effects eruption cycle andrelatedwithin the blast zonewaslimited to helicopters during 1980. ryan...
TRANSCRIPT

APPLIED AND ENVIRONMENTAL MICROBIOLOGY, May 1983, p. 1633-16450099-2240/83/051633-13$02.00/0Copyright 0 1983, American Society for Microbiology
Vol. 45, No. 5
Initial Effects of the Mount St. Helens Eruption on NitrogenCycle and Related Chemical Processes in Ryan Lake
CLIFFORD N. DAHM,1* JOHN A. BAROSS,2 AMELIA K. WARD,3 MARVIN D. LILLEY,2 AND JAMESR. SEDELL4
Department of Fisheries and Wildlife' and School of Oceanography,2 Oregon State University, Corvallis,Oregon 97331; Department ofBiology, University ofAlabama, University, Alabama 354863; and Forestry
Sciences Laboratory, U.S. Forest Service, Corvallis, Oregon 973314
Received 24 September 1982/Accepted 31 January 1983
Ryan Lake, a 1.6-hectare basin lake near the periphery of the tree blowdownarea in the blast zone 19 km north of Mount St. Helens, was studied from Augustto October 1980 to determine the microbial and chemical response of the lake tothe eruption. Nutrient enrichment through the addition of fresh volcanic materialand the organic debris from the surrounding conifer forest stimulated intensemicrobial activity. Concentrations of such nutrients as phosphorus, sulfur,manganese, iron, and dissolved organic carbon were markedly elevated. Nitrogencycle activity was especially important to the lake ecosystem in regulatingbiogeochemical cycling owing to the limiting abundance of nitrogen compounds.Nitrogen fixation, both aerobic and anaerobic, was active from aerobic benthicand planktonic cyanobacteria with rates up to 210 nmol of N2 cm-1 h-1 and 667nmol of N2 liter-1 h-1, respectively, and from anaerobic bacteria with ratesreaching 220 nmol of N2 liter-1 h-1. Nitrification was limited to the aerobicepilimnion and littoral zones where rates were 43 and 261 nmol of NO2 liter-1day-', respectively. Potential denitrification rates were as high as 30 ,umol of N20liter-1 day-1 in the anaerobic hypolimnion. Total bacterial numbers ranged from 1x 106 to 3 x 108 mI-1 with the number of viable sulfur-metal-oxidizing bacteriareaching 2 x 106 ml-1 in the hypolimnion. A general scenario for the microbialcycling of nitrogen, carbon, sulfur, and metals is presented for volcanicallyimpacted lakes. The important role of nitrogen as these lakes recover from thecataclysmic eruption and proceed back towards their prior status as oligotrophicalpine lakes is emphasized.
Numerous alpine and subalpine oligotrophiclakes near Mount St. Helens were affected bythe 18 May 1980 eruption to various degreesdepending upon their location relative to thevolcano. Twenty lakes within the blast zone,ranging in size from less than 1 to 525 hectares,were especially enriched with ash, soil, and theorganic remains from the devastated coniferousforest. Heavily impacted lakes, such as SpiritLake, were reshaped, and numerous new lakesand hydrothermal seeps were created by debrisand pyroclastic flows.The initial microbiological, biological, and
chemical responses in lakes shortly after the 18May 1980 eruption have been documented (3,18, 40, 46, 47; A. K. Ward, J. A. Baross, C. N.Dahm, M. D. Lilly, and J. R. Sedell, J. Phycol.,in press). These were the first studies of thiskind ever made immediately after a catastrophicvolcanic eruption. Intense microbiological activ-ities in the lakes were evident within weeks ofthe initial eruption. These normally oligotrophic
lakes showed various degrees of anaerobiosis,eutrophication, staining, and gas ebullition. Ex-tensive populations of chemolithotrophic (3) andheterotrophic bacteria (40) were found through-out the water column where they were utilizingthe reduced metals, sulfur, gases, and organiccompounds. Nitrogen was found to limit theextent of microbial activities, resulting in highrates of phototrophic and nonphototrophic N2fixation. Anaerobic oxidation coupled with deni-trification occurred throughout the anaerobicwater column. It was predicted that the recoveryof lakes to oligotrophism will occur when in situ02 production and physical processes of atmo-spheric resupply exceed the 02 demand pro-duced by the inputs from the cataclysmic vulca-nism (3).One of the moderately impacted lakes within
the blast zone, Ryan Lake, was selected forfurther study the first summer and fall after theeruption. Ryan Lake, a subalpine oligotrophiclake located in the zone of large-scale tree
1633
on April 29, 2020 by guest
http://aem.asm
.org/D
ownloaded from

APPL. ENVIRON. MICROBIOL.
blowdown, received both heavy ash and organicmatter input. By August 1980, Ryan Lake had a1- to 2-m aerobic epilimnion with an anaerobichypolimnion. Here, we report on the microbio-logical processes and related chemical charac-teristics in Ryan Lake with particular emphasison the nitrogen cycle and the microbiology ofmetal and sulfur oxidation in the water column.Measurements of microbial activities and con-centration of biologically important chemicalconstituents were made on five dates throughoutthe summer of 1980 and at 20-cm intervalsthroughout the oxycline on 26 September 1980.A hypothetical scenario showing the progressionof microbial cycles accompanying lake recoveryis also presented.
MATERIALS AND METHODS
Study area. Ryan Lake is a 1.6-hectare dimicticbasin lake at an elevation of 1,009 m and a distance of19 km from the crater of Mt. St. Helens (Fig. 1).Maximum depth is 7 m. The lake was believed to havebeen a typically oligotrophic Cascade subalpine lakebefore the eruption. This statement is based on datafrom six nearby lakes of similar elevation and geologi-cal setting (8). An old-growth Douglas fir forest existedaround the lake before the eruption. Ashfall depositionin the area near the lake amounted to 10 to 20 cm (Fig.2).
Sampling. Access to Ryan Lake and other siteswithin the blast zone was limited to helicopters during1980. Ryan Lake was deemed to be far enough fromthe crater that on days with fair weather, a 6 to 8 hworking period was allowed. Initial chemical and
RYANLAKE
l BLOWDOWN
[1I1IffiI SCORCH
\\gU, 3MUD FLOW
CRATER
itivr -___,6McBrideLake
MerrillLake
0 5 10KILOMETERS
FIG. 1. Map of blast zone near Mount St. Helens showing location of Ryan Lake.
1634 DAHM ET AL.
on April 29, 2020 by guest
http://aem.asm
.org/D
ownloaded from

VOLCANIC EFFECTS ON LAKE MICROBIOLOGY 1635
FIG. 2. Aerial photo of Ryan Lake at edge of blast zone.
biological data were collected on 30 June 1980 byRobert C. Wissmar, University of Washington, Seattle(46, 47). Subsequent sampling occurred on 10 and 19August and 11 and 26 September 1980. Depth samplingwas done with 5-liter Niskin bottles from a portablerubber raft. The close-interval sampling on September26 was done by using a 50-ml syringe sampler spacedat 20-cm intervals.
Chemical analyses. Temperature (T), pH, and ICO2were measured on site. Temperature was measuredwith a YSI thermistor probe. A Coleman 37A portablepH meter was used in the p-H and ICO2 analyses. Agas-sensing CO2 electrode was used to determine£CO2 (22). Oxygen samples were fixed at the samplingsite (11) and titrated upon return to the lab. Carbonatealkalinity and partial pressure of carbon dioxide(pCO2) were calculated by the method of Park (36)from pH and £CO2 data.
Nutrient concentrations (NO3- + NO2, NH4',P04 , and SiO2) were measured on an AutoanalyzerII (Technicon Corp., Inc.) (2). Total phosphorus wasmeasured colorimetrically on the Autoanalyzer afterdigestion with persulfate (32). Total reduced nitrogen(NH4' plus organic nitrogen) was also determinedcolorimetrically after a modified micro-Kjeldahl diges-tion (1). Dissolved organic carbon (DOC) was ana-lyzed on an Oceanography International instrument byinfrared determination of the CO2 liberated by wetoxidation with potassium persulfate (33). Dissolvedorganic nitrogen (DON) was calculated by subtractingthe concentration of NH3 from total reduced nitrogen.Ferrous iron was measured colorimetrically with fer-rozine added in the field immediately after sample
filtration (42). All samples were filtered at Ryan Lakethrough prefired (450°C, overnight) Whatman GF/Fglass fiber filters and then stored on ice until return tothe lab.Samples for dissolved gases were poisoned by the
addition of 75 mg of Hg per liter as HgCl2 in the fieldand analyzed in the laboratory. Our experience hasshown that the addition of HgCl2 results in stable gasconcentrations over a period of at least 1 week. Themethods described by Lilley et al. (30) were used forthe analyses of hydrogen, methane, and carbon mon-oxide.
Microbial activity. The acetylene reduction methodwas used as an index of N2 fixation (38, 41). Samplesof water (10 to 30 ml) or water and benthic algae wereincubated in situ for at least 2 h in 60-ml serum bottles.The anaerobic measurements were on samples purgedwith argon for 60 to 120 s before incubation. Ethylenecontent was determined with a Hewlett-Packard5830A gas chromatograph with a copper column (2 mby 3 mm) packed with 60/80 mesh Porapak Q (WatersAssociates, Inc., Framingham, Mass.). Controls, ei-ther acidified to below pH 2 with H2SO4 or poisonedwith 0.2 ml of 2% HgCl2, were run to determine theamount of ethylene released abiotically.The rates of N2 fixation reported are only apparent
values. The acetylene reduction method is an indirectassay for fixation, and the rates have not yet beenverified with 15N2. Conversion to nanomoles of N2fixed was done by using the theoretical ratio of 3.0 molof ethylene produced per mol of N2 fixed. The theoret-ical ratio of 3.0 has been reported to vary from 2 to 25(26). A conversion factor for an anaerobic bacterial
VOL. 45, 1983
on April 29, 2020 by guest
http://aem.asm
.org/D
ownloaded from

APPL. ENVIRON. MICROBIOL.
soil population, Microcoleus chthonoplastes, was 5.4(39). Whether this value might more closely match theanaerobic aquatic population of N2 fixers is unknown.If a value near 6 proves to be more appropriate, thevalues reported here would be about double the actualrate.
Potential denitrification rates were measured underin situ conditions by the acetylene inhibition method(29, 43). The N20 formed was analyzed by electroncapture gas chromotography (14). Samples with andwithout added N03- were measured. Controls were
either poisoned with 0.2 ml of 2% HgCl2 or 2% (wt/vol)neutralized Formalin.The potential rates of oxidation of NH4' to N02-
were measured, using 30-ml water samples in 60-mlserum bottles. One set of samples served as controlswith no added NH4', whereas another set received 0.3mg of NH4' ml- l (5). The levels of N02- formed weremeasured colorimetrically (6). All samples for microbi-al activity were either incubated in situ or at in situtemperature in the laboratory for the cases in whichincubation times longer than 4 h were necessary.
Bacterial enumeration. Direct counts were made byepifluorescent microscopy. Samples of water wereimmediately fixed to a final concentration of 2%glutaraldehyde. Various dilutions of the sample were
filtered through 0.2-,.m Nuclepore filters prestainedwith Irgalan black (27). The filtered samples were
stained with acridine orange as described previouslyby Zimmerman and Meyer-Reil (48) and then countedwith a Zeiss IV Fl epifluorescent condenser micro-scope fitted with filters KP 500, IP 490, FT 510, andLP 520. Twenty or thirty representative fields were
counted.Manganese- and sulfur-oxidizing bacteria were enu-
merated with a medium as previously described (3).Confirmation of manganese oxidation was determinedwith a benzidinium reagent (34).
RESULTS
Temperature effects. The heat associated withthe initial blast on 18 May 1980 was estimated tobe >250°C at Ryan Lake as determined by themelting point of plastic deformed in the parkinglights of a truck parked beside the lake. Theblast pyrolized most of the coniferous forestfoliage and toppled the large trunks of treesaround the lake (Fig. 2). However, the intenseheat wave passed quickly, and campers at thelake survived with only minor injuries. Near-surface temperature on 30 June 1980 in RyanLake was 25°C (47). A nearby lake outside theblast zone at a similar elevation, but surroundedand shaded by an extant forest, was 18°C on thesame date. The temperature of Spirit Lake, 8 kmfrom the crater and the most dramatically im-pacted lake in the blast zone, was 32.70C near
the surface on 19 May 1980 (20). Although some
warming of the water in Ryan Lake surelyoccurred, we believe it to have been considera-bly less than that measured in Spirit Lake.
Volcanic inputs. A summary of the inorganicconstituents in the ash from the 18 May 1980eruption is presented in Table 1 (24). The ash
TABLE 1. Major element composition andbiologically important trace elements in Mount St.
Helens ash (24)aCompound(%)Avg SDor element (ppm)
SiO2 65.0 3.3A1203 16.9 0.7Fe2O3 4.81 1.1CaO 4.94 0.92Na2O 4.57 0.17MgO 2.21 0.94K20 1.47 0.28TiO2 0.69 0.13P205 0.37 0.09MnO 0.077 0.020S 840 380Cl 663 182Co 19 5Cu 36 5Zn 53 7Cr 17 8Ni 15 8Mo 2.8 1.1
a Values are given in percentages unless otherwisenoted. The standard deviations are from 9 ash sam-ples.
was of dacitic composition, with silica makingup 65% of the material. Aerobic leaching of theash in distilled water at a ratio by weight of 10:1water to ash yielded mainly SO4, Ca, Cl, Na, K,and Mg with smaller quantities of such essentialplant nutrients as P, Cu, Fe, Zn, Mn, and Mo(16). These experiments also showed no in-crease in acidity. In fact, the majority of theleachates showed a slight increase in pH (16).However, when anaerobic conditions occurred,considerable solubilization of reduced metalsand other nutrients resulted (3, 46; Ward et al.,in press).A second important input into lakes in the
blast zone was organic matter, both dissolvedand particulate, from the devastated forest. Thismaterial produced a dramatic increase in DOC,particularly in lakes in the debris flow, and leftSpirit Lake more than half covered by a floatinglog mass. Conifer material is carbon rich andnitrogen poor. Douglas fir needles have a C/Nratio of about 60, whereas bark, sapwood, andheartwood ratios are near 500 (7). Overall, C, S,metals, and, to a lesser extent, P, were enrichedin Ryan Lake, whereas the relative input ofN bythe eruption was not comparable with the othernutrients.Temporal patterns in lake surface chemistry.
Five sampling trips were made to Ryan Lakefrom June to September 1980. Only a surfacesample was taken on June 30, whereas a depthprofile was sampled on the other trips.
Surface values for T, pH, carbonate alkalin-ity, YCO2, pCO2, and dissolved 02 are shown in
1636 DAHM ET AL.
on April 29, 2020 by guest
http://aem.asm
.org/D
ownloaded from
VOLCANIC EFFECTS ON LAKE MICROBIOLOGY 1637
ICO2(mmol/L)Pco2 (ppm)T (0CM
XpHC.A.02
0* (meq/L)
(mg/L)7.0.306.0
Aug 19
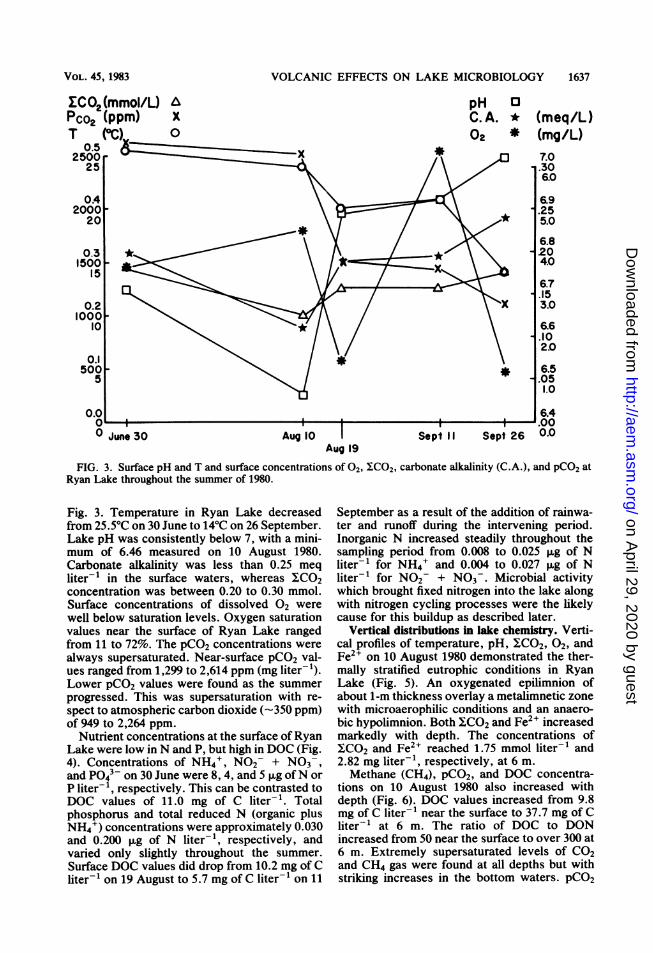
FIG. 3. Surface pH and T and surface concentrations of 02, YCO2, carbonate alkalinity (C.A.), and pCO2 atRyan Lake throughout the summer of 1980.
Fig. 3. Temperature in Ryan Lake decreasedfrom 25.5°C on 30 June to 14°C on 26 September.Lake pH was consistently below 7, with a mini-mum of 6.46 measured on 10 August 1980.Carbonate alkalinity was less than 0.25 meqliter-' in the surface waters, whereas ICO2concentration was between 0.20 to 0.30 mmol.Surface concentrations of dissolved 02 werewell below saturation levels. Oxygen saturationvalues near the surface of Ryan Lake rangedfrom 11 to 72%. The pCO2 concentrations werealways supersaturated. Near-surface pCO2 val-ues ranged from 1,299 to 2,614 ppm (mg liter-').Lower pCO2 values were found as the summerprogressed. This was supersaturation with re-spect to atmospheric carbon dioxide (-350 ppm)of 949 to 2,264 ppm.
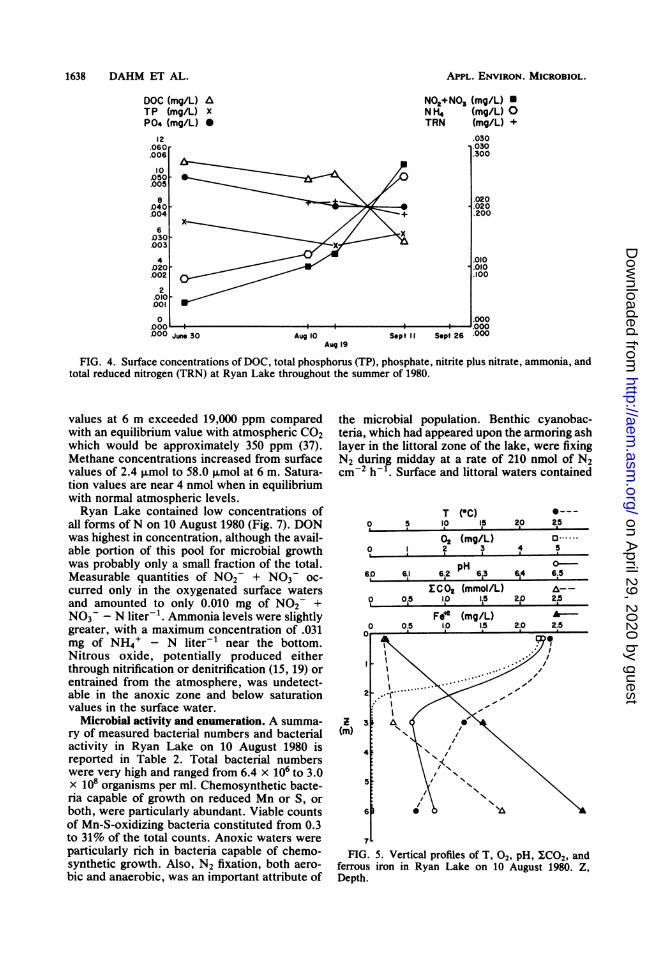
Nutrient concentrations at the surface of RyanLake were low in N and P, but high in DOC (Fig.4). Concentrations of NH4', NO2 + NO3,and P043- on 30 June were 8, 4, and 5 ,ug ofN orP liter-1, respectively. This can be contrasted toDOC values of 11.0 mg of C liter-1. Totalphosphorus and total reduced N (organic plusNH4+) concentrations were approximately 0.030and 0.200 ,ug of N liter-', respectively, andvaried only slightly throughout the summer.Surface DOC values did drop from 10.2 mg of Cliter-1 on 19 August to 5.7 mg of C liter-1 on 11
September as a result of the addition of rainwa-ter and runoff during the intervening period.Inorganic N increased steadily throughout thesampling period from 0.008 to 0.025 ,ug of Nliter- for NH4' and 0.004 to 0.027 ,ug of Nliter-1 for NO2 + NO3. Microbial activitywhich brought fixed nitrogen into the lake alongwith nitrogen cycling processes were the likelycause for this buildup as described later.
Vertical distributions in lake chemistry. Verti-cal profiles of temperature, pH, XCO2, 02, andFe2+ on 10 August 1980 demonstrated the ther-mally stratified eutrophic conditions in RyanLake (Fig. 5). An oxygenated epilimnion ofabout 1-m thickness overlay a metalimnetic zonewith microaerophilic conditions and an anaero-bic hypolimnion. Both IC02 and Fe2+ increasedmarkedly with depth. The concentrations ofZC02 and Fe2+ reached 1.75 mmol liter-1 and2.82 mg liter-1, respectively, at 6 m.Methane (CH4), pCO2, and DOC concentra-
tions on 10 August 1980 also increased withdepth (Fig. 6). DOC values increased from 9.8mg of C liter-1 near the surface to 37.7 mg of Cliter-1 at 6 m. The ratio of DOC to DONincreased from 50 near the surface to over 300 at6 m. Extremely supersaturated levels of CO2and CH4 gas were found at all depths but withstriking increases in the bottom waters. pCO2
VOL. 45, 1983
on April 29, 2020 by guest
http://aem.asm
.org/D
ownloaded from
APPL. ENVIRON. MICROBIOL.
DOC (mg/L) ATP (mg/L) xP04 (mg/L) *
12.060.006
10D50.005
8D40.004
6£030.003
4.020.002
2.010.001
0
.000 June 30
NO2+NN03 (mg/L) UNH4 (mg/L) 0TRN (mg/L) +
.030
.030
.300
.020
.020
.200
.010
.010
.100
.000
Aug 10 Sept II Sept 26 °000Aug 19
FIG. 4. Surface concentrations of DOC, total phosphorus (TP), phosphate, nitrite plus nitrate, ammonia, andtotal reduced nitrogen (TRN) at Ryan Lake throughout the summer of 1980.
values at 6 m exceeded 19,000 ppm comparedwith an equilibrium value with atmospheric CO2which would be approximately 350 ppm (37).Methane concentrations increased from surfacevalues of 2.4 ,umol to 58.0 ,umol at 6 m. Satura-tion values are near 4 nmol when in equilibriumwith normal atmospheric levels.Ryan Lake contained low concentrations of
all forms of N on 10 August 1980 (Fig. 7). DONwas highest in concentration, although the avail-able portion of this pool for microbial growthwas probably only a small fraction of the total.Measurable quantities of NO2 + NO3 oc-curred only in the oxygenated surface watersand amounted to only 0.010 mg of NO2 +N03- - N liter-1. Ammonia levels were slightlygreater, with a maximum concentration of .031mg of NH4+ - N liter-' near the bottom.Nitrous oxide, potentially produced eitherthrough nitrification or denitrification (15, 19) orentrained from the atmosphere, was undetect-able in the anoxic zone and below saturationvalues in the surface water.
Microbial activity and enumeration. A summa-ry of measured bacterial numbers and bacterialactivity in Ryan Lake on 10 August 1980 isreported in Table 2. Total bacterial numberswere very high and ranged from 6.4 x 106 to 3.0x 108 organisms per ml. Chemosynthetic bacte-ria capable of growth on reduced Mn or S, orboth, were particularly abundant. Viable countsof Mn-S-oxidizing bacteria constituted from 0.3to 31% of the total counts. Anoxic waters wereparticularly rich in bacteria capable of chemo-synthetic growth. Also, N2 fixation, both aero-bic and anaerobic, was an important attribute of
the microbial population. Benthic cyanobac-teria, which had appeared upon the armoring ashlayer in the littoral zone of the lake, were fixingN2 during midday at a rate of 210 nmol of N2cm-2 h-V. Surface and littoral waters contained
T (°C) 0--
0 5 10 _,5 20 25
Og (mg/L) O0 2 3 4 5
pH o6.0 6.1 6.2 6.3 614 6.5
EC0Op (mmol/L)0 0.5 ID 1.5 20 25.*J*L. A
Fee (mg/L)
(m)
FIG. 5. Vertical profiles of T, 02, pH, ICO2, andferrous iron in Ryan Lake on 10 August 1980. Z,Depth.
1638 DAHM ET AL.
on April 29, 2020 by guest
http://aem.asm
.org/D
ownloaded from

VOLCANIC EFFECTS ON LAKE MICROBIOLOGY 1639
DOC (mg/L) *---o 1,5 30 45
DOC/DON 0-o 100 200 300
Pcot (ppm)0 6000 12000 18000
CH4 (aimoi/L) A.
CM)
FIG. 6. Vertical profiles of DOC, pCO2, CH4, andratio ofDOC to DON in Ryan Lake on 10 August 1980.Z, Depth.
microbial populations capable of both light aero-bic and dark anaerobic fixation with rates from130 to 667 nmol of N2 liter-' h-1. Dark anaero-bic bacterial fixation was present in all samples,but peaked at a rate of 220 nmol of N2 liter- h-1at 6 m. Addition of new nitrogen, through fixa-tion, was a major response of the microbialpopulation in Ryan Lake. Nitrification was notdetected at 3 or 6 m. The highest rates ofnitrification were 857 nmol of NO2- liter-1day-' in the littoral zone. Nitrification ratesnear the surface of the center of the lake were143 nmol of NO2- liter-' day-'. Potential deni-trification reached 30 ,umol of N20 liter-' day-'in the deepest sample from Ryan Lake, but waslow in the oxygenated portions of the lake.Microscopic examination of water samples fromthe four regions of the lake showed the presenceof the cyanobacteria, Anabaena sp., algae, anddiatoms in the littoral surface water, but not inwaters from deeper strata.
Nitrogen cycle activity was a major concernbecause of the paucity of nitrogen species ascompared with other available nutrients. In ad-dition, the conditions in Ryan Lake werethought to be potentially instructive for futuresuccessional patterns to be seen later in themore devastated lakes on the debris flow. There-fore, we obtained a vertical profile of in siturates of anaerobic N2 fixation, nitrification, de-nitrification, and concentrations ofNH4 at 0.2-m
intervals between 0.4 and 2.2 m in Ryan Lake on26 September 1980 (Fig. 8). Oxygen measure-ments from the small volume (50 ml) and close-interval sampler were inaccurately determinedowing to H2S interference. However, ferrousiron concentrations were relatively constantfrom 0.4 to 1.8 m (0.68 + .07 mg of Fe2+ liter-1)and increased steadily beginning at 2.0 m. Fromthese data, we infer that the interface betweenmicroaerophilic conditions and the anaerobiczone occurred near the 2.0-m depth. Increases inthe rate of anaerobic N2 fixation and potentialdenitrification from 1.8 to 2.2 m supported thisconclusion.Two zones of maximum N2 fixation were
found. One occurred at 0.6 m with rates of 320nmol of N2 liter-' h-1. Surface phytoplanktoncontained low numbers of Anabaena sp. (-3 x103 liter-'), which was dominant in N2-fixingmats in the littoral zone (Ward et al., in press).Because of the low numbers of N2-fixing algae inthe plankton, we suspect the acetylene reduc-tion activity was associated with a combinationof bacterial and algal populations. A secondmaximum was located in the anoxic zone at 5 mwith rates of 232 nmol of N2 liter-' h-1. Con-centrations of NH4', although low throughoutthe water column, were highest at the samedepths as maximum N2 fixation. The highestavailability ofNH4' in the epilimnion also corre-sponded to the narrow zone (0.6 to 1.0 m) of
DOONH4 (mg/L)
.010 .020a
NOt + N03.000 .00DON
.0 .10
(m)
(mg/L).010
(mg/L).200
0--.030
O-.-----.015
0-.300
a- -0
N.N1-
"I.
N1-N1%
IN-"I-
.
FIG. 7. Vertical profiles of ammonia, nitrite plusnitrate, DON, and nitrous oxide (N20) in Ryan Lakeon 10 August 1980. Z, Depth.
VOL. 45, 1983
on April 29, 2020 by guest
http://aem.asm
.org/D
ownloaded from

APPL. ENVIRON. MICROBIOL.
TABLE 2. Microbiological characteristics of Ryan Lake on 10 August 1980
Total S- N2 fixation Denitfica PresenceTotal bac- Mn oxi- N2 fixation (light- (dark aero- tnfica-l Nitrification of cyano-Depth terial no. dizing aerobic) (nmol of bic) (nmol of tion (nmol (nmol of N02 bacteria,
(in) (ml-) bacteria N2 liter-' h-1) N2 liter- ter- day-)l liter-' day') algae, and(ml ') h")te1dy) diatoms
Littoral 3.0 x 108 0.9 x 106 210 (nmol of N2 140 <1 857 +cm-2 h-1)
0 3.0 x 107 5.0 X 104 667 130 <1 143 +3 1.5 x 107 2.0 x 106 No data 180 10 <76 6.4 x 106 2.0 x 106 No data 220 30 <7
nitrifying bacteria (7 to 107 nmol of NO2- liter-'day-'). Potential denitrification was mainly as-sociated with anaerobic bacterial populationsbelow 1.8 m, with rates reaching 32 nmol of N20liter-1 day-1 at 5 m.The trace gases H2, CH4, and CO were also
measured concurrently with the rates of nitrogencycle activity in Ryan Lake on 26 September1980 (Fig. 9). A maximum in H2 at 0.6 m wasmeasured at the same depth as the maximum inN2 fixation. A second H2 maximum occurred at1.2 m, where minimum concentrations of NH4'
NH4 (ug/L)20 40
l60
NITROGEN FIXATION (nmol N2/L/Hour)O 100 200 300
NITRIFICATION (nmol NO-/L/Day)0 50 100 150
DENITRIFICATION (nmol N20/L/Day)10 20 30 0°---
2'
(m) 3
\ '11
I
'0
61
FIG. 8. Concentration of ammonia and the rates ofnitrogen fixation, nitrification, and denitrification at20-cm intervals in Ryan Lake on 26 September 1980.Z, Depth.
5.
6
were present. The concentration of CO peakedat 1,700 nmol liter-1 at a depth of 1.6 m, with 800nmol present at 1.8 m. We have suggestedincomplete CH4 oxidation as the mechanism forincreased levels of CO (Baross, Lilley, Dahm,and Gordon, Abstr. Annu. Am. Geophys. Un.Meet. 1981, EOS, vol. 63, p. 115), but confirma-tory measurements of the rate of CH4 oxidationin Ryan Lake were not undertaken.
Total bacterial numbers in Ryan Lake on 26September 1980 reached 1.5 x 107 bacteria ml-lat 2.2 m (Fig. 10). Bacterial numbers below the
C H,4 (Arno-/L) 0-O 5 10
Co (uJmol/L)2O 2 2
H2 (nmol/L)O 100 200
N2 FIXATION (nmol N2/L/Hour)
FIG. 9. Rate of nitrogen fixation and the concen-
tration of the gases methane, carbon monoxide, andhydrogen at 20-cm intervals in Ryan Lake on 26September 1980. Z, Depth.
0
a
Cm) 3.-
4.
1640 DAHM ET AL.
ILr
on April 29, 2020 by guest
http://aem.asm
.org/D
ownloaded from

VOLCANIC EFFECTS ON LAKE MICROBIOLOGY 1641
PERCENTAGE METAL/SULFUR BACTERIA W*-o 25 50
TOTAL No. VIABLE SULFUR and METALOXIDIZING BACTERIA mr'x 10i A---
o Ip 20
(mI(in)
TOTAL No. BACTERIA ml' x10 CpI)i03 6 9 12 is
FIG. 10. Total bacterial numbers, viable sulfur-metal-oxidizing bacteria, and the percentage of sulfur-metal-oxidizing bacteria at 20-cm intervals in RyanLake on 26 September 1980. Z, Depth.
metalimnion were about 7 x 106 ml-1 and ap-
proximately 3 x 106 in the metalimnion. Viablebacteria capable of Mn-S oxidation again wereabundant, with numbers in the anoxic zone andnear the oxic-anoxic interface as high as 2 x 106.The percentage of the total bacteria having thepotential to oxidize Mn-S ranged from 3 to 57%of the total epifluorescent counts. Since most ofthese bacteria were found to also utilize organiccompounds, it is possible that a high proportionof the previously reported (40, 47) heterotrophicpopulation in Ryan Lake as well as other impact-ed lakes may also have the ability to oxidizemanganese and sulfur.
DISCUSSIONBefore the eruption of Mount St. Helens in
1980, the initial microbial and chemical responseof aquatic ecosystems to cataclysmic terrestrialvulcanism had not been documented. Previousstudies had viewed mainly terrestrial revegeta-tion (e.g., reference 21 and references citedtherein). The 18 May 1980 eruption of Mount St.Helens introduced into nearby lakes an abun-dance of inorganic ash compounds and ele-ments, and organic material from the forest.Although the water temperature of lakes locatedin the blast zone increased immediately after theblast, this was not sufficient to cause the lakes togo anaerobic. In most of the lakes sampledduring the summer of 1980, the surface waterswere oxygenated, and in some lakes located atsimilar distances from the crater as Ryan Lake,various animals including fish, pollywogs, and
insect larvae survived the blast. Within weeksafter the blast, however, and probably as a resultof extensive heterotrophic activity, the lakesbecame anaerobic in the deeper waters. By thethe time we began sampling these lakes, theywere either anaerobic throughout the water col-umn or they had a shallow oxygenated epilimni-on. Evidence of extensive anaerobic microbialactivity included active ebullition of methaneand H2S emanating from the sediments. Anydisturbance of the littoral sediment at RyanLake resulted in extensive degassing.We have postulated that the sequence of mi-
crobial activities that occurred immediately afterthe blast and before we began our samplingincluded heterotrophic activity involving the sol-uble organic compounds from the downed for-est. A major portion of the soluble fractions offorest litter has repeatedly been shown to betaken up by heterotrophic bacteria (17, 45).This, along with related geochemical reactions,utilized the remaining oxygen in the sedimentsand bottom waters. The oxidized elements andcompounds associated with ash gradually be-came reduced owing to microbial activity, re-sulting in high concentrations of H2S and re-duced manganese, iron and other transitionmetals being mobilized into the water column.We have also postulated that in these earlystages nitrogen and perhaps phosphorus becamelimiting nutrients and that this explains why thepotential rates of N2 fixation were so high duringthe summer of 1980 (3). Data from the summers
of 1981 and 1982 showed greatly reduced rates ofanaerobic N2 fixation in Ryan Lake, so that themajor introduction of new N2 into Ryan Lakeoccurred only during the summer of 1980. It ispossible that at the present time phosphorus isthe limiting nutrient in these impacted lakes.The importance of N2 fixation to lakes in the
volcanic blast zone was reflected in the extraor-dinary rates of both aerobic and anaerobic fixa-tion. The potential planktonic aerobic N2 fixa-
TABLE 3. Aerobic N2 fixation in the light insurface waters of various North American lakes
compared with Ryan Lakea
Lake and location N2 fixed (nmol of N2liter' h-1)Crystal Lake,Wis. 0-0.04Trout Lake, Wis.0.4Green Bay, Mich.0.7-7.1Lake Mendota, Wis.0.1-12.5Lake Wingra, Wis.0.3-26.4Little Arbor Vitae Lake, Wis. 0.3-12.5Big Arbor Vitae Lake, Wis. 0.7-104Sanctuary Lake, Pa.7.1-89.3Clear Lake, Calif. 10.0-296Ryan Lake, Wash.130-667
a Modified from Mague (31).
VOL. 45, 1983
on April 29, 2020 by guest
http://aem.asm
.org/D
ownloaded from

APPL. ENVIRON. MICROBIOL.
TABLE 4. Nitrification rates in Ryan Lakecompared with other lake and estuarine watersa
Nitrification rateSiteb (nmol of N liter-
day-')Lake Vanda ...................... 1-136Lake Taupo ...................... 36-286Lake Mendota.................... 50-1,057Blelham Tarn (enriched enclosure) . 3,214Scheldt Estuary .................. 4,071-11,786Scheldt Estuary (potential)......... 1,214-40,786Ryan Lake (potential) ............. 7-857
a Modified from Vincent and Downes (44).b Locations are as follows: Lake Vanda, Antarctica;
Lake Taupo, New Zealand; Lake Mendota, Wis.,Blelham Tarn, England; Scheldt Estuary, The Nether-lands.
tion (algal and bacterial) rate measured in theafternoon 10 August 1980 exceeded all previous-ly reported values in North American lakes(Table 3). Even though the rates could be signifi-cantly overestimated since they were derivedfrom acetylene reduction data and a conversionratio of 3.0 was assumed, a high rate of fixationwas evident. Rates of potential N2 fixation inbenthic algal mats were also substantial, andlittoral-zone cyanobacteria were probably majorfactors providing assimilable nitrogen in thelake.
Similarly, potential anaerobic dark N2 fixationrates were also unusually high. Rates rangingfrom 20 to 232 nmol of N2 liter-' h-1 in RyanLake can be compared with a previously report-ed maximum rate in a lake of 11 nmol liter- h-1(9). Fixation at such high rates under theseenvironmental conditions has been found only inlakes in the blast zone of Mount St. Helens (3).During the summer of 1980, this represented amajor source of reduced nitrogen to Ryan Lake.
Nitrification in Ryan Lake was limited tooxygenated waters in the littoral zone and epi-limnion. In these areas, nitrification rates werecomparable to other eutrophic lake ecosystems(Table 4). Nitrification rates in a small lakeenriched with human waste material and anestuary in an urbanized area had significantlyhigher rates. Although Ryan Lake received highlevels of carbon and inorganic nutrients, verylittle nitrogen was introduced into the lake as aresult of the eruption. All new nitrogen wouldhave to be derived initially from N2 fixation, anddissimilative nitrogen cycle processes would benecessary for net production of oxidized nitro-gen compounds. Competition for NH4' by het-erotrophic bacteria in Ryan Lake possibly limit-ed the numbers of active nitrifying bacteria (4),but a potential nitrifying population was presentin the oxygenated portion of the lake and is
TABLE 5. Rates of potential denitrification in waterand sediment on a volume basis each day
Rate
Place Site (,umol of Refer-N liter- enceday-')
Smith Lake, Water 1.1 25Ala.
Smith Lake, Water and 5.7 25Ala. sediment
Lake Mendota, Water 0.6-1.9 10Wis.
Lake Mendota, SedimentWis. (calcare- 860 13
ous)Trout Lake, Sediment 390 13
Wis.Lake Kinneret, Water 2.9-35.7 12
IsraelGrasmere Tarn, Water (epi- 0-0.03 28England limnion)
Grasmere Tarn, Water (hypo- 0.3-0.4 28England limnion)
Grasmere Tarn, Sediment (top 1,430 28England 1.5 cm)
Denmark Fjord sedi- 12.5 35ment
Ryan Lake, Water 0-32 ThisWash. paper
predicted to be the source of N03- for algae anddenitrifying bacteria.The rates of denitrification measured in the
anaerobic hypolimnia of Ryan Lake demonstrat-ed that a portion of the bacterial community hadthe potential to grow with NO3 as an alterna-tive electron acceptor (Table 5). Only LakeKinneret in Israel has been reported to contain amicrobial population capable of similar potentialdenitrification rates in the water column. TheMn-S-oxidizing bacteria were the most abundantgroup of denitrifying bacteria identified in RyanLake (3). These denitrifying bacteria, whichwere capable of oxidizing manganese and organ-ic compounds, occurred throughout the anaero-bic water column in large numbers. This isunlike other lakes in which denitrifying metaloxidizers are often limited to the oxycline (23). Itis postulated that during the summer of 1980, asa result of denitrification, the concentrations ofNO3 in Ryan Lake were at or near undetect-able concentrations. Although potential denitri-fication was not measured in the sediment ex-cept in the littoral zones, activity in thesediments would be expected to be orders, ofmagnitude above water levels (Table 5; 13, 28)and should continue to be a major sink fornitrogen even as anaerobiosis in the water col-umn dissipates.Based on the chemical and microbial condi-
tions encountered in lakes around Mount St.
1642 DAHM ET AL.
on April 29, 2020 by guest
http://aem.asm
.org/D
ownloaded from

VOLCANIC EFFECTS ON LAKE MICROBIOLOGY 1643
TABLE 6. Dominant ongoing microbial and biogeochemical processes as determined by degree of impact tolakes near Mount St. Helens
Degree of Processes and parametersa Representativeimpact 02 levels Nitrogen cycle Carbon cycle Sulfur cycle Metal cycles lakes
Heavy Anaerobic 1. N2 fixation 1. Fermenta- 1. S04 reduc- 1. Pyrite for- Spirit, North
Moderate Some 02 inepilimnion
(anaerobic)(S, W)
2. NH3 andorganic ni-trogen as-similation(S, W)
1. N2 fixation(S, W)(aerobic,anaerobic,and photo-trophic)
2. Nitrifica-tion (W)
3. Denitrifica-tion (S, W)
tion (S, W)2. Methano-
genesis (S)3. Anaerobic
photosyn-thesis (W)
1. Fermenta-tion (S, W)
2. Hetero-trophic oxi-dation ofDOC (W)
3. CH4 and H2oxidation(W)
4. Aerobic andanaerobicphotosyn-thesis (W)
5. Methano-genesis (S)
tion (S, W)2. Bacterial
photosyn-thesis (W)
1. S04 reduc-tion (S, W)
2. Bacterialphotosyn-thesis (W)
3. H2S oxida-tion (W)
mation (S,W)
1. Mn2' andFe2+ oxi-dation (W)
2. Pyrite for-mation (S)
Coldwater,CastleCreekb
Ryan, St. Hel-ens, Meta,Fawn
Light 02 to sedi-mentsyear-round
1. Hetero-trophic andphoto-trophic N2fixation(aerobic)(S, W)
2. Nitrifica-tion (S, W)
3. Denitrifica-tion (S)
1. Fermenta-tion (S)
2. Hetero-trophic oxi-dation ofDOC (W,S)
3. CH4 and H2oxidation(W)
4. Photosyn-thesis (aer-obic) (W)
5. Methano-genesis (S)
1. S04 reduc-tion (S)
2. H25 oxida-tion (W, S)
1. Dependent McBride, Mer-on metal rill, Juneconcentration
a S, Process occurring in sediments; W, process occurring in the water column.b These lakes were completely anaerobic only during the summer of 1980.
Helens, a general schematic of lake processes asthe lakes progress toward recovery (a return tooligotrophic alpine or subalpine lake conditions)is presented in Table 6. Many of the microbialprocesses listed were either measured directlyor inferred from chemical characteristics ob-tained from several blast zone lakes. We havearbitrarily categorized these lakes according totheir degree of impact as heavily, moderately, orlightly impacted. Ryan Lake is classified asmoderately impacted since it retained an oxy-genated epilimnion. In these lakes, aerobic mi-crobial processes including nitrification, photo-trophic nitrogen fixation, and various oxidativeheterotrophic and chemolithotrophic reactions
contribute to lake recovery by adding nitrogenand oxygen and diminishing the reducing poten-tial in the water column.The eruption of Mount St. Helens induced
ultraeutrophic conditions in numerous lakesnear the volcano. However, unlike other ul-traeutrophic systems, the enrichment processwas dominated by a single event of 1 day and notby cumulative inputs over decades or centuries.In addition, the nutrient enrichment to thesesystems included enormous quantities of sulfurand metals in addition to organic carbon. Bacte-ria responded to these conditions and dominatedthe anaerobic water columns of the lakes. Sincethe input of nutrient material was basically a
VOL. 45, 1983
on April 29, 2020 by guest
http://aem.asm
.org/D
ownloaded from

APPL. ENVIRON. MICROBIOL.
one-time event and the initial microbial activityso intense, a unique opportunity is provided tostudy bacterial and algal activity, succession,and species composition in lakes shifting fromultraeutrophy to oligotrophy in time framesmuch shorter than that encountered in otherecosystems where pollution abatement, but notcessation, is ongoing.
ACKNOWLEDGMENTSWe thank R. Kepler for technical and graphical assistance,
M. DeAngelis for microbiological analyses and field assist-ance, N. Aumen and R. Karlin for help in sampling andchemical analyses, and C. Thomas and M. Davis for manu-script typing.The U.S. Forest Service provided helicopter time and
support for supplies. The National Science Foundation sup-ported manuscript preparation under project DEB-811307.
LITERATURE CITED1. American Public Health Association. 1971. Standard meth-
ods for the examination of water and wastewater. 13th ed.American Public Health Association, Inc., Washington,D.C.
2. Atlas, E. L., L. I. Gordon, S. W. Hager, and P. K. Park.1971. A practical manual for use of the Technicon Autoan-alyzer in seawater nutrient analyses. Oregon State Uni-versity technical report 71-22. Oregon State University,Corvallis.
3. Baross, J. A., C. N. Dahm, A. K. Ward, M. D. Lilley, andJ. R. Sedell. 1982. Initial microbiological response in lakesto the Mt. St. Helens eruption. Nature (London) 296:49-52.
4. Belser, L. W. 1979. Population ecology of nitrifying bacte-ria. Annu. Rev. Microbiol. 33:309-333.
5. Belser, L. W., and E. L. Mays. 1980. Specific inhibition ofnitrite oxidation by chlorate and its use in assessingnitrification in soils and sediments. Appl. Environ. Micro-biol. 39:505-510.
6. Bendschneider, K., and R. J. Robinson. 1952. A newspectrophotometric method for the determination of ni-trite in sea water. J. Mar. Res. 11:87-96.
7. Bollen, W. B. 1953. Carbon and nitrogen in farm andforest products. Agric. Food Chem. 1:379-381.
8. Bortleson, G. C., N. P. Dion, J. B. McConnell, and L. M.Nelson. 1976. Reconnaissance data on lakes in Washing-ton. Washington Department of Ecology Water SupplyBulletin 43, Olympia, Wash.
9. Brezonik, P. L., and C. L. Harper. 1969. Nitrogen fixationin some anoxic lacustrine environments. Science164:1277-1279.
10. Brezonik, P. L., and G. F. Lee. 1968. Denitrification as anitrogen sink in Lake Mendota, Wis. Environ. Sci. Tech-nol. 2:120-125.
11. Carpenter, J. M. 1965. The Chesapeake Bay Institutetechnique for the Winkler dissolved oxygen method.Limnol. Oceanogr. 10:141-143.
12. Cavari, B. Z., and G. Phelps. 1977. Denitrification in LakeKinneret in the presence of oxygen. Freshwater Biol.7:385-391.
13. Chen, R. L., D. R. Keeney, D. A. Graetz, and A. J. Hold-ing. 1972. Denitrification and nitrate reduction in Wiscon-sin lake sediments. J. Environ. Qual. 1:158-162.
14. Cohen, Y. 1977. Shipboard measurement of dissolvednitrous oxide in seawater by electron capture gas chroma-tography. Anal. Chem. 49:1238-1240.
15. Cohen, Y. 1978. Consumption of dissolved nitrous oxidein an anoxic basin, Saanich Inlet, British Columbia.Nature (London) 272:235-237.
16. Cook, R. J., J. C. Barron, R. I. Papendick, and G. J.Williams III. 1981. Impact on agriculture of the Mt. St.Helens eruptions. Science 211:16-22.
17. Dahm, C. N. 1981. Pathways and mechanisms for removalof dissolved organic carbon from leaf leachate in streams.Can. J. Fish. Aquat. Sci. 38:68-76.
18. Dahm, C. N., J. A. Baross, M. D. Lilley, A. K. Ward, andJ. R. Sedeil. 1982. Lakes in the blast zone of Mt. St.Helens: chemical and microbial responses following theMay 18, 1980 eruption, p. 98-137. In W. H. Funk (ed.),Conference on Mt. St. Helens: effects on water resources.Pullman, Wash.
19. Delwiche, C. C. 1978. Biological production and utiliza-tion of N20. Pure Appl. Geophys. 116:414-422.
20. Dion, N. P., and S. S. Embrey. 1981. Effects of Mt. St.Helens eruption on selected lakes in Washington. U.S.Geological Survey Circular: 850-6.
21. Docters vanLeeuwen, W. M. 1936. Krakatua, 1883 to 1933.Ann. Jard. Bot. Buitenz. 46-47:1-506.
22. Driscoll, J. N., E. S. Atwood, J. E. Fowler, and W. Selig.1980. Electrodes measure dissolved ammonia and carbondioxide. Ind. Res. Dev. August:91-94.
23. Dubinina, G. A., V. M. Gorlenko, and Y. I. Suleimanov.1974. A study of microorganisms involved in the circula-tion of manganese, iron, and sulfur in meromictic LakeGek-Gel. Microbiology (U.S.S.R.) 42:817-822.
24. Fruchter, J. S., D. E. Robertson, J. C. Evans, K. B. Ol-sen, E. A. Lepel, J. C. Laul, K. H. Abel, R. W. Sanders,P. O. Jackson, N. S. Wogman, R. W. Perkins, H. H. van-Tuyl, R. H. Beauchamp, J. W. Shade, J. L. Daniel, R. L.Erickson, G. A. Schmel, R. N. Lee, A. V. Robinson, 0. R.Moss, J. K. Briant, and W. C. Cannon. Mt. St. Helens ashfrom the 18 May 1980 eruption: chemical, physical, miner-alogical, and biological properties. Science 209:1116-1125.
25. Goering, J. J., and V. A. Dugdale. 1966. Estimates of therates of denitrification in a subarctic lake. Limnol. Ocean-ogr. 11:113-117.
26. Hardy, R. W. F., R. D. Holstein, E. K. Jackson, andR. C. Burns. 1968. The acetylene-ethylene assay from N2-fixation: laboratory and field evaluation. Plant Physiol.43:1185-1207.
27. Hobble, J. E., R. J. Daley, and S. Jasper. 1977. Use ofNuclepore filters for counting bacteria by fluorescencemicroscopy. Appl. Environ. Microbiol. 33:1225-1228.
28. Jones, J. G. 1979. Microbial nitrate reduction in freshwa-ter sediments. J. Gen. Microbiol. 15:27-35.
29. Knowles, R. 1979. Denitrification, acetylene reduction,and methane metabolism in lake sediment exposed toacetylene. Appl. Environ. Microbiol. 38:486-493.
30. Lilley, M. D., J. A. Baross, and L. I. Gordon. 1982. Dis-solved hydrogen and methane in Saanich Inlet, BritishColumbia. Deep-Sea Res. 29:1471-1484.
31. Mague, T. H. 1977. Ecological aspects of dinitrogen fixa-tion by blue-green algae, p. 85-140. In R. W. F. Hardyand A. H. Gibson (ed.), A treatise on dinitrogen fixation,section IV: agronomy and ecology. Wiley-Interscience,New York.
32. Menzel, D. W., and N. Corwin. 1965. The measurement oftotal phosphorus in seawater based on the liberation oforganically bound fractions by persulfate oxidation. Lim-nol. Oceanogr. 10:280-282.
33. Menzel, D. W., and R. F. Vaccaro. 1964. The measure-ment of dissolved organic and particulate carbon in sea-water. Limnol. Oceanogr. 9:138-142.
34. Nealson, K. H., and J. Ford. 1980. Surface enhancementof bacterial manganese oxidation: implications for aquaticenvironments. Geomicrobiol. 2:21-37.
35. Oren, A., and T. H. Blackburn. 1979. Estimation ofsediment denitrification rates at in situ nitrate concentra-tions. Appl. Environ. Microbiol. 37:174-176.
36. Park, P. K. 1969. Oceanic CO2 system: an evaluation often methods of investigation. Limnol. Oceanogr. 14:179-186.
37. Park, P. K. 1969. Carbon dioxide partial pressure in theColumbia River. Science 166:867-868.
38. Postgate, J. R. 1972. The acetylene reduction test fornitrogen fixation, p. 343-356. In J. R. Norris and D. W.
1644 DAHM ET AL.
on April 29, 2020 by guest
http://aem.asm
.org/D
ownloaded from

VOLCANIC EFFECTS ON LAKE MICROBIOLOGY 1645
Gibbons (ed.), Methods in microbiology, vol. 6b. Aca-demic Press, Inc., London.
39. Potts, M., W. E. Krumbein, and J. Metzger. 1978. Nitro-gen fixation rates in anaerobic sediments determined byacetylene reduction, a new "N field assay, and simulta-neous total N-15N determination, p. 753-769. In W. E.Krumbein (ed.), Environmental biogeochemistry and geo-
microbiology, vol. 3. Ann Arbor Scientific Publications,Inc., Ann Arbor, Mich.
40. Staley, J. T., L. G. Lehmicke, F. E. Palmer, R. W. Peet,and R. C. Wissmar. 1982. Impact of Mount St. Helenseruption on bacteriology of lakes in the blast zone. Appl.Environ. Microbiol. 43:664-670.
41. Stewart, W. D. P., G. P. Fitzgerald, and R. H. Burris.1967. In situ studies on N2 fixation using the acetylenereduction technique. Proc. Natl. Acad. Sci. U.S.A.58:2071-2078.
42. Stookey, L. L. 1970. Ferrozine-a new spectrophotomet-ric reagent for iron. Anal. Chem. 42:779-781.
43. Tiedje, J. M. 1978. Denitrification in soils. p. 362-366. In
D. Schiessinger (ed.), Microbiology-1978. American So-ciety for Microbiology, Washington, D.C.
44. Vincent, W. F., and M. T. Downes. 1981. Nitrate accumu-lation in aerobic hypolimnia: relative importance of ben-thic and planktonic nitrifiers in an oligotrophic lake. Appl.Environ. Microbiol. 42:565-573.
45. Wetzel, R. G., and B. A. Manny. 1972. Decomposition ofdissolved organic carbon and nitrogen compounds fromleaves in an experimental hard-water stream. Limnol.Oceanogr. 17:927-931.
46. Wsnmar, R. C., A. H. Devol, A. E. Nevissi, and J. R.Sedell. 1982. Chemical changes of lakes within the Mt. St.Helens blast zone. Science 216:175-178.
47. Wissmar, R. C., A. H. Devol, J. T. Staley, and J. R.Sedell. 1982. Biological responses of lakes in Mt. St.Helens blast zone. Science 216:178-181.
48. Zimmerman, R., and L. A. Meyer-Reil. 1974. A new
method for fluorescence staining of bacterial populationson membrane filters. Kiel. Meeresforsch. 30:24-27.
VOL. 45, 1983
on April 29, 2020 by guest
http://aem.asm
.org/D
ownloaded from