ecaths1.s3.amazonaws.comecaths1.s3.amazonaws.com/virologiaveterinaria2014... · web viewesquema...
TRANSCRIPT

UNIVERSIDAD JUAN AGUSTÍN MAZAFACULTAD DE CIENCIAS VETERINARIAS Y AMBIENTALESCÁTEDRA DE VIROLOGÍA Y MICOLOGÍA VETERINARIA
CÁTEDRA DE VIROLOGÍA Y MICOLOGÍAVIROLOGÍA
TEORICO Nº 4Temario: Respuesta inmunitaria frente a las infecciones virales. Recuperación de la
infección vírica. Inmunidad frente a las reinfecciones. Inmunidad pasiva.
Respuesta inmunitaria frente a las infecciones virales.
Los vertebrados han desarrollado un grupo elaborado de medidas defensivas en respuesta a las constantes amenazas de invasión de los agentes infecciosos, incluyendo los virus. Este grupo de respuestas son denominadas en conjunto: sistema inmunitario. Durante el encuentro inicial con los virus, el sistema inmunitario del animal hospedador reconoce como extrañas a ciertas macromoléculas virales (proteínas, carbohidratos) llamadas antígenos. Estos provocan varios tipos de respuestas para eliminar al virus y prevenir las re-infecciones. Los linfocitos B (respuesta humoral) responden frente al estimulo antigénico mediante la producción y secreción de inmunoglobulinas o anticuerpos. Los linfocitos T (respuesta celular) secretan citoquinas que regulan la respuesta inmune. Estas moléculas coordinan las actividades de varios tipos celulares involucrados, incluyendo la producción de anticuerpos por los linfocitos B y las funciones citotóxicas directas de los linfocitos T. Los linfocitos T y B expresan receptores altamente específicos que reconocen regiones de las proteínas virales, conocidas como determinantes antigénicos o epitopos.
Las respuestas inmunitarias antígeno-específicas en conjunto con los mecanismos de defensa innatos eliminan muchas infecciones virales antes que estas generen daños importantes en el animal hospedador. Esto suele resultar en una enfermedad leve e incluso sub-clínica.
Componentes celulares del sistema inmunitario
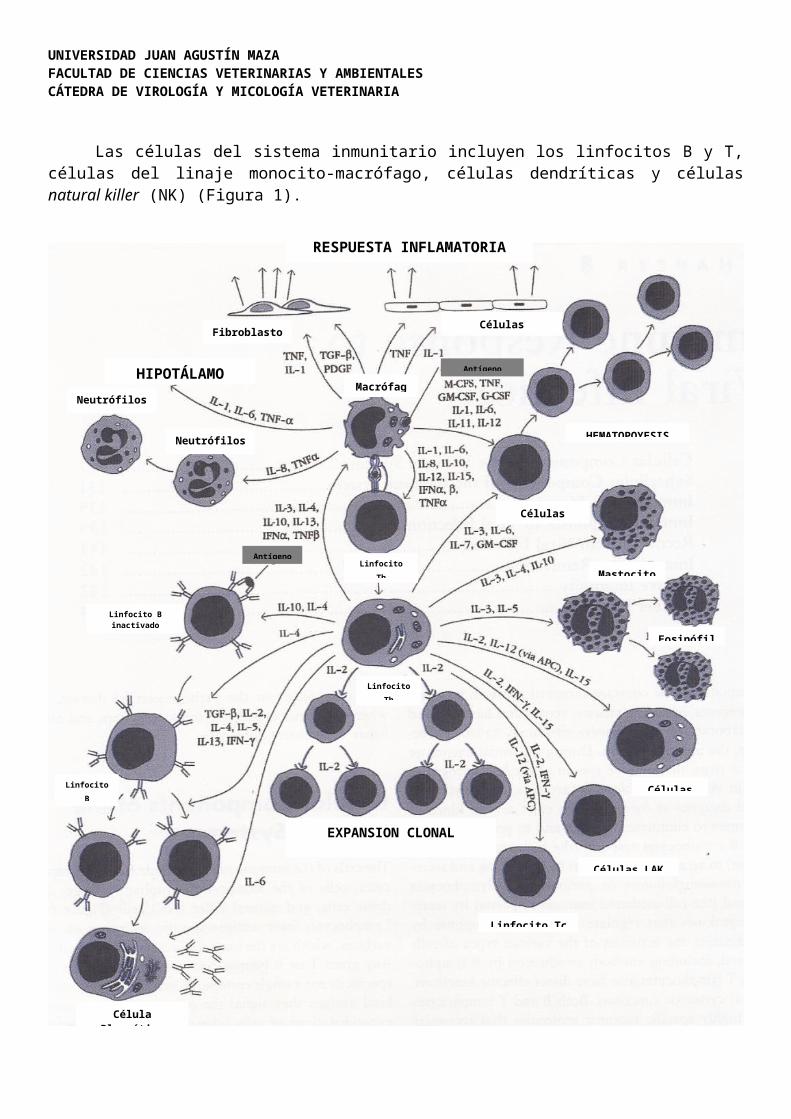
Las células del sistema inmunitario incluyen los linfocitos B y T, células del linaje monocito-macrófago, células dendríticas y células natural killer (NK) (Figura 1).
UNIVERSIDAD JUAN AGUSTÍN MAZAFACULTAD DE CIENCIAS VETERINARIAS Y AMBIENTALESCÁTEDRA DE VIROLOGÍA Y MICOLOGÍA VETERINARIA
Figura 1. Esquema generalizado de la compleja red de interacciones del sistema inmunitario frente a la infección viral. El proceso comienza con la interacción del antígeno viral extracelular con los macrófagos. Estos procesan y presentan a los linfocitos T helper (Th), los productos peptídicos procesados del antígeno, en los receptores del Complejo Mayor de Histocompatibilidad tipo II (CMH-II). Los receptores de los linfocitos Th reconocen específicamente estos péptidos procesados expresados en los receptores CMH-II. Esta interacción induce una serie de señales transduccionales en los Linfocitos Th que conducen a la síntesis y liberación de citoquinas. Estas moléculas proteicas se unen a su receptor específico sobre la superficie de una gran variedad de células. La interacción citoquina-receptor en una célula específica
RESPUESTA INFLAMATORIA
FibroblastosCélulas endoteliales
Antígeno viral
Macrófago
Linfocito Th inactivado
Linfocito Th activado
EXPANSION CLONAL
Antígeno
Células Stem
Mastocitos
Eosinófilos
Células NK
Células LAK
Linfocito Tc
HEMATOPOYESISNeutrófilos
Neutrófilos
HIPOTÁLAMO
Linfocito B inactivado
Linfocito B activado
Célula Plasmática

UNIVERSIDAD JUAN AGUSTÍN MAZAFACULTAD DE CIENCIAS VETERINARIAS Y AMBIENTALESCÁTEDRA DE VIROLOGÍA Y MICOLOGÍA VETERINARIA
induce la síntesis de otras citoquinas o conduce a las células a efectuar acciones más directas como la lisis de las células infectadas con virus desarrollada por los linfocitos T citotóxicos (Tc).
Los linfocitos tienen receptores antígeno-específicos sobre su superficie, los cuales representan la base de la especificidad inmunológica. Tanto los linfocitos B como T poseen receptores con especificidad para un sólo epitopo. Cuando los linfocitos T o B se unen al antígeno, desarrollan una serie de señales que indican a las células a dividirse mitóticamente para formar un clon expandido de células (expansión clonal). Los linfocitos B se diferencian a células plasmáticas, las cuales producen y secretan los anticuerpos. Los linfocitos T secretan factores solubles conocidos como linfoquinas, interleucinas o simplemente citoquinas. Estas moléculas regulan a las células involucradas en la respuesta inmune. Algunos linfocitos B y T se transforman en pequeñas células longevas responsables de la denominada memoria inmunológica. Mientras los anticuerpos y los receptores sobre la superficie de los linfocitos B reconocen epitopos de los antígenos extraños, los receptores de los linfocitos T reconocen pequeños péptidos que son generados por el procesamiento de las proteínas virales. Este reconocimiento se realiza únicamente si dichas péptidos procesados se expresan en los receptores glicoproteicos conocidos como proteínas del Complejo Mayor de Histocompatibilidad (CMH).
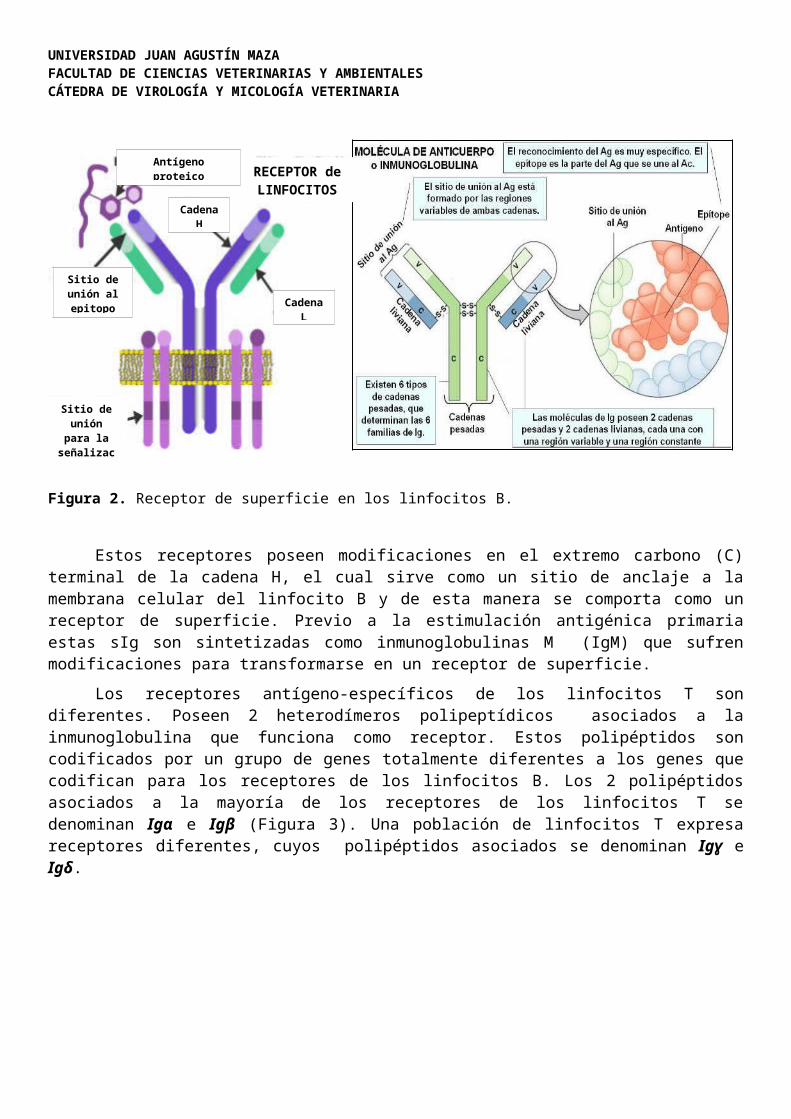
Receptores antígeno-específicos.Los receptores antígeno-específicos presentes sobre la superficie de los linfocitos B son
inmunoglobulinas modificadas compuestas por 4 cadenas polipeptídicas: 2 cadenas livianas (L) y 2 cadenas pesadas (H), denominadas inmunoglobulinas de superficie (sIg) (Figura 2).
Figura 2. Receptor de superficie en los linfocitos B.
Estos receptores poseen modificaciones en el extremo carbono (C) terminal de la cadena H, el cual sirve como un sitio de anclaje a la membrana celular del linfocito B y de esta manera se comporta como un receptor de superficie. Previo a la estimulación antigénica primaria estas sIg
RECEPTOR de LINFOCITOS B
Antígeno proteico inmunogénico
Cadena H
Cadena L
Sitio de unión al epitopo
Sitio de unión para
la señalización intracelular
UNIVERSIDAD JUAN AGUSTÍN MAZAFACULTAD DE CIENCIAS VETERINARIAS Y AMBIENTALESCÁTEDRA DE VIROLOGÍA Y MICOLOGÍA VETERINARIA
son sintetizadas como inmunoglobulinas M (IgM) que sufren modificaciones para transformarse en un receptor de superficie.
Los receptores antígeno-específicos de los linfocitos T son diferentes. Poseen 2 heterodímeros polipeptídicos asociados a la inmunoglobulina que funciona como receptor. Estos polipéptidos son codificados por un grupo de genes totalmente diferentes a los genes que codifican para los receptores de los linfocitos B. Los 2 polipéptidos asociados a la mayoría de los receptores de los linfocitos T se denominan Igα e Igβ (Figura 3). Una población de linfocitos T expresa receptores diferentes, cuyos polipéptidos asociados se denominan Igɣ e Igδ.
Figura 3. Receptor de superficie en los linfocitos T.
El reconocimiento especifico y la unión de los epitopos a los receptores sIg o de linfocitos T genera señales de transducción a través de la membrana plasmática de los linfocitos, produciendo una amplia variedad de procesos efectores que tienen como función atacar y remover a los virus invasores o a las células infectadas con virus (Figura 1). La cascada de interacciones célula-célula y la secreción de citoquinas amplifican la respuesta inmunitaria y establecen una respuesta memoria a largo plazo que capacita al sistema inmune para responder más rápidamente (respuesta amnésica o secundaria) a la reinfección con el mismo virus.
Linfocitos BAlgunas células madre (stem) hematopoyéticas pluripotenciales, que provienen del hígado
fetal o de la médula ósea, se diferencian en linfocitos B. Esto ocurre en la bolsa de Fabricio en las aves o en la misma médula ósea en los mamíferos. Estas células se caracterizan por poseer receptores de unión a antígenos específicos sobre la superficie de la membrana plasmática, como así también receptores de unión al complemento (C3) y receptores de unión a la fracción Fc de las inmunoglobulinas.
Cadena H
Cadena L
Sitio de reconocimiento
Sitio de señalización
RECEPTOR DE LINFOCITO T

UNIVERSIDAD JUAN AGUSTÍN MAZAFACULTAD DE CIENCIAS VETERINARIAS Y AMBIENTALESCÁTEDRA DE VIROLOGÍA Y MICOLOGÍA VETERINARIA
Cada linfocito B individual y su progenie expresan un grupo de genes que codifican para inmunoglobulinas que son específicas para un solo epitopo. Durante el desarrollo de tales células pueden suceder tres posibles acontecimientos: 1) las células pueden reaccionar contra un mismo antígeno y luego ser eliminadas, 2) las células pueden ser inviables para reaccionar y son eliminadas o 3) pueden reaccionar frente al antígeno extraño y proliferar.
Por el contrario a lo que sucede con los linfocitos T, los receptores sIg de los linfocitos B reconocen antígenos en su estado nativo y soluble, en lugar de reconocer a los antígenos expresados en los CMH-II sobre la superficie de los macrófagos. Esto es debido a que los linfocitos B interactúan directamente con las proteínas virales. Cuando una población clonal de linfocitos B expresa receptores complementarios contra alguno de los epitopos presentes en un antígeno, ellos responden y se diferencian en células plasmáticas productoras de anticuerpos, siempre y cuando haya recibido las señales apropiadas desde los linfocitos Th (Figura 1).
Cada célula plasmática secreta anticuerpos específicos, los cuales poseen la misma región variable (V) que los receptores sIg del linfocito B que le dio origen (Figura 2). Inicialmente estos anticuerpos son secretados como inmunoglobulinas M (IgM), pero los genes que codifican estas inmunoglobulinas sufren recombinación genética somática (traslocación), en la cual se combinan la región V con diferentes dominios constantes de las cadenas H. En consecuencia, luego de unos días, se generan IgG, IgA e incluso IgE con la misma especificidad. Varias citoquinas intervienen en la generación de estos anticuerpos específicos. Al comienzo de la respuesta inmune, cuando ingresan grandes cantidades de antígenos, los linfocitos B pueden ser activados incluso si sus receptores se ajustan al epítopo con una afinidad relativamente baja. El resultado de esta primera respuesta es la producción de anticuerpos que se unen a los antígenos con una baja afinidad. Posteriormente, cuando la cantidad de antígenos se ha reducido, los linfocitos B sufren hiper-mutaciones en los genes que codifican para la región V de los anticuerpos producidos. Estos linfocitos se seleccionan y producen anticuerpos que pueden unirse a los antígenos con una elevada afinidad. Este proceso recibe el nombre de afinidad madurativa (Figura 4).
AFINIDAD MADURATIVAEtapa inicial de la respuesta inmune
Linfocito B
Receptor sIg
Antígeno
Activación
Anticuerpos producidos
CLONACION con HIPERMUTACIONES
Muerte delinfocitos B con
baja afinidad
Selección delinfocitos B con
alta afinidad
AFINIDAD MADURATIVAEtapa tardía de la respuesta inmune
Población inicial de
Linfocitos B
Selección clonal e hipermutaciones somáticas
Linfocitos BADAPTADOS

UNIVERSIDAD JUAN AGUSTÍN MAZAFACULTAD DE CIENCIAS VETERINARIAS Y AMBIENTALESCÁTEDRA DE VIROLOGÍA Y MICOLOGÍA VETERINARIA
Figura 4. Proceso de afinidad madurativa en los linfocitos B.
Linfocitos TLos linfocitos T reciben este nombre debido a que maduran a partir de las células stem,
hematopoyéticas, pluripotenciales en el timo (T=timo). Dentro del timo, se produce una selección positiva de aquellos linfocitos T capaces de reconocer péptidos específicos sobre la superficie de las células y una selección negativa de aquellos linfocitos T que reconocen y atacan los antígenos de las propias células del organismo. Sólo el 1 o 2% de los linfocitos producidos en el timo abandonan este órgano y pueblan los tejidos linfoides secundarios. Los linfocitos T se clasifican en dos sub-poblaciones: linfocitos T helper (Th), los cuales se diferencian en linfocitos Th1 y Th2 y linfocitos T citotóxicos (Tc). Los linfocitos Th se caracterizan por sus funciones regulatorias sobre el sistema inmune y los linfocitos Tc por su función efectora directa que consiste en generar lisis de las células blanco. El análisis exhaustivo de los clones de linfocitos T indica que un tipo celular en particular puede realizar tanto las funciones regulatorias como las funciones efectoras y además secretar un amplio rango de citoquinas.
- Linfocitos Th. Los linfocitos Th poseen en la superficie de la membrana celular receptores conocidos
como CD4. Estas moléculas proteicas se asocian a otros receptores presentes en la superficie del linfocito Th (RTC) que pueden reconocer péptidos virales que se expresan en los receptores del CMH-II, normalmente presentes en las células presentadoras de antígenos (CPA) (Figura 5).
Figura 5. Linfocitos Th CD4+ o simplemente linfocitos T4 (LT4), expresando receptores del tipo CD4+ en la superficie de su membrana celular. Estos receptores interactúan con otros receptores presentes en la superficie del linfocito Th denominados RTC. Estos últimos, reconocen los péptidos virales (Ag) que se

UNIVERSIDAD JUAN AGUSTÍN MAZAFACULTAD DE CIENCIAS VETERINARIAS Y AMBIENTALESCÁTEDRA DE VIROLOGÍA Y MICOLOGÍA VETERINARIA
expresan en los receptores del CMH-II, normalmente presentes en las células presentadoras de antígenos (CPA) tales como macrófagos o células dendríticas.
Los linfocitos Th secretan citoquinas que los auto-activan y además activan a linfocitos Tc, linfocitos B e incluso a otros tipos de linfocitos Th. Durante este proceso, los linfocitos Tc son transformados en células citotóxicas y los linfocitos B son inducidos a producir inmunoglobulinas.
Los linfocitos Th1 (células T inflamatorias) se caracterizan por:
1) Secretar citoquinas del tipo interleucina 2 (IL-2), interferón gamma (IFN-ɣ), factor de necrosis tumoral beta (TNF-β), factor estimulante de colonias de granulocitos-macrófagos (GM-CSF) e interleucina 3 (IL-3).
2) Mediar las respuestas de hipersensibilidad del tipo retardada o tipo 4.
3) Promover la secreción de inmunoglubina G tipo 2 (IgG2) por parte de los linfocitos B.
Los linfocitos Th2 se caracterizan por:
1) Secretar IL-3, IL-4, IL-5 e IL-6, además de secretar GM-CSF.
2) Participar de manera indirecta en la generación de las respuestas de hipersensibilidad del tipo retardada o tipo 4.
3) Promover la producción de IgG1, a partir de la IgG2, en los linfocitos B.
Tanto los linfocitos Th1 como Th2 proliferan simultáneamente durante las infecciones virales persistentes crónicas.
Los linfocitos Th1 expresan receptores denominados CD4, sobre la superficie de su membrana celular (aunque algunos linfocitos Th también expresan receptores del tipo CD8). Los linfocitos Th1 CD4+ secretan citoquinas que establecen la respuesta inflamatoria e incrementan enormemente la respuesta inmunitaria atrayendo monocitos/macrófagos y otros linfocitos T al sitio de infección viral. Las citoquinas secretadas actúan sobre el propio linfocito Th1 activándolo, induciendo su proliferación y diferenciación y estimulando la secreción de otras citoquinas. Esta respuesta es la base para comprender las reacciones de hipersensibilidad tipo 4, las cuáles han sido reconocidas como parte de la patogénesis de muchas infecciones virales. El mismo tipo de respuesta celular se produce cuando un antígeno es inyectado intradérmicamente para la realización de test diagnósticos cutáneos. La inoculación intradérmica del antígeno viral genera una reacción celular localizada (hipersensibilidad tipo 4) en aquellos animales que previamente han tenido contacto con ese mismo antígeno viral.
Algunos linfocitos T atenúan las respuestas de los linfocitos B o de otros linfocitos T, lo que sugiere que estos linfocitos pueden pertenecer a otra sub-población celular. Algunos autores han denominado a esta sub-población linfocitos T supresores. Sin embargo, ha resultado difícil clonar estos linfocitos T, que supuestamente ejercen de manera exclusiva esta propiedad, y actualmente se considera que estas funciones supresoras son desarrolladas por los linfocitos Th y Tc y no por una sub-población diferente. Los linfocitos T pueden suprimir varias de las funciones que se desarrollan durante la respuesta inmunitaria, mediante la interacción directa con los linfocitos o produciendo citoquinas inmunosupresoras.
- Linfocitos Tc.Los linfocitos Tc expresan en la superficie de la membrana plasmática receptores del tipo
CD8+ y además poseen receptores que reconocen los péptidos virales expresados en los receptores del Complejo Mayor de Histocompatibilidad tipo I (CMH-I) de aquellas células infectadas con virus (Figura 6).

UNIVERSIDAD JUAN AGUSTÍN MAZAFACULTAD DE CIENCIAS VETERINARIAS Y AMBIENTALESCÁTEDRA DE VIROLOGÍA Y MICOLOGÍA VETERINARIA
Figura 6. Linfocitos Tc CD8+ o simplemente linfocitos T8 (LT8), expresando receptores del tipo CD8+ en la superficie de su membrana celular. Estos receptores interactúan con otros receptores presentes en la superficie del linfocito Tc denominados RTC. Estos últimos, reconocen los péptidos virales que se expresan en los receptores del CMH-I, presentes en las células infectadas con un determinado virus.
La activación de los linfocitos Tc y la posterior citólisis de la célula infectada requieren de un estrecho contacto directo entre el linfocito Tc y la célula infectada, de una manera similar a la cercanía entre dos neuronas para efectuar sinapsis. De hecho, es tan estrecho el contacto producido que este fenómeno se ha denominado “el beso de la muerte”. Los gránulos presentes en el citoplasma del linfocito Tc se polarizan hasta buscar posicionarse en la cercanía de la membrana citoplasmática de la célula infectada. Cuando ambas membranas celulares están en estrecho contacto, estos gránulos liberan su contenido al reducido espacio presente entre ambas células (Figura 7).

UNIVERSIDAD JUAN AGUSTÍN MAZAFACULTAD DE CIENCIAS VETERINARIAS Y AMBIENTALESCÁTEDRA DE VIROLOGÍA Y MICOLOGÍA VETERINARIA
Figura 7. Estrecho contacto entre el linfocito Tc y la célula infectada por virus. Obsérvese el reducido espacio entre las membranas citoplasmáticas de ambas células. Esta unión tan estrecha recibe el nombre de “beso de la muerte”.
El contenido de los gránulos liberados por los linfocitos Tc al espacio intercelular contiene una proteína denominada perforina. La perforina es secretada por el linfocito Tc como un monómero proteico. Este monómero proteico se polimeriza y forma una estructura compleja, que se inserta en la membrana citoplasmática de la célula infectada (Figura 8).
Figura 8. La perforina es secretada por el linfocito Tc al espacio intercelular generado entre esta célula y la célula infectada. La perforina se secreta como un monómero, el cual requiere de calcio extracelular para poder unirse a la superficie externa de la membrana citoplasmática de la célula infectada (1). Posteriormente el monómero de perforina es insertado perpendicularmente a la bicapa lipídica de la membrana citoplasmática de la célula infectada (2). El monómero de perforina se polimeriza, primero en dos subunidades que se disponen de manera adyacente y perpendicular en la membrana citoplasmática (3). La polimerización de la perforina continúa hasta formar una especie de poro (o canal) que genera una comunicación directa entre el espacio extracelular y el citoplasma de la célula infectada (4). Este orifico permite la entrada de líquido extracelular, lo cual genera alteraciones osmóticas en el espacio intracelular, generando la lisis osmótica de la célula infectada.
Mónomero de Perforina
Membrana citoplasmática
Perforina polimerizada
Espacio Extracelular
Espacio Intracelular
Flujo osmótico de líquido extracelular al
interior de la célula infectada.
CELULA INFECTADA

UNIVERSIDAD JUAN AGUSTÍN MAZAFACULTAD DE CIENCIAS VETERINARIAS Y AMBIENTALESCÁTEDRA DE VIROLOGÍA Y MICOLOGÍA VETERINARIA
Una vez insertada, la perforina genera un poro en la membrana de la célula infectada. Este “orificio” creado genera inevitablemente la lisis osmótica de la célula infectada. También se ha demostrado que los gránulos presentes en los linfocitos Tc y en las células NK poseen una elevada concentración de una enzima con actividad serina-esterasa (grazima), la cual induce apoptosis en la célula infectada (Figura 9).
Figura 9. Muerte celular por apoptosis mediada por las granzimas liberadas por los linfocitos Tc. Los gránulos presentes en el linfocito Tc vierten su contenido al espacio intercelular generado entre el linfocito Tc y la célula infectada por virus. El contenido de estos gránulos posee una elevada concentración de una enzima denomina granzima. Existen dos tipos de granzimas: A y B. La granzima B genera liberación mitocondrial de citocromo c (Cyt c), una proteína mitocondrial que participa en la cadena respiratoria mitocondrial. El Cyt c induce la activación de las enzimas caspasas, quienes estimulan a la NADPH oxidasa a generar mayor concentración de especies reactivas del oxígeno (ROS) a partir del oxígeno celular. La elevación intracitoplasmática de las ROS induce la muerte celular programada (apoptosis). La granzima A aumenta la liberación de las ROS quienes activan la Muerte Celular Independiente de la vía de las Caspasas (CICD).
La efectiva respuesta de los linfocitos T es transitoria: en algunas infecciones virales agudas el pico de actividad de los linfocitos Th y Tc sucede 1 semana después del ingreso del virus y dicha actividad desaparece luego de 3 a 4 semanas. La disminución de la actividad citotóxica de los linfocitos Tc se cree puede deberse a la destrucción de las células infectadas, con la consecuente remoción del estímulo antigénico que dispara la actividad de estas células o a las funciones supresoras de algunos linfocitos T sobre los linfocitos Tc.
- Linfocitos T ɣ/δ.
LINFOCITO T CITOTÓXICO
Granzima A Granzima B
CELULA INFECTADA POR VIRUS
Sustrato
Caspasas
ROS
APOPTOSISAPOPTOSIS

UNIVERSIDAD JUAN AGUSTÍN MAZAFACULTAD DE CIENCIAS VETERINARIAS Y AMBIENTALESCÁTEDRA DE VIROLOGÍA Y MICOLOGÍA VETERINARIA
En el epitelio de algunas superficies corporales como la piel e intestino, incluso en los pulmones se encuentra una sub-población característica de linfocitos T denominados Linfocitos T ɣ/δ. Estas células expresan un receptor en la superficie de la membrana citoplasmática del linfocito compuesto por 2 heterodímeros polipetídicos denominados gamma (ɣ) y delta (δ) (Figura 10).
Figura 10. Receptor localizado en la superficie de la membrana citoplasmática de los linfocitos T ɣ/δ. Estos linfocitos poseen dos heterodímeros polipeptídicos (ɣ y δ) asociados a la inmunoglobulina que funciona como receptor del linfocito T.
Existe cada vez más evidencia que esta sub-población de linfocitos T están involucrados en la respuesta inmunitaria frente a aquellas infecciones virales cuyos viriones ingresan por las superficies corporales donde estos linfocitos T se localizan (piel, intestino, pulmones). En cerdos, rumiantes y pollos los linfocitos T ɣ/δ representan aproximadamente el 30% de los linfocitos T totales y se encuentran ampliamente distribuidos en los tejidos cutáneos, intestinales y pulmonares; mientras que en ratones y humanos esta sub-población de linfocitos T representa menos del 5% del total de los linfocitos T presentes en estas especies.
Monocitos, macrófagos y células dendríticasLos monocitos, macrófagos y células dendríticas son importantes grupos celulares que
inician la respuesta inmunitaria frente a infecciones virales. Esto se debe principalmente a la gran motilidad que presentan los monocitos, lo cual incrementa la posibilidad de detectar viriones que intentan ingresar en cualquier superficie corporal y a la variada localización tisular de los macrófagos (macrófagos alveolares en el pulmón o células de Kupffer en el hígado) y las células dendríticas (células de Langerhans en la piel). Estas células participan durante los estadios iniciales de la infección viral: 1) los monocitos se infiltran en los tejidos y se diferencian rápidamente a macrófagos, 2) los macrófagos proliferan y se transforman en las células predominantes en el sitio de infección, a tan sólo 24 hs de la invasión tisular de los viriones y 3) las células dendríticas desempeñan importantes funciones inmunológicas en todas las superficies corporales y en órganos claves como los linfonódulos, bazo e hígado se encargan de remover
RECEPTOR EN EL LINFOCITO T ɣ/δ

UNIVERSIDAD JUAN AGUSTÍN MAZAFACULTAD DE CIENCIAS VETERINARIAS Y AMBIENTALESCÁTEDRA DE VIROLOGÍA Y MICOLOGÍA VETERINARIA
eficientemente cualquier partícula extraña que ingresa. Los tres tipos celulares expresan en la superficie de sus membranas celulares tanto receptores para la fracción Fc de las inmunoglobulinas como receptores para la fracción C3b del complemento (Figura 11).
Figura 11. Receptores para la fracción Fc de las inmunoglobulinas y receptor para la fracción C3b del complemento presentes en la membrana citoplasmática de las células dendríticas (DC).
Estos receptores promueven la fagocitosis de los complejos inmunes, formados por los viriones cubiertos de anticuerpos (Figura 12). Los tres tipos celulares se denominan células profesionales presentadoras de antígenos y ejercen una gran influencia en el control de la rapidez, magnitud y dinámica de la respuesta inmunitaria.
Receptor para la fracción C3b del complemento (CR3):
-Facilita la ingesta del virión recubierto de anticuerpos (ag)Receptor para la fracción Fc de la
inmunoglobulina E (FcƸRI):-Facilita la respuesta inducida por alérgenos.
Receptor para la fracción C1q del complemento (CRT/CD91):
-Induce la quimiotaxis de la célula
Receptor para la fracción Fc de la inmunoglobulina G (FcɣR):
-Induce la maduración de la célula dendrítica- Facilita la fagocitosis del complejo inmune (IgG-ag)

UNIVERSIDAD JUAN AGUSTÍN MAZAFACULTAD DE CIENCIAS VETERINARIAS Y AMBIENTALESCÁTEDRA DE VIROLOGÍA Y MICOLOGÍA VETERINARIA
Figura 12. Fagocitosis de los complejos inmunes por el macrófago.
Las citoquinas liberadas por los linfocitos Th (Figura 1) atraen una mayor cantidad de monocitos al foco infeccioso y los activan para luego diferenciarse a macrófagos. Los macrófagos activados poseen una mayor actividad quimiotáctica y fagocítica y un mayor poder de digestión de los antígenos ingeridos.
Células Natural KillerLas células Natural Killer (NK) son un grupo heterogéneo de linfocitos que expresan
receptores CD3-, CD16+ y CD56+ (Figura 13). Son células granulares, de un linaje celular desconocido, que tienen la capacidad de matar a las células infectadas por virus y a las células tumorales. La base de su selectividad por las células infectadas con virus se relaciona con la síntesis y expresión de CMH-I por estas células infectadas, al inicio de la infección viral (Figura 13).
Los anticuerpos recubren la superficie de los viriones libres, formando el
complejo inmune. De esta manera los virus envueltos no pueden fusionarse
con la membrana de la célula hospedadora.
Anticuerpos
El virus recubierto de anticuerpo es reconocido por los receptores para la fracción Fc de los anticuerpos,
presentes en la superficie del macrófago. Esta interacción induce la fagocitosis del compelo inmune.
Célula hospedadora
Macrófago

UNIVERSIDAD JUAN AGUSTÍN MAZAFACULTAD DE CIENCIAS VETERINARIAS Y AMBIENTALESCÁTEDRA DE VIROLOGÍA Y MICOLOGÍA VETERINARIA
Figura 13. Las células NK expresan receptores CD16 y CD 56 que interactúan con sus ligandos en la célula infectada por virus. Además las NK pueden interaccionar con los antígenos virales expresados en los CMH-I de la célula infectada. Aunque esta interacción no es esencial para que la célula NK libere sus enzimas líticas (contenidas en gránulos citoplasmáticos) y generen la destrucción de la célula infectada.
Las células NK no tienen especificidad inmunológica para un antígeno viral en particular, no tienen memoria, no se limitan a interaccionar sólo con las proteínas CMH-I (Figura 13) y no son dependientes de los anticuerpos para su activación. Desempeñan un importante rol al comienzo de la respuesta inmune, evidenciado por su enorme proliferación dentro del primer o segundo día del comienzo de la infección. La activación de las células NK inducida por los virus es mediada por los interferones, que actúan sinérgicamente con IL-2 (Figura 1). Las células NK secretan citoquinas, incluyendo interferón ɣ y factor de necrosis tumoral alfa (FNT-α).
Componentes subcelulares del sistema inmune.
Complejo Mayor de HistocompatibilidadEs necesario conocer la estructura y la producción celular de las proteínas del Complejo
Mayor de Histocompatibilidad (CMH) para comprender completamente el procesamiento y presentación de los antígenos virales. Durante la ontogenia de linfocitos T, la selección positiva de estas células sucede en el timo, mediante las mismas proteínas CMH, que permiten la proliferación de linfocitos T maduros que pueden reconocer péptidos extraños. Estos péptidos son reconocidos siempre y cuando se expresen en los CMH. Los linfocitos T no reconocen péptidos extraños que se encuentren libres en el espacio extracelular. Este fenómeno se conoce como maduración linfocitaria restringida por los CMH o simplemente restricción CMH (Figura 14).
Interacción de las células NK con las células infectadas que
expresan antígenos virales en los CMH-I
Gránulos líticos
CMH-I
Célula infectada por un virus
Receptores CD16 y CD 56
Ligando
MUERTE CELULAR
Interacción de las células NK con las células infectadas que NO
expresan los antígenos virales en los CMH-I
NKNK
Célula infectada por un virus
Liberación de gránulos líticos
MUERTE CELULAR

UNIVERSIDAD JUAN AGUSTÍN MAZAFACULTAD DE CIENCIAS VETERINARIAS Y AMBIENTALESCÁTEDRA DE VIROLOGÍA Y MICOLOGÍA VETERINARIA
Figura 14. Modelo de la interacción entre las proteínas del CMH, los receptores de los linfocitos T y las moléculas CD4 y CD8. (A) Las proteínas CD8, expresadas sobre la superficie de la membrana citoplasmática de los linfocitos T citotóxicos, interactúan con las proteínas del CMH-I expresados sobre la superficie de las células infectadas. Los CMH-I que expresan el péptido extraño interactúan con los receptores de los linfocitos Tc (TCR). (B) Las proteínas CD4, expresadas sobre la superficie de la membrana citoplasmática de los linfocitos T helper, interactúan con las proteínas del CMH-II expresados sobre la superficie de las células presentadoras de antígenos. Los CMH-II que expresan el péptido extraño interactúan con los receptores de los linfocitos Th (TCR). Los TCR son heterodímeros formados por dos cadenas polipeptidicas (α y β), Cada una de estas cadenas posee un dominio constante anclado en la membrana celular y una región variable que es la que interactúa con el péptido extraño. Los TCR están asociados a una molécula accesoria denominada CD3 (expresada sobre la superficie del linfocito T), la cual se compone de tres polipéptidos (ɣ, δ y Ƹ) y a un homodímero (ζ) el cual sirve como conductor de la señal transductora que se emite hacia el interior del linfocito T. Esta señal fosforila (activa) la proteína tirosina quinasa p56lck, asociada a la proteína CD4 o CD8. Las proteínas CMH-I se componen de una cadena polipeptidica con 3 dominios extracelulares, asociados con una microglobulina (β2); mientras que las proteinas CMH-II se componen de dos cadenas polipeptídicas, cada una de las cuales expresan dos dominios extracelulares.
Existen dos tipos de proteínas CMH: tipo I y II, cuya estructura se muestra en la Figura 15. Las dos sub-poblaciones de linfocitos, Tc y Th, interaccionan con los CMH-I y CMH-II, respectivamente. La forma en la que las células procesan y presentan los péptidos antigénicos a los linfocitos Tc y Th es fundamentalmente diferente. Se denomina vía exógena a la presentación de los péptidos antigénicos en los CMH-II y vía endógena a la presentación de los péptidos antigénicos en los CMH-I.
Los CMH-I pueden expresarse sobre la membrana citoplasmática de muchos tipos celulares (la única excepción son las neuronas) y los CMH-II se expresan en la membrana citoplasmática de las células presentadoras de antígeno. En el dominio distal de estos CMH existe una hendidura donde se expresa y presenta los péptidos antigénicos. En los CMH-I esta hendidura es cerrada y en los CMH-II es abierta (Figura 15). El complejo péptido-CMH es reconocido, con absoluta especificidad, por los receptores de linfocitos T (TCR) expresados en la superficie celular de clones específicos de linfocitos T.
CELULA INFECTADA
LINFOCITO T CITOTÓXICO
CMH-IPEPTIDO
MEMBRANA CITOPLASMÁTICA PEPTIDO
CMH-II
CELULA PRESENTADORA DE
ANTIGENOS
LINFOCITO T HELPER

UNIVERSIDAD JUAN AGUSTÍN MAZAFACULTAD DE CIENCIAS VETERINARIAS Y AMBIENTALESCÁTEDRA DE VIROLOGÍA Y MICOLOGÍA VETERINARIA
Figura 15. Diagrama esquemático de las proteínas del CMH-I y CMH-II que muestra los dominios externos (distal y proximal), transmembrana y la cola citoplasmática.
A pesar que los complejos péptido-CMH son importantes en la evocación de la respuesta inmune frente a las infecciones virales, algunas especies animales carecen de proteínas CMH en algunas de sus células. Estos animales son más susceptibles a padecer las consecuencias de la infección viral.
Presentación de antígenos por las células que expresan CMH-II: vía exógena. Sólo determinados tipos celulares, denominadas células presentadoras de antígenos, pueden procesar y presentar los antígenos virales a los linfocitos T, mediante las proteínas del CMH-II. Las células presentadoras de antígenos son los monocitos/macrófagos, las células dendríticas y en estadios tardíos de la respuesta inmune, los linfocitos B. Las células dendríticas, que incluyen a las células de Langerhans de la piel y a las células dendríticas de los linfonódulos, de las zonas marginales y de la pulpa roja del bazo; se denominan así debido a que emiten prolongaciones celulares que se interdigitan con los linfocitos para favorecer la presentación de los antígenos. Los macrófagos expresan relativamente pocas moléculas del CMH-II cuando están inactivos, en comparación con la expresión natural de estas moléculas en la superficie de las células dendríticas. Cuando los macrófagos son activados, mediante la acción del interferón ɣ, estas células expresan un mayor número de moléculas del CMH-II en la superficie de su membrana celular. En los estadios tardíos de la respuesta inmune o durante las re-infecciones virales, los linfocitos B se convierten en importantes células presentadoras de antígenos. Los antígenos virales, y en algunos casos los viriones completos, se unen a los receptores sIg expresados en la superficie de los linfocitos B. Ambas estructuras son endocitadas y procesadas
CMH-I CMH-IIHendidura donde se une el péptido
antigénico
Microglobulina β2
Dominio distal
Dominio proximal
Segmento transmembrana
Cola citoplasmática

UNIVERSIDAD JUAN AGUSTÍN MAZAFACULTAD DE CIENCIAS VETERINARIAS Y AMBIENTALESCÁTEDRA DE VIROLOGÍA Y MICOLOGÍA VETERINARIA
intracitoplasmáticamente para luego ser expresadas en los CMH-II localizados sobre la superficie del linfocito B (ver vía exógena). Los péptidos antigénicos expresados representan diferentes epitopos de los mismos antígenos reconocidos por los linfocitos B y a partir de los cuales se han producido anticuerpos específicos. Los linfocitos Th CD4+, a los cuales los linfocitos B presentan los antígenos, responden secretando citoquinas que estimulan a los mismos linfocitos B para que ellos produzcan anticuerpos. Esta interacción, que involucra una estrecha asociación entre los linfocitos B y Th, asegura un eficiente transporte de citoquinas desde los linfocitos Th a los linfocitos B.
Las proteínas virales que son incorporadas desde una fuente externa por las células presentadoras de antígenos ingresan en la vía exógena. Estas proteínas exógenas (antígenos virales) son transportadas desde los endosomas tempranos a los endosomas tardíos (acídicos) y desde allí a los pre-lisosomas, donde son destruidas por las enzimas proteolíticas presentes en estas organelas (Figura 16). Los péptidos virales resultantes de esta degradación enzimática, se unen a las cadenas polipeptídicas α y β de los CMH-II sintetizado en el RER. Estas cadenas forman el complejo tetrámerico (péptido antigénico-cadena α-cadena β) que es transportado a la membrana citoplasmática de la célula presentadora de antígenos. Este complejo tetramérico (CT) es reconocido por los linfocitos Th CD4+, estimulando la respuesta de éstas células.
Figura 16. Vía exógena para la producción de los CMH-II. El antígeno viral es endocitado por la célula presentadora de antígeno (CPA). Este es transportado a través de los endosomas hasta ser digerido por las enzimas endosomales y/o pre-lisosomales. Como resultado de esta degradación resultan péptidos antigénicos que serán expresados en los CMH-II en la superficie de la CPA. Las proteinas del CMH-II son sintetizadas en el RER de la CPA. Esta proteína es sintetizada como una pre-proteína que posee una cadena anclada a la hendidura donde se une el antígeno en el CMH-II maduro. Esta cadena se denomina cadena invariante. La cadena invariante es digerida por las enzimas presentes en el aparato de Golgi, de manera que se libera la hendidura donde se unirá el péptido antígenico. En las vesículas exocíticas (VE)
CELULA PRESENTADORA DE ANTIGENOS
Antígeno exógeno
Endosomas
CT
Cadena invariante digerida
Ap. de Golgi
RER Cadena invariante
CMH-II
Núcleo
VÍA EXÓGENA(CMH-II)
VE
CMH-II

UNIVERSIDAD JUAN AGUSTÍN MAZAFACULTAD DE CIENCIAS VETERINARIAS Y AMBIENTALESCÁTEDRA DE VIROLOGÍA Y MICOLOGÍA VETERINARIA
se ensamblan el CMH-II (cadenas α y β) con el péptido antigénico, formando el complejo tetramérico (péptido-cadena α-cadena β). Este complejo se ancla en la membrana citoplasmática de la CPA, expresando el péptido antigénico y formando el CMH-II maduro.
Presentación de antígenos por las células que expresan CMH-I: vía endógena. La mayoría de las células presentes en el organismo animal sintetizan proteínas del CMH-I luego de ser infectadas por virus. La única excepción la constituyen las neuronas. Luego de su síntesis, las cadenas α (1, 2 y 3) y la microglobulina β2 son transportadas al RER donde son ensambladas para formar el CMH-I. En esta formación interviene una proteína chaperona denominada calexina (Figura 17).
Figura 17. Vía endógena para la producción de los CMH-I. Las células infectadas con virus degradan algunas proteínas virales en su citoplasma (proteosoma). Los péptidos resultantes son transportados por una molécula denominada TAP al interior del RER. Aquí se ensamblan las cadenas α, la microglobulina β2 y los péptidos antigénicos para formar el complejo tetramérico (CT). En esta formación interviene una proteína chaperona denominada calexina. El CT es exportado al aparato de Golgi y desde allí es transportado hasta la superficie de la célula para anclarse en la membrana citoplasmática y formar el CMH-I maduro.
Las células infectadas con virus degradan algunas proteínas virales en su citoplasma. En esta degradación interviene una organela llamada proteosoma. Las proteínas virales degradadas ingresan mediante este mecanismo a la vía endógena. Los péptidos resultantes son transportados por una molécula denominada TAP (transportador asociado al antígeno procesado) hacia el interior del RER. Aquí se ensamblan las cadenas α, la microglobulina β2 y los péptidos antigénicos para formar el complejo tetramérico (CT). Este es exportado al aparato de Golgi y
VÍA ENDÓGENA(CMH-I)
PROTEÍNAS
PROTEOSOMA
Péptidos
CALEXINA
RER
Ap. de Golgi
CT
CMH-I
cadenas α 1, 2 y 3 microglobulina β2 péptido antigénico

UNIVERSIDAD JUAN AGUSTÍN MAZAFACULTAD DE CIENCIAS VETERINARIAS Y AMBIENTALESCÁTEDRA DE VIROLOGÍA Y MICOLOGÍA VETERINARIA
desde allí es transportado hasta la superficie de la célula para anclarse en la membrana citoplasmática y formar el CMH-I que expresa el péptido antigénico a los linfocitos Tc.
CitoquinasLas citoquinas son proteínas de bajo peso molecular, similares a hormonas, las cuales
estimulan o inhiben la proliferación, diferenciación y/o maduración de las células inmunitarias (Figura 18a). Estas proteínas difieren de las verdaderas hormonas en que son producidas por células no especializadas. La mayoría de ellas son producidas por los linfocitos T (linfocinas o linfoquinas) o por los monocitos/macrófagos (monocinas o monoquinas) y se utilizan para la regulación de la respuesta inmune coordinando la actividad de los diferentes tipos celulares involucrados en esta respuesta.
Figura 18. (A) Esquema general de la inducción y función de citoquinas. (B) La mayoría de las citoquinas tienen acciones autocrina y/o paracrina y algunas también tienen funciones endocrinas.
Las citoquinas pueden actuar sobre la misma célula que las produce (acción autocrina) o en células vecinas (acción paracrina), especialmente en el espacio intercelular, donde puede ocurrir secreción bidireccional y es necesaria una muy baja concentración de estas moléculas para que la respuesta sea efectiva. También, las citoquinas pueden actuar sobre células localizadas en sitios distantes a los sitios donde las citoquinas son producidas (acción endocrina) (Figura 18b). Las células blanco expresan receptores para citoquinas específicas. Una sóla citoquina puede ejercer una multiplicidad de efectos biológicos y actuar sobre una o varios tipos celulares. Además, varias citoquinas pueden ejercer efectos biológicos similares, aunque para ello utilizan diferentes señales transduccionales para ejercer este efecto (Figura 19).
AB
Estímulo inductor
Célula productora de citoquina
Citoquina
Receptor
Señal
Célula blanco
Efecto Biológico
Acción autocrina
Acción paracrina Célula vecina
Acción endocrina
Circulación
Célula distante

UNIVERSIDAD JUAN AGUSTÍN MAZAFACULTAD DE CIENCIAS VETERINARIAS Y AMBIENTALESCÁTEDRA DE VIROLOGÍA Y MICOLOGÍA VETERINARIA
PLEIOTROPÍA
Linfocito Th activado
CÉLULA BLANCO EFECTO BIOLÓGICO
Linfocito B
Linfocito T en Timo
Mastocito
Linfocito BLinfocito Th activado
REDUNDANCIA
PROLIFERACIÓN
PROLIFERACIÓN
PROLIFERACIÓN
ACTIVACIÓNPROLIFERACIÓNDIFERENCIACIÓN
Linfocito Th activado
SINERGISMO
ANTAGONISMO
Linfocito Th activado
INDUCCIÓN DE IgE
Linfocito B
Linfocito B
BLOQUEO EN LA PRODUCCION DE IgE
mediante la IL-4

UNIVERSIDAD JUAN AGUSTÍN MAZAFACULTAD DE CIENCIAS VETERINARIAS Y AMBIENTALESCÁTEDRA DE VIROLOGÍA Y MICOLOGÍA VETERINARIA
Figura 19. Ejemplos de pleiotropía, redundancia, sinergismo y antagonismo en la acción biológica de las citoquinas.
Las citoquinas estimulan o inhiben a las células blanco y diferentes citoquinas pueden antagonizar a la acción de otras. Normalmente, una citoquina secretada por un tipo particular de célula activa a otro tipo de célula y la induce a secretar diferentes citoquinas o a expresar receptores para una citoquina en particular.
Las citoquinas pueden influenciar la patogénesis viral de diferentes maneras: 1) aumentando la respuesta inmune, por ejemplo: los linfocitos Tc, mediante la producción de FNT-α o interferón ɣ, estimulan la expresión de los CMH-II; 2) regulación del sistema inmune, por ejemplo: las interleucinas 4, 5 y 6 o el interferón ɣ inducen la producción de anticuerpos; 3) supresión de la respuesta inmune, por ejemplo: la IL-10 inhibe la síntesis de interferón ɣ; 4) inhibición de la replicación viral por la acción de los interferones y 5) estimulación de la expresión de genes virales.
AnticuerposEl resultado final de la activación y maduración de los linfocitos B es la producción de
anticuerpos, los cuales reaccionan específicamente contra los epitopos identificados previamente por sus receptores. Los anticuerpos se dividen en 4 clases principales: 2 monómeros, IgE e IgG y 2 polímeros, IgA e IgM. Todas las inmunoglobulinas o anticuerpos tienen una estructura similar, pero ellas varían enormemente en la secuencia de aminoácidos que constituyen el sitio de unión del antígeno. Esto determina su especificidad frente a un determinante antigénico en particular.
La inmunoglobulina más común presente en el suero es la IgG. La misma se compone de dos cadenas H y dos cadenas L y en cada cadena encontramos un dominio variable y otro constante. Las cadenas se mantienen unidas mediante enlances disulfuro. La enzima papaína divide a la molécula en dos fracciones ab (Fab) similares, las cuales contienen el sitio de unión del antígeno y una fracción c (Fc), la cual ejerce diversas funciones tales como la fijación del complemento, la adherencia a células fagocíticas y su transferencia hacia el calostro o la placenta (Figura 20).
Figura 20. Estructura de la IgG. V: dominio variable, C: dominio constante.
La especificidad inmunológica de una molécula de anticuerpo está determinada por su habilidad para unirse específicamente a un epitopo en particular. El sitio de unión, es decir el surco de unión al antígeno, se localiza en el extremo amino terminal de la molécula. Las regiones variables, tanto de las cadenas H como L, están constituidas por más de 100 aminoácidos, dentro de las cuales se encuentran los dominios hipervariables denominados
Antígeno
Cadena Liviana (L)
Cadena Pesada (H)
Fab
Fc

UNIVERSIDAD JUAN AGUSTÍN MAZAFACULTAD DE CIENCIAS VETERINARIAS Y AMBIENTALESCÁTEDRA DE VIROLOGÍA Y MICOLOGÍA VETERINARIA
regiones determinantes complementarias intercaladas entre las cuatro regiones conservadas, denominadas regiones marco. Cuando los péptidos se pliegan para formar la estructura tridimensional de la IgG, las 6 regiones determinantes complementarias (3 en la cadena L y 3 en la cadena H) forman parte del surco de unión al antígeno (Figura 21). Esto permite comprender la variabilidad de las regiones determinantes complementarias que explica el acotado rango de epitopos diferentes reconocidos por estas moléculas.
Figura 21. Estructura tridimensional de la IgG.
Los anticuerpos dirigidos contra ciertos epitopos de la superficie del virión neutralizan la infectividad de esa partícula viral. Estos anticuerpos pueden actuar como opsoninas, facilitando la ingesta y destrucción de los viriones por los macrófagos (Figura 12). Además los anticuerpos pueden atacar a los antígenos virales sobre la superficie de las células infectadas, generando su destrucción seguido de la activación de las vías clásica (Figura 25) y alternativa (Figura 26) del complemento o mediante la activación de los receptores de unión a la fracción Fc localizados en las células NK (Figura 22), leucocitos polimorfonucleares y macrófagos; constituyendo la denominada citotoxidad mediada por células dependiente de anticuerpos.
Estructura tridimensional de la IgG
Surco de unión del antígeno
Cadenas L
Cadenas H

UNIVERSIDAD JUAN AGUSTÍN MAZAFACULTAD DE CIENCIAS VETERINARIAS Y AMBIENTALESCÁTEDRA DE VIROLOGÍA Y MICOLOGÍA VETERINARIA
Figura 22. Citotoxidad mediada por células dependiente de anticuerpos. La unión de la partícula viral expresada en las células infectadas a los anticuerpos IgG, genera la activación de los receptores FCɣR (o CD16) localizados en la superficie de las células NK y su unión a la fracción Fc de la IgG. Esto desencadena una respuesta lítica de las células NK sobre la célula blanco infectada con virus.
Inmunoglobulina G. La principal inmunoglobulina del suero es la IgG, la cual se clasifica en diferentes sub-clases: IgG1, IgG2, IgG3 e IgG4. Luego de una infección viral sistémica, la IgG continúa siendo sintetizada por los linfocitos B durante años y es la principal molécula implicada en la protección contra las reinfecciones. Las diferentes sub-clases de IgG difieren en la región constante de sus cadenas H y por lo tanto en las propiedades biológicas, como por ejemplo la fijación del complemento y la unión a fagocitos.
Inmunoglobulina M. La IgM es un tipo importante de inmunoglobulina. Es un pentámero equivalente a 5 IgG, con 10 fracciones Fab y 10 sitios diferentes de unión a antígeno (Figura 23). Debido a que la IgM se produce al comienzo de una infección viral y luego es reemplazada por la producción de IgG, estos anticuerpos IgM son indicativos de una infección reciente. Niveles bajos de IgM pueden ser encontrados en la segunda mitad de la gestación, cuando el feto comienza a desarrollar su sistema inmune. Ya que la IgM no puede atravesar la placenta, debido su gran tamaño, la presencia de IgM en el animal recién nacido pueden indicar una infección viral intrauterina.

UNIVERSIDAD JUAN AGUSTÍN MAZAFACULTAD DE CIENCIAS VETERINARIAS Y AMBIENTALESCÁTEDRA DE VIROLOGÍA Y MICOLOGÍA VETERINARIA
Figura 23. Inmunoglobulina M. Pentámero equivalente a 5 inmunoglobulinas G, con 10 fracciones Fab y 10 sitios diferentes de unión a antígeno.
Inmunoglobulina A. La IgA es un dímero compuesto por 4 fracciones Fab. Al atravesar las células epiteliales, la IgA adquiere el fragmento J (también llamado fragmento secretorio) y se transforma en IgA secretoria (Figura 24.A). Esta es secretada hacia la luz epitelial del tracto respiratorio, digestivo y genitourinario. La IgA secretoria es más resistente a las proteasas que cualquier otra inmunoglobulina. Es la principal inmunoglobulina en las superficies mucosas y, en algunas especies animales, en el calostro y la leche. Por estas razones, la IgA es un anticuerpo importante para generar resistencia a las infecciones virales del aparato digestivo, respiratorio y urogenital (Figura 24.B).
Figura 24. (A) Estructura de la inmunoglobulina A secretoria. (B) La IgA es la principal inmunoglobulina en las superficies mucosas, por ejemplo del tracto digestivo.
IgA (dímero secretorio)
Fragmento J
A
B

UNIVERSIDAD JUAN AGUSTÍN MAZAFACULTAD DE CIENCIAS VETERINARIAS Y AMBIENTALESCÁTEDRA DE VIROLOGÍA Y MICOLOGÍA VETERINARIA
Inmunoglobulinas D y E. La IgD e IgE son especies menores de inmunoglobulinas, representando menos del 1% del total de las inmunoglobulinas presentes en el suero. La mayor parte de la IgD se encuentra unida en la superficie de los linfocitos B, pero su función es aún desconocida. La IgE, es producida por las células plasmáticas subepiteliales del aparato respiratorio y digestivo y se une firmemente a los mastocitos donde reacciona con ciertos tipos de antígenos (alergenos). Esto estimula la liberación de mediadores (serotonina e histamina) de las respuestas inmunes tipo anafilácticas.
ComplementoEl sistema del complemento está formado por aproximadamente 30 proteínas séricas, las
cuales se activan para “complementar” la respuesta inmune. Existe una vía clásica de activación del complemento (Figura 25), la cual depende de la presencia de complejos antígeno-anticuerpo y una vía alternativa (Figura 26), que es independiente de la presencia de anticuerpos.

UNIVERSIDAD JUAN AGUSTÍN MAZAFACULTAD DE CIENCIAS VETERINARIAS Y AMBIENTALESCÁTEDRA DE VIROLOGÍA Y MICOLOGÍA VETERINARIA
Figura 25. Vía clásica del complemento. Su activación es iniciada por inmunocomplejos formados por IgG e IgM. Esta vía se inicia con la unión de o más moléculas de inmunoglobulinas unidas a los antígenos respectivos, al producirse cambios alostéricos en el extremo Fc. El factor C1 está compuesto por tres subunidades proteicas (q, r y s), que en el momento de la activación del complemento se unen entre sí, formando un complejo. La subunidad C1q se fija al anticuerpo. Los anticuerpos, para activar al complemento, han de encontrarse con la disposición espacial apropiada que permita a C1q acoplarse a varios de ellos al mismo tiempo. El fragmento C1q activa a las dos subunidades C1r, que actuará sobre las dos C1s, responsables de iniciar las fases siguientes. C1s actúa sobre C4 produciendo su escisión en dos moléculas, una pequeña, C4a, que difunde a la fase fluida y otra mayor, C4b, que se une a la superficie celular. Esta fracción C4b unida a la membrana, forma un complejo con la fracción C2. C1s también actúa sobre C2, provocando la escisión de esta molécula en dos fragmentos, uno menor C2b y otro mayor C2a. Este último se une al C4b para formar el complejo C4b2a. El complejo C4b2a, cuyo centro activo se encuentra en el componente C2a, actúa sobre el factor C3 que se transforma por proteólisis en dos fragmentos activos: la anafilotoxina C3a, que pasa al medio líquido, y el fragmento C3b que se une a la membrana celular. El factor C3b unido a la membrana celular puede ser captado por los fagocitos, quienes expresan receptores de membrana para C3b. De ésta forma se facilita el proceso de la fagocitosis (opsonización).
La activación de la vía clásica conduce a la destrucción de los viriones o de las células infectadas por virus. Los viriones son destruidos como resultado de su opsonización, neutralización o lisis de la envoltura viral. La activación del complemento luego de la interacción de los anticuerpos con las partículas virales en los tejidos, contribuye al proceso inflamatorio y a la acumulación de leucocitos.

UNIVERSIDAD JUAN AGUSTÍN MAZAFACULTAD DE CIENCIAS VETERINARIAS Y AMBIENTALESCÁTEDRA DE VIROLOGÍA Y MICOLOGÍA VETERINARIA
Figura 26. Vía alternativa del complemento. Cuando C3 soluble (C3(H2O)) se une a la superficie de una partícula viral envuelta, forma un complejo con el Factor B, el cual se fragmenta por acción del factor D en presencia de Mg++. El complejo C3Bb soluble (C3Bb(H2O)) actúa enzimáticamente sobre moléculas adicionales de C3, amplificando la cascada. C3b se puede unir a la C3 convertasa. Esta convertasa libera más factor C3b que formar C3bBb3b, retroalimenta el circuito y consigue su amplificación. El complejo C3bBb3b actúa sobre C5 e iniciar la vía lítica que lleva a la lisis de los viriones.
La activación del complemento por la vía alternativa ocurre principalmente cuando las células son infectadas con virus envueltos, quienes maduran por gemación a través de la membrana citoplasmática de la célula hospedadora. Por lo tanto, no se requieren de anticuerpos para la activación de la vía alternativa.
Memoria inmunológica
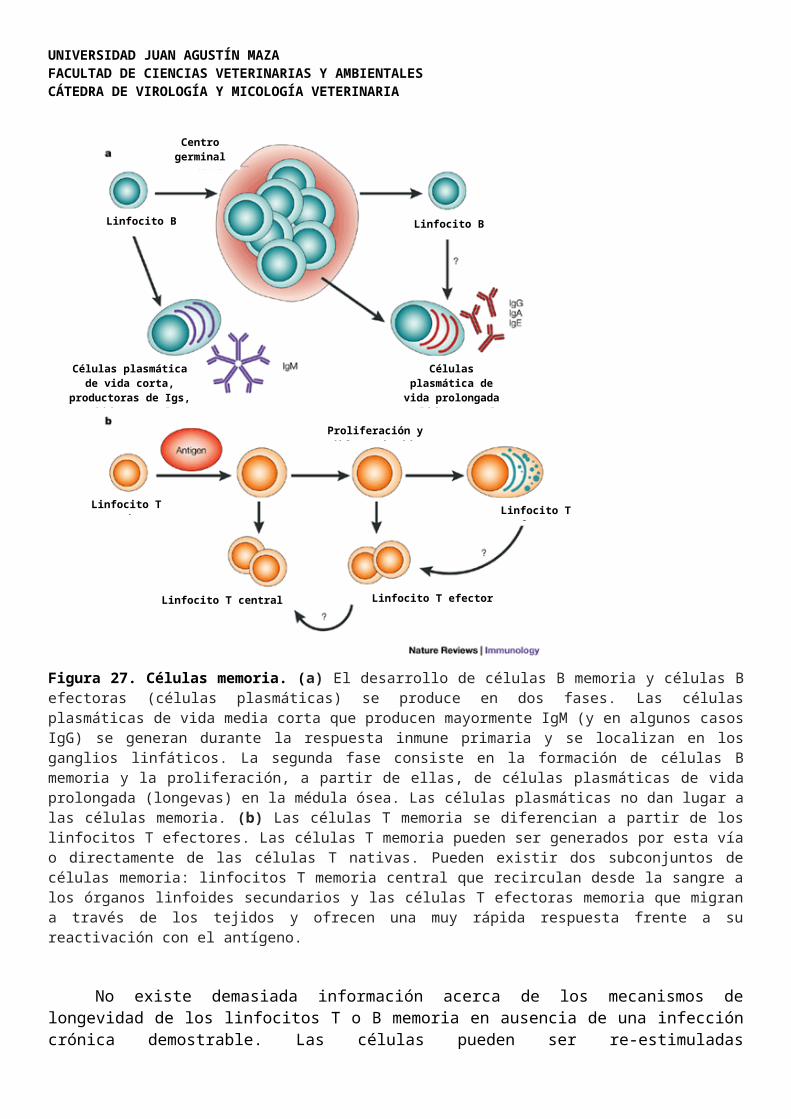
Luego de la estimulación antigénica y expansión clonal de los linfocitos (Figura 1), surge una población persistente de linfocitos memoria. Los linfocitos T memoria se caracterizan por poseer moléculas de superficie particulares (CD45RO) y adhesinas que permiten la recirculación de estas células. Cuando el organismo se re-expone al mismo antígeno, incluso muchos años después del primer contacto, los linfocitos T memoria responden más rápida e intensamente que durante ese primer encuentro. Los linfocitos B memoria también juegan un rol importante en la respuesta amnésica (memoria) y reaccionan produciendo mayores cantidades de anticuerpos específicos (Figura 27).
Envoltura viral

UNIVERSIDAD JUAN AGUSTÍN MAZAFACULTAD DE CIENCIAS VETERINARIAS Y AMBIENTALESCÁTEDRA DE VIROLOGÍA Y MICOLOGÍA VETERINARIA
Figura 27. Células memoria. (a) El desarrollo de células B memoria y células B efectoras (células plasmáticas) se produce en dos fases. Las células plasmáticas de vida media corta que producen mayormente IgM (y en algunos casos IgG) se generan durante la respuesta inmune primaria y se localizan en los ganglios linfáticos. La segunda fase consiste en la formación de células B memoria y la proliferación, a partir de ellas, de células plasmáticas de vida prolongada (longevas) en la médula ósea. Las células plasmáticas no dan lugar a las células memoria. (b) Las células T memoria se diferencian a partir de los linfocitos T efectores. Las células T memoria pueden ser generados por esta vía o directamente de las células T nativas. Pueden existir dos subconjuntos de células memoria: linfocitos T memoria central que recirculan desde la sangre a los órganos linfoides secundarios y las células T efectoras memoria que migran a través de los tejidos y ofrecen una muy rápida respuesta frente a su reactivación con el antígeno.
No existe demasiada información acerca de los mecanismos de longevidad de los linfocitos T o B memoria en ausencia de una infección crónica demostrable. Las células pueden ser re-estimuladas periódicamente por el péptido antigénico original retenido durante largos periodos de tiempo en los CMH expresados en las células dendríticas de los folículos linfoides. Los linfocitos T y B memoria pueden sobrevivir muchos años sin dividirse, hasta que son re-estimulados en la siguiente re-infección.
Respuesta inmune frente a la infección viral
Linfocito B nativo
Centro germinal
Linfocito B memoria
Células plasmática de vida corta, productoras de Igs, proliferan en los
tejidos linfoides.
Células plasmática de vida prolongada
proliferan en la médula ósea
Linfocito T nativo Linfocito T efector
Proliferación y diferenciación
Linfocito T efector memoriaLinfocito T central memoria

UNIVERSIDAD JUAN AGUSTÍN MAZAFACULTAD DE CIENCIAS VETERINARIAS Y AMBIENTALESCÁTEDRA DE VIROLOGÍA Y MICOLOGÍA VETERINARIA
Un resumen de las principales características de la respuesta inmune frente a una infección viral aguda se ilustra en la Figura 1. Como se observa, al menos tres fenómenos contribuyen a la recuperación del organismo frente a una infección viral: 1) destrucción de las células infectadas, 2) producción de interferones y 3) neutralización de la infectividad de los viriones. Inicialmente luego de la infección viral, algunas partículas virales son fagocitadas por los macrófagos. A excepción de algunos virus que son capaces de crecer dentro de los macrófagos, los viriones fagocitados son destruidos. Las proteínas de la cápside de estos viriones son degradadas a péptidos que se expresan en los CMH-II sobre la superficie de los macrófagos. Estos péptidos antigénicos expresados en los CMH-II son reconocidos por los linfocitos Th1 (CD4+). Estos linfocitos responden mediante proliferación clonal y liberan citoquinas, las cuales atraen monocitos sanguíneos al sitio de infección e inducen la proliferación y diferenciación de estas células a macrófagos activados. Esto constituye la respuesta inmune básica. Los linfocitos Th2 responden secretando un gran número de citoquinas que generan clones de linfocitos B. Estos linfocitos B reconocen el antígeno viral, lo endocitan y posteriormente se diferencian a células plasmáticas. Los linfocitos Tc se activan luego del reconocimiento de los péptidos virales expresados en los CMH-I sobre la superficie de las células infectadas. La respuesta de los linfocitos Tc alcanza su pico tras 1 semana del comienzo de la infección, mientras que la producción de anticuerpos por las células plasmáticas sucede luego de 2 a 3 semanas de ocurrida la infección. La actividad de las células NK es máxima a los 2 días post-infección y la actividad pico de los interferones coincide con la entrada de las partículas virales al organismo.
La síntesis de anticuerpos sucede principalmente en el bazo, linfonódulos, tejido linfoide asociado al intestino y en el tejido linfoide asociado a los bronquios. El bazo y los linfonódulos reciben los antígenos virales a través de la circulación sanguínea o linfática, respectivamente. Los primeros anticuerpos en generar son las IgM y luego todas las subclases de IgG. Sin embargo, el tejido linfoide submucoso del tracto respiratorio y digestivo, así como también en las tonsilas y en placas de Peyer, reciben los antígenos virales que atravesaron las células epiteliales y generan la producción local de IgA, principalmente.
Citólisis de las células infectadas por virus.La destrucción de las células infectadas es una necesidad fundamental para lograr la
recuperación del organismo de la infección viral. Esto resulta de 4 maneras diferentes: 1) por acción de los linfocitos Tc, 2) mediante el proceso de citotoxicidad mediado por el complejo anticuerpo-complemento, 3) mediante el proceso de citotoxicidad mediado por células independiente de anticuerpos y 4) por acción de las células NK. Debido a que algunos péptidos virales se expresan en la membrana plasmática antes que se liberen los viriones desde la célula hospedadora, la lisis de estas células impiden la liberación de una progenie de viriones infectivos.
La citotoxicidad mediada por el complejo anticuerpo-complemento sucede aún frente a muy escasas concentraciones de anticuerpos. La activación de la via alternativa del complemento parece tener una importancia particular frente a este fenómeno. La citotoxicidad mediada por células independiente de anticuerpos es desarrollada por aquellos leucocitos (macrófagos, polimorfonucleares y otros tipos de células asesinas) que poseen receptores para la fracción Fc de los anticuerpos sobre sus membranas celulares. Sin embargo, las células NK son activadas directamente por el interferón o por las glicoproteínas virales. Esto demuestra una especificidad no inmunológica para la lisis de las células infectadas con virus. Además, en presencia de anticuerpos, los macrófagos pueden fagocitar y digerir a las células infectadas con virus.
Neutralización de la infectividad viral

UNIVERSIDAD JUAN AGUSTÍN MAZAFACULTAD DE CIENCIAS VETERINARIAS Y AMBIENTALESCÁTEDRA DE VIROLOGÍA Y MICOLOGÍA VETERINARIA
A diferencia de los linfocitos T, los linfocitos B y los anticuerpos generalmente reconocen epitopos que son conformacionales, es decir, epitopos que son relativamente variables en su composición aminoacídica, que se localizan en la superficie de la proteína antigénica y que se presentan como protuberancias (Figura 28). La conformación particular de un epitopo viral es lo genera diferencias antigénicas entre las diferentes cepas de virus.
Figura 28. Los retrovirus poseen glicoproteínas ancladas en la envoltura viral. Esta glicoproteína viral posee diferentes epitopos sobre su superficie, contra los cuales los linfocitos B producen anticuerpos neutralizantes.
Los anticuerpos específicos pueden unirse a cualquier epitopo accesible ubicado sobre la superficie de las proteínas del virión, pero sólo aquellos anticuerpos que se unen con una elevada afinidad a un epitopo en particular sobre las proteínas de la cápside o de la envoltura viral son capaces de neutralizar efectivamente al virus. La proteína viral involucrada es aquella que se comporta como ligando para los receptores ubicados en la superficie de las células hospedadoras. Las mutaciones que ocurren en los epitopos de las proteínas virales permiten a los virus escapar de la neutralización por anticuerpos y la emergencia gradual de las mutaciones en la mayoría de estos epitopos genera la aparición de nuevas cepas virales.
La neutralización no hace referencia sólo a la cobertura del virión con anticuerpos y al bloqueo del ataque del virus contra la célula hospedadora. Los viriones neutralizados por anticuerpos pueden unirse y atacar a la célula hospedadora. En estos casos, el bloqueo generado por la neutralización viral ocurre en algún punto entre la adherencia del virión y su entrada a la célula hospedadora. Una hipótesis es que los anticuerpos que recubren al virión ingresan a la célula junto con el virión, mediante su incorporación en vesículas endociticas. Las enzimas de los lisosomas que se fusionan con estas vesículas endocíticas se encargan de destruir los anticuerpos que bloquean al virión, liberando al mismo al espacio citoplasmático. En el caso de los picornavirus, los anticuerpos neutralizantes aparentan distorsionar la cápside viral, generando
Retrovirus
Envoltura viral
GLICOPROTEINA VIRAL
Superficie de la proteína
Epitopos sobre la superficie de la proteína viral

UNIVERSIDAD JUAN AGUSTÍN MAZAFACULTAD DE CIENCIAS VETERINARIAS Y AMBIENTALESCÁTEDRA DE VIROLOGÍA Y MICOLOGÍA VETERINARIA
la pérdida de proteínas particulares de dicha cápside, lo cual susceptibiliza al virión frente al ataque enzimático. Los virus Influenza pueden sufrir sutiles cambios conformacionales en las hemaglutininas de la envoltura viral lo cual puede evitar la fusión viral que precede a la liberación de la nucleocápside.
Recuperación de la infección viral
La inmunidad mediada por células, los anticuerpos, el complemento, los fagocitos, los interferones y las citoquinas están involucrados en la recuperación del organismo frente a una infección viral. La mayoría de estos elementos del sistema inmune actúan en conjunto.
Rol de los linfocitos T Los linfocitos y los macrófagos predominan en el infiltrado celular de los tejidos infectados
con virus, a diferencia del predominio polimorfonuclear que se observa en las infecciones bacterianas de los tejidos.
Si bien los determinantes antigénicos reconocidos por los linfocitos T y los epitopos sobre la superficie de la proteína viral reconocidos por los linfocitos B muchas veces son las mismas moléculas, el determinante antigénico mayormente reconocido por los linfocitos T se sitúa sobre las proteínas localizadas en el interior del virión o sobre las proteínas no estructurales del virión. Por lo tanto las respuestas de los linfocitos T son generalmente más amplias que las respuestas de los anticuerpos neutralizantes y puede desencadenar una reactividad cruzada entre los serotipos o cepas virales.
Rol de los anticuerposEn las infecciones virales generalizadas que se caracterizan por una viremia importante y
donde una gran cantidad de viriones circulan libres en el plasma, los anticuerpos circulantes juegan un rol muy importante en la recuperación.
Aunque existe bastante evidencia que los anticuerpos juegan un rol importante en la infección con picornavirus, togavirus, flavivirus y parvovirus; esto no implica que los anticuerpos actúen solamente neutralizando las partículas virales. No existe demasiada información en medicina veterinaria acerca del rol exacto de las inmunoglobulinas en la recuperación del organismo frente a una infección viral.
Inmunidad frente a las reinfecciones
Mientras un gran número de fenómenos complejos de interacción contribuyen a la recuperación del organismo frente a una infección viral, los mecanismos de la inmunidad adquirida frente a reinfecciones con el mismo virus son mucho más simples de comprender. La primera línea de defensa la constituyen los anticuerpos. Si estos fueron adquiridos por infección activa con un virus que causan una enfermedad sistémica, continúan siendo sintetizados durante años, proveyendo de una firme protección contra las reinfecciones. El grado de inmunidad adquirida se correlaciona muy bien con los títulos de anticuerpos séricos. Además, la transferencia de anticuerpos ya sea mediante inmunización artificial pasiva (inyección de anticuerpos) o mediante la transferencia materna de anticuerpos al feto (tranferencia placentaria)

UNIVERSIDAD JUAN AGUSTÍN MAZAFACULTAD DE CIENCIAS VETERINARIAS Y AMBIENTALESCÁTEDRA DE VIROLOGÍA Y MICOLOGÍA VETERINARIA
o al animal recién nacido (transferencia calostral) provee de una excelente protección en la mayoría de las infecciones virales. Por todo esto, se puede concluir que los anticuerpos son el factor más influyente en la adquisición de inmunidad ya sea por infección natural o por vacunación. Si la producción de anticuerpos frente a una infección viral es inadecuada, los mecanismos que intervienen en la recuperación de la infección viral vuelven a predominar.
Como regla general, la respuesta de la IgA secretoria es muy corta en comparación con la respuesta de la IgG sérica. Debido a esto, la duración de la resistencia a las reinfecciones con virus respiratorios o digestivos es muy limitada. Por ejemplo, es muy común la reinfección con el mismo serotipo del virus parainfluenza. Además, no existe protección cruzada para los diferentes serotipos de un virus, por lo que estos serotipos pueden repetir el ataque al aparato respiratorio a lo largo de la vida del animal.
La respuesta inmune a la primera infección de un virus puede tener una gran influencia en las posteriores respuestas frente a virus antigénicamente similar, de manera que el segundo virus atacante induce una respuesta que es dirigida principalmente contra los antígenos de la cepa viral original. Por ejemplo, la respuesta humoral frente a la infección secuencial con diferentes cepas del virus Influenza A está dirigida hacia los determinantes antigénicos de la primera cepa que infectó a ese individuo. Este fenómeno es conocido como pecado original antigénico y también ha sido observado en las infecciones producidas por enterovirus, reovirus, paramyxovirus y togavirus. El pecado original antigénico tiene importantes consecuencias para la interpretación de los datos seroepidemiológicos, para la comprensión de fenómenos inmunopatológicos y especialmente para la generación de estrategias de vacunación eficientes.
Inmunidad pasiva
Existe una gran cantidad de información que demuestra la eficacia de los anticuerpos en prevenir las infecciones virales. Por ejemplo, la inmunización artificial pasiva (inyección de anticuerpos) protege temporalmente contra la infección del virus del moquillo canino, panleucopenia felina, cólera porcino y muchas otras infecciones. Además, la inmunización natural pasiva, es decir la transferencia de anticuerpos maternales al feto (transferencia transplacentaria) o al recién nacido (transferencia calostral), protege al animal en los primeros meses de vida contra la mayoría de las infecciones que su madre ha experimentado.
Inmunidad natural pasivaLa inmunidad natural pasiva es importante por dos razones:
1) es esencial para la protección de los animales jóvenes durante sus primeras semanas a meses de vida. Esta protección se establece contra una gran cantidad de microorganismos, incluyendo virus, presentes en el ambiente donde el animal nació.
2) la interferencia de los anticuerpos maternales con los anticuerpos producidos por inmunización activa (vacunación) del animal recién nacido, lo cual debe tomarse en cuenta al momento de diseñar el esquema de vacunación.
- Transferencia de anticuerpos maternos. Los anticuerpos maternos pueden ser transmitidos a través de la yema del huevo en las aves, a través de la placenta en los primates o mediante el calostro y/o leche en otros mamíferos. Las diferentes especies de mamíferos difieren

TEJIDO FETAL
TEJIDO MATERNO
TEJIDO CONECTIVO
VASO SANGUÍNEO
TROFOECTODERMO
EPITELIO UTERINO
VASO SANGUÍNEO
TEJIDO CONECTIVO
EPITELIOCORIAL
UNIVERSIDAD JUAN AGUSTÍN MAZAFACULTAD DE CIENCIAS VETERINARIAS Y AMBIENTALESCÁTEDRA DE VIROLOGÍA Y MICOLOGÍA VETERINARIA
enormemente en la ruta que predomina para la transferencia de anticuerpos maternos, dependiendo de la estructura de la placenta (Tabla 1 y Figura 29).
Tabla 1. Transferencia de inmunidad natural pasiva en los mamíferos.
Especies
Tipo de placentació
n
Número de capas
placentariasTransferenci
a prenatal (vía
placentaria)
Transferencia posnatal
(vía digestiva)
Tiempo límite para la translocació
n (días)Materna
Fetal
Bovina Porcina Equina
Epiteliocorial 3 3 0 +++ 2
OvinaCaprina
Sindesmocorial 2 o 3 3 0 +++ 2
CaninaFelina
Endoteliocorial 1 2 o 3 ± +++ 2
RatónRata
Hemocorial 0 3 ++ + 16-20
Figura 29. Tipos de placentación.
TEJIDO FETAL
TEJIDO FETAL
TEJIDO MATERNO
TEJIDO MATERNO
TEJIDO CONECTIVO
VASO SANGUÍNEO
TROFOECTODERMO
VASO SANGUÍNEO
TEJIDO CONECTIVO
ENDOTELIOCORIAL
MEMBRANA INTERSTICIAL
TEJIDO CONECTIVO
VASO SANGUÍNEO
TROFOECTODERMO
SANGRE MATERNA
HEMOCORIAL
UNIVERSIDAD JUAN AGUSTÍN MAZAFACULTAD DE CIENCIAS VETERINARIAS Y AMBIENTALESCÁTEDRA DE VIROLOGÍA Y MICOLOGÍA VETERINARIA
En ratones y ratas, donde la circulación materna y la fetal están separadas por relativamente pocas capas placentarias, los anticuerpos IgG (no los IgM) pueden atravesar la placenta y la inmunidad materna es transmitida principalmente por esta ruta. Sin embargo, la placenta de la mayoría de los animales domésticos es más compleja (formada por 5 a 6 capas) y se hipotetiza que constituye una barrera para las Igs, incluso la IgG. En estas especies, la inmunidad materna es trasmitida al animal recién nacido mediante el calostro, y en mucha menor cantidad, a través de la leche.
Las diferentes especies animales difieren en la clase o sub-clase de inmunoglobulina que transfieren al recién nacido a través del calostro (Tabla 2), aunque la mayoría de las especies domesticas transfieren principalmente IgG.
Tabla 2. Concentración de inmunoglobulinas IgG, IgA e IgM en el calostro y leche de algunas especies de mamíferos.
Especies
Concentración de Inmunoglobulinas (g/L)
Calostro Leche
IgG IgA IgM IgG IgA IgM
Bovina 36-77ª 4-5 3,2-4,9 1,0-1,8 0,2 0,04
Porcina 62 10 3,2 1,4 3,0 1,9
Equina 80 9 4 0,35 0,8 0,04
Canina 2,0 13,5 0,3 0,01 3,6 0,06
Humana 0,3 120 1,2 0,1 1,5 0,01
ªLos valores subrayados indican los principales componentes.
En la vaca y la oveja se produce una transferencia selectiva de IgG1 desde el suero al epitelio alveolar mamario durante las últimas semanas de gestación, de manera que el nivel de IgG1 en el calostro se eleva a 40 - 70 g/L, comparado con el nivel de 1,0 - 1,8 g/L hallados en la leche y los 13 g/L del suero. Los anticuerpos de la sub-clase IgG1 son importantes en la protección contra las infecciones entéricas durante el periodo de lactancia.
La transferencia selectiva de IgG desde la circulación materna al epitelio alveolar mamario se debe a la fracción Fc de esta molécula. Las grandes cantidades de IgG presentes en el calostro son translocadas en grandes vesículas intracitoplasmáticas por células especializadas presentes en la porción anterior del intestino delgado para alcanzar luego la circulación del recién nacido.

UNIVERSIDAD JUAN AGUSTÍN MAZAFACULTAD DE CIENCIAS VETERINARIAS Y AMBIENTALESCÁTEDRA DE VIROLOGÍA Y MICOLOGÍA VETERINARIA
Pequeñas cantidades de otras inmunoglobulinas (IgM e IgA) presentes en el calostro o la leche, en algunas especies, pueden ser translocadas a través del epitelio intestinal, pero desaparecen rápidamente de la circulación del animal recién nacido. El periodo post-nacimiento durante el cual los anticuerpos ingeridos con el calostro son traslocados (denominado tiempo límite para la translocación) está claramente definido y es muy breve (48 horas) en la mayoría de los animales domésticos (Tabla 1).
En las aves sucede una transferencia selectiva de IgG desde la circulación materna (el nivel de IgG en la yema del huevo es de 25 g/L comparado con los 6 g/L en la circulación materna). Una gallina produce aproximadamente 100 g de IgG por año para ser trasferida a la yema, lo cual es mucho mayor a la cantidad que produce para cubrir sus propias necesidades. La IgG ingresa en la circulación vitelina y se encuentra disponible para el embrión a los 12 días de incubación. Algunas IgG son también transferidas al fluido amniótico y son ingeridas por el embrión. Cerca del momento de la eclosión, el saco vitelino con el remanente de las inmunoglobulinas maternas ingresa a la cavidad abdominal del embrión y es incorporada a la pared intestinal del embrión.
Los anticuerpos maternos en la circulación del recién nacido son destruidos muy rápidamente. La vida media es de 21 días para los terneros y potrillos, 8 a 9 días para los cachorros caninos y felinos y tan sólo 2 días para los ratones. Por lo tanto, el animal recién nacido estará protegido contra las infecciones por un virus en particular solamente si su madre contiene anticuerpos IgG específicos. Tal protección será más prolongada (mayor a la vida media de la molécula de IgG en circulación neonatal) si la madre posee un gran título de estos anticuerpos específicos.
Aunque los niveles de IgA transferidos vía calostro al intestino del animal recién nacido son considerablemente menores a los niveles de IgG, estos anticuerpos IgA son importantes para la protección del neonato frente a virus entéricos contra los cuales la madre haya desarrollado inmunidad. Por otra parte, existe evidencia que luego de transcurrido el tiempo límite para la translocación de inmunoglobulinas, hay transferencia de IgA (principlamnete) y de IgG e IgM a través de la leche para continuar protegiendo al neonato contra las infecciones virales entéricas. El recién nacido se expone permanentemente a los virus contra los cuales está parcialmente protegido. Bajo estas circunstancias los virus pueden replicar, aunque en menor medida, debido en parte, al desarrollo temprano de la respuesta inmune del neonato, no ocasionando una enfermedad grave. Esto sucede ya que el neonato desarrolla la inmunidad activa adquirida en la medida que se encuentra parcialmente protegido por la inmunidad materna.
-Falla en la transferencia de anticuerpos maternos. La falla completa o parcial en la transferencia de anticuerpos maternos es la patología más frecuente que ocasiona inmunodeficiencia en los animales domésticos. Por ejemplo, entre el 10 – 40% de los terneros lactantes y más del 20% de los potrillos no reciben niveles adecuados de anticuerpos maternos. La mortalidad durante el periodo neonatal, debida a enfermedades respiratorias y digestivas, es superior a la mortalidad presente en cualquier otro periodo de la vida del animal y se correlaciona con la falla en la transferencia de anticuerpos maternos. Las razones biológicas que explican esta falla en la transferencia son:
1) nacimiento prematuro de animales débiles,
2) demora en el comienzo de la ingesta de calostro,
3) muerte de la madre,
4) baja producción de calostro por la madre,
5) bajos niveles de anticuerpos en el suero materno y por lo tanto en el calostro,

UNIVERSIDAD JUAN AGUSTÍN MAZAFACULTAD DE CIENCIAS VETERINARIAS Y AMBIENTALESCÁTEDRA DE VIROLOGÍA Y MICOLOGÍA VETERINARIA
6) escaso instinto materno, principalmente en las madres primerizas, con abandono de la cría,
7) lactancia prematura,
8) camada muy numerosa,
9) dominancia de las crías más vigorosa, que impiden calostrar a las crías más débiles.
Entre todas estas razones, los factores más importantes son la cantidad de calostro disponible y las demoras entre el nacimiento de la cría y su primera succión de calostro. El mal manejo de los animales juega un rol importante, ya que puede influir generando condiciones antinaturales al momento del parto y demoras en permitir la ingesta temprana de calostro por la cría. La inmunización materna (vacunación) para generar protección de los animales recién nacidos es una buena estrategia en la práctica médica veterinaria.
BIBLIOGRAFÍA:
Murphy FA, Gibbs EP, Horzinek MC, Studdert MJ. (1999). Veterinary Virology. Third Edition, Elsevier Ed., U.S.A. Traducción realizada por el Dr. Cs. Vet. Diego Grilli (Octubre, 2013).