efecto de la comunidad bacteriana en el …biblio.uabcs.mx/tesis/te3423.pdf · dra. sonia quijano...
TRANSCRIPT

UNIVERSIDAD AUTÓNOMA DE BAJA CALIFORNIA SUR
ÁREA DE CONOCIMIENTO DE CIENCIAS DEL MAR
DEPARTAMENTO ACADÉMICO DE BIOLOGÍA MARINA
TESIS
EFECTO DE LA COMUNIDAD BACTERIANA EN EL DINOFLAGELADO Gymnodinium catenatum (Graham, 1943):
CRECIMIENTO, PERFIL DE PIGMENTOS Y TOXINAS PARALIZANTES
QUE PARA OBTENER EL TÍTULO DE:
BIÓLOGA MARINA
PRESENTA:
DULCE VALERIA RAMÍREZ RODRÍGUEZ
DIRECTOR (A):
DRA. CHRISTINE JOHANNA BAND SCHMIDT
LA PAZ, B.C.S., DICIEMBRE DE 2015.


UNIVERSIDAD AUTÓNOMA DE BAJA CALIFORNIA SUR
ÁREA DE CONOCIMIENTO DE CIENCIAS DEL MAR
DEPARTAMENTO ACADÉMICO DE BIOLOGÍA MARINA
TESIS
EFECTO DE LA COMUNIDAD BACTERIANA EN EL DINOFLAGELADO Gymnodinium catenatum (Graham, 1943):
CRECIMIENTO, PERFIL DE PIGMENTOS Y TOXINAS PARALIZANTES
QUE PARA OBTENER EL TÍTULO DE:
BIÓLOGA MARINA
PRESENTA:
DULCE VALERIA RAMÍREZ RODRÍGUEZ
DIRECTOR (A):
DRA. CHRISTINE JOHANNA BAND SCHMIDT
LA PAZ, B.C.S., DICIEMBRE DE 2015.


AGRADECIMIENTOS
La realización de este trabajo es fruto de mi formación universitaria, por lo que
en primer instancia quiero agradecer a la Universidad Autónoma de Baja California
Sur (UABCS), en especial al Departamento de Biología Marina por fomentar y
fortalecer mi pasión hacia las ciencias marinas, brindándome la oportunidad de
descubrir mi vocación en la investigación científica.
Debo especial gratitud al Centro Interdisciplinario de Ciencias Marinas
(CICIMAR-IPN) por su total apoyo para el uso de sus instalaciones y por ende para la
complementación de mi aprendizaje, además por permitir integrarme al proyecto
institucional SIP20151381. Dentro de esta institución quisiera remarcar mi
agradecimiento hacia el Equipo FAN-CICIMAR y a los integrantes del Laboratorio de
Docencia 1, quienes a lo largo de todo este tiempo se convirtieron en mi familia ya
que el apoyo incondicional, la grata compañía y paciencia durante mi aprendizaje
hicieron más ameno el trayecto. En este sentido, comenzaré agradeciendo a mi Doc
(Dra. Christine J. Band Schmidt), no solo por la apertura de aceptar y apoyar mis
complicadas ideas, sino por darse a la tarea de supervisar, alentar y dirigir mi trabajo
de tesis, siempre con la mejor disposición, además del apoyo cordial, consejos e
importantes observaciones a mi manuscrito. A la Dra. Lorena M. Durán Riveroll, por
quien me incursioné en el mundo de las biotoxinas marinas, por enseñarme el valor
de esforzarse en un trabajo bien hecho, confiar en mis ideas y ayudarme a
encaminarlas hasta obtener el mejor provecho de ellas. A los doctores Ignacio Leyva
Valencia, pero sobre todo a Aldo Aquino Cruz por su invaluable apoyo y disposición,
por sus comentarios alentadores y por las innumerables enseñanzas en el
laboratorio. Además, extiendo mi agradecimiento a todos los miembros de este
maravilloso equipo, a los que van de paso, a los que regresan y a los que nunca se
han ido: Pablo, Nancy, Génesis, Miriam, Tina, Armando y Leyberth, gracias por
contribuir y estar presente en el enriquecimiento de mi trabajo.
Por otra parte, quisiera agradecer al Centro de Investigaciones Biológicas del
Noroeste (CIBNOR) y en especial al Dr. Francisco E. Hernández Sandoval,

responsable del Laboratorio de Ficotoxinas Marinas de esta institución, por su
valiosísima guía y paciencia en la aplicación de las técnicas para la extracción,
identificación y cuantificación de pigmentos y toxinas paralizantes requeridos en este
trabajo. Así mismo, agradezco a la Colección de Dinoflagelados Marinos
(CODIMAR), a la Dra. Lourdes Morquecho Escamilla y a la M. en. C. Amada Reyes
Salinas por proporcionarme algunas de las cepas utilizadas en el presente.
Bajo el mismo concepto, agradezco a la M. en B. Mónica C. Rodríguez Palacio
de la Universidad Autónoma Metropolitana por aportar algunas cepas al trabajo y a la
Dra. Sonia Quijano Scheggia de la Universidad de Colima, quien me dio lo más
valioso de este trabajo: la cepa contaminada con la comunidad bacteriana, además
de mostrar interés e iniciativa en apoyarnos con la parte de biología molecular, que si
bien no se incluye en el presente, no se descarta en un futuro cercano.
Siempre he dicho que un proyecto no es únicamente de quien lo financia, sino
de quien aporta, de manera desinteresada, un granito de arena. Por lo cual, mi total
gratitud a quienes me abrieron un espacio en su sitio de trabajo, me proporcionaron
equipo, material y reactivos para llevar a cabo este proyecto: Dra. Claudia J.
Hernández Guerrero, Dra. Noemí Águila Ramírez, Dra. Griselda M. Rodríguez
Figueroa, Dra. Laura Sánchez Velasco, Dra. Lourdes A. Arriaga Pizano, Dr. Mauricio
Muñoz Ochoa, Dr. Ismael Gárate Lizárraga, Dr. Sergio Hernández Trujillo, Dr.
Rogelio Armas González, Dr. Arturo Cérbulo Vázquez y en especial al Dr. David A.
Siqueiros Beltrones por el microscopio con el que pude evidenciar una parte crucial
de esta investigación.
A lo largo de esta travesía se me permitió participar en diferentes Congresos
Nacionales con un patrocinio inigualable, por lo que solo me resta agradecer al
Programa Interinstitucional para el Fortalecimiento de la Investigación y el Posgrado
del Pacífico (Programa DELFÍN), al CIBNOR con su Programa de Acercamiento de la
Ciencia a la Educación, al Consejo Nacional de Ciencia y Tecnología (CONACyT)
por la beca 21751, así como poder integrarme al proyecto SEP-CONACyT 178227 y
finalmente a la Red Temática de Florecimientos Algales Nocivos (RedFAN) por el
apoyo económico para asistir a los diferentes eventos.

Así también, agradezco a mi extraordinario comité revisor: Dr. Sergio F. Flores
Ramírez, Dr. Rafael Riosmena Rodríguez, por sus importantes observaciones y
aportaciones al presente manuscrito. A mí estimado profesor favorito (Dr. Héctor
Reyes Bonilla), gracias por la paciencia, el interés, la preocupación, pero sobre todo
por siempre tener las palabras necesarias para no dejarme claudicar en esto y a mí
querida Dra. Bárbara González Acosta, porque a pesar de estar tan ocupada,
siempre estuvo al pie del cañón, gracias por dejarme pertenecer a su equipo de
trabajo, por el microscopio de epifluorescencia que olvidaba tapar pero sobre todo
por sentir la misma emoción que yo al ver a los bacilos intracelulares.
Sin lugar a dudas mi apoyo más grande ha sido y será siempre mi familia, por
lo que no solo agradezco a todos sino dedico este trabajo a quienes renunciaron a mí
durante este periodo de estudio para que pudiera cumplir con uno de mis más
grandes sueños: ser bióloga marina. Agradezco a mis padres, Antonio y Jasmín,
quienes siempre se han preocupado por mí, me han dado todo de corazón, han sido
un ejemplo intachable, me han formado y enseñado a “esforzarme y ser valiente” y
nunca han dejado de creer en mí. A mis hermanos, Aldo y Liz, espero que la pérdida
de todo aquello importante para ustedes, pueda ser recompensado ahora que
concluí esta tesis. A mis grandes amores, mi abue, mi mana, Chabela, Víctor, mis
bisa, mis primos y tíos, en verdad no me alcanzan las palabras para agradecer todo
lo que significa para mí que me acompañen en este nuevo logro, que sin su apoyo no
hubiera sido posible.
Dicen que lo mejor de la amistad, es que puedes elegir quien va de paso y
quien se convierte en tu familia por elección, así que mil gracias a todos mis amigos,
de aquí y de allá: Brenda, Emmanuel, César, Wanda, Marcelo, Aurora, Rebeca,
Andrea, Andrés, Marijó, Ricardo, pero sobre todo quien fungió no solo como mi mejor
amiga paceña sino como hermana, confidente, porrista, mamá y de todo, gracias
infinitas pequeña Dan.
Finalmente, me gustaría agradecer a la persona que me ha demostrado el
valor de mis ideas y me ha dado fortaleza durante la realización de este trabajo, la
persona que siempre ha estado como mi mejor amigo, quien me dio alas, me enseñó

a volar y ha volado conmigo; quien nunca dejará de ser ese apoyo aún en los
momentos más difíciles, la persona que admiro por su dedicación e iniciativa y con la
que comparto mis días y sueños, de corazón gracias Miguel. Y obviamente y sin
restarle la importancia que merece, a quien soportó mis desvelos, los acompañó,
contrarrestó el estrés con sonrisas y por quien siempre deseo volver a casa, Romeo.
¡A todos, gracias infinitas!
Dedicado con todo mi amor y cariño a:
Jasmín, Hilda, Aldo, Liz y Miguel
Su fuerza y amor me han dirigido por la vida y me han dado las
alas que necesitaba para volar…
¡Gracias!

Página
I. INTRODUCCIÓN .......................................................................................... 1
II. ANTECEDENTES ......................................................................................... 5
III. JUSTIFICACIÓN ....................................................................................... 8
IV. HIPÓTESIS ............................................................................................... 9
V. OBJETIVOS ............................................................................................ 10
5.1. Objetivo general ................................................................................. 10
5.2. Objetivos específicos ........................................................................ 10
VI. MATERIALES Y MÉTODOS ................................................................... 11
6.1. Cepas .................................................................................................. 11
6.2. Medio de cultivo ................................................................................. 11
6.3. Condiciones de cultivo ...................................................................... 12
6.4. Aislamiento de la cianobacteria ....................................................... 12
6.5. Curvas de crecimiento de la cianobacteria ..................................... 13
6.5.1. Microscopio óptico ........................................................................ 13
6.5.2. Citometría de flujo ......................................................................... 14
6.5.3. Microscopia de epifluorescencia ................................................... 15
6.6. Curvas de crecimiento de G. catenatum .......................................... 17
6.7. Asociación G. catenatum-comunidad bacteriana ........................... 18
6.7.1. Exposición a filtrados celulares ..................................................... 18
6.7.2. Cultivos con contacto celular ........................................................ 18
6.8. Observaciones morfológicas de las cepas de G. catenatum ......... 19
6.9. Determinación de pigmentos ............................................................ 19
6.10. Determinación del perfil de toxinas paralizantes ........................... 20
6.11. Análisis estadísticos ......................................................................... 21
VII. RESULTADOS ........................................................................................ 22
7.1. Aislamiento de la cianobacteria ....................................................... 22
7.2. Curvas de crecimiento de la comunidad bacteriana ...................... 24
7.2.1. Microscopía óptica ........................................................................ 24
7.2.2. Citometría de flujo ......................................................................... 25
7.2.3. Microscopía de epifluorescencia ................................................... 28
ÍNDICE GENERAL

7.3. Curvas de crecimiento de G. catenatum .......................................... 33
7.4. Formación de cadenas celulares ...................................................... 35
7.5. Asociación G. catenatum-comunidad bacteriana ........................... 40
7.5.1. Exposición a filtrados celulares ..................................................... 40
7.5.2. Cultivos con contacto celular ........................................................ 42
7.6. Curvas de crecimiento ...................................................................... 43
7.7. Biomasa máxima ................................................................................ 47
7.8. Tasa de crecimiento .......................................................................... 48
7.9. Tiempo de generación ....................................................................... 49
7.10. Longitud de cadenas celulares ........................................................ 50
7.11. Curvas de crecimiento de la comunidad bacteriana en los cultivos
con contacto celular ............................................................................................ 54
7.12. Cambios morfológicos de Gymnodinium catenatum en presencia
de la comunidad bacteriana ................................................................................ 58
7.13. Perfil de pigmentos ........................................................................ 60
7.13.1. Cianobacteria ................................................................................ 60
7.13.2. Gymnodinium catenatum en presencia/ausencia de la comunidad
bacteriana…....
7.14. Perfil de toxinas paralizantes ........................................................ 64
7.15. Toxicidad......................................................................................... 67
VIII. DISCUSIÓN ............................................................................................ 70
8.1. Aislamiento de la cianobacteria ....................................................... 70
8.2. Curvas de crecimiento de la comunidad bacteriana ...................... 71
8.3. Curvas de crecimiento de G. catenatum .......................................... 74
8.4. Formación de cadenas celulares ...................................................... 75
8.5. Asociación G. catenatum-comunidad bacteriana ........................... 77
8.5.1. Exposición a filtrados celulares ..................................................... 77
8.5.2. Cultivos con contacto celular ........................................................ 78
8.6. Curvas de crecimiento de la comunidad bacteriana en los cultivos
con contacto celular ............................................................................................ 81
8.7. Cambios morfológicos de Gymnodinium catenatum en presencia
de la comunidad bacteriana ................................................................................ 83

8.8. Perfil de pigmentos ............................................................................ 85
8.8.1. Cianobacteria ................................................................................ 85
8.8.2. Gymnodinium catenatum en presencia/ausencia de la comunidad
bacteriana …………………………………………………………………………...86
8.9. Perfil de toxinas paralizantes............................................................ 87
8.10. Toxicidad......................................................................................... 89
IX. CONCLUSIÓN ........................................................................................ 90
X. LITERATURA CITADA ........................................................................... 91
ÍNDICE DE FIGURAS
Página
Figura 1. Distribución global de G. catenatum. .................................................. 3
Figura 2. Ciclo de vida de Gymnodinium catenatum. ......................................... 4
Figura 3. Técnicas microbiológicas para el aislamiento de la cianobacteria. ... 13
Figura 4. Curvas de crecimiento de la cianobacteria……. ............................... 23
Figura 5. Curva de crecimiento de la cianobacteria en medio GSe ................. 25
Figura 6. Ubicación de los bacilos intracelulares ............................................. 25
Figura 7. Citograma de la determinación del tamaño celular de la cianobacteria
a través de citometría de flujo. .................................................................................. 27
Figura 8. Citograma de la determinación de la complejidad celular de la
cianobacteria a través de citometría de flujo. ............................................................ 27
Figura 9. Citograma de identificación de las tres poblaciones de
microorganismos pertenecientes a la comunidad bacteriana .................................... 28
Figura 10. Curvas de crecimiento de la comunidad bacteriana ....................... 30
Figura 11. Patrones de cambio en la abundancia celular de los
microorganismos pertenecientes a la comunidad bacteriana .................................... 31

Figura 12. Micrografía del tamaño celular de los tres micoorganismos de la
comunidad bacteriana asociada a la cepa G7 de Gymnodinium catenatum. ............ 32
Figura 13. Bacterias intracelulares en Gymnodinium catenatum. ................... 32
Figura 14. Curvas de crecimiento de las cinco cepas de Gymnodinium
catenatum en medio GSe a 24 °C, 34 ups, ciclo luz:oscuridad 12:12. ...................... 34
Figura 15. Formación de cadenas durante la curva de crecimiento de la cepa
GCMV-7 de Gymnodinium catenatum en medio GSe. .............................................. 36
Figura 16. Formación de cadenas durante la curva de crecimiento de la cepa
G7 New de Gymnodinium catenatum en medio GSe. ............................................... 37
Figura 17. Formación de cadenas durante la curva de crecimiento de la cepa
G7 de Gymnodinium catenatum en medio GSe. ....................................................... 38
Figura 18. Formación de cadenas durante la curva de crecimiento de la cepa
GCCV-7 de Gymnodinium catenatum en medio GSe. . ............................................ 39
Figura 19. Formación de cadenas durante la curva de crecimiento de la cepa
62L de Gymnodinium catenatum en medio GSe. ...................................................... 40
Figura 20. Curvas de crecimiento de los cultivos de la comunidad bacteriana
con extracto celular de Gymnodinium catenatum .................................................... 41
Figura 21. Curvas de crecimiento de las cuatro cepas de Gymnodinium
catenatum inoculadas con la comunidad bacteriana ................................................. 43
Figura 22. Curvas de crecimiento de las cuatro cepas de Gymnodinium
catenatum en presencia/ausencia de la comunidad bacteriana ................................ 46
Figura 23. Biomasa máxima de las cuatro cepas de Gymnodinium catenatum
inoculadas con la comunidad bacteriana. ................................................................. 47
Figura 24. Tasa de crecimiento de las cuatro cepas de Gymnodinium
catenatum inoculadas con la comunidad bacteriana ................................................. 48
Figura 25. Tiempo de generación de las cuatro cepas de Gymnodinium
catenatum inoculadas con la comunidad bacteriana. ................................................ 49
Figura 26. Formación de cadenas durante la curva de crecimiento de las cepas
GCMV-7 y G7de Gymnodinium catenatum en presencia/ausencia de la comunidad
bacteriana.................................................................................................................. 52

Figura 27. Formación de cadenas durante la curva de crecimiento de las cepas
GCCV-7 y 62L de Gymnodinium catenatum en presencia/ausencia de la comunidad
bacteriana.................................................................................................................. 53
Figura 28. Curvas de crecimiento de la comunidad bacteriana asociada a las
cepas de G. catenatum. ............................................................................................ 56
Figura 29. Tasa de crecimiento de la cianobacteria en los cultivos tratamiento
de G. catenatum ........................................................................................................ 57
Figura 30. Tiempo de generación de la cianobacteria en los cultivos tratamiento
de Gymnodinium catenatum...................................................................................... 57
Figura 31. Cultivos inoculados con la comunidad bacteriana. ......................... 58
Figura 32. Células de G. catenatum en presencia de la comunidad bacteriana.
.................................................................................................................................. 59
Figura 33. Perfil de pigmentos de la cianobacteria asociada a la cepa G7 de
Gymnodinium catenatum. ......................................................................................... 60
Figura 34. Perfil de pigmentos de la cepa GCMV-7 en presencia/ausencia de la
comunidad bacteriana. .............................................................................................. 61
Figura 35. Perfil de pigmentos de la cepa GCCV-7 en presencia/ausencia de la
comunidad bacteriana. .............................................................................................. 62
Figura 36. Perfil de pigmentos de la cepa 62L en presencia/ausencia de la
comunidad bacteriana. .............................................................................................. 63
Figura 37. Perfil de pigmentos de la cepa G7 en presencia/ausencia de la
comunidad bacteriana. .............................................................................................. 64
Figura 38. Perfil de toxinas de la cepa GCMV-7 en presencia/ausencia de la
comunidad bacteriana. .............................................................................................. 65
Figura 39. Perfil de toxinas de la cepa GCCV-7 en presencia/ausencia de la
comunidad bacteriana. .............................................................................................. 66
Figura 40. Perfil de toxinas de la cepa 62L en presencia/ausencia de la
comunidad bacteriana. .............................................................................................. 66
Figura 41. Perfil de toxinas de la cepa G7 en presencia/ausencia de la
comunidad bacteriana. .............................................................................................. 67

Figura 42. Toxicidad (ngSTXeq/cél) de las cepas de G. catenatum en
presencia/ausencia de la comunidad bacteriana. ...................................................... 69
ÍNDICE DE TABLAS
Página
Tabla I. Cepas de G. catenatum aisladas de diversas regiones del Pacífico
mexicano y de la cianobacteria. ................................................................................ 11
Tabla II. Datos de crecimiento de la cianobacteria ........................................... 24
Tabla III. Datos promedio del crecimiento de las cinco cepas de G. catenatum
en medio GSe.. ......................................................................................................... 34
Tabla IV. Porcentaje promedio de cadenas celulares en G. catenatum. .......... 35
Tabla V. Datos promedio del crecimiento de la comunidad bacteriana con
extracto celular de Gymnodinium catenatum en medio GSe. ................................... 41
Tabla VI. Datos promedio del crecimiento de las cepas de Gymnodinium
catenatum inoculadas con la comunidad bacteriana en medio GSe. ........................ 43

RESUMEN
Durante los últimos años, se ha registrado una mayor frecuencia de Florecimientos
Algales Nocivos (FAN), trayendo consigo un incremento en los estudios destinados a
determinar los factores que controlan la dinámica poblacional de las principales
especies formadoras de estos procesos. Sin embargo, un aspecto que ha sido poco
estudiado es la interacción bacteria-microalga, que repercute en diversos aspectos
como el reabastecimiento, síntesis y asimilación de nutrientes, protección ante la
depredación y radiación U.V., en el crecimiento y finalmente en la producción de
toxinas. Gymnodinium catenatum es un dinoflagelado planctónico marino, formador
de cadenas celulares y productor de toxinas paralizantes; es una de las especies
responsables de FAN de las que se tiene mayor información en nuestro país, sin
embargo, poco se conoce sobre las relaciones ecológicas de esta especie con otras
especies de la comunidad bacteriana. Es por ello que el estudio de la interacción
bacteria-microalga proporcionará una mejor comprensión de la influencia de las
bacterias en el crecimiento de G. catenatum. Se trabajó con cinco cepas de G.
catenatum aisladas de Bahía de Santiago, Colima; Lázaro Cárdenas, Michoacán;
Bahía Concepción y Bahía de La Paz, B.C.S., de estas cepas, la de Colima presenta
una asociación simbiótica con una comunidad bacteriana dominada por una
cianobacteria y bacilos, la cianobacteria fue aislada en placas de agar con medio
GSe, a 24°C y con un ciclo de 12:12 horas luz-oscuridad. G. catenatum se cultivó
bajo las mismas condiciones que la cianobacteria y a 34 ups. La interacción entre
microorganismos se evaluó inoculando la comunidad bacteriana en las cepas de G.
catenatum, realizando conteos mediante microscopía de epifluorescencia,
determinándose las tasas y curvas de crecimiento, así como el perfil de pigmentos y
toxinas paralizantes por cromatografía de capa líquida de alta resolución de los
cultivos. La comunidad bacteriana no favoreció el crecimiento de G. catenatum, al
registrarse mayores biomasas y tasas de crecimiento en los monocultivos del
dinoflagelado pero sí repercutió en el perfil de pigmentos al aumentar la
concentración de la clorofila a, peridinina y β-caroteno. En cuanto al perfil de toxinas
paralizantes, no se observaron cambios en el mismo pero sí en la toxicidad celular, la
cual se ve disminuida en presencia de la comunidad bacteriana en tres de las cuatro

cepas, aumentando la concentración de toxinas N-sulfocarbamoil y carbamoil y
disminuyendo la concetración de toxinas carbamatas, por lo que se considera a las
bacterias y a la cianobacteria, agentes estresantes con mecanismos de acción
desconocidos. Con la información obtenida se propone que la interacción entre las
bacterias intracelulares, la cianobacteria extracelular y el dinoflagelado es bimodal:
durante la fase de crecimiento exponencial del cultivo la relación es mutualista al
presentarse un crecimiento óptimo en todos los organismos mientras que al
envejecer el cultivo, esta interacción se vuelve parasitaria, debido a las
modificaciones en el ciclo de vida que presenta G. catenatum, desarrollando quistes
temporales y cambios morfológicos, lo que desencadena una alta mortalidad. Esta
hipótesis nos permitiría explicar la rápida desaparición de los Florecimientos Algales
Nocivos (FAN) en la naturaleza, sin la exclusión de otros factores.
Palabras clave: Gymnodinium catenatum, comunidad bacteriana, crecimiento,
toxinas paralizantes, Pacífico Mexicano.

1
I. INTRODUCCIÓN
El crecimiento del fitoplancton está controlado por una combinación de
factores físicos como la temperatura, salinidad, luz y disposición de nutrientes
esenciales en forma de C, N, P, Si, Fe y diversos metales traza (Rooney-Varga et al.
2005). Aunado a eso, también intervienen factores bióticos como: la alelopatía, la
depredación y/o pastoreo, así como la interacción con una amplia gama de
microorganismos pertenecientes al femto, pico y microplancton, los cuales tienen una
influencia significativa en la regulación del crecimiento de las poblaciones de
fitoplancton (Ferrier et al. 2002).
Durante los últimos años, se ha estudiado la influencia de la comunidad
bacteriana en la dinámica de crecimiento y fisiología de las microalgas,
particularmente el efecto que tienen en la tasa de crecimiento y toxicidad del
fitoplancton nocivo mediante efectos estimulantes o inhibitorios, creándose posibles
asociaciones mutualistas (Doucette y Powell, 1998; Mayali y Azam, 2004).
No obstante, aún se desconocen los mecanismos de acción de la comunidad
bacteriana sobre la microalga, proponiéndose que las bacterias exudan diversos
metabolitos (vitaminas, quelantes orgánicos, formas asimilables de N, nutrientes
inorgánicos) que la microalga puede asimilar. Aunque, también se sabe que las
bacterias tienen una capacidad alguicida que limita el crecimiento de ciertas especies
(Ferrier et al. 2002; Adachi et al. 2003; Croft et al. 2005; Matsumoto, 2011).
De manera específica, los cultivos de dinoflagelados se ven asociados con
comunidades bacterianas inter e intracelulares, representadas por tres phyla:
Alfaproteobacterias, Gamaproteobacterias y Actinobacterias quienes influyen en la
fisiología del dinoflagelado, participan en el proceso del “bucle microbiano” donde
reabastecen y modulan la concentración y aporte de macronutrientes (C, N y P),
vitaminas y hierro, y participan en la producción de citoquininas (Azam, 1998).
Además, por ser organismos fotoheterótrofos aerobios tienen un papel importante en

2
el ciclo del C, lo que en conjunto favorece el crecimiento del dinoflagelado (Adachi et
al. 2001; Hold et al. 2001; Green et al. 2004).
Las bacterias intracelulares participan en la síntesis de nutrientes esenciales y
en la fijación de nitrógeno, mientras que la microalga les provee de un hábitat óptimo
para su crecimiento mediante la producción de compuestos orgánicos y
proporcionando una mayor área de superficie de fijación (Seibold et al. 2001). En
cuanto a las bacterias extracelulares, su colonización en la superficie celular
proporciona un mejor acceso a los nutrientes y en algunos casos ofrece protección
ante la depredación y/o radiación ultravioleta (Dang y Lovell, 2000).
Así, encontramos a Gymnodinium catenatum, dinoflagelado planctónico marino
reportado por primera vez en la primavera de 1939 en el Golfo de California
(Graham, 1943). Esta especie permaneció prácticamente desconocida, siendo
reportada sólo en dos ocasiones entre 1940 y 1967 (Balech, 1964; Hada, 1967).
Posteriormente, no hubo reportes hasta 1976, y de ese año hasta el 2014 se ha
registrado un aumento en su frecuencia, con más de 60 reportes en 23 países:
Australia, Japón, Venezuela, Cuba, Uruguay, Argentina, España, Costa Rica, Egipto,
Portugal, entre otros (Band-Schmidt et al. 2010; Hallegraeff et al. 2011; Quijano-
Scheggia et al. 2012).
En México, su distribución se ha reportado a lo largo de la costa del Pacífico
mexicano (Band-Schmidt et al. 2006; 2010. Fig. 1). Es una especie atecada que
forma cadenas celulares, de las principales productoras de FAN y ha sido muy
estudiada en el país (Blackburn et al. 1989; Band-Schmidt et al. 2010). G. catenatum
tolera un amplio intervalo de temperatura, salinidad y proporciones de N:P lo que
probablemente le permita tener una amplia distribución (Band-Schmidt et al. 2010,
2014).

3
Figura 1. Distribución global de G. catenatum (Durán-Riveroll, 2014).
El dinoflagelado atecado, Gymnodinium catenatum, tiene dos formas de
reproducción, una sexual heterotálica múltiple que se da por fusión de los gametos, y
otra asexual que implica la división vegetativa de las células. Blackburn y
colaboradores (1989) y Figueroa (2005) mostraron que los gametos que se forman
de manera asexual son isogametos, éstos se fusionan y forman un planocigoto
diploide que es biflagelado y mucho más grande en tamaño que las células sexuales
vegetativas.
Eventualmente, el dinoflagelado pierde el flagelo y con ello el planocigoto pierde
su capacidad natatoria, adquiriendo una forma más redondeada con un sulcus y un
cíngulo claramente visibles, que con el paso del tiempo se van desvaneciendo
(Figueroa et al. 2006). En este punto del ciclo de vida existen dos opciones:
enquistarse o dividirse y esto dependerá de las condiciones ambientales. Cuando
son óptimas, el planocigoto se dividirá mediante reproducción sexual, aumentando el
tamaño de la población; pero, si los parámetros ambientales (temperatura, oxígeno,

4
nutrientes, fotoperiodo) no son óptimos, el organismo se enquistará para
transformarse en un hypnocigoto, rodeado por una capa mucilaginosa y con un
cuerpo rojo pigmentado, constituyendo la fase de reposo de su ciclo de vida.
Después de dos semanas de inactividad, el hypnocigoto se puede enquistar,
formando un planomeiocito biflagelado (2n), que después se divide y forma células
vegetativas haploides (Figueroa et al. 2006; 2008; Albinsson, 2011).
La formación de quistes es una estrategia de esta especie ante condiciones
desfavorables y es fundamental en la dinámica de sus eventos de proliferación, ya
que facilita su dispersión, además de promover la recombinación genética. G.
catenatum puede formar, principalmente, dos tipos de quistes: a) de resistencia o
hypnocigotos, formados por reproducción sexual; y b) temporal o pelicicle, a partir de
células móviles cubiertos por una delgada membrana (Bravo, 1986; Anderson y
Garrison, 1997).
Figura 2. Ciclo de vida de Gymnodinium catenatum (Modificado de Albinsson, 2011).

5
El dinoflagelado Gymnodinium catenatum se caracteriza por ser la única
especie marina desnuda productora de toxinas paralizantes (Hallegraeff, 1993).
Estas toxinas pueden acumularse en moluscos bivalvos, caracoles, crustáceos
(Saldate-Castañeda et al. 1991), pulpos (Núñez-Vázquez et al. 2008), peces
planctófagos filtradores (Reis-Costa et al. 2010) y de manera secundaria en el ser
humano. Dado la distribución de G. catenatum, así como el impacto que ocasiona en
la fauna silvestre ha incrementado a nivel mundial, científicos han establecido un
mayor interés en los factores que influyen en el crecimiento y toxicidad de la especie
(Hallegraeff et al. 2011).
Sin embargo, poco se conoce sobre la interacción que existe entre la
comunidad bacteriana y G. catenatum. Reportes recientes demuestran que las
bacterias son esenciales para el crecimiento de esta especie y tienen influencia en su
toxicidad y absorción de nutrientes, pudiendo formar relaciones de mutualismo
mediante mecanismos de acción desconocidos (Amin et al. 2007; Green et al. 2010;
Albinsson, 2011; Bolch et al. 2011; Matsumoto, 2011).
La finalidad de este trabajo es determinar si en presencia de una comunidad
bacteriana asociada a una cepa de G. catenatum aislada de la Bahía de Santiago,
Colima, el dinoflagelado se vea beneficiado en su tasa de crecimiento, en la
formación de cadenas de mayor longitud y en un incremento en su biomasa; además
de determinar los posibles cambios que ocurran en el perfil de toxinas y pigmentos.
II. ANTECEDENTES
Los principales trabajos que se han realizado evaluando la interacción entre
Gymnodinium catenatum y la comunidad bacteriana asociada, se basan en cultivos
experimentales donde se concluye que el dinoflagelado crece en asociación con una
comunidad bacteriana que puede influir en su crecimiento, así como en el contenido
y perfil de toxinas. Específicamente, se ha observado que la influencia bacteriana en
el crecimiento de G. catenatum se da con mayor frecuencia en los quistes de

6
resistencia que en las células vegetativas, mientras que los cambios en el perfil de
toxinas paralizantes (PSP) se atribuyen a tres factores: producción autónoma de
toxinas por las bacterias, modulación de la toxicidad del dinoflagelado por las
bacterias y a través de la biotransformación de los análogos de las toxinas PSP
(Amin et al. 2007; Bolch et al. 2011).
Dichos estudios comenzaron a tener un mayor auge décadas atrás, al evaluar
la influencia de las bacterias en el crecimiento y fisiología del fitoplancton tóxico. De
esta manera encontramos el trabajo de Gallacher y colaboradores (1997), Doucette
y Powell (1998) y Smith y colaboradores (2001) quienes indican que las bacterias
pueden influir en el contenido y producción de toxinas paralizantes, además de
modular la toxicidad del dinoflagelado y la biotransformación de los derivados de las
toxinas.
Uribe y Espejo (2003) tras eliminar la comunidad bacteriana presente en
Alexandrium catenella, observan que en ausencia de la misma, la toxicidad del
dinoflagelado se ve disminuida en una quinta parte, esto tras comparar tres técnicas
diferentes para la cuantificación de las toxinas paralizantes: cromatografía de capa
líquida de alta resolución (HPLC), pruebas electrofisiológicas que miden la inhibición
de la saxitoxina (STX) en los canales de sodio regulados por voltaje y mediante
bioensayos en ratón.
Con la misma temática y llegando a resultados símiles, Maas y colaboradores
(2007), concluyen que la presencia de una comunidad bacteriana asociada a
dinoflagelados productores de toxinas paralizantes es esencial, debido a que su
inhibición con antibióticos refleja un descenso significativo en los niveles de
saxitoxina (STX) y gonyautoxinas (GTX1-3) sin presentarse cambios en la tasa de
crecimiento y en el tiempo de regeneración del dinoflagelado.
Amin y colaboradores (2007) redujeron significativamente la comunidad
bacteriana asociada a cepas aisladas de G. catenatum, percatándose que en
presencia de la comunidad bacteriana se da la formación de sideróforos foto-lábiles,

7
que favorecen la asimilación de Fe en el dinoflagelado a través de pequeños poros
en la membrana celular, incrementando su crecimiento. Se sugiere que la captación
de otros micronutrientes como el boro y selenio pueden mediarse por las bacterias
asociadas y en el caso del selenio, éste puede explicar las pequeñas diferencias
regionales entre cepas debido a que los requerimientos del nutriente no son
generalistas (Amin et al. 2009).
Otro reporte que sustenta lo descrito anteriormente fue el realizado por
Albinsson en 2011, quien mostró una asociación entre la bacteria marina
Marinobacter sp. y una cepa australiana de G. catenatum la cual sobrevivió más de
230 días sin la adición de nutrientes, sugiriendo que era la bacteria quien regulaba la
disponibilidad de los mismos. Además, se demostró que ciertos cambios en la
comunidad bacteriana asociada al dinoflagelado pueden alterar significativamente las
proporciones relativas de toxinas paralizantes producidas por cepas en condiciones
de laboratorio.
En el mismo año, Bolch y colaboradores (2011) reportan que tras inhibir el
crecimiento de las bacterias asociadas a G. catenatum, el dinoflagelado ralentiza su
crecimiento hasta emparejarlo con el de las bacterias, logrando con ello un
crecimiento recíproco. Se ha utilizado el crecimiento bacteriano como variable
determinística del crecimiento del dinoflagelado, estableciendo que la dependencia
bacteria-microalga constituye una relación biológica de mutualismo.
Años después, Albinsson y colaboradores (2014) indican que cepas
australianas de G. catenatum presentan una relación mutualista con dos
proteobacterias marinas: Marinobacter sp. y Alcanivorax sp. En presencia de ambas,
el dinoflagelado tiene un mayor crecimiento, observándose un predominio de toxinas
hidroxibenzoato (GC1/2) y decarbamoil (dcGTX2/3), mientras que en ausencia de las
mismas son las toxinas sulfocarbamoil (C1/2 y C3/4) los análogos dominantes. Esta
observación da la pauta a considerar que las bacterias funcionan como agentes
reguladores de la toxicidad de G. catenatum debido a que la única variable en los
tratamientos fue la presencia/ausencia de ambas bacterias.

8
Cabe destacar que en la mayoría de los casos, se han utilizado antibióticos
para disminuir o eliminar la comunidad bacteriana asociada a cultivos específicos de
microalgas nocivas, comparando los cambios existentes en presencia/ausencia de la
misma y las respuestas observadas en el crecimiento, la fisiología y/o en el nivel de
producción de toxina que han sido atribuidos directa o indirectamente a la comunidad
bacteriana (Maas et al. 2007; Bolch et al. 2011).
Aunque teóricamente parece sencillo, tales experimentos presentan una serie
de dificultades y limitaciones. En primer lugar, la comunidad bacteriana asociada a
los cultivos es compleja, presentan una amplia diversidad filogenética y no se elimina
fácilmente con el uso de uno o varios antibióticos. En segundo lugar, la confirmación
de la ausencia total de una comunidad bacteriana es difícil debido a la existencia de
bacterias intracelulares presentes en la microalga, y en tercer lugar, el dilema
inherente que la prueba de ausencia bacteriana es compleja, laboriosa y necesita de
equipos sofisticados y generalmente costosos (Uribe y Espejo, 2003; Maas et al.
2007).
Por ello, el éxito o fracaso en la obtención de una cepa axénica de microalga
también depende de la respuesta que ésta tenga al tratamiento con el antibiótico,
debido a que en algunos casos no es posible diferenciar entre la toxicidad natural de
la microalga y la ocasionada por el antibiótico (Maas et al. 2007; Matsumoto, 2011).
III. JUSTIFICACIÓN
Los florecimientos de fitoplancton, incluyendo aquellos de especies tóxicas
como Gymnodinium catenatum, son fenómenos frecuentes de las regiones costeras
y de surgencia de los océanos. Dichos eventos, en particular cuando está
involucrada alguna especie nociva, pueden tener efectos negativos en los
ecosistemas marinos, en la salud pública y la economía de la zona (Hallegraeff et al.
1995; Green et al. 2004).

9
Hoy en día, existen reportes que indican que la incidencia de los FAN ha
incrementado (Hallegraeff, 2011) y con ello el interés de estudiar a la comunidad
bacteriana asociada a las microalgas que los producen. Esto debido a que son las
bacterias quienes desempeñan un papel importante en el origen, desarrollo y
duración de los FAN, ya que durante estos eventos es necesaria la presencia de un
agente estimulador del crecimiento celular que promueva la reproducción sexual e
inhiba la formación de quistes. No obstante, los mecanismos implicados en el ciclo
de vida de las diversas especies se desconocen (Albinsson, 2011).
Es por ello que el estudio de la interacción bacteria-microalga proporciona,
como primer paso, una comprensión de la influencia de las bacterias en el
crecimiento de las microalgas, así como brindar información sobre la fisiología y
toxicidad de la especie de interés.
IV. HIPÓTESIS
Se ha observado el crecimiento de una comunidad bacteriana formada
principalmente por bacilos y cianobacterias, en una cepa específica de G. catenatum
en condiciones de laboratorio (Band-Schmidt et al. 2004). La cepa que muestra la
presencia de estos microorganismos, tiene un mayor crecimiento comparado con
otras cepas de la misma especie.
Por lo tanto, se asume que la interacción entre los microorganismos es benéfica
(protocooperación). En presencia de la comunidad bacteriana G. catenatum tendrá
una mayor tasa de crecimiento y biomasa, reflejado en cadenas más largas de
células.
Así mismo, es bien sabido que la comunidad bacteriana asociada a cepas de G.
catenatum desempeña un papel importante en su toxicidad, por lo que, teniendo en
cuenta la estrecha relación entre las bacterias y el dinoflagelado, éstas podrían influir
en la producción y composición de las toxinas y pigmentos de esta especie.

10
V. OBJETIVOS
5.1. Objetivo general
Determinar el tipo de relación existente entre Gymnodinium catenatum
y la comunidad bacteriana asociada a cepas específicas del
dinoflagelado.
5.2. Objetivos específicos
Determinar el perfil de pigmentos de la comunidad bacteriana asociada
a G.catenatum a través de cromatografía de capa líquida de alta
resolución.
Determinar las curvas de crecimiento de la comunidad bacteriana y del
dinoflagelado en cultivos monoespecíficos, así como en cultivos mixtos
de ambas poblaciones.
Determinar el efecto de la interacción entre la comunidad bacteriana y
G. catenatum, evaluando cambios en la tasa de crecimiento, biomasa,
perfil de pigmentos y toxinas paralizantes del dinoflagelado.
Describir los cambios morfológicos, longitud de cadenas y la producción
de quistes temporales de G. catenatum al interactuar con la comunidad
bacteriana.

11
VI. MATERIALES Y MÉTODOS
6.1. Cepas
Se trabajó con cinco cepas de G. catenatum aisladas de diversas regiones del
Pacífico mexicano, las cuales están registradas con las siguientes claves: GCCV-7,
GCMV-7, 62L, G7 y G7 New (Tabla I).
La comunidad bacteriana fue aislada de la cepa G7 proveniente de Bahía de
Santiago, Colima, México.
Tabla I. Cepas de G. catenatum aisladas de diversas regiones del Pacífico mexicano y de la cianobacteria.
Cepa Lugar y año de aislamiento Aislado por
GCCV-7
GCMV-7
G7; G7 New
62L
Bahía Concepción, B.C.S. 2000
Mazatlán, Sinaloa. 2013
Manzanillo, Colima. 2010
Lázaro Cárdenas, Michoacán. 2005
C. Band-Schmidt
L. Morquecho-Escamilla
S. Quijano-Scheggia
M. Rodríguez-Palacio
6.2. Medio de cultivo
Las cepas se mantuvieron en medio GSe modificado con la adición de extracto
de lombricompostaje (Bustillos-Guzmán et al. 2015). Este medio consiste en una
solución de nutrientes (nitratos y fosfatos), metales traza, vitaminas y selenio,
adicionados en agua de mar previamente filtrada y esterilizada.
En cuanto al aislado de la comunidad bacteriana también, se mantuvo, en los
medios de cultivo, los cuales fueron: ASN-III (Rippka, 1988) con y sin aporte de
nitrógeno y el medio Povasoli´s o ES (Provasoli, 1957) que difieren en la composición
de sales minerales pero ambos aportan los requerimientos necesarios de macro y
micronutrientes en agua de mar previamente filtrada y estéril (Torres-Ariño, 2001).

12
6.3. Condiciones de cultivo
Durante todas las condiciones experimentales, los cultivos se mantuvieron a
una temperatura de 24°C ± 1°C, con una iluminación de 150 µmolm-2s-1 en un ciclo
de luz:oscuridad 12:12 y con una salinidad de 34.
6.4. Aislamiento de la cianobacteria
Se realizaron diversas técnicas microbiológicas con la finalidad de obtener una
cepa lo más pura posible de la cianobacteria (microorganismo dominante de la
comunidad bacteriana asociada a G. catenatum). Para tal fin, se utilizó el
fototactismo, la migración vertical (Ramírez-Gama et al. 2006) y estria cruzada
(Olivas, 2004).
Para la técnica de fototactismo y migración vertical, se vertieron 25 mL de la
cepa G7 en probetas previamente cubiertas de aluminio, dejando la parte superior
descubierta. Los cultivos se revisaron diariamente para evaluar el tipo de respuesta
hacia la luz: positiva o negativa. Posteriormente, de la parte superficial de la probeta,
se tomó una alícuota de la muestra que fue sembrada en medio GSe con agar al 1%
en cajas Petri desarrollando el método de siembra de la estría cruzada en placa,
girando la caja al ir rayando hasta formar un pentágono para que al final las células
quedarán separadas unas de otras (Fig. 3).
Una vez obtenidas las colonias de cianobacterias, se resembraron cada tercer
día con un asa bacteriológica realizando el estriado correspondiente. Teniendo la
cepa pura de la cianobacteria, las colonias se trasfirieron nuevamente a medio GSe
líquido.

13
Figura 3. Técnicas microbiológicas para el aislamiento de la cianobacteria. A) Fototactismo y migración vertical. B) Estriado cruzado en cajas Petri con agar bacteriológico al 1% en medio GSe.
6.5. Curvas de crecimiento de la cianobacteria
6.5.1. Microscopio óptico
Se prepararon tres inóculos provenientes de la cepa pura de la cianobacteria,
con una concentración inicial de 9 000 cél/mL en 150 mL de medio GSe en matraces
de 250 mL. Se tomó un mililitro del cultivo cada segundo día hasta la fase de
decaimiento y fueron fijadas con 50 µL de lugol ácido al 1% (Throndsen, 1979). El
conteo celular se determinó por recuento en una cámara de Neubauer de 0.1 mm
bajo un microscopio óptico compuesto (LABOMED CXRII) y de acuerdo a la
siguiente formula:
cél/mL = C
SP · P · Fd
Dónde: C: Número de células observadas; SP: Superficie contada; P:
Profundidad; Fd: Factor de dilución.

14
6.5.2. Citometría de flujo
Para el conteo directo de la comunidad bacteriana y el análisis por citometría de
flujo, se prepararon tres inóculos del cultivo, teniendo una concentración inicial de 9
000 cél/mL en 150 mL de medio GSe en matraces de 250 mL. Se tomó 1 mL del
cultivo cada séptimo día hasta el día 70, las muestras se diluyeron en una proporción
1:2 con la solución salina amortiguada por fosfatos (PBS 1%). Se fijaron
inmediatamente con formol a una concentración final de 4% y fueron refrigeradas a
4°C en oscuridad hasta su análisis.
Adicionalmente, a cada muestra se le agregaron 10 µL de perlas (microesferas
de poliestireno de 10 µm de diámetro) para usarlas como referencia de tamaño.
El análisis de las muestras se realizó mediante un citómetro de flujo modelo
CyAn ADP BeckmanCoulter® equipado con tres láseres (405 nm, 488 nm y 633 nm).
Se contabilizaron 500, 000 eventos y el tiempo de análisis de cada muestra fue de 3
minutos en promedio. Los resultados se registraron en modo logarítmico,
obteniéndose los datos respectivos al tamaño relativo de cada célula (FSC) y de la
complejidad de la misma (SSC).
El principio de la citometría de flujo se basa en que las células suspendidas en
un líquido pasan individualmente al frente de una fuente de luz (láser) y los datos de
la luz reflejada (número total de eventos), son detectados y organizados en un
archivo (Castillo-Navarrete, 2005). Finalmente, la identificación de las diferentes
poblaciones se realizó con el software Infinicyt V1.8 que organiza la información en
citogramas, teniendo como variables: el tamaño relativo (FSC) de la partícula
determinado como la cantidad de luz trasmitida en un ángulo cónico pequeño (0-10°)
y la complejidad interna (SSC), que es la luz dispersada en ángulo recto y compara
la forma y estructura de las partículas con la de una esfera, indicando también la
estructura interna de las células (Gasol y del Giorgio, 2000).

15
6.5.3. Microscopía de epifluorescencia
Se utilizó el método descrito por Kepner y Pratt (1994) utilizando como
fluorocromo SYBR Gold, que tiene afinidad por los ácidos nucleicos y un microscopio
de fluorescencia marca Olympus, modelo BX60. Las muestras se observaron bajo
emisión de luz azul, con una longitud de onda entre 450 y 490 nm a 1 000 x de
magnificación, en un cuarto oscuro.
Se colectaron alícuotas de 1 mL del cultivo de la cianobacteria por 70 días, las
muestras se diluyeron en una proporción 1:2 con la Solución Salina Amortiguada por
Fosfatos (PBS 1%). Se fijaron inmediatamente con formol a una concentración final
de 4% y fueron refrigeradas a 4°C en oscuridad hasta su análisis. Posteriormente, se
realizó una segunda dilución 1:15 con TE 1% previamente filtrado a través de
membranas de 0.22 µm. Las muestras se filtraron en un sistema de filtración de 15
mL de capacidad a través de una membrana de policarbonato de 25 mm de diámetro
y 0.2 µm de tamaño de poro, previamente humedecida con 1 mL de agua destilada.
La filtración se llevó a cabo con una bomba de vacío.
Posteriormente, se colocaron las membranas ya secas en diversos
cubreobjetos, añadiéndoles 80 µL de SYBR Gold en la parte central y dejándolas
reposar durante 15 minutos en total oscuridad. Después se eliminó el exceso de
colorante, lavando las membranas con 4 mL de TE 1%.
Una vez que la membrana se secó, se colocó sobre un portaobjetos,
agregándole una gota de aceite de baja inmersión y se puso la membrana encima,
luego otra gota de aceite y posteriormente el cubreobjetos, presionando la parte
central para cubrir con aceite toda la membrana e ir eliminando las burbujas. Las
preparaciones fueron analizadas inmediatamente o almacenadas en oscuridad y a
4°C hasta su conteo.
La determinación de la densidad de la comunidad bacteriana se realizó
utilizando una retícula de 10 x 10 µm colocada en el ocular del microscopio y debido
a la presencia de tres organismos dominantes con diferente abundancia se utilizaron

16
tres métodos de conteo, todos con una precisión de ±10% a un 95% de intervalo de
confianza (Mecalco-Hernández, 2010).
Para células filamentosas y la cianobacteria (con forma cocoide), se contó el
número de organismos presentes en un total de 30 retículas por muestra, tal como lo
recomienda Kirchman (1993), sin embargo, para esta última, también se enumeraron
un mínimo de 200 células por muestra en campos elegidos aleatoriamente. Se
consideraron todas las células con morfología bien definida, así como todos los
individuos que quedaron en los bordes de la retícula, siempre y cuando más del 50%
de su estructura se encontrara dentro de la misma (Díaz-Hernández, 1997).
Finalmente, para las células con forma de bacilo se contaron 300 células por
muestra en campos elegidos al azar (Venrick, 1978) y siguiendo el método de conteo
antes descrito.
Para calcular la abundancia se utilizó la siguiente ecuación (Kepner y Pratt,
1994):
cél/mL = N ∙ At
D ∙ Vf ∙ G ∙ Ag
Dónde: N: número de células contadas; At: área efectiva de filtración; D: factor
de dilución; Vf: volumen filtrado; G: número de retículas contadas; Ag: área de la
retícula.
Esta técnica, nos permitió dar un seguimiento fotográfico de los principales
cambios en la abundancia de la comunidad bacteriana, a través de micrografías
tomadas con una cámara semi profesional (Cannon) a un aumento final de 1000 X,
Posteriormente y teniendo como referencia de tamaño, la reglilla del
microscopio a 1000 X, se analizaron las micrografías con el software ImageJ V1.49,
quien al promediar la distancia entre dos puntos, logra medir el tamaño celular

17
6.6. Curvas de crecimiento de G. catenatum
Para determinar las curvas de crecimiento del dinoflagelado, se prepararon
inóculos de cada una de las cepas de G. catenatum, iniciando con una concentración
de 500 cél/mL en 150 mL de medio GSe en matraces de 250 mL, por triplicado. Se
tomaron 2 mL de cultivo cada segundo día hasta la fase de decaimiento, y las
muestras se fijaron con lugol ácido al 1% (Throndsen, 1979). Los conteos se
realizaron en una cámara de Sedgwick-Rafter de 1 mL bajo un microscopio óptico
compuesto (LABOMED CXRII). Se registró el número y largo de las cadenas
celulares.
Para la obtención de las curvas de crecimiento, tanto de la comunidad
bacteriana como de G. catenatum, se calculó el promedio y la desviación estándar de
los conteos. Los promedios de cada conteo fueron graficados para obtener la curva
de crecimiento y determinar las fases de crecimiento de las cepas.
La tasa de crecimiento (K) se determinó para cada una de las cepas a partir de
la fase de crecimiento exponencial, utilizando la siguiente ecuación, de acuerdo a
Guillard (1973):
K = ((Ln(C2) – Ln(C1)) / t2-t1).
Donde C1 es el número de células por mililitro al tiempo uno y C2 es el número
de células por mililitro en el tiempo dos.
Se calculó el tiempo de generación (tg) a partir de la tasa de crecimiento
mediante la ecuación (Guillard, 1973):
tg = 0.301/K

18
6.7. Asociación G. catenatum-comunidad bacteriana
6.7.1. Exposición a filtrados celulares
Para evaluar el efecto de los exudados celulares de G. catenatum sobre la
comunidad bacteriana, se mantuvo al dinoflagelado en un cultivo de lote hasta que
alcanzó la fase exponencial tardía, retirando las células mediante una filtración a
través de membranas de fibra de vidrio de 25 mm GF/F. En matraces de 250 mL se
inocularon 50 mL de cultivo de la comunidad bacteriana, exponiendo a las células a
tres volúmenes diferentes (10, 15 y 20 mL) de los filtrados de medio de cultivo con
exudados celulares de G. catenatum por triplicado, teniendo como control, al cultivo
de la comunidad bacteriana sin extracto bajo las mismas condiciones. Se tomaron
muestras cada siete días para conteos celulares y observaciones.
6.7.2. Cultivos con contacto celular
Se escalaron los cinco cultivos de G. catenatum, inoculando cada cepa a una
densidad inicial de 500 cél/mL del dinoflagelado y con 9 000 cél/mL de la
cianobacteria, en un volumen de 150 mL de medio GSe en matraces de 250 mL por
triplicado.
Se tomó una alícuota de 2 mL de cada matraz, previo a una agitación suave, y
se realizaron conteos cada dos días para G. catenatum, fijando las células con una
solución de lugol ácido al 1% (Throndsen, 1979), utilizando una cámara de
Sedgwick-Rafter de 1 mL y registrando la longitud de las cadenas celulares. La
comunidad bacteriana se contabilizó cada cuatro días, obteniendo alícuotas de 500
µL diluidos en una proporción 1:2 con la solución salina amortiguada por fosfatos
(PBS 1%). Se fijaron con formol a una concentración final de 4% y se mantuvieron a
4°C en oscuridad, para su posterior montaje y análisis por microscopia de
epifluorescencia.

19
6.8. Observaciones morfológicas de las cepas de G. catenatum
Las cepas del dinoflagelado inoculadas con la comunidad bacteriana
presentaron la formación de quistes, células deformadas, lisadas o con pérdida de
movilidad. Dichas observaciones, así como el seguimiento de la interacción de todos
los experimentos se realizó en un microscopio óptico Zeiss Axioscop, tomando
microfotografías con una cámara ScopePhoto (3.0v) a diferentes aumentos.
6.9. Determinación de pigmentos
Para la obtención del perfil de pigmentos de la cianobacteria y de G. catenatum
en presencia/ausencia de la comunidad bacteriana, los cultivos se agitaron
suavemente antes de tomar la muestra de agua procedente del cultivo. Se filtraron
40 y 25 mL, respectivamente, a través de una membrana de fibra de vidrio de 25 mm
GF/F, se mantuvieron en tubos eppendorf cubiertos de aluminio y se congelaron
inmediatamente a 4°C hasta su análisis en el Laboratorio de Microalgas Nocivas del
Centro de Investigaciones Biológicas del Noroeste (CIBNOR).
Para los cultivos del dinoflagelado en presencia/ausencia de la comunidad
bacteriana, se tomó una alícuota de 2 mL que se preservó con una solución de lugol
ácido al 1% (Throndsen, 1979) para conteos celulares, los cuales se realizaron como
como se describió anteriormente.
El material retenido en cada filtro se utilizó para identificar y cuantificar los
pigmentos fotosintéticos por cromatografía líquida de alta resolución (HPLC), en un
equipo Agilent HP 1100; extrayendo con pinzas cada filtro y colocándolos dentro de
tubos de plástico de 25 mL, donde fueron macerados con una varilla de vidrio para
posteriormente agregarles 1 mL de acetona y dividir el volumen final a la mitad,
colocando 0.5 mL en dos eppendorf (previamente cubiertos de aluminio).
Posteriormente, los eppendorf se colocaron en una centrifuga (Heraeus Biofuge
Fresco) a 3,000 rpm, con una temperatura de 5°C durante 20 minutos. Al finalizar, el

20
sobrenadante se recuperó y se colocó en viales ámbar, específicos para HPLC,
donde se inyectaron 100 µL al sistema, utilizándose dos longitudes de onda distintas:
440 nm y 667 nm (Vidussi et al. 1996).
La cuantificación de los principales pigmentos se basó en el valor del factor de
respuesta de cada pigmento (concentración de pigmento/absorbancia) de acuerdo a
Vidussi y colaboradores (1996) y Montoura y Repeta (1997), considerándose el
tiempo de retención y las características espectrales con estándares comerciales
certificados (Agencia Internacional para determinaciones de 14C, Dinamarca).
6.10. Determinación del perfil de toxinas paralizantes
La extracción de toxinas se realizó durante la fase exponencial tardía de los
cultivos de G. catenatum en presencia/ausencia de la comunidad bacteriana,
agitando suavemente los matraces antes de tomar la muestra de 50 mL del cultivo
para posteriormente filtrarlos a través de filtros de 25 mm GF/F. A cada filtro,
contenido en un eppendorf individual, se le adicionaron 1.5 mL de ácido acético
(0.03N), sonicando (35 kHz) tres veces por cinco minutos cada muestra en intervalos
de 60 segundos.
El sobrenadante se filtró con un filtro de jeringa desechable (0.45µm de tamaño
de poro). Se tomó una alícuota de 300 µL del extracto clarificado, que se utilizó para
la hidrólisis con HCl (1M) para transformar a toxinas decarbamatadas (tipo B y C) a
sus análogos. Finalmente, 10 µL de ambos extractos (con y sin hidrólisis) se
inyectaron en el sistema HPLC (HP 1100) (Hummert et al. 1997; Yu et al. 1998).
La cromatografía se llevó a cabo como lo sugerido por Hummert y
colaboradores en 1997 y Yu y colaboradores en 1998, utilizando un gradiente
amortiguado de pares de iones (ion-pair-buffer), compuesto de una solución de ácido
octanosulfónico y fosfato de amonio a un pH de 6.9 y acetonitrilo, para separar las
toxinas paralizantes.

21
Después de una oxidación postcolumna con ácido periódico alcalino, los
productos resultantes se detectaron con un detector de fluorescencia (HP 1116) a
una longitud de onda de 330 nm (excitación) y de 395 nm (emisión).
La identificación de toxinas se llevó a cabo comparando cromatogramas de las
muestras con los de sus respectivos estándares (National Research Council,
Canadá) y la cuantificación se realizó comparando las áreas de los picos de
extractos de estándares con el correspondiente factor de respuesta.
6.11. Análisis estadísticos
Las diferencias en la biomasa máxima de las cepas de G. catenatum, así
como de la cianobacteria en los cuatro medios de cultivo se determinaron con
análisis de varianza unifactorial, probando la normalidad de los datos con la prueba
de Kolmogorov-Smirnof y la homoscedasticidad con la prueba de Levene (Zar, 1999).
Si los datos no fueron normales se realizó estadística no paramétrica (prueba de
Kruskal Wallis) (Zar, 1999).
Las diferencias en la longitud de cadenas celulares, así como en el perfil de
toxinas se evaluaron con la prueba estadística X2 , mientras que las diferencias en el
perfil de pigmentos se evaluaron con la prueba t de student (Zar, 1999).
Cuando se encontraron diferencias significativas, se realizaron pruebas a
posteriori de comparación de medias de Tukey (Day y Quinn, 1989) para grupos
homogéneos.
Los análisis se realizaron utilizando el software Statistica v7.0 (StatSoft, Inc.)
con un nivel de significancia de 0.05.

22
VII. RESULTADOS
7.1. Aislamiento de la cianobacteria
Se logró el aislamiento de la cianobacteria mediante las técnicas de
fototactismo y migración vertical ya que de esta manera se eliminó una mayor
cantidad de bacterias heterótrofas asociadas, mientras que el sembrado en placa
resultó efectivo en la limpieza de las cianobacterias, al aislar únicamente las colonias
verdes, con borde circular y aquellas más alejadas a colonias rosadas,
características de bacterias heterótrofas.
A pesar de no registarse diferencias significativas entre la abundancia y el
medio de cultivo, observamos que el medio ASN-III con adición de nitrógeno registró
la mayor abundancia celular (122,222 ± 4,714 cél/mL), seguido del medio GSe. Los
medios ASN-III sin fuentes nitrogenadas y ES tuvieron una abundancia
considerablemente menor, obteniendo, en general, patrones de comportamiento
similares a partir del noveno día en la mayoría de las curvas de crecimiento, no
obstante, únicamente se registraron diferencias significativas en las abundancias
máximas de los medios ASN-III con N2 y ES (H(3, N=12) = 8.4358; p<0.05).
La fase de crecimiento exponencial inició el tercer día en los medios ASN-III con
y sin aporte de N2 y ES, mientras que para GSe dicha fase comenzó hasta el día
nueve. La máxima densidad celular se obtuvo durante la fase exponencial tardía, en
los días 15 (±4.76), 18 (±5.62) y 9 (±3.02) para los medios ASN-III con N2, ASN-III sin
N2 y GSe y finalmente el medio ES, respectivamente (Fig. 4).
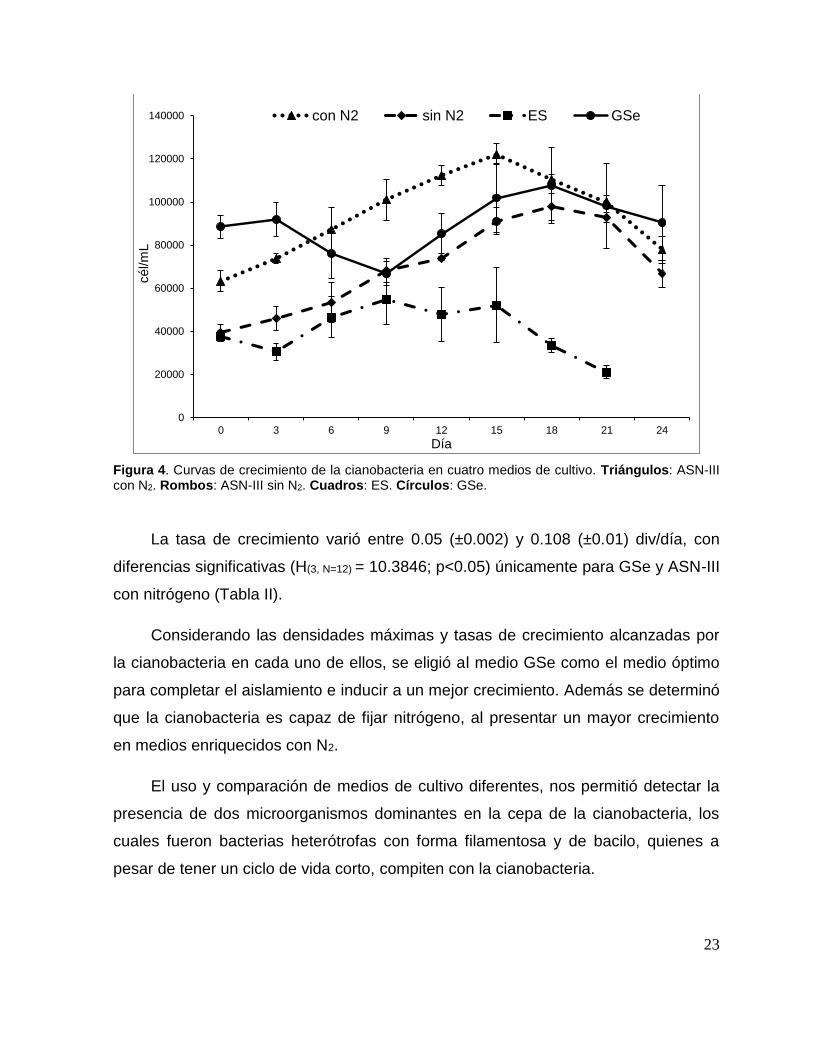
23
Figura 4. Curvas de crecimiento de la cianobacteria en cuatro medios de cultivo. Triángulos: ASN-III con N2. Rombos: ASN-III sin N2. Cuadros: ES. Círculos: GSe.
La tasa de crecimiento varió entre 0.05 (±0.002) y 0.108 (±0.01) div/día, con
diferencias significativas (H(3, N=12) = 10.3846; p<0.05) únicamente para GSe y ASN-III
con nitrógeno (Tabla II).
Considerando las densidades máximas y tasas de crecimiento alcanzadas por
la cianobacteria en cada uno de ellos, se eligió al medio GSe como el medio óptimo
para completar el aislamiento e inducir a un mejor crecimiento. Además se determinó
que la cianobacteria es capaz de fijar nitrógeno, al presentar un mayor crecimiento
en medios enriquecidos con N2.
El uso y comparación de medios de cultivo diferentes, nos permitió detectar la
presencia de dos microorganismos dominantes en la cepa de la cianobacteria, los
cuales fueron bacterias heterótrofas con forma filamentosa y de bacilo, quienes a
pesar de tener un ciclo de vida corto, compiten con la cianobacteria.
0
20000
40000
60000
80000
100000
120000
140000
0 3 6 9 12 15 18 21 24
cél/m
L
Día
con N2 sin N2 ES GSe
24
Tabla II. Datos de crecimiento de la cianobacteria en cuatro medios de cultivo. *diferencias significativas (p<0.05).
Medio de cultivo
Inicio de la fase exponencial
(días)
Fin de la fase exponencial
(días)
Tasa de crecimiento
(div/día)
Abundancia máxima (cél/mL)
Tiempo en alcanzar la biomasa máxima
(días)
ASN-III
con N2 3 15 ± 1.29 0.04 ± 0.001* 122,222 ± 4 714 15 ± 4.76
sin N2 6 24 ± 3.02 0.05 ± 0.002 97,777 ± 6 285 18 ± 5.62
ES 3 21 ± 2.92 0.09 ± 0.001 55,000 ± 17 284 9 ± 3.02
Gse 9 24 ± 3.02 0.10 ± 0.01* 107,592 ± 17 638 18 ± 5.62
7.2. Curvas de crecimiento de la comunidad bacteriana
El crecimiento de la cianobacteria en medio GSe se evaluó con tres métodos: 1)
microscopía óptica, 2) citometría de flujo y 3) microscopía de epifluorescencia; cuya
precisión, duración y grado de sesgo son diferentes.
7.2.1. Microscopía óptica
En la figura 5 mediante conteo por microscopía óptica, se observan cuatro fases
de crecimiento del cultivo de la cianobacteria. La fase de aclimatación dura 4 días, al
quinto día comienza la fase de crecimiento exponencial, que concluye el día 10
obteniéndose una densidad máxima de 109,630 cél/mL. La fase estacionaria dura
dos días, del día 10 al 12 y después el cultivo decae rápidamente.
La tasa de crecimiento en la fase exponencial fue de 0.10 (±0.01) div·día-1, con
un tiempo de generación de 2.76 (±0.02) días.
Se debe resaltar que con esta técnica se visualizó, por primera vez, a un
microorganismo diferente de la cianobacteria y fueron los bacilos, que en conjunto,
rodeaban a las células de G. catenatum formando “parches” tanto en células
individuales como en cadenas (Fig. 6 A, B).

25
Figura 5. Curva de crecimiento de la cianobacteria en medio GSe a 24 °C, 34 ups, ciclo luz:oscuridad 12:12. Líneas verticales: desviación estándar.
Figura 6. Ubicación de los bacilos (flechas) en: A) células individuales y B) en cadenas celulares. Se observa cómo van rodeando al dinoflagelado hasta formar parches a su alrededor conforme el cultivo avanza con el tiempo.
7.2.2. Citometría de flujo
El uso de esta técnica únicamente nos permitió discernir entre poblaciones de
organismos autótrofos y heterótrofos, así como tener una aproximación del tamaño
0
20000
40000
60000
80000
100000
120000
140000
0 2 4 6 8 10 12 14 16 18
cél/m
L
Día

26
celular de los microorganismos debido al uso de microesferas de poliestireno. Sin
embargo, no fue posible llevar a cabo un conteo celular debido al ruido electrónico
(interferencia eléctrica) del equipo y el tamaño de las microesferas (10 µm de
diámetro) que triplicaba en tamaño a los integrantes de la comunidad bacteriana.
A pesar de los inconvenientes en el método de análisis, el uso de la
autoflorescencia de la cianobacteria nos permitió localizar su población dentro de la
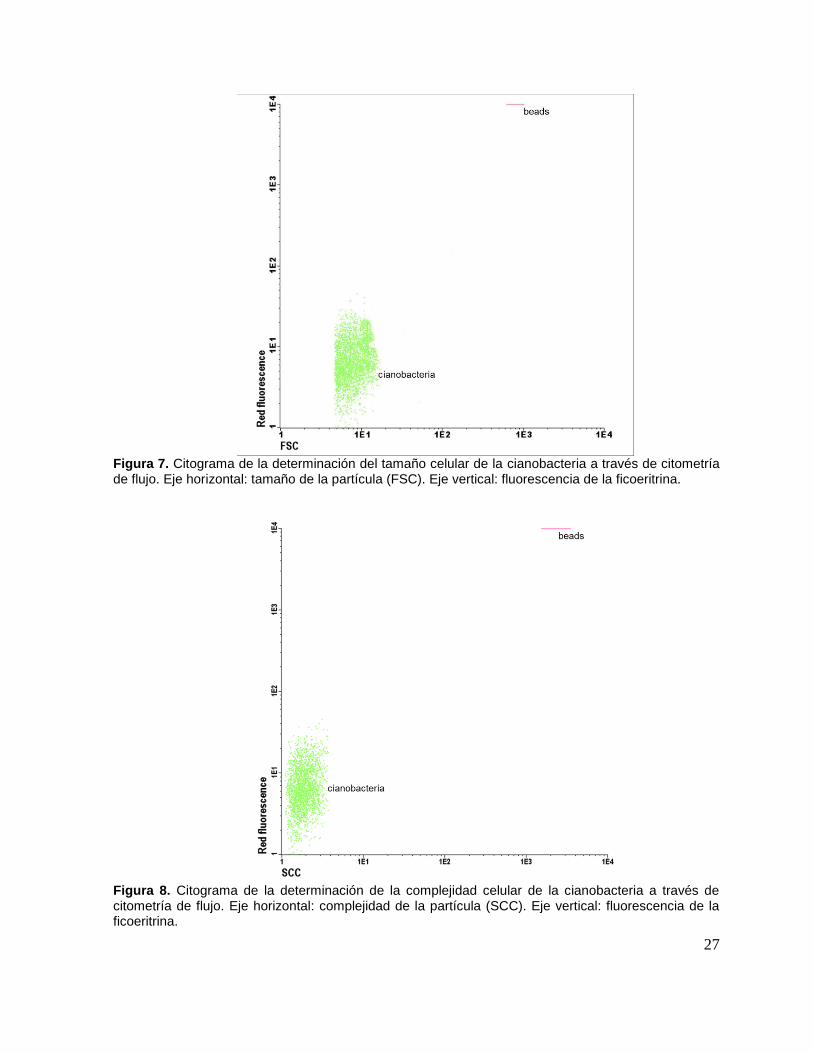
comunidad bacteriana. Utilizando la florescencia de la ficoeritrina y el tamaño celular,
la cianobacteria se encontró en la esquina inferior izquierda, indicando que presenta
poca florescencia y que su tamaño es muchísimo más pequeño que el de las
microesferas, localizadas en la parte superior derecha (Fig. 7). Al graficar la
florescencia roja y la complejidad de la partícula, la población de la cianobacteria se
recorrió horizontalmente hacia el eje de las ordenadas y nuevamente quedó en la
parte inferior, por lo que su autoflorescencia es baja y se comprueba que es una
partícula poco compleja, con forma semejante a una esfera (Fig. 8).
Al analizar la complejidad de la partícula y su tamaño, se logró diferenciar tres
poblaciones diferentes (primer indicio de que la comunidad bacteriana estaba
integrada por tres y no dos microorganismos). La primer población se caracterizó por
tener un tamaño pequeño y ser poco compleja, integrando a la cianobacteria; la
población de mayor tamaño que la cianobacteria y más compleja, integró a los
bacilos y la tercera, más grande y compleja que las anteriores agrupó a las bacterias
heterótrofas filamentosas (Fig. 9).
27
Figura 7. Citograma de la determinación del tamaño celular de la cianobacteria a través de citometría de flujo. Eje horizontal: tamaño de la partícula (FSC). Eje vertical: fluorescencia de la ficoeritrina.
Figura 8. Citograma de la determinación de la complejidad celular de la cianobacteria a través de citometría de flujo. Eje horizontal: complejidad de la partícula (SCC). Eje vertical: fluorescencia de la ficoeritrina.

28
Figura 9. Citograma de identificación de las tres poblaciones de microorganismos pertenecientes a la comunidad bacteriana asociada a la cepa G7 de G. catenatum mediante citometría de flujo. Eje horizontal: tamaño de la partícula (FSC). Eje vertical: complejidad de la partícula (SCC).
7.2.3. Microscopía de epifluorescencia
El conteo y la obtención de la curva de crecimiento de la cianobacteria, así
como de las bacterias heterótrofas filamentosas y bacilos se realizaron mediante
microscopía de epifluorescencia, observándose una mejor delimitación de las fases
de crecimiento.
Para la cianobacteria (línea continua con círculos), la fase de aclimatación tuvo
una duración de 21 días, a partir del día 22 al 63 se observa un crecimiento
exponencial, y posterior a este día, el cultivo decae. La máxima abundancia celular
fue de 75,527 ± 2,516 cél/mL. Inmediatamente, después de la fase exponencial
tardía, comenzó la fase de declive (Fig. 10).

29
La tasa de crecimiento fue de 0.038 (±0.024) div/día y el tiempo de generación
de 7.72 (±0.327) días.
La línea discontinua con cuadros, representa a las bacterias heterótrofas
filamentosas, que tuvieron un crecimiento similar al de la cianobacteria, registrándose
la fase de aclimatación del día cero al día 21, con un crecimiento exponencial hasta
el día 63, día en el que se registró la mayor abundancia (34,963 ± 898 cél/mL). No se
observó una fase estacionaria (Fig. 10). La tasa de crecimiento fue de 0.036 (±0.021)
div/día, mientras que el tiempo de generación fue de 8.26 (±0.541) días.
En el caso de los bacilos, (línea discontinua con triángulos) estos presentaron
una curva de crecimiento peculiar, iniciando con una fase de aclimatación muy corta
e inapreciable. Observamos que del día 14 al 21 se registra la primera fase
exponencial alcanzando una abundancia aproximada de 14,444 (±5,132 cél/mL)
seguida de una fase estacionaria con duración de siete días, para finalmente
presentarse un declive pronunciado, que representa, en el día 35, el comienzo de la
segunda fase exponencial que alberga la mayor densidad obtenida durante el día 49
(214,815 ± 12,830 cél/mL). A partir del día 50 y hasta el día 63 observamos la fase
de declive pero también se aprecia el inicio de una tercera fase exponencial que no
tuvo seguimiento por la muerte celular de los dos microorganismos restantes,
además que el tiempo de muestreo ya era bastante (Fig. 10).
La tasa de crecimiento de los bacilos, se obtuvo del día 14 al día 49 y fue de
0.0303 (±0.028) div/día, con un tiempo de generación de 9.903 (±0.86) días.
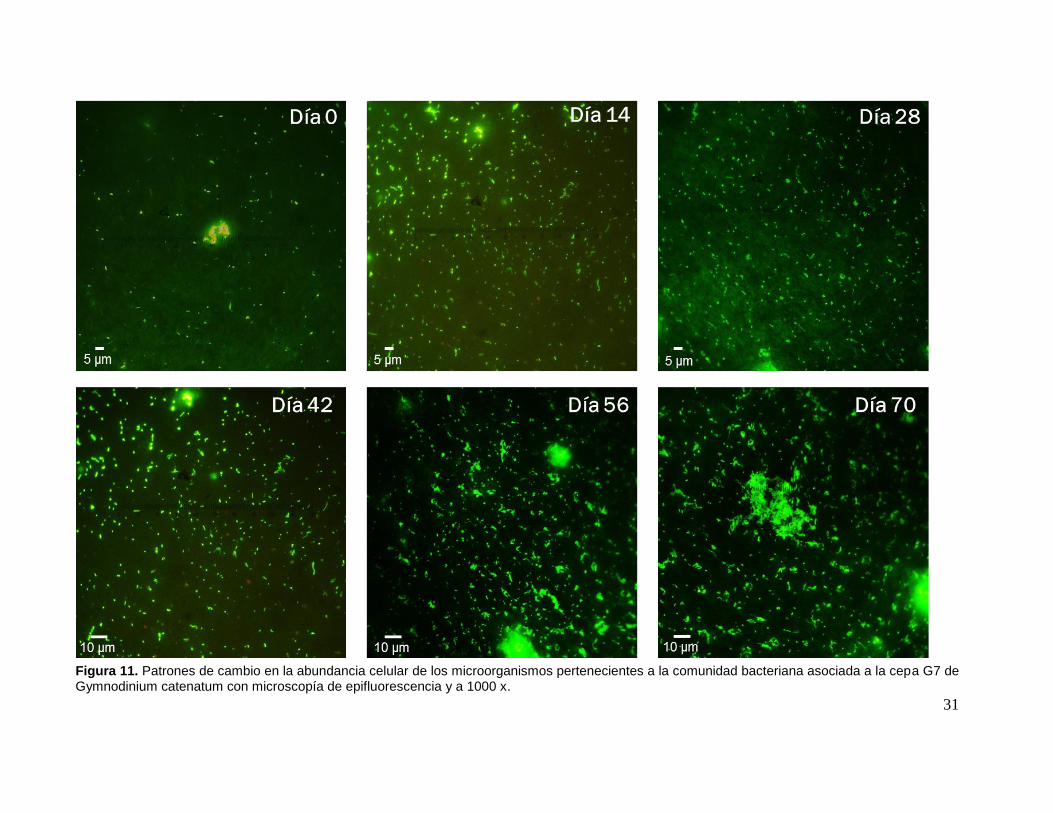
En cuanto a los patrones de cambio reflejados en las micrografías, podemos
observar que el día de siembra (día cero) tuvo una abundancia baja para los tres
microorganismos: cianobacterias, bacterias heterótrofas filamentosas y bacilos, la
cual incrementó con el tiempo, siendo los bacilos quienes lograron no solo obtener la
mayor abundancia celular sino también mantenerla por periodos más prolongados de
tiempo.

30
Durante la segunda semana de muestreo, se observa una fuerte dominancia de
bacilos, mientras que la abundancia de bacterias filamentosas y cianobacterias es
baja. Patrón que se fue invirtiendo durante los siguientes días. A finales del primer
mes ya se observaba un incremento celular a favor de la cianobacteria, el cual se
mantuvo, hasta observarse pequeños conglomerados que generalmente se
encontraban rodeadas de bacilos, contrario a las células filamentosas cuyo
comportamiento era solitario. No obstante, en los últimos días, cuando la abundancia
de las cianobacterias comenzó a disminuir, las bacterias filamentosas aumentaron su
población de tal forma que se les veía como redes enmarañadas al igual que a los
bacilos. Ambas bacterias (filamentosas y bacilos) se caracterizaron por tener ciclos
de crecimiento rápido y por formar patrones de agregación masivos, contrario a la
cianobacteria quien presentó un ciclo de crecimiento lento y con abundancias
relativamente baja (Fig. 11). Haciendo referencia al tamaño celular, del más grande
al menor, encontramos a las bacterias heterótrofas filamentosas, bacilos y
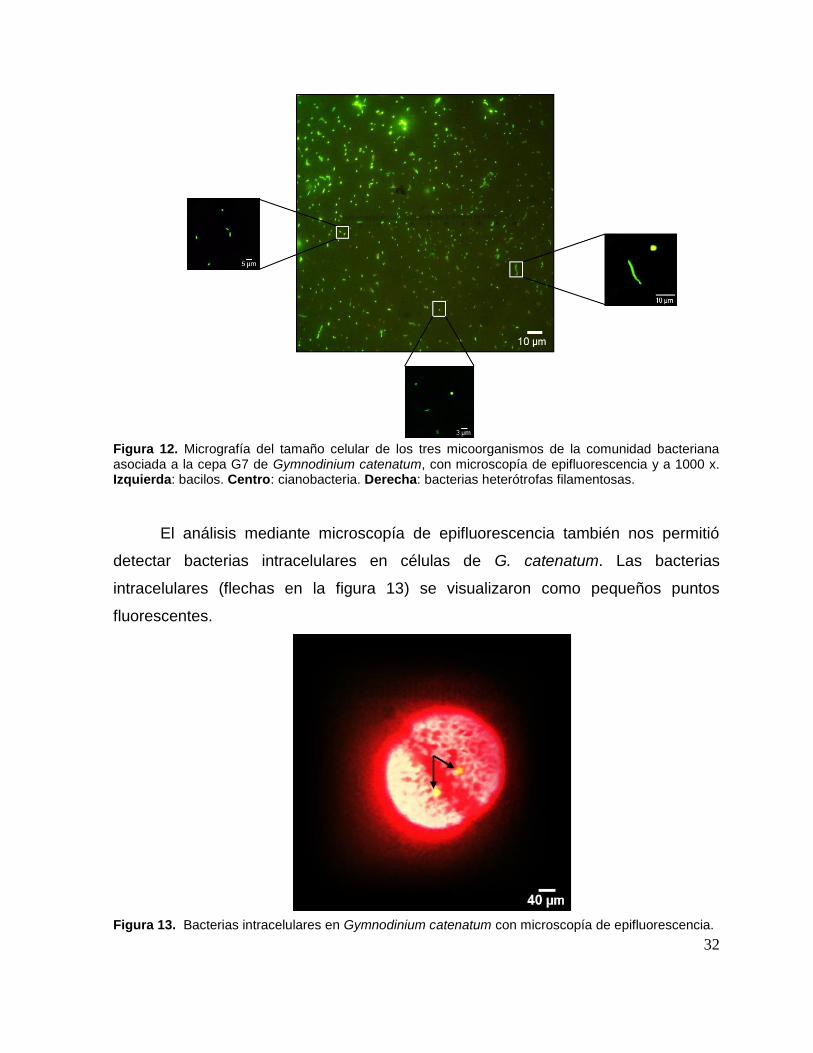
cianobacterias, con 5-10 µm; 4-5 µm y 2-3 µm, respectivamente (Fig. 12).
Figura 10. Curvas de crecimiento de la comunidad bacteriana asociada a la cepa G7 Colima de G. catenatum en medio GSe a 24 °C, 34 ups, ciclo luz:oscuridad 12:12. Círculos: cianobacteria. Cuadros: bacterias heterótrofas filamentosas. Triángulos: bacilos. Líneas verticales: desviación estándar. Eje vertical derecho: bacilos; eje vertical izquierdo: cianobacteria y filamentos.
0
50000
100000
150000
200000
250000
0
10000
20000
30000
40000
50000
60000
70000
80000
90000
0 7 14 21 28 35 42 49 56 63 70
cél/m
L
cél/m
L
Día
Cianobacteria Filamentos Bacilos
31
Figura 11. Patrones de cambio en la abundancia celular de los microorganismos pertenecientes a la comunidad bacteriana asociada a la cepa G7 de Gymnodinium catenatum con microscopía de epifluorescencia y a 1000 x.
32
Figura 12. Micrografía del tamaño celular de los tres micoorganismos de la comunidad bacteriana asociada a la cepa G7 de Gymnodinium catenatum, con microscopía de epifluorescencia y a 1000 x. Izquierda: bacilos. Centro: cianobacteria. Derecha: bacterias heterótrofas filamentosas.
El análisis mediante microscopía de epifluorescencia también nos permitió
detectar bacterias intracelulares en células de G. catenatum. Las bacterias
intracelulares (flechas en la figura 13) se visualizaron como pequeños puntos
fluorescentes.
Figura 13. Bacterias intracelulares en Gymnodinium catenatum con microscopía de epifluorescencia.

33
7.3. Curvas de crecimiento de G. catenatum
Todas las cepas presentaron la típica curva de crecimiento para esta especie,
con una fase de aclimatación corta, seguida de una fase exponencial, contrario a la
fase estacionaria que fue corta o nula debido a que la fase de declive se presentó
rápidamente después de alcanzar la abundancia máxima (Fig. 14).
La cepa G7 fue la que tuvo menor crecimiento, alcanzando la etapa de declive
en menor tiempo que el resto de las cepas, con una biomasa máxima de 1,146 ± 4
cél/mL en el día 14. Contrario a GCCV-7, que mostró la mayor biomasa celular, con
un máximo de 6463 ± 24 cél/mL en el doceavo día, seguido por la cepa michoacana
62L. Las cepas restantes presentaron biomasas máximas intermedias, las cuales se
encontraron entre 2,840 ± 38 y 1,625 ± 4 cél/mL. El inicio de la fase exponencial en
todas las cepas inició el día 2 y finalizó entre los días 10 (±3.02) y 18 (± 4.63), en
cuanto a la biomasa máxima, ésta se alcanzó entre los días 12 (±2.93) y 20 (±1.75)
(Tabla III).
En cuanto a las abundancias máximas, existieron diferencias significativas entre
las cepas GCMV-7, G7 New y G7 (F(4,15)=93292; p<0.05), observándose únicamente
dos grupos con características similares. Mientras que las tasas de crecimiento
variaron entre 0.215 (±0.02) y 0.0924 (±0.002) div/día. Los análisis estadísticos
mostraron diferencias significativas (p<0.05) para tres de las cinco cepas: G7,
GCCV-7 y 62L (H(4, N=20) = 17.15897) (Tabla III).
El tiempo de generación entre cepas varió entre 1.4 (±0.02) y 7 (±0.40) días,
encontrándose diferencias significativas (H(4, N=20) = 17.63979; p<0.05) en GCCV-7,
G7 y 62L, delimitándose dos grupos con características similares entre sí (Tabla III).
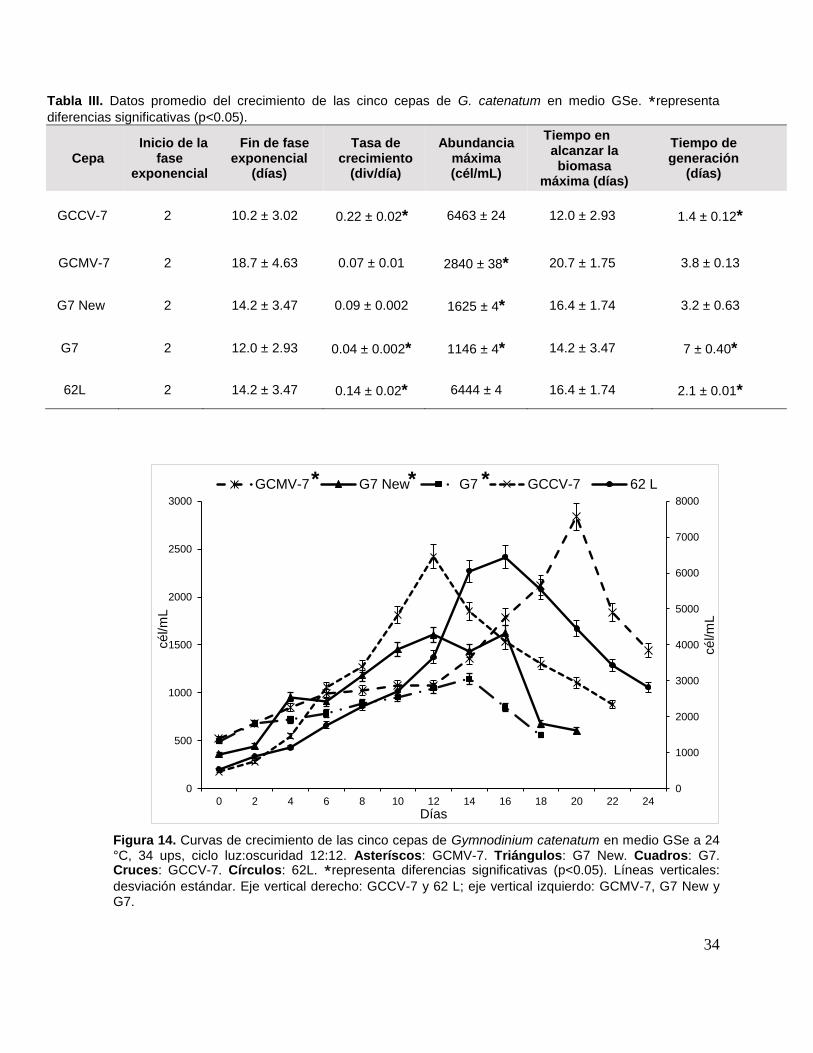
34
Tabla III. Datos promedio del crecimiento de las cinco cepas de G. catenatum en medio GSe. *representa
diferencias significativas (p<0.05).
Cepa Inicio de la
fase exponencial
Fin de fase exponencial
(días)
Tasa de crecimiento
(div/día)
Abundancia máxima (cél/mL)
Tiempo en alcanzar la biomasa
máxima (días)
Tiempo de generación
(días)
GCCV-7 2 10.2 ± 3.02 0.22 ± 0.02* 6463 ± 24 12.0 ± 2.93
1.4 ± 0.12*
GCMV-7 2 18.7 ± 4.63 0.07 ± 0.01 2840 ± 38* 20.7 ± 1.75
3.8 ± 0.13
G7 New 2 14.2 ± 3.47 0.09 ± 0.002 1625 ± 4* 16.4 ± 1.74 3.2 ± 0.63
G7 2 12.0 ± 2.93 0.04 ± 0.002* 1146 ± 4* 14.2 ± 3.47 7 ± 0.40*
62L 2 14.2 ± 3.47 0.14 ± 0.02* 6444 ± 4 16.4 ± 1.74 2.1 ± 0.01*
Figura 14. Curvas de crecimiento de las cinco cepas de Gymnodinium catenatum en medio GSe a 24 °C, 34 ups, ciclo luz:oscuridad 12:12. Asteríscos: GCMV-7. Triángulos: G7 New. Cuadros: G7. Cruces: GCCV-7. Círculos: 62L. *representa diferencias significativas (p<0.05). Líneas verticales:
desviación estándar. Eje vertical derecho: GCCV-7 y 62 L; eje vertical izquierdo: GCMV-7, G7 New y G7.
0
1000
2000
3000
4000
5000
6000
7000
8000
0
500
1000
1500
2000
2500
3000
0 2 4 6 8 10 12 14 16 18 20 22 24
cél/m
L
cél/m
L
Días
GCMV-7 G7 New G7 GCCV-7 62 L* * *

35
7.4. Formación de cadenas celulares
Durante el seguimiento de la curva de crecimiento de cada cepa, se observó el
patrón de las cadenas celulares, que en general mostró una mayor proporción de
células individuales (33.4 ± 8.6%), seguido por cadenas de 4-6 células (26 ± 4.7%),
de 2-3 células (25.8 ± 7.3%) y una menor producción de cadenas de 7 células y
mayores (15 ± 11.8%). Sin embargo, las excepciones fueron la cepa G7 New que
presentó un dominio de cadenas celulares largas, igual y/o mayores a 7 células (36.2
± 17.1%) y la cepa GCMV-7 con una mayor producción de cadenas de 2-3 células
(37.2 ± 15.7%) (Tabla IV).
Tabla IV. Porcentaje promedio de cadenas celulares y día de cultivo con mayor proporción de cadenas de 7 células y mayores en Gymnodinium catenatum. Los números resaltados en negritas, representan los mayores valores en cada categoría.
%
Cepa Células
individuales Cadenas 2-3 Cadenas 4-6 Cadenas ≥7
Día con la mayor proporción de cadenas ≥7 (% / día / fase)
GCMV-7 33 ± 11 37 ± 16 19 ± 11 11 ± 9 26% / 12 / exponencial tardía
G7 New 19 ± 12 18 ± 7 27 ± 6 36 ± 17 67% / 0 / siembra
G7 38 ± 15 27 ± 8 24 ± 13 11 ± 7 26% / 18 / declive
GCCV-7 41 ± 18 21 ± 12 31 ± 16 8 ± 5 15% / 2 / aclimatación
62L 36 ± 11 26 ± 13 29 ± 10 9 ± 5 15% / 4 / exponencial
La cepa GCMV-7, durante la fase de aclimatación mostró una mayor proporción
de células individuales (40.75 ± 12.19%), a partir del día 8 y hasta el declive la
presencia de cadenas celulares más largas fue prevaleciendo, obteniendo una mayor
producción de cadenas de 2-3 células (37.2 ± 15.7%), seguido de cadenas de 4-6
células. Mientras que la producción de cadenas con longitud igual y/o mayor a 7
células fue menor (11.5 ± 9) a pesar de haberse registrado un promedio de 26% de
cadenas con esta longitud durante el día 12, correspondiente al día con mayor
densidad celular (Fig. 15).

36
Figura 15. Formación de cadenas durante la curva de crecimiento de la cepa GCMV-7 de Gymnodinium catenatum en medio GSe. Rayas: células individuales. Gris: cadenas de 2-3 células. Puntos: cadenas de 4-6 células. Azul: cadenas de 7 células y mayores. La línea continua representa la curva de crecimiento.
La cepa G7 New, a diferencia del resto, registró un dominio de cadenas de 7
células y mayores (36.2 ± 17.1%) en todo el ciclo de crecimiento, siendo más notorio
en la fase de aclimatación (67%) y hasta el día 6 (fase exponencial). En cuanto a las
células individuales, estas presentaron una baja producción (19.2 ± 11.6%), seguidas
de las cadenas de 2-3 células (17.6 ± 6.7%); mientras que las cadenas de 4-6 células
comenzaron a observarse más durante la fase exponencial- tardía (27 ± 5.8%) (Fig.
16).
0
1000
2000
3000
0
10
20
30
40
50
60
70
80
90
100
0 2 4 6 8 10 12 14 16 18 20 22 24
cél/m
L
% d
e c
adena
s c
elu
lare
s
Día

37
Figura 16. Formación de cadenas durante la curva de crecimiento de la cepa G7 New de Gymnodinium catenatum en medio GSe. Rayas: células individuales. Gris: cadenas de 2-3 células. Puntos: cadenas de 4-6 células. Azul: cadenas de 7 células y mayores. La línea continua representa la curva de crecimiento.
Caso totalmente inverso al descrito anteriormente es el de la cepa G7, que
presentó oscilaciones en la formación de cadenas celulares, iniciando con un
dominio de células individuales (día de siembra), seguido de una mayor producción
de cadenas de 4-6 células (inicio de la fase exponencial). Posteriormente se observa
una alternancia entre células individuales, cadenas de 2-3 células y de 4-6 células.
En promedio, las células individuales fueron las dominantes (38.1 ± 14.6%), seguidas
por las cadenas de 2-3 células (25.5 ± 7.7%), de 4-6 células (24 ± 13.4%) y
finalmente, las menos representativas, las de 7 células y mayores (11.4 ± 7.5%) que
comenzaban a presentarse en mayor cantidad durante el declive (26%) (Fig. 17).
0
500
1000
1500
0
10
20
30
40
50
60
70
80
90
100
0 2 4 6 8 10 12 14 16 18 20
cél/m
L
% c
adenas c
elu
lare
s
Día

38
Figura 17. Formación de cadenas durante la curva de crecimiento de la cepa G7 de Gymnodinium catenatum en medio GSe. Rayas: células individuales. Gris: cadenas de 2-3 células. Puntos: cadenas de 4-6 células. Azul: cadenas de 7 células y mayores. La línea continua representa la curva de crecimiento.
Otra cepa que mostró un dominio de células individuales a lo largo de todo el
ciclo de crecimiento, fue GCCV-7, con la diferencia que durante la fase de
aclimatación, las cadenas de 4-6 células fueron las dominantes (30.8 ± 16.3%). A
partir del día 6 (fase exponencial) y hasta el día 10 hubo una mayor producción de
células individuales, seguido de una mayor abundancia de cadenas de 2-3 células
(20.8 ± 12.3%). Sin embargo, el declive se caracterizó por presentar, nuevamente
una mayor cantidad de células individuales, que en promedio representaron un 40%.
En cuanto a las cadenas celulares de 7 células y mayores, su presencia fue baja (7.7
± 4.6%) (Fig. 18).
0
400
800
1200
0
10
20
30
40
50
60
70
80
90
100
0 2 4 6 8 10 12 14 16 18
cél/m
L
% c
adena
s c
elu
lare
s
Día
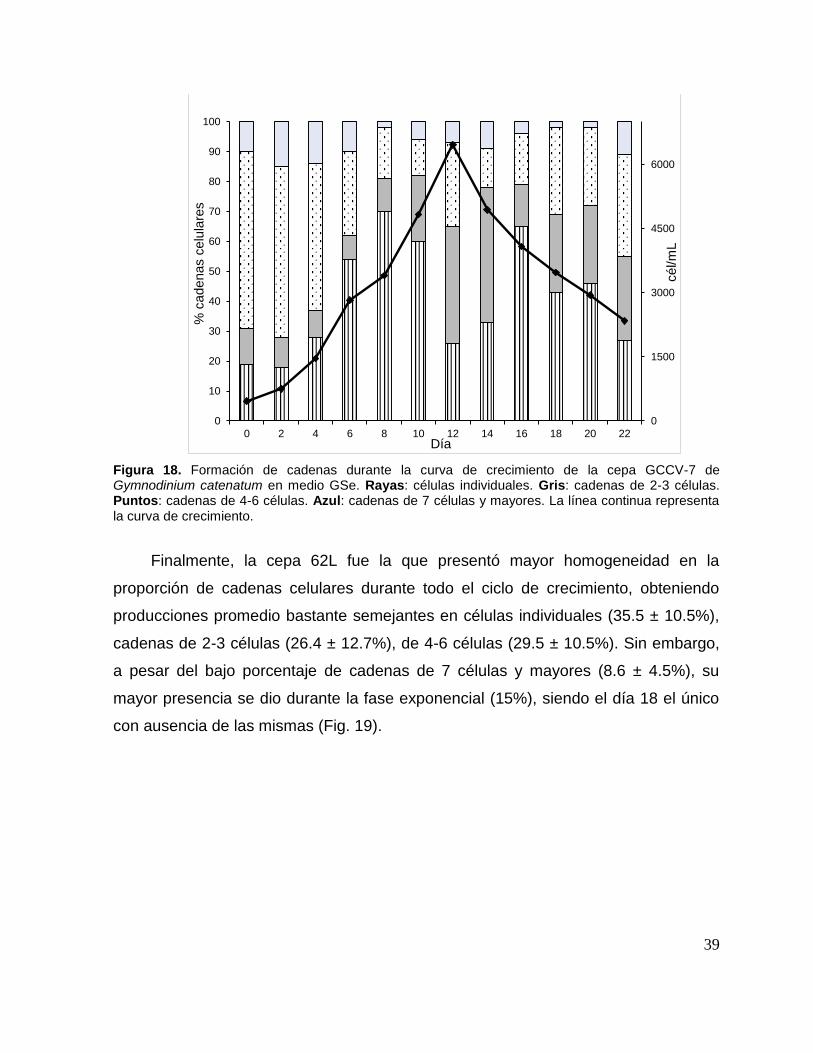
39
Figura 18. Formación de cadenas durante la curva de crecimiento de la cepa GCCV-7 de Gymnodinium catenatum en medio GSe. Rayas: células individuales. Gris: cadenas de 2-3 células. Puntos: cadenas de 4-6 células. Azul: cadenas de 7 células y mayores. La línea continua representa la curva de crecimiento.
Finalmente, la cepa 62L fue la que presentó mayor homogeneidad en la
proporción de cadenas celulares durante todo el ciclo de crecimiento, obteniendo
producciones promedio bastante semejantes en células individuales (35.5 ± 10.5%),
cadenas de 2-3 células (26.4 ± 12.7%), de 4-6 células (29.5 ± 10.5%). Sin embargo,
a pesar del bajo porcentaje de cadenas de 7 células y mayores (8.6 ± 4.5%), su
mayor presencia se dio durante la fase exponencial (15%), siendo el día 18 el único
con ausencia de las mismas (Fig. 19).
0
1500
3000
4500
6000
0
10
20
30
40
50
60
70
80
90
100
0 2 4 6 8 10 12 14 16 18 20 22
cél/m
L
% c
adenas c
elu
lare
s
Día

40
Figura 19. Formación de cadenas durante la curva de crecimiento de la cepa 62L de Gymnodinium catenatum en medio GSe. Puntos: cadenas de 4-6 células. Azul: cadenas de 7 células y mayores. La línea continua representa la curva de crecimiento.
7.5. Asociación G. catenatum-comunidad bacteriana
7.5.1. Exposición a filtrados celulares
En la figura 20 observamos que las curvas de crecimiento de los tratamientos y
el control tuvieron un comportamiento bastante similar, debido a que no se
encontraron diferencias significativas (H(3, N=16) = 4.966814; p<0.05) en la abundancia
celular. No obstante, el control fue el que presentó una mayor abundancia celular
(136,296 ± 34,735 cél/mL), seguido del tratamiento con 20, 15 y 10 mL de extracto.
La fase exponencial en todos los cultivos inició el día siete, mientras que la
biomasa máxima se obtuvo entre los 14 (±3.47) y 20 (±1.75) días. Las tasas de
crecimiento variaron de 0.0400 (± 0.01) a 0.0065 (± 0.0021) div/día, registrando
diferencias significativas (H(3, N=16) = 14.15929; p<0.05) únicamente para el control y
el tratamiento con 15 mL de extracto celular.
0
1500
3000
4500
6000
7500
0
10
20
30
40
50
60
70
80
90
100
0 2 4 6 8 10 12 14 16 18 20 22 24
cél/m
L
% cadenas c
elu
lare
s
Día
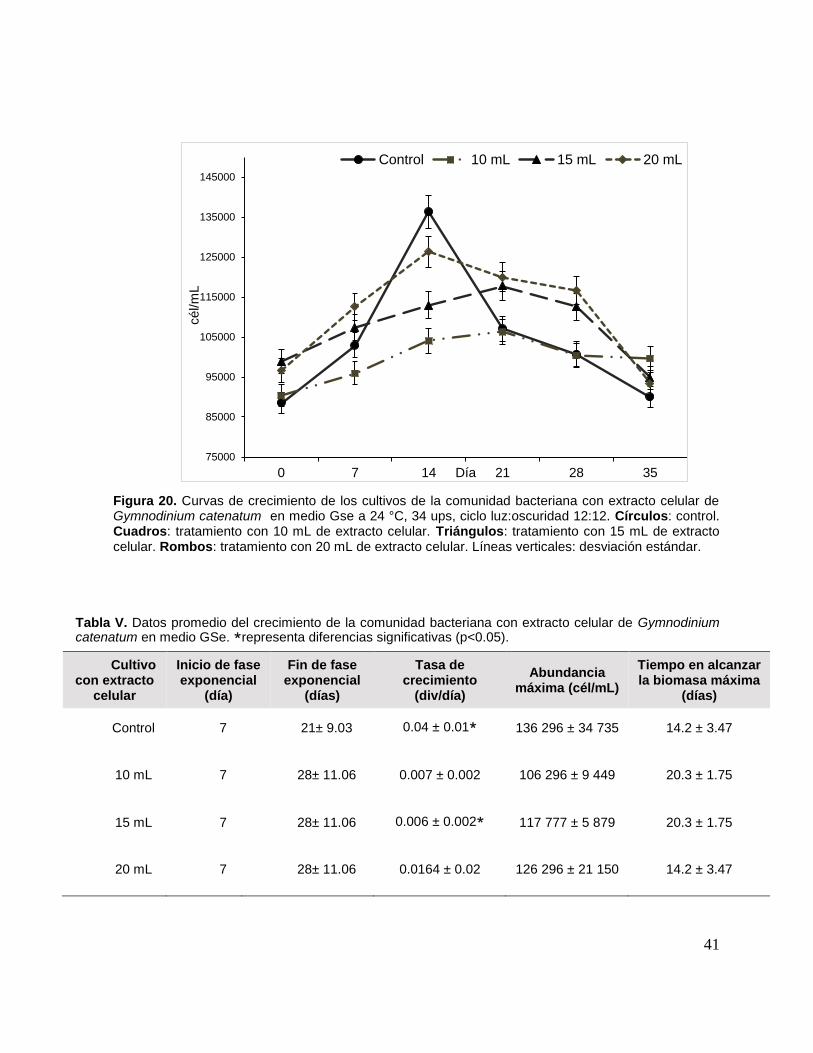
41
Figura 20. Curvas de crecimiento de los cultivos de la comunidad bacteriana con extracto celular de Gymnodinium catenatum en medio Gse a 24 °C, 34 ups, ciclo luz:oscuridad 12:12. Círculos: control. Cuadros: tratamiento con 10 mL de extracto celular. Triángulos: tratamiento con 15 mL de extracto celular. Rombos: tratamiento con 20 mL de extracto celular. Líneas verticales: desviación estándar.
Tabla V. Datos promedio del crecimiento de la comunidad bacteriana con extracto celular de Gymnodinium catenatum en medio GSe. *representa diferencias significativas (p<0.05).
75000
85000
95000
105000
115000
125000
135000
145000
0 7 14 21 28 35
cél/m
L
Día
Control 10 mL 15 mL 20 mL
Cultivo con extracto
celular
Inicio de fase exponencial
(día)
Fin de fase exponencial
(días)
Tasa de crecimiento
(div/día)
Abundancia máxima (cél/mL)
Tiempo en alcanzar la biomasa máxima
(días)
Control 7 21± 9.03 0.04 ± 0.01* 136 296 ± 34 735 14.2 ± 3.47
10 mL 7 28± 11.06 0.007 ± 0.002 106 296 ± 9 449 20.3 ± 1.75
15 mL 7 28± 11.06 0.006 ± 0.002* 117 777 ± 5 879 20.3 ± 1.75
20 mL 7 28± 11.06 0.0164 ± 0.02 126 296 ± 21 150 14.2 ± 3.47

42
7.5.2. Cultivos con contacto celular
Las cepas inoculadas con la comunidad bacteriana presentaron curvas de
crecimiento diferentes entre sí, caracterizadas por una fase de aclimatación corta,
excepto para la cepa GCCV-7 que tardó el doble de tiempo (4 ± 1.58 días) en
presentar la fase de crecimiento exponencial donde todas las cepas alcanzaron su
respectiva biomasa máxima. Finalmente, la fase de decaimiento se presentó rápida y
abruptamente, justo después de la fase exponencial tardía (Fig. 21).
La duración de las curvas fue variable, teniendo un mínimo de 8 (±1.47) y un
máximo de 26 (± 1.47) días, lapso en el que se alcanzaron las biomasas máximas
correspondientes, siendo la cepa GCCV-7 la de mayor densidad celular (6,932 ± 26
cél/mL), seguida de GCMV-7 con 6,334 (±25) cél/mL, G7 con 1146 (±4) cél/mL y
finalmente la cepa michoacana, 62L, con 425 (±11) cél/mL. A pesar de las
diferencias de biomasa en las cepas, el tiempo en alcanzarlas fue más homogéneo,
pues dos de las cuatro cepas obtuvieron su biomasa máxima el día 14 (±3.47)
mientras que GCCV-7 la obtuvo el día 22 (±2.93).
En cuanto a la cepa 62L, no presentó un crecimiento con respecto al tiempo
sino que decreció, registrando su biomasa máxima celular el día cuatro, por lo que
las tasas de crecimiento, quedaron comprendidas entre 0.043 (±0.002) y 0.248
(±0.01) div/día (Tabla VI).

43
Figura 21. Curvas de crecimiento de las cuatro cepas de Gymnodinium catenatum inoculadas con la comunidad bacteriana en medio GSe, a 24 °C, 34 ups, ciclo luz:oscuridad 12:12. Líneas verticales: desviación estándar.
Tabla VI. Datos promedio del crecimiento de las cepas de Gymnodinium catenatum inoculadas con la comunidad bacteriana en medio GSe.
Cepa Inicio de la
fase exponencial
Fin de fase exponencial
(días)
Tasa de crecimiento
(div/día)
Abundancia máxima (cél/mL)
Tiempo en alcanzar la biomasa máxima
(días)
GCCV-7 + CB 4 24 ± 1.47 0.109 ± 0.018 6932 ± 26 22.4 ± 2.93
GCMV-7 + CB 2 16 ± 2.63 0.164 ± 0.01 6334 ± 25 14.2 ± 3.47
G7 + CB 2 16 ± 2.63 0.043 ± 0.002 1146 ± 4 14.2 ± 3.47
62L + CB 2 6 ± 1.47 0.248 ± 0.02 628 ± 316 4.1 ± 1.23
7.6. Curvas de crecimiento
En la cepa GCCV-7, caracterizada por sus mayores abundancias, observamos
que la presencia de la comunidad bacteriana prolongó un par de días más el
0
1000
2000
3000
4000
5000
6000
7000
8000
0 2 4 6 8 10 12 14 16 18 20 22 24 26
cél/m
L
Día
GCCV-7 + CB GCMV-7 + CB G7 + CB 62L + CB

44
crecimiento (2.1 ± 0.9 días), aumentando significativamente la abundancia máxima
alcanzada ( de 6,463 a 6,932 ± 26 cél/mL), no obstante, cabe resaltar que tal
densidad celular se obtuvo en un tiempo mayor (10 ± 2.13 días) además de
presentarse cortas fases estacionarias, caracterizadas por 1) la presencia de quistes
temporales en aproximadamente un 15 % del total de las células mientras que
durante la fase de aclimatación este porcentaje era del 90 % y 2) cambios
estructurales en las células del dinoflagelado: desarreglo del cíngulo y sulcus,
pérdida de flagelos, cambios en los patrones del nado, núcleo prominente con
cromosomas evidentes y lisis celular. Aspecto que prevaleció en ambas curvas fue la
rápida fase de declive, precedida por la fase exponencial tardía (Fig. 22 A).
En el caso de la cepa GCMV-7, la presencia de la comunidad bacteriana le
favoreció significativamente, al obtener una abundancia máxima de casi el doble de
células (de 2,840 a 6,334 ± 25 cél/mL) en un menor tiempo (6 ± 1.47 días),
presentándose una fase de crecimiento exponencial muy marcada al igual que la de
declive. Observamos que la fase de aclimatación en ambos casos fue corta, sin
embargo, nuevamente la presencia de quistes temporales predominó en esta fase
con un 86 %, porcentaje que fue disminuyendo con el paso del tiempo, hasta el día
10 donde ya sólo se observaba que el 2 % del total de células se encontraban en
este estadio de quietud, lo cual coincidió con el inicio de la fase exponencial. Cabe
destacar que esta cepa también presentó deformaciones celulares como: pérdida de
flagelos, lisis de la membrana celular, presencia de una membrana hialina y núcleo
con cromosomas evidentes (Fig. 22 B).
Para la cepa G7, contrario a las dos cepas anteriores, la presencia de la
comunidad bacteriana no favoreció su crecimiento, al registrarse una mayor biomasa
en ausencia de la misma (1,625 ± 4 cél/mL). En cuanto a la fase de aclimatación, en
ambos casos fue de 2 días y no se presenciaron quistes temporales ni cambios
morfológicos en las células. La curva de crecimiento perteneciente al monocultivo de
G. catenatum presentó un ligero crecimiento exponencial en el día dos, seguido por
una corta fase estacionaria que dio lugar a la fase exponencial, contrario al cultivo

45
inoculado con la comunidad bacteriana que presentó un incremento gradual con
respecto al tiempo. Para los dos cultivos, la fase de declive fue rápida, posterior a la
obtención de la máxima biomasa celular (Fig. 22 C).
Al igual que la cepa G7, en 62 L no se vio favorecido su crecimiento en presencia de
la comunidad bacteriana. Todo lo contrario, pues a pesar de haber obtenido su
máxima biomasa al cuarto día, ésta no fue ni el 10 % del total que obtuvo en
ausencia de la misma (628 ± 316 cél/mL). La duración de ambas curvas es
significativamente diferente (16.4 ± 1.74 días; p<0.05), siendo mayor para el
monocultivo del dinoflagelado donde se observa un incremento gradual en la
abundancia celular mientras que el tratamiento mostró un descenso desde el inicio
del experimento. Cabe resaltar que este experimento se realizó en dos ocasiones,
obteniendo el mismo resultado: un descenso abrupto en la densidad celular de G.
catenatum reflejado en 1) altos porcentajes de quistes temporales (95 %) desde el
día de siembra (día cero); 2) deformaciones en la células, como: pérdida de flagelos,
desarreglo de cíngulo y sulcus, desprendimiento de una membrana hialina, núcleo
prominente con cromosomas visibles, o bien, 3) lisis celular (Fig. 22 D).

46
Figura 22. Curvas de crecimiento de las cuatro cepas de Gymnodinium catenatum en presencia (cuadros) / ausencia (círculos) de la comunidad bacteriana en medio GSe. De izquierda a derecha: A) GCCV-7. B) GCMV-7. C) G7. D) 62 L. *representa diferencias significativas (p<0.05). Líneas verticales: desviación estándar.
0
1000
2000
3000
4000
5000
6000
7000
8000
0 2 4 6 8 10 12 14 16 18 20 22 24 26
cél/m
L
Día
GCCV-7 GCCV-7 + CB
0
1000
2000
3000
4000
5000
6000
7000
0 2 4 6 8 10 12 14 16 18 20 22 24
cél/m
L
Día
GCMV-7 GCMV-7 + CB
0
400
800
1200
1600
2000
0 2 4 6 8 10 12 14 16 18 20
cél/m
L
Día
G7 G7 + CB
0
1000
2000
3000
4000
5000
6000
7000
0 2 4 6 8 10 12 14 16 18 20 22 24
cél/m
L
Día
62L 62L + CB
A
*
C
*
*
B
*
A
*
C*
D*
A* B*
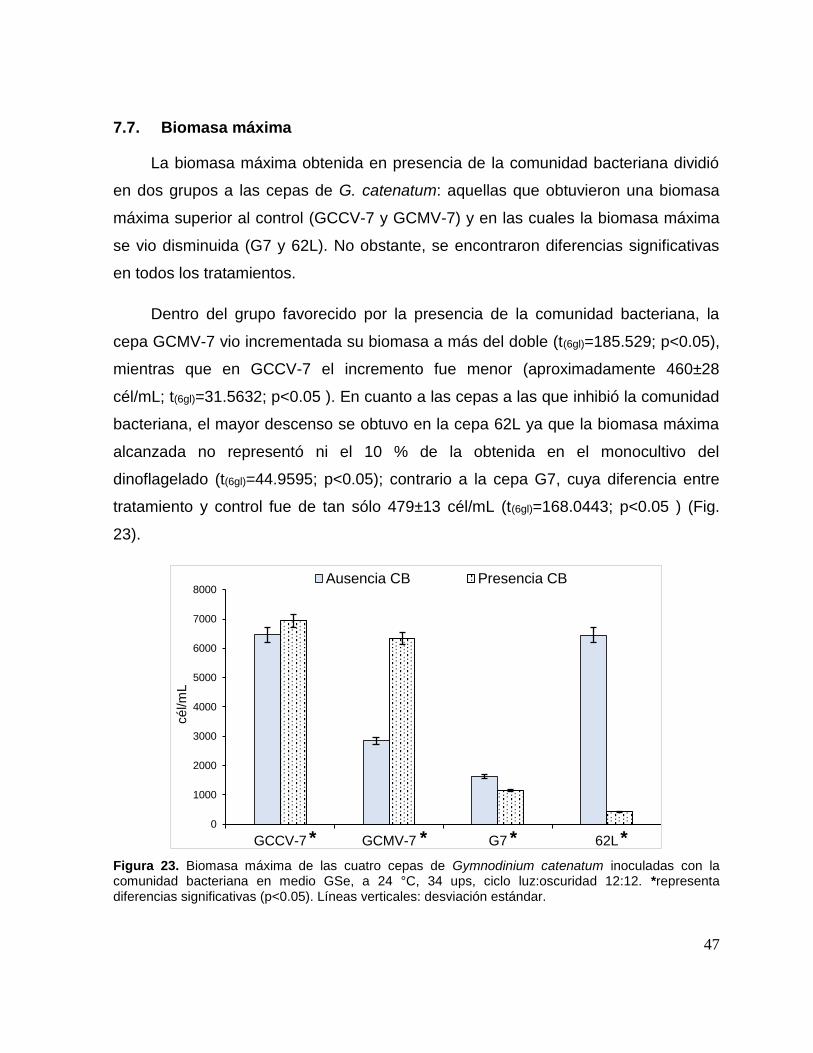
47
7.7. Biomasa máxima
La biomasa máxima obtenida en presencia de la comunidad bacteriana dividió
en dos grupos a las cepas de G. catenatum: aquellas que obtuvieron una biomasa
máxima superior al control (GCCV-7 y GCMV-7) y en las cuales la biomasa máxima
se vio disminuida (G7 y 62L). No obstante, se encontraron diferencias significativas
en todos los tratamientos.
Dentro del grupo favorecido por la presencia de la comunidad bacteriana, la
cepa GCMV-7 vio incrementada su biomasa a más del doble (t(6gl)=185.529; p<0.05),
mientras que en GCCV-7 el incremento fue menor (aproximadamente 460±28
cél/mL; t(6gl)=31.5632; p<0.05 ). En cuanto a las cepas a las que inhibió la comunidad
bacteriana, el mayor descenso se obtuvo en la cepa 62L ya que la biomasa máxima
alcanzada no representó ni el 10 % de la obtenida en el monocultivo del
dinoflagelado (t(6gl)=44.9595; p<0.05); contrario a la cepa G7, cuya diferencia entre
tratamiento y control fue de tan sólo 479±13 cél/mL (t(6gl)=168.0443; p<0.05 ) (Fig.
23).
Figura 23. Biomasa máxima de las cuatro cepas de Gymnodinium catenatum inoculadas con la comunidad bacteriana en medio GSe, a 24 °C, 34 ups, ciclo luz:oscuridad 12:12. *representa diferencias significativas (p<0.05). Líneas verticales: desviación estándar.
0
1000
2000
3000
4000
5000
6000
7000
8000
GCCV-7 GCMV-7 G7 62L
cél/m
L
Ausencia CB Presencia CB
* * * *
*
*
*

48
7.8. Tasa de crecimiento
De manera general, las cepas de G. catenatum inoculadas con la comunidad
bacteriana superaron las 0.1 div/día, con excepción de la cepa G7 (0.043 ± 0.002
div/día), observándose una tasa de crecimiento de entre 0.24 (±0.02) y 0.109
(±0.018) div/día.
Para los monocultivos del dinoflagelado, se obtuvieron tasas de crecimiento
menores a las 0.1 div/día y por tanto, también más bajas que las obtenidas en los
tratamientos de inoculación, con excepción de las cepas GCCV-7 (t(6gl)=1635.174) y
G7 (t(6gl)=7.1637) que registraron una mayor tasa de crecimiento en ausencia de la
comunidad bacteriana, mostrando diferencias significativas en ambos casos
(p<0.05).
En cuanto a las cepas GCMV-7 y 62L, también presentaron diferencias
significativas (p<0.05) (t(6gl)=905.097; t(6gl)=185.329, respectivamente) en las tasas de
crecimiento, siendo mayores en presencia de la comunidad bacteriana (Fig. 24).
Figura 24. Tasa de crecimiento de las cuatro cepas de Gymnodinium catenatum inoculadas con la comunidad bacteriana en medio GSe, a 24 °C, 34 ups, ciclo luz:oscuridad 12:12. *representa diferencias significativas (p<0.05). Líneas verticales: desviación estándar.
0
0.05
0.1
0.15
0.2
0.25
0.3
GCCV-7 GCMV-7 G7 62L
div
/día
Ausencia CB Presencia CB
*
*
*
*
* * * *
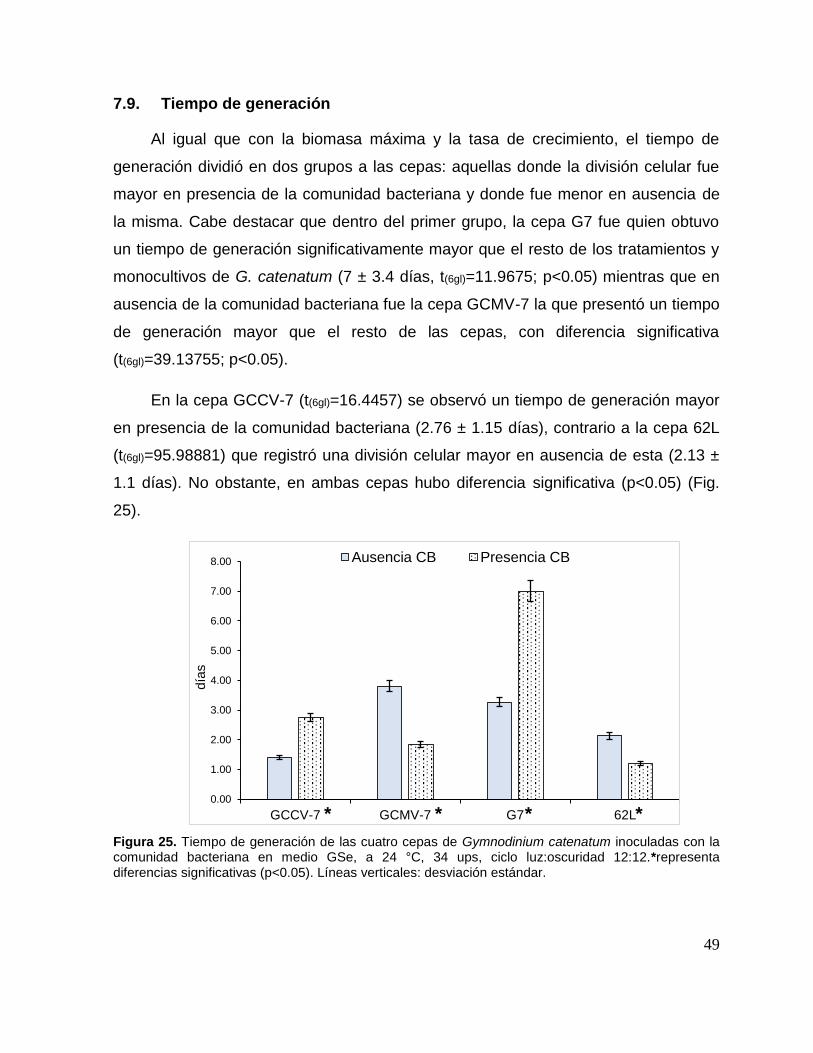
49
7.9. Tiempo de generación
Al igual que con la biomasa máxima y la tasa de crecimiento, el tiempo de
generación dividió en dos grupos a las cepas: aquellas donde la división celular fue
mayor en presencia de la comunidad bacteriana y donde fue menor en ausencia de
la misma. Cabe destacar que dentro del primer grupo, la cepa G7 fue quien obtuvo
un tiempo de generación significativamente mayor que el resto de los tratamientos y
monocultivos de G. catenatum (7 ± 3.4 días, t(6gl)=11.9675; p<0.05) mientras que en
ausencia de la comunidad bacteriana fue la cepa GCMV-7 la que presentó un tiempo
de generación mayor que el resto de las cepas, con diferencia significativa
(t(6gl)=39.13755; p<0.05).
En la cepa GCCV-7 (t(6gl)=16.4457) se observó un tiempo de generación mayor
en presencia de la comunidad bacteriana (2.76 ± 1.15 días), contrario a la cepa 62L
(t(6gl)=95.98881) que registró una división celular mayor en ausencia de esta (2.13 ±
1.1 días). No obstante, en ambas cepas hubo diferencia significativa (p<0.05) (Fig.
25).
Figura 25. Tiempo de generación de las cuatro cepas de Gymnodinium catenatum inoculadas con la comunidad bacteriana en medio GSe, a 24 °C, 34 ups, ciclo luz:oscuridad 12:12.*representa diferencias significativas (p<0.05). Líneas verticales: desviación estándar.
0.00
1.00
2.00
3.00
4.00
5.00
6.00
7.00
8.00
GCCV-7 GCMV-7 G7 62L
día
s
Ausencia CB Presencia CB
* * * *
*
*

50
7.10. Longitud de cadenas celulares
La presencia de la comunidad bacteriana en la cepa GCMV-7 durante todo el
ciclo de crecimiento, disminuyó la proporción de céluas individuales (de 32.7 ± 13.8 a
a 20 ± 11 %) e incrementó, de manera significativa (X2 0.05,3) = 6.4836; p<0.05) la
producción de cadenas celulares de 4-6 células (de 18.5 ± 10.3 a 31 ± 9 %) y de 7
células y mayores (11.4 ± 3.2 a 14 ± 10.2 %). En ambos casos la fase de
aclimatación fue corta, dominando las células individuales en el control mientras que
en el tratamiento las cadenas de 2-3 células, así como de 4-6 células fueron las
predominantes (X2 0.05,3) = 73.2054; p<0.05). Durante la fase de crecimiento
exponencial, en los dos cultivos se fue incrementando la producción de cadenas
celulares más largas, observándose una diferencia significativa (X2 0.05,3) = 21.155;
p<0.05) en las cadenas de 2-3 células, mientras que al llegar al final de esta fase y
comenzar con el declive, la tendencia fue invirtiendose, registrando una mayor
producción de células individuales en el tratamiento y aumentando la proporción de
cadenas de 2-3 y 4-6 células en el control (X2 0.05,3) = 8.9330; p<0.05), quien mantuvo
dicha producción de cadenas celulares por cuatro días más (Fig. 26; A y A’).
Para la cepa G7, en presencia de la comunidad bacteriana se tuvo un
predominio de células individuales, (38.1 ± 14.6 %) contrario al control, donde
dominaron las cadenas de 7 células y mayores (36. 1 ± 17 %). No obstante y a pesar
de haberse mantenido ese patrón durante la mayor parte del ciclo de crecimiento, en
los días de aclimatación e inicio de la fase exponencial, la producción de cadenas de
4-6 células aumentó significativamente (de 25.5 ± 6.3 a 41.5 ± 3.5 %; X2 0.05,3) = 73.
62.7859; p<0.05). Dicho patrón se mantuvo durante la fase exponencial,
incrementando también la producción de cadenas de 2-3 células, sin embargo, las
cadenas de 7 células y mayores no predominaron hasta el declive, donde se tuvo
una producción de células individuales y cadenas celulares homogénea
(aproximadamente del 25 %). Resaltemos que nuevamente el control presentó una
prologación de dos días en su crecimiento, durante los cuales la proporción de
cadenas celulares de 7 células y mayores fueron las dominantes (Fig. 26; B y B’).

51
La cepa GCCV-7 se caracterizó por presentar una mayor variabilidad en la
formación de cadenas celulares a lo largo de su crecimiento, obteniendo una
disminución considerable en la proporción de células individuales (de 40.7 ± 18.1 a
26 ± 21.4 %) y aumentando la producción de cadenas de 7 células y mayores (de 7.6
± 4.5 a 17 ± 10.6 %). Para las cadenas celulares de 2-3 y 4-6 células no se
observaron cambios, logrando una proporción de 23 y 31 %, respectivamente (X2
0.05,3) = 22.6090; p<0.05) (Fig. 27; C y C’).
Finalmente, en la cepa 62L en presencia de la comunidad bacteriana, las
células individuales mantuvieron una baja producción a lo largo del cultivo al igual
que las cadenas de 2-3 células, mientras que las cadenas correspondientes a 4-6 y 7
células y mayores predominaron (X2 0.05,3) = 65.3825; p<0.05) (Fig. 27; D y D’).

52
Figura 26. Formación de cadenas durante la curva de crecimiento de las cepas GCMV-7 (A y A’) y G7 (B y B’) de Gymnodinium catenatum en presencia/ausencia de la comunidad bacteriana. Rayas: células individuales. Gris: cadenas de 2-3 células. Puntos: cadenas de 4-6 células. Azul: cadenas de
7 células y mayores. La línea continua representa la curva de crecimiento del control y la línea semi-continua el tratamiento.
0
1000
2000
3000
0
10
20
30
40
50
60
70
80
90
100
0 2 4 6 8 10 12 14 16 18 20 22 24
cél/m
L
% d
e c
adenas c
elu
lare
s
Día
0
2000
4000
6000
8000
0
10
20
30
40
50
60
70
80
90
100
0 2 4 6 8 10 12 14 16 18 20
cé
l/m
L
% d
e c
adenas c
elu
lare
s
Día
0
500
1000
1500
0
10
20
30
40
50
60
70
80
90
100
0 2 4 6 8 10 12 14 16 18 20
cél/m
L
% c
adenas c
elu
lare
s
Día
0
400
800
1200
0
10
20
30
40
50
60
70
80
90
100
0 2 4 6 8 10 12 14 16 18
cél/m
L
% c
adenas c
elu
lare
s
Día
GCMV-7
GCMV-7 + CB
G
7
G7 + CB
A
A’
B
B’
G7
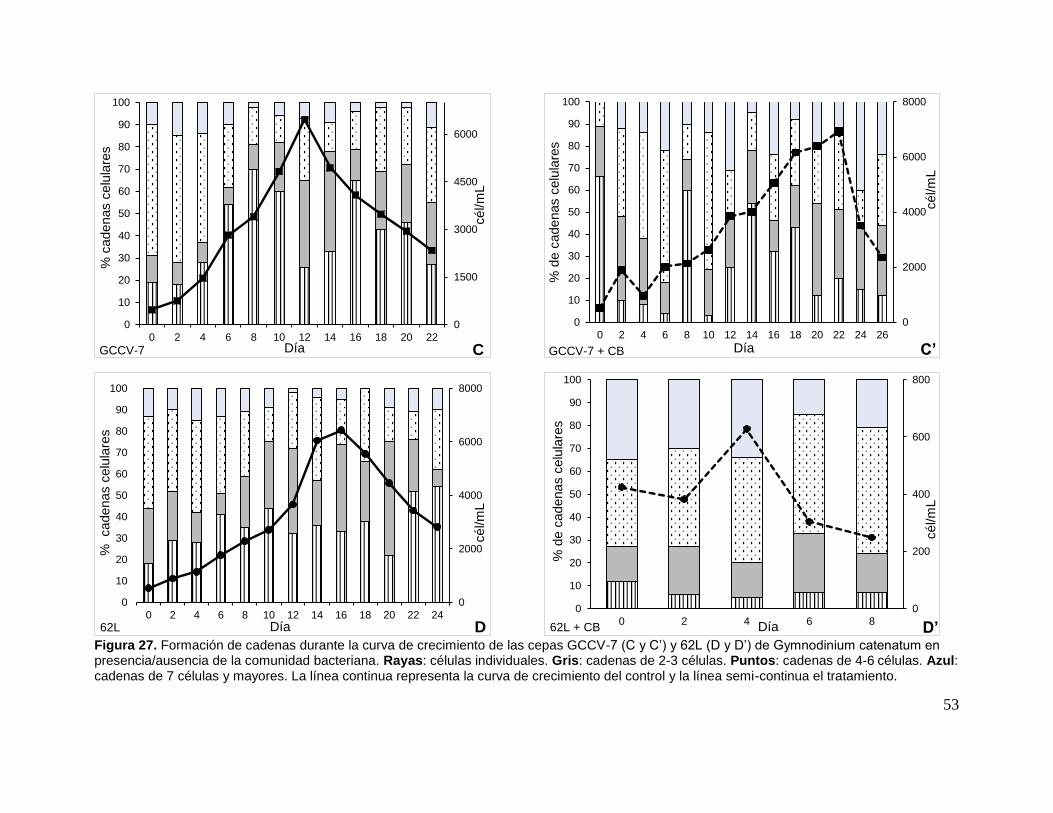
53
Figura 27. Formación de cadenas durante la curva de crecimiento de las cepas GCCV-7 (C y C’) y 62L (D y D’) de Gymnodinium catenatum en presencia/ausencia de la comunidad bacteriana. Rayas: células individuales. Gris: cadenas de 2-3 células. Puntos: cadenas de 4-6 células. Azul: cadenas de 7 células y mayores. La línea continua representa la curva de crecimiento del control y la línea semi-continua el tratamiento.
0
1500
3000
4500
6000
0
10
20
30
40
50
60
70
80
90
100
0 2 4 6 8 10 12 14 16 18 20 22
cél/m
L
% c
adenas c
elu
lare
s
Día
0
2000
4000
6000
8000
0
10
20
30
40
50
60
70
80
90
100
0 2 4 6 8 10 12 14 16 18 20 22 24 26
cél/m
L
% d
e c
adenas c
elu
lare
s
Día
0
2000
4000
6000
8000
0
10
20
30
40
50
60
70
80
90
100
0 2 4 6 8 10 12 14 16 18 20 22 24
cél/m
L
% cadenas c
elu
lare
s
Día
0
200
400
600
800
0
10
20
30
40
50
60
70
80
90
100
0 2 4 6 8
cél/m
L
% d
e c
adenas c
elu
lare
s
Día
GCCV-7
C
GCCV-7 + CB
C’
62L
D
62L + CB
D’

54
7.11. Curvas de crecimiento de la comunidad bacteriana en los cultivos con
contacto celular
Se evaluó el crecimiento de la comunidad bacteriana asociada a las cepas
tratamiento de G. catenatum, observándose un patrón general para todos los
cultivos: la abundancia de la cianobacteria aumentaba, mientras que los bacilos y las
bacterias heterótrofas filamentosas disminuían.
En la cepa GCCV-7, al inicio del ciclo de crecimiento dominaban los bacilos y
filamentos, sin embargo, a partir del día 12, los filamentos comenzaron a decrecer
mientras que los bacilos, al registrar un pequeño declive el día ocho, comenzaron a
recuperar su crecimiento los siguientes días. En cuanto a la cianobacteria, desde el
día de siembra mantuvo un crecimiento constante y gradual, obteniendo la
abundancia máxima el día 24 (Fig. 28 A).
En la cepa GCMV-7 se observa que al iniciar el ciclo de crecimiento, la
cianobacteria tenía una abundancia baja, a partir del día cuatro comienza la fase
exponencial que termina con la obtención de la mayor abundancia el día 20, mientras
que los bacilos inician con una abundancia alta que va en declive a partir del cuarto
día para después mantener constante su crecimiento. Finalmente, los filamentos
presentaron dos puntos mínimos de crecimiento, el primero en el día cuatro y el
segundo al dieciseisavo día para posteriormente registrar un crecimiento exponencial
(Fig. 28 B).
Nuevamente observamos que en la cepa G7, la cianobacteria inicia su
crecimiento con una abundancia más baja que los bacilos y filamentos, la cual va en
incremento hasta el octavo día, donde observamos una fase estacionaria de cuatro
días, seguida de la fase exponencial que alberga la máxima densidad celular en el
día 16. Las bacterias heterótrofas filamentosas presentaron un crecimiento peculiar,
ya que comienzan el ciclo con un crecimiento gradual y a la alza, el cual se ve
interrumpido desde el día ocho dando lugar a la fase estacionaria y después al
declive, caso contrario a los bacilos, que a pesar de registrar un ligero declive el

55
mismo día, vuelven a presentar un crecimiento exponencial que se ve diezmado el
día 12 (Fig. 28 C).
La excepción al patrón de crecimiento antes descrito fue la cepa 62L, debido a
que la cianobacteria presentó una abundancia mayor que la de los bacilos y
filamentos, además que desde el día de siembra comenzó con un crecimiento
exponencial. Los filamentos mantuvieron un crecimiento constante a lo largo del ciclo
mientras que los bacilos, al igual que la cianobacteria, desde el día cero presentaron
un crecimiento exponencial, no obstante, no lograron alcanzar una abundancia
mayor que esta última (Fig. 28 D).
De manera específica para la cianobacteria, únicamente las cepas G7 y GCCV-
7 registraron diferencias significativas (H(3, N=16) = 14.13844; p<0.05) (Fig. 28). La tasa
de crecimiento varió entre 0.040 (±0.021) y 0.12 (±0.01) div/día, mostrando
diferencias significativas (H(3, N=16) = 14.15929; p<0.05) sólo para 62L y GCMV-7 (Fig.
29). El tiempo de generación entre cepas varió entre 2.5 (±1.04) y 7.34 (±2.44) días y
mostró diferencias significativas (H(3, N=16) = 14.13844; p<0.05), entre las cepas 62L y
GCMV-7, observándose tres grupos con características similares (Fig. 30).
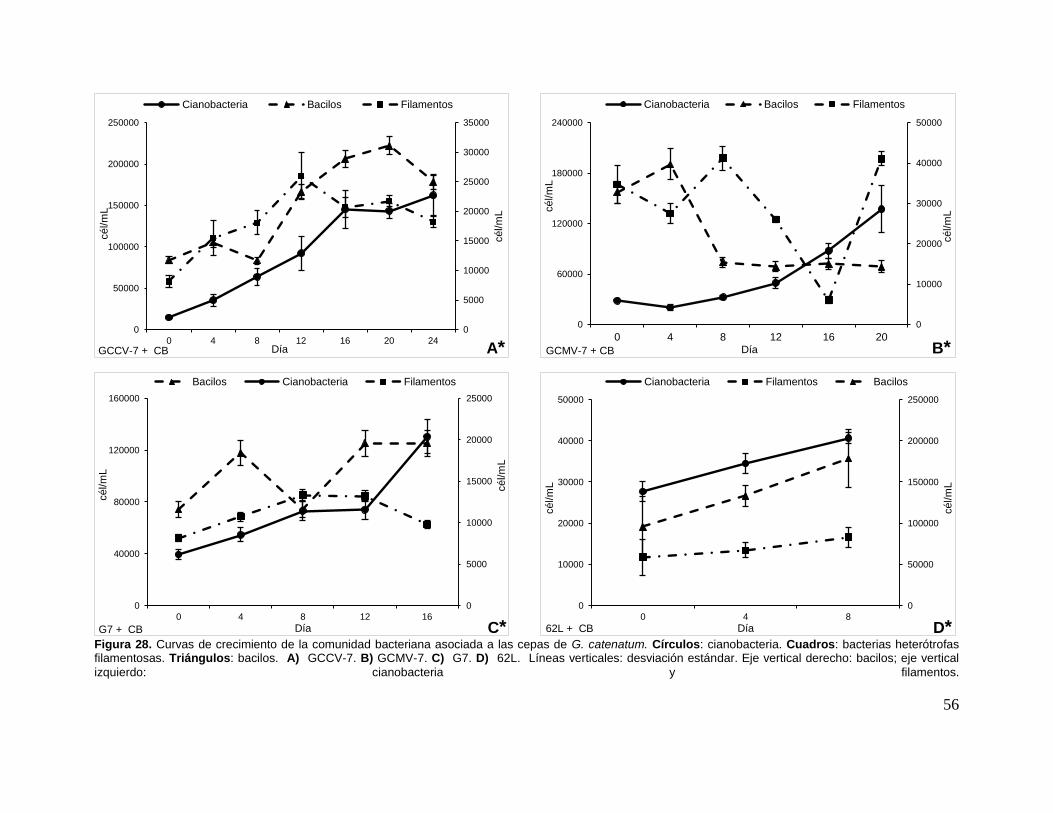
56
Figura 28. Curvas de crecimiento de la comunidad bacteriana asociada a las cepas de G. catenatum. Círculos: cianobacteria. Cuadros: bacterias heterótrofas filamentosas. Triángulos: bacilos. A) GCCV-7. B) GCMV-7. C) G7. D) 62L. Líneas verticales: desviación estándar. Eje vertical derecho: bacilos; eje vertical
izquierdo: cianobacteria y filamentos.
0
5000
10000
15000
20000
25000
30000
35000
0
50000
100000
150000
200000
250000
0 4 8 12 16 20 24
cél/m
L
cél/m
L
Día
Cianobacteria Bacilos Filamentos
0
10000
20000
30000
40000
50000
0
60000
120000
180000
240000
0 4 8 12 16 20
cél/m
Lcé
l/m
L
Día
Cianobacteria Bacilos Filamentos
0
5000
10000
15000
20000
25000
0
40000
80000
120000
160000
0 4 8 12 16
cé
l/m
L
cél/m
L
Día
Bacilos Cianobacteria Filamentos
0
50000
100000
150000
200000
250000
0
10000
20000
30000
40000
50000
0 4 8
cél/m
L
cél/m
L
Día
Cianobacteria Filamentos Bacilos
GCCV-7 + CB
GCMV-7 + CB
G7 + CB
62L + CB
A*
B*
C*
D*

57
Figura 29. Tasa de crecimiento de la cianobacteria en los cultivos tratamiento de G. catenatum en medio GSe, a 24° C, 34 ups, ciclo luz:oscuridad 12:12. * representa diferencias significativas (p<0.05). Líneas verticales: desviación estándar.
Figura 30. Tiempo de generación de la cianobacteria en los cultivos tratamiento de Gymnodinium catenatum en medio GSe, a 24 °C, 34 ups, ciclo luz:oscuridad 12:12. Letras diferentes representan diferencias significativas (p<0.05). Líneas verticales: desviación estándar.
0.000
0.020
0.040
0.060
0.080
0.100
0.120
0.140
GCCV-7 GCMV-7 G7 62L
div
/día
0.00
2.00
4.00
6.00
8.00
10.00
GCCV-7 GCMV-7 G7 62L
día
s
*
*
abc
abc
b
c
*

58
7.12. Cambios morfológicos de Gymnodinium catenatum en presencia de la
comunidad bacteriana
Los principales cambios celulares observados en células de G. catenatum, en
los experimentos con contacto celular fueron: 1) desarreglo en el sulcusy cíngulo, 2)
ruptura de la membrana celular, 3) células con forma aplanada o de trapecio, 4)
desarreglos en el material citoplasmático, 5) núcleos prominentes con cromosomas
condensados, 6) quistes temporales y 7) lisis celular. Estos patrones fueron
visualizados en los tratamientos, con excepción de la cepa G7, a lo largo del ciclo de
crecimiento, presentándose en altos porcentajes (80 – 90 %) en el día de siembra y
en la fase de aclimatación, conforme el cultivo iba creciendo este porcentaje fue
disminuyendo, volviendo a presentarse en la mayoría de las células posterior a la
fase exponencial tardía. En las cepas GCCV-7 y GCMV-7, además de los cambios
morfológicos antes descritos, se observó un cambio en la coloración en los
tratamientos, los cuales se tornaron de un tono verde limón muy evidente. Esta
coloración se presentó a partir del día 8 y 10, respectivamente, coincidiendo con el
inicio de la fase exponencial. Sin embargo, no se vio inhibido el crecimiento de G.
catenatum (Fig. 31 y 32).
Figura 31. Cultivos inoculados con la comunidad bacteriana. A) GCMV-7 en el día seis de cultivo, sin cambio en la coloración. B) GCCV-7 en el día diez de cultivo, con un cambio de color evidente. C) Comparación en la tonalidad del cultivo, día seis del control y del tratamiento de la cepa GCCV-7.

59
Figura 32. Células de G. catenatum en presencia de la comunidad bacteriana. A) Célula individual del control sin cambios aparentes. B) Célula rodeada de una membrana característica de un quiste temporal. C) Ruptura de la membrana, pérdida de flagelo y desarreglo del cíngulo y sulcus. D) Quiste temporal rodeado de bacterias. E) Célula con forma trapezoide. F) Célula redondeada con núcleo evidente y cromosomas condensados. G) Cadena de cuatro células del control. H) Cadena celular rodeada por membrana hialina. I) Desarreglo del cíngulo y sulcus. J) Ruptura de la membrana celular. K) Desarreglos citoplasmáticos. L) Lisis celular.
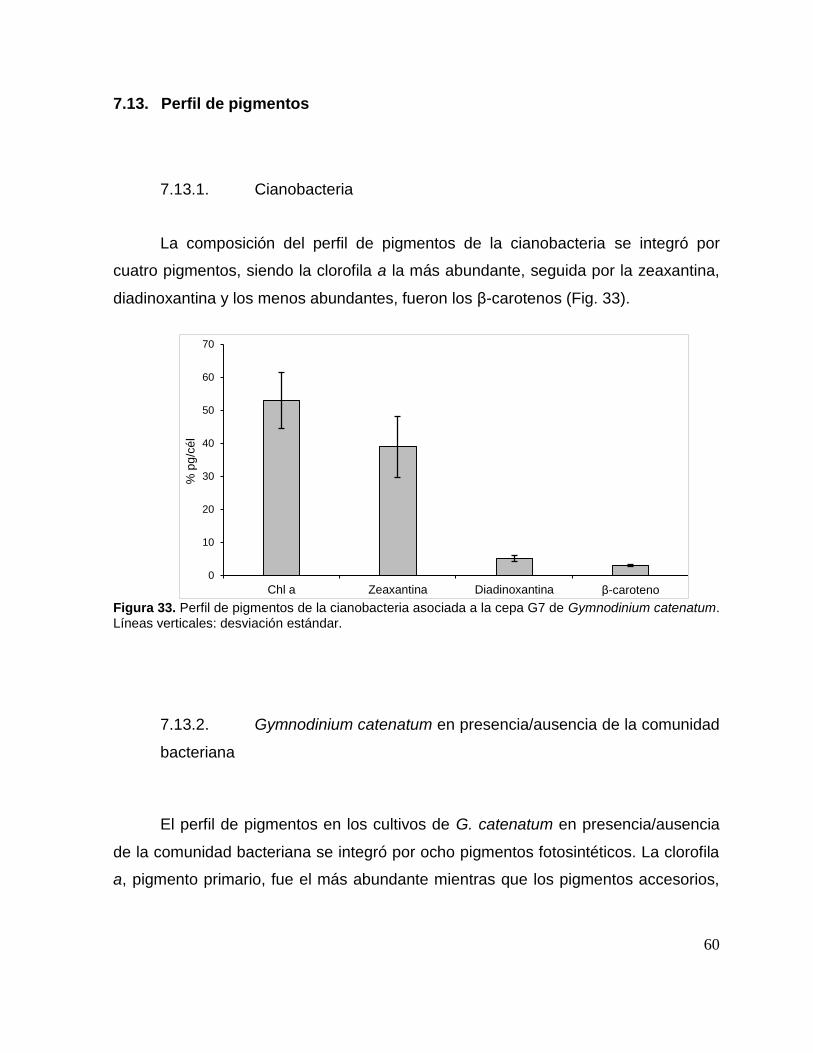
60
7.13. Perfil de pigmentos
7.13.1. Cianobacteria
La composición del perfil de pigmentos de la cianobacteria se integró por
cuatro pigmentos, siendo la clorofila a la más abundante, seguida por la zeaxantina,
diadinoxantina y los menos abundantes, fueron los β-carotenos (Fig. 33).
Figura 33. Perfil de pigmentos de la cianobacteria asociada a la cepa G7 de Gymnodinium catenatum. Líneas verticales: desviación estándar.
7.13.2. Gymnodinium catenatum en presencia/ausencia de la comunidad
bacteriana
El perfil de pigmentos en los cultivos de G. catenatum en presencia/ausencia
de la comunidad bacteriana se integró por ocho pigmentos fotosintéticos. La clorofila
a, pigmento primario, fue el más abundante mientras que los pigmentos accesorios,
0
10
20
30
40
50
60
70
Chl a Zeaxantina Diadinoxantina β-caroteno
% p
g/c
él
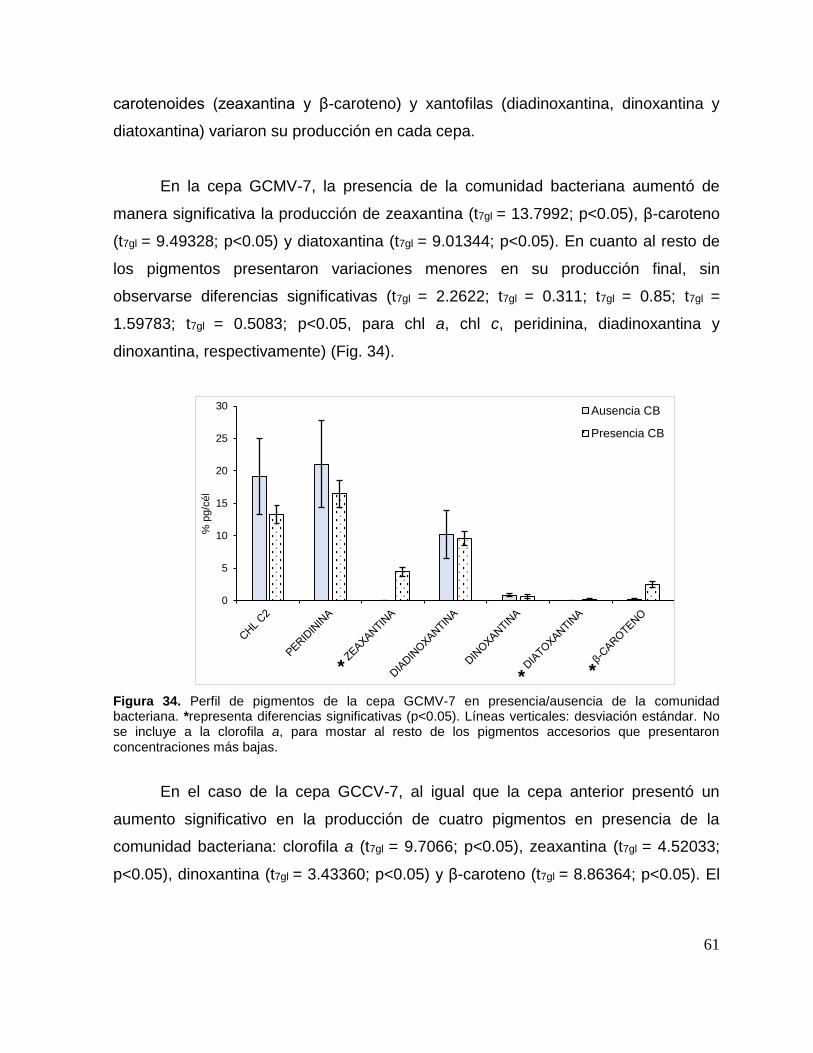
61
carotenoides (zeaxantina y β-caroteno) y xantofilas (diadinoxantina, dinoxantina y
diatoxantina) variaron su producción en cada cepa.
En la cepa GCMV-7, la presencia de la comunidad bacteriana aumentó de
manera significativa la producción de zeaxantina (t7gl = 13.7992; p<0.05), β-caroteno
(t7gl = 9.49328; p<0.05) y diatoxantina (t7gl = 9.01344; p<0.05). En cuanto al resto de
los pigmentos presentaron variaciones menores en su producción final, sin
observarse diferencias significativas (t7gl = 2.2622; t7gl = 0.311; t7gl = 0.85; t7gl =
1.59783; t7gl = 0.5083; p<0.05, para chl a, chl c, peridinina, diadinoxantina y
dinoxantina, respectivamente) (Fig. 34).
Figura 34. Perfil de pigmentos de la cepa GCMV-7 en presencia/ausencia de la comunidad bacteriana. *representa diferencias significativas (p<0.05). Líneas verticales: desviación estándar. No se incluye a la clorofila a, para mostar al resto de los pigmentos accesorios que presentaron concentraciones más bajas.
En el caso de la cepa GCCV-7, al igual que la cepa anterior presentó un
aumento significativo en la producción de cuatro pigmentos en presencia de la
comunidad bacteriana: clorofila a (t7gl = 9.7066; p<0.05), zeaxantina (t7gl = 4.52033;
p<0.05), dinoxantina (t7gl = 3.43360; p<0.05) y β-caroteno (t7gl = 8.86364; p<0.05). El
0
5
10
15
20
25
30
% p
g/c
él
Ausencia CB
Presencia CB
*
*
*
*
*
*

62
resto de los pigmentos tuvo una producción similar en presencia/ausencia de la
comunidad (Fig. 35).
Figura 35. Perfil de pigmentos de la cepa GCCV-7 en presencia/ausencia de la comunidad bacteriana. *representa diferencias significativas (p<0.05). Líneas verticales: desviación estándar. No se incluye a la clorofila a, para mostar al resto de los pigmentos accesorios que presentaron concentraciones más bajas.
La cepa 62L en presencia de la comunidad bacteriana no registró cambios
significativos (t7gl = 0.304539; t7gl = 1.148394; t7gl = 0.504810; t7gl = 2.25283; t7gl =
0.026127; t7gl = 0.893056; t7gl = 0.739977; t7gl = 0.026649; p<0.05, para chl a, chl c,
peridinina, zeaxantina, diadinoxantina, dinoxantina, diatoxantina y β-carotenos,
respectivamente)en la producción de pigmentos, manteniendo su producción
homogénea (Fig. 36).
0
5
10
15
20
25
% p
g/c
él
Ausencia CB
Presencia CB
*
*
*
*
*
*
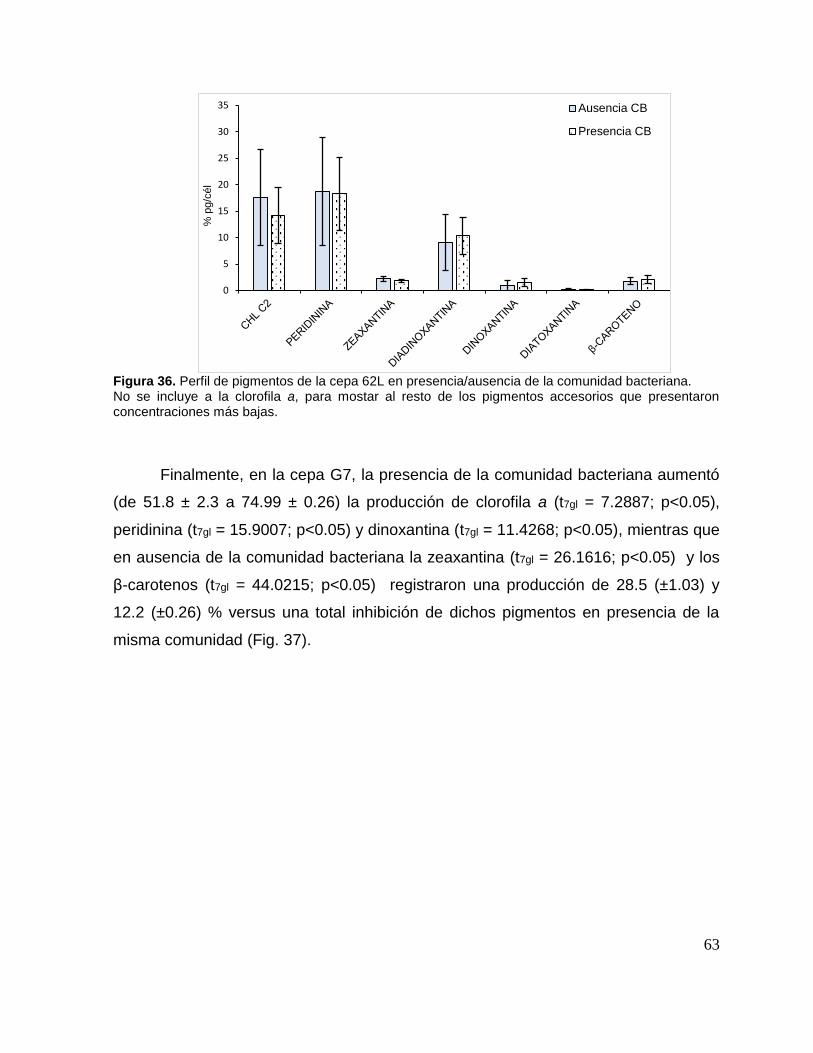
63
Figura 36. Perfil de pigmentos de la cepa 62L en presencia/ausencia de la comunidad bacteriana. No se incluye a la clorofila a, para mostar al resto de los pigmentos accesorios que presentaron concentraciones más bajas.
Finalmente, en la cepa G7, la presencia de la comunidad bacteriana aumentó
(de 51.8 ± 2.3 a 74.99 ± 0.26) la producción de clorofila a (t7gl = 7.2887; p<0.05),
peridinina (t7gl = 15.9007; p<0.05) y dinoxantina (t7gl = 11.4268; p<0.05), mientras que
en ausencia de la comunidad bacteriana la zeaxantina (t7gl = 26.1616; p<0.05) y los
β-carotenos (t7gl = 44.0215; p<0.05) registraron una producción de 28.5 (±1.03) y
12.2 (±0.26) % versus una total inhibición de dichos pigmentos en presencia de la
misma comunidad (Fig. 37).
0
5
10
15
20
25
30
35
% p
g/c
él
Ausencia CB
Presencia CB

64
Figura 37. Perfil de pigmentos de la cepa G7 en presencia/ausencia de la comunidad bacteriana. *representa diferencias significativas (p<0.05). Líneas verticales: desviación estándar. No se incluye a la clorofila a, para mostar al resto de los pigmentos accesorios que presentaron concentraciones más bajas
7.14. Perfil de toxinas paralizantes
Los diversos análogos producidos por Gymnodinium catenatum han sido
divididos en función del grupo sustituyente en la cadena lateral, en carbamoil (STX,
NEO, GTX1-4), decarbamoil (dcSTX, dcNEO, dcGTX1-4) y N-sulfocarbamoil (B1-2 y
C1-2), además que los pares enantioméricos se expresan como sumatorias, ya que
se epimerizan con facilidad bajo las condiciones de extracción y análisis. A pesar de
que pueden ser determinados de manera individual, las cantidades no reflejan
necesariamente la composición original en las células.
Se identificaron ocho toxinas: las N-sulfocarbamoil C1/2; las decarbamoil
dcNEO, dcSTX y dcGTX2/3 y las carbamoil GTX2/3.
Por grupo de toxinas y en la cepa GCMV-7, observamos que las toxinas N-
sulfocarbamoil, en presencia/ausencia de la comunidad bacteriana fueron las
0
5
10
15
20
25
30
35
% p
g/c
él
Ausencia CB
Presencia CB
*
*
*
*
*
*
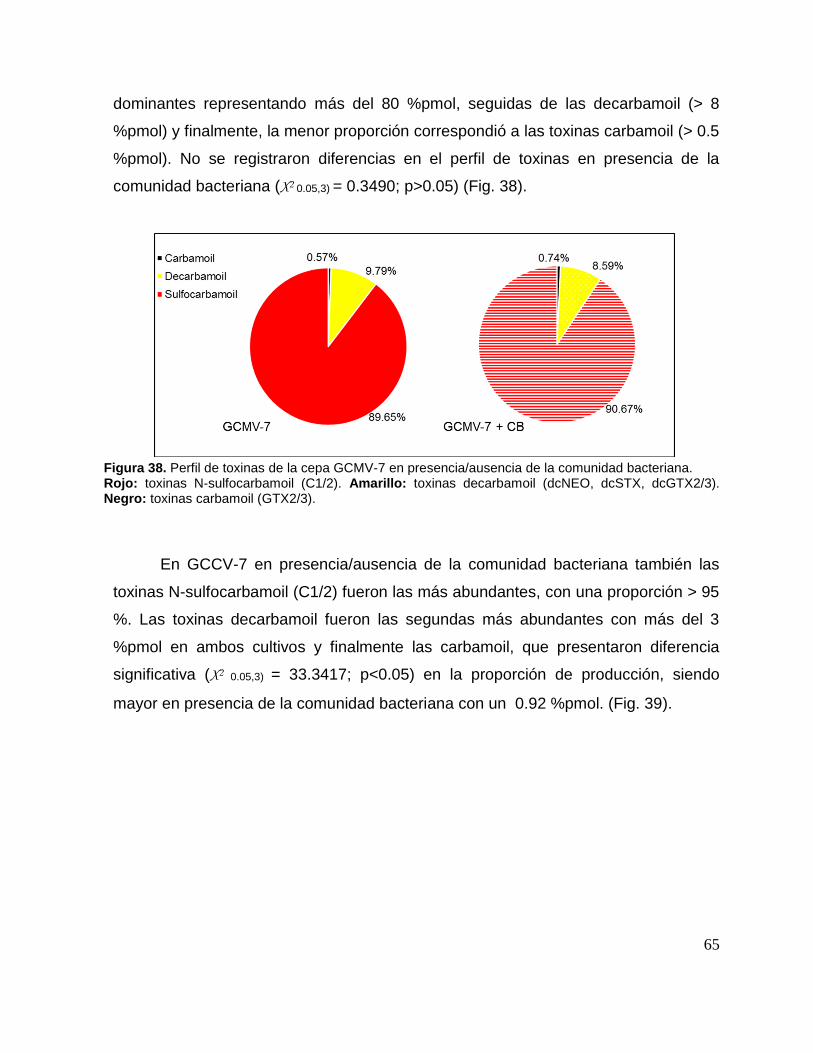
65
dominantes representando más del 80 %pmol, seguidas de las decarbamoil (> 8
%pmol) y finalmente, la menor proporción correspondió a las toxinas carbamoil (> 0.5
%pmol). No se registraron diferencias en el perfil de toxinas en presencia de la
comunidad bacteriana (X2 0.05,3) = 0.3490; p>0.05) (Fig. 38).
Figura 38. Perfil de toxinas de la cepa GCMV-7 en presencia/ausencia de la comunidad bacteriana. Rojo: toxinas N-sulfocarbamoil (C1/2). Amarillo: toxinas decarbamoil (dcNEO, dcSTX, dcGTX2/3). Negro: toxinas carbamoil (GTX2/3).
En GCCV-7 en presencia/ausencia de la comunidad bacteriana también las
toxinas N-sulfocarbamoil (C1/2) fueron las más abundantes, con una proporción > 95
%. Las toxinas decarbamoil fueron las segundas más abundantes con más del 3
%pmol en ambos cultivos y finalmente las carbamoil, que presentaron diferencia
significativa (X2 0.05,3) = 33.3417; p<0.05) en la proporción de producción, siendo
mayor en presencia de la comunidad bacteriana con un 0.92 %pmol. (Fig. 39).
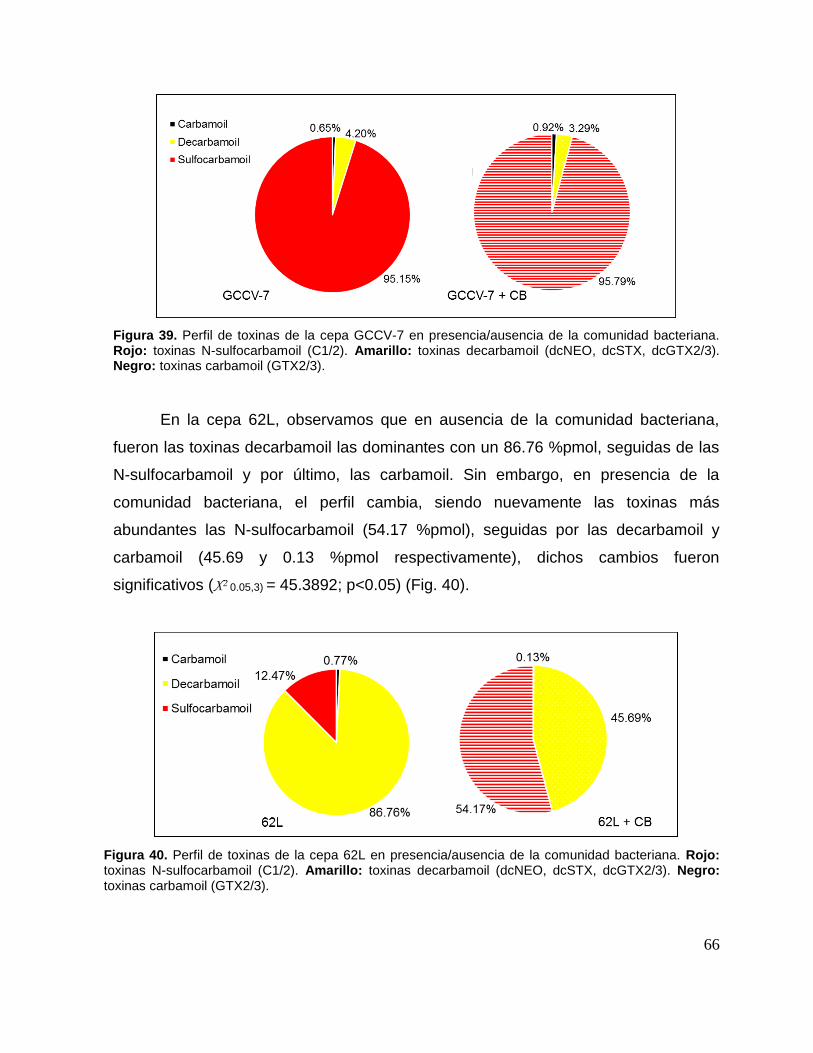
66
Figura 39. Perfil de toxinas de la cepa GCCV-7 en presencia/ausencia de la comunidad bacteriana. Rojo: toxinas N-sulfocarbamoil (C1/2). Amarillo: toxinas decarbamoil (dcNEO, dcSTX, dcGTX2/3). Negro: toxinas carbamoil (GTX2/3).
En la cepa 62L, observamos que en ausencia de la comunidad bacteriana,
fueron las toxinas decarbamoil las dominantes con un 86.76 %pmol, seguidas de las
N-sulfocarbamoil y por último, las carbamoil. Sin embargo, en presencia de la
comunidad bacteriana, el perfil cambia, siendo nuevamente las toxinas más
abundantes las N-sulfocarbamoil (54.17 %pmol), seguidas por las decarbamoil y
carbamoil (45.69 y 0.13 %pmol respectivamente), dichos cambios fueron
significativos (X2 0.05,3) = 45.3892; p<0.05) (Fig. 40).
Figura 40. Perfil de toxinas de la cepa 62L en presencia/ausencia de la comunidad bacteriana. Rojo: toxinas N-sulfocarbamoil (C1/2). Amarillo: toxinas decarbamoil (dcNEO, dcSTX, dcGTX2/3). Negro: toxinas carbamoil (GTX2/3).

67
En la cepa G7 se observa que la comunidad bacteriana aumenta
significativamente (X2 0.05,3) = 0.9650; p<0.05) la proporción de toxinas carbamoil (0.53
a 0.65 %pmol) y N-sulfocarbamoil (70.76 a 87.72 %pmol) y disminuye la producción
de toxinas decarbamoil (Fig. 41).
Figura 41. Perfil de toxinas de la cepa G7 en presencia/ausencia de la comunidad bacteriana. Rojo: toxinas N-sulfocarbamoil (C1/2). Amarillo: toxinas decarbamoil (dcNEO, dcSTX, dcGTX2/3). Negro: toxinas carbamoil (GTX2/3).
7.15. Toxicidad
La toxicidad total (definida como equivalentes de STX/cél) presentó menos
variaciones en función de la cepa y en presencia/ausencia de la comunidad
bacteriana. 62L fue la cepa de mayor toxicidad en ausencia de la comunidad
bacteriana (126 ± 41 ng STX eq/cél) mientras que GCCV-7 fue la cepa de mayor
toxicidad en presencia de la comunidad bacteriana (103 ± 43 ng STX eq/cél). Esta
diferencia se vincula directamente con que en presencia de la comunidad bacteriana,
la cepa 62L disminuyó la producción de toxinas decarbamoil y N-sulfocarbamoil,
mientras que la cepa GCCV-7 incrementó la producción de las toxinas
carbamatadas, decarbamatadas y N-sulfocarbatadas (Fig. 42 B y C).

68
En cuanto a la cepa GCMV-7, no presentó cambios en su toxicidad total pero
si se observó que en presencia de la comunidad bacteriana disminuye la producción
de toxinas decarbamoil y N-sulfocarbamoil (Fig. 42 A).
La cepa G7 fue la que presentó la menor toxicidad tanto en el control como en
el tratamiento y es que en presencia de la comunidad bacteriana disminuye la
producción de todos los grupos de toxinas (carbamoil, decarbamoil y N-
sulfocarbamoil)(Fig.42D).

69
Figura 42. Toxicidad (ngSTXeq/cél) de las cepas de G. catenatum en presencia/ausencia de la comunidad bacteriana. Líneas verticales: desviación estándar.
0
5
10
15
20
25
30
Carbamatadas Decarbamoil Sulfocarbamoil
ng
ST
Xeq
/cél
Ausencia CB Presencia CB
0
10
20
30
40
50
60
70
80
90
100
Carbamatadas Decarbamoil Sulfocarbamoil
ng
ST
Xe
q/c
él
Ausencia CB Presencia CB
0
10
20
30
40
50
60
70
80
90
100
Carbamatadas Decarbamoil Sulfocarbamoil
ng
ST
Xeq/c
él
Ausencia CB
Presencia CB
0
0.5
1
1.5
2
2.5
Carbamatadas Decarbamoil Sulfocarbamoil
ng
ST
Xe
q/c
él
Ausencia CB
Presencia CB
A
B
C
D

70
VIII. DISCUSIÓN
8.1. Aislamiento de la cianobacteria
Para el crecimiento de la cianobacteria aislada de la comunidad bacteriana
asociada a la cepa G7 de G. catenatum, se seleccionó al medio de cultivo GSe
porque en este se mostró un mejor crecimiento y por cumplir con los requerimientos
necesarios en la calidad del agua (esterilizada por filtración), sales minerales (macro
y micronutrientes esterilizados por calor húmedo) y factores de crecimiento
(vitaminas esterilizadas por filtración), además de la adición de extracto de
lombricompostaje que otorga un plus de nutrientes (N:P) y proporciona una fuente de
nitrógeno, ya sea en su forma amoniacal o de nitrito necesarias para un crecimiento
óptimo (González-González, 2010; Mendoza-Guerrero, 2011; Ortíz-Moreno et al.
2012).
Si bien, no se encontró referencia del cultivo de cianobacterias unicelulares
marinas en medio GSe, existen estudios donde utilizan medios de cultivo más
selectivos con la adición de extracto de suelo, obteniendo con ello un mejor
crecimiento. Tal es el caso del medio mineral estándar o SOT (Ogawa y Terui, 1970)
utilizado en el mantenimiento de Spirulina sp., para cianobacterias marinas, son
característicos el medio MN (Fuenmayor et al. 2009) y ES (Provasoli, 1957) por
utilizar agua de mar natural y poseer un menor aporte de sales minerales, y, el medio
ASN-III (Rippka, 1988) para el aislamiento de cianobacterias fijadoras de nitrógeno.
Otra ventaja, es que los macro y micronutrientes no varían tanto entre un medio y
otro, por lo que el aporte nutricional proporcionado es el necesario para el desarrollo
de estas microalgas (Torres-Ariño, 2001; 2004; 2011; Monroy-Sampieri, 2014).
Existen muchas técnicas de aislamiento de cianobacterias, sin embargo, las
más utilizadas son las manuales, entre las que destaca el fototactismo acompañado
de la migración vertical debido a que estratifica la presencia de los diversos
microorganismos, eliminando la mayor cantidad de bacterias, especialmente los
bacilos Gram (-) (Tovar-Ramírez, 1999). Por otra parte, el estriado en placa con agar

71
únicamente nos permitió la purificación del cultivo, pues el crecimiento de la
cianobacteria en dichas condiciones no se favoreció, lo cual podría atribuirse al grado
de sequedad de la superficie del agar, pues a concentraciones mayores de 1.5 % de
agar, éste actúa como inhibidor del crecimiento (Allen, 1968; Olivas, 2004). En el
presente estudio se trabajó con una concentración de 1 % de agar y a pesar de estar
dentro del límite de tolerancia, no se obtuvo un buen crecimiento de la cianobacteria.
8.2. Curvas de crecimiento de la comunidad bacteriana
En los últimos 25 años se han desarrollado una gran variedad de métodos que
evalúan la abundancia y actividad del femto, pico y microplancton, siendo la
microscopía de epifluorescencia una de las técnicas más utilizadas para el conteo
directo de bacterias, demostrando que constituyen una de las comunidades más
abundantes de los océanos (Soto-Castor et al. 2002). Sin embargo, existen otras
técnicas que permiten la visualización y observación puntual de estos
microorganismos, teniendo ventajas y limitaciones dependiendo el enfoque de cada
estudio (Legendre et al. 2001; Bouvier et al. 2007)
En el presente estudio y tal como lo describe Vázquez y colaboradores (2010)
el uso de la microscopía óptica no nos permitió la correcta distinción entre la
cianobacteria y los bacilos. En cuanto al conteo celular con la cámara de Neubauer,
particularmente se tuvieron dos limitaciones: 1) las estimaciones celulares en cada
triplicado eran extremadamente variables conforme el tiempo de cultivo avanzaba y
2) el tiempo de conteo por muestra era demasiado (30-45 minutos), convirtiéndose
en una técnica tediosa, contrario al citometro, que analizó 30 muestras en una hora,
3 minutos por muestra, su precisión difirió en un 2 % entre muestra y muestra y nos
permitió evaluar características intrínsecas de las células, tales como su tamaño y
complejidad (La Ferla et al. 2003; Calvo-Díaz y Morán, 2006; Bouvier et al. 2007).
Sin embargo, a pesar de tantas ventajas que presenta la citometría de flujo
(identificación y aislamiento rápido de organismos, disminución en los tiempos de
procesamiento, análisis y toma de decisiones, análisis de las características

72
intrínsecas de las células) en el análisis celular, en nuestro estudio únicamente nos
permitió separar a los organismos autótrofos de los heterótrofos y tener una
aproximación del tamaño celular de los mismos, esto debido a una mala calibración
del equipo, el exceso de ruido electrónico que arrojaba el citometro, el uso de
microesferas que triplicaban en tamaño a los micoroganismos y la falta de
fluorocromos que facilitarían la identificación de subgrupos específicos dentro de
poblaciones o comunidades celulares (Castillo-Navarrete, 2005; Forget et al. 2010).
En teoría, es posible usar un citometro de flujo para medir los parámetros
intrínsecos tanto en bacterias como en células eucariontes, pero en la práctica, en
una bacteria la señal de estos parámetros es mil veces menor (Castillo-Navarrete,
2005), por lo que su medición es difícil cuando no se tiene experiencia además que
estos organismos presentan variaciones fotoeléctricas que hacen “indetectables”
algunos componentes bacterianos (Shapiro, 2003)
Además, se debe considerar que las microalgas al tener la capacidad de ajustar
su concentración de clorofila en función de las condiciones de iluminación (foto-
aclimatación) el citometro las cuenta como células de mayor tamaño, complejidad o
con mayor fluorescencia (Forget et al. 2010) es por esto que existió una enorme
variabilidad en los conteos entre esta técnica y la microscopía óptica y
epifluorescente.
En cuento al tamaño correcto de las microesferas, Gasol y Del Giorgio (2000) y
Campbell (2001) mencionan que para el estudio del picoplancton es necesario el uso
de microesferas de un tamaño no mayor a 2 µm de diámetro las cuales servirán
como referencia intermedia del tamaño celular, ubicando a los organismos por
debajo y sobre las mismas al momento de graficar el tamaño celular (FSC) y
complejidad (SCC) versus la fluorescencia roja de la ficoeritrina, de lo contrario, la
posible referencia que te proporcionen sub o sobreestimará el tamaño celular, como
en nuestro caso.

73
A pesar del uso de la microscopía óptica y de la citometría de flujo, la tinción
fluorescente y el conteo directo por microscopia de epifluorescencia permanecerá
como el método estándar de enumeración de bacterias en ambientes acuáticos,
siendo el método más eficaz y tomado como base para la interpretación de los datos
ya que se trata de una técnica donde las observaciones son directas, puntuales y
personales, dando realce a las características particulares que necesite cada
investigador (Betancourt et al. 1997; La Ferla et al. 2003; Mecalco-Hernádez, 2010),
por lo que las dos técnicas antes descritas fueron métodos complementarios a la
microscopía de epifluorescencia, aportando características diferentes como la
identificación y cuantificación de las tres fracciones de la comunidad bacteriana.
Debido a que las cianobacterias presentan elevada plasticidad en formas y
tamaños, aquellas que presenten un diámetro de 0.7 a 3 µm y tengan forma esférica
entrarán dentro del grupo de las cianobacterias unicelulares cocoides, por lo que la
presente en la comunidad bacteriana asociada a G. catenatum entra dentro de este
grupo y teóricamente se caracterizaría por 1) preferir irradiancias bajas e
intermedias, comprendidas dentro del intervalo de los 58.8 a 156 µmolm-2s-1; 2)
presentar un crecimiento óptimo con luz blanca; 3) tener un nivel de tolerancia de 0
a 100 ups, y un crecimiento óptimo a 35 ups y 4) una curva de crecimiento con fases
estacionarias cortas de entre seis y nueve días, posterior a las cuales se obtiene la
máxima densidad celular y después el crecimiento se ve inhibido (Díaz-Hernández,
1997; Morales et al. 2002; Rosales-Loaiza et al. 2008; Poot-Delgado et al. 2009).
Sin embargo, a pesar de que las condiciones de cultivo a las que se mantuvo la
cianobacteria de este trabajo están dentro del intervalo de crecimiento descrito en la
bibliografía para cianobacterias marinas, unicelulares y cocoides, no obtuvimos un
buen crecimiento, reflejado en densidades celulares bajas. Así mismo, la tasa de
crecimiento (0.038 - 0.1 div/día) también fue más baja (0.14 - 0.37 div/día) mientras
que el tiempo de generación obtenido (2.76 – 7.72 días) fue mayor al reportado por
Poot-Delgado y colaboradores (2009) el cual fue de 4.85 – 5.89 días.

74
8.3. Curvas de crecimiento de G. catenatum
La curva de crecimiento fue la típica para la especie, caracterizada por una fase
de aclimatación corta, iniciando al segundo día con la fase exponencial. La fase
estacionaria no se presentó ya que rápidamente después de alcanzar la máxima
densidad celular, se observó la fase de declive (Martínez-Tecuapacho, 2009;
Bustillos-Guzmán et al. 2012; Costa et al. 2015).
En 2010, Oh y colaboradores mencionan que las tasas de crecimiento son
características de cada cepa, pues al comparar 83 cepas de G. catenatum de
florecimientos distintos en la Bahía de Inokushi, Japón, obtuvieron que incluso cepas
aisladas el mismo día y bajo las mismas condiciones mostraron diferencias
significativas, concluyendo que la especie presenta respuestas cepa/específica, es
decir, que cada una tendrá un comportamiento diferente a pesar de estar expuestas
a las mismas condiciones de aislamiento, cultivo y crecimiento, lo cual se pudo
confirmar en el presente estudio, al obtener dos cepas favorecidas por la comunidad
bacteriana y dos inhibidas, considerándose la presencia/ausencia de dicha
comunidad como única variable.
En general, se obtuvo una tasa de crecimiento entre 0.04 (± 0.002) y 0.22 (±
0.02) div/día, siendo únicamente GCCV-7 y 62L quienes superaron las 0.1 div/día
con 0.22 (± 0.02) y 0.14 (± 0.02) div/día, respectivamente, teniendo con ello el menor
tiempo de generación (1.4 ± 0.012 y 2.1 ± 0.01 días). Específicamente, para G.
catenatum se ha reportado una tasa de crecimiento mínima de 0.07 div/día (Durán-
Riveroll, 2014) y máxima de 0.34 div/día (Hallegraeff et al. 2011) bajo las mismas
condiciones de cultivo del presente estudio. También existen registros de tasas de
crecimiento intermedias (0.14 div/día) pero en medio f/2, con un intervalo de
temperatura de 15 – 29 °C, 30 ups y 230 µmolm-2s-1 (Band-Schmidt et al. 2004).
Debe reconocerse que las cepas mexicanas han presentado tasas máximas
de crecimiento menores comparadas con aquellas aisladas de otros países, con la
excepción de las cepas de Bahía Concepción, Bahía de La Paz y Mazatlán (Band-
Schmidt et al. 2006) obteniendo tasas de crecimiento de 0.70 a 0.82 div/día. En

75
nuestros experimentos, únicamente la cepa G7 (0.04 ± 0.002 dív/día) obtuvo una
tasa de crecimiento más baja de lo reportado en literatura, el resto quedó dentro del
intervalo establecido para cepas de G. catenatum antes mencionado, aunque con
valores bajos.
Existen numerosos trabajos que evalúan el crecimiento de G. catenatum ante la
variación de diversos factores abióticos. Específicamente y evaluando la
temperatura, salinidad y medio de cultivo no se encontraron diferencias
estadísticamente significativas (Band-Schmidt et al. 2004; Band-Schmidt et al. 2006;
Bustillos-Guzmán et al. 2012; Durán-Riveroll, 2014). Sin embargo, al analizarse
diferentes proporciones de N:P (4:1, 16:1, 32:1 y 64:1) en cultivos semi-continuos
(Hernández-Sandoval, 2010) si se halló una diferencia significativa , reafirmándose la
respuesta cepa/específica antes mencionada ya que nosotros encontramos
diferencias en tres de las cinco cepas (GCCV-7, G7 y 62L), constituyendo el segundo
reporte en donde el crecimiento del dinoflagelado se ve afectado por un factor biótico
(Fernández-Herrera, 2013).
El ecotipo mexicano de G. catenatum se caracteriza por tolerar un amplio
rango de temperatura (11.5 – 30 °C), con un crecimiento entre los 21 y 29 °C, y
dentro de un rango de salinidad entre 28 y 38 ups (Band-Schmidt et al. 2004). Las
condiciones de cultivo de este estudio fueron a 24 °C y a una salinidad de 34 ups,
entrando dentro del rango de crecimiento. No obstante, Blackburn y colaboradores
(1989) establecieron que un buen crecimiento en G. catenatum se verificaba en las
divisiones celulares, siendo las óptimas cada 3 a 4 días, por lo que únicamente las
cepas GCMV-7 y G7 New (3 ± 0.13; 3.2 ± 0.63 días, respectivamente) presentaron
un buen crecimiento, debido a que el resto tuvo un tiempo de generación menor al
descrito.
8.4. Formación de cadenas celulares
Cada cepa mostró un comportamiento distinto, pero en general se observó
una disminución de células individuales al comenzar la fase de crecimiento

76
exponencial, formándose cadenas celulares más largas conforme la edad del cultivo
avanzaba.
Blackburn y colaboradores (1989) señalan que una mayor longitud de cadenas
en el cultivo indica que están en mejores condiciones fisiológicas pues observaron
que durante la fase de crecimiento exponencial, la mayoría de las células formaban
cadenas de 4-16 células y al ir avanzando en el tiempo y bajo condiciones limitantes,
estas cadenas se rompen en células individuales.
Dichas observaciones coinciden con nuestros resultados, ya que a pesar de
que G7 fue la cepa que presentó la menor abundancia celular y tasa de crecimiento
no fue la que tuvo una mayor proporción de células individuales a lo largo de todo el
cultivo (1146 ± 4 cél/mL; 0.04 ± 0.002 div/día; 38 ± 15 %) sino 62L y GCCV-7,
quienes obtuvieron las mayores densidades celulares y tasas de crecimiento (6444 ±
4 cél/mL; 0.14 div/día; 36 ± 11 % y 6463 cél/mL; 0.22 div/día; 41 ± 18 %,
respectivamente).
Aunado a lo anterior, estudios previos han demostrado que el largo de las
cadenas es influenciado también por las condiciones y composición del medio de
cultivo (Blackburn et al. 1989) encontrando que en medio GSe se obtiene una mayor
producción de cadenas celulares largas (Band-Schmidt et al. 2004; Durán-Riveroll,
2014) debido a que este medio tiene extracto de suelo que estimula el crecimiento,
agregando un plus de micronutrientes (González-González, 2010; Ortíz-Moreno et al.
2012). A pesar de que no se evaluó este factor en nuestro estudio, los resultados nos
dan la pauta a decir que no contibuye a la formación de cadenas celulares largas,
debido a que solo en una cepa predominaron las cadenas celulares de siete células y
mayores a lo largo del cultivo (G7 New con 67 %).

77
8.5. Asociación G. catenatum-comunidad bacteriana
8.5.1. Exposición a filtrados celulares
En trabajos previos, donde se han expuesto a otros microoganismos
(diatomeas, copépodos, crisofitas) a filtrados celulares de algún dinoflagelado, se ha
obtenido una total inhibición en el crecimiento de estos organismos y dependiendo el
tiempo y volumen de exposición ha podido recuperarse o no el cultivo (Arzul et al.
1999; Fuentes-Pascacio, 2010; Kim y Park, 2010).
La comunidad bacteriana de nuestro estudio al estar expuesta a los
metabolitos secundarios de G. catenatum disminuyó su densidad celular máxima y
tasa de crecimiento durante todo el cultivo, por lo que a pesar de la posible
disponibilidad de nitrógeno en forma de aminoácidos libres en el medio, no se
favoreció su crecimiento.
La exposición a los filtrados celulares se llevó a cabo porque a través de
microscopía óptica (Fig. 5) se observaba que tanto los bacilos como la cianobacteria
se agrupaban alrededor de las células del dinoflagelado, por lo que se pensó que la
interacción entre las especies se desarrollaba a una escala espacio temporal corta
donde el efecto de los metabolitos secundarios se haría evidente al utilizar los
filtrados de los exudados liberados al medio, detectando cambios en el crecimiento
tal como lo describe Arzul y colaboradores (1999).
Sin embargo, la microscopia de epifluorescencia dilucidó parcialmente por qué
no hubo un incremento en la densidad celular de la comunidad bacteriana, al
evidenciar que los bacilos constituyen bacterias intracelulares (Fig. 11) mientras que
la cianobacteria se encuentra inmersa en el mucus de carbohidratos que produce el
dinoflagelado (Band-Schmidt, com. pers.), creándose un micro hábitat con las
condiciones necesarias para su crecimiento.

78
8.5.2. Cultivos con contacto celular
Curvas, tasas de crecimiento y formación de cadenas celulares
El estudio de la interacción bacteria-microalga ha tomado relevancia de la
última década al presente, siendo descritos varios aspectos como: la producción
autónoma de toxinas por parte de las bacterias, la intervención que tienen en 1) la
síntesis de toxinas 2) en el apareamiento y 3) la influencia directa o indirecta en el
desarrollo de ciertos estadios del ciclo de vida de los dinoflagelados (Albinsson,
2011; Bolch et al. 2011; Albinsson et al. 2014). Sin embargo, estos estudios no
proporcionan información acerca del comportamiento e influencia de la comunidad
bacteriana en el crecimiento, específicamente, en los cambios de densidad celular,
tasas de crecimiento y efectos en el perfil de pigmentos en células vegetativas, por lo
que el presente estudio es el primero para la especie que aborda esos puntos,
además de evaluar los cambios en el perfil de toxinas paralizantes.
En estudios previos se ha concluido que la interacción bacteriana que
presenta G. catenatum es específica y la estimulación o inhibición en su crecimiento
dependerá del tipo de bacteria asociada a sus cepas (Hold et al. 2001; Uribe y
Espejo, 2003; Croft et al. 2005).
Además al cultivar al dinoflagelado con más de una bacteria, su curva de
crecimiento se ve modificada al interrumpirse abruptamente la fase de crecimiento
exponencial por un declive pronunciado, producto de un agotamiento de nutrientes,
principalmente nitratos y fosfatos, siendo estos últimos, utilizados por las bacterias
como agentes estimulantes en su crecimiento (Córdova et al. 2003; Bolch et al.
2004). Dichas características fueron observables en nuestros experimentos (Fig. 22),
siendo la fase de declive repentina y precedida por la obtención de la mayor
densidad celular, un comportamiento constante para todas las cepas. En cuanto al
agotamiento de nutrientes, no se puede concluir al respecto debido a que estos no
fueron cuantificados.

79
Sin embargo, al comparar las curvas de crecimiento de G. catenatum en
presencia/ausencia de la comunidad bacteriana (Fig. 23), nos percatamos que en
presencia de la misma, se obtienen dos patrones de comportamiento: el primero, es
un factor estimulante en el crecimiento de la microalga, traducido en la obtención de
densidades celulares mayores (Fig. 23 B) y en una prolongación en el tiempo del
cultivo (Fig. 23 A) lo que coincide parcialmente con lo reportado para cepas de
Alexandrium spp. donde la presencia de bacterias prolonga la fase de crecimiento
exponencial, modulando la disponibilidad de nutrientes pero no incrementa de
manera significativa las tasas de crecimiento, por el contrario, se registran tasas de
crecimiento bajas producto de un letargo en el crecimiento (Hold et al. 2001; Uribe y
Espejo, 2003; Maas et al. 2007). Esto último solo se observó en la cepa GCCV-7 ya
que GCM-V-7 no solo vio incrementada su densidad celular sino también su tasa de
crecimiento (Fig. 24 y 25).
El segundo patrón de comportamiento fue la inhibición en el crecimiento
de G. catenatum (Fig. 23 C y D) en presencia de la comunidad bacteriana, lo que
podría ser explicado en base a lo descrito por Albinsson (2011) para co-cultivos de
G. catenatum en presencia de la bacteria marina Marinobacter sp. donde observa
que la bacteria presenta cambios en la expresión génica (supresión de genes
reguladores de la producción de toxinas PSP) causando efectos estimulantes en una
cepa e inhibitorios en tres cepas.
Sin embargo, se debe considerar que a pesar de haberse demostrado que las
bacterias presentes en una cepa específica de dinoflagelado son capaces de
asociarse a otra cepa de la misma o de diferente especie, no tienen el mismo efecto
en todas las cepas, incrementando el crecimiento en algunas e inhibiéndolo en otras
como es el caso de los trabajos de Bolch y colaboradores (2011) quienes trabajaron
con la actinobacteria Alcanivorax sp. y Albinsson y colaboradores (2014) quienes
estudiaron el efecto de Marinobacter sp. y Alcanivorax sp. sobre G. catenatum
Dentro de las cepas que vieron interrumpido su crecimiento en presencia de la
comunidad bacteriana, está la cepa michoacana 62L, que mostró un descenso

80
significativo en su densidad celular de más del 90 %. Esto coincide con lo reportado
por Alavi y colaboradores (2001) en cepas del dinoflagelado Pfiesteria que tras
inocular la comunidad bacteriana a cepas específicas de la microalga, esta presentó
descensos significativos en su densidad celular, la cual logró recuperarse al re-
aislarse de la comunidad bacteriana y concluyendo que la inhibición en el crecimiento
por parte de las bacterias se debe a la liberación de metabolitos secundarios al
medio.
Matsumoto (2011) mantuvo a G. catenatum en dos tipos de cultivos: con
filtrados celulares y contacto directo con la bacteria Alcanivorax obteniendo una
inhibición en los primeros y una estimulación significativa en el crecimiento de los
segundos, infiriendo que la sustancia promotora del crecimiento no es una sustancia
intracelular o que la concentración no es lo suficientemente alta para estimular
completamente el crecimiento del dinoflagelado. Aunado a esto, se ha demostrado
que algunas especies de dinoflagelados son capaces de ingerir cianobacterias
unicelulares o algunas bacterias pequeñas (< 2 µm) debido a que estas les
proporcionan factores orgánicos de crecimiento (principalmente aminoácidos)
(Skovgaard, 2000; Jeong et al. 2005a; Skelton et al. 2009).
Específicamente, G. catenatum ingiere células de la cianobacteria
Synochococcus spp. (Jeong et al. 2005b) demostrando que la especie presenta una
alta selectividad por este organismo debido a la calidad nutricional que le provee, sin
embargo, también se menciona que la microalga proporciona formas asimilables de
carbono al producir ácidos grasos, esteroles, lípidos y aceites en abundancia por lo
que la presencia de la cianobacteria de nuestro estudio dentro del mucus producido
por G. catenatum podría establecerse como una relación de mutualismo debido a
que ambos proveen de factores estimulantes en el crecimiento mientras que la
presencia de bacilos intracelulares confirmaría lo descrito por Töbe (2003) al
localizarse en compartimientos ricos en metabolitos, como lo es el citoplasma (Fig.
14).

81
Dado que los trabajos realizados para la especie de interés no han trabajado
con células vegetativas sino con quistes de resistencia, su germinación y
supervivencia, no existen datos comparables que nos eluciden si los resultados
obtenidos son efectos positivos de la comunidad bacteriana asociada pero Kim y
Park (2010) al exponer al dinoflagelado Fragilidium duplocampanaeforme a varias
especies fitoplanctónicas, observó una disminución en la longitud de las cadenas
celulares, muerte celular y la formación de quistes temporales, demostrando que el
contacto directo entre especies diferentes puede modificar completamente la
estructura de la comunidad fitoplanctónica.
Haciendo realce a la formación de cadenas celulares, Fernández-Herrera
(2013) en sus experimentos de alelopatía de Chattonella marina var. marina sobre G.
catenatum menciona que las primeras células en morir del dinoflagelado fueron las
individuales, pero sugiere que las cadenas celulares al tener una mayor superficie de
contacto, serán más susceptibles al efecto por contacto directo, lo que explicaría
porque las cepas 62L y G7 no presentaron un incremento en su crecimiento, al ser
las cepas con mayor proporción de cadenas celulares largas, contrario a GCMV-7 y
GCCV-7 que si incrementaron su crecimiento en presencia de la comunidad
bacteriana pero también presentaron una mayor producción de células individuales.
8.6. Curvas de crecimiento de la comunidad bacteriana en los cultivos con
contacto celular
La comunidad bacteriana asociada a cepas de G. catenatum ha sido
caracterizada filogenética y funcionalmente, determinándose que se integra por 15 a
24 genotipos bacterianos distintos (Green et al. 2004). Sin embargo, a pesar de ser
tan variable, estudios previos han concluido que el efecto de la comunidad bacteriana
en el dinoflagelado se debe a la dominancia de una, máximo dos especies, más no a
la coexistencia de las mismas ya que en los cultivos mixtos, la bacteria dominante
tiene efectos en el crecimiento de la microalga e inhibe el crecimiento del resto de las
bacterias asociadas (Albinsson, 2011; Albinsson et al. 2014). Estas observaciones
podrían explicarnos el por qué la cianobacteria, a diferencia de los bacilos y

82
filamentos, logra un mejor crecimiento en las distintas cepas de G. catenatum aquí
evaluadas (Fig. 29), mostrando un incremento gradual y constante en su densidad
celular, contario a los otros dos microorganismos que presentan amplias oscilaciones
en su crecimiento.
Por otra parte, la dinámica de crecimiento de las distintas bacterias cultivadas
con células de G. catenatum sugiere que el dinoflagelado tiene efectos de interacción
diferentes para cada bacteria, lográndose el establecimiento de diferentes
asociaciones ecológicas dependiendo el impacto de la interacción entre los dos
microorganismos.
Se han reconocido para esta especie, una asociación mutualista con la
bacteria Marinobacter sp. (Bolch et al. 2004; Green et al. 2004; Albinsson, 2011;
Albinsson et al. 2014) y un amensalismo con Alcanivorax borkumensis debido a que
G. catenatum presentó una mortalidad alta (89 %) en presencia de la bacteria,
mientras que esta última duplicó su población en las primeras dos horas de iniciado
el experimento (Green et al. 2004).
Nuestro estudio no identificó a las bacterias asociadas a la cepa G7 de G.
catenatum, sin embargo, existe la posibilidad de que los bacilos pudiesen ser una
especie del género Alcanivorax ya que morfológicamente son similares y se ha
reportado la presencia de esta bacteria en cepas aisladas de diversos ecotipos
(Bolch et al. 2011).
En cuanto a las tasas de crecimiento y densidades celulares máximas
obtenidas por la comunidad bacteriana y aquellas previamente descritas, las
diferencias se atribuyen a tres factores: 1) el método de cuantificación de la
comunidad, ya que en estudios anteriores se ha utilizado el PCR en tiempo real
(Albinsson, 2011) y el conteo por diluciones seriadas (Subramanian, 2008) versus la
microscopía de epifluorescencia del presente. 2) el estadio celular de G. catenatum
ya que previamente se evalúa el porcentaje de eclosión de los quistes de resistencia
y la posterior supervivencia de las células expuestas a la comunidad bacteriana

83
mientras que nosotros trabajamos con células vegetativas que a pesar de no haber
sido aclimatadas, los experimentos fueron viables y 3) la densidad inicial de
células/quistes del dinoflagelado, teniendo en nuestro estudio (500 cél/mL), una
densidad más baja a la reportada (>1000 cél/mL) (Albinsson et al. 2014).
8.7. Cambios morfológicos de Gymnodinium catenatum en presencia de la
comunidad bacteriana
Gymnodinium catenatum es una especie que se caracteriza por la formación de
dos tipos de quistes, por el proceso sexual se obtienen los quistes de resistencia
mientras que en células vegetativas y móviles se llega a desarrollar una membrana
hialina que rodea a la célula, formándose un quiste temporal o pelicle (Figueroa,
2005). Ambos procesos de enquistamiento responden ante condiciones
desfavorables y son parte fundamental del ciclo de vida de la especie, al favorecer
sus eventos de proliferación (Figueroa et al. 2006; 2008).
La interacción bacteria-microalga se ha basado en la influencia que la
comunidad bacteriana tiene en la germinación y supervivencia de quistes de
resistencia en G. catenatum, más no en la formación de estos a partir del contacto
con las bacterias, ya que una de las principales características de estos quistes es la
ausencia de bacterias intracelulares producto de la doble capa que los protege
(Töbe, 2003). Sin embargo, la capacidad de producir quistes como respuesta a la
presencia de otras especies si ha sido reportada (Uchida et al. 1999),
específicamente, la formación de quistes temporales por efectos alelopáticos fue
descrita por primera vez en 2004 por Fistarol y colaboradores quienes expusieron
células del dinoflagelado Scrippsiella trochoidea a filtrados celulares de Alexandrium
tamarense.
En este estudio, no se encontraron quistes de resistencia pero si quistes
temporales, producto del contacto directo con la comunidad bacteriana,
presentándose en altos porcentajes durante la fase de siembra, aclimatación e inicio
del crecimiento exponencial, además de observarse distintos desarreglos

84
morfológicos en las células de G. catenatum modificando con ello la dinámica
poblacional de la especie (Albinsson, 2011).
Son varios los trabajos que han inducido la formación de quistes combinando
cultivos monoalgales (Blackburn, 1989; Green et al. 2004, 2010; Figueroa, 2005;
Figueroa et al. 2006; 2008; Albinsson, 2011; Bolch et al. 2011; Matsumoto, 2011;
Albinsson et al. 2014) y en todos se ha demostrado que G. catenatum presenta
distintas rutas de enquistamiento durante su ciclo de vida pero deben de existir
condiciones limitantes para su crecimiento como la deficiencia en los nutrientes
(N:P), estrés ambiental, cambios en su fotoperiodo, disminución de la temperatura,
entre otros, pero no se ha reportado la producción de quistes temporales por la
interacción con comunidades bacterianas, pero sí por contacto directo con una
rafidoficea (Fernández-Herrera, 2013). Por lo tanto, este es el primer reporte sobre la
inducción de quistes temporales en G. catenatum en presencia de una comunidad
bacteriana.
Los distintos desarreglos que se observaron (Fig. 33) ya habían sido descritos
por Figueroa y colaboradores (2006; 2008), no obstante, nosotros observamos
patrones de cambio característicos de la división sexual del planocigoto: pérdida de
flagelos, desarreglo en el citoplasma, en el cíngulo y sulcus, además de un núcleo
evidente con cromosomas condensados, característico de la fusión de gametos.
Dichas observaciones sustentan una de las hipótesis establecidas por los mismos
autores, que indica que los procesos sexuales se desencadenan como resultado de
una adición de nutrientes, provenientes quizá de los exudados bacterianos.
Otro evento observado fue la presencia de quistes en cadenas celulares, dando
la pauta a pensar que el proceso sexual es una ruta flexible y no unidireccional como
se pensaba ya que el enquistamiento de las cadenas celulares podría deberse a la
fusión de gametos con células vegetativas, a la fusión de dos cadenas o a la fusión
de células pertenecientes a la misma cadena (Figueroa et al. 2008).

85
Finalmente, el tiempo en el que se observaron los cambios estructurales en las
células de G. catenatum no corresponden con los reportados en trabajos previos (72
hrs para la obtención de un quiste del planocigoto) ya que nosotros los observamos 3
hrs después de la inoculación de la comunidad bacteriana.
8.8. Perfil de pigmentos
8.8.1. Cianobacteria
En general, los pigmentos fotosintéticos tienen una distribución específica y
restringida a una clase de microalgas, así como la peridinina es el pigmento
característico de los dinoflagelados, la clorofila a y sus pigmentos accesorios,
comúnmente denominados ficobilisomas, son los pigmentos principales de las
cianobacterias (Ludeña-Hinojosa, 2007).
No obstante, la clorofila a no es el único pigmento encontrado en las
cianobacterias ya que estas necesitan aumentar el rango de absorción de luz
utilizado para la fotosíntesis, en especial, a longitudes de onda donde la clorofila
tiene una absorción baja (Rosales-Loaiza et al. 2008). Es por esta misma razón que
el perfil de pigmentos característico de este grupo de organismos incluye a los β-
carotenos y a la zeaxantina, quienes actúan en el fotosistema I y II, respectivamente,
haciendo más eficiente la captura de la luz.
Como se ha venido mencionando, la comunidad bacteriana presente en cepas
específicas de G. catenatum está formada por bacilos, bacterias heterótrofas
filamentosas y una cianobacteria, evidenciando su presencia con el perfil de
pigmentos descrito en la figura 32, donde se observa la presencia, en un alto
porcentaje, de clorofila a, zeaxantina y β-caroteno. No obstante, también
presenciamos otro pigmento accesorio, la diadinoxantina, que es una xantofila que
está reemplazando a las ficobilinas y su presencia al igual que la de la zeaxantina se

86
ha observado en condiciones de alta intensidad de luz (>150 µmolm-2s-1) (Pérez-
Rontomé, 2000).
En estudios previos se han encontrado estos mismos pigmentos, reportándose
como característicos de la picocianobacteria Synechococcus-DC2, observándose
características de cultivo similares como el fotoperiodo de 12:12 hrs luz:ocuridad,
agua de mar filtrada y estéril adicionada con vitaminas, una temperatura de 23 ± 1 °C
y a 155 µmolm-2s-1 de irradiancia, por lo que Díaz-Hérnández (2010) menciona que
los posibles cambios en el perfil de pigmentos pueden deberse a altas irradiancias,
concentraciones elevadas de N2 y al incremento en nutrientes.
8.8.2. Gymnodinium catenatum en presencia/ausencia de la comunidad
bacteriana
El perfil de pigmentos característico de G. catenatum está conformado por 10
pigmentos: carotenoides polares, clorofila c, peridinina, cis-peridinina, dinoxantina,
diadinoxantina, diatoxantina, carotenoides, clorofila a y β-carotenos, coincidiendo con
el descrito para otros dinoflagelados como Gyrodinium resplendens y Gymnodinium
sanguineum (Hallegraeff et al. 1991).
Estudios anteriores mencionan que el contenido de pigmentos fotosintéticos
puede cambiar como respuesta a factores causantes de estrés, a la capacidad
fotosintética o al estado de desarrollo en el que se encuentre la microalga, no
obstante, dado que de los ocho pigmentos obtenidos en los perfiles (Figs. 36-39)
solamente uno es característico de la cianobacteria, las diferencias encontradas en la
producción de dinoxantina, diatoxantina y peridinina se puede atribuir a las
diferencias en la densidad celular de G. catenatum en presencia/ausencia de la
comunidad bacteriana (Fig. 24).

87
8.9. Perfil de toxinas paralizantes
La presencia de toxinas hidrofílicas, tanto en cepas mexicanas como en cepas
de otros países, ha sido un tema bastante controversial, discutido e investigado. En
el caso de las cepas mexicanas, Band-Schmidt y colaboradores (2010) hicieron una
revisión y concluyen que de manera característica, G. catenatum produce los
análogos STX, neoSTX, GTX2-3, dcGTX2-3, B1-2 y C1-2. Las toxinas decarbamoil
(dcGTX2-3) y N-sulfocarbamoil (C1-2), son generalmente los análogos con mayor
contribución molar, sin embargo, se ha determinado que la presencia, ausencia y
proporción de los análogos varía constantemente entre cepas y poblaciones, y a la
fecha no se conoce una cepa de G. catenatum que produzca todos los análogos,
aunque típicamente todas las cepas sintetizan más de uno (Cembella, 1998).
También se han reportado variaciones dentro de las mismas poblaciones y
diferencias en las proporciones de los análogos en función de factores físicos
(salinidad y temperatura) y nutricionales (medio de cultivo) (Camino-Ordás et al.
2004; Band-Schmidt et al. 2004, 2006, 2014; Gárate-Lizárraga et al. 2005; Oh et al.
2010; Bustillos-Guzmán et al. 2012, 2015; Albinsson et al. 2014). Dado que todos
estos factores fueron controlados en nuestros tratamientos y que la única variable fue
la presencia/ausencia de la comunidad bacteriana podemos atribuir los cambios en el
perfil de toxinas y en la toxicidad a la presencia de este factor biótico.
Son numerosos los trabajos donde se reportan los distintos análogos
producidos por G. catenatum y básicamente dependiendo el método para su
detección y análisis serán los análogos reportados. Específicamente, Negri y
colaboradores (2007) describen la producción de al menos 21 análogos de STX en
G. catenatum, sin embargo, para cepas mexicanas Bustillos-Guzmán y
colaboradores (2015) reportan 25 análogos: 10 hidrofílicos (toxinas carbamoil:
GTX2/3; decarbamoil: dcSTX, dcNEO y dcGTX2/3 y N-sulfocarbamoil: C1/2 y B1/2) y
15 tipo benzoil (GC1-6, GC1a-5a y GC1B, 2b, 4b y 5b). De los análogos descritos, no
fueron detectados los de tipo benzoil y las toxinas B1/2 en este trabajo y no hubo
diferencias en el número de análogos entre cepas, como lo reporta Band-Schmidt y

88
colaboradores (2006), quienes encontraron un mayor número de análogos en cepas
de Bahía Concepción en medio GSe.
En la mayoría de las cepas, con excepción de la 62L (Fig. 39), las toxinas
C1/2 (N-sulfocarbamoil) mostraron abundancias relativas constantemente superiores
a 70 % pmol, llegando a representar hasta el 95.79 % pmol en GCCV-7. La alta
proporción de toxinas C se considera una característica de G. catenatum, con pocas
excepciones (Band-Schmidt et al. 2010), lo que coincide con los resultados obtenidos
en este estudio. La única excepción fue la cepa 62L al mostrar una mayor proporción
de toxinas decarbamoil en ausencia de la comunidad bacteriana lo cual nos
explicaría la mayor toxicidad de esta cepa, ya que el factor de toxicidad relativa es
superior a la de las toxinas C1/2. Este patrón coincide con los de Gárate-Lizárraga y
colaboradores (2005) y Band-Schmidt y colaboradores (2006) quienes encontraron
una mayor proporción de dcNEO en cepas de Bahía de La Paz.
Se ha sugerido que la proporción de dcNEO está directamente relacionada
con el largo de las cadenas celulares (Band-Schmidt et al. 2006), lo cual coincidiría
con este estudio porque 62L fue una de las cepas que presentó mayor proporción de
esta toxina en ausencia de la comunidad bacteriana, pues en presencia de la misma,
la proporción de cadenas celulares largas se inhibió al igual que la producción de
este tipo de toxina.
El resto de las toxinas, pertenecientes al grupo de las carbamoil (GTX2/3) se
encontraron constantemente en muy bajas proporciones, no superando el 2 % del
total, lo cual también ha sido reportado como característico de la especie (Hallegraeff
et al. 2011).
Los perfiles de toxinas permanecieron relativamente constantes en todas las
cepas, algunas diferencias podrían deberse a: deficiencias nutricionales, a un
desacoplamiento en la síntesis de toxina durante la división celular, asincronía en la
división celular en los tratamientos, o bien, modificaciones en el ciclo de vida debido
a que los genes relacionados en la producción de toxinas se expresan en diferentes

89
niveles del mismo, por lo que si este sufre alguna alteración, la producción de toxinas
se afectará (Anderson et al. 1990; Taroncher-Oldenburg et al. 1997, 2000; Zhuang et
al. 2012).
8.10. Toxicidad
Los resultados en este trabajo mostraron un patrón de cambio en la toxicidad
celular de G. catenatum, ya que en presencia de la comunidad bacteriana, disminuye
la producción de toxinas decarbamoil y N-sulfocarbamoil en la mayoría de las cepas,
excepto en la cepa GCCV-7 que fue la más tóxica y a su vez la que presentó una
mayor contribución de toxinas carbamatadas y decarbamoil.
Para cepas de G. catenatum se ha calculado la toxicidad promedio, variando
de una región a otra. Particularmente para cepas mexicanas Bustillos-Guzmán y
colaboradores (2012) reportaron valores de entre 21.75 ± 2.93 y 10 pg STX eq/cél en
cepas de Bahía Concepción; Band-Schmidt y colaboradores (2006) reportan una
toxicidad promedio de 13 a 101 pg STX eq/cél para cepas de Bahía de Mazatlán y
Durán-Riveroll (2014) encuentra valores máximos de 28 ± 5.1 y mínimos de 1.2 ± 0.3
pg STX eq/cél.
Sin embargo, en este estudio se encontraron valores máximos de 103.2 ± 43 y
mínimos de 1.7 ± 0.5 pg STX eq/cél en presencia de la comunidad bacteriana, siendo
el valor máximo superior a lo reportado, lo cual podría asociarse a dos cosas: 1) la
liberación de exudados bacterianos ricos en nitrógeno que favorecen la producción
de toxinas en el dinoflagelado y 2) la producción autónoma de toxinas PSP por parte
de la cianobacteria (Orr et al. 2013).
De acuerdo con Anderson y colaboradores (1990), el contenido de toxina es
un reflejo de la cantidad inicial de toxina en una célula y el equilibrio entre la síntesis
y la pérdida de la misma, ya sea por liberación al medio, catabolismo o división
celular. Sin embargo, en cepas que han sido inoculadas con bacterias específicas se

90
ha observado un patrón de disminución en la toxicidad celular, sugiriéndose que las
bacterias son factores de estrés y/o que liberan al medio metabolitos secundarios
precursores que desencadenan la síntesis de la toxina (Albinsson, 2011; Albinsson et
al. 2014).
IX. CONCLUSIÓN
Este estudio encontró una variación considerable en la dinámica de
crecimiento de Gymnodinium catenatum en presencia de una comunidad bacteriana
integrada por tres distintos microorganismos: bacterias heterótrofas filamentosas,
bacilos intracelulares y una cianobacteria extracelular que rodea a las células del
dinoflagelado en forma de parche. Sin embargo, con la información obtenida en el
presente trabajo no podemos determinar con exactitud el tipo de relación ecológica
entre estos microorganismos y el dinoflagelado pero basados en nuestros resultados,
proponemos la hipótesis de que la interacción entre las bacterias intracelulares, la
cianobacteria extracelular y el dinoflagelado es bimodal: durante la fase de
crecimiento exponencial del cultivo la relación es mutualista al presentarse un
crecimiento óptimo en todos los organismos.
Mientras que al envejecer el cultivo e iniciando la fase de declive, esta
interacción se vuelve parasitaria, debido a las modificaciones en el ciclo de vida que
presenta G. catenatum, desarrollando quistes temporales y cambios morfológicos en
sus células, lo que desencadena una alta mortalidad. Esta hipótesis nos permitiría
explicar la rápida desaparición de los florecimientos algales nocivos en la naturaleza,
sin la exclusión de otros factores.
En cuanto al crecimiento, no queda claro si estos microorganismos lo
favorecen ya que se obtuvo un factor estimulante en dos cepas e inhibitorio en otras
dos pero la formación de cadenas más largas de células si se ve favorecida, al
disminuir la proporción de aparición de células individuales en todas las cepas a lo
largo del cultivo. Además, la presencia de la comunidad bacteriana incrementó la
producción de la clorofila a, zeaxantina y β-carotenos.

91
Finalmente, los resultados de este estudio indican que la comunidad
bacteriana desempeña un papel importante en la producción de toxinas paralizantes
de G. catenatum, debido probablemente a interacciones bioquímicas entre especies.
Si bien es cierto que el perfil de toxinas no presentó modificaciones, si se muestra
que la cantidad total de toxina por célula disminuye, considerando a las bacterias y a
la cianobacteria, agentes estresantes. No obstante, aún se debe seguir estudiando
esta interacción ya que son muchas las interrogantes que quedan sin indicios de
respuesta, además que los mecanismos de acción son totalmente desconocidos.
X. LITERATURA CITADA
Adachi, M., Kanno, T., Okamoto, R., Itakura, S., Yamaguchi, M. y T. Nishijima. 2003. Population structure of Alexandrium (Dinophyceae) cyst formation-promoting bacteria in Hiroshima Bay, Japan. Applied and Environmental Microbiology. 69: 6560-6568.
Adachi, M., Matsubara, T., Okamoto, R., Nishijima, T., Itakura, S. y M. Yamaguchi. 2001. Inhibition of cyst formation in the toxic dinoflagellate Alexandrium (Dinophyceae) by bacteria from Hiroshima Bay, Japan. Aquatic Microbial Ecology 26: 223-233.
Alavi, M., Miller, T., Erlandson, K., Schneider, R. y R. Belas. 2001. Bacterial community associated with Pfiesteria-like dinoflagellate cultures. Applied and Environmental Microbiology. 3: 380-396.
Albinsson, M.E. 2011. Algal-bacterial interactions: a study of Gymnodinium catenatum and its associated bacteria. Tesis de Doctorado. Universidad de Tasmania. Australia 198 p.
Albinsson, M.E., Negri, A.P., Blackburn, S.I. y C.J.S. Bolch. 2014. Bacterial community affects toxin production by Gymnodinium catenatum. Plos one 9(8): e104623.
Allen, M.M. 1968. Simple conditions of growth of unicelular blue-green algae on plates. Journal of. Phycology. 4: 1-4.
Amin, S., Green, D.H., Hart, M.C., Küpper, F.C., Sunda, W.C. y C.J. Carrano. 2009. Photolysis of iron-siderophore chelates promotes bacterial-algal mutualism. Proceedings of the National Academy of Sciences. 106: 17071-17076.
Amin, S., Küpper, F.C., Green, D.H., Harris, W.R. y C.J. Carrano. 2007. Boron binding by a siderophore isolated from marine bacteria associated with the toxic dinoflagellate Gymnodinium catenatum. Journal of the American Chemical Society 129: 478–479.
Anderson, D.M., Kulis, J.J., S. Sullivan, C. Hall y S. Lee, 1990. Dynamics and physiology of saxitoxin production by the dinoflagellates Alexandrium spp. Marine Biology. 104: 511–524.
Anderson, D.M. y O.J. Garrison. 1997. The ecology and oceanography of Harmful Algal Blooms. Limnology Oceanography. 42:1009-1035.

92
Arzul, G., Seguel, M., Guzmán, L. y E. Denn. 1999. Comparison of allelopathic properties in three toxic Alexandrium species. Journal of Experimental Marine Biologyand Ecology. 232: 285-295.
Azam, F. 1998. Microbial control of oceanic carbon flux: the plot thickens. Science 280: 694-696.
Balech, E., 1964. El plancton de Mar del Plata durante el período 1961-1962. Boletín del Instituto de Biología Marina, Universidad Nacional de Buenos Aires, Mar del Plata. 4: 1-49.
Band-Schmidt, C.J., Bustillos-Guzmán, J.J., Morquecho, L., Gárate-Lizárraga, I., Alonso-Rodríguez, R., Reyes-Salinas, A., Erler, K. y B. Luckas, 2006. Variations of PSP toxin profiles during different growth phases in Gymnodinium catenatum (Dinophyceae) strains isolated from three locations in the Gulf of California, Mexico. Journal of Phycology. 42: 757–768.
Band-Schmidt, C.J., Bustillos-Guzmán, J.J., Hernández-Sandoval, F.E., Núñez-Vázquez, E.J. y D.J. López-Cortés, 2014. Effect of temperature on growth and paralytic toxin profiles in isolates of Gymnodinium catenatum (Dinophyceae) of the Pacific coast of Mexico. Toxicon 90:199–212.
Band-Schmidt, C.J., Bustillos-Guzmán, J.J., López-Cortés, D.J., Gárate-Lizárraga, I., Núñez-Vázquez, E.J. y F. Hernández-Sandoval. 2010. Ecological and physiological studies of Gymnodinium catenatum in the Mexican Pacific: a review. Marine Drugs 8: 1935-1961.
Band-Schmidt, C.J., Morquecho, L., Lechuga-Devéze, C.H. y M. Anderson, 2004. Effects of growth medium, temperature, salinity and seawater source on the growth of Gymnodinium catenatum (Dinophyceae) from Bahía Concepción, Gulf of California, Mexico. Journal of Plankton Research. 26: 1459–1470.
Betancourt, L., Cid, A. y J. Abalde. 1997. Variación en el crecimiento y composición pigmentaria de Synechococcus sp. IO9201 en respuesta a la calidad espectral de la luz. Nova Acta Científica Compostelana. 8: 5-13.
Blackburn, S.I., Bolch, C.J., Haskard, C.J.S. y G.M. Hallegraeff. 2001. Reproductive compatibility among four global populations of the toxic dinoflagellate Gymnodinium catenatum (Dinophyceae). Phycologia 40(1): 78-87.
Blackburn, S.I., Hallegraeff, G.M. y C.J. Bolch. 1989. Vegetative reproduction and sexual life cycle of the toxic dinoflagellate Gymnodinium catenatum from Tasmania, Australia. Journal of Phycology. 25: 577-590.
Bolch, C.J.S., Vincent, B., Blackburn, S.I. y D.H. Green. 2004. Host-symbiont range of growth stimulating bacteria associated with Gymnodinium catenatum. Harmful Algae Blooms. 6: 465-485.
Bolch, J.S., Subramanian, T.A. y H.G. Green. 2011. The toxic dinoflagellate Gymnodinium catenatum (Dinophyceae) requires marine bacteria for growth. Phycology. 47: 1009-1022.
Bouvier, T., Del Giorgio, P.A. y J.M. Gasol. 2007. A comparative study of the cytometric characteristics of high and low nucleic-acid bacterioplancton cells from different aquatic ecosystems. Environmental. Microbiology. 9(8): 2050 – 2066.

93
Boyer, G.L., Sullivan, J.J., Andersen, R.J., Harrison, P.J. y F.J.R. Taylor. 1987. Effects of nutrient limitation on toxin production and composition in the marine dinoflagellate Protogonyaulax tamarensis. Marine Biology. 96: 123–128.
Bravo, I., 1986. Germinación de quistes, cultivo y enquistamiento de Gymnodinium catenatum Graham. Scientia Marina. 50(3): 313–321.
Bustillos-Guzmán, J.J., Band-Schmidt, C.J., Durán-Riveroll, L.M., Hernández-Sandoval, F.E., López-Cortés, D.J., Núñez-Vázquez, E.J., Cembella, A. y B. Krock. 2015. Paralytic toxin profile of the marine dinoflagellate Gymnodinium catenatum Graham from the Mexican Pacific as revealed by LC-MS/MS. Food Additives & Contaminants: Part A. 32: 1-15.
Bustillos-Guzmán, J.J., Band-Schmidt, C.J., López-Cortés, D.J., Gárate-Lizárraga, I., Núñez-Vázquez, E.J. y F.E. Hernández-Sandoval. 2012. Variaciones en el crecimiento y toxicidad en Gymnodinium catenatum Graham del Golfo de California bajo diferentes proporciones de nitrógeno y fósforo. Ciencias Marinas. 38: 101-117.
Calvo-Díaz, A. y X.A.G. Morán. 2006. Seasonal dynamics of picoplankton in shelf Waters of the southern Bay of Biscay. Aquatic Microbial Ecology. 42: 159 – 174.
Camino-Ordás, M., Fraga, S., Franco, J.M., Ordás, A. y A. Figuieras, 2004. Toxin and molecular analysis of Gymnodinium catenatum (Dinophyceae) strains from Galicia (NW Spain) and Andalucía (S Spain). Journal of Plankton Research. (26)3: 341–349.
Campbell, L. 2001. Flow Cytometric Analysis of Autotrophic Picoplankton. Texas A&M University, College Station. USA. 27 p.
Castillo-Navarrete, J.L. 2005. Citometría de flujo en la evaluación de potencial de membrana y viabilidad celular de Helicobacter pylori. Tesis de Maestría. Universidad de Concepción. Chile. 144 p.
Cembella, A.D., 1998. Ecophysiology and metabolism of paralytic shellfish toxins in marine microalgae. Phycologia 42(4): 420–447.
Córdova, J.L., Escudero, C. y J. Bustamante. 2003. Blooms inside the Bloom: intracelular bacteria multiplication within toxic dinoflagellates. Biología Marina y Oceanografía. 38(2): 57-67.
Costa, P.R., Robertson, A. y M.A. Quilliam. 2015. Toxin profile of Gymnodinium catenatum (Dinophyceae) from the Portuguese coast, as determined by liquid chromatography tándem mass spectrometry. Marine Drugs. 13: 2046-2062.
Croft, M.T., Lawrence, A.D., Raux-Deery, E., Warren, M.J. y A.G. Smith. 2005. Algae accrue vitamine B12 through a symbiotic relationship with bacteria. Nature. 438: 90-93.
Dang, H. y C.R. Lovell. 2000. Bacterial primary colonization and early succession on surfaces in marine waters as determined by amplified rRNA gene restriction analysis and sequence analysis of 16S rRNA genes. Applied and Environmental Microbiology 66(2): 467-475.
Day, R. y G. Quinn. 1989. Statistical comparisons of the treatments after analysis of variance. Ecological Monographs. 59: 430-460.
Díaz-Hernández, C. 1997. Abundancia de cianobacterias en la corriente de California y Cuenca Delfín y absorción de luz estimada por Synechococcus DC-ZWH7803 como modelo. Tesis de Maestría. CICIMAR-IPN. México. 68 p.

94
Doucette, G.J. y C.L. Powell. 1998. Algal-Bacterial interactions: can they determine the PSP-related toxicity of dinoflagellates. Harmful Algae Blooms. 3: 406-409.
Durán-Riveroll, L.M. 2014. Toxinas paralizantes tipo benzoil en el dinoflagelado Gymnodinium catenatum Graham y modelo teórico de interacción con el canal de sodio. Tesis de Doctorado. Centro Interdisciplinario de Ciencias Marinas – Instituto Politécnico Nacional. México. 213 p.
Fernández-Herrera, L.J. 2013. Efecto alelopático de Chattonella marina var. Marina (Raphydophyceae) sobre Gymnodinium catenatum (Dinophyceae) en condiciones de cultivo. Tesis de Mestría. CICIMAR-IPN. México. 76 p.
Ferrier, M., Martin, J.L. y J.N. Rooney-Varga. 2002. Stimulation of Alexandrium fundyense growth by bacterial assemblages from the Bay of Fundy. Journal of Applied Microbiology 92: 16-706.
Figueroa, R.I. 2005. The significance of sexuality and cyst formation in the life-cycles of four marine dinoflagellate species. Tesis de Doctorado. Lund University. Suecia. 154 p.
Figueroa, R.I., Garcés, E. e I. Ramilo. 2006. Nuclear features and effect of nutrients on Gymnodinium catenatum (Dinophyceae) sexual stages. Journal of Phycology. 42: 67-77.
Figueroa, R.I., Bravo, I., Ramilo, I., Pazos, A. y A. Moroño. 2008. New life-cycle stages of Gymnodinium catenatum (Dinophyceae): laboratory and field observations. Aquatic Microbial Ecology. 52: 13-23.
Fistarol, G.O., Legrand, C., Rengefors, K. y E. Granéli. 2004. Temporary cyst formation in phytoplankton: a response to allelopathic competitors? Environmental Microbiology. 6: 791–798.
Forget, N., Belzile, C., Rioux, P. y C. Nozais. 2010. Teaching the microbial growth curve concept using microalgal cultures and flow cytometry. Journal Biological Education. 44: 4-9.
Fuenmayor, G., Jonte, L., Rosales-Loaiza, N. y N. Morales. 2009. Efecto de la salinidad y la concentración de nutrientes sobre el crecimiento y composición bioquímica de la cianobacteria autóctona Oscillatoria sp. Ciencia marina. 17: 50-57.
Fuentes-Pascacio, J.A. 2010. Efecto de toxinas paralizantes de Gymnodinium catenatum en el copépodo Pseudodiaptomus euryhalinus. Tesis de Licenciatura. UABCS. México. 75 p.
Gallacher, S. y E.A. Smith. 1999. Bacteria and paralytic shellfish toxins. Protist. 150: 245-255.
Gallacher, S., Flynn, K.J., Franco, J.M., Brueggemann, E.E. y H.B. Hines. 1997. Evidence for production of paralytic shellfish toxins by bacteria associated with Alexandrium spp. (Dinophyta) in culture. Applied Environmental Microbiology 63: 239-245.
Gárate-Lizárraga, I., Bustillos-Guzmán, J.J., Morquecho L., Band-Schmidt C.J., Alonso-Rodríguez, R., Erler, K., Luckas, B., Reyes-Salinas, A. y D.T. Góngora-González, 2005. Comparative paralytic shellfish toxin profiles in the strains of Gymnodinium catenatum Graham from the Gulf of California, Mexico. Marine Pollution Bulletin. 50: 208–236.

95
Gasol, J.M. y P.A. del Giorgio. 2000. The flow cytometric determination of bacterioplankton abundance and community structure. Scientia Marina. 64: 197-224.
González-González, L.M. 2010. Influencia de la deficiencia de nitrógeno y fósforo en las interacciones competitivas entre Chlorella vulgaris y Scenedesmus acutus. Tesis de Licenciatura. Universidad Nacional de Colombia. Colombia. 65 p.
Graham, H.W., 1943. Gymnodinium catenatum, a new dinoflagellate from the Gulf of California. Transactionsof the American Microscopical Society. 62: 259–261.
Green, D.H., Llewellyn, L.E., Negri, A.P., Blackburn, S.I. y C.J. Bolch. 2004. Phylogenetic and functional diversity of the cultivable bacterial community associated with the paralytic shellfish poisoning dinoflagellate Gymnodinium catenatum. FEMS Microbiology Ecology. 47: 345-357.
Green, D.H., Hart, M.C., Blackburn, S.I. y C.J.S. Bolch. 2010. Bacterial diversity of Gymnodinium catenatum and its relationship to dinoflagellate toxicity. Aquatic Microbial Ecology 61: 73–87.
Guillard, R.R.L. 1973. Division rates. Handbook of phycological methods. Cambridge University Press. E.U.A. 289-312.
Hada, Y. 1967. Protozoan plankton of the Inland Sea Setonaikai. I. The Mastigophora. Bulletin of Suzugamine Women’s College of Natural Science 13: 1–26.
Hallegraeff, G., Nichols, P., Volkman, J.K., Blackburn, S.I. y D.A. Everitt. 1991. Pigments, fatty acids and sterols of the toxic dinoflagellate Gymnodinium catenatum. Journal of Phycology. 27: 591-599.
Hallegraeff, G.M. 1993. Review of harmful algal blooms and their apparent global increase. Journal of Phycology. 32: 79-99.
Hallegraeff, G.M., Anderson, D.M. y A.D. Cembella. 1995. Manual on Harmful Marine Microalgae. UNESCO. 11: 25-49.
Hallegraeff, G.M., Blackburn, S.I., Doblin, M.A. y C.J.S. Bolch. 2011. Global toxicology, ecophysiology and population relationships of the chain forming PST dinoflagellate Gymnodinium catenatum. Harmful Algae Blooms. 14: 130-143.
Hold, G.L., Smith, E.A., Birkbeck, T.H. y S. Gallacher. 2001. Comparison of paralytic shellfish toxin (PST) production by the dinoflagellates Alexandrium lusitanicum NEPCC 253 and Alexandrium tamarense NEPCC 407 in the presence and absence of bacteria. FEMS Microbiology Ecology. 36: 223-234.
Hummert, C., Ritscher, M., Reinhardt, K. y B. Luckas. 1997. Analysis of the characteristic PSP profiles of Pyrodinium bahamense and several strains of Alexandrium by HPLC based on ion-pair chromatographic separation, post-column oxidation, and fluorescence detection. Chromatographia. 45: 312-316.
Jeong, H. J., Park, J.Y., Nho, J.H., Park, M.O., Ha, J.H., Seong, K.A., Jeng, C., Seong, C.N., Lee, K.Y. y W.H. Yih. 2005a. Feeding by red-tide dinoflagellates on the cyano-bacterium Synechococcus. Aquatic Microbial Ecology. 41:1331–43.
Jeong, H. J., Park, K.Y., Nho, J.H., Park, M.O., Ha, J.H., Seong, K.A., Jeng, C., Seong, C.N., Lee, K.Y. y W.H. Yih. 2005b. Feeding by phototrophic redtide dinoflagellates: five species newly revealed and six species previously known to be mixotrophic. Aquatic Microbial Ecology. 40: 133–50.

96
Kepner, R.L. y J.R. Pratt. 1994. Use of fluorochromes for direct enumeration of total bacteria in environmental samples: past and present. Aquatic Microbial Ecology. 54(4): 603-615.
Kim, M. y M.G. Park. 2010. The thecate mixotrophic dinoflagellate Fragilidium duplocampanaeforme: feeding, prey specificity and allelopathy. 14th International Conference on Harmful Algae. Abstract Book. Grecia. 2010. 110 p.
La Ferla, R., Giudice, A.L. y G. Maimone. 2003. Morphology and LPS content for the estimation of marine bacterioplankton biomass in the Ionian Sea. Scientia Marina. 68: 23 - 32.
Legendre, L., Courties, C y M. Troussellier. 2001. Flow Cytometry in Oceanography 1989-1999. Cytometry. 44(3): 164 -172.
Loreto, C., Rosales, N., Bermúdez, J. y E. Morales. 2003. Producción de pigmentos y proteínas de la cianobacteria Anabaena PCC 7120 en relación a la concentración de nitrógeno e irradiancia. Gayana Botánica. 60: 83-89.
Ludeña-Hinojosa, Y. 2007. Cianobacterias en la Bahía de Mayagüez: abundancia, distribución y su relación con las propiedades bio-ópticas. Tesis de Maestría. Universidad de Puerto Rico. Puerto Rico. 130 p.
Maas, E.W., Latter, R.M., Thiele, J., Waite, A.M., Heather, J. y L. Brooks. 2007. Effect of multiple antibiotic treatments on a paralytic shellfish toxin-producing culture of the dinoflagellate Alexandrium minitum. Aquatic Microbial Ecology. 48: 255-260.
Martínez-Tecuapacho, G. A. 2009. Efecto del medio de cultivo en el crecimiento y toxicidad del dinoflagelado Gymnodinium catenatum Graham, aislado de la Bahía de Mazatlán, Sinaloa, México. Tesis de Licenciatura. Tecnológico de Estudios Superiores Huixquilucan. México. 63 p.
Matsumoto, M. 2011. Aspects of the interaction between the marine bacterium Alcanivorax DG881 and the toxic dinoflagellate Gymnodinium catenatum. Tesis de Doctorado. Universidad de Tasmania. Australia. 96 p.
Mayali, X. y F. Azam. 2000. Dissolver organic carbon from phytoplankton. The Handbook of Environmental Chemistry. Vol. 5. Part D. Springer-Verlag, Berlín, Heidelberg. 112-147 p.
Mecalco-Hernández, A. 2010. Dinámica del bacterioplancton y su relación con la biomasa fitoplanctónica en dos lagunas costeras del estado de Veracruz. Tesis de Maestría. UAM. México. 157 p.
Mendoza-Guerrero, A. 2011. Caracterización de la cinética de crecimiento de cianobacterias rizosféricas de Avicennia germinans. Tesis de Licenciatura. Universidad Veracruzana. México. 48 p.
Monroy-Sampieri, C. 2014. Cultivo de Chrorella vulgaris en medio basal Bold modificado en condiciones de luminosidad baja y caracterización de extracto líquido de lombricompostaje como sustituto de medio de cultivo. Tesina. Universidad Veracruzana. México. 56 p.
Montura, R.F.C. y D. Repeta. 1997. Calibration methods for HPLC. In: Phytoplankton pigments in oceanography: guidelines to modern methods. UNESCO. París. 407-428 p.

97
Morales, E., Rodríguez, M., García, D., Loreto, C. y E. Marco. 2002. Crecimiento, producción de pigmentos y exopolisacáridos de la cianobacteria Anabaena sp. PCC7120 en función del pH y CO2. Interciencia. 27: 7-13.
Mundt, S., Kreitlow, S., Nowotny, A. y U. Effmert. 2001. Biochemical and farmacological investigations of selected cyanobacteria. Journal Environmental Health. 203: 327-334.
Negri, A.P., Bolch, C.J.S., Geier, S., Green, D.H., Park T.G. y S.I. Blackburn. 2007. Widespread presence of hydrophobic paralytic shellfish toxins in Gymnodinium catenatum. Harmful Algae Blooms. 6: 774–780.
Núñez-Vázquez, E.J., Gárate-Lizárraga, I., Band-Schmidt, C.J., Cordero-Tapia, A., López-Cortés, D., Hernández-Sandoval, F., Heredia-Tapia, A. y J. Bustillos-Guzmán. 2008. Impacto de las proliferaciones microalgales nocivas en animales silvestres y de cultivo en el Golfo de California. Instituto Nacional de Ecología. México. 7 p.
Ogawa, T. y G. Terui. (1970). Studies on the growth of Spirulina platensis. On the pure culture of Spirulina platensis. Journal of Fermentation Technology. 48: 361–367.
Oh, S.J., Matsuyama, Y., Yoon, Y.H., Miyamura, K., Choi, C.G., Yang, H.S. e I.J. Kan, 2010. Comparative analysis of paralytic shellfish toxin content and profile produced by dinoflagellate Gymnodinium catenatum isolated from Inokushi Bay, Japan. Journal of the Faculty of Agriculture Kyushu Univ. 55(1): 47–54.
Olivas, E.E. 2004. Manual de Prácticas, Microbiología I, II y Parasitología: Programa de Medicina. Universidad Autónoma de Ciudad Juárez. México. 105 p.
Ortíz-Moreno, M.L., Cortés-Castillo, C.E., Sánchez-Villarraga, J., Padilla, J. y A.M. Otero-Paternina. 2012. Evaluación del crecimiento de la microalga Chlorella sorokiniana en diferentes medios de cultivo en condiciones autotróficas y mixotróficas. Orinoquia. 16: 2-12.
Pérez-Rontomé, M.C. 2000. El ciclo de las xantofilas en plantas sometidas a estrés nutricional o climático. Tesis de Doctorado. Consejo Superior de Investigaciones Científicas. España. 254 p.
Poot-Delgado, C.A., Martínez-López, A. y C.J. Band-Schmidt. 2009. Influencia de la salinidad en el crecimiento de la cepa SNCI_IX04_S (cianobacteria cocoidal), aislada de la laguna de Macapule, Sinaloa, México. XVI Congreso Nacional de Ciencia y Tecnología del Mar. 2 p.
Provasoli, L. 1957. Development of artificial media for marine algae. Archivesof Microbiology. 25: 392-428.
Quijano-Scheggia, S.A., Olivos-Ortíz, J.J., Bustillos-Guzmán, J.J., Gaviño-Rodríguez, J.H., Galicia-Pérez, M.A., Patiño-Barragán, M., Band-Schmidt, C.J. Hernández-Sandoval, F.E. y D.J. López-Cortés, 2012. Bloom of Gymnodinium catenatum in Bahía Santiago and Bahía Manzanillo, Colima, México. BiologíaTropical. 60: 173–186.
Ramírez-Gama, R.M., Luna-Millán, B., Velázquez-Madrazo, O., Vierna-García, L., Mejía-Chávez, A., Tsuzuki Reyes, G., Hernández-Gómez, L., Müggenburg, I., Camacho-Cruz, A y M. del C. Urzúa-Hernández. 2006. Manual de Prácticas de Microbiología General. UNAM. México. 135 p.

98
Reis-Costa, P., João, M. y K.A. Lefebvre. 2010. Characterization of paralytic shellfish toxins in seawater and sardines (Sardina pilchardus) during blooms of Gymnodinium catenatum. Hidrobiología. 655: 89-97.
Rippka, R. 1988. Isolation and purification of cianobacteria. Methods in Enzymology. 167: 3-27.
Rooney-Varga J.N., Giewat, M.W., Savin, M.C., Sood, S., LeGresley, M. y J.L. Martin. 2005. Links between phytoplankton and bacterial community dynamics in a coastal marine environment. FEMS Microbiology Ecology. 49: 75-163.
Rosales-Loaiza, N., Guevara, M., Lodeiros, C. y E. Morales. 2008. Growth and metabolite production of the marine cyanobacterium Synechococcus sp. (Chroococcales) in function to irradiance. Biología Tropical. 56(2): 421-429.
Saldate-Castañeda, O., Vázquez-Castellanos, J.L., Galván, J., Sánchez-Anguiano, A. y A. Nazar. 1991. Intoxicaciones por toxina paralizante de moluscos en Oaxaca. Laboratorio Nacional de Salud Pública. Oaxaca. 8 p.
Seibold, A., Wichels, A. y C. Schütt. 2001. Diversity of endocytic bacteria in the dinoflagellate Noctiluca scintillans. Aquatic Microbial Ecology. 25:229-235.
Shapiro, H.M. 2003. Multiparameter flow cytometry of bacteria: implications for diagnosis and therapeutics. Cytometry. 43: 223-226.
Skelton, H. M., Burkholder, J.M. y M.W. Parrow. 2009. Axenic culture of the heterotrophic dinoflagellate Pfiesteria shumwayae in a semi-defined medium. Journal of Eukaryotic Microbiology. 56: 73–82.
Skovgaard, A. 2000. A phagotrophically derivable growth factor in the plastidic dinoflagellate Gyrodinium resplendens (Dinophyceae) Journal of Phycology. 36:1096-1078.
Smith, E.A., Grant, F., Ferguson, C.M. y S. Gallacher. 2001. Biotransformations of paralytic shellfish toxins by bacteria isolated from bivalve molluscs. Applied Environmental Microbiology. 67: 2345-2353.
Soto-Castor, R., Bulit, C., Esquivel-Herrera, C.A. y A. Pérez-Rojas. 2002. Bacterial abundance and hidrological variation in a tropical lagoon during the rainy season. Oceánides. 17(1): 13-29.
Subramanian, T. A. 2008. Experimental models of algal-bacterial interactions. Tesis de Doctorado. Universidad de Tasmania. Australia. 143 p.
Taroncher-Oldenburg, G. y D.M. Anderson. 2000. Identification and characterization of three differentially expressed genes, encoding S-Adenosylhomocysteine Hydrolase, Methionine Aminopeptidase, and a histone-like protein, in the toxic dinoflagellate Alexandrium fundyense. Applied Environmental Microbiology. 66(5): 2105–2112.
Taroncher-Oldenburg, G., Kulis, D. y D.M. Anderson. 1997. Toxin variability during the cell cycle of the dinoflagellate Alexandrium fundyense. Limnology Oceanography. 42(5, part 2): 1178–1188.
Throndsen, J. 1979. Special methods-micromanipulators. In: Stein, R.J., Ed. Handbook of Phycological Methods: Culture Methods and Growth Measurements. Cambridge. London. 139-144 p.

99
Töbe, K. 2003. Investigations into the role of bacteria/dinoflagellate interactions in Paralytic Shellfish Poisoning. Tesis de Doctorado. Universität Bremen. Bremen. 178 p.
Torres-Ariño, 2001. Uso de cianobacterias en la producción de antibióticos. Ciencia y Mar. 11 p.
Torres-Ariño, A. 2004. Uso de cianobacterias en la producción de antibióticos. Ciencia y Mar. VIII (23): 43-52.
Torres-Ariño, A. 2011. Aislamiento y caracterización de cianobacterias potencialmente tóxicas para Litopenaeus vannamei. Aquacultura. 86: 34-36.
Tovar-Ramírez, D. 1999. Aislamiento y caracterización de Microcoleus (Oscillatoriaceae: cianobacteria) de los tapetes microbianos de los canales de marea de San Carlos, B.C.S., México. Tesis de Licenciatura. Universidad Michoacana de San Nicolás de Hidalgo. Michoacán. 94 p.
Uchida, T., Toda, S., Matsuyama, M., Yamaguchi, M., Kotani, M. y T. Honjo. 1999. Interactions between the red tide dinoflagellates Heterocapsa circularisquama and Gymnodinium mikimotoi in laboratory culture. Journal of Experimental Marine Biology and Ecology. 241: 285–299.
Uribe, P. y R.T. Espejo. 2003. Effects of associated bacteria on the growth and toxicity of Alexandrium catenella. Applied and Environmental Microbiology 69: 659-662.
Vázquez, C., Martín, A., Silóniz, M.I. y S. Serrano. 2010. Técnicas básicas de Microbiología, observación de bacterias. Microbiología. 3(5): 15-38.
Venrick, E.L. 1978. How many cells to count? UNESCO. París, 167-180 p.
Vidussi, F., Claustre, H., Bustillos-Guzmán, J.J., Cailleau, C. y J.C. Marty. 1996. Rapid HPLC method for determination of phytoplankton chemotaxonomic pigments: separation of chlorophyll a from divinyl-chlorophyll a and zeaxanthin from lutein. Journal of Plankton Research. 18: 2377-2382.
Yu, R. C., Hummert, C., Luckas, B., Qian, P. Y. y M.J. Zhou. 1998. Modified HPLC method for analysis of PSP toxins in algae and shellfish from China. Chromatographia. 48: 671-676.
Zar, J.H. 1999. Biostatistical analysis. 4th Ed. Prentice Hall, Upper Saddle River. 1046 p.
Zhuang, Y., Zhang, H., Hannick, L. y S. Lin. 2012. Cell cycle, saxitoxin, and proton pump rhodopsin: insight from gene expression profiling for an Alexandrium fundyense culture and a natural bloom. Phycology. 48(1): 527.