sask
atch
ewan
on
06/2
9/12
For
pers
onal
use
onl
y.

from E. coli (LPS, 2.5 µg/ml, Merck, Germany) was applied,and cells were cultivated for 24 h at 37�C in a 5% CO2
atmosphere. All measurements of cell viability were conductedusing light microscopy and trypan blue staining [20].
Within each separate experiment characteristics of sampleswere concurrently measured in six repetitions to get averagemeans. The averaged values from four experiments were proc-essed to determine the significance of differences betweengroups (n = 4).
Control samples were the cells untreated with LPS, or withinhibitors, or with both of them. Considering that all ofinhibitors were dissolved in DMSO, vehicle control sampleswere incubated for corresponding time intervals with corre-sponding quantity of DMSO. Final DMSO concentrationdid not exceeded 0.1%. In the experiments with LPS- orthymulin/thymopentin- treated cells, PBS additions servedas vehicle controls.
2.2 ELISARAW 264.7 cells were suspended in RPMI 1640 (Sigma,USA) supplemented with gentamycin, Hepes, and 10%FCS, and cultured in 24-well plates with LPS and with/without the inhibitors for 24 h at 37�C in 5% CO2. Lysatesof the cells were obtained by threefold freezing--refreezing.The concentration of cytokines in the cell samples was deter-mined by ELISA. ELISA Development Kits for mouseTNF-a, mouse IL-1, mouse IL-6, mouse IL-10 and mouseIFN-g (the kits were purchased from Peprotech, USA)were used. In order to visualize binding, 100 µl of 2,2¢-azinobis(3-ethylbenzthiazoline-6-sulfonate) (ABTS) greendye (Sigma, USA) dissolved in 0.05 M citrate buffer (pH4.0) with 0.01% H2O2 was applied. The optical densitywas measured at 405 nm with the plate spectrophotometer(Multiscan EX, Thermo Electron Corporation).
2.3 Western blot analysisTo prepare specimens, 5 � 107 cells were lysed in an ultrasonicdisintegrator with constant stirring for 2 min. The concentrationof the total cellular protein was determined by the BradfordmethodwithBradford solution (Sigma,USA). Proteins were pre-cipitated in acetone, solubilized, boiled for 5 min, and storedat -70�C. Proteinswere determined by 10%PAGE.The specific-ity of the analysis was tested by immunoblotting, as describedpreviously [21]. The proteins were transferred from the gel ontoa nitrocellulose membrane in a transblot chamber. Upon block-ing, the membrane was exposed for 2 h to antibodies to themouse proteins: Hsp70 (rabbit anti-mouse HSP 72, cloneSPA-812, StressGen Biotechnologies, inducible form), phos-pho-NF-kB antibody (Phospho-NF-kB p65 (Ser 536, # 3031,Cell Signaling Technology, USA), rabbit phospho-SAPK/JNKantibody to synthetic phospho-peptide SAPK/JNK, (Cell Signal-ing Technology, USA), or phospho-IKKa/b antibody II (Ser176/180) (Cell Signaling Technology, USA). After washing,the nitrocellulose membranes were incubated for 1 h with theanti-rabbit biotinylated antibody (Jackson ImmunoResearch,
USA), and peroxidase-conjugated streptavidin (IMTEK, Russia)was added for 1 h. The loading control was mouse monoclonalanti-human beta tubulin (USBiological,USA). ECLplus chemi-luminescent cocktail (Amersham/GE Healthcare, UK) was usedto develop the immunostaining of blots following the manufac-turer’s instructions, exposing the blot to Kodak film. A positionof proper bandswas determined using amolecular weightmarker(Spectra Multicolor Broad Range Protein Ladder, Fermentas,USA) during the electrophoresis stage and by the specific bindingto antibodies after blotting. Quantitative evaluation of proteinbands was performed using theQapa software. Obtained numer-ical data were normalized to corresponding loading control (thetubulin band) and expressed in relative units as the mean of threeindependent measurements.
2.4 Statistical analysisStatistical analysis was performed using Student’s t-test,one-way ANOVA and Tukey’s post-hoc tests.
3. Results
In experiments with addition of inhibitors that were originallydissolved in DMSO, final DMSO concentration in experimen-tal samples and in corresponding vehicle controls was 0.1%, andanalysis of variance (one-way ANOVA and Tukey’s post-hoctests) did not reveal significant differences between PBS- andDMSO-vehicles in terms of cytokine production as well asinterpassage variations (F(5,18) = 0.185, p = 0.966)
3.1 Effects of thymulin and thymopentin on cytokine
production in unstimulated macrophagesIn order to analyze the effects of thymic peptides on cytokineproduction in un-stimulated RAW 264.7 cells, amounts ofTNF-a, IL-1a, IL-6 and IFN-g were measured in cellstreated with thymopentin or thymulin, as indicated inFigure 1A and B, respectively. The group of bars in Figure 1A
labeled as “thymopentin (tp)” indicates that treatment withthymopentin resulted in a significant increase of TNF-a andIL-6 production, but not IL-1a and IFN-g productionwhen compared to the untreated control. Culturing theRAW 264.7 cells with thymulin produced a similar stimula-tion effect, except that the thymulin slightly increased IL-1asecretion (Figure 1B, bars labeled as “thymulin (tm)”).
3.2 Effects of thymic peptides on cells pretreated
with inhibitorsTo reveal the role of NF-kB, SAPK/JNK, and TLR4 signalingin the stimulating effects of peptides, the levels of the cyto-kines TNF-a, IL-1a, IL-6 and IFN-g were measured insupernatants of macrophages pretreated with each of theinhibitors (IKK Inhibitor XII, SP600125, or CLI-095) andthen cultured with thymopentin or thymulin.
The results in Figure 1A indicate that the stimulating effect ofthymopentin on the production of all studied cytokines wascompletely blocked by either the NF-kB pathway inhibitor
Lunin, Glushkova, Khrenov, Parfenyuk, Novoselova, Fesenko & Novoselova
Expert Opin. Ther. Targets (2011) 15(12) 1339
Exp
ert O
pin.
The
r. T
arge
ts D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Sask
atch
ewan
on
06/2
9/12
For
pers
onal
use
onl
y.

(IKK Inhibitor XII) or SAPK/JNK inhibitor (SP600125). Thethymopentin-induced increase in the secretion of TNF-a andIL-1 was unaffected by the inhibitor of TLR4 signaling(CLI-095), but this inhibitor blocked the thymopentin-induced increase of IL-6 production.The results in Figure 1B indicate that the stimulating effect of
thymulin on cytokine production was also completely blockedby either IKK Inhibitor XII or JNK Inhibitor SP600125. Theinhibitor of TLR4 signaling, CLI-095, blocked the thymulin-induced increase in IL-1a and IL-6 production, but notTNF-a production.It should be noted that IKK Inhibitor XII or JNK Inhibitor
SP600125, added alone into culture media (without thymicpeptides), suppressed production of TNF-a below controlvalues (Figure 2A), but thymulin normalized the levels ofTNF-a in cells treated with the JNK inhibitor SP600125(Figure 1B) (the results are statistically significant, p < 0.05).
3.3 Effects of thymulin and thymopentin on
LPS-stimulated macrophages cultured with or
without inhibitorsInitially, to evaluate the effects of signal cascade inhibitorson stimulated macrophages, secretions of two cytokines, IL-1aand TNF-a, were measured in the presence of LPS from Escher-ichia coliwith or without addition of the signal cascade inhibitorsto the RAW 264.7 cells. Endotoxin induced the expectedincrease of IL-1a and TNF-a production, and RAW 264.7 cellsdemonstrated a significant response to LPS, showing a twofold tothreefold increase in measured cytokine production as comparedwith the unstimulated cells (Figure 2A and B). When LPS wascombined with IKK Inhibitor XII, JNK inhibitor SP600125,or inhibitor of Toll-like receptor signaling CLI-095, thesecretion of cytokines was significantly depressed comparedwith LPS-stimulated cells, but remaining somewhat raisedrelative to the control. The only exception was the effect of
0.4
0.3
ng
ml-1
ng
ml-1
0.2
0.1
0
0.6
0.5
0.4
0.3
0.2
0.1
0Control
Control
*
*
*
‡
‡‡ ‡
‡
‡‡
‡
‡
‡
‡ ‡‡
‡
‡
Thymopentin (tp) tp+CLI-095 tp+SP600125 tp+IKK Inhibitor XII
0.5
Thymulin (tm) tm+CLI-095 tm+SP600125 tm+IKK Inhibitor XII
TNF-alpha
IL1-alpha
IFN-gamma
IL6
TNF-alpha
IL1-alpha
IFN-gamma
IL6
A.
B.
*
*
*
*
Figure 1. The effect of thymopentin (tp) (A) and thymulin (tm) (B) alone and in combinations with inhibitor of TLR4 signaling
(CLI-095), SAPK/JNK pathway inhibitor (SP600125), and NF-kB pathway inhibitor (IKK Inhibitor XII) on production of cytokines
by RAW 264.7 cells. Measurements were made by ELISA. The values are expressed as means (ng/ml) ± SE from four independent
experiments performed separately with four different passages (passages from the fourth to seventh); within each separate
experiment characteristics of samples were concurrently measured in six repetitions to get average means. The averaged
values from four experiments were processed to determine the significance of differences between groups (n = 4). The cells
cultured with corresponding amounts of DMSO vehicle (instead of inhibitors) and PBS vehicle (instead of thymulin/
thymopentin) for corresponding time intervals served as controls.*Significantly more than the control, p < 0.05.zSignificantly less than the control, p < 0.05.
Thymus peptides regulate activity of macrophage cells
1340 Expert Opin. Ther. Targets (2011) 15(12)
Exp
ert O
pin.
The
r. T
arge
ts D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Sask
atch
ewan
on
06/2
9/12
For
pers
onal
use
onl
y.
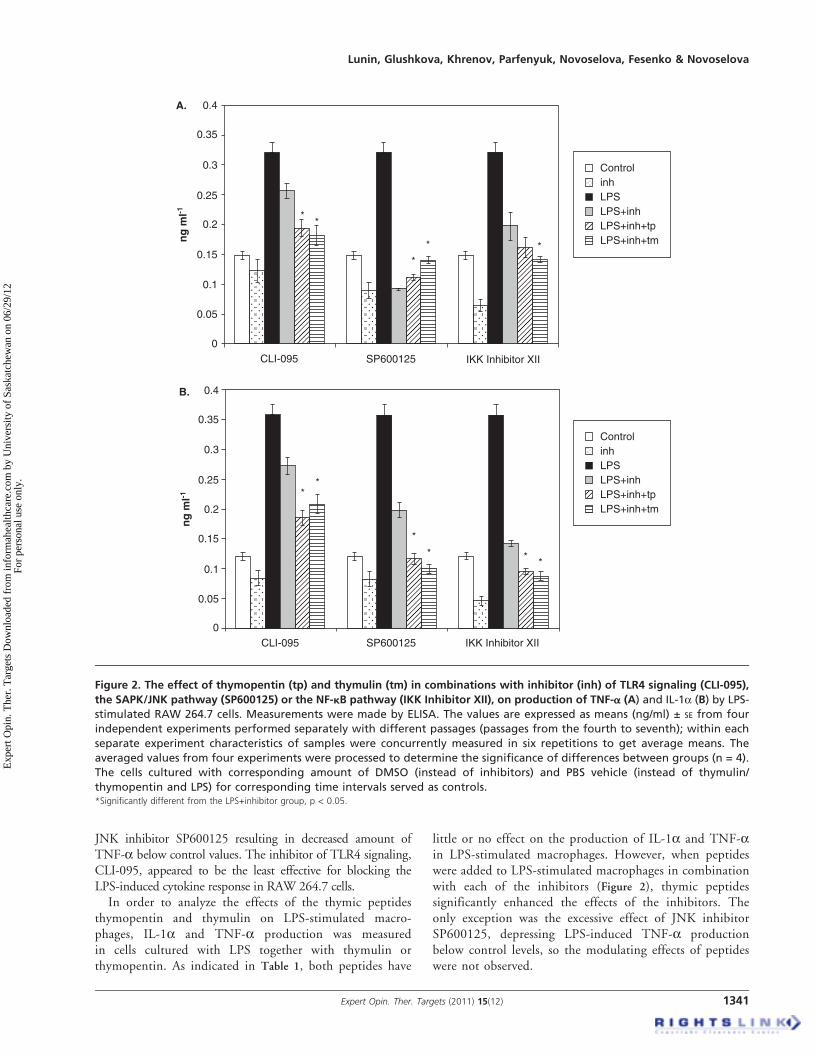
JNK inhibitor SP600125 resulting in decreased amount ofTNF-a below control values. The inhibitor of TLR4 signaling,CLI-095, appeared to be the least effective for blocking theLPS-induced cytokine response in RAW 264.7 cells.
In order to analyze the effects of the thymic peptidesthymopentin and thymulin on LPS-stimulated macro-phages, IL-1a and TNF-a production was measuredin cells cultured with LPS together with thymulin orthymopentin. As indicated in Table 1, both peptides have
little or no effect on the production of IL-1a and TNF-ain LPS-stimulated macrophages. However, when peptideswere added to LPS-stimulated macrophages in combinationwith each of the inhibitors (Figure 2), thymic peptidessignificantly enhanced the effects of the inhibitors. Theonly exception was the excessive effect of JNK inhibitorSP600125, depressing LPS-induced TNF-a productionbelow control levels, so the modulating effects of peptideswere not observed.
0.4
0.35
0.3
0.25
0.2
0.15
0.1
0.05
ng
ml-1
0
0.4
0.35
0.3
0.25
0.2
0.15
0.1
0.05
ng
ml-1
0
CLI-095
A.
B.
SP600125 IKK Inhibitor XII
CLI-095 SP600125 IKK Inhibitor XII
**
*
* *
**
*
* * *
ControlinhLPSLPS+inhLPS+inh+tpLPS+inh+tm
ControlinhLPSLPS+inhLPS+inh+tpLPS+inh+tm
Figure 2. The effect of thymopentin (tp) and thymulin (tm) in combinations with inhibitor (inh) of TLR4 signaling (CLI-095),
the SAPK/JNK pathway (SP600125) or the NF-kB pathway (IKK Inhibitor XII), on production of TNF-a (A) and IL-1a (B) by LPS-
stimulated RAW 264.7 cells. Measurements were made by ELISA. The values are expressed as means (ng/ml) ± SE from four
independent experiments performed separately with different passages (passages from the fourth to seventh); within each
separate experiment characteristics of samples were concurrently measured in six repetitions to get average means. The
averaged values from four experiments were processed to determine the significance of differences between groups (n = 4).
The cells cultured with corresponding amount of DMSO (instead of inhibitors) and PBS vehicle (instead of thymulin/
thymopentin and LPS) for corresponding time intervals served as controls.*Significantly different from the LPS+inhibitor group, p < 0.05.
Lunin, Glushkova, Khrenov, Parfenyuk, Novoselova, Fesenko & Novoselova
Expert Opin. Ther. Targets (2011) 15(12) 1341
Exp
ert O
pin.
The
r. T
arge
ts D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Sask
atch
ewan
on
06/2
9/12
For
pers
onal
use
onl
y.
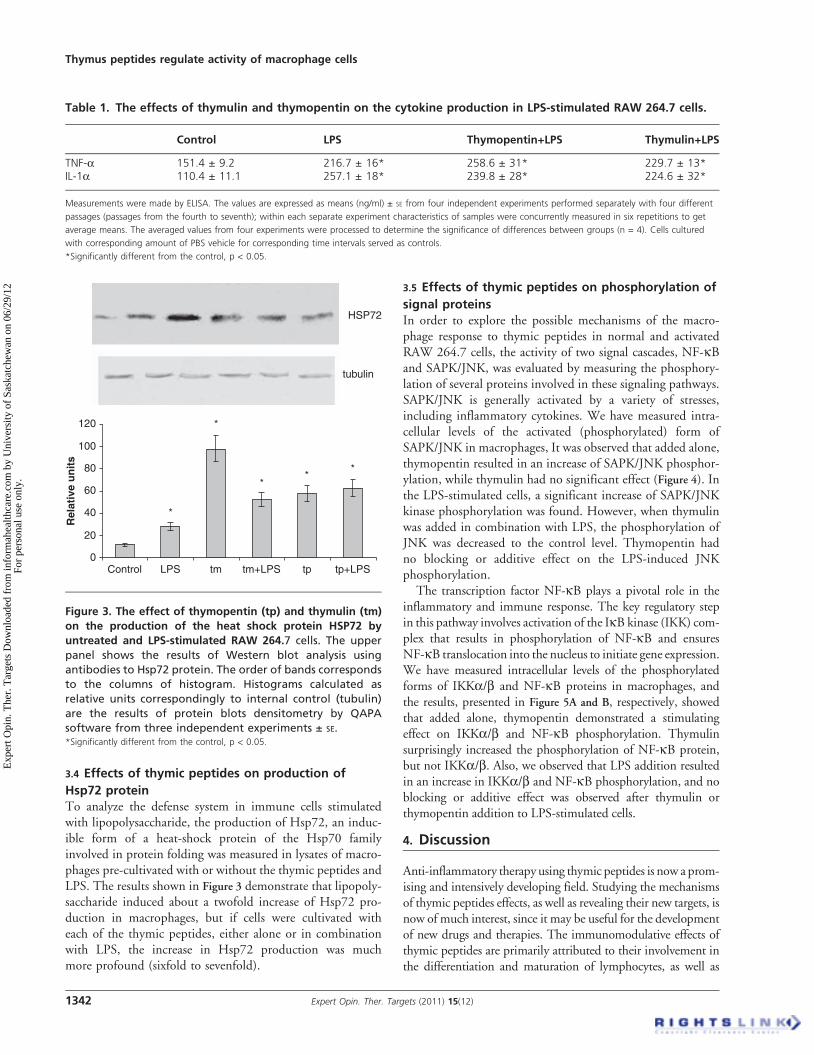
3.4 Effects of thymic peptides on production of
Hsp72 proteinTo analyze the defense system in immune cells stimulatedwith lipopolysaccharide, the production of Hsp72, an induc-ible form of a heat-shock protein of the Hsp70 familyinvolved in protein folding was measured in lysates of macro-phages pre-cultivated with or without the thymic peptides andLPS. The results shown in Figure 3 demonstrate that lipopoly-saccharide induced about a twofold increase of Hsp72 pro-duction in macrophages, but if cells were cultivated witheach of the thymic peptides, either alone or in combinationwith LPS, the increase in Hsp72 production was muchmore profound (sixfold to sevenfold).
3.5 Effects of thymic peptides on phosphorylation of
signal proteinsIn order to explore the possible mechanisms of the macro-phage response to thymic peptides in normal and activatedRAW 264.7 cells, the activity of two signal cascades, NF-kBand SAPK/JNK, was evaluated by measuring the phosphory-lation of several proteins involved in these signaling pathways.SAPK/JNK is generally activated by a variety of stresses,including inflammatory cytokines. We have measured intra-cellular levels of the activated (phosphorylated) form ofSAPK/JNK in macrophages, It was observed that added alone,thymopentin resulted in an increase of SAPK/JNK phosphor-ylation, while thymulin had no significant effect (Figure 4). Inthe LPS-stimulated cells, a significant increase of SAPK/JNKkinase phosphorylation was found. However, when thymulinwas added in combination with LPS, the phosphorylation ofJNK was decreased to the control level. Thymopentin hadno blocking or additive effect on the LPS-induced JNKphosphorylation.
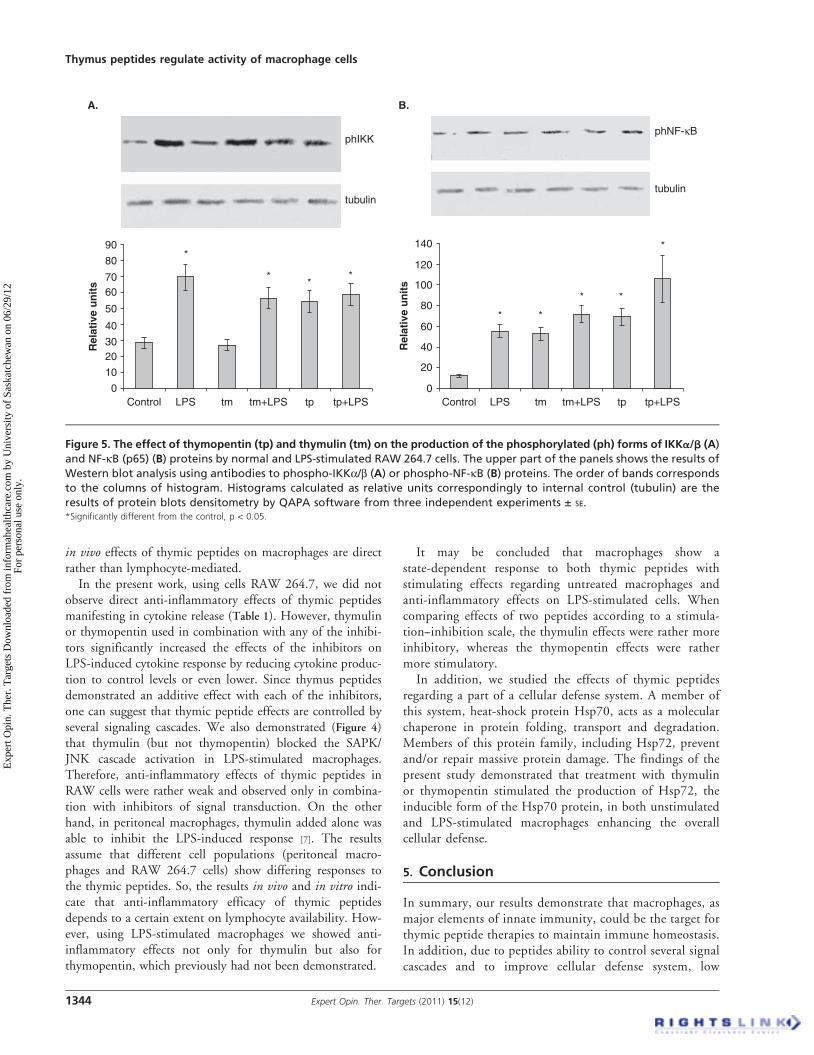
The transcription factor NF-kB plays a pivotal role in theinflammatory and immune response. The key regulatory stepin this pathway involves activation of the IkB kinase (IKK) com-plex that results in phosphorylation of NF-kB and ensuresNF-kB translocation into the nucleus to initiate gene expression.We have measured intracellular levels of the phosphorylatedforms of IKKa/b and NF-kB proteins in macrophages, andthe results, presented in Figure 5A and B, respectively, showedthat added alone, thymopentin demonstrated a stimulatingeffect on IKKa/b and NF-kB phosphorylation. Thymulinsurprisingly increased the phosphorylation of NF-kB protein,but not IKKa/b. Also, we observed that LPS addition resultedin an increase in IKKa/b and NF-kB phosphorylation, and noblocking or additive effect was observed after thymulin orthymopentin addition to LPS-stimulated cells.
4. Discussion
Anti-inflammatory therapy using thymic peptides is now a prom-ising and intensively developing field. Studying the mechanismsof thymic peptides effects, as well as revealing their new targets, isnow of much interest, since it may be useful for the developmentof new drugs and therapies. The immunomodulative effects ofthymic peptides are primarily attributed to their involvement inthe differentiation and maturation of lymphocytes, as well as
Table 1. The effects of thymulin and thymopentin on the cytokine production in LPS-stimulated RAW 264.7 cells.
Control LPS Thymopentin+LPS Thymulin+LPS
TNF-a 151.4 ± 9.2 216.7 ± 16* 258.6 ± 31* 229.7 ± 13*IL-1a 110.4 ± 11.1 257.1 ± 18* 239.8 ± 28* 224.6 ± 32*
Measurements were made by ELISA. The values are expressed as means (ng/ml) ± SE from four independent experiments performed separately with four different
passages (passages from the fourth to seventh); within each separate experiment characteristics of samples were concurrently measured in six repetitions to get
average means. The averaged values from four experiments were processed to determine the significance of differences between groups (n = 4). Cells cultured
with corresponding amount of PBS vehicle for corresponding time intervals served as controls.
*Significantly different from the control, p < 0.05.
120
100
80
60
40
20
0Control LPS
*
*
**
*
tm tm+LPS tp tp+LPS
tubulin
HSP72
Rel
ativ
e u
nit
s
Figure 3. The effect of thymopentin (tp) and thymulin (tm)
on the production of the heat shock protein HSP72 by
untreated and LPS-stimulated RAW 264.7 cells. The upper
panel shows the results of Western blot analysis using
antibodies to Hsp72 protein. The order of bands corresponds
to the columns of histogram. Histograms calculated as
relative units correspondingly to internal control (tubulin)
are the results of protein blots densitometry by QAPA
software from three independent experiments ± SE.*Significantly different from the control, p < 0.05.
Thymus peptides regulate activity of macrophage cells
1342 Expert Opin. Ther. Targets (2011) 15(12)
Exp
ert O
pin.
The
r. T
arge
ts D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Sask
atch
ewan
on
06/2
9/12
For
pers
onal
use
onl
y.

modulation of mature peripheral T-lymphocytes [1]. The cellularmechanisms of these effects and receptors for thymic peptides arenot identified, although it has been shown that T-lymphocytemembranes have specific receptors that bind thymulin withhigh affinity [1]. The present work aimed to study the effects ofthymic peptides on the activity of RAW 264.7 cells in vitro thatwould allow finding of new targets for thymic peptides and clarifythe role of the peptides in the innate immunity control.
The findings of the present work demonstrated thatthe thymic peptides thymulin and thymopentin directlyaffect macrophage cells. Indeed, since experiments wereperformed using macrophage lineage cells, these effects areobviously lymphocyte-independent. It was shown that thepeptides increased the activity of unstimulated macrophagesas was visualized by an enhancement of production of thepro-inflammatory cytokines TNF-a, IL-1a and IL-6.
One of the natural activators of macrophages is LPS fromGram-negative bacteria. In particular, it induces the cell cyto-kine response and leads to activation of several signaling path-ways, particularly the NF-kB and SAPK/JNK cascades,operating through the TLR4, member of the Toll-like receptorprotein family. It has been reported that LPS interacts with
LPS-binding protein and CD14, which presents LPS to Toll-like receptor 4, and so activates inflammatory gene expressionthrough NF-kB signaling [22]. The NF-kB pathway is impor-tant for the expression of genes that are involved in the controlof the host inflammatory response. In unstimulated cells,NF-kB proteins exist as hetero- or homodimers in an inactiveform, complexed with an inhibitory protein from the IkBfamily, thereby preventing the translocation of NF-kB to thenucleus. NF-kB activation is achieved via phosphorylationand degradation of the inhibitory IkB protein through theaction of specific kinases, named IkB kinases (IKKs). Thephosphorylation of NF-kB leads to its nuclear translocationand binding to enhancers or promoters of target genes.
SAPK/JNK is potently and preferentially activated by avariety of environmental stresses. SAPK/JNK, being active asa dimer, can translocate to the nucleus. It has recently beenreported that SAPK/JNK is closely interrelated with NF-kBsignaling pathways [23,24].
The present study demonstrates that the stimulatory effectsof thymic peptides on cytokine production may at least bemediated partly by NF-kB and SAPK/JNK pathway activa-tion. Thymopentin induced the phosphorylation of theIKKa/b kinase and the NF-kB protein, as well as SAPK/JNK kinase, which would lead to pro-inflammatory geneexpression. The thymulin effect was similar in magnitude,except that its effect on IKKa/b kinase phosphorylation wasmuch less pronounced, indicating possible activation ofsome alternative pathway of NF-kB phosphorylation.
The results of the present study suggest that LPS-stimulatedRAW 264.7 cells show a potent cytokine response, accompaniedby NF-kB and SAPK/JNK cascade activation. This effect wasreduced by the novel signal cascade inhibitors IKK InhibitorXII (NF-kB cascade inhibitor), SP600125 (SAPK/JNK cascadeinhibitor), and CLI-095 (TLR4 signaling inhibitor), and theinhibitor CLI-095 demonstrated the least efficacy of the three.It should be noted that the inhibitors used in almost all casesdid not decrease LPS-induced cytokine response to exactlycontrol levels.
In thymulin- or thymopentin-stimulated macrophagesNF-kB or SAPK/JNK cascade inhibitors completely blockedthe cytokine response. The TLR4 signaling inhibitor showedminor activity; nevertheless it blocked the production of severalcytokines. Therefore, thymopentin and thymulin stimulationwas rather mild compared with the stressful LPS stimulation.
Anti-inflammatory effects of thymulin have been shownpreviously by several authors. For example, Dardenne andco-workers, using an endotoxin-induced inflammatory ratmodel, showed that thymulin reduced the inflammatorypain and the levels of cytokines in blood plasma [6].
Also we have shown earlier that thymulin injected in vivo oradded in vitro to peritoneal macrophages increased of macro-phagal TNF-a production in un-stimulated cells, and blockedLPS-induced TNF-a production [7]. The findings of presentstudy observed in RAW 264.7 cells closely correspond withabove mentioned data. One may conclude that at least partly
250
200
150
Rel
ativ
e u
nit
s
100
50
0Control LPS
*
**
tm tm+LPS tp tp+LPS
JNK
tubulin
‡
Figure 4. The effect of thymopentin (tp) and thymulin (tm)
on the production of the phosphorylated (ph) form of
SAPK/JNK prote in by the untreated and LPS-
stimulated RAW 264.7 cells. The upper panel shows the
results of Western blot analysis using antibodies to phospho-
SAPK/JNK protein. The order of bands corresponds to the
columns of histogram. Histograms calculated as relative
units correspondingly to internal control (tubulin) are the
results of protein blots densitometry by QAPA software from
three independent experiments ± SE.*Significantly different from the control, p < 0.05.zSignificantly different from the LPS group, p < 0.05.
Lunin, Glushkova, Khrenov, Parfenyuk, Novoselova, Fesenko & Novoselova
Expert Opin. Ther. Targets (2011) 15(12) 1343
Exp
ert O
pin.
The
r. T
arge
ts D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Sask
atch
ewan
on
06/2
9/12
For
pers
onal
use
onl
y.
in vivo effects of thymic peptides on macrophages are directrather than lymphocyte-mediated.In the present work, using cells RAW 264.7, we did not
observe direct anti-inflammatory effects of thymic peptidesmanifesting in cytokine release (Table 1). However, thymulinor thymopentin used in combination with any of the inhibi-tors significantly increased the effects of the inhibitors onLPS-induced cytokine response by reducing cytokine produc-tion to control levels or even lower. Since thymus peptidesdemonstrated an additive effect with each of the inhibitors,one can suggest that thymic peptide effects are controlled byseveral signaling cascades. We also demonstrated (Figure 4)that thymulin (but not thymopentin) blocked the SAPK/JNK cascade activation in LPS-stimulated macrophages.Therefore, anti-inflammatory effects of thymic peptides inRAW cells were rather weak and observed only in combina-tion with inhibitors of signal transduction. On the otherhand, in peritoneal macrophages, thymulin added alone wasable to inhibit the LPS-induced response [7]. The resultsassume that different cell populations (peritoneal macro-phages and RAW 264.7 cells) show differing responses tothe thymic peptides. So, the results in vivo and in vitro indi-cate that anti-inflammatory efficacy of thymic peptidesdepends to a certain extent on lymphocyte availability. How-ever, using LPS-stimulated macrophages we showed anti-inflammatory effects not only for thymulin but also forthymopentin, which previously had not been demonstrated.
It may be concluded that macrophages show astate-dependent response to both thymic peptides withstimulating effects regarding untreated macrophages andanti-inflammatory effects on LPS-stimulated cells. Whencomparing effects of two peptides according to a stimula-tion--inhibition scale, the thymulin effects were rather moreinhibitory, whereas the thymopentin effects were rathermore stimulatory.
In addition, we studied the effects of thymic peptidesregarding a part of a cellular defense system. A member ofthis system, heat-shock protein Hsp70, acts as a molecularchaperone in protein folding, transport and degradation.Members of this protein family, including Hsp72, preventand/or repair massive protein damage. The findings of thepresent study demonstrated that treatment with thymulinor thymopentin stimulated the production of Hsp72, theinducible form of the Hsp70 protein, in both unstimulatedand LPS-stimulated macrophages enhancing the overallcellular defense.
5. Conclusion
In summary, our results demonstrate that macrophages, asmajor elements of innate immunity, could be the target forthymic peptide therapies to maintain immune homeostasis.In addition, due to peptides ability to control several signalcascades and to improve cellular defense system, low
90 140
120
100
80
60
40
20
0
phIKK
tubulintubulin
phNF-κB
80
7060
50
40
3020
10
0Control LPS
*
* *
* *
*
**
*
Rel
ativ
e u
nit
s
Rel
ativ
e u
nit
stm tm+LPS tp tp+LPS Control LPS tm tm+LPS tp tp+LPS
A. B.
Figure 5. The effect of thymopentin (tp) and thymulin (tm) on the production of the phosphorylated (ph) forms of IKKa/b (A)
and NF-kB (p65) (B) proteins by normal and LPS-stimulated RAW 264.7 cells. The upper part of the panels shows the results of
Western blot analysis using antibodies to phospho-IKKa/b (A) or phospho-NF-kB (B) proteins. The order of bands corresponds
to the columns of histogram. Histograms calculated as relative units correspondingly to internal control (tubulin) are the
results of protein blots densitometry by QAPA software from three independent experiments ± SE.*Significantly different from the control, p < 0.05.
Thymus peptides regulate activity of macrophage cells
1344 Expert Opin. Ther. Targets (2011) 15(12)
Exp
ert O
pin.
The
r. T
arge
ts D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Sask
atch
ewan
on
06/2
9/12
For
pers
onal
use
onl
y.

toxicity thymic peptides may be useful as main or auxiliarytherapeutic agents in combination with other drugs fortreatment of inflammatory disorders.
Acknowledgements
The authors thank Bluepencil Science for help withbasic proofreading.
Declaration of interest
The work was supported by the President’s Foundation forLeading Scientific Schools, project 3202.2010.4; by the Rus-sian Foundation for Fundamental Research, projects No11-04-00023-a and No 10-04-00351-a; and by the Programof Russian Academy of Sciences ‘Fundamental Sciencesfor Medicine’.
BibliographyPapers of special note have been highlighted as
either of interest (�) or of considerable interest(��) to readers.
1. Lunin SM, Novoselova EG. Thymus
hormones as prospective
anti-inflammatory agents. Expert Opin
Ther Targets 2010;14:775-86.. A review of a role of thymic peptides
for the regulation of
inflammatory processes.
2. Dardenne M, Pleau JM,
Nabarra B, et al. Contribution
of zinc and other metals to the
biological activity of the serum thymic
factor. Proc Natl Acad Sci USA
1982;79:5370-3
3. Bene MC, Faure G, Bordigoni P, et al.
In vitro induction of monoclonal
antibody-defined T cell markers in
lymphocytes from immunodeficient
children by synthetic serum thymic factor
(FTS). Clin Exp Immunol
1982;48:423-8
4. Inagaki-Ohara K, Kobayashi N,
Nishimura H, et al. Effects of a
nonapeptide thymic hormone on
intestinal intraepithelial lymphocytes in
mice following administration of
5-fluorouracil. Cell Immunol
1996;171:30-40
5. Safieh-Garabedian B, Dardenne M,
Pleau JM, Saade NE. Potent analgesic
and anti-inflammatory actions of a novel
thymulin-related peptide in the rat.
Br J Pharmacol 2002;136:947-55
6. Dardenne M, Saade N,
Safieh-Garabedian B. Role of thymulin
or its analogue as a new analgesic
molecule. Ann NY Acad Sci
2006;1088:153-63.. This paper presents data on
anti-inflammatory effects of thymulin.
7. Lunin SM, Khrenov MO,
Novoselova TV, et al. Thymulin, a
thymic peptide, prevents the
overproduction of pro-inflammatory
cytokines and heat shock protein
Hsp70 in inflammation-bearing mice.
Immunol Invest 2008;37:858-70.. Our data on effects of thymulin in
vivo and in vitro, in
normal macrophages.
8. Goldstein G, Scheid MP, Boyse EA,
et al. A synthetic pentapeptide with
biological activity characteristic of the
thymic hormone thymopoietin. Science
1979;204:1309-10
9. Audhya T, Scheidt MP, Goldstein G.
Contrasting biological activities of
thymopoietin and splenin, two closely
related polypeptide products of thymus
and spleen. Proc Natl Acad Sci USA
1984;81:2847-9
10. Novoselova EG, Lunin SM,
Khrenov MO, et al. Effect of
thymopentin on production of cytokines,
heat shock proteins, and NF-kappaB
signaling proteins. Izv Akad Nauk
Ser Biol 2008;4:422-8
11. Novoselova EG, Lunin SM,
Khrenov MO, et al. Involvement of
NF-kappaB transcription factor in the
antiinflammatory activity of thymic
peptides. Dokl Biol Sci 2009;428:484-6
12. Orringer DA, Staeheli P, Marsh JA. The
effects of thymulin on macrophage
responsiveness to interferon-gamma.
Dev Comp Immunol 2002;26:95-102
13. Merlino PG, Marsh JA. Evidence for the
direct action of thymulin on avian NK
cells. Dev Comp Immunol
2001;25:337-43
14. Christopher JA, Avitabile BG,
Bamborough P, et al. The
discovery of 2-amino-
3,5-diarylbenzamide inhibitors of
IKK-alpha and IKK-beta kinases.
Bioorg Med Chem Lett 2007;17:3972-7
15. Shin M, Yan C, Boyd D. An inhibitor of
c-jun aminoterminal kinase (SP600125)
represses c-Jun activation, DNA-binding
and PMA-inducible 92-kDa type IV
collagenase expression.
Biochim Biophys Acta 2002;1589:311-16
16. Bennett BL, Sasaki DT, Murray BW,
et al. SP600125, an anthrapyrazolone
inhibitor of Jun N-terminal kinase.
Proc Natl Acad Sci USA
2001;98:13681-6
17. Han Z, Boyle DL, Chang L, et al. c-Jun
N-terminal kinase is required for
metalloproteinase expression and joint
destruction in inflammatory arthritis.
J Clin Invest 2001;108:73-81
18. Li M., Matsunaga N,
Hazeki K, et al. A novel cyclohexene
derivative, ethyl (6R)-6-[N-(2-Chloro-4-
fluorophenyl)sulfamoyl]cyclohex-1-ene-1-
carboxylate (TAK-242), selectively
inhibits Toll-like receptor 4-mediated
cytokine production through suppression
of intracellular signaling. Mol Pharmacol
2006;69:1288-95
19. Kawamoto T, Li M, Kitazaki T, et al.
TAK-242 selectively suppresses Toll-like
receptor 4-signaling mediated by the
intracellular domain. Eur J Pharmacol
2008;584:40-8
20. Strober W. Trypan Blue
Exclusion Test of Cell Viability.
Current Protocols in Immunology
2001;A.3B.1-A.3B.2
21. Towbin H, Staeheln T,
Gordon J. Electrophoretic
transfer of proteins from
polyacrylamide gels to
nitrocellulose sheets: procedure and
some applications. Proc Natl Acad
Sci USA 1979;7:4350-4
22. Bochkov VN, Kadl A, Huber J, et al.
Protective role of phospholipids
oxidation products in endotoxin-induced
tissue damage. Nature 2002;419:77-81
23. Papa S, Zazzeroni F, Pham CG, et al.
Linking JNK signaling to NF-kappaB:
a key to survival. J Cell Sci
2004;117:5197-208
Lunin, Glushkova, Khrenov, Parfenyuk, Novoselova, Fesenko & Novoselova
Expert Opin. Ther. Targets (2011) 15(12) 1345
Exp
ert O
pin.
The
r. T
arge
ts D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Sask
atch
ewan
on
06/2
9/12
For
pers
onal
use
onl
y.

24. Wullaert A, Heynick K, Beyaert R.
Mechanisms of crosstalk between
TNF-induced NF-kappaB and JNK
activation in hepatocytes.
Biochem Pharmacol 2006;72:1090-101
AffiliationSergey M. Lunin†, Olga V. Glushkova,
Maxim O. Khrenov, Svetlana B. Parfenyuk,
Tatyana V. Novoselova, Evgeny E. Fesenko &
Elena G. Novoselova†Author for correspondence
Institute of Cell Biophysics of
Russian Academy of Sciences,
142290, Pushchino,
Moscow Region, Institutskaya Street,
Russia Fax: +7 0967 33 05 89;
E-mail: [email protected]
Thymus peptides regulate activity of macrophage cells
1346 Expert Opin. Ther. Targets (2011) 15(12)
Exp
ert O
pin.
The
r. T
arge
ts D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Sask
atch
ewan
on
06/2
9/12
For
pers
onal
use
onl
y.







