ectopic expression of hoxa-1in the zebrafish alters the...
TRANSCRIPT

735Development 122, 735-746 (1996)Printed in Great Britain © The Company of Biologists Limited 1996DEV2028
Ectopic expression of Hoxa-1 in the zebrafish alters the fate of the mandibular
arch neural crest and phenocopies a retinoic acid-induced phenotype
Daniel Alexandre1,†, Jonathan D. W. Clarke2, Elli Oxtoby3, Yi-Lin Yan4, Trevor Jowett3 and Nigel Holder1,*1Developmental Biology Research Centre, Randall Institute, King’s College, 26-29 Drury Lane, London WC2B 5RL, UK2Department of Anatomy and Developmental Biology, University College, Windeyer Building, Cleveland Street,London, W1P 6DB, UK3Department of Biochemistry and Genetics, Medical School, University of Newcastle upon Tyne, NE2 4HH, UK4Department of Biology, University of Oregon, Eugene, Oregon 97403, USA
*Author for correspondence (email: [email protected])†Author’s current address: Laboratoire de Neurogénétique du Développement, Université Montpellier II, C.C. 103, Place Eugène Bataillon, 34095 Montpellier Cedex 5,France
Considerable evidence has demonstrated that retinoic acidinfluences the formation of the primary body axis in ver-tebrates and that this may occur through the regulation ofHox gene expression. In this study, we show that thephenotype induced by exogenous retinoic acid in thezebrafish can also be generated by the overexpression ofHoxa-1 following injection of synthetic RNA into the fer-tilised egg. The isolation, sequence and expression patternof the zebrafish Hoxa-1 gene is described. We show thatexogenously applied retinoic acid causes the ectopic accu-mulation of Hoxa-1 message during gastrulation in the
hypoblast in the head region. Overexpression of Hoxa-1following injection of RNA causes abnormal growth of theanterior hindbrain, duplication of Mauthner neurons inrhombomere (r) 2 and fate changes of r2 mesenchymal andneurogenic neural crest. These results are discussed interms of the role of Hoxa-1 in controlling anteriorhindbrain patterning and the relationship betweenexpression of Hoxa-1 and retinoic acid.
Key words: Hoxa-1, neural crest, pharyngeal arches, retinoic acid,homeosis
SUMMARY
INTRODUCTION
Genetic studies, principally in Drosophila and in mouse, haveshown that Hox genes are key regulators of positional infor-mation during development (see reviews by McGinnis andKrumlauf, 1992; Krumlauf, 1994). Loss-of-function alleles inthe mouse, for example, generate either homeotic transforma-tions or abnormal development of the region of the embryoassociated with the anterior limit of normal expression of thegene (eg Mark et al., 1993; Rijli et al., 1993; Ramirez-Solis etal., 1993). Homeotic changes are also a consequence of over-expression of a number of different members of the vertebrateHox clusters (eg Kessel et al., 1990; Lufkin et al., 1992; Zhanget al., 1994). The homeotic changes primarily affect thesegmental organisation of the primary body axis such as theformation of inappropriate vertebrae at specific levels (Kesselet al., 1990; Jegalian and DeRobertis, 1992; Ramirez-Solis etal., 1993) or alterations to specific skull bones (Lufkin et al.,1992; Rijli et al., 1993; Gendron-Maguire et al., 1993). It isclear, therefore, that the Hox genes are important in the estab-lishment of the primary body axis. This is best illustrated bythe mouse Hoxa-1 gene which has been studied both by over-expression (Zhang et al., 1994) and by loss-of-functionmutation (Lufkin et al., 1991; Chisaka et al., 1992; Mark et al.,1993; Carpenter et al., 1993) and has been shown to be a key
regulator of the normal formation of the hindbrain. Overex-pression leads predominantly to the conversion of rhombomere(r) 2 to r4 and results in the alteration in expression of a numberof other regulatory genes, including Hoxb-1 and Hoxa-2. Lossof function causes loss of r5 and most of r4.
In addition to the Hox genes, the putative developmental sig-nalling molecule, retinoic acid (RA), has also been implicatedin the patterning of the primary body axis in vertebrates. RAis present during gastrulation in the mouse and the frog (Hoganet al., 1992; Chen and Solursh, 1994) and RA, exogenouslyapplied from the early stages of gastrulation, affects patterningof the anterior-posterior axis in zebrafish (Holder and Hill,1991; Hill et al., 1995), Xenopus (Durston et al., 1989; Sive etal., 1990; Papalopulu et al., 1991) and mammals (Morriss,1972; Morriss-Kay et al., 1991; Kessel and Gruss, 1991;Conlon and Rossant, 1992; Marshall et al., 1992; Simeone etal., 1995). In such experiments, one consistent effect of RA isto alter the formation of the hindbrain, where the segmentalrhombomeres are to some extent posteriorised (Papalopulu etal., 1991; Marshall et al., 1992; Hill et al., 1995). Exogenouslyapplied RA leads to the alteration of expression of a numberof regulatory genes in the hindbrain (Morriss-Kay et al., 1991;Conlon and Rossant, 1992; Marshall et al., 1992) but it remainsunclear how the phenotype is generated. A number of lines ofevidence indicate that Hoxa-1 is a potential target for RA. As

736 D. Alexandre and others
the most 3′ in the Hoxa cluster, it is most sensitive to inductionby RA in NT2 cells (Simeone et al., 1990) and an RA responseelement has been identified in the Hoxa-1 gene that isnecessary for RA induction (Langston and Gudas, 1992). It isalso expressed from the onset of gastrulation (Murphy and Hill,1991), which, as mentioned above, is the period in theembryonic process when formation of the primary body axisis most sensitive to exogenous RA treatment The link betweenthe expression and function of Hoxa-1 and the endogenous roleof RA is further strengthened by similarities in aspects of thephenotypes generated by either Hoxa-1 overexpression orexogenous supply of RA to the embryo. This has been shownin the mouse by Marshall et al. (1992) who demonstrated thatRA causes a transformation of r2 into r4, a result that ismirrored in mouse embryos ectopically expressing Hoxa-1(Zhang et al., 1994). Similarly, RA and ectopic Hoxa-1 bothlead to altered expression of Hoxb-1 and Hoxa-2 in thehindbrain (Conlon and Rossant, 1992; Kessel, 1993; Zhang etal., 1994).
In previous studies, we have characterised the effect ofexogenous RA on the development of the zebrafish hindbrain(Holder and Hill, 1991; Hill et al., 1995). Following the appli-cation of a relatively low concentration of RA, a precisephenotype is generated in the anterior hindbrain. The limits ofthe effect are the caudal midbrain rostrally and the r4/5 bordercaudally and include, in some respects comparable to themouse (Marshall et al., 1992), the partial transformation of r2to r4. For the reasons outlined above, we wished to analyse theexpression and function of Hoxa-1 in the zebrafish in order toestablish whether it is the likely target gene for exogenouslyapplied RA and whether alteration in its expression is partly orwholly responsible for the characteristic RA-induced hindbrainphenotype. To this end, we have isolated and characterised thezebrafish Hoxa-1 cDNA and show that its expression patternis essentially that seen in the mouse. The expression of the geneis affected by exogenously applied RA from the onset of gas-trulation. We analyse the function of Hoxa-1 by RNA injectioninto the fertilised egg and characterise a reliably generatedphenotype that includes alterations to the anterior rhom-bomeres and to the anterior hindbrain neural crest.
The neural crest in the head can be divided into two broadpopulations, the neurogenic precursors, which will differen-tiate into neurons associated with the peripheral head ganglia,and the mesenchymal precursors, which migrate to the pha-ryngeal arches and differentiate into Schwann cells, pigmentcells and cartilage and connective tissue cells (review by LeDouarin et al., 1994). In the head of the zebrafish, the neu-rogenic neural crest is a region of cells located most laterallyin the dorsal crest primordium (Schilling and Kimmel, 1994)and these authors have shown that neural lineage restrictionoccurs prior to migration. The mesenchymal crest lies moremedially in the premigratory mass and is also restricted interms of eventual fate prior to migration. The phenotype thatwe describe, which has not been observed previously in com-parable Hoxa-1 overexpression experiments (Zhang et al.,1994), is an alteration of the pharyngeal skeleton whereby aderivative of the first arch, the mandible, is not formed andthe second arch-derived hyoid cartilage is enlarged. Thisalteration is linked to abnormal migration of the mandibulararch crest. This is coupled with a fusion of cranial ganglia Vand VII. The phenotype generated by Hoxa-1 overexpression
in zebrafish is very similar to that produced by exposure toRA.
MATERIALS AND METHODS
(A) Cloning of the zebrafish Hoxa-1 geneA polymerase chain reaction was performed on zebrafish genomicDNA using primers based on the 5′ and 3′ ends of the labial-likehomeobox of murine Hoxa-1 and Hoxb-1. The 5′ primer wasCTGCAGCGCACCAACTTCACCACNAA(A/G)CA (lab+) and the 3′primer was CTCGAGCTCGCGCTCGCGCTTCTT(T/C)TG(T/C)TT(lab−). 0.5 µg of zebrafish genomic DNA was amplified in 100 µlreaction containing 50 mM KCl, 10 mM Tris-HCl pH 9.0 at 25°C,1.5 mM MgCl2, 0.01% gelatin, 0.1% Triton X-100, 0.2 mM eachdNTPs, 0.5 µg of each primer and 2.5 units of Taq polymerase(Promega). After an initial denaturation of 5 minutes at 94°C, thirtyrounds of amplification were performed, a single cycle being 2minutes at 55°C, 3 minutes at 72°C and 2 minutes at 94°C. A finalextension reaction was performed for 10 minutes at 72°C. The gelpurified products were treated with polynucleotide kinase and thenKlenow before blunt-end ligating into the SmaI site of BluescriptKS−. Seven plasmids were identified as having the same homeoboxsequences and these sequences were used to design primers for per-forming 5′ and 3′ RACE (Frohman and Martin, 1989).
For 3′ RACE, the initial reverse transcription was performed in 50mM Tris-HCl, pH 8.15 at 41°C, 6 mM MgCl2, 40 mM KCl, 1 mMDTT containing 1 mM of each dNTP, 10 units of RNasin, 1 µg ofzebrafish poly(A)+ RNA and 2.5 pmole of the 57-mer (dT)17 -R1-R0primer (Frohman and Martin, 1989) with 10 units of avian myeloblas-tosis virus reverse transcriptase (AMV-RT) for 2 hours. The reactionmixture was diluted to 1 ml with TE (10 mM Tris-HCl pH 8.0, 1 mMEDTA). For first round amplification, 5 µl of the diluted cDNA poolwas mixed with 25 pmol each of a gene-specific primer (3H29A inFig. 1) and R0 (Frohman and Martin, 1989) in a 50 µl PCR cocktailwith the same ingredients as in the original genomic DNA reaction.After an initial denaturation step of 95°C for 7 minutes, the reactionwas cooled to 70°C and 2.5 units of Taq polymerase added, then thirtycycles of 94°C for 2 minutes, 50°C for 2 minutes and 72°C for 3minutes performed with a final extension of 15 minutes at 72°C. Thereaction was diluted 1:20 with TE and a second amplificationperformed with 1 µl of the diluted DNA and 25 pmol each of a secondnested gene-specific primer (3H29B in Fig. 1) and R1 (Frohman andMartin, 1989). The conditions for the amplification were otherwisethe same as for the first round above. A single fragment was amplifiedand this was gel purified, treated with kinase and Klenow before sub-cloning into the SmaI site of Bluescript KS−. Before subcloning theamplified fragment, we confirmed that it was as expected by makinga 10−6 dilution of the purified fragment and showing that it could bereamplified with the 3H29C and R1 primers.
For 5′ RACE, the initial reverse transcription was the same as forthe 3′ reaction except it incorporated 1 pmole of a gene-specificprimer (5H29A in Fig. 1) instead of the 57-mer. The cDNA reactionwas diluted to 1 ml with TE and the excess primer was removed ona Centricon 100 (Amicon Corp.) ultrafiltration spun column. The finalretenate was concentrated to 10 µl before dATP tailing with terminaldeoxynucleotide transferase (BRL) and then diluting to 500 µl withTE. The first round amplification involved 5 µl of diluted tailed cDNAand 25 pmoles each of 5H29A and R0 with an additional 2 pmol ofthe 57-mer in a 50 µl reaction volume. Denaturation followed by thirtycycles was as for 3′ RACE. Second round amplification was with 1µl of a 1:20 dilution of the first reaction and 25 pmol each of a secondnested gene-specific primer (5H29B in Fig. 1) and R1. Amplificationconditions were as for the previous reaction. The final reaction yieldedthree major amplification bands. Before subcloning the amplifiedfragments, we confirmed that they were derived from the same gene

737Ectopic expression of Hoxa-1
uence of the zebrafish Hoxa-1 gene. This is derived from both genomicribed in the Materials and Methods. The positions of the primers usedeactions used to amplify the homeobox region (lab+ and lab−), for 3′ 3H29-C) and 5′RACE (5H29-A, 5H29-B, 5H29-C). The 89 base pairegions are underlined. Putative translations of the two exons, the intronown below the corresponding DNA sequence. The 3′ end of the
olyadenylation site for the transcript. The restriction sites are those usedg the full-length cDNA construct for in vitro transcription.
by making a 10−6 dilution of the purified fragments and showing thatthey were reamplified with the 5H29C and R1 primers. All three werepurified and subcloned as previously described and on sequencingthey were shown to be subsets of the same reverse transcriptionreaction. It would appear that the reverse transcriptase quite frequentlyfell off the RNA template prematurely thereby generating two smallercDNAs.
Primers H16(72) and H16(833), in Fig. 1, were used to PCR ampli-fications of the 3′RACE cDNA pool and also from genomic DNA.Conditions for amplification were thesame as for that with the lab+ and lab−primers. The amplified product derivedfrom the genomic DNA was slightlylarger than the cDNA product.Sequencing of the two amplifiedfragments revealed a single 89 base pairintron in the genomic sequence. ThecDNA amplification product was usedas a probe to a zebrafish genomiclibrary in lambda DASH® II (Strata-gene). A single clone was selected thathad the region of homology in themiddle of the insert. The insert wassubcloned into Bluescript SK− andrestriction mapped. The region ofhomology with the cDNA clone waswithin a single 1335 bp SinI fragment(Fig. 1). This SinI fragment was gelpurified and, after addition of BamHIlinkers, was subcloned into the BglIIsite of the transcription vector pSP64T(Krieg and Melton, 1984). The intronwas removed by excising the XcmI-BssHII fragment (Fig. 1) and replacingit with the equivalent fragment takenfrom the cDNA clone produced byamplification with primers H16(72) andH16(833). The resulting plasmid,zebrafish Hoxa-1 intronless(SinI)BamHI/pSP64T, was used togenerate full-length, capped,polyadenylated transcripts with SP6RNA polymerase. A negative controlfor injections was made by takingthe zebrafish Hoxa-1-intronless(SinI)BamHI/pPSP64T plasmid, lin-earising with XcmI, blunt-ending thefragment with T4 DNA polymerase andrecircularising with T4 ligase. Thischanges the open reading frame suchthat the SP6 RNA polymerase transcriptcontains a stop codon just into thehomeobox region and thus will producea truncated Hoxa-1 polypeptide thatlacks the homeodomain.
(B) Injection of mouse and fishHoxa-1 RNA The mouse Hoxa-1 cDNA p480encoding the 331 amino acid proteinwas excised with EcoRI and PstI andsubcloned initially into Bluescript KS−.BamHI was used to transfer the geneinto the BglII site of the pSP64T vectorand the orientation was checked byrestriction mapping. A negative controlconstruct was produced by removing
Fig. 1. Genomic DNA seqclones and cDNAs as descfor the polymerase chain rRACE (3H29-A, 3H29-B,intron and the homeobox rand primer locations are shgenomic sequence is the pin subcloning and in makin
most of the homeobox by cutting with HincII and BglII. The endswere then blunt ended with mung bean nuclease and ligated producingan in frame fusion which was checked by sequencing.
RNA synthesis, purification and injection into fertilised eggs of 300to 500 pl of RNA at a concentration of 100 µg/ml was carried out asdescribed by Griffin et al. (1995). Prior to injection, all RNAs weretested by in vitro translation (Boehringer Mannheim) according tomanufacturers recommendations. Translation reactions wereseparated by SDS-PAGE and visualised by autoradiography.

738 D. Alexandre and others
(C) Maintenance of fish and RA treatmentBreeding fish were maintained at 28.5°C on a 14 hour light/10 hourdark cycle. Embryos were collected by natural spawning and raisedin aquarium water at 28.5°C and staged according to Westerfield(1995). RA treatment was also performed according to previouslydescribed protocols (Holder and Hill, 1991); in the current study, RAwas given at a concentration of 10−7 M. All exposures were at 50%epiboly, the onset of gastrulation (Westerfield, 1995) for 1 hour at28.5°C.
(D) Antibody and cartilage staining3A10, an antibody originally raised in a screen for rat floor plate tissueby Dr Jane Dodd, recognises exclusively the Mauthner neuron in thezebrafish at the stages used in this study (Hatta, 1992). The anti-acety-lated tubulin and 4D9 antibodies were purchased commercially. Thestaining procedures were those described previously (Holder and Hill,1991). For Alcian blue staining, 4 day old larvae were fixed in 4%paraformaldehyde, rinsed in PBS and stained for 2 hours in 0.01%Alcian blue in 70% ethanol/30% glacial acetic acid. They were thendestained in the acidic alcohol, rinsed once with 40% ethanol in PBSthen in PBS alone and cleared in 0.5% potassium hydroxide for a fewminutes. Specimens were then rinsed and mounted in glycerol.
(E) Whole-mount in situ hybridisationFor single colour in situ mRNA probes to zebrafish krx-20 (Oxtobyand Jowett, 1993), pax 2 (Krauss et al., 1991), dlx 2 (Akimenko et al.,1994) and Hoxa-1 were prepared and used as described by Oxtobyand Jowett (1993). For two colour in situ hybridisation, krx-20 waslabelled with fluorescein and dlx-2 with DIG and detected accordingto the methods described by Jowett and Lettice (1994) as modified byHauptmann and Gerster (1994) except that glycine treatment wasextended to 40 minutes.
(F) Retrograde labelling of reticulospinal neurons4 day embryos were immobilised in 1% low melting point agarose inPBS. The spinal cord was completely transected at the level of thehindgut with a sharpened tungsten needle. The dye (lysinatedrhodamine dextran, Molecular Probes D-1817) was then applied as athick semi-solid with another sharpened tungsten needle to the rostralstump of the cut spinal cord. The embryo was then released into asolution of 10% Hank’s saline for 2 hours to allow transportation ofthe dye. The embryo was then fixed in 4% paraformaldehyde in PBSovernight at 4°C.
The brains from labelled embryos were then carefully dissected freeof surrounding tissue and mounted ventral side uppermost in 90%glycerol/10% PBS on a glass microscope slide, under a glass coverslipsupported by four spots of silicon gel. Permanent mounts were paintedaround the edge of the coverslip with nail varnish to prevent evapo-ration.
Confocal images were viewed on a Leica confocal microscopeusing standard Leica software.
RESULTS
Cloning of the zebrafish homologue of murineHoxa-1We initially used primers complementary to the conservedends of the labial-like homeobox of the murine Hoxa-1 andHoxb-1 genes to perform the polymerase chain reaction onzebrafish genomic DNA. They amplified a 173 bp DNAfragment which when subcloned and sequenced proved to bea DNA species with the homeobox sequence shown in Fig. 1.Although the primers would have been expected to producetwo different labial-like sequences corresponding to Hoxa-1
and Hoxb-1, we only obtained the one type of sequence in 10different clones. We then designed sets of nested primers basedon the sequence of the zebrafish homeobox for use in 5′ and3′ Rapid Amplification of cDNA Ends, RACE (Frohman andMartin, 1989). The starting material for the reactions waspoly(A)+ RNA from 12-24 hour old zebrafish embryos. The 3′RACE produced a single amplified DNA species with thesequence from 3H29B to the 3′ end of the sequence in Fig. 1.The 5′ RACE amplification gave three differently sizedfragments which proved to be derivatives of a single 742 bpfragment with the sequence from the 5H29B primer to base137 in Fig. 1. (Note that these clones also lacked the intronsequence shown in Fig. 1).
We concluded that both the cDNA sequences were derivedfrom the same transcript for the following reasons. Theyhybridised to identically sized transcripts in northern blots ofthe original poly(A)+ RNA population (not shown). They alsogave similar patterns of hybridisation in Southern blots ofgenomic DNA cut with different restriction enzymes (notshown). We designed primers from opposite ends of eachcDNA (H16(72) and H16(833)) and used them to amplifyDNA from a reverse transcribed RNA pool and they gave asingle DNA species with a sequence that was predicted byjoining our original two overlapping cDNAs.
Amplification of genomic DNA using the same two primersgave a slightly larger than predicted fragment, which onsequencing proved to have an 89 bp intron located at the valinecodon upstream from the start of the homeobox (Fig. 1). Theposition of this intron corresponds to the location of the intronsfound in Hoxa-1 (LaRosa and Gudas, 1988), Hoxb-1 (Frohmanet al., 1990) and Hoxd-1 (Frohman and Martin, 1992a) and thechick Hoxb-1 gene (Sundin et al., 1990; Sundin and Eichele,1990) just upstream of the homeobox.
From the translation of the open reading frame harbouringthe homeobox, it appeared that the largest of the 5′RACEclones lacked the start of the protein encoding open readingframe which would be essential for making full-length tran-scripts for injection into embryos. We therefore screened azebrafish genomic DNA library with the cDNA clone andobtained a lambda clone which harboured the region ofhomology in the middle of the insert. The insert was subclonedinto Bluescript SK− and restriction mapped. Analysis of thisclone revealed that the transcribed region was within a 1335base pair SinI fragment. This was subcloned into the in vitrotranscription vector pSP64T and the intron was removed byreplacing the XcmI-BssHII fragment, which harboured theintron with the equivalent fragment from the cDNA clone. Thisplasmid, Hoxa-1 intronless (SinI)BamHI/pSP64T, is suitablefor generating full-length polyadenylated transcripts forinjection into embryos.
A comparison of the putative zebrafish polypeptide showsthat the protein most closely resembles that encoded by theHoxa-1 gene (Fig. 2). There is considerable homology withinthe homeobox and also around the conserved hexapeptideregion (WMKVKR) characteristic of many other members ofthe Hox family (Duboule et al., 1988). The sequence compar-ison alone did not allow us to unambiguously assign thezebrafish gene as being homologous to Hoxa-1.
The zebrafish Hoxa-1 gene maps to the middle of linkagegroup 12 (Gates and Postlethwait, unpublished observation -see Postlethwait et al., 1994).

739Ectopic expression of Hoxa-1
Expression pattern of Hoxa-1 in whole mountsTo confirm our designation of the zebrafish gene as thehomolgue of the murine Hoxa-1 and not Hoxb-1 or Hoxd-1,we performed in situ hybridisations on embryo whole mounts.The original probe used was a mixture of two digoxigenin-labelled antisense RNAs made from the RACE cDNAsubclones into Bluescript. When used separately these probesboth gave the same pattern of signals. (Both probes containpart of the homeobox region; under our conditions of hybridi-sation, this did not cause any problems of cross-hybridisationwith other homeobox-containing transcripts.)
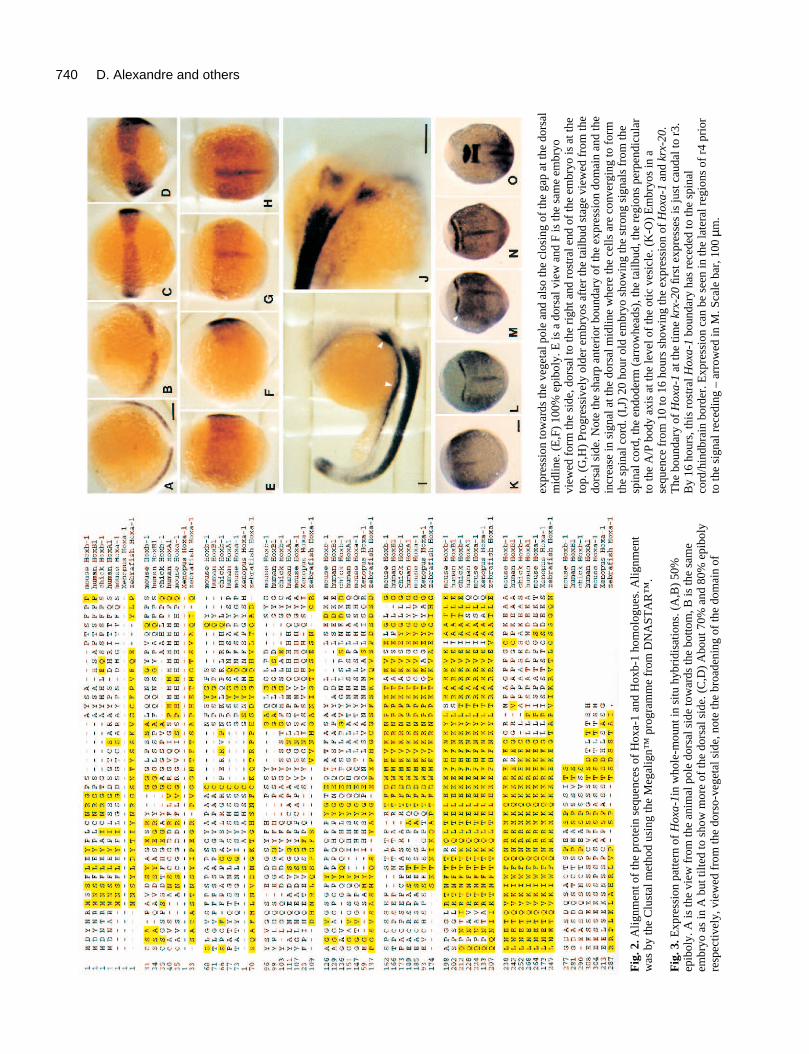
The first appearance of Hoxa-1 is at 50% epiboly as the germring forms (Fig. 3A,B). It produces a semicircle of stain whichis broken in the middle and is located just above the margin.It is centred around the localised thickening of the germ ringknown as the shield, at the dorsal surface of the gastrula. Asthe cells migrate towards the vegetal pole over the surface ofthe yolk and begin to involute, the domain of expressionbroadens. The upper boundary nearest the animal pole remainsrelatively sharp and the gap in the region of the shield beginsto close (Fig. 3C,D). At the tailbud stage, the Hoxa-1 domainis quite broad and indistinct but has a clear anterior border (Fig.3E-H). As cells converge towards the dorsal surface, a stripeappears at the midline. This is probably caused by the con-centration of cells as they gather along the embryonic A/P axisrather than an increase in expression. As somitogenesis pro-gresses, the Hoxa-1 signal is present in the spinal cord and inthe endoderm (Fig. 3I-J). Endodermal expression has a moreanterior boundary than that in the spinal cord. This expressionpattern is consistent with the gene being Hoxa-1 rather thanHoxb-1 since the latter gene in other vertebrates shows adiscrete domain of expression in rhombomere 4 (this designa-tion assumes an expression pattern for fish labial-like Hoxgenes that is similar to that found in other vertebrates).Similarly it is not likely to be the homologue of Hoxd-1because this gene is expressed only in the mesoderm (Frohmanand Martin, 1992b).
In order to assess the anterior border of expression of Hoxa-1 during the neural plate stages, double in situs with Hoxa-1and krx-20 were performed between 10 and 16 hours (Fig. 3K-O). At the time that the anterior (r3) stripe of krx-20 first appearsthere is a gap between it and the anterior boundary of Hoxa-1.The second krx-20 (r5) stripe then appears at the boundary ofHoxa-1, although some Hoxa-1-expressing cells are alsopresent in the lateral regions of r4 (see Fig. 3M). As the cellsconverge on the dorsal midline, the rostral boundary of Hoxa-1 recedes caudally to approximately the boundary between thehindbrain and spinal cord. These results indicate that the mostanterior extent of Hoxa-1 is probably within r4 or at the r3/r4boundary. It is difficult to be certain of this because the Hoxa-1 expression domain may already be receding by the time thatkrx-20 is initially expressed and the expression of Hoxa-1 in r4is only clear in the lateral regions (Fig. 3M).
The effect of RA on expression of Hoxa-1If the effect of RA on hindbrain development is linked to Hoxa-1, it is assumed that RA will alter the expression of the gene.To see if this is the case, embryos were treated with RA at theonset of gastrulation in a manner that is known subsequentlyto cause abnormal development of the anterior hindbrain(Holder and Hill, 1991). Treated and control embryos (treated
with DMSO only) were then fixed at times through gastrula-tion, tail bud and early somite stages and hybridised withHoxa-1 probe under identical conditions. The results, shownin Fig. 4, indicate an upregulation of Hoxa-1 within an hour oftreatment as compared with controls. By 80% epiboly (Fig.4A), in comparison to an identically treated control, Hoxa-1 issignificantly upregulated around the germ ring and expressesin the axial midline in the hypoblast and not in the epiblast(Fig. 4A-F). The increased level of expression in the hypoblastmaintained through tail bud and early somite stages when theectopic expression can be seen to be in the prechordalmesoderm (Fig. 4C-F). In addition, it appears with respect tothe midbrain pax 2 stripe that the expression of Hoxa-1 in theectoderm is not ectopic, the boundary being at the samelocation as in controls (Fig. 4B,C).
Assessing the function of Hoxa-1 by injection ofRNARNA encoding the mouse or zebrafish Hoxa-1 protein wasinjected into the fertilised egg or in both blastomeres of the 2-cell embryo. The majority of the analysis was performed onembryos injected with the mouse wild-type and control RNA,which was lacking the homeobox. In addition, the zebrafishHoxa-1 RNA was used for the analysis of crest derivatives, thevisceral skeleton and cranial ganglia. In general, the survivalrates of injected embryos with either wild-type or control RNAwas close to 100% at 24 hours. After this time, the mortalityof embryos injected with the wild-type RNA increases to reachabout 30% by 5 days. In contrast, the embryos injected withcontrol RNA show no increased mortality compared to unin-jected fish.
In order to assess the degree of mosaic distribution of theinjected RNA, in situ hybridisation was performed using aprobe to the transcribed flanking globin sequences which aregenerated by the pSP64T vector used to make injectedzebrafish Hoxa-1. Following injection into the single cell egg,by the 32-/64-cell stage, the RNA is present in groups of cellswhich vary in size from 10 to 50% of the total blastomeres withthe majority of embryos showing a quarter of blastomerespositive for the RNA (Fig. 5). At this stage, the groups of cellstended to be together rather than mixed with cells not con-taining the RNA. As assessed by in situ hybridisation, thisectopic expression persists at least to tail bud stages whenexpressing cells are present in the derivatives of the epiblastand hypoblast. Distribution of expression over the embryo atthis stage is extensive and random with expressing cells presentin all regions of the embryo (data not shown).
The phenotype of embryos injected with Hoxa-1RNAFor the overall analysis of function by overexpression, a totalof not less than two thousand eggs were injected with Hoxa-1RNA. For a typical experiment, injections were performed onabout 200 embryos and were repeated at least once. Thephenotype described below was never observed in embryosinjected with a control RNA. For illustrative purposes, thespecimens were chosen to show the most complete phenotype,which occurred in approximately 20% of injected embryos.Due to mosaicism, in the remaining injected embryos, the phe-notypes were generally restricted to one side or were presentdorsally and not ventrally.
740 D. Alexandre and others
Fig
. 2.A
lignm
ent o
f th
e pr
otei
n se
quen
ces
of H
oxa-
1 an
d H
oxb-
1 ho
mol
ogue
s. A
lignm
ent
was
by
the
Clu
stal
met
hod
usin
g th
e M
egal
ign™
prog
ram
me
from
DN
AST
AR
™.
Fig
. 3. E
xpre
ssio
n pa
ttern
of
Hox
a-1i
n w
hole
-mou
ntin
situ
hyb
ridi
satio
ns. (
A,B
) 50
%ep
ibol
y. A
is th
e vi
ew f
rom
the
anim
al p
ole
dors
al s
ide
tow
ards
the
botto
m, B
is th
e sa
me
embr
yo a
s in
A b
ut ti
lted
to s
how
mor
e of
the
dors
al s
ide.
(C
,D)
Abo
ut 7
0% a
nd 8
0% e
pibo
lyre
spec
tivel
y, v
iew
ed f
rom
the
dors
o-ve
geta
l sid
e, n
ote
the
broa
deni
ng o
f th
e do
mai
n of
expr
essi
on to
war
ds th
e ve
geta
l pol
e an
d al
so th
e cl
osin
g of
the
gap
at th
e do
rsal
mid
line.
(E
,F)
100%
epi
boly
. E is
a d
orsa
l vie
w a
nd F
is th
e sa
me
embr
yovi
ewed
for
m th
e si
de, d
orsa
l to
the
righ
t and
ros
tral
end
of
the
embr
yo is
at t
heto
p. (
G,H
) Pr
ogre
ssiv
ely
olde
r em
bryo
s af
ter
the
tailb
ud s
tage
vie
wed
fro
m th
edo
rsal
sid
e. N
ote
the
shar
p an
teri
or b
ound
ary
of th
e ex
pres
sion
dom
ain
and
the
incr
ease
in s
igna
l at t
he d
orsa
l mid
line
whe
re th
e ce
lls a
re c
onve
rgin
g to
for
mth
e sp
inal
cor
d. (
I,J)
20
hour
old
em
bryo
sho
win
g th
e st
rong
sig
nals
fro
m th
esp
inal
cor
d, th
e en
dode
rm (
arro
whe
ads)
, the
tailb
ud, t
he r
egio
ns p
erpe
ndic
ular
to th
e A
/P b
ody
axis
at t
he le
vel o
f th
e ot
ic v
esic
le. (
K-O
) E
mbr
yos
in a
sequ
ence
fro
m 1
0 to
16
hour
s sh
owin
g th
e ex
pres
sion
of
Hox
a-1
and
krx-
20.
The
bou
ndar
y of
Hox
a-1
at th
e tim
e kr
x-20
first
exp
ress
es is
just
cau
dal t
o r3
.B
y 16
hou
rs, t
his
rost
ral H
oxa-
1bo
unda
ry h
as r
eced
ed to
the
spin
alco
rd/h
indb
rain
bor
der.
Exp
ress
ion
can
be s
een
in th
e la
tera
l reg
ions
of
r4 p
rior
to th
e si
gnal
rec
edin
g –
arro
wed
in M
. Sca
le b
ar, 1
00 µ
m.

741Ectopic expression of Hoxa-1
Fig. 4. Retinoic acid causes the ectopic expression ofHoxa-1. (A-C) Pairs of embryos that have hybridised toprobes to Hoxa-1 and pax 2 under indenticalconditions. The embryos on the right of each pair havebeen treated with RA on the onset of gastrulation. Atthis dose of RA, Hoxa-1 is ectopically expressed by70% epiboly (A) in the axial midline (black arrow) andaround the germ ring as seen from the animal pole. Bythe 1-somite stage (B - tilted animal pole view) pax 2(white arrow) is first expressed in the prospectivemidbrain and, by this stage, the rostral limit of theHoxa-1 signal in the treated embryos is no furtheranterior than in the control. A small patch ofectopically expressed Hoxa-1 lies anteriorly (blackarrow). (C) The same embryo as seen in B but viewedlaterally with anterior to the top and dorsal to the right.The white arrow indicates the normal pax 2 expressionand the black arrow the ectopic Hoxa-1 expressionwhich is in the prechordal mesoderm. (D-F) A series ofhigh power views of optical sections through themidline of treated embryos showing ectopic expressionof Hoxa-1 only in the hypoblast. (D) Ectopicexpression is evident at the late shield stage.(E) Ectopically expressing cells are seem to be in the
hypoblast. (F) The same embryo as that shown in B and C. Ectopic expression is present in the prechordal mesoderm. The arrow indicatesthe anterior extent of the normal expression of Hoxa-1 in the ectoderm. pcm, prechordal mesoderm; h, hypoblast; e, ectoderm. Scale bar,100 µm.
(1) General appearanceExternally the injected embryos developed normally throughgastrulation and subsequently formed the main parts of thebody axis. Under the dissecting microscope, approximately70% of the injected embryos showed abnormal developmentof the anterior hindbrain in that the neural tube in this regionappeared twisted to one side or expanded (Fig. 6 and seebelow). The remaining CNS, anterior notochord and hatchinggland were normal. The only other obvious abnormalitybecame evident after several days of development at the mostanterior end of the body when the lower jaw was abnormallyformed in 77% of embryos that survived to 5 days (Fig. 8F).
(2) Development of the hindbrainThis was assessed in injected embryos and controls using aprobe for zebrafish krx-20, the antibody 3A10 to locate theMauthner neurons and by analysing the pattern of neurons inthe reticulospinal complex by neuronal tracing using lysinatedrhodamine dextran. krx-20 is normally strongly expressed inr3 and r5 at 20 hours of development (Oxtoby and Jowett,1993); therefore, injected and control embryos were analysedat this time. Control embryos invariably showed the normalpattern of two stripes (Fig. 6A). In contrast, the experimentalembryos showed a dramatic alteration in expression in r3 but
Fig. 5. Embryo at the 132-cell stage hybridised with aprobe to pSP64T vectorsequences followinginjection of Hoxa-1 syntheticRNA into the fertilised egg.The RNA distributes toapproximately one third ofthe blastomeres in this case.Scale bar, 100 µm.
expression in r5 was invariably normal (Fig. 6B). The regionof r3 was distorted and appeared larger than normal. Morpho-logically the midbrain and forebrain appeared normal. In onebatch of injected embryos that showed an abnormal phenotypein the anterior hindbrain, the antibody 4D9 was used to assessthe appearance of the engrailed-expressing region of the caudalmidbrain. In all embryos, 4D9-expressing cells were present ina pattern that appeared essentially normal (data not shown)suggesting the normal development of this region of the brain.
The reticulospinal complex offers an excellent assay for thenormal development of the hindbrain because many of the cellsof this complex are individually identifiable and they arearranged in a segmental manner that reflects their rhombomeric
Fig. 6. Photomicrographs of the anterior hindbrain region of 20 hembryos hybridised with a probe for krx-20. (A) Control showingkrx-20 expressing in r3 and r5. (B,C) Examples of embryos injectedwith mouse Hoxa-1 RNA at the single cell stage. r5 is normal in bothcases but r3 expression of krx-20 is abnormal. The anterior hindbrainappears distorted; thus r3 is split dorsally in B and is laterallydisplaced in C. Ectopic regions of krx-20 expression, such as thatarrowed most anteriorly in C, were occasionally seen. Scale bar,100 µm.

742 D. Alexandre and others
origin (Kimmel et al., 1982, 1985; Metcalfe et al., 1986; Hillet al., 1995). The Mauthner cell is one of the individually iden-tifiable neurons and is characteristic of r4. It is convenientlyidentified in whole-mounted embryos by the 3A10 antibody at32 hours of development (Hatta, 1992; Hill et al., 1995).Analysis of control and Hoxa-1-injected embryos at this timeshowed the Mauthner neuron to be duplicated at more anteriorhindbrain levels (Fig. 7A,C). Without additional markers, it isdifficult to be sure of the rhombomeric identity of the dupli-cated Mauthner cell; therefore, we assessed the reticulospinalcomplex by axonal retrograde labelling. This procedure wascarried out at 4 days of development on control and experi-mental embryos. In approximately 30% of successful prepara-tions, duplication of the Mauthner cell was evident in r2 inHoxa-1-injected embryos but never in controls (Fig. 7D,E);this occurred either on one (Fig. 7B) or both sides (Fig. 7E).Duplicated Mauthner neurons were occasionally found in r4and, in a single case, a Mauthner cell was duplicated in r3. TheMauthner cell was the only identified neuron that we could becertain was duplicated and it is certain that when duplicated inr2 other identified cells in this rhombomere, such as the Rol 2cell, were of r2 character, making it a hybrid structure.
(3) Neural crest-derived structures The derivatives of the mesenchymal crest are altered in theexperimental embryos. This is shown with respect to the devel-oping pharyngeal skeleton, which is derived from the crest cellsin each pharyngeal arch. It is the abnormal development of thejaw apparatus that gives the front of the larvae a truncatedappearance (Fig. 8F). To assess the phenotype, the normalventral or viscerocranium and dorsal head skeleton or neuro-cranium was revealed by staining the cartilage with Alcian bluein 4 day old control embryos (Fig. 8A,C). In the viscerocranium(Fig. 8A), the first (mandibular) arch neural crest gives rise tothe Meckel’s cartilage (mandible) and the palatoquadrate(maxilla). These first arch structures form a characteristic higharch appearance and lie anterior to the shorter and thickersecond (hyoid) arch-derived ceratohyal and hyosymplectic car-tilages. More posteriorly lie a series of five branchial arch car-tilages associated with the gills. The neurocranium (Fig. 8C)differentiates at this time and consists of the posterior andmesoderm-derived parachordals, the anterior limits of which lieparallel to the anterior extent of the notochord and the moreanterior trabeculae, which are neural crest-derived (Langille andHall, 1987, 1988). At their anterior extent, the trabeculae fuseand expand to form the ethmoid plate (Fig. 8C). In experimen-tal embryos (Fig. 8B), the cartilagenous derivatives of the firstarch (Meckel’s cartilage and the palatoquadrate) are absent;however, the second arch-derived ceratohyal cartilages areformed in their normal location but are thicker and partiallyduplicated medially. The hyosymplectic and the 5 branchialarch cartilages were generally normal but were occasionallyunder developed. The neural crest-derived component of theneurocranium is also malformed; the trabeculae are notconnected to the most anteriorly positioned ethmoid plateswhich can be clearly identified by their proximity to theolfactory pits. The trabeculae (arrowed in Fig. 8B) instead curveventrally as they converge rostrally between the eyes (Fig. 8D)and come downwards to the level of the ceratohyals.
Having established that Hoxa-1 overexpression alters theanterior head skeleton, it was necessary to establish if RA has
the same effect. Alcian blue preparations were made ofembryos treated with RA at the onset of gastrulation as before.At 4 days, it is clear that RA causes a distinct and abnormaldifferentiation of the Meckel’s cartilage which is much reducedor absent. The palatoquadrate and the ceratohyals are fused(Fig. 8E) making this phenotype very similar to the jaws thatdevelop following Hoxa-1 overexpression.
The derivatives of the neurogenic crest were assessed byanti-acetylated tubulin antibody staining in injected and controlembryos at 24 hours. At this time, the cranial ganglia formidentifiable populations of cells. The trigeminal (Vth) ganglionlies behind the eye, the facial (VIIth) ganglion immediatelyanterior and at the border of the otic vesicle respectively, andthe posterior lateral line ganglion lying postotically (Fig. 9A).In wild-type embryos, the Vth ganglion crest componentcomes from the most anterior hindbrain stream whereas theVIIth ganglion is generated by the stream opposite r4. In over80% of embryos injected with wild-type zebrafish Hoxa-1RNA, the Vth and VIIth ganglia are fused forming a largestructure just anterior to the otic vesicle (Fig. 9B).
In order to assess whether this abnormal phenotype reflectedan altered pattern of migration, we performed in situs with aprobe for dlx2. Dlx2 has been shown by Akimenko et al. (1994)to be a good marker for migrating hindbrain neural crest. Incontrol embryos with between 10 and 20 somites, three streamsof crest are visible in the hindbrain region (Fig. 10A). Cells inthe most anterior of these enter the mandibular arch and cellsin the preotic (middle) stream enter the hyoid arch (Akimenkoet al., 1994). In Hoxa-1-injected embryos, only two streams areevident, the anteriormost is absent and the stream anterior tothe otic placode is larger than normal (Fig. 10B); the postoticstream is unaffected in experimental embryos. The fusedpreotic stream lies opposite r4 and stretches round the aber-rantly formed r3 as seen in embryos in which double in situshave been performed with krx-20 and dlx-2. (Fig. 10C).
DISCUSSION
Hoxa-1 overexpression following RNA injection into the fer-tilised zebrafish egg results in a defined series of abnormalitiesin the embryo focused on the developing anterior region of thehindbrain and associated neural crest-derived structures. Thelocalised region of the embryo that is affected is despite thewide distribution of the RNA throughout the embryo in cellsderived from 25-50% of blastomeres. Localised effects on theanterior hindbrain suggest that the abnormal expression patternis altering development at the anterior extent of the normalexpression domain, a result that is consistent with studies withother Hox genes, such as Hoxa-2 (Rijli et al., 1993) and Hoxb-4 (Ramirez-Solis et al., 1993). The localised effect of alteringHoxa-1 expression is consistent with the results of overex-pression and loss-of-function experiments with this gene in themouse. The phenotypes that are seen are best understood interms of the normal expression pattern of the gene. In themouse, the anterior extent of expression of Hoxa-1 is the r3/4boundary. The double labels with krx-20 show that this is likelyto be the case also for the fish although it is only clear in thelateral regions of r4. However, this expression is transient andthe gene is subsequently down-regulated in the hindbrain. Inthe mouse, loss of function of Hoxa-1 leads to abnormalities

743Ectopic expression of Hoxa-1
in the caudal hindbrain. This affects the formation of thesensory and motor neurons in this region of the neural tube andthe crest normally derived from this region of the hindbrain. Incontrast to an effect on r4-r6, overexpression of Hoxa-1 in themouse leads to alteration of the anterior hindbrain (Zhang etal., 1994). In this case, there is a transformation of r2 into r4as well as alterations to r3, and effects on expression of otherregulatory genes including Hoxb-1. However, this mouse studydid not observe any effect on the eventual fate of the neuro-genic crest-derived tissues nor was later mesenchymal crest-derived tissues in the pharyngeal region examined. The resultspresented here show that both are dramatically affected byHoxa-1 overexpression in the fish.
A number of new observations relevant to the normal functionof Hoxa-1 have come from the current study. The earliest effectthat we analysed showed an alteration to the expression of krx-20 in r3 but not r5 in contrast to the situation in the mouse whereno effect is reported on krox-20 in Hoxa-1 overexpressedembryos (Zhang et al., 1994). In experimental fish embryos, r3appeared to be enlarged causing the neural keel to becomedistorted. This excess growth suggests that Hoxa-1 may beinvolved in the control of growth within the neural plate.Another consistent phenotype is the absence of the first pharyn-geal arch skeleton where Meckel’s cartilage and the palato-quadrate fail to form. The second arch-derived ceratohyals arepartially duplicated, most commonly appearing as thickenedelements bifurcated at their medial extent. The transformationdoes not affect the more caudally located gill arch skeletal struc-tures but does affect parts of the neurocranium in which themiddle part of the trabeculae are severely malformed indicatingthat this region of the trabeculum is derived from the first archcrest. Furthermore, the most anterior region where the trabecu-lae fuse to form the ethmoid plate is unaffected suggesting aseparate crest origin for this structure. This is consistent with thefate map of the neural crest in the chick embryo (Couly et al.,1993). These results indicate that the neural crest that normallypopulates the first pharyngeal arch has to some extent beenrespecified. In the normal zebrafish embryo, the first arch neuralcrest comes predominantly from r1/2/3, whereas that populatingarch two comes from r3/4/5 (Schilling and Kimmel, 1994).
There are two possible explanations for the abnormality seenin the experimental embryos; the neural crest from the r2 regioncould either (i) enter the first arch but form hyoid rather thanmandible when they differentiate and fuse with the hyoid formedfrom the second arch or (ii) migrate incorrectly and end up inthe second arch where they are patterned accordingly and formadditional hyoid tissue. It appears from the analysis of dlx 2expression (Fig. 10) that the streams of hindbrain crest generat-ing the first and second arches in the wild type are fused into asingle large stream in the Hoxa-1-injected fish. We do not knowwhat causes this fusion of anterior hindbrain streams but thismaybe due to the abnormal development of r3, which normallylies between them. It may also be the case that the neural crestcells within the single large stream may maintain their originalanterior-posterior arrangement and do not intermingle.Similarly, we still do not know at what stage the neural crest pat-terning process occurs. Schilling and Kimmel (1994) suggestedthat crest cells are determined as to their fate (cell type) prior tomigration; however, they did not comment as to the mechanismscontrolling the eventual patterning process. It is also possiblethat RA and Hoxa-1 cause an alteration in the formation of the
arches. Such an alteration could reflect ectopic expression ofHoxa-1 in the hypoblast and could involve the differentiation ofthe pharyngeal pouches which develop from endoderm andseparate the arches. This interpretation is consistent with theobservation in rodent embryos that RA leads to the formation offused first and second arches (Goulding and Pratt, 1986) intowhich neural crest from both r2 and r4 migrate as a single streamin which r2-derived crest populates the anterior half and the r4-derived crest the posterior half (Lee et al., 1995).
Some of our results are similar while others contrast withthe results of a similar Hoxa-1 overexpression study in themouse (Zhang et al., 1994). The striking similarity is the trans-formation or partial transformation of r2 to r4. This wasassessed in our study with respect to the reticulospinal complexwhereas the mouse study analysed this aspect of patterningrelative to a characteristic population of r4 motor neurons. Itwould be very interesting in both systems to learn more of thenature of the transformation in terms of other groups of nuclei.There were specific differences however, including the absencein the transgenic mice of any effect on patterning of the Vthand VIIth ganglion, lack of fusion of the first two pharyngealarch derivatives and Krox 20 expression was normal. Theprimary data for respecification of anterior to posterior neuralcrest in the mouse depends upon alterations to expressionpatterns of patterning genes such as Hoxa-2 and Hoxb-1. Thisstudy thus presents no morphological data suggesting a respec-ification other than the r4 motor neuron population in theneural ectoderm. It is unclear why these differences exist in thetransgenics because it is likely that all cells express Hoxa-1;however, there is no indication of the amount of aberrantprotein and timing of expression. In contrast, in the zebrafish,due to the mosaic distribution of the injected RNA, embryoswill vary in their phenotype. However, it is likely that the RNAis expressed very early and is still broadly expressed during thetail bud stages and it is possible that Hoxa-1 protein isexpressed at high levels in the cells containing the RNA.
The results presented in this paper also shed further light onthe association between RA and Hoxa-1. It is evident that thephenotypes generated by either treating the early zebrafishgastrula with a pulse of RA (Hill et al., 1995) or ectopicexpression of Hoxa-1 give a phenotype that is strikingly similarsuggesting that there maybe a mechanistic link between RAand Hoxa-1 function during hindbrain patterning. It is alsoestablished that treatment with RA causes ectopic expressionof Hoxa-1. However, there are a number of unansweredquestions about this possible functional link. Thus, irrespectiveof whether Hoxa-1 expression is altered by RA treatment or byRNA injection, it is not clear when during development ectopi-cally expressed Hoxa-1 has its effect. It could be during earlygastrula stages when transcripts are evident in the shieldfollowing RA treatment (see Fig. 4A-D) or it could be at latergastrula or early neurula stages when ectopic Hoxa-1 is exclu-sively in the hypoblast (Fig. 4E-F). If it is at the former, earlier,stage it could be affecting the epiblast directly. The region ofthe epiblast in the midline where ectopic Hoxa-1 is evidentafter RA treatment is, at the early gastrula stages, fated to bemidbrain or diencephalon (Woo and Fraser, 1995). Consistentwith the possibility that Hoxa-1 may affect epiblast directly inthe gastrula is that the normal, more lateral, expression ofHoxa-1 (Fig. 3A-B) is in a region of the epiblast fated to behindbrain at 6 hours of development (Woo and Fraser, 1995).
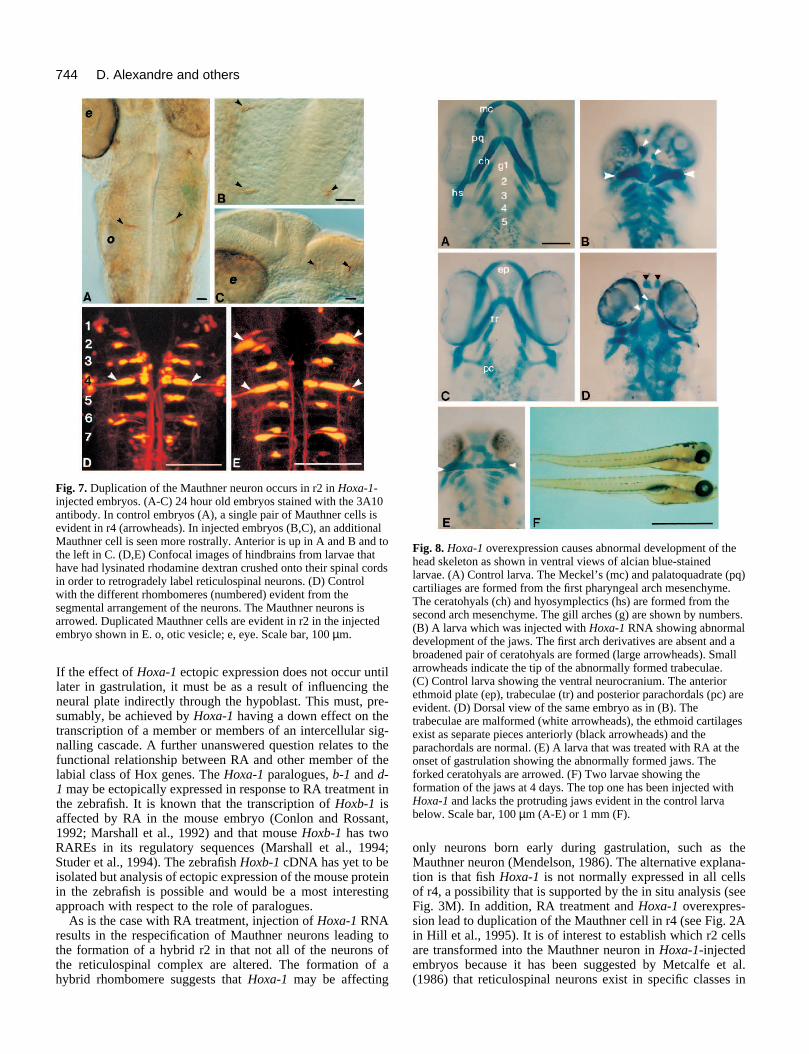
744 D. Alexandre and others
Fig. 7. Duplication of the Mauthner neuron occurs in r2 in Hoxa-1-injected embryos. (A-C) 24 hour old embryos stained with the 3A10antibody. In control embryos (A), a single pair of Mauthner cells isevident in r4 (arrowheads). In injected embryos (B,C), an additionalMauthner cell is seen more rostrally. Anterior is up in A and B and tothe left in C. (D,E) Confocal images of hindbrains from larvae thathave had lysinated rhodamine dextran crushed onto their spinal cordsin order to retrogradely label reticulospinal neurons. (D) Controlwith the different rhombomeres (numbered) evident from thesegmental arrangement of the neurons. The Mauthner neurons isarrowed. Duplicated Mauthner cells are evident in r2 in the injectedembryo shown in E. o, otic vesicle; e, eye. Scale bar, 100 µm.
Fig. 8. Hoxa-1 overexpression causes abnormal development of thehead skeleton as shown in ventral views of alcian blue-stainedlarvae. (A) Control larva. The Meckel’s (mc) and palatoquadrate (pq)cartiliages are formed from the first pharyngeal arch mesenchyme.The ceratohyals (ch) and hyosymplectics (hs) are formed from thesecond arch mesenchyme. The gill arches (g) are shown by numbers.(B) A larva which was injected with Hoxa-1 RNA showing abnormaldevelopment of the jaws. The first arch derivatives are absent and abroadened pair of ceratohyals are formed (large arrowheads). Smallarrowheads indicate the tip of the abnormally formed trabeculae.(C) Control larva showing the ventral neurocranium. The anteriorethmoid plate (ep), trabeculae (tr) and posterior parachordals (pc) areevident. (D) Dorsal view of the same embryo as in (B). Thetrabeculae are malformed (white arrowheads), the ethmoid cartilagesexist as separate pieces anteriorly (black arrowheads) and theparachordals are normal. (E) A larva that was treated with RA at theonset of gastrulation showing the abnormally formed jaws. Theforked ceratohyals are arrowed. (F) Two larvae showing theformation of the jaws at 4 days. The top one has been injected withHoxa-1 and lacks the protruding jaws evident in the control larvabelow. Scale bar, 100 µm (A-E) or 1 mm (F).
If the effect of Hoxa-1 ectopic expression does not occur untillater in gastrulation, it must be as a result of influencing theneural plate indirectly through the hypoblast. This must, pre-sumably, be achieved by Hoxa-1 having a down effect on thetranscription of a member or members of an intercellular sig-nalling cascade. A further unanswered question relates to thefunctional relationship between RA and other member of thelabial class of Hox genes. The Hoxa-1 paralogues, b-1 and d-1 may be ectopically expressed in response to RA treatment inthe zebrafish. It is known that the transcription of Hoxb-1 isaffected by RA in the mouse embryo (Conlon and Rossant,1992; Marshall et al., 1992) and that mouse Hoxb-1 has twoRAREs in its regulatory sequences (Marshall et al., 1994;Studer et al., 1994). The zebrafish Hoxb-1 cDNA has yet to beisolated but analysis of ectopic expression of the mouse proteinin the zebrafish is possible and would be a most interestingapproach with respect to the role of paralogues.
As is the case with RA treatment, injection of Hoxa-1 RNAresults in the respecification of Mauthner neurons leading tothe formation of a hybrid r2 in that not all of the neurons ofthe reticulospinal complex are altered. The formation of ahybrid rhombomere suggests that Hoxa-1 may be affecting
only neurons born early during gastrulation, such as theMauthner neuron (Mendelson, 1986). The alternative explana-tion is that fish Hoxa-1 is not normally expressed in all cellsof r4, a possibility that is supported by the in situ analysis (seeFig. 3M). In addition, RA treatment and Hoxa-1 overexpres-sion lead to duplication of the Mauthner cell in r4 (see Fig. 2Ain Hill et al., 1995). It is of interest to establish which r2 cellsare transformed into the Mauthner neuron in Hoxa-1-injectedembryos because it has been suggested by Metcalfe et al.(1986) that reticulospinal neurons exist in specific classes in

745Ectopic expression of Hoxa-1
Fig. 9. The preotic cranial ganglia are abnormally formed in Hoxa-1-injected embryos. This is seen by comparing 24 hour embryosstained with anti-acetylated tubulin to reveal the VIIth and Vthganglia between the otic vesicle (o) and the eye (e). In controlembryos (A), these ganglia are distinct but, in embryos injected withHoxa-1 (B), they are reduced and fused into one ganglionimmediately anterior to the otic vesicle. Scale bar, 100 µm.
Fig. 10. (A) A 16 somite control embryo probed for expression ofdlx2. Anterior to the left and dorsal up. Three streams of neural crestcells can be seen on the lateral regions of the neural keel two of whichlie anterior to the otic placode (op). The anteriormost stream migratesinto the mandibular arch and the preotic stream into the hyoid arch(see Akimenko et al., 1994). (B) dlx2 expression in a 16-somite-stagedembryo injected with Hoxa-1. Only one preotic stream of crest is nowvisible and it is larger than in controls. This aberrant stream of crestlies immediately anterior to the otic placode and the most anteriorstream of crest is absent. (C) The positioning of the aberrant stream isconfirmed in embryos probed for both dlx2 (blue) and krx-20. (red). Inthis 16-somite example, the single preotic stream stretches round themargin of the abnormally formed r3. Scale bar, 100 µm.
each rhombomere. It is possible therefore that cells of eachclass are specified by particular genes, such as Hox genes andoverexpression of such genes in an abnormal rhombomere willonly affect cells of that class. The association between RA andHoxa-1 makes it a possibility that localised concentrations ofRA act as the trigger for the initiation of Hoxa-1 expression inthe normal embryo. To prove this one needs to know whereRA or other retinoids are in the developing zebrafish embryoprior to gastrulation at the stage that the endogenous Hoxa-1gene is first expressed.
It is a pleasure to thank Robb Krumlauf for his help and encour-agement and for providing the mouse Hoxa-1 clone, which was orig-inally isolated by Baron et al. (1987); to Steve Wilson for his interestand comments on the manuscript and to Monte Westerfield for the dlx2 probe. This work was supported by grants from BBSRC, MRC andthe Human Frontiers Research Program to N. H., who is a BBSRCsenior research fellow and by a grants from the Wellcome Trust to T.J. and J. C.; Y. Y. was supported by grants from NIH (1P01HD22486and 1R01 RR10715) to Professor John Postlethwait. D. A. wassupported by an EMBO long-term fellowship.
REFERENCES
Akimenko, M-A., Ekker, M., Wegmer, J., Lin, W. and Westerfield, M.(1994) Combinatorial expression of three zebrafish genes related to Distal-less: Part of a homeobox gene code for the head. J. Neurosci. 14, 3475-3486
Baron, A., Featherstone, M., Hill. B., Hall, A., Galliot, B. and Duboule, D.(1987) Hox 1-6, a new mouse homeobox containing gene member of theHox-1 complex. EMBO J. 6, 2977-2086
Carpenter, E., Goddard, J., Chisaka, O., Manley, N. and Cappechi, M.(1993) Loss of Hoxa-1 (Hox1.6) function results in reorganisation of themurine hindbrain. Development 118, 1063-1075
Chen, Y. and Solursh, M. (1994) A concentration gradient of retinoids in theearly Xenopus laevis embryo. Dev. Biol. 161, 70-76
Chisaka, O., Musci, T. and Capecchi, M. (1992) Developmental defects ofthe ear, cranial nerves and hindbrain resulting from targetted disruption of themouse homeobox gene Hox 1.6. Nature 355, 516-520
Conlon, R.A. and Rossant, J. (1992). Exogenous retinoic acid rapidly inducesanterior ectopic expression of murine Hox-2 genes in vivo. Development116, 357-368.
Couly, G., Coltey, P. and Le Douarin, N. (1993) The triple origin of thevertebrate skull in higher vertebrates - A study in Quail-Chick chimaeras.Development 117, 409-429.
Duboule, D., Galliot, B., Baron, A. and Featherstone, M. S. (1988). Murinehomeo-genes: some aspects of their organisation and structure. In Cell toCell Signals in Mammalian Development (ed. S. deLaat, J. G. Blueminkand C. L. Mummery), pp. 97-108. Berlin: Springer Verlag, NATO ASISeries.
Durston, A.J., Timmermans, J., Hage, W.J., Hendriks, H.F., de Vries, N.,Heideveld, M. and Nieuwkoop, P. (1989). Retinoic acid causes ananteroposterior transformation in the developing central nervous systemNature 340, 140-144.
Frohman, M. A. and Martin, G. R. (1989). Rapid amplification of cDNA endsusing nested primers. Technique 1, 165-170.
Frohman, M. A., Boyle, M. and Martin, G. R. (1990). Isolation of the mouseHox-2.9 gene; analysis of embryonic expression suggests that positionalinformation along the anterior-posterior axis is specified by mesoderm.Development 110, 589-607.
Frohman, M.A. and Martin, G. (1992a). Isolation and spatial analysis ofHoxd-1, a new murine labial-like gene, reveals that labial subfamilymembers are expressed similarly onbly in early anteroposterior axisformation. Mech. Develop. 38, 55-58.
Frohman, M. A. and Martin, G. R. (1992b). Isolation and analysis ofembryonic expression of Hox-4.9, a member of the murine labial-like genefamily. Mech. Develop. 38, 55-67.
Gendron-Maquire, M., Mallo, M., Zhang, M. and Gridley, T. (1993) Hoxa-2 mutant mice exhibit homeotic transformation of skeletal elements derivedfrom the cranial neural crest. Cell. 75, 1317-1331

746 D. Alexandre and others
Goulding, E. and Pratt R. (1986) Isotreteoin teratogenicity in mouse wholeembryo culture. J. Craniofac. Genet. Dev. Biol. 6, 99-112
Griffin, K., Patient, R. and Holder, N. (1995) Analysis of FGF function innormal and no tail zebrafish embryos reveals separate mechanisms for theformation of the trunk and the tail. Development 121, 2983-2994
Hatta, K. (1992) Role of the floor plate in axonal patterning in the zebrafishCNS. Neuron 9, 629-642.
Hauptmann, G. and Gerster, T. (1994) Two colour whole-mount in situhybridisation to vertebrate and Drosophila embryos. Trends Genet., 10, 266.
Hill, J., Clarke, J.D.W., Vargesson, N., Jowett, T. and Holder, N. (1995)Exogenous retinoic acid causes alterations in the development of thehindbrain and midbrain of the zebrafish embryo including positionalrespecification of the Mauthner neuron. Mech. Develop. 50, 3-16.
Hogan, B., Thaller, C. and Eichele, G. (1992) Evidence that Henson’s node isa site of retinoic acid synthesis. Nature 359, 237-241
Holder, N. and Hill, J. (1991). Retinoic acid modifies development of themidbrain-hindbrain border and affects cranial ganglion formation inzebrafish embryos. Development 113, 1159-1170.
Jegalian, B. and DeRobertis, E. (1992) Homeotic transformations in themouse induced by overexpression of a human Hox3.3 transgene. Cell 71,901-910
Jowett, T. and Lettice, L. (1994) Whole mount in situ hybridizations onzebrafish embryos using a mixture of digoxigenin and flourescein labelledprobes. Trends Genet. 10, 73.
Kessel, M. (1993) Reversal of axonal pathways from rhombomere 3 correlateswith extra Hox expression domains. Neuron 10, 379-393.
Kessel, M., Balling, R. and Gruss, P. (1990) Variations of cervical vertebraeafter expression of a Hox-1.1 transgene in mice. Cell 61, 301-308
Kessel, M. and Gruss, P. (1991) Homeotic transformations of murinevertebrae and concomitant alteration of Hox codes induced by retinoic acid.Cell 67, 89-104.
Kimmel, C.B., Powell, S.L. and Metcalfe, W. (1982). Brain neurons whichproject to the spinal cord in young larvae of the zebrafish J. Comp. Neurol.205, 112-127.
Kimmel, C.B., Metcalfe, W. and Schabtach, E. (1985). T reticularinterneurons: A class of serially repeating cells in the zebrafish hindbrain. J.Comp. Neurol. 233, 365-376.
Krauss, S., Johansen, T., Korzh, V. and Fjose, A. (1991) Expression of thezebrafish paired box gene pax (zf-b) during early neurogenesis. Development113, 1193-1206
Krieg, P. and Melton, D. (1984) Funtional messenger RNAs are produced bySP6 in vitro transcription of cloned cDNAs. Nucl. Acids Res. 12, 7057-7070.
Krumlauf, R. (1994) Hox genes in vertebrate development. Cell 78, 191-201Langille, R. and Hall, B. (1987) Development of the head skeleton of the
Japanese Medaka. J. Morph. 193, 135-158.Langille, R. and Hall, B. (1988) The role of the neural crest in the development
of the cartilagenous cranial and visceral skeleton of the Medaka. Anat.Embryo. 177, 297-305
Langston, A.W. and Gudas, L. (1992) Identification of a retinoic acidresponsive enhancer 3’ of the murine homeobox gene Hox1.6. Mech. Dev.38, 217-228
LaRosa, G. J. and Gudas, L. (1988). Early retinoic acid-induced F9teratocarcinoma stem cell gene ERA-1: alternate splicing creates transcriptsfor a homeobox-containing protein and one lacking the homeobox. Mol. Cell.Biol., 8, 3906-3917.
Lee, Y., Osumi-Yamashita, N., Ninomiya, Y., Moon, C.K., Eriksson, U. andEto, K. (1995) Retinoic acid stage-dependently alters the migration patternand identity of hindbrain neural crest cells. Development 121, 825-837
LeDouarin, N., Dupin, E. and Ziller, C. (1994) Genetic and epigenetic controlin neural crest development. Curr. Biol. 4, 685-695
Lufkin, T., Dierich, A., LeMur, M., Mark, M. and Chambon, P. (1991)Disruption of the Hox1.6 homeobox gene results in defects in a regioncorresponding to its rostral domain of expression. Cell 66, 1105-1119
Lufkin, T., Mark, M., Hart, C., Dolle, P., Lemeur, M. and Chambon, P.(1992) Homeotic transformation of the occipital bones of the skull by ectopicexpression of a homeobox gene. Nature 359, 835-840
Mark, M., Lufkin, T., Vonesch, J-L., Ruberte, E., Olivio, J-O, Dolle, P.,Gorry, P., Lumsden, A. and Chambon. P. (1993) Two rhombomeres arealtered in Hoxa-1 mutant mice. Development 119, 319-338.
Marshall,H., Nonchev, S., Sham, M.H., Muchamore, I., Lumsden, A. andKrumlauf, R. (1992). Retinoic acid alters hindbrain Hox code and inducestransformation of rhombomeres 2/3 to 4/5 identity. Nature 360, 737-741.
Marshall, H., Studer, M., Popperl, H., Aparicio, S., Kuroiwa, A., Brenner,S. and Krunlauf, R. (1994) A conserved retinoic acid response element
required for early expression of the homeobox gene Hoxb-1. Nature 370,567-571
McGinnis, W. and Krumlauf, R. (1992) Homeobox genes and axialpatterning. Cell 68, 283-302
Mendelson, B. (1986). Development of reticulospinal neurons of the zebrafish.I. Time of origin. J. Comp. Neurol. 251, 160-171.
Metcalfe, W., Mendelson, B. and Kimmel, C. B. (1986) Segmentalhomologues among reticulospinal neurons in the hindbrain of the zebrafishlarva. J. Comp. Neurol. 251, 147-159
Morriss, G. (1972) Morphogenesis of the malformations induced in ratembryos by maternal hypervitiminosis A. J. Anat. 113, 241-250
Morriss-Kay, G. M., Murphy, P., Hill, R. E. and Davidson, D. (1991).Effects of retinoic acid excess on expression of Hox-2.9 and Krox-20 and onmorphological segmentation in the hindbrain of mouse embryos. EMBO J.10, 2985-2995.
Murphy, P. and Hill, R. (1991) Expression of the mouse labial-likehomeobox-containing genes, Hox 2.9 and Hox 1.6, during segmentation onthe hindbrain. Development 111, 61-74
Oxtoby, E. and Jowett, T. (1993) Cloning of the zebrafish krox-20 gene (krx-20) and its expression during hindbrain development. Nucl. Acid Res. 21,1087-1095
Papalopulu, N., Clarke, J. D. W., Bradley, L., Wilkinson, D., Krumlauf, R.and Holder, N. (1991). Retinoic acid causes abnormal development andsegmental patterning of the anterior hindbrain in Xenopus Embryos.Development 113, 1145-1158.
Postlethwait, J., Johnson, S., Midson, C., Talbot, W., Gates, M., Ballinger,E., Africa, D., Andrews, R., Carl, T., Eisen, J., Horne, S., Kimmel, C.,Hutchinson, M., Johnson, M, and Rodriguez, A. (1994) A genetic linkagemap for the zebrafish. Science 264, 699-702.
Ramirez-Solis, R., Zheng, H., Whiting, J., Krumlauf, R. and Bradley, A.(1993) Hoxb-4 (Hox-2.6) mutant mice show homeotic transformation of acervical vertebra and defects in the closure of the sternal rediments. Cell 73,279-294.
Rijli, F., Mark, M., Lakkaraju, S., Dierich, A., Dolle, P. and Chambon, P.(1993) A homeotic transformation is generated in the rostral branchial regionof the head by disruption of Hoxa-2, which acts as a selector gene. Cell 75,1333-1349
Schilling, T. and Kimmel, C. (1994) Segment and cell type lineage restrictionsduring pharyngeal arch development in the zebrafish embryo. Development120, 483-494.
Simeone, A., Acampora, D., Arcioni, L., Andrews, P., Boncinelli, E. andMavillo, F. (1990) Sequential activation of Hox2 homeobox genes byretinoic acid in human embryonal carcinoma cells. Nature 346, 763-766
Simeone, A., Avantaggiato, V., Moroni, M., Mavilio, F., Arra, C., Cotelli,F., Nigro, V. and Acamporo, D. (1995) Retinoic acid induces stage-specificantero-posterior transformation of rostral central nervous system. Mech.Develop. 51, 83-98.
Sive, H. L., Draper, B.W., Harland, R. and Weintraub, H. (1990).Identification of a retinoic acid sensitive period during primary axisformation in Xenopus laevis Genes Dev. 4, 932-942.
Sive, H. L. and Cheng, P. F. (1991) Retinoic acid perturbs the expression ofXhox.lab genes and alters mesodermal determination in Xenopus laevisGenes Dev. 5, 1321-1332
Studer, M., Popperl, H., Marshall, H., Kuroiwa, A. and Krumlauf, R.(1994) Role of a conserved retinoic acid response element in rhombomererestriction of Hoxb-1. Science 265, 1728-1732
Sundin, O. H., Busse, H. G., Rogers, M. B., Gudas, L. J. and Eichele, G.(1990). Region specific expression in early chick and mouse embryos ofGhox-lab and Hox 1.6 vertebrate homeobox containing genes related toDrosophila labial. Development 108, 47-58.
Sundin, O. H. and Eichele, G. (1990). A homeo domain protein reveals themetameric nature of the developing chick hindbrain. Genes Dev. 4, 1267-1276.
Westerfield, M. (1995) The Zebrafish Book. Univ. Oregon Press.Woo, K. and Fraser, S. E. (1995). Order and coherence in the fate map of the
zebrafish nervous system. Development 121, 2595-2609.Zhang, M., Kim, H-J., Marshall, H., Gendron-Maguire, M., Lucas, D.,
Baron, A., Gudas, L., Gridley, T., Krumlauf, R. and Grippo, J. (1994)Ectopic Hoxa-1 induces rhombomere transformation in mouse hindbrain.Development 120, 2431-2442.
(Accepted 13 December 1995)