efecto de la dieta en la composiciÓn de … · efecto de la dieta en la composiciÓn de Ácidos...
TRANSCRIPT

INSTITUTO POLITECNICO NACIONAL CENTRO INTERDISCIPLINARIO DE CIENCIAS MARINAS
EFECTO DE LA DIETA EN LA COMPOSICIÓN DE ÁCIDOS GRASOS DEL ALIMENTO VIVO UTILIZADO EN LA
CRIANZA LARVARIA DE PECES MARINOS
TESIS
QUE PARA OBTENER EL GRADO DE MAESTRÍA EN CIENCIAS EN MANEJO DE RECURSOS MARINOS
PRESENTA
IVÁN HERNÁNDEZ ALARCÓN
LA PAZ, B.C.S., DICIEMBRE DE 2016



DEDICATORIA
“No alegrarse de la propia tarea es equivocarse de camino, por muy inteligente
que se sea”
Lao Tse, Tao Te Ching.

AGRADECIMIENTOS Al Instituto Politécnico Nacional y al Centro Interdisciplinario de Ciencias Marinas,
por la formación académica obtenida a través del grupo de investigadores y
docentes.
Al Consejo Nacional de Ciencia y Tecnología (CONACYT) y Beca de Estímulo
Institucional de Formación de Investigadores (BEIFI) del Instituto Politécnico
Nacional, por las becas otorgadas.
Al Dr. Renato Peña y la Dra. Silvie Dumas por darme la oportunidad de trabajar
bajo su dirección y de recibirme como su estudiante.
Al comité revisor Dra. Bárbara Gonzalez Acosta Dr. José de La Cruz Agüero, M.
en C. Mauricio Contreras Olguín por su tiempo, comentarios y revisiones.
Al personal de la Unidad Piloto de Maricultivo: M. en C. Mauricio Contreras
Olguín, Biol. Laura Flores Montijo, técnico Sául Iván León por su apoyo y
colaboración en el desarrollo de este trabajo, y sobretodo por su amistad.
A la Dra. Elena Palacios Mechetnov por permitir el procesamiento de las muestras
para la determinación de ácidos grasos y a todo el laboratorio de Metabolismo de
lípidos del CIBNOR en especial a la M. en C Olivia Arjona López por el apoyo
recibido.

ÍNDICE GENERAL
RELACIÓN DE FIGURAS I RELACIÓN DE TABLAS II LÉXICO Y LISTA DE ACRÓNIMOS IV RESUMEN V ABSTRACT VI 1. INTRODUCCIÓN 1 1.1 Antecedentes 4 1.2 Justificación 9 2.HIPÓTESIS 10 3. OBJETIVO GENERAL 10 3.1 Objetivos Particulares 10 4. MATERIAL Y MÉTODOS 11 4.1 Cultivo de Alimento Vivo 11 4.1.1 Microalgas 11 4.1.2 Rotíferos 12 4.1.3 Copépodos 14 4.1.4 Artemia sp. 15 4.2 Determinación de lípidos totales 16 4.3 Análisis de ácidos grasos 16 5. RESULTADOS 17 6. DISCUSIÒN 32 7. CONCLUSIONES 43 8. RECOMENDACIONES 44 BIBLIOGRAFÌA 45 ANEXOS 60

I
RELACIÓN DE FIGURAS
Figura 1. Proporción de lípidos totales encontrados en las diferentes microalgas
evaluadas…………………………………………………………………17
Figura 2. Proporción de lípidos totales en rotíferos alimentados con diferentes
dietas………………………………………………………………………18
Figura 3. Proporción de lípidos totales en copépodos P. euryhalinus y P.
crassirostris en diferentes estadios de desarrollo y alimentados con
dietas con base en microalgas…………………………………………..19
Figura 4. Proporción de lípidos totales en nauplios de Artemia sp. recién nacidos,
nauplios enriquecidos con Spresso-Selco y juveniles alimentados con
una mezcla de cereales complementada con microalgas…………....19
Figura 5. Proporciones de los ácidos grasos esenciales en las microalgas
Nanochloropsis oculata, Tetraselmis suecica, Chaetoceros calcitrans
e Isochysis galbana. ……………………………………………………..21
Figura 6. Proporciones de los ácidos grasos esenciales en Brachionus plicatilis
alimentados con distintas dietas………………………………………...23
Figura 7. Proporciones de los ácidos grasos esenciales en rotíferos Brachionus
rotundiformis alimentados con distintas dietas……………………...…24
Figura 8. Proporciones de ácidos grasos esenciales en rotífero Proales similis
alimentados con distintas dietas………………………………………...26
Figura 9. Proporciones de ácidos grasos esenciales en diferentes estadios de
desarrollo del copépodo Pseudodiaptomus euryhalinus alimentados
con Chaetoceros calcitrans y una mezcla de C. calcitrans y
Tetraselmis suecica. ……………………………………………………..28
Figura 10. Proporciones de los ácidos grasos esenciales en diferentes estadios
de desarrollo del copépodo Parvocalanus crassirostris alimentados
con I.galbana y una mezcla de I. galbana, C. calcitrans y T.
Suecica............................………………………………………………..30
Figura 11. Proporciones de los ácidos grasos esenciales en diferentes estadios
de desarrollo de Artemia sp.: nauplio recién nacido (sin alimento),
meta-nauplio II-III (enriquecido con Spresso-Selco) y juvenil
(alimentado con una mezcla de cereales complementada con
microalgas. ………………………………………………………………..32

II
RELACIÓN DE TABLAS Tabla I. Requerimientos sugeridos de ácidos grasos y DHA:EPA de distintas
especies de peces marinos. ………………………………………………5
Tabla II. Contenido de ARA, EPA, DHA y la relación DHA:EPA reportados en
diferentes especies de rotíferos producidos con diferentes
alimentos……..……………………………………………………………...6
Tabla III. Contenido de ARA, EPA, DHA y la relación DHA:EPA reportados en
diferentes especies de copépodos. ………………………………………8
Tabla IV. Características de los tratamientos alimenticios utilizados para rotíferos
Brachionus plicatilis, B. rotundiformis y Proales similis. ……………….13
Tabla V. Características de los tratamientos utilizados para los copépodos
Pseudodiaptomus euryhalinus y Parvocalanus crassirostris……….....14
Tabla VI. Características de los tratamientos alimenticios utilizados para Artemia
sp. ……………………………………………………………………………15
Tabla VII. Composición de ácidos grasos (% del total de ácidos grasos) de las
microalgas utilizadas como alimento. ……………………………………20
Tabla VIII. Composición de ácidos grasos (% del total de ácidos grasos) del
rotífero Brachionus plicatilis alimentado con ORI-ONE, N. oculata, N.
oculata y enriquecido con Spresso-Selco, T. suecica y T.suecica
enriquecido con Spresso-Selco. ………………………………………….22
Tabla IX. Composición de ácidos grasos (% del total de ácidos grasos) del rotífero
Brachionus rotundiformis alimentado con ORI-ONE, N. oculata, N.
oculata y enriquecido con Spresso-Selco, T. suecica y T. suecica
enriquecido con Spresso-Selco…………………………………………...24
Tabla X. Composición de ácidos grasos (% del total de ácidos grasos) del rotífero
Proales similis alimentado con ORI-ONE, N. oculata y N.oculata
enriquecido con Spresso-Selco. ………………………………………….25
Tabla XI. Composición de ácidos grasos (% del total de ácidos grasos) del
copépodo Pseudodiaptomus euryhainus, en diferentes estadios de
desarrollo alimentados con C. calcitrans y una mezcla de C. calcitrans y
T. suecica. …………………………………………………………………..27
Tabla XII. Composición de ácidos grasos (% del total de ácidos grasos) del
copépodo Parvocalanus crassirostris en diferentes estadios de

III
desarrollo y alimentados con I. galbana y una mezcla de I. galbana, C.
calcitrans y T. suecica. …………………………………………………….29 Tabla XIII. Composición de ácidos grasos (% del total de ácidos grasos) de
Artemia sp. en diferentes estadios de desarrollo. ………………………31 Tabla XIV. Proporciones reportadas de ácidos grasos esenciales en rotíferos
enriquecidos con diferentes productos comerciales. …………………..37 Tabla XV. Proporciones de ácidos grasos esenciales, así como de supervivencias
en distintas especies de peces marinos utilizando rotíferos. ………….38

IV
LÉXICO Y LISTA DE ACRÓNIMOS
Ácidos grasos altamente insaturados (HUFA por sus siglas en ingles): Ácidos
grasos con 20 o más carbonos y tres dobles enlaces (insaturaciones)
(Gunstone et al., 1994).
Ácidos grasos esenciales (AGE): Ácidos grasos poliinsaturados de las series n-
3 y n-6, que no pueden ser sintetizados o solo en pequeñas cantidades y
deben ser obtenidos del alimento porque son necesarios para la vida de
los organismos (Gunstone et al., 1994).
Ácidos grasos monoinsaturados (MUFA por sus siglas en ingles): Ácidos
grasos con un doble enlace (Gunstone et al., 1994).
Ácidos grasos poliinsaturados (PUFA por sus siglas en ingles): Ácidos grasos
con más de un doble enlace. Los dobles enlaces tienen por lo general
configuración cis y son metilenos interrumpidos (Gunstone et al., 1994).
Ácidos grasos saturados (SFA por sus siglas en ingles): Ácidos grasos sin
doble enlace carbono-carbono (Gunstone et al., 1994).
ARA: Ácido araquidónico, ácido graso poliinsaturado esencial de la serie omega-
6, formado por una cadena de 20 carbonos con cuatro dobles enlaces.
EPA: Àcido eicosapentaenoico, ácido graso poliinsaturado esencial de la serie
omega 3 formado por 20 carbonos con cinco dobles enlaces.
DHA: Ácido docosahexaenoico, ácido graso esencial poliinsaturado de la serie
omega-3, formado por 22 carbonos con 6 dobles enlaces.

V
RESUMEN
Se investigó el efecto de la composición de ácidos grasos de las microalgas
Nannochloropsis oculata, Tetraselmis suecica, Chaetoceros calcitrans e
Isochrysis galbana; de una dieta comercial (ORI-ONE®) así como de un
enriquecedor comercial (Spresso-Selco®) en la composición de ácidos grasos de
las especies de alimento vivo que se utilizan como presas en acuacultura, para
alimentar a los primeros estadios larvarios de peces marinos en CICIMAR - IPN:
rotíferos (Brachionus plicatilis, B. rotundiformis y Proales similis); copépodos
(Pseudodiaptomus euryhalinus y Parvocalanus crassirostris); y Artemia sp. El
análisis de ácidos grasos en las microalgas indica que I. galbana tiene una
concentración superior de ácido docosahexaenoico (DHA), C. calcitrans es a su
vez superior en ácido eicosapentaenoico (EPA); y N. oculata en ácido
araquidónico (ARA). Las dietas multialgales basadas en I. galbana, C. calcitrans y
T. suecica produjeron mayores proporciones de DHA y ARA en el copépodo P.
crassirostris, respecto a las dietas monoalgales de I. galbana. El copépodo P.
euryhalinus presentó mayores proporciones de DHA y ARA cuando se alimenta
con dietas multialgales basadas en C. calcitrans y T. suecica, que con dietas
monoalgales de C. calcitrans. Los rotíferos B. rotundiformis, B. plicatilis y P. similis
alimentados o enriquecidos con productos comerciales incrementaron
notoriamente sus proporciones de DHA y EPA en comparación de dietas
monoalgales de N. oculata y T. suecica, pero disminuyeron sus proporciones de
ARA. Se confirmó que los rotíferos alimentados con la dieta comercial ORI-ONE®
no necesitaron un medio de cultivo adicional ya que sus proporciones de DHA,
EPA y ARA fueron similares a los rotíferos enriquecidos con Spresso-Selco®. Los
juveniles de Artemia sp. presentan valores bajos de ARA, EPA y DHA, por lo cual
se requiere administrar un enriquecedor comercial.

VI
ABSTRACT
The effect of the fatty acid composition of microalgae (Nannochloropsis oculata,
Tetraselmis suecica, Chaetoceros calcitrans and Isochrysis galbana) on rotifers
(Brachionus plicatilis, B. rotundiformis and Proales similis, feed with microalgae
diets, commercial diet ORI-ONE® and a commercial enriching Spresso-Selco®);
copepods (Pseudodiaptomus euryhalinus and Parvocalanus crassirostris, feed
with mono and multialgal diets) and Artemia sp. used in aquaculture, to feed
larvae marine fish in CICIMAR - IPN was investigated. The analysis of fatty acids
in microalgae I. galbana indicates that has a higher concentration of
docosahexaenoic acid (DHA), C. calcitrans is higher in eicosapentaenoic acid
(EPA) and N. oculata on arachidonic acid (ARA). The multialgales based diets I.
galbana, C. calcitrans and T. suecica produced higher proportions of DHA and
ARA in copepods P. crassirostris, regarding monoalgales I. galbana diets. The
copepod P. euryhalinus had higher proportions of DHA and ARA when fed with
diets based multialgaes C. calcitrans and T. suecica, than monoalgales diets C.
calcitrans. B. plicatilis, B. rotundiformis and P. similis rotifers feeds with
commercial products or enriched increased markedly proportions of DHA and EPA
in comparison with monoalgales diets N. oculata and T. suecica, but decreased
proportions of ARA. Fed the commercial diet ORI-ONE® rotifers do not need
additional microalgae cultive and his proportions of DHA, EPA and ARA were
similar to those rotifers enriched with Spresso-Selco®. Juvenile Artemia sp. have
low values of ARA, EPA and DHA if a commercial enrichment is not administered.

1
1. INTRODUCCIÓN
La acuicultura es el área de más rápido desarrollo en el campo de la producción
alimenticia en la actualidad, con un promedio anual de crecimiento mundial
cercano al 10% (FAO, 2014). En el año 1970, se cultivaban en el mundo
alrededor de 3 millones de toneladas de pescado; en 2012 la producción acuícola
mundial llegó a 66.6 millones de toneladas (FAO, 2014). El cultivo de peces
marinos se encuentra en expansión y tiene mucha relevancia, en gran medida,
porque representa una vía de producción de proteínas de origen marino para
satisfacer la demanda de la creciente población humana, lo que en permitirá
establecer un suministro continuo y ordenado de producto que, bajo una
adecuada planificación, estabilice los precios en los mercados y reduzca el
impacto de la actividad humana sobre el medio ambiente protegiendo a especies
sobre explotadas (Llorente-García, 2013). A nivel comercial se han logrado
producir especies como el sabalote (Chanos chanos), el barramundi (Lates
calcarifer), el besugo (Pagrus major), la lubina (Dicentrarchus labrax), la dorada
(Sparus aurata) y la corvina roja (Sciaenops ocellatus) (Tucker, 1998).
Permanece el cultivo larvario como la etapa que presenta mayor problemática
para lograr el desarrollo biotecnológico de las especies sujetas a cultivo (Peña,
2005). Esto ha provocado que el cultivo de larvas en peces marinos se considere
una etapa crítica o “cuello de botella” dentro de los esquemas de producción (Bell
et al., 2003; Bengtson, 2003) ya que uno de los principales problemas en la etapa
larvaria es la alta mortalidad registrada, la cual, salvo algunas excepciones,
sobrepasa el orden del 80% (Subagja et al., 1999; Álvarez-González, 1999;
Cerqueira & Brugger, 2001; Gracia-López et al., 2005). Muchos factores pueden
contribuir a estas elevadas tasas de mortalidad, sin embargo, los relacionados
con la alimentación y nutrición reciben particular atención debido al papel que
juegan durante el desarrollo larvario (Yufera & Darias, 2007). Resaltan dos etapas
dentro de la larvicultura que por su dificultad de sobrellevar limitan el éxito del
cultivo; una de ella es la primera alimentación, donde la larva debe iniciar
exitosamente la búsqueda y captura del alimento exógeno (Civera-Cerecedo et
al., 2004). Y la etapa conocida como “destete” o acondicionamiento (“weaning” en

2
inglés) que implica la transición de un alimento vivo a un alimento inerte
(Yakupitiyage, 2013).
En el medio natural, la primera alimentación se compone de complejas redes
tróficas que incluyen diatomeas, dinoflagelados, flagelados, tintínidos, ciliados,
cladóceros, copépodos, huevos de bivalvos, quetognatos, lamelibranquios,
gasterópodos, poliquetos, decápodos, etc. (Prieto et al., 2006). Desde el punto de
vista alimenticio, la larva se enfrenta a la necesidad de desarrollar las habilidades
institntivas de cazar y asimilar los nutrientes presentes en las presas. Para ello, la
larva debe estar capacitada para: 1) la ingestión del alimento (búsqueda y
captura), 2) la digestión (poner a disponibilidad los nutrientes), y 3) la absorción
de nutrientes (digestión intracelular, asimilación y biosíntesis (Civera-Cerecedo et
al., 2004).
En condiciones de cultivo, es necesario proveer de una presa adecuada como
primer alimento de las larvas. Para ello se deben considerar diferentes criterios
que influyen en el éxito alimenticio de la larva como seleccionar una presa de
tamaño adecuado y acorde al tamaño de la boca y de movimiento lento, para que
sea fácilmente percibido, pues la mayoría de las larvas de peces son cazadores
planctónicos visuales, sin importar los hábitos alimenticios que tendrán cuando
sean adultos (Hunter, 1981). Esta presa debe ser digerida y ser fácilmente
producida a niveles masivos, así como contener un adecuado grado nutricional
(Bengtson, 2003). Bajo estas consideraciones, muchos organismos han sido
utilizados como primer alimento vivo para el cultivo de larvas de peces. Se inició
con protozooarios, sin embargo fueron rápidamente remplazados por larvas
trocóforas de bivalvos, al tener mejor control sobre su producción y limpieza en
los tanques (Castro et al., 2003). Se utilizaron también nauplios de copépodos, los
cuales son muy abundantes en el zooplancton y tienen un valor nutricional mayor
y son la presa principal en el medio natural, lamentablemente muy pocas especies
pueden ser cultivadas masivamente. El nauplio del crustáceo branquiópodo
Artemia sp. también ha sido utilizado como alimento, pero su valor nutricional
cambia de acuerdo al lugar de origen de la cepa, y va disminuyendo a medida que
realiza su proceso de metamorfosis (Bell et al., 2003). A finales de los 60’s se
introdujo el rotífero Brachionus plicatilis, y gracias a su pequeño tamaño,

3
movimiento en el agua y corto ciclo de vida para su cultivo, ha sido una de las
presas más empleadas en el cultivo de larvas de peces (Hagiwara et al., 2001).
Sin embargo, no son utilizables en la larvicultura de algunas especies de peces
marinos como las familias Lutjanidae (Doi et al., 1994; Schipp et al., 1999),
Pomacanthidae (Tucker, 1998) o Serranidae (Toledo et al.,1999) debido al
reducido tamaño de boca de las larvas.
Parte del buen resultado obtenido al emplear alimento vivo en la primera
alimentación está relacionado con la presencia de ciertos nutrientes bioquímicos
indispensables, entre los que destacan los ácidos grasos poliinsaturados de
cadena larga (HUFA, por sus siglas en inglés) (FAO, 2006; Kraul, 1989; Tucker,
1998). Este término incluye a los ácidos grasos de la serie n-6 derivados del ácido
linoléico (18:2 n-6) y de la serie n-3 derivados del ácido linolénico (18:3 n-3),
debido a que los peces marinos no tiene la capacidad para transformarlos a ácido
eicosapentaenoico (20:5n-3, EPA) y ácido docosahexaenoico (22:6n-3, DHA),
éstos se consideran esenciales (AGE), esta limitación se debe a una insuficiencia
metabólica debido a la carencia de las enzimas elongasas del C18 al C20 y de las
Δ 5-desaturasas de ácidos grasos (Bell & Sargent, 2003). Por lo que las series n-3
y n-6, conocidas comúnmente como Omega-3 y Omega-6, al no poder ser
sintetizadas de novo, deben ser incorporados a través de los alimentos, ya sean
presas vivas o dietas inertes (Henderson & Tocher 1987; Sargent et al., 1995;
Sargent et al., 2002).
La importancia de los ácidos grasos en la nutrición de larvas de peces ha sido
investigada extensivamente durante los últimos 30 años. Numerosos estudios
reportan que la deficiencia en ácidos grasos esenciales tiene consecuencias que
van desde afectaciones en el crecimiento, malformaciones, miocarditis e
incrementos en la tasa de mortalidad (Castell et al., 1972; Hardy et al., 1987;
Santha & Gatlin, 1991; Kalogeropoulos et al., 1992; Kennish et al., 1992; Ruyter et
al., 2000; Glencross, 2009). aún cuando la importancia de los HUFAs está bien
reconocida, los requerimientos de éstos en larvas no está bien establecida, ya
que se ha reportado que varían tanto cualitativamente como cuantitativamente
entre las especies de peces (Copeman et al., 2002).

4
Los organismos utilizados como alimento vivo son generalmente producidos de
manera simultanea al cultivo de las larvas, y suelen presentan variaciones
nutricionales considerables (Rasdi et al., 2015). De esta forma, una adecuada
selección de microalgas puede ofrecer un paquete nutricional óptimo para las
presas que a su vez sea transferido a larvas de peces con requerimientos
nutricionales particulares (Brown, 1989). Por lo que este trabajo se enfoca en
evaluar el impacto de diferentes dietas de microalgas y algunos productos
comerciales, en la composición de ácidos grasos en rotíferos, copépodos y
Artemia sp.
1.1 Antecedentes
Los peces necesitan de ácidos grasos poliinsaturados para su desarrollo, la
carencia de éstos en la dieta tiene repercusiones en su crecimiento, desarrollo y
reproducción, además genera enfermedades y consecuentemente, altas
mortalidades (Glencross, 2009). Los requerimientos fluctúan cuantitativamente
durante la ontogenia y varían según las especies; sin embargo, no es sencillo la
cuantificación de requerimientos de ácidos grasos para larvas de peces debido a
limitantes fisiológicas, principalmente dadas a su sistema digestivo rudimentario y
poco desarrollado, por lo que existen solamente para un reducido número de
especies. Tocher (2010) reporta que los requerimientos de juveniles y subadultos
de peces dulceacuícolas y diádromos, pueden ser satisfechos con 1% (peso seco
de la dieta) de los ácidos grasos poliinsaturados (PUFA, por sus sigas en inglés)
de 18-C, ácido alfa- linolénico y el ácido linoleico. Para especies marinas como el
rodaballo Psetta maxima, el besugo, Pagrus major o la lubina europea
Dicentrarchus labrax los requerimientos de HUFA n-3 son menores al 1%, no
obstante, otras especies como Rhabdosargus sarba, Pseudocaranx dentex y
Pleuronectes ferrugineus requieren más del 1% (Kalogeropoulos et al., 1992;
Watanabe, 1993; Ibeas et al., 1994) (Tabla I). En la mayoría de las especies de
larvas de peces marinos cultivados, las microalgas constituyen el primer eslabón
de la cadena trófica (Reitan et al., 1997). Por sus características y respuesta a
condiciones de cultivo, las principales especies de microalgas utilizadas en
acuacultura son Nannochloropsis oculata, por poseer un tamaño pequeño y un
relativamente alto contenido de EPA (Sukenik et al., 1993); Tetraselmis tetrahele
o T. suecica con un alto contenido de EPA (Brown, 1989); Isochrysis galbana, con
5
un elevado contenido de DHA; Chaetoceros muelleri pobre en DHA, pero con
elevados contenidos de EPA y en ARA (Renaud et al., 2002) y Pavlova lutheri con
elevados valores de DHA y EPA (Tonon et al., 2003).
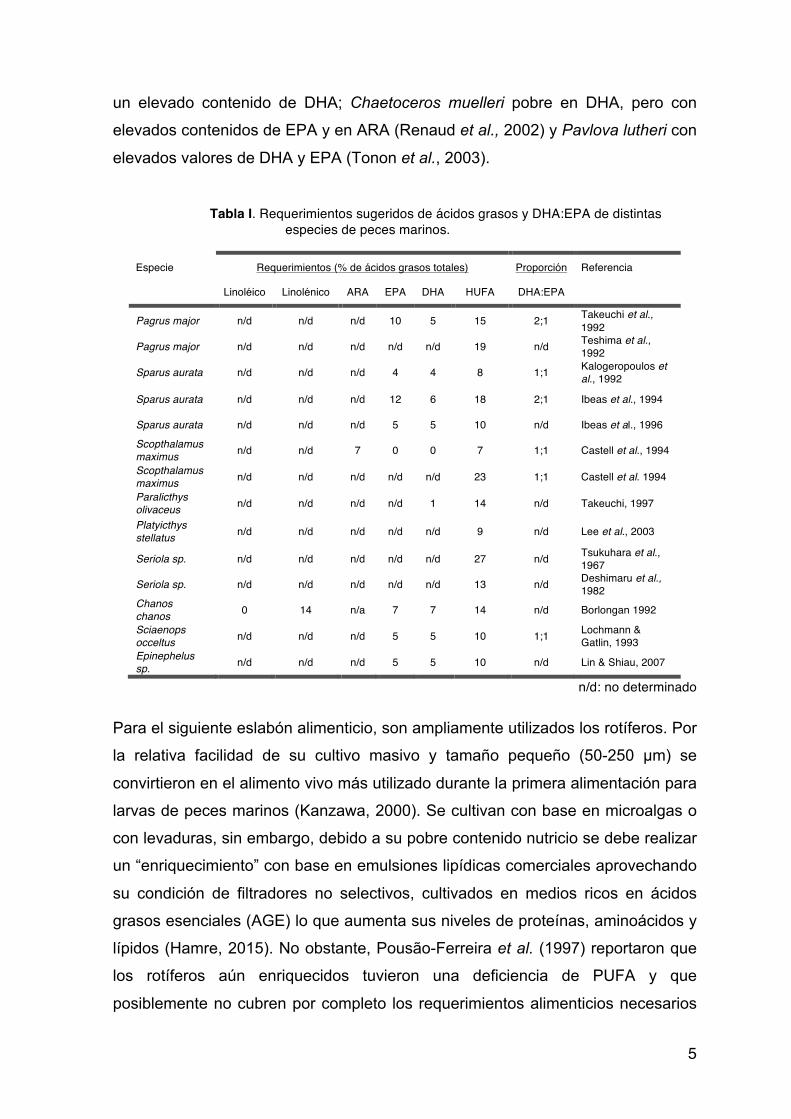
Tabla I. Requerimientos sugeridos de ácidos grasos y DHA:EPA de distintas especies de peces marinos.
Especie Requerimientos (% de ácidos grasos totales) Proporción Referencia Linoléico Linolénico ARA EPA DHA HUFA DHA:EPA
Pagrus major n/d n/d n/d 10 5 15 2;1 Takeuchi et al., 1992
Pagrus major n/d n/d n/d n/d n/d 19 n/d Teshima et al., 1992
Sparus aurata n/d n/d n/d 4 4 8 1;1 Kalogeropoulos et al., 1992
Sparus aurata n/d n/d n/d 12 6 18 2;1 Ibeas et al., 1994 Sparus aurata n/d n/d n/d 5 5 10 n/d Ibeas et al., 1996 Scopthalamus maximus n/d n/d 7 0 0 7 1;1 Castell et al., 1994 Scopthalamus maximus n/d n/d n/d n/d n/d 23 1;1 Castell et al. 1994 Paralicthys olivaceus n/d n/d n/d n/d 1 14 n/d Takeuchi, 1997 Platyicthys stellatus n/d n/d n/d n/d n/d 9 n/d Lee et al., 2003 Seriola sp. n/d n/d n/d n/d n/d 27 n/d Tsukuhara et al.,
1967 Seriola sp. n/d n/d n/d n/d n/d 13 n/d Deshimaru et al.,
1982 Chanos chanos 0 14 n/a 7 7 14 n/d Borlongan 1992 Sciaenops occeltus n/d n/d n/d 5 5 10 1;1 Lochmann &
Gatlin, 1993 Epinephelus sp. n/d n/d n/d 5 5 10 n/d Lin & Shiau, 2007
n/d: no determinado
Para el siguiente eslabón alimenticio, son ampliamente utilizados los rotíferos. Por
la relativa facilidad de su cultivo masivo y tamaño pequeño (50-250 μm) se
convirtieron en el alimento vivo más utilizado durante la primera alimentación para
larvas de peces marinos (Kanzawa, 2000). Se cultivan con base en microalgas o
con levaduras, sin embargo, debido a su pobre contenido nutricio se debe realizar
un “enriquecimiento” con base en emulsiones lipídicas comerciales aprovechando
su condición de filtradores no selectivos, cultivados en medios ricos en ácidos
grasos esenciales (AGE) lo que aumenta sus niveles de proteínas, aminoácidos y
lípidos (Hamre, 2015). No obstante, Pousão-Ferreira et al. (1997) reportaron que
los rotíferos aún enriquecidos tuvieron una deficiencia de PUFA y que
posiblemente no cubren por completo los requerimientos alimenticios necesarios

6
para larvas de la dorada (Sparus aurata). Por su parte, Seychelles et al. (2009)
evaluaron el concentrado, congelado y fresco, de 4 especies de microalgas,
concluyendo que la mezcla congelada de las microalgas Chaetoceros muelleri,
Pavlova lutheri y Nannochloropsis sp. es un enriquecedor adecuado para el
rotífero Brachionus plicatilis (Tabla II).
Tabla II. Contenido de ARA, EPA, DHA y la proporción DHA:EPA reportados en diferentes especies de rotíferos producidos con alimentos diversos.
Especie (% de ácidos grasos totales) Proporción Alimento Referencia ARA EPA DHA DHA:EPA
Brachionus plicatilis 2.90 16 0.3 0.1
Pasta Nannochloropsis oculata
Palmtag et al., 2006
B. plicatilis n/d 0.03 <0.03 1,9 Levadura Rodríguez et al., 1996
B. plicatilis 9.60 14.20 1.8 0.1 Chaetoceros muelleri
Seychelles et al., 2009
B. plicatilis 0.2 0.70 12.8 18.7 Isocrysis galbana Seychelles et al., 2009
B. plicatilis 6.80 27.30 0.2 0 N. oculata Seychelles et al., 2009
B. plicatilis 1.62 7.97 16.5 n/d Protein Selco Plus Can Nhu et al., 2009
B. plicatilis 1.60 2.00 1.0 0.5 Selco® Culture Park et al., 2006
B. plicatilis 0.60 3.80 30.0 1.07 n/d Salhi et al., 1997
Synchaeta cecilia 1 6.8 0.7 0.1 Tetraselmis
suecica Oltra et al., 2000
Brachionus rotundiformis 1.09 0.89 n/d n/d Chlorella Yoshimatsu
et al., 1997 B. rotundiformis <0.05 0.2 0.1 1.9 n/d Rodrìguez et
al., 1997 B. rotundiformis 0.8 6 6.6 1.08 Super fresh C.
vulgaris Hagiwara et al., 2014
Proales similis 5.3 23 n/d 0 N. oculata Hagiwara et al., 2014
P. similis 1 11 17.5 1.59 Super fresh C. vulgaris
Hagiwara et al., 2014
n/d no determinado.
Por otro lado, los copépodos comprenden un gran número de especies, pero son
pocas las que han sido utilizadas en acuacultura. Más específicamente los
calanoideos (Paracalanus, Pseudocalanus, Centropages) y algunos harpacoideos
(Euterpina, Tigriopus y Tisbe) los cuales han sido suministrados como alimento
vivo durante la primera alimentación y se ha reportado que incrementan los
porcentajes de supervivencia y crecimiento de las larvas (Støttrup, 2006). Gran
parte del éxito de su utilización se debe a su calidad nutricional, la cual se

7
caracteriza por altos niveles de proteína (44-52%), un adecuado perfil de
aminoácidos y una composición de ácidos grasos que varia considerablemente
acorde al alimento usado en su cultivo (Støttrup, 2000; Lira, 2002; McKinnon et
al., 2003). Sin embargo, su utilización como alimento presenta limitaciones,
principalmente debido a ciclos de vida largos (comparado con otros alimentos
vivos) y a la problemática que esto genera para su cultivo masivo. Alajmi et al.
(2015) evaluaron al copépodo Parvocalanus crassirostris alimentado con
Isochrysis sp. y reportaron valores más altos de HUFA en poblaciones
domesticadas que en poblaciones naturales.
McKinnon et al. (2003) evaluaron a los copépodos Bestiolina similis, Parvocalanus
crassirostris y Acartia sinjiensis en cuanto a características poblacionales
(producción de huevos, longitud total de organismo, tasa de producción) y en su
perfil de ácidos grasos, concluyendo que el mejor candidato para larvas de peces
es B. similis (Tabla III). Por otra parte, el crustáceo Artemia sp. es el alimento vivo
que por su mayor tamaño (200-500 μm) constituye el nexo del alimento vivo con
el alimento inerte en larvas de peces marinos (Sorgeloos et al., 2001). Además,
su importancia se basa en la practicidad del manejo de sus quistes, que pueden
permanecer mucho tiempo almacenados y listos para su utilización. Sin embargo,
su valor nutricional es bajo y varía según la procedencia de la cepa que se utilice;
por si fuera poco, disminuye a medida que el animal absorbe las reservas
nutricias y realiza su metamorfosis como parte del proceso de crecimiento, hasta
llegar a adulto (Bell et al., 2003).
Es por esta razón que debe enriquecerse con emulsiones lipídicas comerciales.
Malpica-Sanchéz et al. (2003) evaluaron los ácidos grasos de Artemia encontrada
en medio natural, reportando contenidos de ácidos grasos esenciales muy bajos
(EPA 0.2%, ARA 4%, y DHA 5.9%). Watanabe et al. (1983) analizaron los ácidos
grasos de huevos y nauplios de Artemia sp. y mencionan que varía la
composición de ácidos grasos según el origen de la cepa: “Artemia agua dulce”,
con altas concentraciones de ácido linolénico (18:3); y “Artemia marina” con
valores altos de EPA. Rekha et al. (2007) evaluaron nauplios de Artemia sp.
alimentadas con Nannochloropsis salina, Chlorella salina, Chaetoceros calcitrans
y la levadura Saccharomyces cerevisiae, con diferentes tiempos de alimentación
(3, 6, 8 y 24 horas) reportando que con 8 horas de alimentación de la microalga N.

8
salina se producen los más altos valores de ARA, EPA y DHA. Hafezieh et al.
(2009a), reportaron que alimentar a larvas de esturión Persa (Acipenser persicus)
con Artemia sp. enriquecida con elevados niveles de DHA resulta favorable para
su crecimiento y sobrevivencia.
Tabla III Contenido de ARA, EPA, DHA y la proporción DHA:EPA reportados en diferentes especies de copépodos.
Especie (% de ácidos grasos totales) Proporción Alimento Referencia
ARA EPA DHA DHA:EPA
Cyclopina kasignete 5.78 27.50 5.8 0.2 Melosira sp.
Rasdi et al., 2015
C. kasignete 1.36 18.54 2.5 0.13 Schroederiella apiculata, Scenedesmus pectinatus, Tetraedrom minimum, Mesotaenium sp. Desmodesmus sp.
C. kasignete 1.56 25.28 0.5 0.01 Nannochloropsis oculata (deshidratada)
C. kasignete 3.05 25.52 0.5 0.01 N. oculata (Fresca)
C. kasignete 6.02 27.71 8.2 0.3 Tisochrysis lutea
Acartia sinjiensis 0.20 0.6 2.8 4.7 Isochrysis sp, Tetraselmis chuii, Rhodomonas sp. Heterocapsa niei McKinnon et
al., 2003 Parvocalanus crassirostris 0.30 2.8 6.1 2.2 Isochrysis sp, T. chuii,
Rhodomonas sp. H.niei P. crassirostris (domesticado) 0.50 5.40 22.3 4.1 Isochrysis sp. Alajmi et al.,
2015 Temora longicornis n/d 25.50 28.7 1.12 Inanición
Evjemo et al., 2003.
Eurytemora sp. n/d 20.80 39.2 1.88 Inanición
Calanus finmarchicus n/d 17.40 20.8 1.20 Inanición
Pseudodiaptomus euryhalinus 1.35 0.16 1.4 8.75 N. oculata Puello-Cruz et
al., 2015 P. euryhalinus 3 19.44 28.8 1.48 Chaetoceros calcitrans
Flores-Santana, 2008
P. euryhalinus 1.29 9.51 37.1 3.9 Isochrysis galbana
P. euryhalinus 1.81 12 42.0 3.5 C. calcitrans, I. galbana n/d no determinado

9
1.2 Justificación La acuacultura es una actividad de gran importancia como una alternativa para la
obtención de proteína animal, además de ser una vía para prevenir la
sobreexplotación de stocks pesqueros y ser uno de los sectores económicos con
mayor crecimiento. Dentro de los cultivos de peces marinos el principal “cuello de
botella” se debe a la alta mortalidad durante las etapas larvarias; el factor
nutricional del alimento vivo durante la primera alimentación, en particular lo
referente al contenido de ácidos grasos, se presenta como una variable
determinante para incrementar la supervivencia.
Desde 2003, en CICIMAR-IPN en la Unidad Piloto de Maricultivo (UPIMA) se han
implementado esfuerzos para generar conocimiento y aumentar la supervivencia
en larvas de peces marinos Lutjanidos, para ello se ha buscado y experimentado
con diferentes especies de zooplancton para utilizarse como alimento en la etapa
larvaria. Especies como los rotíferos Brachionus plicatilis (Muller 1786),
Brachionus rotundiformis (Tschugunoff, 1921), y Proales similis (De Beauchamp,
1907); los copépodos Pseudodiaptomus euryhalinus (Johnson, 1939) y
Parvocalanus crassirostris (Dahl, 1894), así como el crustáceo Artemia sp; han
sido seleccionados por varios autores como presas potencialmente viables para
larvas de peces marinos (McKinnon et al., 2003; Hagiwara et al., 2014; Alajmi et
al., 2014; Hamre 2015).
Debido a que el contenido de ácidos grasos puede ser modificado por la selección
de las dietas y los procesos de enriquecimiento durante la producción de las
diferentes especies de zooplancton que se utiliza como alimento vivo, la
identificación de dietas que ofrezcan el mejor perfil nutricional en el alimento, es
relevante para aumentar las tasas de supervivencia en larvas. Es por ello que la
presente tesis evalúa el efecto de diferentes protocolos de alimentación en cuanto
al contenido de ácidos grasos utilizados en la producción de alimento vivo y que
han sido suministrados a larvas de diferentes especies de peces marinos en la
trayectoria histórica de CICIMAR-IPN.

10
2. HIPÓTESIS Bajo el marco anterior, resalta una composición diferencial en los ácidos grasos
de las presas utilizadas como alimento vivo durante el cultivo de larvas de peces
marinos, en función del alimento administrado con base en microalgas, una dieta
y/o enriquecedores comerciales. Por lo tanto, se ha planteado la hipótesis de que
existirán diferencias en las proporciones de ácidos grasos de las presas que se
ofrecen a las larvas, alimentadas diferencialmente con microalgas, dietas y/o
enriquecedores comerciales.
3. OBJETIVO GENERAL Determinar el efecto de la dieta en la composición de ácidos grasos de rotíferos
(Brachionus plicatilis, Brachionus rotundiformis y Proales similis), copépodos
(Pseudodiaptomus eurihalynus y Parvocalanus crassirostris) y de Artemia sp. a
través de dietas con base en microalgas, una dieta comercial así cómo el uso de
un enriquecedor comercial.
3.1 Objetivos Particulares
- Determinar el contenido de lípidos totales y de ácidos grasos de las microalgas
Nannochloropsis oculata, Tetraselmis suecica, Chaetoceros calcitrans e
Isochrysis galbana.
- Determinar el contenido de lípidos totales y de ácidos grasos de los rotíferos
Brachionus plicatilis, B. rotundiformis y P. similis alimentados con las
microalgas N. oculata, T. suecica, una dieta comercial y un enriquecedor
comercial.
- Determinar el contenido de lípidos totales y de ácidos grasos en diferentes
estadios de desarrollo de los copépodos Pseudodiaptomus euryhalinus y
Parvocalanus crassirostris alimentado con dietas con base en una microalga
(monoalgales) y con base en la mezcla de microalgas (polialgales).

11
- Determinar el contenido de lípidos totales y de ácidos grasos del nauplio recién
nacido, nauplio alimentado con enriquecedor comercial y juvenil del
branchiopodo Artemia sp.
4. MATERIAL Y MÉTODOS
En el presente trabajo se realizó el cultivo de diferentes organismos empleados
comúnmente como alimento vivo de larvas de peces marinos en CICIMAR-IPN
(rotíferos, copépodos y Artemia). Se evaluaron diferentes tratamientos
alimenticios que consistían ya sea en una sola microalga (dieta monoalgal), una
mezcla de ellas (dieta multialgal) y en el caso de rotíferos también se empleó una
dieta comercial (ORI-ONE®). Adicionalmente, se evaluó el efecto de un
enriquecedor comercial marca Spreso-Selco® utilizado para mejorar la calidad
nutricional en rotíferos y Artemia sp. A continuación se amplía la información
referente a los tratamientos alimentarios y las condiciones de cultivo de cada tipo
de organismo, así como la toma de muestras para la aplicación de las técnicas de
laboratorio para análisis de lípidos totales y ácidos grasos.
4.1 Cultivo de Alimento Vivo Las cepas microalgas, mantenidas en agar inclinado a 21°C y las de rotíferos en
agua marina a 24°C, se obtuvieron del cepario del laboratorio de Biología
Experimental y los cultivos se realizaron en la Unidad Piloto de Maricultivo
(UPIMA) de CICIMAR-IPN. Se utilizó agua de mar (35±1 ups) pasada por filtros
de cartuchos hasta 5 µm, esterilizada con hipoclorito de sodio (NaClO) a una
concentración de 5 ppm durante 24 horas y finalmente neutralizada con tiosulfato
de sodio (Na2S2O3) a una dosis de 5 ppm.
4.1.1 Microalgas Las microalgas utilizadas para este experimento fueron N. oculata, T. suecica, C.
calcitrans e I. galbana. Se cultivaron en un área con temperatura controlada
(24±1ºC) escalando la producción en matraces de 250 y 2000 ml, a bolsas de 50
L, y posteriormente a columnas de 350 L. El medio utilizado para los cultivo fue el
f/2 (Guillard & Ryther, 1962). Todas las microalgas fueron mantenidas con

12
aireación vigorosa. El fotoperiodo se estableció a 16L:8O con una irradiancia
aproximada a 100 µmol m-2s-1 (focos fluorescentes). Los cultivos de microalgas
alcanzaron la fases exponencial y estacionaria antes de ser utilizados para
alimentación de rotíferos, copépodos y Artemia sp. así como para la toma de
muestras para su análisis.
4.1.2 Rotíferos Los rotíferos utilizados como alimento vivo fueron B. plicatilis, B. rotundiformis y
Proales similis. Cada especie fue cultivada en tolvas de 120 L de capacidad con
agua de mar, se inocularon a una densidad de 300 rot ml-1 y contabilizaron
diariamente para manejar densidades máximas de 1000 rot ml-1 al momento de la
cosecha. B. plicatilis y B. rotundiformis se mantuvieron a una temperatura de
27±1ºC y P. similis a 25±1ºC. Los tratamientos monoalgales se suministraron al
inicio del cultivo y cada 48 h se realizó un filtrado, lavado y recambio total del
medio de cultivo, para cosechar una fracción de la población y ser incubados
nuevamente bajo las mismas condiciones. Los tratamientos alimenticios se
detallan en la Tabla IV.
Para el proceso de enriquecimiento con Spresso-Selco®, se cosecharon de las
tolvas en donde fueron cultivados con microalgas y se transfieren a un contenedor
con 18 L de agua de mar con difusor de aire y aireación moderada, a una
densidad de 500 Rot/ml. Se prepararon 7 g de la emulsión Spresso-Selco® en
agua dulce con un triturador y mezclador universal Torrey® (9000 rpm) y se
administró. Se cosecharon 10 horas después, de acuerdo a recomendaciones del
proveedor (350 g m3; máximo de 1,000 rot ml-1; cosecha antes de 12 hrs).
Para la dieta comercial ORI-ONE® se inocularon los rotíferos a una densidad de
500 Rot/ml en una tolva de cultivo con 80 litros de agua de mar. Cada día, se
preparó una mezcla homogénea de la dieta comercial ORI-ONE® con un triturador
y mezclador industrial Torrey®. Se conservó en refrigeración para proporcionar 5
raciones al día con una bomba peristáltica. Se alimentaron así durante 3 días
consecutivos con la siguiente relación: 1er día 0.5 g millón de rotíferos-1; 2do día
0.45 g millón de rotíferos-1; 3er día 0.35 g millón de rotíferos-1. Esto de acuerdo a

13
las instrucciones del proveedor (máximo de 100 g L-1; máximo 2000 rot ml-1 y
cosecha al 4to día).
Tabla IV. Características de los tratamientos alimenticios utilizados para rotíferos.
Especie Tipo de dieta Alimento administrado Densidad
Brachionus plicatilis
& B. rotundiformis
Monoalgal
Nannochloropsis oculata (Nanno) 20 ± 1 x106 cél ml-1 / 48 hrs
Tetraselmis suecica (Tetra) 800 ± 100 x103 cél ml-1 / 48 hrs
Monoalgal y Enriquecedor
Nannochloropsis oculata + Spresso-Selco®
(Nanno - Spresso)
20 ± 1 x106 cél ml-1 / 48 hrs –cosecha-
0.4g Spresso-Selco millónrotíferos-1 / 20hrs
Tetraselmis suecica +
Spresso-Selco® (Tetra+Spresso)
800 ± 100 x103 cél ml-1 / 48 hrs -cosecha-
0.4g Spresso-Selco millónrotíferos-1 / 20hrs
Dieta Comercial ORI-ONE® 0.3-0.5g millónrotíferos-1 / 3 días
Proales similis
Monoalgal
Nannochloropsis oculata (Nanno)
20 ± 1 x106 cél ml-1 /48 hrs
Monoalgal y Enriquecedor Nannochloropsis oculata -
Spresso-Selco® (Nanno-Spresso)
800 ± 100 x103 cél ml-1 /48 hrs –cosecha-
0.4g Spresso-Selco millónrotíferos-1 / 20hrs
Dieta Comercial ORI-ONE® 0.3-0.5g millónrotíferos-1 / 3 días
Cada día se realizó la evaluación del cultivo de rotíferos, para verificar el
crecimiento de la población, la posible contaminación con protozoarios y ajuste de
la ración diaria. Para la cosecha se realizó un pre-filtrado por un tamiz de 100 µm
para eliminar partículas gruesas de materia orgánica. Debido a las diferencias de
tamaño en las especies de rotíferos la recolección de muestras y lavado se realizó
con un tamiz de luz de malla de 50 µm para Brachionus plicatilis y B. rotundiformis
y con un tamiz de luz de malla de 30 µm para P. Similis.

14
4.1.3 Copépodos Para el cultivo de copépodos se estableció un fotoperiodo de 16L:8O con una
aireación reducida, así como una alimentación diaria. Durante el cultivo de
copépodos, para asegurar que las diferencias entre los tratamientos no se
debieran a un cambio drástico en el ambiente experimental y/o alguna situación
de estrés, provocada por el cambio radical de alimentación, las poblaciones de
copépodos fueron sembradas en estadios naupliares y mantenidas a cada
tratamiento hasta alcanzar la madurez reproductiva. Los organismos no fueron
muestreados hasta tener como mínimo 1 ciclo reproductivo en el tratamiento
respectivo. El copépodo Pseudodiaptomus euryhalinus se cultivó a partir de una
población domesticada por más de 100 generaciones, obtenida por medio de
arrastre de zooplanctón en la Bahía de La Paz en julio de 2010, aislada y
eventualmente escalada a niveles de producción. Se cultivaron en tolvas cilindro-
cónicas con capacidad de 140 L a una temperatura de 26±1ºC. El copépodo P.
crassirostris, obtenido de San Diego, California, se cultivó a partir de una
población domesticada durante más de 50 generaciones, en contenedores de
plástico de 1,000 L de capacidad a una temperatura de 27±1ºC.
Los tratamientos alimentarios administrados a cada especie así como los tamaños
de luz de mallas para la separación por estadios de desarrollo los cuales se
detallan en la Tabla V.
Tabla V. Características de los tratamientos utilizados para los copépodos Pseudodiaptomus euryhalinus y Parvocalanus crassirostris.
Especie Tipo de dieta
Alimento administrado Densidad y proporción Luz de malla para toma de muestras
P. euryhalinus
Monoalgal Chaetoceros calcitrans (Cht) 0.8 – 1 x 106 cél ml-1 /72 hrs (1:0)
Nauplio 100 µm Copepodito 180 µm
Adulto 260 µm Multialgal Chaetoceros calcitrans +
Tetraselmis suecica (Cht+Tetra)
0.8 – 1x106 cél ml-1 / 72hrs (9:1)
P. crassirostris
Monoalgal Isochrysis galbana (Iso)
70 – 100 x 103 cél ml-1 (1:0)
Nauplio 40 y 80 µm Copepodito 100µm
Adulto180 µm
Multialgal Isochrysis galbana +
Chaetoceros calcitrans + Tetraselmis suecica
(Iso+Cht+Tetra)
70 – 100 x 103 cél ml-1 (7:2:1)

15
4.1.4 Artemia sp. Se utilizaron quistes de Artemia sp. calidad AAA, pre-tratados en cuanto a la
descapsulación. Se hidrataron 10 g durante 10 minutos en un litro de agua dulce,
se colocaron en una tolva de 120 L de capacidad con agua de mar y aireación
vigorosa. Transcurridas 12 horas se verificó la eclosión de los quistes y se tomó
muestra de los nauplios recién nacidos. Transcurridas 6 horas más, los meta-
nauplios estaban en condiciones para enriquecerlos con Spresso-Selco®, para
esto se cosecharon de las tolvas de eclosión y se colocaron en un contenedor con
18 L de agua de mar, a una densidad de 250 nauplios/ml, se agregaron 14 g de la
emulsión Spresso-Selco® con aireación moderada. Diez horas después de
alimentarse en este medio enriquecedor, se enjuago y tomó la muestra para
análisis de lípidos. Este procedimiento es el recomendado para Artemia sp. por el
proveedor del producto comercial (1,000 g/m3; máximo 300 art ml-1; cosecha
antes de 12 hrs). Cierta cantidad de nauplios fueron sembrados en tanques de
800 L, para alcanzar la fase juvenil, y se alimentaron durante 10 días con una
mezcla en proporciones iguales de cereales (arroz, maíz) y levadura, además de
complementar con mezcla de microalgas C. calcitrans y T. suecica. Las muestras
generadas para Artemia sp. se detallan en el Tabla VI.
Tabla VI. Características de los tratamientos alimenticios utilizados para Artemia sp.
Especie Tipo de alimentación
Alimento administrado Estadio de Desarrollo Luz de malla para toma de muestras
Artemia sp.
Inanición - sin alimento- Nauplio recién nacido
100 µm
Enriquecida Spresso-Selco® Meta - Nauplio II-III
Mezcla de cereales y microalgas
Harina de arroz + harina de maíz + levadura
(1:1:1) Chaetoceros calcitrans +
Tetraselmis suecica (1:1)
Juvenil 260 µm

16
Muestreo De cada uno de los tratamientos evaluados, así como de las microalgas
empleadas como alimento, se obtuvieron aproximadamente 100 mg de muestra.
En el caso partocula de las microalgas, fue necesario utilizar centrífugas
refrigeradas para concetrar las células. Se colocaron en viales de 6 ml de solución
de cloroformo:metanol (2:1). A cada vial se le agregó 10 μl debutilhidroxitoluena
(BHT)(0.5 mg ml–1; B–1378, Sigma, St. Louis, MO, EE.UU) y 10 μl del ácido
graso 23:0 (2 mg ml–1; T–6543, Sigma, St. Louis, MO, EE.UU) como estándar
interno. Se almacenaron a -20ºC para su posterior análisis.
4.2 Determinación de lípidos totales La determinación de lípidos totales se realizó siguiendo el método gravimétrico
como se describe en Palacios y Arjona (2009) en el Centro de Investigación
Biológica del Noreste (CIBNOR), en el laboratorio de Metabolismo de Lípidos. Se
utilizaron tubos de borosilicato de 10 ml y se colocaron en un horno de
temperatura controlada a 80°C por 24 horas (como mínimo), hasta peso
constante. Después, los tubos se colocaron en un desecador, se dejaron enfriar
por 1 hora y se registró el peso de cada uno de ellos. Se agregaron 2 ml del
extracto lipídico y se evaporó a sequedad con gas nitrógeno. Se colocaron los
tubos nuevamente en el horno de temperatura controlada a 30°C por 24 horas y
se pesaron los tubos con el extracto lipídico, y se hicieron los cálculos
correspondientes para sacar el porcentaje de lípidos totales en referencia al peso
total.
4.3 Análisis de ácidos grasos Las muestras se disgregaron usando un homogenizador de vidrio y se sonicaron
por 10 minutos en un baño frío. Se tomaron 2 ml de los extractos que fueron
colocados en cloroformo:metanol (2:1) se evaporaron a aproximadamente 0.5 ml
con nitrógeno gaseoso (N2), posteriormente fueron colocados en microcolumnas
preparadas con pipetas Pasteur rellenas de fibra de vidrio y sílice hidratado con
agua al 6%. Los lípidos fueron separados eluyendo en las microcolumnas 10 ml
de cloroformo:metanol (98:2). Las fracciones obtenidas de cada muestra fueron
evaporadas a sequedad y derivatizadas con trifloruro de boro metanol (BF3;
Supelco, Bellefonte, PA, EE.UU.) por 15 minutos a 95°C. La extracción de los

17
metil-ésteres de los ácidos grasos se realizó con hexano y lavados de agua. Los
metil-ésteres fueron analizados en un cromatógrafo de gases Hewllet-Packard®
(CG 6890–N) equipado con una columna capilar de sílice fundida DB–23 (J & W
Scientific, Folsom, CA, E.U.A) (30 m de largo × 0.25 mm de diámetro interno ×
0.25 μm de espesor de película) y un detector de ionización de flama (FID) a una
temperatura de 280°C. La rampa de temperatura del horno para la separación de
los ácidos grasos fue de 110–220°C a una tasa de incremento de 3°C min–1. Se
utilizó como gas acarreador helio. La identificación de los metil-ésteres de los
ácidos grasos se realizó comparando los tiempos de retención de una mezcla de
ácidos grasos como estándar (47885–U, Supelco, Bellefonte, PA, EE.UU.) y la
cuantificación usando el ácido graso 23:0.
5. RESULTADOS Lípidos Totales La microalga Isochrysis galbana presentó la mayor proporción de lípidos con
9.66%, mientras que N. oculata fue la microalga con menor proporción de lípidos
totales 6.36% (Figura 1).
En el caso de los rotíferos, las mayores proporciones de lípidos totales se
presentaron en B. plicatilis; y en esta especie, la mayor proporción de lípidos
totales se encontró con el tratamiento alimenticio Nanno+Spresso-Selco® (5.90%).
0
2.5
5
7.5
10
I. galbana T. suecica N. oculata
Líp
ido
s to
tale
s (%
)
Figura 1. Proporción de lípidos totales encontrados en las diferentes microalgas evaluadas.

18
B. rotundiformis fue la especie que menor concentración de lípidos totales
presentó, sin importar el tratamiento alimenticio, sin embargo no se observa
mucha diferencia entre los otros tratamientos (Figura 2). P. similis, debido a su
tamaño no puede ser alimentado con T. suecica. Y durante no fue posible obtener
la cuantificación de lípidos totales la muestra de B. plicatilis alimentada con T.
suecica, por problemas durante su manejo.
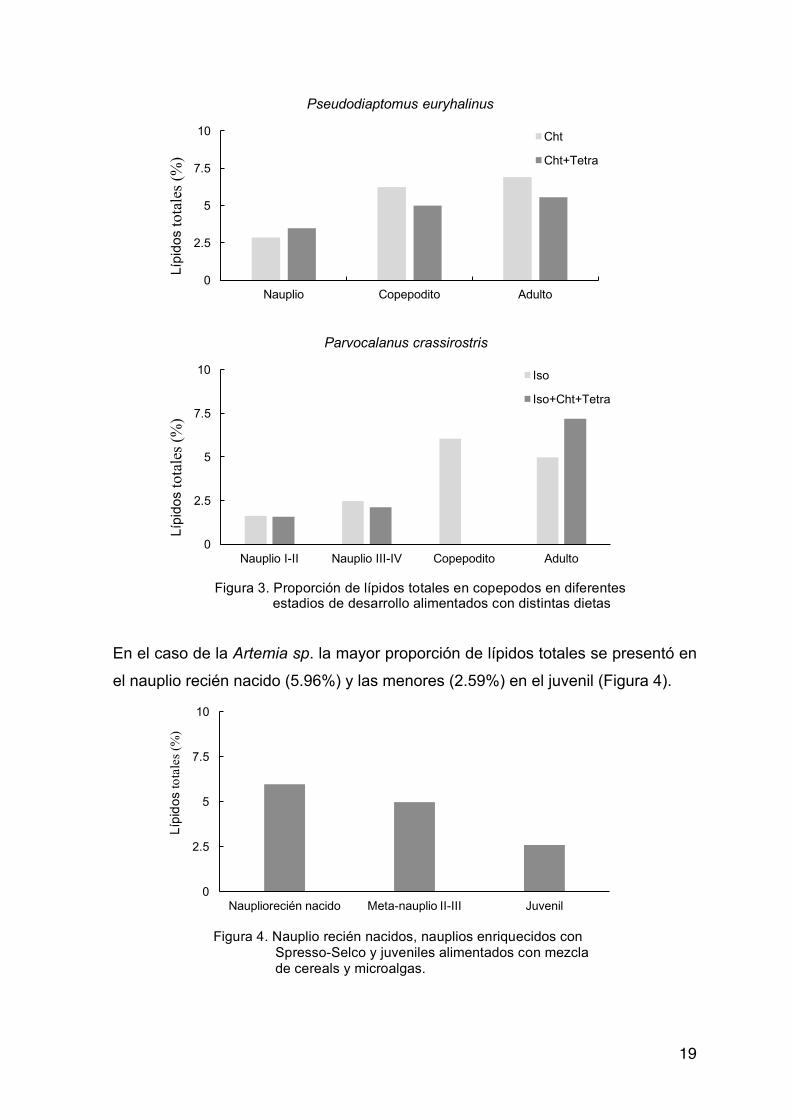
En copépodos, tanto en P. euryhalinus como en P. crassirostris los estadios
copépoditos y adultos presentaron mayores proporciones de lípidos totales que
los estadios naupliares. En ambas especies, los porcentajes de lípidos totales
registrados para estadio nauplio fueron los menores. (Figura 3). Para P.
euryhalinus la dieta monoalgal con base en C. calcitrans generó una mayor
concentración de lípidos totales (excepto en nauplio). Para P. crassirostris la dieta
de mezcla de microalgas obtuvo mayores lípidos totales en adultos. Copepodito
alimentado con mezcla de microalgas no pude ser cuantificado.
Figura 2. Lípidos totales en rotíferos, alimentados con diferentes dietas.
Líp
ido
s to
tale
s (%
)
0
2.5
5
7.5
10
N. oculata Nanno +�Spresso-
Selco®
T. suecica Tetra+�Spresso-Selco®
ORI-ONE®
Brachionus plicatilis
Brachionus rotundiformis
Proales similis
19
En el caso de la Artemia sp. la mayor proporción de lípidos totales se presentó en
el nauplio recién nacido (5.96%) y las menores (2.59%) en el juvenil (Figura 4).
0
2.5
5
7.5
10
Nauplio Copepodito Adulto
Pseudodiaptomus euryhalinus
Cht
Cht+Tetra
0
2.5
5
7.5
10
Nauplio I-II Nauplio III-IV Copepodito Adulto
Parvocalanus crassirostris
Iso
Iso+Cht+Tetra
Figura 3. Proporción de lípidos totales en copepodos en diferentes estadios de desarrollo alimentados con distintas dietas
Líp
ido
s to
tale
s (%
) L
ípid
os
tota
les
(%)
0
2.5
5
7.5
10
Naupliorecién nacido Meta-nauplio II-III Juvenil
Figura 4. Nauplio recién nacidos, nauplios enriquecidos con Spresso-Selco y juveniles alimentados con mezcla de cereals y microalgas.
Líp
ido
s to
tale
s (%
)

20
CONTENIDO DE ÁCIDOS GRASOS TOTALES. Microalgas C. calcitrans presentó la mayor proporción de ácidos grasos saturados (SFA) y
ácidos grasos monoinsaturados (MUFA). Respecto a los ácidos poliinsaturados
(PUFA), I. galbana y T. suecica presentaron las mayores proporciones (65 y 61%,
respectivamente). I. galbana y T. suecica presentaron mejores proporciones de
Omega-3, y N. oculata de omega-6 (Tabla VII).
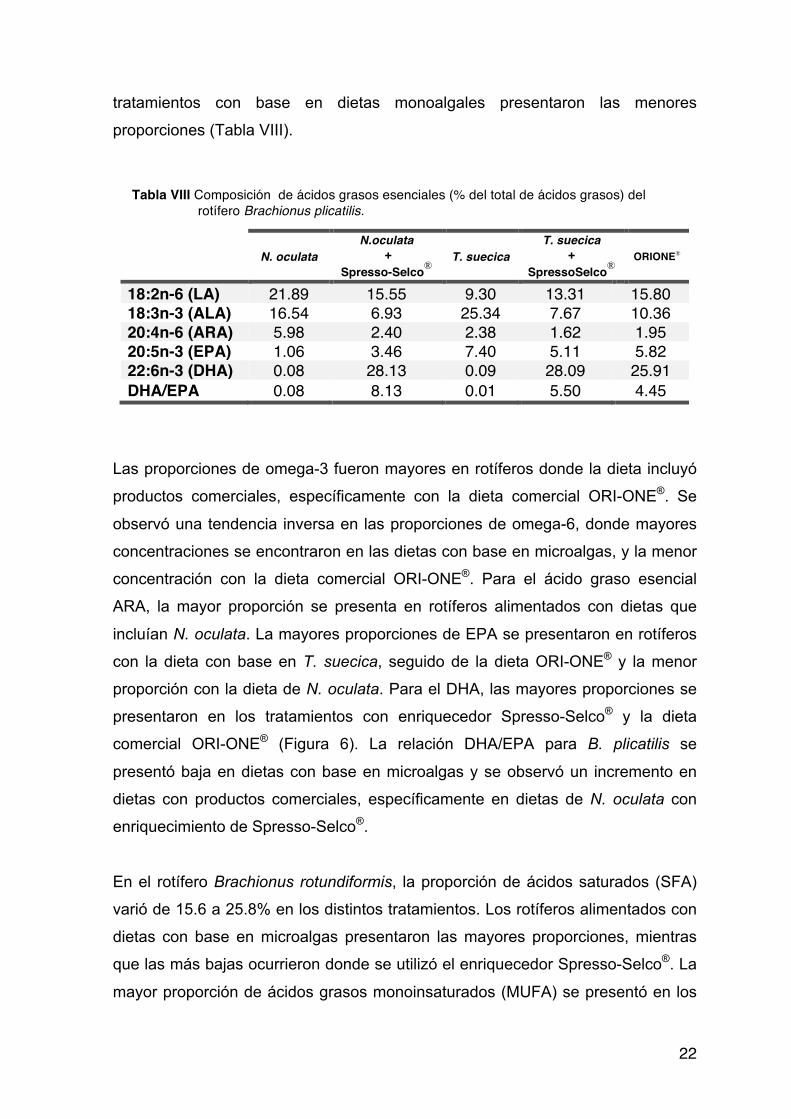
Las diferentes microalgas mostraron también diferencias en cuanto al contenido
de ácido linoleico (LA, 18:2n-6) la mayor proporción se encontró en N. oculata. En
cuanto al ácido linolénico (ALA, 18:3n-3) T. suecica presentó la mayor proporción.
Los ácidos grasos predominantes en C. calcitrans fueron 14:0, 16:0, 16:1n-7 y
20:5n-3 (EPA) los cuales componen un 77.6% del total de ácidos grasos
encontrados. Esta microalga presentó bajos porcentajes de ARA (3.38%) y DHA
(4.41%).
En I. galbana los ácidos grasos predominantes fueron 14:0,18:1n-9, 18:2n-6 (LA),
18:3n-3 (ALA), 18:4n-3 y 22:6n-3 (DHA) que constituyeron el 80.2% del total de
los ácidos grasos. Esta microalga presentó la mayor cantidad de DHA. Por el
contrario, las concentraciones de ARA y EPA fueron muy bajas. (Figura 5).
Tabla VII.Composición de los ácidos grasos esenciales de las microalgas (%del total de AG)
Nannochloropsis oculata
Tetraselmis suecica
Chaetoceros calcitrans
Isochrysis galbana
18:2n-6 (LA) 25.83 6.25 0.54 9.43 18:3n-3 (ALA) 20.14 33.35 3.58 16.15 20:4n-6 (ARA) 4.85 1.48 3.38 0.10 20:5n-3 (EPA) 1.06 9.42 21.82 0.79 22:6n-3 (DHA) 0.02 0.09 4.41 19.42 DHA/EPA 0.02 0.01 0.20 24.68
En T. suecica los ácidos grasos 16:0, 18:1n-9, 18:3n-3 (ALA), 18:4n-3 y 20:5n-3
(EPA), sumaron el 75.05% de todos los ácidos grasos mientras que el 22:6n-3
(DHA) presentó un porcentaje muy bajo. Por último, N. oculata presentó la menor

21
proporción de ácidos grasos altamente insaturados (HUFA) con porcentajes de
DHA y EPA muy bajos (Figura 5). Los ácidos grasos con mayor porcentaje en
esta microalga fueron 16:0, 18:1n-9, 18:2n-6 (LA), 18:3n-3 (ALA) representando el
79.06% del total.
La relación DHA/EPA presente en las 4 distintas microalgas fueron reducidas
desde 0.01% para T. suecica, 0.02% para N. oculata y 0.2 para C. calcitrans
hasta el valor máximo determinado en 24.6% para I.galbana.
Rotíferos La evaluación de ácidos grasos del rotífero Brachionus plicatilis, arrojó las
mayores proporciones de ácidos grasos saturados (SFA) y monoinsaturados
(MUFA) en las dietas monoalgales N. oculata y T. suecica; y los menores
porcentajes de SFA y de MUFA en los tratamientos que incluían productos
comerciales. En cuanto a ácidos grasos poliinsaturados (PUFA), los rotíferos
cultivados con enriquecedor Spresso-Selco® y dieta comercial ORI-ONE® fueron
los que mayores proporciones presentaron con porcentajes similares. Los
Figura 5. Proporciones de los ácidos grasos esenciales en microalgas
% d
el t
ota
l de
áci
do
s g
raso
s
0
5
10
15
20
25
30
N. oculata T. suecica C. calcitrans I. galbana
20:4n-6 (ARA)
20:5n-3(EPA)
22:6n-3 (DHA)
22
tratamientos con base en dietas monoalgales presentaron las menores
proporciones (Tabla VIII).
Tabla VIII Composición de ácidos grasos esenciales (% del total de ácidos grasos) del rotífero Brachionus plicatilis.
N. oculata N.oculata
+ Spresso-Selco®
T. suecica T. suecica
+ SpressoSelco®
ORIONE®
18:2n-6 (LA) 21.89 15.55 9.30 13.31 15.80 18:3n-3 (ALA) 16.54 6.93 25.34 7.67 10.36 20:4n-6 (ARA) 5.98 2.40 2.38 1.62 1.95 20:5n-3 (EPA) 1.06 3.46 7.40 5.11 5.82 22:6n-3 (DHA) 0.08 28.13 0.09 28.09 25.91 DHA/EPA 0.08 8.13 0.01 5.50 4.45
Las proporciones de omega-3 fueron mayores en rotíferos donde la dieta incluyó
productos comerciales, específicamente con la dieta comercial ORI-ONE®. Se
observó una tendencia inversa en las proporciones de omega-6, donde mayores
concentraciones se encontraron en las dietas con base en microalgas, y la menor
concentración con la dieta comercial ORI-ONE®. Para el ácido graso esencial
ARA, la mayor proporción se presenta en rotíferos alimentados con dietas que
incluían N. oculata. La mayores proporciones de EPA se presentaron en rotíferos
con la dieta con base en T. suecica, seguido de la dieta ORI-ONE® y la menor
proporción con la dieta de N. oculata. Para el DHA, las mayores proporciones se
presentaron en los tratamientos con enriquecedor Spresso-Selco® y la dieta
comercial ORI-ONE® (Figura 6). La relación DHA/EPA para B. plicatilis se
presentó baja en dietas con base en microalgas y se observó un incremento en
dietas con productos comerciales, específicamente en dietas de N. oculata con
enriquecimiento de Spresso-Selco®.
En el rotífero Brachionus rotundiformis, la proporción de ácidos saturados (SFA)
varió de 15.6 a 25.8% en los distintos tratamientos. Los rotíferos alimentados con
dietas con base en microalgas presentaron las mayores proporciones, mientras
que las más bajas ocurrieron donde se utilizó el enriquecedor Spresso-Selco®. La
mayor proporción de ácidos grasos monoinsaturados (MUFA) se presentó en los

23
rotíferos alimentados con la dieta de microalgas con base en T. suecica, los
valores más bajos se registraron en las dietas donde se utilizó el enriquecedor
Spresso-Selco®.
En ácidos grasos poliinsaturados (PUFA) la relación se invierte, y son los
tratamientos con Spresso-Selco® y dieta comercial ORI-ONE® los que mayores
proporciones presentan (Tabla IX). En relación a Omega-3, las más bajas
proporciones se encontraron en las dietas con base en la microalga N. oculata;
mientras que la dieta de N. oculata enriquecida con Spresso-Selco® y la dieta
comercial ORI-ONE® presentaron las mayores. Las dietas de microalgas
presentaron valores de DHA muy bajos, lo contrario ocurre en tratamientos con
Spresso-Selco® donde las mayores proporciones de DHA se encontraron (Figura
7). En cuanto a Omega-6, los mayores porcentajes se registraron en los rotíferos
en los que la dieta incluyó a N. oculata, y las menores proporciones se detectaron
en las dietas que incluían a T. suecica (T. suecica, T. suecica + Spresso y ORI-
ONE®).
0
5
10
15
20
25
30
N. oculata N. oculata
+Spresso-Selco®
T. suecica T. suecica
+Spresso-Selco ®
ORI-ONE®
20:4n-6(ARA) 20:5n-3(EPA) 22:6n-3 (DHA)
Figura 6. Proporciones de los ácidos grasos esenciales en Brachionus plicatilis alimentados con distintos tratamientos alimenticios.
% d
el t
ota
l de
áci
do
s g
raso
s

24
Tabla IX Proporciones de ácidos grasos esenciales (% del total de ácidos grasos totales) en Brachionus rotundiformis.
N. oculata N. oculata
+ Spresso-Selco®
T. suecica T. suecica
+ Spresso-Selco®
ORI-ONE®
18:2n-6 (LA) 26.10 16.31 4.56 10.98 14.19 18:3n-3 (ALA) 13.93 7.99 22.00 12.65 5.98 20:4n-6 (ARA) 5.11 2.76 1.81 1.39 3.71 20:5n-3 (EPA) 0.83 3.12 7.88 5.42 6.93 22:6n-3 (DHA) 0.03 28.66 0.25 23.05 20.92 DHA/EPA 0.04 9.19 0.03 4.25 3.02
Para el AGE araquidónico (ARA), las dietas con enriquecedor Spresso-Selco®
presentaron menores proporciones que sus dietas de microalgas respectivas; sin
embargo, en este caso, el tratamiento con mayores proporciones se presentó en
rotíferos alimentados con N. oculata; por el contrario, las mayores proporciones
de EPA se presentaron en rotíferos alimentados con la dieta T. suecica, mientras
que las menores proporciones en rotíferos cultivos con la dieta N. oculata (Figura
7).
0
5
10
15
20
25
30
N. oculata N.
oculata+�Spresso-
Selco®
T. suecica T.
suecica+�Spresso-
selco®
ORI-ONE®
20:4n-6 (ARA) 20:5n-3 (EPA) 22:6n-3 (DHA)
Figura 7. Proporciones de los ácidos grasos esenciales en Brachionus
rotundiformis alimentados con distintos tratamientos alimenticios.
% d
el t
ota
l de
áci
do
s g
raso
s

25
La relación DHA/EPA para B. rotundiformis fue baja en dietas con base en
microalgas (T. suecica y N. oculata; 0.03 y 0.04, respectivamente) e incrementó
en dietas qué contienen productos comerciales (Spresso-Selco® y ORI-ONE®);
específicamente la relación más alta se presentó en rotíferos alimentados con N.
Oculata y enriquecidos con Spresso-Selco® (9.19).
El mayor contenido de ácidos grasos saturados (SFA) en P. similis se presentó en
el tratamiento con N. oculata mientras que el tratamiento con N.oculata y el
enriquecedor Spresso-Selco® presentó la proporción más baja. Los ácidos grasos
monoinsaturados (MUFA) se encontraron con mayor proporción en los rotíferos
cultivados con la dieta comercial ORI-ONE® y los que menor proporción
presentaron fue con el tratamiento de N. oculata + Spresso-Selco®. Se observó
una mayor proporción de los ácidos poliiinsaturados (PUFA) en rotíferos
enriquecidos con Spresso-Selco®, y una menor proporción con la dieta comercial
ORI-ONE®. En ácidos grasos altamente poliinsaturados (HUFA) y en las
proporciones de Omega-3 se observó una mayor proporción en los productos
comerciales, especialmente con Spresso-Selco®. El contenido de ácidos grasos
esenciales se muestra en la Tabla X.
Tabla X Composición de ácidos grasos escenciales (% del total de ácidos grasos) del rotífero Proales similis
N. oculata N. oculata
+ Spresso-Selco®
ORI-ONE®
18:2n-6 (LA) 33.96 23.00 16.33 18:3n-3 (ALA) 15.21 6.28 6.33 20:4n-6 (ARA) 5.15 3.20 2.64 20:5n-3 (EPA) 0.59 2.24 3.95 22:6n-3 (DHA) 0.88 24.79 19.16 DHA/EPA 1.48 11.05 4.85
n/d: No determinado.

26
Las mayores proporciones de EPA se encontraron en rotíferos alimentados con la
dieta comercial ORI-ONE®. Los rotíferos enriquecidos con Spresso-Selco®
presentaron los mayores valores de DHA, al contrario, la dieta de N. oculata arrojó
valores cercanos a 1%, así como las proporciones más bajas de EPA (Figura 8),
sin embargo, esta dieta presentó las mayores proporciones en los AGE linoleico
(18:2n-6, LA), linolénico (18:3n-3, ALA) y araquidónico (20:4n-6, ARA).
La relación DHA/EPA para P. similis varió desde 1.48 hasta 11.05. Con relaciones
más altas en dietas que incluían productos comerciales, particularmente, la mayor
relación se obtuvo con el enriquecedor Spresso-Selco®
Copépodos El contenido de ácidos grasos saturados (SFA) fue muy similar en P. euryhalinus
para todos los tratamientos y estadios, siendo ligeramente más bajo en nauplios,
copépoditos y adultos alimentados con la dieta multialgal de C. calcitrans y T.
0
5
10
15
20
25
30
N. oculata N. oculata�+Spresso-Selco® ORI-ONE®
20:4n-6 (ARA) 20:5n-3 (EPA) 22:6n-3 (DHA)
Figura 8. Proporciones de los ácidos grasos esenciales en Proales similis alimentados con distintos tratamientos alimenticios.
% d
el t
ota
l de
áci
do
s g
raso
s

27
suecica. Los valores detallados de los ácidos grasos se muestran en la Tabla XI.
El porcentaje de ácidos grasos monoinsaturados (MUFA) en los diferentes
tratamientos varió de 16.87 a 28.33%.
Los estadios naupliares y copepodito alimentados con la dieta monoalgal (C.
calcitrans) presentaron porcentajes ligeramente mayores ocurriendo lo contrario
en el caso de los adultos, pero con un mínimo de diferencia (0.33%). En relación a
los ácidos poliinsaturados (PUFA), y altamente insaturados (HUFA), la mayor
proporcion se encontraron en los nauplios alimentados con dieta multialgal; sin
embargo, en la Figura 9, se observa que no hay diferencia en EPA y ARA entre
las dietas para los tres estadios, únicamente en DHA para los nauplios
alimentados con la dieta multialgal.
Tabla XI. Composición de ácidos grasos esenciales (% del total de ácidos grasos) del copépodo Pseudodiaptomus euryhalinus, en diferentes estadios de desarrollo, alimentados con C. calcitrans (Cht) y la mezcla de C.calcitrans y T. suecica (Tetra).
Nauplio Copepodito Adulto
Cht Cht+ Tetra
Cht Cht +
Tetra
Cht Cht + Tetra
18:2n-6 (LA) 0.36 0.72 0.68 1.22 0.47 1.11 18:3n-3 (ALA) 1.27 0.82 1.56 4.72 1.77 4.51 20:4n-6 (ARA) 2.18 4.06 2.83 2.86 2.91 3.33 20:5n-3 (EPA) 20.41 0.00 19.73 19.93 24.28 23.92 22:6n-3 (DHA) 22.46 47.86 14.58 17.17 16.77 12.73 DHA/EPA 1.10 2.00 0.74 0.86 0.69 0.53
La relación DHA/EPA para P. euryhalinus fluctuó desde 0.53 hasta 2%, con los
estadios naupliares con valores mayores de esta proporción.
Por otro lado, el contenido de ácidos grasos del copépodo P. crassirostris. Los
ácidos grasos con mayor proporción sin importar el tipo de dieta (monoalgal y
multialgal) y en todos los estadios fueron: 16:0, 18:2n-6 (LA), 18:3n-3 (ALA) y
22:6n-3 (DHA). En la Tabla XII se muestran los porcentajes del total de ácidos
grasos esenciales.

28
0
10
20
30
40
50
Nauplio Copepodito Adulto
Cht
Cht+Tetra
20:5n-3 (EPA)
% d
el t
ota
l de
áci
do
s g
raso
s
Figura 9. Proporción de ácidos grasos esenciales en estadios de desarrollo del copépodo Pseudodiaptomus euryhalinus
alimentados con Chaetoceros calcitrans (Cht) y la mezcla
de C.calcitrans y Tetraselmis suecica (Cht+Tetra).
0
10
20
30
40
50
Nauplio Copepodito Adulto
Cht
Cht+Tetra
22:6n-3 (DHA)
% d
el t
ota
l de
áci
do
s g
raso
s
0
10
20
30
40
50
Nauplio Copepodito Adulto
Cht
Cht+Tetra
20:4n-6 (ARA)
% d
el t
ota
l de
áci
do
s g
raso
s g
raso
s

29
Los ácidos grasos saturados (SFA) variaron de 17.8 a 27.9%. El estadio
copepodito presentó las mayores proporciones con ambas dietas administradas,
seguido de nauplio I-II, adulto y nauplio III-IV. En cuanto a las concentraciones de
ácidos grasos monoinsaturados (MUFA), en general, la dieta multialgal generó
proporciones ligeramente mayores en todos los estadios de desarrollo; que
fluctuaron de 13.85% a 21.35%
Tabla XII. Composición de ácidos grasos esenciales (% del total de ácidos grasos) del copépodo Parvocalanus crassirostris, en diferentes estadios de desarrollo, alimentados con I. galbana (Iso) y la mezcla de I. galbana, C. calcitrans (Cht) y T. suecica (Tetra).
Nauplio I-II Nauplio III-IV Copepodito Adulto
Iso
Iso + Tetra+
Cht
Iso
Iso + Tetra+
Cht
Iso
Iso + Tetra+
Cht
Iso
Iso + Tetra +
Cht
18:2n-6 (LA) 8.30 4.97 9.30 9.62 7.67 5.91 8.56 6.81 18:3n-3 (ALA) 7.67 10.65 8.33 10.88 9.31 7.41 8.45 12.20 20:4n-6 (ARA) 0.19 1.43 0.63 0.38 0.21 1.19 0.29 0.87 20:5n-3 (EPA) 2.37 12.62 2.56 4.39 1.81 4.34 2.72 7.76 22:6n-3 (DHA) 37.17 24.70 36.15 30.93 23.20 17.42 32.44 20.38 DHA/EPA 15.71 1.96 14.12 7.05 12.83 4.01 11.95 2.63
Los ácidos grasos poliinsaturados (PUFA) con ligeramente mayor proporción los
presentaron, en general, los copépodos alimentados con Isochrysis galbana, el
mayor porcentaje lo presentaron los nauplios I-II y III-IV. Las menores
concentraciones se presentaron en el estadio copepodito alimentados con dieta
multialgal conformada por la mezcla de I. galbana, C. calcitrans y T. suecica
(Tabla XII). Las mayores proporciones de ácidos grasos omega-3 se presentaron
en nauplios I-II y III-IV. En general, la dieta monoalgal de I. galbana provocó
mayores valores de omega-3 y omega-6, además, los copépodos alimentados
con ésta dieta presentaron mayor proporción de 22:6n-3 (DHA), en todos los
estadios de desarrollo. Para los ácidos grasos 20:5n-3 (EPA) y 20:4n-6 (ARA) se
invierte la relación, la dieta multialgal arrojó mayores proporciones para todos los
estadios, donde la mayor proporción se presentó en el estadio nauplio I-II y,
particularmente, en EPA (Figura 10).

30
0
10
20
30
40
50
Nauplio I-II Nauplio III-IV Copepodito Adulto
Iso
Iso+Cht+Tetra
20:5n-3 (EPA)
% d
el t
ota
l de
áci
do
s g
raso
s
0
10
20
30
40
50
Nauplio I-II Nauplio III-IV Copepodito Adulto
Iso
Iso+Cht+ Tetra
22:6n-3 (DHA)
% d
el t
ota
l de
áci
do
s g
raso
s
0
10
20
30
40
50
Nauplio I-II Nauplio III-IV Copepodito Adulto
Iso
Iso+Cht+Tetra
20:4n-6 (ARA)
% d
el t
ota
l de
áci
do
s g
raso
s g
raso
s
Figura 10. Proporción de ácidos grasos esenciales en estadios de desarrollo del copépodo Parvocalanus crassirostris alimentados con I. galbana (Iso) y la mezcla de I. galbana, C. calcitrans y
Tetraselmis suecica (Iso+Cht+Tetra).

31
P. crassirostris presentó proporciones de DHA/EPA que van de 1.96 a 15.7; se
observa que las dietas que incluyen la microalga I. galbana aumentan ésta
relación y se observa la tendencia a disminuir conforme se desarrolla el
copépodo.
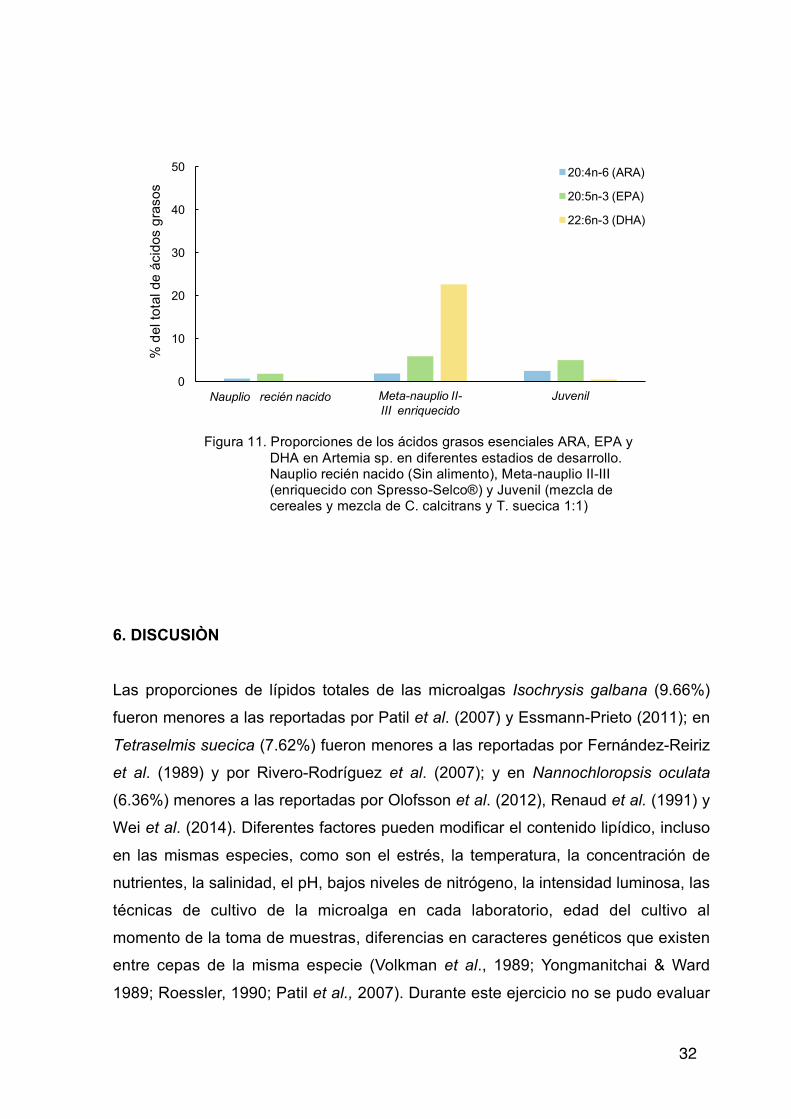
Artemia Los ácidos grasos saturados (SFA) y monoinsaturados (MUFAS) se presentaron
en mayores proporciones en el estadio juvenil. Por el contrario, nauplios
enriquecidos con Spresso-Selco®, presentaron las menores. Sin embargo, en
relación a las concentraciones de ácidos poliinsaturaros (PUFA), omega-3 y
omega-6 las mayores proporciones estuvieron en nauplios enriquecidos con
Spresso-Selco®, y las menores en juveniles. Los resultados de ácidos grasos
esenciales se muestran en la Tabla XIII. Los ácidos grasos ARA, EPA y DHA en
nauplios recién nacidos presentaron las proporciones más bajas, los nauplios
enriquecidos las mayores proporciones de EPA y DHA y los juveniles presentaron
el mayor porcentaje de ARA (Figura 11). La relación DHA/EPA para Artemia sp.
fue muy baja (0.09) sin variar en los diferentes niveles de desarrollo (nauplio o
juvenil) cuando no se administró el enriquecedor Spresso-Selco® que presentó
una relación de 3.85.
Tabla XIII. Composición de ácidos grasos esenciales (% del total de ácidos grasos) de Artemia sp.
Nauplio recién nacido
Meta-nauplio II-III enriquecido Juvenil
18:2n-6 (LA) 5.98 9.35 9.38 18:3n-3 (ALA) 36.07 14.56 4.78 20:4n-6 (ARA) 0.67 1.84 2.47 20:5n-3 (EPA) 1.78 5.87 5.00 22:6n-3 (DHA) 0.15 22.59 0.43 DHA/EPA 0.08 3.85 0.09
32
6. DISCUSIÒN
Las proporciones de lípidos totales de las microalgas Isochrysis galbana (9.66%)
fueron menores a las reportadas por Patil et al. (2007) y Essmann-Prieto (2011); en
Tetraselmis suecica (7.62%) fueron menores a las reportadas por Fernández-Reiriz
et al. (1989) y por Rivero-Rodríguez et al. (2007); y en Nannochloropsis oculata
(6.36%) menores a las reportadas por Olofsson et al. (2012), Renaud et al. (1991) y
Wei et al. (2014). Diferentes factores pueden modificar el contenido lipídico, incluso
en las mismas especies, como son el estrés, la temperatura, la concentración de
nutrientes, la salinidad, el pH, bajos niveles de nitrógeno, la intensidad luminosa, las
técnicas de cultivo de la microalga en cada laboratorio, edad del cultivo al
momento de la toma de muestras, diferencias en caracteres genéticos que existen
entre cepas de la misma especie (Volkman et al., 1989; Yongmanitchai & Ward
1989; Roessler, 1990; Patil et al., 2007). Durante este ejercicio no se pudo evaluar
0
10
20
30
40
50
Nauplio �recién nacido Meta-nauplio II-
III�enriquecido
Juvenil
20:4n-6 (ARA)
20:5n-3 (EPA)
22:6n-3 (DHA)
Figura 11. Proporciones de los ácidos grasos esenciales ARA, EPA y DHA en Artemia sp. en diferentes estadios de desarrollo. Nauplio recién nacido (Sin alimento), Meta-nauplio II-III (enriquecido con Spresso-Selco®) y Juvenil (mezcla de cereales y mezcla de C. calcitrans y T. suecica 1:1)
alimentados con distintos tratamientos alimenticios.
% d
el t
ota
l de
áci
do
s g
raso
s

33
el contenido de lípidos totales para la muestra de C. calcitrans sin embargo
Fernández-Reiriz et al. (1989) y Ohse et al. (2014) reportan proporciones de
lípidos totales de 9 al 12% para esta microalga.
I. galbana presentó mayores proporciones en el ácido graso esencial (AGE)
docosahexaenoico (22:6n-3,DHA), lo cual concuerda con lo reportado por diversos
autores (Volksman et al., 1989; Fidalgo et al., 1998; Rivero-Rodríguez et al., 2007;
Navarro-Hurtado, 2014). T. suecica aporta principalmente los AGE linolénico
(28:3n-3, ALA) y en menor medida eicosahexaenoico (20:5n-3, EPA), esto es
similar a resultados previamente reportados (Volkman et al., 1989; Delaporte et al.,
2005; Rodríguez et al., 2007; Bondioli et al., 2012). Por otra parte, C. calcitrans tuvo
altas proporciones en EPA tal como reportado por (Volkman et al., 1989;
Napolitano et al.,1990; Rivero-Rodríguez et al., 2007) y finalmente N. oculata
destacó únicamente en los AGE linoleico (18:2n-6, LA), ALA y en menor proporción
ARA. Esto concuerda con lo reportado por Olofsson et al. (2012), y para N. atom
por Volkman et al. (1989).
Las 4 microalgas presentaron proporciones bajas de ARA. Según Bigogno et al.
(2002), las únicas microalgas que producen ARA en proporciones significativas son
las Rodofitas. Éstas diferencias de ácidos grasos entre especies se deben a
tamaños y formas diferentes de la células, estructuras y composición de su pared
celular, las condiciones ambientales y sobre todo a su composición bioquímica que
incluye los nutrientes, enzimas y/o toxinas (Brown, 2002).
Por el elevado contenido de ácidos grasos poliinsaturados (PUFA) de I. galbana,
Helm & Bourne (2004) mencionan que es considerada el mejor alimento para
organismos marinos como larvas de moluscos por su alto contenido de DHA así
como por su tamaño (4-6 µm), que además al no poseer una pared celular la hace
más asimilable (Cordoba-Matson et al., 2010). C. calcitrans es una fuente rica de
EPA (Payne & Rippingale, 2000; Renaud et al., 2002) por lo que la mezcla de
éstas microalgas se complementa y puedan proporcionar un adecuado aporte de
los ácidos grasos EPA y DHA. Esto concuerda con lo reportado por Flores-
Santana (2008) quien concluye que al mezclar I.galbana y C. calcitrans se
obtienen mejores perfiles de ácidos grasos en copépodos. Brown (2002)
menciona que, en general, las Clorofitas (T. suecica y N. oculata) no poseen una

34
alta proporción de HUFA y que esta carencia no las hace adecuadas para
utilizarlas como alimento para cultivos de presas vivas. En este estudio N. oculata
y T. suecica presentaron las menores proporciones de HUFA. Sin embargo, la
microalga Nannochloropsis sp. es utilizada al ser más conveniente como fuente
de alimento para cultivo de rotíferos (Støttrup & McEvoy, 2003) por su tamaño (2-
4 !m) y respuesta al cultivo.
El rotífero Brachionus plicatilis presentó mayores porcentajes de lípidos totales que
Brachionus rotundiformis, sin importar el tipo de dieta. Las mayores proporciones se
presentaron en dietas con base en N. oculata y con un proceso de enriquecimiento
(Spresso-Selco®) Evjemo & Olsen (1997) reportan para B. plicatilis enriquecidos con
Super-Selco® un porcentaje de lípidos totales de 6.6%. Por su parte, Kotani et al.
(2009) reportan proporciones más elevadas utilizando N. oculata y el enriquecedor
“DHAProteinSelco®, que van de 13 a 17%, respectivamente. Para Proales similis
alimentado con N. oculata, Wullur et al. (2011) reportan proporciones de lípidos
totales de 2.4% y enriquecidos con “Super Fresh Chlorella vulgaris V-12® de 2.6%;
durante este estudio se encontraron proporciones mayores a las reportadas por
estos autores. El contenido lipídico está considerado como un factor determinante
de su valor nutricional (Fernandez-Reiriz et al., 1993). Las cantidades de lípidos
totales fueron menores a las observadas en las microalgas. Se ha reportado que el
contenido de lípidos es usualmente menor que el encontrado en el alimento
administrado indicando que los lípidos son metabolizados por los rotíferos y/o no
pueden asimilarlos totalmente (Stottrup & McEvoy, 2003).
B. plicatilis y B. rotundiformis al ser alimentados con la dieta compuesta de N.
oculata muestran perfiles de DHA y EPA menores al 1%, mientras que ARA se
presenta con un 5%. Con dieta de T. suecica, las proporciones sólo aumentaron en
EPA, como reflejo del perfil de ésta microalga. Estas proporciones coinciden con los
reportados por Lubzens et al. (1985), Watanabe et al. (1983). Durante este ejercicio
ambas especies de Brachionus presentaron la misma respuesta y tendencia a las
dietas administradas. Esto puede deberse a que B. plicatilis es un complejo de
especies gemelas con una longitud corporal del rango de 90 - 400 !m, y entre estas
especies es B. rotundiformis el más pequeño en tamaño referido comúnmente como
(SS) “super-small” (Wullur et al., 2011), por lo cual comparten muchas

35
características. El perfil de ácidos grasos del rotífero P. similis, que por su menor
tamaño (40-110!m) sólo se logró cultivar con N. oculata como dieta monoalgal,
concuerda con los perfiles reportados por Wullur et al. (2011) y Tomoda et al.
(2014). Presenta tendencias idénticas a los Brachionus spp. cultivados con N.
oculata y sus proporciones de DHA y EPA son bajos aunque presenta un pequeño
aumento de ARA, como reflejo de la dieta.
Por sus bajos niveles de AGE no se recomienda utilizar rotíferos provenientes de
dietas monoalgales por poseer cantidades inadecuadas de DHA y EPA para larvas
de peces marinos (Lubzens et al. 1989; Rainuzzo et al. 1997; Sargent et al. 1997;
Dhert et al., 2001; Støttrup & McEvoy, 2003; Demir & Diken, 2011). El rendimiento
de las dietas depende en gran medida de su digestibilidad, composición química y
contenido energético disponible. El pobre desempeño en dietas con base en N.
oculata puede explicarse por la resistencia y pobre digestibilidad de su pared
celular, su pequeño tamaño y bajo contenido nutritivo (Dhont & Lavens, 1996;
Payne & Rippingale, 2000).
Las tres especies de rotíferos enriquecidos con Spresso-Selco® aumentaron sus
proporciones de PUFA, con especial énfasis en el ácido graso DHA. Esto
concuerda con la mayoría de las publicaciones donde se utiliza algún producto
enriquecedor (Fernandez-Reiriz et al., 1993; Palmtag et al., 2003; Hamre, 2015.
Demir & Diken (2011) evalúan el enriquecedor Spresso® en B. plicatilis y sus
resultados no son diferentes en DHA y ligeramente mayores para ARA y EPA a los
encontrados durante este experimento.
Los rotíferos Brachionus donde la dieta incluía N. oculata, después de enriquecer
presentan mayores proporciones de ARA que rotíferos con dietas que incluían T.
suecica. Sin embargo, rotíferos enriquecidos donde la dieta incluía T. suecica
presenta mejores proporciones de EPA. Esto cobra importancia al estar interesado
en un AGE en particular para administrar a las larvas. Proales similis al ser
enriquecido con Spresso-Selco® presenta un aumento de DHA y EPA, ácidos
grasos que la microalga N. oculata no presenta. Al observar el perfil de ácidos
grasos de las tres especies de rotíferos se puede indicar que la utilización de
Spresso-Selco®, favorece un incremento en la proporción de los ácidos grasos
DHA y EPA; pero no en ARA.

36
Dentro del protocolo de producción de rotíferos para su uso en acuicultura, existe
los procesos de “producción en masa” en el cual se usa la microalga como
alimento para lograr el crecimiento poblacional; seguido del “enriquecimiento”,
para agregar un aporte extra de nutrientes. La principal característica de ORI-
ONE® es fungir como un medio de cultivo para el crecimiento poblacional sin
requerir un posterior enriquecimiento ya que aporta un perfil de AGE más estable
al ser administrado a las larvas de peces. Su estabilidad se debe a que estos
ácidos grasos se incorporan al rotífero. No se han reportado estudios sobre la
dieta comercial ORI-ONE®, no obstante, durante este experimento ambas
especies de los rotíferos Brachionus spp. cultivados en ORI-ONE® arrojaron
proporciones mayores de EPA y ARA que los rotíferos enriquecidos por Spressp-
Selco®, en cuanto a DHA presentaron proporciones similares, dejando claro los
beneficios de las técnicas de enriquecimiento. Para el rotífero Proales similis,
ORI-ONE® presentó mejores proporciones de EPA que con Spresso-Selco y fue
ligeramente menor en cuanto a ARA y DHA. Con la utilización de este tipo de
dieta comercial se puede lograr cultivos de rotíferos con niveles adecuados de
AGE, sin depender de cultivos de microalgas, lo cual se presenta como una
opción muy atractiva para los laboratorios de producción comercial. Los
porcentajes de AGE encontrados en rotíferos enriquecidos con Spresso-Selco® y
con la dieta comercial ORI-ONE® durante este ejercicio son similares a los ya
reportados con diferentes enriquecedores (Tabla XIV) y los rangos de AGES
están dentro de los ya utilizados para diferentes larvas de peces marinos, con
resultados variables de supervivencia (Tabla XV).
Además de un adecuado perfil nutricio cuando esta enriquecido o cultivado con
ORI-ONE®, el rotífero P. similis posee un tamaño de 83μ de largo y 40μ de ancho (38% más pequeño que el rotífero “SS” B. rotundiformis) esto lo convierte en un excelente alimento para larvas de peces marinos con tamaño de boca pequeño
como son Lutjanidos, Epinephelus septemfasciatus, Centropyge ferrugata y
Cheilinus undulatus (Hagiwara et al., 2014). Wullur et al. (2011) mencionan que
larvas de E. septemfasciatus prefirieron P. similis al B. rotundiformis posiblemente
por su tamaño. A su vez, Hirai et al (2012) encuentran que P. similis es
consumido preferentemente por larvas del pez napoléon (Cheilinus undulatus).

37
Tabla XIV. Proporciónes reportadas de AGE en rotíferos con diferentes tipos de enriquecedores comerciales.
Enriquecedor ARA EPA DHA Referencia.
B. plicatilis
Protein Selco Plus® 2.0 7.3 12.1 Cavalin & Weirich (2009)
Algamac 2000® 2.4 4 24 Faulk et al., (2005)
Super-Selco® 1.4 13.8 12.8 Fernandez-Reiriz et al.,1993
B. rotundiformis ORIOGREEN® 1 10 22.1
Hamre, 2016 Spreso®+Multigain®+ Morrón rojo. 1.4 4.3 22.6
P. similis SuperFresh ChlorellaV12® 0.5 11 17.5 Wullur et al. (2011)
Además, Hagiwara et al., (2014) reportan que P. similis fue consumido por larvas de
anguila (Anguilla japonica), un organismo considerado con una crianza larvaria
extremadamente difícil (Tanaka, 2015).
Diversos estudios mencionan mejorías en pigmentación, crecimiento y
supervivencia en larvas de peces cuando se administra copépodos como alimento
vivo, relacionándolo a su contenido y composición lipídica (Evjemo & Olsen, 1997;
Bell et al., 2003; Puello-Cruz et al., 2009; Alajmi et al., 2015). Si bien, en su
ambiente natural las larvas de peces se pueden alimentar de diferentes organismos
zooplanctónicos, los copépodos, especialmente en estadio naupliar, son la presa
virtualmente todas las larvas de peces marinos (Hunter, 1981).
En los copépodos analizados, los estadios más avanzados en su desarrollo
presentaron mayores proporciones de lípidos totales que los estadios neonatos
(nauplios), sin importar qué tipo de dieta fue administrada. Se ha reportado que en
copépodos Calanoideos la acumulación de lípidos ocurre principalmente en los
estadios más grandes con una tendencia a almacenar componentes de cadena más
larga conforme avanza el desarrollo (Kattner et al., 1994).
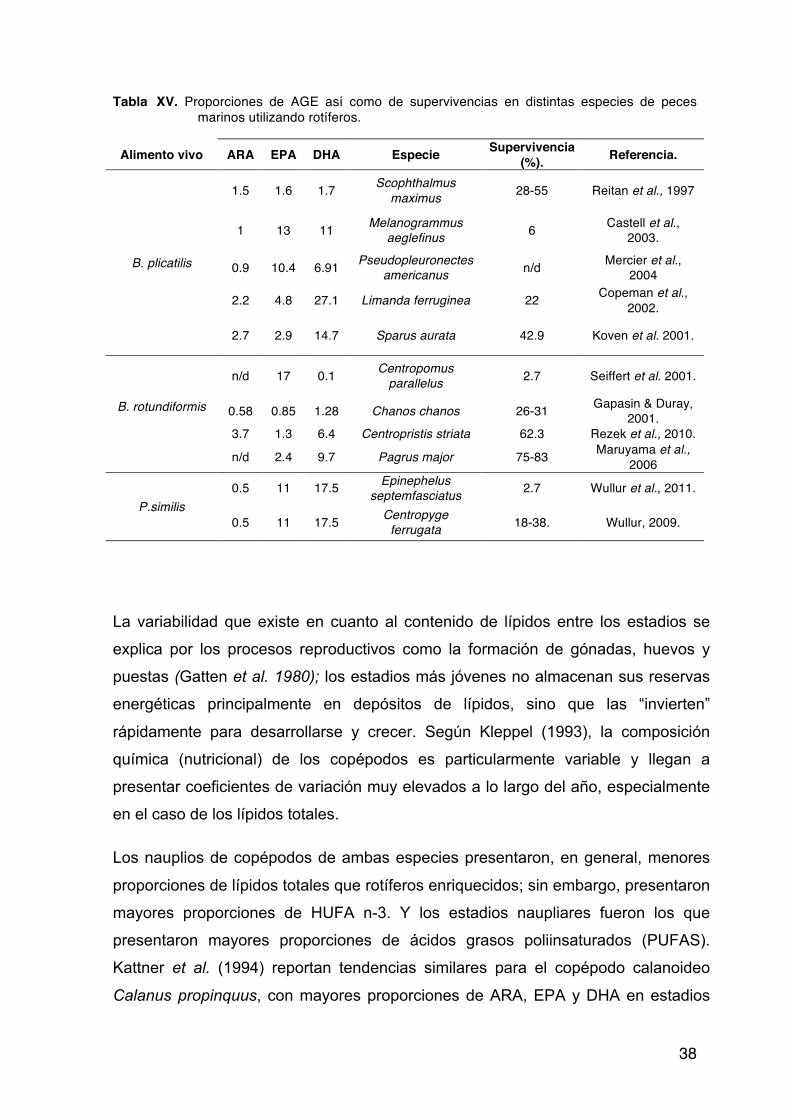
38
Tabla XV. Proporciones de AGE así como de supervivencias en distintas especies de peces marinos utilizando rotíferos.
Alimento vivo ARA EPA DHA Especie Supervivencia (%). Referencia.
B. plicatilis
1.5 1.6 1.7 Scophthalmus maximus
28-55 Reitan et al., 1997
1 13 11 Melanogrammus aeglefinus
6 Castell et al., 2003.
0.9 10.4 6.91 Pseudopleuronectes americanus n/d Mercier et al.,
2004
2.2 4.8 27.1 Limanda ferruginea 22 Copeman et al., 2002.
2.7 2.9 14.7 Sparus aurata 42.9 Koven et al. 2001.
B. rotundiformis
n/d 17 0.1 Centropomus parallelus 2.7 Seiffert et al. 2001.
0.58 0.85 1.28 Chanos chanos 26-31 Gapasin & Duray, 2001.
3.7 1.3 6.4 Centropristis striata 62.3 Rezek et al., 2010.
n/d 2.4 9.7 Pagrus major 75-83 Maruyama et al., 2006
P.similis 0.5 11 17.5 Epinephelus
septemfasciatus 2.7 Wullur et al., 2011.
0.5 11 17.5 Centropyge ferrugata 18-38. Wullur, 2009.
La variabilidad que existe en cuanto al contenido de lípidos entre los estadios se
explica por los procesos reproductivos como la formación de gónadas, huevos y
puestas (Gatten et al. 1980); los estadios más jóvenes no almacenan sus reservas
energéticas principalmente en depósitos de lípidos, sino que las “invierten”
rápidamente para desarrollarse y crecer. Según Kleppel (1993), la composición
química (nutricional) de los copépodos es particularmente variable y llegan a
presentar coeficientes de variación muy elevados a lo largo del año, especialmente
en el caso de los lípidos totales.
Los nauplios de copépodos de ambas especies presentaron, en general, menores
proporciones de lípidos totales que rotíferos enriquecidos; sin embargo, presentaron
mayores proporciones de HUFA n-3. Y los estadios naupliares fueron los que
presentaron mayores proporciones de ácidos grasos poliinsaturados (PUFAS).
Kattner et al. (1994) reportan tendencias similares para el copépodo calanoideo
Calanus propinquus, con mayores proporciones de ARA, EPA y DHA en estadios

39
más jóvenes.
El DHA fue el AGE predomínante en los 4 estadios del copépodo Parvocalanus
crassirostris, especialmente en la dieta monoalgal con base en I. galbana. Esta
elevada proporción en DHA concuerda con los datos reportados por Alajmi et al.
(2015) y refleja el perfil de ácidos grasos de I. galbana, que es reportada como una
fuente rica en DHA (Pillsbury 1985, Dunstan et al., 1993; Payne & Rippingale 2000;
Renaud et.al., 2002; Rivero-Rodríguez et al., 2007). Sin embargo, con I. galbana
presentaron deficiencia en EPA por lo que la mezcla con Chaetoceros calcitrans y
Tetraselmis suecica, ricas en EPA (Napolitano et al., 1990), arrojó un perfil más
equilibrado de AGE. Shields et al. (2005) reportan que el uso combinado de
Chaetoceros sp. e I. galbana favorece la fecundidad y la calidad nutricia de los
nauplios de P. crassirostris lo que favorece la producción en masa. También, Alajmi
et al. (2014) reportan que la mezcla de Isochrysis sp. y Chaetoceros calcitrans
mejora el desempeño en producción de huevos. Schipp et al. (1999) reportan
beneficios similares al utilizar mezclas de microalgas. Debido a esto se ha
identificado a Parvocalanus crassirostris con un alto potencial para especies de
acuacultura debido al pequeño tamaño del nauplio de 80 ± 0.6 μm (Alajmi et al.,
2015) además de su perfil nutricio, considerándose como ideal para la primera
alimentación de varios peces marinos de boca pequeña como pudieran ser
Lutjanidos, Serranidos y Epinephelinos (Støttrup, 2006; Shields et al., 2005)
Pseudodiaptomus euryhalinus, por su parte, presentó las mayores proporciones en
los ácidos grasos esenciales EPA y DHA en todos los estadios sin importar el tipo
de dieta. Sin embargo, en copépodos alimentados con la dieta con base en mezcla
de microalgas éstas proporciones fueron mayores. Flores-Santana (2008), reporta
para la misma especie mejores proporciones de HUFAS con una mezcla de C.
calcitrans e I. galbana, que con una dieta monoalgal. Puello-Cruz et al. (2009),
consideran que Chaetoceros muelleri e I. galbana son los mejores alimentos para
organismos filtradores, debido a sus contenidos de AGE, aún siendo utilizadas de
forma monoalgal. Sin embargo, Knucke et al. (2005) reportan a las dietas mixtas
más convenientes, al complementar las deficiencias nutricionales que tengan las
dietas de una sola microalga, lo cual concuerda con los datos encontrados para P.
euryhalinus en éste estudio ya que la mezcla de las microalgas eleva los contenidos

40
nutricios de DHA y ARA. En relación al EPA, no es tan evidente la mejoría en dietas
multialgales, esto puede deberse a que a pesar que C. calcitrans contiene dos
veces más EPA que T. suecica, fue administrada proporcionalmente en menor
cantidad.
Diversos autores concuerdan que la composición nutricia de copépodos, con
énfasis especial al perfil de ácidos grasos corresponde con los requerimientos
nutricionales de larvas de peces marinos (Watanabe, 1979; Watanabe et al.,
1983; Witt et al., 1984; Shansudin et al., 1997; Toledo et al., 1999). Se atribuye el
éxito de copépodos sobre el de rotíferos a que sus nauplios contienen más del
50% de fosfolípidos, moléculas que pueden ser incorporadas directamente en las
membranas celulares durante el rápido crecimiento de larvas y contienen un perfil
bioactivo de HUFA, especialmente en DHA (Kanazawa, 1985; Kanazawa, 1993;
Geurden et al., 1995). Durante este ejercicio, no se realizó el análisis de clases de
lípidos; sin embargo, se recomienda desarrollar eventualmente estos estudios
para una mejor evaluación del perfil nutricio de copépodos. Algunas especies de
peces marinos han sido criadas utilizando nauplios de copépodos en la primera
alimentación incluyendo a pargos (Doi et al., 1994; Payne et al., 2001; Ogle et al.,
2005; Phelps et al., 2005, Sanchez-Tapia, 2009; Puello-Cruz et al., 2009, meros
(Doi et al., 1997; Toledo et al., 1999), caballitos de mar (Payne & Rippingale,
2000) y angelitos (Shields & Laidley, 2003; Laidley et al., 2004). Sin embargo, a
pesar de los atributos positivos de los copépodos (tamaño, composición
bioquímica), su utilización durante la primera alimentación es aún esporádica,
debido principalmente a las dificultades en su cultivo, destacando su baja
productividad (Drillet et al., 2011).
Aún si Artemia sp. no es el alimento natural de larvas de peces marinos, ha sido
utilizada con éxito para larvas de peces en la acuicultura (Leger et al., 1986); sin
embargo, copépodos y rotíferos enriquecidos presentaron mayores cantidades de
DHA y EPA. Durante este trabajo las mayores proporciones de lípidos totales se
presentan en los estadios naupliares de Artemia sp. y éstos fueron mayores a los
encontrados en copépodos y similares a los encontrados en rotíferos
enriquecidos. Bell et al. (2003) menciona que Artemia posee un alto contenido

41
lipídico, sin embargo, éstos son ácidos grasos estructurales de pobre valor
nutricio.
Artemia sp. presentó las mayores proporciones en los ácidos grasos 16:0, 18:1n-9
y 18:3n-3 (ALA); el DHA presentó proporciones importantes únicamente cuando
se enriqueció con Spresso-Selco®, esto concuerda con lo reportado por distintos
autores para Artemia sp. con algún proceso de enriquecimiento (Shields et al.,
1999; Faulk et al., 2005; Haché & Plante, 2011). El estadio naupliar sin enriquecer
presentó los niveles más bajos de DHA, EPA y ARA. Esto se debe a que el
nauplio no consume alimento externo, sino que se alimenta de sus propias
reservas de energía (Sorgeloos et al., 2001). Seis horas después de eclosionar ya
es nauplio II (Instar II), cerca del 50% más grande y con poca calidad nutricional
porque han agotado sus reservas energéticas, pero ya pueden ser enriquecidos
(Benijts et al., 1977). Sin embargo, se han reportado problemas para el
enriquecimiento de Artemia sp. debido sobre todo, a una rápida retroconversión
de DHA a EPA (Tuncer & Harrell, 1992; Navarro et al., 1999; Bell et al., 2003).
Evjemo et al. (1997) relacionan esto al hecho de que las cantidades de DHA en
Artemia sp. son inestables porque se reducen después de las primeras horas del
enriquecimiento. McEvoy et al. (1995) sugieren que los nauplios enriquecidos no
actúan sólo como vehículos bioencapsuladores, sino que modifiquen activamente
los ácidos grasos en otros menos biodisponibles para las larvas. Además,
contrario a los rotíferos, Artemia sp. es capaz de convertir rapidamente a
trigliceridos y asimilar los PUFA presentes en las emulsiones comerciales
(Navarro et al., 1999). Bell et al. (2003) reportan que los mejores resultados en
supervivencia utilizando rotíferos y copépodos se debe a la digestibilidad y
disponibilidad de HUFAS en forma de fosfolípidos y que Artemia almacena lípidos
como triglicéridos más que en fosfolípidos. Los fosfolípidos dietarios mejoran el
consumo de alimento, actúan como emulsificadores en el intestino, reducen la
peroxidación lipídica, estimulan la síntesis de lipoproteinas, teniendo beneficios
para el crecimiento y desarrollo de larvas de peces (Koven et al., 1993; McEvoy et
al., 1995; Koven et al., 1998; Fontagne et al., 1998; Geurden et al., 1995).
Durante este experimento Artemia sp. enriquecida aumentó sus proporciones de
DHA, EPA y ARA; cabe mencionar que las muestras fueron tomadas

42
inmediatamente después del enriquecimiento, por lo que estos valores de AGE en
las prácticas de cultivo podrían disminuir mientras transcurre el tiempo antes de
ser consumidos por las larvas. El estadio adulto, presentó una deficiencia en
cuanto a PUFA específicamente en DHA, además de ser muy grande para
alimentar durante la primera alimentación a larvas de peces marinos, por lo cual
se enfoca su uso en juveniles de larvas. Por todo lo anterior, los nauplios de
Artemia están aún lejos de ser una dieta óptima para larvas marinas, comparadas
con el plancton marino (Navarro et al., 1999).
Evjemo et al. (1997) reporta que en experimentos de primera alimentación con
larvas de bacalao, éstas consumen Artemia. Además, tienen mayor preferencia
hacia la artemia cuando se alimenta junto con copépodos. Sin embargo, los
problemas de pigmentación de la piel se acrecentan cuando se alimentan
únicamente de nauplios de Artemia. Aunque son usados comúnmente como
alimento de transición entre presas vivas y alimento inerte resultan inadecuados
por su tamaño, en la primera alimentación de especies de peces marinos con
tamaño de boca pequeña (Alajmi & Zeng, 2013). Para estas especies, pequeños
rotíferos como son B. rotundiformis y P. similis se muestran muy prometedores,
así como los nauplios de copépodos, como P. euryhalinus y P. crassirostris.
En el ambiente natural, los peces y los huevos de peces presentan DHA y EPA en
una proporción cercana a 2, debido a esto, diversos autores asumen que el
alimento vivo utilizado durante el cultivo larvario debe proveer una relación similar
de DHA/EPA (Tocher & Sargent, 1984; Rodríguez et al., 1997; Bell et al., 2003)
aunque esta proporción varía según la especie de pez a cultivar. Diversos autores
relacionan que proporciones de DHA/EPA cercanas a 2 en alimento vivo propiciaron
mejores resultados de crecimiento y sobrevivencia de larvas de diferentes especies
(Curé et al., 1996; Rodríguez et al.,1997; Copeman et al., 2002; Hafezieh et al.,
2009). Sin embargo, Dantagnan et al. (2010), al evaluar dos dietas: una alta (2.18) y
otra baja (0.64) en DHA/EPA, a diferentes salinidades (0, 10 y 15 g L-1) en larvas de
Galaxias maculatus reportan que se requiere más DHA en altas salinidades y más
EPA en salinidades bajas. Lo cual indica que incluso pequeños cambios en
salinidad pueden determinar los requerimientos óptimos de EPA, DHA y la relación
DHA/EPA.

43
En este trabajo, las relaciones de DHA/EPA de los rotíferos y Artemia sp. sin
enriquecer fueron muy bajas, porque los contenidos de DHA son cercanos a 0%. En
rotíferos enriquecidos con Spresso-Selco®, las relaciones DHA/EPA aumentaron
marcadamente (B. plicatilis 5.5 a 8.13; B. rotundiformis 4.25 a 9.19; P. similis
11.05), sin embargo, en algunos casos duplican y en otros quintuplican el rango
recomendado (≈2). Los rotíferos alimentados con dieta comercial ORI-ONE®
también superan en el doble el valor DHA/EPA (4.45; 3.02 y 4.85, respectivamente)
y en particular los nauplios de copépodos obtenidos de adultos alimentados con
dietas multialgales presentaron relaciones más cercanas (2 y 1.96,
respectivamente) al valor recomendado para larvas de peces marinos, 2 (Tucker,
1998; Sargent et al., 1999). Lo cual refleja un mejor equilibrio de los AGE.
7. CONCLUSIONES Los resultados obtenidos en este trabajo sugieren que:
1. Los perfiles de ácidos grasos presentes en microalgas, se ven reflejados en el
perfil de ácidos grasos de rotíferos, copépodos con mayor presencia de DHA
en Isochrysis galbana, de EPA en Chaetoceros calcitrans y Tetraselmis
suecica y de ARA en Nannochloropsis oculata
2. Los rotíferos de las tres especies, cultivados solamente con base en
microalgas presentan un perfil de AGE y relación de DHA:EPA muy bajo, por
lo cual no cubren los requerimientos recomendados para larvas de peces
marinos. Sin embargo, una vez enrequecidos con Spresso-Selco® sobrepasa
éstos niveles.
3. Los rotíferos de las tres especies, alimentados con ORI-ONE® presentan
niveles de AGE y relación DHA:EPA ligeramente superiores de las
recomendaciones para larvas de peces marinos.
4. Los estadios naupliares de copépodos de ambas especies, presentan mayores
proporciones de DHA que estadios con mayor grado de desarrollo.

44
5. Las dietas multialgales en copépodos presentan proporciones mejor
equilibradas de AGE que las dietas monoalgales.
6. EL copépodo Parvocalanus crassirostris presenta bajas proporciones de ARA
en todos los estadios indistintamente de la dieta.
7. Los nauplios de Pseudodiaptomus euryhalinus presentan mayores
proporciones de DHA que nauplios de Parvocalanus crassirostris.
8. La Artemia sp. debe ser enriquecida para ser administrada a larvas de peces;
sin embargo, aún así es imprecindible por la fácil disposición, para satisfacer la
necesidad de alimento en larvas, cubrir la sensación de saciedad (evitando el
canibalismo) y para la transición a dietas inertes (destete).
8. RECOMENDACIONES
• Realizar un análisis bromatológico en rotíferos, copépodos y Artemia sp.
para evaluar otros nutrientes como proteínas y carbohidratos.
• Realizar un análisis de fosfolípidos y triacilglicéridos.
• Realizar experimentos de crianza larvaria alimentando con rotiferos
cultivados con ORI-ONE® para evaluar el crecimiento y sobrevivencia y
comparar con los resultados obtenidos con nauplios de copépodos
producidos con base en dietas mono y multialgales.

45
BIBLIOGRAFÌA
Alajmi, F. & C. Zeng. 2013. Evaluation of microalgal diets for the intensive
cultivation of the tropical calanoid copepod, Parvocalanus crassirostris.
Aquacult. Res., 46: 1025–1038.
Alajmi, F., C. Zeng & D. Jerry. 2014. Domestication as a novel approach for
improving the cultivation of calanoid copepods: a case study with
Parvocalanus crassirostris. PLoS ONE, 10(7): e0133269.
Álvarez-González, C. A. 1999. Optimización del proceso de producción de cabrilla
arenera Paralabrax maculatofasciatus (Percoide: Serranidae) en sistema de
circulación cerrada. Tesis de maestría. CICIMAR-IPN. 108p.
Aquaculture topics and activities. Acuicultura. In: Departamento de Pesca y
Acuicultura de la FAO [en línea]. Roma. Actualizado 14 September 2015.
Barclay, W. & S. Zeller. 1996. Nutritional enhancement of n-3 and n-6 fatty acids in
rotifers and Artemia nauplii by feeding spray-dried Schizochytrium sp. J. World
Aquacult. Soc., 27:314-322.
Bell, J. G. & J. R. Sargent. 2003. Arachidonic acid in aquaculture feeds: current
status and future opportunities. Aquaculture, 218: 491–499.
Bell, J. G., F. McGhee, P.J. Campbell & J.R. Sargent. 2003. Rapeseed oil as an
alternative to marine fish oil in diets of post-smolt Atlantic salmon (Salmo
salar): changes in flesh fatty acid composition and effectiveness of subsequent
fish oil “wash out”. Aquaculture, 218: 515–528.
Bengtson, D. 2003. Status of marine aquaculture in relation to live prey: past,
present and future. En: Stottrup, G. J. & M. A. McEvoy (Eds.) Live Feeds in
Marine Aquaculture. Blackwell Science Ltd, Gran Bretaña. 1-16.
Benijts, F., G. Van De Putte & P. Sorgeloos. 1977. Energetic Aspects Of The
Metabolism Of Hydrated cysts. 79-87. En: Jasper E. (Eds.) Fundamental and
Research on the Brine Shrimp Artemia salina (L.) in Belgium. Institute for
Marine Scientific Research. Bredene, Belgica.
Bigogno, C., I. Khozin-Goldberga, S. Boussibaa, A. Vonshaka & Z. Cohena. 2002.
Lipid and fatty acid composition of the green oleaginous alga Parietochloris
incisa, the richest plant source of arachidonic acid. Phytochemistry, 60: 497–
503.

46
Bondioli, P., L. D. Bella, G. Rivolta, G. C. Zittelli, N. Bassi, L. Rodolfi, D. Casini, M.
Prussi, D. Chiaramonti & R. Tredici. 2012. Oil production by the marine
microalgae Nannochloropsis sp. F&M-M24 and Tetraselmis suecica F&M-
M33. Bioresour. Techno., 114: 567–572.
Brown, M. R. 2002. Nutritional value of microalgae for aquaculture. En: Cruz-
Suárez, L. E., D. Ricque-Marie, M. Tapia-Salazar, M. G. Gaxiola-Cortés & N.
Simoes (Eds.) Avances en Nutrición Acuícola VI. Memorias del VI Simposium
Internacional de Nutrición Acuícola. 3 al 6 de Septiembre del 2002. Quintana
Roo, México.
Brown, M. R., S. W. Jeffrey, J. K. Volkman & G. A. Dunstan. 1997. Nutritional
properties of microalgae for mariculture. Aquaculture, 151: 315-331.
Castell, J. D., J. G. Bell, D. R. Tocher & J. R. Sargent. 1994. Effects of purified
diets containing different combinations of arachidonic and docosahexaenoic
acid on survival, growth and fatty acid composition of juvenile turbot
(Scophthalmus maximus). Aquaculture, 128, 315-333.
Castell, J. D., R. O. Sinnhuber, J. H. Wales & D. J. Lee. 1972. Essential fatty acids
in the diet of rainbow trout, Salmo gairdneri, growth, feed conversion and
some gross deficiency symptoms. J. Nutr., 102:77-86.
Castell, J. D., T. Blair, S. Neil, K. Howes, S. Mercer, J. Reid, W. Young-Lai, B.
Gullison, P. Dhert & P. Sorgeloos. 2003. The effect of different HUFA
enrichment emulsions on the nutritional value of rotifers (Brachionus plicatilis)
fed to larval haddock (Melanogrammus aeglefinus). Aquac. Int., 11:109-111.
Castro, T. R., G. DeLara, G. Castro-Mejía, J. Castro & A. Malpica. 2003. Alimento
vivo en la acuicultura. ContactoS, 48: 27-33.
Cavalin, F.G & C.R. Weirich. 2009. Larval performance of aquacultured Florida
pompano (Trachinotus carolinus) fed rotifers (Brachionus plicatilis) enriched
with selected commercial diets. Aquaculture, 292: 67–73.
Cerqueira, V. & A. Brugger. 2001. Effect of light intensity on initial survival of fat
snook (Centropomus parallelus, Pisces: Centropomidae) larvae. Braz. Arch.
Biol. Technol., 44: 343–349.
Civera-Cerecedo, R., C. A. Alvarez-González & F. J. Moyano-López. 2004.
Nutrición y alimentación de larvas de peces marinos. En: Cruz -Suárez, L. E.,
D. Ricque-Marie, M. G. Nieto López, D. Villarreal, U. Scholz & M. González
(Eds.) Avances en Nutrición Acuícola VII. Memorias del VII Simposium

47
Internacional de Nutrición Acuícola. 16-19 Noviembre, 2004. Hermosillo,
Sonora, México.
Copeman, L. A., C. Parrish, J. A Brown & M. Harel. 2002. Effects of
docosahexaenoic, eicosapentaenoic, and arachidonic acids on the early
growth, survival, lipid composition and pigmentation of yellowtail flounder
(Limanda ferruginea): a live food enrichment experiment. Aquaculture, 210:
285–304.
Córdoba-Matson, M., J. Gutiérrez & M. Porta-Gándara. 2010. Evaluation of
Isochrysis galbana (clone T-ISO) cell numbers by digital image analysis of
colour intensity. J. Appl. Phycol., 22: 427–434.
Curé, K., G. Gajardo & P. Coutteau. 1996. The effect of DHA/EPA ratio in live feed
on the fatty acid composition, survival, growth and pigmentation of turbot
larvae Scophthalmus maximus L. 57-67. En: Gajardo, G. & P. Coutteau (Eds.)
Improvement of the Commercial Production of Marine Aquaculture Species.
Proceedings of a workshop on Fish and Mollusc Larviculture. Santiago, Chile.
Dantagnan, P., A. Borquez, A. Hernández & M.S. Izquierdo. 2010. Effect of
EPA/DHA ratios on the growth and survival of Galaxias maculatus (Jenyns,
1842) larvae reared under different salinity regimes. Aquacult. Res., 41: 239-
244.
Delaporte, M., P. Soudant, J. Moal, E. Kraffe, Y. Marty & J. F. Samain. 2005.
Incorporation and modification of dietary fatty acids in gill polar lipids by two
bivalve species Crassostrea gigas and Ruditapes philippinarum. Comp.
Biochem. Physiol. A Mol. Integr. Physiol., 140: 460-470.
Demir, O. & G. Diken. 2011. Effects of commercial enrichment products on
chemical constitutions of rotifer Brachionus plicatilis (O.F. Muller, 1786). J.
Anim. Veter. Adv., 10: 3328-3333.
Dhert, P., G. Rombaut, G. Suantika & P. Sorgeloos. 2001. Advancement of rotifer
culture and manipulation techniques in Europe. Aquaculture, 200: 129-146.
Dhont, J. & P. Lavens. 1996. Tank production and use of ongrown Artemia. 75-82.
En: Sorgeloos, P. & P. Lavens (Eds.) Manual on the Production and Use of
Live Food for Aquaculture. Fisheries Technical Paper No. 361. Food and
Agriculture Organisation of the United Nations, Rome.

48
Doi, M., A. Ohino, Y. Taki, T. Singhagraiwan & H. Kohno. 1997. Nauplii of calanoid
copepod Acartia sinjiensis as an initial food organism for larval red snapper,
Lutjanus argentimaculatus. Sisanzoshoku, 45: 31-40.
Doi, M., H. Khono, A. Ohno & Y. Taki. 1994. Development of mixed feed state
larvae of red snapper, Lutjanus argentimaculatus. Suisanzoshoku 42: 471-
476.
Drillet, G., S. Frouël, M. Sichlau, P M. Jepsen, J. Højgaard, A. K. Joarder & B. W.
Hansen. 2011. Status and recommendations on marine copepod cultivation for
use as live feed. Aquaculture, 315: 155–166.
Essmann-Prieto, M. 2011. Determinación de parámetros biológicos de la microalga
Isochrysis Aff. Galbana: comparación de un fotobioreactor continuo versus un
cultivo batch. Tesis de Licenciatura Escuela de Acuícultura y Pesquerías.
Universidad Austral de Chile, Chile. 57p.
Evjemo, J.O. & Y. Olsen. 1997. Lipid and fatty acid content in cultivated live feed
organisms compared to marine copepods. Hydrobiologia, 358:159-162.
FAO. 2012. El estado mundial de la pesca y la acuicultura. Roma. 231p.
Faulk, C.K., G. J. Holt & D.A. Davies. 2005. Evaluation of fatty acid enrichment of
live food for yellowtail snapper Ocyurus chrysurus larvae. J. World Aquac.
Soc., 36: 271–281.
Fernández-Reiriz, M. J., U. Labarta & M. J. Ferreiro. 1993. Effects of commercial
enrichment diets on the nutritional value of the rotifer (Brachionus plicatilis).
Aquaculture, 112: 195-206.
Fernández-Reiriz, M. J., A. Pérez-Camacho, M. J. Ferreiro, J. Blanco, M. Planas,
M. J. Campos & U. Labarta. 1989. Biomass Production and Variation in the
Biochemical Profile (Total Protein, Carbohydrates, RNA, Lipids and Fatty
acids) of Seven Species of Marine Microalgae. Aquaculture, 83: 17-37.
Fidalgo, J., A. Cid, E. Torres, A. Sukenik & C. Herrero. 1998. Effects of nitrogen
source and growth phase on proximate biochemical composition, lipid classes
and fatty acid profile of the marine microalga Isochrysis galbana. Aquaculture,
166: 105–116.
Flores-Santana, R. 2008. Variación en el contenido de ácidos grasos del copépodo
calanoide Pseudodiaptomus euryhalinus (Johnson, 1939) alimentado con las
microalgas Chaetoceros calcitrans e Isochrysis galbana. Tésis de Maestría.
CIBNOR. La Paz, BCS, México.

49
Fujita, S. 1979. Culture of red sea bream Pagrus major and its food. 183-197. En:
Styczynska-Jurewicz, E., T. Backiel, E. Jaspers & G. Persoone (Eds.)
Cultivation of Fish Fry and its Live Food. European Mariculture Society Special
Publication No. 4, Bredene, Belgium.
Gapasin, R. & M. Duray. 2001. Effects of DHA-enriched live food on growth,
survival and incidence of opercular deformities in milkfish (Chanos chanos).
Aquaculture, 193: 49-63.
Gatten, R. R., J. R. Sargent, T. E. Forsberg, S. C. O'Hara & M. Corner. 1980. On
the nutrition and metabolism of zooplankton. XIV. Utilization of tipid by
Calanus helgolandicos during maturation and reproduction. J. Mar. Biol.
Assoc., 60: 391-399.
Geurden, I., J. Radünz-Neto & P. Bergot. 1995. Essentiality of dietary phospholipids
for carp (Cyprinus carpio L.) larvae. Aquaculture, 131: 303-314.
Glencross, B. 2009. Exploring the nutriotional demand for essential fatty acids by
aquaculture species. Rev. Aquacult., 1: 71-124.
Go, S., S. J. Lee, G. T. Jeong & S. Kim. 2012. Factors affecting the growth and the
oil accumulation of marine microalgae Tetraselmis suecica. Bioprocess.
Biosyst. Eng., 35: 145-150.
Gracia-López, V., M. Kiewek-Martínez, M. Maldonado-García, P. Monsalvo-
Spencer, G. Portillo-Clark, R. Civera-Cerecedo, M. Linares-Aranda, M.
Robles-Mungaray & M. Mazón-Suástegui. 2005. Larvae and juvenile
production of the leopard grouper Mycteroperca rosacea (Streets, 1877).
Aquacult. Res., 36: 110-112.
Guillard, R. R. & J. H. Ryther. 1962. Studies of marine planktonic diatom: Cyclotella
nana (Hustedt) and Detonela Confervacea (Cleve). Can. J. Microbiol., 8: 229-
239.
Gundstone, F., J. Harwood & F. Padley. 1994. The Lipid Handbook (Second
Edition). Chapman & Hall, Londres, Inglaterra. 791p.
Haché, R. & S. Plante. 2011. The relationship between enrichment, fatty acid
profiles and bacterial load in cultured rotifers (Brachionus plicatilis L-strain)
and Artemia (Artemia salina strain Franciscana). Aquaculture, 311: 201-208.
Hafezieh, M., M. S. Kamarudin & N. Agh. 2009a. Nutritional enhancement of total
lipid, n-3 and n-6 fatty acids in artemia urmiana nauplii by enriching with
ICES/30/4. Pak. J. Biol. Sci., 11: 2167-2170.

50
Hafezieh, M., M. S. Kamarudin, C. R. Bin Saad, M.K. Abd Sattar, N. Agh & H.
Hosseinpoar . 2009b. Effect of enriched Artemia urmiana on growth, survival
and composition of larval Persian sturgeon. Turkish J. Fish. Aquat. Sci., 9:
201-207.
Hagiwara, A., H. Wullur, N. Marcial, Y. Hirai & Y. Sakakura. 2014. Euryhaline rotifer
Proales similis as initial live food for rearing fish with small mouth.
Aquaculture, 432: 470-474.
Hagiwara, A., W.G. Gallardo, M. Assavaaree, T. Kotani & A.B. De Araujo. 2001.
Live food production in Japan: recent progress and future aspects.
Aquaculture, 200: 111-127.
Hamre, K. 2015. Nutrient profiles of rotifers (Brachionus sp.) and rotifer diets from
four different marine fish hatcheries. Aquaculture, 450: 136-142.
Helm, M. M., N. Bourne. 2004. Hatchery culture of bivalves. A practical manual.
177p. En: Lovatelli, A. (Eds.). FAO Fisheries Technical Paper 471. Food and
Agriculture Organization of the United Nations Publishing, Roma, Italia,
Hardy, R.W., T. M. Scott & L.W. Harrell. 1987. Replacement of herring oil with
menhaden oil, soybean oil, or tallow in the diets of Atlantic salmon raised in
marine netpens. Aquaculture, 65: 267–277.
Hauville, M. R., K. Main, H. Migaud & J. Gordon. 2014. Fatty acid utilization during
the early larval stages of Florida pompano (Trachinotus carolinus) and
common snook (Centropomus undecimalis). Aquac. Res., 47(5): 1443-1458.
Helm, M. & N. Bourne. 2004. Hatchery culture of bivalves, a practical manual.
Food and Agriculture Organization of the United Nations. Italia, Roma.1-177.
Henderson, R. & D. R Tocher. 1987. The lipid composition and biochemistry of
freshwater fish. Prog. Lipid Res., 26: 281–347.
Hirai, N., M. Koiso, K. Teruya, M. Kobayashi, T. Takebe, T. Sato, K. Nakamura, T.
Goto, & A. Hagiwara. 2012. Rearing conditions for humphead wrasse
Cheilinus undulatus larvae, and Introduction of the minute rotifer Proales
similis as an initial live food. J. Fish. Tech., 4(2): 57-64.
Hunter, J. R. 1981. Feeding ecology and predation of marine fish larvae. 33-77.En:
Lasker, R. (Ed.) Marine fish larvae. Univ. Wash. Press. Seattle.
Ibeas, C., M. S. Izquierdo & A. Lorenzo. 1994. Effect of different levels of n-3 highly
unsaturated fatty acids on growth and fatty acid composition of juvenile
gilthead sea bream (Sparus aurata). Aquaculture, 127: 177-188.

51
Jeffrey, S.W., M. R. Brown & J. K. Volkman. 1994. Haptophyte as feedstocks in
mariculture. 287-302. En: Green, J. C & B. S. Leadbeater (Eds.) The
Haptophyte Algae, Clarendon Press, Oxford.
Kalogeropoulos, N., M. N. Alexis & R. J. Henderson. 1992. Effects of dietary
soybean and cod liver oil levels on growth and body composition of gilthead
bream (Sparus aurata). Aquaculture, 104: 293-308.
Kanazawa, A. 1985. Essential fatty acid and lipid requirement of fish. 281-298. En:
Cowey, C. B., A. M. Mackie & J. G. Bell (Eds.) Nutrition and Feeding in Fish.
Academic Press London. Reino Unido.
Kanazawa, A.1993. Nutritional mechanisms involved in the occurrence of abnormal
pigmentation in hatchery-reared flatfish. J. World Aquacult. Soc., 24: 162-166.
Kanzawa, A. 2000. Nutrición de larvas de peces. 53-64. En: Civera-Cerecedo, R.,
C. J. Pérez-Estrada, D. Ricque-Marie & L. E. Cruz-Suárez (Eds.) Avances en
Nutrición Acuícola IV. Memorias del IV Simposium Internacional de�Nutrición
Acuícola. Noviembre 15-18, 1998. La Paz, B.C.S., México.
Kattner, G. & M. Krause. 1987. Changes in lipids during the development of
Calanus finmarchicuss from copepodid I to adult. Mar. Biol., 96: 511–518.
Kattner, G., M. Graeve & W. Hagen. 1994. Ontogenetic and seasonal changes in
lipid and fatty acid/alcohol compositions of the dominant Antarctic copepods
Calanus propinquus, Calanoides acutus and Rhincalanus gigas. Mar. Biol.,
118: 637–644.
Kennish, J. M., J. L. Sharpdahl, K. A Chambers, F. Thrower & S. Rice. 1992. The
effect of a herring diet on lipid composition, fatty acid composition, and
cholesterol levels in muscle tissue of pen-reared chinook salmon
(Oncorhynchus tshawytscha). Aquaculture, 108: 309-322.
Kleppel, G. S. 1993. On the diets of calanoid copepods. Mar. Ecol. Prog. Ser., 99:
183–195.
Kotani,T., T. Genka, H. Fushimi, M. Hayashi, K. Dierckens & P. Sorgeloos. 2009.
Effect of cultivation methods on nutritional enrichment of euryhaline rotifer
Brachionus plicatilis. Aquaculture, 75: 975-984.
Koven, W., S. Kolkovski, E. Hadas, K. Gamsiz & A. Tandler. 2001. Advances in the
development of MD's for gilthead seabream, Sparus aurata: a review.
Aquaculture, 194: 107–121.

52
Laidley, C.W., R. J. Shields & A. O. Ostrowski. 2004. Progress in amberjack culture
at the Oceanic Institute in Hawaii. Glob. Aqua. Advoc., 7(1): 42-44.
Lee, K. W., S. Raisuddin, D. S. Huang, H. G. Park, H. U Dahm, I. Y. Ahn & J. S.
Lee. 2008. Two generation toxicity study on the copepod model species
Tigriopus japonicus. Chemosphere, 72:1359–1365.
Leger, P., D. A. Bengtson, K. L. Simpson & P. Sorgeloos. 1986. The use and
nutritional value of Artemia as a food source. Oceanogr. Mar. Biol., 24: 521–
623.
Lira, G. 2002. Influence of diet on reproduction and growth of the copepod,
Apocyclops procerus, and its potential as food in larviculture of the fat snook,
Centropomus parallelus. Tesis de Maestría. Universidad Federal de Santa
Catarina, UFSC. Florianópolis, Santa Catarina, Brazil. 41p.
Llorente-García, I. 2013. Análisis de competitividad de las empresas de acuicultura.
aplicaciones empíricas al cultivo de la dorada (Sparus aurata) y la lubina
(Dicentrarchus labrax). Tesis De Doctorado, Facultad de Ciencias económicas
Y empresariales. Departamento de administración de empresas Universidad
de Cantabria, Barcelona. 398p.
Lubzens, E., A. Marko & A. Tietz. 1985. De novo synthesis of fatty acids in the
rotifer Brachionus plicatilis. Aquaculture, 47: 27-37.
Lubzens, E., A. Tandler & G. Minkoff. 1989. Rotifers as food in aquaculture.
Hydrobiologia, 186: 387–400.
Malpica- Sánchez, A., T. Castro-Barrera, H. Sandoval-Trujillo, J. Castro-Mejía, R.
DeLara-Andrade & G. Castro-Mejía. 2003. Composición del contenido de
ácidos grasos en tres poblaciones mexicanas de Artemia franciscana de
aguas epicontinentales. Rev. Biol. Trop., 52(1): 297-300.
Maruyama, S., I. Yamamoto, M. Hayash & O. Mura. 2006. Rotifers fed with n-3
highly unsaturated fatty acid-enriched fish. generally, performed a for
secondary culture of Chlorella vulgaris are suitable for the rearing of larval red
sea bream Pagrus major. Aquacult. Sci., 54(2): 229-230.
McEvoy, L. A., J. C. Navarro, J. G. Bell & J. R. Sargent. 1995. Autoxidation of oil
emulsions during the Artemia enrichment process. Aquaculture, 134: 101–112.
McKinnon, A., S. Duggan, P. Nichols, M. Rimmer, G. Semmens & B. Robino. 2003.
The potential of tropical paracalanoid copepods as live feeds in aquaculture.
Aquaculture, 223: 89-106.

53
Mercier, L., C. Audet, J. de la Noue, B. Parent, C. Parrish & N. W. Ross. 2004.
First feeding of winter flounder (Pseudopleuronectes americanus) larvae: use
of Brachionus plicatilis acclimated at low temperature as live prey.
Aquaculture, 229: 361-376.
Moreno-Figueroa, L. 2011. Efecto de la temperatura en el desarrollo larvario
temprano del huachinango del Pacífico Lutjanus peru (Nichols and Morphy,
1922). Tesis de Maestría. Centro de Investigaciones Biológicas del Noreste,
CIBNOR. La Paz, B.C.S., México. 62p.
Napolitano, G., R. Ackman & W. Ratnayake. 1990. Fatty acid composition of three
cultured algal species (Isochrysis galbana, Chaetoceros gracilis and
Chaetoceros calcitrans) used as food for bivalve larvae. J. World Aquac. Soc.,
21: 122-130.
Navarro-Hurtado, A. 2014. Niveles de HUFA n-3 en el músculo del camarón blanco
del pacífico Litopenaeus Vannamei usando dietas terminales alternativas.
Tesis de Maestría UNAM. México. 130p.
Navarro, J., R. Henderson, L. McEvoy, M. Bell & F. Amat. 1999. Lipid conversions
during enrichment of Artemia. Aquaculture, 174: 155-166.
Ogle, J. T., L. C. Lemus, D. N. Nicholson, J. F. Barnes & D. Lotz. 2005.
Characterization of an extensive zooplankton culture system coupled with
intensive larval rearing of red snapper Lutjanus campechanus. 225–244.En:
Lee, P., J. O’Bryen & N.H. Marcus (Eds.) Copepods in Aquaculture. Blackwell
Science, Oxford. Reino Unido.
Ohse, S., R. Bianchini-Derner, R. Ávila-Ozório, R. Gordo-Correa, E. Badiale & P. C.
Cunha. 2014. Lipid content and fatty acid profiles in ten species of microalgae.
Idesia, 33: 93-101.
Olofsson, M., T. Lamela, E. Nilsson, J. Pascal-Bergé, V. Del-Pino, P. Uronen & C.
Legrand. 2012. Seasonal variation of lipids and fatty acids of the microalgae
Nannochloropsis oculata grown in outdoor large-scale photobioreactors.
Energies. 5: 1577-1592.
Organización De Las Naciones Unidas Para La Agricultura Y La Alimentación
(FAO). 2006. El Estado Mundial De La Pesca y La Acuícultura. Roma: FAO.
Palacios, E. & O. Arjona. 2009. Análisis de ácidos grasos meltilesterificados
(Derivatización directa) en alimento peletizado enriquecido con lípidos.
Informe técnico No. 9. CIBNOR. La Paz, B.C.S., México.

54
Patil, V., T. Källqvist, E. Olsen, G. Vogt & H. Gislerød. 2007. Fatty acid composition
of 12 microalgae for possible use in aquaculture feed. Aquacult. Int., 15: 1-9.
Payne, M. F. & R. J. Rippingale. 2000. Evaluation of diets for culture of the calanoid
copepod Gladioferens imparipes. Aquaculture, 187: 85-96.
Payne, M. F., R. J. Rippingale & J. J. Cleary. 2001. Cultured copepods as food for
West Australian dhufish (Glauscosoma hebraicum) and pink snapper (Pagrus
auratus) larvae. Aquaculture, 194: 137-150.
Peña, R. 2005. Estudio de la funciòn digestiva en la cabrilla arenera Paralabrax
maculatofasciatus: Aspectos alimenticios y sus aplicaciones. Tesis de
Doctorado. CICIMAR-IPN. La Paz, B.C.S., México. 141p.
Phelps, R. P., G. S. Sumiarsa, E. E. Lipman, H. P Lan, K. Kao-Moss & A. D. Davis.
2005. Intensive and extensive production techniques to provide copepod
nauplii for feeding larval red snapper Lutjanus campechanus. 151-168. En:
Lee, C. S., P. J. O'Bryen & N. H. Marcus (Eds.) Copepods in Aquaculture.
Blackwell Publishing, Ames, Iowa, Estados Unidos.
Pousáo-Ferreira, P., F. Cairrão, F. Nery & L. Narciso. 1997. The fatty acid profile of
Spurus uururu larvae is correlated to the composition of the enrichment diets
of Brachionus plicatilis and Artemia sp. Cienc. Mar., 23(l): 83-92.
Prieto, M., L. de la Cruz & M. Morales. 2006. Cultivo experimental del cladócero
Moina sp. alimentado con Ankistrodesmus sp. y Saccharomyces cereviseae.
Rev. MVZ Cordoba, 11(1): 705-714.
Puello-Cruz, A. C., S. Mezo-Villalobos, B. González-Rodríguez & D. Voltolina.
2009. Culture of the calanoid copepod Pseudodiaptomus euryhalinus
(Johnson 1939) with different microalgal diets. Aquaculture, 290: 317–319.
Rainuzzo, J., K. I. Reitan & Y. Olsen. 1997. The significance of lipids at early stages
of marine fish: a review. Aquaculture, 155: 103–115.
Rasdi N., J. Qin & Y. Li. 2015. Effects of dietary microalgae on fatty acids and
digestive enzymes in copepod Cyclopina kasignete, a potential live food for
fish larvae. Aquac. Res., 47(10): 1-11.
Reitan, K. I., J. R. Rainuzzo G. Oie & Y. Olsen. 1997. A review of the nutritional
effects of algae in marine fish larvae. Aquaculture, 155: 207–221.
Rekha, D. C., K. Chakraborty & E. V. Radhakrishnan. 2007. Variation in fatty acid
composition of Artemia salina nauplii enriched with microalgae and baker's
yeast for use in larviculture. Agric. Food Chem., 55: 4043-4051.

55
Renaud, S. M., L. V. Thinh, G. Lambrinidis & D. L. Parry. 2002. Effect of
temperature on growth, chemical composition and fatty acid composition of
tropical Australian microalgae grown in batch cultures. Aquaculture, 211: 195-
214.
Renaud, S., D. Parry, L. Thinh, C. Kuo, A. Padovan & N. Sammy.1991. Effect of
light intensity on the proximate biochemical and fatty acid composition of
Isochrysis sp., and Nannochloropsis oculata for use in tropical aquaculture. J.
Appl. Phycol., 3: 43–53.
Rezek, T. C., W. O. Watanabe, M. Harel & P. J. Seaton. 2010. Effects of dietary
docosahexaenoic acid (22:6n-3) and arachidonic acid (20:4n-6) on the growth,
survival, stress resistance and fatty acid composition in black sea bass
Centropristis striata (Linnaeus 1758) larvae. Aquacult. Res., 41:1302–1314.
Rivero-Rodríguez, S., A. R. Beaumont & M. C. Lora-Vilchi. 2007.The effect of
microalgal diets on growth, biochemical composition, and fatty acid profile of
Crassostrea corteziensis (Hertlein) juveniles. Aquaculture, 263: 199–210.
Rodriguez, C., J. A. Perez, M. Diaz, M. S. Izquierdo, H. Fernandez-Palacios & A.
Lorenzo. 1997. Influence of EPA/DHA ratio in rotifers on gilthead seabream
(Sparus aurata) larval development. Aquaculture, 150: 77–89.
Rodriguez, C., J. A. Perez. M. S. Izquierdo, J. R. Cejas, A. Bolanos & A. Lorenzo.
1996. Improvement of the nutritional value of rotifers by varying the type and
concentration of oil and the enriched period. Aquaculture, 147: 93-105.
Roessler, P. G. 1990. Environmental control of glycerolipid metabolism in
microalgae: commercial implications and future research directions. J. Phycol.,
26: 393–399.
Roman, M. R. 1984. Utilization of detritus by the copepod Acartia tonsa. Limnol.
Oceanogr., 29: 949-959.
Ruyter, B., C. Rùsjù, O. Einen & M. Thomassen. 2000. Essential fatty acids in
Atlantic salmon: efects of increasing dietary doses of n-6 and n-3 fatty acids
on growth, survival and fatty acid composition of liver, blood and carcass.
Aquacult. Nutr., 6: 119-127.
Sánchez-Estudillo, L. 2011. Alimento nutritivo, colorido y en movimiento: los
cultivos de apoyo en acuicultura. Ciencia y Mar, 43:55-60.

56
Santha, C. R. & D. M. Gatlin. 1991. Growth response and fatty acid composition of
channel catfish fry fed practical diets supplemented with menhaden fish oil.
Prog. Fish Cult., 53: 135–140.
Sargent, J. R., D. R Tocher & J. G. Bell. 2002. The lipids. 181–257 En: Halver, J. E.
& R. W. Hardy. (Eds.) Fish Nutrition, 3rd edition. Academic Press, San Diego,
California, Estados Unidos.
Sargent, J. R., J. G. Bell, M. V. Bell, R. J. Henderson & D. R. Tocher. 1995.
Requirement criteria for essential fatty acids. J. Appl. Ichthyol., 11: 183–198.
Sargent, J. R., J. G. Bell, L. A. McEvoy, D. R. Tocher & A. Estevez. 1999. The
essential fatty acid nutrition of early developing fish. Aquaculture, 177: 191-
199.
Sargent, J. R., L. A. McEvoy & J. G. Bell. 1997. Requirements, presentation and
sources of polyunsaturated fatty acids in marine fish larval feeds. Aquaculture,
155: 117-127.
Schipp, G. R., J. M. Bosmans & A. J. Marshall. 1999. A method for hatchery culture
of tropical calanoid copepods, Acartia spp. Aquaculture, 174: 81–88.
Seiffert, M. B., V. R. Cerqueira & L. Madureira. 2001. Effect of dietary (n-3) highly
unsaturated fatty acids (HUFA) on growth and survival of fat snook
(Centropomus parallelus, Pisces: Centropomidae) larvae during first feeding.
Braz. J. Med. Biol. Res., 4: 645–651.
Seychelles, L. H., C. Audet, R. Temblary, R. Fournier & F. Pernet. 2009. Essential
fatty acids enrichment of cultured rotifer (Brachionus plicatilis, Müller) using
frozen-concentrated microalgae. Aquacult. Nutr., 15: 431-439.
Shansudin, L., M. Yusof, A. Azis & Y. Shukri. 1997. The potential of certain
indigenous copepod species as live food for commercial fish larval rearing.
Aquaculture, 151: 351-356.
Shields, R. J. & C. W. Laidley. 2003. Intensively cultured paracalanid copepods. A
high quality diet for small tropical marine fish larvae. Glob. Aqua. Advoc., 6(6):
80-81.
Shields, R. J., J. G. Bell, F. S. Luizi, B. Gara, N. R. Bromage & J. R. Sargent. 1999.
Natural copepods are superior to enriched Artemia nauplii as feed for halibut
larvae (Hippoglossus hippoglossus) in terms of survival, pigmentation and
retinal morphology: relation to dietary essential fatty acids. J. Nutr., 129: 1186-
1194.

57
Shields, R. J., T. Kotani, A. Molnar, K. Marion, J. Kobashigawa & L. Tang. 2005.
Intensive cultivation of a subtropical paracalanid copepod, Parvocalanus sp.,
as a prey for small marine fish larvae. 209-223. En: Lee, C. S., P.J. O’Bryen &
N.H. Marcus (Eds.) Copepods in Aquaculture. Blackwell Publishing. Oxford.
Reino Unido.
Smith, T. I., D. C. McVey, W. E. Jenkins, M. R. Denson, L. D. Heyward, C. V.
Sullivan & D. L. Berlinsky. 1999. Broodstock management and spawning of
southern flounder, Paralichthys lethostigma. Aquaculture, 176: 87-99.
Sorgeloos, P., P. Dhert & P. Candreva. 2001. Use of the brine shrimp, Artemia
spp., in marine fish larviculture. Aquaculture, 200: 147–159.
Støttrup, J. G. & J. Jensen. 1990. Influence of algal diet on feeding and egg
production of the calanoid copepod Acartia Tonsa Dana. J. Exp. Mar. Biol.
Ecol., 141: 87-105.
Stottrup, J. G. & L. A. McEvoy. 2003. Live feeds in Marine aquaculture. Blackwell
Science Ltd. Reino Unido.
Støttrup, J. G. 2000. The elusive copepods: their production and suitability in
marine aquaculture. Aquac. Res., 31: 703–711.
Stottrup, J. G. 2006. Review on the status and progress in rearing copepods for
marine larviculture. Advantages and disvantages among calanoid,
harpacticoid and cyclipoid copepods. 62-83. En: Cruz Suárez, L. E., M. Denis-
Ricque, M. Tapia Salazar, M. G. Nieto-Lopez, D. A. Villareal-Cavazos, A. C.
Puello-Cruz & A. García-Ortega (Eds.) Avances en Nutrición Acuícola VIII.
Monterrey, Nuevo León, México.
Subagja J., J. Slembrouck, L. T. Hung & M. Legendre. 1999. Larval rearing of an
asian catfish, Pangasius hypophthalmus (Siluroidei, Pangasiidae): analysis of
precocious mortality and proposition of appropriate treatments. Aquat. Living
Res., 12: 37-44.
Sukenik, A., P. Zamora & Y. Carmeli. 1993. Biochemical quality of marine
unicellular algae with special emphasis on lipid composition Nannochloropsis
sp. Aquaculture, 117: 313-326.
Tanaka, H. 2015. Progression in artificial seedling production of Japanese eel
Anguilla japonica. Fish. Sci., 81:11-19.
Tocher, D. 2010. Fatty acid requirements in ontogeny of marine and freshwater fish.
Aquac. Res., 41: 717-732.

58
Tocher, D. R. & J. R. Sargent. 1984. Studies on triacylglycerol, wax ester and sterol
ester hydrolases in intestinal caeca of rainbow trout (Salmo gairdneri) fed diets
rich in triacylglycerols and wax esters. Comp. Biochem. Physiol., 77: 561-571.
Toledo, J. D., M. S. Golez, M. Doi & A. Ohno. 1999. Use of copepod nauplii during
early feeding stage of grouper Epinephelus coioides. Fish. Sci., 65: 390-397.
Tomoda, T., H. Furuita, M. Kamoshida, H. Kurogi, T. Shibuno, H. Tanaka & N.
Tezuka. 2014. Nutritional Enrichment Trials of the Minute Rotifer Proales
similis Using Commercial Materials. Fish.Technol., 6(2): 179–184.
Tonon, T., D. Harvey, T. Larson & I. Graham. 2003. Long chain polyunsaturated
fatty acid production and partitioning to triacylglycerols in four microalgae.
Phytochemistry, 61: 15–24.
Tucker, J. W. 1998. Marine Fish Culture. Kluwer Academic Publishers, Norwell
Massachusetts, Estados Unidos. 750p.
Tuncer, H. & R. M. Harrel. 1992. Essential fatty acid nutrition of larval striped bass
(Norone saxatilis) and palmetto bass (M.saxatilis× Mchrysops). Aquaculture,
101: 105–121.
Volkman, J. K., S. W. Jeffrey, P. D. Nichols, G. I. Rogers & C. D. Garland. 1989.
Fatty acid and lipid composition of 10 species of microalgae used in
mariculture. J. Exp. Mar. Biol. Ecol., 128: 219-240.
Watanabe, T. 1979. Nutritional quality of living feeds used in seed production of
fish. 7th Japan-Soviet Joint Symposium Aquaculture, September. Tokai
University Press, Tokyo. 49-57.
Watanabe, T. 1993. Importance of docosahexaenoic acid in marine larval fish. J
World Aquac. Soc., 24: 152–161.
Watanabe, T., C. Kitajima & S. Fujita.1983. Nutritional values of live organisms
used in Japan for mass propagation of fish: a review. Aquaculture, 34: 115-
143.
Wei, L., X. Huang & Z. Huang. 2014. Temperature effects on lipid properties of
microalgae Tetraselmis subcordiformis and Nannochloropsis oculata as
biofuel resources. Chin. J. Oceanol. Limnol., 33(1): 99-106.
Witt, U., G. Quantz & D. Kuhlmann.1984. Survival and growth of turbot larvae
Scophthalmus maximus L. reared on different food organisms with special
regard to long-chain polyunsaturated fatty acids. Aquacult. Eng., 3: 177-190.

59
Wullur, S., Y. Sakakura & A. Hagiwara. 2011. Application of the minute
monogonont rotifer Proales similis de Beauchamp in larval rearing of seven-
band grouper Epinephelus septemfasciatus. Aquaculture, 315: 355-360.
Yakupitiyage, A. 2013. On-farm feeding and feed management strategies in tropical
aquaculture. 361–376 En: Hasan, M. R. & M. B. New (Eds.) On-farm feeding
and feed management in aquaculture. FAO Fisheries and Aquaculture
Technical Paper No. 583. Rome, FAO.
Yongmanitchai, W. & O. P. Ward. 1989. Omega-3 fatty acids: alternative sources of
production. Process Biochem., 24: 117-125.
Yúfera, M. & M. J. Darías. 2007.Changes in the gastrointestinal pH from larvae to
adult in Senegal sole (Solea senegalensis). Aquaculture, 267: 94-99.
Zavala-Leal, I. 2007. Efecto de la temperatura, intensidad de luz, tipo y densidad de
presa en la eficiencia alimenticia durante la ontogenia inicial del huachinango
del Pacífico (Lutjanus peru). Tesis de maestría. CICIMAR-IPN. La Paz,
B.C.S., México. 52p.

60
ANEXOS.
Anexo1. Tipos y nombres comunes de ácidos grasos. Adaptado de Hauville et al., 2014.
Símbolo numérico Nombre Común
Ácidos grasos Saturados 12:0 Laurico 14:0 Mirístico 15:0 Pentadecanoico
16:0 Palmítico
17:0 Heptadecanoico
18:0 Esteárico
Ácidos grasos Mono-Insaturados 15:1 Cis-10 pentadecanoico 16:1n-7 Palmitoleico
18:1n-9 Oleico
18:1n-7 Vaccenico 20:1n-9 Eicosenoico Ácidos Poli-insaturados 16:2n-4 Hexadecadienoico 16:3n-4 Hexadecatrienoico 18:2n-6 Linoleico (LA)
18:3n-6 γ-Linolenico
18:3n-4 Octadecatrienoico
18:3n-3 α- Linolenico (ALA)
18:4n-3 Estearidonico 20:4n-6 Araquidónico (ARA) 20:5n-3 Eicosapentaenoico (EPA)
20:5n-6 ω6-Docosapentaenoico
22:6n-3 Docosahexaenoico (DHA)

61
Anexo 2. Composición de ácidos grasos de las microalgas (% del total de A.G). Se remarcan
los ácidos grasos araquidónico (ARA), eicosapentaenoico (EPA) y docosahexaenoico (DHA).
Nannochloropsis oculata
Tetraselmis suecica
Chaetoceros calcitrans
Isochrysis galbana
14:0 0.41 0.35 19.87 9.86 14:1n-8 0.75 0.00 0.32 0.24 15:0 0.03 0.03 0.45 0.18 16:0 20.46 16.40 8.88 5.82 16:1n-9 4.50 0.64 0.11 0.18 16:1n-5 0.14 0.12 0.44 0.04 16:1n-7 3.12 4.48 27.02 6.11 18:0 0.70 0.23 0.83 0.13 18:1n-9 12.63 7.25 0.52 8.26 18:1n-7 0.79 6.51 1.11 1.04 18:2n-6t 0.03 0.07 0.08 0.00 18:2n-6 (LA) 25.83 6.25 0.54 9.43 18:3n-6 1.00 0.63 0.40 1.61 18:3n-3 (ALA) 20.14 33.35 3.58 16.15 18:4n-3 0.21 8.63 0.20 17.15 20:0 0.01 0.05 0.06 2.00 20:3n-6 0.03 0.09 0.20 0.06 20:4n-6 (ARA) 4.85 1.48 3.38 0.10 20:5n-3 (EPA) 1.06 9.42 21.82 0.79 24:0 0.36 0.03 0.81 0.00 22:6n-3 (DHA) 0.02 0.09 4.41 19.42 Σ Saturados. 23.17 17.12 31.73 18.17 Σ Mono 23.01 22.13 32.57 16.35 Σ PUFA 53.82 60.75 35.70 65.48 Σ HUFA 6.22 20.13 30.91 38.16 Σ n3 21.73 51.69 30.01 53.68 Σ n6 32.30 17.68 5.90 28.96 (n-3)/(n-6) 0.67 2.92 5.09 1.85 DHA/EPA 0.02 0.01 0.20 24.68

62
Anexo 3. Composición de ácidos grasos (% del total de ácidos grasos) del rotífero Brachionus
plicatilis. Se remarcan los ácidos grasos araquidónico (ARA), eicosapentaenoico (EPA) y docosahexaenoico (DHA).
N. oculata N.oculata
+ Spresso-Selco®
T. suecica T. suecica
+ SpressoSelco®
ORIONE®
14:00 1.56 2.23 1.35 2.08 0.71 15:00 0.44 0.31 0.25 0.25 0.36 16:00 16.29 12.11 12.92 11.09 8.81 16:1n-9 2.24 0.62 0.85 0.40 0.62 16:1n-7 1.24 1.36 1.49 1.43 1.49 16:1n-5 0.11 0.09 0.11 0.09 0.09 18:00 4.35 2.13 3.29 1.93 2.93 18:1n-9 8.49 8.34 4.78 8.38 5.83 18:1n-7 1.75 1.11 5.05 1.99 2.37 18:2n-6 (LA) 21.89 15.55 9.30 13.31 15.80 18:3n-6 0.91 0.46 0.69 0.37 0.28 18:3n-3 (ALA) 16.54 6.93 25.34 7.67 10.36 18:4n-3 0.38 0.56 3.32 1.17 0.45 20:00 0.11 0.09 0.08 0.08 0.12 20:1n-9 2.16 1.06 3.04 1.21 1.92 21:00 0.03 0.37 0.01 0.38 0.01 20:3n-6 1.99 0.70 0.76 0.44 0.52 20:4n-6 (ARA) 5.98 2.40 2.38 1.62 1.95 20:5n-3 (EPA) 1.06 3.46 7.40 5.11 5.82 22:1n-9 0.95 0.38 1.21 0.45 0.83 22:4n-6 0.13 5.87 0.13 5.54 1.65 22:5n-3 0.20 0.12 1.50 0.06 0.26 22:5n-6 0.05 0.09 0.11 0.10 0.20 24:1n-9 2.19 0.59 0.80 0.53 0.88 22:6n-3 (DHA) 0.08 28.13 0.09 28.09 25.91 Σ Saturados 23.76 18.56 18.64 17.22 15.52 Σ Mono 21.78 14.61 20.73 15.76 16.69 Σ PUFA 54.46 66.83 60.64 67.02 67.79 Σ HUFA 10.37 41.98 21.84 44.04 38.17 Σ n3 19.79 39.85 39.71 42.75 44.14 Σ n6 35.04 27.54 24.25 25.44 24.10 (n-3)/(n-6) 0.56 1.45 1.64 1.68 1.83 DHA/EPA 0.08 8.13 0.01 5.50 4.45
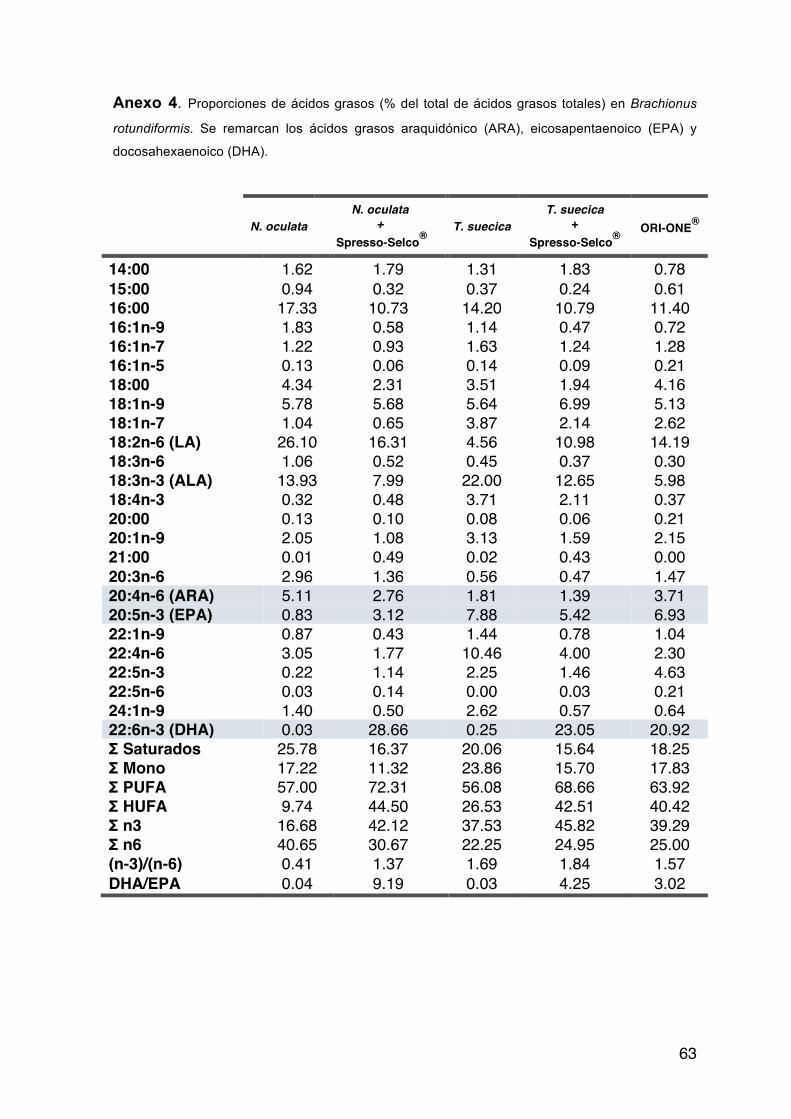
63
Anexo 4. Proporciones de ácidos grasos (% del total de ácidos grasos totales) en Brachionus
rotundiformis. Se remarcan los ácidos grasos araquidónico (ARA), eicosapentaenoico (EPA) y
docosahexaenoico (DHA).
N. oculata N. oculata
+ Spresso-Selco®
T. suecica T. suecica
+ Spresso-Selco®
ORI-ONE®
14:00 1.62 1.79 1.31 1.83 0.78 15:00 0.94 0.32 0.37 0.24 0.61 16:00 17.33 10.73 14.20 10.79 11.40 16:1n-9 1.83 0.58 1.14 0.47 0.72 16:1n-7 1.22 0.93 1.63 1.24 1.28 16:1n-5 0.13 0.06 0.14 0.09 0.21 18:00 4.34 2.31 3.51 1.94 4.16 18:1n-9 5.78 5.68 5.64 6.99 5.13 18:1n-7 1.04 0.65 3.87 2.14 2.62 18:2n-6 (LA) 26.10 16.31 4.56 10.98 14.19 18:3n-6 1.06 0.52 0.45 0.37 0.30 18:3n-3 (ALA) 13.93 7.99 22.00 12.65 5.98 18:4n-3 0.32 0.48 3.71 2.11 0.37 20:00 0.13 0.10 0.08 0.06 0.21 20:1n-9 2.05 1.08 3.13 1.59 2.15 21:00 0.01 0.49 0.02 0.43 0.00 20:3n-6 2.96 1.36 0.56 0.47 1.47 20:4n-6 (ARA) 5.11 2.76 1.81 1.39 3.71 20:5n-3 (EPA) 0.83 3.12 7.88 5.42 6.93 22:1n-9 0.87 0.43 1.44 0.78 1.04 22:4n-6 3.05 1.77 10.46 4.00 2.30 22:5n-3 0.22 1.14 2.25 1.46 4.63 22:5n-6 0.03 0.14 0.00 0.03 0.21 24:1n-9 1.40 0.50 2.62 0.57 0.64 22:6n-3 (DHA) 0.03 28.66 0.25 23.05 20.92 Σ Saturados 25.78 16.37 20.06 15.64 18.25 Σ Mono 17.22 11.32 23.86 15.70 17.83 Σ PUFA 57.00 72.31 56.08 68.66 63.92 Σ HUFA 9.74 44.50 26.53 42.51 40.42 Σ n3 16.68 42.12 37.53 45.82 39.29 Σ n6 40.65 30.67 22.25 24.95 25.00 (n-3)/(n-6) 0.41 1.37 1.69 1.84 1.57 DHA/EPA 0.04 9.19 0.03 4.25 3.02

64
Anexo 5. Composición de ácidos grasos (% del total de ácidos grasos) del rotífero Proales
similis. Se remarcan los ácidos grasos araquidónico (ARA), eicosapentaenoico (EPA) y
docosahexaenoico (DHA).
N. oculata N. oculata
+ Spresso-Selco®
ORI-ONE®
14:0 0.69 1.32 0.85 15:0 0.10 0.20 0.46 16:0 16.37 10.93 11.42 16:1n-9 1.48 0.68 0.44 16:1n-7 1.40 1.19 2.56 16:1n-5 0.12 0.08 0.08 18:0 3.04 2.09 3.74 18:1n-9 9.39 8.22 6.85 18:1n-7 1.33 1.11 8.90 18:2n-6 (LA) 33.96 23.00 16.33 18:3n-6 1.68 1.08 0.56 18:3n-3 (ALA) 15.21 6.28 6.33 18:4n-3 0.92 0.72 0.26 20:0 0.12 0.07 0.31 20:1n-9 3.00 2.16 3.48 21:0 0.02 0.27 0.02 20:3n-6 0.21 0.49 0.34 20:4n-6 (ARA) 5.15 3.20 2.64 20:5n-3 (EPA) 0.59 2.24 3.95 22:1n-9 0.12 0.43 0.67 22:4n-6 0.39 5.42 0.32 22:5n-3 0.12 0.64 1.69 22:5n-6 n/d 0.04 0.07 24:1n-9 1.20 0.96 2.22 22:6n-3 (DHA) 0.88 24.79 19.16 Σ Saturados 20.99 15.27 18.32 Σ Mono 19.17 15.63 28.10 Σ PUFA 59.84 69.10 53.58 Σ HUFA 8.48 37.54 28.89 Σ n3 17.75 34.90 31.77 Σ n6 43.01 34.92 22.07 (n-3)/(n-6) 0.41 1.00 1.44 DHA/EPA 1.48 11.05 4.85
n/d: No determinado.
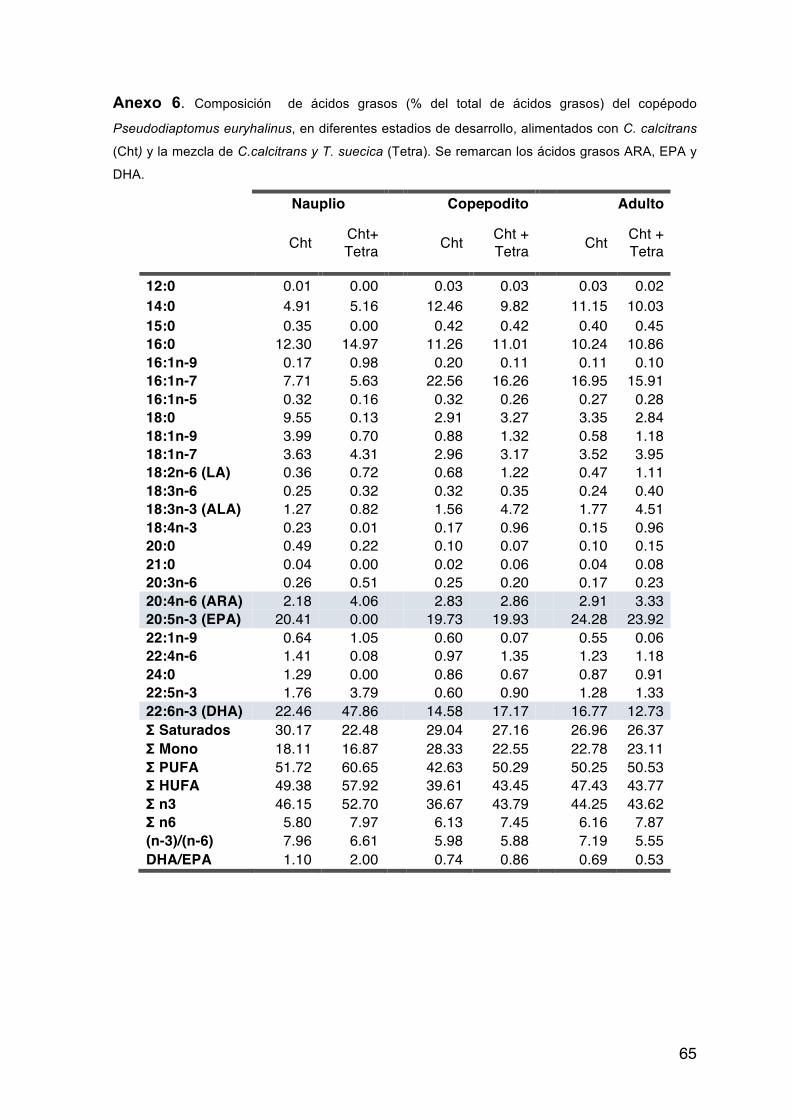
65
Anexo 6. Composición de ácidos grasos (% del total de ácidos grasos) del copépodo
Pseudodiaptomus euryhalinus, en diferentes estadios de desarrollo, alimentados con C. calcitrans
(Cht) y la mezcla de C.calcitrans y T. suecica (Tetra). Se remarcan los ácidos grasos ARA, EPA y
DHA.
Nauplio Copepodito Adulto
Cht Cht+ Tetra
Cht Cht +
Tetra
Cht Cht + Tetra
12:0 0.01 0.00 0.03 0.03 0.03 0.02 14:0 4.91 5.16 12.46 9.82 11.15 10.03 15:0 0.35 0.00 0.42 0.42 0.40 0.45 16:0 12.30 14.97 11.26 11.01 10.24 10.86 16:1n-9 0.17 0.98 0.20 0.11 0.11 0.10 16:1n-7 7.71 5.63 22.56 16.26 16.95 15.91 16:1n-5 0.32 0.16 0.32 0.26 0.27 0.28 18:0 9.55 0.13 2.91 3.27 3.35 2.84 18:1n-9 3.99 0.70 0.88 1.32 0.58 1.18 18:1n-7 3.63 4.31 2.96 3.17 3.52 3.95 18:2n-6 (LA) 0.36 0.72 0.68 1.22 0.47 1.11 18:3n-6 0.25 0.32 0.32 0.35 0.24 0.40 18:3n-3 (ALA) 1.27 0.82 1.56 4.72 1.77 4.51 18:4n-3 0.23 0.01 0.17 0.96 0.15 0.96 20:0 0.49 0.22 0.10 0.07 0.10 0.15 21:0 0.04 0.00 0.02 0.06 0.04 0.08 20:3n-6 0.26 0.51 0.25 0.20 0.17 0.23 20:4n-6 (ARA) 2.18 4.06 2.83 2.86 2.91 3.33 20:5n-3 (EPA) 20.41 0.00 19.73 19.93 24.28 23.92 22:1n-9 0.64 1.05 0.60 0.07 0.55 0.06 22:4n-6 1.41 0.08 0.97 1.35 1.23 1.18 24:0 1.29 0.00 0.86 0.67 0.87 0.91 22:5n-3 1.76 3.79 0.60 0.90 1.28 1.33 22:6n-3 (DHA) 22.46 47.86 14.58 17.17 16.77 12.73 Σ Saturados 30.17 22.48 29.04 27.16 26.96 26.37 Σ Mono 18.11 16.87 28.33 22.55 22.78 23.11 Σ PUFA 51.72 60.65 42.63 50.29 50.25 50.53 Σ HUFA 49.38 57.92 39.61 43.45 47.43 43.77 Σ n3 46.15 52.70 36.67 43.79 44.25 43.62 Σ n6 5.80 7.97 6.13 7.45 6.16 7.87 (n-3)/(n-6) 7.96 6.61 5.98 5.88 7.19 5.55 DHA/EPA 1.10 2.00 0.74 0.86 0.69 0.53

66
Anexo 7. Composición de ácidos grasos (% del total de ácidos grasos) del copépodo
Parvocalanus crassirostris, en diferentes estadios de desarrollo, alimentados con I. galbana (Iso) y
la mezcla de I. galbana, C. calcitrans (Cht) y T. suecica (Tetra). Se remarcan los ácidos grasos
ARA, EPA y DHA.
Nauplio I-II Nauplio III-IV Copepodito Adulto
Iso
Iso + Tetra+
Cht
Iso
Iso + Tetra+
Cht
Iso
Iso + Tetra+
Cht
Iso
Iso + Tetra +
Cht
14:0 5.60 3.42 6.22 6.13 5.83 6.14 5.84 4.73 16:0 9.82 13.94 8.46 8.71 13.62 17.48 10.53 12.83 16:1n-9 0.45 0.57 0.39 0.33 0.38 0.50 0.31 0.30 16:1n-7 3.42 3.52 3.57 4.18 4.69 5.91 4.05 5.56 18:0 1.85 2.34 1.86 1.74 2.32 2.76 1.98 2.29 18:1n-9 5.59 4.45 6.03 5.75 8.16 10.33 5.67 5.99 18:1n-7 1.32 3.10 1.72 1.39 1.56 2.26 1.46 2.63 18:2n-6 (LA) 8.30 4.97 9.30 9.62 7.67 5.91 8.56 6.81 18:3n-6 0.90 0.66 0.90 1.07 1.00 0.83 0.91 0.65 18:3n-3 (ALA) 7.67 10.65 8.33 10.88 9.31 7.41 8.45 12.20 18:4n-3 6.41 4.00 5.04 7.04 10.0 6.78 6.59 4.60 20:1n-9 0.18 0.56 0.16 0.23 0.27 0.38 0.20 0.44 20:3n-6 1.20 0.40 0.99 0.00 1.16 1.36 0.68 0.55 20:4n-6 (ARA) 0.19 1.43 0.63 0.38 0.21 1.19 0.29 0.87 20:5n-3 (EPA) 2.37 12.62 2.56 4.39 1.81 4.34 2.72 7.76 22:1n-9 0.22 0.12 0.30 0.26 0.26 0.10 0.16 0.11 22:4n-6 1.78 0.97 1.31 0.90 0.69 0.63 1.33 0.97 22:5n-3 0.18 0.98 0.17 0.38 0.23 0.78 0.33 1.34 22:6n-3 (DHA) 37.17 24.70 36.15 30.93 23.20 17.42 32.44 20.38 Σ Saturados 18.00 21.86 17.83 18.10 23.30 27.86 19.89 20.90 Σ Mono 13.02 14.10 14.31 13.85 16.97 21.35 13.35 16.67 Σ PUFA 68.98 64.04 67.86 68.05 59.73 50.78 66.76 62.42 Σ HUFA 49.68 45.94 47.23 45.27 38.83 33.38 46.12 38.48 Σ n3 54.55 53.89 52.88 54.32 46.04 38.26 52.01 49.41 Σ n6 20.85 14.15 20.01 20.77 23.78 19.31 21.33 17.61 (n-3)/(n-6) 2.62 3.81 2.64 2.62 1.94 1.98 2.44 2.81 DHA/EPA 15.71 1.96 14.12 7.05 12.83 4.01 11.95 2.63

67
Anexo 8. Composición de ácidos grasos (% del total de ácidos grasos) de Artemia sp. en
diferentes Se remarcan los ácidos grasos ARA, EPA y DHA
Nauplio recién nacido
Meta-nauplio II-III enriquecido Juvenil
14:00 0.48 0.82 0.83 16:00 9.03 7.42 10.39 16:1n-9 0.63 0.40 1.34 16:1n-7 1.99 1.38 10.67 18:00 4.01 2.85 7.00 18:1n-9 18.87 15.71 24.07 18:1n-7 5.20 3.37 12.81 18:2n-6 (LA) 5.98 9.35 9.38 18:3n-3 (ALA) 36.07 14.56 4.78 18:4n-3 6.36 1.97 1.37 20:4n-6 (ARA) 0.67 1.84 2.47 20:5n-3 (EPA) 1.78 5.87 5.00 22:4n-6 0.02 4.97 0.06 22:5n-3 0.01 0.04 0.02 24:1n-9 0.70 0.37 2.55 22:6n-3 (DHA) 0.15 22.59 0.43 Σ Saturados 14.55 12.72 20.75 Σ Mono 31.06 23.63 54.21 Σ PUFA 54.39 63.65 25.03 Σ HUFA 9.91 38.08 9.48 Σ n3 45.38 45.70 11.63 Σ n6 15.37 19.91 14.78 (n-3)/(n-6) 2.95 2.29 0.79 DHA/EPA 0.08 3.85 0.09