electrophoretic study of clostridium species - journal of clinical
TRANSCRIPT

JOURNAL OF CLINICAL MICROBIOLOGY, Apr. 1982, p. 688-702 Vol. 15, No. 40095-1137/82/040688-15$02.00/0
Electrophoretic Study of Clostridium SpeciesELIZABETH P. CATO,* D. E. HASH, LILLIAN V. HOLDEMAN, AND W. E. C. MOORE
Department of Anaerobic Microbiology, Virginia Polytechnic Institute and State Univ,ersity, Blacksburg,Virginia 24061
Received 19 October 1981/Accepted 8 December 1981
Polyacrylamide gel electrophoretic analyses of soluble cellular proteins (with-out sodium dodecyl sulfate) of 70 Clostridium species indicated that the procedurewas readily applicable to the differentiation of species in the genus. The proteinpatterns correlated well with the available DNA homology data and with mostaccepted differential tests. Results indicated that several earlier names for specieswere synonyms of those of accepted species and that two accepted species may besynonymous.
Electrophoretic separation of cellular proteinsappears to be a highly sensitive analysis thatprovides distinctive phenotypic evidence of thesimilarity of strains within species. Variations ofthis technique are being used increasingly intaxonomic studies (3, 9, 13-15, 18, 28, 35; B.Biavati, V. Scardovi, and W. E. C. Moore, Int.J. Syst. Bacteriol., in press; K. Kersters and J.De Ley, Program Abstr. Int. Congr. Microbiol.,Int. Assoc. Microbiol. Soc. 12th, Munich, WestGermany, abstr. no. C25, 1978). Currently, elec-trophoresis is a useful and practical tool forscreening multiple isolates from a site for identi-fication and for verifying the identification ofstrains (22). It also may be useful for primaryidentification when reliable reference patternsare available. The present study represents afurther attempt to determine the reliability andusefulness of this relatively rapid and inexpen-sive procedure for purposes of bacterial identifi-cation and taxonomy.To assay the variation that may occur among
well-documented strains of Clostridium and toevaluate the distinctiveness of protein patternsamong similar and dissimilar species in thisgenus, we determined protein electrophoreticpatterns by the procedure of Moore et al. (22) forthe type strains and for up to 20 well-docu-mented reference strains of 70 Clostridium spe-cies.
MATERIALS AND METHODSBacterial strains. Strains examined included the type
strain of each species as published in the 1980 Ap-proved Lists of Bacterial Names (30) and other strainsrepresentative of these species. Strains were obtainedfrom the American Type Culture Collection (ATCC),the National Collection of Type Cultures, the NationalCollection of Industrial Bacteria, the National Collec-tion of Dairy Organisms, the Deutsche Sammlung vonMikroorganismen, and the Centers for Disease Con-trol from the collections of A. R. Prevot (Pasteur
Institute, Paris), L. S. McClung (University of Indi-ana), L. DS. Smith (Virginia Polytechnic Institute[VPI] Anaerobe Laboratory), and G. Hobbs (TorryResearch Station, Aberdeen, Scotland), or from ourcollection of isolates from normal floras or clinicalspecimens. All strains had been cultured and identifiedby the phenotypic methods described in the VirginiaPolytechnic Institute (VPI) Anaerobe LaboratoryManual (12). Sources of these strains, when known,are listed in the legends accompanying Fig. 1 through28. Unless otherwise stated, strain numbers are thoseof the VPI Anaerobe Laboratory. Names of speciesnot included in the 1980 Approved Lists of BacterialNames (30) are in quotation marks. An index of thespecies studied is given in Table 1.
Analytical methods. Cells from 24-h cultures in 5 mlof supplemented brain heart infusion broth (22) with0.1% CaCO3 and 0.025% Tween 80 were harvested bycentrifugation at 8,000 x g for 10 min. Sedimentedcells in conical tubes were suspended in 0.1 ml of 0.15M Tris-chloride buffer, and 0.15 g of 74- to 110-p.mdiameter glass beads was added. Tubes were placed inan ice bath, and cells were broken by shaking for two2-min periods. The resulting suspensions, with addedsucrose, were placed in 4.7% acrylamide stacking gelwells above an 8.5% acrylamide resolving gel. Nosodium dodecyl sulfate was used in the cell prepara-tions or gel solutions. Electrophoresis was run at roomtemperature at 150 V (constant) and at about 33 mA ofinitial current. Gels were stained with 0.08% Coomas-sie blue, destained with 10% glacial acetic acid (vol/volin water), and photographed. Details of the method aregiven in Moore et al. (22). The similarity or dissimilar-ity of the protein patterns was measured by directcomparison of cut photographs of the gels as describedby Biavati et al. (in press).
RESULTS AND DISCUSSIONThe results are shown in photographs of the
protein patterns obtained by electrophoresis(Fig. 1 to 28). Lane 1 on each gel containedsoluble proteins from Streptococcus faecalisVPI U4-20. Protein bands from this strain, par-ticularly the heavy band at approximately 4.5
688
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
cm o
n 10
Dec
embe
r 20
21 b
y 10
3.17
1.13
.219
.

ELECTROPHORESIS OF CLOSTRIDIUM SPECIES 689
TABLE 1. Index to Figures
Species Fig. no. Species Fig. no. Species Fig. no.
C. absonum 24 C. gboni 27 C. propionicum 21,25C. acidiurici 26 C. glycolicum 24 'C. pseudotetanicum' 19,20C. aminovalericum 23 C. baemolyticum 7,8 C. puniceum 24, 28C. aurantibutyricum 22 C. bastiforme 16, 28 C. purinolyticum 26C. barati 24 C. bistolyticum 23 C. putrefaciens 21C. barkeri 19 'C. imitans' 27,28 C. putrificum 22,27C. beijerinckii 1 'C. incertum' 27,28 C. quercicolum 22C. bifermentans 13 C. indolis 23 C. ramosum 5,17C. botulinum 8,9,10 C. innocuum 4,20 C. rectum 22C. butyricum 1,20 S. inulinus 21 C. sardiniensis 25C. cadaveris 15,25 C. irregularis 24 C. sartagoformum 19C. carnis 25 C. lentoputrescens 18 C. scatologenes 27C. celatum 23 C. leptum 23 C. septicum 6,14,25C. cellobioparum 23 C. limosum 27 C. sordellii 13C. cbauvoei 14,25 C. lituseburense 24,26 C. spbenoides 23C. clostridiiforme 23 C. malenominatum 18 C. spiroforme 17C. coccoides 24 C. mangenotii 24 C. sporogenes 9,11,12C. cocblearium 18 C. nexile 21 C. sporospbaeroides 24C. cocleatum 17 C. novyi 8,11,25 C. sticklandii 24'C. colinum' 24 C. oceanicum 24 C. subterminale 16,28C. difficile 12 C. oroticum 23 C. symbiosum 18C. durum 23 C. paraputrificum 19 C. tertium 23C. fallax 18 C. pasteurianum 25 C. tetani 15,22,25C. felsineum 19 'C. pectinovorum' 28 C. tyrobutyricum 25'C. fluxum' 19,20 C. perfringens 2,3 C. villosum 26C. formicoaceticum 22
cm, served as markers for gel variation. Whenpossible, strains of phenotypically similar spe-cies were placed on the same gel. Results fromtwo or more strains of the same species (Fig. 1 to17) are shown to illustrate the degree of varia-tion that was seen in the patterns. Patterns of thetype strains of single species are shown in Fig.18 through 28.
C. butyricum, the type species of the genusClostridium, and C. beijerinckii share many phe-notypic properties (12). Cummins and Johnson(5) showed that these two species are unrelatedby DNA-DNA homology and differ in their cellwall composition and nutritional requirements.Strains of C. butyricum grow in the third serialtransfer in a glucose-mineral salts-biotin medium(5), whereas strains of C. beijerinckii seldomgrow even in the first or second transfer. Bothspecies have been isolated from human softtissue infections and intestinal contents. Theprotein patterns (Fig. 1) were quite distinctivefor each species.
Pattern variation that may occur among toxintypes A, B, C, D, and E of C. perfringens isshown in Fig. 2 and 3. The only reliable methodthat has been found to identify the toxin type isneutralization with the specific antitoxin(s) (31).Each of these strains was toxic to mice, and the
toxin(s) of each was neutralized by specificantitoxin. The degree of variation in proteinpatterns was comparable to the amount of varia-tion observed in phenotypic characteristics ofthis species (12). With the possible exception ofa pair of fine bands at 9 and 10 mm in type Estrains (Fig. 3, lanes 11-13), the protein electro-phoretic patterns of all C. perfringens toxintypes were similar and could not be distin-guished from one another.Minor variations appeared in the protein pat-
terns of strains of C. innocuum, but the similar-ities were striking (Fig. 4). The pattern of Clos-tridium species strain 3794C was the mostdissimilar, which is not surprising. Johnson andFrancis (16) reported that DNA of strain 3794Chad only 41% DNA homology with DNA of thetype strain of C. innocuum. This indicates thatthis strain is closely related to C. innocuum butprobably should not be considered a member ofthe species. The relationship between the twostrains also is indicated by the 99% homologybetween ribosomal RNA preparations of 3794Cand the 23S rRNA of ATCC 14501, the typestrain of C. innocuum (16). Strain 3794C is oneof less than 10%o of the strains we had identifiedas C. innocuum that did not ferment sucrose.However, another sucrose-negative strain (5792;
VOL. 15, 1982
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
cm o
n 10
Dec
embe
r 20
21 b
y 10
3.17
1.13
.219
.
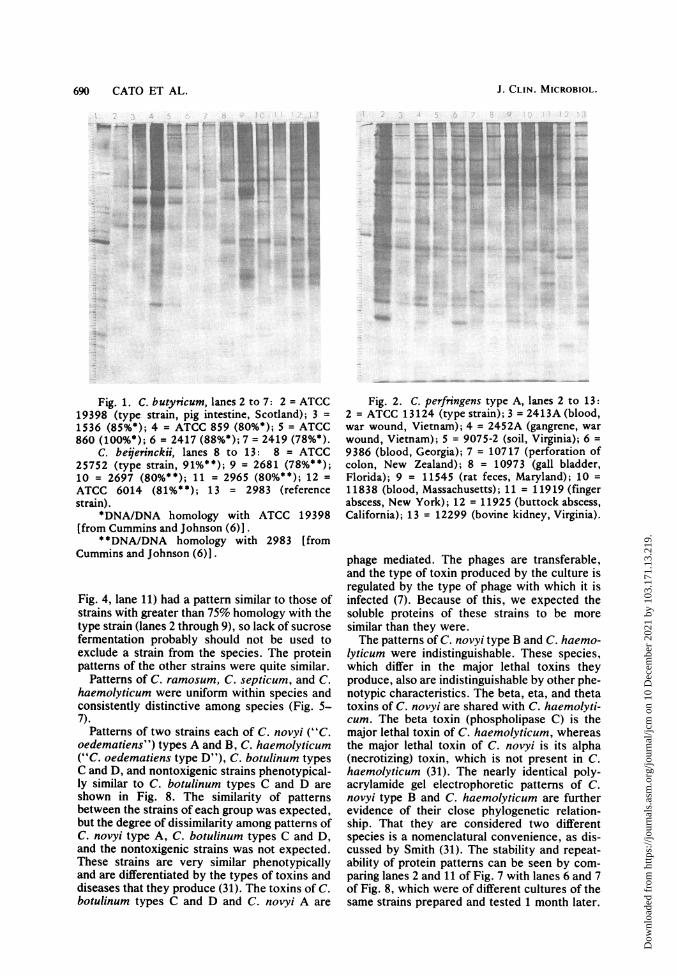
690 CATO ET AL.
Fig. 1. C. butynicum, lanes 2 to 7: 2 = ATCC19398 (type strain, pig intestine, Scotland); 3 =1536 (85%*); 4 ATCC 859 (80*); 5 = ATCC
860 (100%*); 6 =2417 (88%*); 7 = 2419 (78%*).C. beijerinckii, lanes 8 to 13: 8 = ATCC
25752 (type strain, 91%**); 9 = 2681 (78%**);10 = 2697 (80%**); 11 = 2965 (80%**); 12 =ATCC 6014 (81%**); 13 = 2983 (referencestrain).
*DNA/DNA homology with ATCC 19398[from Cummins and Johnson (6)].
* *DNA/DNA homology with 2983 [fromCummins and Johnson (6)].
Fig. 4, lane 11) had a pattern similar to those ofstrains with greater than 75% homology with thetype strain (lanes 2 through 9), so lack of sucrosefermentation probably should not be used toexclude a strain from the species. The proteinpatterns of the other strains were quite similar.
Patterns of C. ramosum, C. septicum, and C.haemolyticum were uniform within species andconsistently distinctive among species (Fig. 5-7).
Patterns of two strains each of C. novyi ("C.oedematiens") types A and B, C. haemolyticum("C. oedematiens type D"), C. botulinum typesC and D, and nontoxigenic strains phenotypical-ly similar to C. botulinum types C and D are
shown in Fig. 8. The similarity of patternsbetween the strains of each group was expected,but the degree of dissimilarity among patterns ofC. novyi type A, C. botulinum types C and D,and the nontoxigenic strains was not expected.These strains are very similar phenotypicallyand are differentiated by the types of toxins anddiseases that they produce (31). The toxins of C.botulinum types C and D and C. novyi A are
Fig. 2. C. perfringens type A, lanes 2 to 13:2 = ATCC 13124 (type strain); 3 = 2413A (blood,war wound, Vietnam); 4 = 2452A (gangrene, warwound, Vietnam); 5 = 9075-2 (soil, Virginia); 6 =9386 (blood, Georgia); 7 = 10717 (perforation ofcolon, New Zealand); 8 = 10973 (gall bladder,Florida); 9 = 11545 (rat feces, Maryland); 10 =11838 (blood, Massachusetts); 11 = 11919 (fingerabscess, New York); 12 = 11925 (buttock abscess,California); 13 = 12299 (bovine kidney, Virginia).
phage mediated. The phages are transferable,and the type of toxin produced by the culture isregulated by the type of phage with which it isinfected (7). Because of this, we expected thesoluble proteins of these strains to be moresimilar than they were.The patterns of C. novyi type B and C. haemo-
lyticum were indistinguishable. These species,which differ in the major lethal toxins theyproduce, also are indistinguishable by other phe-notypic characteristics. The beta, eta, and thetatoxins of C. novyi are shared with C. haemolyti-cum. The beta toxin (phospholipase C) is themajor lethal toxin of C. haemolyticum, whereasthe major lethal toxin of C. novyi is its alpha(necrotizing) toxin, which is not present in C.haemolyticum (31). The nearly identical poly-acrylamide gel electrophoretic patterns of C.novyi type B and C. haemolyticum are furtherevidence of their close phylogenetic relation-ship. That they are considered two differentspecies is a nomenclatural convenience, as dis-cussed by Smith (31). The stability and repeat-ability of protein patterns can be seen by com-paring lanes 2 and 11 of Fig. 7 with lanes 6 and 7of Fig. 8, which were of different cultures of thesame strains prepared and tested 1 month later.
J. CLIN. MICROBIOL.
IF
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
cm o
n 10
Dec
embe
r 20
21 b
y 10
3.17
1.13
.219
.

ELECTROPHORESIS OF CLOSTRIDIUM SPECIES 691
.. .., We x *
Fig. 3. C. perfringens type B, lanes 2 to 4:2 = ATCC 3626 (lamb intestine); 3 = CDC 88; 4 =2528 (bovine enterotoxemia).
C. perfringens type C, lanes 5 to 7: 5 = ATCC3628; 6 = 6800 (bird, Florida); 7 = 7049 (bovine,Virginia).
C. per/ringens type D, lanes 8 to 10: 8 =ATCC 3629 (lamb kidney); 9 = 6442 (bovineintestine, Virginia); 10 = 9192 (lamb intestine,Virginia).
C. perfringens type E, lanes 11 to 13: 11 =ATCC 27324; 12 = 1590 (Montana); 13 = 10339(bovine intestine, Virginia).
Formerly, C. sporogenes and proteolyticstrains of C. botulinum types A, B, and F couldbe differentiated with assurance only by toxinneutralization tests in mice (11, 12, 31). Strainsof C. sporogenes and proteolytic strains of C.botulinum types A, B, and F have high DNA-DNA homology (19, 37). Therefore, the similar-ity among the electrophoretic patterns of theproteolytic strains of C. botulinum types A, B,and F was to be expected. The protein patternswould not distinguish among proteolytic C. bot-ulinum toxin types A, B, and F (Fig. 9). We weresurprised that two bands, one at 4 mm and adark band at 62 mm, were repeatedly present incultures of C. sporogenes (Fig. 9, lanes 12 and13, and Fig. 11, lanes 8 through 13) and absent inthe toxic strains. Farshy and Moss (8) reportedthat C. sporogenes and proteolytic strains of C.botulinum also can be differentiated by gas chro-matographic patterns of trimethylsilyl deriva-tives of whole cell hydrolysates. Because type G(Fig. 9, lane 11) is phenotypically different fromother strains depicted in this figure, its different
Fig. 4. C. innocuum, lanes 2 to 13: 2 =ATCC 14501 (type strain, appendiceal abscess,Tennessee); 3 = 3220E (ruptured appendix,Kentucky, 93%*); 4 = 2808A (rectal abscess,Kentucky, 86%*); 5 = 0120 (Prevot 1666A, Paris,84%*); 6 = 2020 (wound, Georgia, 79%*); 7 =3248 (clinical isolate, Maryland, 76%*); 8 = 2499B(urinary tract, Kentucky, 76%*); 9 = 2019 (pleuralfluid, Georgia, 75%*); 10 = 3794C (pelvic abscess,Kentucky, 41%*); 11 = 5792 (crushing injury,Georgia); 12 = 9251 (blood, Ohio); 13 = 11072(rectal pus, London).
*DNA/DNA homology with ATCC 14501[from Johnson and Francis (17)].
protein banding pattern was to be expected.Also, as was to be expected, the patterns oftoxic saccharolytic strains of C. botulinum typesB, E, and F were different from those of theproteolytic strains. These strains are geneticallyrelated (16, 19) as well as phenotypically similar(11, 32) and could not reliably be identified totype by their gel patterns. There was variation inthe patterns of these strains, although they had ageneral similarity to each other (Fig. 10). Asharp band at 90 mm was distinctive for the typeE strains tested (Fig. 10, lanes 4 to 6), but thetype B and type F strains could not be differenti-ated. There was less growth and less proteinfrom ATCC 27321 (Fig. 10, lane 9), so the bandswere not as dark. Therefore, the film was ex-posed twice with lane 9 blacked out and then athird time after the black strip was removed.
C. sporogenes and C. difficile are morphologi-cally similar and have similar fermentation prod-ucts. They differ in mannitol fermentation, pro-teolytic activity in milk and meat, and lipaseproduction (12). The protein patterns (Fig. 12)
VOL. 15, 1982
9-.- 1- v - olm-1 - - i..q- - --
W.,1:1--
aft-W
1: .... a W.X
0 U"
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
cm o
n 10
Dec
embe
r 20
21 b
y 10
3.17
1.13
.219
.

692 CATO ET AL.
Fig. 5. C. ramosum, lanes 2 to 13: 2 = ATCC25582 (type strain); 3 = 9206 (abdominal wound,Colorado); 4 = 9361 (urinary catheter, Georgia);5 = 9448 (toe wound, Georgia); 6 = 9500 (periton-eal cavity, acute gangrenous appendix, Maryland);7 = 9829 (rectal abscess, London); 8 = 9835(abdominal cavity, London); 9 = 10053 (skingraft, Pennsylvania); 10 = 11456 (peritoneal fluid,Maryland); 11 = 11658 (blood, Delaware); 12 =
12560 (suture wound, Massachusetts); 13 = 12667(colon tissue, N. Dakota).
were quite different and required much lessanalytical time than did conventional tests.
C. bifermentans and C. sordellii traditionallyhave been separated on the basis of ureaseproduction by C. sordellii and, to a lesser extent,on the ability of C. bifermentans to fermentmannose, salicin, and sorbitol in a basal mediumlow in peptone and yeast extract (31). Somestrains of C. sordellii are toxic for mice, whereasno strain of C. bifermentans has been reportedto be toxic (31). Strains of the two species are 50to 70% related by DNA homology studies (24).On the basis of reactions of strains identified byDNA homology, Nakamura et al. (24, 25) report-ed that fermentation of mannose, deamination ofarginine, and the presence of mannose in the cellwalls of C. bifermentans are more reliable thanurease production for differentiating betweenthese two species. The protein patterns of thetwo species are distinct (Fig. 13).
Protein patterns of two important animalpathogens, C. chauvoei and C. septicum, areshown in Fig. 14. C. septicum is pathogenic forhumans as well. Although there were similaritiesin the patterns of these two species, the differ-
Fig. 6. C. septicum, lanes 2 to 13: 2 ATCC12464 (type strain); 3 = 8007 (blood, Indiana);4 = 8356 (blood, Georgia); 5 = 8369 (blood,Pennsylvania); 6 = 8963 (ankle puncture wound,Kansas); 7 = 9069 (gas gangrene, Michigan); 8 =
10119 (liver abscess, Georgia); 9 = 10263 (lung,Minnesota); 10 = 10314 (blood, Minnesota); 11 =
10351 (pus, Quebec); 12 = 10927 (peritonealcavity, Utah); 13 =11947 (blood, Texas).
Fig. 7. C. baemolyticum, lanes 2 to 13: 2=ATCC 9650 (type strain, bovine); 3 = CDC KA25;4 1942A*; 5 = 2166*; 6 = 2167*; 7 = 2168*;8= 2169*; 9 - 2172*; 10 = 2173*; 11 = 2196(alligator, Florida); 12 = 8065*; 13 = 8066*.
* Isolated from cattle in Montana.
J. CLIN. MICROBIOL.
... ...
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
cm o
n 10
Dec
embe
r 20
21 b
y 10
3.17
1.13
.219
.

ELECTROPHORESIS OF CLOSTRIDIUM SPECIES 693
Fig. 8. C. novyi A, lanes 2 and -3: 2= ATCC
17861 (type strain); 3 = ATCC 3540.
C. novyi B, lanes 4 and 5: 4 = ATCC 25758
(sheep); 5 = ATCC 27606.
C. baemolyticum, lanes 6 and 7: 6 = ATCC
9650 (bovine); 7 = 2196 (alligator, Florida).C. botulinum type C, lanes 8 and 9: 8 = ATCC
25766, 9 = NCIB 4218 (duck).
C. botulinum type D, lanes 10 and 11: 10
ATCC 27517, 11 = 6793.
C. botulinum (non-toxic), lanes 12 and 13:
12 = 9026 (dog blood, Seattle), 13 = 9257 (bovine
muscle, Virginia).
ences in the upper third of each of the patterns
clearly separated them.
The patterns of strains of C. tetani, whose
identification was confirmed by mouse toxin
neutralization tests, were more heterogeneous
than those of C. cadaveris (Fig. 15), but the
patterns of the two species were distinct. The
two species differ biochemically in that C. cada-
veris ferments glucose and C. tetani does not.
The electrophoretic pattern of each strain of C.
tetani was constant, and we have successfully
used this property in an epidemiological problem
to establish that two C. tetani infections that
developed in two patients who were treated in
the same emergency room on the same day
resulted from two different strains.
C. hastiforme and C. subterminale share
many phenotypic properties. They both are non-
fermentative and moderately proteolytic. We
previously had confused the two species, and
the description of C. hastiforme given in the 4th
edition of the VPI Anaerobe Laboratory Manual
Fig. 9. C. botulinum type A (proteolytic),lanes 2 to 4: 2 = ATCC 25763 (type strain); 3=7124 (soil, Virginia); 4 = ATCC 17862.
C. botulinum type B (proteolytic), lanes 5 to7: 5 = ATCC 7949; 6 = 2131 (Pasteur Institute,Paris); 7 = 10758 (oats).
C. botulinum type F (proteolytic), lanes 8 to10: 8 = ATCC 25764 (crab); 9 = (CDC KA1, liverpate); 10 = 4257 (soil Argentina).
C. botulinum type G, lane 11: ATCC 27322(soil, Argentina).
C. sporogenes, lanes 12 and 13: 12 = ATCC3584 (type strain, soil); 13 = 12643 (dugong,Australia).
(12) is in error. Strains of C. hastiforme produceterminal spores and do not produce hydrogen;strains of C. subterminale have subterminalspores and produce copious amounts of hydro-gen. C. hastiforme can be distinguished from C.subterminale by the heavy protein bands formedat 72 and 96 mm by C. hastiforme (Fig. 16).Strains of both species have been isolated fromblood cultures, abdominal abscesses, and vari-ous soft tissue infections.
In 1979 Kaneuchi et al. (17) demonstrated byDNA-DNA homology that C. cocleatum and C.spiroforme, both similar biochemically to C.ramosum, were in fact only distantly related.They are 32 to 53% homologous with the typestrain of C. ramosum and 40 to 60% homologouswith each other. Cells of C. ramosum arestraight, whereas those of C. cocleatum and C.spiroforme are coiled, although often somestraight cells are present. C. ramosum fermentsgalactose and a-methyl glucoside, but not inulin,C. cocleatum ferments galactose and inulin, and
VOL. 15, 1982
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
cm o
n 10
Dec
embe
r 20
21 b
y 10
3.17
1.13
.219
.

694 CATO ET AL.
Fig. 10. Saccharolytic strains of C. botulinum.C. botulinum type B, lanes 2 and 3: 2 = ATCC25765; 3 = Prevot 59. C. botulinum type E, lanes4 to 6: 4 = ATCC 9564; 5 = Hobbs 4249; 6 =9817 (soil, Olympic rain forest). C. botulinumtype F, lanes 7 to 10: 7 = Eklund 83F; 8 = ATCC23387 (marine sediment); 9 = ATCC 27321(marine sediment); 10 = 7943 (California).
C. spiroforme ferments only inulin. The proteinpatterns bear out the similarity of these speciesbut point up differences as well (Fig. 17); bandsat 31, 40, and 63 mm in C. ramoslum, one at 34mm in C. spiroforme, and one at 15 mm in C.cocleatum correlated with the biochemical andgenetic differentiations. C. ramosum often ispresent in human clinical specimens and feces.A few strains studied by Kaneuchi et al. (17)were isolated from ceca of rodents. All strains ofC. cocleatum were from ceca of rodents orchickens, and all strains of C. spiroforme werefrom human feces (17).
C. cochlearium, C. lentoputrescens, and C.malenominatum are similar phenotypically, dif-ferentiated principally by the ability of C. lento-putrescens to digest gelatin and casein, the pro-duction of indole by C. lentoputrescens and C.malenominatum, and the reduction of nitrate byC. malenominatum (33). We have not found thatany of these species that reduce nitrate or digestcasein. Gelatin hydrolysis was variable in strainsof these species when tested by the conventionaltube method (12) and by the method of Smith etal. as described by Skerman (29). Gelatin wasusually weakly digested by strains of C. cochle-
Fig. 11. C. novyi type B, lanes 2 to 7: 2 =ATCC 25758 (type strain, sheep); 3 = 8096; 4 =8097 (non-toxic); 5 = 8098; 6 = ATCC 27606; 7 =12207.
C. sporogenes, lanes 8 to 13: 8 = ATCC 3584(type strain, soil); 9 = 11116 (blood, Massachu-setts); 10 = 11125 (blood, Rhode Island); 11 =11248A (bovine muscle, Nebraska); 12 = 11272(post-amputation wound infection, New Zealand);13 = 11440D (snake venom, Costa Rica).
arium and C. lentoputrescens, but not by strainsof C. malenominatum. Further, all strains arenon-saccharoclastic, have terminal spores thatswell the rod, are similar morphologically, andmetabolize peptones to similar volatile acids.They are isolated occasionally from varioustypes of clinical specimens, including blood.Because our strains of C. lentoputrescens wereso similar to those of C. malenominatum andmore nearly fit the description of C. malenomi-natum than that of C. lentoputrescens, we con-sidered them to be C. malenominatum in the 4thedition of the VPI Anaerobe Laboratory Manual(12). Because the names of both species ap-peared in the 1980 Approved Lists of BacterialNames (30) with designated type strains, bothwere included in this study. The protein patterns(Fig. 18) suggest that C. malenominatum isdistinct and that C. lentoputrescens and C. co-chlearium may be the same species. If this isconfirmed by DNA homology studies, it wouldmean that indole production would be variableamong strains of C. cochlearium (C. lento-putrescens). Although indole production usuallyis a stable phenotypic characteristic within aspecies, this is not always the case. DNA homol-
J. CLIN. MICROBIOL.
T-1
oft.,
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
cm o
n 10
Dec
embe
r 20
21 b
y 10
3.17
1.13
.219
.

ELECTROPHORESIS OF CLOSTRIDIUM SPECIES 695
., * - X fit-t i. .t A _ t_t _ . ; ....... e Ft .. ^. . S .W._ v.ii.:Wfi * pbSS:..:br
_
.:..
...
, ils
Fig. 12. C. sporogenes, lanes 2 to 7: 2 =ATCC 3584 (type strain, soil); 3 = 11069 (buttockboil, London); 4 = 11125 (blood, Rhode Island);5 = 12227 (foot drainage, Pennsylvania); 6 =
12434 (blood, Florida); 7 = 12643 (dugong,Australia).
C. difficile, lanes 8 to 13: 8 = ATCC 9689(type strain); 9 = 2003 (blood, Florida); 10 =8484 (abdominal abscess, Georgia); 11 = 10463(pelvic abscess, California); 12 = 12912 (feces,Maryland); 13 = B844-3 (pseudomembranouscolitis, Virginia).
ogy results have shown that indole production isa variable characteristic among strains of Fuso-bacterium varium (23; J. L. Johnson, personalcommunication).
Duplicate patterns of the soluble proteins ofsome less commonly mentioned butyric acid-producing species are shown in Fig. 19 and 20.ATCC strain 25779, labeled "C. pseudotetani-cum," was proposed as a reference strain for thespecies (33). However, its morphological andbiochemical characteristics appeared similar tothose of C. butyricum. Johnson and Francis (16)reported that bulk RNA preparations of ATCC25779 were 100% homologous with the 23S RNAfrom ATCC 19398, the type strain of C. butyri-cum. Although RNA homology does not neces-sarily indicate identity at the species level, theprotein patterns of these two strains were essen-tially identical (Fig. 20, lanes 8, 9, 12, and 13).On the basis of correlations of DNA homology(which does indicate identity at the specieslevel), these patterns indicate that the two spe-cies are synonymous.We received a strain labeled "Plectridium
Fig. 13. C. bifermentans, lanes 2 to 7: 2 =ATCC 638 (type strain, vaccine contaminant); 3 =9809 (ascitic fluid, Georgia); 4 = 9894 (vagina,Quebec); 5 = 11442 (snake venom, Costa Rica);6 = 11860 (ulcer, buttock, Massachusetts); 7 =12025 (abdominal wound, Massachusetts).
C. sordeliii, lanes 8 to 13: 8 = ATCC 9714(type strain); 9 = 4540 (alpaca, Lima, Peru); 10 =5246A (foot drainage, Virginia); 11 = 6516(ruptured appendix, Michigan); 12 = 6899 (analabscess, Ohio); 13 = 7584 (uterine abscess, Penn-sylvania).
fluxum" (VPI 2734, Prevot TB III) from A. R.Prevot, who had first described this species (26,27). Morphologically and biochemically, thisstrain is similar to C. innocuum. Protein patternsof VPI 2734 were identical to those of the typestrain of C. innocuum, ATCC 14501 (Fig. 20,lanes 2 and 3, 6 and 7), indicating that these twospecies are synonymous.
Species with widely divergent properties andproducts gave protein bands that were equallydistinctive (Fig. 21 and 22). The relativelysaccharoclastic organisms C. nexile (major prod-ucts, acetic and formic acids and ethanol) andSporolactobacillus inulinus (major product, lac-tic acid) could be readily differentiated fromeach other as well as from the non-saccharoclas-tic, nonreactive organisms C. putrefaciens (pro-ducing only small amounts of acetic acid) and C.propionicum (producing large amounts of propi-onic and isovaleric acids and small amounts ofacetic and isobutyric acids).
Figures 22a and 22b illustrate an analyticalproblem we encountered occasionally whenstrains with greatly different growth characteris-
VOL. 15, 1982
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
cm o
n 10
Dec
embe
r 20
21 b
y 10
3.17
1.13
.219
.

696 CATO ET AL.
OR t!r --,- i T7nX
Af~~~~''} ..................... ......£4tow::Xl
Fig. 14. C. chauvoei, lanes 2 to 7: 2 = ATCC10092 (type strain, bovine blackleg); 3 = CDC KA-23; 4 = 2454 (internal organs, cattle, California);5 = 9719 (calf muscle, Virginia); 6 = 11685 (sheepblackleg, Ohio); 7 = 11918 (bovine muscle,Virginia).
C. septicum, lanes 8 to 13: 8 = ATCC 12464(type strain); 9 = 9069 (human gas gangrene,Michigan); 10 = 10263 (lung, Minnesota); 11 =
10314 (blood, Minnesota); 12 = 10351 (pus,Quebec); 13 = 10927 (peritoneal cavity, Utah).
tics were placed on the same gel. C. rectumgrows vigorously and quickly reaches maximumturbidity. Broken cell preparations contained somuch material that separate protein bands couldnot be easily distinguished. The problem was notcorrected by standardizing the amount of pro-tein in the samples because often the amount ofprotein was unrelated to the clarity of the bands(James A. Blanks, unpublished data). Ratherthan prepare and reanalyze diluted samples ofsuch cultures (which solves the problem but istime-consuming), we obtained nearly identicalresults by decreasing the camera F-stop from 18(Fig. 22a) to 8 and taking a second photograph(Fig. 22b).The diversity of patterns among type strains
of 34 different species is shown in Fig. 23 to 25.We have tried to place pairs of species that aresometimes difficult to distinguish in the samefigure. C. sphenoides and C. indolis are pheno-typically similar (12), although the percent gua-nine plus cytosine content of the DNA of C.sphenoides ATCC 19403 is 41, whereas that ofC. indolis ATCC 25771 is 44 (16). The similarityof the polyacrylamide gel electrophoretic pat-
Fig. 15. C. cadaveris, lanes 2 to 7: 2 = ATCC25783 (type strain, rabbit cyst); 3 = ATCC 9687;4 = 2038 (Ohio); 5 = 8451 (peritoneal fluid,Wisconsin); 6 = 8531 (abdominal wound, Georgia);7 = 10655 (bovine intestine, Virginia).
C. tetani, lanes 8 to 13: 8 = ATCC 8033; 9 =1608 (normal human feces, Virginia); 10 = 9149(soil, Virginia); 11 = 9768A (knee wound, Ohio);12 = 9776 (soil, Ontario, Canada); 13 = ATCC19406 (type strain).
terns of these two strains (Fig. 23, lanes 4 and 6)confirmed their close relationship, but therewere sharply contrasting bands, particularly be-yond 65 mm from origin, that were more helpfulin distinguishing the two species than any classi-cal phenotypic characteristics we have tested.
"C. colinum," isolated from ulcerative enteri-tis in quail, was described by Berkhoff et al. in1974 (2). The strains originally deposited in theATCC were lost, and the name was not includedin the 1980 Approved Lists of Bacterial Names(30). A replacement of their strain B has nowbeen deposited as ATCC 27769. This strain,which we received from G. A. Berkhoff, had thecharacteristics originally described and pro-duced a distinctive protein pattern (Fig. 24, lane12).Tanner et al. (36) have shown that the type
strains of C. lituseburense and of Eubacteriumtenue are closely related in the oligonucleotidesequence of their 16S ribosomal RNAs as well asin the percentage of guanine plus cytosine oftheir DNAs and the type of murein cross-linkagein their cell walls. Metabolically the two organ-isms are quite distinct (12). C. lituseburense
J. CLIN. MICROBIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
cm o
n 10
Dec
embe
r 20
21 b
y 10
3.17
1.13
.219
.

ELECTROPHORESIS OF CLOSTRIDIUM SPECIES
.a.
Fig. 16. C. bastiforme, lanes 2 to 7: 2 = ATCC33268 (type strain, abdominal abscess, Califomia);3 = 9918B (wound, Virginia); 4 = 10426 (bovinebrain, California); 5 = 11497A (blood, Minnesota);6 = 12486A (leg wound, Texas); 7 = 12627(blood, Texas).
C. subterminale, lanes 8 to 13: 8 = ATCC25774 (type strain, Texas); 9 = 8675A (wound,Maryland); 10 = 8677 (foot wound, Maryland);11 = 8716 (perineum wound, Georgia); 12 = 2491(Lille, France); 13 = 2474 (Lille, France).
ATCC 25759 produces spores and large amountsof butyric acid neither indole nor hydrogen isformed, and fructose, mannose, and sucrose arefermented. E. tenue ATCC 25553 does not pro-duce spores or major amounts of butyric acid,indole and abundant hydrogen are produced,and fructose, mannose, and sucrose are notfermented. Protein patterns of the two strainswere distinct (Fig. 26).The patterns of two purine-degrading clostrid-
ia, C. purinolyticum (6) and C. acidiurici (1), areshown in Fig. 26. Although these two species aredistinct by DNA-DNA homology determina-tions (6), they are very similar phenotypically.They can be differentiated by the ability of C.purinolyticum, but not C. acidiurici, to growreadily in a defined medium, with adenine as thesole source of nitrogen.Biochemical reactions of the type strain of C.
villosum, NCTC 11220, which sporulates poorlyand only after long incubation (20), are similar tothose of Eubacterium combesii, except that allstrains of E. combesii produce large amounts ofhydrogen (12), whereas the strain of C. villosum
I
Fig. 17. C. ramosum, lanes 2 to 5: 2 = ATCC25582 (type strain); 3 = 10053 (skin graft, osteo-myelitis, Pennsylvania); 4 = 12560 (suture wound,Massachusetts); 5 = 12667 (colon tissue,N. Dakota).
C. spiroforme, lanes 6 to 9: 6 = ATCC 29900(type strain, human feces, Hawaii); 7 = UPC 74-1(human feces, Virginia); 8 = C43-37 (human feces,Hawaii); 9 = 8872 (human feces, Texas).
C. cocleatum, lanes 10 and 11: 10 = ATCC29902 (type strain, mouse cecum, Japan); 11 =8723 (mouse cecum, Netherlands).
Fig. 18. Type strains of Clostridium species,lanes 2 to 11: 2,3 = C. symbiosum ATCC 14940;4,5 = C. malenominatum ATCC 25776; 6,7 =
C. fallax ATCC 19400; 8,9 = C. lentoputrescensATCC 17794; 10,11 = C. cocblearium ATCC17787.
VOL . 1 5, 1982 697
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
cm o
n 10
Dec
embe
r 20
21 b
y 10
3.17
1.13
.219
.

698 CATO ET AL.
Fig. 19. Type and reference strains ofClostridium species, lanes 2 to 13: 2,3 = "C.pseudotetanicum" ATCC 25779 (reference strain,septicemia, France); 4,5 = C. sartagoformumATCC 25778 (type strain, vaccine contaminant);6,7 = C. paraputrificum ATCC 25780 (type strain,Ohio); 8,9 = C. felsineum ATCC 17788 (typestrain); 10,11 = C. barkeri ATCC 25849 (typestrain, Potomac River mud); 12,1 3 = "C. fluxum "2734 (reference strain, infant feces, France).
produces only a small amount. Their proteinpatterns, however, were markedly different(Fig. 26), indicating that they probably havelittle or no DNA-DNA homology.We have received many strains of clostridia
with names that have not been accepted as validby the ad hoc Committee of the Judicial Com-mission of the International Committee on Sys-tematic Bacteriology and that have not beenincluded in the Approved Lists of BacterialNames (30). Among these were a strain labeled"Plectridium incertum" and two strains labeled"Clostridium imitans." Broken cell prepara-tions of these organisms were first placed on agel (Fig. 27) with the type strains of threeorganisms, C. ghoni, C. putrificum, and C.scatologenes, having similar fermentation prod-ucts. Major products of each are acetic andbutyric acids and varying amounts of propionic,isobutyric, isovaleric, and isocaproic acids (12).The type strain of an unrelated organism, C.limosum (major products acetic and formic ac-ids), was included for contrast. Differences intheir protein patterns were unmistakable.
Biochemically, "P. incertum" resembled C.
Fig. 20. Type and reference strains of Clos-tridium species, lanes 2 to 13: 2,3 = C. innocuumATCC 14501 (type strain, appendiceal abscess,Tennessee); 4,5 = C. innocuum, 2020 (wound,Georgia); 6,7 = "C. fluxum" 2734 (infant feces,France); 8,9 = C. butyricum ATCC 19398 (typestrain, pig intestine, Scotland); 10,11 = C.butyricum ATCC 860; 12,13 = "C. pseudo-tetanicum" ATCC 25779 (septicemia, France).
Fig. 21. Type strains of Clostridium andSporolactobacillus species, lanes 2 to 9: 2,3 = C.nexile ATCC 27757 (human feces, colon polyppatient, Hawaii); 4,5 = Sporolactobacillus inulinusATCC 15538 (chicken feed, Japan); 6,7 = C.putrefaciens ATCC 25786 (spoiled ham, England);8,9 = C. propionicum ATCC 25522 (San FranciscoBay mud).
J. CLIN. MICROBIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
cm o
n 10
Dec
embe
r 20
21 b
y 10
3.17
1.13
.219
.

ELECTROPHORESIS OF CLOSTRIDIUM SPECIES 699
Fig. 22a. Type and reference strains ofClostridium species, lanes 2 to 13: 2,3 = C. rectumATCC 25751; 4,5 = C. quercicolum ATCC 25974(oak tree); 6,7 = C. aurantibutyricum ATCC17777; 8,9 = C. formicoaceticum ATCC 23439(reference strain, pond mud, West Germany);10,11 = C. putrificum ATCC 25784; 12,13 = C.tetani ATCC 19406.
hastiforme, and "C. imitans" resembled C. sub- As reportterminale. Therefore, the protein patterns of organisms (4these four strains were compared (Fig. 28). The of homogeipattern of "P. incertum" was identical with that among straiof C. hastiforme, indicating synonymy, but C. bore a stronjsubterminale and "C. imitans" were clearly DNA homounrelated. We do not wish to revive the name measuremer"C. imitans" now because we have only two with greateistrains, too few to determine the variation that produced idmay occur among strains. ity (with ovThe recently described organism C. puniceum was often c
(21), isolated from rotting potatoes, was pheno- 70% DNA-Itypically similar to several strains labeled "C. were apparepectinovorum" that we received from A. R. by DNA-D!~Prevot, L. S. McClung, and G. Hobbs. "C. lar phenotyppectinovorum" was most recently described by ic or sacchaHellinger (10) and in the 7th edition of Bergey's similar thanManual ofDeterminative Bacteriology (34), but notypic prolthe name was omitted from the 1980 Approved We have fLists of Bacterial Names (30). Although the unknown stIprotein patterns of these strains (Fig. 28) were exceedinglydistinct, they had general similarity, which notypicallymight represent normal variation in the species. the patternsUntil DNA-DNA homology studies are made, pure culturerevival of the name "C. pectinovorum" for a we can strdistinct species cannot be justified. species.
b
Fig. 22b. Du-plicate of part ofFig. 22a, taken at alower F-stop. Lanes2,3 = C. rectumATCC 25751; 4,5 =
C. querciolum ATCC25974.
ted previously for other groups of4; Biavat et al., in press), the degreeneity of simple protein patternsins varied for different species andg relationship to the degree of DNA-)logy in the instances where bothnts were available. Usually, strainsr than 80% DNA-DNA homologylentical patterns. Minor heterogene-terall similarity of the total pattern)Dbserved among strains with aboutDNA homology. Major differences:nt among species that are unrelated4A homology, but species with simi-pic properties (products or proteolyt-roclastic activities) often were morethose with distinctly different phe-
perties.found that comparing gel patterns ofrains with those of the type strains ishelpful in distinguishing among phe-similar organisms. In many cases,
i obtained within 24 h of isolating ae are sufficiently distinctive so thatongly suspect the identity of the
VOL. 15, 1982
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
cm o
n 10
Dec
embe
r 20
21 b
y 10
3.17
1.13
.219
.

Fig. 23. Type strains of Clostridium species,lanes 2 to 12: 2 = C. tertium ATCC 14573; 3 =
C. durum ATCC 27763 (Black Sea sediment); 4 =
C. sphenoides ATCC 19403; 5 = C. clostridiiformeATCC 25537 (calf rumen); 6 = C. indolis ATCC25771; 7 = C. celatum ATCC 27791 (human feces,Ottawa); 8 = C. oroticum ATCC 13619 (SanFrancisco Bay mud); 9 = C. cellobioparum ATCC15832 (rumen); 10 = C. leptum ATCC 29065(human feces, Texas); 11 = C. aminovalericumATCC 13725 (mud, N. Carolina); 12 = C. histo-lyticum ATCC 19401 (gas gangrene).
Fig. 25. Type and reference strains of Clos-tridium species, lanes 2 to 13: 2 = C. cadaverisATCC 25783 (rabbit cyst); 3 = C. sardiniensisATCC 33455; 4 = C. tetani ATCC 8033 (referencestrain); 5 = C. chauvoei ATCC 10092 (bovineblackleg); 6 = C. septicum ATCC 12464; 7 = C.novyi A ATCC 17861; 8 = C. novyi B ATCC25758 (reference strain); 9 = C. carnis ATCC25777; 10 = C. pasteurianum ATCC 6013; 11 = C.tyrobutyricum ATCC 25755; 12 = C. propionicumATCC 25522 (soil, California).
-4~~~ ~~~~~~~-
Fig. 24. Type strains of Clostridium species,
lanes 2 to 13: 2 = C. puniceum NCIB 11596
(rotting potato, England); 3 = C. absonum ATCC
27555 (soil, Japan); 4 = C. barati ATCC 27638
(canine renal epithelium, France); 5 = C. sporo-
sphaero ides ATCC 25781; 6 = C. sticklandii ATCC
12662 (San Francisco Bay mud); 7 = C. glyco-licum ATCC 14880 (stagnant pond mud); 8 = C.
irregularis ATCC 25756; 9 = C. lituseburense
ATCC 25759 (leaf mold, Africa); 10 = C. mange-
notii ATCC 25761 (African soifl); 1 1 = C. coccoid-
es ATCC 29236 (mouse cecum, Japan); 12 = "C.
colinum" ATCC 27769 (ulcerative enteritis, quail);13 = C. oceanicum ATCC 25647 (marine sedi-
ment).
Fig. 26. Type strains of Clostridium and
Eubacterium species, lanes 2 to 13: 2,3 = C. litus-
eburense ATCC 25759 (soil, Africa); 4,5 = Eu-
bacterium tenue ATCC 25553 (uterine abscess);
6,7 = C. purinolyticum DSM 1384 (soil,
Germany); 8.9 = C. acidiurici ATCC 7906 (soil,
California); 10, 11 = C. villosum NCTC 11220 (catabscess, Australia); 12,13 = E.combesAi ATCC
25545 (soil, Africa).
700
IR
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
cm o
n 10
Dec
embe
r 20
21 b
y 10
3.17
1.13
.219
.

ELECTROPHORESIS OF CLOSTRIDIUM SPECIES 701
Fig. 27. Type and reference strains of Clos-tridium species, lanes 2 to 13: 2,3 = C. gboniATCC 25757 (mud, France); 4,5 = C. putrificumATCC 25784; 6,7 = C. scatologenes ATCC 25775;8,9 = "C. incertum" 2772 (reference strain, brainabscess, France); 10,11 = "C. imitans" 4460(reference strain); 12,13 = C. limosum ATCC25620 (soil, Africa).
ACKNOWLEDGMENTS
We thank Pauletta C. Atkins, Ruth Z. Beyer, Ann P.Donnelly, Luba S. Fabrycky, Jane L. Hungate, Ann C.Mitchell, Debra B. Sinsabaugh, and Dianne M. Wall fortechnical assistance with the culture work.We gratefully acknowledge the financial support of Public
Health Service grant AI-15244 from the National Institute ofAllergy and Infectious Diseases and of Virginia PolytechnicInstitute and State University project 2022820 from the Com-monwealth of Virginia.
LITERATURE CITED
1. Barker, H. A., and J. V. Beck. 1942. Clostridium acidi-urici and Clostridium cylindrosporum, organisms ferment-ing uric acid and some other purines. J. Bacteriol. 43:291-304.
2. Berkhoff, G. A., S. G. Campbell, H. B. Naylor, andL. DS. Smith. 1974. Etiology and pathogenesis of ulcer-ative enteritis ("quail disease"). Characterization of thecausative anaerobe. Avian Dis. 18:195-204.
3. Calhoon, D. A., W. R. Mayberry, and J. Slots. 1981.Cellular fatty acid and soluble protein composition ofActinobacillus actinomycetemcomitans and related or-ganisms. J. Clin. Microbiol. 14:376-382.
4. Cato, E. P., L. V. Holdeman, and W. E. C. Moore. 1982.Clostridium perenne and Clostridium paraperfringens:later subjective synonyms of Clostridium barati. Int. J.Syst. Bacteriol. 32:77-81.
5. Cummins, C. S., and J. L. Johnson. 1971. Taxonomy ofthe clostridia: wall composition and DNA homologies inClostridium butyricum and other butyric acid-producingclostridia. J. Gen. Microbiol. 67:33-46.
6. Durre, P., W. Andersch, and J. R. Andreesen. 1981.Isolation and characterization of an adenine-utilizing,anaerobic sporeformer, Clostridium purinolyticum sp.nov. Int. J. Syst. Bacteriol. 31:184-194.
Fig. 28. Type and reference strains of Clos-tridium species, lanes 2 to 13: 2,3 = C. bastiformeATCC 33268 (abdominal abscess, California);4,5 = "C. incertum" 2772 (brain abscess, France);6,7 = C. puniceum NCIB 11596 (rotting potato,England); 8,9 = "C. pectinovorum" 2430 (rettedflax, Israel); 10,11 = C. subterminale ATCC 25774(Texas); 12,13 = "C. imitans" 4460.
7. Eklund, M. W., T. Poysky, J. A. Meyers, and F. A. Pel-roy. 1974. Interspecies conversion of Clostridium botuli-num type C to Clostridium novyi type A by bacteriophage.Science 186:456-458.
8. Farshy, D. C., and C. W. Moss. 1970. Characterization ofclostridia by gas chromatography. Differentiation of spe-cies by trimethylsilyl derivatives of whole cell hydroly-sates. Appl. Microbiol. 20:78-84.
9. Fox, R. H., and D. E. McClain. 1974. Evaluation of thetaxonomic relationship of Micrococcus cryophilus, Bran-hamella catarrhalis, and Neisseriae by comparative poly-acrylamide gel electrophoresis of soluble proteins. Int. J.Syst. Bacteriol. 24:172-176.
10. Hellinger, E. 1952. A comparative study of certain clos-tridia from flax-retting liquors in Israel. Bull. Res. Counc.tsr. 2:225-238.
11. Holdeman, L. V., and J. B. Brooks. 1970. Variationamong strains of Clostridium botulinum and related clos-tridia, p. 278-286. In M. Herzberg (ed.), Toxic micro-organisms. U.S. Department of the Interior, Washington,D.C.
12. Holdeman, L. V., E. P. Cato, and W. E. C. Moore (ed.).1977. Anaerobe laboratory manual, 4th ed. Virginia Poly-technic Institute and State University, Blacksburg, Va.
13. Holdeman, L. V., W. E. C. Moore, P. J. Churn, and J. L.Johnson. 1982. Descriptions of Bacteroides oris sp. nov.and Bacteroides buccae sp. nov. isolated from humanperiodontitis and other human infections. Int. J. Syst.Bacteriol. 32:125-131.
14. Howard, C. J., D. H. Pocock, and R. N. Gourlay. 1981.Polyacrylamide gel electrophoretic comparison of thepolypeptides and ureaplasmas isolated from cattle andhumans. Int. J. Syst. Bacteriol. 31:128-130.
15. Jarvis, A. W., and J. M. Wolff. 1979. Grouping of lacticstreptococci by gel electrophoresis of soluble cell ex-tracts. Appl. Environ. Microbiol. 37:391-398.
16. Johnson, J. L., and B. S. Francis. 1975. Taxonomy of the
VOL. 15, 1982
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
cm o
n 10
Dec
embe
r 20
21 b
y 10
3.17
1.13
.219
.

702 CATO ET AL.
clostridia: ribosomal ribonucleic acid homologies amongthe species. J. Gen. Microbiol. 88:229-244.
17. Kaneuchi, C., T. Miyazato, T. Shinjo, and T. Mitsuoka.1979. Taxonomic study of helically coiled, sporeforminganaerobes isolated from the intestines of humans andother animals: Clostridium cocleatum sp. nov. and Clos-tridium spiroforme sp. nov. Int. J. Syst. Bacteriol. 29:1-12.
18. Kersters, K., and J. De Ley. 1975. Identification andgrouping of bacteria by numerical analysis of their electro-phoretic protein patterns. J. Gen. Microbiol. 87:333-342.
19. Lee, W. H., and H. Riemann. 1970. Correlation of toxicand non-toxic strains of Clostridium botulinum by DNAcomposition and homology. J. Gen. Microbiol. 60:117-123.
20. Love, D. N., R. F. Jones, and M. Bailey. 1979. Clostridiumvillosum sp. nov. from subcutaneous abscesses in cats.Int. J. Syst. Bacteriol. 29:241-244.
21. Lund, B. M., T. F. Brocklehurst, and G. M. Wyatt. 1981.Characterization of strains of Clostridium puniceum sp.nov., a pink-pigmented pectolytic bacterium. J. Gen.Microbiol. 122:17-26.
22. Moore, W. E. C., D. E. Hash, L. V. Holdeman, and E. P.Cato. 1980. Polyacrylamide slab gel electrophoresis ofsoluble proteins for studies of bacterial floras. AppI.Environ. Microbiol. 39:900-907.
23. Moore, W. E. C., and L. V. Holdeman. 1974. Genus II.Fusobacterium Knorr 1922, p. 407-409. In R. E. Buchan-an and N. E. Gibbons (ed.), Bergey's manual of determi-native bacteriology, 8th ed. The Williams & Wilkins Co..Baltimore.
24. Nakamura, S., T. Shimamura, H. Hayashi, and S. Nishida.1975. Reinvestigation of the taxonomy of Clostridiumbifermentans and Clostridium sordellii. J. Med. Micro-biol. 8:299-309.
25. Nakamura, S., T. Shimamura, and S. Nishida. 1976.Urease-negative strains of Clostridium sordellii. Can. J.Microbiol. 22:673-676.
26. Prevot, A. R. 1938. Etudes de systematique bacterienne.
IV. Critique de la conception actuelle du genre Clostridi-um. Ann. Inst. Pasteur Paris 61:72-91.
27. Prevot, A. R. 1940. Manuel de classification et de determi-nation des bacteries anaerobies. Masson & Co., Paris.
28. Seiter, J. A., and J. M. Jay. 1980. Application of poly-acrylamide gel electrophoresis to the characterization andidentification of Arthrobacter species. Int. J. Syst. Bacte-riol. 30:460-465.
29. Skerman, V. B. D. 1969. Abstracts of microbiologicalmethods, p. 369. John Wiley & Sons, New York.
30. Skerman, V. B. D., V. McGowan, and P. H. A. Sneath(ed.). 1980. Approved lists of bacterial names. Int. J. Syst.Bacteriol. 30:225-420.
31. Smith, L. DS. 1975. The pathogenic anaerobic bacteria,2nd ed. Charles C Thomas, Springfield, Ill.
32. Smith, L. DS. 1977. Botulism, the organism, its toxins, thedisease. Charles C Thomas, Springfield, Ill.
33. Smith, L. DS., and G. Hobbs. 1974. Genus III. ClostridiumPrazmowski, 1880, p. 567. In R. E. Buchanan and N. E.Gibbons (ed.), Bergey's manual of determinative bacteri-ology, 8th ed. The Williams & Wilkins Co., Baltimore.
34. Spray, R. S., L. S. McClung, and E. McCoy. 1957. GenusII. Clostridium Prazmowski, 1880, p. 692-693. In R. S.Breed, E. G. D. Murray, and N. R. Smith (ed.), Bergey'smanual of determinative bacteriology, 7th ed. The Wil-liams & Wilkins Co., Baltimore.
35. Strom, A., J. K. Dyer, C. Marsh, and J. K. Tribble. 1976.Identification and characterization of species of the familyBacteroidaceae by polyacrylamide gel electrophoresis. J.Dent. Res. 55:252-256.
36. Tanner, R. S., E. Stackebrandt, G. E. Fox, and C. R.Woese. 1981. A phylogenetic analysis of Acetobacteriumwoodii, Clostridium barkeri, Clostridium butyricum, Clos-tridium lituseburense, Eubacterium limosum, and Eubac-terium tenue. Curr. Microbiol. 5:35-38.
37. Wu, J. I., H. Reimann, and W. J. Lee. 1972. Thermalstability of the deoxyribonucleic acid hybrids between theproteolytic strains of Clostridium botulinum and Clostridi-um sporogenes. Can. J. Microbiol. 18:97-99.
J. CLIN. MICROBIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
cm o
n 10
Dec
embe
r 20
21 b
y 10
3.17
1.13
.219
.