effect of metformin, rapamycin, and their combination on...
TRANSCRIPT

Research Article
Effect of Metformin, Rapamycin, and TheirCombination on Growth and Progression ofProstate Tumors in HiMyc MiceAchinto Saha1, Jorge Blando1,2, Lisa Tremmel1, and John DiGiovanni1,3
Abstract
In this study, we compared the effect of oral administration ofmetformin (MET) and rapamycin (RAPA) alone or in combina-tion on prostate cancer development and progression in HiMycmice. MET (250mg/kg body weight in the drinking water), RAPA(2.24mg/kg body weight microencapsulated in the diet), and thecombination inhibited progression of prostatic intraepithelialneoplasia lesions to adenocarcinomas in the ventral prostate(VP). RAPA and the combination were more effective than METat the doses used. Inhibition of prostate cancer progression inHiMyc mice by RAPA was associated with a significant reductionin mTORC1 signaling that was further potentiated by the com-bination ofMET andRAPA. In contrast, treatmentwithMET aloneenhancedAMPKactivation, but had little or no effect onmTORC1signaling pathways in the VP of HiMyc mice. Further analyses
revealed a significant effect of all treatments on prostate tissueinflammation as assessed by analysis of the expression of cyto-kines, the presence of inflammatory cells and NFkB signaling.MET at the dose used appeared to reduce prostate cancer pro-gressionprimarily by reducing tissue inflammationwhereasRAPAand the combination appeared to inhibit prostate cancer progres-sion in this mouse model via the combined effects on bothmTORC1 signaling as well as on tissue inflammation. Overall,these data support the hypothesis that blocking mTORC1 signal-ing and/or tissue inflammation can effectively inhibit prostatecancer progression in a relevant mouse model of human prostatecancer. Furthermore, combinatorial approaches that target bothpathwaysmay be highly effective for prevention of prostate cancerprogression in men. Cancer Prev Res; 8(7); 597–606. �2015 AACR.
IntroductionWorldwide, prostate cancer is the second most commonly
diagnosed noncutaneous cancer in men and the sixth mostcommon cause of death (1). In the United States, it is the mostfrequently diagnosed noncutaneous cancer and the second lead-ing cause of cancer-related mortality in this population (2). Anestimated 233,000 new cases will have been diagnosed in 2014 inthe United States with an estimated 29,480 deaths (3). Althoughthemedian age for diagnosis of prostate cancer is 66 (2), the onsetof preclinical diseasemay occur in adults as early as 30 years of age(4). Because there can be a considerable time for the disease toprogress to clinically evident cancer, there is ample opportunityfor chemopreventive strategies to be applied for the successfulmanagement of prostate cancer (reviewed in refs. 4, 5). A numberof agents, including sulforaphane, curcumin, green tea (orEGCG), resveratrol, and various NSAIDS have shown potential
chemopreventive effects in vivo in either animal models of pros-tate cancer or in clinical studies (6–9). Many of these agents targetinflammatory signaling pathways including STAT3 and NFkB, inaddition to other cellular signaling pathways associated withprostate cancer development and progression (10–12).
Metformin (MET) is adrugwidely used for the treatment of typeII diabetes (13) and its use has been shown to be associated withreduced cancer incidence andmortality (14). Several recent in vitroand animal studies also support the anticancer activity of MET indifferent cancer types, including prostate cancer (15–17).MET is apotent activator of AMPK, which is a serine/threonine proteinkinase that acts as a central metabolic sensor involved in cellularenergy homeostasis (18). AMPK activation also leads to inhibi-tion of mTORC1 signaling through phosphorylation of tuberoussclerosis 2. It is now well documented that mTORC1 signalingplays an important role in oncogenic processes (19), and dysre-gulation of this signaling pathway has been reported in variouscancers, including prostate cancer (20, 21).
Rapamycin (RAPA) is a macrolide used clinically as an immu-nosuppressant in organ transplant patients and to treat autoim-mune disorders (22). It acts as a potent inhibitor of mTORC1 bybinding with immunophilin FK506-binding protein (FKBP12;ref. 23). Data in the literature also suggest that RAPA has potentcancer chemopreventive properties in anumber ofmousemodels,including mouse models of prostate cancer (24–26). In addition,data compiled from more than 30,000 kidney transplant recipi-ents found that the use of mTOR inhibitors as maintenanceimmune-suppressive therapy produced a remarkable reductionin non-melanoma skin cancer incidence compared with cyclo-sporine A, a calcineurin inhibitor (27). A dramatic decrease in theincidence of skin malignancies was also observed in transplant
1Division of Pharmacology and Toxicology, Dell Pediatric ResearchInstitute, The University of Texas at Austin, Austin, Texas. 2Immuno-pathology Laboratory Immunotherapy Platform, The University ofTexas MD Anderson Cancer Center, Houston, Texas. 3Department ofNutritional Sciences, Dell Pediatric Research Institute, The Universityof Texas at Austin, Austin, Texas.
Note: Supplementary data for this article are available at Cancer PreventionResearch Online (http://cancerprevres.aacrjournals.org/).
Corresponding Author: John DiGiovanni, The University of Texas at Austin,1400 Barbara Jordan Boulevard, Austin, TX 78723. Phone: 512-495-4726; Fax:512-495-4945; E-mail: [email protected]
doi: 10.1158/1940-6207.CAPR-15-0014
�2015 American Association for Cancer Research.
CancerPreventionResearch
www.aacrjournals.org 597
Research. on May 26, 2019. © 2015 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst April 23, 2015; DOI: 10.1158/1940-6207.CAPR-15-0014

patients who were converted to mTOR inhibitors after 3 monthsof treatment with cyclosporine A (28).
In this study, we evaluated the ability of orally administeredMET, RAPA, and a combination of MET þ RAPA on prostatecancer development and progression in HiMyc mice (29). Inthis model, overexpression of c-Myc in the prostate is directedvia the ARR2Pb probasin promoter resulting in the develop-ment of prostatic lesions that share molecular and histopath-ologic features with human prostate tumors (29). Prostaticepithelial expression of c-Myc in the dorsolateral prostate(DLP), ventral prostate (VP), and anterior prostate (AP) lobesresults in complete penetrance of PIN as early as 2 to 4 weeks ofage, which progressed to locally invasive adenocarcinomaswithin 6 to 12 months of age (29). MET, RAPA, and thecombination inhibited progression of prostatic intraepithelialneoplasia (PIN) lesions to adenocarcinomas in the VP of HiMycmice. RAPA and the combination were more effective than METat the doses used. On the basis of mechanistic studies per-formed, these results suggest that targeting mTORC1 togetherwith inflammatory signaling pathways may be an effectivestrategy for prevention of prostate cancer progression.
Materials and MethodsReagents
RPMI-1640 and FBSwere obtained fromLife Technologies.METand RAPA were purchased from Sigma-Aldrich and LC laborato-ries, respectively. Antibodies against AMPK, pAMPKThr172, mTOR,pmTORSer2448, p70S6K, pp70S6KThr389, S6 ribosomal protein (S6Ribo), pS6 RiboSer235/236, pS6 RiboSer240/244, pNFkBp65Ser536,ULK1, pULK1Ser555, cMyc, GAPDH, cyclin D1, and PARP werepurchased from Cell Signaling Technology. Antibodies for p27,pNFkBp50Ser337, and IkBa were from Santa Cruz Biotechnologyand b-actin from Sigma-Aldrich. Antibodies for CD45 and CD3 (Tlymphocytes) were obtained from Abcam.
Study designHiMyc mice (29) were obtained from the NIHMMRRC on an
FVB/N genetic background and mice were bred in-house for thecurrent experiments. All diets were purchased from Test Dietsand mice were fed chow-based diet containing either 14 mg/kgmicroencapsulated RAPA [RAPA diet equivalent to 2.24 mg/kgbody weight or 14 mg/kg eudragit (control diet; ref. 30)]. Micewere placed on control diet at 4 to 5 weeks of age for a 1-weekequilibration period and then randomized into the followingdietary groups (n¼ 18) for the duration of the study: (i) controldiet fed ad libitum; (ii) RAPA diet fed ad libitum; (iii) control dietfed ad libitum þ MET (250 mg/kg BW) in drinking water; and(iv) RAPA diet fed ad libitumþMET in drinking water. The bodyweight and food consumption were determined every 2 weeks.Groups of mice were terminated by CO2 asphyxiation, and thegenitourinary tract, including urinary bladder, prostate lobes(AP, VP, and DLP), seminal vesicle and urethra, was taken after18 weeks of treatment for histopathologic diagnosis and IHCand immunofluorescence (IF) analyses. An additional set of 10mice from each diet group was used for protein and RNAanalyses (isolated after 10 weeks of treatment). For all thestudies, mice were housed in suspended polycarbonate cageson autoclaved hardwood bedding at room temperatures of20�C to 22�C, relative humidity of 60% to 70%, and 14/10-hour light/dark cycle.
Histologic analysesFor histologic analyses, the genitourinary tract was removed,
fixed in 10% formalin, embedded in paraffin, and transverselysectioned. Sections of 4 mm were stained with hematoxylin andeosin for histopathologic diagnosis as previously described (31).Under the experimental conditions used, HiMyc mice developedlesions primarily in the VP and DLP, with fewer lesions in the AP.Furthermore, at approximately 6months of age,>90%of themicedeveloped invasive tumors primarily in the VP. In contrast, at thesame time point, the number of invasive tumors in the DLP andAP was significantly lower. Thus, the current analyses focused onthe VP that displayed a more homogeneous and consistentdevelopment of the lesions from hyperplasia to invasive adeno-carcinomas within the 6-month time frame of these experiments.
Cell cultureLNCaP cells were purchased from the ATCC. These cells were
maintained in RPMI-1640 medium with 10% FBS. The murineprostate tumor cell line, HMVP2, was derived from the VP of oneyear old HiMyc transgenic mice as previously described andcultured in RPMI-1640 medium containing 10% FBS (32). Celllines were authenticated by genetic biomarkers. Mycoplasma testwas performed by PCR amplification (Applied Biological Materi-als Inc.) and 40,6-diamidino-2-phenylindole staining. All cellswere cultured in 95% air and 5% CO2 at 37�C.
Western blottingLevels of phosphorylated and total proteins were measured by
Western blot analysis with slight modifications of previous meth-ods (31). Briefly, LNCaP and HMVP2 cells were treated with MET(0.5 mmol/L), RAPA (1.0 nmol/L), or MET (0.5 mmol/L)þRAPA(1.0 nmol/L) for 24 hours. After incubation, cells were lysed inRIPA buffer. Alternatively, individual VP lobes were excised,crushed into powder under liquid nitrogen and lysed in RIPAbuffer. Proteinswere separated by SDSPAGE gel and transferred tonitrocellulose membranes. After blocking for 1 hour, the mem-branes were probed with specific primary antibodies overnight at4�C. Following secondary antibody (GE Healthcare) incubation,membranes were visualized using a commercial chemilumines-cent detection kit (Pierce Biotechnology). Except where noted, allresults were confirmed in at least three independent experiments.
Quantitative real-time PCRTotal RNA was isolated from the individual VP lobes by using
the Qiagen RNeasy Mini Kit (Qiagen) according to the manu-facturer's protocol and subjected to reverse-transcription as pre-viously described (31). mRNA levels of genes were quantitativelydetermined by quantitative real-time PCR (qPCR) using the Viia7Real-Time PCR System (Applied Biosystems) with SYBR GreenMaster Mix (Qiagen). The relative abundance of the mRNA wasnormalized against glyceraldehyde 3-phosphate dehydrogenase(GAPDH) mRNA for quantitative evaluation.
IHC and IF analyses and toluidine blue stainingIHC analyses were performed on formalin-fixed, paraffin-
embedded prostate tissue sections (31). Briefly, tissue sectionswere deparaffinized with serial incubation andwashing in xylene,100% to 70% ethanol and water followed by antigen unmaskingwith citrate buffer. The samples were then blocked for 1 hour atroom temperature and incubated with primary antibodies over-night at 4�C. Proteins were detected with biotinylated secondary
Saha et al.
Cancer Prev Res; 8(7) July 2015 Cancer Prevention Research598
Research. on May 26, 2019. © 2015 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst April 23, 2015; DOI: 10.1158/1940-6207.CAPR-15-0014

antibodies, followed by peroxidase-conjugated avidin/biotin(Vector Laboratories) and 3,30-diaminobenzidin substrate(Dako), and then visualized with light microscopy. Quantifica-tion of IHC analyses for pS6Ribo was scored on the basis of a fourpoint scale criteria from negative (0þ) to intense (3þ) asdescribed elsewhere (33). For IF staining (34), paraffin-embeddedprostate tissue sections were detected with fluorochrome-conju-gated secondary antibodies and visualized using an OlympusBX60 fluorescence microscope. Toluidine blue staining was usedto identify mast cells as described elsewhere (35). Briefly, tissuesections were deparaffinized, stained with 0.1% toluidine bluesolution for 1 minute, washed with water, dehydrated with 70%to 100% ethanol and xylene solution. Mast cells were visualizedand counted with light microscopy.
Statistical analysesStatistical analyses were performed for differences in incidence
of prostate lesions between groups using the Fisher exact test.Comparison of changes in protein, protein phosphorylation, andmRNA levels was accomplished using one-way ANOVA followedby Bonferroni multiple comparison tests. Significance was set atP < 0.05.
ResultsEffect of MET, RAPA, and MET þ RAPA on prostate cancerprogression in HiMyc mice
The effect of oral administration of MET, RAPA, and MET þRAPA on prostate cancer progression in HiMyc mice is shownin Fig. 1A. The incidence of hyperplasia and low-grade PIN(lgPIN) in the VP of all the treatment groups was similar after18 weeks of treatment. However, treatment with MET, RAPA, andMETþ RAPA slightly decreased the incidence of hgPIN (Fig. 1A),although this was not statistically significant. The incidence ofboth in situ adenocarcinomas and locally invasive adenocarci-nomas was significantly reduced by treatment with both RAPAand MET þ RAPA. In this regard, RAPA treatment alonedecreased in situ adenocarcinomas by 41% (P < 0.02) andcompletely suppressed formation of locally invasive adenocar-cinomas (P < 0.001), compared with the control group. Thecombination of MET þ RAPA significantly (P < 0.005) decreasedin situ adenocarcinomas by 51% (Fig. 1A) and also completelysuppressed the formation of locally invasive adenocarcinomas (P< 0.001). MET given alone reduced the incidence of in situadenocarcinomas by 27% (not statistically significant) and sig-nificantly decreased the incidence of invasive adenocarcinomasby 50% (P < 0.01; Fig. 1A). The decreases in in situ adenocarci-nomas and adenocarcinomas by the combination were notstatistically significant when compared with the RAPA onlygroup, but the decrease in adenocarcinomas was statisticallysignificant (P < 0.01), when compared with the MET-only treatedgroup. Representative lesions for each of the treatment groupsare shown in Fig. 1B. All mice were closely monitored through-out the experimental period and did not reveal any signs oftoxicity. Furthermore, there were no significant differences inbody weight (Supplementary Fig. S1A) or food consumption(Supplementary Fig. S1B) across the different treatment groupsduring the course of the experiment. Thus, oral administration ofMET, RAPA, and the combination of MET þ RAPA significantlydecreased the incidence of malignant prostate cancer lesions inthe VP of HiMyc mice without apparent toxicity.
Effect of MET, RAPA, and METþ RAPA on AMPK and mTORC1signaling in VP of HiMyc mice
As shown in Fig. 2A, treatment with MET led to increasedphosphorylation of AMPKThr172, and this was greater in micethat received MET þ RAPA. As expected, RAPA alone had littleor no effect on AMPK activation. In contrast, MET alone hadlittle or no effect on mTORC1 signaling (as assessed by phos-phorylation of both p70S6K and S6 Ribo; Fig. 2B). However,treatment with RAPA alone produced a significant reduction ofphospho-p70S6KThr389 and phospho-S6 RiboSer240/244. Inter-estingly, the combination of MET þ RAPA significantlydecreased phospho-mTORSer2448 as well as levels of phos-pho-p70S6K and phospho-S6RiboSer240/244 (Fig. 2A and 2B).The effect on phospho-S6Ribo was further confirmed by IHCstaining of VP sections (Fig. 2C and D). These data indicate thatthe ability of RAPA and METþ RAPA to inhibit prostate cancerprogression in HiMyc mice correlated with significant inhibi-tion of mTORC1 signaling whereas the inhibitory effect of METon prostate cancer progression did not.
Figure 1.Effect of MET, RAPA, and MET þ RAPA on incidence of prostate lesions inHiMyc mice. A, HiMyc mice were administered MET (250mg/kg body weightin drinking water), RAPA (14 mg/kg microencapsulated in the diet), or theircombination for 18 weeks starting at 4 to 5 weeks of age. The incidence ofprostate lesions was quantified as detailed in Materials and Methods (n¼ 18).Significantly different from control; �, P < 0.02; �� , P < 0.01; ��� , P < 0.005;���� , P < 0.001. Significantly different from MET only group; x, P < 0.01. B,representative lesions in the VP of HiMyc mice. Lesions in mice from thecontrol group (a–c); a, low-grade PIN (lgPIN), high-grade PIN (hgPIN); b,in situ adenocarcinoma (AC); c, invasive AC. Lesions in mice from MET onlygroup (d–f); d, lgPIN, hgPIN; e, in situ AC; f, invasive AC. Lesions in mice fromRAPA only group (g–i); g, lgPIN; h, hgPIN; i, in situ AC. Lesions in mice fromMET þ RAPA group (j–l); j, lgPIN; k, hgPIN; l, in situ AC.
Anticancer Activity of Metformin and Rapamycin Combination
www.aacrjournals.org Cancer Prev Res; 8(7) July 2015 599
Research. on May 26, 2019. © 2015 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst April 23, 2015; DOI: 10.1158/1940-6207.CAPR-15-0014

Effect of MET, RAPA, and METþ RAPA on AMPK and mTORC1signaling in cultured mouse and human prostate cancer cells
Similar to the results seen in vivo after administration of thecompounds to HiMyc mice, treatment of HMVP2 cells with MET(0.5mmol/L) alone led to significant activation of AMPK, but didnot produce significant effects onmTORC1 phosphorylation andmTORC1 downstream signaling (again measured by phosphor-ylation of p70S6K and S6 Ribo; Fig. 3A and B). However, bothRAPA (1.0 nmol/L) and the combination ofMET (0.5mmol/L)þ
RAPA (1.0 nmol/L) significantly increased the phosphorylation ofAMPK and decreased the phosphorylation of mTORSer2448,p70S6KThr389, S6 RiboSer240/244 (again see Fig. 3A and B). Asshown in Supplementary Fig. S2A and S2B, the combination ofMET þ RAPA also significantly increased the levels of phospho-ULK1Ser555 and p27. However, no increases were observed inapoptosis marker, cleaved PARP with MET, RAPA, or the combi-nation in these cells. Thus, these results examining the effects ofthe agents on AMPK and mTORC1 signaling in cultured HMVP2
Figure 2.Effect ofMET, RAPA, andMETþRAPAon AMPK and mTORC1 signaling.Protein lysates were prepared fromthe VP of HiMyc mice (pooled sampleof n ¼ 4–5 mice/group) treated withMET, RAPA, or METþRAPA for 10weeks and Western blot analyseswere performed for the followingproteins: A, pAMPKThr172, AMPK,pmTORSer2448, mTOR, and GAPDH.B, pp70S6KThr389, p70S6K, pS6RiboSer240/244, S6 Ribo, and GAPDH.Quantification of Western blotanalyses is shown to the right in A andB and values represent mean � SEMof at least three independentexperiments; a, significantly different(P < 0.05) from the control group; b,significantly different (P < 0.05) fromthe RAPA group; and c, significantlydifferent (P < 0.05) from the METgroup. C, representative IHC analysesof pS6 RiboSer235/236 in sections of VPfrom HiMyc mice treated 18 weekswith MET, RAPA, or MET þ RAPA.D, quantitation of IHC analyses of pS6RiboSer235/236; a, significantly different(P < 0.05) from the control group; andc, significantly different(P < 0.05) from the MET group.
Saha et al.
Cancer Prev Res; 8(7) July 2015 Cancer Prevention Research600
Research. on May 26, 2019. © 2015 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst April 23, 2015; DOI: 10.1158/1940-6207.CAPR-15-0014
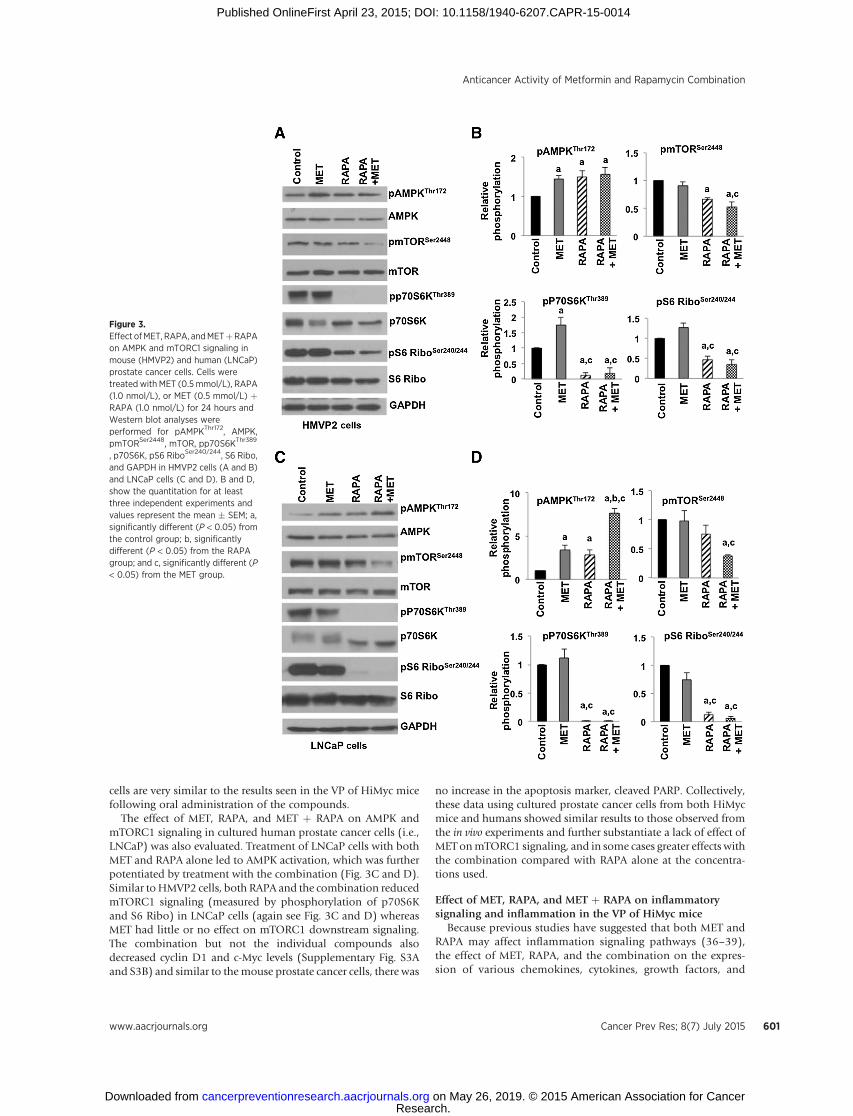
cells are very similar to the results seen in the VP of HiMyc micefollowing oral administration of the compounds.
The effect of MET, RAPA, and MET þ RAPA on AMPK andmTORC1 signaling in cultured human prostate cancer cells (i.e.,LNCaP) was also evaluated. Treatment of LNCaP cells with bothMET and RAPA alone led to AMPK activation, which was furtherpotentiated by treatment with the combination (Fig. 3C and D).Similar toHMVP2 cells, both RAPA and the combination reducedmTORC1 signaling (measured by phosphorylation of p70S6Kand S6 Ribo) in LNCaP cells (again see Fig. 3C and D) whereasMET had little or no effect on mTORC1 downstream signaling.The combination but not the individual compounds alsodecreased cyclin D1 and c-Myc levels (Supplementary Fig. S3Aand S3B) and similar to themouse prostate cancer cells, there was
no increase in the apoptosis marker, cleaved PARP. Collectively,these data using cultured prostate cancer cells from both HiMycmice and humans showed similar results to those observed fromthe in vivo experiments and further substantiate a lack of effect ofMET onmTORC1 signaling, and in some cases greater effects withthe combination compared with RAPA alone at the concentra-tions used.
Effect of MET, RAPA, and MET þ RAPA on inflammatorysignaling and inflammation in the VP of HiMyc mice
Because previous studies have suggested that both MET andRAPA may affect inflammation signaling pathways (36–39),the effect of MET, RAPA, and the combination on the expres-sion of various chemokines, cytokines, growth factors, and
Figure 3.Effect ofMET, RAPA, andMETþRAPAon AMPK and mTORC1 signaling inmouse (HMVP2) and human (LNCaP)prostate cancer cells. Cells weretreated with MET (0.5 mmol/L), RAPA(1.0 nmol/L), or MET (0.5 mmol/L) þRAPA (1.0 nmol/L) for 24 hours andWestern blot analyses wereperformed for pAMPKThr172, AMPK,pmTORSer2448, mTOR, pp70S6KThr389
, p70S6K, pS6 RiboSer240/244, S6 Ribo,and GAPDH in HMVP2 cells (A and B)and LNCaP cells (C and D). B and D,show the quantitation for at leastthree independent experiments andvalues represent the mean � SEM; a,significantly different (P < 0.05) fromthe control group; b, significantlydifferent (P < 0.05) from the RAPAgroup; and c, significantly different (P< 0.05) from the MET group.
Anticancer Activity of Metformin and Rapamycin Combination
www.aacrjournals.org Cancer Prev Res; 8(7) July 2015 601
Research. on May 26, 2019. © 2015 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst April 23, 2015; DOI: 10.1158/1940-6207.CAPR-15-0014

angiogenesis factors associated with inflammation was exam-ined in RNA samples isolated from the VP of HiMyc mice onthe different treatment regimens. As shown in Fig. 4A, treat-ment with MET or RAPA alone significantly decreased themRNA levels of IL1a and IL1b (P < 0.05). MET alone alsosignificantly decreased mRNA levels of IL23, TNFa, andCXCL12 (P < 0.05). The combination of MET þ RAPA appearedto further decrease the expression of these inflammatory med-iators compared with RAPA or MET alone although statisticalsignificance was achieved only for CCL5 (compared with eitherMET or RAPA) and for IL23 and CXCL12 (compared withRAPA). MET treatment alone also significantly (P < 0.05)decreased the expression of both VEGFB and IGF-1 (Fig. 4Band C) whereas treatment with RAPA had little or no effect onmRNA levels for these genes. Again, the combination treatmentproduced greater inhibition of IGF-1, IGF-1R, and VEGFBcompared with RAPA and produced stronger inhibition ofHIF1a and VEGFA compared with both agents.
As shown in Fig. 5, IF staining of prostate tissues fromuntreated HiMyc mice on the control diet showed significantinfiltration of inflammatory cells (measured by CD45þ cells)whereas HiMyc mice treated with MET, RAPA, and MET þRAPA had very few CD45þ cells (Fig. 5A and D). Furtheranalyses confirmed that mice treated with MET, RAPA, andMETþRAPA had significant decreases in the number of both Tlymphocytes (CD3þ cells; Fig 5B and E) and mast cells (O-toluidine blue stained cells; Fig. 5C and F) in the VP comparedwith the control group. Thus, both MET and MET þ RAPAsignificantly reduced expression of many of the genes exam-ined whereas RAPA treatment alone only significantly reducedexpression of IL1a/b. However, all treatments reduced tissueinflammation as assessed by the presence of several types ofinflammatory cells.
Effect of MET, RAPA, andMETþ RAPA onNFkB signaling in VPof HiMyc mice
As shown in Fig. 6A, top,MET, RAPA, andMETþRAPA stronglyinhibited the phosphorylation of the p50 subunit of NFkB asmeasured by IF staining of tissues from the VP of treated mice.Because IkBa acts as an inhibitor of NFkB signaling (40), stainingfor IkBawas also performed in tissues from theVPof treatedmice.As shown in the bottom of Fig. 6A, the level of IkBawas higher inthe VP of HiMyc mice treated with MET, RAPA, and METþ RAPAcompared with the mice in the control group. The status ofpNFkBp65Ser536 was also evaluated byWestern blot analysis (Fig.6B). The level of pNFkBp65Ser536 in VP of HiMyc mice wasdecreased by all three treatments, with the greatest decreases seenwith MET and MET þ RAPA treatment compared with the RAPAonly treated groups. Collectively, these data indicate that treat-ment with MET, RAPA, and MET þ RAPA significantly decreasedNFkB signaling in the VP of HiMyc mice.
DiscussionThis study was designed to evaluate the effect of oral admin-
istration ofMET, RAPA, and their combination on prostate cancerdevelopment and progression in an established mouse model ofprostate cancer. All treatments inhibited prostate cancer progres-sion in HiMyc mice, but had little or no effects on the develop-ment of lgPIN and hgPIN lesions at approximately 6 months ofage (i.e., following 18 weeks of treatment). The effect of MET onprostate cancer progression at the doses used in this study wasobserved primarily on the incidence of locally invasive adeno-carcinomas whereas both RAPA and MET þ RAPA producedstatistically significant decreases in in situ adenocarcinomas aswell as complete inhibition of locally invasive adenocarcinomasin the VPofHiMycmice. BothMET andRAPAhave been shown to
Figure 4.Effect ofMET, RAPA, andMETþRAPAon inflammation and angiogenesismarkers. RNA was isolated from theVPofHiMycmice treated for 10weeks.qPCR analyses of gene expression isshown for CCL5, IL1a, IL1b, IL6, IL23,TNFa, CXCL12 (A); HIF1a, VEGFA,VEGFB, VEGFC (B); and IGF-1, IGF-1R(C). Values represent themean� SEMof at least three independentexperiments; a, significantly different(P < 0.05) from the controlgroup; b, significantly different(P < 0.05) from the RAPA group; andc, significantly different (P < 0.05)from the MET group.
Saha et al.
Cancer Prev Res; 8(7) July 2015 Cancer Prevention Research602
Research. on May 26, 2019. © 2015 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst April 23, 2015; DOI: 10.1158/1940-6207.CAPR-15-0014

decrease prostate cancer cell growth in culture and to inhibitgrowth of prostate cancer cells in xenograft models either alone orin combinationwith other agents (41–45). In addition, bothMETand RAPA have been shown to inhibit development and/orprogression of tumors in a number of mouse primary tumormodels (16, 46, 47). The current results clearly demonstrate theefficacy of both of these compounds and the potential forincreased efficacy with the combination for inhibiting prostatecancer progression in this mouse model. Although the combina-tion of MET þ RAPA was not more effective than RAPA alone interms of effects on tumor incidence, the combination was moreeffective in some cases at reducing relevant signaling pathwaysand gene-expression changes. Therefore, further studies usingdifferent dosing combinations could reveal potential additive orsynergistic effects of this combination. Collectively, these datashow that long-term oral dosing of MET, RAPA, andMETþ RAPAsignificantly reduces prostate cancer progression in HiMyc mice.
Recently, the effect of short-term treatment with MET (200mg/kg body weight in drinking water for 4 weeks) was evalu-ated in HiMyc mice starting at 5 weeks of age (48). This short-term treatment was reported to significantly inhibit formationof both PIN as well as prostate cancer lesions through amechanism involving downregulation of c-myc. Although treat-
ment-related effects on the development of PIN lesions werenot observed in this study, under the experimental conditionsused, MET given alone significantly inhibited progression ofPIN lesions to prostate cancer. In other studies using HiMycmice, a short-term treatment with RAD001 (given daily by oraladministration for 14 days) did not revert mPIN lesions in thesemice even though mTORC1 was inhibited by this treatmentregimen (49). These data suggested that development of mPINlesions in HiMyc mice did not depend on mTORC1 signaling.These data with more long-term treatment using RAPA areconsistent with this earlier study, but also demonstrate adependence of tumor progression in HiMyc mice, at least inpart, on mTORC1 signaling. Finally, short-term treatment withRAPA (given i.p. daily for 14 days) in 12- to 14-month-oldPTENþ/� mice (C57BL/6 genetic background) was shown toreduce mTORC1 signaling and to inhibit proliferation of pros-tate lesions in these mice. A reduction in incidence of alldetectable lesions was seen, but these reductions in incidencewere not statistically significant due to the low numbers ofanimals used in that study (50). These data clearly demonstratethat RAPA given in the diet in a microencapsulated (and morebioavailable) form for an extended period of time inhibited theprogression of prostate cancer in HiMyc mice.
Figure 5.Effect ofMET, RAPA, andMETþRAPAon inflammatory cells in VP of HiMycmice. Representative stained sectionsof the VP of HiMyc mice treated 18weeks with either a control diet ordiets containing MET, RAPA, or RAPAþ MET. A, sections stained with CD45antibody; and D shows thequantitation. B, sections stained withCD3 antibody for T lymphocytes; andE shows the quantitation. C, sectionsstained with o-toluidine blue for mastcells; and F shows the quantitation.Cells were quantified on the basis ofthe average number of positive cellsfrom at least 12 to 16 (4 fields/mouse)randomly selected microscopic fieldsper diet group; a, significantlydifferent (P < 0.05) from the controlgroup.
Anticancer Activity of Metformin and Rapamycin Combination
www.aacrjournals.org Cancer Prev Res; 8(7) July 2015 603
Research. on May 26, 2019. © 2015 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst April 23, 2015; DOI: 10.1158/1940-6207.CAPR-15-0014

Anumber of experiments were performed to examine potentialmechanisms for the effects ofMET, RAPA, and the combination ofMET þ RAPA on prostate cancer progression. As shown in Fig. 2,treatment with RAPA significantly inhibited mTORC1 down-stream signaling to p70S6K and S6 Ribo in the VP of HiMycmice. Importantly, the addition of MET with RAPA further poten-tiated the inhibition of mTORC1 signaling. Further studies usingbothmouse and human prostate cancer cells in culture confirmedthese effects on mTORC1 signaling (Fig. 3). Notably, MET givenalonehad little or no effects onmTORC1 signaling in VPofHiMycmice, although AMPK activation was observed with this com-pound (Figs. 2 and 3). Thus, although MET is known to inhibitmTORC1 through both AMPK-dependent and -independentmechanisms (51), its ability to inhibit prostate cancer progressioncould not be attributed to inhibition ofmTORC1 signaling. Theseobservations were also confirmed in cultured mouse and humanprostate cancer cells whereMET, at relatively high concentrations,failed to significantly inhibit mTORC1 downstream signaling.
The lack of effect of MET on mTORC1 downstream signalingthrough p70S6K and S6 Ribo led to consideration of additionalmechanisms for its inhibitory effects on prostate cancer progres-sion. Previous studies from our laboratory demonstrated a stronglocal anti-inflammatory effect of low-dose RAPA when giventopically to mice during the process of skin tumor promotion(37). Furthermore, other studies have shown anti-inflammatoryeffects of RAPA (36), andmore recently anti-inflammatory effectsof MET (38, 39). Therefore, markers of inflammation were eval-uated in the VP of HiMyc mice treated with MET, RAPA, andMETþ RAPA. As shown in Fig. 4, treatment with MET, RAPA, and thecombination significantly decreased expression of IL1a and IL1b.
MET and the combination also significantly inhibited expressionof IL23, TNFa, CXCL12, VEGFB and IGF-1 whereas the combi-nation also significantly reduced expression of HIF-1a, VEGFA,and IGF-1R. The effect of MET given alone as well as METþ RAPAon CXCL12 is quite interesting. In this regard, CXCL12 is knownto play a role in prostate cancer progression (52). Notably, allthree treatments also significantly decreased the infiltration ofinflammatory cells (CD45þ cells, T lymphocytes, and mast cells)into the VP of HiMyc mice compared with the control animals(Fig.5). These results indicate that all treatments inhibited tissueinflammation and this effect may have contributed to their abilityto inhibit prostate cancer progression inHiMycmice, especially inthe group treated with MET.
Recent evidence suggests that MET has inhibitory effects onNFkB signaling (53). In addition, some of the genes whoseexpression was analyzed in this study are known to be regulatedby NFkB signaling (40). Therefore, the status of NFkB signalingwas also evaluated. The NFkB transcription factor family invertebrates consist of p65 (RelA), RelB, cRel, p50/105 and p52/100 with all having highly conserved dimerization and DNA-binding domains (40). The p50 subunit, which lacks a transacti-vation domain, is essential for NFkB DNA binding, and thephosphorylation of p50 at Ser337 is required for efficient DNAbinding of NFkB (54). IF analysis of prostate tissues from treatedHiMyc mice showed that phospho-p50Ser337 was dramaticallydecreased by all of the treatments compared with tissue from thecontrol mice (Fig. 6A). The most important and common trans-activating p65/p50 heterodimers of NFkB remain bound withinhibitor protein IkBa in the cytoplasm in an inactive form (40).A wide variety of stimuli, including cytokines (TNFa, IL1 etc.),
Figure 6.Effect of MET, RAPA, or MET þ RAPAon NFkB signaling. A, IF staining ofrepresentative sections of VP fromHiMyc mice treated 18 weeks withMET, RAPA or METþRAPA forpNFkBp50 (top) and IkBa (bottom)as described inMaterials andMethods.B, representative Western blotanalysis of lysates from VP tissues(pooled sample of n ¼ 4–5mice/group) for pNFkBp65Ser536
NFkBp65 and GAPDH (left) andquantification of Western blot data(right) showing the average fromtwo separate experiments withnearly identical results.
Saha et al.
Cancer Prev Res; 8(7) July 2015 Cancer Prevention Research604
Research. on May 26, 2019. © 2015 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst April 23, 2015; DOI: 10.1158/1940-6207.CAPR-15-0014

bacterial, and viral products, cause phosphorylation and proteo-somal degradation of IkBa through activation of the upstreaminhibitor of IkBa, leaving the free, active form of NFkB for rapidnuclear translocation (40). As also shown in Fig. 6A, IkBa levelswere higher in the VP ofHiMycmice treated withMET, RAPA, andMET þ RAPA, indicating that the active form of NFkB wascomparatively lower in the VP of these groups compared withthe control group. The phosphorylation of pNFkBp65 at Ser536was also reduced in the VP of HiMyc mice by all treatments (Fig.6B). Because NFkB regulates the expression of various genes,including proinflammatory cytokines, chemokines, growth fac-tors, angiogenesis and adhesion molecules, inhibition of NFkBsignaling (as indicated by decreased phosphorylation of both p50and p65 as well as reduced IkBa degradation) byMET, RAPA, andMET þ RAPA could explain the decreased expression of many ofthe genes examined in Fig. 4. Reduced NFkB signaling could alsoexplain, at least in part, the reduced infiltration of inflammatorycells observed in the VP of treated mice.
In conclusion, the current results demonstrate that long-termoral dosing of HiMyc mice with MET, RAPA, and MET þ RAPAdecreased prostate cancer progression without apparent toxicity.Mechanistic studies revealed that RAPA and the combination ofMET þ RAPA significantly inhibited mTORC1 signaling whereasMET did not. However, all treatments produced significant reduc-tions in inflammatory gene expression, infiltration of inflamma-tory cells, and reduced NFkB signaling in the VP of treated HiMycmice. MET and the combination of MET þ RAPA also reducedexpression of several angiogenesis genes and IGF-1/IGF-1R. Thesedata suggest that RAPA andMETþRAPA inhibited prostate cancerprogression via effects on both mTORC1 and on tissue inflam-mation and inflammation signaling whereas the effects of METwere associated primarily with the latter. Although we wereunable to show that the combination of MET þ RAPA was moreeffective than either agent alone in preventing prostate cancerprogression, this was most likely due to the dose of RAPA used,
which completely suppressed formation of invasive adenocarci-nomas. However, analyses of mTORC1 and inflammation sig-naling suggested the possibility that careful dose selection couldreveal possible additive or synergistic effects of this combinationin future studies. Collectively, the current results suggest thattargeting mTORC1 and/or inflammation signaling may be aneffective strategy for reducing prostate cancer progression- andprostate cancer–specific mortality.
Disclosure of Potential Conflicts of InterestNo potential conflicts of interest were disclosed.
Authors' ContributionsConception and design: A. Saha, J. DiGiovanniDevelopment of methodology: A. Saha, J. DiGiovanniAcquisition of data (provided animals, acquired and managed patients,provided facilities, etc.): A. Saha, J. Blando, L. Tremmel, J. DiGiovanniAnalysis and interpretation of data (e.g., statistical analysis, biostatistics,computational analysis): A. Saha, J. Blando, L. Tremmel, J. DiGiovanniWriting, review, and/or revision of the manuscript: A. Saha, J. Blando,L. Tremmel, J. DiGiovanniAdministrative, technical, or material support (i.e., reporting or organizingdata, constructing databases): J. DiGiovanniStudy supervision: A. Saha, J. DiGiovanni
Grant SupportThis study was supported by NIH grant P50 CA140388 and Start-up funds
from the University of Texas at Austin. A. Saha was supported by CancerPrevention Research Institute of Texas postdoctoral trainee award under grantRP101501 from the State of Texas.
The costs of publication of this article were defrayed in part by thepayment of page charges. This article must therefore be hereby markedadvertisement in accordance with 18 U.S.C. Section 1734 solely to indicatethis fact.
Received January 12, 2015; revised March 31, 2015; accepted April 6, 2015;published OnlineFirst April 23, 2015.
References1. Center MM, Jemal A, Lortet-Tieulent J, Ward E, Ferlay J, Brawley O, et al.
International variation in prostate cancer incidence andmortality rates. EurUrol 2012;61:1079–92.
2. Available from: http://www.cancer.org/cancer/prostatecancer/detailed-guide/prostate-cancer-key-statistics.
3. Siegel R, Ma J, Zou Z, Jemal A. Cancer statistics, 2014. CA Cancer J Clin2014;64:9–29.
4. Klein EA. Chemoprevention of prostate cancer. Annu Rev Med 2006;57:49–63.
5. Van Poppel H, Tombal B. Chemoprevention of prostate cancer withnutrients and supplements. Cancer Manag Res 2011;3:91–100.
6. Barve A, Khor TO,HaoX, KeumYS, YangCS, Reddy B, et al.Murine prostatecancer inhibition by dietary phytochemicals–curcumin and phenyethyli-sothiocyanate. Pharm Res 2008;25:2181–9.
7. Thompson IM Jr, Cabang AB, Wargovich MJ. Future directions in theprevention of prostate cancer. Nat Rev Clin Oncol 2014;11:49–60.
8. Singh SV, Warin R, Xiao D, Powolny AA, Stan SD, Arlotti JA, et al.Sulforaphane inhibits prostate carcinogenesis and pulmonary metastasisin TRAMP mice in association with increased cytotoxicity of natural killercells. Cancer Res 2009;69:2117–25.
9. Raina K, Rajamanickam S, Deep G, Singh M, Agarwal R, Agarwal C.Chemopreventive effects of oral gallic acid feeding on tumor growth andprogression in TRAMP mice. Mol Cancer Ther 2008;7:1258–67.
10. Karin M. NF-kappaB as a critical link between inflammation and cancer.Cold Spring Harb Perspect Biol 2009;1:a000141.
11. KwonKH, BarveA, Yu S,HuangMT, KongAN.Cancer chemopreventionbyphytochemicals: potential molecular targets, biomarkers, and animalmodels. Acta Pharmacol Sin 2007;28:1409–21.
12. SyedDN, KhanN, Afaq F,MukhtarH. Chemoprevention of prostate cancerthrough dietary agents: progress and promise. Cancer Epidemiol Biomar-kers Prev 2007;16:2193–203.
13. Dallaglio K, Bruno A, Cantelmo AR, Esposito AI, Ruggiero L, Orecchioni S,et al. Paradoxic effects ofmetformin on endothelial cells and angiogenesis.Carcinogenesis 2014;35:1055–66.
14. Decensi A, Puntoni M, Goodwin P, Cazzaniga M, Gennari A, Bonanni B,et al. Metformin and cancer risk in diabetic patients: a systematic reviewand meta-analysis. Cancer Prev Res 2010;3:1451–61.
15. Ben Sahra I, Laurent K, Loubat A, Giorgetti-Peraldi S, Colosetti P,Auberger P, et al. The antidiabetic drug metformin exerts an antitumoraleffect in vitro and in vivo through a decrease of cyclin D1 level. Oncogene2008;27:3576–86.
16. Tomimoto A, Endo H, Sugiyama M, Fujisawa T, Hosono K, Takahashi H,et al. Metformin suppresses intestinal polyp growth in ApcMin/þ mice.Cancer Sci 2008;99:2136–41.
17. Zakikhani M, Dowling R, Fantus IG, Sonenberg N, Pollak M. Metformin isan AMP kinase-dependent growth inhibitor for breast cancer cells. CancerRes 2006;66:10269–73.
18. Kahn BB, Alquier T, Carling D, Hardie DG. AMP-activated protein kinase:ancient energy gauge provides clues to modern understanding of metab-olism. Cell Metabol 2005;1:15–25.
www.aacrjournals.org Cancer Prev Res; 8(7) July 2015 605
Anticancer Activity of Metformin and Rapamycin Combination
Research. on May 26, 2019. © 2015 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst April 23, 2015; DOI: 10.1158/1940-6207.CAPR-15-0014

19. Wullschleger S, Loewith R, Hall MN. TOR signaling in growth and metab-olism. Cell 2006;124:471–84.
20. Gibbons JJ, Abraham RT, Yu K. Mammalian target of rapamycin: discoveryof rapamycin reveals a signaling pathway important for normal and cancercell growth. Semin Oncol 2009;36:S3–S17.
21. Kremer CL, Klein RR, Mendelson J, BrowneW, Samadzedeh LK, VanpattenK, et al. Expression of mTOR signaling pathway markers in prostate cancerprogression. Prostate 2006;66:1203–12.
22. Lamming DW, Ye L, Sabatini DM, Baur JA. Rapalogs andmTOR inhibitorsas anti-aging therapeutics. J Clin Invest 2013;123:980–9.
23. Sabatini DM, Barrow RK, Blackshaw S, Burnett PE, Lai MM, Field ME, et al.Interaction of RAFT1 with gephyrin required for rapamycin-sensitivesignaling. Science 1999;284:1161–4.
24. Kinkade CW, Castillo-Martin M, Puzio-Kuter A, Yan J, Foster TH, Gao H,et al. Targeting AKT/mTOR and ERK MAPK signaling inhibits hormone-refractory prostate cancer in a preclinical mouse model. J Clin Invest2008;118:3051–64.
25. Law BK. Rapamycin: an anticancer immunosuppressant? Crit Rev OncolHematol 2005;56:47–60.
26. Kopelovich L, Fay JR, Sigman CC, Crowell JA. The mammalian target ofrapamycin pathway as a potential target for cancer chemoprevention.Cancer Epidemiol Biomarkers Prev 2007;16:1330–40.
27. KauffmanHM, CherikhWS, Cheng Y, Hanto DW, Kahan BD.Maintenanceimmunosuppression with target-of-rapamycin inhibitors is associatedwith a reduced incidence of de novo malignancies. Transplantation 2005;80:883–9.
28. Alberu J, Pascoe MD, Campistol JM, Schena FP, Rial Mdel C, Polinsky M,et al. Lower malignancy rates in renal allograft recipients converted tosirolimus-based, calcineurin inhibitor-free immunotherapy: 24-monthresults from the CONVERT trial. Transplantation 2011;92:303–10.
29. Ellwood-Yen K, Graeber TG, Wongvipat J, Iruela-Arispe ML, Zhang J,Matusik R, et al. Myc-driven murine prostate cancer shares molecularfeatures with human prostate tumors. Cancer Cell 2003;4:223–38.
30. Harrison DE, Strong R, Sharp ZD, Nelson JF, Astle CM, Flurkey K, et al.Rapamycin fed late in life extends lifespan in genetically heterogeneousmice. Nature 2009;460:392–5.
31. Blando J, Moore T, Hursting S, Jiang G, Saha A, Beltran L, et al. Dietaryenergy balance modulates prostate cancer progression in Hi-Myc mice.Cancer Prev Res 2011;4:2002–14.
32. Saha A, Blando J, Silver E, Beltran L, Sessler J, DiGiovanni J. 6-Shogaol fromdried ginger inhibits growth of prostate cancer cells both in vitro and in vivothrough inhibition of STAT3 and NF-kappaB signaling. Cancer Prev Res2014;7:627–38.
33. Maxwell P, McCluggage WG. Audit and internal quality control in immu-nohistochemistry. J Clin Pathol 2000;53:929–32.
34. Mura M, Swain RK, Zhuang X, Vorschmitt H, Reynolds G, Durant S, et al.Identification and angiogenic role of the novel tumor endothelial markerCLEC14A. Oncogene 2012;31:293–305.
35. Heaney LG, Leggett P, Maxwell P, Bharucha H, Ennis M. A comparison ofthree standardmethods of identifyingmast cells in endobronchial biopsiesin normal and asthmatic subjects. Allergy 1997;52:836–43.
36. Attur MG, Patel R, Thakker G, Vyas P, Levartovsky D, Patel P, et al.Differential anti-inflammatory effects of immunosuppressive drugs: cyclo-sporin, rapamycin and FK-506 on inducible nitric oxide synthase, nitricoxide, cyclooxygenase-2 andPGE2production. InflammRes2000;49:20–6.
37. Checkley LA, Rho O, Moore T, Hursting S, DiGiovanni J. Rapamycin is apotent inhibitor of skin tumor promotion by 12-O-tetradecanoylphorbol-13-acetate. Cancer Prev Res 2011;4:1011–20.
38. Hirsch HA, Iliopoulos D, Struhl K. Metformin inhibits the inflammatoryresponse associated with cellular transformation and cancer stem cellgrowth. Proc Natl Acad Sci U S A 2013;110:972–7.
39. Kim SA, Choi HC. Metformin inhibits inflammatory response via AMPK-PTEN pathway in vascular smooth muscle cells. Biochem Biophys ResCommun 2012;425:866–72.
40. Ghosh S, KarinM.Missing pieces in the NF-kappaB puzzle. Cell 2002;109:S81–96.
41. Colquhoun AJ, Venier NA, Vandersluis AD, Besla R, Sugar LM, Kiss A, et al.Metformin enhances the antiproliferative and apoptotic effect of bicalu-tamide in prostate cancer. Prostate Cancer Prostatic Dis 2012;15:346–52.
42. Ben Sahra I, Laurent K, Giuliano S, Larbret F, Ponzio G, Gounon P, et al.Targeting cancer cell metabolism: the combination of metformin and 2-deoxyglucose induces p53-dependent apoptosis in prostate cancer cells.Cancer Res 2010;70:2465–75.
43. Morikawa Y, KoikeH, Sekine Y,MatsuiH, Shibata Y, Ito K, et al. Rapamycinenhances docetaxel-induced cytotoxicity in a androgen-independent pros-tate cancer xenograftmodel by survivin downregulation. BiochemBiophysRes Commun 2012;419:584–9.
44. FagoneP,DoniaM,ManganoK,QuattrocchiC,Mammana S,CocoM, et al.Comparative study of rapamycin and temsirolimus demonstrates super-imposable anti-tumour potency on prostate cancer cells. Basic Clin Phar-macol Toxicol 2013;112:63–9.
45. van der Poel HG, Hanrahan C, Zhong H, Simons JW. Rapamycin inducesSmad activity in prostate cancer cell lines. Urol Res 2003;30:380–6.
46. Athar M, Kopelovich L. Rapamycin andmTORC1 inhibition in themouse:skin cancer prevention. Cancer Prev Res 2011;4:957–61.
47. Checkley LA, Rho O, Angel JM, Cho J, Blando J, Beltran L, et al. Metformininhibits skin tumor promotion in overweight and obese mice. Cancer PrevRes 2014;7:54–64.
48. Akinyeke T, Matsumura S, Wang X, Wu Y, Schalfer ED, Saxena A, et al.Metformin targets c-MYC oncogene to prevent prostate cancer. Carcino-genesis 2013;34:2823–32.
49. CleggNJ, Couto SS,Wongvipat J,HieronymusH,Carver BS, Taylor BS, et al.MYC cooperates with AKT in prostate tumorigenesis and alters sensitivityto mTOR inhibitors. PloS ONE 2011;6:e17449.
50. Blando J, Portis M, Benavides F, Alexander A, Mills G, Dave B, et al. PTENdeficiency is fully penetrant for prostate adenocarcinoma in C57BL/6micevia mTOR-dependent growth. Am J Pathol 2009;174:1869–79.
51. Ben Sahra I, Regazzetti C, Robert G, Laurent K, Le Marchand-Brustel Y,Auberger P, et al. Metformin, independent of AMPK, induces mTORinhibition and cell-cycle arrest through REDD1. Cancer Res 2011;71:4366–72.
52. Sun X, Cheng G,HaoM, Zheng J, Zhou X, Zhang J, et al. CXCL12 / CXCR4 /CXCR7 chemokine axis and cancer progression. Cancer Metastasis Rev2010;29:709–22.
53. Moiseeva O, Deschenes-Simard X, St-Germain E, Igelmann S, Huot G,Cadar AE, et al. Metformin inhibits the senescence-associated secretoryphenotype by interfering with IKK/NF-kappaB activation. Aging Cell2013;12:489–98.
54. Hou S, Guan H, Ricciardi RP. Phosphorylation of serine 337 of NF-kappaBp50 is critical for DNA binding. J Biol Chem 2003;278:45994–8.
Cancer Prev Res; 8(7) July 2015 Cancer Prevention Research606
Saha et al.
Research. on May 26, 2019. © 2015 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst April 23, 2015; DOI: 10.1158/1940-6207.CAPR-15-0014

2015;8:597-606. Published OnlineFirst April 23, 2015.Cancer Prev Res Achinto Saha, Jorge Blando, Lisa Tremmel, et al. and Progression of Prostate Tumors in HiMyc MiceEffect of Metformin, Rapamycin, and Their Combination on Growth
Updated version
10.1158/1940-6207.CAPR-15-0014doi:
Access the most recent version of this article at:
Material
Supplementary
1
http://cancerpreventionresearch.aacrjournals.org/content/suppl/2015/04/25/1940-6207.CAPR-15-0014.DCAccess the most recent supplemental material at:
Cited articles
http://cancerpreventionresearch.aacrjournals.org/content/8/7/597.full#ref-list-1
This article cites 53 articles, 17 of which you can access for free at:
Citing articles
http://cancerpreventionresearch.aacrjournals.org/content/8/7/597.full#related-urls
This article has been cited by 1 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerpreventionresearch.aacrjournals.org/content/8/7/597To request permission to re-use all or part of this article, use this link
Research. on May 26, 2019. © 2015 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst April 23, 2015; DOI: 10.1158/1940-6207.CAPR-15-0014