electrochemical proton gradient in genetic …jb.asm.org/content/157/3/868.full.pdf · role ofthe...
TRANSCRIPT

Vol. 157, No. 3JOURNAL OF BACTERIOLOGY, Mar. 1984, p. 868-8730021-9193/84/030868-06$02.00/0Copyright © 1984, American Society for Microbiology
Role of the Electrochemical Proton Gradient in GeneticTransformation of Haemophilus influenzae
WOUT BREMER,1 JAN KOOISTRA,2 KLAAS J. HELLINGWERF,' AND WIL N. KONINGS1*Departments of Microbiology' and Genetics, 2 Biological Center, University of Groningen, Kerklaan 30, 9751 NN Haren,
The Netherlands
Received 22 August 1983/Accepted 6 December 1983
The uptake of homologous DNA by Haemophilus influenzae was studied as a function of the protonmotive force in completely competent cultures in the pH range of 6 to 8. The composition and magnitude ofthe proton motive force were varied by using the ionophores valinomycin and nigericin (in the presence ofvarious potassium ion concentrations) and by using protonophores. No interaction of the ionophores withthe DNA transformation system itself was observed. Either component of the proton motive force, theelectrical potential or the pH gradient, can drive the uptake ofDNA, and the extent of the uptake of DNA isultimately determined by the total proton motive force. The transformation frequency increases with theproton motive force, which reaches a maximum value at around -130 mV. These results are consistent withan electrogenic proton-DNA symport mechanism, but direct evidence for such a system is not available.The proton motive force was followed during competence development of H. influenzae at pH 8. In theinitial phase (up to 50 min), the proton motive force remained constant at about -90 niV, whereas thetransformation frequency rose steeply. In the second phase, the proton motive force increased. Thetransformattion frequency in this phase increased with the proton motive force, as in completely competentcultures. These observations and the observed inhibition by NAD of both the proton motive force and thetransformation frequency indicate that structural components of the competent state are formed in the initialphase of competence development, whereas the second phase is characterized by an increase of the protonmotive force.
Uptake of DNA by bacterial cells can occur via threedistinct processes: phage infection, conjugation, and genetictransformation. Recently attention has been directed to-wards the energetics of these processes. A growing body ofevidence indicates that the proton motive force plays an
essential role in DNA transport during phage infection (11,12, 17) and bacterial conjugation (7).The role of the proton motive force or its components, the
membrane potential (At) and the pH gradient (ApH), hasalso been studied in genetic transformation in the gram-positive organisth Bacillus subtilis (1, 24) and in the gram-negative organism Escherichia coli (23). The results of thesestudies suggest that the proton motive force plays an essen-tial role in genetic transformation in both organisms. Grinius(6) postulated that DNA transport is driven by the protonmotive force and occurs in essence via an electrogenicproton DNA symport mechanism. This hypothesis, there-fore, places DNA transport within the conceptual frame-work of the chemiosmotic hypothesis.
It should be realized that the experimental evidence insupport of such a proton-DNA symport mechanism for DNAuptake during genetic transformation is not very strong atthis moment. First of all, the role of the proton motive forcein DNA transformation has been studied in cultures in whichonly a fraction of the cells were capable of transporting DNA(in B. subtilis, at the most 20% of the cells of the culturebecome competent for DNA uptake [25]). An estimation ofthe proton motive force in competent cells of these culturesis not possible, and consequently the relationship betweenthe proton motive force and DNA uptake cannot be studieddirectly. Secondly, in our studies on the role of the protonmotive force in transformation in B. subtilis, we observed
* Corresponding author.
868
that the ionophores valinomycin and nigericin not only hadan effect on the components of the proton motive force butalso interacted directly with the binding sites of DNA in thecytoplasmic membrane (24). Recently, it was demonstratedthat these artifacts strongly depend on the concentrationof the ionophores (Griniuviene et al., Abstr. FEMS Int.Symp. Environ. Regulation Microb. Metab., Pushchino,U.S.S.R., 1983, p. 216). Most of the evidence for a role ofthe proton motive force in DNA uptake is based on studieswith these ionophores without taking into account thesepossible side effects (see also reference 23). These experi-mental difficulties make B. subtilis and E. coli less than idealorganisms for studies on the relationship between the protonmotive force and DNA uptake.
Studies on DNA transformation in Haemophilus influen-zae showed that cultures can be obtained in which essential-ly all cells are competent for DNA uptake (9). Here wedemonstrate that in H. influenzae the ionophores valinomy-cin and nigericin do not interfere directly with DNA bindingor DNA transport. H. influenzae therefore appears to meetthe optimal requirements for an investigation on the role ofthe proton motive force in DNA tiptake in the transformationprocess. This paper describes the results of these studies.
MATERIALS AND METHODSBacterial strains. DNA uptake was studied in the recipient
Rd strain of H. influenzae, which is sensitive to 1 ,ug ofnovobiocin, 1 ,ug of erythromycin, and 3 ,ug of streptomycinper ml (16). Donor DNA was isolated from a Novr Eryrstrain of H. influenzae, resistant to 2.5 ,ug of novobiocin and6 jig of erythromycin per ml and sensitive to 3 jig ofstreptomycin per ml. Cells were made competent in an M-IVmedium according to the method of Herriott et al. (9). By thismethod, completely competent cultures could be obtained.
on May 10, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

TRANSFORMATION IN H. INFLUENZAE 869
Media preparation of donor DNA, and transformationassays. The media, preparation of donor DNA, and transfor-mation assays were as described previously (15, 16). Thetransformation assays were performed in the very complexM-IV medium. In experiments with ionophores, controlsamples received equivalent amounts of ethanol.Measurements of the electrical potential and pH gradient.
The electrical potential was determined from the distributionof tritium-labeled tetraphenylphosphonium as described pre-viously (8). Attempts were made to correct for binding oftetraphenylphosphonium to cellular components by sub-tracting the uptake of tetraphenylphosphonium in the pres-ence of the uncoupler carbonylcyanide p-trifluoromethoxy-phenylhydrazone (10 ,uM). This procedure can lead to anoverestimation of the A'V (19). A proper correction forbinding would require a study of tetraphenylphosphonium-binding properties of the different cellular components. Atthis stage of the investigation, these studies have not beenperformed. The obtained AT values are, however, a goodapproximation of the actual APV values (18). The ApH wasestimated by measiacid as described prAt and ApH, a cellused. This volumetritium-labeled wattously (24).
n)J
LLIllJ
(-)
LLI
C-
0
(-)
IL
0
z
0
(-)
U-
TIME (TM Z N
FIG. 1. DevelopmeIV medium (pH 8). Thin samples by incubdouble-stranded Novraddition of DNase (50of donor DNA, cells Imedium, the percentagcells and 2.02% fortransformants was 0.0viable cells per ml wa:
TABLE 1. The effect of ionophores on the transformationpercentage at different potassium concentrationsa
% Transformation at:Addition pH 6.0 pH 8.0of KCI(mM) Con-l Valinomycin Nigericin Con- Valinomycin Nigeri-
10 0.32 0.25 0.14 1.00 0.35 0.9175 0.32 0.13 0.56 x 10-1 0.79 0.56 x 10-1 0.71150 0.20 0.25 x 10-1 0.79 x 10-2 0.35 0.63 x 10-3 0.22
a Competent cells were suspended in M-IV medium (containing80 mM NaCI) at pH 6.0 or 8.0 with the indicated final concentrationsof potassium chloride and incubated for 15 min in the presence orabsence of valinomycin (final concentration, 10 ,uM) or nigericin(final concentration, 4 ,uM). (To the control samples was added anamount of ethanol equivalent to that of the ionophores in the othersamples.) Then cells were incubated for 15 min in the presence ofdonor DNA. DNase (50 ,g/ml) and MgCl2 (10 mM) were added, andthe cells were plated to determine the transformation percentage.
uring the distribution of [7-'4C]benzoic Materials. [7-l4C]benzoic acid (specific activity, 108reviously (24). For the calculation of the Ci/mol), 3H20 (specific activity, 91 mCi/mol), and D-[U-volume of 1.8 l.1/mg of cell protein was 14C]sorbitol (specific activity, 333 Ci/mol) were obtainedwas determined from the distribution of from the Radiochemical Centre, Amersham Corp., Bucking-er and [14C]sorbitol as described previ- hamshire, Great Britain, and [U-3H]tetraphenylphosphon-
ium bromide (specific activity, 2.5 Ci/mmol) was from theNuclear Research Centre, Negev, Beer-Sheva, Israel. 3,5-Di-tert-butyl-4-hydroxybenzylidenemalononitrite (SF-6847)was a gift of Y. Nishizawa (Sumitomo Chemical Industry,Osaka, Japan). All other chemicals were reagent grade andwere obtained from commercial sources.Development of competence. In transformation of H. in-
fluenzae a high degree of competence can be achieved. Wedetermined the fraction of cells in a culture that was compe-tent (fe) from the method first introduced by Goodgal andHerriott (5) and corrected by Porter and Guild (21). Usingthe equation f. = (NA * NB)/(NAB * c * 2), in which NA andNB are the observed numbers of single transformants toindependent markers A and B, NAB is the number of doubletransformants, resulting from the random integration into therecipient genome, and c is the number of viable cells, wefound that f. increased essentially up to 1 during incubationin the competence-inducing M-IV medium (Fig. 1). Thisindicates that under the experimental conditions employedall the cells in the culture became competent.The effect of the proton motive force on DNA uptake. The
proton motive force of many bacteria can be varied by theaddition of the ionophores valinomycin and nigericin in thepresence of potassium ions. Valinomycin mediates the elec-trogenic uniport of potassium ions across the cytoplasmicmembrane; nigericin mediates the electroneutral antiport of
IIf| l potassium ions versus protons. The primary effect of valino-25 50 75 100 mycin is a decrease of the electrical potential, and the
primary effect of nigericin is a decrease of the pH gradientiin) OF INCUBATION IN THE across the cytoplasmic membrane. In a previous publication4EDIUM (24) it was shown that both ionophores can interact with the
binding site of DNA in the cytoplasmic membrane of B.nt of competence in H. influenzae cells in M- subtilis. It was therefore important to investigate the directe transformation percentage was determined effects of these ionophores on the uptake system of DNA inating the recipient cells with homologous H. influenzae. The studies were done at two pH values andEryg DNA (0.2 (1g/ml) for 15 min before the at different concentrations of potassium ions in M-IV medi-adg/ml)andMiba2(tdforM) mi in the M-IV um, which contains 80 mM sodium chloride (Table 1). The
he of single transformants was 2.41% for Novr results show that only in the presence of potassium did bothEryr cells, and the percentage of double ionophores inhibit strongly the transformation frequency at26% for Novr Eryr cells. The total number of pH 6, whereas only valinomycin exerted a strong inhibitions 4.1 x 108. at pH 8. The extent of the inhibition depends on the
VOL. 157, 1984
on May 10, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
870 BREMER ET AL.
potassium concentration and is maximal at the highestpotassium concentration used. These observations stronglyindicate that the ionophores do not interact with the DNAuptake system itself, since the inhibitory effect depends onthe potassium concentration and on the medium pH.The effect of the proton motive force on DNA uptake was
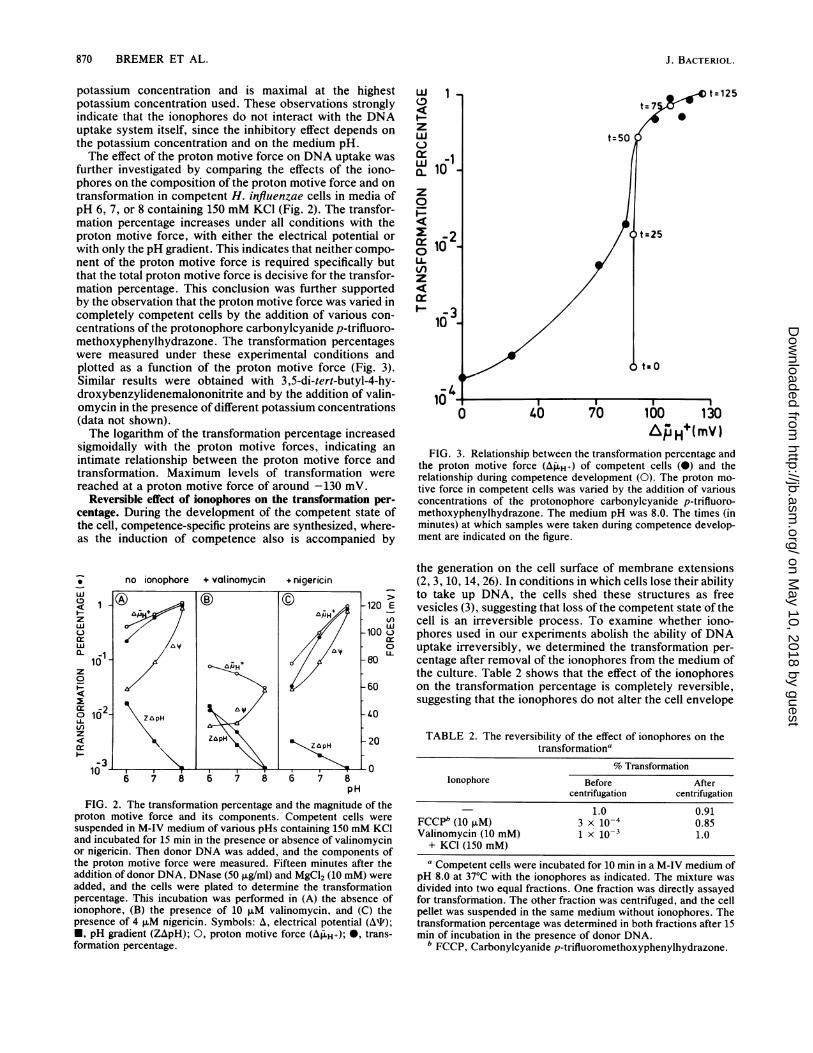
further investigated by comparing the effects of the iono-phores on the composition of the proton motive force and ontransformation in competent H. influenzae cells in media ofpH 6, 7, or 8 containing 150 mM KCI (Fig. 2). The transfor-mation percentage increases under all conditions with theproton motive force, with either the electrical potential orwith only the pH gradient. This indicates that neither compo-nent of the proton motive force is required specifically butthat the total proton motive force is decisive for the transfor-mation percentage. This conclusion was further supportedby the observation that the proton motive force was varied incompletely competent cells by the addition of various con-centrations of the protonophore carbonylcyanide p-trifluoro-methoxyphenylhydrazone. The transformation percentageswere measured under these experimental conditions andplotted as a function of the proton motive force (Fig. 3).Similar results were obtained with 3,5-di-tert-butyl-4-hy-droxybenzylidenemalononitrite and by the addition of valin-omycin in the presence of different potassium concentrations(data not shown).The logarithm of the transformation percentage increased
sigmoidally with the proton motive forces, indicating anintimate relationship between the proton motive force andtransformation. Maximum levels of transformation werereached at a proton motive force of around -130 mV.
Reversible effect of ionophores on the transformation per-centage. During the development of the competent state ofthe cell, competence-specific proteins are synthesized, where-as the induction of competence also is accompanied by
no ionophore + valinomycin0LLIJ
z
cLJL.)ca
a-
z0
0
LA.
z
4r
+ nigericin
7 8 6 7 8 6 7
E
LL~wI v
0
U-
pHFIG. 2. The transformation percentage and the magnitude of the
proton motive force and its components. Competent cells weresuspended in M-IV medium of various pHs containing 150 mM KCland incubated for 15 min in the presence or absence of valinomycinor nigericin. Then donor DNA was added, and the components ofthe proton motive force were measured. Fifteen minutes after theaddition of donor DNA, DNase (50 p.g/ml) and MgCl2 (10 mM) wereadded, and the cells were plated to determine the transformationpercentage. This incubation was performed in (A) the absence ofionophore, (B) the presence of 10 ,uM valinomycin, and (C) thepresence of 4 ,uM nigericin. Symbols: A, electrical potential (A'T);*, pH gradient (ZApH); 0, proton motive force (AjH+); 0, trans-formation percentage.
W 1-
zW
CL, 1
w
0r 10 2.z0
x 2QLL
LAlz
1
-1410
it=25
taO
100 130A H+(mV)
FIG. 3. Relationship between the transformation percentage andthe proton motive force (AfH+) of competent cells (@) and therelationship during competence development (0). The proton mo-tive force in competent cells was varied by the addition of variousconcentrations of the protonophore carbonylcyanide p-trifluoro-methoxyphenylhydrazone. The medium pH was 8.0. The times (inminutes) at which samples were taken during competence develop-ment are indicated on the figure.
the generation on the cell surface of membrane extensions(2, 3, 10, 14, 26). In conditions in which cells lose their abilityto take up DNA, the cells shed these structures as freevesicles (3), suggesting that loss of the competent state of thecell is an irreversible process. To examine whether iono-phores used in our experiments abolish the ability of DNAuptake irreversibly, we determined the transformation per-centage after removal of the ionophores from the medium ofthe culture. Table 2 shows that the effect of the ionophoreson the transformation percentage is completely reversible,suggesting that the ionophores do not alter the cell envelope
TABLE 2. The reversibility of the effect of ionophores on thetransformationa
% TransformationIonophore Before After
centrifugation centrifugation
1.0 0.91FCCPb (10 ,uM) 3 x 10-4 0.85Valinomycin (10 mM) 1 x 10-3 1.0+ KCI (150 mM)a Competent cells were incubated for 10 min in a M-IV medium of
pH 8.0 at 370C with the ionophores as indicated. The mixture wasdivided into two equal fractions. One fraction was directly assayedfor transformation. The other fraction was centrifuged, and the cellpellet was suspended in the same medium without ionophores. Thetransformation percentage was determined in both fractions after 15min of incubation in the presence of donor DNA.
b FCCP, Carbonylcyanide p-trifluoromethoxyphenylhydrazone.
J. BACTERIOL.
on May 10, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

TRANSFORMATION IN H. INFLUENZAE 871
in such a way that its competent structure, required for DNAuptake, is changed irreversibly. The results indicate that forDNA uptake not only the competent structure of the cellenvelope is required but also a certain magnitude of theproton motive force.The proton motive force during competence development.
Since the results obtained so far indicated the essentialfunction of the proton motive force in the uptake of DNA, itwas of interest to compare the proton motive force incompetent and noncompetent cells. These measurementswere made at different pH values (Fig. 4). The results showthat hardly any difference exists in the ApH values betweenboth types of cells but that the AP (and thus the protonmotive force) differs by 10 to 30 mV between competent andnoncompetent cells. The proton motive force at low pHvalues in this experiment was slightly higher than that shownin Fig. 2 as a result of the absence of 150 mM KCl. Duringcompetence development the electrical potential of the pro-ton motive force apparently increases. This aspect wasfurther investigated. The electrical potential was followedduring competence development in a medium of pH 8, acondition in which no ApH exists and the proton motive
E140-
100-
ApH
6 7 8pH
FIG. 4. The proton motive force of competent and noncompe-tent cells of H. influenzae at different medium pHs. Competencewas developed for 100 min in M-IV medium at pH 8.0. Cells were
washed and resuspended in M-IV medium at the pH indicated.Closed symbols, competent cells; open symbols, noncompetentcells. Squares, the proton motive force (AfH+); triangles, electricalpotential (AIP); circles, pH gradient (ZApH).
FIG. 5. Relationship between development of competence andthe proton motive force (AIIH+). Competence development occurredas described in the legend to Fig. 1. NAD (10 p.g/ml) was added after50 or 100 min of competence development. Closed symbols, no
inhibitor added; open symbols, in the presence of NAD. Circles,transformation percentage; squares, the proton motive force.
force is composed solely of a At. In this medium thefraction of component cells increased hyperbolically in time.The At remained constant at about -90 mV in the initialphase of competence development (up to 50 min) andincreased after 50 min gradually to -130 mV, when fullycompetent cultures were obtained (Fig. 5).
All studies described were performed in M-IV medium, inwhich the cells cannot grow and might gradually lose theirviability. Treatment with the ionophores could also lead to a
loss of viability. Since nonviable cells will contribute to thetotal internal volume of the culture but probably will notmaintain a proton motive force, a considerable fraction ofnonviable cells would lead to an underestimation of theproton motive force. The viability was therefore measuredunder the different experimental conditions used. Under no
condition was the viability decreased by more than 30%. Theproton motive force was therefore at the most underestimat-ed by 7 mV (59 log 1.3).Competence development. The results of the previous
section indicate that two processes can be distinguishedduring competence development. The first process, thesynthesis of components of the competent state, is apparent-ly finished within the first 50 min of competence develop-ment. The subsequent increase of competence is due to an
increase of the electrical potential. This conclusion is furtherstrengthened by the results from Fig. 3. The open circles inthat figure represent the relationship between the transfor-mation percentage and the proton motive force during com-
petence development. It is clear that in the first phase ofcompetence development the transformation percentage in-creases, whereas the proton motive force remains constant,indicating that components necessary for competence are
synthesized in this phase (0 to 50 min). In the second phase(50 to 100 min) exactly the same relationship between thetransformation percentage and the proton motive force is
o1 1400l-
0x 100z0
0
60-
20-
LLI
zLL
-10wa.
z0
10 <cr0
LAz
-2<-1Hz
-310
00 125TIME (min)
VOL. 157, 1984
on May 10, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

872 BREMER ET AL.
observed as in fully competent cells. Further evidence for aclose relationship between the increase in the proton motiveforce and the competence development in the second phaseis supplied by the results obtained with NAD, a compoundwhich can inhibit the development of the competent state ofH. influenzae (20). NAD does not inhibit transformationbecause the addition of NAD (10 ,ug/ml) to fully competentcells (after 100 min of incubation in M-IV medium) hardlyaffected the transformation frequency (Fig. 5). In agreementwith this observation, no effect on the proton motive forcewas observed. However, if NAD is added before the protonmotive force increase has occurred, a further increase ofboth the transformation percentage and the proton motiveforce is prevented (Fig. 5).
DISCUSSION
The transformation system of H. influenzae, which isfunctional at neutral pH, is involved in the uptake ofhomologous double-stranded DNA. Below pH 5.5 anothertransformation system becomes operational. This system isnot specific for homologous DNA and is also able totranslocate single-stranded DNA (14). In this paper only theproperties of the first system have been studied. H. influen-zae is an attractive organism for the study of the relationshipbetween the proton motive force and DNA uptake. Culturescan be obtained in which all cells are competent, and theeffects of variations in the magnitude and composition of theproton motive force on DNA uptake can be studied. Anadditional feature is that the ionophores valinomycin andnigericin appear to affect only the components of the protonmotive force and do not interfere with the DNA uptakesystem itself, as was observed in B. subtilis (24). Since theproperties of the DNA transformation systems of H. influen-zae and B. subtilis differ in several aspects (for a review seereference 4) it is not surprising that the interaction of theionophores with these transformation systems also differs.
In H. influenzae both components of the proton motiveforce, the A'T and the ApH, are involved in DNA uptake. Itis not clear at this moment what the exact role of the protonmotive force is in this uptake process. The proton motiveforce can function as a driving force for the translocationprocess across the cytoplasmic membrane. Such a transloca-tion process would be electrogenic and occur in symportwith protons since both the At and the ApH are involved.This would agree with the model proposed by Grinius (6) forDNA uptake in B. subtilis. Another possibility is that theproton motive force affects the structural and functionalproperties of the membrane-bound components of the trans-location system and thus controls the activity of theseproteins. Such a regulatory effect by the proton motive forcehas been demonstrated for several transport systems inwhich the proton motive force controls the redox state ofredox-sensitive groups in the membrane-bound proteins (13,22).A combination of these roles is also possible: the proton
motive force may function as a controlling factor for thefunctional properties and as a driving force for DNA uptake.The DNA uptake system would then behave as the protonmotive force-driven solute transport systems of E. coli (13).A surprising observation made in this paper is the role of
the proton motive force in the development of competence.In the initial phase of competence development, the protonmotive force remains constant, and competence-specificproteins and special cell wall structures (2, 3, 10, 14, 26) aresynthesized. In the second phase, the proton motive force
and especially the electrical potential increases, and thisincrease leads to an increase of the degree of competence.The mechanism behind this increase of the electrical
potential is far from clear at this moment and is subject tofurther investigation.
ACKNOWLEDGMENTThis study was supported by the Netherlands Organization for the
Advancement of Pure Research (ZWO).
LITERATURE CITED1. Chaustova, L. P., L. L. Grinius, B. B. Griniuviene, A. A.
Jasaitis, J. P. Kadziauskas, and R. J. Kiausinyte. 1980. Studieson energy supply for genetic processes. Involvement of mem-brane potential in genetic transformation of Bacillus subtilis.Eur. J. Biochem. 103:349-357.
2. Concino, M. F., and S. H. Goodgal. 1981. Haemophilus influen-zae polypeptides involved in deoxyribonucleic acid uptakedetected by cellular surface protein iodination. J. Bacteriol.148:220-231.
3. Deich, D. A., and L. C. Hoyer. 1982. Generation and release ofDNA-binding vesicles by Haemophilus influenzae during induc-tion and loss of competence. J. Bacteriol. 152:855-864.
4. Goodgal, S. H. 1982. DNA uptake in Haemophilus transforma-tion. Annu. Rev. Genet. 16:169-192.
5. Goodgal, S. H., and R. M. Herriott. 1961. Studies on transfor-mation of Haemophilus influenzae. I. Competence. J. Gen.Physiol. 44:1201-1227.
6. Grinius, L. L. 1980. Nucleic acid transport driven by iongradient across cell membrane. FEBS Lett. 113:1-10.
7. Grinius, L., and J. Berzinskiene. 1976. Studies on DNA trans-port during bacterial conjugation. Role of proton motive force-generating H+-ATPase and respiratory chain. FEBS Lett.72:151-154.
8. Hellingwerf, K. J., J. G. M. Bolscher, and W. N. Konings. 1981.The electrochemical proton gradient generated by the fumaratereductase system in Escherichia coli and its bioenergetic impli-cations. Eur. J. Biochem. 113:369-374.
9. Herriott, R. M., E. M. Meyer, and M. Vogt. 1970. Definednongrowth media for stage II development of competence inHaemophilus influenzae. J. Bacteriol. 101:517-524.
10. Kahn, M. E., G. Maul, and S. H. Goodgal. 1982. Possiblemechanism for donor DNA binding and transport in Haemophi-lus. Proc. Natl. Acad. Sci. U.S.A. 79:6370-6374.
11. Kalasauskaite, E., and L. Grinius. 1979. The role of energy-yielding ATPase and respiratory chain at early stages of bacte-riophage T4 infection. FEBS Lett. 99:287-291.
12. Kalasauskaite, E., L. Grinius, D. Kadisaite, and A. Jasaitis.1980. Electrochemical H' gradient but not phosphate potentialis required for Escherichia coli infection by phage T4. FEBSLett. 117:233-236.
13. Konings, W. N., and G. T. Robillard. 1982. Physical mechanismfor regulation of proton solute symport in Escherichia coli.Proc. Natl. Acad. Sci. U.S.A. 79:5480-5484.
14. Kooistra, J., T. van Boxel, and G. Venema. 1980. Deoxyribonu-cleic acid-binding properties and membrane protein composi-tion of a competence-deficient mutant of Haemophilus influen-zae. J. Bacteriol. 144:22-27.
15. Kooistra, J., and G. Venema. 1970. Fate of donor DNA in somepoorly transformable strains of Haemophilus influenzae. Mutat.Res. 9:245-253.
16. Kooistra, J., and G. Venema. 1974. Fate of donor deoxyribonu-cleic acid in a highly transformation-deficient strain of Haemo-philus influenzae. J. Bacteriol. 119:705-717.
17. Labedan, B., and E. B. Goldberg. 1979. Requirement for mem-brane potential in infection of phage T4 DNA. Proc. Natl. Acad.Sci. U.S.A. 76:4669-4674.
18. Lolkema, J. S., A. Abbing, K. J. Hellingwerf, and W. N.Konings. 1983. The transmembrane electrical potential in Rho-dopseudomonas sphaeroides determined from the distributionof tetraphenyl phosphonium after correction for its binding tocell components. Eur. J. Biochem. 130:287-292.
J. BACTERIOL.
on May 10, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

TRANSFORMATION IN H. INFLUENZAE 873
19. Lolkema, J. S., K. J. Hellingwerf, and W. N. Konings. 1982. Theeffect of "probe binding" on the quantitative determination ofthe proton motive force in bacteria. Biochim. Biophys. Acta681:85-94.
20. Miller, D. H., and P. C. Huang. 1972. Identification of compe-tence-repressing factors during log-phase growth of Haemophi-lus influenzae. J. Bacteriol. 109:560-564.
21. Porter, R. D., and W. R. Guild. 1969. Number of transformableunits per cell in Diplococcus pneumoniae. J. Bacteriol. 97:1033-1035.
22. kobillard, G. T., and W. N. Konings. 1982. A hypothesis for therole of dithioldisulfide interchange in solute transport andenergy transducing processes. Eur. J. Biochem. 127:597-604.
23. Sabelnikov, A. G., and I. V. Domaradsky. i981. Effect ofmetabolic inhibitors on entry of exogenous deoxyribonucleicacid into Ca2 -treated Escherichia coli cells. J. Bacteriol.146:435-443.
24. Van Nieuwenhoven, M. H., K. J. Hellingwerf, G. Venema, andW. N. Konings. 1982. Role of proton motive force in genetictransformation of Bacillus subtilis. J. Bacteriol. 151:771-776.
25. Venema, G. 1979. Bacterial transformation. Adv. Microbiol.Physiol. 19:245-331.
26. Zooni K. C., M. Habersat, and J. J. Scocca. 1976. Synthesis ofenvelope polypeptides by Haemophilus influenzae during devel-opment of competence for genetic transformation. J. Bacteriol.127:545-554.
VOL. 157, 1984
on May 10, 2018 by guest
http://jb.asm.org/
Dow
nloaded from