elodie barbier to cite this version - accueil - tel
TRANSCRIPT

HAL Id tel-01486776httpstelarchives-ouvertesfrtel-01486776
Submitted on 10 Mar 2017
HAL is a multi-disciplinary open accessarchive for the deposit and dissemination of sci-entific research documents whether they are pub-lished or not The documents may come fromteaching and research institutions in France orabroad or from public or private research centers
Lrsquoarchive ouverte pluridisciplinaire HAL estdestineacutee au deacutepocirct et agrave la diffusion de documentsscientifiques de niveau recherche publieacutes ou noneacutemanant des eacutetablissements drsquoenseignement et derecherche franccedilais ou eacutetrangers des laboratoirespublics ou priveacutes
Preacutevalence de Mycobacterium bovis dans lesagroeacutecosystegravemes analyse de reacuteservoirs
environnementaux potentiels (sol eau douce faune dusol et faune aquatique) et traccedilage de la circulation de
cette bacteacuterie entre les diffeacuterents compartimentsElodie Barbier
To cite this versionElodie Barbier Preacutevalence de Mycobacterium bovis dans les agroeacutecosystegravemes analyse de reacuteservoirsenvironnementaux potentiels (sol eau douce faune du sol et faune aquatique) et traccedilage de la circula-tion de cette bacteacuterie entre les diffeacuterents compartiments Biologie animale Universiteacute de Bourgogne2016 Franccedilais NNT 2016DIJOS026 tel-01486776
Universiteacute de Bourgogne Franche-Comteacute - INRA
Uniteacute Mixte de Recherche 1347 Agroeacutecologie
Ecole Doctorale Environnement - Santeacute
THESE
En vue de lrsquoobtention du grade de Docteur de lrsquoUniversiteacute de Bourgogne Discipline Sciences de la Vie Speacutecialiteacute Ecologie microbienne
Preacutevalence de Mycobacterium bovis dans les agroeacutecosystegravemes
analyse de reacuteservoirs environnementaux potentiels (sol eau douce
faune du sol et faune aquatique) et traccedilage de la circulation de
cette bacteacuterie entre les diffeacuterents compartiments
Preacutesenteacutee et soutenue par
Elodie BARBIER
Le 30 mars 2016
Composition du jury
Alain Hartmann Directeur de Recherche INRA Dijon Directeur de thegravese
Maria Laura Boschiroli Directrice de Recherche ANSES Maisons-Alfort Co-Directrice de thegravese
Franccediloise Lucas Professeur Universiteacute Paris Est Creacuteteil (UPEC) Rapporteur
Franck Biet Directeur de Recherche Tours Nouzilly Rapporteur
Sylvie Nazaret Chargeacutee de Recherche CNRS Universiteacute Claude Bernard Lyon 1 Examinateur
Jaquemine Vialard Maitre de confeacuterence Universiteacute Niort ANSES Niort Examinateur
Pascal Simonnet Directeur de Recherche CNRS Ecole Centrale Lyon Examinateur
Jean-Franccedilois Cavin Professeur Universiteacute de Bourgogne AgroSup Dijon Preacutesident du jury
Expert inviteacute
Eric Gueneau Directeur Laboratoire Deacutepartemental de la Cocircte drsquoOr Dijon Co-encadrant
Remerciements
Mes premiers remerciements vont agrave mes encadrants de thegravese Alain Hartmann et Maria-Laura
Boschiroli Merci de mrsquoavoir confieacute ce sujet de thegravese passionnant et de votre accompagnement vos
conseils pertinents et vos encouragements tout au long de ces trois anneacutees
Un grand merci Eric Gueneau co-encadrant de ce travail et directeur du Laboratoire
Deacutepartemental de Cocircte drsquoOr pour son aide et la mise agrave disposition du P3 pour lrsquoincubation de mes
cultures Je remercie eacutegalement chaleureusement toute son eacutequipe de choc en microbiologie et
biologie moleacuteculaire Veacuteronique Julie Virginie et toute lrsquoeacutequipe
Je remercie Franccediloise Lucas et Franck Biet drsquoavoir accepteacute drsquoecirctre les rapporteurs de ce travail
et drsquoen avoir eacutevalueacute la pertinence Merci eacutegalement agrave Sylvie Nazaret Jaquemine Vialard Pascal
Simonet et Jean-Franccedilois Cavin pour leur participation au jury de thegravese
Merci agrave Barbara Dufour Loiumlc Bollache et Jean-Louis Moyen pour leur participation au comiteacute
de pilotage de thegravese et leurs conseils pendant ce travail
Un eacutenorme merci agrave la DDPP21 pour toutes les informations essentielles fournies sur les foyers
bovins et la faune sauvage Aneacutelise Taconnet Fabienne Bartheacuteleacutemy Thierry Grosjean Un merci
particulier agrave Brigitte Biasino pour son aide preacutecieuse lors drsquoune campagne de preacutelegravevements fort
sympathique
Des remerciements chaleureux eacutegalement agrave Fabrice Chevalier et Marina Beacuteral de la CIREV
Bourgogne pour les donneacutees partageacutees mais eacutegalement les magnifiques cartes illustrant mon
manuscrit
Merci aux organismes qui ont contribueacute financiegraverement agrave ce travail en particulier le GDS 21
la Chambre drsquoAgriculture de la Cocircte drsquoOr La MSA Bourgogne et la MSA nationale Interbev et le
Conseil Deacutepartemental de la Cocircte drsquoOr
Un grand merci agrave Violette Sicre (DDPP16 et Sylvatub) pour son accueil chaleureux et son
efficaciteacute lors de mon seacutejour en Charente Merci eacutegalement au directeur du laboratoire de Charente et
agrave son eacutequipe pour leur accueil
Des remerciements au laboratoire de bacteacuteriologie du CHU de Dijon qui a mis son laboratoire
L3 agrave notre disposition pour lrsquoeacutetude sur la survie de M bovis dans le sol Merci agrave Jean-Marie Duez pour
le temps passeacute et sa confiance merci agrave Catherine Neuwirth Julien Bador et toute lrsquoeacutequipe
Un grand merci agrave lrsquoeacutequipe de lrsquoANSES Maisons-Alfort notamment Krystel De Cruz pour le
geacutenotypage des ADN environnementaux mais eacutegalement Claudine Karaoui Lorraine Michelet et le
reste de lrsquoeacutequipe
Merci agrave Franck Biet et Thierry Cochard de lrsquoINRA de Tours qui nous ont fourni les souches de
Map et M smegmatis
Des eacutenormes remerciements pour les lieutenants de louveterie les pieacutegeurs et les chasseurs
qui mrsquoont accompagneacutee dans mes preacutelegravevements sur le terrain et sans qui je nrsquoaurai pas pu preacutelever
tous ces eacutechantillons Jeannick Gautherot Dominique Rigaud Yannick Thibert Georges Balluet Jean-
Luc Moindrot Marcel Guillier qui connaissent les bois comme leur poche et font un travail
consideacuterable
Je remercie les eacuteleveurs qui ont participeacute au travail en nous accompagnant sur leurs pacirctures
Merci agrave Leacuteon Fayolle pour son expertise sur les lombrics et Dominique Meunier pour son
expertise sur les formations peacutedologiques de lrsquoAuxois merci agrave Luc Biju-Duval pour les cartes reacutealiseacutees
Un eacutenorme merci agrave tous les membres du pocircle MERS qui ont fait de ces trois anneacutees un moment
extrecircmement riche en eacutechanges scientifiques humains amicaux et sportifshellipMerci agrave Dominique
Pascal Laurent Jean-Paul Seacutebastien Un merci particulier aux filles de lrsquoeacutequipe Geacuteraldine Ceacutecile
Laetitia pour leur gentillesse leurs compeacutetences et tous les bons moments passeacutes agrave discuterhellipet agrave
marcher Des remerciements tout particuliers agrave Murielle Rochelet pour ses conseils hyper-aviseacutes sa
rigueur son esprit drsquoinnovation son anglais parfait et encore plein plein drsquoautres choses Merci
infiniment pour tout le temps que tu as passeacute agrave me relire et me corrigerhellip
Un grand mais alors tregraves grand merci agrave Benoicirct sans qui lrsquoexpeacuterience lombric nrsquoaurait pas eacuteteacute
aussihellipexceptionnelle Bon courage pour ta thegravese Merci agrave Cleacutement pour les kilos de sols de terriers
ramasseacutes peseacutes analyseacutes lors de ces deux stages Un grand merci agrave Pierre pour sa gentillesse rare son
travail tregraves seacuterieux et son aide inestimable pour la mise en page la relecture et lrsquoimpression de ce
manuscrit
Merci eacutegalement agrave mes collegravegues du Master AE Caroline Alexandre Fam-Ky Justin Marion
et tous les autres qui mrsquoont parfois (souvent) aideacutee agrave supporter de longues heures de cours
Merci agrave tous ceux que jrsquoai croiseacute agrave lrsquoINRA et ailleurs qui ont fait de ces trois ans une expeacuterience
humaine et scientifique inoubliable Ariane Emilie Kadiya Emeline Ceacuteline Heacutelegravene Charline Anne-
Laure Aude Gilles David Marion Nicolas Romain Fabrice Chloeacute Aymeacute Laure Luiz Marie-
Christine Nadine Estelle Amy Amandinehellip et tous ceux que jrsquoai sucircrement oublieacute
Enfin pour finir un grand merci agrave Steacutephane Lola Lucien et Antonin qui ont supporteacute mes hauts
et mes bas pendant ces trois anneacutees agrave ma famille et ma belle-famille qui ont toujours eacuteteacute lagravehellip
Reacutesumeacute
La tuberculose bovine est une maladie infectieuse contagieuse causeacutee par Mycobacterium
bovis Cette maladie touche les bovins et de nombreuses espegraveces de mammifegraveres domestiques et
sauvages ainsi que lrsquohomme La circulation de la bacteacuterie dans des systegravemes multi-hocirctes varieacutes favorise
lrsquoentretien de la maladie et la contamination des bovins vivant agrave proximiteacute des animaux sauvages
infecteacutes En marge de la transmission directe de M bovis par voie respiratoire la transmission indirecte
aux bovins lieacutee agrave lrsquoinhalation ou agrave lrsquoingestion de matrices environnementales contamineacutees par un
animal infecteacute excreacuteteur est suspecteacutee dans plusieurs reacutegions du monde Lrsquoexistence de reacuteservoirs
environnementaux ougrave le bacille M bovis est capable de persister pourrait donc ecirctre un facteur
important de la reacuteeacutemergence puis du maintien de la maladie dans les systegravemes multi-hocirctes
En Cocircte drsquoOr deacutepartement fortement toucheacute par la tuberculose bovine depuis 2004 la
transmission indirecte de la bacteacuterie entre la faune sauvage infecteacutee et les bovins est suspecteacutee dans
plusieurs eacutelevages Pour eacutevaluer la preacutesence et la survie de cette bacteacuterie dans lrsquoenvironnement nous
avons analyseacute un grand nombre drsquoeacutechantillons preacuteleveacutes dans des zones partageacutees par les bovins etou
la faune sauvage infecteacutes dans le but de deacuteterminer la distribution environnementale de M bovis Pour
ce faire nous avons deacuteveloppeacute ou modifieacute des systegravemes de deacutetection moleacuteculaire adapteacutes aux matrices
environnementales complexes Nous avons eacutegalement eacutevalueacute lrsquoimpact de la tempeacuterature et des
proprieacuteteacutes physico-chimiques de deux sols sur la survie de M bovis ainsi que le rocircle de la meacutesofaune
du sol (lombrics en particulier) dans la disseacutemination de la bacteacuterie agrave partir de matiegravere organique
contamineacutee Lrsquoeacutetude environnementale a mis plus particuliegraverement en eacutevidence la contamination de
deux biotopes les zones humides des pacirctures et les sols de terriers de blaireaux De plus les eacutetudes
expeacuterimentales ont montreacute que M bovis pouvait survivre plusieurs mois dans le sol agrave 4degC et que les
lombrics pouvaient disseacuteminer la bacteacuterie dans le sol voire jouer un rocircle potentiel de vecteur pour les
animaux qui les consomment Ces reacutesultats apportent de nouvelles connaissances sur la persistance et
la circulation de M bovis dans lrsquoenvironnement en Cocircte drsquoOr et permettront de proposer des
ameacuteliorations aux mesures de bioseacutecuriteacute deacutejagrave existantes dans les eacutelevages bovins
Mots cleacutes Mycobacterium bovis environnement sol eau fegraveces bovins faune sauvage qPCR culture
Abstract
Bovine tuberculosis is an infectious disease caused by Mycobacterium bovis This disease
affects cattle and many species of domestic and wild mammals and humans The circulation of the
bacteria in various multi-host systems promotes the maintenance of the disease and the contamination
of cattle in the vicinity Beside direct transmission of the bacteria through the respiratory route indirect
transmission through inhalation or ingestion of environmental matrices contaminated by an infected
animal excretory is suspected in several countries Environmental contamination with M bovis appears
to be a crucial factor in the persistence of the infection in multi-host systems
In Cocircte dOr a French department affected by bovine tuberculosis since 2004 the indirect
transmission of the bacteria from infected wildlife to cattle is suspected in several cases To assess this
type of transmission of the bacillus we evaluated the environmental contamination with M bovis on
a large number of samples taken in areas shared by cattle and or wildlife infected For this purpose
we developed or modified molecular detection systems adapted for environmental complex matrices
We also assessed the impact of physicochemical properties of both soil and temperature on survival of
M bovis and the role of earthworms in the spread of the bacteria from contaminated organic material
The environmental study showed the contamination of two media in particular wetlands pastures and
soil badger setts Moreover experimental studies have shown that M bovis can survive in soil for
several months at 4 deg C and the worms could spread the bacteria in the soil or even play a potential
role for vector animals that consume them These results will propose improvements to existing
biosecurity measures on cattle farms and provide new knowledge about the persistence and circulation
of M bovis in the environment in Cocircte dOr
Key words Mycobacterium bovis environment soil water feces cattle wildlife qPCR culture
Table des matiegraveres
INTRODUCTION GENERALE 1
CHAPITRE 1 - CONTEXTE BIBLIOGRAPHIQUE 4
A LES MYCOBACTERIES DIVERSITE PHYLOGENIE SPECTRE DrsquoHOcircTES ET VIRULENCE 4
1 Le genre Mycobacterium taxonomie et meacutethodes de classification 4
2 Caracteacuteristiques geacuteneacuterales des mycobacteacuteries 5
a Morphologie des mycobacteacuteries 5
b Caracteacuteristiques culturales pheacutenotypiques et physiologiques 7
c Phylogeacutenie du genre Mycobacterium 8
d Caracteacuteristiques geacuteneacutetiques des mycobacteacuteries 9
3 Description des diffeacuterents Complexes ou espegraveces de mycobacteacuteries 10
a Les mycobacteacuteries non pathogegravenes potentiellement opportunistes 11
b Les mycobacteacuteries saprophytes non pathogegravenes 12
c Les mycobacteacuteries pathogegravenes obligatoires 12
Les mycobacteacuteries responsables de legravepre 12
Les mycobacteacuteries tuberculeuses le Complexe tuberculosis (MTBC) 13
B LA TUBERCULOSE UNE MALADIE TRES ANCIENNE MAIS UNE MENACE TOUJOURS ACTUELLE 20
1 Introduction 20
2 Histoire de la tuberculose 20
3 La tuberculose bovine 21
a Introduction 21
b Les hocirctes et les communauteacutes drsquohocirctes de Mycobacterium bovis dans le monde et leurs statuts
eacutepideacutemiologiques 22
c Aspect zoonotique de Mycobacterium bovis 23
d Pathogeacutenie de la tuberculose bovine 24
Facteurs influenccedilant le pouvoir pathogegravene de M bovis voies drsquoinfection et dose infectante 24
Facteurs influenccedilant la reacuteceptiviteacute et la sensibiliteacute de lrsquohocircte 25
Les eacutetapes de lrsquoinfection et la reacuteponse immunitaire associeacutee 26
Expression clinique de la maladie et leacutesions observeacutees 27
e Le deacutepistage et le diagnostic de lrsquoinfection chez les animaux 28
f Epideacutemiologie de la tuberculose bovine 30
Evolution de la tuberculose bovine dans le monde et en Europe et mesures de lutte 30
Evolution de la situation sanitaire en France 33
Evolution de la situation sanitaire en Cocircte drsquoOr 38
C ECOLOGIE DE M BOVIS DANS LrsquoENVIRONNEMENT 42
1 Introduction 42
2 Origines de la contamination environnementale par M bovis 42
a Lrsquoexcreacutetion animale de Mycobacterium bovis 42
b Le transfert de Mycobacterium bovis par les effluents drsquoeacutelevage ou urbains 44
3 Deacutetection et survie de Mycobacterium bovis dans lrsquoenvironnement 45
a Deacutetection de Mycobacterium bovis en conditions naturelles 45
b Dureacutees de survie de Mycobacterium bovis en conditions expeacuterimentales 47
4 Strateacutegies de survie des mycobacteacuteries dans lrsquoenvironnement 54
a Lrsquoeacutetat de dormance 54
b La sporulation 55
c Interactions des mycobacteacuteries avec la faune du sol 56
d Interactions des mycobacteacuteries avec les diptegraveres 57
e La formation de biofilms 57
5 Facteurs de risque de contamination de lrsquoenvironnement et de transmission de Mycobacterium
bovis aux animaux par lrsquoenvironnement contamineacute 58
a Risques lieacutes agrave lrsquoeacutecologie des espegraveces sauvages sensibles agrave Mycobacterium bovis 58
b Risques lieacutes aux pratiques drsquoeacutelevage 60
c Risques lieacutes aux pratiques cyneacutegeacutetiques 61
d Risques lieacutes aux facteurs paysagers climatiques et peacutedologiques 62
D PRINCIPAUX OUTILS DE DETECTION DE PHENOTYPAGE ET DE GENOTYPAGE APPLICABLES AU MTBC ET
PLUS PARTICULIEREMENT A MYCOBACTERIUM BOVIS 64
1 La deacutetection des mycobacteacuteries par microscopie 64
2 Les meacutethodes de culture des mycobacteacuteries 65
a Les diffeacuterents traitements preacutealables agrave la mise en culture des eacutechantillons 65
b La culture sur milieu solide 66
c La culture en milieu liquide 67
3 Meacutethodes de deacutetection et drsquoidentification des bacteacuteries du MTBC 68
a Caracteacuterisation pheacutenotypique et biochimique des mycobacteacuteries du MTBC 69
b Lrsquoidentification des mycobacteacuteries par recherche de biomarqueurs 69
c Lrsquoidentification des mycobacteacuteries par des meacutethodes drsquohybridation coupleacutees ou non agrave la PCR 70
d Le seacutequenccedilage 71
e Deacutetection et geacutenotypage des mycobacteacuteries du MTBC les marqueurs utiliseacutes 72
Les seacutequences drsquoinsertion 72
Les courtes seacutequences drsquoADN reacutepeacuteteacutees 73
Reacutegions de Diffeacuterence (RD) et Single Nucleotide Polymorphism (SNP) 78
f Identification par seacutequenccedilage complet des geacutenomes 79
g Meacutethode speacutecifique drsquoidentification des mycobacteacuteries animales utiliseacutee par le Laboratoire National de
reacutefeacuterence de la tuberculose bovine LNR TB 79
OBJECTIFS DE LA THESE 82
CHAPITRE 2 - DEVELOPPEMENT ET OPTIMISATION DrsquoOUTILS POUR LA DETECTION DE MYCOBACTERIUM
BOVIS DANS LES ECHANTILLONS ENVIRONNEMENTAUX 86
A DEVELOPPEMENT DrsquoOUTILS MOLECULAIRES ADAPTES AUX MATRICES ENVIRONNEMENTALES 86
CHAPITRE 3 - ETUDE DE LA PREVALENCE ENVIRONNEMENTALE DE MYCOBACTERIUM BOVIS DANS LA ZONE
ENDEMIQUE DE TUBERCULOSE BOVINE DE COTE DrsquoOR ET RESULTATS OBTENUS EN CHARENTE 93
A AVANT-PROPOS 93
B FIRST MOLECULAR DETECTION OF MYCOBACTERIUM BOVIS IN ENVIRONMENTAL SAMPLES FROM A
FRENCH REGION WITH ENDEMIC BOVINE TUBERCULOSIS 95
C CAMPAGNES DE PRELEVEMENTS SUPPLEMENTAIRES 121
1 Fin des preacutelegravevements en Cocircte drsquoOr 121
2 Preacutelegravevements en Charente 123
3 Compleacutements paysage geacuteomorphologie et peacutedologie 125
D BILAN GLOBAL 130
CHAPITRE 4 - IMPACT DE LA TEMPERATURE ET DE LA NATURE DU SOL SUR LA SURVIE DE MYCOBACTERIUM
BOVIS SB0120 133
A AVANT-PROPOS 133
B EXPERIMENTAL INVESTIGATION OF THE IMPACT OF THE TEMPERATURE AND THE SOIL TYPE ON
MYCOBACTERIUM BOVIS SURVIVAL IN THE ENVIRONMENT 135
C BILAN 150
CHAPITRE 5 - ROLE DE LA FAUNE DU SOL DANS LA CIRCULATION DE MYCOBACTERIUM BOVIS ETUDE DE SA
DISSEMINATION PAR LES LOMBRICS 155
A AVANT-PROPOS 155
B RAPID DISSEMINATION OF M BOVIS FROM CATTLE DUNG TO SOIL BY THE EARTHWORM LUMBRICUS
TERRESTRIS 157
C COMPLEMENT PRELEVEMENT ET ANALYSE DE LOMBRICS DANS UNE PATURE SUSPECTE 174
D BILAN 175
DISCUSSION GENERALE 177
1 Des meacutethodes deacuteveloppeacutees (ou adapteacutees) puis valideacutees dans des matrices environnementales 179
2 Contamination environnementale les zones humides les eaux de ruissellement et les sols de
terriers sont potentiellement agrave risque 180
3 Survie et circulation de M bovis dans lrsquoenvironnement 183
PERSPECTIVES 185
REFERENCES BIBLIOGRAPHIQUES 191
ANNEXES 221
Liste des Figures
Figure 1 Structure et composition de lenveloppe des mycobacteacuteries (drsquoapregraves Marrakchi Laneacuteelle and Daffeacute 2014) AG arabinogalactane PG peptidoglycane TDM treacutehalose dimycolate PL phospholipides TMM treacutehalose monomycolate GL glycolipides 7
Figure 2 Arbre phylogeacutenique baseacute sur lrsquoanalyse de lrsquoARNr 16S des principaux complexes et espegraveces de mycobacteacuteries (drsquoapregraves Magee et Ward 2012) 9
Figure 3 Phylogeacutenie des souches du MTBC baseacutee sur les Reacutegions de Diffeacuterences et diffeacuterents Single Nucleotide Polymorphism (Dippenaar et al 2015) 14
Figure 4 Aspect des colonies de M bovis sur milieu de Lowenstein-Jensen (agrave gauche) et Middlebrook 7H11 suppleacutementeacute (agrave droite) 17
Figure 5 Evolution de la reacuteponse immunitaire chez le bovin infecteacute par M bovis au cours du temps et aux diffeacuterents tests de deacutepistage de la tuberculose bovine (de la Rua-Domenech et al 2006 adapteacute de Vordermeier et al 2004) 27
Figure 6 Carte de distribution de la tuberculose bovine chez les bovins entre juillet et deacutecembre 2014 (Source WAHID Interface) 31
Figure 7 Statuts des pays europeacuteens vis-agrave-vis de la tuberculose bovine en 2012 et proportion de bovins infecteacutes OTF Officially Tuberculosis Free (officiellement indemne de tuberculose bovine) MS Member State (Etat membre) (Source EFSA 2014) 32
Figure 8 Evolution du taux de preacutevalence annuelle national de la tuberculose bovine chez les bovins (Beacutenet et al 2006) 34
Figure 9 Evolution de la preacutevalence et de lrsquoincidence de la tuberculose bovine en France entre 1995 et 2003 (Cavalerie et al 2014) 35
Figure 10 Incidence et distribution de la tuberculose bovine en France dans les anneacutees 50 (en haut) et entre 2000 et 2013 (en bas) (Cavalerie et al 2014) 36
Figure 11 Reacutepartition et reacutesultats des analyses effectueacutees dans le cadre du dispositif Sylvatub chez le grand gibier drsquoaoucirct 2013 agrave aoucirct 2014 et chez les blaireaux en 2014 (Source Sylvatub) 38
Figure 12 Localisation du parcellaire des cheptels infecteacutes par la tuberculose bovine entre 2010 et 2015 et spoligotypes incrimineacutes (Source CIREV) 40
Figure 13 Nombre de nouveaux foyers bovins et de foyers recontamineacutes entre 2002 et 2014 (Donneacutees DDPP21) 40
Figure 14 Nombre de cervideacutes sangliers et blaireaux infecteacutes deacutecouverts en Cocircte drsquoOr entre 2002 et 2014 41
Figure 15 Repreacutesentation scheacutematique de la circulation de M bovis dans lrsquoenvironnement agricole (modifieacute drsquoapregraves Elliott et al 2014) 63
Figure 16 Profils de restriction disolats de M tuberculosis obtenus suite agrave une RFLP-IS6110 Les isolats 3 5 6 9 et 10 ont le mecircme profil et posseacutedaient un lien eacutepideacutemiologique S repreacutesente le marqueur de poids moleacuteculaire (Source web site CDC) 73
Figure 17 Principe du spoligotypage des bacteacuteries du MTBC dapregraves Kamerbeek et al (1997) A Repreacutesentation du locus DR de M tuberculosis H37Rv et M bovis BCG B Amplifications des espaceurs par un couple damorces (a et b) et amplicons obtenus C Profils drsquohybridation sur membrane (un carreacute noir signale la preacutesence de lrsquoespaceur les parties blanches leur absence) 75
Figure 18 Diffeacuterences geacutenomiques de la reacutegion RD1 entre M bovis BCG (A) M microti (C) par comparaison agrave M tuberculosis H37Rv (B) drsquoapregraves Brodin et al 2002 et localisation du locus Rv3866 RD1mic correspond agrave la seacutequence de M tuberculosis deacuteleacuteteacutee chez M microti RD1bcg correspond agrave la seacutequence de M tuberculosis deacuteleacuteteacutee chez M bovis BCG 88
Figure 27 Localisation des communes de Cocircte drsquoOr sur lesquelles de foyers bovins et des animaux sauvages infecteacutes ont eacuteteacute deacutetecteacutes et localisation des preacutelegravevements environnementaux effectueacutes en 2015 (les preacutelegravevements positifs sont repreacutesenteacutes par des eacutetoiles noires) (Reacutealisation Marina Beacuteral CIREV Bourgogne) 123
Figure 28 Localisation des communes de Charente sur lesquelles de foyers bovins et des animaux sauvages infecteacutes ont eacuteteacute deacutetecteacutes et localisation des preacutelegravevements environnementaux effectueacutes en 2015 (les preacutelegravevements positifs sont repreacutesenteacutes par des eacutetoiles noires) (Reacutealisation Marina Beacuteral CIREV Bourgogne) 125
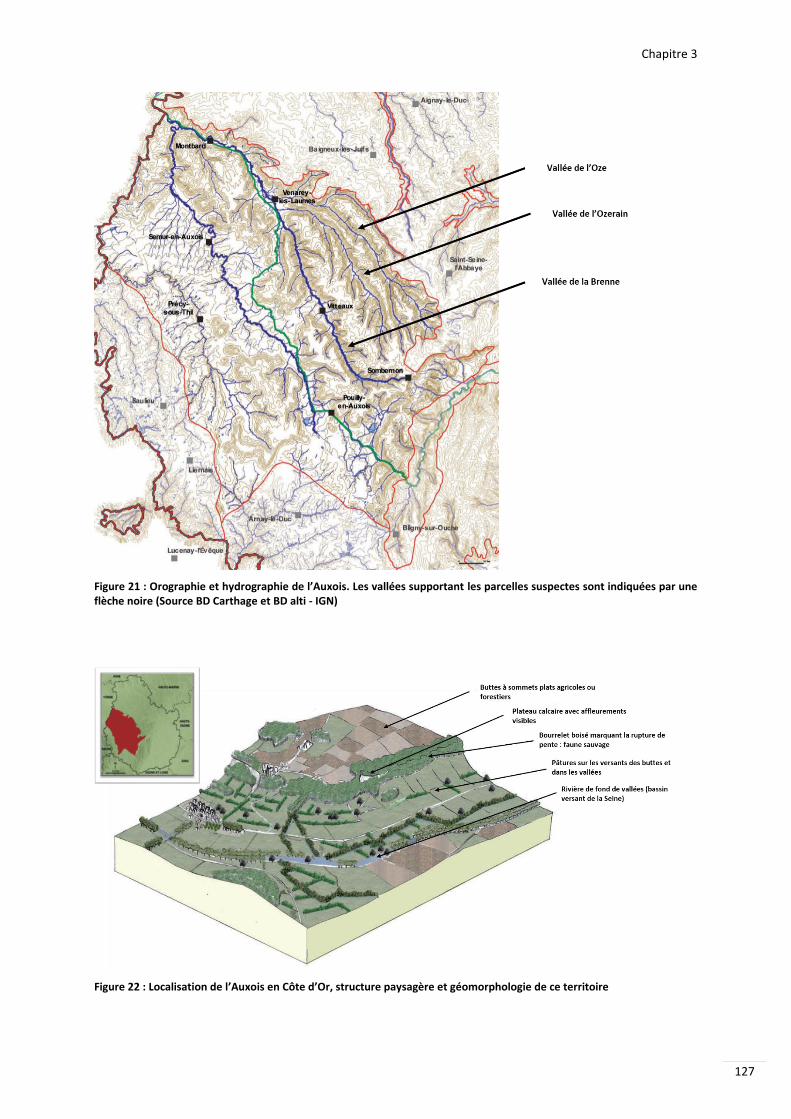
Figure 29 Orographie et hydrographie de lrsquoAuxois Les valleacutees supportant les parcelles suspectes sont indiqueacutees par une flegraveche noire (Source BD Carthage et BD alti - IGN) 127
Figure 30 Localisation de lrsquoAuxois en Cocircte drsquoOr structure paysagegravere et geacuteomorphologie de ce territoire 127
Figure 31 Nombre de vaches allaitantes par canton dans le deacutepartement de la Cocircte drsquoOr (agrave gauche) et parcellaire des foyers bovins de 2010 agrave 2015 (agrave droite) (Source Recensement agricole 2010 ndash Agreste) 128
Figure 32 Reacutepartition spatiale des parcelles des eacutelevages bovins infecteacutes de 2010 agrave 2014 en Cocircte drsquoOr et repreacutesentation des couches peacutedologiques (Reacutealisation Marina Beacuteral CIREV Bourgogne) 129
Figure 33 Tempeacuteratures minimales et maximales observeacutees agrave Dijon-Longvic de 2012 agrave 2015 (Source Meacuteteacuteo France) 151
Liste des Tableaux
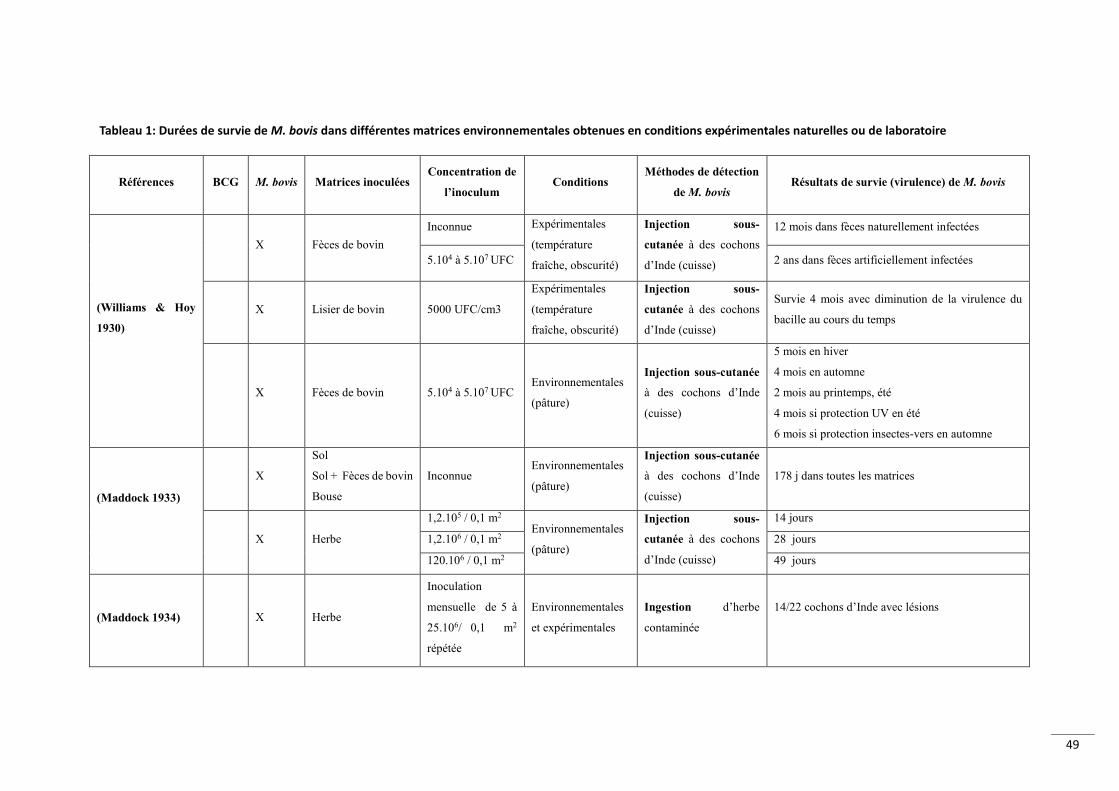
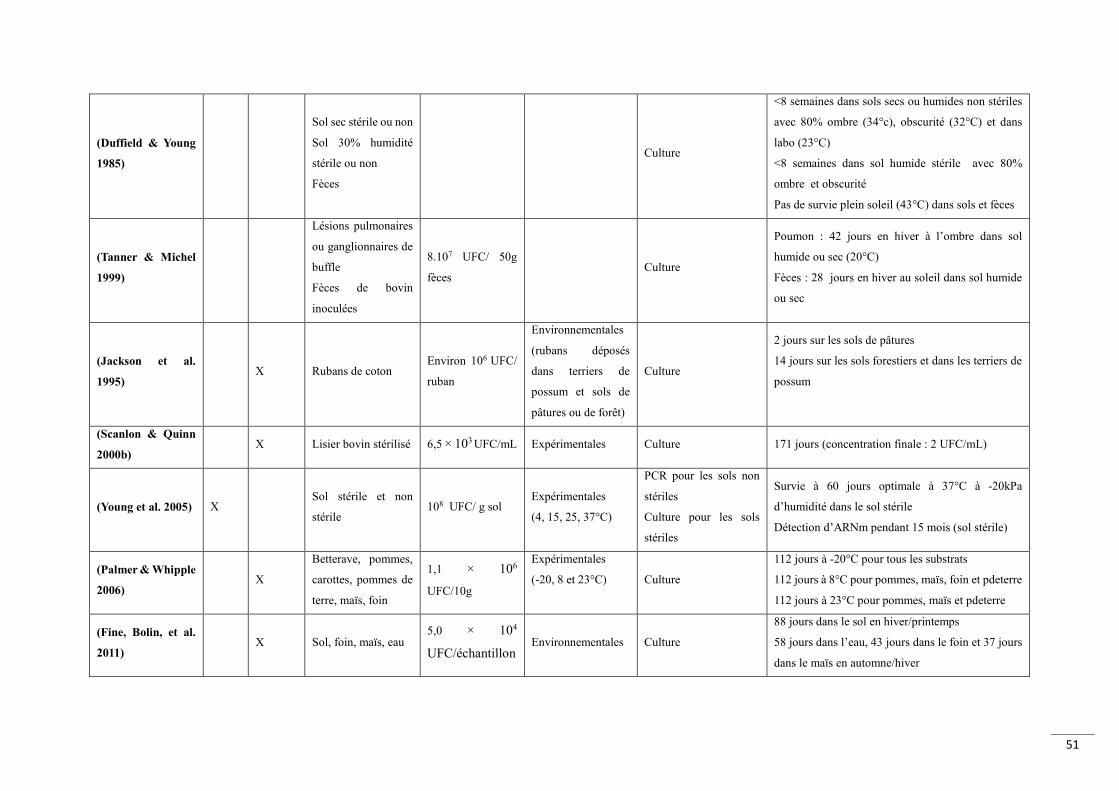
Tableau 1 Dureacutees de survie de M bovis dans diffeacuterentes matrices environnementales obtenues en conditions expeacuterimentales naturelles ou de laboratoire 49
Tableau 2 Identification des principales mycobacteacuteries du MTBC par des observations pheacutenotypiques et des tests biochimiques 69
Tableau 3 Cibles moleacuteculaires deacutecrites dans la litteacuterature pour la deacutetection du Complexe tuberculosis ou de M bovis dans les eacutechantillons environnementaux 76
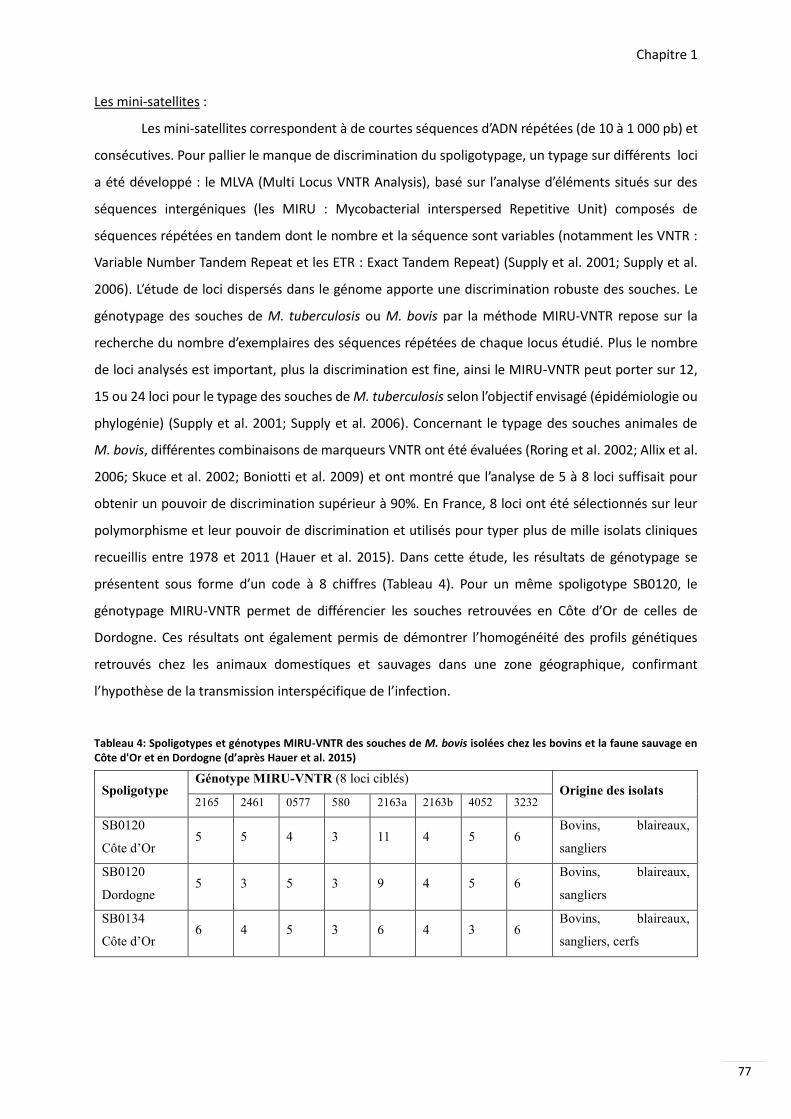
Tableau 4 Spoligotypes et geacutenotypes MIRU-VNTR des souches de M bovis isoleacutees chez les bovins et la faune sauvage en Cocircte dOr et en Dordogne (drsquoapregraves Hauer et al 2015) 77
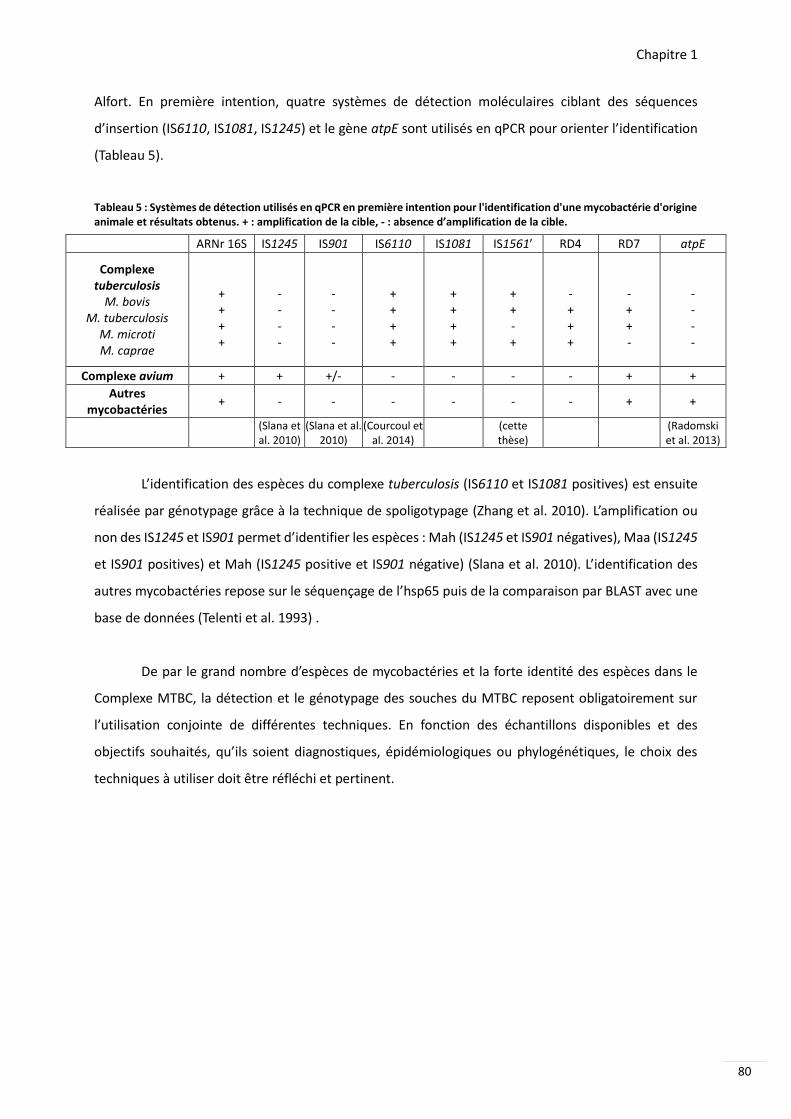
Tableau 5 Systegravemes de deacutetection utiliseacutes en qPCR en premiegravere intention pour lidentification dune mycobacteacuterie dorigine animale et reacutesultats obtenus 80
Liste des Annexes
Annexe 1 Deacutetection de Mycobacterium bovis dans lrsquoenvironnement Premiers reacutesultats des recherches meneacutees en Cocircte-drsquoOr 223
Annexe 2 Preacuteparation du Middlebrook 7H9 235
Annexe 3 Preacuteparation du Middlebrook 7H11 237
Liste des Abreacuteviations
ADC Albumine Dextrose Catalase
ADNDNA Acide deacutesoxyribonucleacuteique
AEEMA Association pour lEtude de lEpideacutemiologie des Maladies Animales
AFNOR Agence Franccedilaise de NORmalisation
ANSES Agence nationale de seacutecuriteacute sanitaire de lrsquoalimentation de lrsquoenvironnement et du travail
ARNRNA Acide ribonucleacuteique
ARNr ARN ribosomique
BAAR Bacille acido-alcoolo reacutesistant
BCG Bacille de Calmette et Gueacuterin
BLAST Basic Local Alignment Search Tool
bTB bovine Tuberculosis
CDC Centers of Disease Control and Prevention
CFP-10 Culture Filtrat Protein 10 kD
CIREV Cellules Inter Reacutegionales dEpideacutemiologie Veacuteteacuterinaire
CRISPR Clustered Regularly Interspersed Short Palindromic Repeats
Ct Cycle Threshold
DDPP Direction Deacutepartementale de la Protection des Populations
DEFRA Department for Environment Food and Rural Affairs
DGAL Direction Geacuteneacuterale de lALimentation
DosR Dormancy Survival Regulator
DR Direct Repeat
EFSA European Food Safety Authority
ELISA Enzyme-Linked ImmunoSorbent Assay
ESAT-6 Early Secreted Antigenic Target 6kD
ETR Exact Tandem Repeat
Fbp Fibronectin binding protein
FISH Fluorescence In Situ Hybridization
GC-MS Gas Chromatography-Mass Spectrometry
GDS Groupements de Deacutefense Sanitaire
HAP Hydrocarbures Aromatiques Polycycliques
HIV Human Immunodeficiency Virus
HPLC High Performance Liquid Chromatography
IDC Intradermotuberculination comparative
IDR Intradermotuberculination
IDS Intradermotuberculination simple
IFN-γ Interfeacuteron γ
INRA Institut National de la Recherche Agronomique
Ion PGM Ion Personal Genome Machine
IS Insertion Sequence
ITS Internal Transcribed Spacer
kDkDa kilo Daltons
LNR Laboratoire National de Reacutefeacuterence
LSP Large Sequence Polymorphism
Maa Mycobacterium avium subspecies avium
MAC Mycobacterium avium complex
MALDI-TOF Matrix-Assisted Laser Desorption Ionization-Time Of Flight
Map Mycobacterium avium subspecies paratuberculosis
MDR Multi Drug Resistant
MIRU Mycobacterial Interspersed Repetitive Unit
MLVA Multi Locus VNTR Analysis
MNT Mycobacteacuteries Non Tuberculeuses
MS Member State
MTBC Mycobacterium tuberculosis complex
NCBI National Center for Biotechnology Information
NF Norme Franccedilaise
NGS Next Generation Sequencing
OADC Oleic acid Albumine Dextrose Catalase
OIE Office International des Epizooties
OMS Organisation Mondiale de la Santeacute
ONCFS Office National de la Chasse et de la Faune Sauvage
ONEMA Office National de lEau et des Milieux Aquatiques
OTF Officialy Tuberculosis Free
pb paire de bases
PCR Polymerase chain reaction
PNA Peptide Nucleic Acid
qPCR quantitative Polymerase Chain Reaction
RD Reacutegions de Diffeacuterence
RFLP Restriction Fragment Lenght Polymorphism
RNS Reactive Nitrogen Species
ROS Reactive Oxygen Species
RT-PCR Reverse-Transcription Polymerase Chain Reaction
SNP Single Nucleotide Polymorphism
ssp subspecies
TDM Treacutehalose dimycolate
TMM Treacutehalose monomycolate
UFCCFU Uniteacute Formant Colonie
VBNC Viable But Non Cultivable
VNTR Variable Number Tandem Repeat
WAHID World Animal Health Information Database
WGS Whole Genome Sequencing
WHO World Health Organization
XDR eXtensively Drug Resistant
1
Introduction geacuteneacuterale
La tuberculose bovine est une maladie drsquoimportance sanitaire majeure en santeacute animale agrave
travers le monde Son agent principal Mycobacterium bovis est un membre du Complexe
Mycobacterium tuberculosis qui regroupe les bacteacuteries tuberculeuses infectant lrsquohomme et les
mammifegraveres Les bovins constituent le reacuteservoir originel de M bovis mais de nombreuses espegraveces de
mammifegraveres peuvent eacutegalement ecirctre infecteacutees dont lrsquohomme Les enjeux sanitaires eacuteconomiques et
sociaux induits par cette maladie ont conduit agrave la mise en place de programmes visant lrsquoeacuteradication de
cette maladie chez les bovins agrave travers le monde Dans les pays deacuteveloppeacutes ces mesures associeacutees agrave
la pasteurisation du lait ont fortement reacuteduit lrsquoincidence de la tuberculose bovine voire eacuteradiqueacute
lrsquoinfection chez les bovins Toutefois dans quelques pays lrsquoinfection persiste dans les cheptels bovins
malgreacute des actions de preacutevention de controcircle et de surveillance Dans les pays en deacuteveloppement ougrave
les controcircles organiseacutes sont quasi- voire totalement absents la preacutevalence et la propagation de la
tuberculose bovine sont inquieacutetantes et repreacutesentent un risque sanitaire et eacuteconomique vis-agrave-vis des
populations sensibles En conseacutequence la tuberculose bovine attire chaque anneacutee un peu plus
lrsquoattention de la communauteacute internationale en raison du nombre croissant de troupeaux infecteacutes et
des effets sur les productions animales ainsi que du risque que la maladie fait planer sur la santeacute
publique
Le passage de lrsquoinfection du reacuteservoir bovin agrave diffeacuterentes populations sauvages qui
entretiennent la maladie et peuvent la transmettre en retour aux bovins a fortement compliqueacute les
scheacutemas drsquoeacuteradication de la maladie dans plusieurs reacutegions du monde et explique lrsquoeacutechec partiel des
programmes de lutte mis en place Alors que la transmission directe du bacille par voie respiratoire
semble ecirctre le mode de transmission preacutepondeacuterant au sein drsquoune mecircme espegravece la transmission
indirecte de M bovis est fortement suspecteacutee entre espegraveces partageant les mecircmes territoires et
ressources compliquant encore lrsquoeacutepideacutemiologie de cette maladie
En France une recrudescence du nombre de cheptels bovins infecteacutes dans certains
deacutepartements dont la Cocircte drsquoOr menace son statut de pays laquo officiellement indemne de tuberculose
bovine raquo obtenu en 2001 Dans ce deacutepartement ougrave plus de 190 foyers bovins ont eacuteteacute deacutetecteacutes depuis
2003 M bovis circule dans un systegraveme multi-hocirctes incluant les bovins les cervideacutes les blaireaux et les
sangliers dans une zone geacuteographique localiseacutee Face agrave des contaminations ou des recontaminations
inexpliqueacutees de cheptels avec des souches identiques agrave celles retrouveacutees dans la faune sauvage
infecteacutee deacutetecteacutee agrave proximiteacute dans des eacutelevages le rocircle de lrsquoenvironnement a eacuteteacute suggeacutereacute dans la
transmission indirecte du bacille
Dans ce contexte mes travaux de thegravese visent agrave eacutevaluer la contamination environnementale
2
des zones partageacutees par les bovins et la faune sauvage infecteacutes gracircce agrave des meacutethodes adapteacutees
utilisables sur un grand nombre de preacutelegravevements et agrave rechercher les facteurs favorisant la survie et la
circulation de M bovis dans lrsquoenvironnement Le premier chapitre de ce manuscrit est une synthegravese
des connaissances relatives aux mycobacteacuteries en geacuteneacuteral et aux mycobacteacuteries tuberculeuses en
particulier notamment en terme drsquoeacutecologie de deacutetection et drsquoidentification Le deuxiegraveme chapitre
deacutetaille le deacuteveloppement des meacutethodes moleacuteculaires utiliseacutees pour le diagnostic environnemental
ainsi que le deacuteveloppement de deux nouveaux protocoles pour isoler les mycobacteacuteries du sol et suivre
leur croissance en milieu liquide Le troisiegraveme chapitre preacutesente les modaliteacutes drsquoeacutetude et les reacutesultats
obtenus entre 2013 et 2015 dans lrsquoeacutevaluation de la contamination environnementale en Cocircte drsquoOr Les
reacutesultats drsquoune eacutetude reacutealiseacutee en Charente sont eacutegalement preacutesenteacutes Le quatriegraveme chapitre traite de
lrsquoinfluence de la tempeacuterature et du type de sol sur la dureacutee de survie de M bovis Enfin le cinquiegraveme
chapitre srsquointeacuteresse au rocircle des lombrics sur la disseacutemination de M bovis dans le sol Pour terminer
une discussion geacuteneacuterale syntheacutetise lrsquoensemble des reacutesultats obtenus lors de ce travail et ouvre de
nombreuses perspectives de recherche
Cette thegravese financeacutee par la Reacutegion Bourgogne entre dans le dispositif Jeunes Chercheurs
Entrepreneurs (JCE) Le travail scientifique de recherche srsquoaccompagne de la reacutealisation drsquoun Master
des Entreprises (Master 2) dans lrsquoobjectif drsquoouvrir les doctorants au monde de lrsquoentreprise et au
deacuteveloppement de projets innovants Ce Master a eacuteteacute lrsquooccasion pour moi drsquoacqueacuterir des compeacutetences
compleacutementaires dans le domaine de lrsquoadministration et de la gestion des entreprises notamment
dans la gestion de projets Mon investissement personnel important en parallegravele de mon travail de
recherche mrsquoa permis drsquoobtenir le Master avec la mention Bien
Dans le cadre de la thegravese JCE lrsquoentreprise LSI (speacutecialiseacutee dans le diagnostic veacuteteacuterinaire) a
accompagneacute le deacutemarrage de la thegravese et mrsquoa permis drsquoeacutechanger avec des industriels autour du
deacuteveloppement de systegravemes de deacutetection baseacutes sur la qPCR (IS1561rsquo et autres systegravemes) Suite au
rachat de lrsquoentreprise par Life Science puis Thermo Fisher le transfert de technologie nrsquoa pas eacuteteacute
poursuivi Cependant ces systegravemes de qPCR sont utiliseacutes en routine par le Laboratoire National de
Reacutefeacuterence pour la tuberculose bovine encadreacute par Maria Laura Boschiroli pour caracteacuteriser les souches
de mycobacteacuteries issues drsquoeacutechantillons animaux Drsquoautre part des travaux et reacutesultats obtenus lors de
ma thegravese vont faire lrsquoobjet drsquoune valorisation sous forme drsquoun brevet en cours de deacutepocirct pour la
deacutetection des mycobacteacuteries et drsquoun savoir-faire concernant lrsquoextraction des mycobacteacuteries du sol par
des meacutethodes chimiques en vue de leur mise en culture Jrsquoai donc au cours de mes travaux de recherche
toujours conserveacute lrsquoesprit drsquoinnovation et de transfert potentiel de technologie
CHAPITRE 1
Contexte bibliographique
Chapitre 1
4
CHAPITRE 1 - Contexte bibliographique
A LES MYCOBACTERIES DIVERSITE PHYLOGENIE SPECTRE DrsquoHOcircTES ET VIRULENCE
1 Le genre Mycobacterium taxonomie et meacutethodes de classification
Les connaissances sur les mycobacteacuteries commencent en 1882 date agrave laquelle Robert Koch
deacutecouvre le bacille responsable de la tuberculose humaine alors baptiseacute Bacterium tuberculosis et
partage son postulat sur lrsquoeacutetiologie infectieuse de la maladie La coloration de Ziehl et Neelsen
deacuteveloppeacutee en 1883 pour la recherche des bacilles acido-alcoolo-reacutesistants permet agrave Smith en 1892 de
mettre en eacutevidence un bacille tregraves proche de Bacterium tuberculosis dans des leacutesions tuberculeuses de
bovins Mycobacterium bovis avec toutefois des diffeacuterences pheacutenotypiques et pathogeacuteniques qui
permettent de distinguer les deux bacilles Le bacille de la tuberculose aviaire M avium ssp avium est
isoleacute en 1894 sur des oiseaux puis M avium ssp paratuberculosis en 1895 sur des leacutesions digestives
de bovins La deacutecouverte des premiegraveres mycobacteacuteries non tuberculeuses a suivi celle de Bacterium
tuberculosis notamment gracircce agrave la coloration Mycobacterium smegmatis a eacuteteacute isoleacutee en 1889 dans
des preacutelegravevements biologiques de patients atteints de syphilis puis dans du beurre par lrsquoeacutequipe de Koch
dans les anneacutees 1896 Les premiegraveres recherches meneacutees dans le sol les plantes puis lrsquoeau ont montreacute
que ces substrats pouvaient ecirctre des sources de bacteacuteries morphologiquement tregraves proches du bacille
de Koch Le genre Mycobacterium (mứkecircs champignon et baktecircrίa bacircton) est proposeacute en 1896 par
Lehmann et Neumann Dans les anneacutees 50 environ 20 espegraveces de mycobacteacuteries eacutetaient deacutecrites Le
deacuteveloppement des meacutethodes de culture puis des outils moleacuteculaires dans les anneacutees 1990 ont permis
drsquoisoler de nouvelles espegraveces dans des niches environnementales varieacutees ainsi que chez lrsquohomme et
les animaux A lrsquoheure actuelle 172 espegraveces de mycobacteacuteries sont identifieacutees (Site bacterionetorg)
La diversiteacute pheacutenotypique physiologique eacutecologique et pathogeacutenique des espegraveces placeacutees au
fur et agrave mesure de leur deacutecouverte dans ce genre a meneacute agrave lrsquoeacutelaboration de plusieurs classifications
Au sein du genre Mycobacterium les espegraveces sont geacuteneacuteralement classeacutees en deux grands groupes un
groupe qui rassemble les mycobacteacuteries pathogegravenes obligatoires que sont les mycobacteacuteries
tuberculeuses et les mycobacteacuteries responsables de la legravepre et un second groupe qui rassemble les
mycobacteacuteries pathogegravenes opportunistes et saprophytes appeleacute mycobacteacuteries atypiques ou
mycobacteacuteries environnementales ou mycobacteacuteries non tuberculeuses (MNT)
Dans ces groupes des complexes drsquoespegraveces ont eacuteteacute deacutefinis sur des bases drsquoidentiteacute geacuteneacutetique Le
Complexe tuberculosis (MTBC) rassemble les mycobacteacuteries tuberculeuses notamment
Chapitre 1
5
M tuberculosis M africanum M canetti M bovis M caprae M microti M pinnipedii Parmi les
nombreuses MNT diffeacuterents complexes sont observeacutes dont le plus important est le Complexe
Mycobacterium avium (MAC) qui regroupe quatre sous-espegraveces distinctes dont trois drsquointeacuterecirct en santeacute
humaine et animale (M avium ssp avium M avium ssp paratuberculosis et M avium ssp hominisuis)
(Radomski et al 2010) ainsi qursquoune dizaine drsquoespegraveces dont M intracellulare
Plus reacutecemment la phylogeacutenie moleacuteculaire de ce genre baseacutee notamment sur lrsquoanalyse des gegravenes de
lrsquoARNr 16S et 23S ainsi que des reacutearrangements de gegravenes a permis drsquoaffiner la position taxonomique
du genre Mycobacterium Drsquoapregraves le Bergeyrsquos manual (Ludwig et al 2008) le genre Mycobacterium
appartient
au Phylum des Actinobacteria (bacteacuteries Gram positif agrave geacutenome riche en acides nucleacuteiques
G et C ()
o agrave la Classe des Actinobacteria
o agrave lrsquoOrdre des Corynebacteriales qui regroupe diffeacuterentes familles de bacteacuteries
caracteacuteriseacutees par la preacutesence drsquoacides mycoliques dans leur paroi et des
proprieacuteteacutes tinctoriales particuliegraveres lieacutees agrave la reacutesistance de leur paroi agrave la
deacutecoloration par un meacutelange alcool-acide (ou BAAR)
agrave la Famille des Mycobacteriaceae qui ne comprend que le genre
Mycobacterium caracteacuteriseacute par une enveloppe riche en acides
mycoliques contenant un grand nombre drsquoatomes de carbone ainsi
que des quinones particuliegraveres
2 Caracteacuteristiques geacuteneacuterales des mycobacteacuteries
a Morphologie des mycobacteacuteries
Les mycobacteacuteries se preacutesentent sous forme de bacirctonnets droits ou leacutegegraverement incurveacutes (02-
06 x 10-10microm) non mobiles et ne sporulant pas Les colonies observeacutees sont de couleur blanche agrave
cregraveme voire jaune agrave orangeacutee pour les mycobacteacuteries productrices de pigments (Magee amp Ward 2012)
Elles fixent difficilement la coloration de Gram et bien qursquoelles soient apparenteacutees aux bacteacuteries Gram
+ leur qualification exacte est laquo Bacilles Acido-Alcoolo-Reacutesistants (BAAR) raquo car lors de la coloration
leur enveloppe particuliegravere reacutesiste agrave la deacutecoloration agrave lrsquoalcool et agrave lrsquoacide
Les mycobacteacuteries possegravedent une enveloppe proche de celle des bacteacuteries Gram + mais dont
lrsquoorganisation la richesse et la diversiteacute des acides gras qui la composent sont uniques et
caracteacuteristiques La mycomembrane est constitueacutee drsquoacides gras agrave longue chaine contenant de 60 agrave 90
atomes de carbone les acides mycoliques qui repreacutesentent 30 agrave 40 de sa masse (Brennan amp Nikaido
Chapitre 1
6
1995) Dans sa couche interne les acides mycoliques sont disposeacutes parrallegravelement les uns aux autres
et sont lieacutes au peptidoglycane de faccedilon covalente par lrsquointermeacutediaire de la couche drsquoarabinogalactane
(polymegravere formeacute drsquoarabinose et de galactose) Sa couche externe contient des lipides extractibles (ie
non fixeacutes de faccedilon covalente au peptidoglycane) tels que le treacutehalose monomycolate ou dimycolate
des phospholipides et des glycolipides (Zuber et al 2008) La mycomembrane est recouverte de
glucanes de proteacuteines et drsquoune faible proportion de lipides
Cette enveloppe particuliegravere tregraves riche en lipides confegravere agrave la bacteacuterie ses proprieacuteteacutes
tinctoriales (BAAR) sa grande reacutesistance aux stress chimiques notamment aux antibiotiques et
deacutetergents aux stress physiques (tempeacuterature) ainsi que biologiques (reacutesistance dans les
macrophages de lrsquohocircte et dans les amibes) Crsquoest eacutegalement un facteur important de virulence chez les
mycobacteacuteries pathogegravenes certains composants lipidiques possegravedent en effet une activiteacute
immunogegravene exceptionnelle (Coulombe et al 2009) Ils sont capables de moduler lrsquoimmuniteacute de lrsquohocircte
voire de deacutesorganiser ses meacutecanismes immunitaires de deacutefense et interviennent dans la pathogeacutenie
comme cela a eacuteteacute deacutemontreacute pour M tuberculosis et M avium ssp paratuberculosis (Libero amp Mori
2014 Momotani et al 2012) La paroi est drsquoailleurs une des cibles des antimicrobiens utiliseacutes contre la
tuberculose humaine ou la legravepre Ces derniers agissent notamment en inhibant la biosynthegravese des
moleacutecules qui la constituent Lrsquoutilisation des antigegravenes lipidiques de la paroi constitue eacutegalement un
axe de recherche pour le deacuteveloppement de vaccins (Libero amp Mori 2014)
Dans les matrices environnementales la paroi hydrophobe des mycobacteacuteries leur permet
drsquoadheacuterer fortement aux particules en suspension dans lrsquoeau aux seacutediments aux surfaces favorisant
la formation de biofilms et la survie des bacteacuteries (Primm et al 2004)
Les acides mycoliques de par leur complexiteacute et leur variabiliteacute structurelle sont eacutegalement utiliseacutes
comme marqueurs taxonomiques car ils permettent drsquoidentifier avec speacutecificiteacute le genre et les espegraveces
Chapitre 1
7
Figure 1 Structure et composition de lenveloppe des mycobacteacuteries (drsquoapregraves Marrakchi Laneacuteelle and Daffeacute 2014) AG arabinogalactane PG peptidoglycane TDM treacutehalose dimycolate PL phospholipides TMM treacutehalose monomycolate GL glycolipides
b Caracteacuteristiques culturales pheacutenotypiques et physiologiques
La plupart des mycobacteacuteries sont aeacuterobies strictes mais quelques espegraveces tolegraverent de faibles
pressions drsquooxygegravene notamment M bovis et M tuberculosis Les substrats utiliseacutes comme sources de
carbone et drsquoazote varient consideacuterablement entre les espegraveces et peuvent servir pour leur
identification La majoriteacute des espegraveces croit sur des milieux de culture classiques mais pour certaines
mycobacteacuteries drsquoimportance clinique la culture est ameacutelioreacutee par lrsquoutilisation de milieux agrave lrsquoœuf
(Loumlwenstein-Jensen et Coletsos) apregraves une deacutecontamination chimique ou de milieu Middlebrook ainsi
que par un enrichissement des milieux (en glyceacuterol pour M tuberculosis en mycobactine pour
M avium subsp paratuberculosis en heacutemine pour M haemophilum) Les mycobacteacuteries responsables
de la legravepre sont cultiveacutees sur tissus vivants (coussinets de souris peau de tatou) et se multiplient tregraves
difficilement sur milieux syntheacutetiques
Les mycobacteacuteries ont des vitesses de croissance tregraves variables on distingue les mycobacteacuteries
agrave croissance rapide dont les colonies sont visibles en moins de 7 jours des mycobacteacuteries agrave croissance
lente (gt7 jours) Les bacteacuteries du MTBC sont toutes des mycobacteacuteries agrave croissance lente avec des
temps de geacuteneacuteration eacuteleveacutes de 14 agrave 15h pour M tuberculosis et environ 20h pour M bovis La
tempeacuterature optimale de croissance des bacteacuteries du MTBC est de 37degC Les MNT regroupent des
mycobacteacuteries agrave croissance rapide et agrave croissance lente avec des taux de geacuteneacuteration extrecircment
variables allant de 2h pour les bacteacuteries agrave croissance rapide comme M smegmatis agrave 21h pour celles
agrave croissance lente comme M aurum voire 7 jours pour M avium ssp paratuberculosis dans des
conditions de laboratoire pourtant optimales Les MNT peuvent se multiplier dans un large eacuteventail de
Chapitre 1
8
tempeacuterature allant de 4degC pour M psychrotolerans (Trujillo et al 2004) agrave plus de 45degC voire agrave 52degC
pour M thermotolerans et M phlei (Stahl amp Urbance 1990) avec une tempeacuterature optimale de
croissance entre 28 et 31degC
c Phylogeacutenie du genre Mycobacterium
La phylogeacutenie baseacutee sur le polymorphisme de lrsquoADN codant pour lrsquoARNr 16S permet de
discriminer les espegraveces de mycobacteacuteries et met en eacutevidence trois grands groupes en fonction de leur
vitesse de croissance Le cladogramme preacutesenteacute dans la Figure 2 regroupe distinctement les
mycobacteacuteries agrave croissance lente (haut du cladogramme) les mycobacteacuteries agrave croissance intermeacutediaire
(au milieu) et les mycobacteacuteries agrave croissance rapide (bas du cladogramme) On peut noter de plus que
les mycobacteacuteries du MTBC et le complexe Mac forment deux groupes phylogeacuteneacutetiquement
homogegravenes au sein du groupe des mycobacteacuteries agrave croissance lente
Myc
ob
acteacute
ries
agrave c
rois
san
ce le
nte
Myc
ob
acteacute
ries
agrave
cro
issa
nce
inte
rmeacute
dia
ire
Chapitre 1
9
Figure 2 Arbre phylogeacutenique baseacute sur lrsquoanalyse de lrsquoARNr 16S des principaux complexes et espegraveces de mycobacteacuteries
(drsquoapregraves Magee et Ward 2012)
d Caracteacuteristiques geacuteneacutetiques des mycobacteacuteries
De nombreux geacutenomes complets (annoteacutes ou en cours drsquoanalyse) de mycobacteacuteries sont
actuellement disponibles (NCBI Genome Resources European Nucleotide archive) Mycobacterium
leprae est la mycobacteacuterie qui possegravede le geacutenome le plus petit avec 33 millions de paires de bases
(Mpb) codant pour 1 605 gegravenes et le G+C (pourcentage Guanine+Cytosine) le plus faible (57) Les
mycobacteacuteries environnementales possegravedent les geacutenomes les plus importants de 483 Mpb (codant
4 350 gegravenes) pour M avium k10 agrave 69 Mpb (codant 6 716 gegravenes) pour M smegmatis Les mycobacteacuteries
tuberculeuses M tuberculosis M africanum et M bovis ont un geacutenome de taille intermeacutediaire (de 43
Mpb agrave 45 Mpb) codant pour 3 850 agrave 4 500 gegravenes Les geacutenomes des mycobacteacuteries preacutesentent des
capaciteacutes codantes importantes de lrsquoordre de 90 pour M tuberculosis M bovis M avium ssp
paratuberculosis et M smegmatis (Garnier et al 2003 Cole et al 1998 Li et al 2005)
La comparaison du geacutenome complet de M tuberculosis M bovis M leprae et M avium ssp
paratuberculosis a montreacute une forte conservation des gegravenes impliqueacutes dans le meacutetabolisme
Myc
ob
acteacute
ries
agrave c
rois
san
ce r
apid
e
Chapitre 1
10
eacutenergeacutetique la biosynthegravese des acides amineacutes et des cofacteurs et le meacutetabolisme des nucleacuteotides et
des proteacuteines (Marri et al 2006) Des plasmides ont eacuteteacute mis en eacutevidence chez quelques espegraveces de
mycobacteacuteries environnementales (Rauzier et al 1988 Stinear et al 2004) notamment le plasmide
pMUM001 isoleacute chez M ulcerans responsable de lrsquoulcegravere de Buruli chez lrsquohomme qui code pour un
macrolide aux proprieacuteteacutes cytotoxiques et immunosuppressives la mycolactone (Stinear et al 2004)
Le nombre drsquoopeacuterons ribosomaux 16S dans le geacutenome des mycobacteacuteries a eacuteteacute suggeacutereacute comme
lrsquoun des facteurs expliquant la diffeacuterence des vitesses de croissance observeacutees La croissance lente des
mycobacteacuteries serait lieacutee agrave la preacutesence drsquoun seul opeacuteron ribosomal les espegraveces agrave croissance rapide en
posseacutedant geacuteneacuteralement deux (rrnA et rrnB) (Bercovier et al 1986) Toutefois des exceptions existent
et drsquoautres facteurs tels que la lenteur des eacutechanges agrave travers leur paroi impermeacuteable riche en lipides
et le coucirct eacutenergeacutetique eacuteleveacute de la synthegravese de leurs acides gras agrave longues chaines seraient eacutegalement
impliqueacutes (Primm et al 2004)
Dans le Complexe tuberculosis les diffeacuterentes espegraveces constituent un modegravele drsquoidentiteacute
geacuteneacutetique interspeacutecifique Leurs geacutenomes preacutesentent une identiteacute supeacuterieure agrave 999 au niveau
nucleacuteotidique et des seacutequences drsquoARNr 16S et drsquoITS identiques (Boumlddinghaus et al 1990 Sreevatsan amp
Pan 1997 Huard et al 2006 Cole et al 1998 Garnier et al 2003 Kasai et al 2000) Lrsquoanalyse des
seacutequences de 56 gegravenes de structure de plusieurs centaines drsquoisolats drsquoorigines geacuteographique et
phylogeacuteneacutetique varieacutees suggegravere que le polymorphisme alleacutelique est exceptionnel dans ce Complexe
(Sreevatsan amp Pan 1997 Musser et al 2000) La recombinaison de tregraves petits fragments drsquoADN est
deacutemontreacutee chez M tuberculosis (Namouchi et al 2012) mais le transfert lateacuteral de gegravenes ne semble
pas ecirctre un facteur important dans lrsquoeacutevolution des bacteacuteries du MTBC qui ont une structure de
population fortement clonale (Pepperell et al 2013) contrairement agrave la plupart des autres bacteacuteries
pathogegravenes Malgreacute leur forte identiteacute geacuteneacutetique les espegraveces du Complexe se diffeacuterencient par des
reacutegions polymorphiques ou LSPs (Large Sequence Polymorphisms) lieacutees agrave des deacuteleacutetions et agrave des
reacutearrangements de seacutequences (Brosch et al 2002 Huard et al 2003) Parmi ces LSPs 14 Reacutegions de
Diffeacuterence ou RD de 2 agrave 127 kpb sont recenseacutees dans le MTBC Ces deacuteleacutetions sont consideacutereacutees comme
une adaptation eacutevolutive agrave lrsquoinfection de nouveaux hocirctes plutocirct qursquoagrave une deacuterive geacuteneacutetique (Brodin et
al 2002 Aranaz et al 2003) Cette variabiliteacute geacutenomique permet agrave la fois de diffeacuterencier les espegraveces
entre elles (Huard et al 2003 Huard et al 2006) et de retracer leur eacutevolution phylogeacuteneacutetique
(Rodriguez-Campos et al 2014)
3 Description des diffeacuterents Complexes ou espegraveces de mycobacteacuteries
Alors que les mycobacteacuteries responsables de la tuberculose et de la legravepre sont des pathogegravenes
obligatoires les MNT rassemblent des mycobacteacuteries saprophytes sans pouvoir pathogegravene jusque-lagrave
Chapitre 1
11
aveacutereacute et des mycobacteacuteries pathogegravenes opportunistes responsables de mycobacteacuterioses humaines ou
animales Depuis la fin du 19egraveme siegravecle jusqursquoaux anneacutees 1950 la tuberculose humaine repreacutesentait un
tel problegraveme de santeacute publique que toutes les eacutetudes eacutetaient centreacutees sur elle et son agent
M tuberculosis Les autres mycobacteacuteries isoleacutees dans lrsquoenvironnement ou dans des preacutelegravevements
biologiques humains et animaux eacutetaient consideacutereacutees comme des bacteacuteries saprophytes sans inteacuterecirct
clinique particulier A partir des anneacutees 1950 la communauteacute scientifique a admis la responsabiliteacute
indiscutable de certaines mycobacteacuteries environnementales dans des processus pathologiques
humains et animaux appeleacutes mycobacteacuterioses amorccedilant des eacutetudes les concernant En lrsquoabsence de
contamination inter-humaine deacutemontreacutee il a eacuteteacute admis que lrsquoorigine de ces mycobacteacuteries est
environnementale et que lrsquohomme se contamine agrave partir de lrsquoenvironnement
a Les mycobacteacuteries non pathogegravenes potentiellement opportunistes
Elles regroupent les mycobacteacuteries environnementales qui preacutesentent un pouvoir pathogegravene
pour lrsquohomme et les animaux tregraves variable en fonction des espegraveces (van Ingen et al 2009 Tortoli 2003)
Lrsquoincidence des infections humaines lieacutees agrave des mycobacteacuteries opportunistes semble augmenter dans
les pays industrialiseacutes (Falkinham 2016) peut-ecirctre en lien avec lrsquoameacutelioration des outils pour les isoler
et les identifier Les mycobacteacuterioses qursquoelles engendrent surviennent geacuteneacuteralement sur un terrain
drsquoimmunodeacutepression locale ou geacuteneacuterale Chez lrsquohomme elles sont responsables drsquoun large eacuteventail de
symptocircmes cliniques majoritairement des infections pulmonaires mais eacutegalement des lymphadeacutenites
chez les enfants des infections cutaneacutees (notamment lieacutees agrave M marinum responsable du granulome
des piscines et M ulcerans responsable de lrsquoulcegravere de Buruli) des infections extra-pulmonaires voire
des infections disseacutemineacutees chez les patients seacutevegraverement immunodeacuteprimeacutes (van Ingen 2013 Biet et al
2005 Hoefsloot et al 2013) Environ 25 espegraveces sont retrouveacutees reacuteguliegraverement dans les eacutechantillons
cliniques les autres beaucoup plus rarement (van Ingen 2013) Parmi elles les mycobacteacuteries agrave
croissance lente semblent plus preacutedisposeacutees agrave provoquer des atteintes pulmonaires ou lymphatiques
alors que celles agrave croissance rapide affectent plutocirct la peau les os et les articulations (Tortoli 2003
2009) Les mycobacteacuteries M avium et M intracellulare sont les plus freacutequemment isoleacutees chez
lrsquohomme majoritairement en cas drsquoatteinte pulmonaire (Hoefsloot et al 2013 Whiley et al 2012) La
distribution des espegraveces pathogegravenes isoleacutees chez lrsquohomme diffegravere selon la zone geacuteographique eacutetudieacutee
probablement en lien avec leurs niches eacutecologiques (Hoefsloot et al 2013)
Chez les animaux M avium ssp paratuberculosis (Map) est lrsquoagent de la maladie de Johne ou
paratuberculose chez les ruminants domestiques et sauvages les primates ainsi que drsquoautres espegraveces
(lapins etc) chez qui elle provoque une enteacuterite chronique contagieuse et incurable agrave lrsquoorigine de
pertes eacuteconomiques importantes dans les cheptels infecteacutes Son rocircle est eacutegalement eacutevoqueacute dans la
Chapitre 1
12
maladie de Crohn chez lrsquohomme (Biet et al 2005 Rowe amp Grant 2006 Momotani et al 2012) sans
toutefois que la causaliteacute ait eacuteteacute deacutemontreacutee Mycobacterium avium ssp avium (Maa) est responsable
de la tuberculose aviaire chez les oiseaux et les petits mammifegraveres terrestres chez qui elle provoque
des leacutesions tuberculeuses majoritairement digestives des lymphadeacutenites et des infections disseacutemineacutees
(Biet et al 2005 Pavlik et al 2000) Mycobacterium avium ssp hominissuis (Mah) est responsable de
leacutesions des nœuds lymphatiques et de diffeacuterents organes chez le porc (Pavlik et al 2000)
b Les mycobacteacuteries saprophytes non pathogegravenes
Parmi toutes les mycobacteacuteries environnementales deacutecrites agrave lrsquoheure actuelle certaines sont
retrouveacutees uniquement dans lrsquoenvironnement et nrsquoont jamais eacuteteacute isoleacutees chez lrsquohomme ou lrsquoanimal
comme M arabiense ou M rufum (Tortoli 2014) Ces mycobacteacuteries sont dites saprophytes Parmi
elles certaines ont eacuteteacute isoleacutees dans des sols et des seacutediments fortement pollueacutes par les HAP
(Hydrocarbures Aromatiques Polycycliques) qursquoelles sont potentiellement capables de deacutegrader
(Hennessee et al 2009 Leys et al 2005 Hormisch et al 2004) Elles repreacutesentent donc une voie
inteacuteressante pour la bioremeacutediation de ces sols
c Les mycobacteacuteries pathogegravenes obligatoires
Les mycobacteacuteries responsables de legravepre
Les mycobacteacuteries responsables de la legravepre humaine et murine sont M lepreae M lepromatosis et
M lepreaemurium Elles provoquent des infections dermatologiques et nerveuses chroniques graves
et mutilantes parfois mortelles Mycobacterium leprae touche environ 15 million de personnes dans
le monde et la maladie reste un problegraveme de santeacute publique preacuteoccupant dans les zones endeacutemiques
drsquoAfrique drsquoAsie et drsquoAmeacuterique du sud (Who 2014) Mycobacterium leprae a longtemps eacuteteacute consideacutereacutee
comme un pathogegravene obligatoire de lrsquohomme avec une voie de transmission interhumaine majoritaire
La deacutecouverte de reacuteservoirs animaux (Truman 2005 Meredith et al 2014) et la deacutetection de M leprae
dans le sol la sphaigne ou lrsquoeau de riviegravere (Matsuoka et al 1999 Mostafa et al 1995 Lavania et al
2006) tendent agrave souligner le rocircle potentiel de certains animaux ou de lrsquoenvironnement comme source
de lrsquoinfection chez lrsquohomme Ces 3 espegraveces sont multiplieacutees sur les coussinets de souris (Shepard 1960)
et sur la peau de tatous agrave 9 bandes (Dasypus novemcinctus) (Kirchheimer amp Storrs 1971) car aucun
milieu syntheacutetique ne permet leur croissance optimale
Chapitre 1
13
Les mycobacteacuteries tuberculeuses le Complexe tuberculosis
(MTBC)
Toutes les mycobacteacuteries tuberculeuses sont des pathogegravenes obligatoires Contrairement aux
mycobacteacuteries environnementales leur croissance dans lrsquoenvironnement nrsquoa jamais eacuteteacute deacutemontreacutee Le
complexe tuberculosis comprend agrave ce jour 13 espegraveces qui malgreacute une forte identiteacute geacuteneacutetique
preacutesentent des pheacutenotypes des hocirctes et des pouvoirs pathogegravenes varieacutes
Phylogeacutenie des mycobacteacuteries du MTBC
Lrsquoeacutetude des RD eacutevoqueacutees preacuteceacutedemment et des mutations agrave lrsquoeacutechelle du nucleacuteotide (ou SNP Single
Nucleacuteotide Polymorphism) sur 7 gegravenes (katG gyrA gyrB rpoB mmpL6 oxyR et ADNr 16S) ont permis
de retracer la phylogeacutenie des bacteacuteries du Complexe agrave partir de leur ancecirctre commun (Brosch et al
2002 Rodriguez-Campos et al 2014 Gordon et al 1999 Mahairas et al 1996 Huard et al 2003) les
deacuteleacutetions et mutations jouant un rocircle drsquohorloge moleacuteculaire permettant de replacer lrsquoapparition et
lrsquoeacutevolution drsquoune espegravece dans le temps Le sceacutenario de lrsquoeacutevolution des bacteacuteries du MTBC est
scheacutematiseacute dans la Figure 3 Selon les auteurs lrsquoancecirctre du MTBC appeleacute M protuberculosis aurait
eacutemergeacute il y a entre 15 000 et 40 000 ans en Afrique de lrsquoEst (Gutierrez et al 2005 Wirth et al 2008
Sreevatsan amp Pan 1997) A cocircteacute de M canetti la bacteacuterie du MTBC geacuteneacutetiquement la plus proche de
lrsquoancecirctre deux clades principaux peuvent ecirctre observeacutes Un clade de souches speacutecifiques de lrsquohomme
caracteacuteriseacute par la deacuteleacutetion TbD1 a donneacute les diffeacuterentes ligneacutees modernes de M tuberculosis (Figure
3) Un second clade caracteacuteriseacute par la perte des reacutegions RD7 RD8 RD9 et RD10 regroupe des souches
endeacutemiques drsquoAfrique speacutecifiques de lrsquohomme (M africanum) et drsquoanimaux (le Chimpanzee bacillus
M mungi le Dassie bacillus et M suricattae) ainsi que des souches adapteacutees agrave diffeacuterentes espegraveces de
mammifegraveres (M orygis M microti M pinnipedii M caprae et M bovis)
Chapitre 1
14
Figure 3 Phylogeacutenie des souches du MTBC baseacutee sur les Reacutegions de Diffeacuterences et diffeacuterents Single Nucleotide Polymorphism (Dippenaar et al 2015)
Virulence des mycobacteacuteries du MTBC
Les mycobacteacuteries du MTBC ne possegravedent pas de facteurs de virulence classiques tels que les
toxines par exemple observeacutes chez drsquoautres bacteacuteries pathogegravenes Pour certains auteurs la virulence
des mycobacteacuteries est restreinte agrave lrsquoaptitude du bacille agrave peacuteneacutetrer dans lrsquohocircte agrave y survivre et srsquoy
multiplier (Pollock amp Neill 2002) De faccedilon plus large la notion de virulence chez les membres du MTBC
a eacuteteacute conceptualiseacutee comme lrsquoassociation de i) la capaciteacute de la bacteacuterie agrave faire face agrave la reacuteponse
immunitaire de lrsquohocircte infecteacute ii) la capaciteacute agrave engendrer des leacutesions iii) la capaciteacute agrave survivre en dehors
de son hocircte notamment lors de lrsquoaeacuterosolisation et iiii) la transmission et lrsquoinfection drsquoun nouvel hocircte
(Coscolla amp Gagneux 2014) La virulence implique donc un ensemble de meacutecanismes complexes et
varieacutes faisant intervenir de nombreux gegravenes et proteacuteines permettant notamment au bacille de peacuteneacutetrer
dans les cellules de lrsquohocirctes et drsquoy survivre en bloquant les diffeacuterents meacutecanismes de deacutefense des
macrophages (inhibition de la fusion phagosome-lysosome et de la production de ROS1 et RNS2
deacutesorganisation de lrsquoimmuniteacute de lrsquohocircte inhibition de lrsquoautophagie et de lrsquoapoptose) (Hmama et al
1 ROS Reactive Oxygen Species 2 RNS Reactive Nitrogen Species
Chapitre 1
15
2015) Plus de cent gegravenes seraient impliqueacutes dans la virulence des bacteacuteries du MTBC et lrsquoutilisation
de mutants permet peu agrave peu de connaitre le rocircle de chacun (Forrellad et al 2013) Lrsquoanalyse du
transcriptome (ARN) de M tuberculosis au cours de lrsquoinfection de macrophages et de cellules
dendritiques a reacuteveacuteleacute lrsquoexpression ou la surexpression de nombreux gegravenes codant des proteacuteines
intervenant notamment dans le meacutetabolisme lipidique la synthegravese de lrsquoenveloppe bacteacuterienne
lrsquoapprovisionnement en fer la reacutesistance au stress oxydatif et lrsquoinhibition de lrsquoapoptose (Mukhopadhyay
et al 2012) Les meacutecanismes de survie en dehors de lrsquohocircte sont inconnus agrave ce jour
Il a eacuteteacute montreacute chez M tuberculosis qursquoen fonction des ligneacutees ou des souches la virulence pouvait
varier entrainant une reacuteponse immunitaire reacuteduite ou retardeacutee chez lrsquohocircte une plus grande seacuteveacuteriteacute
de la maladie et une transmission augmenteacutee (Coscolla amp Gagneux 2014) Ce lien souche-virulence est
eacutegalement suspecteacute chez M bovis pour lequel certains geacutenotypes srsquoaccompagnent drsquoun plus grand
nombre drsquoanimaux agrave leacutesions et ou drsquoun score leacutesionnel plus eacuteleveacute (Wright et al 2013 Gavier-Wideacuten et
al 2009 Garbaccio et al 2014) mais des eacutetudes compleacutementaires restent neacutecessaires
Les mycobacteacuteries speacutecifiques de lrsquohomme
Mycobacterium tuberculosis
Mycobacterium tuberculosis est lrsquoagent de la tuberculose humaine maladie retrouveacutee dans
toutes les reacutegions du monde Crsquoest un problegraveme majeur de santeacute publique puisque 2 milliards
drsquoindividus seraient infecteacutes dans le monde En 2013 lrsquoOMS a eacutevalueacute agrave neuf millions le nombre de
nouveaux cas annuels et agrave un million et demi le nombre de deacutecegraves lieacutes agrave la tuberculose (dont 24 de
personnes seacuteropositives) Les reacutegions de lrsquoAsie du sud-est et du Pacifique occidental regroupent 56
des nouveaux cas lrsquoAfrique 25 lrsquoInde 24 et la Chine 11 (WHO 2014) Lrsquoinfection par le HIV est un
facteur preacutedisposant majeur dans la transmission de la maladie et la progression de lrsquoinfection chez
lrsquohomme de par les effets synergiques deacuteleacutetegraveres des 2 micro-organismes sur le systegraveme immunitaire
(Pawlowski et al 2012) Environ 78 des personnes co-infecteacutees vivent en Afrique (WHO 2014) La
tuberculose se manifeste chez lrsquohomme par des granulomes pulmonaires (tuberculose pulmonaire)
mais peut se deacutevelopper dans drsquoautres organes (tuberculose extra-pulmonaire) Elle se transmet
essentiellement par voie respiratoire par lrsquoinhalation de gouttelettes contamineacutees eacutemises par une
personne infecteacutee lors de toux par exemple Le traitement repose sur lrsquoassociation de plusieurs
antibiotiques adapteacutes tels que lrsquoisoniazide la rifampicine le pyrazinamide et lrsquoeacutethambutol
Lrsquoapparition de souches reacutesistantes aux antibiotiques MDR (Multi Drug Resistant) et XDR
(eXtensively Drug Resistant) constitue agrave lrsquoheure actuelle un problegraveme majeur dans la lutte contre la
maladie Bien que lrsquohomme constitue le reacuteservoir majeur de la bacteacuterie M tuberculosis eacuteteacute isoleacutee chez
Chapitre 1
16
un grand nombre drsquoespegraveces animales telles que les bovins domestiques (Romero et al 2011 Prasad et
al 2005 Ameni et al 2011 Pavlik et al 2005 Cadmus et al 2006) les chegravevres (Cadmus Adesokan et
al 2009) les porcs domestiques (Jenkins et al 2011) les chiens (Pavlik et al 2005) les oiseaux et des
animaux de zoos (Michel et al 2003) dont les eacuteleacutephants
Mycobacterium africanum
Elle est principalement retrouveacutee dans les pays drsquoAfrique de lrsquoouest ougrave elle peut ecirctre impliqueacutee
dans la moitieacute des cas de tuberculose humaine agrave cocircteacute de M tuberculosis (de Jong et al 2010) Isoleacutee
et deacutecrite pour la premiegravere fois au Seacuteneacutegal en 1968 elle preacutesente des caracteacuteristiques biochimiques
variables intermeacutediaires entre celles de M tuberculosis et M bovis (Castets amp Sarrat 1968)
Mycobacterium africanum est parfois isoleacutee chez lrsquohomme en dehors de lrsquoAfrique (Schroumlder 1982
Grange amp Yates 1989 Desmond et al 2004 Frottier et al 1990) La proximiteacute phylogeacuteneacutetique de
M africanum avec les autres membres du MTBC infectant des espegraveces animales a souleveacute la question
drsquoun reacuteservoir animal de la bacteacuterie Mais elle a eacuteteacute isoleacutee de faccedilon sporadique sur des singes (Thorel
1980) des bovins (Rahim et al 2007 Cadmus et al 2006) et du lait (Cadmus et al 2010) infirmant
cette hypothegravese
Mycobacterium canetti et smooth tubercle bacilli
Elle est responsable de tuberculose humaine dans une zone geacuteographique confineacutee agrave la corne de
lrsquoAfrique (Djibouti Somalie Kenya) (Pfyffer et al 1998 Miltgen et al 2002) Elle est la plus proche
geacuteneacutetiquement de lrsquoancecirctre des bacteacuteries du MTBC Elle constitue un taxon divergent des autres
membres du MTBC par son pheacutenotype particulier et son geacutenotype qui preacutesente une large diversiteacute
geacuteneacutetique et des signes de transfert horizontal et de recombinaison de gegravenes contrairement aux autres
bacteacuteries du MTBC (Fabre et al 2004 Supply et al 2013 Gutierrez et al 2005) Sa situation
geacuteographique confineacutee la grande diversiteacute geacuteneacutetique des souches rencontreacutees et le manque de
connaissances sur la transmission de la bacteacuterie ont suggeacutereacute lrsquoexistence drsquoun reacuteservoir environnemental
(Fabre et al 2004 Koeck et al 2011) A ce jour cette hypothegravese nrsquoa pas eacuteteacute confirmeacutee La tuberculose
agrave M canetti est consideacutereacutee aujourdrsquohui comme une maladie eacutemergente
Les mycobacteacuteries speacutecifiques des animaux
Mycobacterium bovis
Mycobacterium bovis est lrsquoagent de la tuberculose bovine maladie preacutesente sur tous les
continents Les bovins constituent le reacuteservoir naturel du bacille mais un grand nombre drsquoespegraveces
Chapitre 1
17
animales domestiques et sauvages peuvent ecirctre infecteacutees ainsi que lrsquohomme (Biet et al 2005 Palmer
2013 Grange amp Yates 1994 OrsquoReilly amp Daborn 1995) Sur le plan microbiologique M bovis se
caracteacuterise par une croissance tregraves lente sur milieu de Loumlwenstein-Jensen (6 agrave 10 semaines) par des
colonies petite taille (1 agrave 2 mm) plates lisses laquo dysgoniques raquo et achromogegravenes Sa croissance est
ameacutelioreacutee par lrsquoajout de pyruvate et inhibeacutee par lrsquoajout de glyceacuterol Sur Middlebrook 7H11 les colonies
sont rugueuses et agrave bords eacutechancreacutes (Figure 4) Le pH optimum de culture de M bovis est compris
entre 58 et 69 mais la bacteacuterie survit 20 jours dans du yaourt agrave 4degC (Mitscherlich amp Marth 2012)
Lrsquoidentification pheacutenotypique repose notamment sur lrsquoabsence de production de niacine lrsquoabsence de
nitrate reacuteductase une reacutesistance au pyrazinamide agrave la D-cycloseacuterine (30mgmL-1) et une croissance
favoriseacutee par la preacutesence de pyruvate (Delaune et al 2012)
Figure 4 Aspect des colonies de M bovis sur milieu de Lowenstein-Jensen (agrave gauche) et Middlebrook 7H11 suppleacutementeacute (agrave droite)
Concernant les caracteacuteristiques geacutenomiques de M bovis le seacutequenccedilage complet de la
premiegravere souche (M bovis AF212297 ou ATCC BAA-935 isoleacutee drsquoune leacutesion caseacuteeuse de bovin en
Grande-Bretagne) en 2003 et son annotation ont enrichi les connaissances sur ce bacille Son geacutenome
comprend 4345 Mpb un pourcentage G-C eacuteleveacute de 656 et 3952 gegravenes codant des proteacuteines dont 1
prophage et 42 seacutequences drsquoinsertion Il preacutesente 11 deacuteleacutetions de 1 agrave 127 kb par comparaison avec le
geacutenome de M tuberculosis H37Rv et 1 locus absent chez M tuberculosis appeleacute TbD1 (Garnier et al
2003) Ne posseacutedant pas de gegravenes propres agrave lrsquoespegravece bovis le geacutenome de M bovis a eacuteteacute faccedilonneacute par
lrsquoapparition de ces deacuteleacutetions conditionnant lrsquoexpression des gegravenes Depuis 2003 une quarantaine de
souches isoleacutees sur des animaux domestiques et sauvages ont eacuteteacute entiegraverement seacutequenceacutees (Sites
NCBI et European Nucleotide archive)
La structure de la population de M bovis reacutesulte de lrsquoexpansion de plusieurs complexes clonaux
issus drsquoun mecircme ancecirctre Quatre complexes clonaux majeurs ont eacuteteacute identifieacutes dans le monde pour
Chapitre 1
18
M bovis en se basant sur la preacutesence de deacuteleacutetions chromosomiques caracteacuteristiques dans le geacutenome
de la bacteacuterie etou sur des SNPs ainsi que sur le spoligotype et lrsquoabsence de certains spacers (Smith
2012) le Complexe African 1 est retrouveacute dans plusieurs pays africains (Mali Cameroun Nigeacuteria
Tchad) (Muumlller et al 2009) le Complexe African 2 est essentiellement retrouveacute chez les bovideacutes
drsquoAfrique de lrsquoEst (Berg et al 2011) le Complexe European 1 est retrouveacute en Europe il est dominant au
Royaume-Uni et beaucoup plus rare en Espagne au Portugal et en France On le retrouve eacutegalement
chez les anciens partenaires commerciaux du Royaume-Uni (USA Afrique du Sud Nouvelle Zeacutelande
Australie Canada) et en Ameacuterique du sud La diffusion geacuteographique de ce Complexe est lieacutee aux
eacutechanges commerciaux de bovins (Smith et al 2011) Enfin le Complexe European 2 est majoritaire au
Portugal et en Espagne et assez freacutequent en France et en Italie (Rodriguez-Campos et al 2012) Drsquoautres
complexes clonaux minoritaires existent En France plusieurs complexes clonaux coexistent certains
autochtones eacutevoluent depuis longtemps sur le territoire alors que drsquoautres ont eacuteteacute importeacutes agrave la faveur
des eacutechanges commerciaux de bovins (Hauer et al 2015)
Une souche atteacutenueacutee obtenue agrave partir drsquoun isolat virulent de M bovis a eacuteteacute creacuteeacutee par Calmette
et Gueacuterin apregraves 230 repiquages sur un milieu bilieacute enrichi en glyceacuterol entre 1908 et 1921 le BCG pour
Bacille de Calmette et Gueacuterin Testeacutee sur des modegraveles animaux cette souche atteacutenueacutee immunisait
contre une infection expeacuterimentale agrave M bovis et M tuberculosis trente jours apregraves injection Utiliseacutee
pour la vaccination elle a eacuteteacute distribueacutee dans le monde entier et maintes fois repiqueacutee ce qui a meneacute
agrave une eacutevolution de la souche initiale en plusieurs souches tregraves heacuteteacuterogegravenes La comparaison geacutenomique
de la souche BCG avec M tuberculosis reacutevegravele la perte de 14 reacutegions (RD1 agrave RD14) chez BCG Lrsquoabsence
de la reacutegion RD1 expliquerait en partie lrsquoatteacutenuation du BCG (Pym et al 2002) toutefois comme la
reacuteintroduction de RD1 dans une souche mutante ne suffit pas agrave restaurer totalement la virulence
drsquoautres reacutegions sont impliqueacutees dans lrsquoatteacutenuation (Brosch et al 2007)
Mycobacterium pinnipedii
Deacutecouverte en 1986 cette mycobacteacuterie touche principalement les diffeacuterentes espegraveces de
phoques et les lions de mer sauvages de lrsquoheacutemisphegravere sud ou en captiviteacute Elle a eacuteteacute isoleacutee en Australie
Nouvelle-Zeacutelande Uruguay Argentine et Grande-Bretagne (Cousins et al 2003) Des infections chez
lrsquohomme ont eacuteteacute rapporteacutees principalement chez les soigneurs en contact avec les animaux malades
en captiviteacute (Kiers et al 2008) La transmission indirecte du bacille par inhalation drsquoaeacuterosols lors du
nettoyage des enclos est fortement suspecteacutee
Mycobacterium caprae
Cette mycobacteacuterie du MTBC a eacuteteacute isoleacutee pour la premiegravere fois sur des chegravevres en Espagne
(Aranaz et al 1999 Aranaz et al 2003) Crsquoest lrsquoun des agents de la tuberculose bovine avec M bovis
Chapitre 1
19
et elle induit des leacutesions identiques agrave la tuberculose agrave M bovis chez les animaux infecteacutes La
tuberculose agrave M caprae touche principalement lrsquoEurope ougrave elle a deacutejagrave eacuteteacute deacutetecteacutee en Espagne
(Rodriacuteguez et al 2011) en France (Haddad et al 2001) en Autriche (Prodinger et al 2002) en
Allemagne (Kubica et al 2003) et dans plusieurs pays drsquoEurope centrale (Pavlik et al 2005 Beširović
et al 2012 Cvetnic et al 2007) Le nombre de cas deacutetecteacutes tend agrave augmenter chez les bovins et les
animaux sauvages notamment en Espagne en Allemagne en Autriche (Rodriacuteguez et al 2011 Schoepf
et al 2012) En Europe centrale elle repreacutesente lrsquoagent principal de la tuberculose bovine notamment
en Reacutepublique Tchegraveque Slovaquie Bosnie Herzeacutegovine et Croatie ougrave elle circule entre les bovins et de
nombreuses espegraveces sauvages (Prodinger et al 2005 Beširović et al 2012) Elle a eacutegalement eacuteteacute isoleacutee
pour la premiegravere fois en Chine en 2013 (Zeng et al 2013) Crsquoest eacutegalement un agent zoonotique
important puisque des infections agrave M caprae sont rapporteacutees chez lrsquohomme dans diffeacuterents pays
drsquoEurope ((Kubica et al 2003 Prodinger et al 2014 Prodinger et al 2002 Muumlller et al 2013)
Mycobacterium microti
Cette mycobacteacuterie est lrsquoagent responsable de la tuberculose des petits rongeurs (campagnols
souris musaraignes hellip) chez qui elle a eacuteteacute isoleacutee dans les anneacutees 30 (Wells 1937 Wells amp Robb-Smith
1946) Elle a eacuteteacute isoleacutee chez plusieurs espegraveces de mammifegraveres dont des chats (Gunn-Moore et al 1996
Gunn-Moore et al 2011) des bovins (Jahans et al 2004) des porcs (Taylor et al 2006) des lamas des
blaireaux (Smith et al 2009) des sangliers (Dondo et al 2007) des suricates (Palgrave et al 2012) En
France la deacutetection de M microti chez des animaux domestiques et sauvages a suggeacutereacute que la bacteacuterie
eacutetait endeacutemique dans certaines reacutegions de France (Michelet et al 2015 Laprie et al 2013) Chez
lrsquohomme une trentaine de cas drsquoinfections agrave M microti principalement agrave localisation pulmonaire ont
eacuteteacute reporteacutes dans la litteacuterature entre 2000 et 2010 (Panteix et al 2010) Bien que les personnes
immunodeacuteprimeacutees soient les plus toucheacutees lrsquoinfection se rencontre eacutegalement chez des sujets
immunocompeacutetents (Niemann et al 2000 Frank et al 2009)
Mycobacterium orygis
Isoleacutee pour la premiegravere fois en 1976 sur des oryxes en Afrique de lrsquoest (Lomme et al 1976) elle
a eacuteteacute isoleacutee dans de rares occasions sur drsquoautres espegraveces de ruminants tels que les gazelles les
antilopes (Van Soolingen et al 1994) les buffles africains (van Pittius et al 2012) en Afrique du sud
dans la Peacuteninsule arabe et dans des zoos des Pays Bas et du Royaume Uni
Des infections agrave Mycobacterium orygis ont eacuteteacute rapporteacutees chez une dizaine de patients humains
(Dawson et al 2012 van Ingen et al 2012)
Chapitre 1
20
Autres mycobacteacuteries agrave reacuteservoir animal
Quatre autres espegraveces de mycobacteacuteries animales phylogeacuteneacutetiquement proches de
M africanum West African 2 ont eacuteteacute isoleacutees sur des animaux en Afrique le Chimpazee bacillus isoleacute
sur un chimpanzeacute de Cocircte drsquoIvoire (Coscolla et al 2013) M mungi isoleacutee sur des mangoustes rayeacutees
(Mungos mungo) du Bostwana (Alexander et al 2010) le Dassie bacillus isoleacute sur un daman du Cap
(Procavia capensis) (Wagner et al 1958) et M suricattae isoleacutee sur des suricates (Suricata suricatta) en
Afrique du sud (Parsons et al 2013) Ces espegraveces nrsquoont agrave ce jour jamais eacuteteacute isoleacutees chez lrsquohomme La
deacutecouverte reacutecente de ces espegraveces rattacheacutees au MTBC suggeacutere que ce Complexe est probablement
plus varieacute que ce qui eacutetait imagineacute
B LA TUBERCULOSE UNE MALADIE TRES ANCIENNE MAIS UNE MENACE TOUJOURS ACTUELLE
1 Introduction
La tuberculose est une maladie bacteacuterienne chronique et contagieuse qui affecte lrsquohomme
ainsi que de nombreux animaux domestiques et sauvages Elle est provoqueacutee par les diffeacuterentes
espegraveces de mycobacteacuteries du MTBC Lrsquoobservation de tubercules granulomes inflammatoires formeacutes
de cellules immunitaires et de caseacuteum (zones neacutecrotiques) dans les organes ou les nœuds
lymphatiques des personnes infecteacutees a donneacute son nom agrave la maladie
2 Histoire de la tuberculose
La tuberculose est une maladie tregraves ancienne puisque lrsquoancecirctre du Complexe tuberculosis serait
apparu il y a 40 000 ans (Wirth et al 2008) La preacutesence exclusive de certaines ligneacutees du MTBC sur le
continent africain suggegravere que le Complexe aurait eacutemergeacute en Afrique de lrsquoEst puis aurait co-eacutevolueacute
avec lrsquohomme et se serait reacutepandu dans le monde entier par le biais des migrations humaines en
Europe en Asie et en Afrique continentale Les populations de mycobacteacuteries tuberculeuses eacutetant
infeacuteodeacutees agrave une zone geacuteographique elles se sont adapteacutees agrave la population locale (Gutierrez et al 2005
Djelouadji et al 2011 Wirth et al 2008) Le passage drsquoun mode de vie nomade agrave une vie seacutedentaire
dans des fermes agricoles de plus en plus peupleacutees ainsi que la domestication des animaux ont
probablement contribueacute agrave la transmission de la maladie agrave divers hocirctes animaux et agrave lrsquoadaptation du
bacille permettant lrsquoapparition de M bovis M caprae et des autres membres du Complexe (Wirth et
al 2008) Les plus anciennes traces drsquoADN du Complexe tuberculosis ont eacuteteacute deacutecouvertes sur des
Chapitre 1
21
ossements de bison vieux de 17 000 ans retrouveacutes dans une grotte drsquoAmeacuterique du Nord (Rothschild et
al 2001) Des traces drsquoADN du Complexe tuberculosis de M tuberculosis ou M bovis ont eacuteteacute
retrouveacutees sur des ossements humains datant de la Preacutehistoire ou de lrsquoAntiquiteacute (les plus vieux ont
9000 ans) deacutecouverts dans diffeacuterentes parties du monde (Chili Royaume Uni Russie Israeumll Indoneacutesie
etc) (Djelouadji et al 2011) Appeleacutee laquo phtisie raquo par les Grecs peste blanche ou consomption (suite au
deacutepeacuterissement des malades) les eacutecrits drsquoHippocrate (5egraveme siegravecle avant JC) sont les premiers agrave deacutecrire la
symptomatologie et les diffeacuterentes formes de la tuberculose Fracastor (15egraveme siegravecle) suggegravere le
caractegravere heacutereacuteditaire de la maladie mais place la phtisie dans les maladies contagieuses au mecircme titre
que la veacuterole hypothegravese reprise en 1722 par Marten puis par Villemin en 1865 qui montre que la
maladie est infectieuse contagieuse et inoculable (Dutau 2005) A la fin du 18egraveme siegravecle et au deacutebut du
19egraveme la tuberculose est la premiegravere cause de mortaliteacute en Europe de lrsquoOuest et en Ameacuterique du Nord
La Reacutevolution industrielle et le deacuteveloppement des villes favorisent lrsquoexpansion de la maladie les
conditions de vie insalubres des taudis de banlieues la promiscuiteacute et la pauvreteacute acceacutelegraverent lrsquoendeacutemie
et agrave cette eacutepoque environ une personne sur 4 est atteinte de tuberculose La fin du 19egraveme siegravecle voit se
succeacuteder les deacutecouvertes agrave un rythme effreacuteneacute dont les principales sont la deacutecouverte du bacille en
1882 par Koch la deacutecouverte du bacille bovin en 1896 lrsquoutilisation de la tuberculine dans le diagnostic
humain en 1907 la creacuteation drsquoune souche atteacutenueacutee M bovis BCG en 1921 et le deacutebut de la vaccination
en 1924 La deacutecouverte de la streptomycine en 1944 le premier antibiotique contre la tuberculose
puis de lrsquoisoniazide en 1952 et la rifamycine en 1957 a marqueacute une eacutetape importante dans le traitement
contre la tuberculose (Daniel 2006)
3 La tuberculose bovine
a Introduction
La tuberculose bovine est une maladie bacteacuterienne contagieuse drsquoeacutevolution chronique causeacutee
par Mycobacterium bovis Cette bacteacuterie a pour hocircte originel les bovins mais peut infecter un grand
nombre drsquoespegraveces de mammifegraveres dont lrsquohomme (Gavier-Wideacuten et al 2009 Broughan Crawshaw et
al 2013 Grange amp Yates 1994) ce qui en fait une maladie zoonotique Elle a un tropisme respiratoire
preacutedominant mais peut infecter un grand nombre drsquoautres organes La tuberculose bovine fait partie
des maladies animales agrave deacuteclaration obligatoire aupreacutes de lrsquoOffice International des Epizooties (OIE)
Le passage de lrsquoinfection dans la faune sauvage a compliqueacute le scheacutema drsquoeacuteradication de la
maladie chez les bovins puisque certaines espegraveces sauvages peuvent entretenir la maladie et
contaminer les bovins en retour A lrsquoheure actuelle M bovis fait partie des six organismes pathogegravenes
prioritaires cibleacutes par les programmes de surveillance europeacuteens (Ciliberti et al 2015)
Chapitre 1
22
b Les hocirctes et les communauteacutes drsquohocirctes de Mycobacterium bovis dans le monde
et leurs statuts eacutepideacutemiologiques
Mycobacterium bovis est la mycobacteacuterie du MTBC qui possegravede le plus large spectre drsquohocirctes
incluant lrsquohomme (Grange 2001 OrsquoReilly amp Daborn 1995) Lrsquoinfection a eacuteteacute retrouveacutee chez de
nombreuses espegraveces de mammifegraveres appartenant aux Ordres des marsupiaux des carnivores des
primates des rongeurs des lagomorphes et des artiodactyles (Broughan Downs et al 2013 OrsquoReilly
amp Daborn 1995 Palmer 2013)
La circulation de lrsquoinfection chez des hocirctes appartenant agrave des espegraveces domestiques et sauvages
diffeacuterentes est agrave lrsquoorigine de systegravemes eacutepideacutemiologiques complexes appeleacutes systegraveme multi-hocirctes
(Haydon et al 2002) qui complexifient fortement les scheacutemas drsquoeacuteradication de la maladie Dans ces
systegravemes les diffeacuterents hocirctes peuvent avoir un statut eacutepideacutemiologique diffeacuterent (Haydon et al 2002
Palmer 2013) ou former une communauteacute drsquohocirctes reacuteservoir (Palmer 2013) Les hocirctes de maintenance
ou reacuteservoirs deacutefinissent une population drsquoanimaux dans laquelle lrsquoinfection est entretenue de faccedilon
autonome Ils sont agrave lrsquoorigine de lrsquoinfection des espegraveces cibles Les hocirctes de liaison sont incapables de
maintenir lrsquoinfection indeacutefiniment au sein de leur population agrave moins qursquoils soient reacuteinfecteacutes par une
autre espegravece Ils sont toutefois capables de transmettre lrsquoinfection agrave drsquoautres populations (Corner 2006
Nugent 2011) Et enfin les hocirctes laquo cul-de-sac raquo deacutefinissent les espegraveces qui ne peuvent pas maintenir
ni transmettre la maladie (Nugent 2011) Diffeacuterentes espegraveces ont ainsi eacuteteacute identifieacutees comme des
reacuteservoirs de lrsquoinfection notamment le blaireau (Meles meles) en Angleterre et en Irlande (Delahay et
al 2001) le sanglier (Sus scrofa) dans le centre et le sud de lrsquoEspagne (Naranjo et al 2008) le cerf
eacutelaphe en France (Zanella et al 2008) le cerf de Virginie (Odocoileus virginianus) dans le Michigan
(USA) (Palmer 2013 OrsquoBrien et al 2006) le bison des bois (Bison bison athabascae) et le wapiti (Cervus
elaphus manitobensis) au Canada (Nishi et al 2006 Wobeser 2009) le phalanger renard (Trichosurus
vulpecula) en Nouvelle-Zeacutelande (Morris et al 1994 Nugent 2011) et le buffle (Syncerus caffer) en
Afrique du Sud (Renwick et al 2007) Drsquoautres espegraveces animales ont eacuteteacute identifieacutees comme hocirctes de
liaison comme le furet le cerf et le sanglier en Nouvelle-Zeacutelande
La transmission retour de lrsquoinfection par les espegraveces sauvages reacuteservoir est une menace pour
les bovins et les espegraveces sauvages sensibles vivant agrave proximiteacute La connaissance du statut
eacutepideacutemiologique de chaque espegravece est important pour comprendre le rocircle de la faune sauvage dans la
maintenance et la transmission de lrsquoinfection et adapter les programmes de lutte et drsquoeacuteradication de la
maladie chez les bovins les animaux sauvages et lrsquohomme (Delahay et al 2001 Corner 2006)
Chapitre 1
23
c Aspect zoonotique de Mycobacterium bovis
Plusieurs mycobacteacuteries du MTBC peuvent engendrer une tuberculose zoonotique chez
lrsquohomme Le principal agent infectieux impliqueacute est M bovis (Thoen et al 2006) dans une moindre
mesure M caprae (EFSA 2014) et M microti (Panteix et al 2010) Cette derniegravere a un pouvoir
zoonotique plus faible mais nrsquoest pas neacutegligeable La tuberculose zoonotique agrave M bovis est
majoritairement lieacutee agrave la consommation de produits issus drsquoanimaux infecteacutes principalement le lait cru
et ses deacuteriveacutes (transmission indirecte) (Evans et al 2007) La transmission par aeacuterosols respiratoires est
eacutegalement deacutecrite lors de contacts rapprocheacutes avec des animaux domestiques ou sauvages infecteacutes
(transmission directe) (Cosivi et al 1999 Ayele et al 2004 Kleeberg 1984) Lrsquoinfection est
sporadiquement rapporteacutee chez des eacuteleveurs de bovins et de cerfs (Smith et al 2004 Fanning amp
Edwards 1991) du personnel veacuteteacuterinaire (Shrikrishna et al 2009) et du personnel drsquoabattoir (Liss et al
1994) La transmission interhumaine de M bovis par voie respiratoire existe chez les patients
immunodeacuteprimeacutes ou immunocompeacutetents mais reste rare (Evans et al 2007 Grange amp Yates 1994 de
la Rua-Domenech 2006 Sunder et al 2009 Shrikrishna et al 2009 Rivero et al 2001)
Les signes cliniques chez lrsquohomme sont identiques agrave ceux retrouveacutes lors de tuberculose agrave
M tuberculosis et se traduisent par une tuberculose pulmonaire etou des atteintes extra-pulmonaires
plus freacutequentes qursquoavec M tuberculosis en raison des diffeacuterences dans le mode de transmission
(atteintes osteacuteo-articulaires ganglionnaires uro-geacutenitales disseacutemineacuteeshellip)(Antoine amp Jarlier 2010
Delaune et al 2012 Mignard et al 2006) Le traitement repose sur lrsquoadministration drsquoantituberculeux
classiques agrave lrsquoexception du pyrazinamide pour lequel M bovis est naturellement reacutesistant Des souches
MDR (Multi Drug Resistant) sont reacuteguliegraverement signaleacutees chez lrsquohomme (McLaughlin et al 2012
Cambau et al 2003) De rares souches XDR (Extensively Drug Resistant) sont eacutegalement deacutecrites
majoritairement chez des patients seacuteropositifs (Rivero et al 2001) Dans les pays industrialiseacutes
lrsquoinfection est principalement diagnostiqueacutee chez des personnes acircgeacutees etou immunodeacuteprimeacutees
(diabegravete VIH traitement immunosuppresseur alcoolismehellip) dont les formes extra-pulmonaires
majoritaires eacutevoquent une contamination ancienne par voie digestive (avant la mise en place de la
pasteurisation du lait) et la reacuteactivation ulteacuterieure de la maladie ainsi que chez des personnes neacutees ou
ayant seacutejourneacute dans des pays ougrave la maladie est endeacutemique (Delaune et al 2012)
Les pays ayant mis en place la pasteurisation du lait et des programmes de lutte efficaces contre
la maladie chez les bovins (deacutepistage et abattage) ont fortement reacuteduit lrsquoincidence de la maladie chez
lrsquohomme et les bovins Malgreacute quelques cas reacuteguliegraverement deacutecrits chez lrsquohomme la tuberculose
zoonotique est une zoonose mineure dans les pays deacuteveloppeacutes notamment en Europe (Torgerson amp
Torgerson 2010 Michel et al 2010) En France lrsquoobligation de pasteurisation du lait en 1955 associeacutee
agrave lrsquoabattage systeacutematique des bovins reacuteagissant agrave la tuberculination agrave partir de 1963 ont permis de faire
Chapitre 1
24
baisser lrsquoincidence de lrsquoinfection agrave M bovis chez lrsquohomme de 15 dans les anneacutees 60 agrave 05 en 1995
(Boulahbal et al 1998) Des eacutetudes locales mettent parfois en eacutevidence des preacutevalences plus eacuteleveacutees
de lrsquoordre de 2 agrave Lyon entre 2000 et 2005 (Mignard et al 2006) et de 29 en Aquitaine entre 2004
et 2008 (Aimeacute et al 2012) Au Royaume-Uni par exemple malgreacute lrsquoincidence eacuteleveacutee chez les bovins
(1806 nouveaux foyers entre janvier et avril 2015)(site DEFRA) la preacutevalence de la maladie nrsquoaugmente
pas chez lrsquohomme gracircce aux programmes de lutte mis en place chez lrsquoanimal (Torgerson amp Torgerson
2010 Jalava et al 2007)
La situation est tregraves diffeacuterente dans les pays en voie de deacuteveloppement (PVD) Lrsquoabsence de
programme de lutte contre la tuberculose bovine et la consommation de lait cru dans des pays ougrave la
maladie est endeacutemique chez les bovins notamment dans certains pays drsquoAfrique drsquoAmeacuterique du sud
et drsquoAsie repreacutesentent un risque de santeacute publique important (Cosivi et al 1999 Ayele et al 2004) La
tuberculose zoonotique est souvent fortement relieacutee agrave la preacutevalence de lrsquoinfection bovine dans le pays
ou la reacutegion (Fetene et al 2011) et touche majoritairement les professions en contact avec les bovins
(eacuteleveurs de bovins personnel drsquoabattoir et drsquousines agro-alimentairehellip) et les consommateurs de lait
cru et de fromage frais dont les enfants drsquoautant plus srsquoils sont immunodeacuteprimeacutes Une eacutetude reacutecente
rapporte que la tuberculose zoonotique repreacutesente environ 28 des cas de tuberculose en Afrique
(de 0 agrave 377 selon les pays) 03 des cas sur le continent ameacutericain (de 0 agrave 339 ) et 04 des cas
en Europe (de 0 agrave 211 ) (Muumlller et al 2013) Lrsquoinfection par le HIV la pauvreteacute et le manque drsquoaccegraves
aux soins sont des facteurs de disseacutemination de la tuberculose humaine et zoonotique dans ces pays
(Ayele et al 2004 WHO 2014) Dans les PVD le diagnostic chez lrsquohomme srsquoarrecircte parfois agrave lrsquoobservation
microscopique de BAAR associeacutes agrave M tuberculosis sous-estimant probablement lrsquooccurrence de
M bovis (Cosivi et al 1999 Michel et al 2010) La transmission retour de lrsquoinfection de lrsquohomme aux
bovins pourrait avoir un impact non neacutegligeable dans les PVD mais aucune donneacutee nrsquoest disponible
(de la Rua-Domenech 2006)
d Pathogeacutenie de la tuberculose bovine
La pathogeacutenie de la maladie est multifactorielle et deacutepend de facteurs lieacutes au pouvoir pathogegravene de
M bovis agrave la reacuteceptiviteacute et la sensibiliteacute de lrsquohocircte exposeacute ainsi qursquoagrave la voie drsquoinfection
Facteurs influenccedilant le pouvoir pathogegravene de M bovis voies
drsquoinfection et dose infectante
La voie respiratoire est la voie de contamination majeure chez les bovins (Pritchard 1988
Morris et al 1994 Phillips et al 2003) Des infections expeacuterimentales de M bovis par voie nasale intra
Chapitre 1
25
tracheacuteale ou intra-tonsillaire ainsi que par la mise en contact drsquoanimaux sains avec des animaux
infecteacutes ont confirmeacute lrsquoefficaciteacute de la transmission de M bovis par cette voie (Palmer et al 1999 Dean
et al 2005 Johnson et al 2007 Collins amp Grange 1983 Cassidy et al 1999) La dose infectante par voie
respiratoire est tregraves faible puisque lrsquoinoculation intra-tracheacuteale de 6 agrave 10 bacilles viables agrave des veaux
suffit agrave provoquer des leacutesions pulmonaires etou ganglionnaires (Dean et al 2005 Johnson et al 2007)
Des reacutesultats identiques ont eacuteteacute obtenus chez le blaireau (Corner et al 2007 Corner et al 2011) Chez
le cerf de Virginie lrsquoinoculation intra-tonsillaire de 300 agrave 500 UFC (Uniteacute Formant Colonie) infecte 100
des animaux (Palmer et al 2002 Griffin et al 2006) Lrsquoinoculation oro-pharyngeacutee de 100 UFC agrave des
sangliers deacuteclenche uniquement des leacutesions amygdaliennes alors que 10 000 UFC entrainent des
leacutesions pulmonaires et ganglionnaires seacutevegraveres (Ballesteros et al 2009) Chez toutes ces espegraveces la dose
infectante affecte la seacuteveacuteriteacute et la progression de la maladie (Neill et al 1994 Palmer et al 2002 Corner
et al 2007)
La transmission de la tuberculose bovine peut eacutegalement se faire par voie digestive suite agrave
lrsquoingestion de matrices contamineacutees La dose infectante neacutecessaire serait bien supeacuterieure agrave celle par
voie respiratoire et selon certains auteurs neacutecessiterait plusieurs millions de bacilles (Phillips et al
2003 OrsquoReilly amp Daborn 1995) Pour les veaux Palmer et al (2004) ont estimeacute agrave 5 000 UFC la dose
infectante par cette voie Lrsquoingestion reacutepeacutetitive de faibles doses de M bovis semble eacutegalement
repreacutesenter un facteur de risque comme cela a eacuteteacute deacutemontreacute chez le cochon drsquoInde (Maddock 1934)
La transmission par voie cutaneacutee ou transcutaneacutee ainsi que la transmission de la megravere au veau par
consommation de lait contamineacute ou par leacutechage sont eacutegalement deacutecrites mais restent rares (Phillips et
al 2003 Morris et al 1994) La transmission du bacille par voie transcutaneacutee (morsure) par le biais de
salive infecteacutee est reacuteguliegraverement deacutecrite chez le blaireau en Angleterre (Corner et al 2011)
Facteurs influenccedilant la reacuteceptiviteacute et la sensibiliteacute de lrsquohocircte
De nombreux facteurs intrinsegraveques agrave lrsquoanimal entrent en jeu dans la reacuteceptiviteacute et la sensibiliteacute
de lrsquohocircte agrave la maladie notamment lrsquoespegravece la race lrsquoacircge le sexe et lrsquoeacutetat geacuteneacuteral (Griffin et al 1996
Menzies amp Neill 2000 Humblet et al 2009 Goodchild amp Clifton-Hadley 2001) Certains animaux
preacutesentent une reacutesistance naturelle ou au contraire une sensibiliteacute exacerbeacutee agrave lrsquoinfection ou agrave la
progression de la maladie (Gavier-Wideacuten et al 2009) Des meacutecanismes modulant la reacuteponse
immunitaire et inflammatoire de lrsquohocircte ont pu ecirctre mis en eacutevidence chez le cerf (Mackintosh et al
2000) Les caracteacuteristiques geacuteneacutetiques de lrsquohocircte pourraient eacutegalement jouer un rocircle important dans la
reacutesistance ou la sensibiliteacute agrave lrsquoinfection (Mackintosh et al 2000 Acevedo-Whitehouse et al 2005
Brotherstone et al 2010) Des eacutetudes reacutecentes sur des geacutenomes complets de bovins ont identifieacute des
Chapitre 1
26
loci particuliers impliqueacutes dans la reacutesistance agrave la maladie (Bermingham et al 2014) ouvrant des
perspectives de seacutelection geacutenomique de populations reacutesistantes agrave la tuberculose bovine (Tsairidou et
al 2014)
Le statut immunitaire de lrsquoanimal est un facteur important puisque certains animaux sont
capables drsquoeacuteliminer le bacille gracircce agrave leur immuniteacute inneacutee etou acquise certains sont capables de
controcircler lrsquoinfection plusieurs anneacutees dans les granulomes et drsquoautres deacuteveloppent une tuberculose
eacutevolutive (Pollock amp Neill 2002) La modulation immunitaire expliquerait pourquoi dans un cheptel
bovin malgreacute la contagiositeacute importante de M bovis seuls quelques animaux sont infecteacutes (Cassidy
2006) Les notions de latence et de reacuteactivation de la maladie deacutecrites chez M tuberculosis sont
discuteacutees dans la tuberculose bovine (Cassidy 2006) et commencent agrave ecirctre appreacutehendeacutees (Aacutelvarez et
al 2009)
Les eacutetapes de lrsquoinfection et la reacuteponse immunitaire associeacutee
Lorsque les bacteacuteries peacutenegravetrent dans lrsquoorganisme le plus souvent au niveau des alveacuteoles
pulmonaires elles sont phagocyteacutees par les macrophages Si la dose infectante est importante ou
lrsquoimmuniteacute insuffisante les mycobacteacuteries eacutechappent agrave la digestion et se multiplient permettant la
formation dans le tissu pulmonaire de la leacutesion primaire appeleacutee chancre drsquoinoculation La primo-
infection est caracteacuteriseacutee par le laquo complexe primaire raquo qui associe le chancre drsquoinoculation et les nœuds
lymphatiques infecteacutes par le drainage de la leacutesion primaire (Neill et al 1994) Lrsquoinfection peut ensuite
eacutevoluer selon 3 modes diffeacuterents la gueacuterison avec une eacutelimination des bacilles la stabilisation
caracteacuteriseacutee par une reacuteactivation possible de lrsquoinfection quelques mois ou anneacutees plus tard ou la
geacuteneacuteralisation preacutecoce de lrsquoinfection lieacutee agrave une multiplication active des bacteacuteries et agrave leur
disseacutemination dans lrsquoorganisme Lrsquoinfection et son eacutevolution sont toutefois fortement deacutependantes de
la dose infectante de la voie drsquoentreacutee de la bacteacuterie de la reacuteceptiviteacute et de la sensibiliteacute de lrsquohocircte
eacutevoqueacutees preacuteceacutedemment
Chez lrsquoanimal infecteacute par M bovis la reacuteponse immunitaire agrave meacutediation cellulaire est la premiegravere
agrave se mettre en place dans les stades preacutecoces de lrsquoinfection par la mobilisation de macrophages et de
lymphocytes T (Figure 5) Ces derniers libegraverent diffeacuterentes cytokines dont lrsquointerfeacuteron-γ (IFN-γ) qui
active les macrophages et la formation de granulomes pour isoler et deacutetruire lrsquoagent infectieux
Lrsquoimmuniteacute agrave meacutediation humorale se met en place tardivement et remplace peu agrave peu lrsquoimmuniteacute agrave
meacutediation cellulaire dans les stades plus avanceacutes de la maladie par la production drsquoanticorps par les
lymphocytes B (Pritchard 1988 Ritacco et al 1991 Fifis et al 1994) Les diffeacuterents tests utiliseacutes pour
le deacutepistage de lrsquoinfection chez les bovins reposent principalement sur lrsquoimmuniteacute agrave meacutediation
cellulaire (Figure 5)
Chapitre 1
27
Figure 5 Evolution de la reacuteponse immunitaire chez le bovin infecteacute par M bovis au cours du temps et aux diffeacuterents tests de deacutepistage de la tuberculose bovine (de la Rua-Domenech et al 2006 adapteacute de Vordermeier et al 2004)
Expression clinique de la maladie et leacutesions observeacutees
La tuberculose bovine est une maladie chronique drsquoeacutevolution lente geacuteneacuteralement
asymptomatique dans ses deacutebuts Lorsque la maladie eacutevolue hormis lrsquoamaigrissement possible les
signes cliniques sont tregraves variables et deacutependent des organes ou tissus atteints mais ne sont pas
pathognomoniques (de la Rua-Domenech et al 2006) La localisation des leacutesions observeacutees chez les
animaux deacutepend geacuteneacuteralement de la voie drsquoentreacutee de la bacteacuterie et de lrsquoeacutevolution de la maladie Chez
les bovins la majoriteacute des leacutesions sont retrouveacutees dans la caviteacute thoracique avec une atteinte des lobes
pulmonaires et des nœuds lymphatiques associeacutes (bronchiques et meacutediastinaux) etou des nœuds
lymphatiques ceacutephaliques (reacutetro-pharyngiens principalement) Les leacutesions sur les nœuds lymphatiques
meacutesenteacuteriques la rate et le foie sont plus rares (Neill et al 1994)
Chez le blaireau infecteacute naturellement la seacuteveacuteriteacute de la maladie est tregraves variable drsquoune
infection latente asymptomatique sans leacutesion macroscopique visible agrave une maladie modeacutereacutee avec des
leacutesions pulmonaires ou extra-pulmonaires de petite taille jusqursquoagrave une maladie geacuteneacuteraliseacutee entrainant
la cachexie puis la mort de lrsquoanimal (Murphy et al 2010) Les leacutesions retrouveacutees sont essentiellement
respiratoires probablement lieacutees agrave lrsquoinhalation drsquoaeacuterosols contamineacutes plus rarement digestives et
cutaneacutees suite agrave des morsures de congeacutenegraveres infecteacutes (Corner et al 2011 Payne et al 2012 Gavier-
Widen et al 2001) Toutefois diffeacuterentes eacutetudes ont montreacute que la majoriteacute des blaireaux infecteacutes ne
Chapitre 1
28
preacutesente pas de leacutesion visible (Gallagher amp Clifton-Hadley 2000 Jenkins et al 2008 Payne et al 2012)
Les blaireaux constituent des hocirctes de persitance efficaces puisqursquoils peuvent supporter lrsquoinfection
plusieurs anneacutees et se reproduire tout en excreacutetant la bacteacuterie (Delahay et al 2001)
Chez le sanglier les leacutesions sont majoritairement observeacutees dans les nœuds lymphatiques
ceacutephaliques notamment les mandibulaires dans les poumons et moins freacutequemment au niveau
meacutesenteacuterique (Zanella et al 2008 Martiacuten-Hernando et al 2007 Naranjo et al 2008) Des formes
geacuteneacuteraliseacutees sont eacutegalement deacutecrites (Naranjo et al 2008)
Le cerf est une espegravece deacutecrite comme tregraves sensible agrave lrsquoinfection par M bovis et deacuteveloppe des
leacutesions ouvertes et disseacutemineacutees pouvant ecirctre agrave lrsquoorigine drsquoune excreacutetion bacillaire importante (Lugton
et al 1998 Palmer et al 2004) Les leacutesions sont geacuteneacuteralement localiseacutees au niveau des nœuds
lymphatiques ceacutephaliques et des poumons (Palmer et al 2004 Delahay et al 2002) des nœuds
lymphatiques meacutesenteacuteriques voire geacuteneacuteraliseacutees (Zanella et al 2008)
e Le deacutepistage et le diagnostic de lrsquoinfection chez les animaux
Le deacutepistage de lrsquoinfection chez les bovins vivants repose sur plusieurs modes de surveillance
(Cavalerie et al 2014)
La surveillance systeacutematique agrave lrsquoabattoir concerne tous les bovins abattus pour la consommation
humaine Il repose sur un examen post mortem de lrsquoanimal avec lrsquoinspection drsquoun certain nombre
drsquoorganes cibles de lrsquoinfection (poumons nœuds lymphatiques reacutetro-pharyngiens tracheacuteo-
bronchiques et meacutediastinaux) Dans le cas de leacutesions suspectes visibles celles-ci sont preacuteleveacutees et
transmises agrave un laboratoire agreacuteeacute pour recherche de mycobacteacuteries par PCR et culture
La surveillance programmeacutee en eacutelevage repose sur le deacutepistage de la maladie par prophylaxie Le
rythme des prophylaxies est fixeacute par le preacutefet en fonction de la situation sanitaire du deacutepartement
Dans les deacutepartements ougrave cette surveillance est absente le deacutepistage deacutepend alors uniquement de la
surveillance agrave lrsquoabattoir Pour quelques deacutepartements agrave fort risque elle est annuelle et peut ecirctre
compleacuteteacutee par un deacutepistage lors de mouvements drsquoanimaux Le test de deacutepistage de reacutefeacuterence est
lrsquointradermotuberculination (IDR) Ce test vise agrave mettre en eacutevidence in vivo la reacuteaction
drsquohypersensibiliteacute retardeacutee conseacutecutive agrave lrsquoinjection intradermique de tuberculine (proteacuteines purifieacutees
de surnageant de culture de mycobacteacuteries) chez un bovin infecteacute (Buddle et al 2011 De la Rua-
Domenech et al 2006) Cette reacuteaction se traduit par une tumeacutefaction cutaneacutee douloureuse et chaude
Deux types drsquoIDR sont utiliseacutes lrsquoIntradermotuberculination simple ou IDS qui consiste en lrsquoinjection
intradermique de tuberculine bovine sur le plat de lrsquoencolure et lrsquoIntradermotuberculination
comparative ou IDC qui associe lrsquoinjection intradermique de tuberculine bovine et de tuberculine
aviaire LrsquoIDC est utiliseacutee dans certains deacutepartements dont la Cocircte drsquoOr ougrave de nombreuses
Chapitre 1
29
mycobacteacuteries environnementales sont retrouveacutees pour augmenter la speacutecificiteacute du test et eacutecarter les
reacuteactions croiseacutees chez les bovins sensibiliseacutes agrave une autre mycobacteacuterie (des antigegravenes sont communs
entre les mycobacteacuteries) (De la Rua-Domenech et al 2006 OIE 2009)
LrsquoIDR peut ecirctre compleacuteteacutee par le dosage de lrsquoInterfeacuteron γ (IFNγ) Cette cytokine qui est libeacutereacutee
par les lymphocytes T activeacutes lors drsquoune stimulation antigeacutenique in vivo mais eacutegalement in vitro permet
entre autre lrsquoactivation de la fonction phagocytaire des macrophages La quantiteacute drsquoIFNγ libeacutereacutee lors de
la culture de sang total avec de la tuberculine bovine ou aviaire est suffisante pour ecirctre mesureacutee par
un test ELISA (Gormley et al 2006 de la Rua-Domenech et al 2006) Le dosage de lrsquoIFNγ est pratiqueacute
pour renforcer lrsquoIDR lorsque la contention des animaux est difficile essentiellement en Camargue et
dans les Landes pour les taureaux de combat Il est eacutegalement utiliseacute agrave titre expeacuterimental dans quelques
deacutepartements comme la Dordogne et la Cocircte drsquoOr (Fediaevsky et al 2014) pour renforcer les reacutesultats
drsquoIDR
Drsquoautres tests notamment seacuterologiques sont en cours drsquoeacutevaluation chez les bovins Les essais
reacutealiseacutes jusqursquoagrave maintenant montrent une sensibiliteacute et une speacutecificiteacute infeacuterieures agrave lrsquoIDR ou lrsquoIFNγ
notamment chez les animaux en deacutebut drsquoinfection mais ils repreacutesentent un axe de recherche important
au vu du faible coucirct et de la simpliciteacute de la technique (Waters et al 2011 de la Rua-Domenech et al
2006)
La seacuterologie et lrsquoIFNγ font lrsquoobjet de nombreux deacuteveloppements pour lrsquoameacutelioration de la
surveillance de la maladie chez les animaux sauvages Reacutecemment des techniques ELISA ont eacuteteacute
deacuteveloppeacutees pour de nombreuses espegraveces dont les sangliers les cerfs les bisons (Boadella et al 2011
Shuralev et al 2012 Boadella et al 2012 Lyashchenko et al 2013 Chapinal et al 2012) LrsquoIFNγ est
eacutegalement testeacute pour le deacutepistage chez les espegraveces sauvages notamment chez les cervideacutes drsquoeacutelevage
et les blaireaux (Palmer et al 2010 Dalley et al 2008) La seacuterologie (kit commercial Brock Stat-Pack
Chembio Diagnostic Systems) et le dosage de lrsquoIFNγ (meacutethode ELISA) sont utiliseacutes en routine apregraves
capture et preacutelegravevement sanguin pour le suivi eacutepideacutemiologique de groupes sociaux de blaireaux agrave
Woodchester Park en Angleterre (Drewe et al 2010 King Murphy James Travis Porter Sawyer et al
2015 King Murphy James Travis Porter Hung et al 2015) La combinaison des deux tests est
conseilleacutee pour ameacuteliorer la sensibiliteacute et la speacutecificiteacute du deacutepistage (Drewe et al 2010) Une meacutethode
de deacutepistage non invasive baseacutee sur la collecte puis lrsquoanalyse par qPCR des latrines a eacuteteacute eacutevalueacutee sur
douze groupes sociaux en parallegravele avec la seacuterologie et le dosage de lrsquoIFNγ (King Murphy James Travis
Porter Sawyer et al 2015 King Murphy James Travis Porter Hung et al 2015) Bien que moins
contraignante et moins couteuse que le suivi par capture lrsquoanalyse des latrines pour lrsquoidentification des
groupes sociaux de blaireaux positifs neacutecessite au moins deux campagnes annuelles de preacutelegravevements
(deacutebut et fin drsquoeacuteteacute) avec un nombre drsquoeacutechantillons minimum variable selon les groupes sociaux (de 10
agrave plus de 20) agrave cause de lrsquoexcreacutetion intermittente du bacille par les blaireaux infecteacutes De plus elle ne
Chapitre 1
30
permet pas un deacutepistage individuel
Concernant le diagnostic de la tuberculose bovine lrsquoinfection chez les bovins par M bovis doit
ecirctre eacutetablie par histologie en cas de leacutesion et confirmeacutee par culture selon la reacuteglementation
europeacuteenne Lrsquohistologie est reacutealiseacutee sur des coupes de nœuds lymphatiques et de leacutesions fixeacutees agrave la
formaline La culture repose sur le protocole deacuteveloppeacute et ameacutelioreacute par le Laboratoire National de
Reacutefeacuterence (LNR) devenu la norme NF U47-104 qui associe une phase de broyage-deacutecontamination
des eacutechantillons puis un ensemencement sur deux milieux agrave lrsquoœuf (Loumlwenstein-Jensen et Coletsos) (OIE
2009) Toutefois ces deux techniques preacutesentent des inconveacutenients car lrsquohistologie manque de
speacutecificiteacute (environ 92 ) par rapport agrave la culture (Varello et al 2008) et le reacutesultat de culture est peu
sensible long (entre trois semaines et trois mois) ce qui pose des problegravemes pratiques en cas de
suspicion (restriction des mouvements du cheptel) Bien que pratiqueacutee depuis quelques anneacutees sans
base regraveglementaire il a eacuteteacute deacutemontreacute que la PCR baseacutee sur la deacutetection de lrsquoIS6110 (kit commercial LSI
VetMAXtrade Mycobacterium tuberculosis Complex PCR kit) serait un outil performant de diagnostic de la
tuberculose chez les bovins avec une bonne speacutecificiteacute (97 ) et une sensibiliteacute supeacuterieure agrave la culture
(877 vs 781 ) (Courcoul et al 2014) En apportant un diagnostic en 48 heures la PCR permettrait
une gestion plus rapide des foyers
Le diagnostic dans la faune sauvage repose sur une PCR systeacutematique sur les leacutesions eacutevocatrices
de tuberculose ou un pool de nœuds lymphatiques en lrsquoabsence de leacutesions En cas de reacutesultat non
neacutegatif lrsquoeacutechantillon est mis en culture pour identification de lrsquoagent infectieux (Source Note de service
DGALSDSPA2015-556 du 26062015) Quelle que soit lrsquoespegravece concerneacutee la culture et la PCR sont
reacutealiseacutees par des laboratoires agreacuteeacutes En cas de PCR etou de culture positive les eacutechantillons sont
systeacutematiquement envoyeacutes au Laboratoire National de Reacutefeacuterence pour la tuberculose bovine pour
confirmation et geacutenotypage des souches
f Epideacutemiologie de la tuberculose bovine
Evolution de la tuberculose bovine dans le monde et en
Europe et mesures de lutte
Avant 1920 la tuberculose bovine eacutetait lrsquoune des maladies majeures des animaux domestiques
agrave lrsquoeacutechelle mondiale A lrsquoheure actuelle elle est toujours largement reacutepandue dans le monde (Cosivi et
al 1999) La maladie est plutocirct sporadique et limiteacutee agrave quelques reacutegions dans les pays industrialiseacutes
toucheacutes (sauf en Irlande au Royaume-Uni en Espagne et au Portugal) et endeacutemique dans les pays agrave
faibles revenus drsquoAfrique drsquoAmeacuterique du sud et drsquoAsie (Figure 6)
Chapitre 1
31
Dans les pays industrialiseacutes la mise en place de la pasteurisation du lait dans les anneacutees 1950-
60 et des programmes drsquoeacuteradication de la maladie baseacutes sur le deacutepistage et lrsquoabattage des bovins
infecteacutes ont permis de reacuteduire fortement lrsquoincidence de lrsquoinfection chez les bovins et lrsquohomme
Aujourdrsquohui dans ces pays la tuberculose bovine repreacutesente principalement un enjeu socio-
eacuteconomique lieacute aux restrictions des eacutechanges internationaux drsquoanimaux et aux coucircts des programmes
de lutte contre la maladie chez les animaux le problegraveme sanitaire est devenu mineur Dans les pays agrave
faibles revenus ougrave lrsquoabsence ou quasi-absence de controcircle de la maladie chez les bovins et la
consommation de lait et produits deacuteriveacutes crus les enjeux de la tuberculose bovine sont agrave la fois
sanitaires et socio-eacuteconomiques
Figure 6 Carte de distribution de la tuberculose bovine chez les bovins entre juillet et deacutecembre 2014 (Source WAHID Interface)
En Europe depuis plus drsquoun siegravecle lrsquoeacuteradication de la maladie a eacuteteacute un objectif majeur pour les
organisations agricoles et les autoriteacutes publiques (Reviriego-Gordejo amp Vermeersch 2006) Les mesures
reacuteglementaires eacutelaboreacutees notamment par lrsquoOIE (Office International des Epizooties aujourdrsquohui appeleacute
Organisation Mondiale pour la Santeacute Animale) visent agrave seacutecuriser les eacutechanges commerciaux drsquoanimaux
lrsquoameacutelioration des techniques de deacutepistage de la maladie les programmes drsquoeacuteradication et leurs aides
financiegraveres Les programmes mis en place chez les bovins ont permis de faire fortement diminuer
lrsquoincidence de la maladie dans la plupart des pays de lrsquoUnion Europeacuteenne (de la Rua-Domenech et al
2006) voire de lrsquoeacuteradiquer leur confeacuterant le statut de pays laquo officiellement indemne de tuberculose
bovine raquo (OTF Officially Tuberculosis Free) Le statut OTF est attribueacute agrave un pays lorsque
- le pourcentage de cheptels bovins confirmeacutes comme infecteacutes nrsquoexcegravede pas 01 par an pour
tous les cheptels pendant 6 ans
Chapitre 1
32
- chaque bovin est identifieacute en accord avec la regraveglementation europeacuteenne
- tous les bovins abattus subissent un examen post-mortem officiel (Reviriego-Gordejo amp
Vermeersch 2006)
Ce statut est tregraves important pour lrsquoeacuteconomie drsquoun pays puisqursquoil facilite le commerce national
et international des animaux par un allegravegement des controcircles sanitaires avant lrsquoexportation de leurs
bovins dans un autre pays membre ou avant leur abattage en les exemptant notamment drsquoune
tuberculination (Reviriego-Gordejo amp Vermeersch 2006) Les pays europeacuteens officiellement indemnes
de tuberculose sont lrsquoAllemagne lrsquoAutriche la Belgique le Danemark lrsquoEstonie la Finlande la France
lrsquoEcosse la Lettonie le Liechtenstein le Luxembourg la Norvegravege les Pays-Bas la Pologne la
Reacutepublique Tchegraveque la Slovaquie la Sloveacutenie la Suegravede la Suisse et une partie de lrsquoItalie (Figure 7) Les
pays drsquoEurope les plus toucheacutes sont la Grande-Bretagne lrsquoIrlande et lrsquoEspagne avec des preacutevalences
respectives de 104 44 et 12 en 2012 (EFSA 2014) Toutefois dans certains pays OTF comme la
France la Belgique lrsquoAllemagne lrsquoAutriche et la Pologne lrsquoeacuteradication de la maladie nrsquoest pas complegravete
malgreacute la preacutevention les controcircles et la surveillance engageacutes Dans ces pays lrsquoeacuteradication de la maladie
est fortement compliqueacutee par la circulation de lrsquoinfection dans des reacuteservoirs sauvages source
potentielle de contamination pour les bovins (Corner 2006 Nugent 2011 Gortaacutezar et al 2012)
Figure 7 Statuts des pays europeacuteens vis-agrave-vis de la tuberculose bovine en 2012 et proportion de bovins infecteacutes OTF Officially Tuberculosis Free (officiellement indemne de tuberculose bovine) MS Member State (Etat membre) (Source EFSA 2014)
Chapitre 1
33
Evolution de la situation sanitaire en France
Chez les bovins
Dans les anneacutees 50 environ 25 des cheptels bovins franccedilais eacutetaient infecteacutes par la tuberculose
bovine (Beacutenet et al 2006) Cette preacutevalence importante avait un fort impact eacuteconomique (saisies de
viande agrave lrsquoabattoir diminution de la productiviteacute en viande et en lait) mais aussi sanitaire puisque 2 agrave
11 des cas de tuberculose chez lrsquohomme eacutetaient imputeacutes agrave M bovis Les programmes de lutte mis en
place ont permis de faire diminuer de faccedilon spectaculaire la preacutevalence cheptel de la maladie de 25
en 1955 agrave moins de 01 degraves 1998 permettant agrave la France drsquoacceacuteder au statut OTF en 2000 (Figure 8)
Ce statut a permis drsquoalleacuteger voire de stopper la prophylaxie sur le territoire drsquoalleacuteger les controcircles mis
en place pour les mouvements de bovins dans lrsquoUE et de fournir une garantie sanitaire aux pays
importateurs de bovins franccedilais dans lrsquoUE et en dehors
Les programmes de lutte contre la tuberculose bovine reposent sur une base reacuteglementaire en
eacutevolution constante eacutelaboreacutee par les services veacuteteacuterinaires (maintenant DDPP Direction
Deacutepartementale de la Protection des Populations) en collaboration technique avec les veacuteteacuterinaires
sanitaires les groupements de deacutefense sanitaire (GDS) et les laboratoires drsquoanalyse agreacuteeacutes Ces
programmes sont baseacutes sur la surveillance systeacutematique et geacuteneacuteraliseacutee en abattoir (inspection post-
mortem recherche et identification de mycobacteacuteries par culture et PCR sur les leacutesions suspectes) et
sur la surveillance programmeacutee en eacutelevage (deacutepistage par intradermo-tuberculination et abattage total
ou partiel des troupeaux infecteacutes) Ils assurent eacutegalement la protection des cheptels indemnes par la
maitrise des facteurs de diffusion de la tuberculose bovine tels que lrsquointroduction drsquoun animal le
voisinage avec des cheptels infecteacutes et la reacutesurgence (Fediaevsky et al 2014) Depuis 2013 deux
Cellules Inter Reacutegionales drsquoEpideacutemiologie Veacuteteacuterinaire (CIREV) ont eacuteteacute creacuteeacutees en Bourgogne et en
Aquitaine afin de renforcer les investigations eacutepideacutemiologiques pour adapter de faccedilon optimale les
mesures de lutte contre la maladie (Fediaevsky et al 2013)
Chapitre 1
34
Figure 8 Evolution du taux de preacutevalence annuelle national de la tuberculose bovine chez les bovins (Beacutenet et al 2006)
Depuis 2004 la France connait une situation preacuteoccupante vis-agrave-vis de la tuberculose bovine
car lrsquoaugmentation du taux de preacutevalence met en danger le statut OTF du pays (Figure 9) Cette
augmentation est lieacutee agrave lrsquoapparition de nouveaux foyers de tuberculose concentreacutes dans des zones
geacuteographiques bien localiseacutees la Cocircte drsquoOr la Dordogne et les Pyreacuteneacutees Atlantiques cumulent 75
des foyers et dans une moindre mesure la Charente la Camargue les Hautes Pyreacuteneacutees lrsquoAriegravege les
Landes et la Corse (Figure 10) Dans ces deacutepartements caracteacuteriseacutes par de lrsquoeacutelevage allaitant
preacutedominant la contamination des troupeaux par le voisinage (par contacts directs avec des bovins
voisins ou des animaux sauvages infecteacutes ou par contacts indirects avec du mateacuteriel ou des matrices
contaminants) et lrsquointroduction drsquoun animal infecteacute dans le cheptel (notamment par lrsquoachat drsquoanimaux
le precirct et la pension) repreacutesentaient la moitieacute des cas en 2011 La reacutesurgence drsquoune ancienne infection
(qui persiste agrave bas bruit chez les animaux ou dans des matrices environnementales souilleacutees) et la faune
sauvage eacutetaient agrave lrsquoorigine preacutesumeacutee drsquoenviron 10 des cas respectivement toutefois sans
deacutemonstration scientifique Pour 35 des foyers bovins lrsquoorigine de lrsquoinfection eacutetait inconnue
(Fediaevsky et al 2012) La contamination des bovins via le voisinage la reacutesurgence et les foyers
drsquoorigine inconnue soulegravevent la question du rocircle de la transmission indirecte de M bovis entre la faune
sauvage et les bovins et donc le rocircle potentiel de lrsquoenvironnement comme reacuteservoir du bacille
25
15
10
5
20
1955
1963
1960
1970
1980
1990
2000
Chapitre 1
35
Figure 9 Evolution de la preacutevalence et de lrsquoincidence de la tuberculose bovine en France entre 1995 et 2014 (Cavalerie et al 2014)
La menace de la perte du statut OTF inquiegravete les pouvoirs publics et les instances agricoles car
la France est le plus gros exportateur de bovins vivants de lrsquoUE elle exporte principalement des bovins
maigres en Italie et en Espagne pour lrsquoengraissement (environ 12 million de tecirctes) mais aussi des gros
bovins destineacutes agrave la boucherie (220 000 tecirctes) et des veaux (128 000 tecirctes) (Source France AgriMer
les filiegraveres de lrsquoeacutelevage franccedilais eacutedition 2013) Les conseacutequences eacuteconomiques sur la filiegravere-
agroalimentaire lieacutees au blocage des animaux seraient consideacuterables De plus la gestion sanitaire de la
tuberculose bovine est tregraves coucircteuse puisque environ 20 millions drsquoeuros sont deacutepenseacutes chaque anneacutee
agrave lrsquoeacutechelle nationale pour lrsquoindemnisation des eacuteleveurs et la reacutealisation des analyses veacuteteacuterinaires
(Fediaevsky et al 2014)
Chapitre 1
36
Figure 10 Incidence et distribution de la tuberculose bovine en France dans les anneacutees 50 (en haut) et entre 2000 et 2014 (en bas) (Cavalerie et al 2014)
Chapitre 1
37
Dans la faune sauvage
En France agrave lrsquoheure actuelle la faune sauvage infecteacutee nrsquoest deacutetecteacutee qursquoagrave proximiteacute de foyers
bovins Le 1er foyer de tuberculose bovine dans la faune sauvage a eacuteteacute deacutecouvert en 2001 dans la forecirct
de Brotonne (Seine-Maritime et Eure) sur des cervideacutes puis des sangliers (Hars et al 2010 Zanella et
al 2008) Lrsquoaggravation de la situation sanitaire en 2005-2006 a neacutecessiteacute un abattage total des cerfs
consideacutereacutes comme reacuteservoir primaire de lrsquoinfection ainsi qursquoune diminution de la densiteacute des sangliers
reacuteservoir secondaire (Zanella et al 2008) Drsquoautres deacutepartements comme la Cocircte drsquoOr la Corse les
Pyreacuteneacutees Atlantiques la Dordogne et la Charente notamment ont par la suite deacutetecteacute des animaux
sauvages infecteacutes essentiellement blaireaux sangliers et dans une moindre mesure cervideacutes avec
toutefois des situations eacutepideacutemiologiques tregraves diffeacuterentes selon les reacutegions toucheacutees Ces situations
deacutependent de la population animale impliqueacutee (espegraveces densiteacutes interactionshellip) et de
lrsquoenvironnement (climat paysagehellip) Dans tous les cas le geacutenotypage (spoligotype et VNTR) des
souches incrimineacutees dans la faune sauvage a montreacute qursquoelles eacutetaient identiques agrave celles retrouveacutees
dans les eacutelevages bovins voisins suggeacuterant un lien eacutepideacutemiologique entre cas domestiques et sauvages
(Zanella et al 2008 Payne et al 2012 Richomme et al 2010)
Depuis 2011 un reacuteseau de surveillance eacutepideacutemiologique de la tuberculose bovine dans la faune
sauvage appeleacute Sylvatub a eacuteteacute mis en place en France Les diffeacuterentes modaliteacutes de surveillance ont
pour but la deacutetection de lrsquoinfection chez les animaux sauvages dans les zones agrave risque et preacutesumeacutees
indemnes ainsi que le suivi de son eacutevolution afin de proposer des mesures de lutte aux autoriteacutes
sanitaires (Riviegravere et al 2011) La surveillance est baseacutee sur un classement du territoire national en 3
niveaux de risque Pour les zones situeacutees en risque 1 (risque faible correspondant agrave une absence de
bovins deacutetecteacutes) il est demandeacute aux chasseurs de signaler toute leacutesion suspecte lors de lrsquoexamen initial
de la venaison Concernant les zones ougrave il existe un risque aveacutereacute (niveau 2 ou 3) la surveillance peut
ecirctre agrave la fois eacuteveacutenementielle (ramassage et analyse des cadavres drsquoanimaux trouveacutes morts) etou
programmeacutee (analyse des cerfs et sangliers tueacutes agrave la chasse et les blaireaux pieacutegeacutes) Les reacutesultats des
analyses effectueacutees en 2013-2014 sont preacutesenteacutes dans la Figure 11
Chapitre 1
38
Figure 11 Reacutepartition et reacutesultats des analyses effectueacutees dans le cadre du dispositif Sylvatub chez le grand gibier drsquoaoucirct 2013 agrave aoucirct 2014 et chez les blaireaux en 2014 (Source Sylvatub)
Evolution de la situation sanitaire en Cocircte drsquoOr
Quelques geacuteneacuteraliteacutes sur lrsquoeacutelevage bovin en Cocircte drsquoOr
La Cocircte drsquoOr est un deacutepartement drsquoeacutelevage bovin principalement allaitant (231 270 tecirctes en 2013
dont 74 565 vaches allaitantes et 15 687 vaches laitiegraveres) Lrsquoeacutelevage allaitant est majoritairement
concentreacute dans les zones herbagegraveres de lrsquoouest du deacutepartement et lrsquoeacutelevage laitier sur le plateau de
Langres au nord de Dijon Lrsquoeacutelevage allaitant est extensif (les bovins sont au preacute drsquoavril agrave octobre) lieacute agrave
la production de broutards maigres destineacutes agrave ecirctre engraisseacutes principalement en Italie En Cocircte drsquoOr le
parcellaire des exploitations de bovins allaitant est souvent tregraves morceleacute
Situation eacutepideacutemiologique chez les bovins et mesures de lutte associeacutees
Historiquement la Cocircte drsquoOr est un deacutepartement en prise avec la tuberculose depuis de
nombreuses anneacutees (Figure 10) A la fin des anneacutees 50 le taux de preacutevalence annuel chez les bovins
atteignait 65 Les mesures de lutte collective mises en place ont permis drsquoabaisser ce taux en dessous
du seuil des 10 en 1969 puis de le faire diminuer lentement et reacuteguliegraverement jusqursquoen 1985 ougrave la
maladie semblait eacuteradiqueacutee A la fin des anneacutees 90 les mesures prophylactiques ont cesseacute dans le
Chapitre 1
39
deacutepartement et la vigilance vis-agrave-vis de la maladie a baisseacute (Communication personnelle DDPP 21)
Les premiers signes drsquoune nouvelle eacutepizootie sont apparus en 2002 sur le canton de Pouilly-en-
Auxois (zone Sud) puis en 2003 dans ceux de Venarey-Les-Laumes et Vitteaux (zone Nord) (Source
DDPP21) Localiseacutee agrave quelques eacutelevages agrave son deacutebut la zone de lrsquoinfection srsquoest peu agrave peu eacutelargie au
cours du temps Un plan de prophylaxie restreint aux communes infecteacutees a eacuteteacute mis en place en 2002
puis eacutelargi aux cantons en 2003 avec de plus une surveillance des cheptels en lien eacutepideacutemiologique
avec les troupeaux infecteacutes et ceux des communes dans lesquelles les bovins infecteacutes avaient patucircreacute
Mais ces mesures nrsquoont pas suffi agrave limiter la progression de lrsquoinfection puisque limiteacute agrave moins de 5 par
an jusqursquoen 2006 le nombre de foyers est passeacute de 11 en 2007 agrave 17 en 2009 (Figure 13) Le
renforcement de la lutte comprenant notamment le deacutepistage annuel de tous les bovins de plus de 12
mois dans tout le deacutepartement a permis la deacutecouverte de 45 foyers en 2010 signe que lrsquoinfection
circulait agrave bas bruit De 2011 agrave 2013 le nombre de foyers a eacuteteacute diviseacute par 2 et pour lrsquoanneacutee 2014 10
foyers seulement ont eacuteteacute mis en eacutevidence ce qui semble conforter une maitrise de lrsquoeacutepizootie (Figure
13)
En Cocircte drsquoOr les foyers bovins sont circonscrits agrave une zone geacuteographique bien deacutelimiteacutee dans le
deacutepartement et partagent le mecircme spoligotype M bovis spoligotype SB0134 dans la zone Sud et
M bovis spoligotype SB0120 dans la zone Nord (Figure 12) Depuis 3 ans toutefois les deux zones ont
tendance agrave se teacutelescoper agrave cause notamment de la deacutecouverte drsquoanimaux sauvages infecteacutes avec
M bovis SB0120 en zone Sud Les deux spoligotypes ont deacutejagrave eacuteteacute retrouveacutes simultaneacutement dans des
cheptels infecteacutes Ces observations associeacutees agrave lrsquoabsence drsquoinstallation peacuterenne de la maladie dans des
zones du deacutepartements ougrave les concentrations de bovins sont aussi eacuteleacuteveacutees que dans la zone infecteacutee
soulegravevent la question de lrsquoexistence de facteurs locaux assurant lrsquoentretien et la propagation de
lrsquoinfection entre les eacutelevages bovins Au vu du morcellement et de lrsquoimbrication des pacirctures la
contamination par voisinage est probablement le facteur primordial chez les bovins Une eacutetude reacutecente
(de type cas-teacutemoin) reacutealiseacutee dans des cheptels de Cocircte drsquoOr de Dordogne et des Ardennes a montreacute
que les contacts de voisinage entre troupeaux eacutetaient le facteur de risque principal drsquoinfection des
troupeaux (Marsot et al Article soumis) Toutefois la deacutetection drsquoanimaux sauvages agrave proximiteacute de
certains foyers bovins interroge sur le rocircle potentiel de la faune sauvage infecteacutee dans la contamination
des bovins et dans la recontamination rapide de plusieurs cheptels apregraves abattage total des bovins et
vide sanitaire (Figure 13) (ANSES 2011)
Chapitre 1
40
Figure 12 Localisation du parcellaire des cheptels infecteacutes par la tuberculose bovine entre 2010 et 2015 et spoligotypes incrimineacutes (Source CIREV)
Figure 13 Nombre de nouveaux foyers bovins et de foyers recontamineacutes entre 2002 et 2014 (Donneacutees DDPP21)
0
5
10
15
20
25
30
35
40
45
50
2002 2003 2004 2005 2006 2007 2008 2009 2010 2011 2012 2013 2014
No
mb
re d
e fo
yers
bo
vin
s
Foyers contamineacutes pour la 1egravere fois Foyers recontamineacutes
Chapitre 1
41
Situation eacutepideacutemiologique dans la faune sauvage et mesures de lutte associeacutees
Le premier animal sauvage infecteacute une biche a eacuteteacute deacutecouvert en 2003 agrave proximiteacute de 2 foyers
bovins de 2002 dans la zone Sud Entre 2003 et 2007 la surveillance active de la faune sauvage mise
en place et porteacutee par lrsquoONCFS (Office National de la Chasse et de la Faune Sauvage) et la DDPP et sur
le terrain par les chasseurs les pieacutegeurs et les veacuteteacuterinaires a permis de mettre en eacutevidence lrsquoinfection
chez 1 cervideacute (n=284) 2 sangliers (n= 160) mais aucun blaireau (n=63) Depuis 2007 des sangliers
infecteacutes sont retrouveacutes tous les ans notamment de jeunes animaux avec des leacutesions eacutevolutives
majoritairement dans la zone Sud ougrave les populations sont plus importantes Depuis 2009 lrsquoinfection
est retrouveacutee chez des blaireaux vivant agrave proximiteacute des cheptels infecteacutes majoritairement dans la zone
Nord (Hars Richomme and Boschiroli 2010 DDPP21)(Figure 14) La maladie circule donc dans un
systegraveme multi-hocirctes complexe et unique incluant les bovins les blaireaux les sangliers et les cervideacutes
(Payne 2014)
Figure 14 Nombre de cervideacutes sangliers et blaireaux infecteacutes deacutecouverts en Cocircte drsquoOr entre 2002 et 2014
Dans la crainte de lrsquoeacutetablissement drsquoun reacuteservoir sauvage chez le sanglier comme en Espagne
(Naranjo et al 2008) ou le blaireau au Royaume-Uni (Delahay et al 2001) des mesures de diminution
des densiteacutes ont eacuteteacute mises en place (augmentation des tableaux de chasse pour le sanglier et les
cervideacutes et intensification du pieacutegeage des blaireaux) le ramassage des viscegraveres drsquoanimaux tueacutes agrave la
chasse a eacuteteacute rendu obligatoire et lrsquoagrainage interdit
0
5
10
15
20
25
30
35
No
mb
re d
an
imau
x in
fect
eacutes
Cervideacutes Sangliers Blaireaux
Chapitre 1
42
C ECOLOGIE DE M BOVIS DANS LrsquoENVIRONNEMENT
1 Introduction
Avant les anneacutees 30 il eacutetait admis sine qua none qursquoen tant que bacteacuteries intracellulaires
obligatoires les bacteacuteries tuberculeuses ne survivaient pas en dehors de leur hocircte Toutefois degraves le
deacutebut du 20egraveme siegravecle la deacutetection de Bacillus tuberculosis virulent dans lrsquoeau et les biofilms des
canalisations drsquoeffluents de diffeacuterents sanatoria en Allemagne et au Royaume-Uni (Musehold en 1900
citeacute par Jensen 1954 et Conroy et al en 1921 et 1922 et Cummins et Ackland en 1929 citeacutes par (Williams
amp Hoy 1930) a souleveacute la question du rocircle potentiel de lrsquoenvironnement dans lrsquoeacutepideacutemiologie des
mycobacteacuteries tuberculeuses Concernant la tuberculose bovine les diffeacuterentes recherches meneacutees
pour deacutetecter et caracteacuteriser la persistance de M bovis dans lrsquoenvironnement ont pour objectif
principal de preacuteciser le rocircle de la transmission indirecte dans leacutepideacutemiologie de la maladie La
transmission indirecte semble en effet prendre toute son importance lorsque des reacuteservoirs sauvages
sont identifieacutes etou des systegravemes multi-hocirctes car ils ont la capaciteacute de transmettre lrsquoinfection aux
bovins (Corner 2006 Payne 2014)
Reacutepondre agrave cette question neacutecessite de connaitre les modaliteacutes de transfert des bacteacuteries de
lrsquoanimal ou lrsquohomme infecteacute vers lrsquoenvironnement la dureacutee de survie dans les diffeacuterentes matrices les
strateacutegies de survie de la bacteacuterie ainsi que les facteurs qui affectent sa survie
2 Origines de la contamination environnementale par M bovis
a Lrsquoexcreacutetion animale de Mycobacterium bovis
Lrsquohomme et les animaux infecteacutes ont la capaciteacute drsquoexcreacuteter le bacille via diffeacuterentes voies et
constituent donc une source de contamination potentielle pour lrsquoenvironnement Comme nous lrsquoavons
indiqueacute preacuteceacutedemment les voies drsquoexcreacutetion ainsi que les niveaux drsquoexcreacutetion deacutependent de la
localisation des leacutesions et de leur seacuteveacuteriteacute Les tableaux leacutesionnels deacutecrits preacuteceacutedemment indiquent
que M bovis est susceptible drsquoecirctre excreacuteteacutee principalement par voie orale ou nasale lors drsquoatteinte
pulmonaire etou des nœuds lymphatiques ceacutephaliques par voie digestive via les matiegraveres feacutecales et
par voie urinaire lors drsquoinfection geacuteneacuteraliseacutee (Phillips et al 2003 Corner et al 2012) Lrsquoexcreacutetion feacutecale
de bacteacuteries du MTBC en lrsquoabsence de leacutesions gastro-intestinales semble majoritairement lieacutee au
passage de mucus respiratoire chargeacute en bacilles dans le tube digestif La reacutesistance aux pH acides
(Vandal et al 2008) des bacteacuteries du MTBC leur permet de reacutesister au passage du tractus digestif et
drsquoecirctre excreacuteteacutees sous forme viable dans les matiegraveres feacutecales comme cela a eacuteteacute deacutemontreacute pour M bovis
dans certaines espegraveces animales (Corner Murphy and Gormley 2011 Gallagher and Clifton-Hadley
Chapitre 1
43
2000 Gavier-Widen et al 2001) pour M tuberculosis (El Kheacutechine et al 2009) et M africanum
(Cadmus et al 2009)
Chez les bovins lrsquoexcreacutetion du bacille a eacuteteacute deacutemontreacutee dans le mucus nasal et tracheacuteal et les
fegraveces drsquoanimaux infecteacutes expeacuterimentalement (Neill et al 1988 McCorry et al 2005 Phillips et al 2003
Maddock 1936) Le deacutelai drsquoexcreacutetion apregraves lrsquoinoculation la dureacutee de lrsquoexcreacutetion (plusieurs semaines) et
la quantiteacute de bacilles excreacuteteacutes sont proportionnels agrave la dose infectieuse utiliseacutee (Neill et al 1988)
Lrsquoexcreacutetion feacutecale apparait peu freacutequente et intermittente chez les bovins (Neill et al 1988 Goodchild
amp Clifton-Hadley 2001) Dans les pays industrialiseacutes ougrave le deacutepistage des animaux infecteacutes est en place
lrsquoexcreacutetion de M bovis dans le lait les fegraveces et les urines est consideacutereacutee comme insignifiante chez les
bovins (Morris et al 1994)
Chez le blaireau infecteacute naturellement ou expeacuterimentalement M bovis est excreacuteteacutee par 25 agrave
50 des animaux par voie respiratoire (mucus tracheacuteal et aeacuterosols) urinaire et feacutecale ainsi que par la
suppuration drsquoabcegraves reacutesultant de morsures (Corner et al 2011 Delahay et al 2000 Clifton-Hadley et
al 1993) Lrsquoisolement de M bovis dans des glandes salivaires de blaireaux suggegravere que la salive
constitue eacutegalement une voie possible drsquoexcreacutetion du bacille (Payne 2014) Les blaireaux preacutesentant un
stade avanceacute de la maladie et des leacutesions macroscopiques semblent ecirctre les plus excreacuteteurs en terme
de dureacutee et drsquointensiteacute drsquoexcreacutetion (Corner et al 2012 Gallagher amp Clifton-Hadley 2000 Jenkins et al
2008) toutefois lrsquoexcreacutetion du bacille a eacuteteacute deacutecrite chez des animaux sans leacutesions visibles (Jenkins et al
2008 Gavier-Wideacuten et al 2009) Dans les stades avanceacutes de la maladie les quantiteacutes excreacuteteacutees peuvent
atteindre 2105 UFCmL1 de mucus bronchique 25105 UFCmL1 drsquourine et 75104 UFCg1 de fegraveces
(Gallagher et al 1998) Le bacille est excreacuteteacute par voie feacutecale de 165 agrave 1305 jours avant la mort de
lrsquoanimal (Little et al 1982) Dans cette espegravece lrsquoexcreacutetion est geacuteneacuteralement intermittente mais des
individus laquo super excreacuteteurs raquo ont eacuteteacute deacutecrits et se caracteacuterisent par une excreacutetion persistante dans le
temps ou par lrsquoexcreacutetion du bacille par plusieurs voies simultaneacutement (Delahay et al 2000) Des
blaireaux laquo super excreacuteteurs raquo ont eacuteteacute mis en eacutevidence en Cocircte drsquoOr la voie feacutecale eacutetant toujours
positive chez ces animaux (Payne 2014) Lrsquointensiteacute de lrsquoexcreacutetion semble varier avec lrsquoacircge (les adultes
excregravetent plus que les jeunes) et la saison puisque lrsquoexcreacutetion semble supeacuterieure au printemps et en
eacuteteacute (Delahay et al 2013 King Murphy James Travis Porter Hung et al 2015)
Chez le sanglier lrsquoexcreacutetion de M bovis se produit par voie orale nasale feacutecale et urinaire
(Martiacuten-Hernando et al 2007 Santos Almeida et al 2015 Barasona et al 2015) Lrsquoexcreacutetion salivaire
est encore discuteacutee (Payne 2014) En Espagne lrsquoexcreacutetion par au moins une des voies a eacuteteacute mise en
eacutevidence par PCR sur 30 des sangliers analyseacutes (n = 39) et 25 des 39 animaux excreacutetaient par les trois
voies (Barasona et al 2015) Lrsquoexcreacutetion eacutetait majoritairement associeacutee agrave une maladie geacuteneacuteraliseacutee Au
Portugal sur 39 sangliers adultes ou subadultes infecteacutes analyseacutes 80 excreacutetaient le bacille par au
moins une voie et 13 eacutetaient des laquo super excreacuteteurs raquo (excreacutetant par trois voies diffeacuterentes) Environ
Chapitre 1
44
28 des sangliers excreacutetaient des quantiteacutes de bacilles supeacuterieures agrave 1 000 UFC par g ou mL drsquoexcreacuteta
par au moins une voie (Santos Almeida et al 2015) Chez le sanglier agrave notre connaissance lrsquoexcreacutetion
nrsquoa jamais eacuteteacute deacutecrite en lrsquoabsence de leacutesions
Chez les cervideacutes diffeacuterentes eacutetudes ont montreacute que le bacille eacutetait excreacuteteacute de faccedilon tregraves
sporadique dans les mucus nasal oro-pharyngeacute et tracheacuteal ainsi que dans les matiegraveres feacutecales lors
drsquoinfections naturelles ou expeacuterimentales (Lugton et al 1998 Santos Almeida et al 2015 Palmer et
al 2001) Dans cette espegravece une excreacutetion intermittente apparait degraves les premiers stades de la
maladie la seacuteveacuteriteacute des tableaux leacutesionnels rencontreacutes explique le niveau drsquoexcreacutetion eacuteleveacute (Palmer et
al 2001 Santos Almeida et al 2015)
Les carcasses drsquoanimaux infecteacutes peuvent constituer une source drsquoinfection directe pour les
animaux charognards qui srsquoen nourrissent mais eacutegalement une source de contamination indirecte pour
les bovins qui pacircturent sur le site de deacutecomposition de la carcasse (Morris et al 1994) Concernant les
animaux tueacutes agrave la chasse lrsquoabandon des viscegraveres ou leur ensevelissement repreacutesente eacutegalement un
risque pour les animaux charognards De ce fait en France la collecte et lrsquoeacutelimination des viscegraveres des
espegraveces sensibles sont obligatoires dans les zones toucheacutees par la tuberculose bovine
b Le transfert de Mycobacterium bovis par les effluents drsquoeacutelevage ou urbains
Les eacutetudes sur la contamination potentielle des effluents drsquoeacutelevage avec M bovis sont tregraves
rares probablement agrave cause de la difficulteacute agrave cultiver le bacille dans ces matrices complexes Les
multiples voies drsquoexcreacutetion du bacille et plus particuliegraverement lrsquoexcreacutetion par voie feacutecale mecircme si elle
est intermittente peuvent probablement contribuer agrave la contamination des effluents drsquoeacutelevage bovin
tels que les fumiers lisiers et eaux useacutees Lrsquoeacutepandage de ces effluents sur les pacirctures ou lrsquoirrigation des
cultures peut constituer un vecteur de contamination du sol des cultures et de lrsquoeau profonde ou de
surface et repreacutesenter un risque pour les animaux et lrsquohomme mais aucune donneacutee chiffreacutee nrsquoexiste
pour ces matrices en conditions naturelles Lrsquoabattage et la deacutecoupe de bovins tuberculeux peut
eacutegalement repreacutesenter une source de contamination des rejets drsquoabattoir (Jensen 1954) Au vu des
mesures actuelles de collecte stockage traitement et controcircle des effluents drsquoabattoir cette
contamination semble tregraves peu probable en France Toutefois elle reste complegravetement valable dans
les zones drsquoendeacutemie des pays nrsquoappliquant pas ces mesures et ne disposant pas de structures
drsquoabattage adeacutequates
Il est inteacuteressant de noter que quelques eacutetudes sont disponibles concernant la contamination
drsquoeffluents urbains avec M tuberculosis Jensen (1954) a montreacute que les effluents bruts de sanatoria
de 5 villes des Pays-Bas les effluents purifieacutes les boues digeacutereacutees et les boues seacutecheacutees (preacuteleveacutees 5
semaines apregraves lrsquoeacutepandage) contenaient des bacilles virulents de M tuberculosis Plus reacutecemment la
Chapitre 1
45
mise en culture de 800 eacutechantillons drsquoeau et 700 eacutechantillons de sol preacuteleveacutes dans 3 quartiers de
Teacuteheacuteran (Iran) entre 2012 et 2014 a deacutemontreacute la preacutesence de M tuberculosis dans 71 des eacutechantillons
drsquoeau (88) et 11 eacutechantillons de sol (15) Les eacutechantillons drsquoeau contamineacutes ont eacuteteacute retrouveacutes
principalement dans les rigoles de ciment acheminant les eaux useacutees et pluviales (5671) et dans les
eaux traiteacutees (1571) La bacteacuterie nrsquoa toutefois pas eacuteteacute isoleacutee du reacuteseau drsquoeau potable Environ 10 des
souches isoleacutees eacutetaient multi-reacutesistants aux antibiotiques (Velayati et al 2015)
3 Deacutetection et survie de Mycobacterium bovis dans lrsquoenvironnement
a Deacutetection de Mycobacterium bovis en conditions naturelles
A ce jour peu drsquoeacutetudes concernant la deacutetection de M bovis dans des substrats naturellement
contamineacutes ont eacuteteacute publieacutees La deacutetection de M bovis dans des eacutechantillons environnementaux
naturellement contamineacutes gracircce agrave des meacutethodes baseacutees sur la culture est exceptionnelle A
Woodchester Park dans le Gloucestershire (Sud-Ouest de lrsquoAngleterre) ougrave les densiteacutes de blaireaux
sont eacuteleveacutees et la preacutevalence importante (20 en 2000 et 35 en 2005) un suivi longitudinal du statut
des populations est reacutealiseacute depuis 1980 (Delahay et al 2000) et diffeacuterentes campagnes de
preacutelegravevements de sols de terriers et de latrines ont eacuteteacute reacutealiseacutees Le traitement drsquoeacutechantillons de sol et
latrines provenant de Woodchester Park traiteacute par immunocapture magneacutetique a permis pour la
premiegravere et seule fois lrsquoisolement de M bovis par culture dans des eacutechantillons environnementaux
(Sweeney et al 2006 Sweeney et al 2007) Cette technique est toutefois peu applicable agrave un grand
nombre drsquoeacutechantillons en termes de coucirct et de temps et nrsquoa jamais eacuteteacute reacuteutiliseacutee par la suite
Drsquoautres tentatives de deacutetection de M bovis par diffeacuterentes meacutethodes baseacutees sur la culture
dans des zones endeacutemiques de tuberculose bovine ont eacutechoueacute malgreacute le grand nombre drsquoeacutechantillons
environnementaux preacuteleveacutes (Pillai et al 2000 Fine Bolin et al 2011 Iivanainen et al 1999 Williams
amp Hoy 1930a Little et al 1982) La deacutetection de M bovis dans des eacutechantillons environnementaux par
la culture est compliqueacutee par la nature des eacutechantillons analyseacutes en effet ces derniers (sol seacutediments
eau stagnante matiegraveres feacutecales etc) contiennent des micro-organismes tregraves divers et abondants La
croissance de M bovis eacutetant tregraves lente les organismes agrave croissance rapide contaminent rapidement les
milieux de culture compliquant voire empecircchant lrsquoisolement de la bacteacuterie Des traitements de
deacutecontamination preacuteliminaires sont indispensables pour eacuteliminer cette microflore mais la
deacutecontamination reacuteduit en geacuteneacuteral fortement la viabiliteacute de M bovis reacuteduisant aussi la sensibiliteacute des
meacutethodes baseacutees sur la culture (Fine Bolin et al 2011) Des microorganismes voisins des
mycobacteacuteries et preacutesents dans les sols peuvent eacutegalement reacutesister aux proceacutedeacutes de deacutecontamination
et ecirctre retrouveacutes dans les isolements De plus lrsquointermittence et le faible niveau drsquoexcreacutetion chez les
Chapitre 1
46
animaux infecteacutes associeacutes au volume limiteacute des preacutelegravevements analyseacutes diminuent les chances de
deacutetection de M bovis dans lrsquoenvironnement (Fine OrsquoBrien et al 2011) Lrsquoameacutelioration des protocoles
de traitement des eacutechantillons avant la culture est fondamentale pour lrsquoisolement des souches
environnementales
Le deacuteveloppement des outils moleacuteculaires a permis de contourner les inconveacutenients lieacutes aux
techniques baseacutees sur la culture deacutecrits preacuteceacutedemment En Irlande lrsquoanalyse de preacutelegravevements reacutealiseacutes
en 2000 dans un eacutelevage bovin toucheacute plusieurs fois par la tuberculose a mis en eacutevidence par des
meacutethodes moleacuteculaires la preacutesence de 1 times 103 agrave 25 times 103 copies de gegravenes cibles de M bovis (mpb64 et
mpb70) dans des pacirctures et des sols drsquoentreacutee de terriers de blaireaux adjacents aux pacirctures (Young et
al 2005) La reacuteiteacuteration des analyses 30 mois plus tard en novembre 2002 a donneacute le mecircme reacutesultat
sur les terriers bien qursquoils soient resteacutes inhabiteacutes Le rocircle des blaireaux dans la contamination des bovins
est fortement suspecteacute dans cette ferme En Angleterre lrsquoanalyse moleacuteculaire de preacutelegravevements
environnementaux (terriers et latrines) reacutealiseacutes en 2003 autour ou dans les pacirctures drsquoeacutelevages bovins
infecteacutes dans six zones endeacutemiques de tuberculose bovine a mis en eacutevidence M bovis dans les sols
de terriers de blaireaux de 78 des fermes analyseacutees (n=60) Dans chaque ferme 47 des terriers et
29 des latrines analyseacutes eacutetaient positifs en PCR une forte proportion eacutetait accessible aux bovins
(Courtenay et al 2006)
Entre 2004 et 2013 lrsquoanalyse par deacutetection moleacuteculaire de latrines appartenantt agrave des groupes sociaux
de blaireaux infecteacutes de Woodchester Park a montreacute que selon les eacutetudes 13 agrave 100 des latrines eacutetaient
contamineacutees avec des quantiteacutes pouvant atteindre 11 times 108 copies de gegravenes par g de latrine
(Courtenay et al 2006 Travis et al 2011 Sweeney et al 2006) Lrsquoamplification de seacutequences ciblant
lrsquoARNr 16S dans certains eacutechantillons des eacutetudes preacutesenteacutees ci-dessus suggegravere que les bacilles deacutetecteacutes
eacutetaient viables (Young et al 2005 Courtenay et al 2006) toutefois la seacutequence de lrsquoARNr 16S cibleacutee
eacutetant speacutecifique de toutes les mycobacteacuteries agrave croissance lente les reacutesultats sont agrave consideacuterer avec
prudence Dans ces eacutetudes le nombre de blaireaux excreacuteteurs et leur niveau drsquoexcreacutetion sont
fortement correacuteleacutes agrave la contamination des terriers et des latrines
Dans le sud du Portugal une eacutetude reacutecente dans une zone endeacutemique a mis en eacutevidence par PCR ciblant
lrsquoIS6110 des bacteacuteries du MTBC dans 38 des 248 eacutechantillons preacuteleveacutes dans des zones drsquoagreacutegation
drsquoanimaux domestiques et sauvages zones drsquoabreuvement utiliseacutees par les bovins et la faune sauvage
(seacutediments et eau drsquoeacutetangs 1358 et 357 et de sources saisonniegraveres 1161) sol des zones de
nourrissage des animaux (528) du sol de boutis de sangliers (416) ainsi que dans le sol des zones de
nourrissage des vautours (119) et dans leurs fegraveces (19) (Santos Santos et al 2015) Lrsquoutilisation drsquoune
seconde cible PCR (RD12) a confirmeacute 4 eacutechantillons comme positifs pour M bovisM caprae (2
eacutechantillons de seacutediments drsquoeacutetang 1 de source et 1 de sol de nourrissage) sans pouvoir diffeacuterencier les
deux espegraveces Le plus grand nombre drsquoeacutechantillons positifs a eacuteteacute retrouveacute au printemps (tempeacuterature
Chapitre 1
47
moyenne de 16degC)
La PCR permet drsquoobtenir un reacutesultat rapide avec une speacutecificiteacute acceptable Lrsquoanalyse
drsquoeacutechantillons de sol drsquoeau de foin et de maiumls inoculeacutes avec M bovis par une meacutethode baseacutee sur la
culture et la PCR a montreacute que cette derniegravere permet de deacutetecter lrsquoADN de M bovis dans lrsquoeacutechantillon
sur une plus longue peacuteriode apregraves la contamination que la culture indeacutependamment du substrat
(Adams et al 2013) Toutefois les seuils de deacutetection eacuteleveacutes des systegravemes speacutecifiques de M bovis (de
4 times 104 agrave 5 times 105 par g ou mL drsquoeacutechantillon Tableau 3) limitent leur utilisation dans le screening
drsquoeacutechantillons environnementaux pour lesquels la charge bacteacuterienne est probablement infeacuterieure au
seuil de deacutetection Enfin la technique PCR ne donne aucune information sur la viabiliteacute et la virulence
des bacteacuteries deacutetecteacutees
b Dureacutees de survie de Mycobacterium bovis en conditions expeacuterimentales
Les eacutetudes expeacuterimentales sur la survie environnementale de M bovis ont deacutebuteacute dans les
anneacutees 1930 en Angleterre dans lrsquoobjectif de preacuteciser le rocircle de lrsquoenvironnement dans lrsquoeacutepideacutemiologie
de la tuberculose bovine et drsquoameacuteliorer les programmes drsquoeacuteradication de la maladie (Maddock 1933
Maddock 1934 Maddock 1936 Williams amp Hoy 1930) Mycobacterium bovis et M bovis BCG sont
capables de survivre dans les fegraveces les effluents le sol et les divers excreacutetas animaux de quelques jours
agrave quelques mois voire jusqursquoagrave deux ans () Les nombreuses conditions drsquoinoculation (concentration de
lrsquoinoculum souche utiliseacutee M bovis ou M bovis BCG origine de lrsquoinoculum culture ou broyat de
leacutesions) la varieacuteteacute des matrices inoculeacutees (matiegraveres feacutecales urine mucus organes sol eau etc) et
des conditions environnementales testeacutees (conditions controcircleacutees de laboratoire ou exposition aux
conditions climatiques) dans ces diffeacuterentes eacutetudes expliquent vraisemblablement de tels eacutecarts de
survie
La compilation de toutes ces donneacutees indique que la tempeacuterature lrsquoexposition aux UV et
lrsquohumiditeacute de la matrice semblent ecirctre les trois facteurs primordiaux intervenant dans la persistance
de M bovis quel que soit le substrat analyseacute Il est montreacute que dans lrsquoheacutemisphegravere Nord la dureacutee de
survie de M bovis est plus longue en automne hiver et printemps (Williams amp Hoy 1930 Tanner amp
Michel 1999 Fine Bolin et al 2011 Duffield amp Young 1985 Jackson et al 1995) saisons caracteacuteriseacutees
par des tempeacuteratures plus basses qursquoen eacuteteacute un ensoleillement minimal et une eacutevapotranspiration par
le couvert veacutegeacutetal limiteacutee Toutefois Fine et al (2011) ont montreacute que M bovis pouvait survivre jusqursquoagrave
48 jours dans lrsquoeau en eacuteteacute malgreacute les tempeacuteratures eacuteleveacutees et lrsquoensoleillement Les eacutetudes reacutealiseacutees en
Afrique du Sud et en Nouvelle-Zeacutelande ougrave les tempeacuteratures sont eacuteleveacutees sur de longues dureacutees avec
un ensoleillement important ont montreacute que la survie de M bovis est ameacutelioreacutee lorsque le sol contient
une humiditeacute suffisante sous une exposition ombrageacutee (Duffield amp Young 1985 Tanner amp Michel 1999)
Chapitre 1
48
Lrsquoexposition des bacilles aux UV diminue en effet fortement leur dureacutee de persistance dans
lrsquoenvironnement (Williams amp Hoy 1930a Genov 1965 Fine Bolin et al 2011) et la dureacutee de survie est
tregraves prolongeacutee lorsque les eacutechantillons sont enterreacutes agrave quelques centimegravetres sous la surface du sol
(Genov 1965) La survie de M bovis est optimale dans les matiegraveres feacutecales ougrave le bacille est proteacutegeacute
dans un environnement humide agrave lrsquoabri des radiations solaires et minimale lorsqursquoil est pulveacuteriseacute sur
les veacutegeacutetaux et soumis aux UV et agrave la dessiccation (Maddock 1933 Van Donsel amp Larkin 1977)
49
Reacutefeacuterences BCG M bovis Matrices inoculeacutees Concentration de
lrsquoinoculum Conditions
Meacutethodes de deacutetection
de M bovis Reacutesultats de survie (virulence) de M bovis
(Williams amp Hoy
1930)
X Fegraveces de bovin
Inconnue Expeacuterimentales
(tempeacuterature
fraicircche obscuriteacute)
Injection sous-
cutaneacutee agrave des cochons
drsquoInde (cuisse)
12 mois dans fegraveces naturellement infecteacutees
5104 agrave 5107 UFC 2 ans dans fegraveces artificiellement infecteacutees
X Lisier de bovin 5000 UFCcm3
Expeacuterimentales
(tempeacuterature
fraicircche obscuriteacute)
Injection sous-
cutaneacutee agrave des cochons
drsquoInde (cuisse)
Survie 4 mois avec diminution de la virulence du
bacille au cours du temps
X Fegraveces de bovin 5104 agrave 5107 UFC Environnementales
(pacircture)
Injection sous-cutaneacutee
agrave des cochons drsquoInde
(cuisse)
5 mois en hiver
4 mois en automne
2 mois au printemps eacuteteacute
4 mois si protection UV en eacuteteacute
6 mois si protection insectes-vers en automne
(Maddock 1933)
X
Sol
Sol + Fegraveces de bovin
Bouse
Inconnue Environnementales
(pacircture)
Injection sous-cutaneacutee
agrave des cochons drsquoInde
(cuisse)
178 j dans toutes les matrices
X Herbe
12105 01 m2 Environnementales
(pacircture)
Injection sous-
cutaneacutee agrave des cochons
drsquoInde (cuisse)
14 jours
12106 01 m2 28 jours
120106 01 m2 49 jours
(Maddock 1934) X Herbe
Inoculation
mensuelle de 5 agrave
25106 01 m2
reacutepeacuteteacutee
Environnementales
et expeacuterimentales
Ingestion drsquoherbe
contamineacutee
1422 cochons drsquoInde avec leacutesions
Tableau 1 Dureacutees de survie de M bovis dans diffeacuterentes matrices environnementales obtenues en conditions expeacuterimentales naturelles ou de laboratoire
50
(Maddock 1936)
X
Veaux infecteacutes
(excreacutetion prouveacutee
par injection agrave des
cochons drsquoInde)
Inconnue Environnementales
Pacircture seacutepareacutee en 3 2
veaux mis 1 mois apregraves
le retrait des veaux
infecteacutes 2 veaux 1 mois
plus tard 2 veaux 2
mois plus tard (laisseacutes 3
sem)
Aucun veau IDR +
Aucune leacutesion tuberculeuse agrave lrsquoautopsie
X
Vache infecteacutee
(excreacutetion prouveacutee
par injection agrave des
cochons drsquoInde)
Pacircture seacutepareacutee en 2 3
veaux mis 1 mois et 12
apregraves le retrait de la
vache infecteacutee (laisseacutes
53j) 3 veaux mis 1 mois
plus tard (laisseacutes 24j)
Abattage et autopsie 1 an plus tard aucune leacutesion
Bouses infectieuses 1 mois et frac12 apregraves retrait de la
vache infecteacutee puis neacutegatif 1 2 et 3 mois plus tard
Sol et herbe sous bouses neacutegatifs 3 4 et 5 mois apregraves
retrait de la vache infecteacutee
(Genov 1965) X
Fegraveces de bovin
Sang
Urine
Environnementales Culture
150 agrave 332 jours agrave 12-24degC sans drsquoUV
18 agrave 31 jours agrave 24-34degC avec UV
2 ans si enterreacute dans sol agrave 5 cm
11 mois si enterreacute agrave 1 cm
452 et 469 jours dans eaubouse et eauurine
(Van Donsel amp
Larkin 1977) X
Effluents animaux
Boues drsquoeacutepuration Environnementales
Abattage 90 en 11 jours pour sols arroseacutes avec
effluents 8 jours avec boues
Isolement apregraves 4 agrave 6 jours sur des radis et jusqursquoagrave
35 jours sur des laitues
Anon 1979 citeacute par
(Morris et al 1994)
Excreacutetas de
blaireaux conserveacutes
dans un terrier
Environnementales Culture
Urine 3 jours en eacuteteacute 28 jours en hiver
Mucus bronchique 7 et 70 jours
Fegraveces naturellement infecteacutes 14 et 28 jours
(Little et al 1982)
Sol foin litiegravere et
eau de blaireaux
inoculeacutes
Inconnue
Expeacuterimentales
Culture 1 culture positive (eau drsquoun abreuvoir)
51
(Duffield amp Young
1985)
Sol sec steacuterile ou non
Sol 30 humiditeacute
steacuterile ou non
Fegraveces
Culture
lt8 semaines dans sols secs ou humides non steacuteriles
avec 80 ombre (34degc) obscuriteacute (32degC) et dans
labo (23degC)
lt8 semaines dans sol humide steacuterile avec 80
ombre et obscuriteacute
Pas de survie plein soleil (43degC) dans sols et fegraveces
(Tanner amp Michel
1999)
Leacutesions pulmonaires
ou ganglionnaires de
buffle
Fegraveces de bovin
inoculeacutees
8107 UFC 50g
fegraveces Culture
Poumon 42 jours en hiver agrave lrsquoombre dans sol
humide ou sec (20degC)
Fegraveces 28 jours en hiver au soleil dans sol humide
ou sec
(Jackson et al
1995) X Rubans de coton
Environ 106 UFC
ruban
Environnementales
(rubans deacuteposeacutes
dans terriers de
possum et sols de
pacirctures ou de forecirct)
Culture
2 jours sur les sols de pacirctures
14 jours sur les sols forestiers et dans les terriers de
possum
(Scanlon amp Quinn
2000b) X Lisier bovin steacuteriliseacute 65 times 103 UFCmL Expeacuterimentales Culture 171 jours (concentration finale 2 UFCmL)
(Young et al 2005) X Sol steacuterile et non
steacuterile 108 UFC g sol
Expeacuterimentales
(4 15 25 37degC)
PCR pour les sols non
steacuteriles
Culture pour les sols
steacuteriles
Survie agrave 60 jours optimale agrave 37degC agrave -20kPa
drsquohumiditeacute dans le sol steacuterile
Deacutetection drsquoARNm pendant 15 mois (sol steacuterile)
(Palmer amp Whipple
2006) X
Betterave pommes
carottes pommes de
terre maiumls foin
11 times 106
UFC10g
Expeacuterimentales
(-20 8 et 23degC)
Culture
112 jours agrave -20degC pour tous les substrats
112 jours agrave 8degC pour pommes maiumls foin et pdeterre
112 jours agrave 23degC pour pommes maiumls et pdeterre
(Fine Bolin et al
2011) X Sol foin maiumls eau
50 times 104
UFCeacutechantillon Environnementales Culture
88 jours dans le sol en hiverprintemps
58 jours dans lrsquoeau 43 jours dans le foin et 37 jours
dans le maiumls en automnehiver
52
(Ghodbane et al
2014) X Sol steacuterile 108 UFC g sol
Expeacuterimentales
(18 agrave 24degC
obscuriteacute)
Culture
Survie 12 mois (concentration finale 150 UFCg
sol)
Virulence conserveacutee sur souris
Chapitre 1
53
Dans les fegraveces drsquoanimaux M bovis survit jusqursquo agrave 2 ans dans les bouses de bovins en conditions
de laboratoire (Williams amp Hoy 1930) et de 14 jours agrave 6 mois lorsque les fegraveces sont exposeacutees dans les
pacirctures (Williams amp Hoy 1930a Genov 1965 Morris et al 1994 Tanner amp Michel 1999) Dans les
effluents drsquoeacutelevages contamineacutes il a eacuteteacute montreacute que M bovis survivait de 4 mois dans du lisier en
conditions de laboratoire (Williams amp Hoy 1930a) et jusqursquoagrave 171 jours dans du lisier steacuteriliseacute conserveacute
agrave tempeacuterature ambiante et agrave lrsquoobscuriteacute (Scanlon amp Quinn 2000b) Lrsquoeacutepandage drsquoeffluents ou de boues
inoculeacutees avec M bovis BCG sur des cultures de radis et laitue a montreacute la reacuteduction de 90 des bacilles
en respectivement 11 et 8 jours M bovis a toutefois eacuteteacute isoleacutee de faccedilon sporadique 35 jours apregraves
arrosage (Van Donsel amp Larkin 1977)
Dans le sol les dureacutees de survie les plus longues pour M bovis et M bovis BCG ont eacuteteacute
observeacutees dans du sol steacuterile (63 de sable pH 65-68) et en conditions de laboratoire (Young et al
2005 Ghodbane et al 2014) Young et al (2005) ont amplifieacute lrsquoARNr 16S de M bovis BCG jusqursquoagrave 15
mois apregraves inoculation du sol alors que Ghodbane et al (2014) ont cultiveacute M bovis apregraves 12 mois de
seacutejour dans le sol (pH 83) Dans les conditions environnementales Fine et al (2011) ont confirmeacute
lrsquoeffet de la saison sur la survie de M bovis dans le sol sur du maiumls et du foin avec une survie maximale
en hiverprintemps (feacutevrier-mai tempeacuterature moyenne 29degC radiations solaires 800 Wm2) une
survie de 34 jours en automnehiver (novembre-janvier tempeacuterature moyenne 04degC radiations
solaires 243 Wm2) et une survie minimale de 12 jours pendant la peacuteriode printempseacuteteacute (mai-aoucirct
tempeacuterature moyenne 217degC radiations solaires 1500 Wm2) De faccedilon contradictoire lrsquoeacutetude de
Young et al (2005) a mis en eacutevidence une dureacutee de survie optimale (60 jours) de M bovis BCG agrave 37degC
et minimale (16 jours) agrave 4degC
La survie de M bovis a eacutegalement eacuteteacute mise en eacutevidence par meacutethode culturale sur diffeacuterents
aliments du beacutetail (Palmer amp Whipple 2006 Fine Bolin et al 2011) Apregraves 112 jours la bacteacuterie est
cultiveacutee sur tous les substrats inoculeacutes conserveacutes agrave -20degC (betteraves pommes carottes pommes de
terre maiumls et foin) sur les pommes le maiumls le foin et les pommes de terre conserveacutes agrave 8degC et
uniquement sur les pommes le maiumls et les pommes de terre conserveacutes agrave 23degC (Palmer amp Whipple
2006) Fine et al (2011) ont eacutegalement montreacute que M bovis survivait plus longuement en
automnehiver sur du maiumls (43 jours) et du foin (37 jours) qursquoagrave la peacuteriode printempseacuteteacute (survie
infeacuterieure agrave 3 jours)
La bacteacuterie peut eacutegalement survivre entre 2 et 6 semaines dans des leacutesions tuberculeuses
macroscopiques de poumons et nœuds lymphatiques de buffle (Tanner amp Michel 1999) la survie
optimale eacutetant observeacutee en hiver dans des situations ombrageacutees et humides Des reacutesultats identiques
sont observeacutes pour des eacutechantillons drsquourine (3 jours en eacuteteacute vs 28 jours en hiver) et des seacutecreacutetions
bronchiques (7 jours vs 70 jours) (Morris et al 1994)
Les diffeacuterentes eacutetudes preacutesenteacutees ci-dessus deacutemontrent que M bovis survit un temps variable dans
Chapitre 1
54
lrsquoenvironnement une fois excreacuteteacute par son hocircte Ce temps est fonction de lrsquoexcreacuteta et du substrat qui le
reccediloit de sa charge bacteacuterienne et des conditions climatiques Il existe donc une peacuteriode appeleacutee
fenecirctre temporelle critique pendant laquelle les animaux qui interagissent avec les matrices
contamineacutees par des bacilles viables preacutesentent un risque accru drsquoinfection Comme nous lrsquoavons vu
preacuteceacutedemment la fenecirctre temporelle critique est fortement deacutependante du pays ou de la reacutegion
eacutetudieacutes et des conditions climatiques
Tregraves peu de donneacutees existent sur la conservation de la virulence de M bovis apregraves un passage
en dehors de son hocircte La virulence du bacille a eacuteteacute confirmeacutee par injection agrave des cobayes dans des
fegraveces de bovins naturellement contamineacutees apregraves 178 jours drsquoexposition dans une pacircture et apregraves 12
mois drsquoincubation en conditions de laboratoire (Maddock 1933) Lrsquoeacutetude reacutecente de Ghodbane et al
(2014) a deacutemontreacute que M tuberculosis M canetti et M bovis conservaient leur virulence apregraves un
seacutejour de 12 mois dans le sol En effet lrsquoinjection intra-peacuteritoneacuteale de suspensions bacteacuteriennes agrave des
souris a entraineacute lrsquoapparition de leacutesions avec M canetti (nodules sur les poumons le foie et la rate) et
M tuberculosis (nodules sur les poumons et la rate) Malgreacute lrsquoabsence de leacutesions lors de lrsquoinjection de
M bovis la bacteacuterie a pu ecirctre isoleacutee en quantiteacute importante des 3 organes Toutefois les injections
eacutetaient reacutealiseacutees avec les colonies de mycobacteacuteries obtenues apregraves culture sur milieu geacuteloseacute induisant
un biais important De plus lrsquoingestion par des souris de sol inoculeacute par M tuberculosis apregraves 12 mois
drsquoincubation a provoqueacute des leacutesions pulmonaires (1 souris5) et des leacutesions de la rate (25) mais la
bacteacuterie a pu ecirctre cultiveacutee agrave partir des poumons de la rate de lrsquoestomac et de lrsquointestin chez toutes les
souris
4 Strateacutegies de survie des mycobacteacuteries dans lrsquoenvironnement
Lrsquoenveloppe particuliegravere des mycobacteacuteries deacutecrite preacuteceacutedemment est probablement un des
facteurs majeurs de la reacutesistance de M bovis en dehors de son hocircte dans des conditions
environnementales deacutefavorables Drsquoautres strateacutegies de survie sont probablement mises en jeu par
M bovis pour persister dans lrsquoenvironnement mais sont totalement inconnues agrave ce jour Les
mycobacteacuteries pathogegravenes et opportunistes ont deacuteveloppeacute diffeacuterents meacutecanismes pour survivre agrave
lrsquointeacuterieur des cellules de lrsquohocircte strateacutegies qui peuvent eacutegalement ecirctre utiliseacutees en dehors de lrsquohocircte
Les meacutecanismes moleacuteculaires de certains processus sont eacutegalement en cours de compreacutehension Les
principales strateacutegies de survie sont preacutesenteacutees dans la partie suivante
a Lrsquoeacutetat de dormance
La dormance est deacutefinie chez les bacteacuteries non sporuleacutees comme un eacutetat physiologique
Chapitre 1
55
reacuteversible permettant la survie de la bacteacuterie pendant une longue peacuteriode sans croissance ni reacuteplication
(Kaprelyants et al 1993) Le programme geacuteneacutetique de la bacteacuterie lui permet drsquoentrer en dormance (en
anglais Non-Replicating Persistence) lorsque les conditions exteacuterieures deviennent deacutefavorables
(notamment en conditions oligotrophiques) puis de retrouver sa capaciteacute agrave se diviser lorsque les
conditions redeviennent favorables La dormance est deacutecrite chez plusieurs espegraveces de mycobacteacuteries
Lrsquoentreacutee en dormance de M smegmatis (Dick et al 1998) M bovis (Lim et al 1999 Boon et al 2001)
et M tuberculosis (Wayne amp Hayes 1996) en culture est provoqueacutee par la rareacutefaction de lrsquooxygegravene
Lrsquoeacutetat de dormance est suggeacutereacute chez Map et pourrait expliquer sa longue survie dans le fumier le lisier
et le sol (Whittington et al 2004) Lrsquoeacutetat de dormance est eacutegalement deacutecrit chez M avium (Archuleta
et al 2005)
Chez les mycobacteacuteries lrsquoentreacutee en eacutetat de dormance est controcircleacutee par le facteur de
transcription DosR (Dormancy Survival Regulator) (Boon amp Dick 2002) Composeacute de 48 gegravenes
(Schnappinger et al 2003 Flores Valdez amp Schoolnik 2010) son expression est activeacutee in vitro par
lrsquooxyde nitrique (NO) (Voskuil et al 2003) le monoxyde de carbone (CO) (Shiloh et al 2008) le deacuteficit
en nutriments (Rodriguez et al 2008) et lrsquohypoxie (Rodriguez et al 2008) Lrsquoexpression de ce reacutegulon a
eacuteteacute deacutemontreacutee lors drsquoinfection expeacuterimentale de macrophages (Schnappinger et al 2003 Rodriguez et
al 2008) et drsquoanimaux modegraveles (Voskuil et al 2003) pour M tuberculosis et dans des nœuds
lymphatiques de bovins positifs en IDR pour M bovis (Jimeacutenez et al 2013) Ce reacutegulon permettrait non
seulement la survie de la bacteacuterie pendant la phase de dormance mais aussi la transition de la
respiration aeacuterobie agrave lrsquoanaeacuterobiose et inversement (Leistikow et al 2010) A lrsquoheure actuelle le rocircle de
certaines proteacuteines codantes et drsquoARN non codants de ce reacutegulon demeure toujours inconnu Une
eacutetude geacutenomique comparative a permis de constater que le reacutegulon de la dormance eacutetait conserveacute
chez les diffeacuterentes espegraveces de mycobacteacuteries y compris chez les mycobacteacuteries environnementales
(Gerasimova et al 2011) Cette donneacutee suggegravere que la dormance est une reacuteponse adaptative
probablement tregraves ancienne chez les mycobacteacuteries pour survivre dans des environnements ougrave les
concentrations en oxygegravene sont fluctuantes (Boon amp Dick 2012) et que cette adaptation a eacuteteacute mise agrave
profit par les mycobacteacuteries pour survivre dans les cellules immunitaires des hocirctes qursquoelles infectent
Aucune eacutetude nrsquoest agrave ce jour disponible concernant lrsquoentreacutee en dormance potentielle de
M bovis dans lrsquoenvironnement Le sol constitue toutefois un environnement hostile ougrave facteurs
biotiques et abiotiques pourraient deacuteclencher le passage de la bacteacuterie drsquoun eacutetat actif agrave un eacutetat
dormant (hypoxie carence en nutriments tempeacuterature hellip)
b La sporulation
Lrsquoeacutevocation de la sporulation chez les mycobacteacuteries est tregraves controverseacutee Ghosh et al (Ghosh
Chapitre 1
56
et al 2009) ont deacutecrit la sporulation expeacuterimentale de M marinum sous forme drsquoendospores dans des
cultures liquides acircgeacutees de 2 mois et dans des cultures solides de M bovis BCG de 8 mois Malgreacute des
veacuterifications pheacutenotypiques biochimiques et geacutenomiques deacutemontrant que ces spores eacutetaient bien
issues de M marinum ces reacutesultats ont eacuteteacute contesteacutes par Traag et al (Traag et al 2010) qui ont eacutechoueacute
agrave les reproduire Lamont et al (Lamont et al 2012) ont pour la premiegravere fois mis en eacutevidence
lrsquoapparition de morphotype spore-like dans des cultures liquides de Map drsquoorigine bovine (K-10) ovine
(7565) et humaine acircgeacutees drsquoun an en conditions oligotrophiques Le seacutequenccedilage de lrsquoARNr 16S et
lrsquoamplification de lrsquoIS900 speacutecifique de Map ont confirmeacute que les colonies bacteacuteriennes issues des
spores correspondaient bien agrave Map Le chauffage des spores obtenues agrave 70degC pendant 30 minutes ne
semble pas affecter sa virulence sur les macrophages Des eacutetudes compleacutementaires sont neacutecessaires
pour confirmer ces observations A notre connaissance la sporulation nrsquoa jamais eacuteteacute eacutevoqueacutee pour les
mycobacteacuteries du MTBC
c Interactions des mycobacteacuteries avec la faune du sol
La microfaune et plus particuliegraverement les amibes libres sont largement preacutesentes dans
lrsquoenvironnement et colonisent les eacutecosystegravemes aquatiques le sol (Khan 2006 Rodriacuteguez-Zaragoza
1994) et lrsquoair Elles ont eacuteteacute eacutegalement isoleacutees chez lrsquohomme Elles vivent preacutefeacuterentiellement dans les
biofilms et aux interfaces eau-sol eau-air et eau-plantes (Rodriacuteguez-Zaragoza 1994) ougrave elles
laquo broutent raquo les bacteacuteries les champignons et les algues dont elles reacutegulent les communauteacutes (Huws
et al 2005) Les mycobacteacuteries environnementales sont largement preacutesentes dans les milieux
aquatiques (Primm et al 2004) ougrave elles peuvent interagir avec les amibes De nombreuses eacutetudes
rapportent la capaciteacute des mycobacteacuteries environnementales agrave survivre et se multiplier dans les
trophozoiumltes etou les kystes drsquoamibes (Adekambi et al 2006 Drancourt et al 2007 Ben Salah amp
Drancourt 2010 Krishna-Prasad amp Gupta 1978 Mura amp Bull 2006) Les amibes pourraient mecircme
constituer un veacuteritable reacuteservoir pour M xenopi (Drancourt et al 2007 Thomas et al 2006)
La plupart des mycobacteacuteries pathogegravenes y compris M bovis sont capables de survivre dans
les amibes voire de srsquoy multiplier (Thomas amp McDonnell 2007 Mba Medie et al 2011 Greub amp Raoult
2004 Cirillo et al 1997 Adekambi et al 2006) La survie de M bovis a eacuteteacute confirmeacutee dans les
trophozoiumltes et les kystes drsquoAcanthamoeba castellanii jusqursquoagrave 5 mois apregraves infection en conditions
expeacuterimentales (Taylor et al 2003 Mardare et al 2013) La virulence vis-agrave-vis des macrophages
humains et des cellules intestinales de souris semble ecirctre exacerbeacutee par la reacuteplication intra-amibienne
(Cirillo et al 1997) En conditions naturelles lrsquoobservation de protozoaires isoleacutes de latrines de
blaireaux infecteacutes a mis en eacutevidence des BAAR sans pouvoir identifier preacuteciseacutement M bovis (Mardare
et al 2013) Des eacutetudes compleacutementaires sont neacutecessaires pour preacuteciser le rocircle de reacuteservoir eacuteventuel
Chapitre 1
57
des amibes pour M bovis Les interactions des mycobacteacuteries pathogegravenes avec les amibes pourraient
ecirctre du mecircme ordre que celles avec les macrophages de lrsquohocircte infecteacute notamment au niveau des
meacutecanismes de peacuteneacutetration (Danelishvili et al 2007) et de lrsquoinhibition de la fusion entre le phagosome
et le lysosome suggeacuterant qursquoune relation eacutevolutive des mycobacteacuteries avec les amibes auraient permis
aux mycobacteacuteries lrsquoacquisition de ces caracteacuteristiques de survie (Cirillo et al 1997)
La localisation intra-amibienne des mycobacteacuteries les protegravege des conditions
environnementales deacutefavorables des biocides tels que le chlore (Adekambi et al 2006 Whan et al
2006) des antibiotiques (Miltner amp Bermudez 2000) des diffeacuterents traitements comme lrsquoozonation de
lrsquoeau (Thomas et al 2008) et favorise leur disseacutemination et leur implication potentielle dans des
maladies humaines (Greub amp Raoult 2004) Lors de la peacuteneacutetration dans lrsquohocircte lrsquoamibe joue un rocircle de
laquo cheval de Troie raquo (Barker amp Brown 1994) proteacutegeant les bacteacuteries contre les premiegraveres deacutefenses de
lrsquohocircte
Lrsquoutilisation des amibes en co-cultures pour isoler des bacteacuteries reacutesistantes aux amibes dans
des eacutechantillons environnementaux et meacutedicaux a permis drsquoisoler des mycobacteacuteries drsquoeacutecosystegravemes
aquatiques (Thomas et al 2006 Thomas amp McDonnell 2007 Pagnier et al 2008) et de sols pollueacutes
(Wang et al 2006)
Les eacutetudes relatant les interactions entre M bovis et la meacuteso- et macrofaune sont rares Une
eacutetude reacutealiseacutee dans des eacutelevages bovins et caprins infecteacutes par Map a mis en eacutevidence la contamination
de 31 des vers de terre preacuteleveacutes sur les pacirctures freacutequenteacutees par les animaux (Fischer et al 2003)
Dans cette eacutetude Map a eacuteteacute retrouveacutee dans le corps et les fegraveces de vers de terre 1 et 2 jours apregraves
lrsquoingestion de bouses infecteacutees expeacuterimentalement suggeacuterant qursquoils peuvent probablement jouer un
rocircle de vecteurs dans la disseacutemination des mycobacteacuteries
Les eacutetudes disponibles bien que tregraves rares suggegraverent que la meacuteso- et macrofaune du sol
peuvent heacuteberger et probablement jouer un rocircle de vecteur des mycobacteacuteries Toutefois des eacutetudes
compleacutementaires sont indispensables concernant M bovis
d Interactions des mycobacteacuteries avec les diptegraveres
Le portage digestif et feacutecal de M bovis par des mouches communes (Musca domestica) en
contact avec des viscegraveres infecteacutes de bovins est deacutecrit La bacteacuterie survit de 12 jours sur leurs corps et
15 jours dans le tractus digestif de mouches mortes (Schlee 1957 citeacute par Kazda et al 2009)
e La formation de biofilms
Lrsquoaptitude agrave former des biofilms est connue pour de nombreuses espegraveces de mycobacteacuteries
Chapitre 1
58
environnementales agrave croissance rapide comme M smegmatis M fortuitum ou M abcessus ou agrave
croissance lente notamment M avium et M intracellulare (Falkinham et al 2001 Beumer et al 2010
Torvinen et al 2007) Lrsquoorganisation des mycobacteacuteries en biofilm leur confegravere une reacutesistance accrue
vis-agrave-vis des conditions deacutefavorables dans lrsquoenvironnement mais eacutegalement chez lrsquohocircte humain ougrave
elles peuvent coloniser les catheacuteters et provoquer des infections sanguines tregraves graves (El Helou et al
2013) Concernant les mycobacteacuteries du MTBC la formation de biofilms est deacutecrite pour
M tuberculosis dans des conditions expeacuterimentales tregraves particuliegraveres tregraves eacuteloigneacutees des conditions
naturelles (Kulka et al 2012) La formation drsquoagreacutegats (clumping ou cording) observeacutes lors de la culture
des bacteacuteries du MTBC en milieu liquide gracircce agrave une matrice extracellulaire riche en acides mycoliques
peut ecirctre assimileacutee agrave une forme organiseacutee de reacutesistance des bacteacuteries puisque sous cette forme les
bacteacuteries survivent agrave des concentrations tregraves eacuteleveacutees en antibiotique (Ojha et al 2008) Il a eacuteteacute montreacute
chez M tuberculosis que la formation de biofilms mettait en jeu des processus geacuteneacutetiques et
physiologiques diffeacuterents de la croissance planctonique (Ojha et al 2008) Toutefois comme il nrsquoa
jamais eacuteteacute deacutemontreacute que ces bacteacuteries se multiplient dans lrsquoenvironnement il semble peu probable
que les biofilms constituent une forme de reacutesistance pour M bovis
5 Facteurs de risque de contamination de lrsquoenvironnement et de transmission de Mycobacterium bovis aux animaux par lrsquoenvironnement contamineacute
Plusieurs eacutetudes portant sur les interactions entre bovins et faune sauvage ont montreacute que les
contacts directs eacutetaient tregraves peu freacutequents entre les bovins et les blaireaux (Benham amp Broom 1989
Drewe et al 2013 Boumlhm et al 2009) ainsi qursquoentre les cerfs les sangliers et les bovins (Cooper et al
2010 Kukielka et al 2013) La transmission indirecte de M bovis semble donc ecirctre le mode de
transmission preacutepondeacuterant entre la faune sauvage et les bovins voire entre les diffeacuterentes espegraveces
sauvages (Cowie et al 2015 Carrasco-Garcia et al 2015) Diffeacuterents facteurs de risque jouent un rocircle
sur ce mode de transmission qui neacutecessite un recouvrement des domaines vitaux des animaux
sauvages infecteacutes et excreacuteteurs avec les sites drsquoeacutelevage bovin (pacirctures et bacirctiments) Ces principaux
facteurs de risque sont deacutetailleacutes ci-dessous
a Risques lieacutes agrave lrsquoeacutecologie des espegraveces sauvages sensibles agrave Mycobacterium bovis
Concernant le blaireau et le sanglier les pacirctures peuvent constituer une grande partie de leur
domaine vital (Hutchings amp Harris 1999 Payne 2014) Elles sont utiliseacutees pour la recherche de
Chapitre 1
59
nourriture essentiellement des lombrics qui repreacutesentent une part importante de leur reacutegime
alimentaire (Tolhurst et al 2009 Cleary et al 2011 Baubet et al 2003) Une eacutetude reacutealiseacutee en Cocircte
drsquoOr a montreacute que la freacutequentation des pacirctures par les blaireaux est optimale au printemps peacuteriode
de mise au preacute des bovins par rapport agrave lrsquoautomne et agrave lrsquohiver Pour les sangliers la freacutequentation des
pacirctures est plus importante au printemps et en eacuteteacute (Payne 2014)
Concernant plus preacuteciseacutement les blaireaux ces derniers urinent et deacutefegravequent dans des latrines
pour deacutelimiter leurs territoires (Hutchings et al 2001) mais ils urinent eacutegalement sur les pacirctures
preacutefeacuterentiellement au niveau des lisiegraveres de forecirct des clocirctures et des haies qursquoils croisent au greacute de
leurs deacuteplacements dans un but de marquage (White et al 1993 Drewe et al 2013) Dans les pacirctures
il a eacuteteacute montreacute que le taux de contact entre bovins et latrines est eacuteleveacute (Hutchings amp Harris 1997 Drewe
et al 2013) Dans lrsquoeacutetude de Hutchings et Harris (1997) 48 des 50 vaches du troupeau ont cumuleacute 416
contacts avec 4 latrines preacutesentes dans la pacircture eacutetudieacutee et 33 animaux ont brouteacute sur les latrines (suivi
sur 1776 heures) Pour Drewe et al (2013) 1716 contacts avec 13 latrines ont eacuteteacute enregistreacutes pour les
33 bovins drsquoun troupeau sur les cinq mois de lrsquoeacutetude Toutefois le rang hieacuterarchique des bovins du
troupeau joue un rocircle important puisque les animaux domineacutes sont les plus enclins agrave brouter au
voisinage des latrines Enfin les bovins consomment lrsquoherbe au voisinage des latrines de faccedilon plus
importante lorsque lrsquoherbe de la pacircture est rase (en cas de surpacircturage ou de fauchage) (Hutchings amp
Harris 1997) Compte tenu du grand nombre de bacilles qui peuvent ecirctre excreacuteteacutes par la voie urinaire
du fort taux de contact bovinslatrines et de la consommation de lrsquoherbe sur les latrines par certains
bovins il semblerait que les latrines et plus particuliegraverement les zones de marquage urinaire dans les
pacirctures repreacutesentent un risque important de transmission indirecte de la maladie par inhalation ou
ingestion (Hutchings amp Harris 1999)
Bien qursquoil nrsquoexiste aucune eacutetude sur le sujet la contamination des pacirctures par des blaireaux
preacutesentant des eacutecoulements purulents de morsures a eacutegalement eacuteteacute envisageacutee (Gallagher amp Clifton-
Hadley 2000)
Les blaireaux vivent dans des terriers qursquoils creusent dans le sol en milieu forestier ou en
dehors notamment dans les pacirctures La contamination des sols drsquoentreacutee de terrier par M bovis
repreacutesente eacutegalement un risque pour les bovins qui peuvent se contaminer en explorant les entreacutees de
terrier localiseacutees dans les pacirctures par ingestion ou inhalation de la litiegravere expulseacutee des tunnels
(Benham 1985) De plus le sol excaveacute des tunnels par les blaireaux est attirant pour les bovins qui
peuvent en ingeacuterer pour combler des carences mineacuterales (Dewes 1996) ou srsquoy frotter la tecircte creacuteant
des aeacuterosols de poussiegravere potentiellement infectant (Phillips et al 2003) La contamination des sols de
terriers repreacutesente eacutegalement un risque important de transmission de lrsquoinfection dans le groupe social
de blaireaux qui partage le terrier Le sol du terrier constitue un biotope adeacutequat pour la survie de
M bovis agrave lrsquoabri des UV sans variation extrecircme de tempeacuterature et avec une humiditeacute relative
Chapitre 1
60
importante (Moore amp Roper 2003) Lrsquoentretien de la maladie dans un groupe social par le terrier
contamineacute a deacutejagrave eacuteteacute eacutevoqueacute (Rogers et al 2003)
La freacutequentation des pacirctures par les sangliers pour la recherche de nourriture constitue
eacutegalement un facteur de risque dans la transmission indirecte de la bacteacuterie aux bovins Lrsquoexploration
du sol avec leurs museaux agrave la recherche de lombrics larves et autre nourriture ainsi que les reliquats
de sol deacuteplaceacutes peuvent constituer un facteur de contamination potentiel de lrsquoenvironnement par la
deacuteposition drsquoexcreacutetas oro-nasaux infecteacutes
Les cervideacutes peuvent eacutegalement utiliser les preacutes pour se nourrir et ecirctre une source de
contamination indirecte pour les bovins (Brook amp McLachlan 2009)
b Risques lieacutes aux pratiques drsquoeacutelevage
Le seacutejour prolongeacute des bovins au preacute est susceptible drsquoaugmenter les contacts directs et
indirects avec la faune sauvage infecteacutee Lrsquoeacutelevage allaitant est caracteacuteriseacute par une longue peacuteriode de
pacircturage entre avril et octobre puis une hivernation en bacirctiments au moment des vecirclages Certaines
races rustiques notamment la race salers qui ont repeupleacute une partie des cheptels bovins infecteacutes
apregraves abattage total passent toute lrsquoanneacutee au pacircturage Ces longues peacuteriodes de pacircturage constituent
un facteur de risque non neacutegligeable pour les bovins qui partagent leur habitat avec des espegraveces
sauvages infecteacutees Le surpacircturage est eacutegalement un facteur de risque pour les bovins qui sont ameneacutes
agrave brouter les latrines de blaireaux et agrave ingeacuterer des quantiteacutes de sol potentiellement contamineacute plus
importantes (Hutchings amp Harris 1999)
Les zones drsquoabreuvement et de nourrissage des bovins dans la pacircture constituent des zones
attractives pour la faune sauvage notamment lrsquohiver lorsque les ressources alimentaires font deacutefaut
Le partage de fourrage entre bovins et cervideacutes est probablement une source de contamination des
bovins dans le Michigan (OrsquoBrien et al 2006 Brook amp McLachlan 2009 Brook et al 2012) drsquoautant plus
que la transmission indirecte de la maladie entre des cerfs et des veaux par le partage du foin a eacuteteacute
prouveacutee expeacuterimentalement (Palmer et al 2010 Palmer et al 2004) La freacutequentation des auges de
bovins et des zones de nourrissage au sol par les blaireaux qui viennent manger les reliquats
alimentaires est reacuteguliegraverement deacutecrite au Royaume-Uni ainsi que celle des reacuteserves de grains
drsquoensilage et de foin dans lesquels ils peuvent uriner et deacutefeacutequer (Garnett et al 2002 Tolhurst et al
2009 Ward et al 2010) Les pierres agrave sel souvent poseacutees au sol dans les preacutes attirent particuliegraverement
les cervideacutes (Payne et al 2015 Brook amp McLachlan 2009) Les abreuvoirs et les zones drsquoabreuvement
(trous drsquoeau ruisseau source) constituent eacutegalement un facteur de risque lorsqursquoils sont utiliseacutes par
les bovins et la faune sauvage la survie de la bacteacuterie excreacuteteacutee eacutetant optimale dans les zones humides
Le comportement de baignade des sangliers dans les trous drsquoeau et les souilles entraine une forte
Chapitre 1
61
freacutequentation de ces zones notamment en eacuteteacute (Payne 2014)
Finalement lrsquoeacutepandage de fumiers frais augmenterait le risque drsquoinfection drsquoun troupeau par
rapport agrave lrsquoeacutepandage drsquoengrais ou de fumiers non frais (stockeacutes au moins deux mois) les animaux se
contaminant par inhalation ou ingestion de substrats contamineacutes dans les deux mois suivant lrsquoeacutepandage
(Griffin et al 1996 Humblet et al 2009) Certaines preacuteconisations conseillent un temps de stockage
minimum des fumiers avant eacutepandage (au moins 6 mois suite aux reacutesultats de Scanlon et Quinn 2000a
et 2000b) ou leur traitement chimique (Scanlon amp Quinn 2000b Scanlon amp Quinn 2000a) un
compostage (McCallan et al 2014) voire lrsquointerdiction drsquoeacutepandage de fumiers pour les cheptels infecteacutes
(Phillips et al 2003)
c Risques lieacutes aux pratiques cyneacutegeacutetiques
Les zones drsquoagreacutegation des animaux sauvages telles que les zones drsquoagrainage ou de
nourrissage agrave points fixes et la reacutealisation de trous drsquoeau artificiels pour lrsquoabreuvement et la baignade
des animaux sauvages sont des zones agrave risque qui augmentent les interactions directes et indirectes
entre les animaux et donc les risques de transmission de M bovis entre eux (Corner 2006 Vicente et
al 2007 Delahay et al 2002 Coleman amp Cooke 2001) En Espagne lrsquoaugmentation des densiteacutes
drsquoonguleacutes sauvages (cervideacutes et sangliers) dans le Sud du pays a entraineacute le deacuteveloppement drsquoactiviteacutes
de chasse agrave but commercial amenant agrave la reacutealisation de parcs gigantesques pour contenir les animaux
sauvages Dans ces zones lrsquoagreacutegation des animaux sauvages de diffeacuterentes espegraveces (sangliers
cervideacutes cochons sauvages et bovins) autour des trous drsquoeau et des zones de nourrissage a favoriseacute la
transmission intra- et interspeacutecifique de la maladie (Vicente et al 2007 Cowie et al 2015) Chez le
sanglier la preacutevalence varie de 52 agrave 70 voire presque 100 dans certaines populations (Gortaacutezar et
al 2008 Naranjo et al 2008) Cowie et al (2015) ont montreacute que 40 agrave 66 des interactions indirectes
avaient lieu dans une fenecirctre temporelle de 3 jours en saison segraveche et 72 agrave 100 dans une fenecirctre
temporelle de 12 jours en saison humide Or dans ces zones la survie de M bovis a eacuteteacute estimeacutee agrave 3
jours pendant la saison segraveche et agrave 12 jours pendant la saison humide (Kukielka et al 2013) suggeacuterant
que si le pathogegravene est preacutesent dans une population lrsquoagreacutegation est un facteur de maintenance et de
disseacutemination de lrsquoinfection dans et entre les espegraveces De plus la preacutesence drsquoeacutelevages bovins agrave
proximiteacute de ces parcs de chasse entraine une freacutequentation importante des structures drsquoeacutelevage par
les animaux sauvages et un risque accru drsquoinfection pour les bovins des eacutelevages visiteacutes (Carrasco-
Garcia et al 2015)
Le nourrissage des cervideacutes dans le Michigan dans le but de maintenir les populations pour la
chasse est la cause du maintien de lrsquoinfection dans les populations sauvages qui se contaminent entre
eux via les aliments distribueacutes (OrsquoBrien et al 2006)
Chapitre 1
62
Comme nous lrsquoavons deacutejagrave dit preacuteceacutedemment les viscegraveres drsquoanimaux infecteacutes laisseacutes agrave la
disposition des animaux sauvages par les chasseurs peuvent repreacutesenter un risque important pour les
animaux charognards (Morris et al 1994)
d Risques lieacutes aux facteurs paysagers climatiques et peacutedologiques
La structure du paysage est un facteur qui influence le recouvrement des domaines vitaux des
animaux sauvages et des bovins La localisation des pacirctures ou des bacirctiments drsquoeacutelevage en lisiegravere de
forecirct augmente les risques drsquointeractions entre animaux sauvages et domestiques (Ward et al 2010)
lrsquointerface pacircture-forecirct repreacutesenteacutee par les lisiegraveres eacutetant un eacutecotone agrave risque pour lrsquoeacutemergence des
agents pathogegravenes multi-hocirctes (Boyard et al 2008) La fragmentation du paysage (preacutes champs forecircts
bosquets) augmente le deacutepocirct drsquourine par les blaireaux aux zones frontiegraveres entre deux milieux
augmentant le risque de contamination de ces sites (White et al 1993 Humblet et al 2009) Les lisiegraveres
sont tregraves freacutequenteacutees par les sangliers et les blaireaux (Payne 2014) et constituent une zone privileacutegieacutee
par les blaireaux pour eacutetablir leurs terriers et marquer leurs territoires par des mictions ou des latrines
(Hutchings et al 2002)
Comme nous lrsquoavons vu preacuteceacutedemment les conditions climatiques sont un facteur essentiel
dans la dureacutee de survie de M bovis dans les diffeacuterents substrats environnementaux contamineacutes et in
fine dans la transmission indirecte de la maladie aux animaux qui interagissent avec ces substrats Les
saisons influencent eacutegalement la freacutequentation des pacirctures par les animaux sauvages avec un risque
accru au printemps et en eacuteteacute vis-agrave-vis des blaireaux et des sangliers qui vermillent dans les pacirctures
notamment lors des nuits humides ougrave les vers de terre remontent en surface appeleacutees laquo nuits agrave
lombrics raquo (Kruuk 1978 Cleary et al 2011 Baubet et al 2003) Lors de temps plus sec voire de
seacutecheresse les points drsquoeau destineacutes aux bovins constituent des zones attractives pour la faune sauvage
(Vicente et al 2007 Santos Santos et al 2015 Barasona et al 2014) favorisant lrsquoagreacutegation des
animaux domestiques et sauvages
Le rocircle des facteurs peacutedologiques dans la survie de M bovis est peu connu agrave ce jour La texture
du sol qui conditionne la structure la porositeacute et le reacutegime hydrique du sol le pH et la matiegravere
organique disponible sont des facteurs importants dans la survie des bacteacuteries A notre connaissance
une seule eacutetude statistique reacutealiseacutee dans le Michigan a montreacute que la nature du sol et plus
preacuteciseacutement le pourcentage de sable eacutetait associeacute agrave un risque drsquoinfection plus eacuteleveacute chez les bovins
(Walter et al 2014) Une augmentation drsquo1 de la quantiteacute de sable dans le sol augmentait de 36
le risque pour des eacutelevages bovins drsquoecirctre infecteacutes Hormis cette eacutetude qui relie indirectement infection
animale et survie de la bacteacuterie dans le sol aucune eacutetude nrsquoest actuellement disponible sur le lien
direct entre nature du sol et survie de M bovis
Chapitre 1
63
A la lumiegravere des informations preacutesenteacutees dans ce chapitre il apparait qursquoune fois excreacuteteacutee en
dehors de son hocircte M bovis peut survivre longuement dans lrsquoenvironnement si les conditions sont
adeacutequates par la mise en jeu de strateacutegies de survie encore inconnues agrave lrsquoheure actuelle Les
connaissances sur les scheacutemas de circulation de la bacteacuterie entre les animaux infecteacutes et
lrsquoenvironnement (et vice et versa) srsquoameacuteliorent et permettent drsquoadapter les programmes de lutte contre
la maladie mais elles sont encore insuffisantes pour permettre la maitrise de tous les facteurs en jeu
Ces diffeacuterents facteurs sont scheacutematiseacutes dans la Figure 15
Figure 15 Repreacutesentation scheacutematique de la circulation de M bovis dans lrsquoenvironnement agricole (modifieacute drsquoapregraves Elliott et al 2014)
Chapitre 1
64
D PRINCIPAUX OUTILS DE DETECTION DE PHENOTYPAGE ET DE GENOTYPAGE APPLICABLES AU MTBC ET PLUS PARTICULIEREMENT A MYCOBACTERIUM BOVIS
Depuis la deacutecouverte du bacille de la tuberculose de nombreux outils ont eacuteteacute deacuteveloppeacutes pour
deacutetecter et discriminer les diffeacuterentes mycobacteacuteries du MTBC Ces meacutethodes microscopiques
pheacutenotypiques et biochimiques essentiellement deacuteveloppeacutees pour le diagnostic meacutedical ont eacuteteacute peu
agrave peu ameacutelioreacutees et associeacutees agrave de nouvelles techniques de deacutetection de caracteacuterisation et
drsquoidentification des bacilles telles que lrsquoamplification geacutenique et le seacutequenccedilage notamment ameacuteliorant
la rapiditeacute du diagnostic la discrimination des espegraveces la pertinence de lrsquoantibiotheacuterapie et permettant
le geacutenotypage des souches agrave des fins eacutepideacutemiologiques Adapteacutees agrave des eacutechantillons biologiques
divers tels que les crachats les liquides de lavage broncho-alveacuteolaire les tissus de biopsies chez
lrsquohomme ou les broyats de nœuds lymphatiques chez les bovins et les animaux sauvages ces outils ne
sont pas tous applicables aux eacutechantillons environnementaux heacutebergeant des communauteacutes
microbiennes tregraves complexes
1 La deacutetection des mycobacteacuteries par microscopie
La microscopie optique
Lrsquoexamen microscopique de frottis ou drsquoeacutetalement drsquoeacutechantillons biologiques est la premiegravere
eacutetape dans le diagnostic bacteacuteriologique de la tuberculose humaine Elle est parfois la seule meacutethode
de diagnostic dans les pays en voie de deacuteveloppement Les proprieacuteteacutes tinctoriales particuliegraveres de la
mycomembrane des mycobacteacuteries sont exploiteacutees dans les colorations utiliseacutees La coloration de
reacutefeacuterence est la coloration de Ziehl et Neelsen Cette coloration agrave chaud est baseacutee sur la reacutesistance de
la paroi des mycobacteacuteries agrave la deacutecoloration agrave lrsquoacide et lrsquoalcool apregraves traitement agrave la fuschine Les
mycobacteacuteries apparaissent comme des bacirctonnets rouges sur un fond bleu Des meacutethodes agrave froid
baseacutees sur les mecircmes reacuteactifs existent eacutegalement (meacutethode de Kinyoun) La coloration agrave lrsquoauramine
un fluorochrome permet drsquoobserver en microscopie agrave fluorescence les bacilles en vert-jaune brillant
sur un fond jaune-orangeacute Cette coloration est plus sensible que la coloration de ZN (Steingart et al
2006)
Ces meacutethodes de coloration ne permettent pas de discriminer les mycobacteacuteries entre elles et
ne sont pas speacutecifiques du genre Mycobacterium puisqursquoelles peuvent colorer drsquoautres actinobacteacuteries
comme les genres bacteacuteriens Nocardia Rhodococcus et Corynebacterium (Shinnick amp Good 1994)
Lrsquoutilisation de la microscopie optique nrsquoest pas deacutecrite pour lrsquoanalyse des eacutechantillons
environnementaux drsquoune part car le seuil de deacutetection est eacuteleveacute (de lrsquoordre de 104 BAARmL) et drsquoautre
Chapitre 1
65
part car la coloration est difficile agrave mettre en œuvre sur des matrices telles que du sol ou des matiegraveres
feacutecales
Lrsquohybridation avec des sondes nucleacuteotidiques FISH (Fluorescence in situ hybridization)
Cette meacutethode est baseacutee sur lrsquohybridation speacutecifique drsquoune sonde marqueacutee avec un
fluorochrome sur une reacutegion cible de lrsquoADN bacteacuterien Les sondes utiliseacutees ADN ARNm ou
oligonucleacuteotides syntheacutetiques (PNA peptide nucleic acid) drsquoenviron 20 bases peacutenegravetrent agrave lrsquointeacuterieur
des cellules preacutealablement permeacuteabiliseacutees ougrave elles srsquohybrident speacutecifiquement avec leur ADN cible
Lrsquoobservation se fait agrave lrsquoaide drsquoun microscope agrave eacutepifluorescence
La meacutethode FISH a surtout eacuteteacute deacuteveloppeacutee pour la deacutetection des mycobacteacuteries dans les
crachats dans des cultures liquides chez des animaux infecteacutes expeacuterimentalement (Hongmanee et al
2001 Stender Mollerup et al 1999 Stender Lund et al 1999) et dans des eacutechantillons biologiques
(Stender Mollerup et al 1999 Lefmann et al 2006 Rodriguez-Nuntildeez et al 2012) La meacutethode FISH
est peu utiliseacutee pour la recherche de mycobacteacuteries dans des eacutechantillons environnementaux mais elle
a permis de mettre en eacutevidence Maa et Map dans de lrsquoeau et des biofilms de canalisations (Lehtola et
al 2006 Torvinen et al 2007) Elle parait toutefois difficilement applicable agrave des matrices telles que le
sol agrave cause de lrsquoauto-fluorescence geacuteneacutereacutee par les particules de sol et du risque de non-speacutecificiteacute des
sondes
2 Les meacutethodes de culture des mycobacteacuteries
Le diagnostic de la tuberculose est indissociable de la culture car historiquement crsquoest la
meacutethode la plus utiliseacutee Elle est sensible et permet drsquoisoler les souches de les caracteacuteriser et de tester
leur sensibiliteacute aux antibiotiques La recherche de mycobacteacuteries par des meacutethodes baseacutees sur la
culture est tregraves standardiseacutee en diagnostic humain et veacuteteacuterinaire tout comme les traitements
preacuteliminaires indispensables A lrsquoopposeacute aucune meacutethode standardiseacutee nrsquoest disponible pour la
recherche de mycobacteacuteries dans lrsquoenvironnement De nombreux protocoles emprunteacutes agrave la
microbiologie clinique ont eacuteteacute adapteacutes pour la recherche de mycobacteacuteries environnementales
notamment dans lrsquoeau (Narang et al 2009 Radomski et al 2011 Norby et al 2007 Pickup et al 2006
Whiley et al 2012 Le Dantec et al 2002 Bland et al 2005) Les meacutethodes utiliseacutees pour cultiver les
mycobacteacuteries pathogegravenes sont moins nombreuses et sont deacutecrites ci-dessous
a Les diffeacuterents traitements preacutealables agrave la mise en culture des eacutechantillons
De nombreuses meacutethodes de deacutecontamination des eacutechantillons biologiques ont eacuteteacute deacutecrites
dans le diagnostic meacutedical de la tuberculose Le choix de la meacutethode agrave utiliser deacutepend essentiellement
Chapitre 1
66
du type drsquoeacutechantillon analyseacute (tissu crachat lait fegraveces eau solhellip) des milieux de culture employeacutes et
de sa compatibiliteacute avec les meacutethodes moleacuteculaires Les meacutethodes les plus utiliseacutees en microbiologie
humaine meacutethodes de Petroff Taquet-Tison et Kubica utilisent la soude comme agent deacutecontaminant
principal (Petroff 1915 Taquet amp Tison 1961 Kubica et al 1963) Lrsquoutilisation drsquoacides comme agent
deacutecontaminant tels que lrsquoacide oxalique (David et al 1989) ou lrsquoacide sulfurique (Hohn amp Lowenstein
1926) ou drsquoammoniums quaternaires comme le chlorure de ceacutetyl pyridinium (Smithwick et al 1975)
ou le chlorure de benzalkonium (Krasnow amp Wayne 1966) est reacuteserveacutee aux eacutechantillons tregraves contamineacutes
(urines fegraveces) ou aux biopsies (cutaneacutees coliques pulmonaires etc) Ces meacutethodes ne sont pas toutes
compatibles avec la culture en milieu liquide et lrsquoamplification geacutenique
Dans le domaine veacuteteacuterinaire les tissus animaux (broyats de ganglions ou de tissus) sont
deacutecontamineacutes selon les preacuteconisations de lrsquoANSES et suivent la norme AFNOR NF U47-104 baseacutee sur
lrsquoutilisation de lrsquoacide sulfurique et de la soude agrave 4 Malgreacute la reacutesistance des mycobacteacuteries aux agents
chimiques la deacutecontamination reacuteduit fortement la population initiale preacutesente dans lrsquoeacutechantillon et
abaisse la sensibiliteacute de la culture (Fine Bolin et al 2011)
Concernant lrsquoisolement de bacteacuteries du MTBC dans les matrices environnementales ou les
excreacutetas animaux les principales meacutethodes utiliseacutees deacutecoulent des meacutethodes citeacutees preacuteceacutedemment
La meacutethode drsquoimmunocapture magneacutetique utiliseacutee par Sweeney (2006 et 2007) a permis lrsquoisolement
de M bovis dans des sols de terriers et des latrines contamineacutes naturellement Cette meacutethode est
baseacutee sur lrsquoutilisation de microbilles magneacutetiques recouvertes avec un ligand speacutecifique drsquoun antigegravene
de lrsquoenveloppe de M bovis Son avantage est qursquoelle permet la capture et la concentration de la bacteacuterie
cible dans un eacutechantillon complexe sans alteacuterer sa viabiliteacute contrairement agrave la deacutecontamination
chimique Cette meacutethode a eacutegalement eacuteteacute testeacutee avec succegraves sur des broyats de nœuds lymphatiques
de bovins (Stewart et al 2012 Stewart et al 2013 Garbaccio amp Cataldi 2010)
b La culture sur milieu solide
Les milieux de culture solides agrave lrsquoœuf (milieux de Loumlwenstein-Jensen et milieu de Coletsos) sont
les plus utiliseacutes en France dans le diagnostic bacteacuteriologique chez lrsquohomme et lrsquoanimal Ils sont sensibles
et reacutepondent aux exigences culturales de la plupart des mycobacteacuteries tuberculeuses et non
tuberculeuses Le milieu de Coletsos est particuliegraverement indiqueacute pour la culture des mycobacteacuteries
exigeantes telles que M bovis et M africanum
Les milieux geacuteloseacutes Middlebrook 7H10 et 7H11 sont des milieux semi-syntheacutetiques non utiliseacutes
en diagnostic de routine en France mais tregraves utiliseacutes aux Etats-Unis Enrichis avec de lrsquoOADC meacutelange
drsquoacide oleacuteique drsquoalbumine bovine de dextrose de chlorure de sodium et de catalase ils permettent
une croissance plus rapide de M tuberculosis (Mejia et al 1999) et de M bovis (Idigoras et al 1995)
Chapitre 1
67
que les milieux agrave base drsquoœuf Lrsquoajout de seacuterum bovin et de sang de mouton lyseacute au milieu 7H11 enrichi
en OADC ameacuteliore fortement la croissance de M bovis agrave partir drsquoeacutechantillons animaux et raccourcit le
deacutelai drsquoobservation des colonies (Gallagher amp Horwill 1977) Ce milieu a eacuteteacute utiliseacute pour la mise en
culture drsquoeacutechantillons environnementaux tels que le sol ou les fegraveces apregraves un traitement preacutealable de
lrsquoeacutechantillon (Sweeney et al 2006 Fine OrsquoBrien et al 2011) mais son manque de seacutelectiviteacute neacutecessite
lrsquoajout drsquoun cocktail drsquoantibiotiques souvent composeacute de lrsquoassociation drsquoune carboxypeacutenicilline de
trimeacutethoprime et de polymyxine B et drsquoun antifongique (amphoteacutericine B) (Sweeney et al 2007 Fine
OrsquoBrien et al 2011) Lrsquoincubation des milieux solides ensemenceacutes est reacutealiseacutee agrave 37degC pendant au moins
8 semaines
c La culture en milieu liquide
Les milieux liquides ont eacuteteacute deacuteveloppeacutes pour pallier la croissance lente des mycobacteacuteries
drsquointeacuterecirct clinique sur les milieux solides afin de confirmer preacutecocement le diagnostic Diffeacuterents
systegravemes manuels ou automatiseacutes existent agrave lrsquoheure actuelle Ils sont essentiellement utiliseacutes dans le
diagnostic chez lrsquohomme plus rarement chez lrsquoanimal
Le systegraveme Bactectrade 460 (Becton Dickinson) baseacute sur la respiromeacutetrie radiomeacutetrique nrsquoest plus
utiliseacute en France mais il lrsquoa eacuteteacute pendant une vingtaine drsquoanneacutees Il eacutetait baseacute sur la mesure du dioxyde
de carbone marqueacute au 14C rejeteacute par les mycobacteacuteries en croissance dans un milieu contenant de
lrsquoacide palmitique marqueacute au 14C La libeacuteration de 14CO2 eacutetait mesureacutee reacuteguliegraverement de faccedilon manuelle
gracircce agrave un appareil adapteacute La contrainte drsquoutilisation de stockage et de recyclage de produits
radioactifs ainsi que lrsquoabsence drsquoautomatisation ont conduit agrave lrsquoarrecirct de sa commercialisation Son
utilisation a eacuteteacute deacutecrite pour la deacutetection de M bovis sur des tissus animaux et des fegraveces (Corner et al
2012 Thacker et al 2015)
Pour eacuteviter les inconveacutenients lieacutes agrave la radioactiviteacute des meacutethodes non radiomeacutetriques ont eacuteteacute
deacuteveloppeacutees La croissance bacteacuterienne en milieu liquide est mise en eacutevidence soit par la
consommation de lrsquooxygegravene soit par le deacutegagement de CO2 dans le tube La mise en eacutevidence de la
diminution de la pression partielle drsquooxygegravene dissous dans le milieu de culture lors de la croissance
bacteacuterienne est utiliseacutee dans les tubes BBLtrade MGITtrade de Becton Dickinson Lorsque lrsquoO2 diminue les sels
de rutheacutenium preacutesents au fond du tube eacutemettent une fluorescence Lrsquoutilisation de systegravemes
automatiseacutes tels que les systegravemes Bactectrade MGITtrade assurent une lecture en continue de la fluorescence
eacutemise Le systegraveme Bactectrade MGITtrade a eacuteteacute utiliseacutee pour la recherche de M bovis dans des broyats de
nœuds lymphatiques de bovins (Hines et al 2006 Stewart et al 2013) des tissus drsquoanimaux sauvages
(Murphy et al 2010 Witmer et al 2010) et dans du fromage frais (Harris et al 2007) Lrsquoaugmentation
du CO2 dans le milieu de culture lors de la croissance bacteacuterienne est agrave la base du systegraveme BacTALERT
Chapitre 1
68
3D de Biomeacuterieux Cette meacutethode colorimeacutetrique est baseacutee sur la diminution du pH suite agrave la libeacuteration
du CO2 Lrsquoautomate eacutevalue en continu le virage de couleur drsquoune pastille fixeacutee au fond du tube Les
systegravemes deacutecrits permettent eacutegalement la recherche de reacutesistance agrave diffeacuterents antibiotiques
(Piersimoni et al 2006) A notre connaissance cette technique nrsquoa jamais eacuteteacute utiliseacutee pour la culture
drsquoeacutechantillons environnementaux
Ces diffeacuterents systegravemes utilisent comme milieu liquide le Middlebrook 7H9 suppleacutementeacute en
OADC ou en ADC (Albumine bovine dextrose catalase) pour ameacuteliorer la croissance des mycobacteacuteries
et rendu plus seacutelectif par lrsquoajout drsquoun meacutelange drsquoantibiotiques et drsquoantifongiques pour eacuteviter la
croissance de bacteacuteries indeacutesirables Le plus utiliseacute est le PANTAtrade (polymyxine B amphoteacutericine acide
nalidixique trimeacutethoprime azlocilline) Il peut ecirctre compleacuteteacute avec de la vancomycine (effet contre les
bacteacuteries gram +) Lrsquoavantage majeur de ces systegravemes en milieu liquide est la diminution du deacutelai moyen
de deacutetection des mycobacteacuteries Avec le systegraveme Bactectrade M tuberculosis est deacutetecteacutee entre 8 et 30
jours en fonction de la charge bacteacuterienne initiale de lrsquoeacutechantillon La deacutetection de M bovis est reacutealiseacutee
en 15 jours contre environ 45 jours sur milieu solide Lorsque la culture est deacuteclareacutee positive la
preacutesence de BAAR doit ecirctre veacuterifieacutee par coloration sub-culture sur milieu de Loumlwenstein-Jensen
identification biochimique ou moleacuteculaire Lrsquoassociation de la culture en milieu liquide et solide donne
les meilleurs reacutesultats avec une limite de deacutetection de 10 agrave 102 bacillesmL drsquoeacutechantillon (Guillet-Caruba
et al 2014)
NB A la fin du 19egraveme siegravecle et au deacutebut du 20egraveme la discrimination des espegraveces M bovis M tuberculosis
et M avium se faisait principalement par inoculation de lrsquoisolat bacteacuterien au cobaye au lapin et au
poulet Des leacutesions pulmonaires heacutepatiques et spleacuteniques importantes chez le cobaye et le lapin
identifiaient M bovis des leacutesions retrouveacutees uniquement chez le cobaye ou le poulet identifiaient
respectivement M tuberculosis et M avium (Kazda et al 2009) Cette meacutethode eacutetait utiliseacutee pour
lrsquoidentification de M bovis dans les tissus animaux et les eacutechantillons environnementaux comme le sol
et lrsquoeau apregraves deacutecontamination avec un meacutelange de javel et de soude (Maddock 1933 Maddock 1934
Maddock 1936 Williams amp Hoy 1930)
3 Meacutethodes de deacutetection et drsquoidentification des bacteacuteries du MTBC
Pour des raisons eacutepideacutemiologiques sanitaires (isolement du malade pour eacuteviter la
transmission) et theacuterapeutiques (mise en place drsquoune antibiotheacuterapie adeacutequate) il est primordial de
diffeacuterencier les membres du MTBC des autres mycobacteacuteries et drsquoidentifier preacuteciseacutement lrsquoespegravece du
MTBC en cause
Chapitre 1
69
a Caracteacuterisation pheacutenotypique et biochimique des mycobacteacuteries du MTBC
Depuis la deacutecouverte du bacille de la tuberculose jusqursquoau deacuteveloppement des outils
modernes les diffeacuterentes espegraveces de mycobacteacuteries ont eacuteteacute identifieacutees gracircce agrave leurs caracteacuteristiques
pheacutenotypiques et biochimiques Certaines de ces caracteacuteristiques sont encore exploiteacutees agrave lrsquoheure
actuelle en compleacutement des meacutethodes de biologie moleacuteculaire plus rapides Lrsquoaspect des colonies sur
un milieu solide leur deacutelai drsquoapparition leur nombre leur aspect rugueux ou lisse et la preacutesence drsquoune
pigmentation ou non permet drsquoorienter lrsquoidentification vers une mycobacteacuterie du MTBC ou une
NTM Dans le MTBC par exemple M tuberculosis apparait en 20 jours sur milieux agrave lrsquoœuf sous forme
de colonies ivoire en chou-fleur drsquoaspect rugueux Les colonies de M bovis apparaissent tardivement
(6 agrave 10 semaines) elles sont non pigmenteacutees petites fines et lisses (Carbonnelle et al 2003) Les
principaux tests biochimiques utiliseacutes pour diffeacuterencier les diffeacuterentes espegraveces du MTBC sont le test agrave
la niacine la reacuteduction des nitrates lrsquohydrolyse du Tween la sensibiliteacute au TCH (acide thiophegravene-2-
carboxylique) agrave la cycloseacuterine et au pyrazinamide Les reacutesultats des observations pheacutenotypiques et
biochimiques permettant de diffeacuterencier les principales espegraveces du MTBC sont preacutesenteacutes dans le
Tableau 2 Les tests biochimiques neacutecessitent des cultures riches et ne sont donc pas reacutealisables avant
la croissance avanceacutee de la bacteacuterie Chronophages et fastidieux ces tests sont remplaceacutes par drsquoautres
techniques plus rapides
Tableau 2 Identification des principales mycobacteacuteries du MTBC par des observations pheacutenotypiques et des tests biochimiques
M tuberculosis M africanum M bovis M bovis BCG
Aspect des colonies Rugueuses Rugueuses Lisses Rugueuses
Pigmentation Non pigmenteacute Non pigmenteacute Non pigmenteacute Non pigmenteacute
Croissance Lente Lente Lente Lente
Niacin test + - - -
Reacuteduction nitrates + +- - -
TCH R S S S
Cycloseacuterine S S R R
Pyrazinamide S S R R
+ test positif - test neacutegatif R reacutesistant S sensible
b Lrsquoidentification des mycobacteacuteries par recherche de biomarqueurs
Diffeacuterentes techniques de recherche de biomarqueurs potentiels de M tuberculosis par
chromatographie ont eacuteteacute deacuteveloppeacutees reacutecemment La chromatographie en phase liquide (HPLC High-
Chapitre 1
70
Performance Liquid Chromatography) a eacuteteacute deacuteveloppeacutee pour lrsquoanalyse des acides mycoliques (Butler amp
Guthertz 2001) La chromatographie en phase gazeuse coupleacutee agrave de la spectromeacutetrie de masse (GC-
MS) est utiliseacutee pour identifier des acides gras speacutecifiques de la paroi de M tuberculosis comme lrsquoacide
tuberculosteacutearique ou lrsquoacide hexacosanoiumlque dans des crachats (Cha et al 2009 Kaal et al 2009)
Lrsquoidentification de biomarqueurs speacutecifiques permet de diffeacuterencier M tuberculosis des NTM en GC-
MS en diagnostic direct sur des crachats (Dang et al 2013)
Depuis quelques anneacutees la spectromeacutetrie de masse de type MALDI-TOF (matrix-assisted laser
desorption ionization-time of flight) est utiliseacutee pour lrsquoidentification rapide des mycobacteacuteries du
complexe tuberculosis et de la majoriteacute des autres mycobacteacuteries agrave partir de cultures en milieu solide
et liquide (El Kheacutechine et al 2011 Bille et al 2012 Lotz et al 2010 Biswas amp Rolain 2013) Cette
technique permet drsquoeacutetablir un spectre de masse des diffeacuterentes proteacuteines constituant la mycobacteacuterie
(profil peptidique obtenu apregraves digestion trypsique des proteacuteines totales) La comparaison du profil
proteacuteique obtenu avec des bases de donneacutees internationales de plus en plus eacutetoffeacutees permet
drsquoidentifier un grand nombre drsquoespegraveces de mycobacteacuteries Elle peut ecirctre eacutegalement utiliseacutee comme
meacutethode de spoligotypage (Shitikov et al 2012) Cette meacutethode est simple rapide reproductible et
neacutecessite des consommables peu coucircteux (hors achat et entretien de lrsquoappareil) La qualiteacute du profil
proteacuteique obtenu est toutefois fortement deacutependante de la qualiteacute du protocole drsquoextraction des
proteacuteines (Saleeb et al 2011) La technique MALDI-TOF est probablement une technique drsquoavenir pour
lrsquoidentification rapide des mycobacteacuteries agrave croissance lente (le deacutelai de croissance reste toutefois le
facteur limitant) et plus geacuteneacuteralement des microorganismes pathogegravenes drsquointeacuterecirct clinique dans les
eacutechantillons A lrsquoheure actuelle elle ne permet pas de diffeacuterencier les diffeacuterentes espegraveces du MTBC
Elle nrsquoest pour lrsquoinstant applicable qursquoagrave des isolats bacteacuteriens issus de culture et non agrave des eacutechantillons
complexes
c Lrsquoidentification des mycobacteacuteries par des meacutethodes drsquohybridation coupleacutees ou
non agrave la PCR
La technique baseacutee sur lrsquohybridation de lrsquoADN est une meacutethode sensible rapide et simple
drsquoemploi couramment utiliseacutee en bacteacuteriologie humaine Elle est reacutealiseacutee sur une culture liquide ou
solide La technique est baseacutee sur lrsquohybridation drsquoune sonde moleacuteculaire conjugueacutee agrave un marqueur
chimioluminescent compleacutementaire de lrsquoARN ribosomal de la bacteacuterie cible Le kit commercial
AccuProbe (Gen Probe) fournit diffeacuterentes sondes permettant drsquoidentifier les Complexes tuberculosis
avium gordonae kansaii (sans discrimination des espegraveces) ainsi que les espegraveces M avium et
M intracellulare
Le systegraveme le plus utiliseacute agrave lrsquoheure actuelle est le GenXpert MTBRIF (Cepheid) utilisant la PCR
Chapitre 1
71
en temps reacuteel multi-colorimeacutetrique Il utilise 5 sondes Beacon (sondes avec une structure en eacutepingle agrave
cheveux qui fluorescent lors de lrsquohybridation) speacutecifiques du locus rpoB (reacutegion de 81 pb deacuteterminant
la reacutesistance agrave la rifampicine) des membres du MTBC Lrsquointeacuterecirct de cette technique est agrave la fois pratique
puisque lrsquoopeacuterateur deacutepose lrsquoeacutechantillon deacutecontamineacute dans une cassette contenant tous les reacuteactifs
lyophiliseacutes lrsquoappareil gegravere toutes les autres eacutetapes et diagnostique car elle couple la deacutetection des
bacteacuteries du MTBC et la mise en eacutevidence drsquoune eacuteventuelle reacutesistance agrave la rifampicine antituberculeux
de premiegravere ligne chez lrsquohomme
Drsquoautres systegravemes tels que lrsquoINNO-Lipareg Mycobacteria v2 (Fujirebio) le GenoType MTBC et le
GenoType Mycobacterium CMAS (Hain Lifescience) sont baseacutes sur lrsquohybridation drsquoune sonde
speacutecifique drsquoun Complexe ou drsquoune espegravece sur la reacutegion intergeacutenique variable 16S-23S apregraves
amplification PCR Ils permettent drsquoidentifier les Complexes tuberculosis et avium ainsi que de 16 agrave 40
espegraveces de NTM
Ces techniques sont essentiellement utiliseacutees dans le diagnostic humain mais elles sont parfois
deacutecrites pour lrsquoidentification du Complexes tuberculosis en diagnostic animal (Oloya et al 2007 Pavlik
et al 2005 Bollo et al 1998 Lieacutebana et al 1995) Bien que deacutejagrave utiliseacutees sur des eacutechantillons
environnementaux comme lrsquoeau (Aronson et al 1999) elles sont agrave utiliser avec preacutecaution car des
erreurs drsquoidentification ont eacuteteacute rapporteacutees (croisement entre des mycobacteacuteries saprophytes et
pathogegravenes) (Turenne 2004) en outre elles ne permettent pas la discrimination des diffeacuterentes
espegraveces du MTBC
d Le seacutequenccedilage
Lrsquooutil geacutenotypique de reacutefeacuterence pour lrsquoidentification des diffeacuterentes espegraveces de bacteacuteries est
le seacutequenccedilage du gegravene codant lrsquoARN de la sous-uniteacute 16S du ribosome (gegravene rrs) De nombreux couples
drsquoamorces ont eacuteteacute proposeacutes dans la litteacuterature pour amplifier entiegraverement ou partiellement le gegravene rrs
avant seacutequenccedilage mais lrsquoidentification des mycobacteacuteries par cette meacutethode nrsquoest pas toujours
possible agrave cause de la forte conservation de cette seacutequence Afin drsquoameacuteliorer la discrimination de ces
espegraveces le seacutequenccedilage total ou partiel drsquoautres gegravenes cibles a eacuteteacute deacuteveloppeacute tels les gegravenes de meacutenage
ciblant rpoB codant la sous-uniteacute β de lrsquoARN polymeacuterase (Adeacutekambi amp Drancourt 2004 Lee et al 2000)
hsp65 codant une proteacuteine de reacuteponse au stress thermique de 65 kDa (McNabb et al 2004 Ringuet et
al 1999 Telenti et al 1993) gyrA et gyrB codant les sous-uniteacutes α et β de lrsquoADN gyrase (enzyme
intervenant dans lrsquoenroulement et le deacuteroulement de lrsquoheacutelice drsquoADN) recA intervenant dans la
recombinaison homologue de lrsquoADN et dans sa reacuteparation (Blackwood et al 2000) sodA codant la super
oxyde dismutase (Zolg amp Philippi-Schulz 1994) secA1 codant la proteacuteine SecA1 intervenant dans le
seacutecreacutetion des proteacuteines agrave travers la membrane plasmique (Zelazny et al 2005) ou la reacutegion inter-
Chapitre 1
72
geacutenique encadreacutee par les ARNr 16S et 23S (appeleacutee ITS Internal Transcribed Spacer) (Roth et al 1998)
Malgreacute une ameacutelioration de la discrimination par lrsquoanalyse combineacutee de ces diffeacuterentes cibles le
seacutequenccedilage ne permet pas drsquoidentifier toutes les diffeacuterentes bacteacuteries du MTBC A cause de la forte
conservation des seacutequences dans le MTBC le seacutequenccedilage drsquoun grand nombre de gegravenes est neacutecessaire
pour discriminer les espegraveces (Kirschner et al 1993 Boumlddinghaus et al 1990) ce qui nrsquoest pas reacutealisable
en routine
Depuis lrsquoidentification de la plupart des gegravenes et des mutations responsables de la reacutesistance
des mycobacteacuteries aux antituberculeux le seacutequenccedilage est un des moyens utiliseacute pour identifier les
souches reacutesistantes La reacutesistance agrave la rifampicine par exemple est lieacutee agrave plusieurs mutations sur le gegravene
rpoB modifiant la structure de la sous-uniteacute β de lrsquoARN polymeacuterase et empecircchant la fixation de la
rifampicine sur lrsquoARN polymeacuterase conduisant agrave son inefficaciteacute (Zhang amp Yew 2009) Ces mutations
peuvent ecirctre deacutetecteacutees par seacutequenccedilage du gegravene amplifieacute par PCR
e Deacutetection et geacutenotypage des mycobacteacuteries du MTBC les marqueurs utiliseacutes
Les seacutequences drsquoinsertion
Les seacutequences drsquoinsertion (IS) correspondent agrave de petits eacuteleacutements geacuteneacutetiques transposables
Elles contiennent uniquement les informations geacuteneacutetiques neacutecessaires agrave leur transposition dans le
geacutenome de la bacteacuterie (transposase) On deacutenombre 42 IS dans le premier geacutenome annoteacute de M bovis
AF212297 (Garnier et al 2003) La seacutequence drsquoinsertion IS6110 speacutecifique du MTBC est la cible
historique utiliseacutee pour la deacutetection moleacuteculaire du Complexe tuberculosis en PCR (Thierry Cave et al
1990 Thierry Brisson-Noeumll et al 1990 Dziadek et al 2001) La meacutethode de reacutefeacuterence pour le
geacutenotypage de M tuberculosis est la RFLP-IS6110 (Restriction Fragment Lenght Polymorphism sur
lrsquoIS6110) Crsquoest la seacutequence drsquoinsertion la plus abondante dans le geacutenome de M tuberculosis qui en
contient de 1 agrave plus de 25 copies (Thierry Cave et al 1990 Thierry Brisson-Noeumll et al 1990) Elle
jouerait un rocircle dans lrsquoadaptation des bacteacuteries du MTBC agrave leur hocircte et dans leur virulence (Otal et al
2008 Safi et al 2004)
De nombreux systegravemes drsquoamorces ont eacuteteacute deacuteveloppeacutes pour la deacutetection de lrsquoIS6110 par PCR
dans les eacutechantillons biologiques (Dziadek et al 2001 El Kheacutechine et al 2009 Lieacutebana et al 1996
Savelkoul et al 2006) Diffeacuterents kits commerciaux utiliseacutes agrave lrsquoheure actuelle sont baseacutes sur la deacutetection
de lrsquoIS6110 notamment dans le diagnostic animal (Courcoul et al 2014) Cette cible a deacutejagrave eacuteteacute utiliseacutee
pour la deacutetection de M bovis dans des eacutechantillons inoculeacutes en PCR nicheacutee (Adams et al 2013) ainsi
que pour la deacutetection moleacuteculaire du MTBC dans du sol de lrsquoeau des seacutediments et des matiegraveres feacutecales
drsquoanimaux (Santos Santos et al 2015) (Tableau 3)
Chapitre 1
73
LrsquoIS6110 est eacutegalement utiliseacutee pour le typage des bacteacuteries du MTBC La comparaison des
souches repose sur le nombre drsquoIS6110 et leurs positions sur le chromosome apregraves extraction de lrsquoADN
digestion par une endonucleacutease (PvuII qui coupe lrsquoIS6110 sur un seul site) eacutelectrophoregravese transfert
sur membrane et hybridation (Van Soolingen et al 1991) Chaque fragment contenant une copie de
lrsquoIS6110 produit une bande (Figure 16) Cette meacutethode standardiseacutee beacuteneacuteficie de banques de donneacutees
de profils drsquohybridation internationales qui permettent des investigations eacutepideacutemiologiques Cette
meacutethode est beaucoup moins discriminante sur les souches agrave faible nombre drsquoIS (Allix et al 2006)
comme M bovis qui nrsquoen contient qursquoune agrave cinq copies et M bovis BCG qui nrsquoen contient qursquoune seule
copie (Van Soolingen et al 1991 Skuce et al 1994) Le reacutesultat est obtenu en une dizaine de jours
neacutecessite un volume important de culture et des manipulations fastidieuses De ce fait il a eacuteteacute
remplaceacute par des meacutethodes plus rapides indeacutependantes de la culture le spoligotypage et le typage
MLVA
Figure 16 Profils de restriction disolats de M tuberculosis obtenus suite agrave une RFLP-IS6110 Les isolats 3 5 6 9 et 10 ont le mecircme profil et posseacutedaient un lien eacutepideacutemiologique S repreacutesente le marqueur de poids moleacuteculaire (Source web site CDC)
Les courtes seacutequences drsquoADN reacutepeacuteteacutees
Le locus DR
Les souches du Complexe tuberculosis contiennent une reacutegion chromosomique particuliegravere
unique et figeacutee caracteacuteriseacutee par des seacutequences reacutepeacuteteacutees de 36 pb appeleacutee locus DR (Direct Repeat)
qui appartient agrave la famille des ADN reacutepeacutetitifs CRISPR (Clustered Regularly Interspersed Short
Chapitre 1
74
Palindromic Repeats) Ces DR sont seacutepareacutes par des espaceurs de 35 agrave 41 pb dont les seacutequences sont
uniques et polymorphes Ce polymorphisme associeacute agrave la deacuteleacutetion de certains espaceurs par des
reacutearrangements geacuteneacutetiques est exploiteacute dans le spoligotypage (en anglais spoligotyping
SPacer OLIGOnucleacuteotide TYPING) (Kamerbeek et al 1997) Par exemple les espaceurs 3 9 16 et 39 agrave
43 sont absents chez M bovis M caprae M microti et M pinnipedii (Smith et al 2006) comme
preacutesenteacute dans la Figure 17 Le geacutenotypage par spoligotypage est baseacute sur lrsquoamplification par PCR du
locus DR le marquage des amplicons et leur hybridation par des oligonucleacuteotides fixeacutes agrave une
membrane (spoligotypage 2D) Le profil final est deacutetecteacute par chimioluminescence (Kamerbeek et al
1997) Dans la technologie Luminex les oligonucleacuteotides sont fixeacutes sur des microbilles agrave signature
spectrale diffeacuterente (spoligotypage 3D) (Zhang et al 2010) Le spolygotypage standard analyse 43
espaceurs et permet drsquoobtenir des donneacutees eacutepideacutemiologiques et phylogeacuteneacutetiques deacuteterminantes (Sola
et al 2001) Il est largement utiliseacute pour typer les souches de M bovis notamment en France (Haddad
et al 2001) Cette meacutethode est reacutealisable sur de faibles quantiteacutes drsquoADN mecircme impur et des bacteacuteries
non cultivables Crsquoest une technique robuste rapide simple de reacutealisation dont le reacutesultat est facile agrave
interpreacuteter car il fournit un code binaire (pour chaque espaceur preacutesent 1 ou absent 0) faisant lrsquoobjet
drsquoune convention internationale (Dale et al 2001) Toutefois le spoligotypage ne preacutesente pas un
pouvoir de reacutesolution suffisant pour la diffeacuterenciation de certains groupes clonaux car il ne cible qursquoun
seul locus sur lrsquoensemble du geacutenome
Chapitre 1
75
C
Figure 17 Principe du spoligotypage des bacteacuteries du MTBC dapregraves Kamerbeek et al (1997) A Repreacutesentation du locus DR de M tuberculosis H37Rv et M bovis BCG B Amplifications des espaceurs par un couple damorces (a et b) et amplicons obtenus C Profils drsquohybridation sur membrane (un carreacute noir signale la preacutesence de lrsquoespaceur les parties blanches leur absence)
76
Gegravene ou locus
cible Espegraveces cibles PCR
Taille du
produit PCR
(pb)
Limites de deacutetection
(bacteacuteries g-1 sol
ou mL-1)
Matrices analyseacutees
Quantification si
deacutetection positive (nb
gegravenes ou UFC g-1 sol
ou mL-1)
Reacutefeacuterence Utiliseacute par
mpb64 MTBC PCR classique 543 101-102 Sols (terriers et preacutes
adjacents) 10 - 25 times 103 (Gormley 1999) (Young et al 2005)
mpb70 MTBC PCR classique 471 101-102 Sols (terriers et preacutes
adjacents) 10 - 25 times 103 (Gormley 1999)
(Young et al 2005)
(Courtenay et al 2006)
Gegravene codant
lrsquoARNr 16S
Mycobacteacuteries
agrave croissance
lente
RT-PCR 605 102-103 Sols (terriers et preacutes
adjacents) -
(Young et al
2005)
(Young et al 2005)
(Courtenay et al 2006)
IS6110 MTBC PCR nicheacutee PCR1 252
PCR2 116 -
Sol Eau
Foin Maiumls -
(Adams et al
2013) (Adams et al 2013)
IS6110 MTBC PCR nicheacutee 110
Sol 40 times 104
Eau 50 times 105
Sol
Seacutediment
Eau et fegraveces
(Soo et al 2006) (Santos Santos et al 2015)
RD12 M bovis
M caprae PCR classique 306
Sol 10 times 106
Eau 50 times 105
Sol
Seacutediment
Eau et fegraveces
(Warren et al
2006) (Santos Santos et al 2015)
RD12
M microti
M tuberculosis
M africanum
M pinnipedii
PCR semi-
nicheacutee 369
Sol 40 times 104
Eau 50 times 105
Sol
Seacutediment
Eau et fegraveces
(Warren et al
2006) (Santos Santos et al 2015)
RD4 (zone
flanquante) M bovis qPCR 142
42 times 105
(Pontiroli et al 2011)
10 times 105
(Pontiroli et al 2011)
Sols de terriers
Latrines
68 times 104 agrave 54 times 106
(Sweeney et al 2007)
20 times 104 agrave 10 times 108
(Travis et al 2011)
(Sweeney et al
2007)
(Sweeney et al 2007)
(King Murphy James Travis
Porter Sawyer et al 2015)
(King Murphy James Travis
Porter Hung et al 2015)
(Pontiroli et al 2011)
(Roug et al 2014)
Rv1510 (RD4) MTBC sauf
M bovis
PCR classique 1 033 - Sols de terriers
Latrines
- (Huard et al
2003)
(Courtenay et al 2006)
(Sweeney et al 2006)
(Sweeney et al 2007)
Tableau 3 Cibles moleacuteculaires deacutecrites dans la litteacuterature pour la deacutetection du Complexe tuberculosis ou de M bovis dans les eacutechantillons environnementaux
Chapitre 1
77
Les mini-satellites
Les mini-satellites correspondent agrave de courtes seacutequences drsquoADN reacutepeacuteteacutees (de 10 agrave 1 000 pb) et
conseacutecutives Pour pallier le manque de discrimination du spoligotypage un typage sur diffeacuterents loci
a eacuteteacute deacuteveloppeacute le MLVA (Multi Locus VNTR Analysis) baseacute sur lrsquoanalyse drsquoeacuteleacutements situeacutes sur des
seacutequences intergeacuteniques (les MIRU Mycobacterial interspersed Repetitive Unit) composeacutes de
seacutequences reacutepeacuteteacutees en tandem dont le nombre et la seacutequence sont variables (notamment les VNTR
Variable Number Tandem Repeat et les ETR Exact Tandem Repeat) (Supply et al 2001 Supply et al
2006) Lrsquoeacutetude de loci disperseacutes dans le geacutenome apporte une discrimination robuste des souches Le
geacutenotypage des souches de M tuberculosis ou M bovis par la meacutethode MIRU-VNTR repose sur la
recherche du nombre drsquoexemplaires des seacutequences reacutepeacuteteacutees de chaque locus eacutetudieacute Plus le nombre
de loci analyseacutes est important plus la discrimination est fine ainsi le MIRU-VNTR peut porter sur 12
15 ou 24 loci pour le typage des souches de M tuberculosis selon lrsquoobjectif envisageacute (eacutepideacutemiologie ou
phylogeacutenie) (Supply et al 2001 Supply et al 2006) Concernant le typage des souches animales de
M bovis diffeacuterentes combinaisons de marqueurs VNTR ont eacuteteacute eacutevalueacutees (Roring et al 2002 Allix et al
2006 Skuce et al 2002 Boniotti et al 2009) et ont montreacute que lrsquoanalyse de 5 agrave 8 loci suffisait pour
obtenir un pouvoir de discrimination supeacuterieur agrave 90 En France 8 loci ont eacuteteacute seacutelectionneacutes sur leur
polymorphisme et leur pouvoir de discrimination et utiliseacutes pour typer plus de mille isolats cliniques
recueillis entre 1978 et 2011 (Hauer et al 2015) Dans cette eacutetude les reacutesultats de geacutenotypage se
preacutesentent sous forme drsquoun code agrave 8 chiffres (Tableau 4) Pour un mecircme spoligotype SB0120 le
geacutenotypage MIRU-VNTR permet de diffeacuterencier les souches retrouveacutees en Cocircte drsquoOr de celles de
Dordogne Ces reacutesultats ont eacutegalement permis de deacutemontrer lrsquohomogeacuteneacuteiteacute des profils geacuteneacutetiques
retrouveacutes chez les animaux domestiques et sauvages dans une zone geacuteographique confirmant
lrsquohypothegravese de la transmission interspeacutecifique de lrsquoinfection
Tableau 4 Spoligotypes et geacutenotypes MIRU-VNTR des souches de M bovis isoleacutees chez les bovins et la faune sauvage en Cocircte dOr et en Dordogne (drsquoapregraves Hauer et al 2015)
Spoligotype Geacutenotype MIRU-VNTR (8 loci cibleacutes)
Origine des isolats 2165 2461 0577 580 2163a 2163b 4052 3232
SB0120
Cocircte drsquoOr 5 5 4 3 11 4 5 6
Bovins blaireaux
sangliers
SB0120
Dordogne 5 3 5 3 9 4 5 6
Bovins blaireaux
sangliers
SB0134
Cocircte drsquoOr 6 4 5 3 6 4 3 6
Bovins blaireaux
sangliers cerfs
Chapitre 1
78
La combinaison du spoligotypage et de la MLVA optimise la discrimination des souches pour
les suivis eacutepideacutemiologiques et apporte des informations phylogeacuteneacutetiques inteacuteressantes Elle permet
eacutegalement de suivre lrsquoeacutemergence de variants clonaux chez un animal ou dans un troupeau infecteacute en
fonction du temps (Navarro et al 2015 Rodriguez-Campos et al 2014) Toutefois les loci agrave analyser
doivent ecirctre adapteacutes en fonction des complexes clonaux rencontreacutes dans la reacutegion ou le pays eacutetudieacutes
(Boniotti et al 2009 Hauer et al 2015) Diffeacuterentes bases de donneacutees internationales sont disponibles
pour la comparaison des reacutesultats de spoligotypage et de MIRU-VNTR comme SpolDB4 SITVITWEB
MIRU-VNTRplus et Mbovisorg (regroupe les spolygotypes des espegraveces deacuteleacuteteacutees pour RD9) (Couvin amp
Rastogi 2014 Smith amp Upton 2012)
Reacutegions de Diffeacuterence (RD) et Single Nucleotide
Polymorphism (SNP)
Les Reacutegions de Diffeacuterences (RD1 agrave RD14) identifieacutees chez les bacteacuteries du MTBC correspondent
agrave des deacuteleacutetions et des reacutearrangements geacuteneacutetiques irreacuteversibles et discriminants Ces RD sont
speacutecifiques drsquoune espegravece ou communes agrave plusieurs espegraveces Le plus grand nombre de RD est observeacute
dans le geacutenome de M bovis BCG qui preacutesente 14 RD par comparaison agrave M tuberculosis H37Rv (Gordon
et al 1999 Brosch et al 2002) En parallegravele six seacutequences deacuteleacuteteacutees chez M tuberculosis H37Rv (les
RvD) et une seacutequence absente de toutes les souches de M tuberculosis (TbD1) sont preacutesentes chez les
autres bacteacuteries du MTBC (Gordon et al 1999 Brosch et al 2002) Lrsquoabsence de RD4 est speacutecifique agrave
M bovis et M bovis BCG et permet de les diffeacuterencier des autres membres du MTBC (Huard et al 2003
Huard et al 2006) La RD4 a eacuteteacute utiliseacutee dans diffeacuterentes eacutetudes pour la deacutetection de M bovis dans les
eacutechantillons environnementaux (Courtenay et al 2006 Sweeney et al 2007 King Murphy James
Travis Porter Sawyer et al 2015 Travis et al 2011 Roug et al 2014 Pontiroli et al 2011) Dans une
eacutetude reacutecente Santos et al (2015) ont associeacute la deacutetection du MTBC par lrsquoIS6110 agrave une identification
de lrsquoespegravece par amplification de deux reacutegions de la RD12 pour diffeacuterencier M bovisM caprae des
autres espegraveces du Complexe (Tableau 3)
Lrsquointeacuterecirct des RD est bien plus phylogeacuteneacutetique que geacutenotypique puisque lrsquoeacutetude de leur
apparition successive au cours de lrsquoeacutevolution a permis drsquoeacutetablir preacuteciseacutement la phylogeacutenie des
diffeacuterentes espegraveces comme preacutesenteacute preacuteceacutedemment dans la Figure 3 (Huard et al 2006 Rodriguez-
Campos et al 2014 Dippenaar et al 2015)
Les Single Nucleotide Polymorphism (SNPs) correspondent agrave des mutations ponctuelles drsquoune
base dans un geacutenome Ils peuvent ecirctre synonymes (le changement de base nrsquoaffecte pas la seacutequence
proteacuteique correspondante) ou non synonymes (la seacutequence proteacuteique est affecteacutee) et repreacutesentent
Chapitre 1
79
une variabiliteacute du geacutenome utile pour la caracteacuterisation de la variabiliteacute drsquoun groupe clonal et lrsquoeacutetude
des relations phylogeacuteneacutetiques entre les souches (Mathema et al 2006)
f Identification par seacutequenccedilage complet des geacutenomes
Le seacutequenccedilage complet des geacutenomes (WGS Whole Genome Sequencing) beacuteneacuteficie depuis
quelques anneacutees de nouvelles technologies (NGS Next Generation Sequencing) plus rapides et moins
coucircteuses que la meacutethode de Sanger Diffeacuterentes technologies coexistent agrave lrsquoheure actuelle dont les
plus utiliseacutees sont le pyroseacutequenccedilage 454 (toutefois en fin drsquoutilisation) la technique Illumina et la
technique Ion PGM (Ion Personal Genome Machine) (Desikan amp Narayanan 2015) Ces techniques ont
reacutevolutionneacute le geacutenotypage en fournissant le plus haut niveau de discrimination geacuteneacutetique possible
Ces techniques parfois coupleacutees au typage des SNPs permettent de suivre la dynamique de
transmission drsquoune maladie de caracteacuteriser lrsquoantibioreacutesistance drsquoune souche et drsquoeacutevaluer la divergence
geacuteneacutetique de souches qui ont les mecircmes profils de restriction ou de geacutenotypage (Schuumlrch amp van
Soolingen 2012) Une eacutetude irlandaise sur des souches de M bovis isoleacutees de bovins et de blaireaux
vivant dans les mecircmes zones a permis gracircce au WGS et agrave lrsquoanalyse de 39 SNPs de deacutemontrer la
circulation et la persistance locales drsquoune souche entre animaux sauvages et domestiques (Biek et al
2012) Une eacutetude reacutecente portant sur le seacutequenccedilage complet de 82 souches de M bovis isoleacutees en
France entre 1978 et 2011 a permis de deacutemontrer la congruence entre le typage par
spoligotypageMIRU-VNTR et le typage des souches par analyse des SNP (Hauer 2015) suggeacuterant que
certains SNP pourraient ecirctre utiliseacutes comme cibles dans le geacutenotypage de M bovis Dans un futur
proche le WGS sera probablement lrsquooutil qui offrira lrsquoinformation la plus complegravete sur le geacutenotype des
souches les liens phylogeacuteneacutetiques entre souches et qui permettra de deacutecoder les scheacutemas de
transmission de lrsquoinfection entre animaux (Biek et al 2012 Pepperell et al 2013 Rodriguez-Campos et
al 2014)
Les puces (microarrays) agrave ADN ou ARN sont utiliseacutees pour la comparaison de geacutenomes complets
des espegraveces du MTBC et ont notamment apporteacute des informations sur lrsquoexpression de gegravenes de la
virulence chez M bovis (Voskuil et al 2004) ainsi que sur lrsquoidentification de seacutequences polymorphiques
distinguant les principaux complexes clonaux (Berg et al 2011 Muumlller et al 2009 Smith et al 2011)
Ces techniques appartiennent toutefois au domaine de la recherche et ne sont pas utiliseacutees en routine
g Meacutethode speacutecifique drsquoidentification des mycobacteacuteries animales utiliseacutee par le
Laboratoire National de reacutefeacuterence de la tuberculose bovine LNR TB
Lrsquoidentification des mycobacteacuteries isoleacutees sur des animaux est reacutealiseacutee au LNR TB de Maisons-
Chapitre 1
80
Alfort En premiegravere intention quatre systegravemes de deacutetection moleacuteculaires ciblant des seacutequences
drsquoinsertion (IS6110 IS1081 IS1245) et le gegravene atpE sont utiliseacutes en qPCR pour orienter lrsquoidentification
(Tableau 5)
Tableau 5 Systegravemes de deacutetection utiliseacutes en qPCR en premiegravere intention pour lidentification dune mycobacteacuterie dorigine animale et reacutesultats obtenus + amplification de la cible - absence drsquoamplification de la cible
ARNr 16S IS1245 IS901 IS6110 IS1081 IS1561rsquo RD4 RD7 atpE
Complexe tuberculosis
M bovis M tuberculosis
M microti M caprae
+ + + +
- - - -
- - - -
+ + + +
+ + + +
+ + - +
- + + +
- + + -
- - - -
Complexe avium + + +- - - - - + +
Autres mycobacteacuteries
+ - - - - - - + +
(Slana et
al 2010) (Slana et al
2010) (Courcoul et
al 2014) (cette thegravese)
(Radomski et al 2013)
Lrsquoidentification des espegraveces du complexe tuberculosis (IS6110 et IS1081 positives) est ensuite
reacutealiseacutee par geacutenotypage gracircce agrave la technique de spoligotypage (Zhang et al 2010) Lrsquoamplification ou
non des IS1245 et IS901 permet drsquoidentifier les espegraveces Mah (IS1245 et IS901 neacutegatives) Maa (IS1245
et IS901 positives) et Mah (IS1245 positive et IS901 neacutegative) (Slana et al 2010) Lrsquoidentification des
autres mycobacteacuteries repose sur le seacutequenccedilage de lrsquohsp65 puis de la comparaison par BLAST avec une
base de donneacutees (Telenti et al 1993)
De par le grand nombre drsquoespegraveces de mycobacteacuteries et la forte identiteacute des espegraveces dans le
Complexe MTBC la deacutetection et le geacutenotypage des souches du MTBC reposent obligatoirement sur
lrsquoutilisation conjointe de diffeacuterentes techniques En fonction des eacutechantillons disponibles et des
objectifs souhaiteacutes qursquoils soient diagnostiques eacutepideacutemiologiques ou phylogeacuteneacutetiques le choix des
techniques agrave utiliser doit ecirctre reacutefleacutechi et pertinent
82
Objectifs de la Thegravese
Depuis 2004 la situation de la Cocircte drsquoOr vis-agrave-vis de la tuberculose bovine est preacuteoccupante
Malgreacute la mise en place de moyens de surveillance et de lutte en premier lieu dans les cheptels bovins
puis dans la faune sauvage lrsquoeacuteradication de la maladie semble difficile Depuis 2011 on observe une
diminution du nombre des foyers bovins et une stabiliteacute du nombre de cas observeacutes dans la faune
sauvage avec une concentration spatiale des foyers dans des petites valleacutees Dans ces valleacutees ougrave
cohabitent faune sauvage et bovins la transmission indirecte du bacille entre animaux sauvages et
domestiques par le biais de lrsquoenvironnement est fortement suspecteacutee pour plusieurs cas bovins Bien
que plusieurs eacutetudes aient deacutemontreacute la preacutesence de M bovis dans diffeacuterentes matrices
environnementales naturelles sur les lieux de vie drsquoanimaux domestiques et sauvages infecteacutes
notamment en Angleterre et en Espagne aucune donneacutee nrsquoest disponible sur la contamination
environnementale potentielle par M bovis dans les zones endeacutemiques de tuberculose bovine en
France et plus particuliegraverement en Cocircte drsquoOr
Pour reacutepondre agrave cette question le premier objectif du travail de thegravese a eacuteteacute de deacutevelopper et de valider
sur des eacutechantillons inoculeacutes artificiellement des outils moleacuteculaires speacutecifiques et sensibles pour la
deacutetection des souches du MTBC et de M bovis dans les substrats environnementaux naturellement
contamineacutes tels que le sol lrsquoeau et les seacutediments et les matiegraveres feacutecales animales Nous avons
eacutegalement deacuteveloppeacute un protocole drsquoextraction-deacutecontamination des mycobacteacuteries du sol coupleacute agrave
une technique originale de suivi de croissance des mycobacteacuteries en milieu liquide dans lrsquoobjectif
drsquoisoler les souches environnementales de M bovis (Chapitre 2)
Les outils moleacuteculaires deacuteveloppeacutes ont eacuteteacute utiliseacutes pour la recherche de M bovis dans un grand nombre
drsquoeacutechantillons environnementaux (sol eau seacutediments fegraveces animales et membres de la micro- ou
meacuteso-faune des sols) preacuteleveacutes dans la zone endeacutemique de Cocircte drsquoOr et plus preacuteciseacutement dans des
zones partageacutees par la faune domestique et sauvage infecteacutee afin de preacuteciser le niveau de
contamination des lieux de vie des bovins et des animaux sauvages (Chapitre 3)
Face aux eacutetudes parfois controverseacutees sur le rocircle des facteurs abiotiques dans la persistance de
M bovis dans le sol nous avons rechercheacute lrsquoeffet de la nature du sol et de la tempeacuterature sur la dureacutee
de survie de M bovis SB0120 en conditions de laboratoire (Chapitre 4)
Nous avons enfin eacutetudieacute le rocircle de la faune du sol et plus particuliegraverement des lombrics tregraves abondants
dans les pacirctures et gros consommateurs de matiegravere organique dans la circulation et la disseacutemination
de M bovis dans les sols (Chapitre 5)
CHAPITRE 2
Deacuteveloppement et optimisation drsquooutils pour la deacutetection
de Mycobacterium bovis dans les eacutechantillons
environnementaux
PARTIE CONFIDENTIELLE agrave lrsquoexception des meacutethodes de deacutetection moleacuteculaire
86
CHAPITRE 2 - Deacuteveloppement et optimisation drsquooutils pour la
deacutetection de Mycobacterium bovis dans les eacutechantillons
environnementaux
A DEVELOPPEMENT DrsquoOUTILS MOLECULAIRES ADAPTES AUX MATRICES ENVIRONNEMENTALES
Les diffeacuterents systegravemes moleacuteculaires utiliseacutes pour la deacutetection de M bovis dans
lrsquoenvironnement citeacutes dans la litteacuterature sont syntheacutetiseacutes dans le Tableau 3 En consideacuterant la diversiteacute
et lrsquoabondance des mycobacteacuteries potentiellement preacutesentes dans lrsquoenvironnement nous avons
eacutelimineacute les systegravemes ciblant toutes les espegraveces du MTBC notamment lrsquoIS6110 car bien que tregraves
sensible et speacutecifique sur les eacutechantillons biologiques et tregraves utiliseacutee en diagnostic humain et
veacuteteacuterinaire nous avons montreacute qursquoelle nrsquoeacutetait pas utilisable en diagnostic environnemental En effet
nous avons effectueacute des essais preacuteliminaires avec le kit LSI VetMAXtrade Mycobacterium tuberculosis PCR
Kit (Life Technologies) utiliseacute en diagnostic veacuteteacuterinaire sur 25 eacutechantillons de sol provenant de zones
geacuteographiques non impacteacutees par la tuberculose bovine Pour tous ces sols lrsquoIS6110 a donneacute un signal
moleacuteculaire quantifiable teacutemoignant de lrsquoabsence de speacutecificiteacute de ce systegraveme pour la matrice sol
De plus M microti membre du MTBC agent de la tuberculose des petits rongeurs est susceptible
drsquoecirctre retrouveacute dans le sol (Courtenay et al 2006) et a deacutejagrave eacuteteacute isoleacute sur des feacutelins et des animaux
sauvages en Cocircte drsquoOr (Laprie et al 2013 Michelet et al 2015) Nous avons donc deacuteveloppeacute des
systegravemes moleacuteculaires qui nrsquoamplifient pas M microti
Enfin M bovis est une mycobacteacuterie pathogegravene de classe III qui neacutecessite une manipulation dans un
laboratoire de confinement L3 Pour des raisons techniques et pour srsquoaffranchir de la contrainte de
confinement dans la mise au point de la technique de PCR nous avons utiliseacute une des souches
atteacutenueacutees de M bovis et plus preacuteciseacutement M bovis BCG Pasteur 1173P2 manipulable dans un
laboratoire de type L2 disponible agrave lrsquoINRA de Dijon Nous avons donc choisi des cibles moleacuteculaires
eacutegalement preacutesentes chez M bovis BCG
Pour augmenter la speacutecificiteacute de la deacutetection moleacuteculaire dans les matrices environnementales
complexes nous avons choisi drsquoutiliser la PCR quantitative avec une sonde TaqManreg qui est formeacutee
par un nucleacuteotide lieacute de faccedilon covalente agrave un fluorophore (reporter) en 5rsquo et agrave un deacutesactivateur
(quencher) en 3rsquo Le clivage de la sonde par la Taq polymeacuterase apregraves lrsquohybridation agrave la seacutequence drsquoADN
compleacutementaire eacuteloigne le quencher du fluorophore qui eacutemet une fluorescence mesureacutee en temps
reacuteel dans un thermocycleur adapteacute
Chapitre 2
87
Le systegraveme moleacuteculaire deacutecrit preacuteceacutedemment par Sweeney et al (2007) ciblant les marges de
la reacutegion RD4 deacuteleacuteteacutee uniquement chez M bovis et M bovis BCG nous a particuliegraverement inteacuteresseacutes
Des amorces internes agrave la seacutequence RD4 ont eacuteteacute utiliseacutees en PCR pour la discrimination de M bovis
avec les autres membres du MTBC (pas de signal pour M bovis mais un signal pour les autres membres
du MTBC) (Huard et al 2003 Courtenay et al 2006) Des amorces de PCR classique ont eacutegalement eacuteteacute
dessineacutees sur les zones flanquantes de la reacutegion RD4 (Brosch et al 2002) Ces amorces ont eacuteteacute adapteacutees
agrave la technologie TaqManreg avec une amorce et une forward localiseacutees sur les marges de la seacutequence
deacuteleacuteteacutee et une sonde agrave cheval sur la zone de deacuteleacutetion (Sweeney et al 2007) La speacutecificiteacute et la
validation de ce systegraveme sur diffeacuterents eacutechantillons environnementaux tels que des sols de terriers
des fegraveces de blaireaux et de buffles africains ont eacuteteacute deacutemontreacutees dans plusieurs eacutetudes (Sweeney et al
2007 Roug et al 2014 Travis et al 2011 Pontiroli et al 2013 King et al 2015) Ce systegraveme de deacutetection
nous paraissant a priori speacutecifique pour la discrimination de M bovis dans le MTBC et adapteacute aux
matrices complexes nous lrsquoavons inclus dans lrsquoeacutetude Toutefois lrsquoamorce Reverse du systegraveme de
deacutetection a eacuteteacute modifieacutee par nos soins afin de reacuteduire lrsquoamplicon obtenu de 146 agrave 116 bases taille plus
adapteacutee agrave la technologie TaqManreg
Face au manque de systegravemes discriminants disponibles dans la litteacuterature nous avons dessineacute
deux nouveaux systegravemes Nous avons choisi de cibler la seacutequence drsquoinsertion IS1561rsquo (Rv3349c) car elle
est preacutesente chez tous les membres du MTBC sauf M microti (Gordon et al 1999 Huard et al 2003
Brodin et al 2002) Elle appartient agrave la famille des ISL3 qui regroupe des copies drsquoIS deacutefectueuses chez
M tuberculosis elles sont alors annoteacutees avec une apostrophe
La troisiegraveme cible a eacuteteacute choisie dans une reacutegion chromosomique de M tuberculosis appeleacutee
RD1 preacutesente chez les souches virulentes de M tuberculosis et M bovis mais partiellement deacuteleacuteteacutee
chez M bovis BCG (Mahairas et al 1996) et M microti (Brodin et al 2002) Cette reacutegion contient
notamment un cluster de 6 gegravenes (Rv3871 agrave Rv3876) qui fait partie des gegravenes codant les proteacuteines
antigeacuteniques ESAT-6 (Early Secreted Antigenic Target 6kD) et CFP-10 (Culture Filtrat Protein 10kD)
responsables de la reacuteponse immunitaire de lrsquohocircte M bovis BCG et M microti sont les deux seuls
membres du MTBC agrave preacutesenter une deacuteleacutetion de ce cluster deacuteleacutetion probablement en partie
responsable de leur absence de virulence et de leur utilisation en vaccination (Pym et al 2002) Chez
M bovis BCG la reacutegion aval de ce cluster est deacuteleacuteteacutee la reacutegion deacuteleacuteteacutee est appeleacutee RD1bcg (Figure 18)
Chez M microti crsquoest la reacutegion amont du cluster qui est absente la reacutegion deacuteleacuteteacutee est appeleacutee RD1mic
Nous avons plus particuliegraverement cibleacute le locus Rv3866 preacutesent chez M tuberculosis et dans toutes les
souches de M bovis et M bovis BCG mais absente chez M microti (Brodin et al 2002) M avium et
M smegmatis (Mahairas et al 1996)
Chapitre 2
88
Figure 18 Diffeacuterences geacutenomiques de la reacutegion RD1 entre M bovis BCG (A) M microti (C) par comparaison agrave M tuberculosis H37Rv (B) drsquoapregraves Brodin et al 2002 et localisation du locus Rv3866 RD1mic correspond agrave la seacutequence de M tuberculosis deacuteleacuteteacutee chez M microti RD1bcg correspond agrave la seacutequence de M tuberculosis deacuteleacuteteacutee chez M bovis BCG
Les seacutequences drsquointeacuterecirct ont eacuteteacute rechercheacutees pour les membres du MTBC et certaines MNT sur
la base de donneacutees GenBank Un alignement de ces seacutequences a eacuteteacute reacutealiseacute gracircce au portail drsquoanalyses
bioinformatiques MOBYLE PASTEUR pour rechercher les reacutegions conserveacutees et les reacutegions de
diffeacuterence Le dessin des oligonucleacuteotides a eacuteteacute reacutealiseacute grace au logiciel en ligne Primer3
(httpbioinfouteeprimer3-040) Les systegravemes de deacutetection ont eacuteteacute testeacutes in silico gracircce agrave lrsquooutil
BLAST (Basic Local Alignment Search Tool) afin drsquoeacutevaluer la speacutecificiteacute de nos systegravemes pour chaque
cible par rapport agrave une base de donneacutees de geacutenomes seacutequenceacutes
La validation (speacutecificiteacute et seuil de deacutetection) de ces trois systegravemes ainsi que leur utilisation
dans le screening drsquoun nombre important drsquoeacutechantillons environnementaux sont preacutesenteacutees dans le
Chapitre 3 dans lrsquoarticle intituleacute laquo First molecular detection of Mycobacterium bovis in environmental
samples from a French region with endemic bovine tuberculosis raquo Les reacutesultats des analyses reacutealiseacutees
en 2013 sont preacutesenteacutes dans un article publieacute en 2014 dans la revue de lrsquoAEEMA (association pour
leacutetude de leacutepideacutemiologie des maladies animales) en Annexe 1
A
B
C
Chapitre 2
89
CHAPITRE 3
Etude de la preacutevalence environnementale de
Mycobacterium bovis dans la zone endeacutemique de
tuberculose bovine de Cocircte drsquoOr et reacutesultats obtenus
en Charente
Elodie Barbier Maria Laura Boschiroli Eric Gueneau Murielle Rochelet Ariane Payne Krystel
de Cruz AnneLaure Blieux Ceacuteline Fossot Alain Hartmann
First molecular detection of Mycobacterium bovis in environmental samples from a French
region with endemic bovine tuberculosis
(Article accepteacute en cours de publication Journal of Applied Microbiology)
93
CHAPITRE 3 - Etude de la preacutevalence environnementale de
Mycobacterium bovis dans la zone endeacutemique de tuberculose
bovine de Cocircte drsquoOr et reacutesultats obtenus en Charente
A AVANT-PROPOS
Dans la synthegravese bibliographique preacutesenteacutee dans le chapitre 1 nous avons deacutecrit la situation
eacutepideacutemiologique complexe de la tuberculose bovine en Cocircte drsquoOr Dans ce deacutepartement
Mycobacterium bovis circule dans un systegraveme multi-hocirctes complexe incluant les bovins les blaireaux
les cervideacutes et les sangliers sur une zone geacuteographique limiteacutee (Payne 2014) La faune sauvage infecteacutee
et potentiellement excreacutetrice est suspecteacutee drsquoecirctre agrave lrsquoorigine de la contamination ou recontamination
de certains foyers bovins dans cette zone (ANSES 2011 Courcoul Moutou amp Vialard 2013) Le mode
de transmission indirecte est suspecteacute car les contacts directs entre bovins et animaux sauvages sont
rarement deacutecrits (Drewe OrsquoConnor Weber McDonald amp Delahay 2013 Kukielka et al 2013) Ce mode
de transmission repose sur la contamination de matrices environnementales par des mycobacteacuteries
viables et virulentes agrave la faveur de lrsquoexcreacutetion animale puis sur lrsquointeraction drsquoanimaux avec ces
matrices Deacutemontreacutee par des meacutethodes moleacuteculaires etou culturales en Angleterre et en Espagne sur
des sites partageacutes par les espegraveces sauvages et les bovins (King et al 2015 Santos et al 2015 Sweeney
et al 2006 2007) la contamination environnementale par M bovis nrsquoa pas encore fait lrsquoobjet drsquoeacutetudes
dans les zones endeacutemiques franccedilaises
Afin drsquoeacutevaluer la preacutevalence et lrsquoexistence de reacuteservoirs potentiels de M bovis dans
lrsquoenvironnement des bovins et des animaux sauvages infecteacutes en Cocircte drsquoOr nous avons deacuteveloppeacute et
valideacute des systegravemes de deacutetection moleacuteculaire ou modifieacute des systegravemes existants adapteacutes aux matrices
environnementales complexes pour deacutetecter les espegraveces du MTBC et plus speacutecifiquement M bovis
Ces systegravemes ont eacuteteacute utiliseacutes pour analyser plus de 400 eacutechantillons environnementaux (sol eau
seacutediments microfaune fegraveces) preacuteleveacutes en Cocircte drsquoOr Notre strateacutegie drsquoeacutechantillonnage a cibleacute des
pacirctures suspectes sur lesquelles des bovins infecteacutes ont transiteacute et autour desquelles des animaux
sauvages infecteacutes ont eacuteteacute deacutetecteacutes et plus particuliegraverement les zones pouvant ecirctre partageacutees par les
diffeacuterentes espegraveces notamment les zones drsquoabreuvement Nous avons eacutegalement analyseacute des
eacutechantillons provenant de sites freacutequenteacutes exclusivement par la faune sauvage notamment des sols
de terriers et des latrines de blaireaux localiseacutes agrave proximiteacute du site de pieacutegeage de blaireaux infecteacutes
ainsi que des seacutediments preacuteleveacutes dans les trous drsquoeau ou souilles freacutequenteacutes par les sangliers et
cervideacutes
Chapitre 3
95
B FIRST MOLECULAR DETECTION OF MYCOBACTERIUM BOVIS IN ENVIRONMENTAL SAMPLES FROM A FRENCH REGION WITH ENDEMIC BOVINE TUBERCULOSIS
E Barbier12 ML Boschiroli3 E Gueneau4 M Rochelet2 A Payne1 Krystel de Cruz3 AL Blieux5 C
Fossot6 A Hartmann1
1 INRA UMR 1347 Agroeacutecologie Dijon Cedex France
2 Universiteacute de Bourgogne Franche Comteacute UMR 1347 Agroeacutecologie Dijon Cedex France
3 Universiteacute Paris-Est Laboratoire National de Reacutefeacuterence de la Tuberculose Uniteacute de Zoonoses
Bacteacuteriennes Laboratoire de Santeacute Animale ANSES Maisons-Alfort Cedex France
4 Laboratoire Deacutepartemental de la Cocircte drsquoOr Dijon France
5 Welience SATT Grand-Est AgrOnov Breteniegravere France
6 Phytocontrol Paris MIN de Rungis Rungis Cedex France
Running head
Environmental detection of Mycobacterium bovis
Corresponding author
Alain Hartmann UMR 1347 Agroeacutecologie INRA 17 rue Sully BP 86510 21065 Dijon Cedex France E-
mail alainhartmanndijoninrafr
Chapitre 3
96
ABSTRACT
Aims
The aim of the study was to determine the prevalence of Mycobacterium bovis (the causative agent of
bovine tuberculosis bTB) in environmental matrices within a French region (Cocircte drsquoOr) affected by this
zoonotic disease
Methods and Results
We report here the development and the use of molecular detection assays based on qPCR (double
fluorescent dye labelled probe) to monitor the occurrence of Mycobacterium tuberculosis complex
(MTBC) or M bovis in environmental samples collected in pastures where infected cattle and wildlife
had been reported Three qPCR assays targeting members of the MTBC (IS1561rsquo and Rv3866 loci) or M
bovis (RD4 locus) were developed or refined from existing assays These tools were validated using
M bovis spiked soil water and feces samples Environmental samples were detected positive for the
presence of MTBC strains and M bovis in the environment of bTB-infected farms in the Cocircte drsquoOr
region
Conclusions
The development of molecular assays permitted testing of several types of environmental samples
including spring water sediment samples and soils from badger setts entrance located in the vicinity
of these farms which were repeatedly contaminated with M bovis (up to 87 times 103 gene copies per g
of badger sett soil) For the first time direct spoligotyping of soil DNA enabled identification of M bovis
genotypes from environmental matrices
Significance and Impact of the Study
All together these results suggest that M bovis occurs at low levels in environmental matrices in Cocircte
drsquoOr within the bTB-infected area Drinking contaminated water or inhaling contaminated bioaerosols
might explain cattle infection in some cases
Keywords
Mycobacterium bovis environment quantitative real-time PCR indirect transmission
Chapitre 3
97
INTRODUCTION
Bovine tuberculosis (bTB) is a worldwide spread zoonosis caused by Mycobacterium bovis a
pathogenic mycobacteria that belongs to the Mycobacterium tuberculosis complex (MTBC) This
disease affects cattle that are the natural host of M bovis but also a wide range of domestic or wild
mammals including humans (Morris et al 1994 Biet et al 2005) Infected wildlife can act as a bTB
reservoir maintaining and spreading M bovis infection in cattle Many studies including French ones
showed the circulation of the bacteria within multi-host systems including cattle and various wild
species as badgers (Meles meles) wild boar (Sus scrofa) or red deer (Cervus elaphus) (Little et al 1982
Aranaz et al 2004 Gortaacutezar et al 2008 Zanella et al 2008 Richomme et al 2010 Payne et al 2012)
The zoonotic nature of bTB may represent a public health threat Moreover animal disease leads to
important economic losses mainly due to cattle testing slaughtering campaigns and wildlife screening
trials
Infected animals can excrete M bovis via sputum feces urine and any type of aerosols (Little
et al 1982 Phillips et al 2003 Palmer et al 2004 Payne 2014) and thus contaminate their environment
(Phillips et al 2003) This may contribute to contamination of cattle pastures through deposition of
M bovis on cattle feedstuffs and into drinking water Indeed many studies confirmed the presence and
the persistence of M bovis in environmental substrates such as soil feces hay corn water (Young et
al 2005 Courtenay et al 2006 Sweeney et al 2006 Fine et al 2011 Ghodbane et al 2014) despite
many technical difficulties in isolating this organism from environmental substrates Altogether these
data strongly suggest that the environment may constitute a reservoir of M bovis from which cattle
might get infected However there is still very little direct evidence concerning these potential M bovis
environmental reservoirs and the way that animals might get infected through them
The European Commission declared that France was bTB-free in 2001 However the disease is
still a persistent problem in several regions including Cocircte drsquoOr (Middle East of France Figure 1)
Despite a very low national prevalence in cattle (0075 since 2012) complete eradication of the
disease seems difficult to achieve (Fediaevsky et al 2014) In Cocircte drsquoOr M bovis circulates in a multi-
host system including badgers wild boar and deer within a restricted area of the region (Payne 2014
Hauer et al 2015) As shown in Figure 1 two genotypes are found in this area M bovis spoligotype
SB0120 VNTR profile 5 5 4 3 11 4 5 6 in the northern bTB endemic area and M bovis spoligotype SB0134
VNTR profile 6 4 5 3 6 4 3 6 in the southern area Between 2002 and 2014 196 cattle breakdowns have
been registered in Cocircte drsquoOr (Cote drsquoOr Regional Veterinary Service personal communication) Wild
animals in the vicinity of infected cattle herds were found to be infected by M bovis with the same
genotypes (Hauer et al 2015)
Direct cross contamination between cattle has historically been considered as the main
Chapitre 3
98
transmission route of M bovis (Morris et al 1994 Phillips et al 2003) but environmental
contamination is suspected in many herd infections when cattle and wildlife share the same resources
(Garnett et al 2002 Palmer et al 2004) However this hypothesis has never been extensively explored
in France This is probably due to the lack of adequate detection tools to monitor M bovis under natural
environmental conditions Indeed most of the studies report the detection of M bovis from
experimentally inoculated environmental samples (Fine et al 2011 Adams et al 2013) Although the
culture method is the gold standard for direct diagnosis of M bovis in animals it is not well suited to
studying environmental samples due to the abundance of soil microbial communities and the slow
growth rate of M bovis To circumvent these drawbacks the use of highly selective media containing
antibiotics and antifungals in combination with sample decontamination is required in order to prevent
competition of fast growing organisms However such pretreatments strongly reduce M bovis viability
(Thornton et al 1998 Fine 2006) On the other hand molecular detection tools such as semi-
quantitative PCR have been used to detect M bovis in soils water and badger feces Following DNA
extraction detection of MTBC members or M bovis was done using primers targeting mpb64 mpb70
hsp65 16SrRNA genes or IS6110 insertion sequence (Young et al 2005 Courtenay et al 2006
Sweeney et al 2006 Adams et al 2013 Santos et al 2015) More recently a M bovis specific qPCR
(based on double fluorescent dye probe detection) was developed to target the RD4 deletion flanking
sequences (Sweeney et al 2007) This molecular system has been validated in several studies to test
environmental substrates such as soil badger feces and cow slurry (Pontiroli et al 2011 Travis et al
2011 King et al 2015a King et al 2015b) This RD4 based qPCR assay allowed to detect the occurrence
of M bovis within environmental matrices in UK (Sweeney et al 2007 King et al 2015a King et al
2015b) Finally Santos et al (Santos et al 2015) successfully used a hemi-nested PCR targeting the
RD12 locus which is common to M bovis and M caprae to monitor these species in environmental
substrates from bTB endemic regions located in the Iberian Peninsula
In this context the objectives of this study were firstly to develop new sensitive qPCR based
tools and to refine an existing one (based on RD4 deletion sequence) for the specific detection of MTBC
or M bovis in environmental matrices The sensitivity and specificity of qPCR based detection systems
were evaluated on a collection of mycobacterial strain DNAs but also on DNAs extracted from samples
(soil water and feces) spiked with known amounts of M bovis Secondly we aimed to screen large
numbers of environmental samples (soil sediments water feces) originating from both bTB endemic
or bTB free areas to assess the presence of MTBC and more specifically the presence of M bovis in
these areas
Chapitre 3
99
Figure 1 Distribution of bTB cumulative outbreaks in cattle in France (picture in the insert above) and per municipality in Cocircte drsquoOr from 2005 to 2013 Geographical location of the sampled areas are indicated by black stars and the location of the six positive areas are indicated by white stars and named A B C D E and F respectively
MATERIALS AND METHODS
Bacterial strains
A collection of bacterial strains belonging to the species M bovis M bovis BCG vaccine strain
and other mycobacterial species (Table 1) was used to test the specificity of the primers to detect
M bovis Crude thermo-lysates of strains corresponding to 20 bacterial spoligotypes of the M bovis
species 4 other species of the MTBC 2 sub-species of the Mycobacterium avium Complex (MAC) and
20 non tuberculous mycobacteria (NTM) Field strains belong to the National Reference Laboratory
collection and were previously identified by several genotyping methods using four qPCR assays done
in parallel and targeting IS6110 IS1081 IS1245 and atpE Namely MTBC members are positive for
Chapitre 3
100
IS6110 and IS1081 targets (Courcoul et al 2014) MAC members are positive for the IS1245 target
(Slana et al 2010) and finally NTM are positive for the atpE target (Radomski et al 2013) Furthermore
spoligotyping was used to discriminate the different MTBC species (Zhang et al 2010) IS901 was used
to discriminate between MAC members (Slana et al 2010) and hsp65 sequencing was used to
discriminate between other mycobacteria species (Telenti et al 1993) Attenuated M bovis BCG strain
Pasteur 1173P2 was used as target organism to validate the specificity and detection limits of the qPCR
assay in soil water and feces microcosms
Origin of environmental samples
Within the bTB endemic zone in Cocircte drsquoOr a total number of 356 environmental samples were
collected from 15 areas (an area corresponds to a municipality territory including at least one bTB
infected cattle farm) between April 2013 and December 2014 In those areas typical landscape is
observed ie limestone plateau with forest or farm fields overhang small hills The hillslopes are
covered with hydromorphic soils containing marl and claylimestone Alluvial valleys set at the bottom
of the slopes Cattle usually graze on those pastures during summer and spring Samples of soil
sediment and water were collected in areas frequented by both wildlife and cattle such as feeding
areas waterholes naturally occurring springs drinking troughs with open access to livestock and
wildlife Knowing that bTB infected badgers were discovered in the vicinity of the pastures grazed by
infected cattle (Cote drsquoOr Regional Veterinary Service personal communication) soils from badger
setts and latrines were also collected
Twenty-four agricultural soils were randomly chosen as control soils in French bTB-free areas
among a panel of soils from the French soil quality monitoring network (Reacuteseau de Mesures de la
Qualiteacute des Sols Infosol INRA Orleacuteans) Among them 11 french soils were collected from Brittany
(North West of France) Yvelines (Middle north of France) Haut Rhin (North east of France) while 13
soils were collected in the bTB-free area of Cocircte drsquoOr
In order to evaluate the detection limit of the qPCR assay we used two agricultural soils from two bTB-
free areas of Cocircte drsquoOr (Soil S1 and soil S2) one pond water and badger feces from animals tested as
bTB-free from non-infected areas Both soils were sampled in a bTB-free cattle pasture in November
2013 They were chosen for their contrasted structural and physico-chemical properties (Table 2)
DNA extraction and purification
DNAs from all soil sediment and latrine samples were extracted from 2 g-samples as following
Briefly 8 ml of lysis buffer (01 mol l-1 Tris pH 80 01 mol l-1 EDTA pH 80 01 mol l-1 NaCl and 2 (wv)
SDS) 4 g of silica beads (100 μm diameter) 5 g of ceramic beads (14 mm diameter) and 8 glass beads
(4 mm diameter) were added to each sample placed in a 15 ml conical centrifugation tube (Falcon
Chapitre 3
101
Dominique Dutscher France) Samples were disrupted for 3 times 30 s at 4 m s-1 in a FastPrep-24 Instrument
(MP Biomedicals Europe France) incubated for 30 min at 70degC and centrifuged for 5 min at 7000 g at
room temperature in a Sigma 3-18K centrifuge (Sigma-Aldrich Germany) Supernatants were
incubated on ice for 10 min with 110 volume of 3 mol l-1 potassium acetate pH 55 and then
centrifuged for 5 min at 14000 g One volume of ice-cold isopropanol was added to the supernatant
for DNA precipitation overnight at minus 20degC DNA was collected by centrifugation (30 min at 14000 g
4degC) and DNA pellets were washed with ice-cold ethanol (70 ) and dissolved in 100 μl of water Crude
DNA extracts (100 μl) were loaded on a Microbiospin column (Biorad France) filled with 100 mg of
polyvinyl polypyrrolidone (Sigma Chemical Co France) After centrifugation (4 min at 1000 g 4degC)
eluted DNA was further purified using the Geneclean Turbo Kit (Qiagen France) according to the
manufacturers instructions For water samples the microbial biomass was concentrated by filtering
from 100 ml to a maximum of 1 l of water sample (depending on water turbidity) through a 045 microm
Millipore HAWP filter (47 mm Merk Millipore Germany) Filters were then ground in liquid nitrogen
and DNA was extracted as described above Nematodes were extracted from soil using an Oostenbrink
elutriator (Oostenbrink 1960) DNA extraction was achieved as described above
Purified DNA concentrations were estimated using a NanoDropreg 2000 spectrophotometer
(Thermo Scientific France) at 260 nm wavelength
Choice of gene targets design of primers and probes for qPCR assays
A first qPCR detection system assay targeting the insertion sequence IS1561rsquo was designed
This insertion sequence is present in all the MTBC members except in M microti (Huard et al 2003
Huard et al 2006) The primers and double dye probe sequences are given in Table 3 IS1561rsquo contains
a defective transposase and belongs to the ISL3 family (Gordon et al 1999) In order to confirm the
specificity of the detection for MTBC a nested PCR protocol was additionally developed the first step
was a classical PCR using primers targeting a sequence of 222-bp sequence in the IS1561rsquo surrounding
the 76-bp qPCR product (Table 3) the second step was a qPCR assay targeting the 76-bp fragment To
confirm the results obtained with IS1561rsquo qPCR assay two supplementary qPCR targets were used The
first one was the Rv3866 locus located in the RD1 locus (Brodin et al 2002) The second one was the
RD4 flanking region (the RD4 locus is specifically deleted in M bovis) To improve the qPCR efficiency
the RD4 based assay was derived from a previously reported qPCR assay (Sweeney et al 2007) by
modifying the reverse primer to reduce the size of the qPCR product from 142 to 116 bp (Table 3) The
RD4 based qPCR assay designed by Sweeney et al (Sweeney et al 2007) has previously been validated
for environmental matrices in several studies (Pontiroli et al 2011 Travis et al 2011 King et al 2015a
King et al 2015b) Primers and probe were designed from alignments of mycobacterial genes from
GenBank (httpwwwncbinlmnihgovgenbank) such as M bovis AF212797 four strains of M bovis
Chapitre 3
102
BCG M tuberculosis H37Rv M africanum GM041182 and Mycobacterium canettii CIPT 140010059
(respective accession numbers NC_0029453 NC_0087691 NC_0168041 NC_0122071
GCA_0004632151 NC_0009623 NC_0157581 NC_0158481) using Primer3web
(httpprimer3utee) The specificity of the assay was tested in silico by BLAST analysis
(httpblastncbinlmnihgovBlastcgi) Primers and probes were synthesized by Eurogentec (Seraing
Belgium)
Quantitative real time PCR (qPCR) and nested PCR conditions
For the three qPCR assays reactions were carried out in a 25 microl reaction mix containing 125 microl
of Absolute QPCR ROX Mix (Thermo Scientific France) at a final concentration of 1X 1 microl of T4
bacteriophage gene 32 product (final concentration 25 ng microl-1 MP Biomedicals Europe France) 1 microl of
each primer (final concentration 400 nmol l-1) 05 microl of each probe (final concentration 200 nmol l-1)
4 microl of ultrapure water and 5 microl of diluted DNAs for the environmental samples screening Absolute
quantification was achieved using standard DNA dilutions A recombinant plasmid containing one copy
of the IS1561rsquo fragment of M bovis BCG str Pasteur 1173P2 cloned in the pCRregII-TOPOreg vector
(Invitrogen France) was used as a standard A calibration curve was obtained by amplification of serial
dilutions of the plasmid ranging from 1 to 106 copies per PCR reaction All the environmental
metagenomic DNA samples were screened with the IS1561rsquo based qPCR assay and final results were
expressed as the gene copy number per g of soil feces or water qPCR assays were performed in a VIIA
7 Real-Time PCR System (Thermo Fischer Scientific France) Initial denaturation was made at 95degC for
15 min followed by 45 cycles with denaturation at 95degC for 15 s and annealing and elongation at 60degC
for 1 min
Positive DNA extracts were then tested by the nested IS1561rsquo PCR protocol as follows the first PCR was
achieved in a 25 microl reaction mixture containing 1725 microl of ultrapure water for molecular biology 25
microl of PCR Buffer 10X with MgCl2 02 microl of dNTP mix at 25 mmol l-1 each 005 microl of Taq DNA polymerase
15 U microl-1 (Taq Core Kit 25 MP Biomedicals Europe France) 125 microl of each primers (final concentration
500 nmol l-1) and 25 microl of environmental DNA The nested PCR was achieved in a Veriti Thermal Cycler
(Applied Biosystems France) with a first step of denaturation at 94degC for 5 min then 30 cycles for 30
s at 94degC for denaturation 30 s at 60degC for annealing and 30 s at 72degC for elongation and 5 min at 72degC
for final elongation The qPCR was then achieved in the same conditions as described above using as a
target a 5 microl aliquot of a 100-fold dilution of the first PCR product
Preliminary screening of all environmental samples was done using the IS1561rsquo based qPCR
assay targeting MTBC species The IS1561rsquo positive samples were then further analysed using the
Rv3866 based qPCR to confirm the presence of MTBC and the RD4 based qPCR assay to confirm the
presence of M bovis All qPCR assays were done in triplicate
Chapitre 3
103
Determination of qPCR assays analytical sensitivity in spiked soil feces or water microcosms
Microcosms consisting of 2 g of fresh soil S1 and S2 sieved to 4 mm 2 g of fresh badger feces
or 100 mL of pond water were spiked in triplicate with 100 microl of 10-fold serially diluted suspensions of
M bovis BCG strain Pasteur to reach M bovis concentrations ranging from 25 to 25 times 106 CFU per
gram of soil or feces and 50 to 50 times 106 CFU per 100 mL of water Briefly inoculum suspensions were
realized as following a thirty days M bovis BCG liquid culture was vortexed for 15 s with glass beads (4
mm diameter) in order to disrupt aggregates Optical bacterial density was then measured at 600 nm
(Biophotometre Eppendorf France) to roughly estimate mycobacterial concentration Appropriate 10-
fold dilutions (10-1 to 10-6) were prepared in sterile water M bovis concentration was estimated by
spreading in triplicate 100 microL-aliquot of 10-4 10-5 and 10-6 diluted suspension onto Middlebrook 7H11
agar enriched with OADC 10 (Sigma-Aldrich France) Plates were incubated at 37degC for two months
and then M bovis colonies were counted Results were reported as CFU per ml of suspension
The spiked microcosms were incubated at room temperature for 3-4 hours prior to DNA
extraction as described in the section 3 To assess the reproducibility all qPCR assays were done in
triplicate and performed as previously using the three detection systems targeting IS1561rsquo Rv3866 and
RD4 Non-inoculated soil feces or water used to prepare the microcosms proved to be negative with
the three qPCR systems in preliminary experiments
Estimation of DNA quality for PCR amplification
Efficiency of qPCR might be impaired by the presence of PCR inhibitors in DNA extracts The
quality of DNA extracted from soil water and feces was assessed by measuring the capacity to amplify
a control DNA target in the presence of various dilutions of soil DNA extracts Briefly qPCR reactions
were done in a final volume of 25 microl containing 25 μl of 106 copies of the circularized pGEM-T Easy
plasmid DNA and 5 μl of water (control) or diluted soil DNA (5- 10- 20-fold dilutions) 1X ABsolute
qPCR SYBR Green ROX Mix (final concentration 500 nmol l-1 Thermo Scientific France) 25 ng microl-1 of T4
bacteriophage gene 32 product qPCR primers were SP6 (5rsquo - ATTTAGGTGACACTATAG - 3rsquo) and T7 (5rsquo -
TAATACGACTCACTATAGGG - 3rsquo) universal primers (final concentration 500 nmol l-1) targeting the
polylinker of the plasmid qPCR and cycling conditions were as following initial denaturation at 95degC
for 15 min followed by 40 cycles with denaturation at 95degC for 15 s annealing at 55degC and elongation
at 72degC for 30 s each The PCR product specificity was assessed by the melting curve analysis A 10-fold
reduction of the copy number amplified by qPCR in the presence of diluted soil DNA with respect to
the copy number obtained in the control reaction (water) was considered as 10 amplification
efficiency or conversely as 90 inhibition Although inhibition tests and detection of specific targets
were conducted with different mixes and primers the DNA polymerases from the ABsolute qPCR SYBR
Chapitre 3
104
Green ROX Mix (used for inhibition testing) and the ABsolute QPCR ROX Mix (used for mycobacterial
detection in environmental samples) are the same (Thermo-Start DNA polymerase) Thus the quality
of DNAs extracted from environmental samples was estimated by this method that is compatible with
acceptable experimental costs associated with the high number of analysed samples
Direct M bovis spoligotyping from environmental DNA
Environmental DNA samples that proved to be positive for the presence of MTBC were
submitted for spoligotyping using the Luminex method described previously by Zhang et al (Zhang et
al 2010) Spoligotypes were named according to an agreed international convention
(wwwmbovisorg)
RESULTS
Validation of the qPCR assays on pure bacterial strains
The results of the specificity test for the three qPCR assays are shown in Table 1 Using IS1561rsquo-
and Rv3866-based qPCR assays all spoligotypes of M bovis were detected with cycle threshold (Ct)
values below 25 cycles as well as all strains from the other species belonging to the MTBC (M caprae
M tuberculosis M pinnipedi) except the M microti species Using the RD4-based qPCR assay only
M bovis strains were detected with Ct values below 25 cycles For the three qPCR assays the 22 strains
of the NTM and MAC members were not detected or gave Ct values above 35 cycles Strains with Ct
values above 35 cycles when using high DNA concentration from pure culture are unlikely to give any
signal when testing soil metagenomic DNA where targets are highly diluted These results indicated
that IS1561rsquo- and Rv3866-based qPCR assays are highly specific for MTBC members and exclude
M microti whereas the RD4-based qPCR assay is specific for M bovis species
The calibration curve used for absolute quantification is presented in Figure 2 for the IS1561rsquo-
based qPCR assay A linear regression was established between the log (gene copy number) and the Ct
value over a wide range of target concentrations from 10 to 106 gene copies per PCR (r2= 09989) Below
10 target gene copies per reaction the detection was still possible but results were not reproducible
The quantification limit of the qPCR detection system was estimated to be 10 gene copies per PCR
reaction thus indicating a high sensitivity Similar calibration curves and detection thresholds were
obtained for the two other qPCR assays targeting Rv3866 and RD4 (data not shown)
Chapitre 3
105
Table 1 Strains used for testing oligonucleotides specificity Results are given with Ct (Cycle Threshold) ND (not detected) means that no signal was observed after 40 cycles
Strains Ct
IS1561rsquo
Ct
Rv3866 Ct RD4
Mycobacterium bovis BCG
BCG str Pasteur 1173P2
BCG str Danish 1331
lt25 lt25 lt25
Mycobacterium bovis
SB0089 SB0120 SB0121 SB0130
SB0134 SB0162 SB0166 SB0821
SB0823 SB0825 SB0826 SB0827
SB0878 SB0885 SB0912 SB0948
SB0999 SB1081
lt25 lt25 lt25
Others MTBC members
Mycobacterium microti ND ND ND
Mycobacterium tuberculosis lt25 lt25 ND
Mycobacterium caprae lt25 lt25 ND
Mycobacterium pinnipedi lt25 lt25 ND
Non tuberculous mycobacteria
Mycobacterium avium ND ND ND
Mycobacterium intracellulare ND ND ND
Mycobacterium aichiense ND ND ND
Mycobacterium arupense ND gt35 gt35
Mycobacterium celatum gt35 ND ND
Mycobacterium chelonae ND ND ND
Mycobacterium chitae ND ND ND
Mycobacterium confluentis gt35 ND gt35
Mycobacterium elephantis ND ND ND
Mycobacterium fortuitum ND ND ND
Mycobacterium intermedium ND ND ND
Mycobacterium kansai ND gt35 ND
Mycobacterium non chromogenicum ND ND ND
Mycobacterium parafinicum gt35 ND ND
Mycobacterium porcinum ND ND ND
Mycobacterium rufum ND ND ND
Mycobacterium rutilum ND ND ND
Mycobacterium terrae ND ND ND
Mycobacterium smegmatis ND gt35 ND
Mycobacterium szulgai ND gt35 ND
Mycobacterium thermoresitibile ND ND ND
Mycobacterium vaccae ND ND ND
Mycobacterium xenopi gt35 gt35 gt35
Chapitre 3
106
Detection threshold of qPCR assays in M bovis BCG spiked environmental matrices
The detection thresholds of the three qPCR assays were evaluated in complex environmental
matrices (soil feces and water) spiked with 25 to 25 times 106 CFU per g of soil or feces and 50 to 50 times
106 CFU per 100 ml of water The quality of DNAs extracted from microcosms was assessed as described
in the MampM section ldquoEstimation of DNA quality for PCR amplificationrdquo The results showed that 20-fold
dilutions of the DNA extracts did not inhibit the qPCR assay and were thus used for all qPCR assays
performed on spiked microcosms Results of M bovis detection in spiked microcosms are given in Table
4 for IS1561rsquo-based qPCR assay No signal was detected in control samples (not inoculated)
For M bovis cell densities above 250 CFU per g of soil 2500 CFU per g of feces and 5 CFU per
g of water (or 500 CFU per 100 ml of water) the detection by the qPCR assay yielded eight to nine
positive reactions out of nine repeats which allowed determination of the numbers of M bovis gene
copy number In contrast although M bovis was sometimes detected in spiked material with lower
densities than those described above quantification was unreliable A strong discrepancy between the
results obtained by the culture-based method and those obtained with the qPCR assay was observed
(10- to 100-fold higher for the qPCR assay) All of the microcosms were analysed with the two
complementary detection systems (targeting the Rv3866 locus and the flanking region of the RD4
deletion) and similar results were obtained (data not shown) Despite soils S1 and S2 being different
based on their physico-chemical characteristics detection limits were equivalent with the three qPCR
systems on these two soils
Extrapolating these results for natural environmental samples suggest that the three qPCR
assays allow a reliable quantification of MTBC (IS1561rsquo and Rv3866) or M bovis (RD4) targets in
environmental samples for Ct values below 39 Above this threshold detection may occur but lacks
reproducibility The word ldquotracesrdquo was then chosen in all further experiments in order to indicate that
MTBC or M bovis targets are present in very low quantities
Chapitre 3
107
Figure 2 Standard curve used for absolute quantification of M bovis in samples with IS1561rsquo qPCR assay (Mean values of ten standard curves) Error bars represent the 95 confidence interval associated to the values
Table 2 Properties of the two soils used for the qPCR sensitivity detection of Mycobacterium bovis
Table 3 Sets of primers tested in this study F forward primer R reverse primer P Double dye probe
Target locus Nucleotide sequence Location Size (bp)
IS1561rsquo F 5rsquo GATCCAGGCCGAGAGAATCTG 3rsquo
R 5rsquo GGACAAAAGCTTCGCCAAAA 3rsquo
P 5rsquo FAM ACGGCGTTGATCCGATTCCGC TAMRA 3rsquo
3742298-3742319
3742354-3782374
3742332-3742353
76
Nested IS1561rsquo F 5rsquo AGGAAGCGGTGTAGGTGATG 3rsquo
R 5rsquo AAGATCGACCCGCAGTG 3rsquo
3742212-3742232
3742416-3742434 222
RD4 F 5rsquo GCAGAAGCGCAACACTCTTG 3rsquo
R 5rsquo ATCTTAGCTGGTCAATAGCCATTTTT 3rsquo
P 5rsquo FAM AGTGGCCTACAACGGCGCTCTCC 3rsquo
1692712-1692736
1692802-1692827
1692743-1692768
116
Rv3866 F 5rsquo ACCCGCCAACGTCGAA 3rsquo
R 5rsquo CAATTGACGCAATACGGTATCG 3rsquo
P 5rsquo FAM CGAGCTAGCCGAATGCACAACCG 3rsquo
4278658-4278673
4278721-4278743
4278693-4278716
86
Location is given by reference to the M bovis AF212297 genome sequence accession ndeg NC_0029453
y = -33818x + 4394Rsup2 = 09989
15
20
25
30
35
40
45
1 2 3 4 5 6
Cyc
le T
hre
sho
ld C
t
DNA concentrations [log(IS1561 copies5microL-1)]
Site Clay Loam Sand Total
Carbon
Total Nitrogen
pH H2O
Organic matter
Land use
S1 397 485 118 361 038 775 0625 Pasture S2 192 463 345 292 0269 546 0506 Pasture
Chapitre 3
108
Molecular detection of M bovis in environmental samples
Our molecular detection systems were first evaluated on twenty-four soils from bTB-free zones
in France Whatever the detection system no signal was observed showing a good specificity of the
IS1561rsquo Rv3866 and RD4 qPCR systems
Between April 2013 and December 2014 63 samples of pasture soil 150 samples of setts soil 21
latrines samples 67 water samples from naturally occurring springs (and subsequent flowing water on
soil surface) within cattle pastures and 55 samples of spring sediments within 15 areas located in the
bTB-endemic zone in Cocircte drsquoOr were analysed Some spots were sampled several times (monthly from
January to June and in November) to assess the persistence of the MTBC signals Altogether 356 DNA
samples were primarily screened using the qPCR assay targeting IS1561rsquo locus Molecular signals
corresponding to MTBC were found in six areas out of 15 analysed and only positive results are
presented in Table 5 Apparent gene copy numbers were the average value from n = 2 to n = 3 repeats
These results indicate a MTBC contamination of the environment in 40 of the studied areas
Among the 36 positive samples 19 were found to be positive for MTBC detection with low
signals (indicated as ldquoTracesrdquo in Table 5) These samples include 7 badger sett soils 3 badger latrines
1 sediment sample and 8 water samples from naturally occurring springs in pastures More
interestingly high positive signals for MTBC were observed for the other 17 positive samples with 58
to 8 700 gene copies per g of soil It is worth noting that these samples only included badger sett soils
in area D and one nematodes sample in area C while no signal was recorded for the pasture soils
In order to confirm these positive results some of them were subjected to the nested IS1561rsquo
qPCR assay Eleven samples including eight badger sett soils two samples of water from springs and
one sample of nematodes isolated from spring sediments were confirmed to be positive for MTBC
detection with this nested qPCR assay Badger sett soils from area D were sampled five times over an
eight month-period The occurrence of MTBC signal was discontinuous in the seven soils from badger
setts that were sampled However MTBC signals were detected in several badger sett soils (badger sett
soils 1 4 and 5 in Table 5) following a seven to eight months period after M bovis was detected for the
first time The IS1561rsquo gene copy numbers for the positive badger sett soils ranged levels from
254 times 102 to 505 times 104 per g of soil The MTBC signal remained at high levels throughout the eight
month-period although the badgers moved out one month after the first sampling of the sett (Table
5) Furthermore confirmation of M bovis detection using the RD4 qPCR detection system was achieved
for a few IS1561rsquo positive samples Only three out of eight tested samples proved to be positive with
RD4-based detection system (see Table 5) This result definitely demonstrates the occurrence of M
bovis DNA in these samples
Spoligotyping was performed on positive DNA samples and the results are shown in Table 6 A
badger sett soil DNA was fully spoligotyped and found to belong to the SB0120 M bovis spoligotype
Chapitre 3
109
which is responsible for cattle and wildlife infection in this area of Cocircte drsquoOr Partial M bovis
spoligotypes were obtained for three badger sett soils DNAs The two other partial spoligotypes did
not allow identification of the species within the MTBC
Table 4 Evaluation of the detection of the IS1561rsquo qPCR assay with DNA extracted from control and spiked test matrices
Traces means that IS1561rsquo signal was detected but that quantification was unreliable (Ct value above 39)
Number of M bovis spiked per g of dry soil or per 100 mL of water
Number of positive PCR 9
reactions
Mean Ct Standard deviation Ct
Mean IS1561rsquo genome-equivalent per g of soil or feces or 100 mL of water
Soil S1
Control soil 0 ND ND ND
25 1 396 ND Traces 25101 1 394 ND Traces 25102 8 389 1 (654 plusmn 366) times103
25103 9 345 05 (217 plusmn 086) times105
25104 9 302 02 (218 plusmn 026) times106
25105 9 276 05 (132 plusmn 039) times107
Soil S2
Control soil 0 ND ND ND 25 0 ND ND ND 25101 5 404 06 Traces 25102 9 381 07 (800 plusmn 319) times103
25103 9 342 04 (141 plusmn 049) times105
25104 9 296 05 (374 plusmn 161) times106
25105 9 267 07 (265 plusmn 185) times107
Water Control water 0 ND ND ND 5101 1 401 ND Traces 5102 9 381 11 (220 plusmn 145) times104
5103 9 351 05 (148 plusmn 058) times105
5104 9 31 16 (320 plusmn 134) times106
5105 9 259 03 (819 plusmn 209) times107
5106 9 228 03 (698 plusmn 131) times108
Badger feces
Control feces 0 ND ND ND 25101 0 ND ND ND 25102 1 399 ND Traces 25103 9 379 105 (232 plusmn 077) times104
25104 9 347 159 (212 plusmn 187) times105
25105 9 312 203 (244 plusmn 286) times106
25106 9 261 059 (336 plusmn 107) times107
Chapitre 3
110
Table 5 Molecular detection of MTBC or M bovis targets in positive environmental samples from 6 areas in Cocircte drsquoOr Only positive samples are presented MTBC gene copy numbers were computed from calibration curve and amounts of DNA extracted from the various samples (n=2 ou n=3)
Area
Type of sample Sampling date Mean MTBC gene copy numbers g-1 of soil or feces (IS1561rsquo qPCR assay)
Confirmation after IS1561rsquo-nested PCR
Confirmation of presence of M bovis after RD4 qPCR assay
A Badger sett soil June 2013 Traces Negative NT dagger
B Spring water 1 January 2014 February 2014 March 2014
Traces
Traces
Traces
Negative Negative Negative
Negative Negative Negative
C Spring water 1 Spring water 2 Sediments Soil nematodes
February 2014 February 2014 February 2014 February 2014
Traces Traces Traces 582101
Positive Negative Negative Positive
NT
NT
NT
NT
D Badger sett soil 1 March 2014 November 2014
193103 577103
Positive NT
Positive NT
Badger sett soil 2 March 2014 May 2014 June 2014
870103 502102 143103
Positive Positive NT
Positive NT
NT Badger sett soil 3 March 2014
May 2014 June 2014
441102
741103 282103
Positive Positive NT
Negative NT
NT Badger sett soil 4 April 2014
May 2014 June 2014 November 2014
446103
236103 502102 607103
Positive NT
NT
NT
Negative NT
NT
NT Badger sett soil 5 April 2014
May 2014 June 2014 November 2014
248103
254102
202103
360103
Positive NT NT
NT
Positive NT
NT
NT Badger sett soil 6 November 2014 Traces NT NT Badger sett soil 7 November 2014 Traces NT NT Latrine 1 March 2014 Traces Negative NT Latrine 2 March 2014 Traces Negative NT
E Spring water 1 March 2014 March 2014 March 2014
Traces Negative NT Spring water 2 Traces Positive NT Spring water 3 Traces Negative NT Badger sett soil 1 March 2014
March 2014 Traces Negative NT
Badger sett soil 2 Traces Negative NT Latrine 1 March 2014 Traces Negative NT
F Badger sett soil 1 June 2014 June 2014
Traces Negative NT Badger sett soil 2 Traces Positive NT
Traces MTBC targets were present but MTBC genome equivalent were not calculated since Ct were close to the detection limit of the method dagger NT not tested
Chapitre 3
111
Table 6 Spoligotyping results of DNA positive samples extracted from environmental matrices Black squares represent positive hybridization signals and dots represent lack of hybridization
Figure 3 Spatial location of MTBC positive water and sediment samples in a pasture (site C) along the run off of one naturally occurring springs Star 1 indicates the sampling zone of positive spring sediment and nematodes sample Stars 2 and 3 indicate the 2 sampling sites of positive source water Contour lines are superimposed on aerial photography (450 m and 425 m contour lines are visible 5 m elevation interval between 2 consecutive lines)
Area Type of sample
Sampling date Hybridization pattern Spoligotype
D Badger sett soil 2
March 2014 M bovis SB0120
D Badger sett soil 1
March 2014 Partial M bovis spoligotype
D Badger sett soil 3
March 2014 Partial M bovis spoligotype
D Badger sett soil 5
November 2014
Partial M bovis spoligotype
C Water February 2014
Partial Mycobacterium spoligotype
Chapitre 3
112
DISCUSSION Mycobacterium bovis is described as an obligate intracellular microorganism transmitted by
direct contact between mammalian hosts However direct contacts between species seem to be scarce
and indirect transmission involving environmental matrices is likely to occur (Boumlhm et al 2009
Kukielka et al 2013) Many studies have clearly demonstrated the presence and the persistence of
M bovis in naturally infected environmental substrates in bTB endemic regions of United Kingdom
(Young et al 2005 Courtenay et al 2006 Sweeney et al 2006 Sweeney et al 2007 King et al 2015a
King et al 2015b) Evaluation of the environment as a reservoir of M bovis could help to understand
its epidemiological significance and its role in the indirect transmission of the disease among livestock
and wildlife To our knowledge the results from our study demonstrate for the first time the
contamination of environmental matrices with MTBC and more specifically M bovis in Cocircte drsquoOr which
is a region where bTB is endemic in cattle and wildlife
qPCR detection is a method of choice for M bovis environmental monitoring since it is a faster
alternative to culture based methods that are hampered by the slow-growing rate of M bovis and the
high number and diversity of microbial communities Moreover PCR is able to detect M bovis DNA in
the environment over a long period following initial contamination with a relatively high sensitivity
(Adams et al 2013)
The detection of M bovis in environmental samples (soil water and feces) that contain highly
diverse microbial communities including numerous described and potentially non-described NTM
(Falkinham 2009) was still a challenge The two qPCR assays developed in this study (IS1561rsquo and
Rv3866 targets) proved to be specific for the MTBC excluding M microti These results are in agreement
with the fact that IS1561rsquo is present in the genomes of all the members of the MTBC except in M microti
(Gordon et al 1999 Brodin et al 2002 Huard et al 2003) Itrsquos worth noting that M microti is quite
widespread in animals in France (Michelet et al 2015) and thus may also occur in environmental
matrices (Courtenay et al 2006) The refined RD4 qPCR assay was shown to be highly specific for
M bovis species in agreement with previous results (King et al 2015a) Although some strains gave Ct
gt 35 when concentrated in pure culture with these molecular assays these targets would not be
detected in natural samples due to extreme dilution in highly complex environmental DNAs
The DNA extraction method used in this study proved to be highly efficient in soils harboring
different physico-chemical characteristics (especially in clayey soils) (Dequiedt et al 2011 Locatelli et
al 2013) The large mass of samples (2 g) allowed a better representativeness of sampling to overcome
the potential heterogeneity of substrates ie a physical heterogeneity as well as a heterogeneous
distribution of mycobacteria in environmental samples The use of an amplification facilitator (T4
bacteriophage gene 32 product) allowed minimizing PCR inhibition by co-extracted contaminants
(Matheson et al 2010 Opel et al 2010) and significantly improving M bovis detection In previous
Chapitre 3
113
published studies (Travis et al 2011 Pontiroli et al 2011 Sweeney et al 2007) DNA extraction was
usually achieved from smaller masses (01 to 05 g) of soil or feces using commercial DNA extraction kit
in order to limit PCR inhibition
The three qPCR assays developed in this study using 20-fold dilutions of DNA extracts of spiked
samples allowed a reliable detection of spiked M bovis (BCG strain) at levels as low as 25 times 102 CFU to
25 times 103 CFU g-1 of sample providing an improved detection limit compared to a recent study where
detection limit was 40 times 104 CFU g-1 soil and 50 times 105 CFU ml-1 water (Santos et al 2015) In other
previous studies detection limits were estimated to 85 times 103 cells g1 for soil and 425 times 105 cells g1 for
fecal samples using the original RD4 qPCR assay (Pontiroli et al 2011 Travis et al 2011) As cell numbers
were estimated by fluorescent dye assisted flow cytometry comparison with our results is difficult
Finally King et al (King et al 2015a King et al 2015b) were able to detect 103 M bovis genome
equivalents per g-1 for badger feces
In our study the detection limits are equivalent for the three qPCR systems developed or refined
whatever the physico-chemical characteristics of the soils
The 10- to 100-fold discrepancy between bacterial numbers obtained using culture based methods and
qPCR based methods that we observed could be explained by the underestimation of cultivable
bacteria due to cell aggregation (linked to a strong hydrophobicity of mycobacteria cell wall) in
inoculum as well as the occurrence of viable but not cultivable cells (VBNC) and cell mortality (with
persistence of DNA) Similar discrepancies have already been observed in previous studies (eg Betelli
et al 2013)
Though similar detection thresholds were observed for the three systems in spiked samples
RD4 and RV3866 based qPCR assays seemed less sensitive than IS1561rsquo-based detection system in
environmental samples (range of 2 Ct between IS1561rsquo and the 2 other systems) However the
occurrence of RD4 based positive signals in several samples gave an additional proof of the specific
detection of M bovis Furthermore the direct spolygotyping of DNA extracted from environmental
matrices confirmed for the first time the presence of M bovis in several environmental samples in
France
All 63-pasture soil samples from the bTB endemic area in Cocircte drsquoOr were found to be negative
for the presence of MTBC (below the detection threshold) Failure to detect MTBC in pasture soil
samples can be related to the low prevalence of bTB in cattle in the sampling areas (herd prevalence
reached 08 in 2014 Cote drsquoOr Regional Veterinary Service) and the low prevalence of bTB infection
in badgers which territories overlap the sampled pastures (Payne et al 2012) This result might reflect
the high dilution rate of M bovis targets in soil DNA extracts which contains high numbers of other
microorganisms (abundant soil microbial communities containing up to 1010 genomes per g of soil)
The discontinuous (spatial and temporal) distribution of M bovis in soil pasture because of
Chapitre 3
114
intermittent andor low shedding (Neill et al 1988 Clifton-Hadley et al 1993) might also limit the
detection of M bovis in soil samples Despite the high number of soil sample analyzed financial and
time constraints have limited more intensive sampling efforts that could have overcome this limit
MTBC signal was detected in 101 (7 positive out of 67) of the water samples taken from
naturally occurring springs with the IS1561rsquo qPCR assay As the level of detection was close to the
detection threshold we were unable to quantify MTBC signals Interestingly one sample of nematodes
originated from sediments located in close vicinity of one of the positive sources was also found
positive for the presence of MTBC Although nematodes are known to host bacterial pathogen no data
is available concerning MTBC This intriguing result would require further investigations These water
springs (and accompanying flowing water) are located in pastures where both cattle and wildlife have
access for drinking In several pastures (four sites) water was sampled along the slope following the
gravitational water flow In two of these pastures we repeatedly found (two to three) positive samples
indicating multiple point contamination of spring and flowing water along these slopes (see Figure 3
for localization of positive samples in one of these pastures) This result is an important new finding of
our study that can support several hypotheses on the role of water in the dissemination of MTBC in
the environment and in animal contamination We can hypothesize from these results that bioaerosols
formation from contaminated water and subsequent inhalation by animals (cattle and wildlife) might
contribute to animal infection with MTBC members especially as the infective dose through this
transmission route is very low (lt 10 CFU) (Dean et al 2005 Corner et al 2007) Infection of animals
might also occur through oral route by repeatedly drinking contaminated water
A special effort was made to monitor the contamination of badger sett soils since bTB infected
badgers were detected in our study area Between 2009 and 2011 878 badgers had been trapped and
analyzed in the infected area of Cocircte drsquoOr (Payne et al 2012) Macroscopic lesions were present in 19
of them despite culture-based method detecting 56 of positive badgers Only one badger showed
widespread tuberculosis Moreover in this infected area numerous badger setts were located either
within pastures or in close vicinity (forest-pasture interface) Our study demonstrated an average
prevalence of MTBC of 73 (11 out of 150) in soils from badger setts with the highest gene copy
number up to 37 times 104 gene copies g-1 of soil observed among environmental samples analysed in our
study The prevalence of MTBC molecular signature in latrines was estimated to 7 (3 positive out of
21) Our results demonstrate that infected badgers might contaminate their close environment for the
first time in France Indeed infected badgers have been shown to excrete M bovis from the respiratory
digestive and urinary tracts and in bite wounds exudates (Gavier-Widen et al 2001 Corner et al 2012)
Our results are in agreement with previous findings from several studies in UK (Young et al 2005
Courtenay et al 2006 Sweeney et al 2007 King et al 2015a) that recovered M bovis from badger
setts latrines and adjacent fields with gene copy numbers ranging from 1 times 103 to 54 times 106 per g of
Chapitre 3
115
soil In Woodchester Park in UK where badgers populations have been extensively studied for more
than 20 years bTB prevalence was estimated around 30 in 2005 which might explain high numbers
of positive setts and latrines observed in this area (Courtenay et al 2006 King et al 2015a) As
suggested by Courtenay et al (Courtenay et al 2006) the low bTB prevalence of infected sett soils
observed in our study may be linked to the low prevalence and the low density of badgers excreting
the bacteria within their social group
Finally in soils of 7 badger setts we were able to detect M bovis signatures (RD4 positive qPCR)
either continuously (4 setts) or discontinuously (3 setts) The presence of M bovis was confirmed by
direct soil DNA spolygotyping (see Table 6) in soils of the setts The presence of M bovis molecular
signatures was proved over an eight month-period (despite the badgers moved out) These results are
in agreement with a previous study where positive molecular detection of M bovis was demonstrated
over a period of 15 months despite the badgers having moved out (Young et al 2005) To our
knowledge this is the first time that spoligotype SB0120 was observed by direct spoligotyping of soil
DNA this spoligotype being identical to the spoligotype of strains isolated from cattle and wildlife in
these sites (Payne et al 2012)
Although the molecular detection of MTBC or M bovis by specific qPCR assay doesnrsquot give any
information about mycobacterial viability Young et al (Young et al 2005) demonstrated that MTBC DNA
did not persist in soil for more than ten days suggesting that DNA detected in our environmental
samples could belong to viable cells
A remaining challenge is to develop adequate methods to cultivate M bovis from
environmental matrices Indeed culture based methods might also be impaired by the occurrence of
VBNC state or dormancy in M bovis present in environmental matrices as previously suggested by
Whittington et al (Whittington et al 2004) for Mycobacterium avium subsp paratuberculosis Further
work is needed to assess if VBNC or dormancy occur in M bovis under environmental conditions
Although we concentrated our sampling efforts on badger environment it is however
acknowledged that in Cocircte drsquoOr M bovis circulates in a multi-host system that includes also wild boar
and deer We can thus hypothesize that these wild animals can also contribute to the shedding of
M bovis in the environment
Further research is needed to understand the ecology of M bovis in the environment and its
epidemiological role in indirect transmission This knowledge would help to improve herd
management in reducing indirect contacts between livestock and wildlife in controlling water access
in areas considered as ldquoat riskrdquo and in preventing contacts with badger setts for example Biosecurity
should thus be an integral part of the long-term strategy for the management of bovine tuberculosis in
Cocircte drsquoOr
Chapitre 3
116
Acknowledgements
The authors thank F Chevalier and M Beacuteral from the CIREV (Cellule Inter-Reacutegionale
drsquoEpideacutemiologie Veacuteteacuterinaire) AE Taconet F Bartheacuteleacutemy and T Grosjean from the Cote drsquoOr Regional
Veterinary Service (DDPP21) We are grateful to the cattle breeders trappers and hunters who helped
us during this study We wish also to thank C Ferrier from ONCFS (National Game Hunting and Wildlife
Agency) Dominique Meunier and Luc Biju-Duval from INRA Dijon Ceacuteline Richomme from ANSES The
work was supported by the Conseil Reacutegional de Bourgogne and Conseil Geacuteneacuteral de Cocircte drsquoOr Interbev
GDS Cocircte drsquoOr
Conflict of interest
No conflict of interest declared
References
Adams AP Bolin SR Fine AE Bolin CA and Kaneene JB (2013) Comparison of PCR versus culture for detection of Mycobacterium bovis after experimental inoculation of various matrices held under environmental conditions for extended periods Appl Environ Microbiol 79 6501-6506
Aranaz A De Juan L Montero N Saacutenchez C Galka M Delso C Alvarez J Romero B Bezos J Vela AI Briones V Mateos A and Domiacutenguez L (2004) Bovine tuberculosis (Mycobacterium bovis) in wildlife in Spain J Clin Microbiol 42 2602-2608
Betelli L Duquenne P Grenouillet F Simon X Scherer E Gehin E and Hartmann A (2013) Development and evaluation of a method for the quantification of airborne Thermoactinomyces vulgaris by real-time PCR J Microbiol Methods 92 25-32
Biet F Boschiroli ML Thorel MF and Guilloteau LA (2005) Zoonotic aspects of Mycobacterium bovis and Mycobacterium avium-intracellulare complex (MAC) Vet Res 36 411-436
Boumlhm M Hutchings MR and White PCL (2009) Contact networks in a wildlife-livestock host community identifying high-risk individuals in the transmission of bovine TB among badgers and cattle PLoS One 4 e5016
Brodin P Eiglmeier K Marmiesse M Billault A Garnier T Niemann S Cole ST and Brosch R (2002) Bacterial artificial chromosome-based comparative genomic analysis identifies Mycobacterium microti as a natural ESAT-6 deletion mutant Infect Immun 70 5568-5578
Clifton-Hadley RS Wilesmith JW and Stuart Fa (1993) Mycobacterium bovis in the European badger (Meles meles) epidemiological findings in tuberculous badgers from a naturally infected population Epidemiol Infect 111 9-19
Corner LA OMeara D Costello E Lesellier S and Gormley E (2012) The distribution of Mycobacterium bovis infection in naturally infected badgers Vet J 194 166-172
Corner LA Costello E Lesellier S OMeara D Sleeman DP and Gormley E (2007) Experimental tuberculosis in the European badger (Meles meles) after endobronchial
Chapitre 3
117
inoculation of Mycobacterium bovis I Pathology and bacteriology Res Vet Sci 83 53-62
Courcoul A Moyen J-L Brugegravere L Faye S Heacutenault S Gares H and Boschiroli M-L (2014) Estimation of sensitivity and specificity of bacteriology histopathology and PCR for the confirmatory diagnosis of bovine tuberculosis using latent class analysis PLoS One 9 e90334
Courtenay O Reilly LA Sweeney FP Hibberd V Bryan S Ul-Hassan A Newman C Macdonald DW Delahay RJ Wilson GJ and Wellington EMH (2006) Is Mycobacterium bovis in the environment important for the persistence of bovine tuberculosis Biol Lett 71 1946-1952
Dean GS Rhodes SG Coad M Adam O Cockle PJ Clifford DJ Hewinson RG and Vordermeier HM (2005) Minimum infective dose of Mycobacterium bovis in cattle Infect Immun 73 6467-6471
Dequiedt S Saby NPa Lelievre M Jolivet C Thioulouse J Toutain B Arrouays D Bispo a Lemanceau P and Ranjard L (2011) Biogeographical patterns of soil molecular microbial biomass as influenced by soil characteristics and management Global Ecol Biogeogr 20 641-652
Falkinham JO (2009) Surrounded by mycobacteria nontuberculous mycobacteria in the human environment J Appl Microbiol 107 356-367
Fediaevsky A Courcoul A Boschiroli M-L and Reveillaud E (2014) Tuberculose bovine en France reacutesultats dune strateacutegie plus offensive Bull Epid Santeacute Anim Alim 64 4-11
Fine AE (2006) The role of indirect transmission in the epidemiology of bovine tuberculosis in cattle and white-tailed deer in Michigan Michigan Bovine Tuberculosis Bibliography and DatabasePaper httpdigitalcommonsunledumichbovinetb33
Fine AE Bolin Ca Gardiner JC and Kaneene JB (2011) A study of the persistence of Mycobacterium bovis in the environment under natural weather conditions in Michigan USA Vet Med Int 2011
Garnett BT Delahay RJ and Roper TJ (2002) Use of cattle farm resources by badgers (Meles meles) and risk of bovine tuberculosis (Mycobacterium bovis) transmission to cattle Proc R Soc Lond B Biol Sci 269 1487-1491
Gavier-Widen D Chambers MA Palmer N Newell DG and Hewinson RG (2001) Pathology of natural Mycobacterium bovis infection in European badgers (Meles meles) and its relationship with bacterial excretion Vet Rec 148 299-304
Ghodbane R Mba Medie F Lepidi H Nappez C and Drancourt M (2014) Long-term survival of tuberculosis complex mycobacteria in soil Microbiology 160 496-501
Gordon SV Heym B Parkhill J Barrell B and Cole ST (1999) New insertion sequences and a novel repeated sequence in the genome of Mycobacterium tuberculosis H37Rv Microbiology 145 881-892
Gortaacutezar C Torres MJ Vicente J Acevedo P Reglero M de la Fuente J Negro JJ and Aznar-Martiacuten J (2008) Bovine tuberculosis in Dontildeana Biosphere Reserve The role of wild ungulates as disease reservoirs in the last Iberian lynx strongholds PLoS One 3 e2776
Chapitre 3
118
Hauer A Cruz KD Cochard T Godreuil S Karaoui C Henault S Bulach T Banuls AL Biet F and Boschiroli M-L (2015) Genetic evolution of Mycobacterium bovis causing tuberculosis in livestock and wildlife in France since 1978 PLoS One 10 e0117103
Huard RC de Oliveira Lazzarini LC Butler WR van Soolingen D and Ho JL (2003) PCR-based method to differentiate the subspecies of the Mycobacterium tuberculosis complex on the basis of genomic deletions J Clin Microbiol 41 1637-1650
Huard RC Fabre M de Haas P Lazzarini LCO van Soolingen D Cousins D and Ho JL (2006) Novel genetic polymorphisms that further delineate the phylogeny of the Mycobacterium tuberculosis complex J Bacteriol 188 4271-4287
King HC Murphy A James P Travis E Porter D Hung Y-J Sawyer J Cork J Delahay RJ Gaze W Courtenay O and Wellington EM (2015a) The variability and seasonality of the environmental reservoir of Mycobacterium bovis shed by wild European badgers Sci Rep 5 1-7
King HC Murphy A James P Travis E Porter D Sawyer J Cork J Delahay RJ Gaze W Courtenay O and Wellington EM (2015b) Performance of a noninvasive test for detecting Mycobacterium bovis shedding in European badger (Meles meles) populations J Clin Microbiol 53 2316-2323
Kukielka E Barasona JA Cowie CE Drewe JA Gortazar C Cotarelo I and Vicente J (2013) Spatial and temporal interactions between livestock and wildlife in South Central Spain assessed by camera traps Prev Vet Med 112 213-221
Little TW Naylor PF and Wilesmith JW (1982) Laboratory study of Mycobacterium bovis infection in badgers and calves Vet Rec 111 550-557
Locatelli A Depret G Jolivet C Henry S Dequiedt S Piveteau P and Hartmann A (2013) Nation-wide study of the occurrence of Listeria monocytogenes in French soils using culture-based and molecular detection methods J Microbiol Methods 93 242-250
Matheson CD Gurney C Esau N and Lehto R (2010) Assessing PCR inhibition from humic substances Open Enzym Inhib J 3 38-45
Michelet L de Cruz K Zanella G Aaziz R Bulach T Karoui C Heacutenault S Joncour G and Boschiroli ML (2015) Infection with Mycobacterium microti in animals in France J Clin Microbiol 53 981-985
Morris RS Pfeiffer DU and Jackson R (1994) The epidemiology of Mycobacterium bovis infections Vet Microbiol 40 153-177
Neill S Hanna J OBrien J and McCracken R (1988) Excretion of Mycobacterium bovis by experimentally infected cattle Vet Rec 123 340-343
Oostenbrink M (1960) Estimating nematode populations by some selected methods Nematology 6(8) 165-2015
Opel KL Chung D and McCord BR (2010) A study of PCR inhibition mechanisms using real time PCR J Forensic Sci 55 25-33
Palmer MV Waters WR and Whipple DL (2004) Investigation of the transmission of
Chapitre 3
119
Mycobacterium bovis from deer to cattle through indirect contact Am J Vet Res 65 1483-1489
Payne A (2014) Rocircle de la faune sauvage dans le systegraveme multi-hocirctes de Mycobacterium bovis et risque de transmission entre faune sauvage et bovins eacutetude expeacuterimentale en Cocircte drsquoOr In Epidemiologie Universiteacute Claude Bernard Lyon 1 httpwwwthesesfr2014LYO10040
Payne A Boschiroli ML Gueneau E Moyen JL Rambaud T Dufour B Gilot-Fromont E and Hars J (2012) Bovine tuberculosis in ldquoEurasianrdquo badgers (Meles meles) in France Eur J Wildlife Res 59 331-339
Phillips CJC Foster CRW Morris PA and Teverson R (2003) The transmission of Mycobacterium bovis infection to cattle Res Vet Sci 74 1-15
Pontiroli A Travis ER Sweeney FP Porter D Gaze WH Mason S Hibberd V Holden J Courtenay O and Wellington EMH (2011) Pathogen quantitation in complex matrices a multi-operator comparison of DNA extraction methods with a novel assessment of PCR inhibition PLoS One 6 e17916
Radomski N Roguet A Lucas FS Veyrier FJ Cambau E Accrombessi H Moilleron R Behr MA and Moulin L (2013) atpE gene as a new useful specific molecular target to quantify Mycobacterium in environmental samples BMC Microbiol 13 277
Richomme C Boschiroli ML Hars J Casabianca F and Ducrot C (2010) Bovine tuberculosis in livestock and wild boar on the Mediterranean island Corsica J Wildl Dis 46 627-631
Santos N Santos C Valente T Gortazar C Almeida V and Correia-Neves M (2015) Widespread environmental contamination with Mycobacterium tuberculosis Complex revealed by a molecular detection protocol PLoS One 10 e0142079
Slana I Kaevska M Kralik P Horvathova A and Pavlik I (2010) Distribution of Mycobacterium avium subsp avium and M a hominissuis in artificially infected pigs studied by culture and IS901 and IS1245 quantitative real time PCR Vet Microbiol 144 437-443
Sweeney FP Courtenay O Hibberd V Hewinson RG Reilly LA Gaze WH and Wellington EMH (2007) Environmental monitoring of Mycobacterium bovis in badger feces and badger sett soil by Real-Time PCR as confirmed by immunofluorescence immunocapture and cultivation Appl Environ Microbiol 73 7471-7473
Sweeney FP Courtenay O Ul-Hassan A Hibberd V Reilly LA and Wellington EMH (2006) Immunomagnetic recovery of Mycobacterium bovis from naturally infected environmental samples Lett Appl Microbiol 43 364-369
Telenti A Marchesi F Balz M Bally F Bottger EC and Bodmer T (1993) Rapid identification of mycobacteria to the species level by polymerase chain reaction and restriction enzyme analysis J Clin Microbiol 31 175-178
Thornton CG MacLellan KM Brink TL Lockwood DE Romagnoli M Turner J Merz WG Schwalbe RS Moody M Lue Y and Passen S (1998) Novel method for processing respiratory specimens for detection of mycobacteria by using C-18-carboxypropylbetaine Blinded study J Clin Microbiol 36 1996-2003
Travis ER Gaze WH Pontiroli A Sweeney FP Porter D Mason S Keeling MJC Jones RM
Chapitre 3
120
Sawyer J Aranaz A Rizaldos EC Cork J Delahay RJ Wilson GJ Hewinson RG Courtenay O and Wellington EMH (2011) An inter-laboratory validation of a real time PCR assay to measure host excretion of bacterial pathogens particularly of Mycobacterium bovis PLoS One 6 e27369
Whittington R Marshall D Nicholls PJ Marsh IB and Reddacliff LA (2004) Survival and dormancy of Mycobacterium avium subsp paratuberculosis in the environment Appl Environ Microbiol 70 2989-3004
Young JS Gormley E and Wellington EM (2005) Molecular detection of Mycobacterium bovis and Mycobacterium bovis BCG (Pasteur) in soil Appl Environ Microbiol 71 1946-1952
Zanella G Durand B Hars J Moutou F Garin-Bastuji B Duvauchelle A Ferme M Karoul C and Boschiroli ML (2008) Mycobacterium bovis in wildlife in France J Wildl Dis 44 99-108
Zhang J Abadia E Refregier G Tafaj S Boschiroli ML Guillard B Andremont A Ruimy R and Sola C (2010) Mycobacterium tuberculosis complex CRISPR genotyping improving efficiency throughput and discriminative power of spoligotyping with new spacers and a microbead-based hybridization assay J Med Microbiol 59 285-294
Chapitre 3
121
C CAMPAGNES DE PRELEVEMENTS SUPPLEMENTAIRES
Entre deacutecembre 2014 et mars 2015 deux campagnes de preacutelegravevement drsquoeacutechantillons
environnementaux suppleacutementaires ont eacuteteacute reacutealiseacutees dans la zone endeacutemique de Cocircte drsquoOr ainsi
qursquoune campagne en Charente ougrave des foyers bovins et sauvages sont recenseacutes respectivement depuis
2006 et 2009
1 Fin des preacutelegravevements en Cocircte drsquoOr
Les preacutelegravevements ont concerneacute drsquoune part une forecirct domaniale agrave cheval sur deux communes
de la zone Sud (eacutetoiles blanches sur la Figure 19) dans laquelle nous nous sommes focaliseacutes sur les
trous drsquoeau freacutequenteacutes par les sangliers et les cervideacutes Cinq trous drsquoeau importants et freacutequenteacutes ont
eacuteteacute cibleacutes et sur chacun trois preacutelegravevements de solsseacutediments et deux preacutelegravevements drsquoeau ont eacuteteacute
reacutealiseacutes en deacutecembre 2014 Drsquoautre part des preacutelegravevements ont eacuteteacute effectueacutes dans les terriers de
blaireaux infecteacutes de la commune D citeacutee dans lrsquoarticle preacuteceacutedent que nous suivons depuis mars 2014
et dans laquelle 1 agrave 3 blaireaux infecteacutes sont pieacutegeacutes tous les ans ainsi que dans des terriers freacutequenteacutes
drsquoune commune limitrophe (commune G) sur laquelle un blaireau infecteacute a eacuteteacute pieacutegeacute en 2014 Quatorze
sols de terriers (5 sur la commune D et 9 sur la commune G) et cinq latrines proches des terriers (3 sur
D et 2 sur G) ont eacuteteacute preacuteleveacutes en mars 2015 Ces deux sites sont repreacutesenteacutes par les eacutetoiles noires sur
la Figure 19 Tous ces eacutechantillons ont eacuteteacute analyseacutes selon les techniques deacutecrites dans lrsquoarticle
preacuteceacutedent (section Mateacuteriel et meacutethodes)
Les vingt-cinq eacutechantillons preacuteleveacutes dans la forecirct domaniale de la zone Sud en deacutecembre 2014
nrsquoont donneacute aucun signal moleacuteculaire avec le systegraveme de deacutetection ciblant lrsquoIS1561rsquo Dans cette zone
forestiegravere aucun animal sauvage infecteacute nrsquoa eacuteteacute deacutetecteacute depuis 2013
Concernant les terriers et latrines de la commune D les cinq eacutechantillons de sols de terriers
analyseacutes et une des deux latrines ont donneacute un signal moleacuteculaire positif pour le MTBC Pour lrsquoun des
sols la quantification nrsquoa pas eacuteteacute possible avec la qPCR (Ct gt 39 traces) Pour les quatre autres sols
des valeurs de 10 times 104 agrave 46 times 104 copies de gegravenes ont eacuteteacute obtenues et de 12 times 104 copies de gegravenes
pour la latrine Trois des eacutechantillons de sol ont donneacute un reacutesultat positif avec la PCR nicheacutee ciblant
lrsquoIS1561rsquo Dans ces terriers les preacutelegravevements reacutealiseacutes en mars avril mai juin et novembre 2014 puis
mars 2015 ont montreacute une persistance du signal moleacuteculaire au cours du temps Pour quatre des
terriers le signal a eacuteteacute deacutetecteacute agrave chaque campagne de preacutelegravevements alors que pour trois autres la
deacutetection a eacuteteacute intermittente Il est inteacuteressant de noter que les valeurs obtenues en mars 2015 sont
supeacuterieures agrave celles obtenues en 2014 (voir article preacuteceacutedent) Plusieurs hypothegraveses sont eacutemises pour
Chapitre 3
122
expliquer ce reacutesultat La premiegravere est lieacutee au fait que les terriers eacutetaient habiteacutes lors de nos
preacutelegravevements en mars 2015 alors qursquoils ne lrsquoeacutetaient plus apregraves les preacutelegravevements de mars 2014
La surveillance videacuteo reacutealiseacutee sur ces terriers a montreacute que les terriers ont agrave nouveau eacuteteacute
occupeacutes agrave partir de septembre 2014 Il est possible que cette occupation ait geacuteneacutereacute lrsquoexteacuteriorisation de
sol profond et de litiegravere potentiellement plus contamineacutes et que nous ayons preacuteleveacute ce sol drsquoorigine
profonde La seconde hypothegravese est lieacutee au fait que nous avons preacuteleveacute plus profondeacutement dans le
terrier que dans les campagnes de preacutelegravevements preacuteceacutedentes (utilisation drsquoun aspirateur) Enfin il est
eacutegalement possible que le groupe de blaireaux peuplant ce terrier possegravede un ou plusieurs animaux
infecteacutes et excreacuteteurs agrave lrsquoorigine drsquoune contamination plus importante du sol du terrier Ces terriers sont
situeacutes dans le bosquet drsquoun preacute qui fournit aux bovins de lrsquoombre et de la fraicheur en eacuteteacute et un abri en
cas de pluie ou de vent La surveillance videacuteo de cette zone nous a permis drsquoobserver que plusieurs
veaux ont reacuteguliegraverement exploreacute les entreacutees de gueules et consommeacute les brins drsquoherbe autour de ces
entreacutees (surveillance videacuteo)
Sur les neuf eacutechantillons de sol preacuteleveacutes dans la commune G deux ont donneacute un signal positif
pour le MTBC Pour lrsquoun drsquoeux seules des traces de M bovis ont eacuteteacute deacutetecteacutees pour lrsquoautre le nombre
de copies de gegravenes a eacuteteacute estimeacute agrave 12 times 104 La PCR nicheacutee srsquoest reacuteveacuteleacutee neacutegative pour les deux
eacutechantillons
Chapitre 3
123
Figure 19 Localisation des communes de Cocircte drsquoOr sur lesquelles de foyers bovins et des animaux sauvages infecteacutes ont eacuteteacute deacutetecteacutes et localisation des preacutelegravevements environnementaux effectueacutes en 2015 (les preacutelegravevements positifs sont repreacutesenteacutes par des eacutetoiles noires) (Reacutealisation Marina Beacuteral CIREV Bourgogne)
2 Preacutelegravevements en Charente
Depuis 2006 25 foyers bovins ont eacuteteacute deacutetecteacutes en Charente avec 12 foyers pour la seule anneacutee
2014 et la deacutecouverte de foyers anciens qui eacutevoluaient agrave bas bruit (communication personnelle
DDPP16) Depuis 2009 lrsquoinfection est mise en eacutevidence chez les blaireaux (6 positifs en 2013 et 11 en
2014) et les sangliers (8 positifs en 2014 donneacutees Sylvatub) et un chevreuil en 2015 (Communication
personnelle Sylvatub) autour des foyers bovins avec les mecircmes spoligotypes Le systegraveme multi-hocirctes
observeacute diffegravere donc de celui de la Cocircte drsquoOr puisqursquoagrave lrsquoheure actuelle lrsquoinfection nrsquoa pas eacuteteacute observeacutee
chez les cerfs dont les densiteacutes sont beaucoup moins importantes qursquoen Cocircte drsquoOr Les foyers bovins
Chapitre 3
124
et sauvages sont majoritairement localiseacutes dans le sud du deacutepartement dans une zone limitrophe de
la Dordogne deacutepartement fortement infecteacute depuis 2005 (Figure 20)
A la demande de la DDPP16 nous sommes alleacutes preacutelever des eacutechantillons environnementaux
dans le Sud du deacutepartement dans sept communes fortement toucheacutees par la maladie (Figure 20) Entre
avril et mai 2015 55 eacutechantillons environnementaux (50 eacutechantillons de sol de terrier de blaireau 3
sols de souille 1 latrine de blaireau et 1 fegraveces de sanglier) ont eacuteteacute preacuteleveacutes sur 7 communes de
Charente Ces communes font lrsquoobjet de la deacuteclaration drsquoun ou plusieurs foyers bovins de tuberculose
bovine etou drsquoanimaux sauvages infecteacutes (Figure 20) De plus 3 eacutechantillons de matiegraveres feacutecales
preacuteleveacutes pendant lrsquoautopsie de 3 blaireaux (BLAIR 7057 BLAIR 7058 et BLAIR 7061) ont eacuteteacute analyseacutes
Six eacutechantillons positifs ont eacuteteacute mis en eacutevidence sur les 55 analyseacutes La qPCR ciblant lrsquoIS1561rsquo a permis
de deacutetecter et de quantifier M bovis dans quatre ADN de sol de terrier avec un nombre de copies de
gegravenes variant de 11 times 103 agrave 54 times 103 par g de sol Deux eacutechantillons suppleacutementaires (1 sol de souille
et 1 sol de terrier) ont donneacute un signal non quantifiable avec la qPCR (traces) Tous les eacutechantillons
positifs ont eacuteteacute preacuteleveacutes sur une mecircme commune sur 2 lieux-dits diffeacuterents Maine-Joli (4 sols de
terrier) et la Tuilerie (1 sol de terrier et 1 sol de souille) A notre connaissance 2 blaireaux et 3 sangliers
infecteacutes ont eacuteteacute deacutecouverts agrave proximiteacute du lieu-dit la Tuilerie deacutebut 2015 (Source DDPP16) Lors de
notre campagne de preacutelegravevement tous les terriers deacutetecteacutes positifs eacutetaient freacutequenteacutes
Les ADN des eacutechantillons positifs ont eacuteteacute envoyeacutes agrave lrsquoANSES pour geacutenotypage Aucun ADN nrsquoa toutefois
pu ecirctre spoligotypeacute
Les fegraveces de blaireaux (BLAIR 7057 BLAIR 7058 et BLAIR 7061) preacuteleveacutes lors des autopsies nrsquoont donneacute
aucun signal avec la qPCR
Chapitre 3
125
Figure 20 Localisation des communes de Charente sur lesquelles de foyers bovins et des animaux sauvages infecteacutes ont eacuteteacute deacutetecteacutes et localisation des preacutelegravevements environnementaux effectueacutes en 2015 (les preacutelegravevements positifs sont repreacutesenteacutes par des eacutetoiles noires) (Reacutealisation Marina Beacuteral CIREV Bourgogne)
3 Compleacutements paysage geacuteomorphologie et peacutedologie
La zone endeacutemique Nord de tuberculose bovine en Cocircte drsquoOr que nous avons principalement
eacutetudieacutee est situeacutee sur un territoire appeleacute lrsquoAuxois caracteacuteriseacute par un paysage bocager ponctueacute de
buttes et de plateaux calcaires ougrave affleure un calcaire dur et karstique Les pacirctures suspectes
preacutesentent un contexte paysager hydrologique et geacuteomorphologique commun Le parcellaire suspect
est localiseacute dans trois petites valleacutees parallegraveles au fond desquelles circule un cours drsquoeau en direction
du Nord-Ouest (Figure 21) Chaque cours drsquoeau est alimenteacute par des sources naissant entre les couches
calcaires et marneuses du sous-sol dans les pentes Ces reacutesurgences drsquoeau proviennent de lrsquoinfiltration
des preacutecipitations dans les plateaux calcaires agrave lrsquoorigine du deacuteveloppement drsquoun reacuteseau karstique
drainant drsquoimportants eacutecoulements souterrains et la reacutesurgence drsquoeau en surface agrave cause des marnes
impermeacuteables du sous-sol Les pacirctures sont situeacutees sur les pentes douces marneuses des buttes ainsi
que dans les valleacutees alluviales humides Au-dessus des pacirctures un bourrelet boiseacute marque la rupture
Chapitre 3
126
de pente il preacutesente en geacuteneacuteral des affleurements plus ou moins importants de calcaire dur (Figure
22) Les pacirctures sont majoritairement situeacutees sur la mecircme formation geacuteologique des marnes du Lias
ou du Jurassique infeacuterieur correspondant agrave des formations peacutedologiques particuliegraveres caracteacuteriseacutees
par des sols de pentes argilo-limoneux localement hydromorphes (sources et laquo mouilles raquo)
Il est inteacuteressant drsquoobserver que les cantons toucheacutes par la tuberculose bovine en Cocircte drsquoOr
possegravedent une densiteacute de bovins allaitant quasi-identique agrave celle des cantons indemnes localiseacutes dans
le sud-ouest du deacutepartement et limitrophe de la zone drsquoendeacutemie de bTB (Figure 23) Ces deux groupes
de cantons sont localiseacutes sur des formations peacutedologiques diffeacuterentes suggeacuterant que le type de sol des
pacirctures pourrait ecirctre un facteur de risque dans le maintien de la maladie et que la densiteacute des bovins
ne semble pas ecirctre un facteur majeur dans la distribution des foyers de bTB Lrsquoobservation de la
distribution du parcellaire des exploitations foyers de tuberculose bovine sur la carte peacutedologique du
deacutepartement montre que 74 des parcelles de ces exploitations sont localiseacutees sur cette uniteacute
paysagegravere (sols argilo-limoneux et hydromorphes des pentes 48 des parcelles sols calcaires humides
sur alluvions des valleacutees 14 et sols argileux des plateaux calcaires 12 ) (Figure 24Figure 24)
Une enquecircte (Etude PatRisk) reacutealiseacutee dans le cadre drsquoun Master drsquoEpideacutemiologie dans neuf
eacutelevages bovins deacuteclareacutes infecteacutes entre 2010 et 2014 a chercheacute agrave identifier les pacirctures laquo agrave risque raquo de
tuberculose et agrave les caracteacuteriser par des variables concernant les pratiques drsquoeacutelevage le voisinage la
faune sauvage et lrsquoenvironnement (pente type de sol relief exposition humiditeacute) Lrsquoanalyse
multivarieacutee a mis en eacutevidence deux expositions indicatrices de risque la pratique du fauchage avant
la mise agrave lrsquoherbe et la preacutesence de zones humides localiseacutees sur la pacircture (Bouchez-Zacria 2015) Ces
reacutesultats sont en accord avec le fait que la survie de M bovis est favoriseacutee par la preacutesence drsquohumiditeacute
et qursquoune faible hauteur drsquoherbe augmente la possibiliteacute de contact avec les mycobacteacuteries du sol Bien
que des eacutetudes plus preacutecises soient recommandeacutees il semble toutefois que les sols argilo-limoneux
hydromorphes des pacirctures suspectes constituent un facteur de risque agrave prendre en compte dans
lrsquoeacutepideacutemiologie de la maladie
Chapitre 3
127
Figure 21 Orographie et hydrographie de lrsquoAuxois Les valleacutees supportant les parcelles suspectes sont indiqueacutees par une flegraveche noire (Source BD Carthage et BD alti - IGN)
Figure 22 Localisation de lrsquoAuxois en Cocircte drsquoOr structure paysagegravere et geacuteomorphologie de ce territoire
Chapitre 3
128
Figure 23 Nombre de vaches allaitantes par canton dans le deacutepartement de la Cocircte drsquoOr (agrave gauche) et parcellaire des foyers bovins de 2010 agrave 2015 (agrave droite) (Source Recensement agricole 2010 ndash Agreste)
Chapitre 3
129
Figure 24 Reacutepartition spatiale des parcelles des eacutelevages bovins infecteacutes de 2010 agrave 2014 en Cocircte drsquoOr et repreacutesentation des couches peacutedologiques (Reacutealisation Marina Beacuteral CIREV Bourgogne)
Chapitre 3
130
D BILAN GLOBAL
Nous avons deacuteveloppeacute deux systegravemes de deacutetection moleacuteculaire adapteacutes aux matrices
environnementales ciblant les espegraveces du MTBC (IS1561rsquo et Rv3866) et modifieacute un systegraveme existant
speacutecifique de M bovis (RD4) Nous avons valideacute ces systegravemes sur trois matrices diffeacuterentes (sol eau
fegraveces) inoculeacutees avec M bovis BCG en deacutemontrant leur speacutecificiteacute soit pour le MTBC (excluant
M microti potentiellement preacutesent dans lrsquoenvironnement) soit pour M bovis Leur sensibiliteacute est
bonne puisque les seuils de deacutetection obtenus sont infeacuterieurs aux seuils deacutecrits dans la litteacuterature (en
comparant avec une meacutethode de deacutenombrement identique agrave la nocirctre) Le systegraveme RD4 est agrave ce jour le
seul systegraveme de deacutetection speacutecifique de M bovis dans les eacutechantillons environnementaux toutefois
drsquoapregraves notre expeacuterience il preacutesente une sensibiliteacute moins importante que les deux autres systegravemes
malgreacute des seuils de deacutetection eacutequivalents sur les eacutechantillons inoculeacutes au laboratoire
La deacutetection du MTBC par lrsquoamplification de lrsquoIS1561rsquo a eacuteteacute effective dans 42 eacutechantillons (sur
plus de 400) la PCR nicheacutee ciblant la mecircme seacutequence a mis en eacutevidence 14 eacutechantillons positifs et le
systegraveme RD4 trois eacutechantillons positifs Les niveaux de deacutetection varient fortement en fonction des
eacutechantillons lrsquoeffet de dilution reste tregraves important dans des matrices comportant des communauteacutes
microbiennes abondantes et diversifieacutees (quantiteacute relative drsquoADN cible faible par rapport agrave des
quantiteacutes eacuteleveacutees drsquoADN non cible) Lrsquoutilisation de la PCR quantitative a permis dans certains cas de
quantifier les cibles preacutesentes dans les eacutechantillons environnementaux Les sols de terriers preacutesentent
les concentrations en mycobacteacuteries du MTBC les plus eacuteleveacutees pouvant atteindre 46 times 104 copies du
gegravene cibleacute ce qui correspond agrave des quantiteacutes de cellules eacuteleveacutees sachant que le nombre de copies du
gegravene cible par geacutenome est de 1
Parmi les diffeacuterents sites preacuteleveacutes la contamination a eacuteteacute retrouveacutee exclusivement dans deux
eacutecosystegravemes particuliers les zones humides de pacirctures (sources et laquo mouilles raquo) et les sols de terrier
et latrines de blaireaux La contamination apparait le plus souvent localiseacutee agrave une zone geacuteographique
et ponctuelle mais un signal moleacuteculaire positif a eacuteteacute retrouveacute pendant un an dans les sols de terriers
de blaireaux localiseacutes sur une pacircture
Ces reacutesultats correacuteleacutes au contexte paysager et peacutedologique des parcelles suspectes nous ont
ameneacute agrave penser que certaines formations peacutedologiques seraient plus susceptibles de favoriser la
persistance de M bovis Nous avons donc eacutevalueacute lrsquoimpact de la nature du sol retrouveacute dans ces
parcelles sur la survie de M bovis Nous avons eacutegalement eacutevalueacute lrsquoimpact de la tempeacuterature sur la
dureacutee de survie de la bacteacuterie dans le sol
CHAPITRE 4
Impact de la tempeacuterature et de la nature du sol sur la
survie de Mycobacterium bovis SB0120
Elodie Barbier Murielle Rochelet Laurent Gal Maria Laura Boschiroli Alain Hartmann
Experimental investigation of the impact of the temperature and the soil type on Mycobacterium
bovis survival in the environment
(Article en cours de soumission)
133
CHAPITRE 4 - Impact de la tempeacuterature et de la nature du sol sur la
survie de Mycobacterium bovis SB0120
A AVANT-PROPOS
La deacutetection moleacuteculaire de Mycobacterium bovis dans les eacutecosystegravemes agricoles de la zone
endeacutemique de Cocircte drsquoOr a confirmeacute que les animaux infecteacutes et excreacuteteurs ont la capaciteacute de
contaminer les sols et lrsquoeau accessibles agrave la faune domestique et sauvage (Chapitre 2) La bacteacuterie ne
se multipliant pas dans lrsquoenvironnement sa survie dans les substrats environnementaux conditionne la
transmission indirecte du bacille entre un animal infecteacute excreacuteteur et un animal sain par le biais de ces
substrats
Des eacutetudes expeacuterimentales sur divers substrats environnementaux notamment du sol
inoculeacutes avec M bovis ou M bovis BCG ont suggeacutereacute que les principaux facteurs influenccedilant la survie
des bacteacuteries sont les facteurs climatiques tels que tempeacuterature humiditeacute et exposition aux UV (Fine
et al 2011 Duffield amp Young 1985 Jackson et al 1995 Young et al 2005) Bien que quelques eacutetudes
aient rechercheacute par la culture ou par des approches statistiques une eacuteventuelle association de la
contamination environnementale ou animale par des mycobacteacuteries avec la nature du sol les reacutesultats
sont difficilement comparables et extrapolables (Walter et al 2012 Dhand et al 2009 Reviriego et al
2000 Johnson-Ifearulundu amp Kaneene 1999 Ward amp Perez 2004)
Face agrave ce manque de connaissances nous avons donc souhaiteacute eacutevaluer la dureacutee de la survie de
M bovis dans deux sols de Cocircte drsquoOr preacutesentant des caracteacuteristiques physico-chimiques contrasteacutees et
ce agrave deux tempeacuteratures diffeacuterentes afin de rechercher un eacuteventuel impact de la tempeacuterature et de la
nature du sol sur la persistance de la bacteacuterie Pour cette eacutetude nous avons utiliseacute une souche
pathogegravene de M bovis (geacutenotype SB0120) ce geacutenotype est retrouveacute majoritairement dans la zone
Nord chez les bovins et dans la faune sauvage Deux meacutethodes de suivi de la population bacteacuterienne
ont eacuteteacute utiliseacutees la culture sur milieu geacuteloseacute reacutealiseacutee agrave partir de sols irradieacutes et la qPCR pour le suivi
des sols irradieacutes et biotiques
135
B EXPERIMENTAL INVESTIGATION OF THE IMPACT OF THE TEMPERATURE AND THE SOIL TYPE ON MYCOBACTERIUM BOVIS SURVIVAL IN THE ENVIRONMENT
Elodie Barbierab Murielle Rocheletab Laurent Galac Maria Laura Boschirolid Alain Hartmanna
a INRA UMR 1347 Agroeacutecologie Dijon Cedex France
b Universiteacute de Bourgogne Franche Comteacute Dijon Cedex France
c AgroSup Dijon UMR1347 Agroeacutecologie Dijon France
d Universiteacute Paris-Est Laboratoire National de Reacutefeacuterence de la Tuberculose Uniteacute des Zoonoses Bacteacuteriennes Laboratoire de
Santeacute Animale ANSES Maisons-Alfort Cedex France
Key words Mycobacterium bovis soil temperature culture quantitative PCR
Corresponding Author Alain Hartmann UMR 1347 Agroeacutecologie INRA 17 rue Sully BP 86510 21065
Dijon Cedex France E-mail alainhartmanndijoninrafr
INRA UMR1347 Agroeacutecologie BP 86510 F-21000 Dijon France
Chapitre 4
136
Abstract
Mycobacterium bovis the causative agent of the bovine tuberculosis (bTB) mainly affect cattle its
natural reservoir but also a wide range of domestic and wild mammals Besides direct transmission via
contaminated aerosols indirect transmission of the disease between wildlife and livestock is thought
to occur by inhalation or ingestion of environmental substrates contaminated by M bovis through
infected animal shedding We monitored the survival of M bovis in two soils chosen for their contrasted
physico-chemical properties at two temperatures The population of M bovis spiked in disinfected soils
was enumerated by a culture-based method after 14 30 60 90 120 and 150 days of incubation at 4degC
and 22degC The fate of M bovis spiked in disinfected or biotic soils was monitored using a qPCR based
assay targeting the IS1561rsquo locus The analysis of survival profiles using culture-based method showed
that M bovis survived longer at cooler temperature but impact of soil characteristics on M bovis
persistence was not obvious Futhermore qPCR based assay detected M bovis longer than the culture
based method with higher gene copy numbers in sterile soils than in biotic ones Impact of soil type on
M bovis persistence need to be further studied in order to shed light on the risk and mechanisms of
indirect transmission of the disease
Chapitre 4
137
INTRODUCTION
Mycobacterium bovis is a pathogenic mycobacteria responsible for bovine tuberculosis (bTB)
Tuberculosis caused by M bovis is a primarily respiratory disease that may also affect various organs in
animals (Morris et al 1994) Even if bTB mainly concerns cattle a wide range of domestic and wild
mammals may be infected as well (Biet et al 2005) On the other hand bTB a zoonotic disease
impacting human health (Peacuterez-Lago et al 2013) Although cattle is considered as the principal host of
M bovis wild fauna such as badgers (Meles meles) wild boar (Sus scrofa) and red deer (Cervus elaphus)
has recently been identified as potential wildlife reservoirs (Palmer 2013) Though the transmission of
M bovis among cattle usually occurs via the inhalation of contaminated aerosols released by infected
animals andor the close contacts (Morris et al 1994 Phillips et al 2003) the indirect transmission
either by inhalation of environmental bioaerosols or ingestion of infected environmental matrices may
be another potential routes of animal infection (Vicente et al 2007 Palmer et al 2004 Phillips et al
2003) Indirect transmission through feed or water has already been demonstrated between deer deer
and cattle (Palmer et al 2004 Palmer et al 2001) and badgers and cattle (Little et al 1982) in laboratory
experiments
Investigating this type of indirect transmission is challenging because it results at least from the
combination of three essential factors iei) the environmental contamination by shedding from
infected animals ii) the persistence of the bacteria under a virulent state in environmental matrices
and finally iii) the interaction between a new susceptible host with the contaminated matrices The
shedding step by animals has already been demonstrated for many species via oro-nasal mucus
sputum urine feces and wound discharges depending on the animal species (Corner et al 2012
Palmer et al 2004 Barasona et al 2015) Secondly a few previous experimental studies using various
environmental substrates spiked with M bovis showed that the bacteria was able to survive for a long
time period outside of its host (Williams amp Hoy 1930 Duffield amp Young 1985 Young et al 2005 Fine et
al 2011 Ghodbane et al 2014) For example using a culture-based method Ghodbane et al (2014)
recovered M bovis bacteria from sterile soils incubated in controlled laboratory conditions 12 months
after their inoculation whereas an incubation under the natural weather conditions of Michigan USA
allowed the persistence of the bacteria in sterile soils for 88 days (Fine et al 2011) Moreover several
studies have identified several climatic factors such as a cool temperature an adequate moisture and
a protection against solar radiation (ultra violet) as positive factors for the persistence of M bovis in
environmental matrices like feces (Williams amp Hoy 1930 Tanner amp Michel 1999) food supply such as
corn hay apples (Palmer amp Whipple 2006 Fine et al 2011) water (Little et al 1982 Fine et al 2011)
and soil (Duffield amp Young 1985 Fine et al 2011) (Duffield amp Young 1985 Williams amp Hoy 1930 Tanner
amp Michel 1999 Jackson et al 1995) When Fine et al (2011) compared the persistence of M bovis in
Chapitre 4
138
four substrates (corn hay water and soil) the longest survival was found in cool and moist soil (88
days in winterspring) suggesting that soil probably ensured better conditions of persistence for
M bovis than the other matrices tested Moreover M bovis was often detected in soils especially in
badger sett soil or in pasture soil (Courtenay et al 2006 Santos et al 2015 Sweeney et al 2007
2006 Young et al 2005 Barbier et al manuscript submitted)
More generally low pH and high iron content are often described as major factors driving
mycobacterial survival in soils especially for Mycobacterium avium ssp paratuberculosis (Johnson-
Ifearulundu amp Kaneene 1999 Ward amp Perez 2004) Concerning M bovis one statistical study
established a relationship between the occurrence of bTB in animals and soil parameters by showing
that every a 1 increase in the sandy content of soil led to an increase in odds of bTB infection by 4
(Walter et al 2014) suggesting that soil type may promote or hinder M bovis environmental
persistence
To our knowledge no comprehensive study focuses on the influence of soil biotic and abiotic
parameters on the persistence M bovis With the aim to further assess the role of the soil
characteristics on the persistence of Mbovis and thus to better understand the impact of the
environment in the epidemiology of the bTB disease the influence of both the physicochemical
characteristics of the soil and the temperature was studied under controlled laboratory conditions For
this purpose two different soils selected for their contrasted properties were spiked with known
concentrations of M bovis and incubated at two different temperatures to mimic seasonal
temperatures The persistence of M bovis over time was monitored using both a culture-based
method for the detection of viable M bovis and a molecular detection method based on a qPCR assay
MATERIAL AND METHODS
All experiments were conducted in a biosafety level 3 laboratory unless otherwise stated
Mycobacterium bovis cultivation and inoculum preparation
The strain Mycobacterium bovis SB0120 VNTR 5 5 4 3 11 4 5 6 that infects cattle and wild
animals such as badgers and wild boar in France was provided by the National Reference Laboratory of
bovine tuberculosis (Maisons-Alfort France) and grown aerobically at 37degC in Middlebrook 7H9
supplemented with 10 Middlebrook ADC Growth Supplement (Sigma-Aldrich France) for thirty days
The M bovis inoculum was prepared as following a 20 mL stationary phase culture was pelleted and
once the supernatant removed the cell aggregates were disrupted by vortexing with glass beads for 15
seconds The pellet was suspended in 1 mL of sterile water constituting the stock solution and serial
10-fold dilutions were subsequently plated in triplicate onto Middlebrook 7H11 supplemented with
10 heat inactivated bovine serum (Dominique Dutscher France) and 10 Middlebrook OADC
Chapitre 4
139
Growth Supplement (Sigma-Aldrich France) for enumeration Colonies were counted following
incubation for two months at 37degC The inoculum for the culture-based assay and the qPCR-based assay
were respectively prepared by diluting 10-fold and 1000-fold the M bovis stock solution
Soil characterization and microcosm preparation
Two different soils were collected in two cattle-grazed pastures at 0-30 cm depth in Cocircte drsquoOr
(a department located in the Middle East of France) The soil A is a clay loamy soil (calcisol) and soil B
is a loamy sandy soil Soils were prepared as following soil samples were gently air dried and sieved to
recover 4 mm aggregates The moisture contents of soils were determined by drying for 24 h at 105degC
and their physico-chemical parameters were determined by the Soil Analysis Laboratory of Arras
France Fresh sieved soil was used to prepare the microcosms
Vials of 15 mL (Falcon Dominique Dutscher France) were filled with 2 g of the soils A and B Half of the
soil vials were sterilized by gamma-irradiation up to an internal minimum dose of 35 KGray (Ionisos
France) All the microcosm soils (biotic and sterile) were then adjusted to 90 of the water holding
capacity We verified that neither soil A nor B were positive for M bovis with the culture-based method
(sterile soils) and with the qPCR assay (sterile and biotic soils) described later All the soil microcosms
were spiked with 100 microL of the M bovis inoculum 10- or 1000-fold diluted (for sterile and biotic soil
respectively) and stored in the dark in a biosafety level 3 laboratory until their analysis Sterile spiked
soils were incubated at 4degC and 22degC while biotic spiked soils were only incubated at 22degC Vials were
opened in a microbiological safety cabinet every fifteen days to regenerate their atmosphere
Schedule of microcosm sampling
In order to monitor M bovis cell numbers over time both plate enumeration and qPCR assay
were performed on irradiated soils while qPCR assay only was carried out for biotic soils At each
sampling date ie day 0 14 30 60 90 120 and 150 three mesocosms of each biotic and sterile soils
were analysed with the qPCR assay after DNA extractionpurification and four mesocosms of each
sterile soil at each temperature were analysed using culture-based method
DNA extraction and purification
DNAs from the soil samples were extracted as follows 8 mL of lysis buffer (100 mM Tris pH 80
100 mM EDTA pH 80 100 mM NaCl and 2 (wv) SDS) 4 g of silica beads (100 μm diameter) 5 g of
ceramic beads (14 mm diameter) and 8 glass beads (4 mm diameter) were added to each vial of soil
However as DNA extraction could not be achieved within the biosafety level 3 laboratory for practical
reason soil was suspended in 8 mL of lysis buffer and heated 2 hours at 80degC to kill viable bacteria The
following steps of DNA extraction were then done in a biosafety level 2 laboratory Samples were
Chapitre 4
140
disrupted for 3 times 30 s at 4 ms in a FastPrepreg-24 Instrument (MP Biomedicals Europe France)
incubated for 30 min at 70degC and centrifuged for 5 min at 7000 timesg at room temperature Supernatants
were incubated on ice for 10 min with 110 volume of 3 M potassium acetate pH 55 and then
centrifuged for 5 min at 14000 times g One volume of ice-cold isopropanol was added to the supernatant
for DNA precipitation overnight at minus 20degC DNA was collected by centrifugation (30 min at 14000 times g)
and DNA pellets were washed with ice-cold ethanol (70 ) and dissolved in 100 μL of water Crude DNA
extracts (100 μL) were loaded on Microbiospintrade columns (Biorad France) filled with 100 mg of
polyvinyl polypyrrolidone (PVPP Sigma Chemical Co France) After centrifugation (4 min at 1000 times g
4 degC) eluted DNA was further purified using the Geneclean Turbo Kit (Qiagen France) according to the
manufacturers instructions Purified DNA concentrations were estimated by spectrophotometry (λ =
260 nm) using a NanoDropreg 2000 spectrophotometer (Thermo Scientific France)
Estimation of DNA quality for PCR amplification
The quality of the purified DNAs extracted from the two soils was assessed by measuring the
capacity to amplify a control DNA target in the presence of various dilutions of soil DNA extracts Briefly
qPCR reactions were done in a final volume of 25 microL containing 25 μl of 106 copies of the circularized
pGEM-T Easy plasmid DNA and 5 μl of water (control) or diluted soil DNA (10- and 20-fold dilutions)
1X Absolute qPCR SYBR Green ROX (500 nM) Mix (Thermo Scientific) 25ngmicroL of T4 bacteriophage
gene 32 product qPCR primers were SP6 (5rsquondashATTTAGGTGACACTATAG-3rsquo) and T7 (5rsquo-
TAATACGACTCACTATAGGG-3rsquo) universal primers (final concentration 500 nM) targeting the polylinker
of the plasmid qPCR and cycling conditions were as following initial denaturation at 95degC for 15 min
followed by 40 cycles with denaturation at 95degC for 15 s annealing at 55degC and elongation at 72degC for
30 s each The analysis of the melting curves allowed to assess PCR specificity A tenfold reduction of
the copy number amplified by qPCR in the presence of diluted soil DNA with respect to the copy
number obtained in the control reaction (water) was considered as 10 amplification efficiency or
conversely as 90 inhibition of the PCR
Cuture-based method
The sterile soil microcosms were suspended in 9 mL of Middlebrook 7H9 supplemented with
ADC by vortexing the vials for 2 min Tenfold dilutions of soil suspensions were achieved and 100 microL
were plated in triplicate onto Middlebrook 7H11 agar enriched as described above and supplemented
with antibiotics and antifungal (100 mgL-1Ticarcillin 10 mgL-1 Trimethoprim 200 000 UIL-1 Polymyxin
B and 100 mgL-1 Cycloheximid Sigma-Aldrich France) for enumeration Plates were then incubated in
gas-permeable bags at 37degC for 2 months and checked every 15 days
Chapitre 4
141
Mycobacterium tuberculosis complex quantitative PCR (qPCR)
Mycobacterium bovis was detected using the Mycobacterium tuberculosis complex (MTBC)
specific qPCR targeting the IS1561rsquo locus (Forward 5rsquo- GATCCAGGCCGAGAGAATCTG -3rsquo Reverse 5rsquo-
GGACAAAAGCTTCGCCAAAA - 3rsquo and probe 5rsquo - FAM ACGGCGTTGATCCGATTCCGC TAMRA - 3rsquo) as
previously described (Barbier et al manuscript submitted for publication)
qPCR reactions were carried out in triplicate with a 25 microL reaction mix containing 125 microL of ABsolutetrade
QPCR ROX Mix (Thermo Scientific France) 1 microL (final concentration 25 ngmicroL-1) of T4 bacteriophage
gene 32 product (MP Biomedicals Europe France) 1 microL of each primer (final concentration 400 nmolL-
1) 05 microL of each probe (final concentration 200 nmol L-1) 4 microL of ultrapure water and 5 microL of 20-fold
diluted DNA Absolute quantification was achieved using standard DNA dilutions A recombinant
plasmid containing one copy of the IS1561rsquo fragment of M bovis BCG strain Pasteur 1173P2 cloned in
the pCRregII-TOPOreg vector (Invitrogen France) was used as a standard A calibration curve was obtained
by amplification of serial dilutions of the plasmid ranging from 1 to 106 copies per PCR reaction All
DNAs extracted from the soil samples were screened with the IS1561rsquo-based qPCR assay and results
were expressed as the number of M bovis gene copy number per g of soil qPCR assay was performed
in a VIIA 7 Real-Time PCR System (Thermo Fischer Scientific France) Initial denaturation was done at
95degC for 15 min followed by fourty-five cycles with denaturation at 95degC for 15 s annealing and
elongation at 60degC for 1 min Detection limits of this molecular system was previously determined in
soil (Barbier et al manuscript submitted for publication)
RESULTS
Physico-chemical characteristics of soils A and B
The physico-chemical characteristics of soils A and B are summarized in Table 1 The soil A is a
clay loamy soil with a slightly basic pH (775) and a high content of clay (clay account for 397 in
particle-size distribution and sand for only 118 ) while the soil B is a loamy sandy soil with an acidic
pH (546) and a high content of sand (sand account for 345 in particle-size distribution and clay for
only 192 ) Silt content was roughly equivalent in the two soils (485 g kg-1 for soil A vs 463 g kg-1 for
soil B) We chose to adjust soil moisture to 90 of their water field holding capacity leading to a
moisture content of 31 for soil A and 21 for soil B Finally the C-to-N ratio was 95 for soil A and
109 for soil B
Chapitre 4
142
Table 1 Physico-chemical parameters of soils A and B
Physical parameters Soil A Soil B Unit
Clay (lt 2 microm) 397 192 g kg-1
Silt 485 463 g kg-1
Sand 118 345 g kg-1
Water field capacity 352 247 g kg-1
Chemical parameters -
pH 775 546
Total C 361 292 g kg-1
Total N 38 269 g kg-1
C- to N- ratio 95 109 -
Organic matter 625 506 g kg-1
P 0031 0025 g kg-1
K 0158 00877 g kg-1
Ca 717 11 g kg-1
Na 00115 00129 g kg-1
Mg 00541 00789 g kg-1
Fe 0526 0227 g kg-1
CECa 183 108 cmol+kg-1b
aCEC Cation Exchange Capacity bcmol+kg-1 centimoles of positive cations per kg of soil
Effect of temperature and soil type on M bovis survival time measured by culture-based method
Theoretical detection limit of our culture-based method was estimated at 15 times 103 CFU g-1 soil
Total cultivable M bovis (CFU g-1 soil) were enumerated by the culture-based method described above
for each sterile soil incubated at 4degC and 22degC at day 0 14 30 60 90 120 and 150 At day 0 number
of M bovis was estimated at 15 (plusmn 06) times 107 CFU g-1 for soil A and 17 (plusmn 07) times 107 CFU g-1 for soil B by
the culture-based method
Two different patterns of mycobacterial survival were observed depending on the temperature
(Figure 1a and 1b) At 22degC M bovis cells decreased over the course of the experiment in the two
sterile soils A and B (Figure 1a) No more cultivable bacteria growth was observed from day 90 in soil B
and day 120 in soil A to the end of the experiment at day 150 Although M bovis SB0120 was detected
by culture for one month longer in soil A than in soil B no statistically significant difference of CFU
numbers was observed between the two soils at 22degC (Student-Newman-Keuls Test p = 0559) thus
no significant difference in M bovis survival was observed due to soil type
At 4degC M bovis cultivable cells were recovered in high concentrations in the two soils at every
sampling date (over the five months of the experiment) The M bovis CFU numbers remained stable
during the first 30 days in the two soils as indicated in Figure 1b We observed a slight decrease of the
Chapitre 4
143
CFU numbers in the two soils starting from day 60 to the end of the study In both soils M bovis CFU
numbers decreased significantly from day 0 to day 150 (Student-Newman-Keuls Test plt0001)
reaching 87 (plusmn 25) times 10 5 CFU g- 1 and 22 (plusmn 11) times 10 5 CFU g- 1 in soil A and B respectively
Effect of soil type and endogenous microbiota on M bovis survival time measured by qPCR assay
Survival of M bovis when biotic or sterile soils A and B were incubated at 22degC was monitored
using the qPCR based assay The M bovis gene copy numbers were estimated with IS1561rsquoqPCR-based
method for each sterile and biotic soils incubated at 22degC at day 0 14 30 60 90 120 and 150 (see
Figure 2) At day 0 number of M bovis gene copies per g of soil yielded 36 (plusmn 12) times 105 and 60 (plusmn 19)
times 105 respectively for sterile soils A and B and 12 (plusmn 07) times 105 and 25 (plusmn 15) times 105 respectively for
biotic soils A and B by the qPCR-based method
Overall high amounts of M bovis gene copies were detected in the four soils at the eight sampling
dates
A statistically significant interaction between time and soil condition (biotic vs sterile) was
observed when considering the two soils independently and dependently (Student-Newman-Keuls
Test plt0001) with higher amounts of gene copies recovered in sterile soils than in biotic ones from
day 0 to day 150 No significant differences were observed when comparing the gene copy numbers in
the two sterile soils or in the two biotic soils at each sampling date
Even though the M bovis gene copy numbers slightly oscillated in the four soils over the course of the
experiment a slight but significant decrease of these numbers was observed between day 0 and day
150 for each soil The highest decreases of M bovis were observed in the biotic soils with a rough loss
of 1 log for soil B and 075 log for soil A between day 0 and day 150 (Figure 2)
Finally qPCR assay allowed a longer M bovis detection in sterile soils A and B incubated at 22degC
than culture-based assay Indeed M bovis DNA was detected by qPCR for up to 150 days in the two
sterile soil samples while bacterial culture detected viable M bovis cells up to 60 days in soil D and 90
days in soil C
Chapitre 4
144
Figure 1 Detection of M bovis SB0120 by culture-based method in sterile soils A and B after incubation at 22degC (figure above) and 4degC (figure below) Error bars represent the standard deviation values on 8 replicates
Figure 2 Detection of M bovis gene copy numbers by IS1561rsquo qPCR-based assay on sterile and biotic soils A and B after incubation at 22degC Error bars represent standard deviation values on 9 replicates
10E+00
10E+01
10E+02
10E+03
10E+04
10E+05
10E+06
10E+07
10E+08
0 14 30 60 90 120 150
M b
ovi
s C
FU n
um
ber
per
g o
f
ster
ile s
oil
Sampling day
22degC
Soil A
Soil B
10E+00
10E+01
10E+02
10E+03
10E+04
10E+05
10E+06
10E+07
10E+08
0 14 30 60 90 120 150
M b
ovi
s C
FU n
um
ber
per
g o
f st
erile
so
il
Sampling day
4degC
Soil A
Soil B
10E+00
10E+01
10E+02
10E+03
10E+04
10E+05
10E+06
0 14 30 60 90 120 150
M b
ovi
s IS
15
61
co
py
nu
mb
er p
er g
of
ster
ile s
oil
Sampling day
Sterile soil A Sterile soil B Soil A Soil B
Chapitre 4
145
DISCUSSION
Contamination of the environment with M bovis is considered as essential for the persistence
of bovine tuberculosis in animals and its interspecific indirect transmission The survival of this
mycobacteria in soil has been documented (Duffield amp Young 1985 Jackson et al 1995 Fine et al 2011
Young et al 2005) but limited information is available on the soils used and the edaphic factors that
may affect M bovis persistence are poorly understood These experiments have dealt with controlled
or environmental parameters that promote or hinder M bovis persistence in soil such as temperature
moisture or UV exposition but to our knowledge impact of the soil type has not been thoroughly
addressed This study was designed to evaluate temperature and soil type impact on M bovis survival
In this study we demonstrated that M bovis SB0120 may survive several months in soil under
different laboratory conditions in two contrasted soils and that cool temperature (4degC) strongly
promotes M bovis persistence whatever the soil type while warmer temperature (22degC) shortens its
survival time Our results are in contradiction with a previous laboratory study on M bovis persistence
(Young et al 2005) where mycobacterial survival was longer at 25degC than at 4degC in a sterile sandy soil
(pH 65-68 60 days vs 16 days) but in agreement with previous field studies where lower temperature
increased M bovis survival time up to 88 days (Fine et al 2011 Jackson et al 1995)
We also pointed out that soil type may impact on M bovis survival time Indeed despite that
no statistical difference was observed the soil A that exhibits high clay and loam content and slightly
basic pH allowed persistence of cultivable M bovis for up to three months (90 days) while this
persistence was only two months in the acidic sandy loamy soil B when soils were incubated at 22degC
On the contrary a difference of M bovis survival was observed at 4degC between the two soils with a
better survival in soil A than in soil B suggesting an impact of soil type on M bovis survival at cool
temperature However we were hampered by the detection limit of the culture-based method in the
experiment on sterile soil at 22degC where CFU numbers and detection limit ovelaped after day 60
Enrichment broth are very useful to recover low number of bacteria in complex matrices such as
Salmonella typhimurium in food or Listeria monocytogenes in soil (Locatelli et al 2013 Zheng et al
2013) but mycobacterial enrichment broths are not yet available for mycobacteria
In our study clay soil with slightly basic pH seemed to promote M bovis SB0120 survival
although mycobacterial culture detection or persistence are usually associated with sandy soils
harbouring acidic pH as previously suggested by positive culture or statistical studies (Walter et al
2014 Johnson-Ifearulundu amp Kaneene 1999 Reviriego et al 2000 Santos et al 2015) suggesting an
improved survival of the mycobacteria such as M avium ssp paratuberculosis and M bovis in these
soils However a one year persistence of cultivable M bovis was demonstrated in a sandy soil with a
basic pH (83) spiked with 108 CFU g- 1 (Ghodbane et al 2014) suggesting that M bovis may survive a
Chapitre 4
146
long time in soils harbouring a wide range of pH Moreover iron content of soils A and B seemed to be
significantly higher than soils described in previous studies
In this study qPCR was able to detect the presence of M bovis DNA over a longer period of
time than bacterial culture in soils samples incubated at 22degC whatever the soil type As one previous
study reported that DNA of dead M bovis cells was no more amplified 10 days after cell death (Young
et al 2005) we hypothesized that our qPCR assay detected live M bovis up to 150 days after
inoculation of soils Viable M bovis cells have already been detected in sterile and biotic soil over a 15
month period targeting the 16S rRNA encoding gene (Young et al 2005) Physiological state of
pathogenic mycobacteria such as M bovis in soil is unknown but dormancy or a viable but non
cultivable state (VBNC) have been suggested to explain their survival in environmental matrices
(Whittington et al 2004 Adams et al 2013) Further analysis of the expression of dormancy-related
genes (Aacutelvarez et al 2009) under soil conditions might allow to support this hypothesis Further
researches are critical to ascertain the long bacterial survival in soil and to understand the genetic
mechanisms involved in soil persistence
Our laboratory conditions were far from field conditions where temperature fluctuations
moisture variations and UV radiation are detrimental to the mycobacteria Mycobacterium bovis
survival time obtained in this study is probably longer than survival time in environmental conditions
As the gap of knowledge is important about how M bovis survive in soils laboratory studies have to
be set out in order to determine the main factors involved in M bovis survival However experiments
dealing with the exposition of M bovis naturally or artificially contaminated samples to environmental
conditions must be carried out in specific open air enclosure with high levels of protection to exclude
mycobacterial dissemination (Fine et al 2011) or require climate chambers with a L3 biosafety level
Significant differences in the recovery of M bovis DNA between biotic and sterile soils were
observed with the qPCR-based assay from the first day of the experiment indicating that qPCR
detection is hampered when soil metagenomic DNA is concentrated although no inhibition was
observed
In our experiment the use two soils gave an insight on the impact of soil type on M bovis
survival However extensive studies on larger number of well-characterised and contrasted soils would
be required to target the key edaphic factors that promote M bovis survival in soil Further work is
needed to identify those key factors and determine if the population detected with qPCR consists of
non-culturable or dormant cells capable of resuscitation under specific growth conditions or following
inhalation or ingestion by animals
Chapitre 4
147
Funding information
The work was supported by the Regional Council of Burgundy (Elodie Barbier PhD fellowship) the Cocircte
drsquoOr General Council Interbev and the Cocircte drsquoOr GDS (Groupement de Deacutefense Sanitaire Livestock
health protection group) The funders had no role in study design data collection and interpretation
or the decision to submit the work for publication
Conflict of interest
The authors have no conflict of interest to declare
Acknowledgements
The authors would like to thank Jean-Marie Duez Julien Bador and Catherine Neuwirth from the
Plateau Technique de Biologie of the Hospital of Dijon
REFERENCES
Adams AP et al 2013 Comparison of PCR versus culture for detection of Mycobacterium bovis after experimental inoculation of various matrices held under environmental conditions for extended periods Applied and Environmental Microbiology 79(20) pp6501ndash6506
Aacutelvarez AH Estrada-Chaacutevez C amp Flores-Valdez MA 2009 Molecular findings and approaches spotlighting Mycobacterium bovis persistence in cattle Veterinary Research 40(3) pp1ndash16
Barasona JA et al 2015 DNA detection reveals Mycobacterium tuberculosis complex shedding routes in its wildlife reservoir the Eurasian wild boar Transboundary and Emerging Diseases (DECEMBER)
Biet F et al 2005 Zoonotic aspects of Mycobacterium bovis and Mycobacterium avium-intracellulare complex (MAC) Veterinary Research 36 pp411ndash436
Corner LAL et al 2012 The distribution of Mycobacterium bovis infection in naturally infected badgers Veterinary journal (London England 1997) 194(2) pp166ndash72
Courtenay O et al 2006 Is Mycobacterium bovis in the environment important for the persistence of bovine tuberculosis Biology letters 2(3) pp460ndash2
Dhand NK et al 2009 Association of farm soil characteristics with ovine Johnersquos disease in Australia Preventive Veterinary Medicine 89 pp110ndash120
Duffield BJ amp Young DA 1985 Survival of Mycobacterium bovis in defined environmental conditions Veterinary Microbiology 10 pp193ndash197
Fine AE et al 2011 A study of the persistence of Mycobacterium bovis in the environment under natural weather conditions in Michigan USA Veterinary medicine international 2011 p765430
Chapitre 4
148
Ghodbane R et al 2014 Long-term survival of tuberculosis complex mycobacteria in soil Microbiology (Reading England) 160(Pt 3) pp496ndash501
Jackson R De Lisle GW amp Morris RS 1995 A study of the environmental survival of Mycobacterium bovis on a farm in New Zealand New Zealand Veterinary Journal 43(7) pp346ndash352
Johnson-Ifearulundu Y amp Kaneene JB 1999 Distribution and environmental risk factors for paratuberculosis in dairy cattle herds in Michigan American journal of veterinary research 60(5) pp589ndash96
Little TW Naylor PF amp Wilesmith JW 1982 Laboratory study of Mycobacterium bovis infection in badgers and calves The Veterinary record 111(24) pp550ndash7
Locatelli A et al 2013 Nation-wide study of the occurrence of Listeria monocytogenes in French soils using culture-based and molecular detection methods Journal of microbiological methods 93(3) pp242ndash50
Morris RS Pfeiffer DU amp Jackson R 1994 The epidemiology of Mycobacterium bovis infections Veterinary microbiology 40(1-2) pp153ndash77
Palmer M V 2013 Mycobacterium bovis Characteristics of wildlife reservoir hosts Transboundary and Emerging Diseases 60(SUPPL1) pp1ndash13
Palmer M V Waters WR amp Whipple DL 2004 Investigation of the transmission of Mycobacterium bovis from deer to cattle through indirect contact American Journal of Veterinary Research 65(11) pp1483ndash1489
Palmer M V amp Whipple DL 2006 Survival of Mycobacterium bovis on feedstuffs commonly used as supplemental feed for white-tailed deer (Odocoileus virginianus) Journal of wildlife diseases 42(4) pp853ndash8
Palmer M V Whipple DL amp Waters WR 2001 Experimental deer-to-deer transmission of Mycobacterium bovis American journal of veterinary research 62(5) pp692ndash6
Peacuterez-Lago L Navarro Y amp Garciacutea-de-Viedma D 2013 Current knowledge and pending challenges in zoonosis caused by Mycobacterium bovis A review Research in Veterinary Science 97 ppS94ndashS100
Phillips CJC et al 2003 The transmission of Mycobacterium bovis infection to cattle Research in veterinary science 74(1) pp1ndash15
Reviriego FJ Moreno M a amp Domiacutenguez L 2000 Soil type as a putative risk factor of ovine and caprine paratuberculosis seropositivity in Spain Preventive Veterinary Medicine 43 pp43ndash51
Santos N et al 2015 Widespread environmental contamination with Mycobacterium tuberculosis Complex revealed by a molecular detection protocol Plos One 10(11) pe0142079
Sweeney FP et al 2007 Environmental monitoring of Mycobacterium bovis in badger feces and badger sett soil by real-time PCR as confirmed by immunofluorescence immunocapture and cultivation Applied and environmental microbiology 73(22) pp7471ndash3
Chapitre 4
149
Sweeney FP et al 2006 Immunomagnetic recovery of Mycobacterium bovis from naturally infected environmental samples Letters in applied microbiology 43(4) pp364ndash9
Tanner M amp Michel AL 1999 Investigation of the viability of M bovis under different environmental conditions in the Kruger National Park Journal of Veterinary Research (66) pp185ndash190
Vicente J et al 2007 Risk factors associated with the prevalence of tuberculosis-like lesions in fenced wild boar and red deer in south central Spain Veterinary Research 38(3) pp451ndash464
Walter WD et al 2014 Linking bovine tuberculosis on cattle farms to white-tailed deer and environmental variables using Bayesian hierarchical analysis PloS one 9(3) pe90925
Walter WD et al 2012 On-farm mitigation of transmission of tuberculosis from white-tailed deer to cattle literature review and recommendations Veterinary medicine international 2012 p616318
Ward MP amp Perez AM 2004 Association between soil type and paratuberculosis in cattle herds American Journal of Veterinary Research 65(1) pp10ndash14
Whittington R et al 2004 Survival and dormancy of Mycobacterium avium subsp paratuberculosis in the environment Applied and environmental microbiology 70(5) pp2989ndash3004
Williams RS amp Hoy WA 1930 The viability of B tuberculosis (bovinus) on pasture land in stored faeces and in liquid manure The Journal of hygiene 30(4) pp413ndash419
Young JS Gormley E amp Wellington EMH 2005 Molecular detection of Mycobacterium bovis and Mycobacterium bovis BCG (Pasteur) in soil Applied and environmental microbiology 71(4) pp1946ndash1952
Zheng Q et al 2013 Comparison of enrichment broths for the recovery of healthy and heat-injured Salmonella typhimurium on raw duck wings Journal of food protection 76(11) pp1963ndash8
Chapitre 4
150
C BILAN
Les reacutesultats preacutesenteacutes dans ce chapitre deacutemontrent la capaciteacute de M bovis SB0120 agrave survivre
plusieurs mois dans les deux sols de Cocircte drsquoOr sous une forme viable cette dureacutee eacutetant impacteacutee par
un facteur abiotique particulier la tempeacuterature Une survie drsquoau moins cinq mois est observeacutee agrave basse
tempeacuterature (4degC) dans les deux sols avec une tregraves faible diminution de la population bacteacuterienne
initialement inoculeacutee alors qursquoelle nrsquoest que de deux agrave trois mois agrave 22degC Bien que M bovis soit cultiveacutee
un mois de plus dans le sol C par rapport au sol D nous nrsquoavons pas mis en eacutevidence drsquoeffet de la nature
du sol sur la survie de la bacteacuterie agrave 22degC Toutefois agrave cause de contraintes techniques notre analyse
nrsquoa porteacute que sur deux sols limitant fortement lrsquointerpreacutetation des reacutesultats Lrsquoanalyse des sols inoculeacutes
et incubeacutes agrave 22degC par qPCR montre un tout autre profil de survie puisque M bovis est deacutetecteacutee en
quantiteacute importante pendant les cinq mois de lrsquoeacutetude sur les deux sols et les deux conditions (biotique
vs steacuterile)
Les reacutesultats divergents obtenus par la culture et la qPCR sur les sols incubeacutes agrave 22degC amegravenent
agrave faire diffeacuterentes hypothegraveses pour expliquer lrsquoabsence de croissance des bacteacuteries puisque la qPCR
deacutetecte lrsquoADN des cellules vivantes et mortes ainsi que lrsquoADN libre Nous pouvons donc envisager que
lrsquoADN deacutetecteacute entre 60 et 150 jours appartienne agrave des cellules viables non cultivables ou agrave des cellules
mortes entiegraveres ou lyseacutees Des eacutetudes compleacutementaires sont indispensables pour preacuteciser lrsquoeacutetat
physiologique des bacteacuteries devenues incultivables apregraves une incubation dans le sol
En extrapolant les reacutesultats obtenus aux conditions de tempeacuterature rencontreacutees en Cocircte drsquoOr
nous pouvons suggeacuterer une fenecirctre temporelle pendant laquelle la survie de M bovis dans le sol est
maximale La Figure 25 repreacutesente les tempeacuteratures minimales et maximales releveacutees agrave Dijon entre
janvier 2012 et deacutecembre 2015 Nous estimons que les tempeacuteratures rencontreacutees dans lrsquoAuxois sont
probablement plus proches des tempeacuteratures minimales observeacutees dans la plaine de Dijon-Longvic ougrave
se trouve la station meacuteteacuteo (anthropisation minimale altitude humiditeacute veacutegeacutetation) que des
maximales Ainsi la peacuteriode la plus agrave risque va de novembre agrave avril (tempeacuteratures minimales de
lrsquoair lt 5degC) et correspond agrave la fin de lrsquoautomne lrsquohiver et le deacutebut du printemps Durant le reste de
lrsquoanneacutee et en particulier en eacuteteacute (tempeacuteratures maximales de lrsquoair gt 22degC) les risques de survie de M
bovis semblent moins importants Ces conclusions restent des hypothegraveses dans la mesure ougrave la
tempeacuterature reacuteelle des sols nrsquoa pas eacuteteacute mesureacutee et la variation des autres facteurs climatiques nrsquoa pas
eacuteteacute prise en compte Cependant on peut penser qursquoen eacuteteacute (sous forte insolation) la tempeacuterature des
sols peut augmenter rapidement dans les quelques premiers centimegravetres de sol limitant la dureacutee de
survie de la bacteacuterie
Chapitre 4
151
Figure 25 Tempeacuteratures minimales et maximales observeacutees agrave Dijon-Longvic de 2012 agrave 2015 (Source Meacuteteacuteo France)
-5
0
5
10
15
20
25
30
35
1 2 3 4 5 6 7 8 9 10 11 12
Tmin2012
Tmin2013
Tmin2014
Tmin2015
Tmax2012
Tmax2013
Tmax2014
Tmax2015
4degC
22degC
CHAPITRE 5
Rocircle de la faune du sol dans la circulation de
Mycobacterium bovis eacutetude de la disseacutemination de
M bovis par les lombrics
E Barbier B Chantemesse M Rochelet L Fayolle L Bollache M L Boschiroli A Hartmann
Rapid dissemination of Mycobacterium bovis from cattle dung to soil by the earthworm
Lumbricus terrestris
(Article accepteacute en cours de publication Veterinary Microbiology)
155
CHAPITRE 5 - Rocircle de la faune du sol dans la circulation de
Mycobacterium bovis eacutetude de sa disseacutemination par les lombrics
A AVANT-PROPOS
Le rocircle de la faune et de la microfaune du sol dans la survie et la disseacutemination de
Mycobacterium bovis dans lrsquoenvironnement est inconnu aujourdrsquohui Comme nous lrsquoavons eacutevoqueacute dans
la synthegravese bibliographique les protozoaires plus particuliegraverement les amibes jouent un rocircle
vraisemblable dans la survie des mycobacteacuteries environnementales (Adekambi et al 2006 Mba Medie
et al 2011 Mura amp Bull 2006) voire dans la virulence des mycobacteacuteries opportunistes (Cirillo et al
1997) In vitro la survie intra-amibienne de M bovis pendant plusieurs mois a eacuteteacute deacutemontreacutee mais
aucune eacutetude de terrain eacutevaluant le rocircle potentiel des amibes comme reacuteservoir de M bovis nrsquoest
disponible Concernant la meacutesofaune les lombrics ont eacuteteacute identifieacutes comme vecteur potentiel dans la
transmission de M avium ssp paratuberculosis dans des eacutelevages bovins et caprins infecteacutes (Fischer
et al 2003) Veacuteritables ingeacutenieurs du sol ces animaux ont un rocircle important dans la circulation des
microorganismes en les transfeacuterant et les seacutelectionnant depuis la matiegravere organique qursquoils ingegraverent et
dont ils sont de gros consommateurs vers le sol qui les entoure par le biais de leurs turricules
Afin drsquoeacutevaluer le rocircle potentiel des lombrics dans la disseacutemination de M bovis dans le sol suite
agrave lrsquoingestion de matiegraveres feacutecales animales contamineacutees nous avons eacutetudieacute le transfert de M bovis BCG
depuis les fegraveces inoculeacutes vers le sol environnant par une espegravece de lombric aneacutecique Lumbricus
terrestris La population bacteacuterienne de M bovis a eacuteteacute deacutetecteacutee et quantifieacutee par qPCR dans le sol
environnant les turricules et le tube digestif des lombrics
Chapitre 5
157
B RAPID DISSEMINATION OF M BOVIS FROM CATTLE DUNG TO SOIL BY THE EARTHWORM LUMBRICUS TERRESTRIS
Elodie Barbierab Benoit Chantemesseab Murielle Rocheletab Leacuteon Fayolle a Loiumlc Bollachebc
Maria Laura Boschiroli d Alain Hartmann a
a INRA UMR 1347 Agroeacutecologie Dijon Cedex France
b Universiteacute de Bourgogne Franche Comteacute Dijon Cedex France
c Laboratoire Chrono-environnement UMR CNRS 6249 Besanccedilon France
d Universiteacute Paris-Est Laboratoire National de Reacutefeacuterence de la Tuberculose Uniteacute de Zoonoses Bacteacuteriennes Laboratoire de
Santeacute Animale ANSES Maisons-Alfort Cedex France
Key words Mycobacterium bovis spreading earthworms Lumbricus terrestris cattle soil quantitative
PCR
Running title Environmental dissemination of Mycobacterium bovis by earthworms
Corresponding Author Alain Hartmann UMR 1347 Agroeacutecologie INRA 17 rue Sully BP 86510 21065
Dijon Cedex France E-mail alainhartmanndijoninrafr
Chapitre 5
158
Abstract
Indirect transmission of the bovine tuberculosis (bTB) between wildlife and livestock is thought to occur
by inhalation or ingestion of environmental substrates contaminated through animal shedding The
role of the soil fauna such as earthworms in the circulation of Mycobacterium bovis from
contaminated animal feces is of interest in the epidemiology of bTB The objective of this study was to
assess the impact of earthworm activity on M bovis transfer from animal dung to castings and the
surrounding soil For this purpose microcosms of soil containing the anecic eathworms Lumbricus
terrestris were prepared and covered with cattle feces spiked with the M bovis BCG strain Pasteur to
carry out two separate experiments The dissemination the gut carriage and the excretion of M bovis
were all monitored using a specific qPCR-based assay Our results showed that the earthworm L
terrestris was able to rapidly disseminate M bovis from the contaminated cattle feces to the
surrounding soil through casting egestion Moreover contaminated earthworms were shown to shed
the bacteria for 4 days when transferred in a bTB-free soil This study highlights for the first time the
possible role of earthworms in the dissemination and the persistence of M bovis in soils in bTB endemic
areas
Chapitre 5
159
INTRODUCTION
Mycobacterium bovis is a pathogenic mycobacteria that can infect and cause bovine
tuberculosis (bTB) in cattle and in a wide range of domestic or wild mammals as well as in humans
(Biet et al 2005) bTB is a primarily respiratory disease but it may affect various organs in animals
(Morris et al 1994) Although cattle are considered as the principal host of M bovis wild fauna such
as badgers (Meles meles) wild boar (Sus scrofa) and red deer (Cervus elaphus) has also been identified
as potential wildlife reservoirs (Palmer 2013) Transmission of bTB between cattle mainly occurs by
inhalation of contaminated aerosols during close contacts with infected animals (Morris et al 1994
Phillips et al 2003) An inoculum of less than ten bacilli is enough to cause respiratory disease in cattle
(Dean et al 2005) Moreover indirect transmission by inhalation of environmental bioaerosols or
ingestion of infected soil and water could also be a potential way of animal contamination (Palmer et
al 2004 Vicente et al 2007)
Infected animals were shown to shed M bovis via oro-nasal mucus sputum urine feces and
wound discharges (Corner et al 2012 Palmer et al 2004) that may contaminate environment
especially pastures shared by cattle and wild animals Thus investigation of soil as a potential reservoir
of M bovis is an important step towards understanding the epidemiology of the bacteria To reach this
goal deciphering the ecological factors that govern the survival and the spread of M bovis is first
required
Among soil fauna earthworms constitute the largest animal biomass in most terrestrial
ecosystems especially in grasslands (Lavelle and Spain 2001) Earthworms play a role of engineers in
soil processes impacting on soil functions and ecosystem services (Blouin et al 2013) affecting soil
structure water movement organic matter decomposition nutrients releasing plant growth and
pollution remediation Earthworms are usually classified in three ecological types according to their
feeding habits i) the epigeic ones are restricted to the surface organic layers where they feed
themselves ii) the endogeic ones live in the subsurface where they burrow lateral tunnels to ingest soil
and organic matter and iii) the anecic ones feed preferentially on surface litter but live in burrows in
the soil (Lee 1985) Earthworms ingest soil particles mixed with organic matter that are mechanically
and chemically digested by both gut fluid and microbiota (Curry and Schmidt 2007) and release
castings (undigested soil egested on or under the soil surface) Earthworm gut microbiota acts as a
selective filter for soil microorganisms shapes microbial communities of soil and either up- or down-
regulates microbial populations (Byzov et al 2007 Egert et al 2004 Goacutemez-Brandoacuten et al 2011) It
has been shown that earthworms may have microbial beneficial activities especially in the transport of
bacterial inocula through soil (Thorpe et al 1996) but earthworms may also act as pests by promoting
the dissemination of plant pathogens such as Phytophtora cactorum and Fusarium reticulatum
Chapitre 5
160
(Montecchio et al 2015) More interestingly a few studies have described that human and animal
pathogenic bacteria viruses and parasites especially Escherichia coli O157H7 foot-and-mouth disease
virus and Cestoda tapeworms may be carried and spread by earthworms (Edwards and Bohlen 1996
Prysor Williams et al 2006)
Although animal dung is a very attractive food for earthworms (Lowe and Butt 2005) few
studies have evaluated the role of earthworms in the survival and the dissemination of potential
pathogenic mycobacteria from animal dung or manure to surrounding soil (Fischer et al 2003) In
addition since earthworms represent an important part of the diet of various animal species including
badgers and wild boar (Granval and Muys 1995 Lee 1985) the consumption of contaminated
earthworms (gut carriage) might be a source of contamination for wild fauna
With the aim to investigate the above hypothesis this current study was first devoted to assess
the impact of earthworm activity on M bovis transfer from animal dung to their released castings and
the surrounding soil Then the M bovis gut carriage and the casting contamination following ingestion
of infected feces were investigated
MATERIAL AND METHODS
Mycobacterium bovis strain preparation
Mycobacterium bovis BCG strain Pasteur 1173P2 was grown aerobically at 37degC in Middlebrook
7H9 broth supplemented with 10 Middlebrook ADC Growth Supplement (Sigma-Aldrich France) The
M bovis inoculum was prepared as following a 10 mL stationary phase culture was pelleted and once
the supernatant removed the cell aggregates were disrupted by vortexing with glass beads for 15
seconds The pellet was suspended in 1 mL of sterile water and serial 10-fold dilutions of the suspension
were spiked on modified Middlebrook 7H11 plates (Middlebrook 7H11 supplemented with 10 heat
inactivated bovine serum (Dominique Dutscher France) and 10 Middlebrook OADC Growth
Supplement (Sigma-Aldrich France) for enumeration The plates were always incubated at 37degC for 2
months A 10-fold dilution of the prepared M bovis suspension was also directly quantified with the
RD4 qPCR assay described in the section ldquoMycobacterium bovis quantitative PCR (qPCR)rdquo
Soil cattle feces and earthworms characteristics
A bTB-free pasture (that has never been grazed by bTB infected cattle) located in Burgundy
(Middle East of France) was selected to sample the soil used in this study This latter is a clayey loamy
soil with a pH of 775 Organic carbon and nitrogen contents were 361 and 38 gkg-1 respectively The
soil was sieved to 4 mm and stored at 4degC until its use
Fresh cattle feces were collected from a bTB-free Charolais cattle in a bTB-free area of Burgundy and
Chapitre 5
161
immediately used
The adult anecic earthworms (Lumbricus terrestris) were provided by LombrirsquoCarraz (France)
and kept at 4degC in large buckets containing soil and organic matter until starting the study Soil
earthworms and cattle feces used in this work were previously proved to be negative for the presence
of M bovis using the qPCR-based assay
DNA extraction and purification
DNAs from all the samples collected were extracted as described previously (Dequiedt et al
2011) Briefly 8 mL of lysis buffer (100 mM Tris pH 80 100 mM EDTA pH 80 100 mM NaCl and 2
(wv) SDS) 4 g of silica beads (100 μm diameter) 5 g of ceramic beads (14 mm diameter) and 8 glass
beads (4 mm diameter) were added to 2 g of sample in a 15 ml sterile vial - whereas 1 mL of lysis
buffer 05 g of silica beads 0625 g of ceramic beads 1 glass beads were added to 250 mg-samples
Samples were disrupted for 3 times 30 s at 4 ms in a FastPrepreg-24 Instrument (MP Biomedicals Europe
Illkirch France) incubated for 30 min at 70 degC and centrifuged for 5 min at 7000 timesg at room
temperature Supernatants were incubated on ice for 10 min with 110 volume of 3 M potassium
acetate pH 55 and then centrifuged for 5 min at 14000 times g One volume of ice-cold isopropanol was
added to the supernatant for DNA precipitation overnight at minus 20 degC DNA was collected by
centrifugation (30 min at 14000 times g) and DNA pellets were washed with ice-cold ethanol (70) and
dissolved in 100 μl of water Crude DNA extracts (100 μl) were loaded on Microbiospintrade columns
(Biorad Marnes-La-Coquette France) filled with 100 mg of polyvinyl polypyrrolidone (PVPP Sigma
Chemical Co France) After centrifugation (4 min 1000 times g 4 degC) eluted DNA was further purified
using the Geneclean Turbo Kit (Qiagen Courtaboeuf France) according to the manufacturers
instructions Purified DNA concentrations were estimated by spectrophotometry (λ = 260 nm) using a
NanoDropreg 2000 spectrophotometer (Thermo Scientific Illkirch France)
Mycobacterium bovis quantitative PCR (qPCR)
Mycobacterium bovis-specific qPCR was set up targeting the RD4 flanking region using
previously described forward RD4 flanking primer (5rsquo-TGTGAATTCATACAAGCCGTAGTCG-3rsquo) and probe
(5rsquo-6-FAM-AGCGCAACACTCTTGGAGTGGCCTAC-3rsquo TAMRA) (Sweeney et al 2007) In order to improve
qPCR efficiency the reverse RD4 flanking primer based assay was modified (5rsquo-
ATCTTAGCTGGTCAATAGCCATTTTT-3rsquo) to reduce the size of the qPCR product to 116 bp
qPCR reactions were carried out in triplicate with a 25 microL reaction mix containing 125 microL of
ABsolutetrade QPCR ROX Mix (Thermo Scientific France) 1 microL (final concentration 25 ngmicroL-1) of T4
bacteriophage gene 32 product (MP Biomedicals Europe France) 1 microL of each primer (final
concentration 400 nmolL-1) 05 microL of each probe (final concentration 200 nmol L-1) 4 microL of ultrapure
Chapitre 5
162
water and 5 microL of pure or diluted (10- or 20-fold) DNA Absolute quantification was achieved using
standard DNA dilutions A recombinant plasmid containing one copy of the RD4 fragment of M bovis
BCG strain Pasteur 1173P2 cloned in the pCRregII-TOPOreg vector (Invitrogen France) was used as a
standard A calibration curve was obtained by amplification of serial dilutions of the plasmid ranging
from 1 to 106 copies per PCR reaction All DNA samples extracted from the soil castings and feces
samples were screened with the RD4-based qPCR assay and results were expressed as the number of
M bovis gene copies per g of soil castings or feces We previously assessed that when Cycle threshold
(Ct) was above 39 detection was possible but unreliable (unpublished results) The word ldquotracesrdquo was
then chosen in all further experiments in order to indicate that M bovis targets are present in very low
quantities (below 250 CFUg-1 of soil) The qPCR assay was performed in a VIIA 7 Real-Time PCR System
(Thermo Fischer Scientific France) Initial denaturation was done at 95degC for 15 min followed by 45
cycles with denaturation at 95degC for 15 s annealing and elongation at 60degC for 1 min
Mesocosms preparation
Plastic jars (15 L) were filled with 1 kg of the clay-loamy soil Sterile water was added to each
soil sample to reach final soil moisture of 80 of the water-holding capacity (WHC) Once the soil was
packed down to obtain a 1 gcm-3 soil density 6 earthworms Lumbricus terrestris were placed into each
jar A 4-days delay was necessary to acclimatize the earthworms Nine control mesocosms where no
earthworms were added were processed and analysed
Evaluation of earthworm dissemination of M bovis to soil mesocosms
Aliquots of 50 g of cattle feces previously spiked with 1 mL of M bovis BCG suspension were
layered on the soil surface of each mesocosm Jars were covered with pierced fitted lids and kept in a
dark room at 20degC in a L2 confinement facility (day 0)
In order to sample fresh earthworm castings all the castings were removed 48 h before each
sampling date A sampling schedule ie 2 4 7 10 14 21 30 45 and 60 days was followed For each
sampling date 2 mesocosms with earthworms and one control mesocosm were analysed as following
First earthworm castings were collected at the surface of each mesocosm containing earthworms
pooled homogenized and then aliquoted in triplicate in 2 g-samples Secondly the cattle feces and
earthworms were removed out of the mesocosms and the remaining soil was mixed and aliquoted in
triplicate in 2 g-samples Finally the six earthworms contained in each mesocosm were rinsed twice
with sterile water dried with a paper towel and individually transferred into a 60 mL empty vessel
After a 48h incubation at 20degC in a dark room to release the gut content one 250 mg-aliquot of fecal
samples was sampled for each earthworm In the control mesocosm the cattle feces were removed
from the top of mesocosm the soil was then mixed sampled and analysed as described above All
Chapitre 5
163
samples were frozen at -20degC until their DNA extraction and RD4 qPCR analysis Gene copy numbers
found in each sample were box-cox transformed before analysis to normalize the data Generalized
Linear Models (GLMs) were used to analyze the effects of sampling date (number of days after the
M bovis spiked cattle feces was layered onto the soil) and treatment (feces castings and soil) on the
number of CFU Tests were performed using JMP 70 Software (SAS Institute Inc) Results were
considered to be significantly different for P values lt 005
Gut carriage and excretion of M bovis by earthworms
Five mesocosms each containing 6 earthworms were prepared as described in the section
ldquomesocosm preparationrdquo and were then covered with 50 g of cattle feces spiked with 1 mL of M bovis
BCG suspension After 7 days of incubation each earthworm was rinsed twice with sterile water dried
with a paper towel To estimate M bovis carriage on earthworm tegument 6 earthworms out of the
30 were deposited in an individual Petri plate containing modified Middlebrook 7H11 media
supplemented with antibiotics and antifungal (100 mgL-1Ticarcillin 10 mgL-1 Trimethoprim 200 000
UIL-1 Polymyxin B and 100 mgL-1 Cycloheximid Sigma-Aldrich France) After gently moving it on the
plate surface for few minutes the earthworms were individually transferred in a 200 mL vessel
containing 100 g of M bovis-free fresh soil as well as the 24 earthworms This step corresponded to
the starting point of the experiment (day 0)
At each sampling date ie day 1 2 3 4 and 5 the castings from series of 6 randomly selected
remaining vessels were collected pooled homogenized and one aliquot of 2 g was sampled Then the
earthworms from 1 series of 6 vessels were removed rinsed twice with sterile water dried with a paper
towel and individually transferred in a 60 mL empty vessel After 48h at 20degC in a dark room the
animals have released their gut contents and a 250 mg-aliquot of fecal sample was recovered from
each vessel containing earthworms All of the samples were stored at -20degC until further analysis
RESULTS
Earthworm dissemination of M bovis from cattle feces spiked with the mycobacteria to soil
mesocosms
For this purpose the protocol sketched in Figure 1 was carried out over a 60 days-period and
the results are summarized in Figure 2 Both the culture-based method and the RD4 qPCR assay were
used to estimate the concentration of the Mycobacterium bovis BCG suspension used to spike the cattle
feces Different values of 125 times 107 CFU per mL and of (65 plusmn 05) times 108 gene copies per mL were
determined with the culture and the molecular methods respectively However given that only the
RD4 qPCR assay was carried out to quantify M bovis throughout this experiment the M bovis genome
Chapitre 5
164
equivalent number contained in the 50 g of cattle feces was estimated to (65 plusmn 05) times 108 at the
beginning of the experiment (day 0 dashed line in Figure 2)
Figure 1 Schematic representation of the procedure followed for the evaluation of M bovis dissemination in
soil by earthworms
Overall M bovis was repeatedly detected in all of the analysed samples ie earthworm castings
fecal samples and soils from 2 days after depositing the cattle feces spiked with M bovis on the surface
of the mesocosms Gene copy number was significantly influenced by the sampling date (df= 8 sup2 =
14508 Plt00001) treatment (df= 2 sup2 = 22430 Plt00001) and interaction between sampling date
and treatment (df= 16 sup2 = 14464 Plt00001) However at the end of the experiment (day 60)
M bovis gene copy numbers were not statistically different (Average and P value) whatever the kind of
samples More precisely the earthworm fecal samples (open circle Figure 2) showed the highest
contamination (up to (57 plusmn 03) times 106 M bovis gene copies per g) at day 2 while the M bovis gene copy
numbers in the fecal samples decreased (20-fold factor) after day 4 to reach a final value of (25 plusmn 22)
times 105 gene copy number per g On the other hand the M bovis gene copy numbers per g of earthworm
castings were up to around 10-fold lower than those per g of earthworm fecal samples until day 45
after which similar M bovis gene concentrations were measured (full circle Figure 2) Although soil
samples from mesocosms also led to a M bovis positive signal from day 2 (44 plusmn 42) times 104 gene copy
number per g) it is worth noting that only 4 out of the 6 analyzed samples were positive at day 2 (full
square Figure 2) Afterwards (ie from day 4) all of the soil samples gave a positive signal for M bovis
with a strong increase (10-fold) of the M bovis gene copy number per g of soil between day 2 and day
(D0 + 2 4 7 10 14 21 30 45)
Mesocosm Deposition of M bovis spiked feces
D0
CASTING POOL
FECAL SAMPLE
48h00
Earthworm
Fresh soil
M bovis spiked cattle feces
SOIL
Sampling day
Chapitre 5
165
7 followed by minor fluctuations until day 60 For the control mesocosms (ie without any
earthworms) no M bovis signal were measured in the soil samples throughout the experiment until
day 60 where a positive signal value of (35 plusmn 19) times 104 M bovis per g of soil was obtained (cross symbol
on Figure 2) thus indicating a very slow spread of M bovis in the bulk soil in the absence of earthworms
Finally it should be noticed that the cattle feces remained undisturbed in the control mesocosms over
the course of the experiment whereas the earthworms totally ingested the spiked cattle feces in all of
the mesocosms containing these animals during the first week of the experiment
Figure 2 Concentration of M bovis DNA determined by RD4 qPCR in soil earthworm castings and fecal samples recovered at each sampling date Concentrations of M bovis DNA in control mesocosms are represented with crosses (times) The dashed line corresponds to the concentration of M bovis spiked in 50 g of cattle feces Error bars represent the standard deviation values of 3 replicates
3
4
5
6
7
8
0 5 10 15 20 25 30 35 40 45 50 55 60
Lo
g1
0(M
b
ovi
sg
ene
cop
y n
um
ber
) in
1 g
of
sam
ple
Days
Chapitre 5
166
Table 1 RD4-based qPCR detection of M bovis gene copy number contained in the earthworm castings recovered according to the protocol of Figure 2
Sampling day Number of analysed
casting poolsa Number of positive
casting poolsb M bovis gene copy number per gc
1 5 5 (196 plusmn 136) times 105
2 4 4 (104 plusmn 060) times 105
3 3 2 Tracesd
4 2 1 Traces
5 1 0 Negative a
From 6 vessels b
Threshold cycle (Ct) between 20 and 39 c Mean value calculated with the signal obtained for the positive earthworm casting pools
d Detected but not quantifiable (below 250 CFUg-1 of soil)
Evaluation of the carriage and the excretion of M bovis by earthworms
In order to check whether earthworms carry M bovis in their gut and are able to excrete the
bacteria the following experiment was realized (see detailed protocol in Figure 3) Earthworms were
artificially contaminated by feeding for 7 days from cattle feces spiked with M bovis then their castings
and their fecal samples were analysed for a 5-days period First no growth of M bovis was observed
from earthworm tegument surfaces suggesting the absence or the low carriage of M bovis by
earthworm teguments The results shown in Table 1 indicated that all of the earthworms produced
castings highly contaminated with M bovis 1 and 2 days after their introduction in the fresh M bovis
free soil (up to around 105 M bovis gene copy number per g) Then the percentage of positive M bovis
casting pools gradually decreased (66 and 50 for sampling day 3 and 4 respectively) until the
sampling at day 5 for which no M bovis molecular signal was detected Although positive signals were
obtained for the sampling days 3 and 4 only traces of M bovis were detected (Ct above 39) With
values ranging from 1 to 5 105 M bovis gene copy number per g of fecal sample (Table 2) the RD4
qPCR assay yielded M bovis positive signals for the sampling days 1 to 4 Nevertheless for each of
these sampling days some earthworms (17-50 ) led to negative M bovis qPCR signal thus indicating
the absence or a very low amount of M bovis in the earthworm feces (below the detection limit of the
qPCR) Finally all of the six earthworm fecal samples analysed at day 5 were M bovis free
Chapitre 5
167
Figure 3 Schematic representation of the procedure followed for the estimation of M bovis gut carriage and excretion by earthworms
Table 2 RD4-based qPCR detection of M bovis gene copy number contained in the earthworm fecal samples recovered according to the protocol of Figure 3
Sampling day Number of positive
fecal samplesa (n = 6) M bovis gene copy number per gb
1 5 (123 plusmn 082) times 105
2 4 (228 plusmn 214) times 105
3 3 (542 plusmn 049) times 105
4 4 (213 plusmn 164) times 105
5 0 Negative
a Threshold cycle (Ct) between 20 and 39 b Mean value calculated with the signal obtained for the positive earthworm fecal samples
Sampling day (D0 + 1 2 3 4 5)Mesocosm
(n = 5)
Deposition of M bovisspiked feces (n = 5 )
Investigation of M bovis tegumentcarriage on 6 earthworms
D0
7 days
Transfer of each earthworm
5 series of 6 vessels(n = 30)
FECAL SAMPLE
CASTING POOL
n = 6
48h00
n = number of remainingseries of 6 vessels
Earthworm
Fresh soil
M bovis spiked cattle feces
Chapitre 5
168
DISCUSSION
The results shown in Figure 1 evidenced for the first time that earthworms were able to widely
transfer M bovis from cattle feces to their castings after ingestion of cattle feces spiked with the
bacteria Owing to the concomitant ingestion of M bovis-free soil and dung remains and thus the
dilution of M bovis cells in the gut content the number of M bovis cells in castings gradually decreased
from day 4 until the end of the experiment These results were in good agreement with earlier reported
laboratory trials with Mycobacterium avium subsp paratuberculosis that is responsible for the Johnersquos
disease in cattle and goat (Fischer et al 2003) Prysor Williams et al (Prysor Williams et al 2006) have
also reported the excretion of the human food-borne pathogen Escherichia coli O157H7 in castings
after ingestion of spiked manure by earthworms
On the other hand this study demonstrated that when earthworms were maintained in the
M bovis infected environment they were able to shed M bovis in their castings for at least 60 days
(Tableau 1) However when transferred in a M bovis-free soil earthworms only shed the bacteria up
to 4 days This data suggests that M bovis is unable to colonize the gut of the earthworm L terrestris
over extended periods Moreover L terrestris may ingest 490 to 3500 mg dry mass g-1 fresh mass of
arable soil per day (Curry and Schmidt 2007) and has a mean gut transit time of about 12 hours (Taylor
and Taylor 2014) leading to the rapid egestion of the bacteria from earthworm guts through castings
Furthermore earthworms were able to transfer M bovis from cattle feces to the bulk soil by
releasing castings in the tunnels they burrow Our results showed that M bovis was still detected in
mixed soil over the course of the experiment (60 days) highlighting a long persistence of M bovis in
soil Rapid dispersal of the bacteria in soil can be fully imputed to earthworm activity since soil from
control mesocosms without earthworms remained free from M bovis-except at day 60 Moreover
bacteria movement was mainly due to M bovis earthworm excretion since no M bovis tegument
carriage was observed The M bovis signal observed in control mesocosms at day 60 is probably due
to the moving of M bovis cells during moisture transfer from dung to soil surface and to the presence
of remaining dung particles in soil (incomplete removal of contaminated dung before soil mixing) As
M bovis is sensitive to solar radiation (UV) and soil dryness the burying of M bovis in deep soil by
earthworms may contribute to enhance its survival in soil environment Indeed survival of M bovis in
soil may be enhanced up to 2 years when the bacteria is buried to a depth of 5 cm (Genov 1965) Our
experimental design based on 1 kg-soil mesocosms did not allow estimating the real spatial distribution
of the bacilli in soil by earthworms It has been shown that earthworms are potentially able to move
several meters apart from the area where they graze organic matter (Mather and Christensen 1988)
thus disseminating soil-borne plan pathogen through large surfaces of soil (Thorpe et al 1996)
Concerning potential spread of animal or human pathogens earthworms were demonstrated in
Chapitre 5
169
experimental boxes to significantly aid vertical and lateral movement of ingested bacteria both up to
40 cm from the inoculation point (Prysor Williams et al 2006)
Finally we showed that earthworms may carry M bovis for many days in their guts even when
transferred in a M bovis-free environment Earthworms are known to host animal parasites and
pathogen such as protozoan tapeworms (Cestoda) and nematodes they can transmit to birds and
mammals when earthworms are ingested (Edwards and Bohlen 1996) However to our knowledge
the fact that earthworms may act as a source of contamination for animals feeding from earthworms
has never been studied Earthworms that constitute a nutritious food resource are an important part
of the diet of badgers and wild boar (Baubet et al 2003 Cleary et al 2011) that often forage in cattle
pastures hosting high earthworm populations (Muldowney et al 2003) Badgers and wild boar are
able to eat up to 200 and 120 earthworms respectively in a night especially during worm nights when
humid and mild weather conditions are optimal for earthworms to ascend to the surface (Baubet et
al 2003 Kruuk and Parish 1981) In endemic area of bTB where fecal M bovis shedding may occur in
infected animals it would be interesting to evaluate M bovis carriage in earthworm gut in the vicinity
of infected animal dung Further studies are needed to assess the role of earthworms harbouring
M bovis gut carriage in the contamination of bTB sensible earthworm feeders
One field study reports the isolation of non-tuberculous mycobacteria such as Mycobacterium
avium complex members (M avium ssp paratuberculosis and M avium ssp hominisuis) as well as M
abcessus M scrofulaceum and M gastri) from 9 out of 109 earthworm feces in sites where cattle
goats and swine were severely infected with the Johnersquos disease or mycobacteriosis (Fischer et al
2003) However this study failed to isolate M avium ssp avium from earthworms feces in aviaries
infected with avian tuberculosis
Cattle feces were spiked with M bovis at a final concentration of 125 times 107 CFU per 50 g of
feces ie 25 times 105 CFU per g (estimated by culture-based enumeration of the inoculum) M bovis
concentration in spiked cattle feces was also measured by the qPCR-based method We observed
numbers of M bovis 52-fold higher when using the molecular detection method than with the culture-
based method This discrepancy is probably due first to the strong aggregation of mycobacteria cells in
the inoculum despite physical disruption of cell aggregates and second to the presence of dead or
viable but non-cultivable cells of M bovis non detected by culture-based method The amount of
M bovis spiked in cattle feces is equivalent or lower than the number of M bovis cells recorded per g
of urine or feces shed by infected badgers (Gallagher and Clifton-Hadley 2000 Sweeney et al 2007)
leading to the presence of millions of bacteria in a latrine (burrowed hole in the soil where badgers
defecate and urinate) But it is probably larger than M bovis cells deposited in infected cattle feces
(Neill et al 1988) For practical reasons we used cattle feces in our study but it could be interesting to
compare if dung from other animals than cattle would be as attractive for earthworms
Chapitre 5
170
Our study was based on a qPCR assay method detecting M bovis DNA in soil castings and feces
samples This molecular tool does not give any information on the viability of the M bovis cells egested
by the earthworms However as mycobacteria harbour a resistant waxy cell envelope (Brennan and
Nikaido 1995) and as Mycobacterium avium ssp paratuberculosis and other environmental
mycobacteria have been showed to largely survive through the gut passage (Fischer et al 2003) these
data strongly suggest that M bovis could resist through the passage of earthworm guts and remain
alive in castings Further research is needed to clarify if tuberculous mycobacteria such as M bovis are
digested or survive during transit through the earthworm gut and if their virulence is conserved
Evidencing M bovis presence with culture-based method in environmental matrices such as soil
castings or animal feces is still a challenge As M bovis is a slow growing mycobacteria environmental
samples have to be strongly decontaminated prior to culturing to overcome contamination with fast
growing microorganisms leading to a decrease in the sensitivity of the method
Surface and subsurface casting shedding of M bovis is all the more relevant because this
pathogen bacteria may survive in soil and feces for long periods (Young et al 2005) Castings are then
within the reach of animals especially cattle that may be infected by inhaling soil aerosols or by
ingesting contaminated casting soil during their exploratory foraging or feeding activities While
Mycobacterium bovis infective dose by the respiratory route is very low since only ten viable bacilli are
able to cause respiratory bovine tuberculosis (Dean et al 2005) 5000 bacilli would be required to
initiate infection through the oral route in cattle (Palmer et al 2004) As cattle usually avoid grazing
close to the feces of other cattle or wild animals (Hutchings and Harris 1997) further dissemination of
M bovis by earthworms away from infected feces might induce contamination risk for cattle grazing in
this area
Further work is still needed to study the occurrence of M bovis in earthworms under natural
field conditions in bTB-contaminated area Cultivation of environmental M bovis is also required to
characterize properly these environmental strains Finally virulence of M bovis contained in
earthworm castings remains to be deciphered
Funding information
The work was supported by the Regional Council of Burgundy (Elodie Barbier PhD fellowship) the Cocircte
drsquoOr General Council Interbev and the Cocircte drsquoOr GDS (Groupement de Deacutefense Sanitaire Livestock
health protection group) The funders had no role in study design data collection and interpretation
or the decision to submit the work for publication
Conflict of interest
The authors have no conflict of interest to declare
Chapitre 5
171
Acknowledgements
The authors would like to thank AE Taconet F Bartheacuteleacutemy and T Grosjean from the Cocircte drsquoOr Regional
Veterinary Service (DDPP21) F Chevalier and M Beacuteral from the CIREV (Cellule Inter-Reacutegionale
drsquoEpideacutemiologie Veacuteteacuterinaire)
References
Baubet E Ropert-Coudert Y Brandt S 2003 Seasonal and annual variations in earthworm consumption by wild boar (Sus scrofa scrofa L) Wildl Res 30 179 doi101071WR00113
Biet F Boschiroli ML Thorel MF Guilloteau LA 2005 Zoonotic aspects of Mycobacterium bovis and Mycobacterium avium-intracellulare complex (MAC) Vet Res doi101051vetres
Blouin M Hodson ME Delgado E a Baker G Brussaard L Butt KR Dai J Dendooven L Peres G Tondoh JE Cluzeau D Brun JJ 2013 A review of earthworm impact on soil function and ecosystem services Eur J Soil Sci 64 161ndash182 doi101111ejss12025
Brennan PJ Nikaido H 1995 The envelope of mycobacteria Annu Rev Biochem 64 29ndash63 doi101146annurevbiochem64129
Byzov BA Khomyakov N V Kharin SA Kurakov A V 2007 Fate of soil bacteria and fungi in the gut of earthworms Eur J Soil Biol 43 149ndash156 doi101016jejsobi200708012
Cleary GP Corner LAL OrsquoKeeffe J Marples NM 2011 Diet of the European badger (Meles meles) in the Republic of Ireland A comparison of results from an analysis of stomach contents and rectal faeces Mamm Biol 76 470ndash475 doi101016jmambio201010012
Corner LAL OrsquoMeara D Costello E Lesellier S Gormley E 2012 The distribution of Mycobacterium bovis infection in naturally infected badgers Vet J 194 166ndash72 doi101016jtvjl201203013
Curry JP Schmidt O 2007 The feeding ecology of earthworms - A review Pedobiologia (Jena) 50 463ndash477 doi101016jpedobi200609001
Dean GS Rhodes SG Coad M Adam O Cockle PJ Clifford DJ Hewinson RG Vordermeier HM 2005 Minimum infective dose of Mycobacterium bovis in cattle Infect Immun 73 6467ndash6471 doi101128IAI73106467
Edwards CA Bohlen PJ 1996 Biology and ecology of earthworms Chapman and Hall London
Egert M Marhan S Wagner B Scheu S Friedrich MW 2004 Molecular profiling of 16S rRNA genes reveals diet-related differences of microbial communities in soil gut and casts of Lumbricus terrestris L (Oligochaeta Lumbricidae) FEMS Microbiol Ecol 48 187ndash197 doi101016jfemsec200401007
Fischer OA Matlova L Bartl J Dvorska L Svastova P Du Maine R Melicharek I Bartos M Pavlik I 2003 Earthworms (Oligochaeta Lumbricidae) and mycobacteria Vet Microbiol 91 325ndash338 doi101016S0378-1135(02)00302-4
Chapitre 5
172
Gallagher J Clifton-Hadley RS 2000 Tuberculosis in badgers a review of the disease and its significance for other animals Res Vet Sci 69 203ndash17 doi101053rvsc20000422
Genov I 1965 The effects of certain physical and chemical agents on Mycobacterium tuberculosis Veteriaacuternı Med Nauk Sofia 2 97ndash107
Goacutemez-Brandoacuten M Aira M Lores M Domiacutenguez J 2011 Epigeic Earthworms exert a bottleneck effect on microbial communities through gut associated processes PLoS One 6 doi101371journalpone0024786
Granval P Muys B 1995 Predation on earthworms by terrestrial vertebrates in XII Congress ldquoThe Game and the Manrdquo Sofia Bulgaria p 480
Hutchings MR Harris S 1997 Effects of farm management practices on cattle grazing behaviour and the potential for transmission of bovine tuberculosis from badgers to cattle Vet J 153 149ndash62
Kruuk H Parish T 1981 Feeding specialization of the European badger Meles meles in Scotland UK J Anim Ecol 50 773ndash788 doi1023074136
Lavelle P Spain A 2001 Soil ecology Kluwer Academic Publishers Dordrecht
Lee KE 1985 Earthworms their ecology and relationship with soils and land use Academic P ed
Lowe CN Butt KR 2005 Culture techniques for soil dwelling earthworms A review Pedobiologia (Jena) 49 401ndash413 doi101016jpedobi200504005
Mather JG Christensen O 1988 Surface movements of earthworms in agricultural land Pedobiologia (Jena) 32 399ndash405
Montecchio L Scattolin L Squartini A Butt K 2015 Potential spread of forest soil-borne fungi through earthworm consumption and casting iForest 8 295ndash301 doi103832ifor1217-008
Morris RS Pfeiffer DU Jackson R 1994 The epidemiology of Mycobacterium bovis infections Vet Microbiol 40 153ndash77
Muldowney J Curry J Okeeffe J Schmidt O 2003 Relationships between earthworm populations grassland management and badger densities in County Kilkenny Ireland Pedobiologia (Jena) 47 913ndash919 doi101016S0031-4056(04)70289-9
Neill SD Hanna J OrsquoBrien J McCracken R 1988 Excretion of Mycobacterium bovis by experimentally infected cattle Vet Rec 123 340ndash343 doi101136vr12313340
Palmer M V 2013 Mycobacterium bovis Characteristics of wildlife reservoir hosts Transbound Emerg Dis 60 1ndash13 doi101111tbed12115
Palmer M V Waters WR Whipple DL 2004 Investigation of the transmission of Mycobacterium bovis from deer to cattle through indirect contact Am J Vet Res 65 1483ndash1489 doi102460ajvr2004651483
Phillips CJC Foster CRW Morris PA Teverson R 2003 The transmission of Mycobacterium bovis infection to cattle Res Vet Sci 74 1ndash15
Chapitre 5
173
Prysor Williams A Roberts P Avery LM Killham K Jones DL 2006 Earthworms as vectors of Escherichia coli O157H7 in soil and vermicomposts FEMS Microbiol Ecol 58 54ndash64 doi101111j1574-6941200600142x
Sweeney FP Courtenay O Hibberd V Hewinson RG Reilly L a Gaze WH Wellington EMH 2007 Environmental monitoring of Mycobacterium bovis in badger feces and badger sett soil by real-time PCR as confirmed by immunofluorescence immunocapture and cultivation Appl Environ Microbiol 73 7471ndash3 doi101128AEM00978-07
Taylor AR Taylor AFS 2014 Assessing daily egestion rates in earthworms using fungal spores as a natural soil marker to estimate gut transit time Biol Fertil Soils 50 179ndash183 doi101007s00374-013-0823-5
Thorpe IS Prosser JI Glover L a Killham K 1996 The role of the earthworm Lumbricus terrestris in the transport of bacterial inocula through soil Biol Fertil Soils 23 132ndash139
Vicente J Houmlfle U Garrido JM Fernaacutedez-De-Mera IG Acevedo P Juste R Barral M Gortazar C 2007 Risk factors associated with the prevalence of tuberculosis-like lesions in fenced wild boar and red deer in south central Spain Vet Res 38 451ndash464 doi101051vetres2007002
Young JS Gormley E Wellington EMH 2005 Molecular detection of Mycobacterium bovis and Mycobacterium bovis BCG (Pasteur) in soil Appl Environ Microbiol 71 1946ndash1952 doi101128AEM7141946
Chapitre 5
174
C COMPLEMENT PRELEVEMENT ET ANALYSE DE LOMBRICS DANS UNE PATURE SUSPECTE
Afin drsquoeacutevaluer la contamination naturelle eacuteventuelle des lombrics par M bovis nous avons
analyseacute 90 lombrics preacuteleveacutes en feacutevrier 2014 agrave proximiteacute drsquoune source permanente drsquoune des pacirctures
suspectes de lrsquoAuxois ougrave des bovins infecteacutes ont pacirctureacute lrsquoeacuteteacute preacuteceacutedent (Site C de lrsquoeacutetude
environnementale Chapitre 2) Les lombrics ont eacuteteacute preacuteleveacutes le mecircme jour que les trois eacutechantillons
positifs de ce site (eau seacutediments neacutematodes) Sur sept points de la pacircture choisis au hasard une
pelleteacutee drsquoenviron 20 kg de sol a eacuteteacute retourneacutee Les vers ont eacuteteacute seacutepareacutes du sol rinceacutes avec de lrsquoeau et
transfeacutereacutes individuellement dans des pots en plastique de 60 mL Les pots ont eacuteteacute stockeacutes 48 h agrave 20degC
dans lrsquoobscuriteacute afin de laisser les vers se vider Pour chaque point de la pacircture preacuteleveacute les fegraveces des
vers ont eacuteteacute pooleacutees et homogeacuteneacuteiseacutees puis 2 g ont eacuteteacute preacuteleveacutes et congeleacutes jusqursquoagrave la reacutealisation de
lrsquoanalyse Apregraves extraction de lrsquoADN chaque eacutechantillon a eacuteteacute analyseacute en qPCR avec le systegraveme de
deacutetection RD4 Les protocoles utiliseacutes sont deacutecrits dans le Mateacuteriel et Meacutethodes de lrsquoarticle preacuteceacutedent
Lrsquoidentification pheacutenotypique des 90 vers collecteacutes dans la pacircture a permis de mettre en
eacutevidence cinq genres de lombric diffeacuterents Lumbricus terrestris Lumbricus castaneus Nicodrilus sp
Octolosium cyaneum Allolobophora sp Aporrectodea caliginosa
Aucun des sept pools de fegraveces analyseacutes nrsquoa donneacute de signal positif en qPCR avec le systegraveme RD4
Chapitre 5
175
D BILAN
Les lombrics sont de gros consommateurs de matiegravere organique naturellement chargeacutee drsquoune
flore abondante et varieacutee et potentiellement drsquoune flore bacteacuterienne pathogegravene Toutefois il existe
tregraves peu de donneacutees sur la disseacutemination des bacteacuteries pathogegravenes pour lrsquohomme et les animaux par
les lombrics dans le sol ainsi que sur les risques lieacutes au portage intestinal de bacteacuteries pathogegravenes par
les lombrics pour les animaux qui les consomment
Lrsquoeacutetude deacutecrite dans lrsquoarticle preacuteceacutedent montre qursquoapregraves avoir ingeacutereacute des matiegraveres feacutecales
fraiches inoculeacutees les lombrics ont la capaciteacute de disseacuteminer rapidement M bovis dans le sol
environnant par le biais des turricules qursquoils eacutemettent agrave la surface du sol et en profondeur dans les
tunnels qursquoils creusent Lrsquoenfouissement de la bacteacuterie dans le sol permet probablement drsquoameacuteliorer la
persistance de la bacteacuterie en la proteacutegeant des UV et de la deacuteshydratation Drsquoautre part le transfert
de la bacteacuterie agrave quelques cm des deacutejections animales contamineacutees par le biais des turricules de surface
la rend plus accessible aux animaux (qui en geacuteneacuteral eacutevitent les fegraveces des autres animaux) Ainsi en
fonction des comportements exploratoires ou alimentaires des diffeacuterentes espegraveces freacutequentant les
pacirctures il existe un risque potentiel de contamination par du sol ou des turricules contamineacutes
(inhalation de bioaeacuterosols ou ingestion)
Nous avons eacutegalement montreacute que le tube digestif des lombrics ne constituait pas un reacuteservoir
peacuterenne de M bovis la bacteacuterie nrsquoeacutetait deacutetecteacutee que transitoirement (4 jours) apregraves avoir remis les
lombrics dans un sol sain Le portage digestif pendant les 4 premiers jours constitue toutefois un risque
pour les animaux qui consomment les lombrics en grande quantiteacute notamment les blaireaux et les
sangliers
Cette eacutetude soulegraveve les mecircmes inconnues que les eacutetudes preacuteceacutedentes principalement sur la
viabiliteacute lrsquoeacutetat physiologique et la conservation de la virulence de M bovis apregraves un transfert dans le
sol ou suite au passage de la bacteacuterie dans le tube digestif des lombrics Ces facteurs sont
indispensables pour permettre lrsquoinfection drsquoanimaux via des matrices environnementales contamineacutees
Les moyens drsquoeacutevaluation de ces diffeacuterents facteurs seront discuteacutes dans la partie Perspectives du
manuscrit
Chapitre 5
176
Discussion geacuteneacuterale
Discussion geacuteneacuterale
179
Depuis 2004 face agrave la recrudescence de la tuberculose bovine en Cocircte drsquoOr la mise en place
drsquoun programme de lutte sanitaire reacuteguliegraverement ameacutelioreacute contre la maladie a permis de controcircler et
drsquoameacuteliorer la situation dans lrsquoeacutelevage bovin depuis 2010 Dans la zone endeacutemique du deacutepartement la
circulation de Mycobacterium bovis dans un systegraveme multi-hocircte speacutecifiques impliquant les bovins et
des espegraveces sauvages complique fortement la lutte contre la maladie notamment par la difficulteacute agrave
geacuterer lrsquoinfection dans la faune sauvage malgreacute les mesures de controcircle entreprises jusqursquoagrave maintenant
La transmission indirecte de la maladie entre animaux sauvages et bovins par le biais de substrats
environnementaux contamineacutes est fortement suspecteacutee dans les zones ougrave bovins et faune sauvage
infecteacutes partagent les mecircmes territoires et ressources La transmission environnementale a deacutejagrave eacuteteacute
deacutemontreacutee en conditions expeacuterimentales (Little et al 1982 Palmer et al 2004) mais elle est tregraves
difficile agrave deacutemontrer en conditions naturelles et passe notamment par lrsquoeacutevaluation du niveau
drsquoexcreacutetion par les animaux infecteacutes du rocircle de lrsquoenvironnement comme reacuteservoir de la bacteacuterie de sa
survie dans un eacutetat viable et du maintien de sa virulence
Dans ce contexte les objectifs de ce travail eacutetaient drsquoune part drsquoeacutevaluer la contamination
environnementale en Cocircte drsquoOr sur des agroeacutecosystegravemes freacutequenteacutes par des bovins etou des animaux
sauvages potentiellement infecteacutes dans le but de mettre en eacutevidence des reacuteservoirs
environnementaux eacuteventuels Drsquoautre part nous avons chercheacute agrave preacuteciser les facteurs impliqueacutes dans
la persistance de M bovis dans le sol et sa disseacutemination par la micro- et meacutesofaune du sol
1 Des meacutethodes deacuteveloppeacutees (ou adapteacutees) puis valideacutees dans des matrices environnementales
Notre objectif initial pour lrsquoanalyse des eacutechantillons environnementaux eacutetait drsquoutiliser en
parallegravele la PCR quantitative apregraves extractionpurification de lrsquoADN pour la deacutetection et la quantification
de M bovis et une meacutethode baseacutee sur la culture afin drsquoisoler les souches environnementales pour les
caracteacuteriser Cependant les protocoles de preacutetraitement et de culture tireacutes de la litteacuterature
(immunocapture et protocoles de deacutecontamination) que nous avons utiliseacutes en premiegravere intention sur
les eacutechantillons environnementaux nrsquoont pas donneacute les reacutesultats escompteacutes du fait de la croissance tregraves
lente de M bovis et de la preacutesence dans les eacutechantillons de tregraves nombreux microorganismes
interfeacuterents agrave croissance rapide qui ont gecircneacute la culture (envahissement rapide des boicirctes en une
semaine) La culture de M bovis agrave partir drsquoeacutechantillons environnementaux reste agrave lrsquoheure actuelle un
challenge (Fine OrsquoBrien et al 2011 Fine Bolin et al 2011 Young et al 2005 King et al 2015)
notamment agrave cause de lrsquoabsence de milieux suffisamment seacutelectifs et de bouillons drsquoenrichissement
Discussion geacuteneacuterale
180
adapteacutes comme il en existe notamment pour lrsquoisolement de bacteacuteries pathogegravenes telluriques telles
que Listeria monocytogenes ou Pseudomonas aeruginosa En lrsquoabsence de meacutethodes disponibles nous
avons donc mis au point deux protocoles tregraves innovants agrave la fin de nos travaux un protocole
drsquoextractiondeacutecontamination des mycobacteacuteries dans le sol et une meacutethode de deacutetection des
mycobacteacuteries en culture liquide Ces deux meacutethodes sont en cours drsquoeacutevaluation par la SATT (Socieacuteteacute
drsquoAcceacuteleacuteration de Transfert de Technologies) en vue drsquoune valorisation sous forme de savoir-faire ou de
brevet Des eacutetudes compleacutementaires seront neacutecessaires pour deacutevelopper ces outils innovants
La recherche de reacuteservoirs environnementaux potentiels de M bovis en Cocircte drsquoOr a donc eacuteteacute
baseacutee sur lrsquoutilisation de meacutethodes moleacuteculaires adapteacutees aux matrices environnementales
Contrairement aux meacutethodes culturales la qPCR permet une deacutetection et une quantification rapide
des ADN cibles sans ecirctre gecircneacutee par la preacutesence de contaminants Face au grand nombre de
mycobacteacuteries potentiellement preacutesentes dans les sols de Cocircte drsquoOr dont M microti (Michelet et al
2015) nous avons choisi drsquoutiliser trois systegravemes ciblant trois seacutequences diffeacuterentes dans le geacutenome
des espegraveces du MTBC ou de M bovis que nous avons deacuteveloppeacutes et valideacutes pour leur speacutecificiteacute et leur
sensibiliteacute Lrsquoutilisation conjointe de ces trois systegravemes nous a permis drsquoaugmenter la speacutecificiteacute de la
deacutetection Toutefois le faible niveau de deacutetection observeacute dans certains eacutechantillons ne nous a pas
permis de quantifier le MTBC ou M bovis dans tous les eacutechantillons
2 Contamination environnementale les zones humides les eaux de ruissellement et les sols de terriers sont potentiellement agrave risque
Les systegravemes moleacuteculaires deacuteveloppeacutes ont eacuteteacute utiliseacutes pour lrsquoanalyse de plus de 400
eacutechantillons environnementaux varieacutes preacuteleveacutes dans les agroeacutecosystegravemes de la zone endeacutemique de
Cocircte drsquoOr et de Charente La contamination environnementale a eacuteteacute deacutetecteacutee de faccedilon reacutepeacuteteacutee dans
deux types drsquoeacutecosystegravemes bien particuliers les zones humides preacutesentes sur les pacirctures (sources et
laquo mouilles raquo) les terriers et latrines de blaireaux
Concernant les zones humides nous avons mis en eacutevidence une contamination dans lrsquoeau
circulante et les seacutediments de deux sources naturelles et permanentes situeacutees sur des pacirctures utiliseacutees
par deux cheptels diffeacuterents dans deux communes diffeacuterentes Cette faible contamination de lrsquoeau
etou des seacutediments retrouveacutee agrave lrsquoendroit de la reacutesurgence ainsi que sur diffeacuterents points en aval est
une donneacutee nouvelle puisqursquoagrave ce jour M bovis nrsquoavait eacuteteacute isoleacutee que dans lrsquoeau stagnante et ses
seacutediments (Santos et al 2015) Lors de nos campagnes de preacutelegravevements nous avons constateacute que ces
reacutesurgences parfois capteacutees dans des abreuvoirs eacutetaient parfois le seul moyen drsquoabreuvement des
bovins dans les pacirctures Ces zones humides semblent tregraves attractives pour les bovins au vu des indices
de preacutesence constateacutes (matiegraveres feacutecales et nombreuses empreintes) La persistance de M bovis eacutetant
Discussion geacuteneacuterale
181
favoriseacutee dans les matrices humides riches en matiegravere organique et dans lrsquoeau (Young et al 2005 Fine
Bolin et al 2011 Duffield amp Young 1985 Wint et al 2002) ces zones constituent un milieu favorable
agrave la survie de la bacteacuterie Dans notre eacutetude la mise en eacutevidence de la contamination des zones humides
dans des pacirctures suspectes corrobore le reacutesultat de lrsquoeacutetude PatRisk dans laquelle une association
significative a eacuteteacute eacutetablie entre la preacutesence de mouilles et le statut laquo agrave risque drsquoune pacircture raquo (Bouchez-
Zacria 2015)
Lrsquoorigine de la contamination de ces sources et laquo mouilles raquo est inconnue toutefois elles sont
accessibles aux bovins et animaux sauvages infecteacutes qui peuvent potentiellement excreacuteter la bacteacuterie
lors de lrsquoabreuvement ou du fouissement Vu le contexte geacuteomorphologique il est eacutegalement
envisageable que les infiltrations drsquoeau dans les plateaux calcaires coiffant les valleacutees entrainent avec
elles des matrices environnementales fortement contamineacutees (latrines sols) qui ressortent au niveau
des pacirctures avec toutefois un effet de dilution tregraves important
Des mesures de bioseacutecuriteacute visant agrave limiter les risques lieacutes aux zones humides sont deacutejagrave
preacuteconiseacutees par la DDPP 21 et le GDS 21 agrave lrsquoeacutechelle de la pacircture Lrsquoameacutenagement drsquoabreuvoirs peu
accessibles aux animaux sauvages et la condamnation par des clocirctures de lrsquoaccegraves des bovins aux
laquo mouilles raquo propices agrave la survie de la bacteacuterie sont fortement conseilleacutes aux eacuteleveurs des zones agrave
risque pour limiter le risque de transmission indirecte de la maladie Le drainage des pacirctures est une
solution envisageacutee pour asseacutecher les zones humides et canaliser les sources dans les pacirctures agrave risque
mais il est strictement reacuteglementeacute et soumis agrave autorisation avec enquecircte publique sous le controcircle de
la Police de lrsquoeau (Direction Deacutepartementale des Territoires) La correacutelation de nos reacutesultats sur la
contamination des zones humides avec le bilan des foyers bovins et des cas sauvages ainsi que les
reacutesultats de lrsquoeacutetude PatRisk ont initieacute une reacuteflexion entre la DDPP21 la CIREV le GDS et le SIRTAVA
(Syndicat Intercommunal pour la Reacutealisation des Travaux drsquoAmeacutenagement de la Valleacutee de lrsquoArmanccedilon)
pour eacutevaluer la faisabiliteacute de travaux de drainage dans les pacirctures agrave risque dans le respect de la
regraveglementation Un problegraveme se pose toutefois pour les pacirctures preacutesentant de tregraves nombreuses
reacutesurgences temporaires sur lesquelles le drainage nrsquoest pas envisageable Lrsquoutilisation de ces pacirctures
pour drsquoautres productions animales (ovins chevaux) ou veacutegeacutetales (production de foin la mise en
culture nrsquoest pas toujours possible agrave cause des pentes) est une solution
Notre eacutetude a eacutegalement mis en eacutevidence la contamination des sols de terriers de blaireaux
par M bovis Le geacutenotype retrouveacute dans les sols de terriers est identique agrave celui retrouveacute chez les
blaireaux infecteacutes pieacutegeacutes agrave proximiteacute confirmant que cette espegravece a la capaciteacute de contaminer son
milieu de vie (Young et al 2005 Courtenay et al 2006 Sweeney et al 2006 Sweeney et al 2007) Dans
cette espegravece des animaux excreacuteteurs et super excreacuteteurs ont eacuteteacute mis en eacutevidence en Cocircte drsquoOr (Payne
2014) expliquant les quantiteacutes importantes de bacilles retrouveacutees dans les sols de terrier (Young et al
Discussion geacuteneacuterale
182
2005 Courtenay et al 2006) et les latrines dans notre eacutetude La deacutetection drsquoun signal moleacuteculaire
pendant 12 mois dans plusieurs entreacutees drsquoun mecircme terrier suggegravere fortement que les sols de terriers
peuvent constituer un reacuteservoir environnemental pour M bovis La survie des mycobacteacuteries est
favoriseacutee dans ces structures complexes et profondes ougrave regravegne un microclimat propice (Moore amp Roper
2003) tant au niveau de la stabiliteacute de la tempeacuterature de lrsquohumiditeacute optimale que de la protection
contre les UV Ce reacuteservoir potentiel repreacutesente un risque pour le maintien de la maladie dans le groupe
social de blaireaux qui partage le terrier notamment pour les blairotins qui y naissent pendant lrsquohiver
mais aussi entre les groupes sociaux qui peuvent reacuteguliegraverement changer de terriers Lorsqursquoils sont
localiseacutes dans les pacirctures et sont accessibles aux bovins le sol des terriers contamineacutes repreacutesenterait
un risque potentiel pour les bovins qui les explorent notamment un risque drsquoinhalation de bioaeacuterosols
contamineacutes geacuteneacutereacutes agrave partir de sol Enfin les sols et latrines contamineacutes souvent en limite des plateaux
calcaires seraient susceptibles drsquoecirctre agrave lrsquoorigine de la contamination des pacirctures par le biais des
infiltrations drsquoeau des pheacutenomegravenes de reacutesurgence drsquoeau et de ruissellement
Les mesures de bioseacutecuriteacute preacuteconisant la clocircture des terriers situeacutes dans les pacirctures semblent
essentielles agrave respecter par les eacuteleveurs si lrsquoon considegravere la longue persistance de la bacteacuterie dans ce
biotope particulier Le repeuplement des terriers contamineacutes ne fait lrsquoobjet drsquoaucun controcircle agrave lrsquoheure
actuelle Sous le controcircle du CSRPN (Conseil Scientifique Reacutegional du Patrimoine Naturel) une eacutetude
est actuellement en cours en Cocircte drsquoOr pour lrsquoeacutetude de faisabiliteacute drsquoun protocole de deacutepeuplement des
terriers (pieacutegeage de tous les individus du groupe) associeacute agrave un traitement des tunnels par des reacutepulsifs
permanents pour eacuteviter la recolonisation immeacutediate
La preacutevalence de M bovis dans lrsquoenvironnement des animaux infecteacutes en Cocircte drsquoOr est
probablement sous-estimeacutee dans notre eacutetude agrave cause de limites temporelles spatiales et
meacutethodologiques auxquelles nous avons fait face En premier lieu il existe un deacutelai incompressible de
plusieurs semaines agrave plusieurs mois entre les trois eacutevegravenements suivants i) la contamination
environnementale par un animal excreacuteteur (bovin ou faune sauvage) ii) le diagnostic de lrsquoinfection chez
cet animal (apregraves prophylaxie et abattage pour les bovins en hiver et pieacutegeage pour les blaireaux au
printemps et en eacuteteacute) et iii) le preacutelegravevement et lrsquoanalyse drsquoeacutechantillons environnementaux dans les zones
de vie de ces animaux Cette limite temporelle est eacutegalement eacutevoqueacutee par drsquoautres auteurs (Fine
OrsquoBrien et al 2011) Il serait ideacuteal drsquoeffectuer les preacutelegravevements environnementaux dans un deacutelai le plus
court possible apregraves lrsquoexcreacutetion du bacille par un animal infecteacute toutefois ce deacutelai deacutepend du rythme
des prophylaxies et de la freacutequence du pieacutegeage des blaireaux De plus sur le terrain malgreacute notre
strateacutegie drsquoeacutechantillonnage ciblant les zones partageacutees preacutefeacuterentiellement par la faune domestique et
sauvage les surfaces agrave preacutelever restent importantes Lrsquoanalyse de la totaliteacute des surfaces cibleacutees est
humainement techniquement et eacuteconomiquement irreacutealisable le nombre de preacutelegravevements a donc ducirc
Discussion geacuteneacuterale
183
ecirctre optimiseacute pour aboutir agrave une analyse pertinente Les reacutesultats obtenus ne donnent de ce fait qursquoune
image partielle de la contamination reacuteelle des zones preacuteleveacutees Enfin la charge bacteacuterienne de certains
preacutelegravevements environnementaux nrsquoa pu ecirctre quantifieacutee signifiant que les quantiteacutes de cibles preacutesentes
eacutetaient proches des limites de deacutetection de nos systegravemes moleacuteculaires Pour ces diffeacuterentes raisons
nous ne pouvons donc pas exclure une contamination environnementale plus large avec des quantiteacutes
tregraves faibles de M bovis
3 Survie et circulation de M bovis dans lrsquoenvironnement
La survie des bacteacuteries dans le sol deacutepend des facteurs biotiques et abiotiques qui caracteacuterisent
ces sols Les diffeacuterentes eacutetudes sur la survie de M bovis ont essentiellement consideacutereacute les facteurs
abiotiques climatiques telles que la tempeacuterature lrsquohumiditeacute et lrsquoexposition aux UV dans un seul type
de sol (Young et al 2005 Jackson et al 1995 Duffield amp Young 1985) Nous avons montreacute que M bovis
SB0120 eacutetait cultivable au moins cinq mois agrave une tempeacuterature basse (4degC) avec une mortaliteacute
bacteacuterienne minime et jusqursquoagrave trois mois agrave 22degC Ces dureacutees de survie sont supeacuterieures agrave celles
rapporteacutees dans de preacuteceacutedentes eacutetudes (Fine Bolin et al 2011 Jackson et al 1995 Duffield amp Young
1985) Les sols retrouveacutes dans lrsquoAuxois ont agrave la fois des caracteacuteristiques diffeacuterentes des sols
habituellement deacutecrits comme favorables agrave la survie des mycobacteacuteries (sols sableux agrave pH acide) et
des caracteacuteristiques communes leur richesse en matiegravere organique et en fer Bien que nous nrsquoayons
pas pu mettre en eacutevidence un effet de la nature du sol dans notre eacutetude sur la survie de M bovis il
nous parait probable que les sols argilo-limoneux peacuterennisent la maladie dans les valleacutees de lrsquoAuxois
Notre expeacuterience limiteacutee par des contraintes techniques lieacutees agrave lrsquoutilisation du laboratoire de
confinement L3 du CHU de Dijon aurait neacutecessiteacute lrsquoutilisation drsquoun plus grand nombre de sols de points
de preacutelegravevements et de reacutepeacutetitions biologiques La concentration des foyers bovins (notamment des
recontaminations de cheptels) dans une zone limiteacutee du deacutepartement en Auxois interroge eacutegalement
sur le rocircle du contexte paysager et geacuteomorphologique commun des pacirctures suspectes dans la survie
de M bovis
Ces dureacutees de survie de M bovis dans les sols doivent ecirctre consideacutereacutees avec preacutecaution car
obtenues dans des conditions expeacuterimentales controcircleacutees qui ne miment que partiellement les
conditions environnementales reacuteelles (sols deacutebarrasseacutees de leur flore endogegravene agrave une tempeacuterature
drsquoincubation constante une humiditeacute optimale et une protection contre les UV) Il est probable qursquoen
conditions naturelles la survie de M bovis soit moins longue agrave cause des variations nyctheacutemeacuterales de
tempeacuteratures des variations des preacutecipitations des UV ainsi que de lrsquoeffet barriegravere potentiel de la flore
endogegravene des matrices environnementales La correacutelation de ces donneacutees avec les courbes de
Discussion geacuteneacuterale
184
tempeacuteratures permettent de montrer que la peacuteriode de novembre agrave avril ougrave les tempeacuteratures sont les
plus basses est compatible avec une survie optimale de M bovis dans les sols
Les reacutesultats obtenus dans notre eacutetude de survie ont souleveacute la question de la persistance de
M bovis dans le sol dans un eacutetat viable mais non cultivable (VNC) puisque la bacteacuterie nrsquoest plus
cultivable mais toujours deacutetecteacutee par qPCR Lrsquoeacutetat VNC de M bovis a deacutejagrave eacuteteacute suggeacutereacute dans des eacutetudes
de survie dans du sol (Adams et al 2013 Young et al 2005) Les mycobacteacuteries possegravedent une capaciteacute
particuliegravere agrave srsquoadapter et agrave survivre dans des conditions deacutefavorables en lrsquoabsence de nutriments
drsquooxygegravene sous diffeacuterents stress exogegravenes notamment dans les phagosomes de lrsquohocircte pour les
mycobacteacuteries pathogegravenes (Gengenbacher amp Kaufmann 2012 Boon amp Dick 2012 Voskuil et al 2003)
Les mycobacteacuteries disposent de diffeacuterents systegravemes agrave deux composants (permettant une transduction
de signal) leur permettant la perception sensorielle des conditions environnementales et en particulier
de la composition du milieu extracellulaire (Cook et al 2009 Cole et al 1998) Dans les substrats
environnementaux la bacteacuterie conserve probablement tout son potentiel de perception du milieu qui
lrsquoentoure et ses capaciteacutes drsquoadaptation mais elle ne peut pas interagir avec lui comme avec les cellules
de lrsquohocircte A lrsquoheure actuelle les meacutecanismes drsquoadaptation mis en jeu par M bovis pour survivre lors de
son excreacutetion par lrsquohocircte dans lrsquoenvironnement sont inconnus
Nous avons montreacute pour la premiegravere fois que les lombrics veacuteritables ingeacutenieurs du sol ont la
capaciteacute de disseacuteminer la bacteacuterie dans leur environnement par le biais des tunnels qursquoils creusent et
des turricules qursquoils eacutemettent agrave et sous la surface du sol Lrsquoenfouissement de la bacteacuterie agrave quelques
centimegravetres sous le sol favorise probablement sa survie Les lombrics peuvent eacutegalement jouer un rocircle
de vecteur quand ils portent du sol contamineacute dans leur tube digestif et qursquoils sont ingeacutereacutes par des
blaireaux ou des sangliers Cette eacutetude soulegraveve la probleacutematique du rocircle de la faune du sol dans la
survie et la circulation de M bovis dans lrsquoenvironnement et le risque que des reacuteservoirs cacheacutes de la
bacteacuterie entretiennent agrave bas bruit une contamination de lrsquoenvironnement
Perspectives
Perspectives
187
Lrsquoeacutevaluation de la contamination environnementale reacutealiseacutee en Cocircte drsquoOr et dans une moindre
mesure en Charente (terriers de blaireaux uniquement) nrsquoest pas extrapolable aux autres reacutegions
franccedilaises toucheacutees par la tuberculose A ce jour aucune investigation environnementale aussi
complegravete nrsquoa eacuteteacute meneacutee dans des deacutepartements fortement infecteacutes comme la Dordogne et les
Pyreacuteneacutees Atlantiques ougrave bovins et faune sauvage cohabitent Il serait donc inteacuteressant drsquoeacutevaluer le
niveau de contamination environnementale des zones partageacutees par les bovins et la faune sauvage afin
de veacuterifier si les zones humides et les terriers sont eacutegalement agrave risque dans ces deacutepartements De plus
les reacutesultats obtenus dans la zone endeacutemique de Cocircte drsquoOr ont suggeacutereacute le rocircle possible du contexte
geacuteomorphologique hydrologique et peacutedologique de lrsquoAuxois dans la contamination environnementale
et la persistance de M bovis Pour veacuterifier cette hypothegravese il serait inteacuteressant de comparer le contexte
des pacirctures suspectes de lrsquoAuxois avec les pacirctures suspectes des deacutepartements les plus toucheacutes par la
tuberculose bovine La caracteacuterisation physico-chimique du sol de ces pacirctures permettrait peut-ecirctre de
mettre en relation drsquoeacuteventuelles proprieacuteteacutes communes entre les sols incrimineacutes et la persistance de la
bacteacuterie La mise en eacutevidence de facteurs de risque paysagers hydrographiques etou peacutedologiques
permettrait eacuteventuellement de mettre au point un outil de preacutevision des risques en fonction du
contexte rencontreacute (pacirctures agrave risque) drsquoadapter au mieux la gestion sanitaire et drsquoameacuteliorer la
bioseacutecuriteacute dans les eacutelevages Si le rocircle des zones humides (laquomouilles raquo) et des sources dans drsquoautres
reacutegions est confirmeacute il conviendra de limiter lrsquoaccegraves des bovins agrave ces zones pour limiter la
contamination En Cocircte drsquoOr des approches de ce type (drainage et clocircture des zones humides) pour
empecirccher lrsquoaccegraves des animaux sont en cours en collaboration avec les eacuteleveurs les agences de bassin
lrsquoONEMA ainsi que les gestionnaires des zones humides agrave lrsquoeacutechelle du bassin versant de la Brenne
La dureacutee de survie deacutemontreacutee de M bovis dans les sols remet partiellement en cause la dureacutee
du deacutelai de carence de 2 mois avant reacuteinstallation drsquoanimaux suite agrave la peacuteriode de vide sanitaire dans
les exploitations qui ont eacuteteacute foyer de tuberculose Nos observations devront ecirctre discuteacutees dans le
cadre de lrsquooptimisation de ces pratiques
Sur le plan expeacuterimental notre expeacuterience de survie sur deux sols eacutetant insuffisante une eacutetude
de la survie de M bovis sur un grand nombre de sols aux caracteacuteristiques physico-chimiques
contrasteacutees permettrait drsquoidentifier les facteurs physico-chimiques du sol associeacutes agrave une survie
optimale de la bacteacuterie comme cela a pu ecirctre fait au sein de notre eacutequipe pour Listeria monocytogenes
(Locatelli et al 2013) Lrsquoidentification des facteurs eacutedaphiques favorisant la persistance de M bovis
permettra de mettre en eacutevidence les sols agrave risque
La recherche de M bovis dans des matrices environnementales se heurte aux limites des
techniques utiliseacutees Concernant la qPCR bien qursquoelle reste une technique de choix des efforts sont
encore neacutecessaires pour abaisser les seuils de deacutetection dans les matrices complexes faiblement
Perspectives
188
chargeacutees en mycobacteacuteries Drsquoautres techniques PCR pourraient ecirctre envisageacutees notamment la PCR
digitale (ou PCR numeacuterique) qui permet de gagner en sensibiliteacute et de limiter lrsquoimpact des inhibiteurs
mais dont lrsquoutilisation est encore limiteacutee par le coucirct De plus la qPCR deacutetecte lrsquoADN bacteacuterien sans
donner aucune indication sur la viabiliteacute de la cellule Devant les difficulteacutes agrave isoler M bovis par la
culture agrave partir de matrices environnementales complexes agrave cause notamment de sa croissance tregraves
lente une approche transcriptomique permettrait de preacuteciser lrsquoeacutetat viable de M bovis Cette approche
pourrait eacutegalement apporter des connaissances encore totalement inconnues agrave lrsquoheure actuelle sur les
meacutecanismes de survie mis en jeu par la bacteacuterie une fois excreacuteteacutee par son hocircte dans lrsquoenvironnement
et mettre en eacutevidence les gegravenes impliqueacutes dans son adaptation aux conditions environnementales et
donc sa persistance Dans lrsquoobjectif drsquoeacuteclaircir ces meacutecanismes qui semblent fondamentaux pour la
compreacutehension de lrsquoadaptation et de la survie de M bovis dans lrsquoenvironnement lrsquoutilisation drsquoune
approche transcriptomique permettrait drsquoidentifier les modifications transcriptionnelles agrave lrsquoeacutechelle du
geacutenome complet par une approche de seacutequenccedilage des ARN (RNA seq) et de certains gegravenes par RT-PCR
lors de lrsquointroduction de M bovis dans le sol La comparaison de ces donneacutees avec les reacutesultats obtenus
in vitro et in vivo dans diffeacuterentes conditions de stress (Rodriguez et al 2008 Jang et al 2009)
permettraient de veacuterifier si les mecircmes meacutecanismes sont impliqueacutes Les gegravenes du reacutegulon de la
dormance preacutesents chez M bovis sont surexprimeacutes dans les macrophages in vitro et in vivo (Rodriguez
et al 2008) il serait particuliegraverement inteacuteressant drsquoeacutevaluer leur niveau drsquoexpression lors de
lrsquointroduction de la bacteacuterie dans le sol Comme il est deacutelicat drsquoextraire lrsquoARN de matrices complexes
comme le sol lrsquoutilisation drsquoextraits aqueux de sol pourrait faciliter lrsquoeacutetude (Piveteau et al 2011) Pour
aller plus loin il serait inteacuteressant de comparer les transcriptomes de M bovis lorsque les conditions
abiotiques (type de sol tempeacuterature humiditeacute etc) et biotiques (comparaison absencepreacutesence du
microbiote endogegravene du sol) varient pour cerner plus finement les meacutecanismes drsquoadaptation deacuteployeacutes
face aux diffeacuterents stress
Les meacutethodes de culture de M bovis agrave partir de matrices environnementales complexes
comme le sol sont encore agrave ameacuteliorer Lrsquoisolement de souches environnementales de M bovis
permettrait gracircce aux nouvelles techniques de seacutequenccedilage (NGS) de faire de la geacutenomique
comparative sur geacutenome complet avec les mecircmes souches isoleacutees chez les animaux et drsquoameacuteliorer les
connaissances sur les meacutecanismes geacuteneacutetiques mis en jeu dans lrsquoadaptation et la persistance de la
bacteacuterie dans lrsquoenvironnement
Concernant la faune la persistance et la circulation des mycobacteacuteries dans lrsquoeau et le sol
pourraient ecirctre favoriseacutees par la micro- et la meacutesofaune preacutesentes dans ces matrices Les protozoaires
et particuliegraverement les amibes ont fait lrsquoobjet drsquoeacutetudes montrant la survie dans leurs trophozoiumltes et
leurs cystes de nombreuses espegraveces dont M bovis (Taylor et al 2003 Mba Medie et al 2011) Dans
notre eacutetude un des eacutechantillons de neacutematodes preacuteleveacute dans les seacutediments drsquoune source a donneacute un
Perspectives
189
signal moleacuteculaire reacuteveacutelant un lien potentiel entre neacutematodes et persistance des mycobacteacuteries Pour
des raisons de temps et de compeacutetences nous nrsquoavons pas exploreacute minutieusement ces eacuteventuels
reacuteservoirs cacheacutes de M bovis Devant lrsquoabondance et la diversiteacute de ces animaux dans les matrices que
nous analysons des recherches compleacutementaires nous semblent neacutecessaires pour preacuteciser leur rocircle
potentiel dans la persistance de M bovis
Les diffeacuterents reacutesultats obtenus pendant ce travail et les implications qui en ressortent ont eacuteteacute
reacuteguliegraverement partageacutes avec les instances sanitaires et agricoles de Cocircte drsquoOr afin drsquoenrichir les
connaissances sur lrsquoeacutepideacutemiologie tregraves complexe de cette maladie et de participer agrave lrsquoameacutelioration des
mesures de bioseacutecuriteacute qui en deacutecoulent Drsquoautres deacutepartements toucheacutes par la tuberculose bovine
inteacuteresseacutes par nos travaux nous ont fait parvenir des demandes drsquoanalyse environnementale dans des
zones ougrave la transmission indirecte de la maladie est fortement suspecteacutee
Les deux protocoles innovants pour lrsquoextraction des mycobacteacuteries du sol et le suivi de leur
croissance en milieu liquide deacuteveloppeacutes dans ce travail de thegravese font lrsquoobjet drsquoune eacutetude de valorisation
(protection intellectuelle et eacutetude de marcheacute) en collaboration avec la SATT Grand Est
Enfin les diffeacuterentes ideacutees et perspectives deacuteveloppeacutees dans cette thegravese ont eacuteteacute reprises en
partie dans le cadre de lrsquoappel drsquooffre Horizon 2020 agrave travers le deacutepocirct drsquoun projet europeacuteen
multipartenaire (scientifiques et entreprises) qui est coordonneacute par lrsquoINRA de Tours Nouzilly et qui
implique de nombreux partenaires en Allemagne (Lionex PME speacutecialiseacutee dans les outils de deacutetection
seacuterologique speacutecifiques) en Hollande (WUR) en UK (APHA) en Irlande (UCD) en Italie (Universiteacute de
Naples) Ce projet intituleacute DEPICT vise entre autre agrave une meilleure compreacutehension des meacutecanismes
infectieux de la reacuteaction inflammatoire et immunologique des animaux mais eacutegalement agrave deacuteterminer
le rocircle de lrsquoenvironnement dans la survie des pathogegravenes comme M bovis et Map entre autres La
diversiteacute et la persistance de M bovis et Map seront eacutetudieacutees dans lrsquoenvironnement afin de deacuteterminer
les facteurs influenccedilant la survie de ces pathogegravenes en particulier dans les sols Au travers de ce
programme et drsquoautres collaborations agrave venir avec lrsquoINRA de Tours le travail deacutebuteacute au cours de cette
thegravese sur M bovis sera eacutelargi agrave Map mycobacteacuterie eacutegalement difficilement cultivable et revecirctant une
grande importance pour la santeacute animale et la santeacute humaine (elle serait impliqueacutee dans la maladie de
Crohn) Les nouveaux outils (culture et deacutetection) deacuteveloppeacutes au cours de cette thegravese seront testeacutes sur
Map pour permettre lrsquoisolement drsquoun plus grand nombre de souches environnementales
Une des questions en suspens reste la deacutemonstration de la contamination de bovins agrave partir
de M bovis preacutesente dans des matrices environnementales Des expeacuteriences de contamination
expeacuterimentale en animalerie P3 agrave partir de sol ou drsquoeau contamineacutes pourraient permettre cette
deacutemonstration Lrsquoutilisation du modegravele cobaye espegravece tregraves sensible agrave lrsquoinfection par M bovis pour
reacutealiser des expeacuteriences preacuteliminaires pourrait ecirctre envisageacutee avant de concevoir des expeacuteriences sur
bovins Bien que ce modegravele animal soit assez eacuteloigneacute du modegravele bovin en ce qui concerne lrsquoeacutetude de la
Perspectives
190
transmission indirecte de M bovis (les cobayes geacutenegraverent par exemple beaucoup moins drsquoaeacuterosols que
les bovins quand ils boivent et ils ne ruminent pas) il constitue un modegravele plus facilement manipulable
et moins coucircteux Il permettrait drsquoaffiner les donneacutees sur les voies de transmission environnementales
potentielles avant de passer sur un modegravele bovin beaucoup plus contraignant de par la taille des
animaux et plus coucircteux
Reacutefeacuterences bibliographiques
Reacutefeacuterences bibliographiques
192
Acevedo-Whitehouse K Vicente J Gortaacutezar C Houmlfle U Fernaacutendez-de-Mera IG Amos W 2005 Genetic resistance to bovine tuberculosis in the Iberian wild boar Mol Ecol 14 3209ndash17
Adams AP Bolin SR Fine AE Bolin CA Kaneene JB 2013 Comparison of PCR versus culture for detection of Mycobacterium bovis after experimental inoculation of various matrices held under environmental conditions for extended periods Appl Environ Microbiol 79 6501ndash6506
Adekambi T Ben Salah S Khlif M Raoult D Drancourt M 2006 Survival of environmental mycobacteria in Acanthamoeba polyphaga Appl Environ Microbiol 72 5974ndash5981
Adeacutekambi T Drancourt M 2004 Dissection of phylogenetic relationships among 19 rapidly growing Mycobacterium species by 16S rRNA hsp65 sodA recA and rpoB gene sequencing Int J Syst Evol Microbiol 54 2095ndash105
Aimeacute B Lequen L Balageas A Haddad N Maugein J 2012 M bovis and M caprae infections in Aquitaine A clinico-epidemiologic study of 15 patients Pathol Biol (Paris) 60 156ndash159
Alexander K A Laver PN Michel AL Williams M van Helden PD Warren RM van Pittius NCG 2010 Novel Mycobacterium tuberculosis complex pathogen M Mungi Emerg Infect Dis 16 1296ndash1299
Allix C Walravens K Saegerman C Godfroid J Supply P Fauville-Dufaux M 2006 Evaluation of the epidemiological relevance of variable-number tandem-repeat genotyping of Mycobacterium bovis and comparison of the method with IS6110 restriction fragment length polymorphism analysis and spoligotyping J Clin Microbiol 44 1951ndash1962
Aacutelvarez AH Estrada-Chaacutevez C Flores-Valdez MA 2009 Molecular findings and approaches spotlighting Mycobacterium bovis persistence in cattle Vet Res 40 1ndash16
Ameni G Vordermeier M Firdessa R Aseffa A Hewinson G Gordon S V Berg S 2011 Mycobacterium tuberculosis infection in grazing cattle in central Ethiopia Vet J 188 359ndash361
ANSES 2011 Tuberculose bovine et faune sauvageRapport Avril 2011
Antoine D Jarlier V 2010 La tuberculose humaine agrave Mycobacterium bovis en France Bull eacutepideacutemiologique santeacute Anim Aliment Septembre
Aranaz A Cousins D Mateos A Domınguez L 2003 Elevation of Mycobacterium tuberculosis subsp caprae (Aranaz et al 1999) to species rank as Mycobacterium caprae comb nov sp Int J Syst Evol Microbiol 53 1785ndash1789
Aranaz A Lieacutebana E Goacutemez-Mampaso Enrique Galaacuten JC Cousins D Ortega A Blaacutezquez J Baquero F Mateos A Suacutearez G Domiacutenguez L 1999 Mycobacterium tuberculosis subsp caprae subsp nov a taxonomic study of a new member of the Mycobacterium tuberculosis complex isolated from goats in Spain Int J Syst Bacteriol 49 1263ndash1273
Archuleta RJ Yvonne Hoppes P Primm TP 2005 Mycobacterium avium enters a state of metabolic dormancy in response to starvation Tuberculosis (Edinb) 85 147ndash58
Aronson T Holtzman A Glover N Boian M Froman S Berlin OGW Hill H Stelma G J 1999 Comparison of large restriction fragments of Mycobacterium avium isolates recovered from AIDS and non-AIDS patients with those of isolates from potable water J Clin Microbiol 37 1008ndash1012
Ayele WY Neill SD Zinsstag J Weiss MG Pavlik I 2004 Bovine tuberculosis an old disease but a new threat to Africa Int J Tuberc Lung Dis 8 924ndash937
Ballesteros C Garrido JM Vicente J Romero B Galindo RC Minguijoacuten E Villar M Martiacuten-Hernando
Reacutefeacuterences bibliographiques
193
MP Sevilla I Juste R Aranaz A de la Fuente J Gortaacutezar C 2009 First data on Eurasian wild boar response to oral immunization with BCG and challenge with a Mycobacterium bovis field strain Vaccine 27 6662ndash6668
Barasona JA Latham MC Acevedo P Armenteros JA Latham ADM Gortazar C Carro F Soriguer RC Vicente J 2014 Spatiotemporal interactions between wild boar and cattle Implications for cross-species disease transmission Vet Res 45 1ndash11
Barasona JA Torres MJ Aznar J Gortaacutezar C Vicente J 2015 DNA detection reveals Mycobacterium tuberculosis complex shedding routes in its wildlife reservoir the Eurasian wild boar Transbound Emerg Dis (December) doi101111tbed12458
Barker J Brown MRW 1994 Trojan Horses of the microbial world protozoa and the survival of bacterial pathogens in the environment Microbiology 140 1253ndash1259
Baubet E Ropert-Coudert Y Brandt S 2003 Seasonal and annual variations in earthworm consumption by wild boar (Sus scrofa scrofa L) Wildl Res 30 179
Ben Salah I Drancourt M 2010 Surviving within the amoebal exocyst the Mycobacterium avium complex paradigm BMC Microbiol 10 99
Beacutenet J Boschiroli ML Dufour B Garin-Bastuji B 2006 Lutte contre la tuberculose bovine en France de 1954 agrave 2005 analyse de la pertinence eacutepideacutemiologique de lrsquoeacutevolution de la reacuteglementation Epideacutemiologie santeacute Anim 50 127ndash143
Benham PF 1985 The behaviour of badgers and cattle and some factors that affect the chance of contact between the species Appl Anim Behav Sci 14 390ndash391
Benham PF Broom DM 1989 Interactions between cattle and badgers at pasture with reference to bovine tuberculosis transmission Br Vet J 145 226ndash241
Bercovier H Kafri O Sela S 1986 Mycobacteria possess a surprisingly small number of ribosomal RNA genes in relation to the size of their genome Biochem Biophys Res Commun 136 1136ndash1141
Berg S Garcia-Pelayo MC Muumlller B Hailu E Asiimwe B Kremer K Dale J Boniotti MB Rodriguez S Hilty M Rigouts L Firdessa R MacHado A Mucavele C Ngandolo BNR Bruchfeld J Boschiroli ML Muumlller A Sahraoui N Pacciarini M Cadmus SI Joloba M Van Soolingen D Michel AL Djoslashnne B Aranaz A Zinsstag J Van Helden P Portaels F Kazwala R Kaumlllenius G Hewinson RG Aseffa A Gordon S V Smith NH 2011 African 2 a clonal complex of Mycobacterium bovis epidemiologically important in East Africa J Bacteriol 193 670ndash678
Bermingham ML Bishop SC Woolliams JA Pong-Wong R Allen AR McBride SH Ryder JJ Wright DM Skuce RA McDowell SW Glass EJ 2014 Genome-wide association study identifies novel loci associated with resistance to bovine tuberculosis Heredity (Edinb) 112 543ndash551
Beširović H Alić A Špičić S Cvetnić Ž Prašović S Velić L 2012 Bovine tuberculosis in Bosnia and Herzegovina caused by Mycobacterium caprae Vet Arh 82 341ndash349
Beumer A King D Donohue M Mistry J Covert T Pfaller S 2010 Detection of Mycobacterium avium subsp paratuberculosis in drinking water and biofilms by quantitative PCR Appl Environ Microbiol 76 7367ndash7370
Biek R OrsquoHare A Wright D Mallon T McCormick C Orton RJ McDowell S Trewby H Skuce RA Kao RR 2012 Whole genome sequencing reveals local transmission patterns of Mycobacterium bovis in sympatric cattle and badger populations PLoS Pathog 8 e1003008
Biet F Boschiroli ML Thorel MF Guilloteau LA 2005 Zoonotic aspects of Mycobacterium bovis and
Reacutefeacuterences bibliographiques
194
Mycobacterium avium-intracellulare complex (MAC) Vet Res36 411-436
Bille E Dauphin B Leto J Bougnoux M-E Beretti J-L Lotz A Suarez S Meyer J Join-Lambert O Descamps P Grall N Mory F Dubreuil L Berche P Nassif X Ferroni A 2012 MALDI-TOF MS Andromas strategy for the routine identification of bacteria mycobacteria yeasts Aspergillus spp and positive blood cultures Clin Microbiol Infect 18 1117ndash1125
Biswas S Rolain JM 2013 Use of MALDI-TOF mass spectrometry for identification of bacteria that are difficult to culture J Microbiol Methods 92 14ndash24
Blackwood KS He C Gunton J Turenne CY Wolfe J Kabani A M 2000 Evaluation of recA sequences for identification of Mycobacterium species J Clin Microbiol 38 2846ndash2852
Bland CS Ireland JM Lozano E Alvarez ME Primm TP 2005 Mycobacterial ecology of the Rio Grande Appl Environ Microbiol 71 5719ndash5727
Boadella M Barasona JA Diaz-Sanchez S Lyashchenko KP Greenwald R Esfandiari J Gortaacutezar C 2012 Performance of immunochromatographic and ELISA tests for detecting fallow deer infected with Mycobacterium bovis Prev Vet Med 104 160ndash164
Boadella M Lyashchenko K Greenwald R Esfandiari J Jaroso R Carta T Garrido JM Vicente J de la Fuente J Gortaacutezar C 2011 Serologic tests for detecting antibodies against Mycobacterium bovis and Mycobacterium avium subspecies paratuberculosis in Eurasian wild boar (Sus scrofa scrofa) J Vet Diagnostic Investig 23 77ndash83
Boumlddinghaus B Rogall T Flohr T Bloumlcker H Boumlttger EC 1990 Detection and identification of mycobacteria by amplification of rRNA J Clin Microbiol 28 1751ndash1759
Boumlhm M Hutchings MR White PCL 2009 Contact networks in a wildlife-livestock host community identifying high-risk individuals in the transmission of bovine TB among badgers and cattle PLoS One 4 e5016
Bollo E Guarda F Capucchio MT Galietti F 1998 Direct detection of Mycobacterium tuberculosis Complex and M avium Complex in tissue specimens from cattle through identification of specific rRNA sequences J Vet Med 45 395ndash400
Boniotti MB Goria M Loda D Garrone A Benedetto A Mondo A Tisato E Zanoni M Zoppi S Dondo A Tagliabue S Bonora S Zanardi G Pacciarini ML 2009 Molecular typing of Mycobacterium bovis strains isolated in Italy from 2000 to 2006 and evaluation of variable-number tandem repeats for geographically optimized genotyping J Clin Microbiol 47 636ndash644
Boon C Dick T 2012 How Mycobacterium tuberculosis goes to sleep the dormancy survival regulator DosR a decade later Future Microbiol 7 513ndash518
Boon C Dick T 2002 Mycobacterium bovis BCG response regulator essential for hypoxic dormancy J Bacteriol 184 6760ndash6767
Boon C Li R Qi R Dick T 2001 Proteins of Mycobacterium bovis BCG induced in the Wayne dormancy model J Bacteriol 183 2672ndash2676
Boulahbal F Robert J Trystram D Benoist A C Vincent V Jarlier V 1998 La tuberculose humaine agrave Mycobacterium bovis en France durant lrsquoanneacutee 1995 Bull eacutepideacutemiologique Hebd 48
Boyard C Vourcrsquoh G Barnouin J 2008 The relationships between Ixodes ricinus and small mammal species at the woodland-pasture interface Exp Appl Acarol 44 61ndash76
Brennan PJ Nikaido H 1995 The envelope of mycobacteria Annu Rev Biochem 64 29ndash63
Reacutefeacuterences bibliographiques
195
Brodin P Eiglmeier K Marmiesse M Billault A Garnier T Niemann S Cole ST Brosch R 2002 Bacterial artificial chromosome-based comparative genomic analysis identifies Mycobacterium microti as a natural ESAT-6 deletion mutant Infect70 5568ndash5578
Brook RK McLachlan SM 2009 Transdisciplinary habitat models for elk and cattle as a proxy for bovine tuberculosis transmission risk Prev Vet Med 91 197ndash208
Brook RK Wal E Vander van Beest FM McLachlan SM 2012 Evaluating use of cattle winter feeding areas by elk and white-tailed deer Implications for managing bovine tuberculosis transmission risk from the ground up Prev Vet Med 1ndash11
Brosch R Gordon SV Marmiesse M Brodin P Buchrieser C Eiglmeier K Garnier T Gutierrez C Hewinson G Kremer K Parsons LM Pym A Samper S van Soolingen D Cole S 2002 A new evolutionary scenario for the Mycobacterium tuberculosis complex Proc Natl Acad Sci U S A 99 3684ndash3689
Brosch R Gordon S V Garnier T Eiglmeier K Frigui W Valenti P Dos Santos S Duthoy S Lacroix C Garcia-Pelayo C Inwald JK Golby P Garcia JN Hewinson RG Behr M A Quail M A Churcher C Barrell BG Parkhill J Cole ST 2007 Genome plasticity of BCG and impact on vaccine efficacy Proc Natl Acad Sci U S A 104 5596ndash5601
Brotherstone S White IMS Coffey M Downs SH Mitchell AP Clifton-Hadley RS More SJ Good M Woolliams JA 2010 Evidence of genetic resistance of cattle to infection with Mycobacterium bovis J Dairy Sci 93 1234ndash1242
Broughan JM Crawshaw TR Downs SH Brewer J Clifton-Hadley RS 2013a Mycobacterium bovis infections in domesticated non-bovine mammalian species Part 2 A review of diagnostic methods Vet J 198 346ndash351
Broughan JM Downs SH Crawshaw TR Upton PA Brewer J Clifton-Hadley RS 2013b Mycobacterium bovis infections in domesticated non-bovine mammalian species Part 1 Review of epidemiology and laboratory submissions in Great Britain 2004-2010 Vet J 198 339ndash345
Buddle BM Wedlock DN Denis M Vordermeier HM Hewinson RG 2011 Update on vaccination of cattle and wildlife populations against tuberculosis Vet Microbiol 151 14ndash22
Butler WR Guthertz LS 2001 Mycolic acid analysis by High-Performance Liquid Chromatography for identification of Mycobacterium species Clin Microbiol Rev 14 704ndash726
Cadmus SI Adesokan HK Jenkins AO van Soolingen D 2009a Mycobacterium bovis and M tuberculosis in goats Nigeria Emerg Infect Dis 15 2066ndash2067
Cadmus SI Jenkins AO Godfroid J Osinusi K Adewole IF Murphy RL Taiwo BO 2009b Mycobacterium tuberculosis and Mycobacterium africanum in stools from children attending an immunization clinic in Ibadan Nigeria Int J Infect Dis 13 740ndash744
Cadmus SI Palmer S Okker M Dale J Gover K Smith N Jahans K Hewinson RG Gordon S V 2006 Molecular analysis of human and bovine tubercle bacilli from a local setting in Nigeria J Clin Microbiol 44 29ndash34
Cadmus SI Yakubu MK Magaji AA Jenkins AO van Soolingen D 2010 Mycobacterium bovis but also M africanum present in raw milk of pastoral cattle in north-central Nigeria Trop Anim Health Prod 42 1047ndash1048
Cambau E Lamaitre N Sougakoff W Jarlier V 2003 Reacutesistance aux antituberculeux Antibiotiques 5 29ndash37
Reacutefeacuterences bibliographiques
196
Carbonnelle B Dailloux M Lebrun L Maugein J Pernot C 2003 Cahier de biologie meacutedicale Ndeg29Mycobacteacuteries mycobacteacuterioses
Carrasco-Garcia R Barasona JA Gort Montoro V Sanchez-Vizcaino JM Vicente J 2015 Wildlife and livestock use of extensive farm resources in South Central Spain implications for disease transmission Eur J Wildl Res62(1) 65-78
Cassidy JP 2006 The pathogenesis and pathology of bovine tuberculosis with insights from studies of tuberculosis in humans and laboratory animal models Vet Microbiol 112 151ndash161
Cassidy JP Bryson DG Pollock JM Evans RT Forster F Neill SD 1999 Lesions in cattle exposed to Mycobacterium bovis-inoculated calves J Comp Pathol 121 321ndash37
Castets M Sarrat H 1968 Bacteriologic aspects of mycobacteria isolated at Dakar in 1967 Bull Soc Med Afr Noire Lang Fr 13 463ndash469
Cavalerie L Courcoul A Boschiroli ML Reacuteveillaud E Gay P 2014 Tuberculose bovine en France en 2014 une situation stable Bull eacutepideacutemiologique santeacute Anim Aliment 4ndash11
Cha D Cheng D Liu M Zeng Z Hu X Guan W 2009 Analysis of fatty acids in sputum from patients with pulmonary tuberculosis using gas chromatography-mass spectrometry preceded by solid-phase microextraction and post-derivatization on the fiber J Chromatogr A 1216 1450ndash1457
Chapinal N Elkin BT Joly DO Schumaker BA Stephen C 2012 Agreement between the caudal fold test and serological tests for the detection of Mycobacterium bovis infection in bison Prev Vet Med 105 326ndash330
Ciliberti A Gavier-Wideacuten D Yon L Hutchings MR Artois M 2015 Prioritisation of wildlife pathogens to be targeted in European surveillance programmes Expert-based risk analysis focus on ruminants Prev Vet Med 118 271ndash284
Cirillo JD Falkow S Tompkins LS Bermudez LE 1997 Interaction of Mycobacterium avium with environmental amoebae enhances virulence Infect Immun 65 3759ndash3767
Cleary GP Corner LAL OrsquoKeeffe J Marples NM 2011 Diet of the European badger (Meles meles) in the Republic of Ireland A comparison of results from an analysis of stomach contents and rectal faeces Mamm Biol 76 470ndash475
Clifton-Hadley RS Wilesmith JW Stuart F A 1993 Mycobacterium bovis in the European badger (Meles meles) epidemiological findings in tuberculous badgers from a naturally infected population Epidemiol Infect 111 9ndash19
Cole ST Brosch R Parkhill J Garnier T Churcher C Harris D Gordon S V Eiglmeier K Gas S Barry CE Tekaia F Badcock K Basham D Brown D Chillingworth T Connor R Davies R Devlin K Feltwell T Gentles S Hamlin N Holroyd S Hornsby T Jagels K Krogh A McLean J Moule S Murphy L Oliver K Osborne J Quail MA Rajandream MA Rogers J Rutter S Seeger K Skelton J Squares R Squares S Sulston JE Taylor K Whitehead S Barrell BG 1998 Deciphering the biology of Mycobacterium tuberculosis from the complete genome sequence Nature 396 190
Coleman JD Cooke MM 2001 Mycobacterium bovis infection in wildlife in New Zealand Tuberculosis (Edinb) 81 191ndash202
Collins CH Grange JM 1983 The bovine tubercle bacillus J Appl Bacteriol 55 13ndash29
Cooper SM Scott HM de la Garza GR Deck AL Cathey JC 2010 Distribution and interspecies contact of feral Swine and cattle on rangeland in South Texas implications for disease transmission J Wildl Dis 46 152ndash164
Reacutefeacuterences bibliographiques
197
Corner LAL 2006 The role of wild animal populations in the epidemiology of tuberculosis in domestic animals How to assess the risk Vet Microbiol 112 303ndash312
Corner LAL Costello E Lesellier S OrsquoMeara D Sleeman DP Gormley E 2007 Experimental tuberculosis in the European badger (Meles meles) after endobronchial inoculation of Mycobacterium bovis I Pathology and bacteriology Res Vet Sci 83 53ndash62
Corner LAL Murphy D Gormley E 2011 Mycobacterium bovis infection in the Eurasian badger (Meles meles) the disease pathogenesis epidemiology and control J Comp Pathol 144 1ndash24
Corner LAL OrsquoMeara D Costello E Lesellier S Gormley E 2012 The distribution of Mycobacterium bovis infection in naturally infected badgers Vet J 194 166ndash172
Coscolla M Gagneux S 2014 Consequences of genomic diversity in Mycobacterium tuberculosis Semin Immunol 26 431ndash444
Coscolla M Lewin A Metzger S Maetz-Rennsing K Calvignac-Spencer S Nitsche A Dabrowski PW Radonic A Niemann S Parkhill J Couacy-Hymann E Feldman J Comas I Boesch C Gagneux S Leendertz FH 2013 Novel Mycobacterium tuberculosis complex isolate from a wild chimpanzee Emerg Infect Dis 19 969ndash976
Cosivi O Grange JM Daborn CJ Raviglione MC Fujikura T Cousins D Robinson R A Huchzermeyer HF de Kantor I Meslin FX 1999 Zoonotic tuberculosis due to Mycobacterium bovis in developing countries Emerg Infect Dis 4 59ndash70
Coulombe F Divangahi M Veyrier F de Leacuteseacuteleuc L Gleason JL Yang Y Kelliher MA Pandey AK Sassetti CM Reed MB Behr M 2009 Increased NOD2-mediated recognition of N-glycolyl muramyl dipeptide J Exp Med 206 1709ndash1716
Courcoul A Moyen J-L Brugegravere L Faye S Heacutenault S Gares H Boschiroli ML 2014 Estimation of sensitivity and specificity of bacteriology histopathology and PCR for the confirmatory diagnosis of bovine tuberculosis using latent class analysis PLoS One 9 e90334
Courtenay O Reilly L A Sweeney FP Hibberd V Bryan S Ul-Hassan A Newman C Macdonald DW Delahay RJ Wilson GJ Wellington EMH 2006 Is Mycobacterium bovis in the environment important for the persistence of bovine tuberculosis Biol Lett 2 460ndash462
Cousins D V Bastida R Cataldi A Quse V Redrobe S Dow S Duignan P Murray A Dupont C Ahmed N Collins DM Butler WR Dawson D Rodriacuteguez D Loureiro J Romano MI Alito A Zumarraga M Bernardelli A 2003 Tuberculosis in seals caused by a novel member of the Mycobacterium tuberculosis complex Mycobacterium pinnipedii sp nov Int J Syst Evol Microbiol 53 1305ndash1314
Couvin D Rastogi N 2014 La mise en place de bases de donneacutees des geacutenotypes circulants du complexe Mycobacterium tuberculosis et des outils web permettant de mieux surveiller comprendre et controcircler lrsquoeacutepideacutemie de la tuberculose dans le monde EuroReference Cahier num 12
Cowie CE Hutchings MR Barasona JA Gortaacutezar C Vicente J White PCL 2015 Interactions between four species in a complex wildlife livestock disease community implications for Mycobacterium bovis maintenance and transmission Eur J Wildl Res62(1) 51-64
Cvetnic Z Katalinic-Jankovic V Sostaric B Spicic S Obrovac M Marjanovic S Benic M Kirin BK Vickovic I 2007 Mycobacterium caprae in cattle and humans in Croatia Int J Tuberc Lung Dis 11 652ndash658
Dale JW Brittain D Cataldi AA Cousins D Crawford JT Driscoll J Heersma H Lillebaek T Quitugua T Rastogi N Skuce RA Sola C Van Soolingen D Vincent V 2001 Spacer oligonucleotide typing of bacteria of the Mycobacterium tuberculosis complex Recommendations for standardised nomenclature
Reacutefeacuterences bibliographiques
198
Int J Tuberc Lung Dis 5 216ndash219
Dalley D Daveacute D Lesellier S Palmer S Crawshaw T Hewinson RG Chambers M 2008 Development and evaluation of a gamma-interferon assay for tuberculosis in badgers (Meles meles) Tuberculosis 88 235ndash243
Danelishvili L Wu M Stang B Harriff M Cirillo SLG Cirillo JD Bildfell R Arbogast B Bermudez LE 2007 Identification of Mycobacterium avium pathogenicity island important for macrophage and amoeba infection Proc Natl Acad Sci U S A 104 11038ndash11043
Dang NA Kuijper S Walters E Claassens M van Soolingen D Vivo-Truyols G Janssen H-G Kolk AHJ 2013 Validation of biomarkers for distinguishing Mycobacterium tuberculosis from non-tuberculous mycobacteria using gas chromatography-mass spectrometry and chemometrics PLoS One 8 e76263
Daniel TM 2006 The history of tuberculosis Respir Med 100 1862ndash1870
David H Levy Frebaut U Thorel MF 1989 Meacutethodes de laboratoire pour mycobacteacuteriologie clinique Institut Pasteur p 35-36
Dawson KL Bell A Kawakami RP Coley K Yates G Collins DM 2012 Transmission of Mycobacterium orygis (M tuberculosis complex species) from a tuberculosis patient to a dairy cow in New Zealand J Clin Microbiol 50 3136ndash3138
de Jong BC Antonio M Gagneux S 2010 Mycobacterium africanum-review of an important cause of human tuberculosis in West Africa PLoS Negl Trop Dis 4(9) e744
de la Rua-Domenech R 2006 Human Mycobacterium bovis infection in the United Kingdom Incidence risks control measures and review of the zoonotic aspects of bovine tuberculosis Tuberculosis (Edinb) 86 77ndash109
de la Rua-Domenech R Goodchild AT Vordermeier HM Hewinson RG Christiansen KH Clifton-Hadley RS 2006 Ante mortem diagnosis of tuberculosis in cattle a review of the tuberculin tests gamma-interferon assay and other ancillary diagnostic techniques Res Vet Sci 81 190ndash210
Dean GS Rhodes SG Coad M Adam O Cockle PJ Clifford DJ Hewinson RG Vordermeier HM 2005 Minimum infective dose of Mycobacterium bovis in cattle Infect Immun 73 6467ndash6471
Delahay RJ Cheeseman CL Clifton-Hadley RS 2001 Wildlife disease reservoirs the epidemiology of Mycobacterium bovis infection in the European badger (Meles meles) and other British mammals Tuberculosis (Edinb) 81 43ndash49
Delahay RJ De Leeuw ANS Barlow AM Clifton-Hadley RS Cheeseman CL 2002 The status of Mycobacterium bovis Infection in UK Wild Mammals A Review Vet J 164 90ndash105
Delahay RJ Langton S Smith GC Clifton-Hadley RS Cheeseman CL 2000 The spatio-temporal distribution of Mycobacterium bovis (bovine tuberculosis) infection in a high-density badger population J Anim Ecol 69 428ndash441
Delahay RJ Walker N Smith G Wilkinson D Clifton-Hadley R Cheeseman C Tomlinson A Chambers M 2013 Long-term temporal trends and estimated transmission rates for Mycobacterium bovis infection in an undisturbed high-density badger (Meles meles) population Epidemiol Infect 141 1445ndash1456
Delaune D Janvier F Rapp C Geacuterocircme P Fabre M Soler C Meacuterens A 2012 Actualiteacutes de lrsquoinfection agrave Mycobacterium bovis en France agrave propos de 4 cas Ann Biol Clin (Paris) 70 231ndash236
Desikan S Narayanan S 2015 Genetic markers genotyping methods amp next generation sequencing in Mycobacterium tuberculosis Indian J Med Res 141 761ndash774
Reacutefeacuterences bibliographiques
199
Desmond E Ahmed AT Probert WS Ely J Jang Y Sanders C A Lin SY Flood J 2004 Mycobacterium africanum Cases California Emerg Infect Dis 10 921ndash923
Dewes HF 1996 The rate of soil ingestion by dairy cows and the effect on available copper calcium sodium and magnesium N Z Vet J 44 199ndash200
Dick T Lee BH Murugasu-Oei B 1998 Oxygen depletion induced dormancy in Mycobacterium smegmatis FEMS Microbiol Lett 163 159ndash164
Dippenaar A Parsons SDC Sampson SL van der Merwe RG Drewe JA Abdallah AM Siame KK Gey van Pittius NC van Helden PD Pain A Warren RM 2015 Whole genome sequence analysis of Mycobacterium suricattae Tuberculosis 95(6) 682-688
Djelouadji Z Raoult D Drancourt M 2011 Palaeogenomics of Mycobacterium tuberculosis Epidemic bursts with a degrading genome Lancet Infect Dis 11 641ndash650
Dondo A Zoppi S Rossi F Chiavacci L Barbaro A Garrone A Benedetto A Goria M 2007 Mycobacteriosis in Wild Boar Results of 2000-2006 Activity in North-Western Italy Epideacutemiologie santeacute Anim 51 35ndash42
Drancourt M Adeacutekambi T Raoult D 2007 Interactions between Mycobacterium xenopi amoeba and human cells J Hosp Infect 65 138ndash142
Drewe JA OrsquoConnor HM Weber N McDonald RA Delahay RJ 2013 Patterns of direct and indirect contact between cattle and badgers naturally infected with tuberculosis Epidemiol Infect 141 1467ndash1475
Drewe JA Tomlinson AJ Walker NJ Delahay RJ 2010 Diagnostic accuracy and optimal use of three tests for tuberculosis in live badgers PLoS One 5 e11196
Duffield BJ Young DA 1985 Survival of Mycobacterium bovis in defined environmental conditions Vet Microbiol 10 193ndash197
Dutau G 2005 Petite histoire illustreacutee de la tuberculose The history of tuberculosis Arch Pediatr 12 88ndash95
Dziadek J Sajduda A Boruń M 2001 Specificity of insertion sequence-based PCR assays for Mycobacterium tuberculosis complex Int J Tuberc Lung Dis 5 569ndash574
EFSA 2014 The European Union summary report on trends ans sources of zoonotic agents and food-borne outbreaks in 2012 EFSA J 12 169
El Helou G Viola GM Hachem R Han XY Raad II 2013 Rapidly growing mycobacterial bloodstream infections Lancet Infect Dis 13 166ndash174
El Kheacutechine A Couderc C Flaudrops C Raoult D Drancourt M 2011 Matrix-assisted laser desorptionionization time-of-flight mass spectrometry identification of mycobacteria in routine clinical practice PLoS One 6 e24720
El Kheacutechine A Henry M Raoult D Drancourt M 2009 Detection of Mycobacterium tuberculosis complex organisms in the stools of patients with pulmonary tuberculosis Microbiology 155 2384ndash2389
Elliott GN Hough RL Avery LM Maltin C A Campbell CD 2014 Environmental risk factors in the incidence of Johnersquos disease Crit Rev Microbiol 7828 1ndash20
Evans JT Smith EG Banerjee A Smith RMM Dale J Innes JA Hunt D Tweddell A Wood A Anderson C Hewinson RG Smith NH Hawkey PM Sonnenberg P 2007 Cluster of human tuberculosis caused by Mycobacterium bovis evidence for person-to-person transmission in the UK Lancet 369 1270ndash1276
Reacutefeacuterences bibliographiques
200
Fabre M Koeck JL Le Flegraveche P Simon F Herveacute V Vergnaud G Pourcel C 2004 High genetic diversity revealed by variable-number tandem repeat genotyping and analysis of hsp65 gene polymorphism in a large collection of ldquoMycobacterium canettiirdquo strains indicates that the M tuberculosis complex is a recently emerged clone of M can J Clin Microbiol 42 3248ndash3255
Falkinham JO 2016 Nontuberculous mycobacteria community and nosocomial waterborne opportunistic pathogens Clin Microbiol Newsl 38 1ndash7
Falkinham JO Norton CD Mark W 2001 Factors influencing numbers of Mycobacterium avium Mycobacterium intracellulare and other Mycobacteria in drinking water distribution systems Appl Environ Microbiol 67 1225ndash1231
Fanning A Edwards S 1991 Mycobacterium bovis infection in human beings in contact with elk (Cervus elaphus) in Alberta Canada Lancet 338 1253ndash1255
Fediaevsky A Beacutenet J-J Boschiroli ML Riviegravere J Hars J 2012 La tuberculose bovine en France en 2011 poursuite de la reacuteduction du nombre de foyers Bull eacutepideacutemiologique santeacute Anim Aliment Novembre 4ndash12
Fediaevsky A Courcoul A Boschiroli ML Reveillaud E 2014 Tuberculose bovine en France reacutesultats drsquoune strateacutegie plus offensive Bull eacutepideacutemiologique santeacute Anim Aliment 64 4ndash11
Fediaevsky A Courcoul A Boschiroli ML Reveillaud E 2013 Tuberculose bovine en France des signaux favorables mais une situation toujours complexe dans certaines zones Bull Epideacutemiologique - Santeacute Anim Aliment 59 4ndash10
Fetene T Kebede N Alem G 2011 Tuberculosis infection in animal and human populations in three districts of Western Gojam Ethiopia Zoonoses Public Health 58 47ndash53
Fifis T Corner LAL Rothel JS Wood PR 1994 Cellular and humoral immune responses of cattle to purified Mycobacterium bovis antigens Scand J Immunol 39 267ndash274
Fine AE Bolin CA Gardiner JC Kaneene JB 2011a A study of the persistence of Mycobacterium bovis in the environment under natural weather conditions in Michigan USA Vet Med Int 2011 ID 765430 1-12
Fine AE OrsquoBrien DJ Winterstein SR Kaneene JB 2011b An effort to isolate Mycobacterium bovis from environmental substrates during investigations of bovine tuberculosis transmission sites (cattle farms and wildlife areas) in Michigan USA ISRN Vet Sci 2011 ID 787181 1-11
Fischer OA Matlova L Bartl J Dvorska L Svastova P Du Maine R Melicharek I Bartos M Pavlik I 2003 Earthworms (Oligochaeta Lumbricidae) and mycobacteria Vet Microbiol 91 325ndash338
Flores Valdez MA Schoolnik GK 2010 DosR-regulon genes induction in Mycobacterium bovis BCG under aerobic conditions Tuberculosis (Edinb) 90 197ndash200
Forrellad M Klepp L Gioffreacute A 2013 Virulence factors of the Mycobacterium tuberculosis complex Virulence 4 3ndash66
Frank W Reisinger EC Brandt-Hamerla W Schwede I Handrick W 2009 Mycobacterium microti--pulmonary tuberculosis in an immunocompetent patient Wien Klin Wochenschr 121 282ndash6
Frottier J Eliaszewicz M Arlet V Gaudillat C 1990 Infections caused by Mycobacterium africanum Bull Acad Natl Med 174 29ndash35
Gallagher J Clifton-Hadley RS 2000 Tuberculosis in badgers a review of the disease and its significance for other animals Res Vet Sci 69 203ndash217
Reacutefeacuterences bibliographiques
201
Gallagher J Horwill DM 1977 A selective oleic acid albumin agar medium for the cultivation of Mycobacterium bovis J Hyg (Lond) 79 155ndash160
Gallagher J Monies R Gavier-Widen M Rule B 1998 Role of infected non-diseased badgers in the pathogenesis of tuberculosis in the badger Vet Rec 142 710ndash714
Garbaccio S Macias A Shimizu E Paolicchi F Pezzone N Magnano G Zapata L Abdala A Tarabla H Peyru M Caimi K Zumaacuterraga M Canal A Cataldi A 2014 Association between spoligotype-VNTR types and virulence of Mycobacterium bovis in cattle Virulence 5 297ndash302
Garbaccio SG Cataldi A 2010 Evaluation of an immunomagnetic capture method followed by PCR to detect Mycobacterium bovis in tissue samples from cattle Rev Argent Microbiol 42 247ndash53
Garnett BT Delahay RJ Roper TJ 2002 Use of cattle farm resources by badgers (Meles meles) and risk of bovine tuberculosis (Mycobacterium bovis) transmission to cattle Proc Biol Sci 269 1487ndash1491
Garnier T Eiglmeier K Camus J-C Medina N Mansoor H Pryor M Duthoy S Grondin S Lacroix C Monsempe C Simon S Harris B Atkin R Doggett J Mayes R Keating L Wheeler PR Parkhill J Barrell BG Cole ST Gordon S V Hewinson RG 2003 The complete genome sequence of Mycobacterium bovis Proc Natl Acad Sci U S A 100 7877ndash7882
Gavier-Widen D Chambers MA Palmer N Newell DG Hewinson RG 2001 Pathology of natural Mycobacterium bovis infection in European badgers (Meles meles) and its relationship with bacterial excretion Vet Rec 148 299ndash304
Gavier-Wideacuten D Cooke MM Gallagher J Chambers MA Gortaacutezar C 2009 A review of infection of wildlife hosts with Mycobacterium bovis and the diagnostic difficulties of the ldquono visible lesionrdquo presentation N Z Vet J 57 122ndash131
Genov I 1965 The effects of certain physical and chemical agents on Mycobacterium tuberculosis Veteriaacuternı Med Nauk Sofia 2 97ndash107
Gerasimova A Kazakov AE Arkin AP Dubchak I Gelfand MS 2011 Comparative genomics of the dormancy regulons in mycobacteria J Bacteriol 193 3446ndash3452
Ghodbane R Mba Medie F Lepidi H Nappez C Drancourt M 2014 Long-term survival of tuberculosis complex mycobacteria in soil Microbiology 160 496ndash501
Ghosh J Larsson P Singh B Pettersson BMF Islam NM Sarkar SN Dasgupta S Kirsebom LA 2009 Sporulation in mycobacteria Proc Natl Acad Sci U S A 106 10781ndash10786
Goodchild A V Clifton-Hadley RS 2001 Cattle-to-cattle transmission of Mycobacterium bovis Tuberculosis (Edinb) 81 23ndash41
Gordon S V Heym B Parkhill J Barrell B Cole ST 1999 New insertion sequences and a novel repeated sequence in the genome of Mycobacterium tuberculosis H37Rv Microbiology 145 Pt 4 881ndash892
Gormley E 1999 Detection of Mycobacterium bovis lymphocyte stimulating antigens in culture filtrates of a recombinant Mycobacterium smegmatis cosmid library Vaccine 17 2792ndash2801
Gormley E Doyle MB Fitzsimons T McGill K Collins JD 2006 Diagnosis of Mycobacterium bovis infection in cattle by use of the gamma-interferon (Bovigam) assay Vet Microbiol 112 171ndash179
Gortaacutezar C Delahay RJ McDonald RA Boadella M Wilson GJ Gavier-Wideacuten D Acevedo P 2012 The status of tuberculosis in European wild mammals Mamm Rev 42 193ndash206
Gortaacutezar C Torres MJ Vicente J Acevedo P Reglero M de la Fuente J Negro JJ Aznar-Martiacuten J
Reacutefeacuterences bibliographiques
202
2008 Bovine tuberculosis in Dontildeana Biosphere Reserve The role of wild ungulates as disease reservoirs in the last Iberian lynx strongholds PLoS One 3(7) e2776
Grange JM 2001 Mycobacterium bovis infection in human beings Tuberculosis (Edinb) 81 71ndash77
Grange JM Yates MD 1994 Zoonotic aspects of Mycobacterium bovis infection Vet Microbiol 40 137ndash151
Grange JM Yates MD 1989 Incidence and nature of human tuberculosis due to Mycobacterium africanum in South-East England 1977-1987 Epidemiol Infect 103 127ndash132
Greub G Raoult D 2004 Microorganisms resistant to free-living amoebae Clin Microbiol Rev 17 413ndash433
Griffin JFT Rodgers CR Liggett S Mackintosh CG 2006 Tuberculosis in ruminants Characteristics of intra-tonsilar Mycobacterium bovis infection models in cattle and deer Tuberculosis 86 404ndash418
Griffin JM Martin SW Thorburn MA Eves JA Hammond RF 1996 A case-control study on the association of selected risk factors with the occurrence of bovine tuberculosis in the Republic of Ireland Prev Vet Med 27 217ndash229
Guillet-Caruba C Martinez V Doucet-Populaire F 2014 The new tools of microbiological diagnosis of tuberculosis La Rev meacutedecine interne 35 794ndash800
Gunn-Moore DA McFarland SE Brewer JI Crawshaw TR Clifton-Hadley RS Kovalik M Shaw DJ 2011 Mycobacterial disease in cats in Great Britain I Culture results geographical distribution and clinical presentation of 339 cases J Feline Med Surg 13 934ndash944
Gunn-Moore DA Jenkins PA Lucke VM 1996 Feline tuberculosis a literature review and discussion of 19 cases caused by an unusual mycobacterial variant Vet Rec 138 53ndash58
Gutierrez MC Brisse S Brosch R Fabre M Omaiumls B Marmiesse M Supply P Vincent V 2005 Ancient origin and gene mosaicism of the progenitor of Mycobacterium tuberculosis PLoS Pathog 1(1) e5
Haddad N Ostyn A Karoui C Masselot M Thorel MF Hughes SL Inwald J Hewinson RG Durand B 2001 Spoligotype diversity of Mycobacterium bovis strains isolated in France from 1979 to 2000 J Clin Microbiol 39 3623ndash3632
Harris NB Payeur J Bravo D Osorio R Stuber T Farrell D Paulson D Treviso S Mikolon A Rodriguez-Lainz A Cernek-Hoskins S Rast R Ginsberg M Kinde H 2007 Recovery of Mycobacterium bovis from soft fresh cheese originating in Mexico Appl Environ Microbiol 73 1025ndash1028
Hars J Richomme C Boschiroli ML 2010 La tuberculose bovine dans la faune sauvage en France Bull Epideacutemiologique - Santeacute Anim Aliment 38 28ndash31
Hauer A Cruz K De Cochard T Godreuil S Karaoui C Henault S Bulach T Banuls A Biet F Boschiroli ML 2015 Genetic evolution of Mycobacterium bovis causing tuberculosis in livestock and wildlife in France since 1978 PLoS One 10(2) e 0117103
Haydon DT Cleaveland S Taylor LH Laurenson MK 2002 Identifying reservoirs of infection A conceptual and practical challenge Emerg Infect Dis 8 1468ndash1473
Hennessee CT Seo JS Alvarez AM Li QX 2009 Polycyclic aromatic hydrocarbon-degrading species isolated from Hawaiian soils Mycobacterium crocinum sp nov Mycobacterium pallens sp nov Mycobacterium rutilum sp nov Mycobacterium rufum sp nov and Mycobacterium aromaticivorans sp nov Int J Syst Evol Microbiol 59 378ndash387
Hines N Payeur JB Hoffman LJ 2006 Comparison of the recovery of Mycobacterium bovis isolates using
Reacutefeacuterences bibliographiques
203
the BACTEC MGIT 960 system BACTEC 460 system and Middlebrook 7H10 and 7H11 solid media J Vet Diagn Invest 18 243ndash250
Hmama Z Pena-Diaz S Sunil J Av-Gay Y 2015 Immunoevasion and immunosuppression of the macrophage by Mycobacterium tuberculosis Immunol Rev 264 220ndash232
Hoefsloot W Van Ingen J Andrejak C Aumlngeby K Bauriaud R Bemer P Beylis N Boeree MJ Cacho J Chihota V Chimara E Churchyard G Cias R Daza R Daley CL Dekhuijzen PNR Domingo D Drobniewski F Esteban J Fauville-Dufaux M Folkvardsen DB Gibbons N Goacutemez-Mampaso E Gonzalez R Hoffmann H Hsueh PR Indra A Jagielski T Jamieson F Jankovic M Jong E Keane J Koh WJ Lange B Leao S Macedo R Mannsaringker T Marras TK Maugein J Milburn HJ Mlinkoacute T Morcillo N Morimoto K Papaventsis D Palenque E Paez-Pentildea M Piersimoni C Polanovaacute M Rastogi N Richter E Ruiz-Serrano MJ Silva A Da Silva MP Simsek H Van Soolingen D Szaboacute N Thomson R Fernandez TT Tortoli E Totten SE Tyrrell G Vasankari T Villar M Walkiewicz R Winthrop KL Wagner D 2013 The geographic diversity of nontuberculous mycobacteria isolated from pulmonary samples An NTM-NET collaborative study Eur Respir J 42 1604ndash1613
Hohn A Lowenstein 1926 Die Kulture des tuberklebacillus zur diagnose der tuberculose Zeut fur BaktI Orig 98 460ndash477
Hongmanee P Stender H Rasmussen OF 2001 Evaluation of a fluorescence in situ hybridization assay for differentiation between tuberculous and nontuberculous Mycobacterium species in smears of Lowenstein-Jensen and Mycobacteria Growth Indicator Tube cultures using peptide nucleic acid prob J Clin Microbiol 39 1032ndash1035
Hormisch D Brost I Kohring G Giffhorn F Kroppenstedt RM Stackebrandt E Faumlrber P 2004 Mycobacterium fluoranthenivorans sp nov a Fluoranthene and Aflatoxin B 1 Degrading Bacterium from Contaminated Soil of a Former Coal Gas Plant Syst Appl Microbiol 27 653ndash660
Huard RC Claudio L Lazzarini DO Butler WR Soolingen D Van Ho JL 2003 PCR-based method to differentiate the subspecies of the Mycobacterium tuberculosis complex on the basis of genomic deletions J Clin Microbiol 41 1637ndash1650
Huard RC Fabre M de Haas P Lazzarini LCO van Soolingen D Cousins D Ho JL 2006 Novel genetic polymorphisms that further delineate the phylogeny of the Mycobacterium tuberculosis complex J Bacteriol 188 4271ndash4287
Humblet MF Boschiroli ML Saegerman C 2009 Classification of worldwide bovine tuberculosis risk factors in cattle A stratified approach Vet Res 40(5) 1-24
Hutchings MR Harris S 1999 Quantifying the risks of TB infection to cattle posed by badger excreta Epidemiol Infect 122 167ndash173
Hutchings MR Harris S 1997 Effects of farm management practices on cattle grazing behaviour and the potential for transmission of bovine tuberculosis from badgers to cattle Vet J 153 149ndash162
Hutchings MR Service KM Harris S 2002 Is population density correlated with faecal and urine scent marking in European badgers (Meles meles) in the UK Mamm Biol 67 286ndash293
Hutchings MR Service KM Harris S 2001 Defecation and urination patterns of badgers Meles meles at low density in south west England- Acta Theriol (Warsz) 46 87ndash96
Huws SA McBain AJ Gilbert P 2005 Protozoan grazing and its impact upon population dynamics in biofilm communities J Appl Microbiol 98 238ndash244
Idigoras P Peacuterez-Trallero E Alcorta M Gutieacuterrez C Muntildeoz-Baroja I 1995 Rapid detection of tuberculous
Reacutefeacuterences bibliographiques
204
and non-tuberculous mycobacteria by microscopic observation of growth on Middlebrook 7H11 agar Eur J Clin Microbiol Infect Dis 14 6ndash10
Iivanainen E Martikainen PJ Vaumlaumlnaumlnen PK Vaananen PK Katila M 1999 Environmental factors affecting the occurrence of mycobacteria in brook waters J Appl Microbiol 86 673ndash681
Jackson R De Lisle GW Morris RS 1995 A study of the environmental survival of Mycobacterium bovis on a farm in New Zealand N Z Vet J 43 346ndash352
Jahans K Palmer S Inwald J Brown J Abayakoon S 2004 Isolation of Mycobacterium microti from a male Charolais-Hereford cross Vet Rec 155 373ndash374
Jalava K Jones JA Goodchild T Clifton-Hadley R Mitchell A Story A Watson JM 2007 No increase in human cases of Mycobacterium bovis disease despite resurgence of infections in cattle in the United Kingdom Epidemiol Infect 135 40ndash45
Jenkins AO Cadmus SI Venter EH Pourcel C Hauk Y Vergnaud G Godfroid J 2011 Molecular epidemiology of human and animal tuberculosis in Ibadan Southwestern Nigeria Vet Microbiol 151 139ndash147
Jenkins HE Morrison WI Cox DR Donnelly CA Johnston WT Bourne FJ Clifton-Hadley RS Gettinby G McInerney JP Watkins GH Woodroffe R 2008 The prevalence distribution and severity of detectable pathological lesions in badgers naturally infected with Mycobacterium bovis Epidemiol Infect 136 1350ndash1361
Jensen KE 1954 Presence and destruction of tubercle bacilli in sewage Bull World Health Organ 10 171ndash179
Jimeacutenez BA Hinojoza-Loza E Flores-Valdez MA Prado-Montes de Oca E Allen K Estrada-Chaacutevez C Herrera-Rodriacuteguez S Flores-Fernaacutendez JM Martiacutenez-Velaacutezquez M Hernaacutendez-Gutieacuterrez R Alvarez AH 2013 Expression of non-replicating persistence associated genes of Mycobacterium bovis in lymph nodes from skin test-reactor cattle Microb Pathog 61-62 23ndash28
Johnson L Dean G Rhodes S Hewinson G Vordermeier M Wangoo A 2007 Low-dose Mycobacterium bovis infection in cattle results in pathology indistinguishable from that of high-dose infection Tuberculosis (Edinb) 87 71ndash76
Kaal E Kolk AHJ Kuijper S Janssen H-G 2009 A fast method for the identification of Mycobacterium tuberculosis in sputum and cultures based on thermally assisted hydrolysis and methylation followed by gas chromatography-mass spectrometry J Chromatogr A 1216 6319ndash6325
Kamerbeek J Schouls L Kolk A van Agterveld M van Soolingen D Kuijper S Bunschoten A Molhuizen H Shaw R Goyal M van Embden J 1997 Simultaneous detection and strain differentiation of Mycobacterium tuberculosis for diagnosis and epidemiology J Clin Microbiol 35 907ndash914
Kaprelyants AS Gottschal JC Kell DB 1993 Dormancy in non-sporulating bacteria FEMS Microbiol Rev 104 271ndash285
Kasai H Ezaki T Harayama S 2000 Differentiation of phylogenetically related slowly growing mycobacteria by their gyrB sequences J Clin Microbiol 38(1) 301
Kazda J Pavlik I Falkinham JO Hruska K 2009 The Ecology of Mycobacteria Impact on Animalrsquos and Humans Health Springer Netherlands Dordrecht
Khan NA 2006 Acanthamoeba biology and increasing importance in human health FEMS Microbiol Rev 30 564ndash595
Kiers A Klarenbeek A Mendelts B Van Soolingen D Koeumlter G 2008 Transmission of Mycobacterium
Reacutefeacuterences bibliographiques
205
pinnipedii to humans in a zoo with marine mammals Int J Tuberc Lung Dis 12 1469ndash1473
King HC Murphy A James P Travis E Porter D Hung Y-J Sawyer J Cork J Delahay RJ Gaze W Courtenay O Wellington EM 2015a The variability and seasonality of the environmental reservoir of Mycobacterium bovis shed by wild European badgers Sci Rep 5 12318
King HC Murphy A James P Travis E Porter D Sawyer J Cork J Delahay RJ Gaze W Courtenay O Wellington EM 2015b Performance of a non-invasive test for detecting Mycobacterium bovis shedding in European badger (Meles meles) populations J Clin Microbiol 53(7) 2316-2323
Kirchheimer WF Storrs EE 1971 Attempts to establish the armadillo (Dasypus novemcinctus Linn) as a model for the study of leprosy I Report of lepromatoid leprosy in an experimentally infected armadillo Int J Lepr Other Mycobact Dis 39 693ndash702
Kirschner P Springer B Vogel U Meier A Wrede A Kiekenbeck M Bange F Bottger EC 1993 Genotypic identification of Mycobacteria by nucleic acid sequence determination report of a 2-year experience in a clinical laboratory J Clin Microbiol 31 2882ndash2889
Kleeberg HH 1984 Tuberculose humaine drsquoorigine bovine et santeacute publique Rev Sci Tech 3 33ndash54
Koeck JL Fabre M Simon F Daffeacute M Garnotel Eacute Matan A B Geacuterocircme P Bernatas JJ Buisson Y Pourcel C 2011 Clinical characteristics of the smooth tubercle bacilli ldquoMycobacterium canettiirdquo infection suggest the existence of an environmental reservoir Clin Microbiol Infect 17 1013ndash1019
Krasnow I Wayne LG 1966 Sputum Digestion I The mortality rate of tubercle bacilli in various digestion systems Am J Clin Pathol 45 352ndash355
Krishna-Prasad BN Gupta SK 1978 Preliminary report on engulfment and retention of mycobacteria by trophozoites of axenically grown Acanthamoeba castellanii Douglas Curr Sci 47 245ndash247
Kruuk H 1978 Foraging and spatial organisation of the European badger Meles meles L Behav Ecol Sociobiol 4 75ndash89
Kubica GP Dye WE Cohn ML Middlebrook GSD and D with N-A-L-CH for C of M 1963 Sputum digestion and decontamination with N-acetyl-L-cysteinemdashsodium hydroxide for culture of mycobacteria Am Rev Respir Dis 87 775ndash779
Kubica T Rusch-Gerdes S Niemann S 2003 Mycobacterium bovis subsp caprae Caused One-Third of Human M bovis-Associated Tuberculosis Cases Reported in Germany between 1999 and 2001 J Clin Microbiol 41 3070ndash3077
Kukielka E Barasona JA Cowie CE Drewe J a Gortaacutezar C Cotarelo I Vicente J 2013 Spatial and temporal interactions between livestock and wildlife in South Central Spain assessed by camera traps Prev Vet Med 112 213ndash221
Kulka K Hatfull G Ojha AK 2012 Growth of Mycobacterium tuberculosis biofilms J Vis Exp (60) e3820
Lamont EA Bannantine JP Armieacuten A Ariyakumar DS Sreevatsan S 2012 Identification and characterization of a spore-like morphotype in chronically starved Mycobacterium avium subsp paratuberculosis cultures PLoS One 7 e30648
Laprie C Duboy J Malik R Fyfe J 2013 Feline cutaneous mycobacteriosis a review of clinical pathological and molecular characterization of one case of Mycobacterium microti skin infection and nine cases of feline leprosy syndrome from France and New Caledonia Vet Dermatol 24(6) 561ndash9 e133ndash4
Lavania M Katoch K Sachan P Dubey A Kapoor S Kashyap M Chauhan DS Singh HB Sharma VD Jadhav RS Katoch VM 2006 Detection of Mycobacterium leprae DNA from soil samples by PCR
Reacutefeacuterences bibliographiques
206
targeting RLEP sequences J Commun Dis 38 269ndash273
Le Dantec C Duguet J-P Montiel A Dumoutier N Dubrou S Vincent V 2002 Occurrence of Mycobacteria in Water Treatment Lines and in Water Distribution Systems Appl Environ Microbiol 68 5318ndash5325
Lee H Park H Cho S Bai G Kim S 2000 Species identification of Mycobacteria by PCR-Restriction Fragment Length Polymorphism of the rpoB gene J Clin Microbiol 38 2966ndash2971
Lefmann M Schweickert B Buchholz P Goumlbel UB Ulrichs T Seiler P Theegarten D Moter A 2006 Evaluation of peptide nucleic acid-fluorescence in situ hybridization for identification of clinically relevant mycobacteria in clinical specimens and tissue sections J Clin Microbiol 44 3760ndash3767
Lehtola MJ Torvinen E Miettinen IT Keevil CW 2006 Fluorescence in situ hybridization using peptide nucleic acid probes for rapid detection of Mycobacterium avium subsp avium and Mycobacterium avium subsp paratuberculosis in potable-water biofilms Appl Environ Microbiol 72 848ndash853
Leistikow RL Morton RA Bartek IL Frimpong I Wagner K Voskuil MI 2010 The Mycobacterium tuberculosis dosR regulon assists in metabolic homeostasis and enables rapid recovery from nonrespiring dormancy J Bacteriol 192 1662ndash1670
Leys NM Ryngaert A Bastiaens L Wattiau P Top EM Verstraete W Springael D 2005 Occurrence and community composition of fast-growing Mycobacterium in soils contaminated with polycyclic aromatic hydrocarbons FEMS Microbiol Ecol 51 375ndash388
Li L Bannantine JP Zhang Q Amonsin A May BJ Alt D Banerji N Kanjilal S Kapur V 2005 The complete genome sequence of Mycobacterium avium subspecies paratuberculosis Proc Natl Acad Sci U S A 102 12344ndash12349
Libero G De Mori L 2014 The T-cell response to lipid antigens of Mycobacterium tuberculosis Front Immunol 5 1ndash9
Lieacutebana E Aranaz A Francis B Cousins D 1996 Assessment of genetic markers for species differentiation within the Mycobacterium tuberculosis complex J Clin Microbiol 34 933ndash938
Lieacutebana E Aranaz A Mateos A Vilafranca M Gomez-Mampaso E Tercero JC Alemany J Suarez G Domingo M Dominguez L 1995 Simple and rapid detection of Mycobacterium tuberculosis complex organisms in bovine tissue samples by PCR J Clin Microbiol 33 33ndash36
Lim A Eleuterio M Hutter B Murugasu-Oei B Dick T 1999 Oxygen depletion-induced dormancy in Mycobacterium bovis BCG J Bacteriol 181 2252ndash2256
Liss GM Wong L Kittle DC Simor A Naus M Martiquet P Misener CR 1994 Occupational exposure to Mycobacterium bovis infection in deer and elk in Ontario Can J Public Heal 85 326ndash329
Little TW Naylor PF Wilesmith JW 1982 Laboratory study of Mycobacterium bovis infection in badgers and calves Vet Rec 111 550ndash557
Lomme JR Thoen CO Himes EM Vinson JW King RE 1976 Mycobacterium tuberculosis infection in two East African oryxes J Am Vet Med Assoc 169 912ndash914
Lotz A Ferroni A Beretti J-L Dauphin B Carbonnelle E Guet-Revillet H Veziris N Heym B Jarlier V Gaillard J-L Pierre-Audigier C Frapy E Berche P Nassif X Bille E 2010 Rapid identification of mycobacterial whole cells in solid and liquid culture media by matrix-assisted laser desorption ionization-time of flight mass spectrometry J Clin Microbiol 48 4481ndash4486
Ludwig W Euzeacuteby J Schumann P Busse H-J Trujillo ME Kaumlmpfer P Whitman WB 2008 Road map
Reacutefeacuterences bibliographiques
207
of the phylum Actinobacteria Road Map of the phylum Actinobacteria Bergeys Manual of Systematic Bacteriology second Edition Volume five Springer Editions
Lugton IW Wilson PR Morris RS Nugent G 1998 Epidemiology and pathogenesis of Mycobacterium bovis infection of red deer (Cervus elaphus) in New Zealand N Z Vet J 46 147ndash156
Lyashchenko KP Greenwald R Esfandiari J OrsquoBrien DJ Schmitt SM Palmer M V Waters WR 2013 Rapid detection of serum antibody by dual-path platform VetTB assay in white-tailed deer infected with Mycobacterium bovis Clin Vaccine Immunol 20 907ndash911
Mackintosh CG Qureshi T Waldrup K Labes RE Dodds KG Griffin JFT 2000 Genetic resistance to experimental infection with Mycobacterium bovis in red deer (Cervus elaphus) Infect Immun 68 1620ndash1625
Maddock EC 1936 Experiments on the infectivity for healthy calves of bovine tubercle bacilli discharged in dung upon pasture Part I From tubercular calves fed with emulsions of Tubercle bacilli 1934-5 Part II From tubercular cows passing Tubercle bacilli in their dun J Hyg (Lond) 36 594ndash601
Maddock EC 1934 Further studies on the survival time of the bovine tubercle bacillus in soil soil and dung in dung and on grass with experiments on feeding guinea-pigs and calves on grass artificially infected with bovine tubercle bacilli J Hyg (Lond) 34 372ndash379
Maddock EC 1933 Studies on the survival time of the bovine tubercle bacillus in soil soil and dung in dung and on grass with experiments on the preliminary treatment of infected organic matter and the cultivation of the organism J Hyg (Lond) 33 103ndash117
Magee JG Ward AC 2012 Mycobacteriaceae Bergeys Manual of Systematic Bacteriology second Edition Volume five Springer Editions
Mahairas GG Sabo PJ Hickey MJ Singh DC Stover CK Mahairas GG Sabo PJ Hickey MJ Singh DC Stover CK 1996 Molecular analysis of genetic differences between Mycobacterium bovis BCG and virulent M bovis J Bacteriol 178 1274ndash1282
Mardare C Delahay RJ Dale JW 2013 Environmental amoebae do not support the long-term survival of virulent mycobacteria J Appl Microbiol 114 1388ndash1394
Marrakchi H Laneacuteelle MA Daffeacute M 2014 Mycolic acids Structures biosynthesis and beyond Chem Biol 21 67ndash85
Marri PR Bannantine JP Golding GB 2006 Comparative genomics of metabolic pathways in Mycobacterium species gene duplication gene decay and lateral gene transfer FEMS Microbiol Rev 30 906ndash925
Martiacuten-Hernando MP Houmlfle U Vicente J Ruiz-Fons F Vidal D Barral M Garrido JM de la Fuente J Gort 2007 Lesions associated with Mycobacterium tuberculosis complex infection in the European wild boar Tuberculosis 87 360ndash367
Mathema B Kurepina NE Bifani PJ Kreiswirth BN 2006 Molecular epidemiology of tuberculosis Current insights Clin Microbiol Rev 19 658ndash685
Matsuoka M Izumi S Budiawan T Nakata N Saeki K 1999 Mycobacterium leprae DNA in daily using water as a possible source of leprosy infection Indian J Lepr 71 61ndash67
Mba Medie F Ben Salah I Henrissat B Raoult D Drancourt M 2011 Mycobacterium tuberculosis complex mycobacteria as amoeba-resistant organisms PLoS One 6 e20499
McCallan L Mcnair J Skuce R 2014 A review of the potential role of cattle slurry in the spread of bovine
Reacutefeacuterences bibliographiques
208
tuberculosis 1ndash48
McCorry T Whelan AO Welsh MD McNair J Walton E Bryson DG Hewinson RG Vordermeier HM Pollock JM 2005 Shedding of Mycobacterium bovis in the nasal mucus of cattle infected experimentally with tuberculosis by the intranasal and intratracheal routes Vet Rec 157 613ndash618
McLaughlin AM Gibbons N Fitzgibbon M Power JT Foley SC Hayes JP Rogers T Keane J 2012 Primary isoniazid resistance in Mycobacterium bovis disease A prospect of concern Am J Respir Crit Care Med 186 110ndash111
McNabb A Eisler D Adie K Amos M Rodrigues M Stephens G Black WA Isaac-Renton J 2004 Assessment of partial sequencing of the 65-kilodalton heat shock protein gene (hsp65) for routine identification of Mycobacterium species isolated from clinical sources J Clin Microbiol 42 3000ndash3011
Mejia GI Castrillon L Trujillo H Robledo J A 1999 Microcolony detection in 7H11 thin layer culture is an alternative for rapid diagnosis of Mycobacterium tuberculosis infection Int J Tuberc Lung Dis 3 138ndash142
Menzies FD Neill SD 2000 Cattle-to-Cattle Transmission of Bovine Tuberculosis Vet J 160 92ndash106
Meredith A Del Pozo J Smith S Milne E Stevenson K McLuckie J 2014 Leprosy in red squirrels in Scotland Vet Rec 175 285ndash286
Michel AL Muumlller B van Helden PD 2010 Mycobacterium bovis at the animal-human interface a problem or not Vet Microbiol 140 371ndash381
Michel AL Venter L Espie IW Coetzee ML 2003 Mycobacterium tuberculosis infections in eight species at the national zoological gardens of South Africa 1991-2001 J Zoo Wildl Med 34 364ndash370
Michelet L de Cruz K Zanella G Aaziz R Bulach T Karoui C Heacutenault S Joncour G Boschiroli ML 2015 Infection with Mycobacterium microti in animals in France J Clin Microbiol 53 981ndash985
Mignard S Pichat C Carret G 2006 Mycobacterium bovis infection Lyon France Emerg Infect Dis 12 1431ndash1433
Miltgen J Morillon M Koeck J-L Varnerot A Briant J-F Nguyen G Verrot D Bonnet D Vincent V 2002 Two cases of pulmonary tuberculosis caused by Mycobacterium tuberculosis subsp canetti Emerg Infect Dis 8 1350ndash1352
Miltner EC Bermudez LE 2000 Mycobacterium avium grown in Acanthamoeba castellanii is protected from the effects of antimicrobials Antimicrob Agents Chemother 44 1990ndash1994
Mitscherlich E Marth EH 2012 Microbial survival in the environment Bacteria and rickettsiae important in human and animal health Springer S ed
Momotani E Ozaki H Hori M Yamamoto S Kuribayashi T Eda S Ikegami M 2012 Mycobacterium avium subsp paratuberculosis lipophilic antigen causes Crohnrsquos disease-type necrotizing colitis in mice Springerplus 1 1ndash10
Moore J Roper T 2003 Temperature and humidity in badger Meles meles setts Mamm Rev 33 308ndash313
Morris RS Pfeiffer DU Jackson R 1994 The epidemiology of Mycobacterium bovis infections Vet Microbiol 40 153ndash177
Mostafa HM Kazda J Irgens LM Luesse HG 1995 Acid-fast bacilli from former leprosy regions in coastal Norway showing PCR positivity for Mycobacterium leprae Int J Lepr other Mycobact Dis 63 97ndash99
Mukhopadhyay S Nair S Ghosh S 2012 Pathogenesis in tuberculosis Transcriptomic approaches to
Reacutefeacuterences bibliographiques
209
unraveling virulence mechanisms and finding new drug targets FEMS Microbiol Rev 36 463ndash485
Muumlller B Duumlrr S Alonso S Hattendorf J Laisse CJM Parsons SDC van Helden PD Zinsstag J 2013 Zoonotic Mycobacterium bovis ndashinduced tuberculosis in humans Emerg Infect Dis 19 899ndash908
Muumlller B Hilty M Berg S Garcia-Pelayo MC Dale J Boschiroli ML Cadmus SI Ngandolo BNR Godreuil S Diguimbaye-Djaibeacute C Kazwala R Bonfoh B Njanpop-Lafourcade BM Sahraoui N Guetarni D Aseffa A Mekonnen MH Razanamparany VR Ramarokoto H Djoslashnne B Oloya J Machado A Mucavele C Skjerve E Portaels F Rigouts L Michel A Muumlller A Kaumlllenius G Van Helden PD Hewinson RG Zinsstag J Gordon S V Smith NH 2009 African 1 an epidemiologically important clonal complex of Mycobacterium bovis dominant in Mali Nigeria Cameroon and Chad J Bacteriol 191 1951ndash1960
Mura M Bull T 2006 Replication and long-term persistence of bovine and human strains of Mycobacterium avium subsp paratuberculosis within Acanthamoeba polyphaga Appl Environ Microbiol 72 854ndash859
Murphy D Gormley E Costello E OrsquoMeara D Corner LAL 2010 The prevalence and distribution of Mycobacterium bovis infection in European badgers (Meles meles) as determined by enhanced post mortem examination and bacteriological culture Res Vet Sci 88 1ndash5
Musser JM Amin A Ramaswamy S 2000 Negligible genetic diversity of Mycobacterium tuberculosis host immune system protein targets Evidence of limited selective pressure Genetics 155 7ndash16
Namouchi A Didelot X Schoumlck U Gicquel B Rocha EPC 2012 After the bottleneck Genome-wide diversification of the Mycobacterium tuberculosis complex by mutation recombination and natural selection Genome Res 22 721ndash734
Narang R Narang P Mendiratta DK 2009 Isolation and identification of nontuberculous mycobacteria from water and soil in central India Indian J Med Microbiol 27 247ndash250
Naranjo V Gortaacutezar C Vicente J de la Fuente J 2008 Evidence of the role of European wild boar as a reservoir of Mycobacterium tuberculosis complex Vet Microbiol 127 1ndash9
Navarro Y Romero B Bouza E Domiacutenguez L Juan L de Garciacutea-de-Viedma D 2015 Detailed chronological analysis of microevolution events in herds infected persistently by Mycobacterium bovis Vet Microbiol 183 97-102
Neill SD Hanna J OrsquoBrien J McCracken R 1988 Excretion of Mycobacterium bovis by experimentally infected cattle Vet Rec 123 340ndash343
Neill SD Pollock JM Bryson DB Hanna J 1994 Pathogenesis of Mycobacterium bovis infection in cattle Vet Microbiol 40 41ndash52
Niemann S Richter E Daluumlgge-Tamm H Schlesinger H Graupner D Koumlnigstein B Gurath G Greinert U Ruumlsch-Gerdes S 2000 Two cases of Mycobacterium microti-derived tuberculosis in HIV-negative immunocompetent patients Emerg Infect Dis 6 539ndash542
Nishi JS Shury T Elkin BT 2006 Wildlife reservoirs for bovine tuberculosis (Mycobacterium bovis) in Canada Strategies for management and research Vet Microbiol 112 325ndash338
Norby B Fosgate GT Manning EJB Collins MT Roussel AJ 2007 Environmental mycobacteria in soil and water on beef ranches Association between presence of cultivable mycobacteria and soil and water physicochemical characteristics Vet Microbiol 124 153ndash159
Nugent G 2011 Maintenance spillover and spillback transmission of bovine tuberculosis in multi-host wildlife complexes a New Zealand case study Vet Microbiol 151 34ndash42
Reacutefeacuterences bibliographiques
210
OrsquoBrien DJ Schmitt SM Fitzgerald SD Berry DE Hickling GJ 2006 Managing the wildlife reservoir of Mycobacterium bovis the Michigan USA experience Vet Microbiol 112 313ndash323
OrsquoReilly LM Daborn CJ 1995 The epidemiology of Mycobacterium bovis infections in animals and man a review Tuber Lung Dis 76 Suppl 1 1ndash46
OIE 2009 Bovine Tuberculosis Terrestrial Manual 2009Chap 247
Ojha AK Baughn AD Sambandan D Hsu T Trivelli X Guerardel Y Alahari A Kremer L Jacobs WR Hatfull GF 2008 Growth of Mycobacterium tuberculosis biofilms containing free mycolic acids and harbouring drug-tolerant bacteria Mol Microbiol 69 164ndash174
Oloya J Kazwala R Lund A Opuda-Asibo J Demelash B Skjerve E Johansen TB Djoslashnne B 2007 Characterisation of mycobacteria isolated from slaughter cattle in pastoral regions of Uganda BMC Microbiol 7 95
Otal I Isabel O Gomez AB Kremer K de Haas P Garciacutea MJ Martiacuten C van Soolingen D 2008 Mapping of IS6110 insertion sites in Mycobacterium bovis isolates in relation to adaptation from the animal to human host Vet Microbiol 129 333ndash341
Pagnier I Raoult D La Scola B 2008 Isolation and identification of amoeba-resisting bacteria from water in human environment by using an Acanthamoeba polyphaga co-culture procedure Environ Microbiol 10 1135ndash1144
Palgrave CJ Benato L Eatwell K Laurenson IF Smith NH 2012 Mycobacterium microti infection in two meerkats (Suricata suricatta) J Comp Pathol 146 278ndash282
Palmer M V 2013 Mycobacterium bovis Characteristics of wildlife reservoir hosts Transbound Emerg Dis 60 1ndash13
Palmer M V C Thacker T Waters WR Robbe-Austerman S Harris BN 2010 Investigations on deer to deer and deer to cattle transmission of the vaccine Mycobacterium bovis bacillus Calmette-Gueacuterin (BCG) J Vaccines Vaccin 01 1ndash5
Palmer M V Waters WR Whipple DL 2004 Investigation of the transmission of Mycobacterium bovis from deer to cattle through indirect contact Am J Vet Res 65 1483ndash1489
Palmer M V Waters WR Whipple DL 2002 Lesion development in white-tailed Deer (Odocoileus virginianus) experimentally infected with Mycobacterium bovis Vet Pathol 39 334ndash340
Palmer M V Whipple DL 2006 Survival of Mycobacterium bovis on feedstuffs commonly used as supplemental feed for white-tailed deer (Odocoileus virginianus) J Wildl Dis 42 853ndash8
Palmer M V Whipple DL Rhyan JC Bolin CA Saari DA 1999 Granuloma development in cattle after intratonsilar inoculation with Mycobacterium bovis Am J Vet Res 60 310ndash315
Palmer M V Whipple DL Waters WR 2001 Experimental deer-to-deer transmission of Mycobacterium bovis Am J Vet Res 62 692ndash696
Panteix G Gutierrez MC Boschiroli ML Rouviere M Plaidy A Pressac D Porcheret H Chyderiotis G Ponsada M Van Oortegem K Salloum S Cabuzel S Bantildeuls AL Van de Perre P Godreuil S 2010 Pulmonary tuberculosis due to Mycobacterium microti a study of six recent cases in France J Med Microbiol 59 984ndash989
Parsons SDC Drewe JA van Pittius NCG Warren RM van Helden PD 2013 Novel cause of tuberculosis in meerkats South Africa Emerg Infect Dis 19 2004ndash2007
Reacutefeacuterences bibliographiques
211
Pavlik I Svastova P Bartl J Dvorska L Rychlik I 2000 Relationship between IS901 in the Mycobacterium avium complex strains isolated from birds animals humans and the environment and virulence for poultry Clin Diagn Lab Immunol 7 212ndash217
Pavlik I Trcka I Parmova I Svobodova J Melicharek I Nagy G Cvetnic Z Ocepek M Pate M Lipiec M 2005 Detection of bovine and human tuberculosis in cattle and other animals in six Central European countries during the years 2000 ndash 2004 Vet Med-Czech 2005 291ndash299
Pawlowski A Jansson M Skoumlld M Rottenberg ME Kaumlllenius G 2012 Tuberculosis and HIV co-infection PLoS Pathog 8(2) e1002464
Payne A 2014 Rocircle de la faune sauvage dans le systegraveme multi-hocirctes de Mycobacterium bovis et risque de transmission entre faune sauvage et bovins Etude expeacuterimentale en Cocircte drsquoOr Thegravese Universiteacute Lyon
Payne A Boschiroli ML Gueneau E Moyen JL Rambaud T Dufour B Gilot-Fromont E Hars J 2012 Bovine tuberculosis in ldquoEurasianrdquo badgers (Meles meles) in France Eur J Wildl Res 59 331ndash339
Payne A Chappa S Hars J Dufour B Gilot-Fromont E 2015 Wildlife visits to farm facilities assessed by camera traps in a bovine tuberculosis-infected area in France Eur J Wildl Res 62(1) 33-42
Pepperell CS Casto AM Kitchen A Granka JM Cornejo OE Holmes EC Holmes EC Birren B Galagan J Feldman MW 2013 The role of selection in shaping diversity of natural M tuberculosis populations PLoS Pathog 9 e1003543
Petroff SA 1915 Some cultural studies on the tubercle bacillus Bull Johns Hopkins Hosp 26 276ndash279
Pfyffer GE Auckenthaler R Van Embden JD a Van Soolingen D 1998 Mycobacterium canettii the smooth variant of M tuberculosis isolated from a Swiss patient exposed in Africa Emerg Infect Dis 4 631ndash634
Phillips CJC Foster CRW Morris PA Teverson R 2003 The transmission of Mycobacterium bovis infection to cattle Res Vet Sci 74 1ndash15
Pickup RW Rhodes G Bull TJ Arnott S Sidi-Boumedine K Hurley M Hermon-Taylor J 2006 Mycobacterium avium subsp paratuberculosis in lake catchments in river water abstracted for domestic use and in effluent from domestic sewage treatment works diverse opportunities for environmental cycling and human exposure Appl Environ Microbiol 72 4067ndash4077
Piersimoni C Olivieri A Benacchio L Scarparo C 2006 Current perspectives on drug susceptibility testing of Mycobacterium tuberculosis complex the automated nonradiometric systems J Clin Microbiol 44 20ndash28
Pillai SD Widmer KW Ivey LJ Coker KC Newman E Lingsweiler S Baca D Kelley M Davis DS Silvy NJ Adams LG 2000 Failure to identify non-bovine reservoirs of Mycobacterium bovis in a region with a history of infected dairy-cattle herds Prev Vet Med 43 53ndash62
Pollock JM Neill SD 2002 Mycobacterium bovis infection and tuberculosis in cattle Vet J 163 115ndash127
Pontiroli A Travis ER Sweeney FP Porter D Gaze WH Mason S Hibberd V Holden J Courtenay O Wellington EMH 2011 Pathogen quantitation in complex matrices a multi-operator comparison of DNA extraction methods with a novel assessment of PCR inhibition PLoS One 6 e17916
Prasad HK Singhal A Mishra A Shah NP Katoch VM Thakral SS Singh D V Chumber S Bal S Aggarwal S Padma M V Kumar S Singh MK Acharya SK 2005 Bovine tuberculosis in India potential basis for zoonosis Tuberculosis (Edinb) 85 421ndash428
Primm TP Lucero CA Falkinham JO 2004 Health impacts of environmental mycobacteria Clin Microbiol Rev 17 98ndash106
Reacutefeacuterences bibliographiques
212
Pritchard DG 1988 A Century of Bovine Tuberculosis 1888-1988 Conquest and Controversy J Comp Pathol 99 357ndash399
Prodinger WM Brandsta A Naumann L Pacciarini M Kubica T Boschiroli ML Aranaz A Cvetnic Z Ocepek M Skrypnyk A Erler W Niemann S Pavlik I Moser I 2005 Characterization of Mycobacterium caprae isolates from Europe by Mycobacterial Interspersed Repetitive Unit Genotyping J Clin Microbiol 43 4984ndash4992
Prodinger WM Eigentler A Allerberger F Schonbauer M Glawischnig W 2002 Infection of Red Deer Cattle and Humans with Mycobacterium bovis subsp caprae in Western Austria J Clin Microbiol 40 2270ndash2272
Prodinger WM Indra A Koksalan OK Kilicaslan Z Richter E 2014 Mycobacterium caprae infection in humans Expert Rev Anti Infect Ther12(12) 1501-1513
Pym AS Brodin P Brosch R Huerre M Cole ST 2002 Loss of RD1 contributed to the attenuation of the live tuberculosis vaccines Mycobacterium bovis BCG and Mycobacterium microti Mol Microbiol 46 709ndash717
Radomski N Betelli L Moilleron R Haenn S Moulin L Cambau E Rocher V Gonccedilalves A Lucas FS 2011 Mycobacterium behavior in wastewater treatment plant a bacterial model distinct from Escherichia coli and Enterococci Environ Sci Technol 45 5380ndash5386
Radomski N Roguet A Lucas FS Veyrier FJ Cambau E Accrombessi H Moilleron R Behr M Moulin L 2013 atpE gene as a new useful specific molecular target to quantify Mycobacterium in environmental samples BMC Microbiol 13 277
Radomski N Thibault VC Karoui C de Cruz K Cochard T Gutierrez C Supply P Biet F Boschiroli ML 2010 Determination of genotypic diversity of Mycobacterium avium subspecies from human and animal origins by Mycobacterial Interspersed Repetitive-Unit-Variable-Number Tandem-Repeat and IS1311 Restriction Fragment Length Polymorphism typing methods J Clin Microbiol 48 1026ndash1034
Rahim Z Moumlllers M Te Koppele-Vije A De Beer J Zaman K Matin MA Kamal M Raquib R Van Soolingen D Baqi M a Heilmann FGC Van Der Zanden AGM 2007 Characterization of Mycobacterium africanum subtype i among cows in a dairy farm in Bangladesh using spoligotyping Southeast Asian J Trop Med Public Health 38 706ndash713
Rauzier J Moniz-Pereira J Gicquel-Sanzey B 1988 Complete nucleotide sequence of pAL5OOO a plasmid from Mycobacterium fortuitum Gene 71 315ndash321
Renwick AR White PCL Bengis RG 2007 Bovine tuberculosis in southern African wildlife a multi-species host-pathogen system Epidemiol Infect 135 529ndash540
Reviriego-Gordejo FJ Vermeersch JP 2006 Towards eradication of bovine tuberculosis in the European Union Vet Microbiol 112 101ndash109
Richomme C Boschiroli ML Hars J Casabianca F Ducrot C 2010 Bovine tuberculosis in livestock and wild boar on the Mediterranean island Corsica J Wildl Dis 46 627ndash631
Ringuet H Akoua-Koffi C Honore S Varnerot A Vincent V Berche P Gaillard JL Pierre-Audigier C 1999 hsp65 sequencing for identification of rapidly growing mycobacteria J Clin Microbiol 37 852ndash857
Ritacco V Loacutepez B De Kantor IN Barrera L Errico F Nader A 1991 Reciprocal cellular and humoral immune responses in bovine tuberculosis Res Vet Sci 50 365ndash367
Rivero A Maacuterquez M Santos J Pinedo A Saacutenchez MA Esteve A Samper S Martiacuten C 2001 High rate of tuberculosis reinfection during a nosocomial outbreak of multidrug-resistant tuberculosis caused by
Reacutefeacuterences bibliographiques
213
Mycobacterium bovis strain B Clin Infect Dis 32 159ndash161
Riviegravere J Fediaevsky A Hars J Richomme C Calavas D Hendrikx P 2011 SYLVATUB Dispositif national de surveillance de la tuberculose bovine dans la faune sauvage Organisation institutionnelle et reacuteseau dacteurs 2011ndash2012
Rodriguez JG Burbano CS Nuntildeez C Gonzaacutelez CE Zambrano MM Garciacutea MJ Del Portillo P 2008 Rv3134cdevRdevS operon of Mycobacterium bovis BCG is differentially transcribed under ldquoin vitrordquo stress conditions Tuberculosis 88 273ndash282
Rodriacuteguez S Bezos J Romero B de Juan L Aacutelvarez J Castellanos E Moya N Lozano F Javed MT Saacuteez-Llorente JL Lieacutebana E Mateos A Domiacutenguez L Aranaz A 2011 Mycobacterium caprae infection in livestock and wildlife Spain Emerg Infect Dis 17 532ndash535
Rodriguez-Campos S Schuumlrch AC Dale J Lohan AJ Cunha M V Botelho A Cruz K Boschiroli ML Boniotti MB Pacciarini M Garcia-Pelayo MC Romero B de Juan L Domiacutenguez L Gordon S V van Soolingen D Loftus B Berg S Hewinson RG Aranaz A Smith NH 2012 European 2 - A clonal complex of Mycobacterium bovis dominant in the Iberian Peninsula Infect Genet Evol 12 866ndash872
Rodriguez-Campos S Smith NH Boniotti MB Aranaz A 2014 Overview and phylogeny of Mycobacterium tuberculosis complex organisms Implications for diagnostics and legislation of bovine tuberculosis Res Vet Sci 97 Suppl S5ndashS19
Rodriguez-Nuntildeez J Avelar FJ Marquez F Rivas-Santiago B Quintildeones C Guerrero-Barrera AL 2012 Mycobacterium tuberculosis complex detected by modified fluorescent in situ hybridization in lymph nodes of clinical samples J Infect Dev Ctries 6(1) 58ndash66
Rodriacuteguez-Zaragoza S 1994 Ecology of free-living Amoebae Crit Rev Microbiol 20(3) 225-241
Rogers LM Forrester GJ Wilson GJ Yarnell RW Cheeseman CL 2003 The role of setts in badger (Meles meles) group size breeding success and status of TB (Mycobacterium bovis) J Zool 260 209ndash215
Romero B Rodriguez S Bezos J Diaz R Francisca Copano M Merediz I Minguez O Marques S Palacios JJ De Viedma DG Saez JL Mateos A Aranaz A Dominguez L De Juan L 2011 Humans as source of Mycobacterium tuberculosis infection in cattle Spain Emerg Infect Dis 17 2393ndash2395
Roring S Scott A Brittain D Walker I Hewinson G Neill SD Skuce R 2002 Development of variable-number tandem repeat typing of Mycobacterium bovis comparison of results with those obtained by using existing exact tandem repeats and spoligotyping J Clin Microbiol 40 2126ndash2133
Roth A Fischer M Hamid ME Michalke S Ludwig W Mauch H 1998 Differentiation of phylogenetically related slowly growing mycobacteria based on 16S-23S rRNA gene internal transcribed spacer sequences J Clin Microbiol 36 139ndash147
Rothschild BM Martin LD Lev G Bercovier H Bar-Gal GK Greenblatt C Donoghue H Spigelman M Brittain D 2001 Mycobacterium tuberculosis complex DNA from an extinct bison dated 17000 years before the present Clin Infect Dis 33 305ndash311
Roug A Geoghegan C Wellington E Miller W A Travis E Porter D Cooper D Clifford DL Mazet J AK Parsons S 2014 Utility of a fecal real-time PCR protocol for detection of Mycobacterium bovis infection in African buffalo (Syncerus caffer) J Wildl Dis 50 140ndash142
Rowe MT Grant IR 2006 Mycobacterium avium ssp paratuberculosis and its potential survival tactics Lett Appl Microbiol 42 305ndash11
Safi H Barnes PF Lakey DL Shams H Samten B Vankayalapati R Howard ST 2004 IS6110 functions as a mobile monocyte-activated promoter in Mycobacterium tuberculosis Mol Microbiol 52 999ndash1012
Reacutefeacuterences bibliographiques
214
Saleeb PG Drake SK Murray PR Zelazny AM 2011 Identification of mycobacteria in solid-culture media by matrix-assisted laser desorption ionization-time of flight mass spectrometry J Clin Microbiol 49 1790ndash1794
Santos N Almeida V Gortaacutezar C Correia-Neves M 2015a Patterns of Mycobacterium tuberculosis-complex excretion and characterization of super-shedders in naturally-infected wild boar and red deer Vet Res 46 129
Santos N Santos C Valente T Gortaacutezar C Almeida V Correia-Neves M 2015b Widespread environmental contamination with Mycobacterium tuberculosis Complex revealed by a molecular detection protocol PLoS One 10 e0142079
Savelkoul PHM Catsburg A Mulder S Oostendorp L Schirm J Wilke H van der Zanden AGM Noordhoek GT 2006 Detection of Mycobacterium tuberculosis complex with Real Time PCR Comparison of different primer-probe sets based on the IS6110 element J Microbiol Methods 66 177ndash180
Scanlon MP Quinn PJ 2000a The survival of Mycobacterium bovis in sterilized cattle slurry and its relevance to the persistence of this pathogen in the environment Ir Vet J 53 412ndash415
Scanlon MP Quinn PJ 2000b Inactivation of Mycobacterium bovis in cattle slurry by five volatile chemicals J Appl Microbiol 89 854ndash861
Schnappinger D Ehrt S Voskuil MI Liu Y Mangan JA Monahan IM Dolganov G Efron B Butcher PD Nathan C Schoolnik GK 2003 Transcriptional adaptation of Mycobacterium tuberculosis within macrophages insights into the phagosomal environment J Exp Med 198 693ndash704
Schoepf K Prodinger WM Glawischnig W Hofer E Revilla-Fernandez S Hofrichter J Fritz J Koumlfer J Schmoll F 2012 A two-yearsrsquo survey on the prevalence of tuberculosis caused by Mycobacterium caprae in red deer (Cervus elaphus) in the Tyrol Austria ISRN Vet Sci 2012 245138
Schroumlder KH 1982 Isolation of M africanum from German patients Bull Int Union Tuberc 57 249ndash53
Schuumlrch AC van Soolingen D 2012 DNA fingerprinting of Mycobacterium tuberculosis from phage typing to whole-genome sequencing Infection Genetics and Evolution 12 602-609
Shepard CC 1960 The experimental disease that follows the injection of human leprosy bacilli into foot-pads of mice J Exp Med 112 445ndash454
Shiloh MU Manzanillo P Cox JS 2008 Mycobacterium tuberculosis Senses Host-Derived Carbon Monoxide during Macrophage Infection Cell Host Microbe 3 323ndash330
Shinnick TM Good RC 1994 Mycobacterial taxonomy Eur J Clin Microbiol Infect Dis 13 884ndash901
Shitikov E Ilina E Chernousova L Borovskaya A Rukin I Afanasrsquoev M Smirnova T Vorobyeva A Larionova E Andreevskaya S Kostrzewa M Govorun V 2012 Mass spectrometry based methods for the discrimination and typing of mycobacteria Infect Genet Evol 12 838ndash845
Shrikrishna D de la Rua-Domenech R Smith NH Colloff A Coutts I 2009 Human and canine pulmonary Mycobacterium bovis infection in the same household re-emergence of an old zoonotic threat Thorax 64 89ndash91
Shuralev E Quinn P Doyle M Duignan A Kwok HF Bezos J Olwill SA Gormley E Aranaz A Good M Davis WC Clarke J Whelan C 2012 Application of the Enfer chemiluminescent multiplex ELISA system for the detection of Mycobacterium bovis infection in goats Vet Microbiol 154 292ndash297
Skuce RA McCorry TP McCarroll JF Roring SMM Scott AN Brittain D Hughes SL Hewinson RG Neill SD 2002 Discrimination of Mycobacterium tuberculosis complex bacteria using novel VNTR-PCR
Reacutefeacuterences bibliographiques
215
targets Microbiology 148 519ndash528
Skuce RA Brittain D Hughes MS Beck LA Neill SD 1994 Genomic fingerprinting of Mycobacterium bovis from cattle by restriction fragment length polymorphism analysis J Clin Microbiol 32 2387ndash2392
Slana I Kaevska M Kralik P Horvathova A Pavlik I 2010 Distribution of Mycobacterium avium subsp avium and M a hominissuis in artificially infected pigs studied by culture and IS901 and IS1245 quantitative real time PCR Vet Microbiol 144 437ndash443
Smith NH 2012 The global distribution and phylogeography of Mycobacterium bovis clonal complexes Infect Genet Evol 12 857ndash865
Smith NH Berg S Dale J Allen AR Rodriguez S Romero B Matos F Ghebremichael S Karoui C Donati C Machado A da C Mucavele C Kazwala RR Hilty M Cadmus SI Ngandolo BNR Habtamu M Oloya J Muller A Milian-Suazo F Andrievskaia O Projahn M Barandiaraacuten S Maciacuteas A Muumlller B Zanini MS Ikuta CY Rodriguez CAR Pinheiro SR Figueroa A Cho S-N Mosavari N Chuang P-C Jou R Zinsstag J van Soolingen D Costello E Aseffa A Proantildeo-Perez F Portaels F Rigouts L Cataldi AA Collins DM Boschiroli ML Hewinson RG Ferreira Neto JS Surujballi O Tadyon K Botelho A Zaacuterraga AM Buller N Skuce R Michel A Aranaz A Gordon S V Jeon B-Y Kaumlllenius G Niemann S Boniotti MB van Helden PD Harris B Zumaacuterraga MJ Kremer K 2011 European 1 a globally important clonal complex of Mycobacterium bovis Infect Genet Evol 11 1340ndash1351
Smith NH Crawshaw T Parry J Birtles RJ 2009 Mycobacterium microti More diverse than previously thought J Clin Microbiol 47 2551ndash2559
Smith NH Kremer K Inwald J Dale J Driscoll JR Gordon S V Van Soolingen D Glyn Hewinson R Maynard Smith J 2006 Ecotypes of the Mycobacterium tuberculosis complex J Theor Biol 239 220ndash225
Smith NH Upton P 2012 Naming spoligotype patterns for the RD9-deleted lineage of the Mycobacterium tuberculosis complex wwwMbovisorg Infect Genet Evol 12 873ndash876
Smith RMM Drobniewski F Gibson A Montague JDE Logan MN Hunt D Hewinson G Salmon RL OrsquoNeill B 2004 Mycobacterium bovis infection United Kingdom Emerg Infect Dis 10 539ndash541
Smithwick RW Stratigos CB David HL 1975 Use of cetylpyridinium chloride and sodium chloride for the decontamination of sputum specimens that are transported to the laboratory for the isolation of Mycobacterium tuberculosis J Clin Microbiol 1 411ndash413
Soini H Musser JM 2001 Molecular diagnosis of mycobacteria Clin Chem 47 809ndash814
Sola C Filliol I Gutierrez MC Mokrousov I Vincent V Rastogi N 2001 Spoligotype database of Mycobacterium tuberculosis biogeographic distribution of shared types and epidemiologic and phylogenetic perspectives Emerg Infect Dis 7 390ndash396
Soo P-C Horng Y-T Hsueh P-R Shen B-J Wang J-Y Tu H-H Wei J-R Hsieh S-C Huang C-C Lai H-C 2006 Direct and simultaneous identification of Mycobacterium tuberculosis complex (MTBC) and Mycobacterium tuberculosis (MTB) by rapid multiplex nested PCR-ICT assay J Microbiol Methods 66 440ndash448
Sreevatsan S Pan X 1997 Restricted structural gene polymorphism in the Mycobacterium tuberculosis complex indicates evolutionarily recent global dissemination ProcNatl Acad Sci USA 94 9869ndash9874
Stahl DA Urbance JW 1990 The division between fast- and slow-growing species corresponds to natural relationships among the mycobacteria J Bacteriol 172 116ndash124
Reacutefeacuterences bibliographiques
216
Steingart KR Ng V Henry M Hopewell PC Ramsay A Cunningham J Urbanczik R Perkins MD Aziz MA Pai M 2006 Sputum processing methods to improve the sensitivity of smear microscopy for tuberculosis a systematic review Lancet Infect Dis 6 664ndash674
Stender H Lund K Petersen KH Rasmussen OF Hongmanee P Miorner H Godtfredsen SE 1999a Fluorescence In Situ Hybridization assay using peptide nucleic acid probes for differentiation between tuberculous and nontuberculous Mycobacterium species in smears of Mycobacterium cultures J Clin Microbiol 37 2760ndash2765
Stender H Mollerup TA Lund K Petersen KH Hongmanee P Godtfredsen SE 1999b Direct detection and identification of Mycobacterium tuberculosis in smear-positive sputum samples by fluorescence in situ hybridization (FISH) using peptide nucleic acid (PNA) probes Int J Tuberc lung Dis 3 830ndash837
Stewart LD McNair J McCallan L Gordon A Grant IR 2013 Improved detection of Mycobacterium bovis infection in bovine lymph node tissue using immunomagnetic separation (IMS)-based methods PLoS One 8 e58374
Stewart LD McNair J McCallan L Thompson S Kulakov LA Grant IR 2012 Production and evaluation of antibodies and phage display-derived peptide ligands for immunomagnetic separation of Mycobacterium bovis J Clin Microbiol 50 1598ndash1605
Stinear TP Mve-Obiang A Small PLC Frigui W Pryor MJ Brosch R Jenkin GA Johnson PDR Davies JK Lee RE Adusumilli S Garnier T Haydock SF Leadlay PF Cole ST 2004 Giant plasmid-encoded polyketide synthases produce the macrolide toxin of Mycobacterium ulcerans Proc Natl Acad Sci U S A 101 1345ndash1349
Sunder S Lanotte P Godreuil S Martin C Boschiroli ML Besnier JM 2009 Human-to-human transmission of tuberculosis caused by Mycobacterium bovis in immunocompetent patients J Clin Microbiol 47 1249ndash1251
Supply P Allix C Lesjean S Cardoso-Oelemann M Rusch-Gerdes S Willery E Savine E de Haas P van Deutekom H Roring S Bifani P Kurepina N Kreiswirth B Sola C Rastogi N Vatin V Gutierrez MC Fauville M Niemann S Skuce R Kremer K Locht C van Soolingen D 2006 Proposal for standardization of optimized Mycobacterial Interspersed Repetitive Unit-Variable-Number Tandem Repeat typing of Mycobacterium tuberculosis J Clin Microbiol 44 4498ndash4510
Supply P Lesjean S Savine E Kremer K Locht C 2001 Automated high-throughput genotyping for study of global epidemiology of Mycobacterium tuberculosis based on Mycobacterial Interspersed Repetitive Units J Clin 39 3563ndash3571
Supply P Marceau M Mangenot S Roche D Rouanet C Khanna V Majlessi L Criscuolo A Tap J Pawlik A Fiette L Orgeur M Fabre M Parmentier C Frigui W Simeone R Boritsch EC Debrie A-S Willery E Walker D Quail M a Ma L Bouchier C Salvignol G Sayes F Cascioferro A Seemann T Barbe V Locht C Gutierrez M-C Leclerc C Bentley SD Stinear TP Brisse S Meacutedigue C Parkhill J Cruveiller S Brosch R 2013 Genomic analysis of smooth tubercle bacilli provides insights into ancestry and pathoadaptation of Mycobacterium tuberculosis Nat Genet 45 172ndash179
Sweeney FP Courtenay O Hibberd V Hewinson RG Reilly LA Gaze WH Wellington EMH 2007 Environmental monitoring of Mycobacterium bovis in badger feces and badger sett soil by real-time PCR as confirmed by immunofluorescence immunocapture and cultivation Appl Environ Microbiol 73 7471ndash7473
Sweeney FP Courtenay O Ul-Hassan A Hibberd V Reilly LA Wellington EMH 2006 Immunomagnetic recovery of Mycobacterium bovis from naturally infected environmental samples Lett Appl Microbiol 43 364ndash369
Reacutefeacuterences bibliographiques
217
Tanner M Michel AL 1999 Investigation of the viability of M bovis under different environmental conditions in the Kruger National Park J Vet Res 185ndash190
Taquet A Tison F 1961 Nouvelle technique drsquoisolement des mycobacteacuteries par le lauryl sulfate de sodium Ann Inst Pasteur 100 676ndash680
Taylor C Jahans K Palmer S Okker M Brown J Steer K 2006 Mycobacterium microti isolated from two pigs Vet Rec 159 59ndash60
Taylor SJ Ahonen LJ de Leij FAAM Dale JW 2003 Infection of Acanthamoeba castellanii with Mycobacterium bovis BCG and survival of M bovis within the amoeba Appl Environ Microbiol 69 4316ndash4319
Telenti A Marchesi F Balz M Bally F Botrger EC Bodmer T 1993 Rapid identification of mycobacteria to the species level by Polymerase Chain Reaction and Restriction Enzyme Analysis J Clin Microbiol 31 175ndash178
Thacker TC Palmer M V Robbe-Austerman S Stuber TP Waters WR 2015 Anatomical distribution of Mycobacterium bovis genotypes in experimentally infected white-tailed deer Vet Microbiol180(1-2) 75-81
Thierry D Brisson-Noeumll A Vincent-Leacutevy-Freacutebault V Nguyen S Guesdon JL Gicquel B 1990a Characterization of a Mycobacterium tuberculosis insertion sequence IS6110 and its application in diagnosis J Clin Microbiol 28 2668ndash2673
Thierry D Cave MD Eisenach KD Crawford JT Bates JH Gicquel B Guesdon JL 1990b IS6110 an IS-like element of Mycobacterium tuberculosis complex Nucleic Acids Res 18 188
Thoen C LoBue P De Kantor I 2006 The importance of Mycobacterium bovis as a zoonosis Vet Microbiol 112 339ndash345
Thomas V Blanc D Bille J Greub G 2006 Biodiversity of amoebae resistant micro-organisms in an hospital water network Appl Environ Microbiol 72 2428ndash2438
Thomas V Loret J-F Jousset M Greub G 2008 Biodiversity of amoebae and amoebae-resisting bacteria in a drinking water treatment plant Environ Microbiol 10 2728ndash2745
Thomas V McDonnell G 2007 Relationship between mycobacteria and amoebae ecological and epidemiological concerns Lett Appl Microbiol 45 349ndash357
Thorel MF 1980 Isolation of Mycobacterium africanum from monkeys Tubercle 61 101ndash104
Tolhurst BA Delahay RJ Walker NJ Ward AI Roper TJ 2009 Behaviour of badgers (Meles meles) in farm buildings Opportunities for the transmission of Mycobacterium bovis to cattle Appl Anim Behav Sci 117 103ndash113
Torgerson PR Torgerson DJ 2010 Public health and bovine tuberculosis whatrsquos all the fuss about Trends Microbiol 18 67ndash72
Tortoli E 2014 Microbiological features and clinical relevance of new species of the genus Mycobacterium Clin Microbiol Rev 27 727ndash752
Tortoli E 2009 Clinical manifestations of nontuberculous mycobacteria Clin Microbiol Infect 15 906ndash910
Tortoli E 2003 Impact of genotypic studies on mycobacterial taxonomy the new mycobacteria of the 1990s Clin Microbiol Rev 16 319ndash354
Reacutefeacuterences bibliographiques
218
Torvinen E Lehtola MJ Martikainen PJ Miettinen IT 2007 Survival of Mycobacterium avium in drinking water biofilms as affected by water flow velocity availability of phosphorus and temperature Appl Environ Microbiol 73 6201ndash6207
Traag BA Driks A Stragier P Bitter W Broussard G Hatfull G Chu F Adams KN Ramakrishnan L Losick R 2010 Do mycobacteria produce endospores Proc Natl Acad Sci 107 878ndash881
Travis ER Gaze WH Pontiroli A Sweeney FP Porter D Mason S Keeling MJC Jones RM Sawyer J Aranaz A Rizaldos EC Cork J Delahay RJ Wilson GJ Hewinson RG Courtenay O Wellington EMH 2011 An inter-laboratory validation of a real time PCR assay to measure host excretion of bacterial pathogens particularly of Mycobacterium bovis PLoS One 6 e27369
Trujillo ME Velaacutezquez E Kroppenstedt RM Schumann P Rivas R Mateos PF Martiacutenez-Molina E 2004 Mycobacterium psychrotolerans sp nov isolated from pond water near a uranium mine Int J Syst Evol Microbiol 54 1459ndash1463
Truman R 2005 Leprosy in wild armadillos Lepr Rev 76 198ndash208
Tsairidou S Woolliams JA Allen AR Skuce RA McBride SH Wright DM Bermingham ML Pong-Wong R Matika O McDowell SWJ Glass EJ Bishop SC 2014 Genomic prediction for tuberculosis resistance in dairy cattle PLoS One 9(5) e96728
Turenne CY 2004 Mycobacterium saskatchewanense sp nov a novel slowly growing scotochromogenic species from human clinical isolates related to Mycobacterium interjectum and Accuprobe-positive for Mycobacterium avium complex Int J Syst Evol Microbiol 54 659ndash667
Van Donsel DJ Larkin EP 1977 Persistence of Mycobacterium bovis BCG in soil and on vegetables spray irrigated with sewage effluent and sludge J Food Prot 40 160ndash163
Van Ingen J Bendien SA de Lange WCM Hoefsloot W Dekhuijzen PNR Boeree MJ van Soolingen D 2009 Clinical relevance of non-tuberculous mycobacteria isolated in the Nijmegen-Arnhem region The Netherlands Thorax 64 502ndash506
Van Ingen J Rahim Z Mulder A Boeree MJ Simeone R Brosch R van Soolingen D 2012 Characterization of Mycobacterium orygis as M tuberculosis complex subspecies Emerg Infect Dis 18 653ndash655
Van Ingen J Van 2013 Diagnosis of nontuberculous mycobacterial infections Semin Respir Crit Care Med 34 103ndash109
Van Pittius NCG Perrett KD Michel AL Keet DF Hlokwe T Streicher EM Warren RM van Helden PD 2012 Infection of African Buffalo (Syncerus Caffer) by Oryx Bacillus A rare member of the antelope clade of the Mycobacterium Tuberculosis Complex J Wildl Dis 48 849ndash857
Van Soolingen D De Haas PEW Haagsma J Eger T Hermans PWM Ritacco V Alito A Van Embden JDA 1994 Use of various genetic markers in differentiation of Mycobacterium bovis strains from animals and humans and for studying epidemiology of bovine tuberculosis J Clin Microbiol 32 2425ndash2433
Van Soolingen D Hermans PWM De Haas PEW Soll DR Van Embden JDA 1991 Occurrence and stability of insertion sequences in Mycobacterium tuberculosis complex strains Evaluation of an insertion sequence-dependent DNA polymorphism as a tool in the epidemiology of tuberculosis J Clin Microbiol 29 2578ndash2586
Vandal OH Pierini LM Schnappinger D Nathan CF Ehrt S 2008 A membrane protein preserves intrabacterial pH in intraphagosomal Mycobacterium tuberculosis Nat Med 14 849ndash854
Varello K Pezzolato M Mascarino D Ingravalle F Caramelli M Bozzetta E 2008 Comparison of
Reacutefeacuterences bibliographiques
219
histologic techniques for the diagnosis of bovine tuberculosis in the framework of eradication programs J Vet Diagnostic Investig 20 164ndash169
Velayati AA Farnia P Mozafari M Malekshahian D Farahbod AM Seif S Rahideh S Mirsaeidi M 2015 Identification and genotyping of Mycobacterium tuberculosis isolated from water and soil samples of a metropolitan city CHEST J 147 1094
Vicente J Houmlfle U Garrido JM Fernaacutedez-De-Mera IG Acevedo P Juste R Barral M Gortaacutezar C 2007 Risk factors associated with the prevalence of tuberculosis-like lesions in fenced wild boar and red deer in south central Spain Vet Res 38 451ndash464
Voskuil MI Schnappinger D Rutherford R Liu Y Schoolnik GK 2004 Regulation of the Mycobacterium tuberculosis PEPPE genes Tuberculosis (Edinb) 84 256ndash262
Voskuil MI Schnappinger D Visconti KC Harrell MI Dolganov GM Sherman DR Schoolnik GK 2003 Inhibition of respiration by nitric oxide induces a Mycobacterium tuberculosis dormancy program J Exp Med 198 705ndash713
Wagner JC Buchanan G Bokkenheuser V Leviseur S 1958 An acid-fast bacillus isolated from the lungs of the Cape Hyrax Procavia capensis (Pallas) Nature 181 284ndash285
Walter WD Smith R Vanderklok M VerCauteren KC 2014 Linking bovine tuberculosis on cattle farms to white-tailed deer and environmental variables using Bayesian hierarchical analysis PLoS One 9 e90925
Wang Y Ogawa M Fukuda K Miyamoto H Taniguchi H 2006 Isolation and identification of mycobacteria from soils at an illegal dumping site and landfills in Japan Microbiol Immunol 50 513ndash524
Ward AI Judge J Delahay RJ 2010 Farm husbandry and badger behaviour Opportunities to manage badger to cattle transmission of Mycobacterium bovis Prev Vet Med 93 2ndash10
Warren RM Gey Van Pittius NC Barnard M Hesseling A Engelke E De Kock M Gutierrez MC Chege GK Victor TC Hoal EG Van Helden PD 2006 Differentiation of Mycobacterium tuberculosis complex by PCR amplification of genomic regions of difference Int J Tuberc Lung Dis 10 818ndash822
Waters WR Buddle BM Vordermeier HM Gormley E Palmer M V Thacker TC Bannantine JP Stabel JR Linscott R Martel E Milian F Foshaug W Lawrence JC 2011 Development and evaluation of an enzyme-linked immunosorbent assay for use in the detection of bovine tuberculosis in cattle Clin Vaccine Immunol 18 1882ndash1888
Wayne LG Hayes LG 1996 An in vitro model for sequential study of shiftdown of Mycobacterium tuberculosis through two stages of nonreplicating persistence Infect Immun 64 2062ndash2069
Wells AQ 1937 Tuberculosis in wild voles Lancet 229 1221
Wells AQ Robb-Smith AHT 1946 Murine type of tubercle bacillus (the vole acid-fast bacillus) Spec Rep Ser Med Res Cou London 259
Whan L Grant IR Rowe MT 2006 Interaction between Mycobacterium avium subsp paratuberculosis and environmental protozoa BMC Microbiol 6 63
Whiley H Keegan A Giglio S Bentham R 2012 Mycobacterium avium complex - the role of potable water in disease transmission J Appl Microbiol 113 223ndash32
White PCL Brown JA Harris S 1993 Badgers (Meles meles) cattle and bovine tuberculosis (Mycobacterium bovis) a hypothesis to explain the influence of habitat on the risk of disease transmission in southwest England Proc R Soc London B 253 277ndash284
Reacutefeacuterences bibliographiques
220
Whittington R Marshall D Nicholls PJ Marsh IB Reddacliff LA 2004 Survival and dormancy of Mycobacterium avium subsp paratuberculosis in the environment Appl Environ Microbiol 70 2989ndash3004
WHO 2014 Global leprosy update 2013 Wkly Epidemiol Rec 89 389ndash400
WHO 2014 Global Tuberculosis Report 2014
Williams RS Hoy WA 1930 The viability of B tuberculosis (bovinus) on pasture land in stored faeces and in liquid manure J Hyg (Lond) 30 413ndash419
Wirth T Hildebrand F Allix-Beacuteguec C Woumllbeling F Kubica T Kremer K Van Soolingen D Ruumlsch-Gerdes S Locht C Brisse S Meyer A Supply P Niemann S 2008 Origin spread and demography of the Mycobacterium tuberculosis complex PLoS Pathog 4(9) e1000160
Witmer G Fine AE Gionfriddo J Pipas M Shively K Piccolo K Burke P 2010 Epizootiologic survey of Mycobacterium bovis in wildlife and farm environments in northern Michigan J Wildl Dis 46 368ndash378
Wobeser G 2009 Bovine tuberculosis in Canadian wildlife an updated history Can Vet J 50 1169ndash1176
Wright DM Allen AR Mallon TR McDowell SWJ Bishop SC Glass EJ Bermingham ML Woolliams JA Skuce RA 2013 Field-isolated genotypes of Mycobacterium bovis vary in virulence and influence case pathology but do not affect outbreak size PLoS One 8(9) e74503
Young JS Gormley E Wellington EMH 2005 Molecular detection of Mycobacterium bovis and Mycobacterium bovis BCG (Pasteur) in soil Appl Environ Microbiol 71 1946ndash1952
Zanella G Durand B Hars J Moutou F Garin-Bastuji B Duvauchelle A Fermeacute M Karoui C Boschiroli ML 2008 Mycobacterium bovis in wildlife in France J Wildl Dis 44 99ndash108
Zelazny AM Calhoun LB L Shea YR Fischer SH 2005 Identification of Mycobacterium species by secA1 sequences J Clin Microbiol 43 1051ndash1058
Zeng W Zhang Y Zhao X Huang G Jiang Y Dong H Li X Wan K He C 2013 Occurrence of non-tuberculous mycobacteria species in livestock from northern China and first isolation of Mycobacterium caprae Epidemiol Infect 141 1545ndash1551
Zhang J Abadia E Refregier G Tafaj S Boschiroli ML Guillard B Andremont A Ruimy R Sola C 2010 Mycobacterium tuberculosis complex CRISPR genotyping Improving efficiency throughput and discriminative power of ldquospoligotypingrdquo with new spacers and a microbead-based hybridization assay J Med Microbiol 59 285ndash294
Zhang Y Yew W 2009 Mechanisms of drug resistance in Mycobacterium tuberculosis Int J Tuberc Lung Dis 13 1320ndash1330
Zolg JW Philippi-Schulz S 1994 The superoxide dismutase gene a target for detection and identification of mycobacteria by PCR J Clin Microbiol 32 2801ndash2812
Zuber B Chami M Houssin C Dubochet J Griffiths G Daffeacute M 2008 Direct visualization of the outer membrane of mycobacteria and corynebacteria in their native state J Bacteriol 190 5672ndash5680
Annexes
223
Annexe 1 Deacutetection de Mycobacterium bovis dans lrsquoenvironnement Premiers
reacutesultats des recherches meneacutees en Cocircte-drsquoOr
Deacutetection de Mycobacterium bovis dans lrsquoenvironnement
Premiers reacutesultats des recherches meneacutees en Cocircte-drsquoOr
Elodie Barbier1 et Alain Hartmann1
RESUME
Depuis les anneacutees 2000 en Cocircte-drsquoOr on assiste agrave une recrudescence des cas de tuberculose agrave
Mycobacterium bovis dans les cheptels bovins associeacutee agrave une contamination de la faune sauvage agrave
proximiteacute des eacutelevages contamineacutes Dans certaines zones du deacutepartement la faune sauvage infecteacutee
serait impliqueacutee dans le retour de la maladie aux bovins La transmission indirecte de la maladie par
lrsquoenvironnement semblerait une voie importante de la contamination entre faune sauvage et
domestique
Afin drsquoeacutevaluer la preacutevalence de M bovis dans lrsquoenvironnement et drsquoeacutevaluer le rocircle de lrsquoenvironnement
dans la transmission indirecte de la maladie nous avons deacuteveloppeacute des outils moleacuteculaires de
deacutetection adapteacutes aux matrices environnementales complexes (sol eau matiegraveres feacutecales) Ces outils agrave
la fois speacutecifiques et sensibles nous ont permis drsquoanalyser de nombreux preacutelegravevements reacutealiseacutes en 2013
Seuls deux eacutechantillons de sol se sont reacuteveacuteleacutes faiblement positifs
Mots-cleacutes tuberculose bovine bovins faune sauvage environnement PCR en temps reacuteel
SUMMARY
Since 2000 an increasing number of cases of bovine tuberculosis were found in cattle in Cocircte-drsquoOr and
in wild animals infected by Mycobacterium bovis in the vicinity of contaminated farms In some areas
wildlife could be considered as the likely source of cattle contamination We assume that
contamination between wild and domestic animals may occur indirectly by environmental factors (soil
water feces)
To quantify the prevalence of M bovis in the environment and to evaluate its role in indirect
transmission of the disease between species we developed molecular detection tools These are
adapted to complex environmental matrices (soil water faces) and are both specific and sensitive
They allowed us to analyze numerous samples collected in 2013 Two samples gave slight positive
results
Keywords Bovine tuberculosis Cattle Wildlife Environment Real time PCR
Texte de la confeacuterence preacutesenteacutee au cours de la Journeacutee scientifique AEEMA-RFSA 18 mars 2014
1 INRA UMR Agroeacutecologie 16 rue de Sully BP 86510 21065 Dijon France
Eacutepideacutemiol et santeacute anim 2014 65 Deacutetection de Mycobacterium bovis dans lrsquoenvironnement Premiers reacutesultats des recherches meneacutees en Cocircte-drsquoOr
224
I - Introduction
Deacuteclareacutee indemne de tuberculose bovine en
2000 la France connait depuis 2002 une
reacuteeacutemergence de lrsquoinfection dans certains
deacutepartements notamment en Cocircte-drsquoOr Entre
2002 et aujourdrsquohui 178 foyers ont eacuteteacute
successivement deacuteclareacutes dans ce deacutepartement
Pour la saison 2012-2013 la moitieacute des
cheptels infecteacutes avaient deacutejagrave eacuteteacute deacuteclareacutes
foyer une deux voire trois fois auparavant La
reacutepartition des cas bovins sur le deacutepartement
nrsquoest pas aleacuteatoire mais concentreacutee sur une
zone assez reacuteduite
Deux spoligotypes principaux sont retrouveacutes
chez les bovins SB0120 (VNTR 5544) appeleacute
plus simplement BCG-like dans la reacutegion de
Veacutenarey-Les-Laumes et Vitteaux et SB0134
(VNTR 5355) appeleacute GB35 dans la reacutegion de
Pouilly-en-Auxois
Parallegravelement agrave lrsquoinfection bovine et agrave
proximiteacute drsquoun cheptel foyer lrsquoinfection a eacuteteacute
deacutetecteacutee pour la premiegravere fois chez un animal
sauvage en 2003 chez un cerf Entre 2003 et
2007 seulement deux sangliers en plus du
cervideacute ont eacuteteacute trouveacutes infecteacutes Depuis 2007
et chaque anneacutee la maladie est reacuteguliegraverement
retrouveacutee chez des blaireaux et des sangliers
moins reacuteguliegraverement chez les cerfs Les
animaux sauvages infecteacutes sont
majoritairement trouveacutes agrave proximiteacute de
cheptels bovins infecteacutes
Lrsquohypothegravese drsquoun cycle eacutepideacutemiologique de
transmission de la maladie entre bovins et
faune sauvage est maintenant reconnue
[Richomme et al 2010] mecircme si celui-ci est
encore mal connu En France les exemples de
la forecirct de Brotonne [Zanella et al 2008] de la
Cocircte-drsquoOr de la Dordogne [Payne et al 2012]
de la Corse [Richomme et al 2010] et ceux de
lrsquoEspagne [Aranaz et al 2004] ont montreacute que
les spoligotypes et les profils VNTR (Variable
Number Tandem Repeat) des souches isoleacutees
chez les animaux sauvages tueacutes ou pieacutegeacutes agrave
proximiteacute des cheptels bovins infecteacutes sont
exactement les mecircmes que ceux des souches
bovines Cette identiteacute des souches reacutesulterait
drsquoune transmission directe ou indirecte entre
faune domestique et sauvage et vice et versa
Bien que quelques contacts directs soient
possibles entre faune sauvage et bovins [Boumlhm
et al 2009] il semble probable que la
transmission indirecte de M bovis soit la voie
de transmission preacutepondeacuterante entre bovin et
faune sauvage et inversement Il semble eacutetabli
que la transmission indirecte de M bovis soit la
principale voie de transmission entre bovins et
faune sauvage et inversement La transmission
indirecte ferait intervenir une excreacutetion de M
bovis par les animaux infecteacutes au niveau
respiratoire salivaire urinaire etou feacutecal les
animaux excreacuteteurs deacuteposeraient donc le
bacille dans lrsquoenvironnement par le biais de
leurs excreacutetats lrsquoenvironnement deviendrait
alors un reacuteservoir potentiel pour le bacille La
contamination drsquoanimaux sains pourrait se
produire par ingestion ou inhalation drsquoaeacuterosols
produits agrave partir des matrices
environnementales souilleacutees Par exemple la
formation de bioaeacuterosols contamineacutes par des
souches bacteacuteriennes drsquoactinomycegravetes
thermotoleacuterants est maintenant bien
documenteacutee agrave partir de matrices solides ou
liquides [Betteli et al 2013] Dans
lrsquoenvironnement des bovins ces bioaeacuterosols
pourraient ecirctre formeacutes agrave partir drsquoeau ou de
meacutelanges eau-seacutediments dans les zones
drsquoabreuvement ou agrave partir de particules fines
de sol (argiles)
Lrsquoexcreacutetion respiratoire salivaire urinaire et
feacutecale de M bovis par des blaireaux
tuberculeux a eacuteteacute documenteacutee en Angleterre
[Gallagher et al 1998 Gavier-Widen et al
2001 Corner et al 2011] De plus
Mycobacterium bovis a la capaciteacute de survivre
en dehors de ses hocirctes pendant des dureacutees
Eacutepideacutemiol et santeacute anim 2014 65 Deacutetection de Mycobacterium bovis dans lrsquoenvironnement Premiers reacutesultats des recherches meneacutees en Cocircte-drsquoOr
225
assez longues Cette dureacutee varie en fonction de
la nature du support contamineacute (eau sol foin
maiumls etc) de la tempeacuterature de lrsquohumiditeacute et
de lrsquoexposition aux UV [Courtenay et al 2006
Fine et al 2011 Ghodbane et al 2014] La
survie de M bovis dans des microcosmes de
sols contamineacutes soumis aux facteurs
climatiques a mecircme eacuteteacute deacutemontreacutee aux Etats-
Unis pour des dureacutees allant jusqursquoagrave environ 88
jours [Fine et al 2011]
La recherche de M bovis par une meacutethode
culturale dans lrsquoenvironnement drsquoanimaux
infecteacutes est souvent infructueuse [Pillai et al
2000 Michel et al 2007 Fine et al 2011] car
elle se heurte agrave plusieurs difficulteacutes i) la vitesse
de croissance tregraves lente de M bovis de lrsquoordre
de 45 jours qui impose un traitement preacutealable
des eacutechantillons de faccedilon agrave deacutetruire les micro-
organismes agrave croissance plus rapide ii) la
richesse des matrices analyseacutees (sol
seacutediments eau matiegraveres feacutecales etc) en
bacteacuteries saprophytes et en champignons tregraves
diversifieacutes qui rendent la deacutecontamination des
eacutechantillons tregraves difficile La deacutecontamination a
toutefois un effet non neacutegligeable sur la
viabiliteacute de M bovis et interfegravere avec la
sensibiliteacute de deacutetection par la mise en culture
[Fine 2006] Lrsquoimmunocapture magneacutetique a
eacuteteacute testeacutee avec succegraves en Angleterre pour isoler
et concentrer le bacille avant la mise en culture
[Sweeney et al 2006 et 2007]
La technique de PCR (Polymerase Chain
Reaction) est une technique indeacutependante de
la croissance de la bacteacuterie Quelques eacutetudes
ont permis la deacutetection par PCR du bacille dans
des sols et des latrines de blaireaux au
Royaume-Uni et en Irlande agrave proximiteacute de
cheptels bovins infecteacutes [Young et al 2005
Courtenay et al 2006 Sweeney et al 2006 et
Les objectifs de ce travail visaient en premier
lieu agrave mettre au point un systegraveme de deacutetection
moleacuteculaire agrave la fois speacutecifique du bacille et
assez sensible pour deacutetecter de faibles
concentrations dans les diffeacuterentes matrices
environnementales suspecteacutees pour leur rocircle
de reacuteservoir potentiel Ces nouveaux outils
moleacuteculaires nous ont permis drsquoanalyser les
eacutechantillons preacuteleveacutes dans des parcelles
partageacutees par les bovins et la faune sauvage
Ces parcelles ont eacuteteacute choisies car une
transmission indirecte de lrsquoinfection entre
animaux sauvages et domestiques eacutetait
suspecteacutee (origine de lrsquoinfection bovine
indeacutetermineacutee suite agrave lrsquoenquecircte
eacutepideacutemiologique et preacutesence drsquoanimaux
sauvages infecteacutes agrave proximiteacute) Cette approche
permettra drsquoeacutevaluer lrsquoimportance de la
transmission indirecte du bacille entre les
espegraveces et drsquoappreacutecier le rocircle joueacute par
lrsquoenvironnement
Cet article ne traite que de la deacutetection
moleacuteculaire la meacutethode fondeacutee sur la culture
eacutetant toujours en cours de validation
II - Mateacuteriel et meacutethodes
Souches bacteacuteriennes utiliseacutees
La souche vaccinale Mycobacterium bovis BCG
strain 1331 (SSI Copenhague Danemark) a eacuteteacute
utiliseacutee pour lrsquoeacutevaluation de la sensibiliteacute de la
PCR en temps reacuteel (qPCR) sur souche pure et
sur microcosmes de sol inoculeacute ainsi que pour
la reacutealisation des gammes eacutetalon de qPCR (PCR
quantitative) Cette souche a eacuteteacute cultiveacutee sur
geacutelose Coletsos pendant 30 jours puis remise
en suspension dans du milieu Middlebrook 7H9
enrichi en ADC (Sigma Aldrich) Apregraves mesure
de la densiteacute optique agrave 600nm la suspension a
eacuteteacute dilueacutee en cascade au 110 pour inoculer les
microcosmes de sol et drsquoeau Un
Eacutepideacutemiol et santeacute anim 2014 65 Deacutetection de Mycobacterium bovis dans lrsquoenvironnement Premiers reacutesultats des recherches meneacutees en Cocircte-drsquoOr
226
deacutenombrement des dilutions est ensuite
reacutealiseacute sur milieu Middlebrook 7H11 enrichi en
OADC (Sigma Aldrich)
Les ADN de 20 spoligotypes de M bovis et de
30 souches de mycobacteacuteries issus drsquoisolats
cliniques drsquoanimaux infecteacutes ont eacuteteacute fournis par
lrsquoANSES Maisons-Alfort pour les tests de
speacutecificiteacute (inclusion - exclusion) de la PCR en
temps reacuteel (tableau 1)
Extraction de lrsquoADN et purification
Lrsquoextraction drsquoADN et la purification des
microcosmes de sol drsquoeau et des eacutechantillons
environnementaux a eacuteteacute reacutealiseacutee selon le
protocole modifieacute deacutecrit par Ranjard et al
[2003]
Les eacutechantillons drsquoeau ont eacuteteacute filtreacutes sur des
membranes drsquoester de cellulose de 045 microm de
porositeacute Les filtres ont eacuteteacute broyeacutes avec de
lrsquoazote liquide puis lrsquoADN extrait selon la
meacutethode preacuteceacutedemment citeacutee agrave lrsquoexception de
la purification sur colonnes PVPP
(polyvinylpolypyrrolidone insoluble)
Les ADN extraits ont eacuteteacute quantifieacutes avec un
spectrophotomegravetre (NanoDropreg 2000 Thermo
Scientific) et dilueacutes au 15 110 et 120 pour la
PCR en temps reacuteel
Deacuteveloppement de trois systegravemes de
deacutetection moleacuteculaire
La bibliographie nous a permis de seacutelectionner
trois cibles geacuteneacutetiques situeacutees sur des locus
diffeacuterents tregraves speacutecifiques de M bovis [Huard
et al 2002 Brodin et al 2002] Les geacutenomes
entiegraverement seacutequenceacutes des mycobacteacuteries du
MTBC (complexe M tuberculosis comprenant
M bovis) et des MNT (mycobacteacuteries non
tuberculeuses) sur le site NCBI nous ont permis
drsquoaligner les seacutequences de ces trois gegravenes agrave la
recherche de zones tregraves conserveacutees dans le
MTBC Chaque systegraveme de deacutetection
moleacuteculaire en PCR temps reacuteel creacuteeacute comprend
une amorce Forward une amorce Reverse et
une sonde oligonucleacuteotidique marqueacutee par
deux fluorochromes (sonde TaqMan) choisies
agrave lrsquoaide du logiciel Primer3 (Simgenecom) Pour
chaque systegraveme nous avons eacutegalement conccedilu
des amorces de PCR agrave lrsquoexteacuterieur des seacutequences
preacuteceacutedentes de faccedilon agrave reacutealiser des PCR
nicheacutees (utiliseacutees uniquement en confirmation
des PCR en temps reacuteel positives)
Speacutecificiteacute et sensibiliteacute des systegravemes de
deacutetection sur souches pures sol et eau
La speacutecificiteacute des trois systegravemes de deacutetection a
eacuteteacute testeacutee par PCR en temps reacuteel sur lrsquoADN de
19 spoligotypes diffeacuterents de M bovis quatre
souches de mycobacteacuteries appartenant au
MTBC deux au Complexe avium et 20 MNT
(tableau 1) Nous avons eacutegalement veacuterifieacute que
la PCR en temps reacuteel ne donnait pas de signal
sur des ADN extraits de 22 sols provenant du
Morbihan des Yvelines du Haut-Rhin et de
Cocircte-drsquoOr preacuteleveacutes en dehors des zones ougrave
seacutevit la tuberculose bovine Ces sols preacutesentent
des caracteacuteristiques physico-chimiques varieacutees
Pour deacuteterminer la sensibiliteacute de deacutetection de
la PCR en temps reacuteel de chacun des systegravemes
la quantification a eacuteteacute reacutealiseacutee sur des dilutions
du gegravene cible cloneacute dans un plasmide (gamme
allant de 1 copie agrave 106 copies du gegravene rechercheacute
dans 5 microl) La limite de deacutetection de la PCR en
temps reacuteel dans le sol a eacuteteacute eacutevalueacutee par
lrsquoinoculation de suspension bacteacuterienne de M
bovis BCG dans un sol dont les caracteacuteristiques
physico-chimiques sont identiques aux sols
incrimineacutes en Cocircte-drsquoOr Chaque microcosme
de sol de 2 g a eacuteteacute inoculeacute avec des suspensions
bacteacuteriennes pour obtenir une concentration
finale en bacteacuteries variant de 10 agrave 105 CFU par
g de sol Des aliquotes de 100 ml drsquoeau non
steacuterile issue drsquoun ruisseau hors zone de
tuberculose ont eacuteteacute inoculeacutes avec ces mecircmes
concentrations
Eacutepideacutemiol et santeacute anim 2014 65 Deacutetection de Mycobacterium bovis dans lrsquoenvironnement Premiers reacutesultats des recherches meneacutees en Cocircte-drsquoOr
227
Tableau 1
Souches bacteacuteriennes et spoligotypes utiliseacutes pour eacutevaluer la speacutecificiteacute des trois systegravemes
de deacutetection moleacuteculaire
M bovis (spoligotypes) Mycobacteacuteries du MTBC Mycobacteacuteries atypiques Mycobacteacuteries du
Complexe avium
GB35 M microti M chitae Mavium
BCG M tuberculosis M elephantis M intracellulare
F1 M caprae M confluentis
F2 M pinnipedi M fortuitum
F5 M rufum
F6 M non chromogenicum
F7 M aichiense
F23 M thermoresitibile
F41 M xenopi
F57 M kansai
F61 M vaccae
F105 M arupense
F151 M chelonae
GB21 M porcinum
GB54 M parafinicum
SB0166 M celatum
SB0885 M intermedium
SB0999 M rutilum
SB1081 M smegmatis
M szulgai
Choix des sites de preacutelegravevements
Les eacutechantillons environnementaux ont eacuteteacute
preacuteleveacutes sur huit communes de Cocircte-drsquoOr
situeacutees dans la zone Nord entre mai et juillet
2013 Nous avons cibleacute avec lrsquoaide de la DDPP21
des parcelles
Sur lesquelles un ou plusieurs bovins
deacutecouverts infecteacutes lors de la prophylaxie
2012-2013 ont pacirctureacute en 2012 et pour
lesquels lrsquoorigine de la contamination reste
inexpliqueacutee
Autour desquelles la surveillance de la
faune sauvage a mis en eacutevidence lrsquoinfection
agrave M bovis chez des blaireaux pieacutegeacutes ou des
sangliers tueacutes agrave la chasse
Sur les pacirctures nous avons privileacutegieacute les zones
susceptibles drsquoecirctre freacutequenteacutees par les bovins
et la faune sauvage zones drsquoabreuvement
(trous drsquoeau sources abreuvoirs) zones de
nourrissage au sol De lrsquoeau du sol et des
seacutediments ont eacuteteacute reacutecolteacutes Autour des
parcelles les terriers et matiegraveres feacutecales de
blaireaux (latrines) ainsi que les zones
drsquoabreuvement de la faune sauvage ont eacuteteacute
preacuteleveacutes
Eacutepideacutemiol et santeacute anim 2014 65 Deacutetection de Mycobacterium bovis dans lrsquoenvironnement Premiers reacutesultats des recherches meneacutees en Cocircte-drsquoOr
228
III - Reacutesultats
Les systegravemes de deacutetection conccedilus sont
speacutecifiques des espegraveces appartenant au MTBC
in silico
Les gegravenes ou locus cibles ont eacuteteacute choisis soit
pour leur speacutecificiteacute agrave M bovis soit pour leur
absence chez M microti seule mycobacteacuterie
du complexe MTBC agrave avoir eacuteteacute deacutecrite dans
lrsquoenvironnement avec M bovis Les systegravemes de
deacutetection ciblant lrsquoIS1561 et la reacutegion Rv3866
ont eacuteteacute choisis car ces locus sont preacutesents chez
tous les membres du MTBC mais absents chez
M microti chez les mycobacteacuteries du complexe
avium ainsi que chez les MNT [Huard et al
2002 Brodin et al 2002] La reacutegion RD4
preacutesente chez tous les membres du MTBC est
absente chez M bovis [Huard et al 2002]
Deux systegravemes de deacutetection moleacuteculaire
ciblant lrsquoIS1561 et la reacutegion Rv3866 ont eacuteteacute
reacutealiseacutes agrave lrsquoaide du logiciel Primer3 Le troisiegraveme
systegraveme ciblant la zone encadrant la RD4 utilise
une amorce Forward et une sonde deacutejagrave deacutecrite
[Sweeney et al 2007] seule lrsquoamorce Reverse
a eacuteteacute redessineacutee afin drsquoobtenir un produit PCR
plus petit La sonde utiliseacutee ne peut se fixer que
sur lrsquoADN des souches preacutesentant la deacuteleacutetion La
speacutecificiteacute des systegravemes de deacutetection a eacuteteacute
deacutemontreacutee in silico par alignement de
seacutequences agrave lrsquoaide du programme BLASTreg
Aucune deacutetection croiseacutee avec drsquoautres
espegraveces que celles appartenant au MTBC nrsquoa
eacuteteacute mise en eacutevidence in silico pour les systegravemes
de deacutetection utiliseacutes
Speacutecificiteacute et sensibiliteacute des systegravemes de
deacutetection sur souches pures et microcosmes
de sol inoculeacutes
Sur les 18 spoligotypes de M bovis testeacutes tous
ont donneacute un signal fort et conserveacute lors de la
PCR en temps reacuteel (Cycle threshold = 20)
Dans le complexe MTBC nous avons testeacute nos
systegravemes sur M microti M tuberculosis M
caprae et M pinnipedii Le systegraveme RD4
preacutesente la plus grande speacutecificiteacute puisqursquoil
nrsquoamplifie que M bovis parmi les membres du
complexe testeacutes LrsquoIS1561 et la reacutegion Rv3866
donnent un signal identique pour M bovis M
caprae M tuberculosis et M pinnipedii mais
ces systegravemes nrsquoamplifient pas M microti
Mecircme srsquoil a eacuteteacute montreacute que M tuberculosis
peut survivre expeacuterimentalement dans le sol
(Ghodbane et al 2014) seuls M microti et M
bovis y ont eacuteteacute deacutejagrave eacuteteacute deacutecrits [Courtenay et
al 2006 Sweeney et al 2006 et 2007]
Sur les 20 souches de MNT testeacutees cinq ont
donneacute un tregraves faible signal (Ct gt 38) avec les
trois systegravemes de deacutetection Pour le complexe
avium seul M avium a eacuteteacute deacutetecteacute avec un
Ctgt38
Aucun des 22 sols testeacutes nrsquoa donneacute de signal en
PCR en temps reacuteel quel que soit le systegraveme de
deacutetection utiliseacute ce qui indique une tregraves bonne
speacutecificiteacute de ces systegravemes de deacutetection sur
une large gamme de sols aux caracteacuteristiques
physico-chimiques diffeacuterentes et contenant des
communauteacutes microbiennes varieacutees
En ce qui concerne la sensibiliteacute des systegravemes
la limite de deacutetection des cibles se situe entre 1
et 10 copies lorsqursquoon utilise des solutions de
plasmide dilueacutees La reacutepeacutetabiliteacute de la
quantification des dilutions de plasmides est
tregraves bonne entre 10 et 106 copies de gegravenes avec
les trois systegravemes En revanche en dessous de
10 copies la deacutetection est moins reacutepeacutetable
(non deacutetection dans certaines reacutepeacutetitions)
Sur les microcosmes de sol et drsquoeau inoculeacutes la
limite de deacutetection des cibles se situe agrave 103
copies du gegravene par g de sol et agrave 10 copies du
gegravene par ml drsquoeau
Le systegraveme ciblant lrsquoIS1561 semble ecirctre tregraves
leacutegegraverement plus sensible que les deux autres
Nous lrsquoavons donc utiliseacute en 1egravere intention pour
Eacutepideacutemiol et santeacute anim 2014 65 Deacutetection de Mycobacterium bovis dans lrsquoenvironnement Premiers reacutesultats des recherches meneacutees en Cocircte-drsquoOr
229
cribler lrsquoensemble des eacutechantillons
environnementaux
Premiers reacutesultats sur les eacutechantillons
environnementaux
Sur les huit communes cibleacutees 150
eacutechantillons de sol 18 eacutechantillons de
seacutediments cinq eacutechantillons drsquoeau et huit
eacutechantillons de fegraveces de blaireaux ont eacuteteacute
preacuteleveacutes Parmi les 150 sols preacuteleveacutes 28 lrsquoont
eacuteteacute dans des terriers de blaireaux et de renards
Parmi les terriers de blaireaux preacuteleveacutes
certains eacutetaient potentiellement des terriers de
blaireaux infecteacutes par M bovis car des
animaux pieacutegeacutes agrave proximiteacute se sont reacuteveacuteleacutes ecirctre
porteurs de M bovis Cependant en 2013 nous
nrsquoavons pu obtenir aucune certitude sur les
terriers reacuteellement occupeacutes par les blaireaux
porteurs de M bovis (pieacutegeage des blaireaux en
couleacutee et non pas en gueule de terrier)
Sur les 181 eacutechantillons analyseacutes seuls deux
eacutechantillons de sol dans deux communes
diffeacuterentes ont donneacute un signal en PCR en
temps reacuteel avec lrsquoIS1561 agrave lrsquoune des dilutions
seulement Ce signal est toutefois faible et nrsquoa
pas pu ecirctre reproduit avec les autres systegravemes
de deacutetection Un des eacutechantillons positifs
provient drsquoun terrier agrave proximiteacute duquel un
blaireau infecteacute a eacuteteacute pieacutegeacute Le second
eacutechantillon de sol a eacuteteacute preacuteleveacute dans une latrine
de blaireau creuseacutee sous une clocircture de preacute
Lrsquoanalyse des fegraveces de cette latrine nrsquoa toutefois
pas donneacute de signal
La campagne de preacutelegravevement 2014 sera
reacutealiseacutee pour augmenter le nombre
drsquoeacutechantillons preacuteleveacutes sur une zone plus
restreinte en particulier ougrave des contaminations
reacutecurrentes des cheptels bovins ont eacuteteacute et sont
observeacutees
IV - Discussion
Mycobacterium bovis lrsquoagent de la tuberculose
bovine est une bacteacuterie pathogegravene qui infecte
les bovins et certains mammifegraveres sauvages en
France La maicirctrise de la maladie chez les
bovins dans certains deacutepartements franccedilais
semble ecirctre entraveacutee par la proximiteacute
drsquoanimaux sauvages infecteacutes La transmission
indirecte du bacille entre faune sauvage et
domestique par lrsquointermeacutediaire de
lrsquoenvironnement permettrait drsquoexpliquer
certains cas de re-contaminations chez les
bovins Lrsquoenvironnement pourrait servir de
reacuteservoir de la bacteacuterie par le biais de
lrsquoexcreacutetion animale il pourrait en retour
infecter des animaux sains soit par inhalation
drsquoaeacuterosols contamineacutes soit par ingestion
(abreuvement) Quelques eacutetudes en
Angleterre et en Irlande ont mis en eacutevidence la
preacutesence de M bovis dans des sols de preacutes des
sols de terriers et des latrines de blaireaux agrave
proximiteacute de cheptels bovins infecteacutes
[Courtenay et al 2006 Sweeney et al 2006
et 2007 Young et al 2005 Travis et al 2011]
Des eacutetudes expeacuterimentales ont montreacute que M
bovis pouvait survivre plusieurs mois dans le sol
[Maddock 1933 Young et al 2005 Fine et al
2011 Ghodbane et al 2014] tout en restant
virulent [Maddock 1933 Ghodbane et al
2014] La deacutetection du bacille dans
lrsquoenvironnement freacutequenteacute par les animaux
domestiques ou sauvages infecteacutes nrsquoa jamais
eacuteteacute deacutecrite en France Dans certaines zones de
Cocircte-drsquoOr la transmission indirecte du bacille
aux bovins par lrsquoenvironnement est fortement
suspecteacutee
Dans cette eacutetude nous avons deacuteveloppeacute trois
systegravemes de deacutetection moleacuteculaire afin
drsquooptimiser la mise en eacutevidence de M bovis
Eacutepideacutemiol et santeacute anim 2014 65 Deacutetection de Mycobacterium bovis dans lrsquoenvironnement Premiers reacutesultats des recherches meneacutees en Cocircte-drsquoOr
230
dans des eacutechantillons environnementaux
complexes afin de srsquoaffranchir de la culture En
effet lrsquoisolement de M bovis dans les matrices
environnementales par la culture est long car
drsquoune part un traitement preacutealable est
neacutecessaire pour srsquoaffranchir de la flore
microbienne de contamination et drsquoautre part
le bacille a une croissance tregraves lente
Contrairement agrave la culture la technique de PCR
en temps reacuteel permet une deacutetection rapide du
bacille sous reacuteserve que le ou les gegravenes cibleacutes
soient tregraves speacutecifiques de faccedilon agrave deacutetecter et
quantifier une espegravece bacteacuterienne particuliegravere
dans un meacutelange drsquoADN complexe Mais cette
technique rencontre neacuteanmoins quelques
limites techniques
Les eacutechantillons environnementaux
contiennent une flore microbienne et animale
abondante et varieacutee lrsquoADN extrait contient une
grande diversiteacute de seacutequences geacuteneacutetiques agrave
lrsquoorigine de faux positifs Ainsi des espegraveces
proches de M bovis notamment les
mycobacteacuteries environnementales ainsi que
des espegraveces encore inconnues peuvent ecirctre
preacutesentes dans les eacutechantillons Il est donc
neacutecessaire de srsquoassurer de la speacutecificiteacute des
systegravemes de deacutetection La deacutetection du bacille
par la recherche associeacutee de trois cibles
geacuteneacutetiques avec le systegraveme TaqMan nous a
permis de deacutevelopper une meacutethode de
deacutetection moleacuteculaire speacutecifique et sensible de
M bovis dans des matrices complexes
Lrsquoextraction drsquoADN de matrices environ-
nementales srsquoaccompagne drsquoune co-extraction
drsquoinhibiteurs potentiels de la PCR (acides
humiques polypheacutenols polysaccharides et
meacutetaux) Lrsquoacide humique par exemple inhibe
lrsquoactiviteacute de la Taq polymeacuterase [Kermekchiev et
al 2009] et se lie sur des sites speacutecifiques de
lrsquoADN empecircchant son amplification [Opel et al
2010] La dilution des eacutechantillons drsquoADN
utiliseacutes en PCR en temps reacuteel permet la dilution
des inhibiteurs et limite leur action Toutefois
lorsque la cible rechercheacutee est preacutesente en
petit nombre de copies la dilution de lrsquoADN
rend la deacutetection moins efficace Crsquoest
pourquoi nous analysons lrsquoADN extrait pur et
dilueacute au 15 au 110 et au 120 en PCR en
temps reacuteel de faccedilon agrave rechercher la quantiteacute
drsquoinhibiteurs minimale permettant drsquoamplifier
la cible
Les limites de deacutetection de la PCR se situent au
mieux autour de 1000 copies de gegravenes par g de
sol Les seuils de deacutetection des trois systegravemes
que nous avons creacuteeacutes se situent au-dessus de
1000 copies du gegravene par g de sol ce qui
correspond agrave des valeurs en adeacutequation avec
drsquoautres reacutesultats obtenus sur la deacutetection de
bacteacuteries dans les sols [Ibekwe et Grieve
2003]
En 2013 seulement deux des eacutechantillons
analyseacutes ont donneacute un signal faible avec un des
systegravemes de deacutetection moleacuteculaire Il srsquoagissait
dans les deux cas drsquoeacutechantillons de sol un sol
de terrier probablement habiteacute par un blaireau
infecteacute et un sol de latrine Nous nrsquoavons pu
reacutepeacuteter les reacutesultats probablement agrave cause de
la faible charge en M bovis des eacutechantillons
analyseacutes en dessous de la limite de deacutetection
de nos outils Il reste difficile drsquoisoler M bovis
de lrsquoenvironnement malgreacute la sensibiliteacute et la
speacutecificiteacute des techniques utiliseacutees Plusieurs
eacutetudes malgreacute lrsquoabondance de preacutelegravevements
reacutealiseacutes ont eacuteteacute infructueuses [Pillai et al
2000 Michel et al 2007 Fine et al 2011]
Cette difficulteacute reacutesulte des limites techniques
preacuteceacutedemment citeacutees mais eacutegalement du
caractegravere variable et intermittent de lrsquoexcreacutetion
de M bovis par les animaux infecteacutes
Lrsquoexcreacutetion de la bacteacuterie par la voie
respiratoire les urines les matiegraveres feacutecales ou
les plaies de morsure nrsquoest pas systeacutematique
chez les blaireaux infecteacutes Le niveau
drsquoexcreacutetion deacutepend de la localisation et de
lrsquointensiteacute des leacutesions Il semble que lrsquoexcreacutetion
la plus importante soit trouveacutee chez des
animaux agrave un stade avanceacute de la maladie
[Gallagher et al 1998 Corner et al 2012] En
Eacutepideacutemiol et santeacute anim 2014 65 Deacutetection de Mycobacterium bovis dans lrsquoenvironnement Premiers reacutesultats des recherches meneacutees en Cocircte-drsquoOr
231
Cocircte-drsquoOr une eacutetude meneacutee entre 2009 et
2011 a montreacute que seuls 17 des blaireaux
pieacutegeacutes et analyseacutes (19 sur 1 146) preacutesentaient
des leacutesions visibles alors que la bacteacuterie eacutetait
retrouveacutee sur 56 des blaireaux (49 sur 878)
dans la zone de tuberculose bovine [Payne et
al 2012] Lrsquoabsence de leacutesion visible sur la
majoriteacute des blaireaux analyseacutes pourrait
expliquer une absence drsquoexcreacutetion ou une
faible excreacutetion de M bovis dans
lrsquoenvironnement
Une autre difficulteacute reacuteside dans la reacutealisation
spatiale et temporelle de lrsquoeacutechantillonnage Les
bacilles sont excreacuteteacutes par les animaux sur de
petites zones bien localiseacutees Hormis les
latrines de blaireaux bien visibles les
preacutelegravevements agrave lrsquoeacutechelle drsquoune parcelle drsquoun
terrier drsquoune source sont reacutealiseacutes de maniegravere
ponctuelle et au hasard ce qui diminue la
probabiliteacute de deacutetection de cette bacteacuterie De
plus nous avons toujours un temps de
deacutecalage entre le deacutepistage des bovins ou des
animaux sauvages infecteacutes et la date des
preacutelegravevements car la certitude du diagnostic
animal neacutecessite plusieurs semaines
V - Conclusion et perspectives
Nous avons deacuteveloppeacute des outils de PCR en
temps reacuteel agrave la fois speacutecifiques et sensibles
pour la deacutetection de M bovis dans les matrices
environnementales Malgreacute le nombre
important drsquoeacutechantillons analyseacutes seuls deux
eacutechantillons peu chargeacutes en M bovis ont pu
ecirctre mis en eacutevidence Ces outils de PCR en
temps reacuteel vont encore ecirctre optimiseacutes en 2014
et permettre lrsquoanalyse des nombreux
preacutelegravevements preacutevus dans des zones plus
cibleacutees de Cocircte-drsquoOr dans lesquelles bovins et
faune sauvage infecteacutes ont eacuteteacute retrouveacutes
Plusieurs meacutethodes de culture sont en cours
drsquoessai et seront utiliseacutees sur les eacutechantillons
preacuteleveacutes afin de montrer la viabiliteacute des
bacteacuteries dans lrsquoenvironnement
Eacutepideacutemiol et santeacute anim 2014 65 Deacutetection de Mycobacterium bovis dans lrsquoenvironnement Premiers reacutesultats des recherches meneacutees en Cocircte-drsquoOr
232
Bibliographie
Aranaz A De Juan L Montero N Saacutenchez C Galka M Delso C Alvarez J Romero B Bezos J Vela A I Briones V Mateos A Domiacutenguez L - Bovine tuberculosis (Mycobacterium bovis) in wildlife in Spain Journal of clinical microbiology 2004 42 2602-2608
Betelli L Duquenne P Grenouillet F Simon X Scherer E Geacutehin E Hartmann A - Development and evaluation of a method for the quantification of airborne Thermoactinomyces vulgaris by real-time PCR Journal of microbiological methods 2013 92 25-32
Boumlhm M Hutchings MR White PCL - Contact networks in a wildlife-livestock host community identifying high-risk individuals in the transmission of bovine TB among badgers and cattle PloS One 2009 4(4) e5016
Brodin P Eiglmeier K Marmiesse M Garnier T Niemann S Cole S T Brosch R Billault A - Bacterial Artificial Chromosome-Based Comparative Genomic Analysis Identifies Mycobacterium microti as a Natural ESAT-6 Deletion Mutant Bacterial Artificial Chromosome-Based Comparative Genomic Analysis Identifies Mycobacterium microti as a Natural ESAT-6 Infection and immunity 2002 70 10 5568-5578
Colinon C Deredjian A Hien E Brothier E Bouziri L Cournoyer B Hartman A Henry S Jolivet C Ranjard L Nazaret S - Detection and enumeration of Pseudomonas aeruginosa in soil and manure assessed by an ecfX qPCR assay Journal of applied microbiology 2013 114 1734-1749
Corner L A L Murphy D Gormley E - Mycobacterium bovis infection in the Eurasian badger (Meles meles) the disease pathogenesis epidemiology and control Journal of comparative pathology 2011 144 1-24
Corner L A L OrsquoMeara D Costello E Lesellier S Gormley E - The distribution of Mycobacterium bovis infection in naturally infected badgers Veterinary journal (London England 1997) 2012 194 166-172
Courtenay O Reilly L A Sweeney F P Hibberd V Bryan S Ul-Hassan A Newman C Macdonald D W Delahay R J Wilson G J Wellington E M H - Is Mycobacterium bovis in the environment important for the persistence of bovine tuberculosis Biology letters 2006 2 460-462
Fine AE - The Role of Indirect Transmission in the Epidemiology of Bovine Tuberculosis in Cattle and White-Tailed Deer in Michigan 2006 Dissertation for the degree of Doctor of Philosophy Michigan State University
Fine A E Bolin C A Gardiner J C Kaneene J B - A Study of the Persistence of Mycobacterium bovis in the Environment under Natural Weather Conditions in Michigan USA Veterinary medicine international 2011 2011 765430
Gallagher J Clifton-Hadley R S - Tuberculosis in badgers a review of the disease and its significance for other animals Research in veterinary science 2000 69 203-217
Gallagher J Monies R Gavier-Widen M Rule B - Role of infected non-diseased badgers in the pathogenesis of tuberculosis in the badger Veterinary Record 1998 142 710-714
Gavier-Widen D Chambers M A Palmer N Newell DG Hewinson R G - Pathology of natural Mycobacterium bovis infection in European badgers (Meles meles) and its relationship with bacterial excretion Veterinary Record 2001 148(10) 299-304
Ghodbane R Mba Medie F Lepidi H Nappez C Drancourt M - Long-term survival of tuberculosis complex mycobacteria in soil Microbiology (Reading England) 2014 160(Pt 3) 496-501
Goodchild A V Clifton-Hadley R S - Cattle-to-cattle transmission of Mycobacterium bovis Tuberculosis (Edinburgh Scotland) 2001 81 23-41
Huard R C Claudio L Lazzarini D O Butler W R Soolingen D Van Ho J L - PCR-Based Method To Differentiate the Subspecies of the Mycobacterium tuberculosis Complex on the Basis of Genomic Deletions Journal of clinical microbiology 2003 41 1637-1650
Ibekwe AM Grieve CM - Detection and quantification of Escherichia coli O157H7 in environmental samples by real-time PCR Journal of Applied Microbiology 2003 94(3) 421-431
Kermekchiev M B Kirilova L I Vail E E Barnes W M - Mutants of Taq DNA polymerase resistant to PCR inhibitors allow DNA amplification from whole blood and crude soil samples Nucleic acids research 2009 37 e40
Maddock EC - Studies on the survival time of the bovine tubercle bacillus in soil soil and dung in dung and on grass with experiments on the
Eacutepideacutemiol et santeacute anim 2014 65 Deacutetection de Mycobacterium bovis dans lrsquoenvironnement Premiers reacutesultats des recherches meneacutees en Cocircte-drsquoOr
233
preliminary treatment of infected organic matter and the cultivation of the organism The Journal of Hygiene 1933 33(1) 103-117
Michel AL de Klerk LM Gey van Pittius NC Warren RM van Helden PD - Bovine tuberculosis in African buffaloes observations regarding Mycobacterium bovis shedding into water and exposure to environmental mycobacteria BMC Veterinary Research 2007 3 23
Neill S Hanna J OrsquoBrien J McCracken R - Excretion of Mycobacterium bovis by experimentally infected cattle Veterinary Record 1988 123 340-343
Opel K L Chung D McCord B R - A study of PCR inhibition mechanisms using real time PCR Journal of forensic sciences 2010 55 25-33
Palmer M V Whipple D L Waters W R Mitchell V Diana L Ray W Transmission E D - Experimental Deer-To-Deer Transmission of Mycobacterium bovis Experimental deer-to-deer transmission of Mycobacterium bovis American journal of veterinary research 2001 62 692-696
Payne A Boschiroli M L Gueneau E Moyen J L Rambaud T Dufour B Gilot-Fromont E Hars J - Bovine tuberculosis in ldquoEurasianrdquo badgers (Meles meles) in France European Journal of Wildlife Research 2012 59 331-339
Pillai S D Widmer K W Ivey L J Coker K C Newman E Lingsweiler S Baca D Kelley M Davis D S Silvy N J Adams L G - Failure to identify non-bovine reservoirs of Mycobacterium bovis in a region with a history of infected dairy-cattle herds Preventive veterinary medicine 2000 43 53-62
Ranjard L Lejon DPH Mougel C Schehrer L Merdinoglu D Chaussod R - Sampling strategy in molecular microbial ecology influence of soil sample size on DNA fingerprinting analysis of fungal and bacterial communities Environmental Microbiology 2003 5(11) 1111-1120
Richomme C Boschiroli M L Hars J Casabianca F Ducrot C - Bovine tuberculosis in livestock and wild boar on the Mediterranean island Corsica Journal of wildlife diseases 2010 46 627-631
Sweeney F P Courtenay O Ul-Hassan A Hibberd V Reilly L A Wellington E M H - Immunomagnetic recovery of Mycobacterium bovis from naturally infected environmental
samples Letters in applied microbiology 2006 43 364-369
Sweeney F P Courtenay O Hibberd V Hewinson R G Reilly L A Gaze W H Wellington E M H - Environmental monitoring of Mycobacterium bovis in badger feces and badger sett soil by real-time PCR as confirmed by immunofluorescence immunocapture and cultivation Applied and environmental microbiology 2007 73 7471-7473
Travis ER Gaze WH Pontiroli A Sweeney FP Porter D Mason S Keeling MJC Jones RM Sawyer J Aranaz A Rizaldos EC Cork J Delahay RJ Wilson GJ Hewinson RG Courtenay O Wellington EMH - An inter-laboratory validation of a real time PCR assay to measure host excretion of bacterial pathogens particularly of Mycobacterium bovis PloS One 2011 6(11) e27369
Walter W D Anderson C W Smith R Vanderklok M Averill J J Vercauteren K C - On-farm mitigation of transmission of tuberculosis from white-tailed deer to cattle literature review and recommendations Veterinary medicine international 2012 2012 616318
Young J S Gormley E amp Wellington E M H - Molecular Detection of Mycobacterium bovis and Mycobacterium bovis BCG ( Pasteur ) in Soil Applied and environmental microbiology 2005 71 1946-1952
Zanella G Durand B Hars J Moutou F Garin-Bastuji B Duvauchelle A Fermeacute M Karoui C Boschiroli M L - Mycobacterium bovis in wildlife in France Journal of wildlife diseases 2008 44 99-108
235
Annexe 2 Preacuteparation du Middlebrook 7H9
Middlebrook 7H9 suppleacutementeacute pour la culture liquide de Mycobacterium bovis
Preacuteparation de la base 7H9 (pour 500 mL de milieu)
1 Diluer 235 g de Middlebrook 7H9 (Fluka analytical) dans 450 mL drsquoeau osmoseacutee 2 Autoclaver 15 min agrave 120degC
Preacuteparation de lrsquoenrichissement ADC (pour 50 mL)
1 Peser 25 g de BSA 1 g de Dextrose et 0005 g de catalase bovine 2 Ajouter 50 mL drsquoeau osmoseacutee 3 Agiter environ 30 min (agitation magneacutetique modeacutereacutee pour eacuteviter la formation de mousse) 4 Filtrer lrsquoenrichissement sur une membrane filtrante de porositeacute 022 microM (Conservation 1 mois agrave 4degC)
Preacuteparation du Middlebrook 7H9 enrichi
1 Apregraves lrsquoautoclavage une fois le milieu refroidi ajouter 50 mL drsquoADC (conservation 1 mois agrave 4degC)
237
Annexe 3 Preacuteparation du Middlebrook 7H11
Middlebrook 7H11 suppleacutementeacute pour la culture de Mycobacterium bovis
Preacuteparation des solutions stock (tous les produits ont eacuteteacute fournis par le laboratoire Sigma-Aldrich)
Vert malachite 06 dissoudre 03 g dans 50 mL drsquoeau osmoseacutee
TicarcillineTrimeacutethoprime peser 1 g ticarcilline et 01 g de trimeacutethoprime lactate ajouter 10 mL drsquoeau osmoseacutee steacuterile Vortexer jusqursquoagrave dissolution reacutepartir en Eppendorf de 1 mL congeler (conservation 6 mois agrave -20degC)
Cycloheximide peser 500 mg et dissoudre dans 10 mL drsquoeacutethanol absolu (conservation 1 an agrave 4degC)
Polymyxine B dilution dans eau osmoseacutee steacuterile pour concentration finale de 100 000 UI par mL
Sang mouton lyseacute Meacutelanger 25 mL de sang avec 25 mL drsquoeau osmoseacutee steacuterile bain-marie 10 min agrave 60degC puis utilisation immeacutediate
Preacuteparation de la base 7H11 (pour 1 L de milieu)
1 Diluer 189 g de Middlebrook 7H11 (Fluka analytical) dans 781 mL drsquoeau 2 Ajouter 1 mL de Vert de malachite 06 3 Ajouter un barreau aimanteacute dans la bouteille 4 Autoclaver 15 min agrave 120degC
Preacuteparation de lrsquoenrichissement OADC (pour 100mL)
1 Peser 5 g de BSA 2 g de Dextrose 085 g de NaCl et 0005g de catalase bovine 2 Ajouter 100 mL drsquoeau osmoseacutee 3 Agiter environ 30 min (agitation magneacutetique modeacutereacutee pour eacuteviter la formation de mousse) 4 Ajouter 125 microL drsquoacide oleacuteique 5 Filtrer lrsquoenrichissement sur une membrane filtrante de porositeacute 022 microM (Conservation 1 mois agrave 4degC)
Preacuteparation du Middlebrook 7H11 enrichi
1 Apregraves lrsquoautoclavage porter le milieu agrave 45-50degC 2 Ajouter 100 mL de seacuterum bovin inactiveacute (agitation magneacutetique) 3 Ajouter 100 mL drsquoOADC (agitation magneacutetique) 4 Ajouter 5 mL de sang de mouton lyseacute 5 Ajouter 1 mL de TicarcillineTrimeacutethoprime 2 mL de Polymyxine B 2 mL de cycloheximide
(agitation magneacutetique) (facultatif si ensemencement de cultures pures)(Concentrations finales respectives 100 mgL 10 mgL 200 000 UIL et 100 mgL)
6 Reacutepartition de 25 mL de milieu par boite (Conservation 1 mois agrave 4degC)
Reacutesumeacute La tuberculose bovine est une maladie infectieuse contagieuse causeacutee par Mycobacterium bovis Cette maladie touche les bovins
et de nombreuses espegraveces de mammifegraveres domestiques et sauvages ainsi que lrsquohomme La circulation de la bacteacuterie dans des systegravemes
multi-hocirctes varieacutes favorise lrsquoentretien de la maladie et la contamination des bovins vivant agrave proximiteacute des animaux sauvages infecteacutes En
marge de la transmission directe de M bovis par voie respiratoire la transmission indirecte aux bovins lieacutee agrave lrsquoinhalation ou agrave lrsquoingestion de
matrices environnementales contamineacutees par un animal infecteacute excreacuteteur est suspecteacutee dans plusieurs reacutegions du monde Lrsquoexistence de
reacuteservoirs environnementaux ougrave le bacille M bovis est capable de persister pourrait donc ecirctre un facteur important de la reacuteeacutemergence puis
du maintien de la maladie dans les systegravemes multi-hocirctes
En Cocircte drsquoOr deacutepartement fortement toucheacute par la tuberculose bovine depuis 2004 la transmission indirecte de la bacteacuterie entre la faune
sauvage infecteacutee et les bovins est suspecteacutee dans plusieurs eacutelevages Pour eacutevaluer la preacutesence et la survie de cette bacteacuterie dans
lrsquoenvironnement nous avons analyseacute un grand nombre drsquoeacutechantillons preacuteleveacutes dans des zones partageacutees par les bovins etou la faune sauvage
infecteacutes dans le but de deacuteterminer la distribution environnementale de M bovis Pour ce faire nous avons deacuteveloppeacute ou modifieacute des systegravemes
de deacutetection moleacuteculaire adapteacutes aux matrices environnementales complexes Nous avons eacutegalement eacutevalueacute lrsquoimpact de la tempeacuterature et
des proprieacuteteacutes physico-chimiques de deux sols sur la survie de M bovis ainsi que le rocircle de la meacutesofaune du sol (lombrics en particulier)
dans la disseacutemination de la bacteacuterie agrave partir de matiegravere organique contamineacutee Lrsquoeacutetude environnementale a mis plus particuliegraverement en
eacutevidence la contamination de deux biotopes les zones humides des pacirctures et les sols de terriers de blaireaux De plus les eacutetudes
expeacuterimentales ont montreacute que M bovis pouvait survivre plusieurs mois dans le sol agrave 4degC et que les lombrics pouvaient disseacuteminer la bacteacuterie
dans le sol voire jouer un rocircle potentiel de vecteur pour les animaux qui les consomment Ces reacutesultats apportent de nouvelles connaissances
sur la persistance et la circulation de M bovis dans lrsquoenvironnement en Cocircte drsquoOr et permettront de proposer des ameacuteliorations aux mesures
de bioseacutecuriteacute deacutejagrave existantes dans les eacutelevages bovins
Mots Cleacutes Mycobacterium bovis environnement sol eau fegraveces bovins faune sauvage qPCR culture
Abstract Bovine tuberculosis is an infectious disease caused by Mycobacterium bovis This disease affects multiple species of domestic
livestock notably cattle wild mammals and humans The circulation of the bacteria within complex multi-host systems promotes the
maintenance of the disease and the reinfection of cattle Besides the classical direct transmission of the bacteria through the respiratory
route indirect transmission through inhalation or ingestion of environmental substrates contaminated by the excretions of infected animals
is suspect in several countries Environmental contamination with M bovis appears to be a crucial factor in the persistence of the infection in
multi-host systems
In Cocircte dOr a French department impacted by bovine tuberculosis since 2004 the indirect transmission of the bacteria from infected wildlife
to cattle is now suspect in several cases To assess this mode of transmission we evaluated environmental contamination with M bovis on a
large number of samples collected in areas shared by cattle and infected wildlife For this purpose we both developed and modified molecular
detection systems adapted for complex environmental substrates We also assessed the impact of temperature and soil physicochemical
properties on the survival of M bovis as well as the role of earthworms in the dissemination of the bacteria from contaminated organic
material The environmental study showed contamination of two habitats in particular wetland pastures and soil badger setts Moreover
results indicated that M bovis can survive in soil for several months at 4degC and that worms may disseminate the bacteria in the soil and even
serve as a vector for the mammals that consume them This study increases our understanding of the persistence and the circulation of
M bovis in the environment in Cocircte dOr In addition these results will be used to improve the existing biosecurity measures on cattle farms
in the region
Key words Mycobacterium bovis environment soil water feces cattle wildlife qPCR culture

Universiteacute de Bourgogne Franche-Comteacute - INRA
Uniteacute Mixte de Recherche 1347 Agroeacutecologie
Ecole Doctorale Environnement - Santeacute
THESE
En vue de lrsquoobtention du grade de Docteur de lrsquoUniversiteacute de Bourgogne Discipline Sciences de la Vie Speacutecialiteacute Ecologie microbienne
Preacutevalence de Mycobacterium bovis dans les agroeacutecosystegravemes
analyse de reacuteservoirs environnementaux potentiels (sol eau douce
faune du sol et faune aquatique) et traccedilage de la circulation de
cette bacteacuterie entre les diffeacuterents compartiments
Preacutesenteacutee et soutenue par
Elodie BARBIER
Le 30 mars 2016
Composition du jury
Alain Hartmann Directeur de Recherche INRA Dijon Directeur de thegravese
Maria Laura Boschiroli Directrice de Recherche ANSES Maisons-Alfort Co-Directrice de thegravese
Franccediloise Lucas Professeur Universiteacute Paris Est Creacuteteil (UPEC) Rapporteur
Franck Biet Directeur de Recherche Tours Nouzilly Rapporteur
Sylvie Nazaret Chargeacutee de Recherche CNRS Universiteacute Claude Bernard Lyon 1 Examinateur
Jaquemine Vialard Maitre de confeacuterence Universiteacute Niort ANSES Niort Examinateur
Pascal Simonnet Directeur de Recherche CNRS Ecole Centrale Lyon Examinateur
Jean-Franccedilois Cavin Professeur Universiteacute de Bourgogne AgroSup Dijon Preacutesident du jury
Expert inviteacute
Eric Gueneau Directeur Laboratoire Deacutepartemental de la Cocircte drsquoOr Dijon Co-encadrant
Remerciements
Mes premiers remerciements vont agrave mes encadrants de thegravese Alain Hartmann et Maria-Laura
Boschiroli Merci de mrsquoavoir confieacute ce sujet de thegravese passionnant et de votre accompagnement vos
conseils pertinents et vos encouragements tout au long de ces trois anneacutees
Un grand merci Eric Gueneau co-encadrant de ce travail et directeur du Laboratoire
Deacutepartemental de Cocircte drsquoOr pour son aide et la mise agrave disposition du P3 pour lrsquoincubation de mes
cultures Je remercie eacutegalement chaleureusement toute son eacutequipe de choc en microbiologie et
biologie moleacuteculaire Veacuteronique Julie Virginie et toute lrsquoeacutequipe
Je remercie Franccediloise Lucas et Franck Biet drsquoavoir accepteacute drsquoecirctre les rapporteurs de ce travail
et drsquoen avoir eacutevalueacute la pertinence Merci eacutegalement agrave Sylvie Nazaret Jaquemine Vialard Pascal
Simonet et Jean-Franccedilois Cavin pour leur participation au jury de thegravese
Merci agrave Barbara Dufour Loiumlc Bollache et Jean-Louis Moyen pour leur participation au comiteacute
de pilotage de thegravese et leurs conseils pendant ce travail
Un eacutenorme merci agrave la DDPP21 pour toutes les informations essentielles fournies sur les foyers
bovins et la faune sauvage Aneacutelise Taconnet Fabienne Bartheacuteleacutemy Thierry Grosjean Un merci
particulier agrave Brigitte Biasino pour son aide preacutecieuse lors drsquoune campagne de preacutelegravevements fort
sympathique
Des remerciements chaleureux eacutegalement agrave Fabrice Chevalier et Marina Beacuteral de la CIREV
Bourgogne pour les donneacutees partageacutees mais eacutegalement les magnifiques cartes illustrant mon
manuscrit
Merci aux organismes qui ont contribueacute financiegraverement agrave ce travail en particulier le GDS 21
la Chambre drsquoAgriculture de la Cocircte drsquoOr La MSA Bourgogne et la MSA nationale Interbev et le
Conseil Deacutepartemental de la Cocircte drsquoOr
Un grand merci agrave Violette Sicre (DDPP16 et Sylvatub) pour son accueil chaleureux et son
efficaciteacute lors de mon seacutejour en Charente Merci eacutegalement au directeur du laboratoire de Charente et
agrave son eacutequipe pour leur accueil
Des remerciements au laboratoire de bacteacuteriologie du CHU de Dijon qui a mis son laboratoire
L3 agrave notre disposition pour lrsquoeacutetude sur la survie de M bovis dans le sol Merci agrave Jean-Marie Duez pour
le temps passeacute et sa confiance merci agrave Catherine Neuwirth Julien Bador et toute lrsquoeacutequipe
Un grand merci agrave lrsquoeacutequipe de lrsquoANSES Maisons-Alfort notamment Krystel De Cruz pour le
geacutenotypage des ADN environnementaux mais eacutegalement Claudine Karaoui Lorraine Michelet et le
reste de lrsquoeacutequipe
Merci agrave Franck Biet et Thierry Cochard de lrsquoINRA de Tours qui nous ont fourni les souches de
Map et M smegmatis
Des eacutenormes remerciements pour les lieutenants de louveterie les pieacutegeurs et les chasseurs
qui mrsquoont accompagneacutee dans mes preacutelegravevements sur le terrain et sans qui je nrsquoaurai pas pu preacutelever
tous ces eacutechantillons Jeannick Gautherot Dominique Rigaud Yannick Thibert Georges Balluet Jean-
Luc Moindrot Marcel Guillier qui connaissent les bois comme leur poche et font un travail
consideacuterable
Je remercie les eacuteleveurs qui ont participeacute au travail en nous accompagnant sur leurs pacirctures
Merci agrave Leacuteon Fayolle pour son expertise sur les lombrics et Dominique Meunier pour son
expertise sur les formations peacutedologiques de lrsquoAuxois merci agrave Luc Biju-Duval pour les cartes reacutealiseacutees
Un eacutenorme merci agrave tous les membres du pocircle MERS qui ont fait de ces trois anneacutees un moment
extrecircmement riche en eacutechanges scientifiques humains amicaux et sportifshellipMerci agrave Dominique
Pascal Laurent Jean-Paul Seacutebastien Un merci particulier aux filles de lrsquoeacutequipe Geacuteraldine Ceacutecile
Laetitia pour leur gentillesse leurs compeacutetences et tous les bons moments passeacutes agrave discuterhellipet agrave
marcher Des remerciements tout particuliers agrave Murielle Rochelet pour ses conseils hyper-aviseacutes sa
rigueur son esprit drsquoinnovation son anglais parfait et encore plein plein drsquoautres choses Merci
infiniment pour tout le temps que tu as passeacute agrave me relire et me corrigerhellip
Un grand mais alors tregraves grand merci agrave Benoicirct sans qui lrsquoexpeacuterience lombric nrsquoaurait pas eacuteteacute
aussihellipexceptionnelle Bon courage pour ta thegravese Merci agrave Cleacutement pour les kilos de sols de terriers
ramasseacutes peseacutes analyseacutes lors de ces deux stages Un grand merci agrave Pierre pour sa gentillesse rare son
travail tregraves seacuterieux et son aide inestimable pour la mise en page la relecture et lrsquoimpression de ce
manuscrit
Merci eacutegalement agrave mes collegravegues du Master AE Caroline Alexandre Fam-Ky Justin Marion
et tous les autres qui mrsquoont parfois (souvent) aideacutee agrave supporter de longues heures de cours
Merci agrave tous ceux que jrsquoai croiseacute agrave lrsquoINRA et ailleurs qui ont fait de ces trois ans une expeacuterience
humaine et scientifique inoubliable Ariane Emilie Kadiya Emeline Ceacuteline Heacutelegravene Charline Anne-
Laure Aude Gilles David Marion Nicolas Romain Fabrice Chloeacute Aymeacute Laure Luiz Marie-
Christine Nadine Estelle Amy Amandinehellip et tous ceux que jrsquoai sucircrement oublieacute
Enfin pour finir un grand merci agrave Steacutephane Lola Lucien et Antonin qui ont supporteacute mes hauts
et mes bas pendant ces trois anneacutees agrave ma famille et ma belle-famille qui ont toujours eacuteteacute lagravehellip
Reacutesumeacute
La tuberculose bovine est une maladie infectieuse contagieuse causeacutee par Mycobacterium
bovis Cette maladie touche les bovins et de nombreuses espegraveces de mammifegraveres domestiques et
sauvages ainsi que lrsquohomme La circulation de la bacteacuterie dans des systegravemes multi-hocirctes varieacutes favorise
lrsquoentretien de la maladie et la contamination des bovins vivant agrave proximiteacute des animaux sauvages
infecteacutes En marge de la transmission directe de M bovis par voie respiratoire la transmission indirecte
aux bovins lieacutee agrave lrsquoinhalation ou agrave lrsquoingestion de matrices environnementales contamineacutees par un
animal infecteacute excreacuteteur est suspecteacutee dans plusieurs reacutegions du monde Lrsquoexistence de reacuteservoirs
environnementaux ougrave le bacille M bovis est capable de persister pourrait donc ecirctre un facteur
important de la reacuteeacutemergence puis du maintien de la maladie dans les systegravemes multi-hocirctes
En Cocircte drsquoOr deacutepartement fortement toucheacute par la tuberculose bovine depuis 2004 la
transmission indirecte de la bacteacuterie entre la faune sauvage infecteacutee et les bovins est suspecteacutee dans
plusieurs eacutelevages Pour eacutevaluer la preacutesence et la survie de cette bacteacuterie dans lrsquoenvironnement nous
avons analyseacute un grand nombre drsquoeacutechantillons preacuteleveacutes dans des zones partageacutees par les bovins etou
la faune sauvage infecteacutes dans le but de deacuteterminer la distribution environnementale de M bovis Pour
ce faire nous avons deacuteveloppeacute ou modifieacute des systegravemes de deacutetection moleacuteculaire adapteacutes aux matrices
environnementales complexes Nous avons eacutegalement eacutevalueacute lrsquoimpact de la tempeacuterature et des
proprieacuteteacutes physico-chimiques de deux sols sur la survie de M bovis ainsi que le rocircle de la meacutesofaune
du sol (lombrics en particulier) dans la disseacutemination de la bacteacuterie agrave partir de matiegravere organique
contamineacutee Lrsquoeacutetude environnementale a mis plus particuliegraverement en eacutevidence la contamination de
deux biotopes les zones humides des pacirctures et les sols de terriers de blaireaux De plus les eacutetudes
expeacuterimentales ont montreacute que M bovis pouvait survivre plusieurs mois dans le sol agrave 4degC et que les
lombrics pouvaient disseacuteminer la bacteacuterie dans le sol voire jouer un rocircle potentiel de vecteur pour les
animaux qui les consomment Ces reacutesultats apportent de nouvelles connaissances sur la persistance et
la circulation de M bovis dans lrsquoenvironnement en Cocircte drsquoOr et permettront de proposer des
ameacuteliorations aux mesures de bioseacutecuriteacute deacutejagrave existantes dans les eacutelevages bovins
Mots cleacutes Mycobacterium bovis environnement sol eau fegraveces bovins faune sauvage qPCR culture
Abstract
Bovine tuberculosis is an infectious disease caused by Mycobacterium bovis This disease
affects cattle and many species of domestic and wild mammals and humans The circulation of the
bacteria in various multi-host systems promotes the maintenance of the disease and the contamination
of cattle in the vicinity Beside direct transmission of the bacteria through the respiratory route indirect
transmission through inhalation or ingestion of environmental matrices contaminated by an infected
animal excretory is suspected in several countries Environmental contamination with M bovis appears
to be a crucial factor in the persistence of the infection in multi-host systems
In Cocircte dOr a French department affected by bovine tuberculosis since 2004 the indirect
transmission of the bacteria from infected wildlife to cattle is suspected in several cases To assess this
type of transmission of the bacillus we evaluated the environmental contamination with M bovis on
a large number of samples taken in areas shared by cattle and or wildlife infected For this purpose
we developed or modified molecular detection systems adapted for environmental complex matrices
We also assessed the impact of physicochemical properties of both soil and temperature on survival of
M bovis and the role of earthworms in the spread of the bacteria from contaminated organic material
The environmental study showed the contamination of two media in particular wetlands pastures and
soil badger setts Moreover experimental studies have shown that M bovis can survive in soil for
several months at 4 deg C and the worms could spread the bacteria in the soil or even play a potential
role for vector animals that consume them These results will propose improvements to existing
biosecurity measures on cattle farms and provide new knowledge about the persistence and circulation
of M bovis in the environment in Cocircte dOr
Key words Mycobacterium bovis environment soil water feces cattle wildlife qPCR culture
Table des matiegraveres
INTRODUCTION GENERALE 1
CHAPITRE 1 - CONTEXTE BIBLIOGRAPHIQUE 4
A LES MYCOBACTERIES DIVERSITE PHYLOGENIE SPECTRE DrsquoHOcircTES ET VIRULENCE 4
1 Le genre Mycobacterium taxonomie et meacutethodes de classification 4
2 Caracteacuteristiques geacuteneacuterales des mycobacteacuteries 5
a Morphologie des mycobacteacuteries 5
b Caracteacuteristiques culturales pheacutenotypiques et physiologiques 7
c Phylogeacutenie du genre Mycobacterium 8
d Caracteacuteristiques geacuteneacutetiques des mycobacteacuteries 9
3 Description des diffeacuterents Complexes ou espegraveces de mycobacteacuteries 10
a Les mycobacteacuteries non pathogegravenes potentiellement opportunistes 11
b Les mycobacteacuteries saprophytes non pathogegravenes 12
c Les mycobacteacuteries pathogegravenes obligatoires 12
Les mycobacteacuteries responsables de legravepre 12
Les mycobacteacuteries tuberculeuses le Complexe tuberculosis (MTBC) 13
B LA TUBERCULOSE UNE MALADIE TRES ANCIENNE MAIS UNE MENACE TOUJOURS ACTUELLE 20
1 Introduction 20
2 Histoire de la tuberculose 20
3 La tuberculose bovine 21
a Introduction 21
b Les hocirctes et les communauteacutes drsquohocirctes de Mycobacterium bovis dans le monde et leurs statuts
eacutepideacutemiologiques 22
c Aspect zoonotique de Mycobacterium bovis 23
d Pathogeacutenie de la tuberculose bovine 24
Facteurs influenccedilant le pouvoir pathogegravene de M bovis voies drsquoinfection et dose infectante 24
Facteurs influenccedilant la reacuteceptiviteacute et la sensibiliteacute de lrsquohocircte 25
Les eacutetapes de lrsquoinfection et la reacuteponse immunitaire associeacutee 26
Expression clinique de la maladie et leacutesions observeacutees 27
e Le deacutepistage et le diagnostic de lrsquoinfection chez les animaux 28
f Epideacutemiologie de la tuberculose bovine 30
Evolution de la tuberculose bovine dans le monde et en Europe et mesures de lutte 30
Evolution de la situation sanitaire en France 33
Evolution de la situation sanitaire en Cocircte drsquoOr 38
C ECOLOGIE DE M BOVIS DANS LrsquoENVIRONNEMENT 42
1 Introduction 42
2 Origines de la contamination environnementale par M bovis 42
a Lrsquoexcreacutetion animale de Mycobacterium bovis 42
b Le transfert de Mycobacterium bovis par les effluents drsquoeacutelevage ou urbains 44
3 Deacutetection et survie de Mycobacterium bovis dans lrsquoenvironnement 45
a Deacutetection de Mycobacterium bovis en conditions naturelles 45
b Dureacutees de survie de Mycobacterium bovis en conditions expeacuterimentales 47
4 Strateacutegies de survie des mycobacteacuteries dans lrsquoenvironnement 54
a Lrsquoeacutetat de dormance 54
b La sporulation 55
c Interactions des mycobacteacuteries avec la faune du sol 56
d Interactions des mycobacteacuteries avec les diptegraveres 57
e La formation de biofilms 57
5 Facteurs de risque de contamination de lrsquoenvironnement et de transmission de Mycobacterium
bovis aux animaux par lrsquoenvironnement contamineacute 58
a Risques lieacutes agrave lrsquoeacutecologie des espegraveces sauvages sensibles agrave Mycobacterium bovis 58
b Risques lieacutes aux pratiques drsquoeacutelevage 60
c Risques lieacutes aux pratiques cyneacutegeacutetiques 61
d Risques lieacutes aux facteurs paysagers climatiques et peacutedologiques 62
D PRINCIPAUX OUTILS DE DETECTION DE PHENOTYPAGE ET DE GENOTYPAGE APPLICABLES AU MTBC ET
PLUS PARTICULIEREMENT A MYCOBACTERIUM BOVIS 64
1 La deacutetection des mycobacteacuteries par microscopie 64
2 Les meacutethodes de culture des mycobacteacuteries 65
a Les diffeacuterents traitements preacutealables agrave la mise en culture des eacutechantillons 65
b La culture sur milieu solide 66
c La culture en milieu liquide 67
3 Meacutethodes de deacutetection et drsquoidentification des bacteacuteries du MTBC 68
a Caracteacuterisation pheacutenotypique et biochimique des mycobacteacuteries du MTBC 69
b Lrsquoidentification des mycobacteacuteries par recherche de biomarqueurs 69
c Lrsquoidentification des mycobacteacuteries par des meacutethodes drsquohybridation coupleacutees ou non agrave la PCR 70
d Le seacutequenccedilage 71
e Deacutetection et geacutenotypage des mycobacteacuteries du MTBC les marqueurs utiliseacutes 72
Les seacutequences drsquoinsertion 72
Les courtes seacutequences drsquoADN reacutepeacuteteacutees 73
Reacutegions de Diffeacuterence (RD) et Single Nucleotide Polymorphism (SNP) 78
f Identification par seacutequenccedilage complet des geacutenomes 79
g Meacutethode speacutecifique drsquoidentification des mycobacteacuteries animales utiliseacutee par le Laboratoire National de
reacutefeacuterence de la tuberculose bovine LNR TB 79
OBJECTIFS DE LA THESE 82
CHAPITRE 2 - DEVELOPPEMENT ET OPTIMISATION DrsquoOUTILS POUR LA DETECTION DE MYCOBACTERIUM
BOVIS DANS LES ECHANTILLONS ENVIRONNEMENTAUX 86
A DEVELOPPEMENT DrsquoOUTILS MOLECULAIRES ADAPTES AUX MATRICES ENVIRONNEMENTALES 86
CHAPITRE 3 - ETUDE DE LA PREVALENCE ENVIRONNEMENTALE DE MYCOBACTERIUM BOVIS DANS LA ZONE
ENDEMIQUE DE TUBERCULOSE BOVINE DE COTE DrsquoOR ET RESULTATS OBTENUS EN CHARENTE 93
A AVANT-PROPOS 93
B FIRST MOLECULAR DETECTION OF MYCOBACTERIUM BOVIS IN ENVIRONMENTAL SAMPLES FROM A
FRENCH REGION WITH ENDEMIC BOVINE TUBERCULOSIS 95
C CAMPAGNES DE PRELEVEMENTS SUPPLEMENTAIRES 121
1 Fin des preacutelegravevements en Cocircte drsquoOr 121
2 Preacutelegravevements en Charente 123
3 Compleacutements paysage geacuteomorphologie et peacutedologie 125
D BILAN GLOBAL 130
CHAPITRE 4 - IMPACT DE LA TEMPERATURE ET DE LA NATURE DU SOL SUR LA SURVIE DE MYCOBACTERIUM
BOVIS SB0120 133
A AVANT-PROPOS 133
B EXPERIMENTAL INVESTIGATION OF THE IMPACT OF THE TEMPERATURE AND THE SOIL TYPE ON
MYCOBACTERIUM BOVIS SURVIVAL IN THE ENVIRONMENT 135
C BILAN 150
CHAPITRE 5 - ROLE DE LA FAUNE DU SOL DANS LA CIRCULATION DE MYCOBACTERIUM BOVIS ETUDE DE SA
DISSEMINATION PAR LES LOMBRICS 155
A AVANT-PROPOS 155
B RAPID DISSEMINATION OF M BOVIS FROM CATTLE DUNG TO SOIL BY THE EARTHWORM LUMBRICUS
TERRESTRIS 157
C COMPLEMENT PRELEVEMENT ET ANALYSE DE LOMBRICS DANS UNE PATURE SUSPECTE 174
D BILAN 175
DISCUSSION GENERALE 177
1 Des meacutethodes deacuteveloppeacutees (ou adapteacutees) puis valideacutees dans des matrices environnementales 179
2 Contamination environnementale les zones humides les eaux de ruissellement et les sols de
terriers sont potentiellement agrave risque 180
3 Survie et circulation de M bovis dans lrsquoenvironnement 183
PERSPECTIVES 185
REFERENCES BIBLIOGRAPHIQUES 191
ANNEXES 221
Liste des Figures
Figure 1 Structure et composition de lenveloppe des mycobacteacuteries (drsquoapregraves Marrakchi Laneacuteelle and Daffeacute 2014) AG arabinogalactane PG peptidoglycane TDM treacutehalose dimycolate PL phospholipides TMM treacutehalose monomycolate GL glycolipides 7
Figure 2 Arbre phylogeacutenique baseacute sur lrsquoanalyse de lrsquoARNr 16S des principaux complexes et espegraveces de mycobacteacuteries (drsquoapregraves Magee et Ward 2012) 9
Figure 3 Phylogeacutenie des souches du MTBC baseacutee sur les Reacutegions de Diffeacuterences et diffeacuterents Single Nucleotide Polymorphism (Dippenaar et al 2015) 14
Figure 4 Aspect des colonies de M bovis sur milieu de Lowenstein-Jensen (agrave gauche) et Middlebrook 7H11 suppleacutementeacute (agrave droite) 17
Figure 5 Evolution de la reacuteponse immunitaire chez le bovin infecteacute par M bovis au cours du temps et aux diffeacuterents tests de deacutepistage de la tuberculose bovine (de la Rua-Domenech et al 2006 adapteacute de Vordermeier et al 2004) 27
Figure 6 Carte de distribution de la tuberculose bovine chez les bovins entre juillet et deacutecembre 2014 (Source WAHID Interface) 31
Figure 7 Statuts des pays europeacuteens vis-agrave-vis de la tuberculose bovine en 2012 et proportion de bovins infecteacutes OTF Officially Tuberculosis Free (officiellement indemne de tuberculose bovine) MS Member State (Etat membre) (Source EFSA 2014) 32
Figure 8 Evolution du taux de preacutevalence annuelle national de la tuberculose bovine chez les bovins (Beacutenet et al 2006) 34
Figure 9 Evolution de la preacutevalence et de lrsquoincidence de la tuberculose bovine en France entre 1995 et 2003 (Cavalerie et al 2014) 35
Figure 10 Incidence et distribution de la tuberculose bovine en France dans les anneacutees 50 (en haut) et entre 2000 et 2013 (en bas) (Cavalerie et al 2014) 36
Figure 11 Reacutepartition et reacutesultats des analyses effectueacutees dans le cadre du dispositif Sylvatub chez le grand gibier drsquoaoucirct 2013 agrave aoucirct 2014 et chez les blaireaux en 2014 (Source Sylvatub) 38
Figure 12 Localisation du parcellaire des cheptels infecteacutes par la tuberculose bovine entre 2010 et 2015 et spoligotypes incrimineacutes (Source CIREV) 40
Figure 13 Nombre de nouveaux foyers bovins et de foyers recontamineacutes entre 2002 et 2014 (Donneacutees DDPP21) 40
Figure 14 Nombre de cervideacutes sangliers et blaireaux infecteacutes deacutecouverts en Cocircte drsquoOr entre 2002 et 2014 41
Figure 15 Repreacutesentation scheacutematique de la circulation de M bovis dans lrsquoenvironnement agricole (modifieacute drsquoapregraves Elliott et al 2014) 63
Figure 16 Profils de restriction disolats de M tuberculosis obtenus suite agrave une RFLP-IS6110 Les isolats 3 5 6 9 et 10 ont le mecircme profil et posseacutedaient un lien eacutepideacutemiologique S repreacutesente le marqueur de poids moleacuteculaire (Source web site CDC) 73
Figure 17 Principe du spoligotypage des bacteacuteries du MTBC dapregraves Kamerbeek et al (1997) A Repreacutesentation du locus DR de M tuberculosis H37Rv et M bovis BCG B Amplifications des espaceurs par un couple damorces (a et b) et amplicons obtenus C Profils drsquohybridation sur membrane (un carreacute noir signale la preacutesence de lrsquoespaceur les parties blanches leur absence) 75
Figure 18 Diffeacuterences geacutenomiques de la reacutegion RD1 entre M bovis BCG (A) M microti (C) par comparaison agrave M tuberculosis H37Rv (B) drsquoapregraves Brodin et al 2002 et localisation du locus Rv3866 RD1mic correspond agrave la seacutequence de M tuberculosis deacuteleacuteteacutee chez M microti RD1bcg correspond agrave la seacutequence de M tuberculosis deacuteleacuteteacutee chez M bovis BCG 88
Figure 27 Localisation des communes de Cocircte drsquoOr sur lesquelles de foyers bovins et des animaux sauvages infecteacutes ont eacuteteacute deacutetecteacutes et localisation des preacutelegravevements environnementaux effectueacutes en 2015 (les preacutelegravevements positifs sont repreacutesenteacutes par des eacutetoiles noires) (Reacutealisation Marina Beacuteral CIREV Bourgogne) 123
Figure 28 Localisation des communes de Charente sur lesquelles de foyers bovins et des animaux sauvages infecteacutes ont eacuteteacute deacutetecteacutes et localisation des preacutelegravevements environnementaux effectueacutes en 2015 (les preacutelegravevements positifs sont repreacutesenteacutes par des eacutetoiles noires) (Reacutealisation Marina Beacuteral CIREV Bourgogne) 125
Figure 29 Orographie et hydrographie de lrsquoAuxois Les valleacutees supportant les parcelles suspectes sont indiqueacutees par une flegraveche noire (Source BD Carthage et BD alti - IGN) 127
Figure 30 Localisation de lrsquoAuxois en Cocircte drsquoOr structure paysagegravere et geacuteomorphologie de ce territoire 127
Figure 31 Nombre de vaches allaitantes par canton dans le deacutepartement de la Cocircte drsquoOr (agrave gauche) et parcellaire des foyers bovins de 2010 agrave 2015 (agrave droite) (Source Recensement agricole 2010 ndash Agreste) 128
Figure 32 Reacutepartition spatiale des parcelles des eacutelevages bovins infecteacutes de 2010 agrave 2014 en Cocircte drsquoOr et repreacutesentation des couches peacutedologiques (Reacutealisation Marina Beacuteral CIREV Bourgogne) 129
Figure 33 Tempeacuteratures minimales et maximales observeacutees agrave Dijon-Longvic de 2012 agrave 2015 (Source Meacuteteacuteo France) 151
Liste des Tableaux
Tableau 1 Dureacutees de survie de M bovis dans diffeacuterentes matrices environnementales obtenues en conditions expeacuterimentales naturelles ou de laboratoire 49
Tableau 2 Identification des principales mycobacteacuteries du MTBC par des observations pheacutenotypiques et des tests biochimiques 69
Tableau 3 Cibles moleacuteculaires deacutecrites dans la litteacuterature pour la deacutetection du Complexe tuberculosis ou de M bovis dans les eacutechantillons environnementaux 76
Tableau 4 Spoligotypes et geacutenotypes MIRU-VNTR des souches de M bovis isoleacutees chez les bovins et la faune sauvage en Cocircte dOr et en Dordogne (drsquoapregraves Hauer et al 2015) 77
Tableau 5 Systegravemes de deacutetection utiliseacutes en qPCR en premiegravere intention pour lidentification dune mycobacteacuterie dorigine animale et reacutesultats obtenus 80
Liste des Annexes
Annexe 1 Deacutetection de Mycobacterium bovis dans lrsquoenvironnement Premiers reacutesultats des recherches meneacutees en Cocircte-drsquoOr 223
Annexe 2 Preacuteparation du Middlebrook 7H9 235
Annexe 3 Preacuteparation du Middlebrook 7H11 237
Liste des Abreacuteviations
ADC Albumine Dextrose Catalase
ADNDNA Acide deacutesoxyribonucleacuteique
AEEMA Association pour lEtude de lEpideacutemiologie des Maladies Animales
AFNOR Agence Franccedilaise de NORmalisation
ANSES Agence nationale de seacutecuriteacute sanitaire de lrsquoalimentation de lrsquoenvironnement et du travail
ARNRNA Acide ribonucleacuteique
ARNr ARN ribosomique
BAAR Bacille acido-alcoolo reacutesistant
BCG Bacille de Calmette et Gueacuterin
BLAST Basic Local Alignment Search Tool
bTB bovine Tuberculosis
CDC Centers of Disease Control and Prevention
CFP-10 Culture Filtrat Protein 10 kD
CIREV Cellules Inter Reacutegionales dEpideacutemiologie Veacuteteacuterinaire
CRISPR Clustered Regularly Interspersed Short Palindromic Repeats
Ct Cycle Threshold
DDPP Direction Deacutepartementale de la Protection des Populations
DEFRA Department for Environment Food and Rural Affairs
DGAL Direction Geacuteneacuterale de lALimentation
DosR Dormancy Survival Regulator
DR Direct Repeat
EFSA European Food Safety Authority
ELISA Enzyme-Linked ImmunoSorbent Assay
ESAT-6 Early Secreted Antigenic Target 6kD
ETR Exact Tandem Repeat
Fbp Fibronectin binding protein
FISH Fluorescence In Situ Hybridization
GC-MS Gas Chromatography-Mass Spectrometry
GDS Groupements de Deacutefense Sanitaire
HAP Hydrocarbures Aromatiques Polycycliques
HIV Human Immunodeficiency Virus
HPLC High Performance Liquid Chromatography
IDC Intradermotuberculination comparative
IDR Intradermotuberculination
IDS Intradermotuberculination simple
IFN-γ Interfeacuteron γ
INRA Institut National de la Recherche Agronomique
Ion PGM Ion Personal Genome Machine
IS Insertion Sequence
ITS Internal Transcribed Spacer
kDkDa kilo Daltons
LNR Laboratoire National de Reacutefeacuterence
LSP Large Sequence Polymorphism
Maa Mycobacterium avium subspecies avium
MAC Mycobacterium avium complex
MALDI-TOF Matrix-Assisted Laser Desorption Ionization-Time Of Flight
Map Mycobacterium avium subspecies paratuberculosis
MDR Multi Drug Resistant
MIRU Mycobacterial Interspersed Repetitive Unit
MLVA Multi Locus VNTR Analysis
MNT Mycobacteacuteries Non Tuberculeuses
MS Member State
MTBC Mycobacterium tuberculosis complex
NCBI National Center for Biotechnology Information
NF Norme Franccedilaise
NGS Next Generation Sequencing
OADC Oleic acid Albumine Dextrose Catalase
OIE Office International des Epizooties
OMS Organisation Mondiale de la Santeacute
ONCFS Office National de la Chasse et de la Faune Sauvage
ONEMA Office National de lEau et des Milieux Aquatiques
OTF Officialy Tuberculosis Free
pb paire de bases
PCR Polymerase chain reaction
PNA Peptide Nucleic Acid
qPCR quantitative Polymerase Chain Reaction
RD Reacutegions de Diffeacuterence
RFLP Restriction Fragment Lenght Polymorphism
RNS Reactive Nitrogen Species
ROS Reactive Oxygen Species
RT-PCR Reverse-Transcription Polymerase Chain Reaction
SNP Single Nucleotide Polymorphism
ssp subspecies
TDM Treacutehalose dimycolate
TMM Treacutehalose monomycolate
UFCCFU Uniteacute Formant Colonie
VBNC Viable But Non Cultivable
VNTR Variable Number Tandem Repeat
WAHID World Animal Health Information Database
WGS Whole Genome Sequencing
WHO World Health Organization
XDR eXtensively Drug Resistant
1
Introduction geacuteneacuterale
La tuberculose bovine est une maladie drsquoimportance sanitaire majeure en santeacute animale agrave
travers le monde Son agent principal Mycobacterium bovis est un membre du Complexe
Mycobacterium tuberculosis qui regroupe les bacteacuteries tuberculeuses infectant lrsquohomme et les
mammifegraveres Les bovins constituent le reacuteservoir originel de M bovis mais de nombreuses espegraveces de
mammifegraveres peuvent eacutegalement ecirctre infecteacutees dont lrsquohomme Les enjeux sanitaires eacuteconomiques et
sociaux induits par cette maladie ont conduit agrave la mise en place de programmes visant lrsquoeacuteradication de
cette maladie chez les bovins agrave travers le monde Dans les pays deacuteveloppeacutes ces mesures associeacutees agrave
la pasteurisation du lait ont fortement reacuteduit lrsquoincidence de la tuberculose bovine voire eacuteradiqueacute
lrsquoinfection chez les bovins Toutefois dans quelques pays lrsquoinfection persiste dans les cheptels bovins
malgreacute des actions de preacutevention de controcircle et de surveillance Dans les pays en deacuteveloppement ougrave
les controcircles organiseacutes sont quasi- voire totalement absents la preacutevalence et la propagation de la
tuberculose bovine sont inquieacutetantes et repreacutesentent un risque sanitaire et eacuteconomique vis-agrave-vis des
populations sensibles En conseacutequence la tuberculose bovine attire chaque anneacutee un peu plus
lrsquoattention de la communauteacute internationale en raison du nombre croissant de troupeaux infecteacutes et
des effets sur les productions animales ainsi que du risque que la maladie fait planer sur la santeacute
publique
Le passage de lrsquoinfection du reacuteservoir bovin agrave diffeacuterentes populations sauvages qui
entretiennent la maladie et peuvent la transmettre en retour aux bovins a fortement compliqueacute les
scheacutemas drsquoeacuteradication de la maladie dans plusieurs reacutegions du monde et explique lrsquoeacutechec partiel des
programmes de lutte mis en place Alors que la transmission directe du bacille par voie respiratoire
semble ecirctre le mode de transmission preacutepondeacuterant au sein drsquoune mecircme espegravece la transmission
indirecte de M bovis est fortement suspecteacutee entre espegraveces partageant les mecircmes territoires et
ressources compliquant encore lrsquoeacutepideacutemiologie de cette maladie
En France une recrudescence du nombre de cheptels bovins infecteacutes dans certains
deacutepartements dont la Cocircte drsquoOr menace son statut de pays laquo officiellement indemne de tuberculose
bovine raquo obtenu en 2001 Dans ce deacutepartement ougrave plus de 190 foyers bovins ont eacuteteacute deacutetecteacutes depuis
2003 M bovis circule dans un systegraveme multi-hocirctes incluant les bovins les cervideacutes les blaireaux et les
sangliers dans une zone geacuteographique localiseacutee Face agrave des contaminations ou des recontaminations
inexpliqueacutees de cheptels avec des souches identiques agrave celles retrouveacutees dans la faune sauvage
infecteacutee deacutetecteacutee agrave proximiteacute dans des eacutelevages le rocircle de lrsquoenvironnement a eacuteteacute suggeacutereacute dans la
transmission indirecte du bacille
Dans ce contexte mes travaux de thegravese visent agrave eacutevaluer la contamination environnementale
2
des zones partageacutees par les bovins et la faune sauvage infecteacutes gracircce agrave des meacutethodes adapteacutees
utilisables sur un grand nombre de preacutelegravevements et agrave rechercher les facteurs favorisant la survie et la
circulation de M bovis dans lrsquoenvironnement Le premier chapitre de ce manuscrit est une synthegravese
des connaissances relatives aux mycobacteacuteries en geacuteneacuteral et aux mycobacteacuteries tuberculeuses en
particulier notamment en terme drsquoeacutecologie de deacutetection et drsquoidentification Le deuxiegraveme chapitre
deacutetaille le deacuteveloppement des meacutethodes moleacuteculaires utiliseacutees pour le diagnostic environnemental
ainsi que le deacuteveloppement de deux nouveaux protocoles pour isoler les mycobacteacuteries du sol et suivre
leur croissance en milieu liquide Le troisiegraveme chapitre preacutesente les modaliteacutes drsquoeacutetude et les reacutesultats
obtenus entre 2013 et 2015 dans lrsquoeacutevaluation de la contamination environnementale en Cocircte drsquoOr Les
reacutesultats drsquoune eacutetude reacutealiseacutee en Charente sont eacutegalement preacutesenteacutes Le quatriegraveme chapitre traite de
lrsquoinfluence de la tempeacuterature et du type de sol sur la dureacutee de survie de M bovis Enfin le cinquiegraveme
chapitre srsquointeacuteresse au rocircle des lombrics sur la disseacutemination de M bovis dans le sol Pour terminer
une discussion geacuteneacuterale syntheacutetise lrsquoensemble des reacutesultats obtenus lors de ce travail et ouvre de
nombreuses perspectives de recherche
Cette thegravese financeacutee par la Reacutegion Bourgogne entre dans le dispositif Jeunes Chercheurs
Entrepreneurs (JCE) Le travail scientifique de recherche srsquoaccompagne de la reacutealisation drsquoun Master
des Entreprises (Master 2) dans lrsquoobjectif drsquoouvrir les doctorants au monde de lrsquoentreprise et au
deacuteveloppement de projets innovants Ce Master a eacuteteacute lrsquooccasion pour moi drsquoacqueacuterir des compeacutetences
compleacutementaires dans le domaine de lrsquoadministration et de la gestion des entreprises notamment
dans la gestion de projets Mon investissement personnel important en parallegravele de mon travail de
recherche mrsquoa permis drsquoobtenir le Master avec la mention Bien
Dans le cadre de la thegravese JCE lrsquoentreprise LSI (speacutecialiseacutee dans le diagnostic veacuteteacuterinaire) a
accompagneacute le deacutemarrage de la thegravese et mrsquoa permis drsquoeacutechanger avec des industriels autour du
deacuteveloppement de systegravemes de deacutetection baseacutes sur la qPCR (IS1561rsquo et autres systegravemes) Suite au
rachat de lrsquoentreprise par Life Science puis Thermo Fisher le transfert de technologie nrsquoa pas eacuteteacute
poursuivi Cependant ces systegravemes de qPCR sont utiliseacutes en routine par le Laboratoire National de
Reacutefeacuterence pour la tuberculose bovine encadreacute par Maria Laura Boschiroli pour caracteacuteriser les souches
de mycobacteacuteries issues drsquoeacutechantillons animaux Drsquoautre part des travaux et reacutesultats obtenus lors de
ma thegravese vont faire lrsquoobjet drsquoune valorisation sous forme drsquoun brevet en cours de deacutepocirct pour la
deacutetection des mycobacteacuteries et drsquoun savoir-faire concernant lrsquoextraction des mycobacteacuteries du sol par
des meacutethodes chimiques en vue de leur mise en culture Jrsquoai donc au cours de mes travaux de recherche
toujours conserveacute lrsquoesprit drsquoinnovation et de transfert potentiel de technologie
CHAPITRE 1
Contexte bibliographique
Chapitre 1
4
CHAPITRE 1 - Contexte bibliographique
A LES MYCOBACTERIES DIVERSITE PHYLOGENIE SPECTRE DrsquoHOcircTES ET VIRULENCE
1 Le genre Mycobacterium taxonomie et meacutethodes de classification
Les connaissances sur les mycobacteacuteries commencent en 1882 date agrave laquelle Robert Koch
deacutecouvre le bacille responsable de la tuberculose humaine alors baptiseacute Bacterium tuberculosis et
partage son postulat sur lrsquoeacutetiologie infectieuse de la maladie La coloration de Ziehl et Neelsen
deacuteveloppeacutee en 1883 pour la recherche des bacilles acido-alcoolo-reacutesistants permet agrave Smith en 1892 de
mettre en eacutevidence un bacille tregraves proche de Bacterium tuberculosis dans des leacutesions tuberculeuses de
bovins Mycobacterium bovis avec toutefois des diffeacuterences pheacutenotypiques et pathogeacuteniques qui
permettent de distinguer les deux bacilles Le bacille de la tuberculose aviaire M avium ssp avium est
isoleacute en 1894 sur des oiseaux puis M avium ssp paratuberculosis en 1895 sur des leacutesions digestives
de bovins La deacutecouverte des premiegraveres mycobacteacuteries non tuberculeuses a suivi celle de Bacterium
tuberculosis notamment gracircce agrave la coloration Mycobacterium smegmatis a eacuteteacute isoleacutee en 1889 dans
des preacutelegravevements biologiques de patients atteints de syphilis puis dans du beurre par lrsquoeacutequipe de Koch
dans les anneacutees 1896 Les premiegraveres recherches meneacutees dans le sol les plantes puis lrsquoeau ont montreacute
que ces substrats pouvaient ecirctre des sources de bacteacuteries morphologiquement tregraves proches du bacille
de Koch Le genre Mycobacterium (mứkecircs champignon et baktecircrίa bacircton) est proposeacute en 1896 par
Lehmann et Neumann Dans les anneacutees 50 environ 20 espegraveces de mycobacteacuteries eacutetaient deacutecrites Le
deacuteveloppement des meacutethodes de culture puis des outils moleacuteculaires dans les anneacutees 1990 ont permis
drsquoisoler de nouvelles espegraveces dans des niches environnementales varieacutees ainsi que chez lrsquohomme et
les animaux A lrsquoheure actuelle 172 espegraveces de mycobacteacuteries sont identifieacutees (Site bacterionetorg)
La diversiteacute pheacutenotypique physiologique eacutecologique et pathogeacutenique des espegraveces placeacutees au
fur et agrave mesure de leur deacutecouverte dans ce genre a meneacute agrave lrsquoeacutelaboration de plusieurs classifications
Au sein du genre Mycobacterium les espegraveces sont geacuteneacuteralement classeacutees en deux grands groupes un
groupe qui rassemble les mycobacteacuteries pathogegravenes obligatoires que sont les mycobacteacuteries
tuberculeuses et les mycobacteacuteries responsables de la legravepre et un second groupe qui rassemble les
mycobacteacuteries pathogegravenes opportunistes et saprophytes appeleacute mycobacteacuteries atypiques ou
mycobacteacuteries environnementales ou mycobacteacuteries non tuberculeuses (MNT)
Dans ces groupes des complexes drsquoespegraveces ont eacuteteacute deacutefinis sur des bases drsquoidentiteacute geacuteneacutetique Le
Complexe tuberculosis (MTBC) rassemble les mycobacteacuteries tuberculeuses notamment
Chapitre 1
5
M tuberculosis M africanum M canetti M bovis M caprae M microti M pinnipedii Parmi les
nombreuses MNT diffeacuterents complexes sont observeacutes dont le plus important est le Complexe
Mycobacterium avium (MAC) qui regroupe quatre sous-espegraveces distinctes dont trois drsquointeacuterecirct en santeacute
humaine et animale (M avium ssp avium M avium ssp paratuberculosis et M avium ssp hominisuis)
(Radomski et al 2010) ainsi qursquoune dizaine drsquoespegraveces dont M intracellulare
Plus reacutecemment la phylogeacutenie moleacuteculaire de ce genre baseacutee notamment sur lrsquoanalyse des gegravenes de
lrsquoARNr 16S et 23S ainsi que des reacutearrangements de gegravenes a permis drsquoaffiner la position taxonomique
du genre Mycobacterium Drsquoapregraves le Bergeyrsquos manual (Ludwig et al 2008) le genre Mycobacterium
appartient
au Phylum des Actinobacteria (bacteacuteries Gram positif agrave geacutenome riche en acides nucleacuteiques
G et C ()
o agrave la Classe des Actinobacteria
o agrave lrsquoOrdre des Corynebacteriales qui regroupe diffeacuterentes familles de bacteacuteries
caracteacuteriseacutees par la preacutesence drsquoacides mycoliques dans leur paroi et des
proprieacuteteacutes tinctoriales particuliegraveres lieacutees agrave la reacutesistance de leur paroi agrave la
deacutecoloration par un meacutelange alcool-acide (ou BAAR)
agrave la Famille des Mycobacteriaceae qui ne comprend que le genre
Mycobacterium caracteacuteriseacute par une enveloppe riche en acides
mycoliques contenant un grand nombre drsquoatomes de carbone ainsi
que des quinones particuliegraveres
2 Caracteacuteristiques geacuteneacuterales des mycobacteacuteries
a Morphologie des mycobacteacuteries
Les mycobacteacuteries se preacutesentent sous forme de bacirctonnets droits ou leacutegegraverement incurveacutes (02-
06 x 10-10microm) non mobiles et ne sporulant pas Les colonies observeacutees sont de couleur blanche agrave
cregraveme voire jaune agrave orangeacutee pour les mycobacteacuteries productrices de pigments (Magee amp Ward 2012)
Elles fixent difficilement la coloration de Gram et bien qursquoelles soient apparenteacutees aux bacteacuteries Gram
+ leur qualification exacte est laquo Bacilles Acido-Alcoolo-Reacutesistants (BAAR) raquo car lors de la coloration
leur enveloppe particuliegravere reacutesiste agrave la deacutecoloration agrave lrsquoalcool et agrave lrsquoacide
Les mycobacteacuteries possegravedent une enveloppe proche de celle des bacteacuteries Gram + mais dont
lrsquoorganisation la richesse et la diversiteacute des acides gras qui la composent sont uniques et
caracteacuteristiques La mycomembrane est constitueacutee drsquoacides gras agrave longue chaine contenant de 60 agrave 90
atomes de carbone les acides mycoliques qui repreacutesentent 30 agrave 40 de sa masse (Brennan amp Nikaido
Chapitre 1
6
1995) Dans sa couche interne les acides mycoliques sont disposeacutes parrallegravelement les uns aux autres
et sont lieacutes au peptidoglycane de faccedilon covalente par lrsquointermeacutediaire de la couche drsquoarabinogalactane
(polymegravere formeacute drsquoarabinose et de galactose) Sa couche externe contient des lipides extractibles (ie
non fixeacutes de faccedilon covalente au peptidoglycane) tels que le treacutehalose monomycolate ou dimycolate
des phospholipides et des glycolipides (Zuber et al 2008) La mycomembrane est recouverte de
glucanes de proteacuteines et drsquoune faible proportion de lipides
Cette enveloppe particuliegravere tregraves riche en lipides confegravere agrave la bacteacuterie ses proprieacuteteacutes
tinctoriales (BAAR) sa grande reacutesistance aux stress chimiques notamment aux antibiotiques et
deacutetergents aux stress physiques (tempeacuterature) ainsi que biologiques (reacutesistance dans les
macrophages de lrsquohocircte et dans les amibes) Crsquoest eacutegalement un facteur important de virulence chez les
mycobacteacuteries pathogegravenes certains composants lipidiques possegravedent en effet une activiteacute
immunogegravene exceptionnelle (Coulombe et al 2009) Ils sont capables de moduler lrsquoimmuniteacute de lrsquohocircte
voire de deacutesorganiser ses meacutecanismes immunitaires de deacutefense et interviennent dans la pathogeacutenie
comme cela a eacuteteacute deacutemontreacute pour M tuberculosis et M avium ssp paratuberculosis (Libero amp Mori
2014 Momotani et al 2012) La paroi est drsquoailleurs une des cibles des antimicrobiens utiliseacutes contre la
tuberculose humaine ou la legravepre Ces derniers agissent notamment en inhibant la biosynthegravese des
moleacutecules qui la constituent Lrsquoutilisation des antigegravenes lipidiques de la paroi constitue eacutegalement un
axe de recherche pour le deacuteveloppement de vaccins (Libero amp Mori 2014)
Dans les matrices environnementales la paroi hydrophobe des mycobacteacuteries leur permet
drsquoadheacuterer fortement aux particules en suspension dans lrsquoeau aux seacutediments aux surfaces favorisant
la formation de biofilms et la survie des bacteacuteries (Primm et al 2004)
Les acides mycoliques de par leur complexiteacute et leur variabiliteacute structurelle sont eacutegalement utiliseacutes
comme marqueurs taxonomiques car ils permettent drsquoidentifier avec speacutecificiteacute le genre et les espegraveces
Chapitre 1
7
Figure 1 Structure et composition de lenveloppe des mycobacteacuteries (drsquoapregraves Marrakchi Laneacuteelle and Daffeacute 2014) AG arabinogalactane PG peptidoglycane TDM treacutehalose dimycolate PL phospholipides TMM treacutehalose monomycolate GL glycolipides
b Caracteacuteristiques culturales pheacutenotypiques et physiologiques
La plupart des mycobacteacuteries sont aeacuterobies strictes mais quelques espegraveces tolegraverent de faibles
pressions drsquooxygegravene notamment M bovis et M tuberculosis Les substrats utiliseacutes comme sources de
carbone et drsquoazote varient consideacuterablement entre les espegraveces et peuvent servir pour leur
identification La majoriteacute des espegraveces croit sur des milieux de culture classiques mais pour certaines
mycobacteacuteries drsquoimportance clinique la culture est ameacutelioreacutee par lrsquoutilisation de milieux agrave lrsquoœuf
(Loumlwenstein-Jensen et Coletsos) apregraves une deacutecontamination chimique ou de milieu Middlebrook ainsi
que par un enrichissement des milieux (en glyceacuterol pour M tuberculosis en mycobactine pour
M avium subsp paratuberculosis en heacutemine pour M haemophilum) Les mycobacteacuteries responsables
de la legravepre sont cultiveacutees sur tissus vivants (coussinets de souris peau de tatou) et se multiplient tregraves
difficilement sur milieux syntheacutetiques
Les mycobacteacuteries ont des vitesses de croissance tregraves variables on distingue les mycobacteacuteries
agrave croissance rapide dont les colonies sont visibles en moins de 7 jours des mycobacteacuteries agrave croissance
lente (gt7 jours) Les bacteacuteries du MTBC sont toutes des mycobacteacuteries agrave croissance lente avec des
temps de geacuteneacuteration eacuteleveacutes de 14 agrave 15h pour M tuberculosis et environ 20h pour M bovis La
tempeacuterature optimale de croissance des bacteacuteries du MTBC est de 37degC Les MNT regroupent des
mycobacteacuteries agrave croissance rapide et agrave croissance lente avec des taux de geacuteneacuteration extrecircment
variables allant de 2h pour les bacteacuteries agrave croissance rapide comme M smegmatis agrave 21h pour celles
agrave croissance lente comme M aurum voire 7 jours pour M avium ssp paratuberculosis dans des
conditions de laboratoire pourtant optimales Les MNT peuvent se multiplier dans un large eacuteventail de
Chapitre 1
8
tempeacuterature allant de 4degC pour M psychrotolerans (Trujillo et al 2004) agrave plus de 45degC voire agrave 52degC
pour M thermotolerans et M phlei (Stahl amp Urbance 1990) avec une tempeacuterature optimale de
croissance entre 28 et 31degC
c Phylogeacutenie du genre Mycobacterium
La phylogeacutenie baseacutee sur le polymorphisme de lrsquoADN codant pour lrsquoARNr 16S permet de
discriminer les espegraveces de mycobacteacuteries et met en eacutevidence trois grands groupes en fonction de leur
vitesse de croissance Le cladogramme preacutesenteacute dans la Figure 2 regroupe distinctement les
mycobacteacuteries agrave croissance lente (haut du cladogramme) les mycobacteacuteries agrave croissance intermeacutediaire
(au milieu) et les mycobacteacuteries agrave croissance rapide (bas du cladogramme) On peut noter de plus que
les mycobacteacuteries du MTBC et le complexe Mac forment deux groupes phylogeacuteneacutetiquement
homogegravenes au sein du groupe des mycobacteacuteries agrave croissance lente
Myc
ob
acteacute
ries
agrave c
rois
san
ce le
nte
Myc
ob
acteacute
ries
agrave
cro
issa
nce
inte
rmeacute
dia
ire
Chapitre 1
9
Figure 2 Arbre phylogeacutenique baseacute sur lrsquoanalyse de lrsquoARNr 16S des principaux complexes et espegraveces de mycobacteacuteries
(drsquoapregraves Magee et Ward 2012)
d Caracteacuteristiques geacuteneacutetiques des mycobacteacuteries
De nombreux geacutenomes complets (annoteacutes ou en cours drsquoanalyse) de mycobacteacuteries sont
actuellement disponibles (NCBI Genome Resources European Nucleotide archive) Mycobacterium
leprae est la mycobacteacuterie qui possegravede le geacutenome le plus petit avec 33 millions de paires de bases
(Mpb) codant pour 1 605 gegravenes et le G+C (pourcentage Guanine+Cytosine) le plus faible (57) Les
mycobacteacuteries environnementales possegravedent les geacutenomes les plus importants de 483 Mpb (codant
4 350 gegravenes) pour M avium k10 agrave 69 Mpb (codant 6 716 gegravenes) pour M smegmatis Les mycobacteacuteries
tuberculeuses M tuberculosis M africanum et M bovis ont un geacutenome de taille intermeacutediaire (de 43
Mpb agrave 45 Mpb) codant pour 3 850 agrave 4 500 gegravenes Les geacutenomes des mycobacteacuteries preacutesentent des
capaciteacutes codantes importantes de lrsquoordre de 90 pour M tuberculosis M bovis M avium ssp
paratuberculosis et M smegmatis (Garnier et al 2003 Cole et al 1998 Li et al 2005)
La comparaison du geacutenome complet de M tuberculosis M bovis M leprae et M avium ssp
paratuberculosis a montreacute une forte conservation des gegravenes impliqueacutes dans le meacutetabolisme
Myc
ob
acteacute
ries
agrave c
rois
san
ce r
apid
e
Chapitre 1
10
eacutenergeacutetique la biosynthegravese des acides amineacutes et des cofacteurs et le meacutetabolisme des nucleacuteotides et
des proteacuteines (Marri et al 2006) Des plasmides ont eacuteteacute mis en eacutevidence chez quelques espegraveces de
mycobacteacuteries environnementales (Rauzier et al 1988 Stinear et al 2004) notamment le plasmide
pMUM001 isoleacute chez M ulcerans responsable de lrsquoulcegravere de Buruli chez lrsquohomme qui code pour un
macrolide aux proprieacuteteacutes cytotoxiques et immunosuppressives la mycolactone (Stinear et al 2004)
Le nombre drsquoopeacuterons ribosomaux 16S dans le geacutenome des mycobacteacuteries a eacuteteacute suggeacutereacute comme
lrsquoun des facteurs expliquant la diffeacuterence des vitesses de croissance observeacutees La croissance lente des
mycobacteacuteries serait lieacutee agrave la preacutesence drsquoun seul opeacuteron ribosomal les espegraveces agrave croissance rapide en
posseacutedant geacuteneacuteralement deux (rrnA et rrnB) (Bercovier et al 1986) Toutefois des exceptions existent
et drsquoautres facteurs tels que la lenteur des eacutechanges agrave travers leur paroi impermeacuteable riche en lipides
et le coucirct eacutenergeacutetique eacuteleveacute de la synthegravese de leurs acides gras agrave longues chaines seraient eacutegalement
impliqueacutes (Primm et al 2004)
Dans le Complexe tuberculosis les diffeacuterentes espegraveces constituent un modegravele drsquoidentiteacute
geacuteneacutetique interspeacutecifique Leurs geacutenomes preacutesentent une identiteacute supeacuterieure agrave 999 au niveau
nucleacuteotidique et des seacutequences drsquoARNr 16S et drsquoITS identiques (Boumlddinghaus et al 1990 Sreevatsan amp
Pan 1997 Huard et al 2006 Cole et al 1998 Garnier et al 2003 Kasai et al 2000) Lrsquoanalyse des
seacutequences de 56 gegravenes de structure de plusieurs centaines drsquoisolats drsquoorigines geacuteographique et
phylogeacuteneacutetique varieacutees suggegravere que le polymorphisme alleacutelique est exceptionnel dans ce Complexe
(Sreevatsan amp Pan 1997 Musser et al 2000) La recombinaison de tregraves petits fragments drsquoADN est
deacutemontreacutee chez M tuberculosis (Namouchi et al 2012) mais le transfert lateacuteral de gegravenes ne semble
pas ecirctre un facteur important dans lrsquoeacutevolution des bacteacuteries du MTBC qui ont une structure de
population fortement clonale (Pepperell et al 2013) contrairement agrave la plupart des autres bacteacuteries
pathogegravenes Malgreacute leur forte identiteacute geacuteneacutetique les espegraveces du Complexe se diffeacuterencient par des
reacutegions polymorphiques ou LSPs (Large Sequence Polymorphisms) lieacutees agrave des deacuteleacutetions et agrave des
reacutearrangements de seacutequences (Brosch et al 2002 Huard et al 2003) Parmi ces LSPs 14 Reacutegions de
Diffeacuterence ou RD de 2 agrave 127 kpb sont recenseacutees dans le MTBC Ces deacuteleacutetions sont consideacutereacutees comme
une adaptation eacutevolutive agrave lrsquoinfection de nouveaux hocirctes plutocirct qursquoagrave une deacuterive geacuteneacutetique (Brodin et
al 2002 Aranaz et al 2003) Cette variabiliteacute geacutenomique permet agrave la fois de diffeacuterencier les espegraveces
entre elles (Huard et al 2003 Huard et al 2006) et de retracer leur eacutevolution phylogeacuteneacutetique
(Rodriguez-Campos et al 2014)
3 Description des diffeacuterents Complexes ou espegraveces de mycobacteacuteries
Alors que les mycobacteacuteries responsables de la tuberculose et de la legravepre sont des pathogegravenes
obligatoires les MNT rassemblent des mycobacteacuteries saprophytes sans pouvoir pathogegravene jusque-lagrave
Chapitre 1
11
aveacutereacute et des mycobacteacuteries pathogegravenes opportunistes responsables de mycobacteacuterioses humaines ou
animales Depuis la fin du 19egraveme siegravecle jusqursquoaux anneacutees 1950 la tuberculose humaine repreacutesentait un
tel problegraveme de santeacute publique que toutes les eacutetudes eacutetaient centreacutees sur elle et son agent
M tuberculosis Les autres mycobacteacuteries isoleacutees dans lrsquoenvironnement ou dans des preacutelegravevements
biologiques humains et animaux eacutetaient consideacutereacutees comme des bacteacuteries saprophytes sans inteacuterecirct
clinique particulier A partir des anneacutees 1950 la communauteacute scientifique a admis la responsabiliteacute
indiscutable de certaines mycobacteacuteries environnementales dans des processus pathologiques
humains et animaux appeleacutes mycobacteacuterioses amorccedilant des eacutetudes les concernant En lrsquoabsence de
contamination inter-humaine deacutemontreacutee il a eacuteteacute admis que lrsquoorigine de ces mycobacteacuteries est
environnementale et que lrsquohomme se contamine agrave partir de lrsquoenvironnement
a Les mycobacteacuteries non pathogegravenes potentiellement opportunistes
Elles regroupent les mycobacteacuteries environnementales qui preacutesentent un pouvoir pathogegravene
pour lrsquohomme et les animaux tregraves variable en fonction des espegraveces (van Ingen et al 2009 Tortoli 2003)
Lrsquoincidence des infections humaines lieacutees agrave des mycobacteacuteries opportunistes semble augmenter dans
les pays industrialiseacutes (Falkinham 2016) peut-ecirctre en lien avec lrsquoameacutelioration des outils pour les isoler
et les identifier Les mycobacteacuterioses qursquoelles engendrent surviennent geacuteneacuteralement sur un terrain
drsquoimmunodeacutepression locale ou geacuteneacuterale Chez lrsquohomme elles sont responsables drsquoun large eacuteventail de
symptocircmes cliniques majoritairement des infections pulmonaires mais eacutegalement des lymphadeacutenites
chez les enfants des infections cutaneacutees (notamment lieacutees agrave M marinum responsable du granulome
des piscines et M ulcerans responsable de lrsquoulcegravere de Buruli) des infections extra-pulmonaires voire
des infections disseacutemineacutees chez les patients seacutevegraverement immunodeacuteprimeacutes (van Ingen 2013 Biet et al
2005 Hoefsloot et al 2013) Environ 25 espegraveces sont retrouveacutees reacuteguliegraverement dans les eacutechantillons
cliniques les autres beaucoup plus rarement (van Ingen 2013) Parmi elles les mycobacteacuteries agrave
croissance lente semblent plus preacutedisposeacutees agrave provoquer des atteintes pulmonaires ou lymphatiques
alors que celles agrave croissance rapide affectent plutocirct la peau les os et les articulations (Tortoli 2003
2009) Les mycobacteacuteries M avium et M intracellulare sont les plus freacutequemment isoleacutees chez
lrsquohomme majoritairement en cas drsquoatteinte pulmonaire (Hoefsloot et al 2013 Whiley et al 2012) La
distribution des espegraveces pathogegravenes isoleacutees chez lrsquohomme diffegravere selon la zone geacuteographique eacutetudieacutee
probablement en lien avec leurs niches eacutecologiques (Hoefsloot et al 2013)
Chez les animaux M avium ssp paratuberculosis (Map) est lrsquoagent de la maladie de Johne ou
paratuberculose chez les ruminants domestiques et sauvages les primates ainsi que drsquoautres espegraveces
(lapins etc) chez qui elle provoque une enteacuterite chronique contagieuse et incurable agrave lrsquoorigine de
pertes eacuteconomiques importantes dans les cheptels infecteacutes Son rocircle est eacutegalement eacutevoqueacute dans la
Chapitre 1
12
maladie de Crohn chez lrsquohomme (Biet et al 2005 Rowe amp Grant 2006 Momotani et al 2012) sans
toutefois que la causaliteacute ait eacuteteacute deacutemontreacutee Mycobacterium avium ssp avium (Maa) est responsable
de la tuberculose aviaire chez les oiseaux et les petits mammifegraveres terrestres chez qui elle provoque
des leacutesions tuberculeuses majoritairement digestives des lymphadeacutenites et des infections disseacutemineacutees
(Biet et al 2005 Pavlik et al 2000) Mycobacterium avium ssp hominissuis (Mah) est responsable de
leacutesions des nœuds lymphatiques et de diffeacuterents organes chez le porc (Pavlik et al 2000)
b Les mycobacteacuteries saprophytes non pathogegravenes
Parmi toutes les mycobacteacuteries environnementales deacutecrites agrave lrsquoheure actuelle certaines sont
retrouveacutees uniquement dans lrsquoenvironnement et nrsquoont jamais eacuteteacute isoleacutees chez lrsquohomme ou lrsquoanimal
comme M arabiense ou M rufum (Tortoli 2014) Ces mycobacteacuteries sont dites saprophytes Parmi
elles certaines ont eacuteteacute isoleacutees dans des sols et des seacutediments fortement pollueacutes par les HAP
(Hydrocarbures Aromatiques Polycycliques) qursquoelles sont potentiellement capables de deacutegrader
(Hennessee et al 2009 Leys et al 2005 Hormisch et al 2004) Elles repreacutesentent donc une voie
inteacuteressante pour la bioremeacutediation de ces sols
c Les mycobacteacuteries pathogegravenes obligatoires
Les mycobacteacuteries responsables de legravepre
Les mycobacteacuteries responsables de la legravepre humaine et murine sont M lepreae M lepromatosis et
M lepreaemurium Elles provoquent des infections dermatologiques et nerveuses chroniques graves
et mutilantes parfois mortelles Mycobacterium leprae touche environ 15 million de personnes dans
le monde et la maladie reste un problegraveme de santeacute publique preacuteoccupant dans les zones endeacutemiques
drsquoAfrique drsquoAsie et drsquoAmeacuterique du sud (Who 2014) Mycobacterium leprae a longtemps eacuteteacute consideacutereacutee
comme un pathogegravene obligatoire de lrsquohomme avec une voie de transmission interhumaine majoritaire
La deacutecouverte de reacuteservoirs animaux (Truman 2005 Meredith et al 2014) et la deacutetection de M leprae
dans le sol la sphaigne ou lrsquoeau de riviegravere (Matsuoka et al 1999 Mostafa et al 1995 Lavania et al
2006) tendent agrave souligner le rocircle potentiel de certains animaux ou de lrsquoenvironnement comme source
de lrsquoinfection chez lrsquohomme Ces 3 espegraveces sont multiplieacutees sur les coussinets de souris (Shepard 1960)
et sur la peau de tatous agrave 9 bandes (Dasypus novemcinctus) (Kirchheimer amp Storrs 1971) car aucun
milieu syntheacutetique ne permet leur croissance optimale
Chapitre 1
13
Les mycobacteacuteries tuberculeuses le Complexe tuberculosis
(MTBC)
Toutes les mycobacteacuteries tuberculeuses sont des pathogegravenes obligatoires Contrairement aux
mycobacteacuteries environnementales leur croissance dans lrsquoenvironnement nrsquoa jamais eacuteteacute deacutemontreacutee Le
complexe tuberculosis comprend agrave ce jour 13 espegraveces qui malgreacute une forte identiteacute geacuteneacutetique
preacutesentent des pheacutenotypes des hocirctes et des pouvoirs pathogegravenes varieacutes
Phylogeacutenie des mycobacteacuteries du MTBC
Lrsquoeacutetude des RD eacutevoqueacutees preacuteceacutedemment et des mutations agrave lrsquoeacutechelle du nucleacuteotide (ou SNP Single
Nucleacuteotide Polymorphism) sur 7 gegravenes (katG gyrA gyrB rpoB mmpL6 oxyR et ADNr 16S) ont permis
de retracer la phylogeacutenie des bacteacuteries du Complexe agrave partir de leur ancecirctre commun (Brosch et al
2002 Rodriguez-Campos et al 2014 Gordon et al 1999 Mahairas et al 1996 Huard et al 2003) les
deacuteleacutetions et mutations jouant un rocircle drsquohorloge moleacuteculaire permettant de replacer lrsquoapparition et
lrsquoeacutevolution drsquoune espegravece dans le temps Le sceacutenario de lrsquoeacutevolution des bacteacuteries du MTBC est
scheacutematiseacute dans la Figure 3 Selon les auteurs lrsquoancecirctre du MTBC appeleacute M protuberculosis aurait
eacutemergeacute il y a entre 15 000 et 40 000 ans en Afrique de lrsquoEst (Gutierrez et al 2005 Wirth et al 2008
Sreevatsan amp Pan 1997) A cocircteacute de M canetti la bacteacuterie du MTBC geacuteneacutetiquement la plus proche de
lrsquoancecirctre deux clades principaux peuvent ecirctre observeacutes Un clade de souches speacutecifiques de lrsquohomme
caracteacuteriseacute par la deacuteleacutetion TbD1 a donneacute les diffeacuterentes ligneacutees modernes de M tuberculosis (Figure
3) Un second clade caracteacuteriseacute par la perte des reacutegions RD7 RD8 RD9 et RD10 regroupe des souches
endeacutemiques drsquoAfrique speacutecifiques de lrsquohomme (M africanum) et drsquoanimaux (le Chimpanzee bacillus
M mungi le Dassie bacillus et M suricattae) ainsi que des souches adapteacutees agrave diffeacuterentes espegraveces de
mammifegraveres (M orygis M microti M pinnipedii M caprae et M bovis)
Chapitre 1
14
Figure 3 Phylogeacutenie des souches du MTBC baseacutee sur les Reacutegions de Diffeacuterences et diffeacuterents Single Nucleotide Polymorphism (Dippenaar et al 2015)
Virulence des mycobacteacuteries du MTBC
Les mycobacteacuteries du MTBC ne possegravedent pas de facteurs de virulence classiques tels que les
toxines par exemple observeacutes chez drsquoautres bacteacuteries pathogegravenes Pour certains auteurs la virulence
des mycobacteacuteries est restreinte agrave lrsquoaptitude du bacille agrave peacuteneacutetrer dans lrsquohocircte agrave y survivre et srsquoy
multiplier (Pollock amp Neill 2002) De faccedilon plus large la notion de virulence chez les membres du MTBC
a eacuteteacute conceptualiseacutee comme lrsquoassociation de i) la capaciteacute de la bacteacuterie agrave faire face agrave la reacuteponse
immunitaire de lrsquohocircte infecteacute ii) la capaciteacute agrave engendrer des leacutesions iii) la capaciteacute agrave survivre en dehors
de son hocircte notamment lors de lrsquoaeacuterosolisation et iiii) la transmission et lrsquoinfection drsquoun nouvel hocircte
(Coscolla amp Gagneux 2014) La virulence implique donc un ensemble de meacutecanismes complexes et
varieacutes faisant intervenir de nombreux gegravenes et proteacuteines permettant notamment au bacille de peacuteneacutetrer
dans les cellules de lrsquohocirctes et drsquoy survivre en bloquant les diffeacuterents meacutecanismes de deacutefense des
macrophages (inhibition de la fusion phagosome-lysosome et de la production de ROS1 et RNS2
deacutesorganisation de lrsquoimmuniteacute de lrsquohocircte inhibition de lrsquoautophagie et de lrsquoapoptose) (Hmama et al
1 ROS Reactive Oxygen Species 2 RNS Reactive Nitrogen Species
Chapitre 1
15
2015) Plus de cent gegravenes seraient impliqueacutes dans la virulence des bacteacuteries du MTBC et lrsquoutilisation
de mutants permet peu agrave peu de connaitre le rocircle de chacun (Forrellad et al 2013) Lrsquoanalyse du
transcriptome (ARN) de M tuberculosis au cours de lrsquoinfection de macrophages et de cellules
dendritiques a reacuteveacuteleacute lrsquoexpression ou la surexpression de nombreux gegravenes codant des proteacuteines
intervenant notamment dans le meacutetabolisme lipidique la synthegravese de lrsquoenveloppe bacteacuterienne
lrsquoapprovisionnement en fer la reacutesistance au stress oxydatif et lrsquoinhibition de lrsquoapoptose (Mukhopadhyay
et al 2012) Les meacutecanismes de survie en dehors de lrsquohocircte sont inconnus agrave ce jour
Il a eacuteteacute montreacute chez M tuberculosis qursquoen fonction des ligneacutees ou des souches la virulence pouvait
varier entrainant une reacuteponse immunitaire reacuteduite ou retardeacutee chez lrsquohocircte une plus grande seacuteveacuteriteacute
de la maladie et une transmission augmenteacutee (Coscolla amp Gagneux 2014) Ce lien souche-virulence est
eacutegalement suspecteacute chez M bovis pour lequel certains geacutenotypes srsquoaccompagnent drsquoun plus grand
nombre drsquoanimaux agrave leacutesions et ou drsquoun score leacutesionnel plus eacuteleveacute (Wright et al 2013 Gavier-Wideacuten et
al 2009 Garbaccio et al 2014) mais des eacutetudes compleacutementaires restent neacutecessaires
Les mycobacteacuteries speacutecifiques de lrsquohomme
Mycobacterium tuberculosis
Mycobacterium tuberculosis est lrsquoagent de la tuberculose humaine maladie retrouveacutee dans
toutes les reacutegions du monde Crsquoest un problegraveme majeur de santeacute publique puisque 2 milliards
drsquoindividus seraient infecteacutes dans le monde En 2013 lrsquoOMS a eacutevalueacute agrave neuf millions le nombre de
nouveaux cas annuels et agrave un million et demi le nombre de deacutecegraves lieacutes agrave la tuberculose (dont 24 de
personnes seacuteropositives) Les reacutegions de lrsquoAsie du sud-est et du Pacifique occidental regroupent 56
des nouveaux cas lrsquoAfrique 25 lrsquoInde 24 et la Chine 11 (WHO 2014) Lrsquoinfection par le HIV est un
facteur preacutedisposant majeur dans la transmission de la maladie et la progression de lrsquoinfection chez
lrsquohomme de par les effets synergiques deacuteleacutetegraveres des 2 micro-organismes sur le systegraveme immunitaire
(Pawlowski et al 2012) Environ 78 des personnes co-infecteacutees vivent en Afrique (WHO 2014) La
tuberculose se manifeste chez lrsquohomme par des granulomes pulmonaires (tuberculose pulmonaire)
mais peut se deacutevelopper dans drsquoautres organes (tuberculose extra-pulmonaire) Elle se transmet
essentiellement par voie respiratoire par lrsquoinhalation de gouttelettes contamineacutees eacutemises par une
personne infecteacutee lors de toux par exemple Le traitement repose sur lrsquoassociation de plusieurs
antibiotiques adapteacutes tels que lrsquoisoniazide la rifampicine le pyrazinamide et lrsquoeacutethambutol
Lrsquoapparition de souches reacutesistantes aux antibiotiques MDR (Multi Drug Resistant) et XDR
(eXtensively Drug Resistant) constitue agrave lrsquoheure actuelle un problegraveme majeur dans la lutte contre la
maladie Bien que lrsquohomme constitue le reacuteservoir majeur de la bacteacuterie M tuberculosis eacuteteacute isoleacutee chez
Chapitre 1
16
un grand nombre drsquoespegraveces animales telles que les bovins domestiques (Romero et al 2011 Prasad et
al 2005 Ameni et al 2011 Pavlik et al 2005 Cadmus et al 2006) les chegravevres (Cadmus Adesokan et
al 2009) les porcs domestiques (Jenkins et al 2011) les chiens (Pavlik et al 2005) les oiseaux et des
animaux de zoos (Michel et al 2003) dont les eacuteleacutephants
Mycobacterium africanum
Elle est principalement retrouveacutee dans les pays drsquoAfrique de lrsquoouest ougrave elle peut ecirctre impliqueacutee
dans la moitieacute des cas de tuberculose humaine agrave cocircteacute de M tuberculosis (de Jong et al 2010) Isoleacutee
et deacutecrite pour la premiegravere fois au Seacuteneacutegal en 1968 elle preacutesente des caracteacuteristiques biochimiques
variables intermeacutediaires entre celles de M tuberculosis et M bovis (Castets amp Sarrat 1968)
Mycobacterium africanum est parfois isoleacutee chez lrsquohomme en dehors de lrsquoAfrique (Schroumlder 1982
Grange amp Yates 1989 Desmond et al 2004 Frottier et al 1990) La proximiteacute phylogeacuteneacutetique de
M africanum avec les autres membres du MTBC infectant des espegraveces animales a souleveacute la question
drsquoun reacuteservoir animal de la bacteacuterie Mais elle a eacuteteacute isoleacutee de faccedilon sporadique sur des singes (Thorel
1980) des bovins (Rahim et al 2007 Cadmus et al 2006) et du lait (Cadmus et al 2010) infirmant
cette hypothegravese
Mycobacterium canetti et smooth tubercle bacilli
Elle est responsable de tuberculose humaine dans une zone geacuteographique confineacutee agrave la corne de
lrsquoAfrique (Djibouti Somalie Kenya) (Pfyffer et al 1998 Miltgen et al 2002) Elle est la plus proche
geacuteneacutetiquement de lrsquoancecirctre des bacteacuteries du MTBC Elle constitue un taxon divergent des autres
membres du MTBC par son pheacutenotype particulier et son geacutenotype qui preacutesente une large diversiteacute
geacuteneacutetique et des signes de transfert horizontal et de recombinaison de gegravenes contrairement aux autres
bacteacuteries du MTBC (Fabre et al 2004 Supply et al 2013 Gutierrez et al 2005) Sa situation
geacuteographique confineacutee la grande diversiteacute geacuteneacutetique des souches rencontreacutees et le manque de
connaissances sur la transmission de la bacteacuterie ont suggeacutereacute lrsquoexistence drsquoun reacuteservoir environnemental
(Fabre et al 2004 Koeck et al 2011) A ce jour cette hypothegravese nrsquoa pas eacuteteacute confirmeacutee La tuberculose
agrave M canetti est consideacutereacutee aujourdrsquohui comme une maladie eacutemergente
Les mycobacteacuteries speacutecifiques des animaux
Mycobacterium bovis
Mycobacterium bovis est lrsquoagent de la tuberculose bovine maladie preacutesente sur tous les
continents Les bovins constituent le reacuteservoir naturel du bacille mais un grand nombre drsquoespegraveces
Chapitre 1
17
animales domestiques et sauvages peuvent ecirctre infecteacutees ainsi que lrsquohomme (Biet et al 2005 Palmer
2013 Grange amp Yates 1994 OrsquoReilly amp Daborn 1995) Sur le plan microbiologique M bovis se
caracteacuterise par une croissance tregraves lente sur milieu de Loumlwenstein-Jensen (6 agrave 10 semaines) par des
colonies petite taille (1 agrave 2 mm) plates lisses laquo dysgoniques raquo et achromogegravenes Sa croissance est
ameacutelioreacutee par lrsquoajout de pyruvate et inhibeacutee par lrsquoajout de glyceacuterol Sur Middlebrook 7H11 les colonies
sont rugueuses et agrave bords eacutechancreacutes (Figure 4) Le pH optimum de culture de M bovis est compris
entre 58 et 69 mais la bacteacuterie survit 20 jours dans du yaourt agrave 4degC (Mitscherlich amp Marth 2012)
Lrsquoidentification pheacutenotypique repose notamment sur lrsquoabsence de production de niacine lrsquoabsence de
nitrate reacuteductase une reacutesistance au pyrazinamide agrave la D-cycloseacuterine (30mgmL-1) et une croissance
favoriseacutee par la preacutesence de pyruvate (Delaune et al 2012)
Figure 4 Aspect des colonies de M bovis sur milieu de Lowenstein-Jensen (agrave gauche) et Middlebrook 7H11 suppleacutementeacute (agrave droite)
Concernant les caracteacuteristiques geacutenomiques de M bovis le seacutequenccedilage complet de la
premiegravere souche (M bovis AF212297 ou ATCC BAA-935 isoleacutee drsquoune leacutesion caseacuteeuse de bovin en
Grande-Bretagne) en 2003 et son annotation ont enrichi les connaissances sur ce bacille Son geacutenome
comprend 4345 Mpb un pourcentage G-C eacuteleveacute de 656 et 3952 gegravenes codant des proteacuteines dont 1
prophage et 42 seacutequences drsquoinsertion Il preacutesente 11 deacuteleacutetions de 1 agrave 127 kb par comparaison avec le
geacutenome de M tuberculosis H37Rv et 1 locus absent chez M tuberculosis appeleacute TbD1 (Garnier et al
2003) Ne posseacutedant pas de gegravenes propres agrave lrsquoespegravece bovis le geacutenome de M bovis a eacuteteacute faccedilonneacute par
lrsquoapparition de ces deacuteleacutetions conditionnant lrsquoexpression des gegravenes Depuis 2003 une quarantaine de
souches isoleacutees sur des animaux domestiques et sauvages ont eacuteteacute entiegraverement seacutequenceacutees (Sites
NCBI et European Nucleotide archive)
La structure de la population de M bovis reacutesulte de lrsquoexpansion de plusieurs complexes clonaux
issus drsquoun mecircme ancecirctre Quatre complexes clonaux majeurs ont eacuteteacute identifieacutes dans le monde pour
Chapitre 1
18
M bovis en se basant sur la preacutesence de deacuteleacutetions chromosomiques caracteacuteristiques dans le geacutenome
de la bacteacuterie etou sur des SNPs ainsi que sur le spoligotype et lrsquoabsence de certains spacers (Smith
2012) le Complexe African 1 est retrouveacute dans plusieurs pays africains (Mali Cameroun Nigeacuteria
Tchad) (Muumlller et al 2009) le Complexe African 2 est essentiellement retrouveacute chez les bovideacutes
drsquoAfrique de lrsquoEst (Berg et al 2011) le Complexe European 1 est retrouveacute en Europe il est dominant au
Royaume-Uni et beaucoup plus rare en Espagne au Portugal et en France On le retrouve eacutegalement
chez les anciens partenaires commerciaux du Royaume-Uni (USA Afrique du Sud Nouvelle Zeacutelande
Australie Canada) et en Ameacuterique du sud La diffusion geacuteographique de ce Complexe est lieacutee aux
eacutechanges commerciaux de bovins (Smith et al 2011) Enfin le Complexe European 2 est majoritaire au
Portugal et en Espagne et assez freacutequent en France et en Italie (Rodriguez-Campos et al 2012) Drsquoautres
complexes clonaux minoritaires existent En France plusieurs complexes clonaux coexistent certains
autochtones eacutevoluent depuis longtemps sur le territoire alors que drsquoautres ont eacuteteacute importeacutes agrave la faveur
des eacutechanges commerciaux de bovins (Hauer et al 2015)
Une souche atteacutenueacutee obtenue agrave partir drsquoun isolat virulent de M bovis a eacuteteacute creacuteeacutee par Calmette
et Gueacuterin apregraves 230 repiquages sur un milieu bilieacute enrichi en glyceacuterol entre 1908 et 1921 le BCG pour
Bacille de Calmette et Gueacuterin Testeacutee sur des modegraveles animaux cette souche atteacutenueacutee immunisait
contre une infection expeacuterimentale agrave M bovis et M tuberculosis trente jours apregraves injection Utiliseacutee
pour la vaccination elle a eacuteteacute distribueacutee dans le monde entier et maintes fois repiqueacutee ce qui a meneacute
agrave une eacutevolution de la souche initiale en plusieurs souches tregraves heacuteteacuterogegravenes La comparaison geacutenomique
de la souche BCG avec M tuberculosis reacutevegravele la perte de 14 reacutegions (RD1 agrave RD14) chez BCG Lrsquoabsence
de la reacutegion RD1 expliquerait en partie lrsquoatteacutenuation du BCG (Pym et al 2002) toutefois comme la
reacuteintroduction de RD1 dans une souche mutante ne suffit pas agrave restaurer totalement la virulence
drsquoautres reacutegions sont impliqueacutees dans lrsquoatteacutenuation (Brosch et al 2007)
Mycobacterium pinnipedii
Deacutecouverte en 1986 cette mycobacteacuterie touche principalement les diffeacuterentes espegraveces de
phoques et les lions de mer sauvages de lrsquoheacutemisphegravere sud ou en captiviteacute Elle a eacuteteacute isoleacutee en Australie
Nouvelle-Zeacutelande Uruguay Argentine et Grande-Bretagne (Cousins et al 2003) Des infections chez
lrsquohomme ont eacuteteacute rapporteacutees principalement chez les soigneurs en contact avec les animaux malades
en captiviteacute (Kiers et al 2008) La transmission indirecte du bacille par inhalation drsquoaeacuterosols lors du
nettoyage des enclos est fortement suspecteacutee
Mycobacterium caprae
Cette mycobacteacuterie du MTBC a eacuteteacute isoleacutee pour la premiegravere fois sur des chegravevres en Espagne
(Aranaz et al 1999 Aranaz et al 2003) Crsquoest lrsquoun des agents de la tuberculose bovine avec M bovis
Chapitre 1
19
et elle induit des leacutesions identiques agrave la tuberculose agrave M bovis chez les animaux infecteacutes La
tuberculose agrave M caprae touche principalement lrsquoEurope ougrave elle a deacutejagrave eacuteteacute deacutetecteacutee en Espagne
(Rodriacuteguez et al 2011) en France (Haddad et al 2001) en Autriche (Prodinger et al 2002) en
Allemagne (Kubica et al 2003) et dans plusieurs pays drsquoEurope centrale (Pavlik et al 2005 Beširović
et al 2012 Cvetnic et al 2007) Le nombre de cas deacutetecteacutes tend agrave augmenter chez les bovins et les
animaux sauvages notamment en Espagne en Allemagne en Autriche (Rodriacuteguez et al 2011 Schoepf
et al 2012) En Europe centrale elle repreacutesente lrsquoagent principal de la tuberculose bovine notamment
en Reacutepublique Tchegraveque Slovaquie Bosnie Herzeacutegovine et Croatie ougrave elle circule entre les bovins et de
nombreuses espegraveces sauvages (Prodinger et al 2005 Beširović et al 2012) Elle a eacutegalement eacuteteacute isoleacutee
pour la premiegravere fois en Chine en 2013 (Zeng et al 2013) Crsquoest eacutegalement un agent zoonotique
important puisque des infections agrave M caprae sont rapporteacutees chez lrsquohomme dans diffeacuterents pays
drsquoEurope ((Kubica et al 2003 Prodinger et al 2014 Prodinger et al 2002 Muumlller et al 2013)
Mycobacterium microti
Cette mycobacteacuterie est lrsquoagent responsable de la tuberculose des petits rongeurs (campagnols
souris musaraignes hellip) chez qui elle a eacuteteacute isoleacutee dans les anneacutees 30 (Wells 1937 Wells amp Robb-Smith
1946) Elle a eacuteteacute isoleacutee chez plusieurs espegraveces de mammifegraveres dont des chats (Gunn-Moore et al 1996
Gunn-Moore et al 2011) des bovins (Jahans et al 2004) des porcs (Taylor et al 2006) des lamas des
blaireaux (Smith et al 2009) des sangliers (Dondo et al 2007) des suricates (Palgrave et al 2012) En
France la deacutetection de M microti chez des animaux domestiques et sauvages a suggeacutereacute que la bacteacuterie
eacutetait endeacutemique dans certaines reacutegions de France (Michelet et al 2015 Laprie et al 2013) Chez
lrsquohomme une trentaine de cas drsquoinfections agrave M microti principalement agrave localisation pulmonaire ont
eacuteteacute reporteacutes dans la litteacuterature entre 2000 et 2010 (Panteix et al 2010) Bien que les personnes
immunodeacuteprimeacutees soient les plus toucheacutees lrsquoinfection se rencontre eacutegalement chez des sujets
immunocompeacutetents (Niemann et al 2000 Frank et al 2009)
Mycobacterium orygis
Isoleacutee pour la premiegravere fois en 1976 sur des oryxes en Afrique de lrsquoest (Lomme et al 1976) elle
a eacuteteacute isoleacutee dans de rares occasions sur drsquoautres espegraveces de ruminants tels que les gazelles les
antilopes (Van Soolingen et al 1994) les buffles africains (van Pittius et al 2012) en Afrique du sud
dans la Peacuteninsule arabe et dans des zoos des Pays Bas et du Royaume Uni
Des infections agrave Mycobacterium orygis ont eacuteteacute rapporteacutees chez une dizaine de patients humains
(Dawson et al 2012 van Ingen et al 2012)
Chapitre 1
20
Autres mycobacteacuteries agrave reacuteservoir animal
Quatre autres espegraveces de mycobacteacuteries animales phylogeacuteneacutetiquement proches de
M africanum West African 2 ont eacuteteacute isoleacutees sur des animaux en Afrique le Chimpazee bacillus isoleacute
sur un chimpanzeacute de Cocircte drsquoIvoire (Coscolla et al 2013) M mungi isoleacutee sur des mangoustes rayeacutees
(Mungos mungo) du Bostwana (Alexander et al 2010) le Dassie bacillus isoleacute sur un daman du Cap
(Procavia capensis) (Wagner et al 1958) et M suricattae isoleacutee sur des suricates (Suricata suricatta) en
Afrique du sud (Parsons et al 2013) Ces espegraveces nrsquoont agrave ce jour jamais eacuteteacute isoleacutees chez lrsquohomme La
deacutecouverte reacutecente de ces espegraveces rattacheacutees au MTBC suggeacutere que ce Complexe est probablement
plus varieacute que ce qui eacutetait imagineacute
B LA TUBERCULOSE UNE MALADIE TRES ANCIENNE MAIS UNE MENACE TOUJOURS ACTUELLE
1 Introduction
La tuberculose est une maladie bacteacuterienne chronique et contagieuse qui affecte lrsquohomme
ainsi que de nombreux animaux domestiques et sauvages Elle est provoqueacutee par les diffeacuterentes
espegraveces de mycobacteacuteries du MTBC Lrsquoobservation de tubercules granulomes inflammatoires formeacutes
de cellules immunitaires et de caseacuteum (zones neacutecrotiques) dans les organes ou les nœuds
lymphatiques des personnes infecteacutees a donneacute son nom agrave la maladie
2 Histoire de la tuberculose
La tuberculose est une maladie tregraves ancienne puisque lrsquoancecirctre du Complexe tuberculosis serait
apparu il y a 40 000 ans (Wirth et al 2008) La preacutesence exclusive de certaines ligneacutees du MTBC sur le
continent africain suggegravere que le Complexe aurait eacutemergeacute en Afrique de lrsquoEst puis aurait co-eacutevolueacute
avec lrsquohomme et se serait reacutepandu dans le monde entier par le biais des migrations humaines en
Europe en Asie et en Afrique continentale Les populations de mycobacteacuteries tuberculeuses eacutetant
infeacuteodeacutees agrave une zone geacuteographique elles se sont adapteacutees agrave la population locale (Gutierrez et al 2005
Djelouadji et al 2011 Wirth et al 2008) Le passage drsquoun mode de vie nomade agrave une vie seacutedentaire
dans des fermes agricoles de plus en plus peupleacutees ainsi que la domestication des animaux ont
probablement contribueacute agrave la transmission de la maladie agrave divers hocirctes animaux et agrave lrsquoadaptation du
bacille permettant lrsquoapparition de M bovis M caprae et des autres membres du Complexe (Wirth et
al 2008) Les plus anciennes traces drsquoADN du Complexe tuberculosis ont eacuteteacute deacutecouvertes sur des
Chapitre 1
21
ossements de bison vieux de 17 000 ans retrouveacutes dans une grotte drsquoAmeacuterique du Nord (Rothschild et
al 2001) Des traces drsquoADN du Complexe tuberculosis de M tuberculosis ou M bovis ont eacuteteacute
retrouveacutees sur des ossements humains datant de la Preacutehistoire ou de lrsquoAntiquiteacute (les plus vieux ont
9000 ans) deacutecouverts dans diffeacuterentes parties du monde (Chili Royaume Uni Russie Israeumll Indoneacutesie
etc) (Djelouadji et al 2011) Appeleacutee laquo phtisie raquo par les Grecs peste blanche ou consomption (suite au
deacutepeacuterissement des malades) les eacutecrits drsquoHippocrate (5egraveme siegravecle avant JC) sont les premiers agrave deacutecrire la
symptomatologie et les diffeacuterentes formes de la tuberculose Fracastor (15egraveme siegravecle) suggegravere le
caractegravere heacutereacuteditaire de la maladie mais place la phtisie dans les maladies contagieuses au mecircme titre
que la veacuterole hypothegravese reprise en 1722 par Marten puis par Villemin en 1865 qui montre que la
maladie est infectieuse contagieuse et inoculable (Dutau 2005) A la fin du 18egraveme siegravecle et au deacutebut du
19egraveme la tuberculose est la premiegravere cause de mortaliteacute en Europe de lrsquoOuest et en Ameacuterique du Nord
La Reacutevolution industrielle et le deacuteveloppement des villes favorisent lrsquoexpansion de la maladie les
conditions de vie insalubres des taudis de banlieues la promiscuiteacute et la pauvreteacute acceacutelegraverent lrsquoendeacutemie
et agrave cette eacutepoque environ une personne sur 4 est atteinte de tuberculose La fin du 19egraveme siegravecle voit se
succeacuteder les deacutecouvertes agrave un rythme effreacuteneacute dont les principales sont la deacutecouverte du bacille en
1882 par Koch la deacutecouverte du bacille bovin en 1896 lrsquoutilisation de la tuberculine dans le diagnostic
humain en 1907 la creacuteation drsquoune souche atteacutenueacutee M bovis BCG en 1921 et le deacutebut de la vaccination
en 1924 La deacutecouverte de la streptomycine en 1944 le premier antibiotique contre la tuberculose
puis de lrsquoisoniazide en 1952 et la rifamycine en 1957 a marqueacute une eacutetape importante dans le traitement
contre la tuberculose (Daniel 2006)
3 La tuberculose bovine
a Introduction
La tuberculose bovine est une maladie bacteacuterienne contagieuse drsquoeacutevolution chronique causeacutee
par Mycobacterium bovis Cette bacteacuterie a pour hocircte originel les bovins mais peut infecter un grand
nombre drsquoespegraveces de mammifegraveres dont lrsquohomme (Gavier-Wideacuten et al 2009 Broughan Crawshaw et
al 2013 Grange amp Yates 1994) ce qui en fait une maladie zoonotique Elle a un tropisme respiratoire
preacutedominant mais peut infecter un grand nombre drsquoautres organes La tuberculose bovine fait partie
des maladies animales agrave deacuteclaration obligatoire aupreacutes de lrsquoOffice International des Epizooties (OIE)
Le passage de lrsquoinfection dans la faune sauvage a compliqueacute le scheacutema drsquoeacuteradication de la
maladie chez les bovins puisque certaines espegraveces sauvages peuvent entretenir la maladie et
contaminer les bovins en retour A lrsquoheure actuelle M bovis fait partie des six organismes pathogegravenes
prioritaires cibleacutes par les programmes de surveillance europeacuteens (Ciliberti et al 2015)
Chapitre 1
22
b Les hocirctes et les communauteacutes drsquohocirctes de Mycobacterium bovis dans le monde
et leurs statuts eacutepideacutemiologiques
Mycobacterium bovis est la mycobacteacuterie du MTBC qui possegravede le plus large spectre drsquohocirctes
incluant lrsquohomme (Grange 2001 OrsquoReilly amp Daborn 1995) Lrsquoinfection a eacuteteacute retrouveacutee chez de
nombreuses espegraveces de mammifegraveres appartenant aux Ordres des marsupiaux des carnivores des
primates des rongeurs des lagomorphes et des artiodactyles (Broughan Downs et al 2013 OrsquoReilly
amp Daborn 1995 Palmer 2013)
La circulation de lrsquoinfection chez des hocirctes appartenant agrave des espegraveces domestiques et sauvages
diffeacuterentes est agrave lrsquoorigine de systegravemes eacutepideacutemiologiques complexes appeleacutes systegraveme multi-hocirctes
(Haydon et al 2002) qui complexifient fortement les scheacutemas drsquoeacuteradication de la maladie Dans ces
systegravemes les diffeacuterents hocirctes peuvent avoir un statut eacutepideacutemiologique diffeacuterent (Haydon et al 2002
Palmer 2013) ou former une communauteacute drsquohocirctes reacuteservoir (Palmer 2013) Les hocirctes de maintenance
ou reacuteservoirs deacutefinissent une population drsquoanimaux dans laquelle lrsquoinfection est entretenue de faccedilon
autonome Ils sont agrave lrsquoorigine de lrsquoinfection des espegraveces cibles Les hocirctes de liaison sont incapables de
maintenir lrsquoinfection indeacutefiniment au sein de leur population agrave moins qursquoils soient reacuteinfecteacutes par une
autre espegravece Ils sont toutefois capables de transmettre lrsquoinfection agrave drsquoautres populations (Corner 2006
Nugent 2011) Et enfin les hocirctes laquo cul-de-sac raquo deacutefinissent les espegraveces qui ne peuvent pas maintenir
ni transmettre la maladie (Nugent 2011) Diffeacuterentes espegraveces ont ainsi eacuteteacute identifieacutees comme des
reacuteservoirs de lrsquoinfection notamment le blaireau (Meles meles) en Angleterre et en Irlande (Delahay et
al 2001) le sanglier (Sus scrofa) dans le centre et le sud de lrsquoEspagne (Naranjo et al 2008) le cerf
eacutelaphe en France (Zanella et al 2008) le cerf de Virginie (Odocoileus virginianus) dans le Michigan
(USA) (Palmer 2013 OrsquoBrien et al 2006) le bison des bois (Bison bison athabascae) et le wapiti (Cervus
elaphus manitobensis) au Canada (Nishi et al 2006 Wobeser 2009) le phalanger renard (Trichosurus
vulpecula) en Nouvelle-Zeacutelande (Morris et al 1994 Nugent 2011) et le buffle (Syncerus caffer) en
Afrique du Sud (Renwick et al 2007) Drsquoautres espegraveces animales ont eacuteteacute identifieacutees comme hocirctes de
liaison comme le furet le cerf et le sanglier en Nouvelle-Zeacutelande
La transmission retour de lrsquoinfection par les espegraveces sauvages reacuteservoir est une menace pour
les bovins et les espegraveces sauvages sensibles vivant agrave proximiteacute La connaissance du statut
eacutepideacutemiologique de chaque espegravece est important pour comprendre le rocircle de la faune sauvage dans la
maintenance et la transmission de lrsquoinfection et adapter les programmes de lutte et drsquoeacuteradication de la
maladie chez les bovins les animaux sauvages et lrsquohomme (Delahay et al 2001 Corner 2006)
Chapitre 1
23
c Aspect zoonotique de Mycobacterium bovis
Plusieurs mycobacteacuteries du MTBC peuvent engendrer une tuberculose zoonotique chez
lrsquohomme Le principal agent infectieux impliqueacute est M bovis (Thoen et al 2006) dans une moindre
mesure M caprae (EFSA 2014) et M microti (Panteix et al 2010) Cette derniegravere a un pouvoir
zoonotique plus faible mais nrsquoest pas neacutegligeable La tuberculose zoonotique agrave M bovis est
majoritairement lieacutee agrave la consommation de produits issus drsquoanimaux infecteacutes principalement le lait cru
et ses deacuteriveacutes (transmission indirecte) (Evans et al 2007) La transmission par aeacuterosols respiratoires est
eacutegalement deacutecrite lors de contacts rapprocheacutes avec des animaux domestiques ou sauvages infecteacutes
(transmission directe) (Cosivi et al 1999 Ayele et al 2004 Kleeberg 1984) Lrsquoinfection est
sporadiquement rapporteacutee chez des eacuteleveurs de bovins et de cerfs (Smith et al 2004 Fanning amp
Edwards 1991) du personnel veacuteteacuterinaire (Shrikrishna et al 2009) et du personnel drsquoabattoir (Liss et al
1994) La transmission interhumaine de M bovis par voie respiratoire existe chez les patients
immunodeacuteprimeacutes ou immunocompeacutetents mais reste rare (Evans et al 2007 Grange amp Yates 1994 de
la Rua-Domenech 2006 Sunder et al 2009 Shrikrishna et al 2009 Rivero et al 2001)
Les signes cliniques chez lrsquohomme sont identiques agrave ceux retrouveacutes lors de tuberculose agrave
M tuberculosis et se traduisent par une tuberculose pulmonaire etou des atteintes extra-pulmonaires
plus freacutequentes qursquoavec M tuberculosis en raison des diffeacuterences dans le mode de transmission
(atteintes osteacuteo-articulaires ganglionnaires uro-geacutenitales disseacutemineacuteeshellip)(Antoine amp Jarlier 2010
Delaune et al 2012 Mignard et al 2006) Le traitement repose sur lrsquoadministration drsquoantituberculeux
classiques agrave lrsquoexception du pyrazinamide pour lequel M bovis est naturellement reacutesistant Des souches
MDR (Multi Drug Resistant) sont reacuteguliegraverement signaleacutees chez lrsquohomme (McLaughlin et al 2012
Cambau et al 2003) De rares souches XDR (Extensively Drug Resistant) sont eacutegalement deacutecrites
majoritairement chez des patients seacuteropositifs (Rivero et al 2001) Dans les pays industrialiseacutes
lrsquoinfection est principalement diagnostiqueacutee chez des personnes acircgeacutees etou immunodeacuteprimeacutees
(diabegravete VIH traitement immunosuppresseur alcoolismehellip) dont les formes extra-pulmonaires
majoritaires eacutevoquent une contamination ancienne par voie digestive (avant la mise en place de la
pasteurisation du lait) et la reacuteactivation ulteacuterieure de la maladie ainsi que chez des personnes neacutees ou
ayant seacutejourneacute dans des pays ougrave la maladie est endeacutemique (Delaune et al 2012)
Les pays ayant mis en place la pasteurisation du lait et des programmes de lutte efficaces contre
la maladie chez les bovins (deacutepistage et abattage) ont fortement reacuteduit lrsquoincidence de la maladie chez
lrsquohomme et les bovins Malgreacute quelques cas reacuteguliegraverement deacutecrits chez lrsquohomme la tuberculose
zoonotique est une zoonose mineure dans les pays deacuteveloppeacutes notamment en Europe (Torgerson amp
Torgerson 2010 Michel et al 2010) En France lrsquoobligation de pasteurisation du lait en 1955 associeacutee
agrave lrsquoabattage systeacutematique des bovins reacuteagissant agrave la tuberculination agrave partir de 1963 ont permis de faire
Chapitre 1
24
baisser lrsquoincidence de lrsquoinfection agrave M bovis chez lrsquohomme de 15 dans les anneacutees 60 agrave 05 en 1995
(Boulahbal et al 1998) Des eacutetudes locales mettent parfois en eacutevidence des preacutevalences plus eacuteleveacutees
de lrsquoordre de 2 agrave Lyon entre 2000 et 2005 (Mignard et al 2006) et de 29 en Aquitaine entre 2004
et 2008 (Aimeacute et al 2012) Au Royaume-Uni par exemple malgreacute lrsquoincidence eacuteleveacutee chez les bovins
(1806 nouveaux foyers entre janvier et avril 2015)(site DEFRA) la preacutevalence de la maladie nrsquoaugmente
pas chez lrsquohomme gracircce aux programmes de lutte mis en place chez lrsquoanimal (Torgerson amp Torgerson
2010 Jalava et al 2007)
La situation est tregraves diffeacuterente dans les pays en voie de deacuteveloppement (PVD) Lrsquoabsence de
programme de lutte contre la tuberculose bovine et la consommation de lait cru dans des pays ougrave la
maladie est endeacutemique chez les bovins notamment dans certains pays drsquoAfrique drsquoAmeacuterique du sud
et drsquoAsie repreacutesentent un risque de santeacute publique important (Cosivi et al 1999 Ayele et al 2004) La
tuberculose zoonotique est souvent fortement relieacutee agrave la preacutevalence de lrsquoinfection bovine dans le pays
ou la reacutegion (Fetene et al 2011) et touche majoritairement les professions en contact avec les bovins
(eacuteleveurs de bovins personnel drsquoabattoir et drsquousines agro-alimentairehellip) et les consommateurs de lait
cru et de fromage frais dont les enfants drsquoautant plus srsquoils sont immunodeacuteprimeacutes Une eacutetude reacutecente
rapporte que la tuberculose zoonotique repreacutesente environ 28 des cas de tuberculose en Afrique
(de 0 agrave 377 selon les pays) 03 des cas sur le continent ameacutericain (de 0 agrave 339 ) et 04 des cas
en Europe (de 0 agrave 211 ) (Muumlller et al 2013) Lrsquoinfection par le HIV la pauvreteacute et le manque drsquoaccegraves
aux soins sont des facteurs de disseacutemination de la tuberculose humaine et zoonotique dans ces pays
(Ayele et al 2004 WHO 2014) Dans les PVD le diagnostic chez lrsquohomme srsquoarrecircte parfois agrave lrsquoobservation
microscopique de BAAR associeacutes agrave M tuberculosis sous-estimant probablement lrsquooccurrence de
M bovis (Cosivi et al 1999 Michel et al 2010) La transmission retour de lrsquoinfection de lrsquohomme aux
bovins pourrait avoir un impact non neacutegligeable dans les PVD mais aucune donneacutee nrsquoest disponible
(de la Rua-Domenech 2006)
d Pathogeacutenie de la tuberculose bovine
La pathogeacutenie de la maladie est multifactorielle et deacutepend de facteurs lieacutes au pouvoir pathogegravene de
M bovis agrave la reacuteceptiviteacute et la sensibiliteacute de lrsquohocircte exposeacute ainsi qursquoagrave la voie drsquoinfection
Facteurs influenccedilant le pouvoir pathogegravene de M bovis voies
drsquoinfection et dose infectante
La voie respiratoire est la voie de contamination majeure chez les bovins (Pritchard 1988
Morris et al 1994 Phillips et al 2003) Des infections expeacuterimentales de M bovis par voie nasale intra
Chapitre 1
25
tracheacuteale ou intra-tonsillaire ainsi que par la mise en contact drsquoanimaux sains avec des animaux
infecteacutes ont confirmeacute lrsquoefficaciteacute de la transmission de M bovis par cette voie (Palmer et al 1999 Dean
et al 2005 Johnson et al 2007 Collins amp Grange 1983 Cassidy et al 1999) La dose infectante par voie
respiratoire est tregraves faible puisque lrsquoinoculation intra-tracheacuteale de 6 agrave 10 bacilles viables agrave des veaux
suffit agrave provoquer des leacutesions pulmonaires etou ganglionnaires (Dean et al 2005 Johnson et al 2007)
Des reacutesultats identiques ont eacuteteacute obtenus chez le blaireau (Corner et al 2007 Corner et al 2011) Chez
le cerf de Virginie lrsquoinoculation intra-tonsillaire de 300 agrave 500 UFC (Uniteacute Formant Colonie) infecte 100
des animaux (Palmer et al 2002 Griffin et al 2006) Lrsquoinoculation oro-pharyngeacutee de 100 UFC agrave des
sangliers deacuteclenche uniquement des leacutesions amygdaliennes alors que 10 000 UFC entrainent des
leacutesions pulmonaires et ganglionnaires seacutevegraveres (Ballesteros et al 2009) Chez toutes ces espegraveces la dose
infectante affecte la seacuteveacuteriteacute et la progression de la maladie (Neill et al 1994 Palmer et al 2002 Corner
et al 2007)
La transmission de la tuberculose bovine peut eacutegalement se faire par voie digestive suite agrave
lrsquoingestion de matrices contamineacutees La dose infectante neacutecessaire serait bien supeacuterieure agrave celle par
voie respiratoire et selon certains auteurs neacutecessiterait plusieurs millions de bacilles (Phillips et al
2003 OrsquoReilly amp Daborn 1995) Pour les veaux Palmer et al (2004) ont estimeacute agrave 5 000 UFC la dose
infectante par cette voie Lrsquoingestion reacutepeacutetitive de faibles doses de M bovis semble eacutegalement
repreacutesenter un facteur de risque comme cela a eacuteteacute deacutemontreacute chez le cochon drsquoInde (Maddock 1934)
La transmission par voie cutaneacutee ou transcutaneacutee ainsi que la transmission de la megravere au veau par
consommation de lait contamineacute ou par leacutechage sont eacutegalement deacutecrites mais restent rares (Phillips et
al 2003 Morris et al 1994) La transmission du bacille par voie transcutaneacutee (morsure) par le biais de
salive infecteacutee est reacuteguliegraverement deacutecrite chez le blaireau en Angleterre (Corner et al 2011)
Facteurs influenccedilant la reacuteceptiviteacute et la sensibiliteacute de lrsquohocircte
De nombreux facteurs intrinsegraveques agrave lrsquoanimal entrent en jeu dans la reacuteceptiviteacute et la sensibiliteacute
de lrsquohocircte agrave la maladie notamment lrsquoespegravece la race lrsquoacircge le sexe et lrsquoeacutetat geacuteneacuteral (Griffin et al 1996
Menzies amp Neill 2000 Humblet et al 2009 Goodchild amp Clifton-Hadley 2001) Certains animaux
preacutesentent une reacutesistance naturelle ou au contraire une sensibiliteacute exacerbeacutee agrave lrsquoinfection ou agrave la
progression de la maladie (Gavier-Wideacuten et al 2009) Des meacutecanismes modulant la reacuteponse
immunitaire et inflammatoire de lrsquohocircte ont pu ecirctre mis en eacutevidence chez le cerf (Mackintosh et al
2000) Les caracteacuteristiques geacuteneacutetiques de lrsquohocircte pourraient eacutegalement jouer un rocircle important dans la
reacutesistance ou la sensibiliteacute agrave lrsquoinfection (Mackintosh et al 2000 Acevedo-Whitehouse et al 2005
Brotherstone et al 2010) Des eacutetudes reacutecentes sur des geacutenomes complets de bovins ont identifieacute des
Chapitre 1
26
loci particuliers impliqueacutes dans la reacutesistance agrave la maladie (Bermingham et al 2014) ouvrant des
perspectives de seacutelection geacutenomique de populations reacutesistantes agrave la tuberculose bovine (Tsairidou et
al 2014)
Le statut immunitaire de lrsquoanimal est un facteur important puisque certains animaux sont
capables drsquoeacuteliminer le bacille gracircce agrave leur immuniteacute inneacutee etou acquise certains sont capables de
controcircler lrsquoinfection plusieurs anneacutees dans les granulomes et drsquoautres deacuteveloppent une tuberculose
eacutevolutive (Pollock amp Neill 2002) La modulation immunitaire expliquerait pourquoi dans un cheptel
bovin malgreacute la contagiositeacute importante de M bovis seuls quelques animaux sont infecteacutes (Cassidy
2006) Les notions de latence et de reacuteactivation de la maladie deacutecrites chez M tuberculosis sont
discuteacutees dans la tuberculose bovine (Cassidy 2006) et commencent agrave ecirctre appreacutehendeacutees (Aacutelvarez et
al 2009)
Les eacutetapes de lrsquoinfection et la reacuteponse immunitaire associeacutee
Lorsque les bacteacuteries peacutenegravetrent dans lrsquoorganisme le plus souvent au niveau des alveacuteoles
pulmonaires elles sont phagocyteacutees par les macrophages Si la dose infectante est importante ou
lrsquoimmuniteacute insuffisante les mycobacteacuteries eacutechappent agrave la digestion et se multiplient permettant la
formation dans le tissu pulmonaire de la leacutesion primaire appeleacutee chancre drsquoinoculation La primo-
infection est caracteacuteriseacutee par le laquo complexe primaire raquo qui associe le chancre drsquoinoculation et les nœuds
lymphatiques infecteacutes par le drainage de la leacutesion primaire (Neill et al 1994) Lrsquoinfection peut ensuite
eacutevoluer selon 3 modes diffeacuterents la gueacuterison avec une eacutelimination des bacilles la stabilisation
caracteacuteriseacutee par une reacuteactivation possible de lrsquoinfection quelques mois ou anneacutees plus tard ou la
geacuteneacuteralisation preacutecoce de lrsquoinfection lieacutee agrave une multiplication active des bacteacuteries et agrave leur
disseacutemination dans lrsquoorganisme Lrsquoinfection et son eacutevolution sont toutefois fortement deacutependantes de
la dose infectante de la voie drsquoentreacutee de la bacteacuterie de la reacuteceptiviteacute et de la sensibiliteacute de lrsquohocircte
eacutevoqueacutees preacuteceacutedemment
Chez lrsquoanimal infecteacute par M bovis la reacuteponse immunitaire agrave meacutediation cellulaire est la premiegravere
agrave se mettre en place dans les stades preacutecoces de lrsquoinfection par la mobilisation de macrophages et de
lymphocytes T (Figure 5) Ces derniers libegraverent diffeacuterentes cytokines dont lrsquointerfeacuteron-γ (IFN-γ) qui
active les macrophages et la formation de granulomes pour isoler et deacutetruire lrsquoagent infectieux
Lrsquoimmuniteacute agrave meacutediation humorale se met en place tardivement et remplace peu agrave peu lrsquoimmuniteacute agrave
meacutediation cellulaire dans les stades plus avanceacutes de la maladie par la production drsquoanticorps par les
lymphocytes B (Pritchard 1988 Ritacco et al 1991 Fifis et al 1994) Les diffeacuterents tests utiliseacutes pour
le deacutepistage de lrsquoinfection chez les bovins reposent principalement sur lrsquoimmuniteacute agrave meacutediation
cellulaire (Figure 5)
Chapitre 1
27
Figure 5 Evolution de la reacuteponse immunitaire chez le bovin infecteacute par M bovis au cours du temps et aux diffeacuterents tests de deacutepistage de la tuberculose bovine (de la Rua-Domenech et al 2006 adapteacute de Vordermeier et al 2004)
Expression clinique de la maladie et leacutesions observeacutees
La tuberculose bovine est une maladie chronique drsquoeacutevolution lente geacuteneacuteralement
asymptomatique dans ses deacutebuts Lorsque la maladie eacutevolue hormis lrsquoamaigrissement possible les
signes cliniques sont tregraves variables et deacutependent des organes ou tissus atteints mais ne sont pas
pathognomoniques (de la Rua-Domenech et al 2006) La localisation des leacutesions observeacutees chez les
animaux deacutepend geacuteneacuteralement de la voie drsquoentreacutee de la bacteacuterie et de lrsquoeacutevolution de la maladie Chez
les bovins la majoriteacute des leacutesions sont retrouveacutees dans la caviteacute thoracique avec une atteinte des lobes
pulmonaires et des nœuds lymphatiques associeacutes (bronchiques et meacutediastinaux) etou des nœuds
lymphatiques ceacutephaliques (reacutetro-pharyngiens principalement) Les leacutesions sur les nœuds lymphatiques
meacutesenteacuteriques la rate et le foie sont plus rares (Neill et al 1994)
Chez le blaireau infecteacute naturellement la seacuteveacuteriteacute de la maladie est tregraves variable drsquoune
infection latente asymptomatique sans leacutesion macroscopique visible agrave une maladie modeacutereacutee avec des
leacutesions pulmonaires ou extra-pulmonaires de petite taille jusqursquoagrave une maladie geacuteneacuteraliseacutee entrainant
la cachexie puis la mort de lrsquoanimal (Murphy et al 2010) Les leacutesions retrouveacutees sont essentiellement
respiratoires probablement lieacutees agrave lrsquoinhalation drsquoaeacuterosols contamineacutes plus rarement digestives et
cutaneacutees suite agrave des morsures de congeacutenegraveres infecteacutes (Corner et al 2011 Payne et al 2012 Gavier-
Widen et al 2001) Toutefois diffeacuterentes eacutetudes ont montreacute que la majoriteacute des blaireaux infecteacutes ne
Chapitre 1
28
preacutesente pas de leacutesion visible (Gallagher amp Clifton-Hadley 2000 Jenkins et al 2008 Payne et al 2012)
Les blaireaux constituent des hocirctes de persitance efficaces puisqursquoils peuvent supporter lrsquoinfection
plusieurs anneacutees et se reproduire tout en excreacutetant la bacteacuterie (Delahay et al 2001)
Chez le sanglier les leacutesions sont majoritairement observeacutees dans les nœuds lymphatiques
ceacutephaliques notamment les mandibulaires dans les poumons et moins freacutequemment au niveau
meacutesenteacuterique (Zanella et al 2008 Martiacuten-Hernando et al 2007 Naranjo et al 2008) Des formes
geacuteneacuteraliseacutees sont eacutegalement deacutecrites (Naranjo et al 2008)
Le cerf est une espegravece deacutecrite comme tregraves sensible agrave lrsquoinfection par M bovis et deacuteveloppe des
leacutesions ouvertes et disseacutemineacutees pouvant ecirctre agrave lrsquoorigine drsquoune excreacutetion bacillaire importante (Lugton
et al 1998 Palmer et al 2004) Les leacutesions sont geacuteneacuteralement localiseacutees au niveau des nœuds
lymphatiques ceacutephaliques et des poumons (Palmer et al 2004 Delahay et al 2002) des nœuds
lymphatiques meacutesenteacuteriques voire geacuteneacuteraliseacutees (Zanella et al 2008)
e Le deacutepistage et le diagnostic de lrsquoinfection chez les animaux
Le deacutepistage de lrsquoinfection chez les bovins vivants repose sur plusieurs modes de surveillance
(Cavalerie et al 2014)
La surveillance systeacutematique agrave lrsquoabattoir concerne tous les bovins abattus pour la consommation
humaine Il repose sur un examen post mortem de lrsquoanimal avec lrsquoinspection drsquoun certain nombre
drsquoorganes cibles de lrsquoinfection (poumons nœuds lymphatiques reacutetro-pharyngiens tracheacuteo-
bronchiques et meacutediastinaux) Dans le cas de leacutesions suspectes visibles celles-ci sont preacuteleveacutees et
transmises agrave un laboratoire agreacuteeacute pour recherche de mycobacteacuteries par PCR et culture
La surveillance programmeacutee en eacutelevage repose sur le deacutepistage de la maladie par prophylaxie Le
rythme des prophylaxies est fixeacute par le preacutefet en fonction de la situation sanitaire du deacutepartement
Dans les deacutepartements ougrave cette surveillance est absente le deacutepistage deacutepend alors uniquement de la
surveillance agrave lrsquoabattoir Pour quelques deacutepartements agrave fort risque elle est annuelle et peut ecirctre
compleacuteteacutee par un deacutepistage lors de mouvements drsquoanimaux Le test de deacutepistage de reacutefeacuterence est
lrsquointradermotuberculination (IDR) Ce test vise agrave mettre en eacutevidence in vivo la reacuteaction
drsquohypersensibiliteacute retardeacutee conseacutecutive agrave lrsquoinjection intradermique de tuberculine (proteacuteines purifieacutees
de surnageant de culture de mycobacteacuteries) chez un bovin infecteacute (Buddle et al 2011 De la Rua-
Domenech et al 2006) Cette reacuteaction se traduit par une tumeacutefaction cutaneacutee douloureuse et chaude
Deux types drsquoIDR sont utiliseacutes lrsquoIntradermotuberculination simple ou IDS qui consiste en lrsquoinjection
intradermique de tuberculine bovine sur le plat de lrsquoencolure et lrsquoIntradermotuberculination
comparative ou IDC qui associe lrsquoinjection intradermique de tuberculine bovine et de tuberculine
aviaire LrsquoIDC est utiliseacutee dans certains deacutepartements dont la Cocircte drsquoOr ougrave de nombreuses
Chapitre 1
29
mycobacteacuteries environnementales sont retrouveacutees pour augmenter la speacutecificiteacute du test et eacutecarter les
reacuteactions croiseacutees chez les bovins sensibiliseacutes agrave une autre mycobacteacuterie (des antigegravenes sont communs
entre les mycobacteacuteries) (De la Rua-Domenech et al 2006 OIE 2009)
LrsquoIDR peut ecirctre compleacuteteacutee par le dosage de lrsquoInterfeacuteron γ (IFNγ) Cette cytokine qui est libeacutereacutee
par les lymphocytes T activeacutes lors drsquoune stimulation antigeacutenique in vivo mais eacutegalement in vitro permet
entre autre lrsquoactivation de la fonction phagocytaire des macrophages La quantiteacute drsquoIFNγ libeacutereacutee lors de
la culture de sang total avec de la tuberculine bovine ou aviaire est suffisante pour ecirctre mesureacutee par
un test ELISA (Gormley et al 2006 de la Rua-Domenech et al 2006) Le dosage de lrsquoIFNγ est pratiqueacute
pour renforcer lrsquoIDR lorsque la contention des animaux est difficile essentiellement en Camargue et
dans les Landes pour les taureaux de combat Il est eacutegalement utiliseacute agrave titre expeacuterimental dans quelques
deacutepartements comme la Dordogne et la Cocircte drsquoOr (Fediaevsky et al 2014) pour renforcer les reacutesultats
drsquoIDR
Drsquoautres tests notamment seacuterologiques sont en cours drsquoeacutevaluation chez les bovins Les essais
reacutealiseacutes jusqursquoagrave maintenant montrent une sensibiliteacute et une speacutecificiteacute infeacuterieures agrave lrsquoIDR ou lrsquoIFNγ
notamment chez les animaux en deacutebut drsquoinfection mais ils repreacutesentent un axe de recherche important
au vu du faible coucirct et de la simpliciteacute de la technique (Waters et al 2011 de la Rua-Domenech et al
2006)
La seacuterologie et lrsquoIFNγ font lrsquoobjet de nombreux deacuteveloppements pour lrsquoameacutelioration de la
surveillance de la maladie chez les animaux sauvages Reacutecemment des techniques ELISA ont eacuteteacute
deacuteveloppeacutees pour de nombreuses espegraveces dont les sangliers les cerfs les bisons (Boadella et al 2011
Shuralev et al 2012 Boadella et al 2012 Lyashchenko et al 2013 Chapinal et al 2012) LrsquoIFNγ est
eacutegalement testeacute pour le deacutepistage chez les espegraveces sauvages notamment chez les cervideacutes drsquoeacutelevage
et les blaireaux (Palmer et al 2010 Dalley et al 2008) La seacuterologie (kit commercial Brock Stat-Pack
Chembio Diagnostic Systems) et le dosage de lrsquoIFNγ (meacutethode ELISA) sont utiliseacutes en routine apregraves
capture et preacutelegravevement sanguin pour le suivi eacutepideacutemiologique de groupes sociaux de blaireaux agrave
Woodchester Park en Angleterre (Drewe et al 2010 King Murphy James Travis Porter Sawyer et al
2015 King Murphy James Travis Porter Hung et al 2015) La combinaison des deux tests est
conseilleacutee pour ameacuteliorer la sensibiliteacute et la speacutecificiteacute du deacutepistage (Drewe et al 2010) Une meacutethode
de deacutepistage non invasive baseacutee sur la collecte puis lrsquoanalyse par qPCR des latrines a eacuteteacute eacutevalueacutee sur
douze groupes sociaux en parallegravele avec la seacuterologie et le dosage de lrsquoIFNγ (King Murphy James Travis
Porter Sawyer et al 2015 King Murphy James Travis Porter Hung et al 2015) Bien que moins
contraignante et moins couteuse que le suivi par capture lrsquoanalyse des latrines pour lrsquoidentification des
groupes sociaux de blaireaux positifs neacutecessite au moins deux campagnes annuelles de preacutelegravevements
(deacutebut et fin drsquoeacuteteacute) avec un nombre drsquoeacutechantillons minimum variable selon les groupes sociaux (de 10
agrave plus de 20) agrave cause de lrsquoexcreacutetion intermittente du bacille par les blaireaux infecteacutes De plus elle ne
Chapitre 1
30
permet pas un deacutepistage individuel
Concernant le diagnostic de la tuberculose bovine lrsquoinfection chez les bovins par M bovis doit
ecirctre eacutetablie par histologie en cas de leacutesion et confirmeacutee par culture selon la reacuteglementation
europeacuteenne Lrsquohistologie est reacutealiseacutee sur des coupes de nœuds lymphatiques et de leacutesions fixeacutees agrave la
formaline La culture repose sur le protocole deacuteveloppeacute et ameacutelioreacute par le Laboratoire National de
Reacutefeacuterence (LNR) devenu la norme NF U47-104 qui associe une phase de broyage-deacutecontamination
des eacutechantillons puis un ensemencement sur deux milieux agrave lrsquoœuf (Loumlwenstein-Jensen et Coletsos) (OIE
2009) Toutefois ces deux techniques preacutesentent des inconveacutenients car lrsquohistologie manque de
speacutecificiteacute (environ 92 ) par rapport agrave la culture (Varello et al 2008) et le reacutesultat de culture est peu
sensible long (entre trois semaines et trois mois) ce qui pose des problegravemes pratiques en cas de
suspicion (restriction des mouvements du cheptel) Bien que pratiqueacutee depuis quelques anneacutees sans
base regraveglementaire il a eacuteteacute deacutemontreacute que la PCR baseacutee sur la deacutetection de lrsquoIS6110 (kit commercial LSI
VetMAXtrade Mycobacterium tuberculosis Complex PCR kit) serait un outil performant de diagnostic de la
tuberculose chez les bovins avec une bonne speacutecificiteacute (97 ) et une sensibiliteacute supeacuterieure agrave la culture
(877 vs 781 ) (Courcoul et al 2014) En apportant un diagnostic en 48 heures la PCR permettrait
une gestion plus rapide des foyers
Le diagnostic dans la faune sauvage repose sur une PCR systeacutematique sur les leacutesions eacutevocatrices
de tuberculose ou un pool de nœuds lymphatiques en lrsquoabsence de leacutesions En cas de reacutesultat non
neacutegatif lrsquoeacutechantillon est mis en culture pour identification de lrsquoagent infectieux (Source Note de service
DGALSDSPA2015-556 du 26062015) Quelle que soit lrsquoespegravece concerneacutee la culture et la PCR sont
reacutealiseacutees par des laboratoires agreacuteeacutes En cas de PCR etou de culture positive les eacutechantillons sont
systeacutematiquement envoyeacutes au Laboratoire National de Reacutefeacuterence pour la tuberculose bovine pour
confirmation et geacutenotypage des souches
f Epideacutemiologie de la tuberculose bovine
Evolution de la tuberculose bovine dans le monde et en
Europe et mesures de lutte
Avant 1920 la tuberculose bovine eacutetait lrsquoune des maladies majeures des animaux domestiques
agrave lrsquoeacutechelle mondiale A lrsquoheure actuelle elle est toujours largement reacutepandue dans le monde (Cosivi et
al 1999) La maladie est plutocirct sporadique et limiteacutee agrave quelques reacutegions dans les pays industrialiseacutes
toucheacutes (sauf en Irlande au Royaume-Uni en Espagne et au Portugal) et endeacutemique dans les pays agrave
faibles revenus drsquoAfrique drsquoAmeacuterique du sud et drsquoAsie (Figure 6)
Chapitre 1
31
Dans les pays industrialiseacutes la mise en place de la pasteurisation du lait dans les anneacutees 1950-
60 et des programmes drsquoeacuteradication de la maladie baseacutes sur le deacutepistage et lrsquoabattage des bovins
infecteacutes ont permis de reacuteduire fortement lrsquoincidence de lrsquoinfection chez les bovins et lrsquohomme
Aujourdrsquohui dans ces pays la tuberculose bovine repreacutesente principalement un enjeu socio-
eacuteconomique lieacute aux restrictions des eacutechanges internationaux drsquoanimaux et aux coucircts des programmes
de lutte contre la maladie chez les animaux le problegraveme sanitaire est devenu mineur Dans les pays agrave
faibles revenus ougrave lrsquoabsence ou quasi-absence de controcircle de la maladie chez les bovins et la
consommation de lait et produits deacuteriveacutes crus les enjeux de la tuberculose bovine sont agrave la fois
sanitaires et socio-eacuteconomiques
Figure 6 Carte de distribution de la tuberculose bovine chez les bovins entre juillet et deacutecembre 2014 (Source WAHID Interface)
En Europe depuis plus drsquoun siegravecle lrsquoeacuteradication de la maladie a eacuteteacute un objectif majeur pour les
organisations agricoles et les autoriteacutes publiques (Reviriego-Gordejo amp Vermeersch 2006) Les mesures
reacuteglementaires eacutelaboreacutees notamment par lrsquoOIE (Office International des Epizooties aujourdrsquohui appeleacute
Organisation Mondiale pour la Santeacute Animale) visent agrave seacutecuriser les eacutechanges commerciaux drsquoanimaux
lrsquoameacutelioration des techniques de deacutepistage de la maladie les programmes drsquoeacuteradication et leurs aides
financiegraveres Les programmes mis en place chez les bovins ont permis de faire fortement diminuer
lrsquoincidence de la maladie dans la plupart des pays de lrsquoUnion Europeacuteenne (de la Rua-Domenech et al
2006) voire de lrsquoeacuteradiquer leur confeacuterant le statut de pays laquo officiellement indemne de tuberculose
bovine raquo (OTF Officially Tuberculosis Free) Le statut OTF est attribueacute agrave un pays lorsque
- le pourcentage de cheptels bovins confirmeacutes comme infecteacutes nrsquoexcegravede pas 01 par an pour
tous les cheptels pendant 6 ans
Chapitre 1
32
- chaque bovin est identifieacute en accord avec la regraveglementation europeacuteenne
- tous les bovins abattus subissent un examen post-mortem officiel (Reviriego-Gordejo amp
Vermeersch 2006)
Ce statut est tregraves important pour lrsquoeacuteconomie drsquoun pays puisqursquoil facilite le commerce national
et international des animaux par un allegravegement des controcircles sanitaires avant lrsquoexportation de leurs
bovins dans un autre pays membre ou avant leur abattage en les exemptant notamment drsquoune
tuberculination (Reviriego-Gordejo amp Vermeersch 2006) Les pays europeacuteens officiellement indemnes
de tuberculose sont lrsquoAllemagne lrsquoAutriche la Belgique le Danemark lrsquoEstonie la Finlande la France
lrsquoEcosse la Lettonie le Liechtenstein le Luxembourg la Norvegravege les Pays-Bas la Pologne la
Reacutepublique Tchegraveque la Slovaquie la Sloveacutenie la Suegravede la Suisse et une partie de lrsquoItalie (Figure 7) Les
pays drsquoEurope les plus toucheacutes sont la Grande-Bretagne lrsquoIrlande et lrsquoEspagne avec des preacutevalences
respectives de 104 44 et 12 en 2012 (EFSA 2014) Toutefois dans certains pays OTF comme la
France la Belgique lrsquoAllemagne lrsquoAutriche et la Pologne lrsquoeacuteradication de la maladie nrsquoest pas complegravete
malgreacute la preacutevention les controcircles et la surveillance engageacutes Dans ces pays lrsquoeacuteradication de la maladie
est fortement compliqueacutee par la circulation de lrsquoinfection dans des reacuteservoirs sauvages source
potentielle de contamination pour les bovins (Corner 2006 Nugent 2011 Gortaacutezar et al 2012)
Figure 7 Statuts des pays europeacuteens vis-agrave-vis de la tuberculose bovine en 2012 et proportion de bovins infecteacutes OTF Officially Tuberculosis Free (officiellement indemne de tuberculose bovine) MS Member State (Etat membre) (Source EFSA 2014)
Chapitre 1
33
Evolution de la situation sanitaire en France
Chez les bovins
Dans les anneacutees 50 environ 25 des cheptels bovins franccedilais eacutetaient infecteacutes par la tuberculose
bovine (Beacutenet et al 2006) Cette preacutevalence importante avait un fort impact eacuteconomique (saisies de
viande agrave lrsquoabattoir diminution de la productiviteacute en viande et en lait) mais aussi sanitaire puisque 2 agrave
11 des cas de tuberculose chez lrsquohomme eacutetaient imputeacutes agrave M bovis Les programmes de lutte mis en
place ont permis de faire diminuer de faccedilon spectaculaire la preacutevalence cheptel de la maladie de 25
en 1955 agrave moins de 01 degraves 1998 permettant agrave la France drsquoacceacuteder au statut OTF en 2000 (Figure 8)
Ce statut a permis drsquoalleacuteger voire de stopper la prophylaxie sur le territoire drsquoalleacuteger les controcircles mis
en place pour les mouvements de bovins dans lrsquoUE et de fournir une garantie sanitaire aux pays
importateurs de bovins franccedilais dans lrsquoUE et en dehors
Les programmes de lutte contre la tuberculose bovine reposent sur une base reacuteglementaire en
eacutevolution constante eacutelaboreacutee par les services veacuteteacuterinaires (maintenant DDPP Direction
Deacutepartementale de la Protection des Populations) en collaboration technique avec les veacuteteacuterinaires
sanitaires les groupements de deacutefense sanitaire (GDS) et les laboratoires drsquoanalyse agreacuteeacutes Ces
programmes sont baseacutes sur la surveillance systeacutematique et geacuteneacuteraliseacutee en abattoir (inspection post-
mortem recherche et identification de mycobacteacuteries par culture et PCR sur les leacutesions suspectes) et
sur la surveillance programmeacutee en eacutelevage (deacutepistage par intradermo-tuberculination et abattage total
ou partiel des troupeaux infecteacutes) Ils assurent eacutegalement la protection des cheptels indemnes par la
maitrise des facteurs de diffusion de la tuberculose bovine tels que lrsquointroduction drsquoun animal le
voisinage avec des cheptels infecteacutes et la reacutesurgence (Fediaevsky et al 2014) Depuis 2013 deux
Cellules Inter Reacutegionales drsquoEpideacutemiologie Veacuteteacuterinaire (CIREV) ont eacuteteacute creacuteeacutees en Bourgogne et en
Aquitaine afin de renforcer les investigations eacutepideacutemiologiques pour adapter de faccedilon optimale les
mesures de lutte contre la maladie (Fediaevsky et al 2013)
Chapitre 1
34
Figure 8 Evolution du taux de preacutevalence annuelle national de la tuberculose bovine chez les bovins (Beacutenet et al 2006)
Depuis 2004 la France connait une situation preacuteoccupante vis-agrave-vis de la tuberculose bovine
car lrsquoaugmentation du taux de preacutevalence met en danger le statut OTF du pays (Figure 9) Cette
augmentation est lieacutee agrave lrsquoapparition de nouveaux foyers de tuberculose concentreacutes dans des zones
geacuteographiques bien localiseacutees la Cocircte drsquoOr la Dordogne et les Pyreacuteneacutees Atlantiques cumulent 75
des foyers et dans une moindre mesure la Charente la Camargue les Hautes Pyreacuteneacutees lrsquoAriegravege les
Landes et la Corse (Figure 10) Dans ces deacutepartements caracteacuteriseacutes par de lrsquoeacutelevage allaitant
preacutedominant la contamination des troupeaux par le voisinage (par contacts directs avec des bovins
voisins ou des animaux sauvages infecteacutes ou par contacts indirects avec du mateacuteriel ou des matrices
contaminants) et lrsquointroduction drsquoun animal infecteacute dans le cheptel (notamment par lrsquoachat drsquoanimaux
le precirct et la pension) repreacutesentaient la moitieacute des cas en 2011 La reacutesurgence drsquoune ancienne infection
(qui persiste agrave bas bruit chez les animaux ou dans des matrices environnementales souilleacutees) et la faune
sauvage eacutetaient agrave lrsquoorigine preacutesumeacutee drsquoenviron 10 des cas respectivement toutefois sans
deacutemonstration scientifique Pour 35 des foyers bovins lrsquoorigine de lrsquoinfection eacutetait inconnue
(Fediaevsky et al 2012) La contamination des bovins via le voisinage la reacutesurgence et les foyers
drsquoorigine inconnue soulegravevent la question du rocircle de la transmission indirecte de M bovis entre la faune
sauvage et les bovins et donc le rocircle potentiel de lrsquoenvironnement comme reacuteservoir du bacille
25
15
10
5
20
1955
1963
1960
1970
1980
1990
2000
Chapitre 1
35
Figure 9 Evolution de la preacutevalence et de lrsquoincidence de la tuberculose bovine en France entre 1995 et 2014 (Cavalerie et al 2014)
La menace de la perte du statut OTF inquiegravete les pouvoirs publics et les instances agricoles car
la France est le plus gros exportateur de bovins vivants de lrsquoUE elle exporte principalement des bovins
maigres en Italie et en Espagne pour lrsquoengraissement (environ 12 million de tecirctes) mais aussi des gros
bovins destineacutes agrave la boucherie (220 000 tecirctes) et des veaux (128 000 tecirctes) (Source France AgriMer
les filiegraveres de lrsquoeacutelevage franccedilais eacutedition 2013) Les conseacutequences eacuteconomiques sur la filiegravere-
agroalimentaire lieacutees au blocage des animaux seraient consideacuterables De plus la gestion sanitaire de la
tuberculose bovine est tregraves coucircteuse puisque environ 20 millions drsquoeuros sont deacutepenseacutes chaque anneacutee
agrave lrsquoeacutechelle nationale pour lrsquoindemnisation des eacuteleveurs et la reacutealisation des analyses veacuteteacuterinaires
(Fediaevsky et al 2014)
Chapitre 1
36
Figure 10 Incidence et distribution de la tuberculose bovine en France dans les anneacutees 50 (en haut) et entre 2000 et 2014 (en bas) (Cavalerie et al 2014)
Chapitre 1
37
Dans la faune sauvage
En France agrave lrsquoheure actuelle la faune sauvage infecteacutee nrsquoest deacutetecteacutee qursquoagrave proximiteacute de foyers
bovins Le 1er foyer de tuberculose bovine dans la faune sauvage a eacuteteacute deacutecouvert en 2001 dans la forecirct
de Brotonne (Seine-Maritime et Eure) sur des cervideacutes puis des sangliers (Hars et al 2010 Zanella et
al 2008) Lrsquoaggravation de la situation sanitaire en 2005-2006 a neacutecessiteacute un abattage total des cerfs
consideacutereacutes comme reacuteservoir primaire de lrsquoinfection ainsi qursquoune diminution de la densiteacute des sangliers
reacuteservoir secondaire (Zanella et al 2008) Drsquoautres deacutepartements comme la Cocircte drsquoOr la Corse les
Pyreacuteneacutees Atlantiques la Dordogne et la Charente notamment ont par la suite deacutetecteacute des animaux
sauvages infecteacutes essentiellement blaireaux sangliers et dans une moindre mesure cervideacutes avec
toutefois des situations eacutepideacutemiologiques tregraves diffeacuterentes selon les reacutegions toucheacutees Ces situations
deacutependent de la population animale impliqueacutee (espegraveces densiteacutes interactionshellip) et de
lrsquoenvironnement (climat paysagehellip) Dans tous les cas le geacutenotypage (spoligotype et VNTR) des
souches incrimineacutees dans la faune sauvage a montreacute qursquoelles eacutetaient identiques agrave celles retrouveacutees
dans les eacutelevages bovins voisins suggeacuterant un lien eacutepideacutemiologique entre cas domestiques et sauvages
(Zanella et al 2008 Payne et al 2012 Richomme et al 2010)
Depuis 2011 un reacuteseau de surveillance eacutepideacutemiologique de la tuberculose bovine dans la faune
sauvage appeleacute Sylvatub a eacuteteacute mis en place en France Les diffeacuterentes modaliteacutes de surveillance ont
pour but la deacutetection de lrsquoinfection chez les animaux sauvages dans les zones agrave risque et preacutesumeacutees
indemnes ainsi que le suivi de son eacutevolution afin de proposer des mesures de lutte aux autoriteacutes
sanitaires (Riviegravere et al 2011) La surveillance est baseacutee sur un classement du territoire national en 3
niveaux de risque Pour les zones situeacutees en risque 1 (risque faible correspondant agrave une absence de
bovins deacutetecteacutes) il est demandeacute aux chasseurs de signaler toute leacutesion suspecte lors de lrsquoexamen initial
de la venaison Concernant les zones ougrave il existe un risque aveacutereacute (niveau 2 ou 3) la surveillance peut
ecirctre agrave la fois eacuteveacutenementielle (ramassage et analyse des cadavres drsquoanimaux trouveacutes morts) etou
programmeacutee (analyse des cerfs et sangliers tueacutes agrave la chasse et les blaireaux pieacutegeacutes) Les reacutesultats des
analyses effectueacutees en 2013-2014 sont preacutesenteacutes dans la Figure 11
Chapitre 1
38
Figure 11 Reacutepartition et reacutesultats des analyses effectueacutees dans le cadre du dispositif Sylvatub chez le grand gibier drsquoaoucirct 2013 agrave aoucirct 2014 et chez les blaireaux en 2014 (Source Sylvatub)
Evolution de la situation sanitaire en Cocircte drsquoOr
Quelques geacuteneacuteraliteacutes sur lrsquoeacutelevage bovin en Cocircte drsquoOr
La Cocircte drsquoOr est un deacutepartement drsquoeacutelevage bovin principalement allaitant (231 270 tecirctes en 2013
dont 74 565 vaches allaitantes et 15 687 vaches laitiegraveres) Lrsquoeacutelevage allaitant est majoritairement
concentreacute dans les zones herbagegraveres de lrsquoouest du deacutepartement et lrsquoeacutelevage laitier sur le plateau de
Langres au nord de Dijon Lrsquoeacutelevage allaitant est extensif (les bovins sont au preacute drsquoavril agrave octobre) lieacute agrave
la production de broutards maigres destineacutes agrave ecirctre engraisseacutes principalement en Italie En Cocircte drsquoOr le
parcellaire des exploitations de bovins allaitant est souvent tregraves morceleacute
Situation eacutepideacutemiologique chez les bovins et mesures de lutte associeacutees
Historiquement la Cocircte drsquoOr est un deacutepartement en prise avec la tuberculose depuis de
nombreuses anneacutees (Figure 10) A la fin des anneacutees 50 le taux de preacutevalence annuel chez les bovins
atteignait 65 Les mesures de lutte collective mises en place ont permis drsquoabaisser ce taux en dessous
du seuil des 10 en 1969 puis de le faire diminuer lentement et reacuteguliegraverement jusqursquoen 1985 ougrave la
maladie semblait eacuteradiqueacutee A la fin des anneacutees 90 les mesures prophylactiques ont cesseacute dans le
Chapitre 1
39
deacutepartement et la vigilance vis-agrave-vis de la maladie a baisseacute (Communication personnelle DDPP 21)
Les premiers signes drsquoune nouvelle eacutepizootie sont apparus en 2002 sur le canton de Pouilly-en-
Auxois (zone Sud) puis en 2003 dans ceux de Venarey-Les-Laumes et Vitteaux (zone Nord) (Source
DDPP21) Localiseacutee agrave quelques eacutelevages agrave son deacutebut la zone de lrsquoinfection srsquoest peu agrave peu eacutelargie au
cours du temps Un plan de prophylaxie restreint aux communes infecteacutees a eacuteteacute mis en place en 2002
puis eacutelargi aux cantons en 2003 avec de plus une surveillance des cheptels en lien eacutepideacutemiologique
avec les troupeaux infecteacutes et ceux des communes dans lesquelles les bovins infecteacutes avaient patucircreacute
Mais ces mesures nrsquoont pas suffi agrave limiter la progression de lrsquoinfection puisque limiteacute agrave moins de 5 par
an jusqursquoen 2006 le nombre de foyers est passeacute de 11 en 2007 agrave 17 en 2009 (Figure 13) Le
renforcement de la lutte comprenant notamment le deacutepistage annuel de tous les bovins de plus de 12
mois dans tout le deacutepartement a permis la deacutecouverte de 45 foyers en 2010 signe que lrsquoinfection
circulait agrave bas bruit De 2011 agrave 2013 le nombre de foyers a eacuteteacute diviseacute par 2 et pour lrsquoanneacutee 2014 10
foyers seulement ont eacuteteacute mis en eacutevidence ce qui semble conforter une maitrise de lrsquoeacutepizootie (Figure
13)
En Cocircte drsquoOr les foyers bovins sont circonscrits agrave une zone geacuteographique bien deacutelimiteacutee dans le
deacutepartement et partagent le mecircme spoligotype M bovis spoligotype SB0134 dans la zone Sud et
M bovis spoligotype SB0120 dans la zone Nord (Figure 12) Depuis 3 ans toutefois les deux zones ont
tendance agrave se teacutelescoper agrave cause notamment de la deacutecouverte drsquoanimaux sauvages infecteacutes avec
M bovis SB0120 en zone Sud Les deux spoligotypes ont deacutejagrave eacuteteacute retrouveacutes simultaneacutement dans des
cheptels infecteacutes Ces observations associeacutees agrave lrsquoabsence drsquoinstallation peacuterenne de la maladie dans des
zones du deacutepartements ougrave les concentrations de bovins sont aussi eacuteleacuteveacutees que dans la zone infecteacutee
soulegravevent la question de lrsquoexistence de facteurs locaux assurant lrsquoentretien et la propagation de
lrsquoinfection entre les eacutelevages bovins Au vu du morcellement et de lrsquoimbrication des pacirctures la
contamination par voisinage est probablement le facteur primordial chez les bovins Une eacutetude reacutecente
(de type cas-teacutemoin) reacutealiseacutee dans des cheptels de Cocircte drsquoOr de Dordogne et des Ardennes a montreacute
que les contacts de voisinage entre troupeaux eacutetaient le facteur de risque principal drsquoinfection des
troupeaux (Marsot et al Article soumis) Toutefois la deacutetection drsquoanimaux sauvages agrave proximiteacute de
certains foyers bovins interroge sur le rocircle potentiel de la faune sauvage infecteacutee dans la contamination
des bovins et dans la recontamination rapide de plusieurs cheptels apregraves abattage total des bovins et
vide sanitaire (Figure 13) (ANSES 2011)
Chapitre 1
40
Figure 12 Localisation du parcellaire des cheptels infecteacutes par la tuberculose bovine entre 2010 et 2015 et spoligotypes incrimineacutes (Source CIREV)
Figure 13 Nombre de nouveaux foyers bovins et de foyers recontamineacutes entre 2002 et 2014 (Donneacutees DDPP21)
0
5
10
15
20
25
30
35
40
45
50
2002 2003 2004 2005 2006 2007 2008 2009 2010 2011 2012 2013 2014
No
mb
re d
e fo
yers
bo
vin
s
Foyers contamineacutes pour la 1egravere fois Foyers recontamineacutes
Chapitre 1
41
Situation eacutepideacutemiologique dans la faune sauvage et mesures de lutte associeacutees
Le premier animal sauvage infecteacute une biche a eacuteteacute deacutecouvert en 2003 agrave proximiteacute de 2 foyers
bovins de 2002 dans la zone Sud Entre 2003 et 2007 la surveillance active de la faune sauvage mise
en place et porteacutee par lrsquoONCFS (Office National de la Chasse et de la Faune Sauvage) et la DDPP et sur
le terrain par les chasseurs les pieacutegeurs et les veacuteteacuterinaires a permis de mettre en eacutevidence lrsquoinfection
chez 1 cervideacute (n=284) 2 sangliers (n= 160) mais aucun blaireau (n=63) Depuis 2007 des sangliers
infecteacutes sont retrouveacutes tous les ans notamment de jeunes animaux avec des leacutesions eacutevolutives
majoritairement dans la zone Sud ougrave les populations sont plus importantes Depuis 2009 lrsquoinfection
est retrouveacutee chez des blaireaux vivant agrave proximiteacute des cheptels infecteacutes majoritairement dans la zone
Nord (Hars Richomme and Boschiroli 2010 DDPP21)(Figure 14) La maladie circule donc dans un
systegraveme multi-hocirctes complexe et unique incluant les bovins les blaireaux les sangliers et les cervideacutes
(Payne 2014)
Figure 14 Nombre de cervideacutes sangliers et blaireaux infecteacutes deacutecouverts en Cocircte drsquoOr entre 2002 et 2014
Dans la crainte de lrsquoeacutetablissement drsquoun reacuteservoir sauvage chez le sanglier comme en Espagne
(Naranjo et al 2008) ou le blaireau au Royaume-Uni (Delahay et al 2001) des mesures de diminution
des densiteacutes ont eacuteteacute mises en place (augmentation des tableaux de chasse pour le sanglier et les
cervideacutes et intensification du pieacutegeage des blaireaux) le ramassage des viscegraveres drsquoanimaux tueacutes agrave la
chasse a eacuteteacute rendu obligatoire et lrsquoagrainage interdit
0
5
10
15
20
25
30
35
No
mb
re d
an
imau
x in
fect
eacutes
Cervideacutes Sangliers Blaireaux
Chapitre 1
42
C ECOLOGIE DE M BOVIS DANS LrsquoENVIRONNEMENT
1 Introduction
Avant les anneacutees 30 il eacutetait admis sine qua none qursquoen tant que bacteacuteries intracellulaires
obligatoires les bacteacuteries tuberculeuses ne survivaient pas en dehors de leur hocircte Toutefois degraves le
deacutebut du 20egraveme siegravecle la deacutetection de Bacillus tuberculosis virulent dans lrsquoeau et les biofilms des
canalisations drsquoeffluents de diffeacuterents sanatoria en Allemagne et au Royaume-Uni (Musehold en 1900
citeacute par Jensen 1954 et Conroy et al en 1921 et 1922 et Cummins et Ackland en 1929 citeacutes par (Williams
amp Hoy 1930) a souleveacute la question du rocircle potentiel de lrsquoenvironnement dans lrsquoeacutepideacutemiologie des
mycobacteacuteries tuberculeuses Concernant la tuberculose bovine les diffeacuterentes recherches meneacutees
pour deacutetecter et caracteacuteriser la persistance de M bovis dans lrsquoenvironnement ont pour objectif
principal de preacuteciser le rocircle de la transmission indirecte dans leacutepideacutemiologie de la maladie La
transmission indirecte semble en effet prendre toute son importance lorsque des reacuteservoirs sauvages
sont identifieacutes etou des systegravemes multi-hocirctes car ils ont la capaciteacute de transmettre lrsquoinfection aux
bovins (Corner 2006 Payne 2014)
Reacutepondre agrave cette question neacutecessite de connaitre les modaliteacutes de transfert des bacteacuteries de
lrsquoanimal ou lrsquohomme infecteacute vers lrsquoenvironnement la dureacutee de survie dans les diffeacuterentes matrices les
strateacutegies de survie de la bacteacuterie ainsi que les facteurs qui affectent sa survie
2 Origines de la contamination environnementale par M bovis
a Lrsquoexcreacutetion animale de Mycobacterium bovis
Lrsquohomme et les animaux infecteacutes ont la capaciteacute drsquoexcreacuteter le bacille via diffeacuterentes voies et
constituent donc une source de contamination potentielle pour lrsquoenvironnement Comme nous lrsquoavons
indiqueacute preacuteceacutedemment les voies drsquoexcreacutetion ainsi que les niveaux drsquoexcreacutetion deacutependent de la
localisation des leacutesions et de leur seacuteveacuteriteacute Les tableaux leacutesionnels deacutecrits preacuteceacutedemment indiquent
que M bovis est susceptible drsquoecirctre excreacuteteacutee principalement par voie orale ou nasale lors drsquoatteinte
pulmonaire etou des nœuds lymphatiques ceacutephaliques par voie digestive via les matiegraveres feacutecales et
par voie urinaire lors drsquoinfection geacuteneacuteraliseacutee (Phillips et al 2003 Corner et al 2012) Lrsquoexcreacutetion feacutecale
de bacteacuteries du MTBC en lrsquoabsence de leacutesions gastro-intestinales semble majoritairement lieacutee au
passage de mucus respiratoire chargeacute en bacilles dans le tube digestif La reacutesistance aux pH acides
(Vandal et al 2008) des bacteacuteries du MTBC leur permet de reacutesister au passage du tractus digestif et
drsquoecirctre excreacuteteacutees sous forme viable dans les matiegraveres feacutecales comme cela a eacuteteacute deacutemontreacute pour M bovis
dans certaines espegraveces animales (Corner Murphy and Gormley 2011 Gallagher and Clifton-Hadley
Chapitre 1
43
2000 Gavier-Widen et al 2001) pour M tuberculosis (El Kheacutechine et al 2009) et M africanum
(Cadmus et al 2009)
Chez les bovins lrsquoexcreacutetion du bacille a eacuteteacute deacutemontreacutee dans le mucus nasal et tracheacuteal et les
fegraveces drsquoanimaux infecteacutes expeacuterimentalement (Neill et al 1988 McCorry et al 2005 Phillips et al 2003
Maddock 1936) Le deacutelai drsquoexcreacutetion apregraves lrsquoinoculation la dureacutee de lrsquoexcreacutetion (plusieurs semaines) et
la quantiteacute de bacilles excreacuteteacutes sont proportionnels agrave la dose infectieuse utiliseacutee (Neill et al 1988)
Lrsquoexcreacutetion feacutecale apparait peu freacutequente et intermittente chez les bovins (Neill et al 1988 Goodchild
amp Clifton-Hadley 2001) Dans les pays industrialiseacutes ougrave le deacutepistage des animaux infecteacutes est en place
lrsquoexcreacutetion de M bovis dans le lait les fegraveces et les urines est consideacutereacutee comme insignifiante chez les
bovins (Morris et al 1994)
Chez le blaireau infecteacute naturellement ou expeacuterimentalement M bovis est excreacuteteacutee par 25 agrave
50 des animaux par voie respiratoire (mucus tracheacuteal et aeacuterosols) urinaire et feacutecale ainsi que par la
suppuration drsquoabcegraves reacutesultant de morsures (Corner et al 2011 Delahay et al 2000 Clifton-Hadley et
al 1993) Lrsquoisolement de M bovis dans des glandes salivaires de blaireaux suggegravere que la salive
constitue eacutegalement une voie possible drsquoexcreacutetion du bacille (Payne 2014) Les blaireaux preacutesentant un
stade avanceacute de la maladie et des leacutesions macroscopiques semblent ecirctre les plus excreacuteteurs en terme
de dureacutee et drsquointensiteacute drsquoexcreacutetion (Corner et al 2012 Gallagher amp Clifton-Hadley 2000 Jenkins et al
2008) toutefois lrsquoexcreacutetion du bacille a eacuteteacute deacutecrite chez des animaux sans leacutesions visibles (Jenkins et al
2008 Gavier-Wideacuten et al 2009) Dans les stades avanceacutes de la maladie les quantiteacutes excreacuteteacutees peuvent
atteindre 2105 UFCmL1 de mucus bronchique 25105 UFCmL1 drsquourine et 75104 UFCg1 de fegraveces
(Gallagher et al 1998) Le bacille est excreacuteteacute par voie feacutecale de 165 agrave 1305 jours avant la mort de
lrsquoanimal (Little et al 1982) Dans cette espegravece lrsquoexcreacutetion est geacuteneacuteralement intermittente mais des
individus laquo super excreacuteteurs raquo ont eacuteteacute deacutecrits et se caracteacuterisent par une excreacutetion persistante dans le
temps ou par lrsquoexcreacutetion du bacille par plusieurs voies simultaneacutement (Delahay et al 2000) Des
blaireaux laquo super excreacuteteurs raquo ont eacuteteacute mis en eacutevidence en Cocircte drsquoOr la voie feacutecale eacutetant toujours
positive chez ces animaux (Payne 2014) Lrsquointensiteacute de lrsquoexcreacutetion semble varier avec lrsquoacircge (les adultes
excregravetent plus que les jeunes) et la saison puisque lrsquoexcreacutetion semble supeacuterieure au printemps et en
eacuteteacute (Delahay et al 2013 King Murphy James Travis Porter Hung et al 2015)
Chez le sanglier lrsquoexcreacutetion de M bovis se produit par voie orale nasale feacutecale et urinaire
(Martiacuten-Hernando et al 2007 Santos Almeida et al 2015 Barasona et al 2015) Lrsquoexcreacutetion salivaire
est encore discuteacutee (Payne 2014) En Espagne lrsquoexcreacutetion par au moins une des voies a eacuteteacute mise en
eacutevidence par PCR sur 30 des sangliers analyseacutes (n = 39) et 25 des 39 animaux excreacutetaient par les trois
voies (Barasona et al 2015) Lrsquoexcreacutetion eacutetait majoritairement associeacutee agrave une maladie geacuteneacuteraliseacutee Au
Portugal sur 39 sangliers adultes ou subadultes infecteacutes analyseacutes 80 excreacutetaient le bacille par au
moins une voie et 13 eacutetaient des laquo super excreacuteteurs raquo (excreacutetant par trois voies diffeacuterentes) Environ
Chapitre 1
44
28 des sangliers excreacutetaient des quantiteacutes de bacilles supeacuterieures agrave 1 000 UFC par g ou mL drsquoexcreacuteta
par au moins une voie (Santos Almeida et al 2015) Chez le sanglier agrave notre connaissance lrsquoexcreacutetion
nrsquoa jamais eacuteteacute deacutecrite en lrsquoabsence de leacutesions
Chez les cervideacutes diffeacuterentes eacutetudes ont montreacute que le bacille eacutetait excreacuteteacute de faccedilon tregraves
sporadique dans les mucus nasal oro-pharyngeacute et tracheacuteal ainsi que dans les matiegraveres feacutecales lors
drsquoinfections naturelles ou expeacuterimentales (Lugton et al 1998 Santos Almeida et al 2015 Palmer et
al 2001) Dans cette espegravece une excreacutetion intermittente apparait degraves les premiers stades de la
maladie la seacuteveacuteriteacute des tableaux leacutesionnels rencontreacutes explique le niveau drsquoexcreacutetion eacuteleveacute (Palmer et
al 2001 Santos Almeida et al 2015)
Les carcasses drsquoanimaux infecteacutes peuvent constituer une source drsquoinfection directe pour les
animaux charognards qui srsquoen nourrissent mais eacutegalement une source de contamination indirecte pour
les bovins qui pacircturent sur le site de deacutecomposition de la carcasse (Morris et al 1994) Concernant les
animaux tueacutes agrave la chasse lrsquoabandon des viscegraveres ou leur ensevelissement repreacutesente eacutegalement un
risque pour les animaux charognards De ce fait en France la collecte et lrsquoeacutelimination des viscegraveres des
espegraveces sensibles sont obligatoires dans les zones toucheacutees par la tuberculose bovine
b Le transfert de Mycobacterium bovis par les effluents drsquoeacutelevage ou urbains
Les eacutetudes sur la contamination potentielle des effluents drsquoeacutelevage avec M bovis sont tregraves
rares probablement agrave cause de la difficulteacute agrave cultiver le bacille dans ces matrices complexes Les
multiples voies drsquoexcreacutetion du bacille et plus particuliegraverement lrsquoexcreacutetion par voie feacutecale mecircme si elle
est intermittente peuvent probablement contribuer agrave la contamination des effluents drsquoeacutelevage bovin
tels que les fumiers lisiers et eaux useacutees Lrsquoeacutepandage de ces effluents sur les pacirctures ou lrsquoirrigation des
cultures peut constituer un vecteur de contamination du sol des cultures et de lrsquoeau profonde ou de
surface et repreacutesenter un risque pour les animaux et lrsquohomme mais aucune donneacutee chiffreacutee nrsquoexiste
pour ces matrices en conditions naturelles Lrsquoabattage et la deacutecoupe de bovins tuberculeux peut
eacutegalement repreacutesenter une source de contamination des rejets drsquoabattoir (Jensen 1954) Au vu des
mesures actuelles de collecte stockage traitement et controcircle des effluents drsquoabattoir cette
contamination semble tregraves peu probable en France Toutefois elle reste complegravetement valable dans
les zones drsquoendeacutemie des pays nrsquoappliquant pas ces mesures et ne disposant pas de structures
drsquoabattage adeacutequates
Il est inteacuteressant de noter que quelques eacutetudes sont disponibles concernant la contamination
drsquoeffluents urbains avec M tuberculosis Jensen (1954) a montreacute que les effluents bruts de sanatoria
de 5 villes des Pays-Bas les effluents purifieacutes les boues digeacutereacutees et les boues seacutecheacutees (preacuteleveacutees 5
semaines apregraves lrsquoeacutepandage) contenaient des bacilles virulents de M tuberculosis Plus reacutecemment la
Chapitre 1
45
mise en culture de 800 eacutechantillons drsquoeau et 700 eacutechantillons de sol preacuteleveacutes dans 3 quartiers de
Teacuteheacuteran (Iran) entre 2012 et 2014 a deacutemontreacute la preacutesence de M tuberculosis dans 71 des eacutechantillons
drsquoeau (88) et 11 eacutechantillons de sol (15) Les eacutechantillons drsquoeau contamineacutes ont eacuteteacute retrouveacutes
principalement dans les rigoles de ciment acheminant les eaux useacutees et pluviales (5671) et dans les
eaux traiteacutees (1571) La bacteacuterie nrsquoa toutefois pas eacuteteacute isoleacutee du reacuteseau drsquoeau potable Environ 10 des
souches isoleacutees eacutetaient multi-reacutesistants aux antibiotiques (Velayati et al 2015)
3 Deacutetection et survie de Mycobacterium bovis dans lrsquoenvironnement
a Deacutetection de Mycobacterium bovis en conditions naturelles
A ce jour peu drsquoeacutetudes concernant la deacutetection de M bovis dans des substrats naturellement
contamineacutes ont eacuteteacute publieacutees La deacutetection de M bovis dans des eacutechantillons environnementaux
naturellement contamineacutes gracircce agrave des meacutethodes baseacutees sur la culture est exceptionnelle A
Woodchester Park dans le Gloucestershire (Sud-Ouest de lrsquoAngleterre) ougrave les densiteacutes de blaireaux
sont eacuteleveacutees et la preacutevalence importante (20 en 2000 et 35 en 2005) un suivi longitudinal du statut
des populations est reacutealiseacute depuis 1980 (Delahay et al 2000) et diffeacuterentes campagnes de
preacutelegravevements de sols de terriers et de latrines ont eacuteteacute reacutealiseacutees Le traitement drsquoeacutechantillons de sol et
latrines provenant de Woodchester Park traiteacute par immunocapture magneacutetique a permis pour la
premiegravere et seule fois lrsquoisolement de M bovis par culture dans des eacutechantillons environnementaux
(Sweeney et al 2006 Sweeney et al 2007) Cette technique est toutefois peu applicable agrave un grand
nombre drsquoeacutechantillons en termes de coucirct et de temps et nrsquoa jamais eacuteteacute reacuteutiliseacutee par la suite
Drsquoautres tentatives de deacutetection de M bovis par diffeacuterentes meacutethodes baseacutees sur la culture
dans des zones endeacutemiques de tuberculose bovine ont eacutechoueacute malgreacute le grand nombre drsquoeacutechantillons
environnementaux preacuteleveacutes (Pillai et al 2000 Fine Bolin et al 2011 Iivanainen et al 1999 Williams
amp Hoy 1930a Little et al 1982) La deacutetection de M bovis dans des eacutechantillons environnementaux par
la culture est compliqueacutee par la nature des eacutechantillons analyseacutes en effet ces derniers (sol seacutediments
eau stagnante matiegraveres feacutecales etc) contiennent des micro-organismes tregraves divers et abondants La
croissance de M bovis eacutetant tregraves lente les organismes agrave croissance rapide contaminent rapidement les
milieux de culture compliquant voire empecircchant lrsquoisolement de la bacteacuterie Des traitements de
deacutecontamination preacuteliminaires sont indispensables pour eacuteliminer cette microflore mais la
deacutecontamination reacuteduit en geacuteneacuteral fortement la viabiliteacute de M bovis reacuteduisant aussi la sensibiliteacute des
meacutethodes baseacutees sur la culture (Fine Bolin et al 2011) Des microorganismes voisins des
mycobacteacuteries et preacutesents dans les sols peuvent eacutegalement reacutesister aux proceacutedeacutes de deacutecontamination
et ecirctre retrouveacutes dans les isolements De plus lrsquointermittence et le faible niveau drsquoexcreacutetion chez les
Chapitre 1
46
animaux infecteacutes associeacutes au volume limiteacute des preacutelegravevements analyseacutes diminuent les chances de
deacutetection de M bovis dans lrsquoenvironnement (Fine OrsquoBrien et al 2011) Lrsquoameacutelioration des protocoles
de traitement des eacutechantillons avant la culture est fondamentale pour lrsquoisolement des souches
environnementales
Le deacuteveloppement des outils moleacuteculaires a permis de contourner les inconveacutenients lieacutes aux
techniques baseacutees sur la culture deacutecrits preacuteceacutedemment En Irlande lrsquoanalyse de preacutelegravevements reacutealiseacutes
en 2000 dans un eacutelevage bovin toucheacute plusieurs fois par la tuberculose a mis en eacutevidence par des
meacutethodes moleacuteculaires la preacutesence de 1 times 103 agrave 25 times 103 copies de gegravenes cibles de M bovis (mpb64 et
mpb70) dans des pacirctures et des sols drsquoentreacutee de terriers de blaireaux adjacents aux pacirctures (Young et
al 2005) La reacuteiteacuteration des analyses 30 mois plus tard en novembre 2002 a donneacute le mecircme reacutesultat
sur les terriers bien qursquoils soient resteacutes inhabiteacutes Le rocircle des blaireaux dans la contamination des bovins
est fortement suspecteacute dans cette ferme En Angleterre lrsquoanalyse moleacuteculaire de preacutelegravevements
environnementaux (terriers et latrines) reacutealiseacutes en 2003 autour ou dans les pacirctures drsquoeacutelevages bovins
infecteacutes dans six zones endeacutemiques de tuberculose bovine a mis en eacutevidence M bovis dans les sols
de terriers de blaireaux de 78 des fermes analyseacutees (n=60) Dans chaque ferme 47 des terriers et
29 des latrines analyseacutes eacutetaient positifs en PCR une forte proportion eacutetait accessible aux bovins
(Courtenay et al 2006)
Entre 2004 et 2013 lrsquoanalyse par deacutetection moleacuteculaire de latrines appartenantt agrave des groupes sociaux
de blaireaux infecteacutes de Woodchester Park a montreacute que selon les eacutetudes 13 agrave 100 des latrines eacutetaient
contamineacutees avec des quantiteacutes pouvant atteindre 11 times 108 copies de gegravenes par g de latrine
(Courtenay et al 2006 Travis et al 2011 Sweeney et al 2006) Lrsquoamplification de seacutequences ciblant
lrsquoARNr 16S dans certains eacutechantillons des eacutetudes preacutesenteacutees ci-dessus suggegravere que les bacilles deacutetecteacutes
eacutetaient viables (Young et al 2005 Courtenay et al 2006) toutefois la seacutequence de lrsquoARNr 16S cibleacutee
eacutetant speacutecifique de toutes les mycobacteacuteries agrave croissance lente les reacutesultats sont agrave consideacuterer avec
prudence Dans ces eacutetudes le nombre de blaireaux excreacuteteurs et leur niveau drsquoexcreacutetion sont
fortement correacuteleacutes agrave la contamination des terriers et des latrines
Dans le sud du Portugal une eacutetude reacutecente dans une zone endeacutemique a mis en eacutevidence par PCR ciblant
lrsquoIS6110 des bacteacuteries du MTBC dans 38 des 248 eacutechantillons preacuteleveacutes dans des zones drsquoagreacutegation
drsquoanimaux domestiques et sauvages zones drsquoabreuvement utiliseacutees par les bovins et la faune sauvage
(seacutediments et eau drsquoeacutetangs 1358 et 357 et de sources saisonniegraveres 1161) sol des zones de
nourrissage des animaux (528) du sol de boutis de sangliers (416) ainsi que dans le sol des zones de
nourrissage des vautours (119) et dans leurs fegraveces (19) (Santos Santos et al 2015) Lrsquoutilisation drsquoune
seconde cible PCR (RD12) a confirmeacute 4 eacutechantillons comme positifs pour M bovisM caprae (2
eacutechantillons de seacutediments drsquoeacutetang 1 de source et 1 de sol de nourrissage) sans pouvoir diffeacuterencier les
deux espegraveces Le plus grand nombre drsquoeacutechantillons positifs a eacuteteacute retrouveacute au printemps (tempeacuterature
Chapitre 1
47
moyenne de 16degC)
La PCR permet drsquoobtenir un reacutesultat rapide avec une speacutecificiteacute acceptable Lrsquoanalyse
drsquoeacutechantillons de sol drsquoeau de foin et de maiumls inoculeacutes avec M bovis par une meacutethode baseacutee sur la
culture et la PCR a montreacute que cette derniegravere permet de deacutetecter lrsquoADN de M bovis dans lrsquoeacutechantillon
sur une plus longue peacuteriode apregraves la contamination que la culture indeacutependamment du substrat
(Adams et al 2013) Toutefois les seuils de deacutetection eacuteleveacutes des systegravemes speacutecifiques de M bovis (de
4 times 104 agrave 5 times 105 par g ou mL drsquoeacutechantillon Tableau 3) limitent leur utilisation dans le screening
drsquoeacutechantillons environnementaux pour lesquels la charge bacteacuterienne est probablement infeacuterieure au
seuil de deacutetection Enfin la technique PCR ne donne aucune information sur la viabiliteacute et la virulence
des bacteacuteries deacutetecteacutees
b Dureacutees de survie de Mycobacterium bovis en conditions expeacuterimentales
Les eacutetudes expeacuterimentales sur la survie environnementale de M bovis ont deacutebuteacute dans les
anneacutees 1930 en Angleterre dans lrsquoobjectif de preacuteciser le rocircle de lrsquoenvironnement dans lrsquoeacutepideacutemiologie
de la tuberculose bovine et drsquoameacuteliorer les programmes drsquoeacuteradication de la maladie (Maddock 1933
Maddock 1934 Maddock 1936 Williams amp Hoy 1930) Mycobacterium bovis et M bovis BCG sont
capables de survivre dans les fegraveces les effluents le sol et les divers excreacutetas animaux de quelques jours
agrave quelques mois voire jusqursquoagrave deux ans () Les nombreuses conditions drsquoinoculation (concentration de
lrsquoinoculum souche utiliseacutee M bovis ou M bovis BCG origine de lrsquoinoculum culture ou broyat de
leacutesions) la varieacuteteacute des matrices inoculeacutees (matiegraveres feacutecales urine mucus organes sol eau etc) et
des conditions environnementales testeacutees (conditions controcircleacutees de laboratoire ou exposition aux
conditions climatiques) dans ces diffeacuterentes eacutetudes expliquent vraisemblablement de tels eacutecarts de
survie
La compilation de toutes ces donneacutees indique que la tempeacuterature lrsquoexposition aux UV et
lrsquohumiditeacute de la matrice semblent ecirctre les trois facteurs primordiaux intervenant dans la persistance
de M bovis quel que soit le substrat analyseacute Il est montreacute que dans lrsquoheacutemisphegravere Nord la dureacutee de
survie de M bovis est plus longue en automne hiver et printemps (Williams amp Hoy 1930 Tanner amp
Michel 1999 Fine Bolin et al 2011 Duffield amp Young 1985 Jackson et al 1995) saisons caracteacuteriseacutees
par des tempeacuteratures plus basses qursquoen eacuteteacute un ensoleillement minimal et une eacutevapotranspiration par
le couvert veacutegeacutetal limiteacutee Toutefois Fine et al (2011) ont montreacute que M bovis pouvait survivre jusqursquoagrave
48 jours dans lrsquoeau en eacuteteacute malgreacute les tempeacuteratures eacuteleveacutees et lrsquoensoleillement Les eacutetudes reacutealiseacutees en
Afrique du Sud et en Nouvelle-Zeacutelande ougrave les tempeacuteratures sont eacuteleveacutees sur de longues dureacutees avec
un ensoleillement important ont montreacute que la survie de M bovis est ameacutelioreacutee lorsque le sol contient
une humiditeacute suffisante sous une exposition ombrageacutee (Duffield amp Young 1985 Tanner amp Michel 1999)
Chapitre 1
48
Lrsquoexposition des bacilles aux UV diminue en effet fortement leur dureacutee de persistance dans
lrsquoenvironnement (Williams amp Hoy 1930a Genov 1965 Fine Bolin et al 2011) et la dureacutee de survie est
tregraves prolongeacutee lorsque les eacutechantillons sont enterreacutes agrave quelques centimegravetres sous la surface du sol
(Genov 1965) La survie de M bovis est optimale dans les matiegraveres feacutecales ougrave le bacille est proteacutegeacute
dans un environnement humide agrave lrsquoabri des radiations solaires et minimale lorsqursquoil est pulveacuteriseacute sur
les veacutegeacutetaux et soumis aux UV et agrave la dessiccation (Maddock 1933 Van Donsel amp Larkin 1977)
49
Reacutefeacuterences BCG M bovis Matrices inoculeacutees Concentration de
lrsquoinoculum Conditions
Meacutethodes de deacutetection
de M bovis Reacutesultats de survie (virulence) de M bovis
(Williams amp Hoy
1930)
X Fegraveces de bovin
Inconnue Expeacuterimentales
(tempeacuterature
fraicircche obscuriteacute)
Injection sous-
cutaneacutee agrave des cochons
drsquoInde (cuisse)
12 mois dans fegraveces naturellement infecteacutees
5104 agrave 5107 UFC 2 ans dans fegraveces artificiellement infecteacutees
X Lisier de bovin 5000 UFCcm3
Expeacuterimentales
(tempeacuterature
fraicircche obscuriteacute)
Injection sous-
cutaneacutee agrave des cochons
drsquoInde (cuisse)
Survie 4 mois avec diminution de la virulence du
bacille au cours du temps
X Fegraveces de bovin 5104 agrave 5107 UFC Environnementales
(pacircture)
Injection sous-cutaneacutee
agrave des cochons drsquoInde
(cuisse)
5 mois en hiver
4 mois en automne
2 mois au printemps eacuteteacute
4 mois si protection UV en eacuteteacute
6 mois si protection insectes-vers en automne
(Maddock 1933)
X
Sol
Sol + Fegraveces de bovin
Bouse
Inconnue Environnementales
(pacircture)
Injection sous-cutaneacutee
agrave des cochons drsquoInde
(cuisse)
178 j dans toutes les matrices
X Herbe
12105 01 m2 Environnementales
(pacircture)
Injection sous-
cutaneacutee agrave des cochons
drsquoInde (cuisse)
14 jours
12106 01 m2 28 jours
120106 01 m2 49 jours
(Maddock 1934) X Herbe
Inoculation
mensuelle de 5 agrave
25106 01 m2
reacutepeacuteteacutee
Environnementales
et expeacuterimentales
Ingestion drsquoherbe
contamineacutee
1422 cochons drsquoInde avec leacutesions
Tableau 1 Dureacutees de survie de M bovis dans diffeacuterentes matrices environnementales obtenues en conditions expeacuterimentales naturelles ou de laboratoire
50
(Maddock 1936)
X
Veaux infecteacutes
(excreacutetion prouveacutee
par injection agrave des
cochons drsquoInde)
Inconnue Environnementales
Pacircture seacutepareacutee en 3 2
veaux mis 1 mois apregraves
le retrait des veaux
infecteacutes 2 veaux 1 mois
plus tard 2 veaux 2
mois plus tard (laisseacutes 3
sem)
Aucun veau IDR +
Aucune leacutesion tuberculeuse agrave lrsquoautopsie
X
Vache infecteacutee
(excreacutetion prouveacutee
par injection agrave des
cochons drsquoInde)
Pacircture seacutepareacutee en 2 3
veaux mis 1 mois et 12
apregraves le retrait de la
vache infecteacutee (laisseacutes
53j) 3 veaux mis 1 mois
plus tard (laisseacutes 24j)
Abattage et autopsie 1 an plus tard aucune leacutesion
Bouses infectieuses 1 mois et frac12 apregraves retrait de la
vache infecteacutee puis neacutegatif 1 2 et 3 mois plus tard
Sol et herbe sous bouses neacutegatifs 3 4 et 5 mois apregraves
retrait de la vache infecteacutee
(Genov 1965) X
Fegraveces de bovin
Sang
Urine
Environnementales Culture
150 agrave 332 jours agrave 12-24degC sans drsquoUV
18 agrave 31 jours agrave 24-34degC avec UV
2 ans si enterreacute dans sol agrave 5 cm
11 mois si enterreacute agrave 1 cm
452 et 469 jours dans eaubouse et eauurine
(Van Donsel amp
Larkin 1977) X
Effluents animaux
Boues drsquoeacutepuration Environnementales
Abattage 90 en 11 jours pour sols arroseacutes avec
effluents 8 jours avec boues
Isolement apregraves 4 agrave 6 jours sur des radis et jusqursquoagrave
35 jours sur des laitues
Anon 1979 citeacute par
(Morris et al 1994)
Excreacutetas de
blaireaux conserveacutes
dans un terrier
Environnementales Culture
Urine 3 jours en eacuteteacute 28 jours en hiver
Mucus bronchique 7 et 70 jours
Fegraveces naturellement infecteacutes 14 et 28 jours
(Little et al 1982)
Sol foin litiegravere et
eau de blaireaux
inoculeacutes
Inconnue
Expeacuterimentales
Culture 1 culture positive (eau drsquoun abreuvoir)
51
(Duffield amp Young
1985)
Sol sec steacuterile ou non
Sol 30 humiditeacute
steacuterile ou non
Fegraveces
Culture
lt8 semaines dans sols secs ou humides non steacuteriles
avec 80 ombre (34degc) obscuriteacute (32degC) et dans
labo (23degC)
lt8 semaines dans sol humide steacuterile avec 80
ombre et obscuriteacute
Pas de survie plein soleil (43degC) dans sols et fegraveces
(Tanner amp Michel
1999)
Leacutesions pulmonaires
ou ganglionnaires de
buffle
Fegraveces de bovin
inoculeacutees
8107 UFC 50g
fegraveces Culture
Poumon 42 jours en hiver agrave lrsquoombre dans sol
humide ou sec (20degC)
Fegraveces 28 jours en hiver au soleil dans sol humide
ou sec
(Jackson et al
1995) X Rubans de coton
Environ 106 UFC
ruban
Environnementales
(rubans deacuteposeacutes
dans terriers de
possum et sols de
pacirctures ou de forecirct)
Culture
2 jours sur les sols de pacirctures
14 jours sur les sols forestiers et dans les terriers de
possum
(Scanlon amp Quinn
2000b) X Lisier bovin steacuteriliseacute 65 times 103 UFCmL Expeacuterimentales Culture 171 jours (concentration finale 2 UFCmL)
(Young et al 2005) X Sol steacuterile et non
steacuterile 108 UFC g sol
Expeacuterimentales
(4 15 25 37degC)
PCR pour les sols non
steacuteriles
Culture pour les sols
steacuteriles
Survie agrave 60 jours optimale agrave 37degC agrave -20kPa
drsquohumiditeacute dans le sol steacuterile
Deacutetection drsquoARNm pendant 15 mois (sol steacuterile)
(Palmer amp Whipple
2006) X
Betterave pommes
carottes pommes de
terre maiumls foin
11 times 106
UFC10g
Expeacuterimentales
(-20 8 et 23degC)
Culture
112 jours agrave -20degC pour tous les substrats
112 jours agrave 8degC pour pommes maiumls foin et pdeterre
112 jours agrave 23degC pour pommes maiumls et pdeterre
(Fine Bolin et al
2011) X Sol foin maiumls eau
50 times 104
UFCeacutechantillon Environnementales Culture
88 jours dans le sol en hiverprintemps
58 jours dans lrsquoeau 43 jours dans le foin et 37 jours
dans le maiumls en automnehiver
52
(Ghodbane et al
2014) X Sol steacuterile 108 UFC g sol
Expeacuterimentales
(18 agrave 24degC
obscuriteacute)
Culture
Survie 12 mois (concentration finale 150 UFCg
sol)
Virulence conserveacutee sur souris
Chapitre 1
53
Dans les fegraveces drsquoanimaux M bovis survit jusqursquo agrave 2 ans dans les bouses de bovins en conditions
de laboratoire (Williams amp Hoy 1930) et de 14 jours agrave 6 mois lorsque les fegraveces sont exposeacutees dans les
pacirctures (Williams amp Hoy 1930a Genov 1965 Morris et al 1994 Tanner amp Michel 1999) Dans les
effluents drsquoeacutelevages contamineacutes il a eacuteteacute montreacute que M bovis survivait de 4 mois dans du lisier en
conditions de laboratoire (Williams amp Hoy 1930a) et jusqursquoagrave 171 jours dans du lisier steacuteriliseacute conserveacute
agrave tempeacuterature ambiante et agrave lrsquoobscuriteacute (Scanlon amp Quinn 2000b) Lrsquoeacutepandage drsquoeffluents ou de boues
inoculeacutees avec M bovis BCG sur des cultures de radis et laitue a montreacute la reacuteduction de 90 des bacilles
en respectivement 11 et 8 jours M bovis a toutefois eacuteteacute isoleacutee de faccedilon sporadique 35 jours apregraves
arrosage (Van Donsel amp Larkin 1977)
Dans le sol les dureacutees de survie les plus longues pour M bovis et M bovis BCG ont eacuteteacute
observeacutees dans du sol steacuterile (63 de sable pH 65-68) et en conditions de laboratoire (Young et al
2005 Ghodbane et al 2014) Young et al (2005) ont amplifieacute lrsquoARNr 16S de M bovis BCG jusqursquoagrave 15
mois apregraves inoculation du sol alors que Ghodbane et al (2014) ont cultiveacute M bovis apregraves 12 mois de
seacutejour dans le sol (pH 83) Dans les conditions environnementales Fine et al (2011) ont confirmeacute
lrsquoeffet de la saison sur la survie de M bovis dans le sol sur du maiumls et du foin avec une survie maximale
en hiverprintemps (feacutevrier-mai tempeacuterature moyenne 29degC radiations solaires 800 Wm2) une
survie de 34 jours en automnehiver (novembre-janvier tempeacuterature moyenne 04degC radiations
solaires 243 Wm2) et une survie minimale de 12 jours pendant la peacuteriode printempseacuteteacute (mai-aoucirct
tempeacuterature moyenne 217degC radiations solaires 1500 Wm2) De faccedilon contradictoire lrsquoeacutetude de
Young et al (2005) a mis en eacutevidence une dureacutee de survie optimale (60 jours) de M bovis BCG agrave 37degC
et minimale (16 jours) agrave 4degC
La survie de M bovis a eacutegalement eacuteteacute mise en eacutevidence par meacutethode culturale sur diffeacuterents
aliments du beacutetail (Palmer amp Whipple 2006 Fine Bolin et al 2011) Apregraves 112 jours la bacteacuterie est
cultiveacutee sur tous les substrats inoculeacutes conserveacutes agrave -20degC (betteraves pommes carottes pommes de
terre maiumls et foin) sur les pommes le maiumls le foin et les pommes de terre conserveacutes agrave 8degC et
uniquement sur les pommes le maiumls et les pommes de terre conserveacutes agrave 23degC (Palmer amp Whipple
2006) Fine et al (2011) ont eacutegalement montreacute que M bovis survivait plus longuement en
automnehiver sur du maiumls (43 jours) et du foin (37 jours) qursquoagrave la peacuteriode printempseacuteteacute (survie
infeacuterieure agrave 3 jours)
La bacteacuterie peut eacutegalement survivre entre 2 et 6 semaines dans des leacutesions tuberculeuses
macroscopiques de poumons et nœuds lymphatiques de buffle (Tanner amp Michel 1999) la survie
optimale eacutetant observeacutee en hiver dans des situations ombrageacutees et humides Des reacutesultats identiques
sont observeacutes pour des eacutechantillons drsquourine (3 jours en eacuteteacute vs 28 jours en hiver) et des seacutecreacutetions
bronchiques (7 jours vs 70 jours) (Morris et al 1994)
Les diffeacuterentes eacutetudes preacutesenteacutees ci-dessus deacutemontrent que M bovis survit un temps variable dans
Chapitre 1
54
lrsquoenvironnement une fois excreacuteteacute par son hocircte Ce temps est fonction de lrsquoexcreacuteta et du substrat qui le
reccediloit de sa charge bacteacuterienne et des conditions climatiques Il existe donc une peacuteriode appeleacutee
fenecirctre temporelle critique pendant laquelle les animaux qui interagissent avec les matrices
contamineacutees par des bacilles viables preacutesentent un risque accru drsquoinfection Comme nous lrsquoavons vu
preacuteceacutedemment la fenecirctre temporelle critique est fortement deacutependante du pays ou de la reacutegion
eacutetudieacutes et des conditions climatiques
Tregraves peu de donneacutees existent sur la conservation de la virulence de M bovis apregraves un passage
en dehors de son hocircte La virulence du bacille a eacuteteacute confirmeacutee par injection agrave des cobayes dans des
fegraveces de bovins naturellement contamineacutees apregraves 178 jours drsquoexposition dans une pacircture et apregraves 12
mois drsquoincubation en conditions de laboratoire (Maddock 1933) Lrsquoeacutetude reacutecente de Ghodbane et al
(2014) a deacutemontreacute que M tuberculosis M canetti et M bovis conservaient leur virulence apregraves un
seacutejour de 12 mois dans le sol En effet lrsquoinjection intra-peacuteritoneacuteale de suspensions bacteacuteriennes agrave des
souris a entraineacute lrsquoapparition de leacutesions avec M canetti (nodules sur les poumons le foie et la rate) et
M tuberculosis (nodules sur les poumons et la rate) Malgreacute lrsquoabsence de leacutesions lors de lrsquoinjection de
M bovis la bacteacuterie a pu ecirctre isoleacutee en quantiteacute importante des 3 organes Toutefois les injections
eacutetaient reacutealiseacutees avec les colonies de mycobacteacuteries obtenues apregraves culture sur milieu geacuteloseacute induisant
un biais important De plus lrsquoingestion par des souris de sol inoculeacute par M tuberculosis apregraves 12 mois
drsquoincubation a provoqueacute des leacutesions pulmonaires (1 souris5) et des leacutesions de la rate (25) mais la
bacteacuterie a pu ecirctre cultiveacutee agrave partir des poumons de la rate de lrsquoestomac et de lrsquointestin chez toutes les
souris
4 Strateacutegies de survie des mycobacteacuteries dans lrsquoenvironnement
Lrsquoenveloppe particuliegravere des mycobacteacuteries deacutecrite preacuteceacutedemment est probablement un des
facteurs majeurs de la reacutesistance de M bovis en dehors de son hocircte dans des conditions
environnementales deacutefavorables Drsquoautres strateacutegies de survie sont probablement mises en jeu par
M bovis pour persister dans lrsquoenvironnement mais sont totalement inconnues agrave ce jour Les
mycobacteacuteries pathogegravenes et opportunistes ont deacuteveloppeacute diffeacuterents meacutecanismes pour survivre agrave
lrsquointeacuterieur des cellules de lrsquohocircte strateacutegies qui peuvent eacutegalement ecirctre utiliseacutees en dehors de lrsquohocircte
Les meacutecanismes moleacuteculaires de certains processus sont eacutegalement en cours de compreacutehension Les
principales strateacutegies de survie sont preacutesenteacutees dans la partie suivante
a Lrsquoeacutetat de dormance
La dormance est deacutefinie chez les bacteacuteries non sporuleacutees comme un eacutetat physiologique
Chapitre 1
55
reacuteversible permettant la survie de la bacteacuterie pendant une longue peacuteriode sans croissance ni reacuteplication
(Kaprelyants et al 1993) Le programme geacuteneacutetique de la bacteacuterie lui permet drsquoentrer en dormance (en
anglais Non-Replicating Persistence) lorsque les conditions exteacuterieures deviennent deacutefavorables
(notamment en conditions oligotrophiques) puis de retrouver sa capaciteacute agrave se diviser lorsque les
conditions redeviennent favorables La dormance est deacutecrite chez plusieurs espegraveces de mycobacteacuteries
Lrsquoentreacutee en dormance de M smegmatis (Dick et al 1998) M bovis (Lim et al 1999 Boon et al 2001)
et M tuberculosis (Wayne amp Hayes 1996) en culture est provoqueacutee par la rareacutefaction de lrsquooxygegravene
Lrsquoeacutetat de dormance est suggeacutereacute chez Map et pourrait expliquer sa longue survie dans le fumier le lisier
et le sol (Whittington et al 2004) Lrsquoeacutetat de dormance est eacutegalement deacutecrit chez M avium (Archuleta
et al 2005)
Chez les mycobacteacuteries lrsquoentreacutee en eacutetat de dormance est controcircleacutee par le facteur de
transcription DosR (Dormancy Survival Regulator) (Boon amp Dick 2002) Composeacute de 48 gegravenes
(Schnappinger et al 2003 Flores Valdez amp Schoolnik 2010) son expression est activeacutee in vitro par
lrsquooxyde nitrique (NO) (Voskuil et al 2003) le monoxyde de carbone (CO) (Shiloh et al 2008) le deacuteficit
en nutriments (Rodriguez et al 2008) et lrsquohypoxie (Rodriguez et al 2008) Lrsquoexpression de ce reacutegulon a
eacuteteacute deacutemontreacutee lors drsquoinfection expeacuterimentale de macrophages (Schnappinger et al 2003 Rodriguez et
al 2008) et drsquoanimaux modegraveles (Voskuil et al 2003) pour M tuberculosis et dans des nœuds
lymphatiques de bovins positifs en IDR pour M bovis (Jimeacutenez et al 2013) Ce reacutegulon permettrait non
seulement la survie de la bacteacuterie pendant la phase de dormance mais aussi la transition de la
respiration aeacuterobie agrave lrsquoanaeacuterobiose et inversement (Leistikow et al 2010) A lrsquoheure actuelle le rocircle de
certaines proteacuteines codantes et drsquoARN non codants de ce reacutegulon demeure toujours inconnu Une
eacutetude geacutenomique comparative a permis de constater que le reacutegulon de la dormance eacutetait conserveacute
chez les diffeacuterentes espegraveces de mycobacteacuteries y compris chez les mycobacteacuteries environnementales
(Gerasimova et al 2011) Cette donneacutee suggegravere que la dormance est une reacuteponse adaptative
probablement tregraves ancienne chez les mycobacteacuteries pour survivre dans des environnements ougrave les
concentrations en oxygegravene sont fluctuantes (Boon amp Dick 2012) et que cette adaptation a eacuteteacute mise agrave
profit par les mycobacteacuteries pour survivre dans les cellules immunitaires des hocirctes qursquoelles infectent
Aucune eacutetude nrsquoest agrave ce jour disponible concernant lrsquoentreacutee en dormance potentielle de
M bovis dans lrsquoenvironnement Le sol constitue toutefois un environnement hostile ougrave facteurs
biotiques et abiotiques pourraient deacuteclencher le passage de la bacteacuterie drsquoun eacutetat actif agrave un eacutetat
dormant (hypoxie carence en nutriments tempeacuterature hellip)
b La sporulation
Lrsquoeacutevocation de la sporulation chez les mycobacteacuteries est tregraves controverseacutee Ghosh et al (Ghosh
Chapitre 1
56
et al 2009) ont deacutecrit la sporulation expeacuterimentale de M marinum sous forme drsquoendospores dans des
cultures liquides acircgeacutees de 2 mois et dans des cultures solides de M bovis BCG de 8 mois Malgreacute des
veacuterifications pheacutenotypiques biochimiques et geacutenomiques deacutemontrant que ces spores eacutetaient bien
issues de M marinum ces reacutesultats ont eacuteteacute contesteacutes par Traag et al (Traag et al 2010) qui ont eacutechoueacute
agrave les reproduire Lamont et al (Lamont et al 2012) ont pour la premiegravere fois mis en eacutevidence
lrsquoapparition de morphotype spore-like dans des cultures liquides de Map drsquoorigine bovine (K-10) ovine
(7565) et humaine acircgeacutees drsquoun an en conditions oligotrophiques Le seacutequenccedilage de lrsquoARNr 16S et
lrsquoamplification de lrsquoIS900 speacutecifique de Map ont confirmeacute que les colonies bacteacuteriennes issues des
spores correspondaient bien agrave Map Le chauffage des spores obtenues agrave 70degC pendant 30 minutes ne
semble pas affecter sa virulence sur les macrophages Des eacutetudes compleacutementaires sont neacutecessaires
pour confirmer ces observations A notre connaissance la sporulation nrsquoa jamais eacuteteacute eacutevoqueacutee pour les
mycobacteacuteries du MTBC
c Interactions des mycobacteacuteries avec la faune du sol
La microfaune et plus particuliegraverement les amibes libres sont largement preacutesentes dans
lrsquoenvironnement et colonisent les eacutecosystegravemes aquatiques le sol (Khan 2006 Rodriacuteguez-Zaragoza
1994) et lrsquoair Elles ont eacuteteacute eacutegalement isoleacutees chez lrsquohomme Elles vivent preacutefeacuterentiellement dans les
biofilms et aux interfaces eau-sol eau-air et eau-plantes (Rodriacuteguez-Zaragoza 1994) ougrave elles
laquo broutent raquo les bacteacuteries les champignons et les algues dont elles reacutegulent les communauteacutes (Huws
et al 2005) Les mycobacteacuteries environnementales sont largement preacutesentes dans les milieux
aquatiques (Primm et al 2004) ougrave elles peuvent interagir avec les amibes De nombreuses eacutetudes
rapportent la capaciteacute des mycobacteacuteries environnementales agrave survivre et se multiplier dans les
trophozoiumltes etou les kystes drsquoamibes (Adekambi et al 2006 Drancourt et al 2007 Ben Salah amp
Drancourt 2010 Krishna-Prasad amp Gupta 1978 Mura amp Bull 2006) Les amibes pourraient mecircme
constituer un veacuteritable reacuteservoir pour M xenopi (Drancourt et al 2007 Thomas et al 2006)
La plupart des mycobacteacuteries pathogegravenes y compris M bovis sont capables de survivre dans
les amibes voire de srsquoy multiplier (Thomas amp McDonnell 2007 Mba Medie et al 2011 Greub amp Raoult
2004 Cirillo et al 1997 Adekambi et al 2006) La survie de M bovis a eacuteteacute confirmeacutee dans les
trophozoiumltes et les kystes drsquoAcanthamoeba castellanii jusqursquoagrave 5 mois apregraves infection en conditions
expeacuterimentales (Taylor et al 2003 Mardare et al 2013) La virulence vis-agrave-vis des macrophages
humains et des cellules intestinales de souris semble ecirctre exacerbeacutee par la reacuteplication intra-amibienne
(Cirillo et al 1997) En conditions naturelles lrsquoobservation de protozoaires isoleacutes de latrines de
blaireaux infecteacutes a mis en eacutevidence des BAAR sans pouvoir identifier preacuteciseacutement M bovis (Mardare
et al 2013) Des eacutetudes compleacutementaires sont neacutecessaires pour preacuteciser le rocircle de reacuteservoir eacuteventuel
Chapitre 1
57
des amibes pour M bovis Les interactions des mycobacteacuteries pathogegravenes avec les amibes pourraient
ecirctre du mecircme ordre que celles avec les macrophages de lrsquohocircte infecteacute notamment au niveau des
meacutecanismes de peacuteneacutetration (Danelishvili et al 2007) et de lrsquoinhibition de la fusion entre le phagosome
et le lysosome suggeacuterant qursquoune relation eacutevolutive des mycobacteacuteries avec les amibes auraient permis
aux mycobacteacuteries lrsquoacquisition de ces caracteacuteristiques de survie (Cirillo et al 1997)
La localisation intra-amibienne des mycobacteacuteries les protegravege des conditions
environnementales deacutefavorables des biocides tels que le chlore (Adekambi et al 2006 Whan et al
2006) des antibiotiques (Miltner amp Bermudez 2000) des diffeacuterents traitements comme lrsquoozonation de
lrsquoeau (Thomas et al 2008) et favorise leur disseacutemination et leur implication potentielle dans des
maladies humaines (Greub amp Raoult 2004) Lors de la peacuteneacutetration dans lrsquohocircte lrsquoamibe joue un rocircle de
laquo cheval de Troie raquo (Barker amp Brown 1994) proteacutegeant les bacteacuteries contre les premiegraveres deacutefenses de
lrsquohocircte
Lrsquoutilisation des amibes en co-cultures pour isoler des bacteacuteries reacutesistantes aux amibes dans
des eacutechantillons environnementaux et meacutedicaux a permis drsquoisoler des mycobacteacuteries drsquoeacutecosystegravemes
aquatiques (Thomas et al 2006 Thomas amp McDonnell 2007 Pagnier et al 2008) et de sols pollueacutes
(Wang et al 2006)
Les eacutetudes relatant les interactions entre M bovis et la meacuteso- et macrofaune sont rares Une
eacutetude reacutealiseacutee dans des eacutelevages bovins et caprins infecteacutes par Map a mis en eacutevidence la contamination
de 31 des vers de terre preacuteleveacutes sur les pacirctures freacutequenteacutees par les animaux (Fischer et al 2003)
Dans cette eacutetude Map a eacuteteacute retrouveacutee dans le corps et les fegraveces de vers de terre 1 et 2 jours apregraves
lrsquoingestion de bouses infecteacutees expeacuterimentalement suggeacuterant qursquoils peuvent probablement jouer un
rocircle de vecteurs dans la disseacutemination des mycobacteacuteries
Les eacutetudes disponibles bien que tregraves rares suggegraverent que la meacuteso- et macrofaune du sol
peuvent heacuteberger et probablement jouer un rocircle de vecteur des mycobacteacuteries Toutefois des eacutetudes
compleacutementaires sont indispensables concernant M bovis
d Interactions des mycobacteacuteries avec les diptegraveres
Le portage digestif et feacutecal de M bovis par des mouches communes (Musca domestica) en
contact avec des viscegraveres infecteacutes de bovins est deacutecrit La bacteacuterie survit de 12 jours sur leurs corps et
15 jours dans le tractus digestif de mouches mortes (Schlee 1957 citeacute par Kazda et al 2009)
e La formation de biofilms
Lrsquoaptitude agrave former des biofilms est connue pour de nombreuses espegraveces de mycobacteacuteries
Chapitre 1
58
environnementales agrave croissance rapide comme M smegmatis M fortuitum ou M abcessus ou agrave
croissance lente notamment M avium et M intracellulare (Falkinham et al 2001 Beumer et al 2010
Torvinen et al 2007) Lrsquoorganisation des mycobacteacuteries en biofilm leur confegravere une reacutesistance accrue
vis-agrave-vis des conditions deacutefavorables dans lrsquoenvironnement mais eacutegalement chez lrsquohocircte humain ougrave
elles peuvent coloniser les catheacuteters et provoquer des infections sanguines tregraves graves (El Helou et al
2013) Concernant les mycobacteacuteries du MTBC la formation de biofilms est deacutecrite pour
M tuberculosis dans des conditions expeacuterimentales tregraves particuliegraveres tregraves eacuteloigneacutees des conditions
naturelles (Kulka et al 2012) La formation drsquoagreacutegats (clumping ou cording) observeacutes lors de la culture
des bacteacuteries du MTBC en milieu liquide gracircce agrave une matrice extracellulaire riche en acides mycoliques
peut ecirctre assimileacutee agrave une forme organiseacutee de reacutesistance des bacteacuteries puisque sous cette forme les
bacteacuteries survivent agrave des concentrations tregraves eacuteleveacutees en antibiotique (Ojha et al 2008) Il a eacuteteacute montreacute
chez M tuberculosis que la formation de biofilms mettait en jeu des processus geacuteneacutetiques et
physiologiques diffeacuterents de la croissance planctonique (Ojha et al 2008) Toutefois comme il nrsquoa
jamais eacuteteacute deacutemontreacute que ces bacteacuteries se multiplient dans lrsquoenvironnement il semble peu probable
que les biofilms constituent une forme de reacutesistance pour M bovis
5 Facteurs de risque de contamination de lrsquoenvironnement et de transmission de Mycobacterium bovis aux animaux par lrsquoenvironnement contamineacute
Plusieurs eacutetudes portant sur les interactions entre bovins et faune sauvage ont montreacute que les
contacts directs eacutetaient tregraves peu freacutequents entre les bovins et les blaireaux (Benham amp Broom 1989
Drewe et al 2013 Boumlhm et al 2009) ainsi qursquoentre les cerfs les sangliers et les bovins (Cooper et al
2010 Kukielka et al 2013) La transmission indirecte de M bovis semble donc ecirctre le mode de
transmission preacutepondeacuterant entre la faune sauvage et les bovins voire entre les diffeacuterentes espegraveces
sauvages (Cowie et al 2015 Carrasco-Garcia et al 2015) Diffeacuterents facteurs de risque jouent un rocircle
sur ce mode de transmission qui neacutecessite un recouvrement des domaines vitaux des animaux
sauvages infecteacutes et excreacuteteurs avec les sites drsquoeacutelevage bovin (pacirctures et bacirctiments) Ces principaux
facteurs de risque sont deacutetailleacutes ci-dessous
a Risques lieacutes agrave lrsquoeacutecologie des espegraveces sauvages sensibles agrave Mycobacterium bovis
Concernant le blaireau et le sanglier les pacirctures peuvent constituer une grande partie de leur
domaine vital (Hutchings amp Harris 1999 Payne 2014) Elles sont utiliseacutees pour la recherche de
Chapitre 1
59
nourriture essentiellement des lombrics qui repreacutesentent une part importante de leur reacutegime
alimentaire (Tolhurst et al 2009 Cleary et al 2011 Baubet et al 2003) Une eacutetude reacutealiseacutee en Cocircte
drsquoOr a montreacute que la freacutequentation des pacirctures par les blaireaux est optimale au printemps peacuteriode
de mise au preacute des bovins par rapport agrave lrsquoautomne et agrave lrsquohiver Pour les sangliers la freacutequentation des
pacirctures est plus importante au printemps et en eacuteteacute (Payne 2014)
Concernant plus preacuteciseacutement les blaireaux ces derniers urinent et deacutefegravequent dans des latrines
pour deacutelimiter leurs territoires (Hutchings et al 2001) mais ils urinent eacutegalement sur les pacirctures
preacutefeacuterentiellement au niveau des lisiegraveres de forecirct des clocirctures et des haies qursquoils croisent au greacute de
leurs deacuteplacements dans un but de marquage (White et al 1993 Drewe et al 2013) Dans les pacirctures
il a eacuteteacute montreacute que le taux de contact entre bovins et latrines est eacuteleveacute (Hutchings amp Harris 1997 Drewe
et al 2013) Dans lrsquoeacutetude de Hutchings et Harris (1997) 48 des 50 vaches du troupeau ont cumuleacute 416
contacts avec 4 latrines preacutesentes dans la pacircture eacutetudieacutee et 33 animaux ont brouteacute sur les latrines (suivi
sur 1776 heures) Pour Drewe et al (2013) 1716 contacts avec 13 latrines ont eacuteteacute enregistreacutes pour les
33 bovins drsquoun troupeau sur les cinq mois de lrsquoeacutetude Toutefois le rang hieacuterarchique des bovins du
troupeau joue un rocircle important puisque les animaux domineacutes sont les plus enclins agrave brouter au
voisinage des latrines Enfin les bovins consomment lrsquoherbe au voisinage des latrines de faccedilon plus
importante lorsque lrsquoherbe de la pacircture est rase (en cas de surpacircturage ou de fauchage) (Hutchings amp
Harris 1997) Compte tenu du grand nombre de bacilles qui peuvent ecirctre excreacuteteacutes par la voie urinaire
du fort taux de contact bovinslatrines et de la consommation de lrsquoherbe sur les latrines par certains
bovins il semblerait que les latrines et plus particuliegraverement les zones de marquage urinaire dans les
pacirctures repreacutesentent un risque important de transmission indirecte de la maladie par inhalation ou
ingestion (Hutchings amp Harris 1999)
Bien qursquoil nrsquoexiste aucune eacutetude sur le sujet la contamination des pacirctures par des blaireaux
preacutesentant des eacutecoulements purulents de morsures a eacutegalement eacuteteacute envisageacutee (Gallagher amp Clifton-
Hadley 2000)
Les blaireaux vivent dans des terriers qursquoils creusent dans le sol en milieu forestier ou en
dehors notamment dans les pacirctures La contamination des sols drsquoentreacutee de terrier par M bovis
repreacutesente eacutegalement un risque pour les bovins qui peuvent se contaminer en explorant les entreacutees de
terrier localiseacutees dans les pacirctures par ingestion ou inhalation de la litiegravere expulseacutee des tunnels
(Benham 1985) De plus le sol excaveacute des tunnels par les blaireaux est attirant pour les bovins qui
peuvent en ingeacuterer pour combler des carences mineacuterales (Dewes 1996) ou srsquoy frotter la tecircte creacuteant
des aeacuterosols de poussiegravere potentiellement infectant (Phillips et al 2003) La contamination des sols de
terriers repreacutesente eacutegalement un risque important de transmission de lrsquoinfection dans le groupe social
de blaireaux qui partage le terrier Le sol du terrier constitue un biotope adeacutequat pour la survie de
M bovis agrave lrsquoabri des UV sans variation extrecircme de tempeacuterature et avec une humiditeacute relative
Chapitre 1
60
importante (Moore amp Roper 2003) Lrsquoentretien de la maladie dans un groupe social par le terrier
contamineacute a deacutejagrave eacuteteacute eacutevoqueacute (Rogers et al 2003)
La freacutequentation des pacirctures par les sangliers pour la recherche de nourriture constitue
eacutegalement un facteur de risque dans la transmission indirecte de la bacteacuterie aux bovins Lrsquoexploration
du sol avec leurs museaux agrave la recherche de lombrics larves et autre nourriture ainsi que les reliquats
de sol deacuteplaceacutes peuvent constituer un facteur de contamination potentiel de lrsquoenvironnement par la
deacuteposition drsquoexcreacutetas oro-nasaux infecteacutes
Les cervideacutes peuvent eacutegalement utiliser les preacutes pour se nourrir et ecirctre une source de
contamination indirecte pour les bovins (Brook amp McLachlan 2009)
b Risques lieacutes aux pratiques drsquoeacutelevage
Le seacutejour prolongeacute des bovins au preacute est susceptible drsquoaugmenter les contacts directs et
indirects avec la faune sauvage infecteacutee Lrsquoeacutelevage allaitant est caracteacuteriseacute par une longue peacuteriode de
pacircturage entre avril et octobre puis une hivernation en bacirctiments au moment des vecirclages Certaines
races rustiques notamment la race salers qui ont repeupleacute une partie des cheptels bovins infecteacutes
apregraves abattage total passent toute lrsquoanneacutee au pacircturage Ces longues peacuteriodes de pacircturage constituent
un facteur de risque non neacutegligeable pour les bovins qui partagent leur habitat avec des espegraveces
sauvages infecteacutees Le surpacircturage est eacutegalement un facteur de risque pour les bovins qui sont ameneacutes
agrave brouter les latrines de blaireaux et agrave ingeacuterer des quantiteacutes de sol potentiellement contamineacute plus
importantes (Hutchings amp Harris 1999)
Les zones drsquoabreuvement et de nourrissage des bovins dans la pacircture constituent des zones
attractives pour la faune sauvage notamment lrsquohiver lorsque les ressources alimentaires font deacutefaut
Le partage de fourrage entre bovins et cervideacutes est probablement une source de contamination des
bovins dans le Michigan (OrsquoBrien et al 2006 Brook amp McLachlan 2009 Brook et al 2012) drsquoautant plus
que la transmission indirecte de la maladie entre des cerfs et des veaux par le partage du foin a eacuteteacute
prouveacutee expeacuterimentalement (Palmer et al 2010 Palmer et al 2004) La freacutequentation des auges de
bovins et des zones de nourrissage au sol par les blaireaux qui viennent manger les reliquats
alimentaires est reacuteguliegraverement deacutecrite au Royaume-Uni ainsi que celle des reacuteserves de grains
drsquoensilage et de foin dans lesquels ils peuvent uriner et deacutefeacutequer (Garnett et al 2002 Tolhurst et al
2009 Ward et al 2010) Les pierres agrave sel souvent poseacutees au sol dans les preacutes attirent particuliegraverement
les cervideacutes (Payne et al 2015 Brook amp McLachlan 2009) Les abreuvoirs et les zones drsquoabreuvement
(trous drsquoeau ruisseau source) constituent eacutegalement un facteur de risque lorsqursquoils sont utiliseacutes par
les bovins et la faune sauvage la survie de la bacteacuterie excreacuteteacutee eacutetant optimale dans les zones humides
Le comportement de baignade des sangliers dans les trous drsquoeau et les souilles entraine une forte
Chapitre 1
61
freacutequentation de ces zones notamment en eacuteteacute (Payne 2014)
Finalement lrsquoeacutepandage de fumiers frais augmenterait le risque drsquoinfection drsquoun troupeau par
rapport agrave lrsquoeacutepandage drsquoengrais ou de fumiers non frais (stockeacutes au moins deux mois) les animaux se
contaminant par inhalation ou ingestion de substrats contamineacutes dans les deux mois suivant lrsquoeacutepandage
(Griffin et al 1996 Humblet et al 2009) Certaines preacuteconisations conseillent un temps de stockage
minimum des fumiers avant eacutepandage (au moins 6 mois suite aux reacutesultats de Scanlon et Quinn 2000a
et 2000b) ou leur traitement chimique (Scanlon amp Quinn 2000b Scanlon amp Quinn 2000a) un
compostage (McCallan et al 2014) voire lrsquointerdiction drsquoeacutepandage de fumiers pour les cheptels infecteacutes
(Phillips et al 2003)
c Risques lieacutes aux pratiques cyneacutegeacutetiques
Les zones drsquoagreacutegation des animaux sauvages telles que les zones drsquoagrainage ou de
nourrissage agrave points fixes et la reacutealisation de trous drsquoeau artificiels pour lrsquoabreuvement et la baignade
des animaux sauvages sont des zones agrave risque qui augmentent les interactions directes et indirectes
entre les animaux et donc les risques de transmission de M bovis entre eux (Corner 2006 Vicente et
al 2007 Delahay et al 2002 Coleman amp Cooke 2001) En Espagne lrsquoaugmentation des densiteacutes
drsquoonguleacutes sauvages (cervideacutes et sangliers) dans le Sud du pays a entraineacute le deacuteveloppement drsquoactiviteacutes
de chasse agrave but commercial amenant agrave la reacutealisation de parcs gigantesques pour contenir les animaux
sauvages Dans ces zones lrsquoagreacutegation des animaux sauvages de diffeacuterentes espegraveces (sangliers
cervideacutes cochons sauvages et bovins) autour des trous drsquoeau et des zones de nourrissage a favoriseacute la
transmission intra- et interspeacutecifique de la maladie (Vicente et al 2007 Cowie et al 2015) Chez le
sanglier la preacutevalence varie de 52 agrave 70 voire presque 100 dans certaines populations (Gortaacutezar et
al 2008 Naranjo et al 2008) Cowie et al (2015) ont montreacute que 40 agrave 66 des interactions indirectes
avaient lieu dans une fenecirctre temporelle de 3 jours en saison segraveche et 72 agrave 100 dans une fenecirctre
temporelle de 12 jours en saison humide Or dans ces zones la survie de M bovis a eacuteteacute estimeacutee agrave 3
jours pendant la saison segraveche et agrave 12 jours pendant la saison humide (Kukielka et al 2013) suggeacuterant
que si le pathogegravene est preacutesent dans une population lrsquoagreacutegation est un facteur de maintenance et de
disseacutemination de lrsquoinfection dans et entre les espegraveces De plus la preacutesence drsquoeacutelevages bovins agrave
proximiteacute de ces parcs de chasse entraine une freacutequentation importante des structures drsquoeacutelevage par
les animaux sauvages et un risque accru drsquoinfection pour les bovins des eacutelevages visiteacutes (Carrasco-
Garcia et al 2015)
Le nourrissage des cervideacutes dans le Michigan dans le but de maintenir les populations pour la
chasse est la cause du maintien de lrsquoinfection dans les populations sauvages qui se contaminent entre
eux via les aliments distribueacutes (OrsquoBrien et al 2006)
Chapitre 1
62
Comme nous lrsquoavons deacutejagrave dit preacuteceacutedemment les viscegraveres drsquoanimaux infecteacutes laisseacutes agrave la
disposition des animaux sauvages par les chasseurs peuvent repreacutesenter un risque important pour les
animaux charognards (Morris et al 1994)
d Risques lieacutes aux facteurs paysagers climatiques et peacutedologiques
La structure du paysage est un facteur qui influence le recouvrement des domaines vitaux des
animaux sauvages et des bovins La localisation des pacirctures ou des bacirctiments drsquoeacutelevage en lisiegravere de
forecirct augmente les risques drsquointeractions entre animaux sauvages et domestiques (Ward et al 2010)
lrsquointerface pacircture-forecirct repreacutesenteacutee par les lisiegraveres eacutetant un eacutecotone agrave risque pour lrsquoeacutemergence des
agents pathogegravenes multi-hocirctes (Boyard et al 2008) La fragmentation du paysage (preacutes champs forecircts
bosquets) augmente le deacutepocirct drsquourine par les blaireaux aux zones frontiegraveres entre deux milieux
augmentant le risque de contamination de ces sites (White et al 1993 Humblet et al 2009) Les lisiegraveres
sont tregraves freacutequenteacutees par les sangliers et les blaireaux (Payne 2014) et constituent une zone privileacutegieacutee
par les blaireaux pour eacutetablir leurs terriers et marquer leurs territoires par des mictions ou des latrines
(Hutchings et al 2002)
Comme nous lrsquoavons vu preacuteceacutedemment les conditions climatiques sont un facteur essentiel
dans la dureacutee de survie de M bovis dans les diffeacuterents substrats environnementaux contamineacutes et in
fine dans la transmission indirecte de la maladie aux animaux qui interagissent avec ces substrats Les
saisons influencent eacutegalement la freacutequentation des pacirctures par les animaux sauvages avec un risque
accru au printemps et en eacuteteacute vis-agrave-vis des blaireaux et des sangliers qui vermillent dans les pacirctures
notamment lors des nuits humides ougrave les vers de terre remontent en surface appeleacutees laquo nuits agrave
lombrics raquo (Kruuk 1978 Cleary et al 2011 Baubet et al 2003) Lors de temps plus sec voire de
seacutecheresse les points drsquoeau destineacutes aux bovins constituent des zones attractives pour la faune sauvage
(Vicente et al 2007 Santos Santos et al 2015 Barasona et al 2014) favorisant lrsquoagreacutegation des
animaux domestiques et sauvages
Le rocircle des facteurs peacutedologiques dans la survie de M bovis est peu connu agrave ce jour La texture
du sol qui conditionne la structure la porositeacute et le reacutegime hydrique du sol le pH et la matiegravere
organique disponible sont des facteurs importants dans la survie des bacteacuteries A notre connaissance
une seule eacutetude statistique reacutealiseacutee dans le Michigan a montreacute que la nature du sol et plus
preacuteciseacutement le pourcentage de sable eacutetait associeacute agrave un risque drsquoinfection plus eacuteleveacute chez les bovins
(Walter et al 2014) Une augmentation drsquo1 de la quantiteacute de sable dans le sol augmentait de 36
le risque pour des eacutelevages bovins drsquoecirctre infecteacutes Hormis cette eacutetude qui relie indirectement infection
animale et survie de la bacteacuterie dans le sol aucune eacutetude nrsquoest actuellement disponible sur le lien
direct entre nature du sol et survie de M bovis
Chapitre 1
63
A la lumiegravere des informations preacutesenteacutees dans ce chapitre il apparait qursquoune fois excreacuteteacutee en
dehors de son hocircte M bovis peut survivre longuement dans lrsquoenvironnement si les conditions sont
adeacutequates par la mise en jeu de strateacutegies de survie encore inconnues agrave lrsquoheure actuelle Les
connaissances sur les scheacutemas de circulation de la bacteacuterie entre les animaux infecteacutes et
lrsquoenvironnement (et vice et versa) srsquoameacuteliorent et permettent drsquoadapter les programmes de lutte contre
la maladie mais elles sont encore insuffisantes pour permettre la maitrise de tous les facteurs en jeu
Ces diffeacuterents facteurs sont scheacutematiseacutes dans la Figure 15
Figure 15 Repreacutesentation scheacutematique de la circulation de M bovis dans lrsquoenvironnement agricole (modifieacute drsquoapregraves Elliott et al 2014)
Chapitre 1
64
D PRINCIPAUX OUTILS DE DETECTION DE PHENOTYPAGE ET DE GENOTYPAGE APPLICABLES AU MTBC ET PLUS PARTICULIEREMENT A MYCOBACTERIUM BOVIS
Depuis la deacutecouverte du bacille de la tuberculose de nombreux outils ont eacuteteacute deacuteveloppeacutes pour
deacutetecter et discriminer les diffeacuterentes mycobacteacuteries du MTBC Ces meacutethodes microscopiques
pheacutenotypiques et biochimiques essentiellement deacuteveloppeacutees pour le diagnostic meacutedical ont eacuteteacute peu
agrave peu ameacutelioreacutees et associeacutees agrave de nouvelles techniques de deacutetection de caracteacuterisation et
drsquoidentification des bacilles telles que lrsquoamplification geacutenique et le seacutequenccedilage notamment ameacuteliorant
la rapiditeacute du diagnostic la discrimination des espegraveces la pertinence de lrsquoantibiotheacuterapie et permettant
le geacutenotypage des souches agrave des fins eacutepideacutemiologiques Adapteacutees agrave des eacutechantillons biologiques
divers tels que les crachats les liquides de lavage broncho-alveacuteolaire les tissus de biopsies chez
lrsquohomme ou les broyats de nœuds lymphatiques chez les bovins et les animaux sauvages ces outils ne
sont pas tous applicables aux eacutechantillons environnementaux heacutebergeant des communauteacutes
microbiennes tregraves complexes
1 La deacutetection des mycobacteacuteries par microscopie
La microscopie optique
Lrsquoexamen microscopique de frottis ou drsquoeacutetalement drsquoeacutechantillons biologiques est la premiegravere
eacutetape dans le diagnostic bacteacuteriologique de la tuberculose humaine Elle est parfois la seule meacutethode
de diagnostic dans les pays en voie de deacuteveloppement Les proprieacuteteacutes tinctoriales particuliegraveres de la
mycomembrane des mycobacteacuteries sont exploiteacutees dans les colorations utiliseacutees La coloration de
reacutefeacuterence est la coloration de Ziehl et Neelsen Cette coloration agrave chaud est baseacutee sur la reacutesistance de
la paroi des mycobacteacuteries agrave la deacutecoloration agrave lrsquoacide et lrsquoalcool apregraves traitement agrave la fuschine Les
mycobacteacuteries apparaissent comme des bacirctonnets rouges sur un fond bleu Des meacutethodes agrave froid
baseacutees sur les mecircmes reacuteactifs existent eacutegalement (meacutethode de Kinyoun) La coloration agrave lrsquoauramine
un fluorochrome permet drsquoobserver en microscopie agrave fluorescence les bacilles en vert-jaune brillant
sur un fond jaune-orangeacute Cette coloration est plus sensible que la coloration de ZN (Steingart et al
2006)
Ces meacutethodes de coloration ne permettent pas de discriminer les mycobacteacuteries entre elles et
ne sont pas speacutecifiques du genre Mycobacterium puisqursquoelles peuvent colorer drsquoautres actinobacteacuteries
comme les genres bacteacuteriens Nocardia Rhodococcus et Corynebacterium (Shinnick amp Good 1994)
Lrsquoutilisation de la microscopie optique nrsquoest pas deacutecrite pour lrsquoanalyse des eacutechantillons
environnementaux drsquoune part car le seuil de deacutetection est eacuteleveacute (de lrsquoordre de 104 BAARmL) et drsquoautre
Chapitre 1
65
part car la coloration est difficile agrave mettre en œuvre sur des matrices telles que du sol ou des matiegraveres
feacutecales
Lrsquohybridation avec des sondes nucleacuteotidiques FISH (Fluorescence in situ hybridization)
Cette meacutethode est baseacutee sur lrsquohybridation speacutecifique drsquoune sonde marqueacutee avec un
fluorochrome sur une reacutegion cible de lrsquoADN bacteacuterien Les sondes utiliseacutees ADN ARNm ou
oligonucleacuteotides syntheacutetiques (PNA peptide nucleic acid) drsquoenviron 20 bases peacutenegravetrent agrave lrsquointeacuterieur
des cellules preacutealablement permeacuteabiliseacutees ougrave elles srsquohybrident speacutecifiquement avec leur ADN cible
Lrsquoobservation se fait agrave lrsquoaide drsquoun microscope agrave eacutepifluorescence
La meacutethode FISH a surtout eacuteteacute deacuteveloppeacutee pour la deacutetection des mycobacteacuteries dans les
crachats dans des cultures liquides chez des animaux infecteacutes expeacuterimentalement (Hongmanee et al
2001 Stender Mollerup et al 1999 Stender Lund et al 1999) et dans des eacutechantillons biologiques
(Stender Mollerup et al 1999 Lefmann et al 2006 Rodriguez-Nuntildeez et al 2012) La meacutethode FISH
est peu utiliseacutee pour la recherche de mycobacteacuteries dans des eacutechantillons environnementaux mais elle
a permis de mettre en eacutevidence Maa et Map dans de lrsquoeau et des biofilms de canalisations (Lehtola et
al 2006 Torvinen et al 2007) Elle parait toutefois difficilement applicable agrave des matrices telles que le
sol agrave cause de lrsquoauto-fluorescence geacuteneacutereacutee par les particules de sol et du risque de non-speacutecificiteacute des
sondes
2 Les meacutethodes de culture des mycobacteacuteries
Le diagnostic de la tuberculose est indissociable de la culture car historiquement crsquoest la
meacutethode la plus utiliseacutee Elle est sensible et permet drsquoisoler les souches de les caracteacuteriser et de tester
leur sensibiliteacute aux antibiotiques La recherche de mycobacteacuteries par des meacutethodes baseacutees sur la
culture est tregraves standardiseacutee en diagnostic humain et veacuteteacuterinaire tout comme les traitements
preacuteliminaires indispensables A lrsquoopposeacute aucune meacutethode standardiseacutee nrsquoest disponible pour la
recherche de mycobacteacuteries dans lrsquoenvironnement De nombreux protocoles emprunteacutes agrave la
microbiologie clinique ont eacuteteacute adapteacutes pour la recherche de mycobacteacuteries environnementales
notamment dans lrsquoeau (Narang et al 2009 Radomski et al 2011 Norby et al 2007 Pickup et al 2006
Whiley et al 2012 Le Dantec et al 2002 Bland et al 2005) Les meacutethodes utiliseacutees pour cultiver les
mycobacteacuteries pathogegravenes sont moins nombreuses et sont deacutecrites ci-dessous
a Les diffeacuterents traitements preacutealables agrave la mise en culture des eacutechantillons
De nombreuses meacutethodes de deacutecontamination des eacutechantillons biologiques ont eacuteteacute deacutecrites
dans le diagnostic meacutedical de la tuberculose Le choix de la meacutethode agrave utiliser deacutepend essentiellement
Chapitre 1
66
du type drsquoeacutechantillon analyseacute (tissu crachat lait fegraveces eau solhellip) des milieux de culture employeacutes et
de sa compatibiliteacute avec les meacutethodes moleacuteculaires Les meacutethodes les plus utiliseacutees en microbiologie
humaine meacutethodes de Petroff Taquet-Tison et Kubica utilisent la soude comme agent deacutecontaminant
principal (Petroff 1915 Taquet amp Tison 1961 Kubica et al 1963) Lrsquoutilisation drsquoacides comme agent
deacutecontaminant tels que lrsquoacide oxalique (David et al 1989) ou lrsquoacide sulfurique (Hohn amp Lowenstein
1926) ou drsquoammoniums quaternaires comme le chlorure de ceacutetyl pyridinium (Smithwick et al 1975)
ou le chlorure de benzalkonium (Krasnow amp Wayne 1966) est reacuteserveacutee aux eacutechantillons tregraves contamineacutes
(urines fegraveces) ou aux biopsies (cutaneacutees coliques pulmonaires etc) Ces meacutethodes ne sont pas toutes
compatibles avec la culture en milieu liquide et lrsquoamplification geacutenique
Dans le domaine veacuteteacuterinaire les tissus animaux (broyats de ganglions ou de tissus) sont
deacutecontamineacutes selon les preacuteconisations de lrsquoANSES et suivent la norme AFNOR NF U47-104 baseacutee sur
lrsquoutilisation de lrsquoacide sulfurique et de la soude agrave 4 Malgreacute la reacutesistance des mycobacteacuteries aux agents
chimiques la deacutecontamination reacuteduit fortement la population initiale preacutesente dans lrsquoeacutechantillon et
abaisse la sensibiliteacute de la culture (Fine Bolin et al 2011)
Concernant lrsquoisolement de bacteacuteries du MTBC dans les matrices environnementales ou les
excreacutetas animaux les principales meacutethodes utiliseacutees deacutecoulent des meacutethodes citeacutees preacuteceacutedemment
La meacutethode drsquoimmunocapture magneacutetique utiliseacutee par Sweeney (2006 et 2007) a permis lrsquoisolement
de M bovis dans des sols de terriers et des latrines contamineacutes naturellement Cette meacutethode est
baseacutee sur lrsquoutilisation de microbilles magneacutetiques recouvertes avec un ligand speacutecifique drsquoun antigegravene
de lrsquoenveloppe de M bovis Son avantage est qursquoelle permet la capture et la concentration de la bacteacuterie
cible dans un eacutechantillon complexe sans alteacuterer sa viabiliteacute contrairement agrave la deacutecontamination
chimique Cette meacutethode a eacutegalement eacuteteacute testeacutee avec succegraves sur des broyats de nœuds lymphatiques
de bovins (Stewart et al 2012 Stewart et al 2013 Garbaccio amp Cataldi 2010)
b La culture sur milieu solide
Les milieux de culture solides agrave lrsquoœuf (milieux de Loumlwenstein-Jensen et milieu de Coletsos) sont
les plus utiliseacutes en France dans le diagnostic bacteacuteriologique chez lrsquohomme et lrsquoanimal Ils sont sensibles
et reacutepondent aux exigences culturales de la plupart des mycobacteacuteries tuberculeuses et non
tuberculeuses Le milieu de Coletsos est particuliegraverement indiqueacute pour la culture des mycobacteacuteries
exigeantes telles que M bovis et M africanum
Les milieux geacuteloseacutes Middlebrook 7H10 et 7H11 sont des milieux semi-syntheacutetiques non utiliseacutes
en diagnostic de routine en France mais tregraves utiliseacutes aux Etats-Unis Enrichis avec de lrsquoOADC meacutelange
drsquoacide oleacuteique drsquoalbumine bovine de dextrose de chlorure de sodium et de catalase ils permettent
une croissance plus rapide de M tuberculosis (Mejia et al 1999) et de M bovis (Idigoras et al 1995)
Chapitre 1
67
que les milieux agrave base drsquoœuf Lrsquoajout de seacuterum bovin et de sang de mouton lyseacute au milieu 7H11 enrichi
en OADC ameacuteliore fortement la croissance de M bovis agrave partir drsquoeacutechantillons animaux et raccourcit le
deacutelai drsquoobservation des colonies (Gallagher amp Horwill 1977) Ce milieu a eacuteteacute utiliseacute pour la mise en
culture drsquoeacutechantillons environnementaux tels que le sol ou les fegraveces apregraves un traitement preacutealable de
lrsquoeacutechantillon (Sweeney et al 2006 Fine OrsquoBrien et al 2011) mais son manque de seacutelectiviteacute neacutecessite
lrsquoajout drsquoun cocktail drsquoantibiotiques souvent composeacute de lrsquoassociation drsquoune carboxypeacutenicilline de
trimeacutethoprime et de polymyxine B et drsquoun antifongique (amphoteacutericine B) (Sweeney et al 2007 Fine
OrsquoBrien et al 2011) Lrsquoincubation des milieux solides ensemenceacutes est reacutealiseacutee agrave 37degC pendant au moins
8 semaines
c La culture en milieu liquide
Les milieux liquides ont eacuteteacute deacuteveloppeacutes pour pallier la croissance lente des mycobacteacuteries
drsquointeacuterecirct clinique sur les milieux solides afin de confirmer preacutecocement le diagnostic Diffeacuterents
systegravemes manuels ou automatiseacutes existent agrave lrsquoheure actuelle Ils sont essentiellement utiliseacutes dans le
diagnostic chez lrsquohomme plus rarement chez lrsquoanimal
Le systegraveme Bactectrade 460 (Becton Dickinson) baseacute sur la respiromeacutetrie radiomeacutetrique nrsquoest plus
utiliseacute en France mais il lrsquoa eacuteteacute pendant une vingtaine drsquoanneacutees Il eacutetait baseacute sur la mesure du dioxyde
de carbone marqueacute au 14C rejeteacute par les mycobacteacuteries en croissance dans un milieu contenant de
lrsquoacide palmitique marqueacute au 14C La libeacuteration de 14CO2 eacutetait mesureacutee reacuteguliegraverement de faccedilon manuelle
gracircce agrave un appareil adapteacute La contrainte drsquoutilisation de stockage et de recyclage de produits
radioactifs ainsi que lrsquoabsence drsquoautomatisation ont conduit agrave lrsquoarrecirct de sa commercialisation Son
utilisation a eacuteteacute deacutecrite pour la deacutetection de M bovis sur des tissus animaux et des fegraveces (Corner et al
2012 Thacker et al 2015)
Pour eacuteviter les inconveacutenients lieacutes agrave la radioactiviteacute des meacutethodes non radiomeacutetriques ont eacuteteacute
deacuteveloppeacutees La croissance bacteacuterienne en milieu liquide est mise en eacutevidence soit par la
consommation de lrsquooxygegravene soit par le deacutegagement de CO2 dans le tube La mise en eacutevidence de la
diminution de la pression partielle drsquooxygegravene dissous dans le milieu de culture lors de la croissance
bacteacuterienne est utiliseacutee dans les tubes BBLtrade MGITtrade de Becton Dickinson Lorsque lrsquoO2 diminue les sels
de rutheacutenium preacutesents au fond du tube eacutemettent une fluorescence Lrsquoutilisation de systegravemes
automatiseacutes tels que les systegravemes Bactectrade MGITtrade assurent une lecture en continue de la fluorescence
eacutemise Le systegraveme Bactectrade MGITtrade a eacuteteacute utiliseacutee pour la recherche de M bovis dans des broyats de
nœuds lymphatiques de bovins (Hines et al 2006 Stewart et al 2013) des tissus drsquoanimaux sauvages
(Murphy et al 2010 Witmer et al 2010) et dans du fromage frais (Harris et al 2007) Lrsquoaugmentation
du CO2 dans le milieu de culture lors de la croissance bacteacuterienne est agrave la base du systegraveme BacTALERT
Chapitre 1
68
3D de Biomeacuterieux Cette meacutethode colorimeacutetrique est baseacutee sur la diminution du pH suite agrave la libeacuteration
du CO2 Lrsquoautomate eacutevalue en continu le virage de couleur drsquoune pastille fixeacutee au fond du tube Les
systegravemes deacutecrits permettent eacutegalement la recherche de reacutesistance agrave diffeacuterents antibiotiques
(Piersimoni et al 2006) A notre connaissance cette technique nrsquoa jamais eacuteteacute utiliseacutee pour la culture
drsquoeacutechantillons environnementaux
Ces diffeacuterents systegravemes utilisent comme milieu liquide le Middlebrook 7H9 suppleacutementeacute en
OADC ou en ADC (Albumine bovine dextrose catalase) pour ameacuteliorer la croissance des mycobacteacuteries
et rendu plus seacutelectif par lrsquoajout drsquoun meacutelange drsquoantibiotiques et drsquoantifongiques pour eacuteviter la
croissance de bacteacuteries indeacutesirables Le plus utiliseacute est le PANTAtrade (polymyxine B amphoteacutericine acide
nalidixique trimeacutethoprime azlocilline) Il peut ecirctre compleacuteteacute avec de la vancomycine (effet contre les
bacteacuteries gram +) Lrsquoavantage majeur de ces systegravemes en milieu liquide est la diminution du deacutelai moyen
de deacutetection des mycobacteacuteries Avec le systegraveme Bactectrade M tuberculosis est deacutetecteacutee entre 8 et 30
jours en fonction de la charge bacteacuterienne initiale de lrsquoeacutechantillon La deacutetection de M bovis est reacutealiseacutee
en 15 jours contre environ 45 jours sur milieu solide Lorsque la culture est deacuteclareacutee positive la
preacutesence de BAAR doit ecirctre veacuterifieacutee par coloration sub-culture sur milieu de Loumlwenstein-Jensen
identification biochimique ou moleacuteculaire Lrsquoassociation de la culture en milieu liquide et solide donne
les meilleurs reacutesultats avec une limite de deacutetection de 10 agrave 102 bacillesmL drsquoeacutechantillon (Guillet-Caruba
et al 2014)
NB A la fin du 19egraveme siegravecle et au deacutebut du 20egraveme la discrimination des espegraveces M bovis M tuberculosis
et M avium se faisait principalement par inoculation de lrsquoisolat bacteacuterien au cobaye au lapin et au
poulet Des leacutesions pulmonaires heacutepatiques et spleacuteniques importantes chez le cobaye et le lapin
identifiaient M bovis des leacutesions retrouveacutees uniquement chez le cobaye ou le poulet identifiaient
respectivement M tuberculosis et M avium (Kazda et al 2009) Cette meacutethode eacutetait utiliseacutee pour
lrsquoidentification de M bovis dans les tissus animaux et les eacutechantillons environnementaux comme le sol
et lrsquoeau apregraves deacutecontamination avec un meacutelange de javel et de soude (Maddock 1933 Maddock 1934
Maddock 1936 Williams amp Hoy 1930)
3 Meacutethodes de deacutetection et drsquoidentification des bacteacuteries du MTBC
Pour des raisons eacutepideacutemiologiques sanitaires (isolement du malade pour eacuteviter la
transmission) et theacuterapeutiques (mise en place drsquoune antibiotheacuterapie adeacutequate) il est primordial de
diffeacuterencier les membres du MTBC des autres mycobacteacuteries et drsquoidentifier preacuteciseacutement lrsquoespegravece du
MTBC en cause
Chapitre 1
69
a Caracteacuterisation pheacutenotypique et biochimique des mycobacteacuteries du MTBC
Depuis la deacutecouverte du bacille de la tuberculose jusqursquoau deacuteveloppement des outils
modernes les diffeacuterentes espegraveces de mycobacteacuteries ont eacuteteacute identifieacutees gracircce agrave leurs caracteacuteristiques
pheacutenotypiques et biochimiques Certaines de ces caracteacuteristiques sont encore exploiteacutees agrave lrsquoheure
actuelle en compleacutement des meacutethodes de biologie moleacuteculaire plus rapides Lrsquoaspect des colonies sur
un milieu solide leur deacutelai drsquoapparition leur nombre leur aspect rugueux ou lisse et la preacutesence drsquoune
pigmentation ou non permet drsquoorienter lrsquoidentification vers une mycobacteacuterie du MTBC ou une
NTM Dans le MTBC par exemple M tuberculosis apparait en 20 jours sur milieux agrave lrsquoœuf sous forme
de colonies ivoire en chou-fleur drsquoaspect rugueux Les colonies de M bovis apparaissent tardivement
(6 agrave 10 semaines) elles sont non pigmenteacutees petites fines et lisses (Carbonnelle et al 2003) Les
principaux tests biochimiques utiliseacutes pour diffeacuterencier les diffeacuterentes espegraveces du MTBC sont le test agrave
la niacine la reacuteduction des nitrates lrsquohydrolyse du Tween la sensibiliteacute au TCH (acide thiophegravene-2-
carboxylique) agrave la cycloseacuterine et au pyrazinamide Les reacutesultats des observations pheacutenotypiques et
biochimiques permettant de diffeacuterencier les principales espegraveces du MTBC sont preacutesenteacutes dans le
Tableau 2 Les tests biochimiques neacutecessitent des cultures riches et ne sont donc pas reacutealisables avant
la croissance avanceacutee de la bacteacuterie Chronophages et fastidieux ces tests sont remplaceacutes par drsquoautres
techniques plus rapides
Tableau 2 Identification des principales mycobacteacuteries du MTBC par des observations pheacutenotypiques et des tests biochimiques
M tuberculosis M africanum M bovis M bovis BCG
Aspect des colonies Rugueuses Rugueuses Lisses Rugueuses
Pigmentation Non pigmenteacute Non pigmenteacute Non pigmenteacute Non pigmenteacute
Croissance Lente Lente Lente Lente
Niacin test + - - -
Reacuteduction nitrates + +- - -
TCH R S S S
Cycloseacuterine S S R R
Pyrazinamide S S R R
+ test positif - test neacutegatif R reacutesistant S sensible
b Lrsquoidentification des mycobacteacuteries par recherche de biomarqueurs
Diffeacuterentes techniques de recherche de biomarqueurs potentiels de M tuberculosis par
chromatographie ont eacuteteacute deacuteveloppeacutees reacutecemment La chromatographie en phase liquide (HPLC High-
Chapitre 1
70
Performance Liquid Chromatography) a eacuteteacute deacuteveloppeacutee pour lrsquoanalyse des acides mycoliques (Butler amp
Guthertz 2001) La chromatographie en phase gazeuse coupleacutee agrave de la spectromeacutetrie de masse (GC-
MS) est utiliseacutee pour identifier des acides gras speacutecifiques de la paroi de M tuberculosis comme lrsquoacide
tuberculosteacutearique ou lrsquoacide hexacosanoiumlque dans des crachats (Cha et al 2009 Kaal et al 2009)
Lrsquoidentification de biomarqueurs speacutecifiques permet de diffeacuterencier M tuberculosis des NTM en GC-
MS en diagnostic direct sur des crachats (Dang et al 2013)
Depuis quelques anneacutees la spectromeacutetrie de masse de type MALDI-TOF (matrix-assisted laser
desorption ionization-time of flight) est utiliseacutee pour lrsquoidentification rapide des mycobacteacuteries du
complexe tuberculosis et de la majoriteacute des autres mycobacteacuteries agrave partir de cultures en milieu solide
et liquide (El Kheacutechine et al 2011 Bille et al 2012 Lotz et al 2010 Biswas amp Rolain 2013) Cette
technique permet drsquoeacutetablir un spectre de masse des diffeacuterentes proteacuteines constituant la mycobacteacuterie
(profil peptidique obtenu apregraves digestion trypsique des proteacuteines totales) La comparaison du profil
proteacuteique obtenu avec des bases de donneacutees internationales de plus en plus eacutetoffeacutees permet
drsquoidentifier un grand nombre drsquoespegraveces de mycobacteacuteries Elle peut ecirctre eacutegalement utiliseacutee comme
meacutethode de spoligotypage (Shitikov et al 2012) Cette meacutethode est simple rapide reproductible et
neacutecessite des consommables peu coucircteux (hors achat et entretien de lrsquoappareil) La qualiteacute du profil
proteacuteique obtenu est toutefois fortement deacutependante de la qualiteacute du protocole drsquoextraction des
proteacuteines (Saleeb et al 2011) La technique MALDI-TOF est probablement une technique drsquoavenir pour
lrsquoidentification rapide des mycobacteacuteries agrave croissance lente (le deacutelai de croissance reste toutefois le
facteur limitant) et plus geacuteneacuteralement des microorganismes pathogegravenes drsquointeacuterecirct clinique dans les
eacutechantillons A lrsquoheure actuelle elle ne permet pas de diffeacuterencier les diffeacuterentes espegraveces du MTBC
Elle nrsquoest pour lrsquoinstant applicable qursquoagrave des isolats bacteacuteriens issus de culture et non agrave des eacutechantillons
complexes
c Lrsquoidentification des mycobacteacuteries par des meacutethodes drsquohybridation coupleacutees ou
non agrave la PCR
La technique baseacutee sur lrsquohybridation de lrsquoADN est une meacutethode sensible rapide et simple
drsquoemploi couramment utiliseacutee en bacteacuteriologie humaine Elle est reacutealiseacutee sur une culture liquide ou
solide La technique est baseacutee sur lrsquohybridation drsquoune sonde moleacuteculaire conjugueacutee agrave un marqueur
chimioluminescent compleacutementaire de lrsquoARN ribosomal de la bacteacuterie cible Le kit commercial
AccuProbe (Gen Probe) fournit diffeacuterentes sondes permettant drsquoidentifier les Complexes tuberculosis
avium gordonae kansaii (sans discrimination des espegraveces) ainsi que les espegraveces M avium et
M intracellulare
Le systegraveme le plus utiliseacute agrave lrsquoheure actuelle est le GenXpert MTBRIF (Cepheid) utilisant la PCR
Chapitre 1
71
en temps reacuteel multi-colorimeacutetrique Il utilise 5 sondes Beacon (sondes avec une structure en eacutepingle agrave
cheveux qui fluorescent lors de lrsquohybridation) speacutecifiques du locus rpoB (reacutegion de 81 pb deacuteterminant
la reacutesistance agrave la rifampicine) des membres du MTBC Lrsquointeacuterecirct de cette technique est agrave la fois pratique
puisque lrsquoopeacuterateur deacutepose lrsquoeacutechantillon deacutecontamineacute dans une cassette contenant tous les reacuteactifs
lyophiliseacutes lrsquoappareil gegravere toutes les autres eacutetapes et diagnostique car elle couple la deacutetection des
bacteacuteries du MTBC et la mise en eacutevidence drsquoune eacuteventuelle reacutesistance agrave la rifampicine antituberculeux
de premiegravere ligne chez lrsquohomme
Drsquoautres systegravemes tels que lrsquoINNO-Lipareg Mycobacteria v2 (Fujirebio) le GenoType MTBC et le
GenoType Mycobacterium CMAS (Hain Lifescience) sont baseacutes sur lrsquohybridation drsquoune sonde
speacutecifique drsquoun Complexe ou drsquoune espegravece sur la reacutegion intergeacutenique variable 16S-23S apregraves
amplification PCR Ils permettent drsquoidentifier les Complexes tuberculosis et avium ainsi que de 16 agrave 40
espegraveces de NTM
Ces techniques sont essentiellement utiliseacutees dans le diagnostic humain mais elles sont parfois
deacutecrites pour lrsquoidentification du Complexes tuberculosis en diagnostic animal (Oloya et al 2007 Pavlik
et al 2005 Bollo et al 1998 Lieacutebana et al 1995) Bien que deacutejagrave utiliseacutees sur des eacutechantillons
environnementaux comme lrsquoeau (Aronson et al 1999) elles sont agrave utiliser avec preacutecaution car des
erreurs drsquoidentification ont eacuteteacute rapporteacutees (croisement entre des mycobacteacuteries saprophytes et
pathogegravenes) (Turenne 2004) en outre elles ne permettent pas la discrimination des diffeacuterentes
espegraveces du MTBC
d Le seacutequenccedilage
Lrsquooutil geacutenotypique de reacutefeacuterence pour lrsquoidentification des diffeacuterentes espegraveces de bacteacuteries est
le seacutequenccedilage du gegravene codant lrsquoARN de la sous-uniteacute 16S du ribosome (gegravene rrs) De nombreux couples
drsquoamorces ont eacuteteacute proposeacutes dans la litteacuterature pour amplifier entiegraverement ou partiellement le gegravene rrs
avant seacutequenccedilage mais lrsquoidentification des mycobacteacuteries par cette meacutethode nrsquoest pas toujours
possible agrave cause de la forte conservation de cette seacutequence Afin drsquoameacuteliorer la discrimination de ces
espegraveces le seacutequenccedilage total ou partiel drsquoautres gegravenes cibles a eacuteteacute deacuteveloppeacute tels les gegravenes de meacutenage
ciblant rpoB codant la sous-uniteacute β de lrsquoARN polymeacuterase (Adeacutekambi amp Drancourt 2004 Lee et al 2000)
hsp65 codant une proteacuteine de reacuteponse au stress thermique de 65 kDa (McNabb et al 2004 Ringuet et
al 1999 Telenti et al 1993) gyrA et gyrB codant les sous-uniteacutes α et β de lrsquoADN gyrase (enzyme
intervenant dans lrsquoenroulement et le deacuteroulement de lrsquoheacutelice drsquoADN) recA intervenant dans la
recombinaison homologue de lrsquoADN et dans sa reacuteparation (Blackwood et al 2000) sodA codant la super
oxyde dismutase (Zolg amp Philippi-Schulz 1994) secA1 codant la proteacuteine SecA1 intervenant dans le
seacutecreacutetion des proteacuteines agrave travers la membrane plasmique (Zelazny et al 2005) ou la reacutegion inter-
Chapitre 1
72
geacutenique encadreacutee par les ARNr 16S et 23S (appeleacutee ITS Internal Transcribed Spacer) (Roth et al 1998)
Malgreacute une ameacutelioration de la discrimination par lrsquoanalyse combineacutee de ces diffeacuterentes cibles le
seacutequenccedilage ne permet pas drsquoidentifier toutes les diffeacuterentes bacteacuteries du MTBC A cause de la forte
conservation des seacutequences dans le MTBC le seacutequenccedilage drsquoun grand nombre de gegravenes est neacutecessaire
pour discriminer les espegraveces (Kirschner et al 1993 Boumlddinghaus et al 1990) ce qui nrsquoest pas reacutealisable
en routine
Depuis lrsquoidentification de la plupart des gegravenes et des mutations responsables de la reacutesistance
des mycobacteacuteries aux antituberculeux le seacutequenccedilage est un des moyens utiliseacute pour identifier les
souches reacutesistantes La reacutesistance agrave la rifampicine par exemple est lieacutee agrave plusieurs mutations sur le gegravene
rpoB modifiant la structure de la sous-uniteacute β de lrsquoARN polymeacuterase et empecircchant la fixation de la
rifampicine sur lrsquoARN polymeacuterase conduisant agrave son inefficaciteacute (Zhang amp Yew 2009) Ces mutations
peuvent ecirctre deacutetecteacutees par seacutequenccedilage du gegravene amplifieacute par PCR
e Deacutetection et geacutenotypage des mycobacteacuteries du MTBC les marqueurs utiliseacutes
Les seacutequences drsquoinsertion
Les seacutequences drsquoinsertion (IS) correspondent agrave de petits eacuteleacutements geacuteneacutetiques transposables
Elles contiennent uniquement les informations geacuteneacutetiques neacutecessaires agrave leur transposition dans le
geacutenome de la bacteacuterie (transposase) On deacutenombre 42 IS dans le premier geacutenome annoteacute de M bovis
AF212297 (Garnier et al 2003) La seacutequence drsquoinsertion IS6110 speacutecifique du MTBC est la cible
historique utiliseacutee pour la deacutetection moleacuteculaire du Complexe tuberculosis en PCR (Thierry Cave et al
1990 Thierry Brisson-Noeumll et al 1990 Dziadek et al 2001) La meacutethode de reacutefeacuterence pour le
geacutenotypage de M tuberculosis est la RFLP-IS6110 (Restriction Fragment Lenght Polymorphism sur
lrsquoIS6110) Crsquoest la seacutequence drsquoinsertion la plus abondante dans le geacutenome de M tuberculosis qui en
contient de 1 agrave plus de 25 copies (Thierry Cave et al 1990 Thierry Brisson-Noeumll et al 1990) Elle
jouerait un rocircle dans lrsquoadaptation des bacteacuteries du MTBC agrave leur hocircte et dans leur virulence (Otal et al
2008 Safi et al 2004)
De nombreux systegravemes drsquoamorces ont eacuteteacute deacuteveloppeacutes pour la deacutetection de lrsquoIS6110 par PCR
dans les eacutechantillons biologiques (Dziadek et al 2001 El Kheacutechine et al 2009 Lieacutebana et al 1996
Savelkoul et al 2006) Diffeacuterents kits commerciaux utiliseacutes agrave lrsquoheure actuelle sont baseacutes sur la deacutetection
de lrsquoIS6110 notamment dans le diagnostic animal (Courcoul et al 2014) Cette cible a deacutejagrave eacuteteacute utiliseacutee
pour la deacutetection de M bovis dans des eacutechantillons inoculeacutes en PCR nicheacutee (Adams et al 2013) ainsi
que pour la deacutetection moleacuteculaire du MTBC dans du sol de lrsquoeau des seacutediments et des matiegraveres feacutecales
drsquoanimaux (Santos Santos et al 2015) (Tableau 3)
Chapitre 1
73
LrsquoIS6110 est eacutegalement utiliseacutee pour le typage des bacteacuteries du MTBC La comparaison des
souches repose sur le nombre drsquoIS6110 et leurs positions sur le chromosome apregraves extraction de lrsquoADN
digestion par une endonucleacutease (PvuII qui coupe lrsquoIS6110 sur un seul site) eacutelectrophoregravese transfert
sur membrane et hybridation (Van Soolingen et al 1991) Chaque fragment contenant une copie de
lrsquoIS6110 produit une bande (Figure 16) Cette meacutethode standardiseacutee beacuteneacuteficie de banques de donneacutees
de profils drsquohybridation internationales qui permettent des investigations eacutepideacutemiologiques Cette
meacutethode est beaucoup moins discriminante sur les souches agrave faible nombre drsquoIS (Allix et al 2006)
comme M bovis qui nrsquoen contient qursquoune agrave cinq copies et M bovis BCG qui nrsquoen contient qursquoune seule
copie (Van Soolingen et al 1991 Skuce et al 1994) Le reacutesultat est obtenu en une dizaine de jours
neacutecessite un volume important de culture et des manipulations fastidieuses De ce fait il a eacuteteacute
remplaceacute par des meacutethodes plus rapides indeacutependantes de la culture le spoligotypage et le typage
MLVA
Figure 16 Profils de restriction disolats de M tuberculosis obtenus suite agrave une RFLP-IS6110 Les isolats 3 5 6 9 et 10 ont le mecircme profil et posseacutedaient un lien eacutepideacutemiologique S repreacutesente le marqueur de poids moleacuteculaire (Source web site CDC)
Les courtes seacutequences drsquoADN reacutepeacuteteacutees
Le locus DR
Les souches du Complexe tuberculosis contiennent une reacutegion chromosomique particuliegravere
unique et figeacutee caracteacuteriseacutee par des seacutequences reacutepeacuteteacutees de 36 pb appeleacutee locus DR (Direct Repeat)
qui appartient agrave la famille des ADN reacutepeacutetitifs CRISPR (Clustered Regularly Interspersed Short
Chapitre 1
74
Palindromic Repeats) Ces DR sont seacutepareacutes par des espaceurs de 35 agrave 41 pb dont les seacutequences sont
uniques et polymorphes Ce polymorphisme associeacute agrave la deacuteleacutetion de certains espaceurs par des
reacutearrangements geacuteneacutetiques est exploiteacute dans le spoligotypage (en anglais spoligotyping
SPacer OLIGOnucleacuteotide TYPING) (Kamerbeek et al 1997) Par exemple les espaceurs 3 9 16 et 39 agrave
43 sont absents chez M bovis M caprae M microti et M pinnipedii (Smith et al 2006) comme
preacutesenteacute dans la Figure 17 Le geacutenotypage par spoligotypage est baseacute sur lrsquoamplification par PCR du
locus DR le marquage des amplicons et leur hybridation par des oligonucleacuteotides fixeacutes agrave une
membrane (spoligotypage 2D) Le profil final est deacutetecteacute par chimioluminescence (Kamerbeek et al
1997) Dans la technologie Luminex les oligonucleacuteotides sont fixeacutes sur des microbilles agrave signature
spectrale diffeacuterente (spoligotypage 3D) (Zhang et al 2010) Le spolygotypage standard analyse 43
espaceurs et permet drsquoobtenir des donneacutees eacutepideacutemiologiques et phylogeacuteneacutetiques deacuteterminantes (Sola
et al 2001) Il est largement utiliseacute pour typer les souches de M bovis notamment en France (Haddad
et al 2001) Cette meacutethode est reacutealisable sur de faibles quantiteacutes drsquoADN mecircme impur et des bacteacuteries
non cultivables Crsquoest une technique robuste rapide simple de reacutealisation dont le reacutesultat est facile agrave
interpreacuteter car il fournit un code binaire (pour chaque espaceur preacutesent 1 ou absent 0) faisant lrsquoobjet
drsquoune convention internationale (Dale et al 2001) Toutefois le spoligotypage ne preacutesente pas un
pouvoir de reacutesolution suffisant pour la diffeacuterenciation de certains groupes clonaux car il ne cible qursquoun
seul locus sur lrsquoensemble du geacutenome
Chapitre 1
75
C
Figure 17 Principe du spoligotypage des bacteacuteries du MTBC dapregraves Kamerbeek et al (1997) A Repreacutesentation du locus DR de M tuberculosis H37Rv et M bovis BCG B Amplifications des espaceurs par un couple damorces (a et b) et amplicons obtenus C Profils drsquohybridation sur membrane (un carreacute noir signale la preacutesence de lrsquoespaceur les parties blanches leur absence)
76
Gegravene ou locus
cible Espegraveces cibles PCR
Taille du
produit PCR
(pb)
Limites de deacutetection
(bacteacuteries g-1 sol
ou mL-1)
Matrices analyseacutees
Quantification si
deacutetection positive (nb
gegravenes ou UFC g-1 sol
ou mL-1)
Reacutefeacuterence Utiliseacute par
mpb64 MTBC PCR classique 543 101-102 Sols (terriers et preacutes
adjacents) 10 - 25 times 103 (Gormley 1999) (Young et al 2005)
mpb70 MTBC PCR classique 471 101-102 Sols (terriers et preacutes
adjacents) 10 - 25 times 103 (Gormley 1999)
(Young et al 2005)
(Courtenay et al 2006)
Gegravene codant
lrsquoARNr 16S
Mycobacteacuteries
agrave croissance
lente
RT-PCR 605 102-103 Sols (terriers et preacutes
adjacents) -
(Young et al
2005)
(Young et al 2005)
(Courtenay et al 2006)
IS6110 MTBC PCR nicheacutee PCR1 252
PCR2 116 -
Sol Eau
Foin Maiumls -
(Adams et al
2013) (Adams et al 2013)
IS6110 MTBC PCR nicheacutee 110
Sol 40 times 104
Eau 50 times 105
Sol
Seacutediment
Eau et fegraveces
(Soo et al 2006) (Santos Santos et al 2015)
RD12 M bovis
M caprae PCR classique 306
Sol 10 times 106
Eau 50 times 105
Sol
Seacutediment
Eau et fegraveces
(Warren et al
2006) (Santos Santos et al 2015)
RD12
M microti
M tuberculosis
M africanum
M pinnipedii
PCR semi-
nicheacutee 369
Sol 40 times 104
Eau 50 times 105
Sol
Seacutediment
Eau et fegraveces
(Warren et al
2006) (Santos Santos et al 2015)
RD4 (zone
flanquante) M bovis qPCR 142
42 times 105
(Pontiroli et al 2011)
10 times 105
(Pontiroli et al 2011)
Sols de terriers
Latrines
68 times 104 agrave 54 times 106
(Sweeney et al 2007)
20 times 104 agrave 10 times 108
(Travis et al 2011)
(Sweeney et al
2007)
(Sweeney et al 2007)
(King Murphy James Travis
Porter Sawyer et al 2015)
(King Murphy James Travis
Porter Hung et al 2015)
(Pontiroli et al 2011)
(Roug et al 2014)
Rv1510 (RD4) MTBC sauf
M bovis
PCR classique 1 033 - Sols de terriers
Latrines
- (Huard et al
2003)
(Courtenay et al 2006)
(Sweeney et al 2006)
(Sweeney et al 2007)
Tableau 3 Cibles moleacuteculaires deacutecrites dans la litteacuterature pour la deacutetection du Complexe tuberculosis ou de M bovis dans les eacutechantillons environnementaux
Chapitre 1
77
Les mini-satellites
Les mini-satellites correspondent agrave de courtes seacutequences drsquoADN reacutepeacuteteacutees (de 10 agrave 1 000 pb) et
conseacutecutives Pour pallier le manque de discrimination du spoligotypage un typage sur diffeacuterents loci
a eacuteteacute deacuteveloppeacute le MLVA (Multi Locus VNTR Analysis) baseacute sur lrsquoanalyse drsquoeacuteleacutements situeacutes sur des
seacutequences intergeacuteniques (les MIRU Mycobacterial interspersed Repetitive Unit) composeacutes de
seacutequences reacutepeacuteteacutees en tandem dont le nombre et la seacutequence sont variables (notamment les VNTR
Variable Number Tandem Repeat et les ETR Exact Tandem Repeat) (Supply et al 2001 Supply et al
2006) Lrsquoeacutetude de loci disperseacutes dans le geacutenome apporte une discrimination robuste des souches Le
geacutenotypage des souches de M tuberculosis ou M bovis par la meacutethode MIRU-VNTR repose sur la
recherche du nombre drsquoexemplaires des seacutequences reacutepeacuteteacutees de chaque locus eacutetudieacute Plus le nombre
de loci analyseacutes est important plus la discrimination est fine ainsi le MIRU-VNTR peut porter sur 12
15 ou 24 loci pour le typage des souches de M tuberculosis selon lrsquoobjectif envisageacute (eacutepideacutemiologie ou
phylogeacutenie) (Supply et al 2001 Supply et al 2006) Concernant le typage des souches animales de
M bovis diffeacuterentes combinaisons de marqueurs VNTR ont eacuteteacute eacutevalueacutees (Roring et al 2002 Allix et al
2006 Skuce et al 2002 Boniotti et al 2009) et ont montreacute que lrsquoanalyse de 5 agrave 8 loci suffisait pour
obtenir un pouvoir de discrimination supeacuterieur agrave 90 En France 8 loci ont eacuteteacute seacutelectionneacutes sur leur
polymorphisme et leur pouvoir de discrimination et utiliseacutes pour typer plus de mille isolats cliniques
recueillis entre 1978 et 2011 (Hauer et al 2015) Dans cette eacutetude les reacutesultats de geacutenotypage se
preacutesentent sous forme drsquoun code agrave 8 chiffres (Tableau 4) Pour un mecircme spoligotype SB0120 le
geacutenotypage MIRU-VNTR permet de diffeacuterencier les souches retrouveacutees en Cocircte drsquoOr de celles de
Dordogne Ces reacutesultats ont eacutegalement permis de deacutemontrer lrsquohomogeacuteneacuteiteacute des profils geacuteneacutetiques
retrouveacutes chez les animaux domestiques et sauvages dans une zone geacuteographique confirmant
lrsquohypothegravese de la transmission interspeacutecifique de lrsquoinfection
Tableau 4 Spoligotypes et geacutenotypes MIRU-VNTR des souches de M bovis isoleacutees chez les bovins et la faune sauvage en Cocircte dOr et en Dordogne (drsquoapregraves Hauer et al 2015)
Spoligotype Geacutenotype MIRU-VNTR (8 loci cibleacutes)
Origine des isolats 2165 2461 0577 580 2163a 2163b 4052 3232
SB0120
Cocircte drsquoOr 5 5 4 3 11 4 5 6
Bovins blaireaux
sangliers
SB0120
Dordogne 5 3 5 3 9 4 5 6
Bovins blaireaux
sangliers
SB0134
Cocircte drsquoOr 6 4 5 3 6 4 3 6
Bovins blaireaux
sangliers cerfs
Chapitre 1
78
La combinaison du spoligotypage et de la MLVA optimise la discrimination des souches pour
les suivis eacutepideacutemiologiques et apporte des informations phylogeacuteneacutetiques inteacuteressantes Elle permet
eacutegalement de suivre lrsquoeacutemergence de variants clonaux chez un animal ou dans un troupeau infecteacute en
fonction du temps (Navarro et al 2015 Rodriguez-Campos et al 2014) Toutefois les loci agrave analyser
doivent ecirctre adapteacutes en fonction des complexes clonaux rencontreacutes dans la reacutegion ou le pays eacutetudieacutes
(Boniotti et al 2009 Hauer et al 2015) Diffeacuterentes bases de donneacutees internationales sont disponibles
pour la comparaison des reacutesultats de spoligotypage et de MIRU-VNTR comme SpolDB4 SITVITWEB
MIRU-VNTRplus et Mbovisorg (regroupe les spolygotypes des espegraveces deacuteleacuteteacutees pour RD9) (Couvin amp
Rastogi 2014 Smith amp Upton 2012)
Reacutegions de Diffeacuterence (RD) et Single Nucleotide
Polymorphism (SNP)
Les Reacutegions de Diffeacuterences (RD1 agrave RD14) identifieacutees chez les bacteacuteries du MTBC correspondent
agrave des deacuteleacutetions et des reacutearrangements geacuteneacutetiques irreacuteversibles et discriminants Ces RD sont
speacutecifiques drsquoune espegravece ou communes agrave plusieurs espegraveces Le plus grand nombre de RD est observeacute
dans le geacutenome de M bovis BCG qui preacutesente 14 RD par comparaison agrave M tuberculosis H37Rv (Gordon
et al 1999 Brosch et al 2002) En parallegravele six seacutequences deacuteleacuteteacutees chez M tuberculosis H37Rv (les
RvD) et une seacutequence absente de toutes les souches de M tuberculosis (TbD1) sont preacutesentes chez les
autres bacteacuteries du MTBC (Gordon et al 1999 Brosch et al 2002) Lrsquoabsence de RD4 est speacutecifique agrave
M bovis et M bovis BCG et permet de les diffeacuterencier des autres membres du MTBC (Huard et al 2003
Huard et al 2006) La RD4 a eacuteteacute utiliseacutee dans diffeacuterentes eacutetudes pour la deacutetection de M bovis dans les
eacutechantillons environnementaux (Courtenay et al 2006 Sweeney et al 2007 King Murphy James
Travis Porter Sawyer et al 2015 Travis et al 2011 Roug et al 2014 Pontiroli et al 2011) Dans une
eacutetude reacutecente Santos et al (2015) ont associeacute la deacutetection du MTBC par lrsquoIS6110 agrave une identification
de lrsquoespegravece par amplification de deux reacutegions de la RD12 pour diffeacuterencier M bovisM caprae des
autres espegraveces du Complexe (Tableau 3)
Lrsquointeacuterecirct des RD est bien plus phylogeacuteneacutetique que geacutenotypique puisque lrsquoeacutetude de leur
apparition successive au cours de lrsquoeacutevolution a permis drsquoeacutetablir preacuteciseacutement la phylogeacutenie des
diffeacuterentes espegraveces comme preacutesenteacute preacuteceacutedemment dans la Figure 3 (Huard et al 2006 Rodriguez-
Campos et al 2014 Dippenaar et al 2015)
Les Single Nucleotide Polymorphism (SNPs) correspondent agrave des mutations ponctuelles drsquoune
base dans un geacutenome Ils peuvent ecirctre synonymes (le changement de base nrsquoaffecte pas la seacutequence
proteacuteique correspondante) ou non synonymes (la seacutequence proteacuteique est affecteacutee) et repreacutesentent
Chapitre 1
79
une variabiliteacute du geacutenome utile pour la caracteacuterisation de la variabiliteacute drsquoun groupe clonal et lrsquoeacutetude
des relations phylogeacuteneacutetiques entre les souches (Mathema et al 2006)
f Identification par seacutequenccedilage complet des geacutenomes
Le seacutequenccedilage complet des geacutenomes (WGS Whole Genome Sequencing) beacuteneacuteficie depuis
quelques anneacutees de nouvelles technologies (NGS Next Generation Sequencing) plus rapides et moins
coucircteuses que la meacutethode de Sanger Diffeacuterentes technologies coexistent agrave lrsquoheure actuelle dont les
plus utiliseacutees sont le pyroseacutequenccedilage 454 (toutefois en fin drsquoutilisation) la technique Illumina et la
technique Ion PGM (Ion Personal Genome Machine) (Desikan amp Narayanan 2015) Ces techniques ont
reacutevolutionneacute le geacutenotypage en fournissant le plus haut niveau de discrimination geacuteneacutetique possible
Ces techniques parfois coupleacutees au typage des SNPs permettent de suivre la dynamique de
transmission drsquoune maladie de caracteacuteriser lrsquoantibioreacutesistance drsquoune souche et drsquoeacutevaluer la divergence
geacuteneacutetique de souches qui ont les mecircmes profils de restriction ou de geacutenotypage (Schuumlrch amp van
Soolingen 2012) Une eacutetude irlandaise sur des souches de M bovis isoleacutees de bovins et de blaireaux
vivant dans les mecircmes zones a permis gracircce au WGS et agrave lrsquoanalyse de 39 SNPs de deacutemontrer la
circulation et la persistance locales drsquoune souche entre animaux sauvages et domestiques (Biek et al
2012) Une eacutetude reacutecente portant sur le seacutequenccedilage complet de 82 souches de M bovis isoleacutees en
France entre 1978 et 2011 a permis de deacutemontrer la congruence entre le typage par
spoligotypageMIRU-VNTR et le typage des souches par analyse des SNP (Hauer 2015) suggeacuterant que
certains SNP pourraient ecirctre utiliseacutes comme cibles dans le geacutenotypage de M bovis Dans un futur
proche le WGS sera probablement lrsquooutil qui offrira lrsquoinformation la plus complegravete sur le geacutenotype des
souches les liens phylogeacuteneacutetiques entre souches et qui permettra de deacutecoder les scheacutemas de
transmission de lrsquoinfection entre animaux (Biek et al 2012 Pepperell et al 2013 Rodriguez-Campos et
al 2014)
Les puces (microarrays) agrave ADN ou ARN sont utiliseacutees pour la comparaison de geacutenomes complets
des espegraveces du MTBC et ont notamment apporteacute des informations sur lrsquoexpression de gegravenes de la
virulence chez M bovis (Voskuil et al 2004) ainsi que sur lrsquoidentification de seacutequences polymorphiques
distinguant les principaux complexes clonaux (Berg et al 2011 Muumlller et al 2009 Smith et al 2011)
Ces techniques appartiennent toutefois au domaine de la recherche et ne sont pas utiliseacutees en routine
g Meacutethode speacutecifique drsquoidentification des mycobacteacuteries animales utiliseacutee par le
Laboratoire National de reacutefeacuterence de la tuberculose bovine LNR TB
Lrsquoidentification des mycobacteacuteries isoleacutees sur des animaux est reacutealiseacutee au LNR TB de Maisons-
Chapitre 1
80
Alfort En premiegravere intention quatre systegravemes de deacutetection moleacuteculaires ciblant des seacutequences
drsquoinsertion (IS6110 IS1081 IS1245) et le gegravene atpE sont utiliseacutes en qPCR pour orienter lrsquoidentification
(Tableau 5)
Tableau 5 Systegravemes de deacutetection utiliseacutes en qPCR en premiegravere intention pour lidentification dune mycobacteacuterie dorigine animale et reacutesultats obtenus + amplification de la cible - absence drsquoamplification de la cible
ARNr 16S IS1245 IS901 IS6110 IS1081 IS1561rsquo RD4 RD7 atpE
Complexe tuberculosis
M bovis M tuberculosis
M microti M caprae
+ + + +
- - - -
- - - -
+ + + +
+ + + +
+ + - +
- + + +
- + + -
- - - -
Complexe avium + + +- - - - - + +
Autres mycobacteacuteries
+ - - - - - - + +
(Slana et
al 2010) (Slana et al
2010) (Courcoul et
al 2014) (cette thegravese)
(Radomski et al 2013)
Lrsquoidentification des espegraveces du complexe tuberculosis (IS6110 et IS1081 positives) est ensuite
reacutealiseacutee par geacutenotypage gracircce agrave la technique de spoligotypage (Zhang et al 2010) Lrsquoamplification ou
non des IS1245 et IS901 permet drsquoidentifier les espegraveces Mah (IS1245 et IS901 neacutegatives) Maa (IS1245
et IS901 positives) et Mah (IS1245 positive et IS901 neacutegative) (Slana et al 2010) Lrsquoidentification des
autres mycobacteacuteries repose sur le seacutequenccedilage de lrsquohsp65 puis de la comparaison par BLAST avec une
base de donneacutees (Telenti et al 1993)
De par le grand nombre drsquoespegraveces de mycobacteacuteries et la forte identiteacute des espegraveces dans le
Complexe MTBC la deacutetection et le geacutenotypage des souches du MTBC reposent obligatoirement sur
lrsquoutilisation conjointe de diffeacuterentes techniques En fonction des eacutechantillons disponibles et des
objectifs souhaiteacutes qursquoils soient diagnostiques eacutepideacutemiologiques ou phylogeacuteneacutetiques le choix des
techniques agrave utiliser doit ecirctre reacutefleacutechi et pertinent
82
Objectifs de la Thegravese
Depuis 2004 la situation de la Cocircte drsquoOr vis-agrave-vis de la tuberculose bovine est preacuteoccupante
Malgreacute la mise en place de moyens de surveillance et de lutte en premier lieu dans les cheptels bovins
puis dans la faune sauvage lrsquoeacuteradication de la maladie semble difficile Depuis 2011 on observe une
diminution du nombre des foyers bovins et une stabiliteacute du nombre de cas observeacutes dans la faune
sauvage avec une concentration spatiale des foyers dans des petites valleacutees Dans ces valleacutees ougrave
cohabitent faune sauvage et bovins la transmission indirecte du bacille entre animaux sauvages et
domestiques par le biais de lrsquoenvironnement est fortement suspecteacutee pour plusieurs cas bovins Bien
que plusieurs eacutetudes aient deacutemontreacute la preacutesence de M bovis dans diffeacuterentes matrices
environnementales naturelles sur les lieux de vie drsquoanimaux domestiques et sauvages infecteacutes
notamment en Angleterre et en Espagne aucune donneacutee nrsquoest disponible sur la contamination
environnementale potentielle par M bovis dans les zones endeacutemiques de tuberculose bovine en
France et plus particuliegraverement en Cocircte drsquoOr
Pour reacutepondre agrave cette question le premier objectif du travail de thegravese a eacuteteacute de deacutevelopper et de valider
sur des eacutechantillons inoculeacutes artificiellement des outils moleacuteculaires speacutecifiques et sensibles pour la
deacutetection des souches du MTBC et de M bovis dans les substrats environnementaux naturellement
contamineacutes tels que le sol lrsquoeau et les seacutediments et les matiegraveres feacutecales animales Nous avons
eacutegalement deacuteveloppeacute un protocole drsquoextraction-deacutecontamination des mycobacteacuteries du sol coupleacute agrave
une technique originale de suivi de croissance des mycobacteacuteries en milieu liquide dans lrsquoobjectif
drsquoisoler les souches environnementales de M bovis (Chapitre 2)
Les outils moleacuteculaires deacuteveloppeacutes ont eacuteteacute utiliseacutes pour la recherche de M bovis dans un grand nombre
drsquoeacutechantillons environnementaux (sol eau seacutediments fegraveces animales et membres de la micro- ou
meacuteso-faune des sols) preacuteleveacutes dans la zone endeacutemique de Cocircte drsquoOr et plus preacuteciseacutement dans des
zones partageacutees par la faune domestique et sauvage infecteacutee afin de preacuteciser le niveau de
contamination des lieux de vie des bovins et des animaux sauvages (Chapitre 3)
Face aux eacutetudes parfois controverseacutees sur le rocircle des facteurs abiotiques dans la persistance de
M bovis dans le sol nous avons rechercheacute lrsquoeffet de la nature du sol et de la tempeacuterature sur la dureacutee
de survie de M bovis SB0120 en conditions de laboratoire (Chapitre 4)
Nous avons enfin eacutetudieacute le rocircle de la faune du sol et plus particuliegraverement des lombrics tregraves abondants
dans les pacirctures et gros consommateurs de matiegravere organique dans la circulation et la disseacutemination
de M bovis dans les sols (Chapitre 5)
CHAPITRE 2
Deacuteveloppement et optimisation drsquooutils pour la deacutetection
de Mycobacterium bovis dans les eacutechantillons
environnementaux
PARTIE CONFIDENTIELLE agrave lrsquoexception des meacutethodes de deacutetection moleacuteculaire
86
CHAPITRE 2 - Deacuteveloppement et optimisation drsquooutils pour la
deacutetection de Mycobacterium bovis dans les eacutechantillons
environnementaux
A DEVELOPPEMENT DrsquoOUTILS MOLECULAIRES ADAPTES AUX MATRICES ENVIRONNEMENTALES
Les diffeacuterents systegravemes moleacuteculaires utiliseacutes pour la deacutetection de M bovis dans
lrsquoenvironnement citeacutes dans la litteacuterature sont syntheacutetiseacutes dans le Tableau 3 En consideacuterant la diversiteacute
et lrsquoabondance des mycobacteacuteries potentiellement preacutesentes dans lrsquoenvironnement nous avons
eacutelimineacute les systegravemes ciblant toutes les espegraveces du MTBC notamment lrsquoIS6110 car bien que tregraves
sensible et speacutecifique sur les eacutechantillons biologiques et tregraves utiliseacutee en diagnostic humain et
veacuteteacuterinaire nous avons montreacute qursquoelle nrsquoeacutetait pas utilisable en diagnostic environnemental En effet
nous avons effectueacute des essais preacuteliminaires avec le kit LSI VetMAXtrade Mycobacterium tuberculosis PCR
Kit (Life Technologies) utiliseacute en diagnostic veacuteteacuterinaire sur 25 eacutechantillons de sol provenant de zones
geacuteographiques non impacteacutees par la tuberculose bovine Pour tous ces sols lrsquoIS6110 a donneacute un signal
moleacuteculaire quantifiable teacutemoignant de lrsquoabsence de speacutecificiteacute de ce systegraveme pour la matrice sol
De plus M microti membre du MTBC agent de la tuberculose des petits rongeurs est susceptible
drsquoecirctre retrouveacute dans le sol (Courtenay et al 2006) et a deacutejagrave eacuteteacute isoleacute sur des feacutelins et des animaux
sauvages en Cocircte drsquoOr (Laprie et al 2013 Michelet et al 2015) Nous avons donc deacuteveloppeacute des
systegravemes moleacuteculaires qui nrsquoamplifient pas M microti
Enfin M bovis est une mycobacteacuterie pathogegravene de classe III qui neacutecessite une manipulation dans un
laboratoire de confinement L3 Pour des raisons techniques et pour srsquoaffranchir de la contrainte de
confinement dans la mise au point de la technique de PCR nous avons utiliseacute une des souches
atteacutenueacutees de M bovis et plus preacuteciseacutement M bovis BCG Pasteur 1173P2 manipulable dans un
laboratoire de type L2 disponible agrave lrsquoINRA de Dijon Nous avons donc choisi des cibles moleacuteculaires
eacutegalement preacutesentes chez M bovis BCG
Pour augmenter la speacutecificiteacute de la deacutetection moleacuteculaire dans les matrices environnementales
complexes nous avons choisi drsquoutiliser la PCR quantitative avec une sonde TaqManreg qui est formeacutee
par un nucleacuteotide lieacute de faccedilon covalente agrave un fluorophore (reporter) en 5rsquo et agrave un deacutesactivateur
(quencher) en 3rsquo Le clivage de la sonde par la Taq polymeacuterase apregraves lrsquohybridation agrave la seacutequence drsquoADN
compleacutementaire eacuteloigne le quencher du fluorophore qui eacutemet une fluorescence mesureacutee en temps
reacuteel dans un thermocycleur adapteacute
Chapitre 2
87
Le systegraveme moleacuteculaire deacutecrit preacuteceacutedemment par Sweeney et al (2007) ciblant les marges de
la reacutegion RD4 deacuteleacuteteacutee uniquement chez M bovis et M bovis BCG nous a particuliegraverement inteacuteresseacutes
Des amorces internes agrave la seacutequence RD4 ont eacuteteacute utiliseacutees en PCR pour la discrimination de M bovis
avec les autres membres du MTBC (pas de signal pour M bovis mais un signal pour les autres membres
du MTBC) (Huard et al 2003 Courtenay et al 2006) Des amorces de PCR classique ont eacutegalement eacuteteacute
dessineacutees sur les zones flanquantes de la reacutegion RD4 (Brosch et al 2002) Ces amorces ont eacuteteacute adapteacutees
agrave la technologie TaqManreg avec une amorce et une forward localiseacutees sur les marges de la seacutequence
deacuteleacuteteacutee et une sonde agrave cheval sur la zone de deacuteleacutetion (Sweeney et al 2007) La speacutecificiteacute et la
validation de ce systegraveme sur diffeacuterents eacutechantillons environnementaux tels que des sols de terriers
des fegraveces de blaireaux et de buffles africains ont eacuteteacute deacutemontreacutees dans plusieurs eacutetudes (Sweeney et al
2007 Roug et al 2014 Travis et al 2011 Pontiroli et al 2013 King et al 2015) Ce systegraveme de deacutetection
nous paraissant a priori speacutecifique pour la discrimination de M bovis dans le MTBC et adapteacute aux
matrices complexes nous lrsquoavons inclus dans lrsquoeacutetude Toutefois lrsquoamorce Reverse du systegraveme de
deacutetection a eacuteteacute modifieacutee par nos soins afin de reacuteduire lrsquoamplicon obtenu de 146 agrave 116 bases taille plus
adapteacutee agrave la technologie TaqManreg
Face au manque de systegravemes discriminants disponibles dans la litteacuterature nous avons dessineacute
deux nouveaux systegravemes Nous avons choisi de cibler la seacutequence drsquoinsertion IS1561rsquo (Rv3349c) car elle
est preacutesente chez tous les membres du MTBC sauf M microti (Gordon et al 1999 Huard et al 2003
Brodin et al 2002) Elle appartient agrave la famille des ISL3 qui regroupe des copies drsquoIS deacutefectueuses chez
M tuberculosis elles sont alors annoteacutees avec une apostrophe
La troisiegraveme cible a eacuteteacute choisie dans une reacutegion chromosomique de M tuberculosis appeleacutee
RD1 preacutesente chez les souches virulentes de M tuberculosis et M bovis mais partiellement deacuteleacuteteacutee
chez M bovis BCG (Mahairas et al 1996) et M microti (Brodin et al 2002) Cette reacutegion contient
notamment un cluster de 6 gegravenes (Rv3871 agrave Rv3876) qui fait partie des gegravenes codant les proteacuteines
antigeacuteniques ESAT-6 (Early Secreted Antigenic Target 6kD) et CFP-10 (Culture Filtrat Protein 10kD)
responsables de la reacuteponse immunitaire de lrsquohocircte M bovis BCG et M microti sont les deux seuls
membres du MTBC agrave preacutesenter une deacuteleacutetion de ce cluster deacuteleacutetion probablement en partie
responsable de leur absence de virulence et de leur utilisation en vaccination (Pym et al 2002) Chez
M bovis BCG la reacutegion aval de ce cluster est deacuteleacuteteacutee la reacutegion deacuteleacuteteacutee est appeleacutee RD1bcg (Figure 18)
Chez M microti crsquoest la reacutegion amont du cluster qui est absente la reacutegion deacuteleacuteteacutee est appeleacutee RD1mic
Nous avons plus particuliegraverement cibleacute le locus Rv3866 preacutesent chez M tuberculosis et dans toutes les
souches de M bovis et M bovis BCG mais absente chez M microti (Brodin et al 2002) M avium et
M smegmatis (Mahairas et al 1996)
Chapitre 2
88
Figure 18 Diffeacuterences geacutenomiques de la reacutegion RD1 entre M bovis BCG (A) M microti (C) par comparaison agrave M tuberculosis H37Rv (B) drsquoapregraves Brodin et al 2002 et localisation du locus Rv3866 RD1mic correspond agrave la seacutequence de M tuberculosis deacuteleacuteteacutee chez M microti RD1bcg correspond agrave la seacutequence de M tuberculosis deacuteleacuteteacutee chez M bovis BCG
Les seacutequences drsquointeacuterecirct ont eacuteteacute rechercheacutees pour les membres du MTBC et certaines MNT sur
la base de donneacutees GenBank Un alignement de ces seacutequences a eacuteteacute reacutealiseacute gracircce au portail drsquoanalyses
bioinformatiques MOBYLE PASTEUR pour rechercher les reacutegions conserveacutees et les reacutegions de
diffeacuterence Le dessin des oligonucleacuteotides a eacuteteacute reacutealiseacute grace au logiciel en ligne Primer3
(httpbioinfouteeprimer3-040) Les systegravemes de deacutetection ont eacuteteacute testeacutes in silico gracircce agrave lrsquooutil
BLAST (Basic Local Alignment Search Tool) afin drsquoeacutevaluer la speacutecificiteacute de nos systegravemes pour chaque
cible par rapport agrave une base de donneacutees de geacutenomes seacutequenceacutes
La validation (speacutecificiteacute et seuil de deacutetection) de ces trois systegravemes ainsi que leur utilisation
dans le screening drsquoun nombre important drsquoeacutechantillons environnementaux sont preacutesenteacutees dans le
Chapitre 3 dans lrsquoarticle intituleacute laquo First molecular detection of Mycobacterium bovis in environmental
samples from a French region with endemic bovine tuberculosis raquo Les reacutesultats des analyses reacutealiseacutees
en 2013 sont preacutesenteacutes dans un article publieacute en 2014 dans la revue de lrsquoAEEMA (association pour
leacutetude de leacutepideacutemiologie des maladies animales) en Annexe 1
A
B
C
Chapitre 2
89
CHAPITRE 3
Etude de la preacutevalence environnementale de
Mycobacterium bovis dans la zone endeacutemique de
tuberculose bovine de Cocircte drsquoOr et reacutesultats obtenus
en Charente
Elodie Barbier Maria Laura Boschiroli Eric Gueneau Murielle Rochelet Ariane Payne Krystel
de Cruz AnneLaure Blieux Ceacuteline Fossot Alain Hartmann
First molecular detection of Mycobacterium bovis in environmental samples from a French
region with endemic bovine tuberculosis
(Article accepteacute en cours de publication Journal of Applied Microbiology)
93
CHAPITRE 3 - Etude de la preacutevalence environnementale de
Mycobacterium bovis dans la zone endeacutemique de tuberculose
bovine de Cocircte drsquoOr et reacutesultats obtenus en Charente
A AVANT-PROPOS
Dans la synthegravese bibliographique preacutesenteacutee dans le chapitre 1 nous avons deacutecrit la situation
eacutepideacutemiologique complexe de la tuberculose bovine en Cocircte drsquoOr Dans ce deacutepartement
Mycobacterium bovis circule dans un systegraveme multi-hocirctes complexe incluant les bovins les blaireaux
les cervideacutes et les sangliers sur une zone geacuteographique limiteacutee (Payne 2014) La faune sauvage infecteacutee
et potentiellement excreacutetrice est suspecteacutee drsquoecirctre agrave lrsquoorigine de la contamination ou recontamination
de certains foyers bovins dans cette zone (ANSES 2011 Courcoul Moutou amp Vialard 2013) Le mode
de transmission indirecte est suspecteacute car les contacts directs entre bovins et animaux sauvages sont
rarement deacutecrits (Drewe OrsquoConnor Weber McDonald amp Delahay 2013 Kukielka et al 2013) Ce mode
de transmission repose sur la contamination de matrices environnementales par des mycobacteacuteries
viables et virulentes agrave la faveur de lrsquoexcreacutetion animale puis sur lrsquointeraction drsquoanimaux avec ces
matrices Deacutemontreacutee par des meacutethodes moleacuteculaires etou culturales en Angleterre et en Espagne sur
des sites partageacutes par les espegraveces sauvages et les bovins (King et al 2015 Santos et al 2015 Sweeney
et al 2006 2007) la contamination environnementale par M bovis nrsquoa pas encore fait lrsquoobjet drsquoeacutetudes
dans les zones endeacutemiques franccedilaises
Afin drsquoeacutevaluer la preacutevalence et lrsquoexistence de reacuteservoirs potentiels de M bovis dans
lrsquoenvironnement des bovins et des animaux sauvages infecteacutes en Cocircte drsquoOr nous avons deacuteveloppeacute et
valideacute des systegravemes de deacutetection moleacuteculaire ou modifieacute des systegravemes existants adapteacutes aux matrices
environnementales complexes pour deacutetecter les espegraveces du MTBC et plus speacutecifiquement M bovis
Ces systegravemes ont eacuteteacute utiliseacutes pour analyser plus de 400 eacutechantillons environnementaux (sol eau
seacutediments microfaune fegraveces) preacuteleveacutes en Cocircte drsquoOr Notre strateacutegie drsquoeacutechantillonnage a cibleacute des
pacirctures suspectes sur lesquelles des bovins infecteacutes ont transiteacute et autour desquelles des animaux
sauvages infecteacutes ont eacuteteacute deacutetecteacutes et plus particuliegraverement les zones pouvant ecirctre partageacutees par les
diffeacuterentes espegraveces notamment les zones drsquoabreuvement Nous avons eacutegalement analyseacute des
eacutechantillons provenant de sites freacutequenteacutes exclusivement par la faune sauvage notamment des sols
de terriers et des latrines de blaireaux localiseacutes agrave proximiteacute du site de pieacutegeage de blaireaux infecteacutes
ainsi que des seacutediments preacuteleveacutes dans les trous drsquoeau ou souilles freacutequenteacutes par les sangliers et
cervideacutes
Chapitre 3
95
B FIRST MOLECULAR DETECTION OF MYCOBACTERIUM BOVIS IN ENVIRONMENTAL SAMPLES FROM A FRENCH REGION WITH ENDEMIC BOVINE TUBERCULOSIS
E Barbier12 ML Boschiroli3 E Gueneau4 M Rochelet2 A Payne1 Krystel de Cruz3 AL Blieux5 C
Fossot6 A Hartmann1
1 INRA UMR 1347 Agroeacutecologie Dijon Cedex France
2 Universiteacute de Bourgogne Franche Comteacute UMR 1347 Agroeacutecologie Dijon Cedex France
3 Universiteacute Paris-Est Laboratoire National de Reacutefeacuterence de la Tuberculose Uniteacute de Zoonoses
Bacteacuteriennes Laboratoire de Santeacute Animale ANSES Maisons-Alfort Cedex France
4 Laboratoire Deacutepartemental de la Cocircte drsquoOr Dijon France
5 Welience SATT Grand-Est AgrOnov Breteniegravere France
6 Phytocontrol Paris MIN de Rungis Rungis Cedex France
Running head
Environmental detection of Mycobacterium bovis
Corresponding author
Alain Hartmann UMR 1347 Agroeacutecologie INRA 17 rue Sully BP 86510 21065 Dijon Cedex France E-
mail alainhartmanndijoninrafr
Chapitre 3
96
ABSTRACT
Aims
The aim of the study was to determine the prevalence of Mycobacterium bovis (the causative agent of
bovine tuberculosis bTB) in environmental matrices within a French region (Cocircte drsquoOr) affected by this
zoonotic disease
Methods and Results
We report here the development and the use of molecular detection assays based on qPCR (double
fluorescent dye labelled probe) to monitor the occurrence of Mycobacterium tuberculosis complex
(MTBC) or M bovis in environmental samples collected in pastures where infected cattle and wildlife
had been reported Three qPCR assays targeting members of the MTBC (IS1561rsquo and Rv3866 loci) or M
bovis (RD4 locus) were developed or refined from existing assays These tools were validated using
M bovis spiked soil water and feces samples Environmental samples were detected positive for the
presence of MTBC strains and M bovis in the environment of bTB-infected farms in the Cocircte drsquoOr
region
Conclusions
The development of molecular assays permitted testing of several types of environmental samples
including spring water sediment samples and soils from badger setts entrance located in the vicinity
of these farms which were repeatedly contaminated with M bovis (up to 87 times 103 gene copies per g
of badger sett soil) For the first time direct spoligotyping of soil DNA enabled identification of M bovis
genotypes from environmental matrices
Significance and Impact of the Study
All together these results suggest that M bovis occurs at low levels in environmental matrices in Cocircte
drsquoOr within the bTB-infected area Drinking contaminated water or inhaling contaminated bioaerosols
might explain cattle infection in some cases
Keywords
Mycobacterium bovis environment quantitative real-time PCR indirect transmission
Chapitre 3
97
INTRODUCTION
Bovine tuberculosis (bTB) is a worldwide spread zoonosis caused by Mycobacterium bovis a
pathogenic mycobacteria that belongs to the Mycobacterium tuberculosis complex (MTBC) This
disease affects cattle that are the natural host of M bovis but also a wide range of domestic or wild
mammals including humans (Morris et al 1994 Biet et al 2005) Infected wildlife can act as a bTB
reservoir maintaining and spreading M bovis infection in cattle Many studies including French ones
showed the circulation of the bacteria within multi-host systems including cattle and various wild
species as badgers (Meles meles) wild boar (Sus scrofa) or red deer (Cervus elaphus) (Little et al 1982
Aranaz et al 2004 Gortaacutezar et al 2008 Zanella et al 2008 Richomme et al 2010 Payne et al 2012)
The zoonotic nature of bTB may represent a public health threat Moreover animal disease leads to
important economic losses mainly due to cattle testing slaughtering campaigns and wildlife screening
trials
Infected animals can excrete M bovis via sputum feces urine and any type of aerosols (Little
et al 1982 Phillips et al 2003 Palmer et al 2004 Payne 2014) and thus contaminate their environment
(Phillips et al 2003) This may contribute to contamination of cattle pastures through deposition of
M bovis on cattle feedstuffs and into drinking water Indeed many studies confirmed the presence and
the persistence of M bovis in environmental substrates such as soil feces hay corn water (Young et
al 2005 Courtenay et al 2006 Sweeney et al 2006 Fine et al 2011 Ghodbane et al 2014) despite
many technical difficulties in isolating this organism from environmental substrates Altogether these
data strongly suggest that the environment may constitute a reservoir of M bovis from which cattle
might get infected However there is still very little direct evidence concerning these potential M bovis
environmental reservoirs and the way that animals might get infected through them
The European Commission declared that France was bTB-free in 2001 However the disease is
still a persistent problem in several regions including Cocircte drsquoOr (Middle East of France Figure 1)
Despite a very low national prevalence in cattle (0075 since 2012) complete eradication of the
disease seems difficult to achieve (Fediaevsky et al 2014) In Cocircte drsquoOr M bovis circulates in a multi-
host system including badgers wild boar and deer within a restricted area of the region (Payne 2014
Hauer et al 2015) As shown in Figure 1 two genotypes are found in this area M bovis spoligotype
SB0120 VNTR profile 5 5 4 3 11 4 5 6 in the northern bTB endemic area and M bovis spoligotype SB0134
VNTR profile 6 4 5 3 6 4 3 6 in the southern area Between 2002 and 2014 196 cattle breakdowns have
been registered in Cocircte drsquoOr (Cote drsquoOr Regional Veterinary Service personal communication) Wild
animals in the vicinity of infected cattle herds were found to be infected by M bovis with the same
genotypes (Hauer et al 2015)
Direct cross contamination between cattle has historically been considered as the main
Chapitre 3
98
transmission route of M bovis (Morris et al 1994 Phillips et al 2003) but environmental
contamination is suspected in many herd infections when cattle and wildlife share the same resources
(Garnett et al 2002 Palmer et al 2004) However this hypothesis has never been extensively explored
in France This is probably due to the lack of adequate detection tools to monitor M bovis under natural
environmental conditions Indeed most of the studies report the detection of M bovis from
experimentally inoculated environmental samples (Fine et al 2011 Adams et al 2013) Although the
culture method is the gold standard for direct diagnosis of M bovis in animals it is not well suited to
studying environmental samples due to the abundance of soil microbial communities and the slow
growth rate of M bovis To circumvent these drawbacks the use of highly selective media containing
antibiotics and antifungals in combination with sample decontamination is required in order to prevent
competition of fast growing organisms However such pretreatments strongly reduce M bovis viability
(Thornton et al 1998 Fine 2006) On the other hand molecular detection tools such as semi-
quantitative PCR have been used to detect M bovis in soils water and badger feces Following DNA
extraction detection of MTBC members or M bovis was done using primers targeting mpb64 mpb70
hsp65 16SrRNA genes or IS6110 insertion sequence (Young et al 2005 Courtenay et al 2006
Sweeney et al 2006 Adams et al 2013 Santos et al 2015) More recently a M bovis specific qPCR
(based on double fluorescent dye probe detection) was developed to target the RD4 deletion flanking
sequences (Sweeney et al 2007) This molecular system has been validated in several studies to test
environmental substrates such as soil badger feces and cow slurry (Pontiroli et al 2011 Travis et al
2011 King et al 2015a King et al 2015b) This RD4 based qPCR assay allowed to detect the occurrence
of M bovis within environmental matrices in UK (Sweeney et al 2007 King et al 2015a King et al
2015b) Finally Santos et al (Santos et al 2015) successfully used a hemi-nested PCR targeting the
RD12 locus which is common to M bovis and M caprae to monitor these species in environmental
substrates from bTB endemic regions located in the Iberian Peninsula
In this context the objectives of this study were firstly to develop new sensitive qPCR based
tools and to refine an existing one (based on RD4 deletion sequence) for the specific detection of MTBC
or M bovis in environmental matrices The sensitivity and specificity of qPCR based detection systems
were evaluated on a collection of mycobacterial strain DNAs but also on DNAs extracted from samples
(soil water and feces) spiked with known amounts of M bovis Secondly we aimed to screen large
numbers of environmental samples (soil sediments water feces) originating from both bTB endemic
or bTB free areas to assess the presence of MTBC and more specifically the presence of M bovis in
these areas
Chapitre 3
99
Figure 1 Distribution of bTB cumulative outbreaks in cattle in France (picture in the insert above) and per municipality in Cocircte drsquoOr from 2005 to 2013 Geographical location of the sampled areas are indicated by black stars and the location of the six positive areas are indicated by white stars and named A B C D E and F respectively
MATERIALS AND METHODS
Bacterial strains
A collection of bacterial strains belonging to the species M bovis M bovis BCG vaccine strain
and other mycobacterial species (Table 1) was used to test the specificity of the primers to detect
M bovis Crude thermo-lysates of strains corresponding to 20 bacterial spoligotypes of the M bovis
species 4 other species of the MTBC 2 sub-species of the Mycobacterium avium Complex (MAC) and
20 non tuberculous mycobacteria (NTM) Field strains belong to the National Reference Laboratory
collection and were previously identified by several genotyping methods using four qPCR assays done
in parallel and targeting IS6110 IS1081 IS1245 and atpE Namely MTBC members are positive for
Chapitre 3
100
IS6110 and IS1081 targets (Courcoul et al 2014) MAC members are positive for the IS1245 target
(Slana et al 2010) and finally NTM are positive for the atpE target (Radomski et al 2013) Furthermore
spoligotyping was used to discriminate the different MTBC species (Zhang et al 2010) IS901 was used
to discriminate between MAC members (Slana et al 2010) and hsp65 sequencing was used to
discriminate between other mycobacteria species (Telenti et al 1993) Attenuated M bovis BCG strain
Pasteur 1173P2 was used as target organism to validate the specificity and detection limits of the qPCR
assay in soil water and feces microcosms
Origin of environmental samples
Within the bTB endemic zone in Cocircte drsquoOr a total number of 356 environmental samples were
collected from 15 areas (an area corresponds to a municipality territory including at least one bTB
infected cattle farm) between April 2013 and December 2014 In those areas typical landscape is
observed ie limestone plateau with forest or farm fields overhang small hills The hillslopes are
covered with hydromorphic soils containing marl and claylimestone Alluvial valleys set at the bottom
of the slopes Cattle usually graze on those pastures during summer and spring Samples of soil
sediment and water were collected in areas frequented by both wildlife and cattle such as feeding
areas waterholes naturally occurring springs drinking troughs with open access to livestock and
wildlife Knowing that bTB infected badgers were discovered in the vicinity of the pastures grazed by
infected cattle (Cote drsquoOr Regional Veterinary Service personal communication) soils from badger
setts and latrines were also collected
Twenty-four agricultural soils were randomly chosen as control soils in French bTB-free areas
among a panel of soils from the French soil quality monitoring network (Reacuteseau de Mesures de la
Qualiteacute des Sols Infosol INRA Orleacuteans) Among them 11 french soils were collected from Brittany
(North West of France) Yvelines (Middle north of France) Haut Rhin (North east of France) while 13
soils were collected in the bTB-free area of Cocircte drsquoOr
In order to evaluate the detection limit of the qPCR assay we used two agricultural soils from two bTB-
free areas of Cocircte drsquoOr (Soil S1 and soil S2) one pond water and badger feces from animals tested as
bTB-free from non-infected areas Both soils were sampled in a bTB-free cattle pasture in November
2013 They were chosen for their contrasted structural and physico-chemical properties (Table 2)
DNA extraction and purification
DNAs from all soil sediment and latrine samples were extracted from 2 g-samples as following
Briefly 8 ml of lysis buffer (01 mol l-1 Tris pH 80 01 mol l-1 EDTA pH 80 01 mol l-1 NaCl and 2 (wv)
SDS) 4 g of silica beads (100 μm diameter) 5 g of ceramic beads (14 mm diameter) and 8 glass beads
(4 mm diameter) were added to each sample placed in a 15 ml conical centrifugation tube (Falcon
Chapitre 3
101
Dominique Dutscher France) Samples were disrupted for 3 times 30 s at 4 m s-1 in a FastPrep-24 Instrument
(MP Biomedicals Europe France) incubated for 30 min at 70degC and centrifuged for 5 min at 7000 g at
room temperature in a Sigma 3-18K centrifuge (Sigma-Aldrich Germany) Supernatants were
incubated on ice for 10 min with 110 volume of 3 mol l-1 potassium acetate pH 55 and then
centrifuged for 5 min at 14000 g One volume of ice-cold isopropanol was added to the supernatant
for DNA precipitation overnight at minus 20degC DNA was collected by centrifugation (30 min at 14000 g
4degC) and DNA pellets were washed with ice-cold ethanol (70 ) and dissolved in 100 μl of water Crude
DNA extracts (100 μl) were loaded on a Microbiospin column (Biorad France) filled with 100 mg of
polyvinyl polypyrrolidone (Sigma Chemical Co France) After centrifugation (4 min at 1000 g 4degC)
eluted DNA was further purified using the Geneclean Turbo Kit (Qiagen France) according to the
manufacturers instructions For water samples the microbial biomass was concentrated by filtering
from 100 ml to a maximum of 1 l of water sample (depending on water turbidity) through a 045 microm
Millipore HAWP filter (47 mm Merk Millipore Germany) Filters were then ground in liquid nitrogen
and DNA was extracted as described above Nematodes were extracted from soil using an Oostenbrink
elutriator (Oostenbrink 1960) DNA extraction was achieved as described above
Purified DNA concentrations were estimated using a NanoDropreg 2000 spectrophotometer
(Thermo Scientific France) at 260 nm wavelength
Choice of gene targets design of primers and probes for qPCR assays
A first qPCR detection system assay targeting the insertion sequence IS1561rsquo was designed
This insertion sequence is present in all the MTBC members except in M microti (Huard et al 2003
Huard et al 2006) The primers and double dye probe sequences are given in Table 3 IS1561rsquo contains
a defective transposase and belongs to the ISL3 family (Gordon et al 1999) In order to confirm the
specificity of the detection for MTBC a nested PCR protocol was additionally developed the first step
was a classical PCR using primers targeting a sequence of 222-bp sequence in the IS1561rsquo surrounding
the 76-bp qPCR product (Table 3) the second step was a qPCR assay targeting the 76-bp fragment To
confirm the results obtained with IS1561rsquo qPCR assay two supplementary qPCR targets were used The
first one was the Rv3866 locus located in the RD1 locus (Brodin et al 2002) The second one was the
RD4 flanking region (the RD4 locus is specifically deleted in M bovis) To improve the qPCR efficiency
the RD4 based assay was derived from a previously reported qPCR assay (Sweeney et al 2007) by
modifying the reverse primer to reduce the size of the qPCR product from 142 to 116 bp (Table 3) The
RD4 based qPCR assay designed by Sweeney et al (Sweeney et al 2007) has previously been validated
for environmental matrices in several studies (Pontiroli et al 2011 Travis et al 2011 King et al 2015a
King et al 2015b) Primers and probe were designed from alignments of mycobacterial genes from
GenBank (httpwwwncbinlmnihgovgenbank) such as M bovis AF212797 four strains of M bovis
Chapitre 3
102
BCG M tuberculosis H37Rv M africanum GM041182 and Mycobacterium canettii CIPT 140010059
(respective accession numbers NC_0029453 NC_0087691 NC_0168041 NC_0122071
GCA_0004632151 NC_0009623 NC_0157581 NC_0158481) using Primer3web
(httpprimer3utee) The specificity of the assay was tested in silico by BLAST analysis
(httpblastncbinlmnihgovBlastcgi) Primers and probes were synthesized by Eurogentec (Seraing
Belgium)
Quantitative real time PCR (qPCR) and nested PCR conditions
For the three qPCR assays reactions were carried out in a 25 microl reaction mix containing 125 microl
of Absolute QPCR ROX Mix (Thermo Scientific France) at a final concentration of 1X 1 microl of T4
bacteriophage gene 32 product (final concentration 25 ng microl-1 MP Biomedicals Europe France) 1 microl of
each primer (final concentration 400 nmol l-1) 05 microl of each probe (final concentration 200 nmol l-1)
4 microl of ultrapure water and 5 microl of diluted DNAs for the environmental samples screening Absolute
quantification was achieved using standard DNA dilutions A recombinant plasmid containing one copy
of the IS1561rsquo fragment of M bovis BCG str Pasteur 1173P2 cloned in the pCRregII-TOPOreg vector
(Invitrogen France) was used as a standard A calibration curve was obtained by amplification of serial
dilutions of the plasmid ranging from 1 to 106 copies per PCR reaction All the environmental
metagenomic DNA samples were screened with the IS1561rsquo based qPCR assay and final results were
expressed as the gene copy number per g of soil feces or water qPCR assays were performed in a VIIA
7 Real-Time PCR System (Thermo Fischer Scientific France) Initial denaturation was made at 95degC for
15 min followed by 45 cycles with denaturation at 95degC for 15 s and annealing and elongation at 60degC
for 1 min
Positive DNA extracts were then tested by the nested IS1561rsquo PCR protocol as follows the first PCR was
achieved in a 25 microl reaction mixture containing 1725 microl of ultrapure water for molecular biology 25
microl of PCR Buffer 10X with MgCl2 02 microl of dNTP mix at 25 mmol l-1 each 005 microl of Taq DNA polymerase
15 U microl-1 (Taq Core Kit 25 MP Biomedicals Europe France) 125 microl of each primers (final concentration
500 nmol l-1) and 25 microl of environmental DNA The nested PCR was achieved in a Veriti Thermal Cycler
(Applied Biosystems France) with a first step of denaturation at 94degC for 5 min then 30 cycles for 30
s at 94degC for denaturation 30 s at 60degC for annealing and 30 s at 72degC for elongation and 5 min at 72degC
for final elongation The qPCR was then achieved in the same conditions as described above using as a
target a 5 microl aliquot of a 100-fold dilution of the first PCR product
Preliminary screening of all environmental samples was done using the IS1561rsquo based qPCR
assay targeting MTBC species The IS1561rsquo positive samples were then further analysed using the
Rv3866 based qPCR to confirm the presence of MTBC and the RD4 based qPCR assay to confirm the
presence of M bovis All qPCR assays were done in triplicate
Chapitre 3
103
Determination of qPCR assays analytical sensitivity in spiked soil feces or water microcosms
Microcosms consisting of 2 g of fresh soil S1 and S2 sieved to 4 mm 2 g of fresh badger feces
or 100 mL of pond water were spiked in triplicate with 100 microl of 10-fold serially diluted suspensions of
M bovis BCG strain Pasteur to reach M bovis concentrations ranging from 25 to 25 times 106 CFU per
gram of soil or feces and 50 to 50 times 106 CFU per 100 mL of water Briefly inoculum suspensions were
realized as following a thirty days M bovis BCG liquid culture was vortexed for 15 s with glass beads (4
mm diameter) in order to disrupt aggregates Optical bacterial density was then measured at 600 nm
(Biophotometre Eppendorf France) to roughly estimate mycobacterial concentration Appropriate 10-
fold dilutions (10-1 to 10-6) were prepared in sterile water M bovis concentration was estimated by
spreading in triplicate 100 microL-aliquot of 10-4 10-5 and 10-6 diluted suspension onto Middlebrook 7H11
agar enriched with OADC 10 (Sigma-Aldrich France) Plates were incubated at 37degC for two months
and then M bovis colonies were counted Results were reported as CFU per ml of suspension
The spiked microcosms were incubated at room temperature for 3-4 hours prior to DNA
extraction as described in the section 3 To assess the reproducibility all qPCR assays were done in
triplicate and performed as previously using the three detection systems targeting IS1561rsquo Rv3866 and
RD4 Non-inoculated soil feces or water used to prepare the microcosms proved to be negative with
the three qPCR systems in preliminary experiments
Estimation of DNA quality for PCR amplification
Efficiency of qPCR might be impaired by the presence of PCR inhibitors in DNA extracts The
quality of DNA extracted from soil water and feces was assessed by measuring the capacity to amplify
a control DNA target in the presence of various dilutions of soil DNA extracts Briefly qPCR reactions
were done in a final volume of 25 microl containing 25 μl of 106 copies of the circularized pGEM-T Easy
plasmid DNA and 5 μl of water (control) or diluted soil DNA (5- 10- 20-fold dilutions) 1X ABsolute
qPCR SYBR Green ROX Mix (final concentration 500 nmol l-1 Thermo Scientific France) 25 ng microl-1 of T4
bacteriophage gene 32 product qPCR primers were SP6 (5rsquo - ATTTAGGTGACACTATAG - 3rsquo) and T7 (5rsquo -
TAATACGACTCACTATAGGG - 3rsquo) universal primers (final concentration 500 nmol l-1) targeting the
polylinker of the plasmid qPCR and cycling conditions were as following initial denaturation at 95degC
for 15 min followed by 40 cycles with denaturation at 95degC for 15 s annealing at 55degC and elongation
at 72degC for 30 s each The PCR product specificity was assessed by the melting curve analysis A 10-fold
reduction of the copy number amplified by qPCR in the presence of diluted soil DNA with respect to
the copy number obtained in the control reaction (water) was considered as 10 amplification
efficiency or conversely as 90 inhibition Although inhibition tests and detection of specific targets
were conducted with different mixes and primers the DNA polymerases from the ABsolute qPCR SYBR
Chapitre 3
104
Green ROX Mix (used for inhibition testing) and the ABsolute QPCR ROX Mix (used for mycobacterial
detection in environmental samples) are the same (Thermo-Start DNA polymerase) Thus the quality
of DNAs extracted from environmental samples was estimated by this method that is compatible with
acceptable experimental costs associated with the high number of analysed samples
Direct M bovis spoligotyping from environmental DNA
Environmental DNA samples that proved to be positive for the presence of MTBC were
submitted for spoligotyping using the Luminex method described previously by Zhang et al (Zhang et
al 2010) Spoligotypes were named according to an agreed international convention
(wwwmbovisorg)
RESULTS
Validation of the qPCR assays on pure bacterial strains
The results of the specificity test for the three qPCR assays are shown in Table 1 Using IS1561rsquo-
and Rv3866-based qPCR assays all spoligotypes of M bovis were detected with cycle threshold (Ct)
values below 25 cycles as well as all strains from the other species belonging to the MTBC (M caprae
M tuberculosis M pinnipedi) except the M microti species Using the RD4-based qPCR assay only
M bovis strains were detected with Ct values below 25 cycles For the three qPCR assays the 22 strains
of the NTM and MAC members were not detected or gave Ct values above 35 cycles Strains with Ct
values above 35 cycles when using high DNA concentration from pure culture are unlikely to give any
signal when testing soil metagenomic DNA where targets are highly diluted These results indicated
that IS1561rsquo- and Rv3866-based qPCR assays are highly specific for MTBC members and exclude
M microti whereas the RD4-based qPCR assay is specific for M bovis species
The calibration curve used for absolute quantification is presented in Figure 2 for the IS1561rsquo-
based qPCR assay A linear regression was established between the log (gene copy number) and the Ct
value over a wide range of target concentrations from 10 to 106 gene copies per PCR (r2= 09989) Below
10 target gene copies per reaction the detection was still possible but results were not reproducible
The quantification limit of the qPCR detection system was estimated to be 10 gene copies per PCR
reaction thus indicating a high sensitivity Similar calibration curves and detection thresholds were
obtained for the two other qPCR assays targeting Rv3866 and RD4 (data not shown)
Chapitre 3
105
Table 1 Strains used for testing oligonucleotides specificity Results are given with Ct (Cycle Threshold) ND (not detected) means that no signal was observed after 40 cycles
Strains Ct
IS1561rsquo
Ct
Rv3866 Ct RD4
Mycobacterium bovis BCG
BCG str Pasteur 1173P2
BCG str Danish 1331
lt25 lt25 lt25
Mycobacterium bovis
SB0089 SB0120 SB0121 SB0130
SB0134 SB0162 SB0166 SB0821
SB0823 SB0825 SB0826 SB0827
SB0878 SB0885 SB0912 SB0948
SB0999 SB1081
lt25 lt25 lt25
Others MTBC members
Mycobacterium microti ND ND ND
Mycobacterium tuberculosis lt25 lt25 ND
Mycobacterium caprae lt25 lt25 ND
Mycobacterium pinnipedi lt25 lt25 ND
Non tuberculous mycobacteria
Mycobacterium avium ND ND ND
Mycobacterium intracellulare ND ND ND
Mycobacterium aichiense ND ND ND
Mycobacterium arupense ND gt35 gt35
Mycobacterium celatum gt35 ND ND
Mycobacterium chelonae ND ND ND
Mycobacterium chitae ND ND ND
Mycobacterium confluentis gt35 ND gt35
Mycobacterium elephantis ND ND ND
Mycobacterium fortuitum ND ND ND
Mycobacterium intermedium ND ND ND
Mycobacterium kansai ND gt35 ND
Mycobacterium non chromogenicum ND ND ND
Mycobacterium parafinicum gt35 ND ND
Mycobacterium porcinum ND ND ND
Mycobacterium rufum ND ND ND
Mycobacterium rutilum ND ND ND
Mycobacterium terrae ND ND ND
Mycobacterium smegmatis ND gt35 ND
Mycobacterium szulgai ND gt35 ND
Mycobacterium thermoresitibile ND ND ND
Mycobacterium vaccae ND ND ND
Mycobacterium xenopi gt35 gt35 gt35
Chapitre 3
106
Detection threshold of qPCR assays in M bovis BCG spiked environmental matrices
The detection thresholds of the three qPCR assays were evaluated in complex environmental
matrices (soil feces and water) spiked with 25 to 25 times 106 CFU per g of soil or feces and 50 to 50 times
106 CFU per 100 ml of water The quality of DNAs extracted from microcosms was assessed as described
in the MampM section ldquoEstimation of DNA quality for PCR amplificationrdquo The results showed that 20-fold
dilutions of the DNA extracts did not inhibit the qPCR assay and were thus used for all qPCR assays
performed on spiked microcosms Results of M bovis detection in spiked microcosms are given in Table
4 for IS1561rsquo-based qPCR assay No signal was detected in control samples (not inoculated)
For M bovis cell densities above 250 CFU per g of soil 2500 CFU per g of feces and 5 CFU per
g of water (or 500 CFU per 100 ml of water) the detection by the qPCR assay yielded eight to nine
positive reactions out of nine repeats which allowed determination of the numbers of M bovis gene
copy number In contrast although M bovis was sometimes detected in spiked material with lower
densities than those described above quantification was unreliable A strong discrepancy between the
results obtained by the culture-based method and those obtained with the qPCR assay was observed
(10- to 100-fold higher for the qPCR assay) All of the microcosms were analysed with the two
complementary detection systems (targeting the Rv3866 locus and the flanking region of the RD4
deletion) and similar results were obtained (data not shown) Despite soils S1 and S2 being different
based on their physico-chemical characteristics detection limits were equivalent with the three qPCR
systems on these two soils
Extrapolating these results for natural environmental samples suggest that the three qPCR
assays allow a reliable quantification of MTBC (IS1561rsquo and Rv3866) or M bovis (RD4) targets in
environmental samples for Ct values below 39 Above this threshold detection may occur but lacks
reproducibility The word ldquotracesrdquo was then chosen in all further experiments in order to indicate that
MTBC or M bovis targets are present in very low quantities
Chapitre 3
107
Figure 2 Standard curve used for absolute quantification of M bovis in samples with IS1561rsquo qPCR assay (Mean values of ten standard curves) Error bars represent the 95 confidence interval associated to the values
Table 2 Properties of the two soils used for the qPCR sensitivity detection of Mycobacterium bovis
Table 3 Sets of primers tested in this study F forward primer R reverse primer P Double dye probe
Target locus Nucleotide sequence Location Size (bp)
IS1561rsquo F 5rsquo GATCCAGGCCGAGAGAATCTG 3rsquo
R 5rsquo GGACAAAAGCTTCGCCAAAA 3rsquo
P 5rsquo FAM ACGGCGTTGATCCGATTCCGC TAMRA 3rsquo
3742298-3742319
3742354-3782374
3742332-3742353
76
Nested IS1561rsquo F 5rsquo AGGAAGCGGTGTAGGTGATG 3rsquo
R 5rsquo AAGATCGACCCGCAGTG 3rsquo
3742212-3742232
3742416-3742434 222
RD4 F 5rsquo GCAGAAGCGCAACACTCTTG 3rsquo
R 5rsquo ATCTTAGCTGGTCAATAGCCATTTTT 3rsquo
P 5rsquo FAM AGTGGCCTACAACGGCGCTCTCC 3rsquo
1692712-1692736
1692802-1692827
1692743-1692768
116
Rv3866 F 5rsquo ACCCGCCAACGTCGAA 3rsquo
R 5rsquo CAATTGACGCAATACGGTATCG 3rsquo
P 5rsquo FAM CGAGCTAGCCGAATGCACAACCG 3rsquo
4278658-4278673
4278721-4278743
4278693-4278716
86
Location is given by reference to the M bovis AF212297 genome sequence accession ndeg NC_0029453
y = -33818x + 4394Rsup2 = 09989
15
20
25
30
35
40
45
1 2 3 4 5 6
Cyc
le T
hre
sho
ld C
t
DNA concentrations [log(IS1561 copies5microL-1)]
Site Clay Loam Sand Total
Carbon
Total Nitrogen
pH H2O
Organic matter
Land use
S1 397 485 118 361 038 775 0625 Pasture S2 192 463 345 292 0269 546 0506 Pasture
Chapitre 3
108
Molecular detection of M bovis in environmental samples
Our molecular detection systems were first evaluated on twenty-four soils from bTB-free zones
in France Whatever the detection system no signal was observed showing a good specificity of the
IS1561rsquo Rv3866 and RD4 qPCR systems
Between April 2013 and December 2014 63 samples of pasture soil 150 samples of setts soil 21
latrines samples 67 water samples from naturally occurring springs (and subsequent flowing water on
soil surface) within cattle pastures and 55 samples of spring sediments within 15 areas located in the
bTB-endemic zone in Cocircte drsquoOr were analysed Some spots were sampled several times (monthly from
January to June and in November) to assess the persistence of the MTBC signals Altogether 356 DNA
samples were primarily screened using the qPCR assay targeting IS1561rsquo locus Molecular signals
corresponding to MTBC were found in six areas out of 15 analysed and only positive results are
presented in Table 5 Apparent gene copy numbers were the average value from n = 2 to n = 3 repeats
These results indicate a MTBC contamination of the environment in 40 of the studied areas
Among the 36 positive samples 19 were found to be positive for MTBC detection with low
signals (indicated as ldquoTracesrdquo in Table 5) These samples include 7 badger sett soils 3 badger latrines
1 sediment sample and 8 water samples from naturally occurring springs in pastures More
interestingly high positive signals for MTBC were observed for the other 17 positive samples with 58
to 8 700 gene copies per g of soil It is worth noting that these samples only included badger sett soils
in area D and one nematodes sample in area C while no signal was recorded for the pasture soils
In order to confirm these positive results some of them were subjected to the nested IS1561rsquo
qPCR assay Eleven samples including eight badger sett soils two samples of water from springs and
one sample of nematodes isolated from spring sediments were confirmed to be positive for MTBC
detection with this nested qPCR assay Badger sett soils from area D were sampled five times over an
eight month-period The occurrence of MTBC signal was discontinuous in the seven soils from badger
setts that were sampled However MTBC signals were detected in several badger sett soils (badger sett
soils 1 4 and 5 in Table 5) following a seven to eight months period after M bovis was detected for the
first time The IS1561rsquo gene copy numbers for the positive badger sett soils ranged levels from
254 times 102 to 505 times 104 per g of soil The MTBC signal remained at high levels throughout the eight
month-period although the badgers moved out one month after the first sampling of the sett (Table
5) Furthermore confirmation of M bovis detection using the RD4 qPCR detection system was achieved
for a few IS1561rsquo positive samples Only three out of eight tested samples proved to be positive with
RD4-based detection system (see Table 5) This result definitely demonstrates the occurrence of M
bovis DNA in these samples
Spoligotyping was performed on positive DNA samples and the results are shown in Table 6 A
badger sett soil DNA was fully spoligotyped and found to belong to the SB0120 M bovis spoligotype
Chapitre 3
109
which is responsible for cattle and wildlife infection in this area of Cocircte drsquoOr Partial M bovis
spoligotypes were obtained for three badger sett soils DNAs The two other partial spoligotypes did
not allow identification of the species within the MTBC
Table 4 Evaluation of the detection of the IS1561rsquo qPCR assay with DNA extracted from control and spiked test matrices
Traces means that IS1561rsquo signal was detected but that quantification was unreliable (Ct value above 39)
Number of M bovis spiked per g of dry soil or per 100 mL of water
Number of positive PCR 9
reactions
Mean Ct Standard deviation Ct
Mean IS1561rsquo genome-equivalent per g of soil or feces or 100 mL of water
Soil S1
Control soil 0 ND ND ND
25 1 396 ND Traces 25101 1 394 ND Traces 25102 8 389 1 (654 plusmn 366) times103
25103 9 345 05 (217 plusmn 086) times105
25104 9 302 02 (218 plusmn 026) times106
25105 9 276 05 (132 plusmn 039) times107
Soil S2
Control soil 0 ND ND ND 25 0 ND ND ND 25101 5 404 06 Traces 25102 9 381 07 (800 plusmn 319) times103
25103 9 342 04 (141 plusmn 049) times105
25104 9 296 05 (374 plusmn 161) times106
25105 9 267 07 (265 plusmn 185) times107
Water Control water 0 ND ND ND 5101 1 401 ND Traces 5102 9 381 11 (220 plusmn 145) times104
5103 9 351 05 (148 plusmn 058) times105
5104 9 31 16 (320 plusmn 134) times106
5105 9 259 03 (819 plusmn 209) times107
5106 9 228 03 (698 plusmn 131) times108
Badger feces
Control feces 0 ND ND ND 25101 0 ND ND ND 25102 1 399 ND Traces 25103 9 379 105 (232 plusmn 077) times104
25104 9 347 159 (212 plusmn 187) times105
25105 9 312 203 (244 plusmn 286) times106
25106 9 261 059 (336 plusmn 107) times107
Chapitre 3
110
Table 5 Molecular detection of MTBC or M bovis targets in positive environmental samples from 6 areas in Cocircte drsquoOr Only positive samples are presented MTBC gene copy numbers were computed from calibration curve and amounts of DNA extracted from the various samples (n=2 ou n=3)
Area
Type of sample Sampling date Mean MTBC gene copy numbers g-1 of soil or feces (IS1561rsquo qPCR assay)
Confirmation after IS1561rsquo-nested PCR
Confirmation of presence of M bovis after RD4 qPCR assay
A Badger sett soil June 2013 Traces Negative NT dagger
B Spring water 1 January 2014 February 2014 March 2014
Traces
Traces
Traces
Negative Negative Negative
Negative Negative Negative
C Spring water 1 Spring water 2 Sediments Soil nematodes
February 2014 February 2014 February 2014 February 2014
Traces Traces Traces 582101
Positive Negative Negative Positive
NT
NT
NT
NT
D Badger sett soil 1 March 2014 November 2014
193103 577103
Positive NT
Positive NT
Badger sett soil 2 March 2014 May 2014 June 2014
870103 502102 143103
Positive Positive NT
Positive NT
NT Badger sett soil 3 March 2014
May 2014 June 2014
441102
741103 282103
Positive Positive NT
Negative NT
NT Badger sett soil 4 April 2014
May 2014 June 2014 November 2014
446103
236103 502102 607103
Positive NT
NT
NT
Negative NT
NT
NT Badger sett soil 5 April 2014
May 2014 June 2014 November 2014
248103
254102
202103
360103
Positive NT NT
NT
Positive NT
NT
NT Badger sett soil 6 November 2014 Traces NT NT Badger sett soil 7 November 2014 Traces NT NT Latrine 1 March 2014 Traces Negative NT Latrine 2 March 2014 Traces Negative NT
E Spring water 1 March 2014 March 2014 March 2014
Traces Negative NT Spring water 2 Traces Positive NT Spring water 3 Traces Negative NT Badger sett soil 1 March 2014
March 2014 Traces Negative NT
Badger sett soil 2 Traces Negative NT Latrine 1 March 2014 Traces Negative NT
F Badger sett soil 1 June 2014 June 2014
Traces Negative NT Badger sett soil 2 Traces Positive NT
Traces MTBC targets were present but MTBC genome equivalent were not calculated since Ct were close to the detection limit of the method dagger NT not tested
Chapitre 3
111
Table 6 Spoligotyping results of DNA positive samples extracted from environmental matrices Black squares represent positive hybridization signals and dots represent lack of hybridization
Figure 3 Spatial location of MTBC positive water and sediment samples in a pasture (site C) along the run off of one naturally occurring springs Star 1 indicates the sampling zone of positive spring sediment and nematodes sample Stars 2 and 3 indicate the 2 sampling sites of positive source water Contour lines are superimposed on aerial photography (450 m and 425 m contour lines are visible 5 m elevation interval between 2 consecutive lines)
Area Type of sample
Sampling date Hybridization pattern Spoligotype
D Badger sett soil 2
March 2014 M bovis SB0120
D Badger sett soil 1
March 2014 Partial M bovis spoligotype
D Badger sett soil 3
March 2014 Partial M bovis spoligotype
D Badger sett soil 5
November 2014
Partial M bovis spoligotype
C Water February 2014
Partial Mycobacterium spoligotype
Chapitre 3
112
DISCUSSION Mycobacterium bovis is described as an obligate intracellular microorganism transmitted by
direct contact between mammalian hosts However direct contacts between species seem to be scarce
and indirect transmission involving environmental matrices is likely to occur (Boumlhm et al 2009
Kukielka et al 2013) Many studies have clearly demonstrated the presence and the persistence of
M bovis in naturally infected environmental substrates in bTB endemic regions of United Kingdom
(Young et al 2005 Courtenay et al 2006 Sweeney et al 2006 Sweeney et al 2007 King et al 2015a
King et al 2015b) Evaluation of the environment as a reservoir of M bovis could help to understand
its epidemiological significance and its role in the indirect transmission of the disease among livestock
and wildlife To our knowledge the results from our study demonstrate for the first time the
contamination of environmental matrices with MTBC and more specifically M bovis in Cocircte drsquoOr which
is a region where bTB is endemic in cattle and wildlife
qPCR detection is a method of choice for M bovis environmental monitoring since it is a faster
alternative to culture based methods that are hampered by the slow-growing rate of M bovis and the
high number and diversity of microbial communities Moreover PCR is able to detect M bovis DNA in
the environment over a long period following initial contamination with a relatively high sensitivity
(Adams et al 2013)
The detection of M bovis in environmental samples (soil water and feces) that contain highly
diverse microbial communities including numerous described and potentially non-described NTM
(Falkinham 2009) was still a challenge The two qPCR assays developed in this study (IS1561rsquo and
Rv3866 targets) proved to be specific for the MTBC excluding M microti These results are in agreement
with the fact that IS1561rsquo is present in the genomes of all the members of the MTBC except in M microti
(Gordon et al 1999 Brodin et al 2002 Huard et al 2003) Itrsquos worth noting that M microti is quite
widespread in animals in France (Michelet et al 2015) and thus may also occur in environmental
matrices (Courtenay et al 2006) The refined RD4 qPCR assay was shown to be highly specific for
M bovis species in agreement with previous results (King et al 2015a) Although some strains gave Ct
gt 35 when concentrated in pure culture with these molecular assays these targets would not be
detected in natural samples due to extreme dilution in highly complex environmental DNAs
The DNA extraction method used in this study proved to be highly efficient in soils harboring
different physico-chemical characteristics (especially in clayey soils) (Dequiedt et al 2011 Locatelli et
al 2013) The large mass of samples (2 g) allowed a better representativeness of sampling to overcome
the potential heterogeneity of substrates ie a physical heterogeneity as well as a heterogeneous
distribution of mycobacteria in environmental samples The use of an amplification facilitator (T4
bacteriophage gene 32 product) allowed minimizing PCR inhibition by co-extracted contaminants
(Matheson et al 2010 Opel et al 2010) and significantly improving M bovis detection In previous
Chapitre 3
113
published studies (Travis et al 2011 Pontiroli et al 2011 Sweeney et al 2007) DNA extraction was
usually achieved from smaller masses (01 to 05 g) of soil or feces using commercial DNA extraction kit
in order to limit PCR inhibition
The three qPCR assays developed in this study using 20-fold dilutions of DNA extracts of spiked
samples allowed a reliable detection of spiked M bovis (BCG strain) at levels as low as 25 times 102 CFU to
25 times 103 CFU g-1 of sample providing an improved detection limit compared to a recent study where
detection limit was 40 times 104 CFU g-1 soil and 50 times 105 CFU ml-1 water (Santos et al 2015) In other
previous studies detection limits were estimated to 85 times 103 cells g1 for soil and 425 times 105 cells g1 for
fecal samples using the original RD4 qPCR assay (Pontiroli et al 2011 Travis et al 2011) As cell numbers
were estimated by fluorescent dye assisted flow cytometry comparison with our results is difficult
Finally King et al (King et al 2015a King et al 2015b) were able to detect 103 M bovis genome
equivalents per g-1 for badger feces
In our study the detection limits are equivalent for the three qPCR systems developed or refined
whatever the physico-chemical characteristics of the soils
The 10- to 100-fold discrepancy between bacterial numbers obtained using culture based methods and
qPCR based methods that we observed could be explained by the underestimation of cultivable
bacteria due to cell aggregation (linked to a strong hydrophobicity of mycobacteria cell wall) in
inoculum as well as the occurrence of viable but not cultivable cells (VBNC) and cell mortality (with
persistence of DNA) Similar discrepancies have already been observed in previous studies (eg Betelli
et al 2013)
Though similar detection thresholds were observed for the three systems in spiked samples
RD4 and RV3866 based qPCR assays seemed less sensitive than IS1561rsquo-based detection system in
environmental samples (range of 2 Ct between IS1561rsquo and the 2 other systems) However the
occurrence of RD4 based positive signals in several samples gave an additional proof of the specific
detection of M bovis Furthermore the direct spolygotyping of DNA extracted from environmental
matrices confirmed for the first time the presence of M bovis in several environmental samples in
France
All 63-pasture soil samples from the bTB endemic area in Cocircte drsquoOr were found to be negative
for the presence of MTBC (below the detection threshold) Failure to detect MTBC in pasture soil
samples can be related to the low prevalence of bTB in cattle in the sampling areas (herd prevalence
reached 08 in 2014 Cote drsquoOr Regional Veterinary Service) and the low prevalence of bTB infection
in badgers which territories overlap the sampled pastures (Payne et al 2012) This result might reflect
the high dilution rate of M bovis targets in soil DNA extracts which contains high numbers of other
microorganisms (abundant soil microbial communities containing up to 1010 genomes per g of soil)
The discontinuous (spatial and temporal) distribution of M bovis in soil pasture because of
Chapitre 3
114
intermittent andor low shedding (Neill et al 1988 Clifton-Hadley et al 1993) might also limit the
detection of M bovis in soil samples Despite the high number of soil sample analyzed financial and
time constraints have limited more intensive sampling efforts that could have overcome this limit
MTBC signal was detected in 101 (7 positive out of 67) of the water samples taken from
naturally occurring springs with the IS1561rsquo qPCR assay As the level of detection was close to the
detection threshold we were unable to quantify MTBC signals Interestingly one sample of nematodes
originated from sediments located in close vicinity of one of the positive sources was also found
positive for the presence of MTBC Although nematodes are known to host bacterial pathogen no data
is available concerning MTBC This intriguing result would require further investigations These water
springs (and accompanying flowing water) are located in pastures where both cattle and wildlife have
access for drinking In several pastures (four sites) water was sampled along the slope following the
gravitational water flow In two of these pastures we repeatedly found (two to three) positive samples
indicating multiple point contamination of spring and flowing water along these slopes (see Figure 3
for localization of positive samples in one of these pastures) This result is an important new finding of
our study that can support several hypotheses on the role of water in the dissemination of MTBC in
the environment and in animal contamination We can hypothesize from these results that bioaerosols
formation from contaminated water and subsequent inhalation by animals (cattle and wildlife) might
contribute to animal infection with MTBC members especially as the infective dose through this
transmission route is very low (lt 10 CFU) (Dean et al 2005 Corner et al 2007) Infection of animals
might also occur through oral route by repeatedly drinking contaminated water
A special effort was made to monitor the contamination of badger sett soils since bTB infected
badgers were detected in our study area Between 2009 and 2011 878 badgers had been trapped and
analyzed in the infected area of Cocircte drsquoOr (Payne et al 2012) Macroscopic lesions were present in 19
of them despite culture-based method detecting 56 of positive badgers Only one badger showed
widespread tuberculosis Moreover in this infected area numerous badger setts were located either
within pastures or in close vicinity (forest-pasture interface) Our study demonstrated an average
prevalence of MTBC of 73 (11 out of 150) in soils from badger setts with the highest gene copy
number up to 37 times 104 gene copies g-1 of soil observed among environmental samples analysed in our
study The prevalence of MTBC molecular signature in latrines was estimated to 7 (3 positive out of
21) Our results demonstrate that infected badgers might contaminate their close environment for the
first time in France Indeed infected badgers have been shown to excrete M bovis from the respiratory
digestive and urinary tracts and in bite wounds exudates (Gavier-Widen et al 2001 Corner et al 2012)
Our results are in agreement with previous findings from several studies in UK (Young et al 2005
Courtenay et al 2006 Sweeney et al 2007 King et al 2015a) that recovered M bovis from badger
setts latrines and adjacent fields with gene copy numbers ranging from 1 times 103 to 54 times 106 per g of
Chapitre 3
115
soil In Woodchester Park in UK where badgers populations have been extensively studied for more
than 20 years bTB prevalence was estimated around 30 in 2005 which might explain high numbers
of positive setts and latrines observed in this area (Courtenay et al 2006 King et al 2015a) As
suggested by Courtenay et al (Courtenay et al 2006) the low bTB prevalence of infected sett soils
observed in our study may be linked to the low prevalence and the low density of badgers excreting
the bacteria within their social group
Finally in soils of 7 badger setts we were able to detect M bovis signatures (RD4 positive qPCR)
either continuously (4 setts) or discontinuously (3 setts) The presence of M bovis was confirmed by
direct soil DNA spolygotyping (see Table 6) in soils of the setts The presence of M bovis molecular
signatures was proved over an eight month-period (despite the badgers moved out) These results are
in agreement with a previous study where positive molecular detection of M bovis was demonstrated
over a period of 15 months despite the badgers having moved out (Young et al 2005) To our
knowledge this is the first time that spoligotype SB0120 was observed by direct spoligotyping of soil
DNA this spoligotype being identical to the spoligotype of strains isolated from cattle and wildlife in
these sites (Payne et al 2012)
Although the molecular detection of MTBC or M bovis by specific qPCR assay doesnrsquot give any
information about mycobacterial viability Young et al (Young et al 2005) demonstrated that MTBC DNA
did not persist in soil for more than ten days suggesting that DNA detected in our environmental
samples could belong to viable cells
A remaining challenge is to develop adequate methods to cultivate M bovis from
environmental matrices Indeed culture based methods might also be impaired by the occurrence of
VBNC state or dormancy in M bovis present in environmental matrices as previously suggested by
Whittington et al (Whittington et al 2004) for Mycobacterium avium subsp paratuberculosis Further
work is needed to assess if VBNC or dormancy occur in M bovis under environmental conditions
Although we concentrated our sampling efforts on badger environment it is however
acknowledged that in Cocircte drsquoOr M bovis circulates in a multi-host system that includes also wild boar
and deer We can thus hypothesize that these wild animals can also contribute to the shedding of
M bovis in the environment
Further research is needed to understand the ecology of M bovis in the environment and its
epidemiological role in indirect transmission This knowledge would help to improve herd
management in reducing indirect contacts between livestock and wildlife in controlling water access
in areas considered as ldquoat riskrdquo and in preventing contacts with badger setts for example Biosecurity
should thus be an integral part of the long-term strategy for the management of bovine tuberculosis in
Cocircte drsquoOr
Chapitre 3
116
Acknowledgements
The authors thank F Chevalier and M Beacuteral from the CIREV (Cellule Inter-Reacutegionale
drsquoEpideacutemiologie Veacuteteacuterinaire) AE Taconet F Bartheacuteleacutemy and T Grosjean from the Cote drsquoOr Regional
Veterinary Service (DDPP21) We are grateful to the cattle breeders trappers and hunters who helped
us during this study We wish also to thank C Ferrier from ONCFS (National Game Hunting and Wildlife
Agency) Dominique Meunier and Luc Biju-Duval from INRA Dijon Ceacuteline Richomme from ANSES The
work was supported by the Conseil Reacutegional de Bourgogne and Conseil Geacuteneacuteral de Cocircte drsquoOr Interbev
GDS Cocircte drsquoOr
Conflict of interest
No conflict of interest declared
References
Adams AP Bolin SR Fine AE Bolin CA and Kaneene JB (2013) Comparison of PCR versus culture for detection of Mycobacterium bovis after experimental inoculation of various matrices held under environmental conditions for extended periods Appl Environ Microbiol 79 6501-6506
Aranaz A De Juan L Montero N Saacutenchez C Galka M Delso C Alvarez J Romero B Bezos J Vela AI Briones V Mateos A and Domiacutenguez L (2004) Bovine tuberculosis (Mycobacterium bovis) in wildlife in Spain J Clin Microbiol 42 2602-2608
Betelli L Duquenne P Grenouillet F Simon X Scherer E Gehin E and Hartmann A (2013) Development and evaluation of a method for the quantification of airborne Thermoactinomyces vulgaris by real-time PCR J Microbiol Methods 92 25-32
Biet F Boschiroli ML Thorel MF and Guilloteau LA (2005) Zoonotic aspects of Mycobacterium bovis and Mycobacterium avium-intracellulare complex (MAC) Vet Res 36 411-436
Boumlhm M Hutchings MR and White PCL (2009) Contact networks in a wildlife-livestock host community identifying high-risk individuals in the transmission of bovine TB among badgers and cattle PLoS One 4 e5016
Brodin P Eiglmeier K Marmiesse M Billault A Garnier T Niemann S Cole ST and Brosch R (2002) Bacterial artificial chromosome-based comparative genomic analysis identifies Mycobacterium microti as a natural ESAT-6 deletion mutant Infect Immun 70 5568-5578
Clifton-Hadley RS Wilesmith JW and Stuart Fa (1993) Mycobacterium bovis in the European badger (Meles meles) epidemiological findings in tuberculous badgers from a naturally infected population Epidemiol Infect 111 9-19
Corner LA OMeara D Costello E Lesellier S and Gormley E (2012) The distribution of Mycobacterium bovis infection in naturally infected badgers Vet J 194 166-172
Corner LA Costello E Lesellier S OMeara D Sleeman DP and Gormley E (2007) Experimental tuberculosis in the European badger (Meles meles) after endobronchial
Chapitre 3
117
inoculation of Mycobacterium bovis I Pathology and bacteriology Res Vet Sci 83 53-62
Courcoul A Moyen J-L Brugegravere L Faye S Heacutenault S Gares H and Boschiroli M-L (2014) Estimation of sensitivity and specificity of bacteriology histopathology and PCR for the confirmatory diagnosis of bovine tuberculosis using latent class analysis PLoS One 9 e90334
Courtenay O Reilly LA Sweeney FP Hibberd V Bryan S Ul-Hassan A Newman C Macdonald DW Delahay RJ Wilson GJ and Wellington EMH (2006) Is Mycobacterium bovis in the environment important for the persistence of bovine tuberculosis Biol Lett 71 1946-1952
Dean GS Rhodes SG Coad M Adam O Cockle PJ Clifford DJ Hewinson RG and Vordermeier HM (2005) Minimum infective dose of Mycobacterium bovis in cattle Infect Immun 73 6467-6471
Dequiedt S Saby NPa Lelievre M Jolivet C Thioulouse J Toutain B Arrouays D Bispo a Lemanceau P and Ranjard L (2011) Biogeographical patterns of soil molecular microbial biomass as influenced by soil characteristics and management Global Ecol Biogeogr 20 641-652
Falkinham JO (2009) Surrounded by mycobacteria nontuberculous mycobacteria in the human environment J Appl Microbiol 107 356-367
Fediaevsky A Courcoul A Boschiroli M-L and Reveillaud E (2014) Tuberculose bovine en France reacutesultats dune strateacutegie plus offensive Bull Epid Santeacute Anim Alim 64 4-11
Fine AE (2006) The role of indirect transmission in the epidemiology of bovine tuberculosis in cattle and white-tailed deer in Michigan Michigan Bovine Tuberculosis Bibliography and DatabasePaper httpdigitalcommonsunledumichbovinetb33
Fine AE Bolin Ca Gardiner JC and Kaneene JB (2011) A study of the persistence of Mycobacterium bovis in the environment under natural weather conditions in Michigan USA Vet Med Int 2011
Garnett BT Delahay RJ and Roper TJ (2002) Use of cattle farm resources by badgers (Meles meles) and risk of bovine tuberculosis (Mycobacterium bovis) transmission to cattle Proc R Soc Lond B Biol Sci 269 1487-1491
Gavier-Widen D Chambers MA Palmer N Newell DG and Hewinson RG (2001) Pathology of natural Mycobacterium bovis infection in European badgers (Meles meles) and its relationship with bacterial excretion Vet Rec 148 299-304
Ghodbane R Mba Medie F Lepidi H Nappez C and Drancourt M (2014) Long-term survival of tuberculosis complex mycobacteria in soil Microbiology 160 496-501
Gordon SV Heym B Parkhill J Barrell B and Cole ST (1999) New insertion sequences and a novel repeated sequence in the genome of Mycobacterium tuberculosis H37Rv Microbiology 145 881-892
Gortaacutezar C Torres MJ Vicente J Acevedo P Reglero M de la Fuente J Negro JJ and Aznar-Martiacuten J (2008) Bovine tuberculosis in Dontildeana Biosphere Reserve The role of wild ungulates as disease reservoirs in the last Iberian lynx strongholds PLoS One 3 e2776
Chapitre 3
118
Hauer A Cruz KD Cochard T Godreuil S Karaoui C Henault S Bulach T Banuls AL Biet F and Boschiroli M-L (2015) Genetic evolution of Mycobacterium bovis causing tuberculosis in livestock and wildlife in France since 1978 PLoS One 10 e0117103
Huard RC de Oliveira Lazzarini LC Butler WR van Soolingen D and Ho JL (2003) PCR-based method to differentiate the subspecies of the Mycobacterium tuberculosis complex on the basis of genomic deletions J Clin Microbiol 41 1637-1650
Huard RC Fabre M de Haas P Lazzarini LCO van Soolingen D Cousins D and Ho JL (2006) Novel genetic polymorphisms that further delineate the phylogeny of the Mycobacterium tuberculosis complex J Bacteriol 188 4271-4287
King HC Murphy A James P Travis E Porter D Hung Y-J Sawyer J Cork J Delahay RJ Gaze W Courtenay O and Wellington EM (2015a) The variability and seasonality of the environmental reservoir of Mycobacterium bovis shed by wild European badgers Sci Rep 5 1-7
King HC Murphy A James P Travis E Porter D Sawyer J Cork J Delahay RJ Gaze W Courtenay O and Wellington EM (2015b) Performance of a noninvasive test for detecting Mycobacterium bovis shedding in European badger (Meles meles) populations J Clin Microbiol 53 2316-2323
Kukielka E Barasona JA Cowie CE Drewe JA Gortazar C Cotarelo I and Vicente J (2013) Spatial and temporal interactions between livestock and wildlife in South Central Spain assessed by camera traps Prev Vet Med 112 213-221
Little TW Naylor PF and Wilesmith JW (1982) Laboratory study of Mycobacterium bovis infection in badgers and calves Vet Rec 111 550-557
Locatelli A Depret G Jolivet C Henry S Dequiedt S Piveteau P and Hartmann A (2013) Nation-wide study of the occurrence of Listeria monocytogenes in French soils using culture-based and molecular detection methods J Microbiol Methods 93 242-250
Matheson CD Gurney C Esau N and Lehto R (2010) Assessing PCR inhibition from humic substances Open Enzym Inhib J 3 38-45
Michelet L de Cruz K Zanella G Aaziz R Bulach T Karoui C Heacutenault S Joncour G and Boschiroli ML (2015) Infection with Mycobacterium microti in animals in France J Clin Microbiol 53 981-985
Morris RS Pfeiffer DU and Jackson R (1994) The epidemiology of Mycobacterium bovis infections Vet Microbiol 40 153-177
Neill S Hanna J OBrien J and McCracken R (1988) Excretion of Mycobacterium bovis by experimentally infected cattle Vet Rec 123 340-343
Oostenbrink M (1960) Estimating nematode populations by some selected methods Nematology 6(8) 165-2015
Opel KL Chung D and McCord BR (2010) A study of PCR inhibition mechanisms using real time PCR J Forensic Sci 55 25-33
Palmer MV Waters WR and Whipple DL (2004) Investigation of the transmission of
Chapitre 3
119
Mycobacterium bovis from deer to cattle through indirect contact Am J Vet Res 65 1483-1489
Payne A (2014) Rocircle de la faune sauvage dans le systegraveme multi-hocirctes de Mycobacterium bovis et risque de transmission entre faune sauvage et bovins eacutetude expeacuterimentale en Cocircte drsquoOr In Epidemiologie Universiteacute Claude Bernard Lyon 1 httpwwwthesesfr2014LYO10040
Payne A Boschiroli ML Gueneau E Moyen JL Rambaud T Dufour B Gilot-Fromont E and Hars J (2012) Bovine tuberculosis in ldquoEurasianrdquo badgers (Meles meles) in France Eur J Wildlife Res 59 331-339
Phillips CJC Foster CRW Morris PA and Teverson R (2003) The transmission of Mycobacterium bovis infection to cattle Res Vet Sci 74 1-15
Pontiroli A Travis ER Sweeney FP Porter D Gaze WH Mason S Hibberd V Holden J Courtenay O and Wellington EMH (2011) Pathogen quantitation in complex matrices a multi-operator comparison of DNA extraction methods with a novel assessment of PCR inhibition PLoS One 6 e17916
Radomski N Roguet A Lucas FS Veyrier FJ Cambau E Accrombessi H Moilleron R Behr MA and Moulin L (2013) atpE gene as a new useful specific molecular target to quantify Mycobacterium in environmental samples BMC Microbiol 13 277
Richomme C Boschiroli ML Hars J Casabianca F and Ducrot C (2010) Bovine tuberculosis in livestock and wild boar on the Mediterranean island Corsica J Wildl Dis 46 627-631
Santos N Santos C Valente T Gortazar C Almeida V and Correia-Neves M (2015) Widespread environmental contamination with Mycobacterium tuberculosis Complex revealed by a molecular detection protocol PLoS One 10 e0142079
Slana I Kaevska M Kralik P Horvathova A and Pavlik I (2010) Distribution of Mycobacterium avium subsp avium and M a hominissuis in artificially infected pigs studied by culture and IS901 and IS1245 quantitative real time PCR Vet Microbiol 144 437-443
Sweeney FP Courtenay O Hibberd V Hewinson RG Reilly LA Gaze WH and Wellington EMH (2007) Environmental monitoring of Mycobacterium bovis in badger feces and badger sett soil by Real-Time PCR as confirmed by immunofluorescence immunocapture and cultivation Appl Environ Microbiol 73 7471-7473
Sweeney FP Courtenay O Ul-Hassan A Hibberd V Reilly LA and Wellington EMH (2006) Immunomagnetic recovery of Mycobacterium bovis from naturally infected environmental samples Lett Appl Microbiol 43 364-369
Telenti A Marchesi F Balz M Bally F Bottger EC and Bodmer T (1993) Rapid identification of mycobacteria to the species level by polymerase chain reaction and restriction enzyme analysis J Clin Microbiol 31 175-178
Thornton CG MacLellan KM Brink TL Lockwood DE Romagnoli M Turner J Merz WG Schwalbe RS Moody M Lue Y and Passen S (1998) Novel method for processing respiratory specimens for detection of mycobacteria by using C-18-carboxypropylbetaine Blinded study J Clin Microbiol 36 1996-2003
Travis ER Gaze WH Pontiroli A Sweeney FP Porter D Mason S Keeling MJC Jones RM
Chapitre 3
120
Sawyer J Aranaz A Rizaldos EC Cork J Delahay RJ Wilson GJ Hewinson RG Courtenay O and Wellington EMH (2011) An inter-laboratory validation of a real time PCR assay to measure host excretion of bacterial pathogens particularly of Mycobacterium bovis PLoS One 6 e27369
Whittington R Marshall D Nicholls PJ Marsh IB and Reddacliff LA (2004) Survival and dormancy of Mycobacterium avium subsp paratuberculosis in the environment Appl Environ Microbiol 70 2989-3004
Young JS Gormley E and Wellington EM (2005) Molecular detection of Mycobacterium bovis and Mycobacterium bovis BCG (Pasteur) in soil Appl Environ Microbiol 71 1946-1952
Zanella G Durand B Hars J Moutou F Garin-Bastuji B Duvauchelle A Ferme M Karoul C and Boschiroli ML (2008) Mycobacterium bovis in wildlife in France J Wildl Dis 44 99-108
Zhang J Abadia E Refregier G Tafaj S Boschiroli ML Guillard B Andremont A Ruimy R and Sola C (2010) Mycobacterium tuberculosis complex CRISPR genotyping improving efficiency throughput and discriminative power of spoligotyping with new spacers and a microbead-based hybridization assay J Med Microbiol 59 285-294
Chapitre 3
121
C CAMPAGNES DE PRELEVEMENTS SUPPLEMENTAIRES
Entre deacutecembre 2014 et mars 2015 deux campagnes de preacutelegravevement drsquoeacutechantillons
environnementaux suppleacutementaires ont eacuteteacute reacutealiseacutees dans la zone endeacutemique de Cocircte drsquoOr ainsi
qursquoune campagne en Charente ougrave des foyers bovins et sauvages sont recenseacutes respectivement depuis
2006 et 2009
1 Fin des preacutelegravevements en Cocircte drsquoOr
Les preacutelegravevements ont concerneacute drsquoune part une forecirct domaniale agrave cheval sur deux communes
de la zone Sud (eacutetoiles blanches sur la Figure 19) dans laquelle nous nous sommes focaliseacutes sur les
trous drsquoeau freacutequenteacutes par les sangliers et les cervideacutes Cinq trous drsquoeau importants et freacutequenteacutes ont
eacuteteacute cibleacutes et sur chacun trois preacutelegravevements de solsseacutediments et deux preacutelegravevements drsquoeau ont eacuteteacute
reacutealiseacutes en deacutecembre 2014 Drsquoautre part des preacutelegravevements ont eacuteteacute effectueacutes dans les terriers de
blaireaux infecteacutes de la commune D citeacutee dans lrsquoarticle preacuteceacutedent que nous suivons depuis mars 2014
et dans laquelle 1 agrave 3 blaireaux infecteacutes sont pieacutegeacutes tous les ans ainsi que dans des terriers freacutequenteacutes
drsquoune commune limitrophe (commune G) sur laquelle un blaireau infecteacute a eacuteteacute pieacutegeacute en 2014 Quatorze
sols de terriers (5 sur la commune D et 9 sur la commune G) et cinq latrines proches des terriers (3 sur
D et 2 sur G) ont eacuteteacute preacuteleveacutes en mars 2015 Ces deux sites sont repreacutesenteacutes par les eacutetoiles noires sur
la Figure 19 Tous ces eacutechantillons ont eacuteteacute analyseacutes selon les techniques deacutecrites dans lrsquoarticle
preacuteceacutedent (section Mateacuteriel et meacutethodes)
Les vingt-cinq eacutechantillons preacuteleveacutes dans la forecirct domaniale de la zone Sud en deacutecembre 2014
nrsquoont donneacute aucun signal moleacuteculaire avec le systegraveme de deacutetection ciblant lrsquoIS1561rsquo Dans cette zone
forestiegravere aucun animal sauvage infecteacute nrsquoa eacuteteacute deacutetecteacute depuis 2013
Concernant les terriers et latrines de la commune D les cinq eacutechantillons de sols de terriers
analyseacutes et une des deux latrines ont donneacute un signal moleacuteculaire positif pour le MTBC Pour lrsquoun des
sols la quantification nrsquoa pas eacuteteacute possible avec la qPCR (Ct gt 39 traces) Pour les quatre autres sols
des valeurs de 10 times 104 agrave 46 times 104 copies de gegravenes ont eacuteteacute obtenues et de 12 times 104 copies de gegravenes
pour la latrine Trois des eacutechantillons de sol ont donneacute un reacutesultat positif avec la PCR nicheacutee ciblant
lrsquoIS1561rsquo Dans ces terriers les preacutelegravevements reacutealiseacutes en mars avril mai juin et novembre 2014 puis
mars 2015 ont montreacute une persistance du signal moleacuteculaire au cours du temps Pour quatre des
terriers le signal a eacuteteacute deacutetecteacute agrave chaque campagne de preacutelegravevements alors que pour trois autres la
deacutetection a eacuteteacute intermittente Il est inteacuteressant de noter que les valeurs obtenues en mars 2015 sont
supeacuterieures agrave celles obtenues en 2014 (voir article preacuteceacutedent) Plusieurs hypothegraveses sont eacutemises pour
Chapitre 3
122
expliquer ce reacutesultat La premiegravere est lieacutee au fait que les terriers eacutetaient habiteacutes lors de nos
preacutelegravevements en mars 2015 alors qursquoils ne lrsquoeacutetaient plus apregraves les preacutelegravevements de mars 2014
La surveillance videacuteo reacutealiseacutee sur ces terriers a montreacute que les terriers ont agrave nouveau eacuteteacute
occupeacutes agrave partir de septembre 2014 Il est possible que cette occupation ait geacuteneacutereacute lrsquoexteacuteriorisation de
sol profond et de litiegravere potentiellement plus contamineacutes et que nous ayons preacuteleveacute ce sol drsquoorigine
profonde La seconde hypothegravese est lieacutee au fait que nous avons preacuteleveacute plus profondeacutement dans le
terrier que dans les campagnes de preacutelegravevements preacuteceacutedentes (utilisation drsquoun aspirateur) Enfin il est
eacutegalement possible que le groupe de blaireaux peuplant ce terrier possegravede un ou plusieurs animaux
infecteacutes et excreacuteteurs agrave lrsquoorigine drsquoune contamination plus importante du sol du terrier Ces terriers sont
situeacutes dans le bosquet drsquoun preacute qui fournit aux bovins de lrsquoombre et de la fraicheur en eacuteteacute et un abri en
cas de pluie ou de vent La surveillance videacuteo de cette zone nous a permis drsquoobserver que plusieurs
veaux ont reacuteguliegraverement exploreacute les entreacutees de gueules et consommeacute les brins drsquoherbe autour de ces
entreacutees (surveillance videacuteo)
Sur les neuf eacutechantillons de sol preacuteleveacutes dans la commune G deux ont donneacute un signal positif
pour le MTBC Pour lrsquoun drsquoeux seules des traces de M bovis ont eacuteteacute deacutetecteacutees pour lrsquoautre le nombre
de copies de gegravenes a eacuteteacute estimeacute agrave 12 times 104 La PCR nicheacutee srsquoest reacuteveacuteleacutee neacutegative pour les deux
eacutechantillons
Chapitre 3
123
Figure 19 Localisation des communes de Cocircte drsquoOr sur lesquelles de foyers bovins et des animaux sauvages infecteacutes ont eacuteteacute deacutetecteacutes et localisation des preacutelegravevements environnementaux effectueacutes en 2015 (les preacutelegravevements positifs sont repreacutesenteacutes par des eacutetoiles noires) (Reacutealisation Marina Beacuteral CIREV Bourgogne)
2 Preacutelegravevements en Charente
Depuis 2006 25 foyers bovins ont eacuteteacute deacutetecteacutes en Charente avec 12 foyers pour la seule anneacutee
2014 et la deacutecouverte de foyers anciens qui eacutevoluaient agrave bas bruit (communication personnelle
DDPP16) Depuis 2009 lrsquoinfection est mise en eacutevidence chez les blaireaux (6 positifs en 2013 et 11 en
2014) et les sangliers (8 positifs en 2014 donneacutees Sylvatub) et un chevreuil en 2015 (Communication
personnelle Sylvatub) autour des foyers bovins avec les mecircmes spoligotypes Le systegraveme multi-hocirctes
observeacute diffegravere donc de celui de la Cocircte drsquoOr puisqursquoagrave lrsquoheure actuelle lrsquoinfection nrsquoa pas eacuteteacute observeacutee
chez les cerfs dont les densiteacutes sont beaucoup moins importantes qursquoen Cocircte drsquoOr Les foyers bovins
Chapitre 3
124
et sauvages sont majoritairement localiseacutes dans le sud du deacutepartement dans une zone limitrophe de
la Dordogne deacutepartement fortement infecteacute depuis 2005 (Figure 20)
A la demande de la DDPP16 nous sommes alleacutes preacutelever des eacutechantillons environnementaux
dans le Sud du deacutepartement dans sept communes fortement toucheacutees par la maladie (Figure 20) Entre
avril et mai 2015 55 eacutechantillons environnementaux (50 eacutechantillons de sol de terrier de blaireau 3
sols de souille 1 latrine de blaireau et 1 fegraveces de sanglier) ont eacuteteacute preacuteleveacutes sur 7 communes de
Charente Ces communes font lrsquoobjet de la deacuteclaration drsquoun ou plusieurs foyers bovins de tuberculose
bovine etou drsquoanimaux sauvages infecteacutes (Figure 20) De plus 3 eacutechantillons de matiegraveres feacutecales
preacuteleveacutes pendant lrsquoautopsie de 3 blaireaux (BLAIR 7057 BLAIR 7058 et BLAIR 7061) ont eacuteteacute analyseacutes
Six eacutechantillons positifs ont eacuteteacute mis en eacutevidence sur les 55 analyseacutes La qPCR ciblant lrsquoIS1561rsquo a permis
de deacutetecter et de quantifier M bovis dans quatre ADN de sol de terrier avec un nombre de copies de
gegravenes variant de 11 times 103 agrave 54 times 103 par g de sol Deux eacutechantillons suppleacutementaires (1 sol de souille
et 1 sol de terrier) ont donneacute un signal non quantifiable avec la qPCR (traces) Tous les eacutechantillons
positifs ont eacuteteacute preacuteleveacutes sur une mecircme commune sur 2 lieux-dits diffeacuterents Maine-Joli (4 sols de
terrier) et la Tuilerie (1 sol de terrier et 1 sol de souille) A notre connaissance 2 blaireaux et 3 sangliers
infecteacutes ont eacuteteacute deacutecouverts agrave proximiteacute du lieu-dit la Tuilerie deacutebut 2015 (Source DDPP16) Lors de
notre campagne de preacutelegravevement tous les terriers deacutetecteacutes positifs eacutetaient freacutequenteacutes
Les ADN des eacutechantillons positifs ont eacuteteacute envoyeacutes agrave lrsquoANSES pour geacutenotypage Aucun ADN nrsquoa toutefois
pu ecirctre spoligotypeacute
Les fegraveces de blaireaux (BLAIR 7057 BLAIR 7058 et BLAIR 7061) preacuteleveacutes lors des autopsies nrsquoont donneacute
aucun signal avec la qPCR
Chapitre 3
125
Figure 20 Localisation des communes de Charente sur lesquelles de foyers bovins et des animaux sauvages infecteacutes ont eacuteteacute deacutetecteacutes et localisation des preacutelegravevements environnementaux effectueacutes en 2015 (les preacutelegravevements positifs sont repreacutesenteacutes par des eacutetoiles noires) (Reacutealisation Marina Beacuteral CIREV Bourgogne)
3 Compleacutements paysage geacuteomorphologie et peacutedologie
La zone endeacutemique Nord de tuberculose bovine en Cocircte drsquoOr que nous avons principalement
eacutetudieacutee est situeacutee sur un territoire appeleacute lrsquoAuxois caracteacuteriseacute par un paysage bocager ponctueacute de
buttes et de plateaux calcaires ougrave affleure un calcaire dur et karstique Les pacirctures suspectes
preacutesentent un contexte paysager hydrologique et geacuteomorphologique commun Le parcellaire suspect
est localiseacute dans trois petites valleacutees parallegraveles au fond desquelles circule un cours drsquoeau en direction
du Nord-Ouest (Figure 21) Chaque cours drsquoeau est alimenteacute par des sources naissant entre les couches
calcaires et marneuses du sous-sol dans les pentes Ces reacutesurgences drsquoeau proviennent de lrsquoinfiltration
des preacutecipitations dans les plateaux calcaires agrave lrsquoorigine du deacuteveloppement drsquoun reacuteseau karstique
drainant drsquoimportants eacutecoulements souterrains et la reacutesurgence drsquoeau en surface agrave cause des marnes
impermeacuteables du sous-sol Les pacirctures sont situeacutees sur les pentes douces marneuses des buttes ainsi
que dans les valleacutees alluviales humides Au-dessus des pacirctures un bourrelet boiseacute marque la rupture
Chapitre 3
126
de pente il preacutesente en geacuteneacuteral des affleurements plus ou moins importants de calcaire dur (Figure
22) Les pacirctures sont majoritairement situeacutees sur la mecircme formation geacuteologique des marnes du Lias
ou du Jurassique infeacuterieur correspondant agrave des formations peacutedologiques particuliegraveres caracteacuteriseacutees
par des sols de pentes argilo-limoneux localement hydromorphes (sources et laquo mouilles raquo)
Il est inteacuteressant drsquoobserver que les cantons toucheacutes par la tuberculose bovine en Cocircte drsquoOr
possegravedent une densiteacute de bovins allaitant quasi-identique agrave celle des cantons indemnes localiseacutes dans
le sud-ouest du deacutepartement et limitrophe de la zone drsquoendeacutemie de bTB (Figure 23) Ces deux groupes
de cantons sont localiseacutes sur des formations peacutedologiques diffeacuterentes suggeacuterant que le type de sol des
pacirctures pourrait ecirctre un facteur de risque dans le maintien de la maladie et que la densiteacute des bovins
ne semble pas ecirctre un facteur majeur dans la distribution des foyers de bTB Lrsquoobservation de la
distribution du parcellaire des exploitations foyers de tuberculose bovine sur la carte peacutedologique du
deacutepartement montre que 74 des parcelles de ces exploitations sont localiseacutees sur cette uniteacute
paysagegravere (sols argilo-limoneux et hydromorphes des pentes 48 des parcelles sols calcaires humides
sur alluvions des valleacutees 14 et sols argileux des plateaux calcaires 12 ) (Figure 24Figure 24)
Une enquecircte (Etude PatRisk) reacutealiseacutee dans le cadre drsquoun Master drsquoEpideacutemiologie dans neuf
eacutelevages bovins deacuteclareacutes infecteacutes entre 2010 et 2014 a chercheacute agrave identifier les pacirctures laquo agrave risque raquo de
tuberculose et agrave les caracteacuteriser par des variables concernant les pratiques drsquoeacutelevage le voisinage la
faune sauvage et lrsquoenvironnement (pente type de sol relief exposition humiditeacute) Lrsquoanalyse
multivarieacutee a mis en eacutevidence deux expositions indicatrices de risque la pratique du fauchage avant
la mise agrave lrsquoherbe et la preacutesence de zones humides localiseacutees sur la pacircture (Bouchez-Zacria 2015) Ces
reacutesultats sont en accord avec le fait que la survie de M bovis est favoriseacutee par la preacutesence drsquohumiditeacute
et qursquoune faible hauteur drsquoherbe augmente la possibiliteacute de contact avec les mycobacteacuteries du sol Bien
que des eacutetudes plus preacutecises soient recommandeacutees il semble toutefois que les sols argilo-limoneux
hydromorphes des pacirctures suspectes constituent un facteur de risque agrave prendre en compte dans
lrsquoeacutepideacutemiologie de la maladie
Chapitre 3
127
Figure 21 Orographie et hydrographie de lrsquoAuxois Les valleacutees supportant les parcelles suspectes sont indiqueacutees par une flegraveche noire (Source BD Carthage et BD alti - IGN)
Figure 22 Localisation de lrsquoAuxois en Cocircte drsquoOr structure paysagegravere et geacuteomorphologie de ce territoire
Chapitre 3
128
Figure 23 Nombre de vaches allaitantes par canton dans le deacutepartement de la Cocircte drsquoOr (agrave gauche) et parcellaire des foyers bovins de 2010 agrave 2015 (agrave droite) (Source Recensement agricole 2010 ndash Agreste)
Chapitre 3
129
Figure 24 Reacutepartition spatiale des parcelles des eacutelevages bovins infecteacutes de 2010 agrave 2014 en Cocircte drsquoOr et repreacutesentation des couches peacutedologiques (Reacutealisation Marina Beacuteral CIREV Bourgogne)
Chapitre 3
130
D BILAN GLOBAL
Nous avons deacuteveloppeacute deux systegravemes de deacutetection moleacuteculaire adapteacutes aux matrices
environnementales ciblant les espegraveces du MTBC (IS1561rsquo et Rv3866) et modifieacute un systegraveme existant
speacutecifique de M bovis (RD4) Nous avons valideacute ces systegravemes sur trois matrices diffeacuterentes (sol eau
fegraveces) inoculeacutees avec M bovis BCG en deacutemontrant leur speacutecificiteacute soit pour le MTBC (excluant
M microti potentiellement preacutesent dans lrsquoenvironnement) soit pour M bovis Leur sensibiliteacute est
bonne puisque les seuils de deacutetection obtenus sont infeacuterieurs aux seuils deacutecrits dans la litteacuterature (en
comparant avec une meacutethode de deacutenombrement identique agrave la nocirctre) Le systegraveme RD4 est agrave ce jour le
seul systegraveme de deacutetection speacutecifique de M bovis dans les eacutechantillons environnementaux toutefois
drsquoapregraves notre expeacuterience il preacutesente une sensibiliteacute moins importante que les deux autres systegravemes
malgreacute des seuils de deacutetection eacutequivalents sur les eacutechantillons inoculeacutes au laboratoire
La deacutetection du MTBC par lrsquoamplification de lrsquoIS1561rsquo a eacuteteacute effective dans 42 eacutechantillons (sur
plus de 400) la PCR nicheacutee ciblant la mecircme seacutequence a mis en eacutevidence 14 eacutechantillons positifs et le
systegraveme RD4 trois eacutechantillons positifs Les niveaux de deacutetection varient fortement en fonction des
eacutechantillons lrsquoeffet de dilution reste tregraves important dans des matrices comportant des communauteacutes
microbiennes abondantes et diversifieacutees (quantiteacute relative drsquoADN cible faible par rapport agrave des
quantiteacutes eacuteleveacutees drsquoADN non cible) Lrsquoutilisation de la PCR quantitative a permis dans certains cas de
quantifier les cibles preacutesentes dans les eacutechantillons environnementaux Les sols de terriers preacutesentent
les concentrations en mycobacteacuteries du MTBC les plus eacuteleveacutees pouvant atteindre 46 times 104 copies du
gegravene cibleacute ce qui correspond agrave des quantiteacutes de cellules eacuteleveacutees sachant que le nombre de copies du
gegravene cible par geacutenome est de 1
Parmi les diffeacuterents sites preacuteleveacutes la contamination a eacuteteacute retrouveacutee exclusivement dans deux
eacutecosystegravemes particuliers les zones humides de pacirctures (sources et laquo mouilles raquo) et les sols de terrier
et latrines de blaireaux La contamination apparait le plus souvent localiseacutee agrave une zone geacuteographique
et ponctuelle mais un signal moleacuteculaire positif a eacuteteacute retrouveacute pendant un an dans les sols de terriers
de blaireaux localiseacutes sur une pacircture
Ces reacutesultats correacuteleacutes au contexte paysager et peacutedologique des parcelles suspectes nous ont
ameneacute agrave penser que certaines formations peacutedologiques seraient plus susceptibles de favoriser la
persistance de M bovis Nous avons donc eacutevalueacute lrsquoimpact de la nature du sol retrouveacute dans ces
parcelles sur la survie de M bovis Nous avons eacutegalement eacutevalueacute lrsquoimpact de la tempeacuterature sur la
dureacutee de survie de la bacteacuterie dans le sol
CHAPITRE 4
Impact de la tempeacuterature et de la nature du sol sur la
survie de Mycobacterium bovis SB0120
Elodie Barbier Murielle Rochelet Laurent Gal Maria Laura Boschiroli Alain Hartmann
Experimental investigation of the impact of the temperature and the soil type on Mycobacterium
bovis survival in the environment
(Article en cours de soumission)
133
CHAPITRE 4 - Impact de la tempeacuterature et de la nature du sol sur la
survie de Mycobacterium bovis SB0120
A AVANT-PROPOS
La deacutetection moleacuteculaire de Mycobacterium bovis dans les eacutecosystegravemes agricoles de la zone
endeacutemique de Cocircte drsquoOr a confirmeacute que les animaux infecteacutes et excreacuteteurs ont la capaciteacute de
contaminer les sols et lrsquoeau accessibles agrave la faune domestique et sauvage (Chapitre 2) La bacteacuterie ne
se multipliant pas dans lrsquoenvironnement sa survie dans les substrats environnementaux conditionne la
transmission indirecte du bacille entre un animal infecteacute excreacuteteur et un animal sain par le biais de ces
substrats
Des eacutetudes expeacuterimentales sur divers substrats environnementaux notamment du sol
inoculeacutes avec M bovis ou M bovis BCG ont suggeacutereacute que les principaux facteurs influenccedilant la survie
des bacteacuteries sont les facteurs climatiques tels que tempeacuterature humiditeacute et exposition aux UV (Fine
et al 2011 Duffield amp Young 1985 Jackson et al 1995 Young et al 2005) Bien que quelques eacutetudes
aient rechercheacute par la culture ou par des approches statistiques une eacuteventuelle association de la
contamination environnementale ou animale par des mycobacteacuteries avec la nature du sol les reacutesultats
sont difficilement comparables et extrapolables (Walter et al 2012 Dhand et al 2009 Reviriego et al
2000 Johnson-Ifearulundu amp Kaneene 1999 Ward amp Perez 2004)
Face agrave ce manque de connaissances nous avons donc souhaiteacute eacutevaluer la dureacutee de la survie de
M bovis dans deux sols de Cocircte drsquoOr preacutesentant des caracteacuteristiques physico-chimiques contrasteacutees et
ce agrave deux tempeacuteratures diffeacuterentes afin de rechercher un eacuteventuel impact de la tempeacuterature et de la
nature du sol sur la persistance de la bacteacuterie Pour cette eacutetude nous avons utiliseacute une souche
pathogegravene de M bovis (geacutenotype SB0120) ce geacutenotype est retrouveacute majoritairement dans la zone
Nord chez les bovins et dans la faune sauvage Deux meacutethodes de suivi de la population bacteacuterienne
ont eacuteteacute utiliseacutees la culture sur milieu geacuteloseacute reacutealiseacutee agrave partir de sols irradieacutes et la qPCR pour le suivi
des sols irradieacutes et biotiques
135
B EXPERIMENTAL INVESTIGATION OF THE IMPACT OF THE TEMPERATURE AND THE SOIL TYPE ON MYCOBACTERIUM BOVIS SURVIVAL IN THE ENVIRONMENT
Elodie Barbierab Murielle Rocheletab Laurent Galac Maria Laura Boschirolid Alain Hartmanna
a INRA UMR 1347 Agroeacutecologie Dijon Cedex France
b Universiteacute de Bourgogne Franche Comteacute Dijon Cedex France
c AgroSup Dijon UMR1347 Agroeacutecologie Dijon France
d Universiteacute Paris-Est Laboratoire National de Reacutefeacuterence de la Tuberculose Uniteacute des Zoonoses Bacteacuteriennes Laboratoire de
Santeacute Animale ANSES Maisons-Alfort Cedex France
Key words Mycobacterium bovis soil temperature culture quantitative PCR
Corresponding Author Alain Hartmann UMR 1347 Agroeacutecologie INRA 17 rue Sully BP 86510 21065
Dijon Cedex France E-mail alainhartmanndijoninrafr
INRA UMR1347 Agroeacutecologie BP 86510 F-21000 Dijon France
Chapitre 4
136
Abstract
Mycobacterium bovis the causative agent of the bovine tuberculosis (bTB) mainly affect cattle its
natural reservoir but also a wide range of domestic and wild mammals Besides direct transmission via
contaminated aerosols indirect transmission of the disease between wildlife and livestock is thought
to occur by inhalation or ingestion of environmental substrates contaminated by M bovis through
infected animal shedding We monitored the survival of M bovis in two soils chosen for their contrasted
physico-chemical properties at two temperatures The population of M bovis spiked in disinfected soils
was enumerated by a culture-based method after 14 30 60 90 120 and 150 days of incubation at 4degC
and 22degC The fate of M bovis spiked in disinfected or biotic soils was monitored using a qPCR based
assay targeting the IS1561rsquo locus The analysis of survival profiles using culture-based method showed
that M bovis survived longer at cooler temperature but impact of soil characteristics on M bovis
persistence was not obvious Futhermore qPCR based assay detected M bovis longer than the culture
based method with higher gene copy numbers in sterile soils than in biotic ones Impact of soil type on
M bovis persistence need to be further studied in order to shed light on the risk and mechanisms of
indirect transmission of the disease
Chapitre 4
137
INTRODUCTION
Mycobacterium bovis is a pathogenic mycobacteria responsible for bovine tuberculosis (bTB)
Tuberculosis caused by M bovis is a primarily respiratory disease that may also affect various organs in
animals (Morris et al 1994) Even if bTB mainly concerns cattle a wide range of domestic and wild
mammals may be infected as well (Biet et al 2005) On the other hand bTB a zoonotic disease
impacting human health (Peacuterez-Lago et al 2013) Although cattle is considered as the principal host of
M bovis wild fauna such as badgers (Meles meles) wild boar (Sus scrofa) and red deer (Cervus elaphus)
has recently been identified as potential wildlife reservoirs (Palmer 2013) Though the transmission of
M bovis among cattle usually occurs via the inhalation of contaminated aerosols released by infected
animals andor the close contacts (Morris et al 1994 Phillips et al 2003) the indirect transmission
either by inhalation of environmental bioaerosols or ingestion of infected environmental matrices may
be another potential routes of animal infection (Vicente et al 2007 Palmer et al 2004 Phillips et al
2003) Indirect transmission through feed or water has already been demonstrated between deer deer
and cattle (Palmer et al 2004 Palmer et al 2001) and badgers and cattle (Little et al 1982) in laboratory
experiments
Investigating this type of indirect transmission is challenging because it results at least from the
combination of three essential factors iei) the environmental contamination by shedding from
infected animals ii) the persistence of the bacteria under a virulent state in environmental matrices
and finally iii) the interaction between a new susceptible host with the contaminated matrices The
shedding step by animals has already been demonstrated for many species via oro-nasal mucus
sputum urine feces and wound discharges depending on the animal species (Corner et al 2012
Palmer et al 2004 Barasona et al 2015) Secondly a few previous experimental studies using various
environmental substrates spiked with M bovis showed that the bacteria was able to survive for a long
time period outside of its host (Williams amp Hoy 1930 Duffield amp Young 1985 Young et al 2005 Fine et
al 2011 Ghodbane et al 2014) For example using a culture-based method Ghodbane et al (2014)
recovered M bovis bacteria from sterile soils incubated in controlled laboratory conditions 12 months
after their inoculation whereas an incubation under the natural weather conditions of Michigan USA
allowed the persistence of the bacteria in sterile soils for 88 days (Fine et al 2011) Moreover several
studies have identified several climatic factors such as a cool temperature an adequate moisture and
a protection against solar radiation (ultra violet) as positive factors for the persistence of M bovis in
environmental matrices like feces (Williams amp Hoy 1930 Tanner amp Michel 1999) food supply such as
corn hay apples (Palmer amp Whipple 2006 Fine et al 2011) water (Little et al 1982 Fine et al 2011)
and soil (Duffield amp Young 1985 Fine et al 2011) (Duffield amp Young 1985 Williams amp Hoy 1930 Tanner
amp Michel 1999 Jackson et al 1995) When Fine et al (2011) compared the persistence of M bovis in
Chapitre 4
138
four substrates (corn hay water and soil) the longest survival was found in cool and moist soil (88
days in winterspring) suggesting that soil probably ensured better conditions of persistence for
M bovis than the other matrices tested Moreover M bovis was often detected in soils especially in
badger sett soil or in pasture soil (Courtenay et al 2006 Santos et al 2015 Sweeney et al 2007
2006 Young et al 2005 Barbier et al manuscript submitted)
More generally low pH and high iron content are often described as major factors driving
mycobacterial survival in soils especially for Mycobacterium avium ssp paratuberculosis (Johnson-
Ifearulundu amp Kaneene 1999 Ward amp Perez 2004) Concerning M bovis one statistical study
established a relationship between the occurrence of bTB in animals and soil parameters by showing
that every a 1 increase in the sandy content of soil led to an increase in odds of bTB infection by 4
(Walter et al 2014) suggesting that soil type may promote or hinder M bovis environmental
persistence
To our knowledge no comprehensive study focuses on the influence of soil biotic and abiotic
parameters on the persistence M bovis With the aim to further assess the role of the soil
characteristics on the persistence of Mbovis and thus to better understand the impact of the
environment in the epidemiology of the bTB disease the influence of both the physicochemical
characteristics of the soil and the temperature was studied under controlled laboratory conditions For
this purpose two different soils selected for their contrasted properties were spiked with known
concentrations of M bovis and incubated at two different temperatures to mimic seasonal
temperatures The persistence of M bovis over time was monitored using both a culture-based
method for the detection of viable M bovis and a molecular detection method based on a qPCR assay
MATERIAL AND METHODS
All experiments were conducted in a biosafety level 3 laboratory unless otherwise stated
Mycobacterium bovis cultivation and inoculum preparation
The strain Mycobacterium bovis SB0120 VNTR 5 5 4 3 11 4 5 6 that infects cattle and wild
animals such as badgers and wild boar in France was provided by the National Reference Laboratory of
bovine tuberculosis (Maisons-Alfort France) and grown aerobically at 37degC in Middlebrook 7H9
supplemented with 10 Middlebrook ADC Growth Supplement (Sigma-Aldrich France) for thirty days
The M bovis inoculum was prepared as following a 20 mL stationary phase culture was pelleted and
once the supernatant removed the cell aggregates were disrupted by vortexing with glass beads for 15
seconds The pellet was suspended in 1 mL of sterile water constituting the stock solution and serial
10-fold dilutions were subsequently plated in triplicate onto Middlebrook 7H11 supplemented with
10 heat inactivated bovine serum (Dominique Dutscher France) and 10 Middlebrook OADC
Chapitre 4
139
Growth Supplement (Sigma-Aldrich France) for enumeration Colonies were counted following
incubation for two months at 37degC The inoculum for the culture-based assay and the qPCR-based assay
were respectively prepared by diluting 10-fold and 1000-fold the M bovis stock solution
Soil characterization and microcosm preparation
Two different soils were collected in two cattle-grazed pastures at 0-30 cm depth in Cocircte drsquoOr
(a department located in the Middle East of France) The soil A is a clay loamy soil (calcisol) and soil B
is a loamy sandy soil Soils were prepared as following soil samples were gently air dried and sieved to
recover 4 mm aggregates The moisture contents of soils were determined by drying for 24 h at 105degC
and their physico-chemical parameters were determined by the Soil Analysis Laboratory of Arras
France Fresh sieved soil was used to prepare the microcosms
Vials of 15 mL (Falcon Dominique Dutscher France) were filled with 2 g of the soils A and B Half of the
soil vials were sterilized by gamma-irradiation up to an internal minimum dose of 35 KGray (Ionisos
France) All the microcosm soils (biotic and sterile) were then adjusted to 90 of the water holding
capacity We verified that neither soil A nor B were positive for M bovis with the culture-based method
(sterile soils) and with the qPCR assay (sterile and biotic soils) described later All the soil microcosms
were spiked with 100 microL of the M bovis inoculum 10- or 1000-fold diluted (for sterile and biotic soil
respectively) and stored in the dark in a biosafety level 3 laboratory until their analysis Sterile spiked
soils were incubated at 4degC and 22degC while biotic spiked soils were only incubated at 22degC Vials were
opened in a microbiological safety cabinet every fifteen days to regenerate their atmosphere
Schedule of microcosm sampling
In order to monitor M bovis cell numbers over time both plate enumeration and qPCR assay
were performed on irradiated soils while qPCR assay only was carried out for biotic soils At each
sampling date ie day 0 14 30 60 90 120 and 150 three mesocosms of each biotic and sterile soils
were analysed with the qPCR assay after DNA extractionpurification and four mesocosms of each
sterile soil at each temperature were analysed using culture-based method
DNA extraction and purification
DNAs from the soil samples were extracted as follows 8 mL of lysis buffer (100 mM Tris pH 80
100 mM EDTA pH 80 100 mM NaCl and 2 (wv) SDS) 4 g of silica beads (100 μm diameter) 5 g of
ceramic beads (14 mm diameter) and 8 glass beads (4 mm diameter) were added to each vial of soil
However as DNA extraction could not be achieved within the biosafety level 3 laboratory for practical
reason soil was suspended in 8 mL of lysis buffer and heated 2 hours at 80degC to kill viable bacteria The
following steps of DNA extraction were then done in a biosafety level 2 laboratory Samples were
Chapitre 4
140
disrupted for 3 times 30 s at 4 ms in a FastPrepreg-24 Instrument (MP Biomedicals Europe France)
incubated for 30 min at 70degC and centrifuged for 5 min at 7000 timesg at room temperature Supernatants
were incubated on ice for 10 min with 110 volume of 3 M potassium acetate pH 55 and then
centrifuged for 5 min at 14000 times g One volume of ice-cold isopropanol was added to the supernatant
for DNA precipitation overnight at minus 20degC DNA was collected by centrifugation (30 min at 14000 times g)
and DNA pellets were washed with ice-cold ethanol (70 ) and dissolved in 100 μL of water Crude DNA
extracts (100 μL) were loaded on Microbiospintrade columns (Biorad France) filled with 100 mg of
polyvinyl polypyrrolidone (PVPP Sigma Chemical Co France) After centrifugation (4 min at 1000 times g
4 degC) eluted DNA was further purified using the Geneclean Turbo Kit (Qiagen France) according to the
manufacturers instructions Purified DNA concentrations were estimated by spectrophotometry (λ =
260 nm) using a NanoDropreg 2000 spectrophotometer (Thermo Scientific France)
Estimation of DNA quality for PCR amplification
The quality of the purified DNAs extracted from the two soils was assessed by measuring the
capacity to amplify a control DNA target in the presence of various dilutions of soil DNA extracts Briefly
qPCR reactions were done in a final volume of 25 microL containing 25 μl of 106 copies of the circularized
pGEM-T Easy plasmid DNA and 5 μl of water (control) or diluted soil DNA (10- and 20-fold dilutions)
1X Absolute qPCR SYBR Green ROX (500 nM) Mix (Thermo Scientific) 25ngmicroL of T4 bacteriophage
gene 32 product qPCR primers were SP6 (5rsquondashATTTAGGTGACACTATAG-3rsquo) and T7 (5rsquo-
TAATACGACTCACTATAGGG-3rsquo) universal primers (final concentration 500 nM) targeting the polylinker
of the plasmid qPCR and cycling conditions were as following initial denaturation at 95degC for 15 min
followed by 40 cycles with denaturation at 95degC for 15 s annealing at 55degC and elongation at 72degC for
30 s each The analysis of the melting curves allowed to assess PCR specificity A tenfold reduction of
the copy number amplified by qPCR in the presence of diluted soil DNA with respect to the copy
number obtained in the control reaction (water) was considered as 10 amplification efficiency or
conversely as 90 inhibition of the PCR
Cuture-based method
The sterile soil microcosms were suspended in 9 mL of Middlebrook 7H9 supplemented with
ADC by vortexing the vials for 2 min Tenfold dilutions of soil suspensions were achieved and 100 microL
were plated in triplicate onto Middlebrook 7H11 agar enriched as described above and supplemented
with antibiotics and antifungal (100 mgL-1Ticarcillin 10 mgL-1 Trimethoprim 200 000 UIL-1 Polymyxin
B and 100 mgL-1 Cycloheximid Sigma-Aldrich France) for enumeration Plates were then incubated in
gas-permeable bags at 37degC for 2 months and checked every 15 days
Chapitre 4
141
Mycobacterium tuberculosis complex quantitative PCR (qPCR)
Mycobacterium bovis was detected using the Mycobacterium tuberculosis complex (MTBC)
specific qPCR targeting the IS1561rsquo locus (Forward 5rsquo- GATCCAGGCCGAGAGAATCTG -3rsquo Reverse 5rsquo-
GGACAAAAGCTTCGCCAAAA - 3rsquo and probe 5rsquo - FAM ACGGCGTTGATCCGATTCCGC TAMRA - 3rsquo) as
previously described (Barbier et al manuscript submitted for publication)
qPCR reactions were carried out in triplicate with a 25 microL reaction mix containing 125 microL of ABsolutetrade
QPCR ROX Mix (Thermo Scientific France) 1 microL (final concentration 25 ngmicroL-1) of T4 bacteriophage
gene 32 product (MP Biomedicals Europe France) 1 microL of each primer (final concentration 400 nmolL-
1) 05 microL of each probe (final concentration 200 nmol L-1) 4 microL of ultrapure water and 5 microL of 20-fold
diluted DNA Absolute quantification was achieved using standard DNA dilutions A recombinant
plasmid containing one copy of the IS1561rsquo fragment of M bovis BCG strain Pasteur 1173P2 cloned in
the pCRregII-TOPOreg vector (Invitrogen France) was used as a standard A calibration curve was obtained
by amplification of serial dilutions of the plasmid ranging from 1 to 106 copies per PCR reaction All
DNAs extracted from the soil samples were screened with the IS1561rsquo-based qPCR assay and results
were expressed as the number of M bovis gene copy number per g of soil qPCR assay was performed
in a VIIA 7 Real-Time PCR System (Thermo Fischer Scientific France) Initial denaturation was done at
95degC for 15 min followed by fourty-five cycles with denaturation at 95degC for 15 s annealing and
elongation at 60degC for 1 min Detection limits of this molecular system was previously determined in
soil (Barbier et al manuscript submitted for publication)
RESULTS
Physico-chemical characteristics of soils A and B
The physico-chemical characteristics of soils A and B are summarized in Table 1 The soil A is a
clay loamy soil with a slightly basic pH (775) and a high content of clay (clay account for 397 in
particle-size distribution and sand for only 118 ) while the soil B is a loamy sandy soil with an acidic
pH (546) and a high content of sand (sand account for 345 in particle-size distribution and clay for
only 192 ) Silt content was roughly equivalent in the two soils (485 g kg-1 for soil A vs 463 g kg-1 for
soil B) We chose to adjust soil moisture to 90 of their water field holding capacity leading to a
moisture content of 31 for soil A and 21 for soil B Finally the C-to-N ratio was 95 for soil A and
109 for soil B
Chapitre 4
142
Table 1 Physico-chemical parameters of soils A and B
Physical parameters Soil A Soil B Unit
Clay (lt 2 microm) 397 192 g kg-1
Silt 485 463 g kg-1
Sand 118 345 g kg-1
Water field capacity 352 247 g kg-1
Chemical parameters -
pH 775 546
Total C 361 292 g kg-1
Total N 38 269 g kg-1
C- to N- ratio 95 109 -
Organic matter 625 506 g kg-1
P 0031 0025 g kg-1
K 0158 00877 g kg-1
Ca 717 11 g kg-1
Na 00115 00129 g kg-1
Mg 00541 00789 g kg-1
Fe 0526 0227 g kg-1
CECa 183 108 cmol+kg-1b
aCEC Cation Exchange Capacity bcmol+kg-1 centimoles of positive cations per kg of soil
Effect of temperature and soil type on M bovis survival time measured by culture-based method
Theoretical detection limit of our culture-based method was estimated at 15 times 103 CFU g-1 soil
Total cultivable M bovis (CFU g-1 soil) were enumerated by the culture-based method described above
for each sterile soil incubated at 4degC and 22degC at day 0 14 30 60 90 120 and 150 At day 0 number
of M bovis was estimated at 15 (plusmn 06) times 107 CFU g-1 for soil A and 17 (plusmn 07) times 107 CFU g-1 for soil B by
the culture-based method
Two different patterns of mycobacterial survival were observed depending on the temperature
(Figure 1a and 1b) At 22degC M bovis cells decreased over the course of the experiment in the two
sterile soils A and B (Figure 1a) No more cultivable bacteria growth was observed from day 90 in soil B
and day 120 in soil A to the end of the experiment at day 150 Although M bovis SB0120 was detected
by culture for one month longer in soil A than in soil B no statistically significant difference of CFU
numbers was observed between the two soils at 22degC (Student-Newman-Keuls Test p = 0559) thus
no significant difference in M bovis survival was observed due to soil type
At 4degC M bovis cultivable cells were recovered in high concentrations in the two soils at every
sampling date (over the five months of the experiment) The M bovis CFU numbers remained stable
during the first 30 days in the two soils as indicated in Figure 1b We observed a slight decrease of the
Chapitre 4
143
CFU numbers in the two soils starting from day 60 to the end of the study In both soils M bovis CFU
numbers decreased significantly from day 0 to day 150 (Student-Newman-Keuls Test plt0001)
reaching 87 (plusmn 25) times 10 5 CFU g- 1 and 22 (plusmn 11) times 10 5 CFU g- 1 in soil A and B respectively
Effect of soil type and endogenous microbiota on M bovis survival time measured by qPCR assay
Survival of M bovis when biotic or sterile soils A and B were incubated at 22degC was monitored
using the qPCR based assay The M bovis gene copy numbers were estimated with IS1561rsquoqPCR-based
method for each sterile and biotic soils incubated at 22degC at day 0 14 30 60 90 120 and 150 (see
Figure 2) At day 0 number of M bovis gene copies per g of soil yielded 36 (plusmn 12) times 105 and 60 (plusmn 19)
times 105 respectively for sterile soils A and B and 12 (plusmn 07) times 105 and 25 (plusmn 15) times 105 respectively for
biotic soils A and B by the qPCR-based method
Overall high amounts of M bovis gene copies were detected in the four soils at the eight sampling
dates
A statistically significant interaction between time and soil condition (biotic vs sterile) was
observed when considering the two soils independently and dependently (Student-Newman-Keuls
Test plt0001) with higher amounts of gene copies recovered in sterile soils than in biotic ones from
day 0 to day 150 No significant differences were observed when comparing the gene copy numbers in
the two sterile soils or in the two biotic soils at each sampling date
Even though the M bovis gene copy numbers slightly oscillated in the four soils over the course of the
experiment a slight but significant decrease of these numbers was observed between day 0 and day
150 for each soil The highest decreases of M bovis were observed in the biotic soils with a rough loss
of 1 log for soil B and 075 log for soil A between day 0 and day 150 (Figure 2)
Finally qPCR assay allowed a longer M bovis detection in sterile soils A and B incubated at 22degC
than culture-based assay Indeed M bovis DNA was detected by qPCR for up to 150 days in the two
sterile soil samples while bacterial culture detected viable M bovis cells up to 60 days in soil D and 90
days in soil C
Chapitre 4
144
Figure 1 Detection of M bovis SB0120 by culture-based method in sterile soils A and B after incubation at 22degC (figure above) and 4degC (figure below) Error bars represent the standard deviation values on 8 replicates
Figure 2 Detection of M bovis gene copy numbers by IS1561rsquo qPCR-based assay on sterile and biotic soils A and B after incubation at 22degC Error bars represent standard deviation values on 9 replicates
10E+00
10E+01
10E+02
10E+03
10E+04
10E+05
10E+06
10E+07
10E+08
0 14 30 60 90 120 150
M b
ovi
s C
FU n
um
ber
per
g o
f
ster
ile s
oil
Sampling day
22degC
Soil A
Soil B
10E+00
10E+01
10E+02
10E+03
10E+04
10E+05
10E+06
10E+07
10E+08
0 14 30 60 90 120 150
M b
ovi
s C
FU n
um
ber
per
g o
f st
erile
so
il
Sampling day
4degC
Soil A
Soil B
10E+00
10E+01
10E+02
10E+03
10E+04
10E+05
10E+06
0 14 30 60 90 120 150
M b
ovi
s IS
15
61
co
py
nu
mb
er p
er g
of
ster
ile s
oil
Sampling day
Sterile soil A Sterile soil B Soil A Soil B
Chapitre 4
145
DISCUSSION
Contamination of the environment with M bovis is considered as essential for the persistence
of bovine tuberculosis in animals and its interspecific indirect transmission The survival of this
mycobacteria in soil has been documented (Duffield amp Young 1985 Jackson et al 1995 Fine et al 2011
Young et al 2005) but limited information is available on the soils used and the edaphic factors that
may affect M bovis persistence are poorly understood These experiments have dealt with controlled
or environmental parameters that promote or hinder M bovis persistence in soil such as temperature
moisture or UV exposition but to our knowledge impact of the soil type has not been thoroughly
addressed This study was designed to evaluate temperature and soil type impact on M bovis survival
In this study we demonstrated that M bovis SB0120 may survive several months in soil under
different laboratory conditions in two contrasted soils and that cool temperature (4degC) strongly
promotes M bovis persistence whatever the soil type while warmer temperature (22degC) shortens its
survival time Our results are in contradiction with a previous laboratory study on M bovis persistence
(Young et al 2005) where mycobacterial survival was longer at 25degC than at 4degC in a sterile sandy soil
(pH 65-68 60 days vs 16 days) but in agreement with previous field studies where lower temperature
increased M bovis survival time up to 88 days (Fine et al 2011 Jackson et al 1995)
We also pointed out that soil type may impact on M bovis survival time Indeed despite that
no statistical difference was observed the soil A that exhibits high clay and loam content and slightly
basic pH allowed persistence of cultivable M bovis for up to three months (90 days) while this
persistence was only two months in the acidic sandy loamy soil B when soils were incubated at 22degC
On the contrary a difference of M bovis survival was observed at 4degC between the two soils with a
better survival in soil A than in soil B suggesting an impact of soil type on M bovis survival at cool
temperature However we were hampered by the detection limit of the culture-based method in the
experiment on sterile soil at 22degC where CFU numbers and detection limit ovelaped after day 60
Enrichment broth are very useful to recover low number of bacteria in complex matrices such as
Salmonella typhimurium in food or Listeria monocytogenes in soil (Locatelli et al 2013 Zheng et al
2013) but mycobacterial enrichment broths are not yet available for mycobacteria
In our study clay soil with slightly basic pH seemed to promote M bovis SB0120 survival
although mycobacterial culture detection or persistence are usually associated with sandy soils
harbouring acidic pH as previously suggested by positive culture or statistical studies (Walter et al
2014 Johnson-Ifearulundu amp Kaneene 1999 Reviriego et al 2000 Santos et al 2015) suggesting an
improved survival of the mycobacteria such as M avium ssp paratuberculosis and M bovis in these
soils However a one year persistence of cultivable M bovis was demonstrated in a sandy soil with a
basic pH (83) spiked with 108 CFU g- 1 (Ghodbane et al 2014) suggesting that M bovis may survive a
Chapitre 4
146
long time in soils harbouring a wide range of pH Moreover iron content of soils A and B seemed to be
significantly higher than soils described in previous studies
In this study qPCR was able to detect the presence of M bovis DNA over a longer period of
time than bacterial culture in soils samples incubated at 22degC whatever the soil type As one previous
study reported that DNA of dead M bovis cells was no more amplified 10 days after cell death (Young
et al 2005) we hypothesized that our qPCR assay detected live M bovis up to 150 days after
inoculation of soils Viable M bovis cells have already been detected in sterile and biotic soil over a 15
month period targeting the 16S rRNA encoding gene (Young et al 2005) Physiological state of
pathogenic mycobacteria such as M bovis in soil is unknown but dormancy or a viable but non
cultivable state (VBNC) have been suggested to explain their survival in environmental matrices
(Whittington et al 2004 Adams et al 2013) Further analysis of the expression of dormancy-related
genes (Aacutelvarez et al 2009) under soil conditions might allow to support this hypothesis Further
researches are critical to ascertain the long bacterial survival in soil and to understand the genetic
mechanisms involved in soil persistence
Our laboratory conditions were far from field conditions where temperature fluctuations
moisture variations and UV radiation are detrimental to the mycobacteria Mycobacterium bovis
survival time obtained in this study is probably longer than survival time in environmental conditions
As the gap of knowledge is important about how M bovis survive in soils laboratory studies have to
be set out in order to determine the main factors involved in M bovis survival However experiments
dealing with the exposition of M bovis naturally or artificially contaminated samples to environmental
conditions must be carried out in specific open air enclosure with high levels of protection to exclude
mycobacterial dissemination (Fine et al 2011) or require climate chambers with a L3 biosafety level
Significant differences in the recovery of M bovis DNA between biotic and sterile soils were
observed with the qPCR-based assay from the first day of the experiment indicating that qPCR
detection is hampered when soil metagenomic DNA is concentrated although no inhibition was
observed
In our experiment the use two soils gave an insight on the impact of soil type on M bovis
survival However extensive studies on larger number of well-characterised and contrasted soils would
be required to target the key edaphic factors that promote M bovis survival in soil Further work is
needed to identify those key factors and determine if the population detected with qPCR consists of
non-culturable or dormant cells capable of resuscitation under specific growth conditions or following
inhalation or ingestion by animals
Chapitre 4
147
Funding information
The work was supported by the Regional Council of Burgundy (Elodie Barbier PhD fellowship) the Cocircte
drsquoOr General Council Interbev and the Cocircte drsquoOr GDS (Groupement de Deacutefense Sanitaire Livestock
health protection group) The funders had no role in study design data collection and interpretation
or the decision to submit the work for publication
Conflict of interest
The authors have no conflict of interest to declare
Acknowledgements
The authors would like to thank Jean-Marie Duez Julien Bador and Catherine Neuwirth from the
Plateau Technique de Biologie of the Hospital of Dijon
REFERENCES
Adams AP et al 2013 Comparison of PCR versus culture for detection of Mycobacterium bovis after experimental inoculation of various matrices held under environmental conditions for extended periods Applied and Environmental Microbiology 79(20) pp6501ndash6506
Aacutelvarez AH Estrada-Chaacutevez C amp Flores-Valdez MA 2009 Molecular findings and approaches spotlighting Mycobacterium bovis persistence in cattle Veterinary Research 40(3) pp1ndash16
Barasona JA et al 2015 DNA detection reveals Mycobacterium tuberculosis complex shedding routes in its wildlife reservoir the Eurasian wild boar Transboundary and Emerging Diseases (DECEMBER)
Biet F et al 2005 Zoonotic aspects of Mycobacterium bovis and Mycobacterium avium-intracellulare complex (MAC) Veterinary Research 36 pp411ndash436
Corner LAL et al 2012 The distribution of Mycobacterium bovis infection in naturally infected badgers Veterinary journal (London England 1997) 194(2) pp166ndash72
Courtenay O et al 2006 Is Mycobacterium bovis in the environment important for the persistence of bovine tuberculosis Biology letters 2(3) pp460ndash2
Dhand NK et al 2009 Association of farm soil characteristics with ovine Johnersquos disease in Australia Preventive Veterinary Medicine 89 pp110ndash120
Duffield BJ amp Young DA 1985 Survival of Mycobacterium bovis in defined environmental conditions Veterinary Microbiology 10 pp193ndash197
Fine AE et al 2011 A study of the persistence of Mycobacterium bovis in the environment under natural weather conditions in Michigan USA Veterinary medicine international 2011 p765430
Chapitre 4
148
Ghodbane R et al 2014 Long-term survival of tuberculosis complex mycobacteria in soil Microbiology (Reading England) 160(Pt 3) pp496ndash501
Jackson R De Lisle GW amp Morris RS 1995 A study of the environmental survival of Mycobacterium bovis on a farm in New Zealand New Zealand Veterinary Journal 43(7) pp346ndash352
Johnson-Ifearulundu Y amp Kaneene JB 1999 Distribution and environmental risk factors for paratuberculosis in dairy cattle herds in Michigan American journal of veterinary research 60(5) pp589ndash96
Little TW Naylor PF amp Wilesmith JW 1982 Laboratory study of Mycobacterium bovis infection in badgers and calves The Veterinary record 111(24) pp550ndash7
Locatelli A et al 2013 Nation-wide study of the occurrence of Listeria monocytogenes in French soils using culture-based and molecular detection methods Journal of microbiological methods 93(3) pp242ndash50
Morris RS Pfeiffer DU amp Jackson R 1994 The epidemiology of Mycobacterium bovis infections Veterinary microbiology 40(1-2) pp153ndash77
Palmer M V 2013 Mycobacterium bovis Characteristics of wildlife reservoir hosts Transboundary and Emerging Diseases 60(SUPPL1) pp1ndash13
Palmer M V Waters WR amp Whipple DL 2004 Investigation of the transmission of Mycobacterium bovis from deer to cattle through indirect contact American Journal of Veterinary Research 65(11) pp1483ndash1489
Palmer M V amp Whipple DL 2006 Survival of Mycobacterium bovis on feedstuffs commonly used as supplemental feed for white-tailed deer (Odocoileus virginianus) Journal of wildlife diseases 42(4) pp853ndash8
Palmer M V Whipple DL amp Waters WR 2001 Experimental deer-to-deer transmission of Mycobacterium bovis American journal of veterinary research 62(5) pp692ndash6
Peacuterez-Lago L Navarro Y amp Garciacutea-de-Viedma D 2013 Current knowledge and pending challenges in zoonosis caused by Mycobacterium bovis A review Research in Veterinary Science 97 ppS94ndashS100
Phillips CJC et al 2003 The transmission of Mycobacterium bovis infection to cattle Research in veterinary science 74(1) pp1ndash15
Reviriego FJ Moreno M a amp Domiacutenguez L 2000 Soil type as a putative risk factor of ovine and caprine paratuberculosis seropositivity in Spain Preventive Veterinary Medicine 43 pp43ndash51
Santos N et al 2015 Widespread environmental contamination with Mycobacterium tuberculosis Complex revealed by a molecular detection protocol Plos One 10(11) pe0142079
Sweeney FP et al 2007 Environmental monitoring of Mycobacterium bovis in badger feces and badger sett soil by real-time PCR as confirmed by immunofluorescence immunocapture and cultivation Applied and environmental microbiology 73(22) pp7471ndash3
Chapitre 4
149
Sweeney FP et al 2006 Immunomagnetic recovery of Mycobacterium bovis from naturally infected environmental samples Letters in applied microbiology 43(4) pp364ndash9
Tanner M amp Michel AL 1999 Investigation of the viability of M bovis under different environmental conditions in the Kruger National Park Journal of Veterinary Research (66) pp185ndash190
Vicente J et al 2007 Risk factors associated with the prevalence of tuberculosis-like lesions in fenced wild boar and red deer in south central Spain Veterinary Research 38(3) pp451ndash464
Walter WD et al 2014 Linking bovine tuberculosis on cattle farms to white-tailed deer and environmental variables using Bayesian hierarchical analysis PloS one 9(3) pe90925
Walter WD et al 2012 On-farm mitigation of transmission of tuberculosis from white-tailed deer to cattle literature review and recommendations Veterinary medicine international 2012 p616318
Ward MP amp Perez AM 2004 Association between soil type and paratuberculosis in cattle herds American Journal of Veterinary Research 65(1) pp10ndash14
Whittington R et al 2004 Survival and dormancy of Mycobacterium avium subsp paratuberculosis in the environment Applied and environmental microbiology 70(5) pp2989ndash3004
Williams RS amp Hoy WA 1930 The viability of B tuberculosis (bovinus) on pasture land in stored faeces and in liquid manure The Journal of hygiene 30(4) pp413ndash419
Young JS Gormley E amp Wellington EMH 2005 Molecular detection of Mycobacterium bovis and Mycobacterium bovis BCG (Pasteur) in soil Applied and environmental microbiology 71(4) pp1946ndash1952
Zheng Q et al 2013 Comparison of enrichment broths for the recovery of healthy and heat-injured Salmonella typhimurium on raw duck wings Journal of food protection 76(11) pp1963ndash8
Chapitre 4
150
C BILAN
Les reacutesultats preacutesenteacutes dans ce chapitre deacutemontrent la capaciteacute de M bovis SB0120 agrave survivre
plusieurs mois dans les deux sols de Cocircte drsquoOr sous une forme viable cette dureacutee eacutetant impacteacutee par
un facteur abiotique particulier la tempeacuterature Une survie drsquoau moins cinq mois est observeacutee agrave basse
tempeacuterature (4degC) dans les deux sols avec une tregraves faible diminution de la population bacteacuterienne
initialement inoculeacutee alors qursquoelle nrsquoest que de deux agrave trois mois agrave 22degC Bien que M bovis soit cultiveacutee
un mois de plus dans le sol C par rapport au sol D nous nrsquoavons pas mis en eacutevidence drsquoeffet de la nature
du sol sur la survie de la bacteacuterie agrave 22degC Toutefois agrave cause de contraintes techniques notre analyse
nrsquoa porteacute que sur deux sols limitant fortement lrsquointerpreacutetation des reacutesultats Lrsquoanalyse des sols inoculeacutes
et incubeacutes agrave 22degC par qPCR montre un tout autre profil de survie puisque M bovis est deacutetecteacutee en
quantiteacute importante pendant les cinq mois de lrsquoeacutetude sur les deux sols et les deux conditions (biotique
vs steacuterile)
Les reacutesultats divergents obtenus par la culture et la qPCR sur les sols incubeacutes agrave 22degC amegravenent
agrave faire diffeacuterentes hypothegraveses pour expliquer lrsquoabsence de croissance des bacteacuteries puisque la qPCR
deacutetecte lrsquoADN des cellules vivantes et mortes ainsi que lrsquoADN libre Nous pouvons donc envisager que
lrsquoADN deacutetecteacute entre 60 et 150 jours appartienne agrave des cellules viables non cultivables ou agrave des cellules
mortes entiegraveres ou lyseacutees Des eacutetudes compleacutementaires sont indispensables pour preacuteciser lrsquoeacutetat
physiologique des bacteacuteries devenues incultivables apregraves une incubation dans le sol
En extrapolant les reacutesultats obtenus aux conditions de tempeacuterature rencontreacutees en Cocircte drsquoOr
nous pouvons suggeacuterer une fenecirctre temporelle pendant laquelle la survie de M bovis dans le sol est
maximale La Figure 25 repreacutesente les tempeacuteratures minimales et maximales releveacutees agrave Dijon entre
janvier 2012 et deacutecembre 2015 Nous estimons que les tempeacuteratures rencontreacutees dans lrsquoAuxois sont
probablement plus proches des tempeacuteratures minimales observeacutees dans la plaine de Dijon-Longvic ougrave
se trouve la station meacuteteacuteo (anthropisation minimale altitude humiditeacute veacutegeacutetation) que des
maximales Ainsi la peacuteriode la plus agrave risque va de novembre agrave avril (tempeacuteratures minimales de
lrsquoair lt 5degC) et correspond agrave la fin de lrsquoautomne lrsquohiver et le deacutebut du printemps Durant le reste de
lrsquoanneacutee et en particulier en eacuteteacute (tempeacuteratures maximales de lrsquoair gt 22degC) les risques de survie de M
bovis semblent moins importants Ces conclusions restent des hypothegraveses dans la mesure ougrave la
tempeacuterature reacuteelle des sols nrsquoa pas eacuteteacute mesureacutee et la variation des autres facteurs climatiques nrsquoa pas
eacuteteacute prise en compte Cependant on peut penser qursquoen eacuteteacute (sous forte insolation) la tempeacuterature des
sols peut augmenter rapidement dans les quelques premiers centimegravetres de sol limitant la dureacutee de
survie de la bacteacuterie
Chapitre 4
151
Figure 25 Tempeacuteratures minimales et maximales observeacutees agrave Dijon-Longvic de 2012 agrave 2015 (Source Meacuteteacuteo France)
-5
0
5
10
15
20
25
30
35
1 2 3 4 5 6 7 8 9 10 11 12
Tmin2012
Tmin2013
Tmin2014
Tmin2015
Tmax2012
Tmax2013
Tmax2014
Tmax2015
4degC
22degC
CHAPITRE 5
Rocircle de la faune du sol dans la circulation de
Mycobacterium bovis eacutetude de la disseacutemination de
M bovis par les lombrics
E Barbier B Chantemesse M Rochelet L Fayolle L Bollache M L Boschiroli A Hartmann
Rapid dissemination of Mycobacterium bovis from cattle dung to soil by the earthworm
Lumbricus terrestris
(Article accepteacute en cours de publication Veterinary Microbiology)
155
CHAPITRE 5 - Rocircle de la faune du sol dans la circulation de
Mycobacterium bovis eacutetude de sa disseacutemination par les lombrics
A AVANT-PROPOS
Le rocircle de la faune et de la microfaune du sol dans la survie et la disseacutemination de
Mycobacterium bovis dans lrsquoenvironnement est inconnu aujourdrsquohui Comme nous lrsquoavons eacutevoqueacute dans
la synthegravese bibliographique les protozoaires plus particuliegraverement les amibes jouent un rocircle
vraisemblable dans la survie des mycobacteacuteries environnementales (Adekambi et al 2006 Mba Medie
et al 2011 Mura amp Bull 2006) voire dans la virulence des mycobacteacuteries opportunistes (Cirillo et al
1997) In vitro la survie intra-amibienne de M bovis pendant plusieurs mois a eacuteteacute deacutemontreacutee mais
aucune eacutetude de terrain eacutevaluant le rocircle potentiel des amibes comme reacuteservoir de M bovis nrsquoest
disponible Concernant la meacutesofaune les lombrics ont eacuteteacute identifieacutes comme vecteur potentiel dans la
transmission de M avium ssp paratuberculosis dans des eacutelevages bovins et caprins infecteacutes (Fischer
et al 2003) Veacuteritables ingeacutenieurs du sol ces animaux ont un rocircle important dans la circulation des
microorganismes en les transfeacuterant et les seacutelectionnant depuis la matiegravere organique qursquoils ingegraverent et
dont ils sont de gros consommateurs vers le sol qui les entoure par le biais de leurs turricules
Afin drsquoeacutevaluer le rocircle potentiel des lombrics dans la disseacutemination de M bovis dans le sol suite
agrave lrsquoingestion de matiegraveres feacutecales animales contamineacutees nous avons eacutetudieacute le transfert de M bovis BCG
depuis les fegraveces inoculeacutes vers le sol environnant par une espegravece de lombric aneacutecique Lumbricus
terrestris La population bacteacuterienne de M bovis a eacuteteacute deacutetecteacutee et quantifieacutee par qPCR dans le sol
environnant les turricules et le tube digestif des lombrics
Chapitre 5
157
B RAPID DISSEMINATION OF M BOVIS FROM CATTLE DUNG TO SOIL BY THE EARTHWORM LUMBRICUS TERRESTRIS
Elodie Barbierab Benoit Chantemesseab Murielle Rocheletab Leacuteon Fayolle a Loiumlc Bollachebc
Maria Laura Boschiroli d Alain Hartmann a
a INRA UMR 1347 Agroeacutecologie Dijon Cedex France
b Universiteacute de Bourgogne Franche Comteacute Dijon Cedex France
c Laboratoire Chrono-environnement UMR CNRS 6249 Besanccedilon France
d Universiteacute Paris-Est Laboratoire National de Reacutefeacuterence de la Tuberculose Uniteacute de Zoonoses Bacteacuteriennes Laboratoire de
Santeacute Animale ANSES Maisons-Alfort Cedex France
Key words Mycobacterium bovis spreading earthworms Lumbricus terrestris cattle soil quantitative
PCR
Running title Environmental dissemination of Mycobacterium bovis by earthworms
Corresponding Author Alain Hartmann UMR 1347 Agroeacutecologie INRA 17 rue Sully BP 86510 21065
Dijon Cedex France E-mail alainhartmanndijoninrafr
Chapitre 5
158
Abstract
Indirect transmission of the bovine tuberculosis (bTB) between wildlife and livestock is thought to occur
by inhalation or ingestion of environmental substrates contaminated through animal shedding The
role of the soil fauna such as earthworms in the circulation of Mycobacterium bovis from
contaminated animal feces is of interest in the epidemiology of bTB The objective of this study was to
assess the impact of earthworm activity on M bovis transfer from animal dung to castings and the
surrounding soil For this purpose microcosms of soil containing the anecic eathworms Lumbricus
terrestris were prepared and covered with cattle feces spiked with the M bovis BCG strain Pasteur to
carry out two separate experiments The dissemination the gut carriage and the excretion of M bovis
were all monitored using a specific qPCR-based assay Our results showed that the earthworm L
terrestris was able to rapidly disseminate M bovis from the contaminated cattle feces to the
surrounding soil through casting egestion Moreover contaminated earthworms were shown to shed
the bacteria for 4 days when transferred in a bTB-free soil This study highlights for the first time the
possible role of earthworms in the dissemination and the persistence of M bovis in soils in bTB endemic
areas
Chapitre 5
159
INTRODUCTION
Mycobacterium bovis is a pathogenic mycobacteria that can infect and cause bovine
tuberculosis (bTB) in cattle and in a wide range of domestic or wild mammals as well as in humans
(Biet et al 2005) bTB is a primarily respiratory disease but it may affect various organs in animals
(Morris et al 1994) Although cattle are considered as the principal host of M bovis wild fauna such
as badgers (Meles meles) wild boar (Sus scrofa) and red deer (Cervus elaphus) has also been identified
as potential wildlife reservoirs (Palmer 2013) Transmission of bTB between cattle mainly occurs by
inhalation of contaminated aerosols during close contacts with infected animals (Morris et al 1994
Phillips et al 2003) An inoculum of less than ten bacilli is enough to cause respiratory disease in cattle
(Dean et al 2005) Moreover indirect transmission by inhalation of environmental bioaerosols or
ingestion of infected soil and water could also be a potential way of animal contamination (Palmer et
al 2004 Vicente et al 2007)
Infected animals were shown to shed M bovis via oro-nasal mucus sputum urine feces and
wound discharges (Corner et al 2012 Palmer et al 2004) that may contaminate environment
especially pastures shared by cattle and wild animals Thus investigation of soil as a potential reservoir
of M bovis is an important step towards understanding the epidemiology of the bacteria To reach this
goal deciphering the ecological factors that govern the survival and the spread of M bovis is first
required
Among soil fauna earthworms constitute the largest animal biomass in most terrestrial
ecosystems especially in grasslands (Lavelle and Spain 2001) Earthworms play a role of engineers in
soil processes impacting on soil functions and ecosystem services (Blouin et al 2013) affecting soil
structure water movement organic matter decomposition nutrients releasing plant growth and
pollution remediation Earthworms are usually classified in three ecological types according to their
feeding habits i) the epigeic ones are restricted to the surface organic layers where they feed
themselves ii) the endogeic ones live in the subsurface where they burrow lateral tunnels to ingest soil
and organic matter and iii) the anecic ones feed preferentially on surface litter but live in burrows in
the soil (Lee 1985) Earthworms ingest soil particles mixed with organic matter that are mechanically
and chemically digested by both gut fluid and microbiota (Curry and Schmidt 2007) and release
castings (undigested soil egested on or under the soil surface) Earthworm gut microbiota acts as a
selective filter for soil microorganisms shapes microbial communities of soil and either up- or down-
regulates microbial populations (Byzov et al 2007 Egert et al 2004 Goacutemez-Brandoacuten et al 2011) It
has been shown that earthworms may have microbial beneficial activities especially in the transport of
bacterial inocula through soil (Thorpe et al 1996) but earthworms may also act as pests by promoting
the dissemination of plant pathogens such as Phytophtora cactorum and Fusarium reticulatum
Chapitre 5
160
(Montecchio et al 2015) More interestingly a few studies have described that human and animal
pathogenic bacteria viruses and parasites especially Escherichia coli O157H7 foot-and-mouth disease
virus and Cestoda tapeworms may be carried and spread by earthworms (Edwards and Bohlen 1996
Prysor Williams et al 2006)
Although animal dung is a very attractive food for earthworms (Lowe and Butt 2005) few
studies have evaluated the role of earthworms in the survival and the dissemination of potential
pathogenic mycobacteria from animal dung or manure to surrounding soil (Fischer et al 2003) In
addition since earthworms represent an important part of the diet of various animal species including
badgers and wild boar (Granval and Muys 1995 Lee 1985) the consumption of contaminated
earthworms (gut carriage) might be a source of contamination for wild fauna
With the aim to investigate the above hypothesis this current study was first devoted to assess
the impact of earthworm activity on M bovis transfer from animal dung to their released castings and
the surrounding soil Then the M bovis gut carriage and the casting contamination following ingestion
of infected feces were investigated
MATERIAL AND METHODS
Mycobacterium bovis strain preparation
Mycobacterium bovis BCG strain Pasteur 1173P2 was grown aerobically at 37degC in Middlebrook
7H9 broth supplemented with 10 Middlebrook ADC Growth Supplement (Sigma-Aldrich France) The
M bovis inoculum was prepared as following a 10 mL stationary phase culture was pelleted and once
the supernatant removed the cell aggregates were disrupted by vortexing with glass beads for 15
seconds The pellet was suspended in 1 mL of sterile water and serial 10-fold dilutions of the suspension
were spiked on modified Middlebrook 7H11 plates (Middlebrook 7H11 supplemented with 10 heat
inactivated bovine serum (Dominique Dutscher France) and 10 Middlebrook OADC Growth
Supplement (Sigma-Aldrich France) for enumeration The plates were always incubated at 37degC for 2
months A 10-fold dilution of the prepared M bovis suspension was also directly quantified with the
RD4 qPCR assay described in the section ldquoMycobacterium bovis quantitative PCR (qPCR)rdquo
Soil cattle feces and earthworms characteristics
A bTB-free pasture (that has never been grazed by bTB infected cattle) located in Burgundy
(Middle East of France) was selected to sample the soil used in this study This latter is a clayey loamy
soil with a pH of 775 Organic carbon and nitrogen contents were 361 and 38 gkg-1 respectively The
soil was sieved to 4 mm and stored at 4degC until its use
Fresh cattle feces were collected from a bTB-free Charolais cattle in a bTB-free area of Burgundy and
Chapitre 5
161
immediately used
The adult anecic earthworms (Lumbricus terrestris) were provided by LombrirsquoCarraz (France)
and kept at 4degC in large buckets containing soil and organic matter until starting the study Soil
earthworms and cattle feces used in this work were previously proved to be negative for the presence
of M bovis using the qPCR-based assay
DNA extraction and purification
DNAs from all the samples collected were extracted as described previously (Dequiedt et al
2011) Briefly 8 mL of lysis buffer (100 mM Tris pH 80 100 mM EDTA pH 80 100 mM NaCl and 2
(wv) SDS) 4 g of silica beads (100 μm diameter) 5 g of ceramic beads (14 mm diameter) and 8 glass
beads (4 mm diameter) were added to 2 g of sample in a 15 ml sterile vial - whereas 1 mL of lysis
buffer 05 g of silica beads 0625 g of ceramic beads 1 glass beads were added to 250 mg-samples
Samples were disrupted for 3 times 30 s at 4 ms in a FastPrepreg-24 Instrument (MP Biomedicals Europe
Illkirch France) incubated for 30 min at 70 degC and centrifuged for 5 min at 7000 timesg at room
temperature Supernatants were incubated on ice for 10 min with 110 volume of 3 M potassium
acetate pH 55 and then centrifuged for 5 min at 14000 times g One volume of ice-cold isopropanol was
added to the supernatant for DNA precipitation overnight at minus 20 degC DNA was collected by
centrifugation (30 min at 14000 times g) and DNA pellets were washed with ice-cold ethanol (70) and
dissolved in 100 μl of water Crude DNA extracts (100 μl) were loaded on Microbiospintrade columns
(Biorad Marnes-La-Coquette France) filled with 100 mg of polyvinyl polypyrrolidone (PVPP Sigma
Chemical Co France) After centrifugation (4 min 1000 times g 4 degC) eluted DNA was further purified
using the Geneclean Turbo Kit (Qiagen Courtaboeuf France) according to the manufacturers
instructions Purified DNA concentrations were estimated by spectrophotometry (λ = 260 nm) using a
NanoDropreg 2000 spectrophotometer (Thermo Scientific Illkirch France)
Mycobacterium bovis quantitative PCR (qPCR)
Mycobacterium bovis-specific qPCR was set up targeting the RD4 flanking region using
previously described forward RD4 flanking primer (5rsquo-TGTGAATTCATACAAGCCGTAGTCG-3rsquo) and probe
(5rsquo-6-FAM-AGCGCAACACTCTTGGAGTGGCCTAC-3rsquo TAMRA) (Sweeney et al 2007) In order to improve
qPCR efficiency the reverse RD4 flanking primer based assay was modified (5rsquo-
ATCTTAGCTGGTCAATAGCCATTTTT-3rsquo) to reduce the size of the qPCR product to 116 bp
qPCR reactions were carried out in triplicate with a 25 microL reaction mix containing 125 microL of
ABsolutetrade QPCR ROX Mix (Thermo Scientific France) 1 microL (final concentration 25 ngmicroL-1) of T4
bacteriophage gene 32 product (MP Biomedicals Europe France) 1 microL of each primer (final
concentration 400 nmolL-1) 05 microL of each probe (final concentration 200 nmol L-1) 4 microL of ultrapure
Chapitre 5
162
water and 5 microL of pure or diluted (10- or 20-fold) DNA Absolute quantification was achieved using
standard DNA dilutions A recombinant plasmid containing one copy of the RD4 fragment of M bovis
BCG strain Pasteur 1173P2 cloned in the pCRregII-TOPOreg vector (Invitrogen France) was used as a
standard A calibration curve was obtained by amplification of serial dilutions of the plasmid ranging
from 1 to 106 copies per PCR reaction All DNA samples extracted from the soil castings and feces
samples were screened with the RD4-based qPCR assay and results were expressed as the number of
M bovis gene copies per g of soil castings or feces We previously assessed that when Cycle threshold
(Ct) was above 39 detection was possible but unreliable (unpublished results) The word ldquotracesrdquo was
then chosen in all further experiments in order to indicate that M bovis targets are present in very low
quantities (below 250 CFUg-1 of soil) The qPCR assay was performed in a VIIA 7 Real-Time PCR System
(Thermo Fischer Scientific France) Initial denaturation was done at 95degC for 15 min followed by 45
cycles with denaturation at 95degC for 15 s annealing and elongation at 60degC for 1 min
Mesocosms preparation
Plastic jars (15 L) were filled with 1 kg of the clay-loamy soil Sterile water was added to each
soil sample to reach final soil moisture of 80 of the water-holding capacity (WHC) Once the soil was
packed down to obtain a 1 gcm-3 soil density 6 earthworms Lumbricus terrestris were placed into each
jar A 4-days delay was necessary to acclimatize the earthworms Nine control mesocosms where no
earthworms were added were processed and analysed
Evaluation of earthworm dissemination of M bovis to soil mesocosms
Aliquots of 50 g of cattle feces previously spiked with 1 mL of M bovis BCG suspension were
layered on the soil surface of each mesocosm Jars were covered with pierced fitted lids and kept in a
dark room at 20degC in a L2 confinement facility (day 0)
In order to sample fresh earthworm castings all the castings were removed 48 h before each
sampling date A sampling schedule ie 2 4 7 10 14 21 30 45 and 60 days was followed For each
sampling date 2 mesocosms with earthworms and one control mesocosm were analysed as following
First earthworm castings were collected at the surface of each mesocosm containing earthworms
pooled homogenized and then aliquoted in triplicate in 2 g-samples Secondly the cattle feces and
earthworms were removed out of the mesocosms and the remaining soil was mixed and aliquoted in
triplicate in 2 g-samples Finally the six earthworms contained in each mesocosm were rinsed twice
with sterile water dried with a paper towel and individually transferred into a 60 mL empty vessel
After a 48h incubation at 20degC in a dark room to release the gut content one 250 mg-aliquot of fecal
samples was sampled for each earthworm In the control mesocosm the cattle feces were removed
from the top of mesocosm the soil was then mixed sampled and analysed as described above All
Chapitre 5
163
samples were frozen at -20degC until their DNA extraction and RD4 qPCR analysis Gene copy numbers
found in each sample were box-cox transformed before analysis to normalize the data Generalized
Linear Models (GLMs) were used to analyze the effects of sampling date (number of days after the
M bovis spiked cattle feces was layered onto the soil) and treatment (feces castings and soil) on the
number of CFU Tests were performed using JMP 70 Software (SAS Institute Inc) Results were
considered to be significantly different for P values lt 005
Gut carriage and excretion of M bovis by earthworms
Five mesocosms each containing 6 earthworms were prepared as described in the section
ldquomesocosm preparationrdquo and were then covered with 50 g of cattle feces spiked with 1 mL of M bovis
BCG suspension After 7 days of incubation each earthworm was rinsed twice with sterile water dried
with a paper towel To estimate M bovis carriage on earthworm tegument 6 earthworms out of the
30 were deposited in an individual Petri plate containing modified Middlebrook 7H11 media
supplemented with antibiotics and antifungal (100 mgL-1Ticarcillin 10 mgL-1 Trimethoprim 200 000
UIL-1 Polymyxin B and 100 mgL-1 Cycloheximid Sigma-Aldrich France) After gently moving it on the
plate surface for few minutes the earthworms were individually transferred in a 200 mL vessel
containing 100 g of M bovis-free fresh soil as well as the 24 earthworms This step corresponded to
the starting point of the experiment (day 0)
At each sampling date ie day 1 2 3 4 and 5 the castings from series of 6 randomly selected
remaining vessels were collected pooled homogenized and one aliquot of 2 g was sampled Then the
earthworms from 1 series of 6 vessels were removed rinsed twice with sterile water dried with a paper
towel and individually transferred in a 60 mL empty vessel After 48h at 20degC in a dark room the
animals have released their gut contents and a 250 mg-aliquot of fecal sample was recovered from
each vessel containing earthworms All of the samples were stored at -20degC until further analysis
RESULTS
Earthworm dissemination of M bovis from cattle feces spiked with the mycobacteria to soil
mesocosms
For this purpose the protocol sketched in Figure 1 was carried out over a 60 days-period and
the results are summarized in Figure 2 Both the culture-based method and the RD4 qPCR assay were
used to estimate the concentration of the Mycobacterium bovis BCG suspension used to spike the cattle
feces Different values of 125 times 107 CFU per mL and of (65 plusmn 05) times 108 gene copies per mL were
determined with the culture and the molecular methods respectively However given that only the
RD4 qPCR assay was carried out to quantify M bovis throughout this experiment the M bovis genome
Chapitre 5
164
equivalent number contained in the 50 g of cattle feces was estimated to (65 plusmn 05) times 108 at the
beginning of the experiment (day 0 dashed line in Figure 2)
Figure 1 Schematic representation of the procedure followed for the evaluation of M bovis dissemination in
soil by earthworms
Overall M bovis was repeatedly detected in all of the analysed samples ie earthworm castings
fecal samples and soils from 2 days after depositing the cattle feces spiked with M bovis on the surface
of the mesocosms Gene copy number was significantly influenced by the sampling date (df= 8 sup2 =
14508 Plt00001) treatment (df= 2 sup2 = 22430 Plt00001) and interaction between sampling date
and treatment (df= 16 sup2 = 14464 Plt00001) However at the end of the experiment (day 60)
M bovis gene copy numbers were not statistically different (Average and P value) whatever the kind of
samples More precisely the earthworm fecal samples (open circle Figure 2) showed the highest
contamination (up to (57 plusmn 03) times 106 M bovis gene copies per g) at day 2 while the M bovis gene copy
numbers in the fecal samples decreased (20-fold factor) after day 4 to reach a final value of (25 plusmn 22)
times 105 gene copy number per g On the other hand the M bovis gene copy numbers per g of earthworm
castings were up to around 10-fold lower than those per g of earthworm fecal samples until day 45
after which similar M bovis gene concentrations were measured (full circle Figure 2) Although soil
samples from mesocosms also led to a M bovis positive signal from day 2 (44 plusmn 42) times 104 gene copy
number per g) it is worth noting that only 4 out of the 6 analyzed samples were positive at day 2 (full
square Figure 2) Afterwards (ie from day 4) all of the soil samples gave a positive signal for M bovis
with a strong increase (10-fold) of the M bovis gene copy number per g of soil between day 2 and day
(D0 + 2 4 7 10 14 21 30 45)
Mesocosm Deposition of M bovis spiked feces
D0
CASTING POOL
FECAL SAMPLE
48h00
Earthworm
Fresh soil
M bovis spiked cattle feces
SOIL
Sampling day
Chapitre 5
165
7 followed by minor fluctuations until day 60 For the control mesocosms (ie without any
earthworms) no M bovis signal were measured in the soil samples throughout the experiment until
day 60 where a positive signal value of (35 plusmn 19) times 104 M bovis per g of soil was obtained (cross symbol
on Figure 2) thus indicating a very slow spread of M bovis in the bulk soil in the absence of earthworms
Finally it should be noticed that the cattle feces remained undisturbed in the control mesocosms over
the course of the experiment whereas the earthworms totally ingested the spiked cattle feces in all of
the mesocosms containing these animals during the first week of the experiment
Figure 2 Concentration of M bovis DNA determined by RD4 qPCR in soil earthworm castings and fecal samples recovered at each sampling date Concentrations of M bovis DNA in control mesocosms are represented with crosses (times) The dashed line corresponds to the concentration of M bovis spiked in 50 g of cattle feces Error bars represent the standard deviation values of 3 replicates
3
4
5
6
7
8
0 5 10 15 20 25 30 35 40 45 50 55 60
Lo
g1
0(M
b
ovi
sg
ene
cop
y n
um
ber
) in
1 g
of
sam
ple
Days
Chapitre 5
166
Table 1 RD4-based qPCR detection of M bovis gene copy number contained in the earthworm castings recovered according to the protocol of Figure 2
Sampling day Number of analysed
casting poolsa Number of positive
casting poolsb M bovis gene copy number per gc
1 5 5 (196 plusmn 136) times 105
2 4 4 (104 plusmn 060) times 105
3 3 2 Tracesd
4 2 1 Traces
5 1 0 Negative a
From 6 vessels b
Threshold cycle (Ct) between 20 and 39 c Mean value calculated with the signal obtained for the positive earthworm casting pools
d Detected but not quantifiable (below 250 CFUg-1 of soil)
Evaluation of the carriage and the excretion of M bovis by earthworms
In order to check whether earthworms carry M bovis in their gut and are able to excrete the
bacteria the following experiment was realized (see detailed protocol in Figure 3) Earthworms were
artificially contaminated by feeding for 7 days from cattle feces spiked with M bovis then their castings
and their fecal samples were analysed for a 5-days period First no growth of M bovis was observed
from earthworm tegument surfaces suggesting the absence or the low carriage of M bovis by
earthworm teguments The results shown in Table 1 indicated that all of the earthworms produced
castings highly contaminated with M bovis 1 and 2 days after their introduction in the fresh M bovis
free soil (up to around 105 M bovis gene copy number per g) Then the percentage of positive M bovis
casting pools gradually decreased (66 and 50 for sampling day 3 and 4 respectively) until the
sampling at day 5 for which no M bovis molecular signal was detected Although positive signals were
obtained for the sampling days 3 and 4 only traces of M bovis were detected (Ct above 39) With
values ranging from 1 to 5 105 M bovis gene copy number per g of fecal sample (Table 2) the RD4
qPCR assay yielded M bovis positive signals for the sampling days 1 to 4 Nevertheless for each of
these sampling days some earthworms (17-50 ) led to negative M bovis qPCR signal thus indicating
the absence or a very low amount of M bovis in the earthworm feces (below the detection limit of the
qPCR) Finally all of the six earthworm fecal samples analysed at day 5 were M bovis free
Chapitre 5
167
Figure 3 Schematic representation of the procedure followed for the estimation of M bovis gut carriage and excretion by earthworms
Table 2 RD4-based qPCR detection of M bovis gene copy number contained in the earthworm fecal samples recovered according to the protocol of Figure 3
Sampling day Number of positive
fecal samplesa (n = 6) M bovis gene copy number per gb
1 5 (123 plusmn 082) times 105
2 4 (228 plusmn 214) times 105
3 3 (542 plusmn 049) times 105
4 4 (213 plusmn 164) times 105
5 0 Negative
a Threshold cycle (Ct) between 20 and 39 b Mean value calculated with the signal obtained for the positive earthworm fecal samples
Sampling day (D0 + 1 2 3 4 5)Mesocosm
(n = 5)
Deposition of M bovisspiked feces (n = 5 )
Investigation of M bovis tegumentcarriage on 6 earthworms
D0
7 days
Transfer of each earthworm
5 series of 6 vessels(n = 30)
FECAL SAMPLE
CASTING POOL
n = 6
48h00
n = number of remainingseries of 6 vessels
Earthworm
Fresh soil
M bovis spiked cattle feces
Chapitre 5
168
DISCUSSION
The results shown in Figure 1 evidenced for the first time that earthworms were able to widely
transfer M bovis from cattle feces to their castings after ingestion of cattle feces spiked with the
bacteria Owing to the concomitant ingestion of M bovis-free soil and dung remains and thus the
dilution of M bovis cells in the gut content the number of M bovis cells in castings gradually decreased
from day 4 until the end of the experiment These results were in good agreement with earlier reported
laboratory trials with Mycobacterium avium subsp paratuberculosis that is responsible for the Johnersquos
disease in cattle and goat (Fischer et al 2003) Prysor Williams et al (Prysor Williams et al 2006) have
also reported the excretion of the human food-borne pathogen Escherichia coli O157H7 in castings
after ingestion of spiked manure by earthworms
On the other hand this study demonstrated that when earthworms were maintained in the
M bovis infected environment they were able to shed M bovis in their castings for at least 60 days
(Tableau 1) However when transferred in a M bovis-free soil earthworms only shed the bacteria up
to 4 days This data suggests that M bovis is unable to colonize the gut of the earthworm L terrestris
over extended periods Moreover L terrestris may ingest 490 to 3500 mg dry mass g-1 fresh mass of
arable soil per day (Curry and Schmidt 2007) and has a mean gut transit time of about 12 hours (Taylor
and Taylor 2014) leading to the rapid egestion of the bacteria from earthworm guts through castings
Furthermore earthworms were able to transfer M bovis from cattle feces to the bulk soil by
releasing castings in the tunnels they burrow Our results showed that M bovis was still detected in
mixed soil over the course of the experiment (60 days) highlighting a long persistence of M bovis in
soil Rapid dispersal of the bacteria in soil can be fully imputed to earthworm activity since soil from
control mesocosms without earthworms remained free from M bovis-except at day 60 Moreover
bacteria movement was mainly due to M bovis earthworm excretion since no M bovis tegument
carriage was observed The M bovis signal observed in control mesocosms at day 60 is probably due
to the moving of M bovis cells during moisture transfer from dung to soil surface and to the presence
of remaining dung particles in soil (incomplete removal of contaminated dung before soil mixing) As
M bovis is sensitive to solar radiation (UV) and soil dryness the burying of M bovis in deep soil by
earthworms may contribute to enhance its survival in soil environment Indeed survival of M bovis in
soil may be enhanced up to 2 years when the bacteria is buried to a depth of 5 cm (Genov 1965) Our
experimental design based on 1 kg-soil mesocosms did not allow estimating the real spatial distribution
of the bacilli in soil by earthworms It has been shown that earthworms are potentially able to move
several meters apart from the area where they graze organic matter (Mather and Christensen 1988)
thus disseminating soil-borne plan pathogen through large surfaces of soil (Thorpe et al 1996)
Concerning potential spread of animal or human pathogens earthworms were demonstrated in
Chapitre 5
169
experimental boxes to significantly aid vertical and lateral movement of ingested bacteria both up to
40 cm from the inoculation point (Prysor Williams et al 2006)
Finally we showed that earthworms may carry M bovis for many days in their guts even when
transferred in a M bovis-free environment Earthworms are known to host animal parasites and
pathogen such as protozoan tapeworms (Cestoda) and nematodes they can transmit to birds and
mammals when earthworms are ingested (Edwards and Bohlen 1996) However to our knowledge
the fact that earthworms may act as a source of contamination for animals feeding from earthworms
has never been studied Earthworms that constitute a nutritious food resource are an important part
of the diet of badgers and wild boar (Baubet et al 2003 Cleary et al 2011) that often forage in cattle
pastures hosting high earthworm populations (Muldowney et al 2003) Badgers and wild boar are
able to eat up to 200 and 120 earthworms respectively in a night especially during worm nights when
humid and mild weather conditions are optimal for earthworms to ascend to the surface (Baubet et
al 2003 Kruuk and Parish 1981) In endemic area of bTB where fecal M bovis shedding may occur in
infected animals it would be interesting to evaluate M bovis carriage in earthworm gut in the vicinity
of infected animal dung Further studies are needed to assess the role of earthworms harbouring
M bovis gut carriage in the contamination of bTB sensible earthworm feeders
One field study reports the isolation of non-tuberculous mycobacteria such as Mycobacterium
avium complex members (M avium ssp paratuberculosis and M avium ssp hominisuis) as well as M
abcessus M scrofulaceum and M gastri) from 9 out of 109 earthworm feces in sites where cattle
goats and swine were severely infected with the Johnersquos disease or mycobacteriosis (Fischer et al
2003) However this study failed to isolate M avium ssp avium from earthworms feces in aviaries
infected with avian tuberculosis
Cattle feces were spiked with M bovis at a final concentration of 125 times 107 CFU per 50 g of
feces ie 25 times 105 CFU per g (estimated by culture-based enumeration of the inoculum) M bovis
concentration in spiked cattle feces was also measured by the qPCR-based method We observed
numbers of M bovis 52-fold higher when using the molecular detection method than with the culture-
based method This discrepancy is probably due first to the strong aggregation of mycobacteria cells in
the inoculum despite physical disruption of cell aggregates and second to the presence of dead or
viable but non-cultivable cells of M bovis non detected by culture-based method The amount of
M bovis spiked in cattle feces is equivalent or lower than the number of M bovis cells recorded per g
of urine or feces shed by infected badgers (Gallagher and Clifton-Hadley 2000 Sweeney et al 2007)
leading to the presence of millions of bacteria in a latrine (burrowed hole in the soil where badgers
defecate and urinate) But it is probably larger than M bovis cells deposited in infected cattle feces
(Neill et al 1988) For practical reasons we used cattle feces in our study but it could be interesting to
compare if dung from other animals than cattle would be as attractive for earthworms
Chapitre 5
170
Our study was based on a qPCR assay method detecting M bovis DNA in soil castings and feces
samples This molecular tool does not give any information on the viability of the M bovis cells egested
by the earthworms However as mycobacteria harbour a resistant waxy cell envelope (Brennan and
Nikaido 1995) and as Mycobacterium avium ssp paratuberculosis and other environmental
mycobacteria have been showed to largely survive through the gut passage (Fischer et al 2003) these
data strongly suggest that M bovis could resist through the passage of earthworm guts and remain
alive in castings Further research is needed to clarify if tuberculous mycobacteria such as M bovis are
digested or survive during transit through the earthworm gut and if their virulence is conserved
Evidencing M bovis presence with culture-based method in environmental matrices such as soil
castings or animal feces is still a challenge As M bovis is a slow growing mycobacteria environmental
samples have to be strongly decontaminated prior to culturing to overcome contamination with fast
growing microorganisms leading to a decrease in the sensitivity of the method
Surface and subsurface casting shedding of M bovis is all the more relevant because this
pathogen bacteria may survive in soil and feces for long periods (Young et al 2005) Castings are then
within the reach of animals especially cattle that may be infected by inhaling soil aerosols or by
ingesting contaminated casting soil during their exploratory foraging or feeding activities While
Mycobacterium bovis infective dose by the respiratory route is very low since only ten viable bacilli are
able to cause respiratory bovine tuberculosis (Dean et al 2005) 5000 bacilli would be required to
initiate infection through the oral route in cattle (Palmer et al 2004) As cattle usually avoid grazing
close to the feces of other cattle or wild animals (Hutchings and Harris 1997) further dissemination of
M bovis by earthworms away from infected feces might induce contamination risk for cattle grazing in
this area
Further work is still needed to study the occurrence of M bovis in earthworms under natural
field conditions in bTB-contaminated area Cultivation of environmental M bovis is also required to
characterize properly these environmental strains Finally virulence of M bovis contained in
earthworm castings remains to be deciphered
Funding information
The work was supported by the Regional Council of Burgundy (Elodie Barbier PhD fellowship) the Cocircte
drsquoOr General Council Interbev and the Cocircte drsquoOr GDS (Groupement de Deacutefense Sanitaire Livestock
health protection group) The funders had no role in study design data collection and interpretation
or the decision to submit the work for publication
Conflict of interest
The authors have no conflict of interest to declare
Chapitre 5
171
Acknowledgements
The authors would like to thank AE Taconet F Bartheacuteleacutemy and T Grosjean from the Cocircte drsquoOr Regional
Veterinary Service (DDPP21) F Chevalier and M Beacuteral from the CIREV (Cellule Inter-Reacutegionale
drsquoEpideacutemiologie Veacuteteacuterinaire)
References
Baubet E Ropert-Coudert Y Brandt S 2003 Seasonal and annual variations in earthworm consumption by wild boar (Sus scrofa scrofa L) Wildl Res 30 179 doi101071WR00113
Biet F Boschiroli ML Thorel MF Guilloteau LA 2005 Zoonotic aspects of Mycobacterium bovis and Mycobacterium avium-intracellulare complex (MAC) Vet Res doi101051vetres
Blouin M Hodson ME Delgado E a Baker G Brussaard L Butt KR Dai J Dendooven L Peres G Tondoh JE Cluzeau D Brun JJ 2013 A review of earthworm impact on soil function and ecosystem services Eur J Soil Sci 64 161ndash182 doi101111ejss12025
Brennan PJ Nikaido H 1995 The envelope of mycobacteria Annu Rev Biochem 64 29ndash63 doi101146annurevbiochem64129
Byzov BA Khomyakov N V Kharin SA Kurakov A V 2007 Fate of soil bacteria and fungi in the gut of earthworms Eur J Soil Biol 43 149ndash156 doi101016jejsobi200708012
Cleary GP Corner LAL OrsquoKeeffe J Marples NM 2011 Diet of the European badger (Meles meles) in the Republic of Ireland A comparison of results from an analysis of stomach contents and rectal faeces Mamm Biol 76 470ndash475 doi101016jmambio201010012
Corner LAL OrsquoMeara D Costello E Lesellier S Gormley E 2012 The distribution of Mycobacterium bovis infection in naturally infected badgers Vet J 194 166ndash72 doi101016jtvjl201203013
Curry JP Schmidt O 2007 The feeding ecology of earthworms - A review Pedobiologia (Jena) 50 463ndash477 doi101016jpedobi200609001
Dean GS Rhodes SG Coad M Adam O Cockle PJ Clifford DJ Hewinson RG Vordermeier HM 2005 Minimum infective dose of Mycobacterium bovis in cattle Infect Immun 73 6467ndash6471 doi101128IAI73106467
Edwards CA Bohlen PJ 1996 Biology and ecology of earthworms Chapman and Hall London
Egert M Marhan S Wagner B Scheu S Friedrich MW 2004 Molecular profiling of 16S rRNA genes reveals diet-related differences of microbial communities in soil gut and casts of Lumbricus terrestris L (Oligochaeta Lumbricidae) FEMS Microbiol Ecol 48 187ndash197 doi101016jfemsec200401007
Fischer OA Matlova L Bartl J Dvorska L Svastova P Du Maine R Melicharek I Bartos M Pavlik I 2003 Earthworms (Oligochaeta Lumbricidae) and mycobacteria Vet Microbiol 91 325ndash338 doi101016S0378-1135(02)00302-4
Chapitre 5
172
Gallagher J Clifton-Hadley RS 2000 Tuberculosis in badgers a review of the disease and its significance for other animals Res Vet Sci 69 203ndash17 doi101053rvsc20000422
Genov I 1965 The effects of certain physical and chemical agents on Mycobacterium tuberculosis Veteriaacuternı Med Nauk Sofia 2 97ndash107
Goacutemez-Brandoacuten M Aira M Lores M Domiacutenguez J 2011 Epigeic Earthworms exert a bottleneck effect on microbial communities through gut associated processes PLoS One 6 doi101371journalpone0024786
Granval P Muys B 1995 Predation on earthworms by terrestrial vertebrates in XII Congress ldquoThe Game and the Manrdquo Sofia Bulgaria p 480
Hutchings MR Harris S 1997 Effects of farm management practices on cattle grazing behaviour and the potential for transmission of bovine tuberculosis from badgers to cattle Vet J 153 149ndash62
Kruuk H Parish T 1981 Feeding specialization of the European badger Meles meles in Scotland UK J Anim Ecol 50 773ndash788 doi1023074136
Lavelle P Spain A 2001 Soil ecology Kluwer Academic Publishers Dordrecht
Lee KE 1985 Earthworms their ecology and relationship with soils and land use Academic P ed
Lowe CN Butt KR 2005 Culture techniques for soil dwelling earthworms A review Pedobiologia (Jena) 49 401ndash413 doi101016jpedobi200504005
Mather JG Christensen O 1988 Surface movements of earthworms in agricultural land Pedobiologia (Jena) 32 399ndash405
Montecchio L Scattolin L Squartini A Butt K 2015 Potential spread of forest soil-borne fungi through earthworm consumption and casting iForest 8 295ndash301 doi103832ifor1217-008
Morris RS Pfeiffer DU Jackson R 1994 The epidemiology of Mycobacterium bovis infections Vet Microbiol 40 153ndash77
Muldowney J Curry J Okeeffe J Schmidt O 2003 Relationships between earthworm populations grassland management and badger densities in County Kilkenny Ireland Pedobiologia (Jena) 47 913ndash919 doi101016S0031-4056(04)70289-9
Neill SD Hanna J OrsquoBrien J McCracken R 1988 Excretion of Mycobacterium bovis by experimentally infected cattle Vet Rec 123 340ndash343 doi101136vr12313340
Palmer M V 2013 Mycobacterium bovis Characteristics of wildlife reservoir hosts Transbound Emerg Dis 60 1ndash13 doi101111tbed12115
Palmer M V Waters WR Whipple DL 2004 Investigation of the transmission of Mycobacterium bovis from deer to cattle through indirect contact Am J Vet Res 65 1483ndash1489 doi102460ajvr2004651483
Phillips CJC Foster CRW Morris PA Teverson R 2003 The transmission of Mycobacterium bovis infection to cattle Res Vet Sci 74 1ndash15
Chapitre 5
173
Prysor Williams A Roberts P Avery LM Killham K Jones DL 2006 Earthworms as vectors of Escherichia coli O157H7 in soil and vermicomposts FEMS Microbiol Ecol 58 54ndash64 doi101111j1574-6941200600142x
Sweeney FP Courtenay O Hibberd V Hewinson RG Reilly L a Gaze WH Wellington EMH 2007 Environmental monitoring of Mycobacterium bovis in badger feces and badger sett soil by real-time PCR as confirmed by immunofluorescence immunocapture and cultivation Appl Environ Microbiol 73 7471ndash3 doi101128AEM00978-07
Taylor AR Taylor AFS 2014 Assessing daily egestion rates in earthworms using fungal spores as a natural soil marker to estimate gut transit time Biol Fertil Soils 50 179ndash183 doi101007s00374-013-0823-5
Thorpe IS Prosser JI Glover L a Killham K 1996 The role of the earthworm Lumbricus terrestris in the transport of bacterial inocula through soil Biol Fertil Soils 23 132ndash139
Vicente J Houmlfle U Garrido JM Fernaacutedez-De-Mera IG Acevedo P Juste R Barral M Gortazar C 2007 Risk factors associated with the prevalence of tuberculosis-like lesions in fenced wild boar and red deer in south central Spain Vet Res 38 451ndash464 doi101051vetres2007002
Young JS Gormley E Wellington EMH 2005 Molecular detection of Mycobacterium bovis and Mycobacterium bovis BCG (Pasteur) in soil Appl Environ Microbiol 71 1946ndash1952 doi101128AEM7141946
Chapitre 5
174
C COMPLEMENT PRELEVEMENT ET ANALYSE DE LOMBRICS DANS UNE PATURE SUSPECTE
Afin drsquoeacutevaluer la contamination naturelle eacuteventuelle des lombrics par M bovis nous avons
analyseacute 90 lombrics preacuteleveacutes en feacutevrier 2014 agrave proximiteacute drsquoune source permanente drsquoune des pacirctures
suspectes de lrsquoAuxois ougrave des bovins infecteacutes ont pacirctureacute lrsquoeacuteteacute preacuteceacutedent (Site C de lrsquoeacutetude
environnementale Chapitre 2) Les lombrics ont eacuteteacute preacuteleveacutes le mecircme jour que les trois eacutechantillons
positifs de ce site (eau seacutediments neacutematodes) Sur sept points de la pacircture choisis au hasard une
pelleteacutee drsquoenviron 20 kg de sol a eacuteteacute retourneacutee Les vers ont eacuteteacute seacutepareacutes du sol rinceacutes avec de lrsquoeau et
transfeacutereacutes individuellement dans des pots en plastique de 60 mL Les pots ont eacuteteacute stockeacutes 48 h agrave 20degC
dans lrsquoobscuriteacute afin de laisser les vers se vider Pour chaque point de la pacircture preacuteleveacute les fegraveces des
vers ont eacuteteacute pooleacutees et homogeacuteneacuteiseacutees puis 2 g ont eacuteteacute preacuteleveacutes et congeleacutes jusqursquoagrave la reacutealisation de
lrsquoanalyse Apregraves extraction de lrsquoADN chaque eacutechantillon a eacuteteacute analyseacute en qPCR avec le systegraveme de
deacutetection RD4 Les protocoles utiliseacutes sont deacutecrits dans le Mateacuteriel et Meacutethodes de lrsquoarticle preacuteceacutedent
Lrsquoidentification pheacutenotypique des 90 vers collecteacutes dans la pacircture a permis de mettre en
eacutevidence cinq genres de lombric diffeacuterents Lumbricus terrestris Lumbricus castaneus Nicodrilus sp
Octolosium cyaneum Allolobophora sp Aporrectodea caliginosa
Aucun des sept pools de fegraveces analyseacutes nrsquoa donneacute de signal positif en qPCR avec le systegraveme RD4
Chapitre 5
175
D BILAN
Les lombrics sont de gros consommateurs de matiegravere organique naturellement chargeacutee drsquoune
flore abondante et varieacutee et potentiellement drsquoune flore bacteacuterienne pathogegravene Toutefois il existe
tregraves peu de donneacutees sur la disseacutemination des bacteacuteries pathogegravenes pour lrsquohomme et les animaux par
les lombrics dans le sol ainsi que sur les risques lieacutes au portage intestinal de bacteacuteries pathogegravenes par
les lombrics pour les animaux qui les consomment
Lrsquoeacutetude deacutecrite dans lrsquoarticle preacuteceacutedent montre qursquoapregraves avoir ingeacutereacute des matiegraveres feacutecales
fraiches inoculeacutees les lombrics ont la capaciteacute de disseacuteminer rapidement M bovis dans le sol
environnant par le biais des turricules qursquoils eacutemettent agrave la surface du sol et en profondeur dans les
tunnels qursquoils creusent Lrsquoenfouissement de la bacteacuterie dans le sol permet probablement drsquoameacuteliorer la
persistance de la bacteacuterie en la proteacutegeant des UV et de la deacuteshydratation Drsquoautre part le transfert
de la bacteacuterie agrave quelques cm des deacutejections animales contamineacutees par le biais des turricules de surface
la rend plus accessible aux animaux (qui en geacuteneacuteral eacutevitent les fegraveces des autres animaux) Ainsi en
fonction des comportements exploratoires ou alimentaires des diffeacuterentes espegraveces freacutequentant les
pacirctures il existe un risque potentiel de contamination par du sol ou des turricules contamineacutes
(inhalation de bioaeacuterosols ou ingestion)
Nous avons eacutegalement montreacute que le tube digestif des lombrics ne constituait pas un reacuteservoir
peacuterenne de M bovis la bacteacuterie nrsquoeacutetait deacutetecteacutee que transitoirement (4 jours) apregraves avoir remis les
lombrics dans un sol sain Le portage digestif pendant les 4 premiers jours constitue toutefois un risque
pour les animaux qui consomment les lombrics en grande quantiteacute notamment les blaireaux et les
sangliers
Cette eacutetude soulegraveve les mecircmes inconnues que les eacutetudes preacuteceacutedentes principalement sur la
viabiliteacute lrsquoeacutetat physiologique et la conservation de la virulence de M bovis apregraves un transfert dans le
sol ou suite au passage de la bacteacuterie dans le tube digestif des lombrics Ces facteurs sont
indispensables pour permettre lrsquoinfection drsquoanimaux via des matrices environnementales contamineacutees
Les moyens drsquoeacutevaluation de ces diffeacuterents facteurs seront discuteacutes dans la partie Perspectives du
manuscrit
Chapitre 5
176
Discussion geacuteneacuterale
Discussion geacuteneacuterale
179
Depuis 2004 face agrave la recrudescence de la tuberculose bovine en Cocircte drsquoOr la mise en place
drsquoun programme de lutte sanitaire reacuteguliegraverement ameacutelioreacute contre la maladie a permis de controcircler et
drsquoameacuteliorer la situation dans lrsquoeacutelevage bovin depuis 2010 Dans la zone endeacutemique du deacutepartement la
circulation de Mycobacterium bovis dans un systegraveme multi-hocircte speacutecifiques impliquant les bovins et
des espegraveces sauvages complique fortement la lutte contre la maladie notamment par la difficulteacute agrave
geacuterer lrsquoinfection dans la faune sauvage malgreacute les mesures de controcircle entreprises jusqursquoagrave maintenant
La transmission indirecte de la maladie entre animaux sauvages et bovins par le biais de substrats
environnementaux contamineacutes est fortement suspecteacutee dans les zones ougrave bovins et faune sauvage
infecteacutes partagent les mecircmes territoires et ressources La transmission environnementale a deacutejagrave eacuteteacute
deacutemontreacutee en conditions expeacuterimentales (Little et al 1982 Palmer et al 2004) mais elle est tregraves
difficile agrave deacutemontrer en conditions naturelles et passe notamment par lrsquoeacutevaluation du niveau
drsquoexcreacutetion par les animaux infecteacutes du rocircle de lrsquoenvironnement comme reacuteservoir de la bacteacuterie de sa
survie dans un eacutetat viable et du maintien de sa virulence
Dans ce contexte les objectifs de ce travail eacutetaient drsquoune part drsquoeacutevaluer la contamination
environnementale en Cocircte drsquoOr sur des agroeacutecosystegravemes freacutequenteacutes par des bovins etou des animaux
sauvages potentiellement infecteacutes dans le but de mettre en eacutevidence des reacuteservoirs
environnementaux eacuteventuels Drsquoautre part nous avons chercheacute agrave preacuteciser les facteurs impliqueacutes dans
la persistance de M bovis dans le sol et sa disseacutemination par la micro- et meacutesofaune du sol
1 Des meacutethodes deacuteveloppeacutees (ou adapteacutees) puis valideacutees dans des matrices environnementales
Notre objectif initial pour lrsquoanalyse des eacutechantillons environnementaux eacutetait drsquoutiliser en
parallegravele la PCR quantitative apregraves extractionpurification de lrsquoADN pour la deacutetection et la quantification
de M bovis et une meacutethode baseacutee sur la culture afin drsquoisoler les souches environnementales pour les
caracteacuteriser Cependant les protocoles de preacutetraitement et de culture tireacutes de la litteacuterature
(immunocapture et protocoles de deacutecontamination) que nous avons utiliseacutes en premiegravere intention sur
les eacutechantillons environnementaux nrsquoont pas donneacute les reacutesultats escompteacutes du fait de la croissance tregraves
lente de M bovis et de la preacutesence dans les eacutechantillons de tregraves nombreux microorganismes
interfeacuterents agrave croissance rapide qui ont gecircneacute la culture (envahissement rapide des boicirctes en une
semaine) La culture de M bovis agrave partir drsquoeacutechantillons environnementaux reste agrave lrsquoheure actuelle un
challenge (Fine OrsquoBrien et al 2011 Fine Bolin et al 2011 Young et al 2005 King et al 2015)
notamment agrave cause de lrsquoabsence de milieux suffisamment seacutelectifs et de bouillons drsquoenrichissement
Discussion geacuteneacuterale
180
adapteacutes comme il en existe notamment pour lrsquoisolement de bacteacuteries pathogegravenes telluriques telles
que Listeria monocytogenes ou Pseudomonas aeruginosa En lrsquoabsence de meacutethodes disponibles nous
avons donc mis au point deux protocoles tregraves innovants agrave la fin de nos travaux un protocole
drsquoextractiondeacutecontamination des mycobacteacuteries dans le sol et une meacutethode de deacutetection des
mycobacteacuteries en culture liquide Ces deux meacutethodes sont en cours drsquoeacutevaluation par la SATT (Socieacuteteacute
drsquoAcceacuteleacuteration de Transfert de Technologies) en vue drsquoune valorisation sous forme de savoir-faire ou de
brevet Des eacutetudes compleacutementaires seront neacutecessaires pour deacutevelopper ces outils innovants
La recherche de reacuteservoirs environnementaux potentiels de M bovis en Cocircte drsquoOr a donc eacuteteacute
baseacutee sur lrsquoutilisation de meacutethodes moleacuteculaires adapteacutees aux matrices environnementales
Contrairement aux meacutethodes culturales la qPCR permet une deacutetection et une quantification rapide
des ADN cibles sans ecirctre gecircneacutee par la preacutesence de contaminants Face au grand nombre de
mycobacteacuteries potentiellement preacutesentes dans les sols de Cocircte drsquoOr dont M microti (Michelet et al
2015) nous avons choisi drsquoutiliser trois systegravemes ciblant trois seacutequences diffeacuterentes dans le geacutenome
des espegraveces du MTBC ou de M bovis que nous avons deacuteveloppeacutes et valideacutes pour leur speacutecificiteacute et leur
sensibiliteacute Lrsquoutilisation conjointe de ces trois systegravemes nous a permis drsquoaugmenter la speacutecificiteacute de la
deacutetection Toutefois le faible niveau de deacutetection observeacute dans certains eacutechantillons ne nous a pas
permis de quantifier le MTBC ou M bovis dans tous les eacutechantillons
2 Contamination environnementale les zones humides les eaux de ruissellement et les sols de terriers sont potentiellement agrave risque
Les systegravemes moleacuteculaires deacuteveloppeacutes ont eacuteteacute utiliseacutes pour lrsquoanalyse de plus de 400
eacutechantillons environnementaux varieacutes preacuteleveacutes dans les agroeacutecosystegravemes de la zone endeacutemique de
Cocircte drsquoOr et de Charente La contamination environnementale a eacuteteacute deacutetecteacutee de faccedilon reacutepeacuteteacutee dans
deux types drsquoeacutecosystegravemes bien particuliers les zones humides preacutesentes sur les pacirctures (sources et
laquo mouilles raquo) les terriers et latrines de blaireaux
Concernant les zones humides nous avons mis en eacutevidence une contamination dans lrsquoeau
circulante et les seacutediments de deux sources naturelles et permanentes situeacutees sur des pacirctures utiliseacutees
par deux cheptels diffeacuterents dans deux communes diffeacuterentes Cette faible contamination de lrsquoeau
etou des seacutediments retrouveacutee agrave lrsquoendroit de la reacutesurgence ainsi que sur diffeacuterents points en aval est
une donneacutee nouvelle puisqursquoagrave ce jour M bovis nrsquoavait eacuteteacute isoleacutee que dans lrsquoeau stagnante et ses
seacutediments (Santos et al 2015) Lors de nos campagnes de preacutelegravevements nous avons constateacute que ces
reacutesurgences parfois capteacutees dans des abreuvoirs eacutetaient parfois le seul moyen drsquoabreuvement des
bovins dans les pacirctures Ces zones humides semblent tregraves attractives pour les bovins au vu des indices
de preacutesence constateacutes (matiegraveres feacutecales et nombreuses empreintes) La persistance de M bovis eacutetant
Discussion geacuteneacuterale
181
favoriseacutee dans les matrices humides riches en matiegravere organique et dans lrsquoeau (Young et al 2005 Fine
Bolin et al 2011 Duffield amp Young 1985 Wint et al 2002) ces zones constituent un milieu favorable
agrave la survie de la bacteacuterie Dans notre eacutetude la mise en eacutevidence de la contamination des zones humides
dans des pacirctures suspectes corrobore le reacutesultat de lrsquoeacutetude PatRisk dans laquelle une association
significative a eacuteteacute eacutetablie entre la preacutesence de mouilles et le statut laquo agrave risque drsquoune pacircture raquo (Bouchez-
Zacria 2015)
Lrsquoorigine de la contamination de ces sources et laquo mouilles raquo est inconnue toutefois elles sont
accessibles aux bovins et animaux sauvages infecteacutes qui peuvent potentiellement excreacuteter la bacteacuterie
lors de lrsquoabreuvement ou du fouissement Vu le contexte geacuteomorphologique il est eacutegalement
envisageable que les infiltrations drsquoeau dans les plateaux calcaires coiffant les valleacutees entrainent avec
elles des matrices environnementales fortement contamineacutees (latrines sols) qui ressortent au niveau
des pacirctures avec toutefois un effet de dilution tregraves important
Des mesures de bioseacutecuriteacute visant agrave limiter les risques lieacutes aux zones humides sont deacutejagrave
preacuteconiseacutees par la DDPP 21 et le GDS 21 agrave lrsquoeacutechelle de la pacircture Lrsquoameacutenagement drsquoabreuvoirs peu
accessibles aux animaux sauvages et la condamnation par des clocirctures de lrsquoaccegraves des bovins aux
laquo mouilles raquo propices agrave la survie de la bacteacuterie sont fortement conseilleacutes aux eacuteleveurs des zones agrave
risque pour limiter le risque de transmission indirecte de la maladie Le drainage des pacirctures est une
solution envisageacutee pour asseacutecher les zones humides et canaliser les sources dans les pacirctures agrave risque
mais il est strictement reacuteglementeacute et soumis agrave autorisation avec enquecircte publique sous le controcircle de
la Police de lrsquoeau (Direction Deacutepartementale des Territoires) La correacutelation de nos reacutesultats sur la
contamination des zones humides avec le bilan des foyers bovins et des cas sauvages ainsi que les
reacutesultats de lrsquoeacutetude PatRisk ont initieacute une reacuteflexion entre la DDPP21 la CIREV le GDS et le SIRTAVA
(Syndicat Intercommunal pour la Reacutealisation des Travaux drsquoAmeacutenagement de la Valleacutee de lrsquoArmanccedilon)
pour eacutevaluer la faisabiliteacute de travaux de drainage dans les pacirctures agrave risque dans le respect de la
regraveglementation Un problegraveme se pose toutefois pour les pacirctures preacutesentant de tregraves nombreuses
reacutesurgences temporaires sur lesquelles le drainage nrsquoest pas envisageable Lrsquoutilisation de ces pacirctures
pour drsquoautres productions animales (ovins chevaux) ou veacutegeacutetales (production de foin la mise en
culture nrsquoest pas toujours possible agrave cause des pentes) est une solution
Notre eacutetude a eacutegalement mis en eacutevidence la contamination des sols de terriers de blaireaux
par M bovis Le geacutenotype retrouveacute dans les sols de terriers est identique agrave celui retrouveacute chez les
blaireaux infecteacutes pieacutegeacutes agrave proximiteacute confirmant que cette espegravece a la capaciteacute de contaminer son
milieu de vie (Young et al 2005 Courtenay et al 2006 Sweeney et al 2006 Sweeney et al 2007) Dans
cette espegravece des animaux excreacuteteurs et super excreacuteteurs ont eacuteteacute mis en eacutevidence en Cocircte drsquoOr (Payne
2014) expliquant les quantiteacutes importantes de bacilles retrouveacutees dans les sols de terrier (Young et al
Discussion geacuteneacuterale
182
2005 Courtenay et al 2006) et les latrines dans notre eacutetude La deacutetection drsquoun signal moleacuteculaire
pendant 12 mois dans plusieurs entreacutees drsquoun mecircme terrier suggegravere fortement que les sols de terriers
peuvent constituer un reacuteservoir environnemental pour M bovis La survie des mycobacteacuteries est
favoriseacutee dans ces structures complexes et profondes ougrave regravegne un microclimat propice (Moore amp Roper
2003) tant au niveau de la stabiliteacute de la tempeacuterature de lrsquohumiditeacute optimale que de la protection
contre les UV Ce reacuteservoir potentiel repreacutesente un risque pour le maintien de la maladie dans le groupe
social de blaireaux qui partage le terrier notamment pour les blairotins qui y naissent pendant lrsquohiver
mais aussi entre les groupes sociaux qui peuvent reacuteguliegraverement changer de terriers Lorsqursquoils sont
localiseacutes dans les pacirctures et sont accessibles aux bovins le sol des terriers contamineacutes repreacutesenterait
un risque potentiel pour les bovins qui les explorent notamment un risque drsquoinhalation de bioaeacuterosols
contamineacutes geacuteneacutereacutes agrave partir de sol Enfin les sols et latrines contamineacutes souvent en limite des plateaux
calcaires seraient susceptibles drsquoecirctre agrave lrsquoorigine de la contamination des pacirctures par le biais des
infiltrations drsquoeau des pheacutenomegravenes de reacutesurgence drsquoeau et de ruissellement
Les mesures de bioseacutecuriteacute preacuteconisant la clocircture des terriers situeacutes dans les pacirctures semblent
essentielles agrave respecter par les eacuteleveurs si lrsquoon considegravere la longue persistance de la bacteacuterie dans ce
biotope particulier Le repeuplement des terriers contamineacutes ne fait lrsquoobjet drsquoaucun controcircle agrave lrsquoheure
actuelle Sous le controcircle du CSRPN (Conseil Scientifique Reacutegional du Patrimoine Naturel) une eacutetude
est actuellement en cours en Cocircte drsquoOr pour lrsquoeacutetude de faisabiliteacute drsquoun protocole de deacutepeuplement des
terriers (pieacutegeage de tous les individus du groupe) associeacute agrave un traitement des tunnels par des reacutepulsifs
permanents pour eacuteviter la recolonisation immeacutediate
La preacutevalence de M bovis dans lrsquoenvironnement des animaux infecteacutes en Cocircte drsquoOr est
probablement sous-estimeacutee dans notre eacutetude agrave cause de limites temporelles spatiales et
meacutethodologiques auxquelles nous avons fait face En premier lieu il existe un deacutelai incompressible de
plusieurs semaines agrave plusieurs mois entre les trois eacutevegravenements suivants i) la contamination
environnementale par un animal excreacuteteur (bovin ou faune sauvage) ii) le diagnostic de lrsquoinfection chez
cet animal (apregraves prophylaxie et abattage pour les bovins en hiver et pieacutegeage pour les blaireaux au
printemps et en eacuteteacute) et iii) le preacutelegravevement et lrsquoanalyse drsquoeacutechantillons environnementaux dans les zones
de vie de ces animaux Cette limite temporelle est eacutegalement eacutevoqueacutee par drsquoautres auteurs (Fine
OrsquoBrien et al 2011) Il serait ideacuteal drsquoeffectuer les preacutelegravevements environnementaux dans un deacutelai le plus
court possible apregraves lrsquoexcreacutetion du bacille par un animal infecteacute toutefois ce deacutelai deacutepend du rythme
des prophylaxies et de la freacutequence du pieacutegeage des blaireaux De plus sur le terrain malgreacute notre
strateacutegie drsquoeacutechantillonnage ciblant les zones partageacutees preacutefeacuterentiellement par la faune domestique et
sauvage les surfaces agrave preacutelever restent importantes Lrsquoanalyse de la totaliteacute des surfaces cibleacutees est
humainement techniquement et eacuteconomiquement irreacutealisable le nombre de preacutelegravevements a donc ducirc
Discussion geacuteneacuterale
183
ecirctre optimiseacute pour aboutir agrave une analyse pertinente Les reacutesultats obtenus ne donnent de ce fait qursquoune
image partielle de la contamination reacuteelle des zones preacuteleveacutees Enfin la charge bacteacuterienne de certains
preacutelegravevements environnementaux nrsquoa pu ecirctre quantifieacutee signifiant que les quantiteacutes de cibles preacutesentes
eacutetaient proches des limites de deacutetection de nos systegravemes moleacuteculaires Pour ces diffeacuterentes raisons
nous ne pouvons donc pas exclure une contamination environnementale plus large avec des quantiteacutes
tregraves faibles de M bovis
3 Survie et circulation de M bovis dans lrsquoenvironnement
La survie des bacteacuteries dans le sol deacutepend des facteurs biotiques et abiotiques qui caracteacuterisent
ces sols Les diffeacuterentes eacutetudes sur la survie de M bovis ont essentiellement consideacutereacute les facteurs
abiotiques climatiques telles que la tempeacuterature lrsquohumiditeacute et lrsquoexposition aux UV dans un seul type
de sol (Young et al 2005 Jackson et al 1995 Duffield amp Young 1985) Nous avons montreacute que M bovis
SB0120 eacutetait cultivable au moins cinq mois agrave une tempeacuterature basse (4degC) avec une mortaliteacute
bacteacuterienne minime et jusqursquoagrave trois mois agrave 22degC Ces dureacutees de survie sont supeacuterieures agrave celles
rapporteacutees dans de preacuteceacutedentes eacutetudes (Fine Bolin et al 2011 Jackson et al 1995 Duffield amp Young
1985) Les sols retrouveacutes dans lrsquoAuxois ont agrave la fois des caracteacuteristiques diffeacuterentes des sols
habituellement deacutecrits comme favorables agrave la survie des mycobacteacuteries (sols sableux agrave pH acide) et
des caracteacuteristiques communes leur richesse en matiegravere organique et en fer Bien que nous nrsquoayons
pas pu mettre en eacutevidence un effet de la nature du sol dans notre eacutetude sur la survie de M bovis il
nous parait probable que les sols argilo-limoneux peacuterennisent la maladie dans les valleacutees de lrsquoAuxois
Notre expeacuterience limiteacutee par des contraintes techniques lieacutees agrave lrsquoutilisation du laboratoire de
confinement L3 du CHU de Dijon aurait neacutecessiteacute lrsquoutilisation drsquoun plus grand nombre de sols de points
de preacutelegravevements et de reacutepeacutetitions biologiques La concentration des foyers bovins (notamment des
recontaminations de cheptels) dans une zone limiteacutee du deacutepartement en Auxois interroge eacutegalement
sur le rocircle du contexte paysager et geacuteomorphologique commun des pacirctures suspectes dans la survie
de M bovis
Ces dureacutees de survie de M bovis dans les sols doivent ecirctre consideacutereacutees avec preacutecaution car
obtenues dans des conditions expeacuterimentales controcircleacutees qui ne miment que partiellement les
conditions environnementales reacuteelles (sols deacutebarrasseacutees de leur flore endogegravene agrave une tempeacuterature
drsquoincubation constante une humiditeacute optimale et une protection contre les UV) Il est probable qursquoen
conditions naturelles la survie de M bovis soit moins longue agrave cause des variations nyctheacutemeacuterales de
tempeacuteratures des variations des preacutecipitations des UV ainsi que de lrsquoeffet barriegravere potentiel de la flore
endogegravene des matrices environnementales La correacutelation de ces donneacutees avec les courbes de
Discussion geacuteneacuterale
184
tempeacuteratures permettent de montrer que la peacuteriode de novembre agrave avril ougrave les tempeacuteratures sont les
plus basses est compatible avec une survie optimale de M bovis dans les sols
Les reacutesultats obtenus dans notre eacutetude de survie ont souleveacute la question de la persistance de
M bovis dans le sol dans un eacutetat viable mais non cultivable (VNC) puisque la bacteacuterie nrsquoest plus
cultivable mais toujours deacutetecteacutee par qPCR Lrsquoeacutetat VNC de M bovis a deacutejagrave eacuteteacute suggeacutereacute dans des eacutetudes
de survie dans du sol (Adams et al 2013 Young et al 2005) Les mycobacteacuteries possegravedent une capaciteacute
particuliegravere agrave srsquoadapter et agrave survivre dans des conditions deacutefavorables en lrsquoabsence de nutriments
drsquooxygegravene sous diffeacuterents stress exogegravenes notamment dans les phagosomes de lrsquohocircte pour les
mycobacteacuteries pathogegravenes (Gengenbacher amp Kaufmann 2012 Boon amp Dick 2012 Voskuil et al 2003)
Les mycobacteacuteries disposent de diffeacuterents systegravemes agrave deux composants (permettant une transduction
de signal) leur permettant la perception sensorielle des conditions environnementales et en particulier
de la composition du milieu extracellulaire (Cook et al 2009 Cole et al 1998) Dans les substrats
environnementaux la bacteacuterie conserve probablement tout son potentiel de perception du milieu qui
lrsquoentoure et ses capaciteacutes drsquoadaptation mais elle ne peut pas interagir avec lui comme avec les cellules
de lrsquohocircte A lrsquoheure actuelle les meacutecanismes drsquoadaptation mis en jeu par M bovis pour survivre lors de
son excreacutetion par lrsquohocircte dans lrsquoenvironnement sont inconnus
Nous avons montreacute pour la premiegravere fois que les lombrics veacuteritables ingeacutenieurs du sol ont la
capaciteacute de disseacuteminer la bacteacuterie dans leur environnement par le biais des tunnels qursquoils creusent et
des turricules qursquoils eacutemettent agrave et sous la surface du sol Lrsquoenfouissement de la bacteacuterie agrave quelques
centimegravetres sous le sol favorise probablement sa survie Les lombrics peuvent eacutegalement jouer un rocircle
de vecteur quand ils portent du sol contamineacute dans leur tube digestif et qursquoils sont ingeacutereacutes par des
blaireaux ou des sangliers Cette eacutetude soulegraveve la probleacutematique du rocircle de la faune du sol dans la
survie et la circulation de M bovis dans lrsquoenvironnement et le risque que des reacuteservoirs cacheacutes de la
bacteacuterie entretiennent agrave bas bruit une contamination de lrsquoenvironnement
Perspectives
Perspectives
187
Lrsquoeacutevaluation de la contamination environnementale reacutealiseacutee en Cocircte drsquoOr et dans une moindre
mesure en Charente (terriers de blaireaux uniquement) nrsquoest pas extrapolable aux autres reacutegions
franccedilaises toucheacutees par la tuberculose A ce jour aucune investigation environnementale aussi
complegravete nrsquoa eacuteteacute meneacutee dans des deacutepartements fortement infecteacutes comme la Dordogne et les
Pyreacuteneacutees Atlantiques ougrave bovins et faune sauvage cohabitent Il serait donc inteacuteressant drsquoeacutevaluer le
niveau de contamination environnementale des zones partageacutees par les bovins et la faune sauvage afin
de veacuterifier si les zones humides et les terriers sont eacutegalement agrave risque dans ces deacutepartements De plus
les reacutesultats obtenus dans la zone endeacutemique de Cocircte drsquoOr ont suggeacutereacute le rocircle possible du contexte
geacuteomorphologique hydrologique et peacutedologique de lrsquoAuxois dans la contamination environnementale
et la persistance de M bovis Pour veacuterifier cette hypothegravese il serait inteacuteressant de comparer le contexte
des pacirctures suspectes de lrsquoAuxois avec les pacirctures suspectes des deacutepartements les plus toucheacutes par la
tuberculose bovine La caracteacuterisation physico-chimique du sol de ces pacirctures permettrait peut-ecirctre de
mettre en relation drsquoeacuteventuelles proprieacuteteacutes communes entre les sols incrimineacutes et la persistance de la
bacteacuterie La mise en eacutevidence de facteurs de risque paysagers hydrographiques etou peacutedologiques
permettrait eacuteventuellement de mettre au point un outil de preacutevision des risques en fonction du
contexte rencontreacute (pacirctures agrave risque) drsquoadapter au mieux la gestion sanitaire et drsquoameacuteliorer la
bioseacutecuriteacute dans les eacutelevages Si le rocircle des zones humides (laquomouilles raquo) et des sources dans drsquoautres
reacutegions est confirmeacute il conviendra de limiter lrsquoaccegraves des bovins agrave ces zones pour limiter la
contamination En Cocircte drsquoOr des approches de ce type (drainage et clocircture des zones humides) pour
empecirccher lrsquoaccegraves des animaux sont en cours en collaboration avec les eacuteleveurs les agences de bassin
lrsquoONEMA ainsi que les gestionnaires des zones humides agrave lrsquoeacutechelle du bassin versant de la Brenne
La dureacutee de survie deacutemontreacutee de M bovis dans les sols remet partiellement en cause la dureacutee
du deacutelai de carence de 2 mois avant reacuteinstallation drsquoanimaux suite agrave la peacuteriode de vide sanitaire dans
les exploitations qui ont eacuteteacute foyer de tuberculose Nos observations devront ecirctre discuteacutees dans le
cadre de lrsquooptimisation de ces pratiques
Sur le plan expeacuterimental notre expeacuterience de survie sur deux sols eacutetant insuffisante une eacutetude
de la survie de M bovis sur un grand nombre de sols aux caracteacuteristiques physico-chimiques
contrasteacutees permettrait drsquoidentifier les facteurs physico-chimiques du sol associeacutes agrave une survie
optimale de la bacteacuterie comme cela a pu ecirctre fait au sein de notre eacutequipe pour Listeria monocytogenes
(Locatelli et al 2013) Lrsquoidentification des facteurs eacutedaphiques favorisant la persistance de M bovis
permettra de mettre en eacutevidence les sols agrave risque
La recherche de M bovis dans des matrices environnementales se heurte aux limites des
techniques utiliseacutees Concernant la qPCR bien qursquoelle reste une technique de choix des efforts sont
encore neacutecessaires pour abaisser les seuils de deacutetection dans les matrices complexes faiblement
Perspectives
188
chargeacutees en mycobacteacuteries Drsquoautres techniques PCR pourraient ecirctre envisageacutees notamment la PCR
digitale (ou PCR numeacuterique) qui permet de gagner en sensibiliteacute et de limiter lrsquoimpact des inhibiteurs
mais dont lrsquoutilisation est encore limiteacutee par le coucirct De plus la qPCR deacutetecte lrsquoADN bacteacuterien sans
donner aucune indication sur la viabiliteacute de la cellule Devant les difficulteacutes agrave isoler M bovis par la
culture agrave partir de matrices environnementales complexes agrave cause notamment de sa croissance tregraves
lente une approche transcriptomique permettrait de preacuteciser lrsquoeacutetat viable de M bovis Cette approche
pourrait eacutegalement apporter des connaissances encore totalement inconnues agrave lrsquoheure actuelle sur les
meacutecanismes de survie mis en jeu par la bacteacuterie une fois excreacuteteacutee par son hocircte dans lrsquoenvironnement
et mettre en eacutevidence les gegravenes impliqueacutes dans son adaptation aux conditions environnementales et
donc sa persistance Dans lrsquoobjectif drsquoeacuteclaircir ces meacutecanismes qui semblent fondamentaux pour la
compreacutehension de lrsquoadaptation et de la survie de M bovis dans lrsquoenvironnement lrsquoutilisation drsquoune
approche transcriptomique permettrait drsquoidentifier les modifications transcriptionnelles agrave lrsquoeacutechelle du
geacutenome complet par une approche de seacutequenccedilage des ARN (RNA seq) et de certains gegravenes par RT-PCR
lors de lrsquointroduction de M bovis dans le sol La comparaison de ces donneacutees avec les reacutesultats obtenus
in vitro et in vivo dans diffeacuterentes conditions de stress (Rodriguez et al 2008 Jang et al 2009)
permettraient de veacuterifier si les mecircmes meacutecanismes sont impliqueacutes Les gegravenes du reacutegulon de la
dormance preacutesents chez M bovis sont surexprimeacutes dans les macrophages in vitro et in vivo (Rodriguez
et al 2008) il serait particuliegraverement inteacuteressant drsquoeacutevaluer leur niveau drsquoexpression lors de
lrsquointroduction de la bacteacuterie dans le sol Comme il est deacutelicat drsquoextraire lrsquoARN de matrices complexes
comme le sol lrsquoutilisation drsquoextraits aqueux de sol pourrait faciliter lrsquoeacutetude (Piveteau et al 2011) Pour
aller plus loin il serait inteacuteressant de comparer les transcriptomes de M bovis lorsque les conditions
abiotiques (type de sol tempeacuterature humiditeacute etc) et biotiques (comparaison absencepreacutesence du
microbiote endogegravene du sol) varient pour cerner plus finement les meacutecanismes drsquoadaptation deacuteployeacutes
face aux diffeacuterents stress
Les meacutethodes de culture de M bovis agrave partir de matrices environnementales complexes
comme le sol sont encore agrave ameacuteliorer Lrsquoisolement de souches environnementales de M bovis
permettrait gracircce aux nouvelles techniques de seacutequenccedilage (NGS) de faire de la geacutenomique
comparative sur geacutenome complet avec les mecircmes souches isoleacutees chez les animaux et drsquoameacuteliorer les
connaissances sur les meacutecanismes geacuteneacutetiques mis en jeu dans lrsquoadaptation et la persistance de la
bacteacuterie dans lrsquoenvironnement
Concernant la faune la persistance et la circulation des mycobacteacuteries dans lrsquoeau et le sol
pourraient ecirctre favoriseacutees par la micro- et la meacutesofaune preacutesentes dans ces matrices Les protozoaires
et particuliegraverement les amibes ont fait lrsquoobjet drsquoeacutetudes montrant la survie dans leurs trophozoiumltes et
leurs cystes de nombreuses espegraveces dont M bovis (Taylor et al 2003 Mba Medie et al 2011) Dans
notre eacutetude un des eacutechantillons de neacutematodes preacuteleveacute dans les seacutediments drsquoune source a donneacute un
Perspectives
189
signal moleacuteculaire reacuteveacutelant un lien potentiel entre neacutematodes et persistance des mycobacteacuteries Pour
des raisons de temps et de compeacutetences nous nrsquoavons pas exploreacute minutieusement ces eacuteventuels
reacuteservoirs cacheacutes de M bovis Devant lrsquoabondance et la diversiteacute de ces animaux dans les matrices que
nous analysons des recherches compleacutementaires nous semblent neacutecessaires pour preacuteciser leur rocircle
potentiel dans la persistance de M bovis
Les diffeacuterents reacutesultats obtenus pendant ce travail et les implications qui en ressortent ont eacuteteacute
reacuteguliegraverement partageacutes avec les instances sanitaires et agricoles de Cocircte drsquoOr afin drsquoenrichir les
connaissances sur lrsquoeacutepideacutemiologie tregraves complexe de cette maladie et de participer agrave lrsquoameacutelioration des
mesures de bioseacutecuriteacute qui en deacutecoulent Drsquoautres deacutepartements toucheacutes par la tuberculose bovine
inteacuteresseacutes par nos travaux nous ont fait parvenir des demandes drsquoanalyse environnementale dans des
zones ougrave la transmission indirecte de la maladie est fortement suspecteacutee
Les deux protocoles innovants pour lrsquoextraction des mycobacteacuteries du sol et le suivi de leur
croissance en milieu liquide deacuteveloppeacutes dans ce travail de thegravese font lrsquoobjet drsquoune eacutetude de valorisation
(protection intellectuelle et eacutetude de marcheacute) en collaboration avec la SATT Grand Est
Enfin les diffeacuterentes ideacutees et perspectives deacuteveloppeacutees dans cette thegravese ont eacuteteacute reprises en
partie dans le cadre de lrsquoappel drsquooffre Horizon 2020 agrave travers le deacutepocirct drsquoun projet europeacuteen
multipartenaire (scientifiques et entreprises) qui est coordonneacute par lrsquoINRA de Tours Nouzilly et qui
implique de nombreux partenaires en Allemagne (Lionex PME speacutecialiseacutee dans les outils de deacutetection
seacuterologique speacutecifiques) en Hollande (WUR) en UK (APHA) en Irlande (UCD) en Italie (Universiteacute de
Naples) Ce projet intituleacute DEPICT vise entre autre agrave une meilleure compreacutehension des meacutecanismes
infectieux de la reacuteaction inflammatoire et immunologique des animaux mais eacutegalement agrave deacuteterminer
le rocircle de lrsquoenvironnement dans la survie des pathogegravenes comme M bovis et Map entre autres La
diversiteacute et la persistance de M bovis et Map seront eacutetudieacutees dans lrsquoenvironnement afin de deacuteterminer
les facteurs influenccedilant la survie de ces pathogegravenes en particulier dans les sols Au travers de ce
programme et drsquoautres collaborations agrave venir avec lrsquoINRA de Tours le travail deacutebuteacute au cours de cette
thegravese sur M bovis sera eacutelargi agrave Map mycobacteacuterie eacutegalement difficilement cultivable et revecirctant une
grande importance pour la santeacute animale et la santeacute humaine (elle serait impliqueacutee dans la maladie de
Crohn) Les nouveaux outils (culture et deacutetection) deacuteveloppeacutes au cours de cette thegravese seront testeacutes sur
Map pour permettre lrsquoisolement drsquoun plus grand nombre de souches environnementales
Une des questions en suspens reste la deacutemonstration de la contamination de bovins agrave partir
de M bovis preacutesente dans des matrices environnementales Des expeacuteriences de contamination
expeacuterimentale en animalerie P3 agrave partir de sol ou drsquoeau contamineacutes pourraient permettre cette
deacutemonstration Lrsquoutilisation du modegravele cobaye espegravece tregraves sensible agrave lrsquoinfection par M bovis pour
reacutealiser des expeacuteriences preacuteliminaires pourrait ecirctre envisageacutee avant de concevoir des expeacuteriences sur
bovins Bien que ce modegravele animal soit assez eacuteloigneacute du modegravele bovin en ce qui concerne lrsquoeacutetude de la
Perspectives
190
transmission indirecte de M bovis (les cobayes geacutenegraverent par exemple beaucoup moins drsquoaeacuterosols que
les bovins quand ils boivent et ils ne ruminent pas) il constitue un modegravele plus facilement manipulable
et moins coucircteux Il permettrait drsquoaffiner les donneacutees sur les voies de transmission environnementales
potentielles avant de passer sur un modegravele bovin beaucoup plus contraignant de par la taille des
animaux et plus coucircteux
Reacutefeacuterences bibliographiques
Reacutefeacuterences bibliographiques
192
Acevedo-Whitehouse K Vicente J Gortaacutezar C Houmlfle U Fernaacutendez-de-Mera IG Amos W 2005 Genetic resistance to bovine tuberculosis in the Iberian wild boar Mol Ecol 14 3209ndash17
Adams AP Bolin SR Fine AE Bolin CA Kaneene JB 2013 Comparison of PCR versus culture for detection of Mycobacterium bovis after experimental inoculation of various matrices held under environmental conditions for extended periods Appl Environ Microbiol 79 6501ndash6506
Adekambi T Ben Salah S Khlif M Raoult D Drancourt M 2006 Survival of environmental mycobacteria in Acanthamoeba polyphaga Appl Environ Microbiol 72 5974ndash5981
Adeacutekambi T Drancourt M 2004 Dissection of phylogenetic relationships among 19 rapidly growing Mycobacterium species by 16S rRNA hsp65 sodA recA and rpoB gene sequencing Int J Syst Evol Microbiol 54 2095ndash105
Aimeacute B Lequen L Balageas A Haddad N Maugein J 2012 M bovis and M caprae infections in Aquitaine A clinico-epidemiologic study of 15 patients Pathol Biol (Paris) 60 156ndash159
Alexander K A Laver PN Michel AL Williams M van Helden PD Warren RM van Pittius NCG 2010 Novel Mycobacterium tuberculosis complex pathogen M Mungi Emerg Infect Dis 16 1296ndash1299
Allix C Walravens K Saegerman C Godfroid J Supply P Fauville-Dufaux M 2006 Evaluation of the epidemiological relevance of variable-number tandem-repeat genotyping of Mycobacterium bovis and comparison of the method with IS6110 restriction fragment length polymorphism analysis and spoligotyping J Clin Microbiol 44 1951ndash1962
Aacutelvarez AH Estrada-Chaacutevez C Flores-Valdez MA 2009 Molecular findings and approaches spotlighting Mycobacterium bovis persistence in cattle Vet Res 40 1ndash16
Ameni G Vordermeier M Firdessa R Aseffa A Hewinson G Gordon S V Berg S 2011 Mycobacterium tuberculosis infection in grazing cattle in central Ethiopia Vet J 188 359ndash361
ANSES 2011 Tuberculose bovine et faune sauvageRapport Avril 2011
Antoine D Jarlier V 2010 La tuberculose humaine agrave Mycobacterium bovis en France Bull eacutepideacutemiologique santeacute Anim Aliment Septembre
Aranaz A Cousins D Mateos A Domınguez L 2003 Elevation of Mycobacterium tuberculosis subsp caprae (Aranaz et al 1999) to species rank as Mycobacterium caprae comb nov sp Int J Syst Evol Microbiol 53 1785ndash1789
Aranaz A Lieacutebana E Goacutemez-Mampaso Enrique Galaacuten JC Cousins D Ortega A Blaacutezquez J Baquero F Mateos A Suacutearez G Domiacutenguez L 1999 Mycobacterium tuberculosis subsp caprae subsp nov a taxonomic study of a new member of the Mycobacterium tuberculosis complex isolated from goats in Spain Int J Syst Bacteriol 49 1263ndash1273
Archuleta RJ Yvonne Hoppes P Primm TP 2005 Mycobacterium avium enters a state of metabolic dormancy in response to starvation Tuberculosis (Edinb) 85 147ndash58
Aronson T Holtzman A Glover N Boian M Froman S Berlin OGW Hill H Stelma G J 1999 Comparison of large restriction fragments of Mycobacterium avium isolates recovered from AIDS and non-AIDS patients with those of isolates from potable water J Clin Microbiol 37 1008ndash1012
Ayele WY Neill SD Zinsstag J Weiss MG Pavlik I 2004 Bovine tuberculosis an old disease but a new threat to Africa Int J Tuberc Lung Dis 8 924ndash937
Ballesteros C Garrido JM Vicente J Romero B Galindo RC Minguijoacuten E Villar M Martiacuten-Hernando
Reacutefeacuterences bibliographiques
193
MP Sevilla I Juste R Aranaz A de la Fuente J Gortaacutezar C 2009 First data on Eurasian wild boar response to oral immunization with BCG and challenge with a Mycobacterium bovis field strain Vaccine 27 6662ndash6668
Barasona JA Latham MC Acevedo P Armenteros JA Latham ADM Gortazar C Carro F Soriguer RC Vicente J 2014 Spatiotemporal interactions between wild boar and cattle Implications for cross-species disease transmission Vet Res 45 1ndash11
Barasona JA Torres MJ Aznar J Gortaacutezar C Vicente J 2015 DNA detection reveals Mycobacterium tuberculosis complex shedding routes in its wildlife reservoir the Eurasian wild boar Transbound Emerg Dis (December) doi101111tbed12458
Barker J Brown MRW 1994 Trojan Horses of the microbial world protozoa and the survival of bacterial pathogens in the environment Microbiology 140 1253ndash1259
Baubet E Ropert-Coudert Y Brandt S 2003 Seasonal and annual variations in earthworm consumption by wild boar (Sus scrofa scrofa L) Wildl Res 30 179
Ben Salah I Drancourt M 2010 Surviving within the amoebal exocyst the Mycobacterium avium complex paradigm BMC Microbiol 10 99
Beacutenet J Boschiroli ML Dufour B Garin-Bastuji B 2006 Lutte contre la tuberculose bovine en France de 1954 agrave 2005 analyse de la pertinence eacutepideacutemiologique de lrsquoeacutevolution de la reacuteglementation Epideacutemiologie santeacute Anim 50 127ndash143
Benham PF 1985 The behaviour of badgers and cattle and some factors that affect the chance of contact between the species Appl Anim Behav Sci 14 390ndash391
Benham PF Broom DM 1989 Interactions between cattle and badgers at pasture with reference to bovine tuberculosis transmission Br Vet J 145 226ndash241
Bercovier H Kafri O Sela S 1986 Mycobacteria possess a surprisingly small number of ribosomal RNA genes in relation to the size of their genome Biochem Biophys Res Commun 136 1136ndash1141
Berg S Garcia-Pelayo MC Muumlller B Hailu E Asiimwe B Kremer K Dale J Boniotti MB Rodriguez S Hilty M Rigouts L Firdessa R MacHado A Mucavele C Ngandolo BNR Bruchfeld J Boschiroli ML Muumlller A Sahraoui N Pacciarini M Cadmus SI Joloba M Van Soolingen D Michel AL Djoslashnne B Aranaz A Zinsstag J Van Helden P Portaels F Kazwala R Kaumlllenius G Hewinson RG Aseffa A Gordon S V Smith NH 2011 African 2 a clonal complex of Mycobacterium bovis epidemiologically important in East Africa J Bacteriol 193 670ndash678
Bermingham ML Bishop SC Woolliams JA Pong-Wong R Allen AR McBride SH Ryder JJ Wright DM Skuce RA McDowell SW Glass EJ 2014 Genome-wide association study identifies novel loci associated with resistance to bovine tuberculosis Heredity (Edinb) 112 543ndash551
Beširović H Alić A Špičić S Cvetnić Ž Prašović S Velić L 2012 Bovine tuberculosis in Bosnia and Herzegovina caused by Mycobacterium caprae Vet Arh 82 341ndash349
Beumer A King D Donohue M Mistry J Covert T Pfaller S 2010 Detection of Mycobacterium avium subsp paratuberculosis in drinking water and biofilms by quantitative PCR Appl Environ Microbiol 76 7367ndash7370
Biek R OrsquoHare A Wright D Mallon T McCormick C Orton RJ McDowell S Trewby H Skuce RA Kao RR 2012 Whole genome sequencing reveals local transmission patterns of Mycobacterium bovis in sympatric cattle and badger populations PLoS Pathog 8 e1003008
Biet F Boschiroli ML Thorel MF Guilloteau LA 2005 Zoonotic aspects of Mycobacterium bovis and
Reacutefeacuterences bibliographiques
194
Mycobacterium avium-intracellulare complex (MAC) Vet Res36 411-436
Bille E Dauphin B Leto J Bougnoux M-E Beretti J-L Lotz A Suarez S Meyer J Join-Lambert O Descamps P Grall N Mory F Dubreuil L Berche P Nassif X Ferroni A 2012 MALDI-TOF MS Andromas strategy for the routine identification of bacteria mycobacteria yeasts Aspergillus spp and positive blood cultures Clin Microbiol Infect 18 1117ndash1125
Biswas S Rolain JM 2013 Use of MALDI-TOF mass spectrometry for identification of bacteria that are difficult to culture J Microbiol Methods 92 14ndash24
Blackwood KS He C Gunton J Turenne CY Wolfe J Kabani A M 2000 Evaluation of recA sequences for identification of Mycobacterium species J Clin Microbiol 38 2846ndash2852
Bland CS Ireland JM Lozano E Alvarez ME Primm TP 2005 Mycobacterial ecology of the Rio Grande Appl Environ Microbiol 71 5719ndash5727
Boadella M Barasona JA Diaz-Sanchez S Lyashchenko KP Greenwald R Esfandiari J Gortaacutezar C 2012 Performance of immunochromatographic and ELISA tests for detecting fallow deer infected with Mycobacterium bovis Prev Vet Med 104 160ndash164
Boadella M Lyashchenko K Greenwald R Esfandiari J Jaroso R Carta T Garrido JM Vicente J de la Fuente J Gortaacutezar C 2011 Serologic tests for detecting antibodies against Mycobacterium bovis and Mycobacterium avium subspecies paratuberculosis in Eurasian wild boar (Sus scrofa scrofa) J Vet Diagnostic Investig 23 77ndash83
Boumlddinghaus B Rogall T Flohr T Bloumlcker H Boumlttger EC 1990 Detection and identification of mycobacteria by amplification of rRNA J Clin Microbiol 28 1751ndash1759
Boumlhm M Hutchings MR White PCL 2009 Contact networks in a wildlife-livestock host community identifying high-risk individuals in the transmission of bovine TB among badgers and cattle PLoS One 4 e5016
Bollo E Guarda F Capucchio MT Galietti F 1998 Direct detection of Mycobacterium tuberculosis Complex and M avium Complex in tissue specimens from cattle through identification of specific rRNA sequences J Vet Med 45 395ndash400
Boniotti MB Goria M Loda D Garrone A Benedetto A Mondo A Tisato E Zanoni M Zoppi S Dondo A Tagliabue S Bonora S Zanardi G Pacciarini ML 2009 Molecular typing of Mycobacterium bovis strains isolated in Italy from 2000 to 2006 and evaluation of variable-number tandem repeats for geographically optimized genotyping J Clin Microbiol 47 636ndash644
Boon C Dick T 2012 How Mycobacterium tuberculosis goes to sleep the dormancy survival regulator DosR a decade later Future Microbiol 7 513ndash518
Boon C Dick T 2002 Mycobacterium bovis BCG response regulator essential for hypoxic dormancy J Bacteriol 184 6760ndash6767
Boon C Li R Qi R Dick T 2001 Proteins of Mycobacterium bovis BCG induced in the Wayne dormancy model J Bacteriol 183 2672ndash2676
Boulahbal F Robert J Trystram D Benoist A C Vincent V Jarlier V 1998 La tuberculose humaine agrave Mycobacterium bovis en France durant lrsquoanneacutee 1995 Bull eacutepideacutemiologique Hebd 48
Boyard C Vourcrsquoh G Barnouin J 2008 The relationships between Ixodes ricinus and small mammal species at the woodland-pasture interface Exp Appl Acarol 44 61ndash76
Brennan PJ Nikaido H 1995 The envelope of mycobacteria Annu Rev Biochem 64 29ndash63
Reacutefeacuterences bibliographiques
195
Brodin P Eiglmeier K Marmiesse M Billault A Garnier T Niemann S Cole ST Brosch R 2002 Bacterial artificial chromosome-based comparative genomic analysis identifies Mycobacterium microti as a natural ESAT-6 deletion mutant Infect70 5568ndash5578
Brook RK McLachlan SM 2009 Transdisciplinary habitat models for elk and cattle as a proxy for bovine tuberculosis transmission risk Prev Vet Med 91 197ndash208
Brook RK Wal E Vander van Beest FM McLachlan SM 2012 Evaluating use of cattle winter feeding areas by elk and white-tailed deer Implications for managing bovine tuberculosis transmission risk from the ground up Prev Vet Med 1ndash11
Brosch R Gordon SV Marmiesse M Brodin P Buchrieser C Eiglmeier K Garnier T Gutierrez C Hewinson G Kremer K Parsons LM Pym A Samper S van Soolingen D Cole S 2002 A new evolutionary scenario for the Mycobacterium tuberculosis complex Proc Natl Acad Sci U S A 99 3684ndash3689
Brosch R Gordon S V Garnier T Eiglmeier K Frigui W Valenti P Dos Santos S Duthoy S Lacroix C Garcia-Pelayo C Inwald JK Golby P Garcia JN Hewinson RG Behr M A Quail M A Churcher C Barrell BG Parkhill J Cole ST 2007 Genome plasticity of BCG and impact on vaccine efficacy Proc Natl Acad Sci U S A 104 5596ndash5601
Brotherstone S White IMS Coffey M Downs SH Mitchell AP Clifton-Hadley RS More SJ Good M Woolliams JA 2010 Evidence of genetic resistance of cattle to infection with Mycobacterium bovis J Dairy Sci 93 1234ndash1242
Broughan JM Crawshaw TR Downs SH Brewer J Clifton-Hadley RS 2013a Mycobacterium bovis infections in domesticated non-bovine mammalian species Part 2 A review of diagnostic methods Vet J 198 346ndash351
Broughan JM Downs SH Crawshaw TR Upton PA Brewer J Clifton-Hadley RS 2013b Mycobacterium bovis infections in domesticated non-bovine mammalian species Part 1 Review of epidemiology and laboratory submissions in Great Britain 2004-2010 Vet J 198 339ndash345
Buddle BM Wedlock DN Denis M Vordermeier HM Hewinson RG 2011 Update on vaccination of cattle and wildlife populations against tuberculosis Vet Microbiol 151 14ndash22
Butler WR Guthertz LS 2001 Mycolic acid analysis by High-Performance Liquid Chromatography for identification of Mycobacterium species Clin Microbiol Rev 14 704ndash726
Cadmus SI Adesokan HK Jenkins AO van Soolingen D 2009a Mycobacterium bovis and M tuberculosis in goats Nigeria Emerg Infect Dis 15 2066ndash2067
Cadmus SI Jenkins AO Godfroid J Osinusi K Adewole IF Murphy RL Taiwo BO 2009b Mycobacterium tuberculosis and Mycobacterium africanum in stools from children attending an immunization clinic in Ibadan Nigeria Int J Infect Dis 13 740ndash744
Cadmus SI Palmer S Okker M Dale J Gover K Smith N Jahans K Hewinson RG Gordon S V 2006 Molecular analysis of human and bovine tubercle bacilli from a local setting in Nigeria J Clin Microbiol 44 29ndash34
Cadmus SI Yakubu MK Magaji AA Jenkins AO van Soolingen D 2010 Mycobacterium bovis but also M africanum present in raw milk of pastoral cattle in north-central Nigeria Trop Anim Health Prod 42 1047ndash1048
Cambau E Lamaitre N Sougakoff W Jarlier V 2003 Reacutesistance aux antituberculeux Antibiotiques 5 29ndash37
Reacutefeacuterences bibliographiques
196
Carbonnelle B Dailloux M Lebrun L Maugein J Pernot C 2003 Cahier de biologie meacutedicale Ndeg29Mycobacteacuteries mycobacteacuterioses
Carrasco-Garcia R Barasona JA Gort Montoro V Sanchez-Vizcaino JM Vicente J 2015 Wildlife and livestock use of extensive farm resources in South Central Spain implications for disease transmission Eur J Wildl Res62(1) 65-78
Cassidy JP 2006 The pathogenesis and pathology of bovine tuberculosis with insights from studies of tuberculosis in humans and laboratory animal models Vet Microbiol 112 151ndash161
Cassidy JP Bryson DG Pollock JM Evans RT Forster F Neill SD 1999 Lesions in cattle exposed to Mycobacterium bovis-inoculated calves J Comp Pathol 121 321ndash37
Castets M Sarrat H 1968 Bacteriologic aspects of mycobacteria isolated at Dakar in 1967 Bull Soc Med Afr Noire Lang Fr 13 463ndash469
Cavalerie L Courcoul A Boschiroli ML Reacuteveillaud E Gay P 2014 Tuberculose bovine en France en 2014 une situation stable Bull eacutepideacutemiologique santeacute Anim Aliment 4ndash11
Cha D Cheng D Liu M Zeng Z Hu X Guan W 2009 Analysis of fatty acids in sputum from patients with pulmonary tuberculosis using gas chromatography-mass spectrometry preceded by solid-phase microextraction and post-derivatization on the fiber J Chromatogr A 1216 1450ndash1457
Chapinal N Elkin BT Joly DO Schumaker BA Stephen C 2012 Agreement between the caudal fold test and serological tests for the detection of Mycobacterium bovis infection in bison Prev Vet Med 105 326ndash330
Ciliberti A Gavier-Wideacuten D Yon L Hutchings MR Artois M 2015 Prioritisation of wildlife pathogens to be targeted in European surveillance programmes Expert-based risk analysis focus on ruminants Prev Vet Med 118 271ndash284
Cirillo JD Falkow S Tompkins LS Bermudez LE 1997 Interaction of Mycobacterium avium with environmental amoebae enhances virulence Infect Immun 65 3759ndash3767
Cleary GP Corner LAL OrsquoKeeffe J Marples NM 2011 Diet of the European badger (Meles meles) in the Republic of Ireland A comparison of results from an analysis of stomach contents and rectal faeces Mamm Biol 76 470ndash475
Clifton-Hadley RS Wilesmith JW Stuart F A 1993 Mycobacterium bovis in the European badger (Meles meles) epidemiological findings in tuberculous badgers from a naturally infected population Epidemiol Infect 111 9ndash19
Cole ST Brosch R Parkhill J Garnier T Churcher C Harris D Gordon S V Eiglmeier K Gas S Barry CE Tekaia F Badcock K Basham D Brown D Chillingworth T Connor R Davies R Devlin K Feltwell T Gentles S Hamlin N Holroyd S Hornsby T Jagels K Krogh A McLean J Moule S Murphy L Oliver K Osborne J Quail MA Rajandream MA Rogers J Rutter S Seeger K Skelton J Squares R Squares S Sulston JE Taylor K Whitehead S Barrell BG 1998 Deciphering the biology of Mycobacterium tuberculosis from the complete genome sequence Nature 396 190
Coleman JD Cooke MM 2001 Mycobacterium bovis infection in wildlife in New Zealand Tuberculosis (Edinb) 81 191ndash202
Collins CH Grange JM 1983 The bovine tubercle bacillus J Appl Bacteriol 55 13ndash29
Cooper SM Scott HM de la Garza GR Deck AL Cathey JC 2010 Distribution and interspecies contact of feral Swine and cattle on rangeland in South Texas implications for disease transmission J Wildl Dis 46 152ndash164
Reacutefeacuterences bibliographiques
197
Corner LAL 2006 The role of wild animal populations in the epidemiology of tuberculosis in domestic animals How to assess the risk Vet Microbiol 112 303ndash312
Corner LAL Costello E Lesellier S OrsquoMeara D Sleeman DP Gormley E 2007 Experimental tuberculosis in the European badger (Meles meles) after endobronchial inoculation of Mycobacterium bovis I Pathology and bacteriology Res Vet Sci 83 53ndash62
Corner LAL Murphy D Gormley E 2011 Mycobacterium bovis infection in the Eurasian badger (Meles meles) the disease pathogenesis epidemiology and control J Comp Pathol 144 1ndash24
Corner LAL OrsquoMeara D Costello E Lesellier S Gormley E 2012 The distribution of Mycobacterium bovis infection in naturally infected badgers Vet J 194 166ndash172
Coscolla M Gagneux S 2014 Consequences of genomic diversity in Mycobacterium tuberculosis Semin Immunol 26 431ndash444
Coscolla M Lewin A Metzger S Maetz-Rennsing K Calvignac-Spencer S Nitsche A Dabrowski PW Radonic A Niemann S Parkhill J Couacy-Hymann E Feldman J Comas I Boesch C Gagneux S Leendertz FH 2013 Novel Mycobacterium tuberculosis complex isolate from a wild chimpanzee Emerg Infect Dis 19 969ndash976
Cosivi O Grange JM Daborn CJ Raviglione MC Fujikura T Cousins D Robinson R A Huchzermeyer HF de Kantor I Meslin FX 1999 Zoonotic tuberculosis due to Mycobacterium bovis in developing countries Emerg Infect Dis 4 59ndash70
Coulombe F Divangahi M Veyrier F de Leacuteseacuteleuc L Gleason JL Yang Y Kelliher MA Pandey AK Sassetti CM Reed MB Behr M 2009 Increased NOD2-mediated recognition of N-glycolyl muramyl dipeptide J Exp Med 206 1709ndash1716
Courcoul A Moyen J-L Brugegravere L Faye S Heacutenault S Gares H Boschiroli ML 2014 Estimation of sensitivity and specificity of bacteriology histopathology and PCR for the confirmatory diagnosis of bovine tuberculosis using latent class analysis PLoS One 9 e90334
Courtenay O Reilly L A Sweeney FP Hibberd V Bryan S Ul-Hassan A Newman C Macdonald DW Delahay RJ Wilson GJ Wellington EMH 2006 Is Mycobacterium bovis in the environment important for the persistence of bovine tuberculosis Biol Lett 2 460ndash462
Cousins D V Bastida R Cataldi A Quse V Redrobe S Dow S Duignan P Murray A Dupont C Ahmed N Collins DM Butler WR Dawson D Rodriacuteguez D Loureiro J Romano MI Alito A Zumarraga M Bernardelli A 2003 Tuberculosis in seals caused by a novel member of the Mycobacterium tuberculosis complex Mycobacterium pinnipedii sp nov Int J Syst Evol Microbiol 53 1305ndash1314
Couvin D Rastogi N 2014 La mise en place de bases de donneacutees des geacutenotypes circulants du complexe Mycobacterium tuberculosis et des outils web permettant de mieux surveiller comprendre et controcircler lrsquoeacutepideacutemie de la tuberculose dans le monde EuroReference Cahier num 12
Cowie CE Hutchings MR Barasona JA Gortaacutezar C Vicente J White PCL 2015 Interactions between four species in a complex wildlife livestock disease community implications for Mycobacterium bovis maintenance and transmission Eur J Wildl Res62(1) 51-64
Cvetnic Z Katalinic-Jankovic V Sostaric B Spicic S Obrovac M Marjanovic S Benic M Kirin BK Vickovic I 2007 Mycobacterium caprae in cattle and humans in Croatia Int J Tuberc Lung Dis 11 652ndash658
Dale JW Brittain D Cataldi AA Cousins D Crawford JT Driscoll J Heersma H Lillebaek T Quitugua T Rastogi N Skuce RA Sola C Van Soolingen D Vincent V 2001 Spacer oligonucleotide typing of bacteria of the Mycobacterium tuberculosis complex Recommendations for standardised nomenclature
Reacutefeacuterences bibliographiques
198
Int J Tuberc Lung Dis 5 216ndash219
Dalley D Daveacute D Lesellier S Palmer S Crawshaw T Hewinson RG Chambers M 2008 Development and evaluation of a gamma-interferon assay for tuberculosis in badgers (Meles meles) Tuberculosis 88 235ndash243
Danelishvili L Wu M Stang B Harriff M Cirillo SLG Cirillo JD Bildfell R Arbogast B Bermudez LE 2007 Identification of Mycobacterium avium pathogenicity island important for macrophage and amoeba infection Proc Natl Acad Sci U S A 104 11038ndash11043
Dang NA Kuijper S Walters E Claassens M van Soolingen D Vivo-Truyols G Janssen H-G Kolk AHJ 2013 Validation of biomarkers for distinguishing Mycobacterium tuberculosis from non-tuberculous mycobacteria using gas chromatography-mass spectrometry and chemometrics PLoS One 8 e76263
Daniel TM 2006 The history of tuberculosis Respir Med 100 1862ndash1870
David H Levy Frebaut U Thorel MF 1989 Meacutethodes de laboratoire pour mycobacteacuteriologie clinique Institut Pasteur p 35-36
Dawson KL Bell A Kawakami RP Coley K Yates G Collins DM 2012 Transmission of Mycobacterium orygis (M tuberculosis complex species) from a tuberculosis patient to a dairy cow in New Zealand J Clin Microbiol 50 3136ndash3138
de Jong BC Antonio M Gagneux S 2010 Mycobacterium africanum-review of an important cause of human tuberculosis in West Africa PLoS Negl Trop Dis 4(9) e744
de la Rua-Domenech R 2006 Human Mycobacterium bovis infection in the United Kingdom Incidence risks control measures and review of the zoonotic aspects of bovine tuberculosis Tuberculosis (Edinb) 86 77ndash109
de la Rua-Domenech R Goodchild AT Vordermeier HM Hewinson RG Christiansen KH Clifton-Hadley RS 2006 Ante mortem diagnosis of tuberculosis in cattle a review of the tuberculin tests gamma-interferon assay and other ancillary diagnostic techniques Res Vet Sci 81 190ndash210
Dean GS Rhodes SG Coad M Adam O Cockle PJ Clifford DJ Hewinson RG Vordermeier HM 2005 Minimum infective dose of Mycobacterium bovis in cattle Infect Immun 73 6467ndash6471
Delahay RJ Cheeseman CL Clifton-Hadley RS 2001 Wildlife disease reservoirs the epidemiology of Mycobacterium bovis infection in the European badger (Meles meles) and other British mammals Tuberculosis (Edinb) 81 43ndash49
Delahay RJ De Leeuw ANS Barlow AM Clifton-Hadley RS Cheeseman CL 2002 The status of Mycobacterium bovis Infection in UK Wild Mammals A Review Vet J 164 90ndash105
Delahay RJ Langton S Smith GC Clifton-Hadley RS Cheeseman CL 2000 The spatio-temporal distribution of Mycobacterium bovis (bovine tuberculosis) infection in a high-density badger population J Anim Ecol 69 428ndash441
Delahay RJ Walker N Smith G Wilkinson D Clifton-Hadley R Cheeseman C Tomlinson A Chambers M 2013 Long-term temporal trends and estimated transmission rates for Mycobacterium bovis infection in an undisturbed high-density badger (Meles meles) population Epidemiol Infect 141 1445ndash1456
Delaune D Janvier F Rapp C Geacuterocircme P Fabre M Soler C Meacuterens A 2012 Actualiteacutes de lrsquoinfection agrave Mycobacterium bovis en France agrave propos de 4 cas Ann Biol Clin (Paris) 70 231ndash236
Desikan S Narayanan S 2015 Genetic markers genotyping methods amp next generation sequencing in Mycobacterium tuberculosis Indian J Med Res 141 761ndash774
Reacutefeacuterences bibliographiques
199
Desmond E Ahmed AT Probert WS Ely J Jang Y Sanders C A Lin SY Flood J 2004 Mycobacterium africanum Cases California Emerg Infect Dis 10 921ndash923
Dewes HF 1996 The rate of soil ingestion by dairy cows and the effect on available copper calcium sodium and magnesium N Z Vet J 44 199ndash200
Dick T Lee BH Murugasu-Oei B 1998 Oxygen depletion induced dormancy in Mycobacterium smegmatis FEMS Microbiol Lett 163 159ndash164
Dippenaar A Parsons SDC Sampson SL van der Merwe RG Drewe JA Abdallah AM Siame KK Gey van Pittius NC van Helden PD Pain A Warren RM 2015 Whole genome sequence analysis of Mycobacterium suricattae Tuberculosis 95(6) 682-688
Djelouadji Z Raoult D Drancourt M 2011 Palaeogenomics of Mycobacterium tuberculosis Epidemic bursts with a degrading genome Lancet Infect Dis 11 641ndash650
Dondo A Zoppi S Rossi F Chiavacci L Barbaro A Garrone A Benedetto A Goria M 2007 Mycobacteriosis in Wild Boar Results of 2000-2006 Activity in North-Western Italy Epideacutemiologie santeacute Anim 51 35ndash42
Drancourt M Adeacutekambi T Raoult D 2007 Interactions between Mycobacterium xenopi amoeba and human cells J Hosp Infect 65 138ndash142
Drewe JA OrsquoConnor HM Weber N McDonald RA Delahay RJ 2013 Patterns of direct and indirect contact between cattle and badgers naturally infected with tuberculosis Epidemiol Infect 141 1467ndash1475
Drewe JA Tomlinson AJ Walker NJ Delahay RJ 2010 Diagnostic accuracy and optimal use of three tests for tuberculosis in live badgers PLoS One 5 e11196
Duffield BJ Young DA 1985 Survival of Mycobacterium bovis in defined environmental conditions Vet Microbiol 10 193ndash197
Dutau G 2005 Petite histoire illustreacutee de la tuberculose The history of tuberculosis Arch Pediatr 12 88ndash95
Dziadek J Sajduda A Boruń M 2001 Specificity of insertion sequence-based PCR assays for Mycobacterium tuberculosis complex Int J Tuberc Lung Dis 5 569ndash574
EFSA 2014 The European Union summary report on trends ans sources of zoonotic agents and food-borne outbreaks in 2012 EFSA J 12 169
El Helou G Viola GM Hachem R Han XY Raad II 2013 Rapidly growing mycobacterial bloodstream infections Lancet Infect Dis 13 166ndash174
El Kheacutechine A Couderc C Flaudrops C Raoult D Drancourt M 2011 Matrix-assisted laser desorptionionization time-of-flight mass spectrometry identification of mycobacteria in routine clinical practice PLoS One 6 e24720
El Kheacutechine A Henry M Raoult D Drancourt M 2009 Detection of Mycobacterium tuberculosis complex organisms in the stools of patients with pulmonary tuberculosis Microbiology 155 2384ndash2389
Elliott GN Hough RL Avery LM Maltin C A Campbell CD 2014 Environmental risk factors in the incidence of Johnersquos disease Crit Rev Microbiol 7828 1ndash20
Evans JT Smith EG Banerjee A Smith RMM Dale J Innes JA Hunt D Tweddell A Wood A Anderson C Hewinson RG Smith NH Hawkey PM Sonnenberg P 2007 Cluster of human tuberculosis caused by Mycobacterium bovis evidence for person-to-person transmission in the UK Lancet 369 1270ndash1276
Reacutefeacuterences bibliographiques
200
Fabre M Koeck JL Le Flegraveche P Simon F Herveacute V Vergnaud G Pourcel C 2004 High genetic diversity revealed by variable-number tandem repeat genotyping and analysis of hsp65 gene polymorphism in a large collection of ldquoMycobacterium canettiirdquo strains indicates that the M tuberculosis complex is a recently emerged clone of M can J Clin Microbiol 42 3248ndash3255
Falkinham JO 2016 Nontuberculous mycobacteria community and nosocomial waterborne opportunistic pathogens Clin Microbiol Newsl 38 1ndash7
Falkinham JO Norton CD Mark W 2001 Factors influencing numbers of Mycobacterium avium Mycobacterium intracellulare and other Mycobacteria in drinking water distribution systems Appl Environ Microbiol 67 1225ndash1231
Fanning A Edwards S 1991 Mycobacterium bovis infection in human beings in contact with elk (Cervus elaphus) in Alberta Canada Lancet 338 1253ndash1255
Fediaevsky A Beacutenet J-J Boschiroli ML Riviegravere J Hars J 2012 La tuberculose bovine en France en 2011 poursuite de la reacuteduction du nombre de foyers Bull eacutepideacutemiologique santeacute Anim Aliment Novembre 4ndash12
Fediaevsky A Courcoul A Boschiroli ML Reveillaud E 2014 Tuberculose bovine en France reacutesultats drsquoune strateacutegie plus offensive Bull eacutepideacutemiologique santeacute Anim Aliment 64 4ndash11
Fediaevsky A Courcoul A Boschiroli ML Reveillaud E 2013 Tuberculose bovine en France des signaux favorables mais une situation toujours complexe dans certaines zones Bull Epideacutemiologique - Santeacute Anim Aliment 59 4ndash10
Fetene T Kebede N Alem G 2011 Tuberculosis infection in animal and human populations in three districts of Western Gojam Ethiopia Zoonoses Public Health 58 47ndash53
Fifis T Corner LAL Rothel JS Wood PR 1994 Cellular and humoral immune responses of cattle to purified Mycobacterium bovis antigens Scand J Immunol 39 267ndash274
Fine AE Bolin CA Gardiner JC Kaneene JB 2011a A study of the persistence of Mycobacterium bovis in the environment under natural weather conditions in Michigan USA Vet Med Int 2011 ID 765430 1-12
Fine AE OrsquoBrien DJ Winterstein SR Kaneene JB 2011b An effort to isolate Mycobacterium bovis from environmental substrates during investigations of bovine tuberculosis transmission sites (cattle farms and wildlife areas) in Michigan USA ISRN Vet Sci 2011 ID 787181 1-11
Fischer OA Matlova L Bartl J Dvorska L Svastova P Du Maine R Melicharek I Bartos M Pavlik I 2003 Earthworms (Oligochaeta Lumbricidae) and mycobacteria Vet Microbiol 91 325ndash338
Flores Valdez MA Schoolnik GK 2010 DosR-regulon genes induction in Mycobacterium bovis BCG under aerobic conditions Tuberculosis (Edinb) 90 197ndash200
Forrellad M Klepp L Gioffreacute A 2013 Virulence factors of the Mycobacterium tuberculosis complex Virulence 4 3ndash66
Frank W Reisinger EC Brandt-Hamerla W Schwede I Handrick W 2009 Mycobacterium microti--pulmonary tuberculosis in an immunocompetent patient Wien Klin Wochenschr 121 282ndash6
Frottier J Eliaszewicz M Arlet V Gaudillat C 1990 Infections caused by Mycobacterium africanum Bull Acad Natl Med 174 29ndash35
Gallagher J Clifton-Hadley RS 2000 Tuberculosis in badgers a review of the disease and its significance for other animals Res Vet Sci 69 203ndash217
Reacutefeacuterences bibliographiques
201
Gallagher J Horwill DM 1977 A selective oleic acid albumin agar medium for the cultivation of Mycobacterium bovis J Hyg (Lond) 79 155ndash160
Gallagher J Monies R Gavier-Widen M Rule B 1998 Role of infected non-diseased badgers in the pathogenesis of tuberculosis in the badger Vet Rec 142 710ndash714
Garbaccio S Macias A Shimizu E Paolicchi F Pezzone N Magnano G Zapata L Abdala A Tarabla H Peyru M Caimi K Zumaacuterraga M Canal A Cataldi A 2014 Association between spoligotype-VNTR types and virulence of Mycobacterium bovis in cattle Virulence 5 297ndash302
Garbaccio SG Cataldi A 2010 Evaluation of an immunomagnetic capture method followed by PCR to detect Mycobacterium bovis in tissue samples from cattle Rev Argent Microbiol 42 247ndash53
Garnett BT Delahay RJ Roper TJ 2002 Use of cattle farm resources by badgers (Meles meles) and risk of bovine tuberculosis (Mycobacterium bovis) transmission to cattle Proc Biol Sci 269 1487ndash1491
Garnier T Eiglmeier K Camus J-C Medina N Mansoor H Pryor M Duthoy S Grondin S Lacroix C Monsempe C Simon S Harris B Atkin R Doggett J Mayes R Keating L Wheeler PR Parkhill J Barrell BG Cole ST Gordon S V Hewinson RG 2003 The complete genome sequence of Mycobacterium bovis Proc Natl Acad Sci U S A 100 7877ndash7882
Gavier-Widen D Chambers MA Palmer N Newell DG Hewinson RG 2001 Pathology of natural Mycobacterium bovis infection in European badgers (Meles meles) and its relationship with bacterial excretion Vet Rec 148 299ndash304
Gavier-Wideacuten D Cooke MM Gallagher J Chambers MA Gortaacutezar C 2009 A review of infection of wildlife hosts with Mycobacterium bovis and the diagnostic difficulties of the ldquono visible lesionrdquo presentation N Z Vet J 57 122ndash131
Genov I 1965 The effects of certain physical and chemical agents on Mycobacterium tuberculosis Veteriaacuternı Med Nauk Sofia 2 97ndash107
Gerasimova A Kazakov AE Arkin AP Dubchak I Gelfand MS 2011 Comparative genomics of the dormancy regulons in mycobacteria J Bacteriol 193 3446ndash3452
Ghodbane R Mba Medie F Lepidi H Nappez C Drancourt M 2014 Long-term survival of tuberculosis complex mycobacteria in soil Microbiology 160 496ndash501
Ghosh J Larsson P Singh B Pettersson BMF Islam NM Sarkar SN Dasgupta S Kirsebom LA 2009 Sporulation in mycobacteria Proc Natl Acad Sci U S A 106 10781ndash10786
Goodchild A V Clifton-Hadley RS 2001 Cattle-to-cattle transmission of Mycobacterium bovis Tuberculosis (Edinb) 81 23ndash41
Gordon S V Heym B Parkhill J Barrell B Cole ST 1999 New insertion sequences and a novel repeated sequence in the genome of Mycobacterium tuberculosis H37Rv Microbiology 145 Pt 4 881ndash892
Gormley E 1999 Detection of Mycobacterium bovis lymphocyte stimulating antigens in culture filtrates of a recombinant Mycobacterium smegmatis cosmid library Vaccine 17 2792ndash2801
Gormley E Doyle MB Fitzsimons T McGill K Collins JD 2006 Diagnosis of Mycobacterium bovis infection in cattle by use of the gamma-interferon (Bovigam) assay Vet Microbiol 112 171ndash179
Gortaacutezar C Delahay RJ McDonald RA Boadella M Wilson GJ Gavier-Wideacuten D Acevedo P 2012 The status of tuberculosis in European wild mammals Mamm Rev 42 193ndash206
Gortaacutezar C Torres MJ Vicente J Acevedo P Reglero M de la Fuente J Negro JJ Aznar-Martiacuten J
Reacutefeacuterences bibliographiques
202
2008 Bovine tuberculosis in Dontildeana Biosphere Reserve The role of wild ungulates as disease reservoirs in the last Iberian lynx strongholds PLoS One 3(7) e2776
Grange JM 2001 Mycobacterium bovis infection in human beings Tuberculosis (Edinb) 81 71ndash77
Grange JM Yates MD 1994 Zoonotic aspects of Mycobacterium bovis infection Vet Microbiol 40 137ndash151
Grange JM Yates MD 1989 Incidence and nature of human tuberculosis due to Mycobacterium africanum in South-East England 1977-1987 Epidemiol Infect 103 127ndash132
Greub G Raoult D 2004 Microorganisms resistant to free-living amoebae Clin Microbiol Rev 17 413ndash433
Griffin JFT Rodgers CR Liggett S Mackintosh CG 2006 Tuberculosis in ruminants Characteristics of intra-tonsilar Mycobacterium bovis infection models in cattle and deer Tuberculosis 86 404ndash418
Griffin JM Martin SW Thorburn MA Eves JA Hammond RF 1996 A case-control study on the association of selected risk factors with the occurrence of bovine tuberculosis in the Republic of Ireland Prev Vet Med 27 217ndash229
Guillet-Caruba C Martinez V Doucet-Populaire F 2014 The new tools of microbiological diagnosis of tuberculosis La Rev meacutedecine interne 35 794ndash800
Gunn-Moore DA McFarland SE Brewer JI Crawshaw TR Clifton-Hadley RS Kovalik M Shaw DJ 2011 Mycobacterial disease in cats in Great Britain I Culture results geographical distribution and clinical presentation of 339 cases J Feline Med Surg 13 934ndash944
Gunn-Moore DA Jenkins PA Lucke VM 1996 Feline tuberculosis a literature review and discussion of 19 cases caused by an unusual mycobacterial variant Vet Rec 138 53ndash58
Gutierrez MC Brisse S Brosch R Fabre M Omaiumls B Marmiesse M Supply P Vincent V 2005 Ancient origin and gene mosaicism of the progenitor of Mycobacterium tuberculosis PLoS Pathog 1(1) e5
Haddad N Ostyn A Karoui C Masselot M Thorel MF Hughes SL Inwald J Hewinson RG Durand B 2001 Spoligotype diversity of Mycobacterium bovis strains isolated in France from 1979 to 2000 J Clin Microbiol 39 3623ndash3632
Harris NB Payeur J Bravo D Osorio R Stuber T Farrell D Paulson D Treviso S Mikolon A Rodriguez-Lainz A Cernek-Hoskins S Rast R Ginsberg M Kinde H 2007 Recovery of Mycobacterium bovis from soft fresh cheese originating in Mexico Appl Environ Microbiol 73 1025ndash1028
Hars J Richomme C Boschiroli ML 2010 La tuberculose bovine dans la faune sauvage en France Bull Epideacutemiologique - Santeacute Anim Aliment 38 28ndash31
Hauer A Cruz K De Cochard T Godreuil S Karaoui C Henault S Bulach T Banuls A Biet F Boschiroli ML 2015 Genetic evolution of Mycobacterium bovis causing tuberculosis in livestock and wildlife in France since 1978 PLoS One 10(2) e 0117103
Haydon DT Cleaveland S Taylor LH Laurenson MK 2002 Identifying reservoirs of infection A conceptual and practical challenge Emerg Infect Dis 8 1468ndash1473
Hennessee CT Seo JS Alvarez AM Li QX 2009 Polycyclic aromatic hydrocarbon-degrading species isolated from Hawaiian soils Mycobacterium crocinum sp nov Mycobacterium pallens sp nov Mycobacterium rutilum sp nov Mycobacterium rufum sp nov and Mycobacterium aromaticivorans sp nov Int J Syst Evol Microbiol 59 378ndash387
Hines N Payeur JB Hoffman LJ 2006 Comparison of the recovery of Mycobacterium bovis isolates using
Reacutefeacuterences bibliographiques
203
the BACTEC MGIT 960 system BACTEC 460 system and Middlebrook 7H10 and 7H11 solid media J Vet Diagn Invest 18 243ndash250
Hmama Z Pena-Diaz S Sunil J Av-Gay Y 2015 Immunoevasion and immunosuppression of the macrophage by Mycobacterium tuberculosis Immunol Rev 264 220ndash232
Hoefsloot W Van Ingen J Andrejak C Aumlngeby K Bauriaud R Bemer P Beylis N Boeree MJ Cacho J Chihota V Chimara E Churchyard G Cias R Daza R Daley CL Dekhuijzen PNR Domingo D Drobniewski F Esteban J Fauville-Dufaux M Folkvardsen DB Gibbons N Goacutemez-Mampaso E Gonzalez R Hoffmann H Hsueh PR Indra A Jagielski T Jamieson F Jankovic M Jong E Keane J Koh WJ Lange B Leao S Macedo R Mannsaringker T Marras TK Maugein J Milburn HJ Mlinkoacute T Morcillo N Morimoto K Papaventsis D Palenque E Paez-Pentildea M Piersimoni C Polanovaacute M Rastogi N Richter E Ruiz-Serrano MJ Silva A Da Silva MP Simsek H Van Soolingen D Szaboacute N Thomson R Fernandez TT Tortoli E Totten SE Tyrrell G Vasankari T Villar M Walkiewicz R Winthrop KL Wagner D 2013 The geographic diversity of nontuberculous mycobacteria isolated from pulmonary samples An NTM-NET collaborative study Eur Respir J 42 1604ndash1613
Hohn A Lowenstein 1926 Die Kulture des tuberklebacillus zur diagnose der tuberculose Zeut fur BaktI Orig 98 460ndash477
Hongmanee P Stender H Rasmussen OF 2001 Evaluation of a fluorescence in situ hybridization assay for differentiation between tuberculous and nontuberculous Mycobacterium species in smears of Lowenstein-Jensen and Mycobacteria Growth Indicator Tube cultures using peptide nucleic acid prob J Clin Microbiol 39 1032ndash1035
Hormisch D Brost I Kohring G Giffhorn F Kroppenstedt RM Stackebrandt E Faumlrber P 2004 Mycobacterium fluoranthenivorans sp nov a Fluoranthene and Aflatoxin B 1 Degrading Bacterium from Contaminated Soil of a Former Coal Gas Plant Syst Appl Microbiol 27 653ndash660
Huard RC Claudio L Lazzarini DO Butler WR Soolingen D Van Ho JL 2003 PCR-based method to differentiate the subspecies of the Mycobacterium tuberculosis complex on the basis of genomic deletions J Clin Microbiol 41 1637ndash1650
Huard RC Fabre M de Haas P Lazzarini LCO van Soolingen D Cousins D Ho JL 2006 Novel genetic polymorphisms that further delineate the phylogeny of the Mycobacterium tuberculosis complex J Bacteriol 188 4271ndash4287
Humblet MF Boschiroli ML Saegerman C 2009 Classification of worldwide bovine tuberculosis risk factors in cattle A stratified approach Vet Res 40(5) 1-24
Hutchings MR Harris S 1999 Quantifying the risks of TB infection to cattle posed by badger excreta Epidemiol Infect 122 167ndash173
Hutchings MR Harris S 1997 Effects of farm management practices on cattle grazing behaviour and the potential for transmission of bovine tuberculosis from badgers to cattle Vet J 153 149ndash162
Hutchings MR Service KM Harris S 2002 Is population density correlated with faecal and urine scent marking in European badgers (Meles meles) in the UK Mamm Biol 67 286ndash293
Hutchings MR Service KM Harris S 2001 Defecation and urination patterns of badgers Meles meles at low density in south west England- Acta Theriol (Warsz) 46 87ndash96
Huws SA McBain AJ Gilbert P 2005 Protozoan grazing and its impact upon population dynamics in biofilm communities J Appl Microbiol 98 238ndash244
Idigoras P Peacuterez-Trallero E Alcorta M Gutieacuterrez C Muntildeoz-Baroja I 1995 Rapid detection of tuberculous
Reacutefeacuterences bibliographiques
204
and non-tuberculous mycobacteria by microscopic observation of growth on Middlebrook 7H11 agar Eur J Clin Microbiol Infect Dis 14 6ndash10
Iivanainen E Martikainen PJ Vaumlaumlnaumlnen PK Vaananen PK Katila M 1999 Environmental factors affecting the occurrence of mycobacteria in brook waters J Appl Microbiol 86 673ndash681
Jackson R De Lisle GW Morris RS 1995 A study of the environmental survival of Mycobacterium bovis on a farm in New Zealand N Z Vet J 43 346ndash352
Jahans K Palmer S Inwald J Brown J Abayakoon S 2004 Isolation of Mycobacterium microti from a male Charolais-Hereford cross Vet Rec 155 373ndash374
Jalava K Jones JA Goodchild T Clifton-Hadley R Mitchell A Story A Watson JM 2007 No increase in human cases of Mycobacterium bovis disease despite resurgence of infections in cattle in the United Kingdom Epidemiol Infect 135 40ndash45
Jenkins AO Cadmus SI Venter EH Pourcel C Hauk Y Vergnaud G Godfroid J 2011 Molecular epidemiology of human and animal tuberculosis in Ibadan Southwestern Nigeria Vet Microbiol 151 139ndash147
Jenkins HE Morrison WI Cox DR Donnelly CA Johnston WT Bourne FJ Clifton-Hadley RS Gettinby G McInerney JP Watkins GH Woodroffe R 2008 The prevalence distribution and severity of detectable pathological lesions in badgers naturally infected with Mycobacterium bovis Epidemiol Infect 136 1350ndash1361
Jensen KE 1954 Presence and destruction of tubercle bacilli in sewage Bull World Health Organ 10 171ndash179
Jimeacutenez BA Hinojoza-Loza E Flores-Valdez MA Prado-Montes de Oca E Allen K Estrada-Chaacutevez C Herrera-Rodriacuteguez S Flores-Fernaacutendez JM Martiacutenez-Velaacutezquez M Hernaacutendez-Gutieacuterrez R Alvarez AH 2013 Expression of non-replicating persistence associated genes of Mycobacterium bovis in lymph nodes from skin test-reactor cattle Microb Pathog 61-62 23ndash28
Johnson L Dean G Rhodes S Hewinson G Vordermeier M Wangoo A 2007 Low-dose Mycobacterium bovis infection in cattle results in pathology indistinguishable from that of high-dose infection Tuberculosis (Edinb) 87 71ndash76
Kaal E Kolk AHJ Kuijper S Janssen H-G 2009 A fast method for the identification of Mycobacterium tuberculosis in sputum and cultures based on thermally assisted hydrolysis and methylation followed by gas chromatography-mass spectrometry J Chromatogr A 1216 6319ndash6325
Kamerbeek J Schouls L Kolk A van Agterveld M van Soolingen D Kuijper S Bunschoten A Molhuizen H Shaw R Goyal M van Embden J 1997 Simultaneous detection and strain differentiation of Mycobacterium tuberculosis for diagnosis and epidemiology J Clin Microbiol 35 907ndash914
Kaprelyants AS Gottschal JC Kell DB 1993 Dormancy in non-sporulating bacteria FEMS Microbiol Rev 104 271ndash285
Kasai H Ezaki T Harayama S 2000 Differentiation of phylogenetically related slowly growing mycobacteria by their gyrB sequences J Clin Microbiol 38(1) 301
Kazda J Pavlik I Falkinham JO Hruska K 2009 The Ecology of Mycobacteria Impact on Animalrsquos and Humans Health Springer Netherlands Dordrecht
Khan NA 2006 Acanthamoeba biology and increasing importance in human health FEMS Microbiol Rev 30 564ndash595
Kiers A Klarenbeek A Mendelts B Van Soolingen D Koeumlter G 2008 Transmission of Mycobacterium
Reacutefeacuterences bibliographiques
205
pinnipedii to humans in a zoo with marine mammals Int J Tuberc Lung Dis 12 1469ndash1473
King HC Murphy A James P Travis E Porter D Hung Y-J Sawyer J Cork J Delahay RJ Gaze W Courtenay O Wellington EM 2015a The variability and seasonality of the environmental reservoir of Mycobacterium bovis shed by wild European badgers Sci Rep 5 12318
King HC Murphy A James P Travis E Porter D Sawyer J Cork J Delahay RJ Gaze W Courtenay O Wellington EM 2015b Performance of a non-invasive test for detecting Mycobacterium bovis shedding in European badger (Meles meles) populations J Clin Microbiol 53(7) 2316-2323
Kirchheimer WF Storrs EE 1971 Attempts to establish the armadillo (Dasypus novemcinctus Linn) as a model for the study of leprosy I Report of lepromatoid leprosy in an experimentally infected armadillo Int J Lepr Other Mycobact Dis 39 693ndash702
Kirschner P Springer B Vogel U Meier A Wrede A Kiekenbeck M Bange F Bottger EC 1993 Genotypic identification of Mycobacteria by nucleic acid sequence determination report of a 2-year experience in a clinical laboratory J Clin Microbiol 31 2882ndash2889
Kleeberg HH 1984 Tuberculose humaine drsquoorigine bovine et santeacute publique Rev Sci Tech 3 33ndash54
Koeck JL Fabre M Simon F Daffeacute M Garnotel Eacute Matan A B Geacuterocircme P Bernatas JJ Buisson Y Pourcel C 2011 Clinical characteristics of the smooth tubercle bacilli ldquoMycobacterium canettiirdquo infection suggest the existence of an environmental reservoir Clin Microbiol Infect 17 1013ndash1019
Krasnow I Wayne LG 1966 Sputum Digestion I The mortality rate of tubercle bacilli in various digestion systems Am J Clin Pathol 45 352ndash355
Krishna-Prasad BN Gupta SK 1978 Preliminary report on engulfment and retention of mycobacteria by trophozoites of axenically grown Acanthamoeba castellanii Douglas Curr Sci 47 245ndash247
Kruuk H 1978 Foraging and spatial organisation of the European badger Meles meles L Behav Ecol Sociobiol 4 75ndash89
Kubica GP Dye WE Cohn ML Middlebrook GSD and D with N-A-L-CH for C of M 1963 Sputum digestion and decontamination with N-acetyl-L-cysteinemdashsodium hydroxide for culture of mycobacteria Am Rev Respir Dis 87 775ndash779
Kubica T Rusch-Gerdes S Niemann S 2003 Mycobacterium bovis subsp caprae Caused One-Third of Human M bovis-Associated Tuberculosis Cases Reported in Germany between 1999 and 2001 J Clin Microbiol 41 3070ndash3077
Kukielka E Barasona JA Cowie CE Drewe J a Gortaacutezar C Cotarelo I Vicente J 2013 Spatial and temporal interactions between livestock and wildlife in South Central Spain assessed by camera traps Prev Vet Med 112 213ndash221
Kulka K Hatfull G Ojha AK 2012 Growth of Mycobacterium tuberculosis biofilms J Vis Exp (60) e3820
Lamont EA Bannantine JP Armieacuten A Ariyakumar DS Sreevatsan S 2012 Identification and characterization of a spore-like morphotype in chronically starved Mycobacterium avium subsp paratuberculosis cultures PLoS One 7 e30648
Laprie C Duboy J Malik R Fyfe J 2013 Feline cutaneous mycobacteriosis a review of clinical pathological and molecular characterization of one case of Mycobacterium microti skin infection and nine cases of feline leprosy syndrome from France and New Caledonia Vet Dermatol 24(6) 561ndash9 e133ndash4
Lavania M Katoch K Sachan P Dubey A Kapoor S Kashyap M Chauhan DS Singh HB Sharma VD Jadhav RS Katoch VM 2006 Detection of Mycobacterium leprae DNA from soil samples by PCR
Reacutefeacuterences bibliographiques
206
targeting RLEP sequences J Commun Dis 38 269ndash273
Le Dantec C Duguet J-P Montiel A Dumoutier N Dubrou S Vincent V 2002 Occurrence of Mycobacteria in Water Treatment Lines and in Water Distribution Systems Appl Environ Microbiol 68 5318ndash5325
Lee H Park H Cho S Bai G Kim S 2000 Species identification of Mycobacteria by PCR-Restriction Fragment Length Polymorphism of the rpoB gene J Clin Microbiol 38 2966ndash2971
Lefmann M Schweickert B Buchholz P Goumlbel UB Ulrichs T Seiler P Theegarten D Moter A 2006 Evaluation of peptide nucleic acid-fluorescence in situ hybridization for identification of clinically relevant mycobacteria in clinical specimens and tissue sections J Clin Microbiol 44 3760ndash3767
Lehtola MJ Torvinen E Miettinen IT Keevil CW 2006 Fluorescence in situ hybridization using peptide nucleic acid probes for rapid detection of Mycobacterium avium subsp avium and Mycobacterium avium subsp paratuberculosis in potable-water biofilms Appl Environ Microbiol 72 848ndash853
Leistikow RL Morton RA Bartek IL Frimpong I Wagner K Voskuil MI 2010 The Mycobacterium tuberculosis dosR regulon assists in metabolic homeostasis and enables rapid recovery from nonrespiring dormancy J Bacteriol 192 1662ndash1670
Leys NM Ryngaert A Bastiaens L Wattiau P Top EM Verstraete W Springael D 2005 Occurrence and community composition of fast-growing Mycobacterium in soils contaminated with polycyclic aromatic hydrocarbons FEMS Microbiol Ecol 51 375ndash388
Li L Bannantine JP Zhang Q Amonsin A May BJ Alt D Banerji N Kanjilal S Kapur V 2005 The complete genome sequence of Mycobacterium avium subspecies paratuberculosis Proc Natl Acad Sci U S A 102 12344ndash12349
Libero G De Mori L 2014 The T-cell response to lipid antigens of Mycobacterium tuberculosis Front Immunol 5 1ndash9
Lieacutebana E Aranaz A Francis B Cousins D 1996 Assessment of genetic markers for species differentiation within the Mycobacterium tuberculosis complex J Clin Microbiol 34 933ndash938
Lieacutebana E Aranaz A Mateos A Vilafranca M Gomez-Mampaso E Tercero JC Alemany J Suarez G Domingo M Dominguez L 1995 Simple and rapid detection of Mycobacterium tuberculosis complex organisms in bovine tissue samples by PCR J Clin Microbiol 33 33ndash36
Lim A Eleuterio M Hutter B Murugasu-Oei B Dick T 1999 Oxygen depletion-induced dormancy in Mycobacterium bovis BCG J Bacteriol 181 2252ndash2256
Liss GM Wong L Kittle DC Simor A Naus M Martiquet P Misener CR 1994 Occupational exposure to Mycobacterium bovis infection in deer and elk in Ontario Can J Public Heal 85 326ndash329
Little TW Naylor PF Wilesmith JW 1982 Laboratory study of Mycobacterium bovis infection in badgers and calves Vet Rec 111 550ndash557
Lomme JR Thoen CO Himes EM Vinson JW King RE 1976 Mycobacterium tuberculosis infection in two East African oryxes J Am Vet Med Assoc 169 912ndash914
Lotz A Ferroni A Beretti J-L Dauphin B Carbonnelle E Guet-Revillet H Veziris N Heym B Jarlier V Gaillard J-L Pierre-Audigier C Frapy E Berche P Nassif X Bille E 2010 Rapid identification of mycobacterial whole cells in solid and liquid culture media by matrix-assisted laser desorption ionization-time of flight mass spectrometry J Clin Microbiol 48 4481ndash4486
Ludwig W Euzeacuteby J Schumann P Busse H-J Trujillo ME Kaumlmpfer P Whitman WB 2008 Road map
Reacutefeacuterences bibliographiques
207
of the phylum Actinobacteria Road Map of the phylum Actinobacteria Bergeys Manual of Systematic Bacteriology second Edition Volume five Springer Editions
Lugton IW Wilson PR Morris RS Nugent G 1998 Epidemiology and pathogenesis of Mycobacterium bovis infection of red deer (Cervus elaphus) in New Zealand N Z Vet J 46 147ndash156
Lyashchenko KP Greenwald R Esfandiari J OrsquoBrien DJ Schmitt SM Palmer M V Waters WR 2013 Rapid detection of serum antibody by dual-path platform VetTB assay in white-tailed deer infected with Mycobacterium bovis Clin Vaccine Immunol 20 907ndash911
Mackintosh CG Qureshi T Waldrup K Labes RE Dodds KG Griffin JFT 2000 Genetic resistance to experimental infection with Mycobacterium bovis in red deer (Cervus elaphus) Infect Immun 68 1620ndash1625
Maddock EC 1936 Experiments on the infectivity for healthy calves of bovine tubercle bacilli discharged in dung upon pasture Part I From tubercular calves fed with emulsions of Tubercle bacilli 1934-5 Part II From tubercular cows passing Tubercle bacilli in their dun J Hyg (Lond) 36 594ndash601
Maddock EC 1934 Further studies on the survival time of the bovine tubercle bacillus in soil soil and dung in dung and on grass with experiments on feeding guinea-pigs and calves on grass artificially infected with bovine tubercle bacilli J Hyg (Lond) 34 372ndash379
Maddock EC 1933 Studies on the survival time of the bovine tubercle bacillus in soil soil and dung in dung and on grass with experiments on the preliminary treatment of infected organic matter and the cultivation of the organism J Hyg (Lond) 33 103ndash117
Magee JG Ward AC 2012 Mycobacteriaceae Bergeys Manual of Systematic Bacteriology second Edition Volume five Springer Editions
Mahairas GG Sabo PJ Hickey MJ Singh DC Stover CK Mahairas GG Sabo PJ Hickey MJ Singh DC Stover CK 1996 Molecular analysis of genetic differences between Mycobacterium bovis BCG and virulent M bovis J Bacteriol 178 1274ndash1282
Mardare C Delahay RJ Dale JW 2013 Environmental amoebae do not support the long-term survival of virulent mycobacteria J Appl Microbiol 114 1388ndash1394
Marrakchi H Laneacuteelle MA Daffeacute M 2014 Mycolic acids Structures biosynthesis and beyond Chem Biol 21 67ndash85
Marri PR Bannantine JP Golding GB 2006 Comparative genomics of metabolic pathways in Mycobacterium species gene duplication gene decay and lateral gene transfer FEMS Microbiol Rev 30 906ndash925
Martiacuten-Hernando MP Houmlfle U Vicente J Ruiz-Fons F Vidal D Barral M Garrido JM de la Fuente J Gort 2007 Lesions associated with Mycobacterium tuberculosis complex infection in the European wild boar Tuberculosis 87 360ndash367
Mathema B Kurepina NE Bifani PJ Kreiswirth BN 2006 Molecular epidemiology of tuberculosis Current insights Clin Microbiol Rev 19 658ndash685
Matsuoka M Izumi S Budiawan T Nakata N Saeki K 1999 Mycobacterium leprae DNA in daily using water as a possible source of leprosy infection Indian J Lepr 71 61ndash67
Mba Medie F Ben Salah I Henrissat B Raoult D Drancourt M 2011 Mycobacterium tuberculosis complex mycobacteria as amoeba-resistant organisms PLoS One 6 e20499
McCallan L Mcnair J Skuce R 2014 A review of the potential role of cattle slurry in the spread of bovine
Reacutefeacuterences bibliographiques
208
tuberculosis 1ndash48
McCorry T Whelan AO Welsh MD McNair J Walton E Bryson DG Hewinson RG Vordermeier HM Pollock JM 2005 Shedding of Mycobacterium bovis in the nasal mucus of cattle infected experimentally with tuberculosis by the intranasal and intratracheal routes Vet Rec 157 613ndash618
McLaughlin AM Gibbons N Fitzgibbon M Power JT Foley SC Hayes JP Rogers T Keane J 2012 Primary isoniazid resistance in Mycobacterium bovis disease A prospect of concern Am J Respir Crit Care Med 186 110ndash111
McNabb A Eisler D Adie K Amos M Rodrigues M Stephens G Black WA Isaac-Renton J 2004 Assessment of partial sequencing of the 65-kilodalton heat shock protein gene (hsp65) for routine identification of Mycobacterium species isolated from clinical sources J Clin Microbiol 42 3000ndash3011
Mejia GI Castrillon L Trujillo H Robledo J A 1999 Microcolony detection in 7H11 thin layer culture is an alternative for rapid diagnosis of Mycobacterium tuberculosis infection Int J Tuberc Lung Dis 3 138ndash142
Menzies FD Neill SD 2000 Cattle-to-Cattle Transmission of Bovine Tuberculosis Vet J 160 92ndash106
Meredith A Del Pozo J Smith S Milne E Stevenson K McLuckie J 2014 Leprosy in red squirrels in Scotland Vet Rec 175 285ndash286
Michel AL Muumlller B van Helden PD 2010 Mycobacterium bovis at the animal-human interface a problem or not Vet Microbiol 140 371ndash381
Michel AL Venter L Espie IW Coetzee ML 2003 Mycobacterium tuberculosis infections in eight species at the national zoological gardens of South Africa 1991-2001 J Zoo Wildl Med 34 364ndash370
Michelet L de Cruz K Zanella G Aaziz R Bulach T Karoui C Heacutenault S Joncour G Boschiroli ML 2015 Infection with Mycobacterium microti in animals in France J Clin Microbiol 53 981ndash985
Mignard S Pichat C Carret G 2006 Mycobacterium bovis infection Lyon France Emerg Infect Dis 12 1431ndash1433
Miltgen J Morillon M Koeck J-L Varnerot A Briant J-F Nguyen G Verrot D Bonnet D Vincent V 2002 Two cases of pulmonary tuberculosis caused by Mycobacterium tuberculosis subsp canetti Emerg Infect Dis 8 1350ndash1352
Miltner EC Bermudez LE 2000 Mycobacterium avium grown in Acanthamoeba castellanii is protected from the effects of antimicrobials Antimicrob Agents Chemother 44 1990ndash1994
Mitscherlich E Marth EH 2012 Microbial survival in the environment Bacteria and rickettsiae important in human and animal health Springer S ed
Momotani E Ozaki H Hori M Yamamoto S Kuribayashi T Eda S Ikegami M 2012 Mycobacterium avium subsp paratuberculosis lipophilic antigen causes Crohnrsquos disease-type necrotizing colitis in mice Springerplus 1 1ndash10
Moore J Roper T 2003 Temperature and humidity in badger Meles meles setts Mamm Rev 33 308ndash313
Morris RS Pfeiffer DU Jackson R 1994 The epidemiology of Mycobacterium bovis infections Vet Microbiol 40 153ndash177
Mostafa HM Kazda J Irgens LM Luesse HG 1995 Acid-fast bacilli from former leprosy regions in coastal Norway showing PCR positivity for Mycobacterium leprae Int J Lepr other Mycobact Dis 63 97ndash99
Mukhopadhyay S Nair S Ghosh S 2012 Pathogenesis in tuberculosis Transcriptomic approaches to
Reacutefeacuterences bibliographiques
209
unraveling virulence mechanisms and finding new drug targets FEMS Microbiol Rev 36 463ndash485
Muumlller B Duumlrr S Alonso S Hattendorf J Laisse CJM Parsons SDC van Helden PD Zinsstag J 2013 Zoonotic Mycobacterium bovis ndashinduced tuberculosis in humans Emerg Infect Dis 19 899ndash908
Muumlller B Hilty M Berg S Garcia-Pelayo MC Dale J Boschiroli ML Cadmus SI Ngandolo BNR Godreuil S Diguimbaye-Djaibeacute C Kazwala R Bonfoh B Njanpop-Lafourcade BM Sahraoui N Guetarni D Aseffa A Mekonnen MH Razanamparany VR Ramarokoto H Djoslashnne B Oloya J Machado A Mucavele C Skjerve E Portaels F Rigouts L Michel A Muumlller A Kaumlllenius G Van Helden PD Hewinson RG Zinsstag J Gordon S V Smith NH 2009 African 1 an epidemiologically important clonal complex of Mycobacterium bovis dominant in Mali Nigeria Cameroon and Chad J Bacteriol 191 1951ndash1960
Mura M Bull T 2006 Replication and long-term persistence of bovine and human strains of Mycobacterium avium subsp paratuberculosis within Acanthamoeba polyphaga Appl Environ Microbiol 72 854ndash859
Murphy D Gormley E Costello E OrsquoMeara D Corner LAL 2010 The prevalence and distribution of Mycobacterium bovis infection in European badgers (Meles meles) as determined by enhanced post mortem examination and bacteriological culture Res Vet Sci 88 1ndash5
Musser JM Amin A Ramaswamy S 2000 Negligible genetic diversity of Mycobacterium tuberculosis host immune system protein targets Evidence of limited selective pressure Genetics 155 7ndash16
Namouchi A Didelot X Schoumlck U Gicquel B Rocha EPC 2012 After the bottleneck Genome-wide diversification of the Mycobacterium tuberculosis complex by mutation recombination and natural selection Genome Res 22 721ndash734
Narang R Narang P Mendiratta DK 2009 Isolation and identification of nontuberculous mycobacteria from water and soil in central India Indian J Med Microbiol 27 247ndash250
Naranjo V Gortaacutezar C Vicente J de la Fuente J 2008 Evidence of the role of European wild boar as a reservoir of Mycobacterium tuberculosis complex Vet Microbiol 127 1ndash9
Navarro Y Romero B Bouza E Domiacutenguez L Juan L de Garciacutea-de-Viedma D 2015 Detailed chronological analysis of microevolution events in herds infected persistently by Mycobacterium bovis Vet Microbiol 183 97-102
Neill SD Hanna J OrsquoBrien J McCracken R 1988 Excretion of Mycobacterium bovis by experimentally infected cattle Vet Rec 123 340ndash343
Neill SD Pollock JM Bryson DB Hanna J 1994 Pathogenesis of Mycobacterium bovis infection in cattle Vet Microbiol 40 41ndash52
Niemann S Richter E Daluumlgge-Tamm H Schlesinger H Graupner D Koumlnigstein B Gurath G Greinert U Ruumlsch-Gerdes S 2000 Two cases of Mycobacterium microti-derived tuberculosis in HIV-negative immunocompetent patients Emerg Infect Dis 6 539ndash542
Nishi JS Shury T Elkin BT 2006 Wildlife reservoirs for bovine tuberculosis (Mycobacterium bovis) in Canada Strategies for management and research Vet Microbiol 112 325ndash338
Norby B Fosgate GT Manning EJB Collins MT Roussel AJ 2007 Environmental mycobacteria in soil and water on beef ranches Association between presence of cultivable mycobacteria and soil and water physicochemical characteristics Vet Microbiol 124 153ndash159
Nugent G 2011 Maintenance spillover and spillback transmission of bovine tuberculosis in multi-host wildlife complexes a New Zealand case study Vet Microbiol 151 34ndash42
Reacutefeacuterences bibliographiques
210
OrsquoBrien DJ Schmitt SM Fitzgerald SD Berry DE Hickling GJ 2006 Managing the wildlife reservoir of Mycobacterium bovis the Michigan USA experience Vet Microbiol 112 313ndash323
OrsquoReilly LM Daborn CJ 1995 The epidemiology of Mycobacterium bovis infections in animals and man a review Tuber Lung Dis 76 Suppl 1 1ndash46
OIE 2009 Bovine Tuberculosis Terrestrial Manual 2009Chap 247
Ojha AK Baughn AD Sambandan D Hsu T Trivelli X Guerardel Y Alahari A Kremer L Jacobs WR Hatfull GF 2008 Growth of Mycobacterium tuberculosis biofilms containing free mycolic acids and harbouring drug-tolerant bacteria Mol Microbiol 69 164ndash174
Oloya J Kazwala R Lund A Opuda-Asibo J Demelash B Skjerve E Johansen TB Djoslashnne B 2007 Characterisation of mycobacteria isolated from slaughter cattle in pastoral regions of Uganda BMC Microbiol 7 95
Otal I Isabel O Gomez AB Kremer K de Haas P Garciacutea MJ Martiacuten C van Soolingen D 2008 Mapping of IS6110 insertion sites in Mycobacterium bovis isolates in relation to adaptation from the animal to human host Vet Microbiol 129 333ndash341
Pagnier I Raoult D La Scola B 2008 Isolation and identification of amoeba-resisting bacteria from water in human environment by using an Acanthamoeba polyphaga co-culture procedure Environ Microbiol 10 1135ndash1144
Palgrave CJ Benato L Eatwell K Laurenson IF Smith NH 2012 Mycobacterium microti infection in two meerkats (Suricata suricatta) J Comp Pathol 146 278ndash282
Palmer M V 2013 Mycobacterium bovis Characteristics of wildlife reservoir hosts Transbound Emerg Dis 60 1ndash13
Palmer M V C Thacker T Waters WR Robbe-Austerman S Harris BN 2010 Investigations on deer to deer and deer to cattle transmission of the vaccine Mycobacterium bovis bacillus Calmette-Gueacuterin (BCG) J Vaccines Vaccin 01 1ndash5
Palmer M V Waters WR Whipple DL 2004 Investigation of the transmission of Mycobacterium bovis from deer to cattle through indirect contact Am J Vet Res 65 1483ndash1489
Palmer M V Waters WR Whipple DL 2002 Lesion development in white-tailed Deer (Odocoileus virginianus) experimentally infected with Mycobacterium bovis Vet Pathol 39 334ndash340
Palmer M V Whipple DL 2006 Survival of Mycobacterium bovis on feedstuffs commonly used as supplemental feed for white-tailed deer (Odocoileus virginianus) J Wildl Dis 42 853ndash8
Palmer M V Whipple DL Rhyan JC Bolin CA Saari DA 1999 Granuloma development in cattle after intratonsilar inoculation with Mycobacterium bovis Am J Vet Res 60 310ndash315
Palmer M V Whipple DL Waters WR 2001 Experimental deer-to-deer transmission of Mycobacterium bovis Am J Vet Res 62 692ndash696
Panteix G Gutierrez MC Boschiroli ML Rouviere M Plaidy A Pressac D Porcheret H Chyderiotis G Ponsada M Van Oortegem K Salloum S Cabuzel S Bantildeuls AL Van de Perre P Godreuil S 2010 Pulmonary tuberculosis due to Mycobacterium microti a study of six recent cases in France J Med Microbiol 59 984ndash989
Parsons SDC Drewe JA van Pittius NCG Warren RM van Helden PD 2013 Novel cause of tuberculosis in meerkats South Africa Emerg Infect Dis 19 2004ndash2007
Reacutefeacuterences bibliographiques
211
Pavlik I Svastova P Bartl J Dvorska L Rychlik I 2000 Relationship between IS901 in the Mycobacterium avium complex strains isolated from birds animals humans and the environment and virulence for poultry Clin Diagn Lab Immunol 7 212ndash217
Pavlik I Trcka I Parmova I Svobodova J Melicharek I Nagy G Cvetnic Z Ocepek M Pate M Lipiec M 2005 Detection of bovine and human tuberculosis in cattle and other animals in six Central European countries during the years 2000 ndash 2004 Vet Med-Czech 2005 291ndash299
Pawlowski A Jansson M Skoumlld M Rottenberg ME Kaumlllenius G 2012 Tuberculosis and HIV co-infection PLoS Pathog 8(2) e1002464
Payne A 2014 Rocircle de la faune sauvage dans le systegraveme multi-hocirctes de Mycobacterium bovis et risque de transmission entre faune sauvage et bovins Etude expeacuterimentale en Cocircte drsquoOr Thegravese Universiteacute Lyon
Payne A Boschiroli ML Gueneau E Moyen JL Rambaud T Dufour B Gilot-Fromont E Hars J 2012 Bovine tuberculosis in ldquoEurasianrdquo badgers (Meles meles) in France Eur J Wildl Res 59 331ndash339
Payne A Chappa S Hars J Dufour B Gilot-Fromont E 2015 Wildlife visits to farm facilities assessed by camera traps in a bovine tuberculosis-infected area in France Eur J Wildl Res 62(1) 33-42
Pepperell CS Casto AM Kitchen A Granka JM Cornejo OE Holmes EC Holmes EC Birren B Galagan J Feldman MW 2013 The role of selection in shaping diversity of natural M tuberculosis populations PLoS Pathog 9 e1003543
Petroff SA 1915 Some cultural studies on the tubercle bacillus Bull Johns Hopkins Hosp 26 276ndash279
Pfyffer GE Auckenthaler R Van Embden JD a Van Soolingen D 1998 Mycobacterium canettii the smooth variant of M tuberculosis isolated from a Swiss patient exposed in Africa Emerg Infect Dis 4 631ndash634
Phillips CJC Foster CRW Morris PA Teverson R 2003 The transmission of Mycobacterium bovis infection to cattle Res Vet Sci 74 1ndash15
Pickup RW Rhodes G Bull TJ Arnott S Sidi-Boumedine K Hurley M Hermon-Taylor J 2006 Mycobacterium avium subsp paratuberculosis in lake catchments in river water abstracted for domestic use and in effluent from domestic sewage treatment works diverse opportunities for environmental cycling and human exposure Appl Environ Microbiol 72 4067ndash4077
Piersimoni C Olivieri A Benacchio L Scarparo C 2006 Current perspectives on drug susceptibility testing of Mycobacterium tuberculosis complex the automated nonradiometric systems J Clin Microbiol 44 20ndash28
Pillai SD Widmer KW Ivey LJ Coker KC Newman E Lingsweiler S Baca D Kelley M Davis DS Silvy NJ Adams LG 2000 Failure to identify non-bovine reservoirs of Mycobacterium bovis in a region with a history of infected dairy-cattle herds Prev Vet Med 43 53ndash62
Pollock JM Neill SD 2002 Mycobacterium bovis infection and tuberculosis in cattle Vet J 163 115ndash127
Pontiroli A Travis ER Sweeney FP Porter D Gaze WH Mason S Hibberd V Holden J Courtenay O Wellington EMH 2011 Pathogen quantitation in complex matrices a multi-operator comparison of DNA extraction methods with a novel assessment of PCR inhibition PLoS One 6 e17916
Prasad HK Singhal A Mishra A Shah NP Katoch VM Thakral SS Singh D V Chumber S Bal S Aggarwal S Padma M V Kumar S Singh MK Acharya SK 2005 Bovine tuberculosis in India potential basis for zoonosis Tuberculosis (Edinb) 85 421ndash428
Primm TP Lucero CA Falkinham JO 2004 Health impacts of environmental mycobacteria Clin Microbiol Rev 17 98ndash106
Reacutefeacuterences bibliographiques
212
Pritchard DG 1988 A Century of Bovine Tuberculosis 1888-1988 Conquest and Controversy J Comp Pathol 99 357ndash399
Prodinger WM Brandsta A Naumann L Pacciarini M Kubica T Boschiroli ML Aranaz A Cvetnic Z Ocepek M Skrypnyk A Erler W Niemann S Pavlik I Moser I 2005 Characterization of Mycobacterium caprae isolates from Europe by Mycobacterial Interspersed Repetitive Unit Genotyping J Clin Microbiol 43 4984ndash4992
Prodinger WM Eigentler A Allerberger F Schonbauer M Glawischnig W 2002 Infection of Red Deer Cattle and Humans with Mycobacterium bovis subsp caprae in Western Austria J Clin Microbiol 40 2270ndash2272
Prodinger WM Indra A Koksalan OK Kilicaslan Z Richter E 2014 Mycobacterium caprae infection in humans Expert Rev Anti Infect Ther12(12) 1501-1513
Pym AS Brodin P Brosch R Huerre M Cole ST 2002 Loss of RD1 contributed to the attenuation of the live tuberculosis vaccines Mycobacterium bovis BCG and Mycobacterium microti Mol Microbiol 46 709ndash717
Radomski N Betelli L Moilleron R Haenn S Moulin L Cambau E Rocher V Gonccedilalves A Lucas FS 2011 Mycobacterium behavior in wastewater treatment plant a bacterial model distinct from Escherichia coli and Enterococci Environ Sci Technol 45 5380ndash5386
Radomski N Roguet A Lucas FS Veyrier FJ Cambau E Accrombessi H Moilleron R Behr M Moulin L 2013 atpE gene as a new useful specific molecular target to quantify Mycobacterium in environmental samples BMC Microbiol 13 277
Radomski N Thibault VC Karoui C de Cruz K Cochard T Gutierrez C Supply P Biet F Boschiroli ML 2010 Determination of genotypic diversity of Mycobacterium avium subspecies from human and animal origins by Mycobacterial Interspersed Repetitive-Unit-Variable-Number Tandem-Repeat and IS1311 Restriction Fragment Length Polymorphism typing methods J Clin Microbiol 48 1026ndash1034
Rahim Z Moumlllers M Te Koppele-Vije A De Beer J Zaman K Matin MA Kamal M Raquib R Van Soolingen D Baqi M a Heilmann FGC Van Der Zanden AGM 2007 Characterization of Mycobacterium africanum subtype i among cows in a dairy farm in Bangladesh using spoligotyping Southeast Asian J Trop Med Public Health 38 706ndash713
Rauzier J Moniz-Pereira J Gicquel-Sanzey B 1988 Complete nucleotide sequence of pAL5OOO a plasmid from Mycobacterium fortuitum Gene 71 315ndash321
Renwick AR White PCL Bengis RG 2007 Bovine tuberculosis in southern African wildlife a multi-species host-pathogen system Epidemiol Infect 135 529ndash540
Reviriego-Gordejo FJ Vermeersch JP 2006 Towards eradication of bovine tuberculosis in the European Union Vet Microbiol 112 101ndash109
Richomme C Boschiroli ML Hars J Casabianca F Ducrot C 2010 Bovine tuberculosis in livestock and wild boar on the Mediterranean island Corsica J Wildl Dis 46 627ndash631
Ringuet H Akoua-Koffi C Honore S Varnerot A Vincent V Berche P Gaillard JL Pierre-Audigier C 1999 hsp65 sequencing for identification of rapidly growing mycobacteria J Clin Microbiol 37 852ndash857
Ritacco V Loacutepez B De Kantor IN Barrera L Errico F Nader A 1991 Reciprocal cellular and humoral immune responses in bovine tuberculosis Res Vet Sci 50 365ndash367
Rivero A Maacuterquez M Santos J Pinedo A Saacutenchez MA Esteve A Samper S Martiacuten C 2001 High rate of tuberculosis reinfection during a nosocomial outbreak of multidrug-resistant tuberculosis caused by
Reacutefeacuterences bibliographiques
213
Mycobacterium bovis strain B Clin Infect Dis 32 159ndash161
Riviegravere J Fediaevsky A Hars J Richomme C Calavas D Hendrikx P 2011 SYLVATUB Dispositif national de surveillance de la tuberculose bovine dans la faune sauvage Organisation institutionnelle et reacuteseau dacteurs 2011ndash2012
Rodriguez JG Burbano CS Nuntildeez C Gonzaacutelez CE Zambrano MM Garciacutea MJ Del Portillo P 2008 Rv3134cdevRdevS operon of Mycobacterium bovis BCG is differentially transcribed under ldquoin vitrordquo stress conditions Tuberculosis 88 273ndash282
Rodriacuteguez S Bezos J Romero B de Juan L Aacutelvarez J Castellanos E Moya N Lozano F Javed MT Saacuteez-Llorente JL Lieacutebana E Mateos A Domiacutenguez L Aranaz A 2011 Mycobacterium caprae infection in livestock and wildlife Spain Emerg Infect Dis 17 532ndash535
Rodriguez-Campos S Schuumlrch AC Dale J Lohan AJ Cunha M V Botelho A Cruz K Boschiroli ML Boniotti MB Pacciarini M Garcia-Pelayo MC Romero B de Juan L Domiacutenguez L Gordon S V van Soolingen D Loftus B Berg S Hewinson RG Aranaz A Smith NH 2012 European 2 - A clonal complex of Mycobacterium bovis dominant in the Iberian Peninsula Infect Genet Evol 12 866ndash872
Rodriguez-Campos S Smith NH Boniotti MB Aranaz A 2014 Overview and phylogeny of Mycobacterium tuberculosis complex organisms Implications for diagnostics and legislation of bovine tuberculosis Res Vet Sci 97 Suppl S5ndashS19
Rodriguez-Nuntildeez J Avelar FJ Marquez F Rivas-Santiago B Quintildeones C Guerrero-Barrera AL 2012 Mycobacterium tuberculosis complex detected by modified fluorescent in situ hybridization in lymph nodes of clinical samples J Infect Dev Ctries 6(1) 58ndash66
Rodriacuteguez-Zaragoza S 1994 Ecology of free-living Amoebae Crit Rev Microbiol 20(3) 225-241
Rogers LM Forrester GJ Wilson GJ Yarnell RW Cheeseman CL 2003 The role of setts in badger (Meles meles) group size breeding success and status of TB (Mycobacterium bovis) J Zool 260 209ndash215
Romero B Rodriguez S Bezos J Diaz R Francisca Copano M Merediz I Minguez O Marques S Palacios JJ De Viedma DG Saez JL Mateos A Aranaz A Dominguez L De Juan L 2011 Humans as source of Mycobacterium tuberculosis infection in cattle Spain Emerg Infect Dis 17 2393ndash2395
Roring S Scott A Brittain D Walker I Hewinson G Neill SD Skuce R 2002 Development of variable-number tandem repeat typing of Mycobacterium bovis comparison of results with those obtained by using existing exact tandem repeats and spoligotyping J Clin Microbiol 40 2126ndash2133
Roth A Fischer M Hamid ME Michalke S Ludwig W Mauch H 1998 Differentiation of phylogenetically related slowly growing mycobacteria based on 16S-23S rRNA gene internal transcribed spacer sequences J Clin Microbiol 36 139ndash147
Rothschild BM Martin LD Lev G Bercovier H Bar-Gal GK Greenblatt C Donoghue H Spigelman M Brittain D 2001 Mycobacterium tuberculosis complex DNA from an extinct bison dated 17000 years before the present Clin Infect Dis 33 305ndash311
Roug A Geoghegan C Wellington E Miller W A Travis E Porter D Cooper D Clifford DL Mazet J AK Parsons S 2014 Utility of a fecal real-time PCR protocol for detection of Mycobacterium bovis infection in African buffalo (Syncerus caffer) J Wildl Dis 50 140ndash142
Rowe MT Grant IR 2006 Mycobacterium avium ssp paratuberculosis and its potential survival tactics Lett Appl Microbiol 42 305ndash11
Safi H Barnes PF Lakey DL Shams H Samten B Vankayalapati R Howard ST 2004 IS6110 functions as a mobile monocyte-activated promoter in Mycobacterium tuberculosis Mol Microbiol 52 999ndash1012
Reacutefeacuterences bibliographiques
214
Saleeb PG Drake SK Murray PR Zelazny AM 2011 Identification of mycobacteria in solid-culture media by matrix-assisted laser desorption ionization-time of flight mass spectrometry J Clin Microbiol 49 1790ndash1794
Santos N Almeida V Gortaacutezar C Correia-Neves M 2015a Patterns of Mycobacterium tuberculosis-complex excretion and characterization of super-shedders in naturally-infected wild boar and red deer Vet Res 46 129
Santos N Santos C Valente T Gortaacutezar C Almeida V Correia-Neves M 2015b Widespread environmental contamination with Mycobacterium tuberculosis Complex revealed by a molecular detection protocol PLoS One 10 e0142079
Savelkoul PHM Catsburg A Mulder S Oostendorp L Schirm J Wilke H van der Zanden AGM Noordhoek GT 2006 Detection of Mycobacterium tuberculosis complex with Real Time PCR Comparison of different primer-probe sets based on the IS6110 element J Microbiol Methods 66 177ndash180
Scanlon MP Quinn PJ 2000a The survival of Mycobacterium bovis in sterilized cattle slurry and its relevance to the persistence of this pathogen in the environment Ir Vet J 53 412ndash415
Scanlon MP Quinn PJ 2000b Inactivation of Mycobacterium bovis in cattle slurry by five volatile chemicals J Appl Microbiol 89 854ndash861
Schnappinger D Ehrt S Voskuil MI Liu Y Mangan JA Monahan IM Dolganov G Efron B Butcher PD Nathan C Schoolnik GK 2003 Transcriptional adaptation of Mycobacterium tuberculosis within macrophages insights into the phagosomal environment J Exp Med 198 693ndash704
Schoepf K Prodinger WM Glawischnig W Hofer E Revilla-Fernandez S Hofrichter J Fritz J Koumlfer J Schmoll F 2012 A two-yearsrsquo survey on the prevalence of tuberculosis caused by Mycobacterium caprae in red deer (Cervus elaphus) in the Tyrol Austria ISRN Vet Sci 2012 245138
Schroumlder KH 1982 Isolation of M africanum from German patients Bull Int Union Tuberc 57 249ndash53
Schuumlrch AC van Soolingen D 2012 DNA fingerprinting of Mycobacterium tuberculosis from phage typing to whole-genome sequencing Infection Genetics and Evolution 12 602-609
Shepard CC 1960 The experimental disease that follows the injection of human leprosy bacilli into foot-pads of mice J Exp Med 112 445ndash454
Shiloh MU Manzanillo P Cox JS 2008 Mycobacterium tuberculosis Senses Host-Derived Carbon Monoxide during Macrophage Infection Cell Host Microbe 3 323ndash330
Shinnick TM Good RC 1994 Mycobacterial taxonomy Eur J Clin Microbiol Infect Dis 13 884ndash901
Shitikov E Ilina E Chernousova L Borovskaya A Rukin I Afanasrsquoev M Smirnova T Vorobyeva A Larionova E Andreevskaya S Kostrzewa M Govorun V 2012 Mass spectrometry based methods for the discrimination and typing of mycobacteria Infect Genet Evol 12 838ndash845
Shrikrishna D de la Rua-Domenech R Smith NH Colloff A Coutts I 2009 Human and canine pulmonary Mycobacterium bovis infection in the same household re-emergence of an old zoonotic threat Thorax 64 89ndash91
Shuralev E Quinn P Doyle M Duignan A Kwok HF Bezos J Olwill SA Gormley E Aranaz A Good M Davis WC Clarke J Whelan C 2012 Application of the Enfer chemiluminescent multiplex ELISA system for the detection of Mycobacterium bovis infection in goats Vet Microbiol 154 292ndash297
Skuce RA McCorry TP McCarroll JF Roring SMM Scott AN Brittain D Hughes SL Hewinson RG Neill SD 2002 Discrimination of Mycobacterium tuberculosis complex bacteria using novel VNTR-PCR
Reacutefeacuterences bibliographiques
215
targets Microbiology 148 519ndash528
Skuce RA Brittain D Hughes MS Beck LA Neill SD 1994 Genomic fingerprinting of Mycobacterium bovis from cattle by restriction fragment length polymorphism analysis J Clin Microbiol 32 2387ndash2392
Slana I Kaevska M Kralik P Horvathova A Pavlik I 2010 Distribution of Mycobacterium avium subsp avium and M a hominissuis in artificially infected pigs studied by culture and IS901 and IS1245 quantitative real time PCR Vet Microbiol 144 437ndash443
Smith NH 2012 The global distribution and phylogeography of Mycobacterium bovis clonal complexes Infect Genet Evol 12 857ndash865
Smith NH Berg S Dale J Allen AR Rodriguez S Romero B Matos F Ghebremichael S Karoui C Donati C Machado A da C Mucavele C Kazwala RR Hilty M Cadmus SI Ngandolo BNR Habtamu M Oloya J Muller A Milian-Suazo F Andrievskaia O Projahn M Barandiaraacuten S Maciacuteas A Muumlller B Zanini MS Ikuta CY Rodriguez CAR Pinheiro SR Figueroa A Cho S-N Mosavari N Chuang P-C Jou R Zinsstag J van Soolingen D Costello E Aseffa A Proantildeo-Perez F Portaels F Rigouts L Cataldi AA Collins DM Boschiroli ML Hewinson RG Ferreira Neto JS Surujballi O Tadyon K Botelho A Zaacuterraga AM Buller N Skuce R Michel A Aranaz A Gordon S V Jeon B-Y Kaumlllenius G Niemann S Boniotti MB van Helden PD Harris B Zumaacuterraga MJ Kremer K 2011 European 1 a globally important clonal complex of Mycobacterium bovis Infect Genet Evol 11 1340ndash1351
Smith NH Crawshaw T Parry J Birtles RJ 2009 Mycobacterium microti More diverse than previously thought J Clin Microbiol 47 2551ndash2559
Smith NH Kremer K Inwald J Dale J Driscoll JR Gordon S V Van Soolingen D Glyn Hewinson R Maynard Smith J 2006 Ecotypes of the Mycobacterium tuberculosis complex J Theor Biol 239 220ndash225
Smith NH Upton P 2012 Naming spoligotype patterns for the RD9-deleted lineage of the Mycobacterium tuberculosis complex wwwMbovisorg Infect Genet Evol 12 873ndash876
Smith RMM Drobniewski F Gibson A Montague JDE Logan MN Hunt D Hewinson G Salmon RL OrsquoNeill B 2004 Mycobacterium bovis infection United Kingdom Emerg Infect Dis 10 539ndash541
Smithwick RW Stratigos CB David HL 1975 Use of cetylpyridinium chloride and sodium chloride for the decontamination of sputum specimens that are transported to the laboratory for the isolation of Mycobacterium tuberculosis J Clin Microbiol 1 411ndash413
Soini H Musser JM 2001 Molecular diagnosis of mycobacteria Clin Chem 47 809ndash814
Sola C Filliol I Gutierrez MC Mokrousov I Vincent V Rastogi N 2001 Spoligotype database of Mycobacterium tuberculosis biogeographic distribution of shared types and epidemiologic and phylogenetic perspectives Emerg Infect Dis 7 390ndash396
Soo P-C Horng Y-T Hsueh P-R Shen B-J Wang J-Y Tu H-H Wei J-R Hsieh S-C Huang C-C Lai H-C 2006 Direct and simultaneous identification of Mycobacterium tuberculosis complex (MTBC) and Mycobacterium tuberculosis (MTB) by rapid multiplex nested PCR-ICT assay J Microbiol Methods 66 440ndash448
Sreevatsan S Pan X 1997 Restricted structural gene polymorphism in the Mycobacterium tuberculosis complex indicates evolutionarily recent global dissemination ProcNatl Acad Sci USA 94 9869ndash9874
Stahl DA Urbance JW 1990 The division between fast- and slow-growing species corresponds to natural relationships among the mycobacteria J Bacteriol 172 116ndash124
Reacutefeacuterences bibliographiques
216
Steingart KR Ng V Henry M Hopewell PC Ramsay A Cunningham J Urbanczik R Perkins MD Aziz MA Pai M 2006 Sputum processing methods to improve the sensitivity of smear microscopy for tuberculosis a systematic review Lancet Infect Dis 6 664ndash674
Stender H Lund K Petersen KH Rasmussen OF Hongmanee P Miorner H Godtfredsen SE 1999a Fluorescence In Situ Hybridization assay using peptide nucleic acid probes for differentiation between tuberculous and nontuberculous Mycobacterium species in smears of Mycobacterium cultures J Clin Microbiol 37 2760ndash2765
Stender H Mollerup TA Lund K Petersen KH Hongmanee P Godtfredsen SE 1999b Direct detection and identification of Mycobacterium tuberculosis in smear-positive sputum samples by fluorescence in situ hybridization (FISH) using peptide nucleic acid (PNA) probes Int J Tuberc lung Dis 3 830ndash837
Stewart LD McNair J McCallan L Gordon A Grant IR 2013 Improved detection of Mycobacterium bovis infection in bovine lymph node tissue using immunomagnetic separation (IMS)-based methods PLoS One 8 e58374
Stewart LD McNair J McCallan L Thompson S Kulakov LA Grant IR 2012 Production and evaluation of antibodies and phage display-derived peptide ligands for immunomagnetic separation of Mycobacterium bovis J Clin Microbiol 50 1598ndash1605
Stinear TP Mve-Obiang A Small PLC Frigui W Pryor MJ Brosch R Jenkin GA Johnson PDR Davies JK Lee RE Adusumilli S Garnier T Haydock SF Leadlay PF Cole ST 2004 Giant plasmid-encoded polyketide synthases produce the macrolide toxin of Mycobacterium ulcerans Proc Natl Acad Sci U S A 101 1345ndash1349
Sunder S Lanotte P Godreuil S Martin C Boschiroli ML Besnier JM 2009 Human-to-human transmission of tuberculosis caused by Mycobacterium bovis in immunocompetent patients J Clin Microbiol 47 1249ndash1251
Supply P Allix C Lesjean S Cardoso-Oelemann M Rusch-Gerdes S Willery E Savine E de Haas P van Deutekom H Roring S Bifani P Kurepina N Kreiswirth B Sola C Rastogi N Vatin V Gutierrez MC Fauville M Niemann S Skuce R Kremer K Locht C van Soolingen D 2006 Proposal for standardization of optimized Mycobacterial Interspersed Repetitive Unit-Variable-Number Tandem Repeat typing of Mycobacterium tuberculosis J Clin Microbiol 44 4498ndash4510
Supply P Lesjean S Savine E Kremer K Locht C 2001 Automated high-throughput genotyping for study of global epidemiology of Mycobacterium tuberculosis based on Mycobacterial Interspersed Repetitive Units J Clin 39 3563ndash3571
Supply P Marceau M Mangenot S Roche D Rouanet C Khanna V Majlessi L Criscuolo A Tap J Pawlik A Fiette L Orgeur M Fabre M Parmentier C Frigui W Simeone R Boritsch EC Debrie A-S Willery E Walker D Quail M a Ma L Bouchier C Salvignol G Sayes F Cascioferro A Seemann T Barbe V Locht C Gutierrez M-C Leclerc C Bentley SD Stinear TP Brisse S Meacutedigue C Parkhill J Cruveiller S Brosch R 2013 Genomic analysis of smooth tubercle bacilli provides insights into ancestry and pathoadaptation of Mycobacterium tuberculosis Nat Genet 45 172ndash179
Sweeney FP Courtenay O Hibberd V Hewinson RG Reilly LA Gaze WH Wellington EMH 2007 Environmental monitoring of Mycobacterium bovis in badger feces and badger sett soil by real-time PCR as confirmed by immunofluorescence immunocapture and cultivation Appl Environ Microbiol 73 7471ndash7473
Sweeney FP Courtenay O Ul-Hassan A Hibberd V Reilly LA Wellington EMH 2006 Immunomagnetic recovery of Mycobacterium bovis from naturally infected environmental samples Lett Appl Microbiol 43 364ndash369
Reacutefeacuterences bibliographiques
217
Tanner M Michel AL 1999 Investigation of the viability of M bovis under different environmental conditions in the Kruger National Park J Vet Res 185ndash190
Taquet A Tison F 1961 Nouvelle technique drsquoisolement des mycobacteacuteries par le lauryl sulfate de sodium Ann Inst Pasteur 100 676ndash680
Taylor C Jahans K Palmer S Okker M Brown J Steer K 2006 Mycobacterium microti isolated from two pigs Vet Rec 159 59ndash60
Taylor SJ Ahonen LJ de Leij FAAM Dale JW 2003 Infection of Acanthamoeba castellanii with Mycobacterium bovis BCG and survival of M bovis within the amoeba Appl Environ Microbiol 69 4316ndash4319
Telenti A Marchesi F Balz M Bally F Botrger EC Bodmer T 1993 Rapid identification of mycobacteria to the species level by Polymerase Chain Reaction and Restriction Enzyme Analysis J Clin Microbiol 31 175ndash178
Thacker TC Palmer M V Robbe-Austerman S Stuber TP Waters WR 2015 Anatomical distribution of Mycobacterium bovis genotypes in experimentally infected white-tailed deer Vet Microbiol180(1-2) 75-81
Thierry D Brisson-Noeumll A Vincent-Leacutevy-Freacutebault V Nguyen S Guesdon JL Gicquel B 1990a Characterization of a Mycobacterium tuberculosis insertion sequence IS6110 and its application in diagnosis J Clin Microbiol 28 2668ndash2673
Thierry D Cave MD Eisenach KD Crawford JT Bates JH Gicquel B Guesdon JL 1990b IS6110 an IS-like element of Mycobacterium tuberculosis complex Nucleic Acids Res 18 188
Thoen C LoBue P De Kantor I 2006 The importance of Mycobacterium bovis as a zoonosis Vet Microbiol 112 339ndash345
Thomas V Blanc D Bille J Greub G 2006 Biodiversity of amoebae resistant micro-organisms in an hospital water network Appl Environ Microbiol 72 2428ndash2438
Thomas V Loret J-F Jousset M Greub G 2008 Biodiversity of amoebae and amoebae-resisting bacteria in a drinking water treatment plant Environ Microbiol 10 2728ndash2745
Thomas V McDonnell G 2007 Relationship between mycobacteria and amoebae ecological and epidemiological concerns Lett Appl Microbiol 45 349ndash357
Thorel MF 1980 Isolation of Mycobacterium africanum from monkeys Tubercle 61 101ndash104
Tolhurst BA Delahay RJ Walker NJ Ward AI Roper TJ 2009 Behaviour of badgers (Meles meles) in farm buildings Opportunities for the transmission of Mycobacterium bovis to cattle Appl Anim Behav Sci 117 103ndash113
Torgerson PR Torgerson DJ 2010 Public health and bovine tuberculosis whatrsquos all the fuss about Trends Microbiol 18 67ndash72
Tortoli E 2014 Microbiological features and clinical relevance of new species of the genus Mycobacterium Clin Microbiol Rev 27 727ndash752
Tortoli E 2009 Clinical manifestations of nontuberculous mycobacteria Clin Microbiol Infect 15 906ndash910
Tortoli E 2003 Impact of genotypic studies on mycobacterial taxonomy the new mycobacteria of the 1990s Clin Microbiol Rev 16 319ndash354
Reacutefeacuterences bibliographiques
218
Torvinen E Lehtola MJ Martikainen PJ Miettinen IT 2007 Survival of Mycobacterium avium in drinking water biofilms as affected by water flow velocity availability of phosphorus and temperature Appl Environ Microbiol 73 6201ndash6207
Traag BA Driks A Stragier P Bitter W Broussard G Hatfull G Chu F Adams KN Ramakrishnan L Losick R 2010 Do mycobacteria produce endospores Proc Natl Acad Sci 107 878ndash881
Travis ER Gaze WH Pontiroli A Sweeney FP Porter D Mason S Keeling MJC Jones RM Sawyer J Aranaz A Rizaldos EC Cork J Delahay RJ Wilson GJ Hewinson RG Courtenay O Wellington EMH 2011 An inter-laboratory validation of a real time PCR assay to measure host excretion of bacterial pathogens particularly of Mycobacterium bovis PLoS One 6 e27369
Trujillo ME Velaacutezquez E Kroppenstedt RM Schumann P Rivas R Mateos PF Martiacutenez-Molina E 2004 Mycobacterium psychrotolerans sp nov isolated from pond water near a uranium mine Int J Syst Evol Microbiol 54 1459ndash1463
Truman R 2005 Leprosy in wild armadillos Lepr Rev 76 198ndash208
Tsairidou S Woolliams JA Allen AR Skuce RA McBride SH Wright DM Bermingham ML Pong-Wong R Matika O McDowell SWJ Glass EJ Bishop SC 2014 Genomic prediction for tuberculosis resistance in dairy cattle PLoS One 9(5) e96728
Turenne CY 2004 Mycobacterium saskatchewanense sp nov a novel slowly growing scotochromogenic species from human clinical isolates related to Mycobacterium interjectum and Accuprobe-positive for Mycobacterium avium complex Int J Syst Evol Microbiol 54 659ndash667
Van Donsel DJ Larkin EP 1977 Persistence of Mycobacterium bovis BCG in soil and on vegetables spray irrigated with sewage effluent and sludge J Food Prot 40 160ndash163
Van Ingen J Bendien SA de Lange WCM Hoefsloot W Dekhuijzen PNR Boeree MJ van Soolingen D 2009 Clinical relevance of non-tuberculous mycobacteria isolated in the Nijmegen-Arnhem region The Netherlands Thorax 64 502ndash506
Van Ingen J Rahim Z Mulder A Boeree MJ Simeone R Brosch R van Soolingen D 2012 Characterization of Mycobacterium orygis as M tuberculosis complex subspecies Emerg Infect Dis 18 653ndash655
Van Ingen J Van 2013 Diagnosis of nontuberculous mycobacterial infections Semin Respir Crit Care Med 34 103ndash109
Van Pittius NCG Perrett KD Michel AL Keet DF Hlokwe T Streicher EM Warren RM van Helden PD 2012 Infection of African Buffalo (Syncerus Caffer) by Oryx Bacillus A rare member of the antelope clade of the Mycobacterium Tuberculosis Complex J Wildl Dis 48 849ndash857
Van Soolingen D De Haas PEW Haagsma J Eger T Hermans PWM Ritacco V Alito A Van Embden JDA 1994 Use of various genetic markers in differentiation of Mycobacterium bovis strains from animals and humans and for studying epidemiology of bovine tuberculosis J Clin Microbiol 32 2425ndash2433
Van Soolingen D Hermans PWM De Haas PEW Soll DR Van Embden JDA 1991 Occurrence and stability of insertion sequences in Mycobacterium tuberculosis complex strains Evaluation of an insertion sequence-dependent DNA polymorphism as a tool in the epidemiology of tuberculosis J Clin Microbiol 29 2578ndash2586
Vandal OH Pierini LM Schnappinger D Nathan CF Ehrt S 2008 A membrane protein preserves intrabacterial pH in intraphagosomal Mycobacterium tuberculosis Nat Med 14 849ndash854
Varello K Pezzolato M Mascarino D Ingravalle F Caramelli M Bozzetta E 2008 Comparison of
Reacutefeacuterences bibliographiques
219
histologic techniques for the diagnosis of bovine tuberculosis in the framework of eradication programs J Vet Diagnostic Investig 20 164ndash169
Velayati AA Farnia P Mozafari M Malekshahian D Farahbod AM Seif S Rahideh S Mirsaeidi M 2015 Identification and genotyping of Mycobacterium tuberculosis isolated from water and soil samples of a metropolitan city CHEST J 147 1094
Vicente J Houmlfle U Garrido JM Fernaacutedez-De-Mera IG Acevedo P Juste R Barral M Gortaacutezar C 2007 Risk factors associated with the prevalence of tuberculosis-like lesions in fenced wild boar and red deer in south central Spain Vet Res 38 451ndash464
Voskuil MI Schnappinger D Rutherford R Liu Y Schoolnik GK 2004 Regulation of the Mycobacterium tuberculosis PEPPE genes Tuberculosis (Edinb) 84 256ndash262
Voskuil MI Schnappinger D Visconti KC Harrell MI Dolganov GM Sherman DR Schoolnik GK 2003 Inhibition of respiration by nitric oxide induces a Mycobacterium tuberculosis dormancy program J Exp Med 198 705ndash713
Wagner JC Buchanan G Bokkenheuser V Leviseur S 1958 An acid-fast bacillus isolated from the lungs of the Cape Hyrax Procavia capensis (Pallas) Nature 181 284ndash285
Walter WD Smith R Vanderklok M VerCauteren KC 2014 Linking bovine tuberculosis on cattle farms to white-tailed deer and environmental variables using Bayesian hierarchical analysis PLoS One 9 e90925
Wang Y Ogawa M Fukuda K Miyamoto H Taniguchi H 2006 Isolation and identification of mycobacteria from soils at an illegal dumping site and landfills in Japan Microbiol Immunol 50 513ndash524
Ward AI Judge J Delahay RJ 2010 Farm husbandry and badger behaviour Opportunities to manage badger to cattle transmission of Mycobacterium bovis Prev Vet Med 93 2ndash10
Warren RM Gey Van Pittius NC Barnard M Hesseling A Engelke E De Kock M Gutierrez MC Chege GK Victor TC Hoal EG Van Helden PD 2006 Differentiation of Mycobacterium tuberculosis complex by PCR amplification of genomic regions of difference Int J Tuberc Lung Dis 10 818ndash822
Waters WR Buddle BM Vordermeier HM Gormley E Palmer M V Thacker TC Bannantine JP Stabel JR Linscott R Martel E Milian F Foshaug W Lawrence JC 2011 Development and evaluation of an enzyme-linked immunosorbent assay for use in the detection of bovine tuberculosis in cattle Clin Vaccine Immunol 18 1882ndash1888
Wayne LG Hayes LG 1996 An in vitro model for sequential study of shiftdown of Mycobacterium tuberculosis through two stages of nonreplicating persistence Infect Immun 64 2062ndash2069
Wells AQ 1937 Tuberculosis in wild voles Lancet 229 1221
Wells AQ Robb-Smith AHT 1946 Murine type of tubercle bacillus (the vole acid-fast bacillus) Spec Rep Ser Med Res Cou London 259
Whan L Grant IR Rowe MT 2006 Interaction between Mycobacterium avium subsp paratuberculosis and environmental protozoa BMC Microbiol 6 63
Whiley H Keegan A Giglio S Bentham R 2012 Mycobacterium avium complex - the role of potable water in disease transmission J Appl Microbiol 113 223ndash32
White PCL Brown JA Harris S 1993 Badgers (Meles meles) cattle and bovine tuberculosis (Mycobacterium bovis) a hypothesis to explain the influence of habitat on the risk of disease transmission in southwest England Proc R Soc London B 253 277ndash284
Reacutefeacuterences bibliographiques
220
Whittington R Marshall D Nicholls PJ Marsh IB Reddacliff LA 2004 Survival and dormancy of Mycobacterium avium subsp paratuberculosis in the environment Appl Environ Microbiol 70 2989ndash3004
WHO 2014 Global leprosy update 2013 Wkly Epidemiol Rec 89 389ndash400
WHO 2014 Global Tuberculosis Report 2014
Williams RS Hoy WA 1930 The viability of B tuberculosis (bovinus) on pasture land in stored faeces and in liquid manure J Hyg (Lond) 30 413ndash419
Wirth T Hildebrand F Allix-Beacuteguec C Woumllbeling F Kubica T Kremer K Van Soolingen D Ruumlsch-Gerdes S Locht C Brisse S Meyer A Supply P Niemann S 2008 Origin spread and demography of the Mycobacterium tuberculosis complex PLoS Pathog 4(9) e1000160
Witmer G Fine AE Gionfriddo J Pipas M Shively K Piccolo K Burke P 2010 Epizootiologic survey of Mycobacterium bovis in wildlife and farm environments in northern Michigan J Wildl Dis 46 368ndash378
Wobeser G 2009 Bovine tuberculosis in Canadian wildlife an updated history Can Vet J 50 1169ndash1176
Wright DM Allen AR Mallon TR McDowell SWJ Bishop SC Glass EJ Bermingham ML Woolliams JA Skuce RA 2013 Field-isolated genotypes of Mycobacterium bovis vary in virulence and influence case pathology but do not affect outbreak size PLoS One 8(9) e74503
Young JS Gormley E Wellington EMH 2005 Molecular detection of Mycobacterium bovis and Mycobacterium bovis BCG (Pasteur) in soil Appl Environ Microbiol 71 1946ndash1952
Zanella G Durand B Hars J Moutou F Garin-Bastuji B Duvauchelle A Fermeacute M Karoui C Boschiroli ML 2008 Mycobacterium bovis in wildlife in France J Wildl Dis 44 99ndash108
Zelazny AM Calhoun LB L Shea YR Fischer SH 2005 Identification of Mycobacterium species by secA1 sequences J Clin Microbiol 43 1051ndash1058
Zeng W Zhang Y Zhao X Huang G Jiang Y Dong H Li X Wan K He C 2013 Occurrence of non-tuberculous mycobacteria species in livestock from northern China and first isolation of Mycobacterium caprae Epidemiol Infect 141 1545ndash1551
Zhang J Abadia E Refregier G Tafaj S Boschiroli ML Guillard B Andremont A Ruimy R Sola C 2010 Mycobacterium tuberculosis complex CRISPR genotyping Improving efficiency throughput and discriminative power of ldquospoligotypingrdquo with new spacers and a microbead-based hybridization assay J Med Microbiol 59 285ndash294
Zhang Y Yew W 2009 Mechanisms of drug resistance in Mycobacterium tuberculosis Int J Tuberc Lung Dis 13 1320ndash1330
Zolg JW Philippi-Schulz S 1994 The superoxide dismutase gene a target for detection and identification of mycobacteria by PCR J Clin Microbiol 32 2801ndash2812
Zuber B Chami M Houssin C Dubochet J Griffiths G Daffeacute M 2008 Direct visualization of the outer membrane of mycobacteria and corynebacteria in their native state J Bacteriol 190 5672ndash5680
Annexes
223
Annexe 1 Deacutetection de Mycobacterium bovis dans lrsquoenvironnement Premiers
reacutesultats des recherches meneacutees en Cocircte-drsquoOr
Deacutetection de Mycobacterium bovis dans lrsquoenvironnement
Premiers reacutesultats des recherches meneacutees en Cocircte-drsquoOr
Elodie Barbier1 et Alain Hartmann1
RESUME
Depuis les anneacutees 2000 en Cocircte-drsquoOr on assiste agrave une recrudescence des cas de tuberculose agrave
Mycobacterium bovis dans les cheptels bovins associeacutee agrave une contamination de la faune sauvage agrave
proximiteacute des eacutelevages contamineacutes Dans certaines zones du deacutepartement la faune sauvage infecteacutee
serait impliqueacutee dans le retour de la maladie aux bovins La transmission indirecte de la maladie par
lrsquoenvironnement semblerait une voie importante de la contamination entre faune sauvage et
domestique
Afin drsquoeacutevaluer la preacutevalence de M bovis dans lrsquoenvironnement et drsquoeacutevaluer le rocircle de lrsquoenvironnement
dans la transmission indirecte de la maladie nous avons deacuteveloppeacute des outils moleacuteculaires de
deacutetection adapteacutes aux matrices environnementales complexes (sol eau matiegraveres feacutecales) Ces outils agrave
la fois speacutecifiques et sensibles nous ont permis drsquoanalyser de nombreux preacutelegravevements reacutealiseacutes en 2013
Seuls deux eacutechantillons de sol se sont reacuteveacuteleacutes faiblement positifs
Mots-cleacutes tuberculose bovine bovins faune sauvage environnement PCR en temps reacuteel
SUMMARY
Since 2000 an increasing number of cases of bovine tuberculosis were found in cattle in Cocircte-drsquoOr and
in wild animals infected by Mycobacterium bovis in the vicinity of contaminated farms In some areas
wildlife could be considered as the likely source of cattle contamination We assume that
contamination between wild and domestic animals may occur indirectly by environmental factors (soil
water feces)
To quantify the prevalence of M bovis in the environment and to evaluate its role in indirect
transmission of the disease between species we developed molecular detection tools These are
adapted to complex environmental matrices (soil water faces) and are both specific and sensitive
They allowed us to analyze numerous samples collected in 2013 Two samples gave slight positive
results
Keywords Bovine tuberculosis Cattle Wildlife Environment Real time PCR
Texte de la confeacuterence preacutesenteacutee au cours de la Journeacutee scientifique AEEMA-RFSA 18 mars 2014
1 INRA UMR Agroeacutecologie 16 rue de Sully BP 86510 21065 Dijon France
Eacutepideacutemiol et santeacute anim 2014 65 Deacutetection de Mycobacterium bovis dans lrsquoenvironnement Premiers reacutesultats des recherches meneacutees en Cocircte-drsquoOr
224
I - Introduction
Deacuteclareacutee indemne de tuberculose bovine en
2000 la France connait depuis 2002 une
reacuteeacutemergence de lrsquoinfection dans certains
deacutepartements notamment en Cocircte-drsquoOr Entre
2002 et aujourdrsquohui 178 foyers ont eacuteteacute
successivement deacuteclareacutes dans ce deacutepartement
Pour la saison 2012-2013 la moitieacute des
cheptels infecteacutes avaient deacutejagrave eacuteteacute deacuteclareacutes
foyer une deux voire trois fois auparavant La
reacutepartition des cas bovins sur le deacutepartement
nrsquoest pas aleacuteatoire mais concentreacutee sur une
zone assez reacuteduite
Deux spoligotypes principaux sont retrouveacutes
chez les bovins SB0120 (VNTR 5544) appeleacute
plus simplement BCG-like dans la reacutegion de
Veacutenarey-Les-Laumes et Vitteaux et SB0134
(VNTR 5355) appeleacute GB35 dans la reacutegion de
Pouilly-en-Auxois
Parallegravelement agrave lrsquoinfection bovine et agrave
proximiteacute drsquoun cheptel foyer lrsquoinfection a eacuteteacute
deacutetecteacutee pour la premiegravere fois chez un animal
sauvage en 2003 chez un cerf Entre 2003 et
2007 seulement deux sangliers en plus du
cervideacute ont eacuteteacute trouveacutes infecteacutes Depuis 2007
et chaque anneacutee la maladie est reacuteguliegraverement
retrouveacutee chez des blaireaux et des sangliers
moins reacuteguliegraverement chez les cerfs Les
animaux sauvages infecteacutes sont
majoritairement trouveacutes agrave proximiteacute de
cheptels bovins infecteacutes
Lrsquohypothegravese drsquoun cycle eacutepideacutemiologique de
transmission de la maladie entre bovins et
faune sauvage est maintenant reconnue
[Richomme et al 2010] mecircme si celui-ci est
encore mal connu En France les exemples de
la forecirct de Brotonne [Zanella et al 2008] de la
Cocircte-drsquoOr de la Dordogne [Payne et al 2012]
de la Corse [Richomme et al 2010] et ceux de
lrsquoEspagne [Aranaz et al 2004] ont montreacute que
les spoligotypes et les profils VNTR (Variable
Number Tandem Repeat) des souches isoleacutees
chez les animaux sauvages tueacutes ou pieacutegeacutes agrave
proximiteacute des cheptels bovins infecteacutes sont
exactement les mecircmes que ceux des souches
bovines Cette identiteacute des souches reacutesulterait
drsquoune transmission directe ou indirecte entre
faune domestique et sauvage et vice et versa
Bien que quelques contacts directs soient
possibles entre faune sauvage et bovins [Boumlhm
et al 2009] il semble probable que la
transmission indirecte de M bovis soit la voie
de transmission preacutepondeacuterante entre bovin et
faune sauvage et inversement Il semble eacutetabli
que la transmission indirecte de M bovis soit la
principale voie de transmission entre bovins et
faune sauvage et inversement La transmission
indirecte ferait intervenir une excreacutetion de M
bovis par les animaux infecteacutes au niveau
respiratoire salivaire urinaire etou feacutecal les
animaux excreacuteteurs deacuteposeraient donc le
bacille dans lrsquoenvironnement par le biais de
leurs excreacutetats lrsquoenvironnement deviendrait
alors un reacuteservoir potentiel pour le bacille La
contamination drsquoanimaux sains pourrait se
produire par ingestion ou inhalation drsquoaeacuterosols
produits agrave partir des matrices
environnementales souilleacutees Par exemple la
formation de bioaeacuterosols contamineacutes par des
souches bacteacuteriennes drsquoactinomycegravetes
thermotoleacuterants est maintenant bien
documenteacutee agrave partir de matrices solides ou
liquides [Betteli et al 2013] Dans
lrsquoenvironnement des bovins ces bioaeacuterosols
pourraient ecirctre formeacutes agrave partir drsquoeau ou de
meacutelanges eau-seacutediments dans les zones
drsquoabreuvement ou agrave partir de particules fines
de sol (argiles)
Lrsquoexcreacutetion respiratoire salivaire urinaire et
feacutecale de M bovis par des blaireaux
tuberculeux a eacuteteacute documenteacutee en Angleterre
[Gallagher et al 1998 Gavier-Widen et al
2001 Corner et al 2011] De plus
Mycobacterium bovis a la capaciteacute de survivre
en dehors de ses hocirctes pendant des dureacutees
Eacutepideacutemiol et santeacute anim 2014 65 Deacutetection de Mycobacterium bovis dans lrsquoenvironnement Premiers reacutesultats des recherches meneacutees en Cocircte-drsquoOr
225
assez longues Cette dureacutee varie en fonction de
la nature du support contamineacute (eau sol foin
maiumls etc) de la tempeacuterature de lrsquohumiditeacute et
de lrsquoexposition aux UV [Courtenay et al 2006
Fine et al 2011 Ghodbane et al 2014] La
survie de M bovis dans des microcosmes de
sols contamineacutes soumis aux facteurs
climatiques a mecircme eacuteteacute deacutemontreacutee aux Etats-
Unis pour des dureacutees allant jusqursquoagrave environ 88
jours [Fine et al 2011]
La recherche de M bovis par une meacutethode
culturale dans lrsquoenvironnement drsquoanimaux
infecteacutes est souvent infructueuse [Pillai et al
2000 Michel et al 2007 Fine et al 2011] car
elle se heurte agrave plusieurs difficulteacutes i) la vitesse
de croissance tregraves lente de M bovis de lrsquoordre
de 45 jours qui impose un traitement preacutealable
des eacutechantillons de faccedilon agrave deacutetruire les micro-
organismes agrave croissance plus rapide ii) la
richesse des matrices analyseacutees (sol
seacutediments eau matiegraveres feacutecales etc) en
bacteacuteries saprophytes et en champignons tregraves
diversifieacutes qui rendent la deacutecontamination des
eacutechantillons tregraves difficile La deacutecontamination a
toutefois un effet non neacutegligeable sur la
viabiliteacute de M bovis et interfegravere avec la
sensibiliteacute de deacutetection par la mise en culture
[Fine 2006] Lrsquoimmunocapture magneacutetique a
eacuteteacute testeacutee avec succegraves en Angleterre pour isoler
et concentrer le bacille avant la mise en culture
[Sweeney et al 2006 et 2007]
La technique de PCR (Polymerase Chain
Reaction) est une technique indeacutependante de
la croissance de la bacteacuterie Quelques eacutetudes
ont permis la deacutetection par PCR du bacille dans
des sols et des latrines de blaireaux au
Royaume-Uni et en Irlande agrave proximiteacute de
cheptels bovins infecteacutes [Young et al 2005
Courtenay et al 2006 Sweeney et al 2006 et
Les objectifs de ce travail visaient en premier
lieu agrave mettre au point un systegraveme de deacutetection
moleacuteculaire agrave la fois speacutecifique du bacille et
assez sensible pour deacutetecter de faibles
concentrations dans les diffeacuterentes matrices
environnementales suspecteacutees pour leur rocircle
de reacuteservoir potentiel Ces nouveaux outils
moleacuteculaires nous ont permis drsquoanalyser les
eacutechantillons preacuteleveacutes dans des parcelles
partageacutees par les bovins et la faune sauvage
Ces parcelles ont eacuteteacute choisies car une
transmission indirecte de lrsquoinfection entre
animaux sauvages et domestiques eacutetait
suspecteacutee (origine de lrsquoinfection bovine
indeacutetermineacutee suite agrave lrsquoenquecircte
eacutepideacutemiologique et preacutesence drsquoanimaux
sauvages infecteacutes agrave proximiteacute) Cette approche
permettra drsquoeacutevaluer lrsquoimportance de la
transmission indirecte du bacille entre les
espegraveces et drsquoappreacutecier le rocircle joueacute par
lrsquoenvironnement
Cet article ne traite que de la deacutetection
moleacuteculaire la meacutethode fondeacutee sur la culture
eacutetant toujours en cours de validation
II - Mateacuteriel et meacutethodes
Souches bacteacuteriennes utiliseacutees
La souche vaccinale Mycobacterium bovis BCG
strain 1331 (SSI Copenhague Danemark) a eacuteteacute
utiliseacutee pour lrsquoeacutevaluation de la sensibiliteacute de la
PCR en temps reacuteel (qPCR) sur souche pure et
sur microcosmes de sol inoculeacute ainsi que pour
la reacutealisation des gammes eacutetalon de qPCR (PCR
quantitative) Cette souche a eacuteteacute cultiveacutee sur
geacutelose Coletsos pendant 30 jours puis remise
en suspension dans du milieu Middlebrook 7H9
enrichi en ADC (Sigma Aldrich) Apregraves mesure
de la densiteacute optique agrave 600nm la suspension a
eacuteteacute dilueacutee en cascade au 110 pour inoculer les
microcosmes de sol et drsquoeau Un
Eacutepideacutemiol et santeacute anim 2014 65 Deacutetection de Mycobacterium bovis dans lrsquoenvironnement Premiers reacutesultats des recherches meneacutees en Cocircte-drsquoOr
226
deacutenombrement des dilutions est ensuite
reacutealiseacute sur milieu Middlebrook 7H11 enrichi en
OADC (Sigma Aldrich)
Les ADN de 20 spoligotypes de M bovis et de
30 souches de mycobacteacuteries issus drsquoisolats
cliniques drsquoanimaux infecteacutes ont eacuteteacute fournis par
lrsquoANSES Maisons-Alfort pour les tests de
speacutecificiteacute (inclusion - exclusion) de la PCR en
temps reacuteel (tableau 1)
Extraction de lrsquoADN et purification
Lrsquoextraction drsquoADN et la purification des
microcosmes de sol drsquoeau et des eacutechantillons
environnementaux a eacuteteacute reacutealiseacutee selon le
protocole modifieacute deacutecrit par Ranjard et al
[2003]
Les eacutechantillons drsquoeau ont eacuteteacute filtreacutes sur des
membranes drsquoester de cellulose de 045 microm de
porositeacute Les filtres ont eacuteteacute broyeacutes avec de
lrsquoazote liquide puis lrsquoADN extrait selon la
meacutethode preacuteceacutedemment citeacutee agrave lrsquoexception de
la purification sur colonnes PVPP
(polyvinylpolypyrrolidone insoluble)
Les ADN extraits ont eacuteteacute quantifieacutes avec un
spectrophotomegravetre (NanoDropreg 2000 Thermo
Scientific) et dilueacutes au 15 110 et 120 pour la
PCR en temps reacuteel
Deacuteveloppement de trois systegravemes de
deacutetection moleacuteculaire
La bibliographie nous a permis de seacutelectionner
trois cibles geacuteneacutetiques situeacutees sur des locus
diffeacuterents tregraves speacutecifiques de M bovis [Huard
et al 2002 Brodin et al 2002] Les geacutenomes
entiegraverement seacutequenceacutes des mycobacteacuteries du
MTBC (complexe M tuberculosis comprenant
M bovis) et des MNT (mycobacteacuteries non
tuberculeuses) sur le site NCBI nous ont permis
drsquoaligner les seacutequences de ces trois gegravenes agrave la
recherche de zones tregraves conserveacutees dans le
MTBC Chaque systegraveme de deacutetection
moleacuteculaire en PCR temps reacuteel creacuteeacute comprend
une amorce Forward une amorce Reverse et
une sonde oligonucleacuteotidique marqueacutee par
deux fluorochromes (sonde TaqMan) choisies
agrave lrsquoaide du logiciel Primer3 (Simgenecom) Pour
chaque systegraveme nous avons eacutegalement conccedilu
des amorces de PCR agrave lrsquoexteacuterieur des seacutequences
preacuteceacutedentes de faccedilon agrave reacutealiser des PCR
nicheacutees (utiliseacutees uniquement en confirmation
des PCR en temps reacuteel positives)
Speacutecificiteacute et sensibiliteacute des systegravemes de
deacutetection sur souches pures sol et eau
La speacutecificiteacute des trois systegravemes de deacutetection a
eacuteteacute testeacutee par PCR en temps reacuteel sur lrsquoADN de
19 spoligotypes diffeacuterents de M bovis quatre
souches de mycobacteacuteries appartenant au
MTBC deux au Complexe avium et 20 MNT
(tableau 1) Nous avons eacutegalement veacuterifieacute que
la PCR en temps reacuteel ne donnait pas de signal
sur des ADN extraits de 22 sols provenant du
Morbihan des Yvelines du Haut-Rhin et de
Cocircte-drsquoOr preacuteleveacutes en dehors des zones ougrave
seacutevit la tuberculose bovine Ces sols preacutesentent
des caracteacuteristiques physico-chimiques varieacutees
Pour deacuteterminer la sensibiliteacute de deacutetection de
la PCR en temps reacuteel de chacun des systegravemes
la quantification a eacuteteacute reacutealiseacutee sur des dilutions
du gegravene cible cloneacute dans un plasmide (gamme
allant de 1 copie agrave 106 copies du gegravene rechercheacute
dans 5 microl) La limite de deacutetection de la PCR en
temps reacuteel dans le sol a eacuteteacute eacutevalueacutee par
lrsquoinoculation de suspension bacteacuterienne de M
bovis BCG dans un sol dont les caracteacuteristiques
physico-chimiques sont identiques aux sols
incrimineacutes en Cocircte-drsquoOr Chaque microcosme
de sol de 2 g a eacuteteacute inoculeacute avec des suspensions
bacteacuteriennes pour obtenir une concentration
finale en bacteacuteries variant de 10 agrave 105 CFU par
g de sol Des aliquotes de 100 ml drsquoeau non
steacuterile issue drsquoun ruisseau hors zone de
tuberculose ont eacuteteacute inoculeacutes avec ces mecircmes
concentrations
Eacutepideacutemiol et santeacute anim 2014 65 Deacutetection de Mycobacterium bovis dans lrsquoenvironnement Premiers reacutesultats des recherches meneacutees en Cocircte-drsquoOr
227
Tableau 1
Souches bacteacuteriennes et spoligotypes utiliseacutes pour eacutevaluer la speacutecificiteacute des trois systegravemes
de deacutetection moleacuteculaire
M bovis (spoligotypes) Mycobacteacuteries du MTBC Mycobacteacuteries atypiques Mycobacteacuteries du
Complexe avium
GB35 M microti M chitae Mavium
BCG M tuberculosis M elephantis M intracellulare
F1 M caprae M confluentis
F2 M pinnipedi M fortuitum
F5 M rufum
F6 M non chromogenicum
F7 M aichiense
F23 M thermoresitibile
F41 M xenopi
F57 M kansai
F61 M vaccae
F105 M arupense
F151 M chelonae
GB21 M porcinum
GB54 M parafinicum
SB0166 M celatum
SB0885 M intermedium
SB0999 M rutilum
SB1081 M smegmatis
M szulgai
Choix des sites de preacutelegravevements
Les eacutechantillons environnementaux ont eacuteteacute
preacuteleveacutes sur huit communes de Cocircte-drsquoOr
situeacutees dans la zone Nord entre mai et juillet
2013 Nous avons cibleacute avec lrsquoaide de la DDPP21
des parcelles
Sur lesquelles un ou plusieurs bovins
deacutecouverts infecteacutes lors de la prophylaxie
2012-2013 ont pacirctureacute en 2012 et pour
lesquels lrsquoorigine de la contamination reste
inexpliqueacutee
Autour desquelles la surveillance de la
faune sauvage a mis en eacutevidence lrsquoinfection
agrave M bovis chez des blaireaux pieacutegeacutes ou des
sangliers tueacutes agrave la chasse
Sur les pacirctures nous avons privileacutegieacute les zones
susceptibles drsquoecirctre freacutequenteacutees par les bovins
et la faune sauvage zones drsquoabreuvement
(trous drsquoeau sources abreuvoirs) zones de
nourrissage au sol De lrsquoeau du sol et des
seacutediments ont eacuteteacute reacutecolteacutes Autour des
parcelles les terriers et matiegraveres feacutecales de
blaireaux (latrines) ainsi que les zones
drsquoabreuvement de la faune sauvage ont eacuteteacute
preacuteleveacutes
Eacutepideacutemiol et santeacute anim 2014 65 Deacutetection de Mycobacterium bovis dans lrsquoenvironnement Premiers reacutesultats des recherches meneacutees en Cocircte-drsquoOr
228
III - Reacutesultats
Les systegravemes de deacutetection conccedilus sont
speacutecifiques des espegraveces appartenant au MTBC
in silico
Les gegravenes ou locus cibles ont eacuteteacute choisis soit
pour leur speacutecificiteacute agrave M bovis soit pour leur
absence chez M microti seule mycobacteacuterie
du complexe MTBC agrave avoir eacuteteacute deacutecrite dans
lrsquoenvironnement avec M bovis Les systegravemes de
deacutetection ciblant lrsquoIS1561 et la reacutegion Rv3866
ont eacuteteacute choisis car ces locus sont preacutesents chez
tous les membres du MTBC mais absents chez
M microti chez les mycobacteacuteries du complexe
avium ainsi que chez les MNT [Huard et al
2002 Brodin et al 2002] La reacutegion RD4
preacutesente chez tous les membres du MTBC est
absente chez M bovis [Huard et al 2002]
Deux systegravemes de deacutetection moleacuteculaire
ciblant lrsquoIS1561 et la reacutegion Rv3866 ont eacuteteacute
reacutealiseacutes agrave lrsquoaide du logiciel Primer3 Le troisiegraveme
systegraveme ciblant la zone encadrant la RD4 utilise
une amorce Forward et une sonde deacutejagrave deacutecrite
[Sweeney et al 2007] seule lrsquoamorce Reverse
a eacuteteacute redessineacutee afin drsquoobtenir un produit PCR
plus petit La sonde utiliseacutee ne peut se fixer que
sur lrsquoADN des souches preacutesentant la deacuteleacutetion La
speacutecificiteacute des systegravemes de deacutetection a eacuteteacute
deacutemontreacutee in silico par alignement de
seacutequences agrave lrsquoaide du programme BLASTreg
Aucune deacutetection croiseacutee avec drsquoautres
espegraveces que celles appartenant au MTBC nrsquoa
eacuteteacute mise en eacutevidence in silico pour les systegravemes
de deacutetection utiliseacutes
Speacutecificiteacute et sensibiliteacute des systegravemes de
deacutetection sur souches pures et microcosmes
de sol inoculeacutes
Sur les 18 spoligotypes de M bovis testeacutes tous
ont donneacute un signal fort et conserveacute lors de la
PCR en temps reacuteel (Cycle threshold = 20)
Dans le complexe MTBC nous avons testeacute nos
systegravemes sur M microti M tuberculosis M
caprae et M pinnipedii Le systegraveme RD4
preacutesente la plus grande speacutecificiteacute puisqursquoil
nrsquoamplifie que M bovis parmi les membres du
complexe testeacutes LrsquoIS1561 et la reacutegion Rv3866
donnent un signal identique pour M bovis M
caprae M tuberculosis et M pinnipedii mais
ces systegravemes nrsquoamplifient pas M microti
Mecircme srsquoil a eacuteteacute montreacute que M tuberculosis
peut survivre expeacuterimentalement dans le sol
(Ghodbane et al 2014) seuls M microti et M
bovis y ont eacuteteacute deacutejagrave eacuteteacute deacutecrits [Courtenay et
al 2006 Sweeney et al 2006 et 2007]
Sur les 20 souches de MNT testeacutees cinq ont
donneacute un tregraves faible signal (Ct gt 38) avec les
trois systegravemes de deacutetection Pour le complexe
avium seul M avium a eacuteteacute deacutetecteacute avec un
Ctgt38
Aucun des 22 sols testeacutes nrsquoa donneacute de signal en
PCR en temps reacuteel quel que soit le systegraveme de
deacutetection utiliseacute ce qui indique une tregraves bonne
speacutecificiteacute de ces systegravemes de deacutetection sur
une large gamme de sols aux caracteacuteristiques
physico-chimiques diffeacuterentes et contenant des
communauteacutes microbiennes varieacutees
En ce qui concerne la sensibiliteacute des systegravemes
la limite de deacutetection des cibles se situe entre 1
et 10 copies lorsqursquoon utilise des solutions de
plasmide dilueacutees La reacutepeacutetabiliteacute de la
quantification des dilutions de plasmides est
tregraves bonne entre 10 et 106 copies de gegravenes avec
les trois systegravemes En revanche en dessous de
10 copies la deacutetection est moins reacutepeacutetable
(non deacutetection dans certaines reacutepeacutetitions)
Sur les microcosmes de sol et drsquoeau inoculeacutes la
limite de deacutetection des cibles se situe agrave 103
copies du gegravene par g de sol et agrave 10 copies du
gegravene par ml drsquoeau
Le systegraveme ciblant lrsquoIS1561 semble ecirctre tregraves
leacutegegraverement plus sensible que les deux autres
Nous lrsquoavons donc utiliseacute en 1egravere intention pour
Eacutepideacutemiol et santeacute anim 2014 65 Deacutetection de Mycobacterium bovis dans lrsquoenvironnement Premiers reacutesultats des recherches meneacutees en Cocircte-drsquoOr
229
cribler lrsquoensemble des eacutechantillons
environnementaux
Premiers reacutesultats sur les eacutechantillons
environnementaux
Sur les huit communes cibleacutees 150
eacutechantillons de sol 18 eacutechantillons de
seacutediments cinq eacutechantillons drsquoeau et huit
eacutechantillons de fegraveces de blaireaux ont eacuteteacute
preacuteleveacutes Parmi les 150 sols preacuteleveacutes 28 lrsquoont
eacuteteacute dans des terriers de blaireaux et de renards
Parmi les terriers de blaireaux preacuteleveacutes
certains eacutetaient potentiellement des terriers de
blaireaux infecteacutes par M bovis car des
animaux pieacutegeacutes agrave proximiteacute se sont reacuteveacuteleacutes ecirctre
porteurs de M bovis Cependant en 2013 nous
nrsquoavons pu obtenir aucune certitude sur les
terriers reacuteellement occupeacutes par les blaireaux
porteurs de M bovis (pieacutegeage des blaireaux en
couleacutee et non pas en gueule de terrier)
Sur les 181 eacutechantillons analyseacutes seuls deux
eacutechantillons de sol dans deux communes
diffeacuterentes ont donneacute un signal en PCR en
temps reacuteel avec lrsquoIS1561 agrave lrsquoune des dilutions
seulement Ce signal est toutefois faible et nrsquoa
pas pu ecirctre reproduit avec les autres systegravemes
de deacutetection Un des eacutechantillons positifs
provient drsquoun terrier agrave proximiteacute duquel un
blaireau infecteacute a eacuteteacute pieacutegeacute Le second
eacutechantillon de sol a eacuteteacute preacuteleveacute dans une latrine
de blaireau creuseacutee sous une clocircture de preacute
Lrsquoanalyse des fegraveces de cette latrine nrsquoa toutefois
pas donneacute de signal
La campagne de preacutelegravevement 2014 sera
reacutealiseacutee pour augmenter le nombre
drsquoeacutechantillons preacuteleveacutes sur une zone plus
restreinte en particulier ougrave des contaminations
reacutecurrentes des cheptels bovins ont eacuteteacute et sont
observeacutees
IV - Discussion
Mycobacterium bovis lrsquoagent de la tuberculose
bovine est une bacteacuterie pathogegravene qui infecte
les bovins et certains mammifegraveres sauvages en
France La maicirctrise de la maladie chez les
bovins dans certains deacutepartements franccedilais
semble ecirctre entraveacutee par la proximiteacute
drsquoanimaux sauvages infecteacutes La transmission
indirecte du bacille entre faune sauvage et
domestique par lrsquointermeacutediaire de
lrsquoenvironnement permettrait drsquoexpliquer
certains cas de re-contaminations chez les
bovins Lrsquoenvironnement pourrait servir de
reacuteservoir de la bacteacuterie par le biais de
lrsquoexcreacutetion animale il pourrait en retour
infecter des animaux sains soit par inhalation
drsquoaeacuterosols contamineacutes soit par ingestion
(abreuvement) Quelques eacutetudes en
Angleterre et en Irlande ont mis en eacutevidence la
preacutesence de M bovis dans des sols de preacutes des
sols de terriers et des latrines de blaireaux agrave
proximiteacute de cheptels bovins infecteacutes
[Courtenay et al 2006 Sweeney et al 2006
et 2007 Young et al 2005 Travis et al 2011]
Des eacutetudes expeacuterimentales ont montreacute que M
bovis pouvait survivre plusieurs mois dans le sol
[Maddock 1933 Young et al 2005 Fine et al
2011 Ghodbane et al 2014] tout en restant
virulent [Maddock 1933 Ghodbane et al
2014] La deacutetection du bacille dans
lrsquoenvironnement freacutequenteacute par les animaux
domestiques ou sauvages infecteacutes nrsquoa jamais
eacuteteacute deacutecrite en France Dans certaines zones de
Cocircte-drsquoOr la transmission indirecte du bacille
aux bovins par lrsquoenvironnement est fortement
suspecteacutee
Dans cette eacutetude nous avons deacuteveloppeacute trois
systegravemes de deacutetection moleacuteculaire afin
drsquooptimiser la mise en eacutevidence de M bovis
Eacutepideacutemiol et santeacute anim 2014 65 Deacutetection de Mycobacterium bovis dans lrsquoenvironnement Premiers reacutesultats des recherches meneacutees en Cocircte-drsquoOr
230
dans des eacutechantillons environnementaux
complexes afin de srsquoaffranchir de la culture En
effet lrsquoisolement de M bovis dans les matrices
environnementales par la culture est long car
drsquoune part un traitement preacutealable est
neacutecessaire pour srsquoaffranchir de la flore
microbienne de contamination et drsquoautre part
le bacille a une croissance tregraves lente
Contrairement agrave la culture la technique de PCR
en temps reacuteel permet une deacutetection rapide du
bacille sous reacuteserve que le ou les gegravenes cibleacutes
soient tregraves speacutecifiques de faccedilon agrave deacutetecter et
quantifier une espegravece bacteacuterienne particuliegravere
dans un meacutelange drsquoADN complexe Mais cette
technique rencontre neacuteanmoins quelques
limites techniques
Les eacutechantillons environnementaux
contiennent une flore microbienne et animale
abondante et varieacutee lrsquoADN extrait contient une
grande diversiteacute de seacutequences geacuteneacutetiques agrave
lrsquoorigine de faux positifs Ainsi des espegraveces
proches de M bovis notamment les
mycobacteacuteries environnementales ainsi que
des espegraveces encore inconnues peuvent ecirctre
preacutesentes dans les eacutechantillons Il est donc
neacutecessaire de srsquoassurer de la speacutecificiteacute des
systegravemes de deacutetection La deacutetection du bacille
par la recherche associeacutee de trois cibles
geacuteneacutetiques avec le systegraveme TaqMan nous a
permis de deacutevelopper une meacutethode de
deacutetection moleacuteculaire speacutecifique et sensible de
M bovis dans des matrices complexes
Lrsquoextraction drsquoADN de matrices environ-
nementales srsquoaccompagne drsquoune co-extraction
drsquoinhibiteurs potentiels de la PCR (acides
humiques polypheacutenols polysaccharides et
meacutetaux) Lrsquoacide humique par exemple inhibe
lrsquoactiviteacute de la Taq polymeacuterase [Kermekchiev et
al 2009] et se lie sur des sites speacutecifiques de
lrsquoADN empecircchant son amplification [Opel et al
2010] La dilution des eacutechantillons drsquoADN
utiliseacutes en PCR en temps reacuteel permet la dilution
des inhibiteurs et limite leur action Toutefois
lorsque la cible rechercheacutee est preacutesente en
petit nombre de copies la dilution de lrsquoADN
rend la deacutetection moins efficace Crsquoest
pourquoi nous analysons lrsquoADN extrait pur et
dilueacute au 15 au 110 et au 120 en PCR en
temps reacuteel de faccedilon agrave rechercher la quantiteacute
drsquoinhibiteurs minimale permettant drsquoamplifier
la cible
Les limites de deacutetection de la PCR se situent au
mieux autour de 1000 copies de gegravenes par g de
sol Les seuils de deacutetection des trois systegravemes
que nous avons creacuteeacutes se situent au-dessus de
1000 copies du gegravene par g de sol ce qui
correspond agrave des valeurs en adeacutequation avec
drsquoautres reacutesultats obtenus sur la deacutetection de
bacteacuteries dans les sols [Ibekwe et Grieve
2003]
En 2013 seulement deux des eacutechantillons
analyseacutes ont donneacute un signal faible avec un des
systegravemes de deacutetection moleacuteculaire Il srsquoagissait
dans les deux cas drsquoeacutechantillons de sol un sol
de terrier probablement habiteacute par un blaireau
infecteacute et un sol de latrine Nous nrsquoavons pu
reacutepeacuteter les reacutesultats probablement agrave cause de
la faible charge en M bovis des eacutechantillons
analyseacutes en dessous de la limite de deacutetection
de nos outils Il reste difficile drsquoisoler M bovis
de lrsquoenvironnement malgreacute la sensibiliteacute et la
speacutecificiteacute des techniques utiliseacutees Plusieurs
eacutetudes malgreacute lrsquoabondance de preacutelegravevements
reacutealiseacutes ont eacuteteacute infructueuses [Pillai et al
2000 Michel et al 2007 Fine et al 2011]
Cette difficulteacute reacutesulte des limites techniques
preacuteceacutedemment citeacutees mais eacutegalement du
caractegravere variable et intermittent de lrsquoexcreacutetion
de M bovis par les animaux infecteacutes
Lrsquoexcreacutetion de la bacteacuterie par la voie
respiratoire les urines les matiegraveres feacutecales ou
les plaies de morsure nrsquoest pas systeacutematique
chez les blaireaux infecteacutes Le niveau
drsquoexcreacutetion deacutepend de la localisation et de
lrsquointensiteacute des leacutesions Il semble que lrsquoexcreacutetion
la plus importante soit trouveacutee chez des
animaux agrave un stade avanceacute de la maladie
[Gallagher et al 1998 Corner et al 2012] En
Eacutepideacutemiol et santeacute anim 2014 65 Deacutetection de Mycobacterium bovis dans lrsquoenvironnement Premiers reacutesultats des recherches meneacutees en Cocircte-drsquoOr
231
Cocircte-drsquoOr une eacutetude meneacutee entre 2009 et
2011 a montreacute que seuls 17 des blaireaux
pieacutegeacutes et analyseacutes (19 sur 1 146) preacutesentaient
des leacutesions visibles alors que la bacteacuterie eacutetait
retrouveacutee sur 56 des blaireaux (49 sur 878)
dans la zone de tuberculose bovine [Payne et
al 2012] Lrsquoabsence de leacutesion visible sur la
majoriteacute des blaireaux analyseacutes pourrait
expliquer une absence drsquoexcreacutetion ou une
faible excreacutetion de M bovis dans
lrsquoenvironnement
Une autre difficulteacute reacuteside dans la reacutealisation
spatiale et temporelle de lrsquoeacutechantillonnage Les
bacilles sont excreacuteteacutes par les animaux sur de
petites zones bien localiseacutees Hormis les
latrines de blaireaux bien visibles les
preacutelegravevements agrave lrsquoeacutechelle drsquoune parcelle drsquoun
terrier drsquoune source sont reacutealiseacutes de maniegravere
ponctuelle et au hasard ce qui diminue la
probabiliteacute de deacutetection de cette bacteacuterie De
plus nous avons toujours un temps de
deacutecalage entre le deacutepistage des bovins ou des
animaux sauvages infecteacutes et la date des
preacutelegravevements car la certitude du diagnostic
animal neacutecessite plusieurs semaines
V - Conclusion et perspectives
Nous avons deacuteveloppeacute des outils de PCR en
temps reacuteel agrave la fois speacutecifiques et sensibles
pour la deacutetection de M bovis dans les matrices
environnementales Malgreacute le nombre
important drsquoeacutechantillons analyseacutes seuls deux
eacutechantillons peu chargeacutes en M bovis ont pu
ecirctre mis en eacutevidence Ces outils de PCR en
temps reacuteel vont encore ecirctre optimiseacutes en 2014
et permettre lrsquoanalyse des nombreux
preacutelegravevements preacutevus dans des zones plus
cibleacutees de Cocircte-drsquoOr dans lesquelles bovins et
faune sauvage infecteacutes ont eacuteteacute retrouveacutes
Plusieurs meacutethodes de culture sont en cours
drsquoessai et seront utiliseacutees sur les eacutechantillons
preacuteleveacutes afin de montrer la viabiliteacute des
bacteacuteries dans lrsquoenvironnement
Eacutepideacutemiol et santeacute anim 2014 65 Deacutetection de Mycobacterium bovis dans lrsquoenvironnement Premiers reacutesultats des recherches meneacutees en Cocircte-drsquoOr
232
Bibliographie
Aranaz A De Juan L Montero N Saacutenchez C Galka M Delso C Alvarez J Romero B Bezos J Vela A I Briones V Mateos A Domiacutenguez L - Bovine tuberculosis (Mycobacterium bovis) in wildlife in Spain Journal of clinical microbiology 2004 42 2602-2608
Betelli L Duquenne P Grenouillet F Simon X Scherer E Geacutehin E Hartmann A - Development and evaluation of a method for the quantification of airborne Thermoactinomyces vulgaris by real-time PCR Journal of microbiological methods 2013 92 25-32
Boumlhm M Hutchings MR White PCL - Contact networks in a wildlife-livestock host community identifying high-risk individuals in the transmission of bovine TB among badgers and cattle PloS One 2009 4(4) e5016
Brodin P Eiglmeier K Marmiesse M Garnier T Niemann S Cole S T Brosch R Billault A - Bacterial Artificial Chromosome-Based Comparative Genomic Analysis Identifies Mycobacterium microti as a Natural ESAT-6 Deletion Mutant Bacterial Artificial Chromosome-Based Comparative Genomic Analysis Identifies Mycobacterium microti as a Natural ESAT-6 Infection and immunity 2002 70 10 5568-5578
Colinon C Deredjian A Hien E Brothier E Bouziri L Cournoyer B Hartman A Henry S Jolivet C Ranjard L Nazaret S - Detection and enumeration of Pseudomonas aeruginosa in soil and manure assessed by an ecfX qPCR assay Journal of applied microbiology 2013 114 1734-1749
Corner L A L Murphy D Gormley E - Mycobacterium bovis infection in the Eurasian badger (Meles meles) the disease pathogenesis epidemiology and control Journal of comparative pathology 2011 144 1-24
Corner L A L OrsquoMeara D Costello E Lesellier S Gormley E - The distribution of Mycobacterium bovis infection in naturally infected badgers Veterinary journal (London England 1997) 2012 194 166-172
Courtenay O Reilly L A Sweeney F P Hibberd V Bryan S Ul-Hassan A Newman C Macdonald D W Delahay R J Wilson G J Wellington E M H - Is Mycobacterium bovis in the environment important for the persistence of bovine tuberculosis Biology letters 2006 2 460-462
Fine AE - The Role of Indirect Transmission in the Epidemiology of Bovine Tuberculosis in Cattle and White-Tailed Deer in Michigan 2006 Dissertation for the degree of Doctor of Philosophy Michigan State University
Fine A E Bolin C A Gardiner J C Kaneene J B - A Study of the Persistence of Mycobacterium bovis in the Environment under Natural Weather Conditions in Michigan USA Veterinary medicine international 2011 2011 765430
Gallagher J Clifton-Hadley R S - Tuberculosis in badgers a review of the disease and its significance for other animals Research in veterinary science 2000 69 203-217
Gallagher J Monies R Gavier-Widen M Rule B - Role of infected non-diseased badgers in the pathogenesis of tuberculosis in the badger Veterinary Record 1998 142 710-714
Gavier-Widen D Chambers M A Palmer N Newell DG Hewinson R G - Pathology of natural Mycobacterium bovis infection in European badgers (Meles meles) and its relationship with bacterial excretion Veterinary Record 2001 148(10) 299-304
Ghodbane R Mba Medie F Lepidi H Nappez C Drancourt M - Long-term survival of tuberculosis complex mycobacteria in soil Microbiology (Reading England) 2014 160(Pt 3) 496-501
Goodchild A V Clifton-Hadley R S - Cattle-to-cattle transmission of Mycobacterium bovis Tuberculosis (Edinburgh Scotland) 2001 81 23-41
Huard R C Claudio L Lazzarini D O Butler W R Soolingen D Van Ho J L - PCR-Based Method To Differentiate the Subspecies of the Mycobacterium tuberculosis Complex on the Basis of Genomic Deletions Journal of clinical microbiology 2003 41 1637-1650
Ibekwe AM Grieve CM - Detection and quantification of Escherichia coli O157H7 in environmental samples by real-time PCR Journal of Applied Microbiology 2003 94(3) 421-431
Kermekchiev M B Kirilova L I Vail E E Barnes W M - Mutants of Taq DNA polymerase resistant to PCR inhibitors allow DNA amplification from whole blood and crude soil samples Nucleic acids research 2009 37 e40
Maddock EC - Studies on the survival time of the bovine tubercle bacillus in soil soil and dung in dung and on grass with experiments on the
Eacutepideacutemiol et santeacute anim 2014 65 Deacutetection de Mycobacterium bovis dans lrsquoenvironnement Premiers reacutesultats des recherches meneacutees en Cocircte-drsquoOr
233
preliminary treatment of infected organic matter and the cultivation of the organism The Journal of Hygiene 1933 33(1) 103-117
Michel AL de Klerk LM Gey van Pittius NC Warren RM van Helden PD - Bovine tuberculosis in African buffaloes observations regarding Mycobacterium bovis shedding into water and exposure to environmental mycobacteria BMC Veterinary Research 2007 3 23
Neill S Hanna J OrsquoBrien J McCracken R - Excretion of Mycobacterium bovis by experimentally infected cattle Veterinary Record 1988 123 340-343
Opel K L Chung D McCord B R - A study of PCR inhibition mechanisms using real time PCR Journal of forensic sciences 2010 55 25-33
Palmer M V Whipple D L Waters W R Mitchell V Diana L Ray W Transmission E D - Experimental Deer-To-Deer Transmission of Mycobacterium bovis Experimental deer-to-deer transmission of Mycobacterium bovis American journal of veterinary research 2001 62 692-696
Payne A Boschiroli M L Gueneau E Moyen J L Rambaud T Dufour B Gilot-Fromont E Hars J - Bovine tuberculosis in ldquoEurasianrdquo badgers (Meles meles) in France European Journal of Wildlife Research 2012 59 331-339
Pillai S D Widmer K W Ivey L J Coker K C Newman E Lingsweiler S Baca D Kelley M Davis D S Silvy N J Adams L G - Failure to identify non-bovine reservoirs of Mycobacterium bovis in a region with a history of infected dairy-cattle herds Preventive veterinary medicine 2000 43 53-62
Ranjard L Lejon DPH Mougel C Schehrer L Merdinoglu D Chaussod R - Sampling strategy in molecular microbial ecology influence of soil sample size on DNA fingerprinting analysis of fungal and bacterial communities Environmental Microbiology 2003 5(11) 1111-1120
Richomme C Boschiroli M L Hars J Casabianca F Ducrot C - Bovine tuberculosis in livestock and wild boar on the Mediterranean island Corsica Journal of wildlife diseases 2010 46 627-631
Sweeney F P Courtenay O Ul-Hassan A Hibberd V Reilly L A Wellington E M H - Immunomagnetic recovery of Mycobacterium bovis from naturally infected environmental
samples Letters in applied microbiology 2006 43 364-369
Sweeney F P Courtenay O Hibberd V Hewinson R G Reilly L A Gaze W H Wellington E M H - Environmental monitoring of Mycobacterium bovis in badger feces and badger sett soil by real-time PCR as confirmed by immunofluorescence immunocapture and cultivation Applied and environmental microbiology 2007 73 7471-7473
Travis ER Gaze WH Pontiroli A Sweeney FP Porter D Mason S Keeling MJC Jones RM Sawyer J Aranaz A Rizaldos EC Cork J Delahay RJ Wilson GJ Hewinson RG Courtenay O Wellington EMH - An inter-laboratory validation of a real time PCR assay to measure host excretion of bacterial pathogens particularly of Mycobacterium bovis PloS One 2011 6(11) e27369
Walter W D Anderson C W Smith R Vanderklok M Averill J J Vercauteren K C - On-farm mitigation of transmission of tuberculosis from white-tailed deer to cattle literature review and recommendations Veterinary medicine international 2012 2012 616318
Young J S Gormley E amp Wellington E M H - Molecular Detection of Mycobacterium bovis and Mycobacterium bovis BCG ( Pasteur ) in Soil Applied and environmental microbiology 2005 71 1946-1952
Zanella G Durand B Hars J Moutou F Garin-Bastuji B Duvauchelle A Fermeacute M Karoui C Boschiroli M L - Mycobacterium bovis in wildlife in France Journal of wildlife diseases 2008 44 99-108
235
Annexe 2 Preacuteparation du Middlebrook 7H9
Middlebrook 7H9 suppleacutementeacute pour la culture liquide de Mycobacterium bovis
Preacuteparation de la base 7H9 (pour 500 mL de milieu)
1 Diluer 235 g de Middlebrook 7H9 (Fluka analytical) dans 450 mL drsquoeau osmoseacutee 2 Autoclaver 15 min agrave 120degC
Preacuteparation de lrsquoenrichissement ADC (pour 50 mL)
1 Peser 25 g de BSA 1 g de Dextrose et 0005 g de catalase bovine 2 Ajouter 50 mL drsquoeau osmoseacutee 3 Agiter environ 30 min (agitation magneacutetique modeacutereacutee pour eacuteviter la formation de mousse) 4 Filtrer lrsquoenrichissement sur une membrane filtrante de porositeacute 022 microM (Conservation 1 mois agrave 4degC)
Preacuteparation du Middlebrook 7H9 enrichi
1 Apregraves lrsquoautoclavage une fois le milieu refroidi ajouter 50 mL drsquoADC (conservation 1 mois agrave 4degC)
237
Annexe 3 Preacuteparation du Middlebrook 7H11
Middlebrook 7H11 suppleacutementeacute pour la culture de Mycobacterium bovis
Preacuteparation des solutions stock (tous les produits ont eacuteteacute fournis par le laboratoire Sigma-Aldrich)
Vert malachite 06 dissoudre 03 g dans 50 mL drsquoeau osmoseacutee
TicarcillineTrimeacutethoprime peser 1 g ticarcilline et 01 g de trimeacutethoprime lactate ajouter 10 mL drsquoeau osmoseacutee steacuterile Vortexer jusqursquoagrave dissolution reacutepartir en Eppendorf de 1 mL congeler (conservation 6 mois agrave -20degC)
Cycloheximide peser 500 mg et dissoudre dans 10 mL drsquoeacutethanol absolu (conservation 1 an agrave 4degC)
Polymyxine B dilution dans eau osmoseacutee steacuterile pour concentration finale de 100 000 UI par mL
Sang mouton lyseacute Meacutelanger 25 mL de sang avec 25 mL drsquoeau osmoseacutee steacuterile bain-marie 10 min agrave 60degC puis utilisation immeacutediate
Preacuteparation de la base 7H11 (pour 1 L de milieu)
1 Diluer 189 g de Middlebrook 7H11 (Fluka analytical) dans 781 mL drsquoeau 2 Ajouter 1 mL de Vert de malachite 06 3 Ajouter un barreau aimanteacute dans la bouteille 4 Autoclaver 15 min agrave 120degC
Preacuteparation de lrsquoenrichissement OADC (pour 100mL)
1 Peser 5 g de BSA 2 g de Dextrose 085 g de NaCl et 0005g de catalase bovine 2 Ajouter 100 mL drsquoeau osmoseacutee 3 Agiter environ 30 min (agitation magneacutetique modeacutereacutee pour eacuteviter la formation de mousse) 4 Ajouter 125 microL drsquoacide oleacuteique 5 Filtrer lrsquoenrichissement sur une membrane filtrante de porositeacute 022 microM (Conservation 1 mois agrave 4degC)
Preacuteparation du Middlebrook 7H11 enrichi
1 Apregraves lrsquoautoclavage porter le milieu agrave 45-50degC 2 Ajouter 100 mL de seacuterum bovin inactiveacute (agitation magneacutetique) 3 Ajouter 100 mL drsquoOADC (agitation magneacutetique) 4 Ajouter 5 mL de sang de mouton lyseacute 5 Ajouter 1 mL de TicarcillineTrimeacutethoprime 2 mL de Polymyxine B 2 mL de cycloheximide
(agitation magneacutetique) (facultatif si ensemencement de cultures pures)(Concentrations finales respectives 100 mgL 10 mgL 200 000 UIL et 100 mgL)
6 Reacutepartition de 25 mL de milieu par boite (Conservation 1 mois agrave 4degC)
Reacutesumeacute La tuberculose bovine est une maladie infectieuse contagieuse causeacutee par Mycobacterium bovis Cette maladie touche les bovins
et de nombreuses espegraveces de mammifegraveres domestiques et sauvages ainsi que lrsquohomme La circulation de la bacteacuterie dans des systegravemes
multi-hocirctes varieacutes favorise lrsquoentretien de la maladie et la contamination des bovins vivant agrave proximiteacute des animaux sauvages infecteacutes En
marge de la transmission directe de M bovis par voie respiratoire la transmission indirecte aux bovins lieacutee agrave lrsquoinhalation ou agrave lrsquoingestion de
matrices environnementales contamineacutees par un animal infecteacute excreacuteteur est suspecteacutee dans plusieurs reacutegions du monde Lrsquoexistence de
reacuteservoirs environnementaux ougrave le bacille M bovis est capable de persister pourrait donc ecirctre un facteur important de la reacuteeacutemergence puis
du maintien de la maladie dans les systegravemes multi-hocirctes
En Cocircte drsquoOr deacutepartement fortement toucheacute par la tuberculose bovine depuis 2004 la transmission indirecte de la bacteacuterie entre la faune
sauvage infecteacutee et les bovins est suspecteacutee dans plusieurs eacutelevages Pour eacutevaluer la preacutesence et la survie de cette bacteacuterie dans
lrsquoenvironnement nous avons analyseacute un grand nombre drsquoeacutechantillons preacuteleveacutes dans des zones partageacutees par les bovins etou la faune sauvage
infecteacutes dans le but de deacuteterminer la distribution environnementale de M bovis Pour ce faire nous avons deacuteveloppeacute ou modifieacute des systegravemes
de deacutetection moleacuteculaire adapteacutes aux matrices environnementales complexes Nous avons eacutegalement eacutevalueacute lrsquoimpact de la tempeacuterature et
des proprieacuteteacutes physico-chimiques de deux sols sur la survie de M bovis ainsi que le rocircle de la meacutesofaune du sol (lombrics en particulier)
dans la disseacutemination de la bacteacuterie agrave partir de matiegravere organique contamineacutee Lrsquoeacutetude environnementale a mis plus particuliegraverement en
eacutevidence la contamination de deux biotopes les zones humides des pacirctures et les sols de terriers de blaireaux De plus les eacutetudes
expeacuterimentales ont montreacute que M bovis pouvait survivre plusieurs mois dans le sol agrave 4degC et que les lombrics pouvaient disseacuteminer la bacteacuterie
dans le sol voire jouer un rocircle potentiel de vecteur pour les animaux qui les consomment Ces reacutesultats apportent de nouvelles connaissances
sur la persistance et la circulation de M bovis dans lrsquoenvironnement en Cocircte drsquoOr et permettront de proposer des ameacuteliorations aux mesures
de bioseacutecuriteacute deacutejagrave existantes dans les eacutelevages bovins
Mots Cleacutes Mycobacterium bovis environnement sol eau fegraveces bovins faune sauvage qPCR culture
Abstract Bovine tuberculosis is an infectious disease caused by Mycobacterium bovis This disease affects multiple species of domestic
livestock notably cattle wild mammals and humans The circulation of the bacteria within complex multi-host systems promotes the
maintenance of the disease and the reinfection of cattle Besides the classical direct transmission of the bacteria through the respiratory
route indirect transmission through inhalation or ingestion of environmental substrates contaminated by the excretions of infected animals
is suspect in several countries Environmental contamination with M bovis appears to be a crucial factor in the persistence of the infection in
multi-host systems
In Cocircte dOr a French department impacted by bovine tuberculosis since 2004 the indirect transmission of the bacteria from infected wildlife
to cattle is now suspect in several cases To assess this mode of transmission we evaluated environmental contamination with M bovis on a
large number of samples collected in areas shared by cattle and infected wildlife For this purpose we both developed and modified molecular
detection systems adapted for complex environmental substrates We also assessed the impact of temperature and soil physicochemical
properties on the survival of M bovis as well as the role of earthworms in the dissemination of the bacteria from contaminated organic
material The environmental study showed contamination of two habitats in particular wetland pastures and soil badger setts Moreover
results indicated that M bovis can survive in soil for several months at 4degC and that worms may disseminate the bacteria in the soil and even
serve as a vector for the mammals that consume them This study increases our understanding of the persistence and the circulation of
M bovis in the environment in Cocircte dOr In addition these results will be used to improve the existing biosecurity measures on cattle farms
in the region
Key words Mycobacterium bovis environment soil water feces cattle wildlife qPCR culture

Remerciements
Mes premiers remerciements vont agrave mes encadrants de thegravese Alain Hartmann et Maria-Laura
Boschiroli Merci de mrsquoavoir confieacute ce sujet de thegravese passionnant et de votre accompagnement vos
conseils pertinents et vos encouragements tout au long de ces trois anneacutees
Un grand merci Eric Gueneau co-encadrant de ce travail et directeur du Laboratoire
Deacutepartemental de Cocircte drsquoOr pour son aide et la mise agrave disposition du P3 pour lrsquoincubation de mes
cultures Je remercie eacutegalement chaleureusement toute son eacutequipe de choc en microbiologie et
biologie moleacuteculaire Veacuteronique Julie Virginie et toute lrsquoeacutequipe
Je remercie Franccediloise Lucas et Franck Biet drsquoavoir accepteacute drsquoecirctre les rapporteurs de ce travail
et drsquoen avoir eacutevalueacute la pertinence Merci eacutegalement agrave Sylvie Nazaret Jaquemine Vialard Pascal
Simonet et Jean-Franccedilois Cavin pour leur participation au jury de thegravese
Merci agrave Barbara Dufour Loiumlc Bollache et Jean-Louis Moyen pour leur participation au comiteacute
de pilotage de thegravese et leurs conseils pendant ce travail
Un eacutenorme merci agrave la DDPP21 pour toutes les informations essentielles fournies sur les foyers
bovins et la faune sauvage Aneacutelise Taconnet Fabienne Bartheacuteleacutemy Thierry Grosjean Un merci
particulier agrave Brigitte Biasino pour son aide preacutecieuse lors drsquoune campagne de preacutelegravevements fort
sympathique
Des remerciements chaleureux eacutegalement agrave Fabrice Chevalier et Marina Beacuteral de la CIREV
Bourgogne pour les donneacutees partageacutees mais eacutegalement les magnifiques cartes illustrant mon
manuscrit
Merci aux organismes qui ont contribueacute financiegraverement agrave ce travail en particulier le GDS 21
la Chambre drsquoAgriculture de la Cocircte drsquoOr La MSA Bourgogne et la MSA nationale Interbev et le
Conseil Deacutepartemental de la Cocircte drsquoOr
Un grand merci agrave Violette Sicre (DDPP16 et Sylvatub) pour son accueil chaleureux et son
efficaciteacute lors de mon seacutejour en Charente Merci eacutegalement au directeur du laboratoire de Charente et
agrave son eacutequipe pour leur accueil
Des remerciements au laboratoire de bacteacuteriologie du CHU de Dijon qui a mis son laboratoire
L3 agrave notre disposition pour lrsquoeacutetude sur la survie de M bovis dans le sol Merci agrave Jean-Marie Duez pour
le temps passeacute et sa confiance merci agrave Catherine Neuwirth Julien Bador et toute lrsquoeacutequipe
Un grand merci agrave lrsquoeacutequipe de lrsquoANSES Maisons-Alfort notamment Krystel De Cruz pour le
geacutenotypage des ADN environnementaux mais eacutegalement Claudine Karaoui Lorraine Michelet et le
reste de lrsquoeacutequipe
Merci agrave Franck Biet et Thierry Cochard de lrsquoINRA de Tours qui nous ont fourni les souches de
Map et M smegmatis
Des eacutenormes remerciements pour les lieutenants de louveterie les pieacutegeurs et les chasseurs
qui mrsquoont accompagneacutee dans mes preacutelegravevements sur le terrain et sans qui je nrsquoaurai pas pu preacutelever
tous ces eacutechantillons Jeannick Gautherot Dominique Rigaud Yannick Thibert Georges Balluet Jean-
Luc Moindrot Marcel Guillier qui connaissent les bois comme leur poche et font un travail
consideacuterable
Je remercie les eacuteleveurs qui ont participeacute au travail en nous accompagnant sur leurs pacirctures
Merci agrave Leacuteon Fayolle pour son expertise sur les lombrics et Dominique Meunier pour son
expertise sur les formations peacutedologiques de lrsquoAuxois merci agrave Luc Biju-Duval pour les cartes reacutealiseacutees
Un eacutenorme merci agrave tous les membres du pocircle MERS qui ont fait de ces trois anneacutees un moment
extrecircmement riche en eacutechanges scientifiques humains amicaux et sportifshellipMerci agrave Dominique
Pascal Laurent Jean-Paul Seacutebastien Un merci particulier aux filles de lrsquoeacutequipe Geacuteraldine Ceacutecile
Laetitia pour leur gentillesse leurs compeacutetences et tous les bons moments passeacutes agrave discuterhellipet agrave
marcher Des remerciements tout particuliers agrave Murielle Rochelet pour ses conseils hyper-aviseacutes sa
rigueur son esprit drsquoinnovation son anglais parfait et encore plein plein drsquoautres choses Merci
infiniment pour tout le temps que tu as passeacute agrave me relire et me corrigerhellip
Un grand mais alors tregraves grand merci agrave Benoicirct sans qui lrsquoexpeacuterience lombric nrsquoaurait pas eacuteteacute
aussihellipexceptionnelle Bon courage pour ta thegravese Merci agrave Cleacutement pour les kilos de sols de terriers
ramasseacutes peseacutes analyseacutes lors de ces deux stages Un grand merci agrave Pierre pour sa gentillesse rare son
travail tregraves seacuterieux et son aide inestimable pour la mise en page la relecture et lrsquoimpression de ce
manuscrit
Merci eacutegalement agrave mes collegravegues du Master AE Caroline Alexandre Fam-Ky Justin Marion
et tous les autres qui mrsquoont parfois (souvent) aideacutee agrave supporter de longues heures de cours
Merci agrave tous ceux que jrsquoai croiseacute agrave lrsquoINRA et ailleurs qui ont fait de ces trois ans une expeacuterience
humaine et scientifique inoubliable Ariane Emilie Kadiya Emeline Ceacuteline Heacutelegravene Charline Anne-
Laure Aude Gilles David Marion Nicolas Romain Fabrice Chloeacute Aymeacute Laure Luiz Marie-
Christine Nadine Estelle Amy Amandinehellip et tous ceux que jrsquoai sucircrement oublieacute
Enfin pour finir un grand merci agrave Steacutephane Lola Lucien et Antonin qui ont supporteacute mes hauts
et mes bas pendant ces trois anneacutees agrave ma famille et ma belle-famille qui ont toujours eacuteteacute lagravehellip
Reacutesumeacute
La tuberculose bovine est une maladie infectieuse contagieuse causeacutee par Mycobacterium
bovis Cette maladie touche les bovins et de nombreuses espegraveces de mammifegraveres domestiques et
sauvages ainsi que lrsquohomme La circulation de la bacteacuterie dans des systegravemes multi-hocirctes varieacutes favorise
lrsquoentretien de la maladie et la contamination des bovins vivant agrave proximiteacute des animaux sauvages
infecteacutes En marge de la transmission directe de M bovis par voie respiratoire la transmission indirecte
aux bovins lieacutee agrave lrsquoinhalation ou agrave lrsquoingestion de matrices environnementales contamineacutees par un
animal infecteacute excreacuteteur est suspecteacutee dans plusieurs reacutegions du monde Lrsquoexistence de reacuteservoirs
environnementaux ougrave le bacille M bovis est capable de persister pourrait donc ecirctre un facteur
important de la reacuteeacutemergence puis du maintien de la maladie dans les systegravemes multi-hocirctes
En Cocircte drsquoOr deacutepartement fortement toucheacute par la tuberculose bovine depuis 2004 la
transmission indirecte de la bacteacuterie entre la faune sauvage infecteacutee et les bovins est suspecteacutee dans
plusieurs eacutelevages Pour eacutevaluer la preacutesence et la survie de cette bacteacuterie dans lrsquoenvironnement nous
avons analyseacute un grand nombre drsquoeacutechantillons preacuteleveacutes dans des zones partageacutees par les bovins etou
la faune sauvage infecteacutes dans le but de deacuteterminer la distribution environnementale de M bovis Pour
ce faire nous avons deacuteveloppeacute ou modifieacute des systegravemes de deacutetection moleacuteculaire adapteacutes aux matrices
environnementales complexes Nous avons eacutegalement eacutevalueacute lrsquoimpact de la tempeacuterature et des
proprieacuteteacutes physico-chimiques de deux sols sur la survie de M bovis ainsi que le rocircle de la meacutesofaune
du sol (lombrics en particulier) dans la disseacutemination de la bacteacuterie agrave partir de matiegravere organique
contamineacutee Lrsquoeacutetude environnementale a mis plus particuliegraverement en eacutevidence la contamination de
deux biotopes les zones humides des pacirctures et les sols de terriers de blaireaux De plus les eacutetudes
expeacuterimentales ont montreacute que M bovis pouvait survivre plusieurs mois dans le sol agrave 4degC et que les
lombrics pouvaient disseacuteminer la bacteacuterie dans le sol voire jouer un rocircle potentiel de vecteur pour les
animaux qui les consomment Ces reacutesultats apportent de nouvelles connaissances sur la persistance et
la circulation de M bovis dans lrsquoenvironnement en Cocircte drsquoOr et permettront de proposer des
ameacuteliorations aux mesures de bioseacutecuriteacute deacutejagrave existantes dans les eacutelevages bovins
Mots cleacutes Mycobacterium bovis environnement sol eau fegraveces bovins faune sauvage qPCR culture
Abstract
Bovine tuberculosis is an infectious disease caused by Mycobacterium bovis This disease
affects cattle and many species of domestic and wild mammals and humans The circulation of the
bacteria in various multi-host systems promotes the maintenance of the disease and the contamination
of cattle in the vicinity Beside direct transmission of the bacteria through the respiratory route indirect
transmission through inhalation or ingestion of environmental matrices contaminated by an infected
animal excretory is suspected in several countries Environmental contamination with M bovis appears
to be a crucial factor in the persistence of the infection in multi-host systems
In Cocircte dOr a French department affected by bovine tuberculosis since 2004 the indirect
transmission of the bacteria from infected wildlife to cattle is suspected in several cases To assess this
type of transmission of the bacillus we evaluated the environmental contamination with M bovis on
a large number of samples taken in areas shared by cattle and or wildlife infected For this purpose
we developed or modified molecular detection systems adapted for environmental complex matrices
We also assessed the impact of physicochemical properties of both soil and temperature on survival of
M bovis and the role of earthworms in the spread of the bacteria from contaminated organic material
The environmental study showed the contamination of two media in particular wetlands pastures and
soil badger setts Moreover experimental studies have shown that M bovis can survive in soil for
several months at 4 deg C and the worms could spread the bacteria in the soil or even play a potential
role for vector animals that consume them These results will propose improvements to existing
biosecurity measures on cattle farms and provide new knowledge about the persistence and circulation
of M bovis in the environment in Cocircte dOr
Key words Mycobacterium bovis environment soil water feces cattle wildlife qPCR culture
Table des matiegraveres
INTRODUCTION GENERALE 1
CHAPITRE 1 - CONTEXTE BIBLIOGRAPHIQUE 4
A LES MYCOBACTERIES DIVERSITE PHYLOGENIE SPECTRE DrsquoHOcircTES ET VIRULENCE 4
1 Le genre Mycobacterium taxonomie et meacutethodes de classification 4
2 Caracteacuteristiques geacuteneacuterales des mycobacteacuteries 5
a Morphologie des mycobacteacuteries 5
b Caracteacuteristiques culturales pheacutenotypiques et physiologiques 7
c Phylogeacutenie du genre Mycobacterium 8
d Caracteacuteristiques geacuteneacutetiques des mycobacteacuteries 9
3 Description des diffeacuterents Complexes ou espegraveces de mycobacteacuteries 10
a Les mycobacteacuteries non pathogegravenes potentiellement opportunistes 11
b Les mycobacteacuteries saprophytes non pathogegravenes 12
c Les mycobacteacuteries pathogegravenes obligatoires 12
Les mycobacteacuteries responsables de legravepre 12
Les mycobacteacuteries tuberculeuses le Complexe tuberculosis (MTBC) 13
B LA TUBERCULOSE UNE MALADIE TRES ANCIENNE MAIS UNE MENACE TOUJOURS ACTUELLE 20
1 Introduction 20
2 Histoire de la tuberculose 20
3 La tuberculose bovine 21
a Introduction 21
b Les hocirctes et les communauteacutes drsquohocirctes de Mycobacterium bovis dans le monde et leurs statuts
eacutepideacutemiologiques 22
c Aspect zoonotique de Mycobacterium bovis 23
d Pathogeacutenie de la tuberculose bovine 24
Facteurs influenccedilant le pouvoir pathogegravene de M bovis voies drsquoinfection et dose infectante 24
Facteurs influenccedilant la reacuteceptiviteacute et la sensibiliteacute de lrsquohocircte 25
Les eacutetapes de lrsquoinfection et la reacuteponse immunitaire associeacutee 26
Expression clinique de la maladie et leacutesions observeacutees 27
e Le deacutepistage et le diagnostic de lrsquoinfection chez les animaux 28
f Epideacutemiologie de la tuberculose bovine 30
Evolution de la tuberculose bovine dans le monde et en Europe et mesures de lutte 30
Evolution de la situation sanitaire en France 33
Evolution de la situation sanitaire en Cocircte drsquoOr 38
C ECOLOGIE DE M BOVIS DANS LrsquoENVIRONNEMENT 42
1 Introduction 42
2 Origines de la contamination environnementale par M bovis 42
a Lrsquoexcreacutetion animale de Mycobacterium bovis 42
b Le transfert de Mycobacterium bovis par les effluents drsquoeacutelevage ou urbains 44
3 Deacutetection et survie de Mycobacterium bovis dans lrsquoenvironnement 45
a Deacutetection de Mycobacterium bovis en conditions naturelles 45
b Dureacutees de survie de Mycobacterium bovis en conditions expeacuterimentales 47
4 Strateacutegies de survie des mycobacteacuteries dans lrsquoenvironnement 54
a Lrsquoeacutetat de dormance 54
b La sporulation 55
c Interactions des mycobacteacuteries avec la faune du sol 56
d Interactions des mycobacteacuteries avec les diptegraveres 57
e La formation de biofilms 57
5 Facteurs de risque de contamination de lrsquoenvironnement et de transmission de Mycobacterium
bovis aux animaux par lrsquoenvironnement contamineacute 58
a Risques lieacutes agrave lrsquoeacutecologie des espegraveces sauvages sensibles agrave Mycobacterium bovis 58
b Risques lieacutes aux pratiques drsquoeacutelevage 60
c Risques lieacutes aux pratiques cyneacutegeacutetiques 61
d Risques lieacutes aux facteurs paysagers climatiques et peacutedologiques 62
D PRINCIPAUX OUTILS DE DETECTION DE PHENOTYPAGE ET DE GENOTYPAGE APPLICABLES AU MTBC ET
PLUS PARTICULIEREMENT A MYCOBACTERIUM BOVIS 64
1 La deacutetection des mycobacteacuteries par microscopie 64
2 Les meacutethodes de culture des mycobacteacuteries 65
a Les diffeacuterents traitements preacutealables agrave la mise en culture des eacutechantillons 65
b La culture sur milieu solide 66
c La culture en milieu liquide 67
3 Meacutethodes de deacutetection et drsquoidentification des bacteacuteries du MTBC 68
a Caracteacuterisation pheacutenotypique et biochimique des mycobacteacuteries du MTBC 69
b Lrsquoidentification des mycobacteacuteries par recherche de biomarqueurs 69
c Lrsquoidentification des mycobacteacuteries par des meacutethodes drsquohybridation coupleacutees ou non agrave la PCR 70
d Le seacutequenccedilage 71
e Deacutetection et geacutenotypage des mycobacteacuteries du MTBC les marqueurs utiliseacutes 72
Les seacutequences drsquoinsertion 72
Les courtes seacutequences drsquoADN reacutepeacuteteacutees 73
Reacutegions de Diffeacuterence (RD) et Single Nucleotide Polymorphism (SNP) 78
f Identification par seacutequenccedilage complet des geacutenomes 79
g Meacutethode speacutecifique drsquoidentification des mycobacteacuteries animales utiliseacutee par le Laboratoire National de
reacutefeacuterence de la tuberculose bovine LNR TB 79
OBJECTIFS DE LA THESE 82
CHAPITRE 2 - DEVELOPPEMENT ET OPTIMISATION DrsquoOUTILS POUR LA DETECTION DE MYCOBACTERIUM
BOVIS DANS LES ECHANTILLONS ENVIRONNEMENTAUX 86
A DEVELOPPEMENT DrsquoOUTILS MOLECULAIRES ADAPTES AUX MATRICES ENVIRONNEMENTALES 86
CHAPITRE 3 - ETUDE DE LA PREVALENCE ENVIRONNEMENTALE DE MYCOBACTERIUM BOVIS DANS LA ZONE
ENDEMIQUE DE TUBERCULOSE BOVINE DE COTE DrsquoOR ET RESULTATS OBTENUS EN CHARENTE 93
A AVANT-PROPOS 93
B FIRST MOLECULAR DETECTION OF MYCOBACTERIUM BOVIS IN ENVIRONMENTAL SAMPLES FROM A
FRENCH REGION WITH ENDEMIC BOVINE TUBERCULOSIS 95
C CAMPAGNES DE PRELEVEMENTS SUPPLEMENTAIRES 121
1 Fin des preacutelegravevements en Cocircte drsquoOr 121
2 Preacutelegravevements en Charente 123
3 Compleacutements paysage geacuteomorphologie et peacutedologie 125
D BILAN GLOBAL 130
CHAPITRE 4 - IMPACT DE LA TEMPERATURE ET DE LA NATURE DU SOL SUR LA SURVIE DE MYCOBACTERIUM
BOVIS SB0120 133
A AVANT-PROPOS 133
B EXPERIMENTAL INVESTIGATION OF THE IMPACT OF THE TEMPERATURE AND THE SOIL TYPE ON
MYCOBACTERIUM BOVIS SURVIVAL IN THE ENVIRONMENT 135
C BILAN 150
CHAPITRE 5 - ROLE DE LA FAUNE DU SOL DANS LA CIRCULATION DE MYCOBACTERIUM BOVIS ETUDE DE SA
DISSEMINATION PAR LES LOMBRICS 155
A AVANT-PROPOS 155
B RAPID DISSEMINATION OF M BOVIS FROM CATTLE DUNG TO SOIL BY THE EARTHWORM LUMBRICUS
TERRESTRIS 157
C COMPLEMENT PRELEVEMENT ET ANALYSE DE LOMBRICS DANS UNE PATURE SUSPECTE 174
D BILAN 175
DISCUSSION GENERALE 177
1 Des meacutethodes deacuteveloppeacutees (ou adapteacutees) puis valideacutees dans des matrices environnementales 179
2 Contamination environnementale les zones humides les eaux de ruissellement et les sols de
terriers sont potentiellement agrave risque 180
3 Survie et circulation de M bovis dans lrsquoenvironnement 183
PERSPECTIVES 185
REFERENCES BIBLIOGRAPHIQUES 191
ANNEXES 221
Liste des Figures
Figure 1 Structure et composition de lenveloppe des mycobacteacuteries (drsquoapregraves Marrakchi Laneacuteelle and Daffeacute 2014) AG arabinogalactane PG peptidoglycane TDM treacutehalose dimycolate PL phospholipides TMM treacutehalose monomycolate GL glycolipides 7
Figure 2 Arbre phylogeacutenique baseacute sur lrsquoanalyse de lrsquoARNr 16S des principaux complexes et espegraveces de mycobacteacuteries (drsquoapregraves Magee et Ward 2012) 9
Figure 3 Phylogeacutenie des souches du MTBC baseacutee sur les Reacutegions de Diffeacuterences et diffeacuterents Single Nucleotide Polymorphism (Dippenaar et al 2015) 14
Figure 4 Aspect des colonies de M bovis sur milieu de Lowenstein-Jensen (agrave gauche) et Middlebrook 7H11 suppleacutementeacute (agrave droite) 17
Figure 5 Evolution de la reacuteponse immunitaire chez le bovin infecteacute par M bovis au cours du temps et aux diffeacuterents tests de deacutepistage de la tuberculose bovine (de la Rua-Domenech et al 2006 adapteacute de Vordermeier et al 2004) 27
Figure 6 Carte de distribution de la tuberculose bovine chez les bovins entre juillet et deacutecembre 2014 (Source WAHID Interface) 31
Figure 7 Statuts des pays europeacuteens vis-agrave-vis de la tuberculose bovine en 2012 et proportion de bovins infecteacutes OTF Officially Tuberculosis Free (officiellement indemne de tuberculose bovine) MS Member State (Etat membre) (Source EFSA 2014) 32
Figure 8 Evolution du taux de preacutevalence annuelle national de la tuberculose bovine chez les bovins (Beacutenet et al 2006) 34
Figure 9 Evolution de la preacutevalence et de lrsquoincidence de la tuberculose bovine en France entre 1995 et 2003 (Cavalerie et al 2014) 35
Figure 10 Incidence et distribution de la tuberculose bovine en France dans les anneacutees 50 (en haut) et entre 2000 et 2013 (en bas) (Cavalerie et al 2014) 36
Figure 11 Reacutepartition et reacutesultats des analyses effectueacutees dans le cadre du dispositif Sylvatub chez le grand gibier drsquoaoucirct 2013 agrave aoucirct 2014 et chez les blaireaux en 2014 (Source Sylvatub) 38
Figure 12 Localisation du parcellaire des cheptels infecteacutes par la tuberculose bovine entre 2010 et 2015 et spoligotypes incrimineacutes (Source CIREV) 40
Figure 13 Nombre de nouveaux foyers bovins et de foyers recontamineacutes entre 2002 et 2014 (Donneacutees DDPP21) 40
Figure 14 Nombre de cervideacutes sangliers et blaireaux infecteacutes deacutecouverts en Cocircte drsquoOr entre 2002 et 2014 41
Figure 15 Repreacutesentation scheacutematique de la circulation de M bovis dans lrsquoenvironnement agricole (modifieacute drsquoapregraves Elliott et al 2014) 63
Figure 16 Profils de restriction disolats de M tuberculosis obtenus suite agrave une RFLP-IS6110 Les isolats 3 5 6 9 et 10 ont le mecircme profil et posseacutedaient un lien eacutepideacutemiologique S repreacutesente le marqueur de poids moleacuteculaire (Source web site CDC) 73
Figure 17 Principe du spoligotypage des bacteacuteries du MTBC dapregraves Kamerbeek et al (1997) A Repreacutesentation du locus DR de M tuberculosis H37Rv et M bovis BCG B Amplifications des espaceurs par un couple damorces (a et b) et amplicons obtenus C Profils drsquohybridation sur membrane (un carreacute noir signale la preacutesence de lrsquoespaceur les parties blanches leur absence) 75
Figure 18 Diffeacuterences geacutenomiques de la reacutegion RD1 entre M bovis BCG (A) M microti (C) par comparaison agrave M tuberculosis H37Rv (B) drsquoapregraves Brodin et al 2002 et localisation du locus Rv3866 RD1mic correspond agrave la seacutequence de M tuberculosis deacuteleacuteteacutee chez M microti RD1bcg correspond agrave la seacutequence de M tuberculosis deacuteleacuteteacutee chez M bovis BCG 88
Figure 27 Localisation des communes de Cocircte drsquoOr sur lesquelles de foyers bovins et des animaux sauvages infecteacutes ont eacuteteacute deacutetecteacutes et localisation des preacutelegravevements environnementaux effectueacutes en 2015 (les preacutelegravevements positifs sont repreacutesenteacutes par des eacutetoiles noires) (Reacutealisation Marina Beacuteral CIREV Bourgogne) 123
Figure 28 Localisation des communes de Charente sur lesquelles de foyers bovins et des animaux sauvages infecteacutes ont eacuteteacute deacutetecteacutes et localisation des preacutelegravevements environnementaux effectueacutes en 2015 (les preacutelegravevements positifs sont repreacutesenteacutes par des eacutetoiles noires) (Reacutealisation Marina Beacuteral CIREV Bourgogne) 125
Figure 29 Orographie et hydrographie de lrsquoAuxois Les valleacutees supportant les parcelles suspectes sont indiqueacutees par une flegraveche noire (Source BD Carthage et BD alti - IGN) 127
Figure 30 Localisation de lrsquoAuxois en Cocircte drsquoOr structure paysagegravere et geacuteomorphologie de ce territoire 127
Figure 31 Nombre de vaches allaitantes par canton dans le deacutepartement de la Cocircte drsquoOr (agrave gauche) et parcellaire des foyers bovins de 2010 agrave 2015 (agrave droite) (Source Recensement agricole 2010 ndash Agreste) 128
Figure 32 Reacutepartition spatiale des parcelles des eacutelevages bovins infecteacutes de 2010 agrave 2014 en Cocircte drsquoOr et repreacutesentation des couches peacutedologiques (Reacutealisation Marina Beacuteral CIREV Bourgogne) 129
Figure 33 Tempeacuteratures minimales et maximales observeacutees agrave Dijon-Longvic de 2012 agrave 2015 (Source Meacuteteacuteo France) 151
Liste des Tableaux
Tableau 1 Dureacutees de survie de M bovis dans diffeacuterentes matrices environnementales obtenues en conditions expeacuterimentales naturelles ou de laboratoire 49
Tableau 2 Identification des principales mycobacteacuteries du MTBC par des observations pheacutenotypiques et des tests biochimiques 69
Tableau 3 Cibles moleacuteculaires deacutecrites dans la litteacuterature pour la deacutetection du Complexe tuberculosis ou de M bovis dans les eacutechantillons environnementaux 76
Tableau 4 Spoligotypes et geacutenotypes MIRU-VNTR des souches de M bovis isoleacutees chez les bovins et la faune sauvage en Cocircte dOr et en Dordogne (drsquoapregraves Hauer et al 2015) 77
Tableau 5 Systegravemes de deacutetection utiliseacutes en qPCR en premiegravere intention pour lidentification dune mycobacteacuterie dorigine animale et reacutesultats obtenus 80
Liste des Annexes
Annexe 1 Deacutetection de Mycobacterium bovis dans lrsquoenvironnement Premiers reacutesultats des recherches meneacutees en Cocircte-drsquoOr 223
Annexe 2 Preacuteparation du Middlebrook 7H9 235
Annexe 3 Preacuteparation du Middlebrook 7H11 237
Liste des Abreacuteviations
ADC Albumine Dextrose Catalase
ADNDNA Acide deacutesoxyribonucleacuteique
AEEMA Association pour lEtude de lEpideacutemiologie des Maladies Animales
AFNOR Agence Franccedilaise de NORmalisation
ANSES Agence nationale de seacutecuriteacute sanitaire de lrsquoalimentation de lrsquoenvironnement et du travail
ARNRNA Acide ribonucleacuteique
ARNr ARN ribosomique
BAAR Bacille acido-alcoolo reacutesistant
BCG Bacille de Calmette et Gueacuterin
BLAST Basic Local Alignment Search Tool
bTB bovine Tuberculosis
CDC Centers of Disease Control and Prevention
CFP-10 Culture Filtrat Protein 10 kD
CIREV Cellules Inter Reacutegionales dEpideacutemiologie Veacuteteacuterinaire
CRISPR Clustered Regularly Interspersed Short Palindromic Repeats
Ct Cycle Threshold
DDPP Direction Deacutepartementale de la Protection des Populations
DEFRA Department for Environment Food and Rural Affairs
DGAL Direction Geacuteneacuterale de lALimentation
DosR Dormancy Survival Regulator
DR Direct Repeat
EFSA European Food Safety Authority
ELISA Enzyme-Linked ImmunoSorbent Assay
ESAT-6 Early Secreted Antigenic Target 6kD
ETR Exact Tandem Repeat
Fbp Fibronectin binding protein
FISH Fluorescence In Situ Hybridization
GC-MS Gas Chromatography-Mass Spectrometry
GDS Groupements de Deacutefense Sanitaire
HAP Hydrocarbures Aromatiques Polycycliques
HIV Human Immunodeficiency Virus
HPLC High Performance Liquid Chromatography
IDC Intradermotuberculination comparative
IDR Intradermotuberculination
IDS Intradermotuberculination simple
IFN-γ Interfeacuteron γ
INRA Institut National de la Recherche Agronomique
Ion PGM Ion Personal Genome Machine
IS Insertion Sequence
ITS Internal Transcribed Spacer
kDkDa kilo Daltons
LNR Laboratoire National de Reacutefeacuterence
LSP Large Sequence Polymorphism
Maa Mycobacterium avium subspecies avium
MAC Mycobacterium avium complex
MALDI-TOF Matrix-Assisted Laser Desorption Ionization-Time Of Flight
Map Mycobacterium avium subspecies paratuberculosis
MDR Multi Drug Resistant
MIRU Mycobacterial Interspersed Repetitive Unit
MLVA Multi Locus VNTR Analysis
MNT Mycobacteacuteries Non Tuberculeuses
MS Member State
MTBC Mycobacterium tuberculosis complex
NCBI National Center for Biotechnology Information
NF Norme Franccedilaise
NGS Next Generation Sequencing
OADC Oleic acid Albumine Dextrose Catalase
OIE Office International des Epizooties
OMS Organisation Mondiale de la Santeacute
ONCFS Office National de la Chasse et de la Faune Sauvage
ONEMA Office National de lEau et des Milieux Aquatiques
OTF Officialy Tuberculosis Free
pb paire de bases
PCR Polymerase chain reaction
PNA Peptide Nucleic Acid
qPCR quantitative Polymerase Chain Reaction
RD Reacutegions de Diffeacuterence
RFLP Restriction Fragment Lenght Polymorphism
RNS Reactive Nitrogen Species
ROS Reactive Oxygen Species
RT-PCR Reverse-Transcription Polymerase Chain Reaction
SNP Single Nucleotide Polymorphism
ssp subspecies
TDM Treacutehalose dimycolate
TMM Treacutehalose monomycolate
UFCCFU Uniteacute Formant Colonie
VBNC Viable But Non Cultivable
VNTR Variable Number Tandem Repeat
WAHID World Animal Health Information Database
WGS Whole Genome Sequencing
WHO World Health Organization
XDR eXtensively Drug Resistant
1
Introduction geacuteneacuterale
La tuberculose bovine est une maladie drsquoimportance sanitaire majeure en santeacute animale agrave
travers le monde Son agent principal Mycobacterium bovis est un membre du Complexe
Mycobacterium tuberculosis qui regroupe les bacteacuteries tuberculeuses infectant lrsquohomme et les
mammifegraveres Les bovins constituent le reacuteservoir originel de M bovis mais de nombreuses espegraveces de
mammifegraveres peuvent eacutegalement ecirctre infecteacutees dont lrsquohomme Les enjeux sanitaires eacuteconomiques et
sociaux induits par cette maladie ont conduit agrave la mise en place de programmes visant lrsquoeacuteradication de
cette maladie chez les bovins agrave travers le monde Dans les pays deacuteveloppeacutes ces mesures associeacutees agrave
la pasteurisation du lait ont fortement reacuteduit lrsquoincidence de la tuberculose bovine voire eacuteradiqueacute
lrsquoinfection chez les bovins Toutefois dans quelques pays lrsquoinfection persiste dans les cheptels bovins
malgreacute des actions de preacutevention de controcircle et de surveillance Dans les pays en deacuteveloppement ougrave
les controcircles organiseacutes sont quasi- voire totalement absents la preacutevalence et la propagation de la
tuberculose bovine sont inquieacutetantes et repreacutesentent un risque sanitaire et eacuteconomique vis-agrave-vis des
populations sensibles En conseacutequence la tuberculose bovine attire chaque anneacutee un peu plus
lrsquoattention de la communauteacute internationale en raison du nombre croissant de troupeaux infecteacutes et
des effets sur les productions animales ainsi que du risque que la maladie fait planer sur la santeacute
publique
Le passage de lrsquoinfection du reacuteservoir bovin agrave diffeacuterentes populations sauvages qui
entretiennent la maladie et peuvent la transmettre en retour aux bovins a fortement compliqueacute les
scheacutemas drsquoeacuteradication de la maladie dans plusieurs reacutegions du monde et explique lrsquoeacutechec partiel des
programmes de lutte mis en place Alors que la transmission directe du bacille par voie respiratoire
semble ecirctre le mode de transmission preacutepondeacuterant au sein drsquoune mecircme espegravece la transmission
indirecte de M bovis est fortement suspecteacutee entre espegraveces partageant les mecircmes territoires et
ressources compliquant encore lrsquoeacutepideacutemiologie de cette maladie
En France une recrudescence du nombre de cheptels bovins infecteacutes dans certains
deacutepartements dont la Cocircte drsquoOr menace son statut de pays laquo officiellement indemne de tuberculose
bovine raquo obtenu en 2001 Dans ce deacutepartement ougrave plus de 190 foyers bovins ont eacuteteacute deacutetecteacutes depuis
2003 M bovis circule dans un systegraveme multi-hocirctes incluant les bovins les cervideacutes les blaireaux et les
sangliers dans une zone geacuteographique localiseacutee Face agrave des contaminations ou des recontaminations
inexpliqueacutees de cheptels avec des souches identiques agrave celles retrouveacutees dans la faune sauvage
infecteacutee deacutetecteacutee agrave proximiteacute dans des eacutelevages le rocircle de lrsquoenvironnement a eacuteteacute suggeacutereacute dans la
transmission indirecte du bacille
Dans ce contexte mes travaux de thegravese visent agrave eacutevaluer la contamination environnementale
2
des zones partageacutees par les bovins et la faune sauvage infecteacutes gracircce agrave des meacutethodes adapteacutees
utilisables sur un grand nombre de preacutelegravevements et agrave rechercher les facteurs favorisant la survie et la
circulation de M bovis dans lrsquoenvironnement Le premier chapitre de ce manuscrit est une synthegravese
des connaissances relatives aux mycobacteacuteries en geacuteneacuteral et aux mycobacteacuteries tuberculeuses en
particulier notamment en terme drsquoeacutecologie de deacutetection et drsquoidentification Le deuxiegraveme chapitre
deacutetaille le deacuteveloppement des meacutethodes moleacuteculaires utiliseacutees pour le diagnostic environnemental
ainsi que le deacuteveloppement de deux nouveaux protocoles pour isoler les mycobacteacuteries du sol et suivre
leur croissance en milieu liquide Le troisiegraveme chapitre preacutesente les modaliteacutes drsquoeacutetude et les reacutesultats
obtenus entre 2013 et 2015 dans lrsquoeacutevaluation de la contamination environnementale en Cocircte drsquoOr Les
reacutesultats drsquoune eacutetude reacutealiseacutee en Charente sont eacutegalement preacutesenteacutes Le quatriegraveme chapitre traite de
lrsquoinfluence de la tempeacuterature et du type de sol sur la dureacutee de survie de M bovis Enfin le cinquiegraveme
chapitre srsquointeacuteresse au rocircle des lombrics sur la disseacutemination de M bovis dans le sol Pour terminer
une discussion geacuteneacuterale syntheacutetise lrsquoensemble des reacutesultats obtenus lors de ce travail et ouvre de
nombreuses perspectives de recherche
Cette thegravese financeacutee par la Reacutegion Bourgogne entre dans le dispositif Jeunes Chercheurs
Entrepreneurs (JCE) Le travail scientifique de recherche srsquoaccompagne de la reacutealisation drsquoun Master
des Entreprises (Master 2) dans lrsquoobjectif drsquoouvrir les doctorants au monde de lrsquoentreprise et au
deacuteveloppement de projets innovants Ce Master a eacuteteacute lrsquooccasion pour moi drsquoacqueacuterir des compeacutetences
compleacutementaires dans le domaine de lrsquoadministration et de la gestion des entreprises notamment
dans la gestion de projets Mon investissement personnel important en parallegravele de mon travail de
recherche mrsquoa permis drsquoobtenir le Master avec la mention Bien
Dans le cadre de la thegravese JCE lrsquoentreprise LSI (speacutecialiseacutee dans le diagnostic veacuteteacuterinaire) a
accompagneacute le deacutemarrage de la thegravese et mrsquoa permis drsquoeacutechanger avec des industriels autour du
deacuteveloppement de systegravemes de deacutetection baseacutes sur la qPCR (IS1561rsquo et autres systegravemes) Suite au
rachat de lrsquoentreprise par Life Science puis Thermo Fisher le transfert de technologie nrsquoa pas eacuteteacute
poursuivi Cependant ces systegravemes de qPCR sont utiliseacutes en routine par le Laboratoire National de
Reacutefeacuterence pour la tuberculose bovine encadreacute par Maria Laura Boschiroli pour caracteacuteriser les souches
de mycobacteacuteries issues drsquoeacutechantillons animaux Drsquoautre part des travaux et reacutesultats obtenus lors de
ma thegravese vont faire lrsquoobjet drsquoune valorisation sous forme drsquoun brevet en cours de deacutepocirct pour la
deacutetection des mycobacteacuteries et drsquoun savoir-faire concernant lrsquoextraction des mycobacteacuteries du sol par
des meacutethodes chimiques en vue de leur mise en culture Jrsquoai donc au cours de mes travaux de recherche
toujours conserveacute lrsquoesprit drsquoinnovation et de transfert potentiel de technologie
CHAPITRE 1
Contexte bibliographique
Chapitre 1
4
CHAPITRE 1 - Contexte bibliographique
A LES MYCOBACTERIES DIVERSITE PHYLOGENIE SPECTRE DrsquoHOcircTES ET VIRULENCE
1 Le genre Mycobacterium taxonomie et meacutethodes de classification
Les connaissances sur les mycobacteacuteries commencent en 1882 date agrave laquelle Robert Koch
deacutecouvre le bacille responsable de la tuberculose humaine alors baptiseacute Bacterium tuberculosis et
partage son postulat sur lrsquoeacutetiologie infectieuse de la maladie La coloration de Ziehl et Neelsen
deacuteveloppeacutee en 1883 pour la recherche des bacilles acido-alcoolo-reacutesistants permet agrave Smith en 1892 de
mettre en eacutevidence un bacille tregraves proche de Bacterium tuberculosis dans des leacutesions tuberculeuses de
bovins Mycobacterium bovis avec toutefois des diffeacuterences pheacutenotypiques et pathogeacuteniques qui
permettent de distinguer les deux bacilles Le bacille de la tuberculose aviaire M avium ssp avium est
isoleacute en 1894 sur des oiseaux puis M avium ssp paratuberculosis en 1895 sur des leacutesions digestives
de bovins La deacutecouverte des premiegraveres mycobacteacuteries non tuberculeuses a suivi celle de Bacterium
tuberculosis notamment gracircce agrave la coloration Mycobacterium smegmatis a eacuteteacute isoleacutee en 1889 dans
des preacutelegravevements biologiques de patients atteints de syphilis puis dans du beurre par lrsquoeacutequipe de Koch
dans les anneacutees 1896 Les premiegraveres recherches meneacutees dans le sol les plantes puis lrsquoeau ont montreacute
que ces substrats pouvaient ecirctre des sources de bacteacuteries morphologiquement tregraves proches du bacille
de Koch Le genre Mycobacterium (mứkecircs champignon et baktecircrίa bacircton) est proposeacute en 1896 par
Lehmann et Neumann Dans les anneacutees 50 environ 20 espegraveces de mycobacteacuteries eacutetaient deacutecrites Le
deacuteveloppement des meacutethodes de culture puis des outils moleacuteculaires dans les anneacutees 1990 ont permis
drsquoisoler de nouvelles espegraveces dans des niches environnementales varieacutees ainsi que chez lrsquohomme et
les animaux A lrsquoheure actuelle 172 espegraveces de mycobacteacuteries sont identifieacutees (Site bacterionetorg)
La diversiteacute pheacutenotypique physiologique eacutecologique et pathogeacutenique des espegraveces placeacutees au
fur et agrave mesure de leur deacutecouverte dans ce genre a meneacute agrave lrsquoeacutelaboration de plusieurs classifications
Au sein du genre Mycobacterium les espegraveces sont geacuteneacuteralement classeacutees en deux grands groupes un
groupe qui rassemble les mycobacteacuteries pathogegravenes obligatoires que sont les mycobacteacuteries
tuberculeuses et les mycobacteacuteries responsables de la legravepre et un second groupe qui rassemble les
mycobacteacuteries pathogegravenes opportunistes et saprophytes appeleacute mycobacteacuteries atypiques ou
mycobacteacuteries environnementales ou mycobacteacuteries non tuberculeuses (MNT)
Dans ces groupes des complexes drsquoespegraveces ont eacuteteacute deacutefinis sur des bases drsquoidentiteacute geacuteneacutetique Le
Complexe tuberculosis (MTBC) rassemble les mycobacteacuteries tuberculeuses notamment
Chapitre 1
5
M tuberculosis M africanum M canetti M bovis M caprae M microti M pinnipedii Parmi les
nombreuses MNT diffeacuterents complexes sont observeacutes dont le plus important est le Complexe
Mycobacterium avium (MAC) qui regroupe quatre sous-espegraveces distinctes dont trois drsquointeacuterecirct en santeacute
humaine et animale (M avium ssp avium M avium ssp paratuberculosis et M avium ssp hominisuis)
(Radomski et al 2010) ainsi qursquoune dizaine drsquoespegraveces dont M intracellulare
Plus reacutecemment la phylogeacutenie moleacuteculaire de ce genre baseacutee notamment sur lrsquoanalyse des gegravenes de
lrsquoARNr 16S et 23S ainsi que des reacutearrangements de gegravenes a permis drsquoaffiner la position taxonomique
du genre Mycobacterium Drsquoapregraves le Bergeyrsquos manual (Ludwig et al 2008) le genre Mycobacterium
appartient
au Phylum des Actinobacteria (bacteacuteries Gram positif agrave geacutenome riche en acides nucleacuteiques
G et C ()
o agrave la Classe des Actinobacteria
o agrave lrsquoOrdre des Corynebacteriales qui regroupe diffeacuterentes familles de bacteacuteries
caracteacuteriseacutees par la preacutesence drsquoacides mycoliques dans leur paroi et des
proprieacuteteacutes tinctoriales particuliegraveres lieacutees agrave la reacutesistance de leur paroi agrave la
deacutecoloration par un meacutelange alcool-acide (ou BAAR)
agrave la Famille des Mycobacteriaceae qui ne comprend que le genre
Mycobacterium caracteacuteriseacute par une enveloppe riche en acides
mycoliques contenant un grand nombre drsquoatomes de carbone ainsi
que des quinones particuliegraveres
2 Caracteacuteristiques geacuteneacuterales des mycobacteacuteries
a Morphologie des mycobacteacuteries
Les mycobacteacuteries se preacutesentent sous forme de bacirctonnets droits ou leacutegegraverement incurveacutes (02-
06 x 10-10microm) non mobiles et ne sporulant pas Les colonies observeacutees sont de couleur blanche agrave
cregraveme voire jaune agrave orangeacutee pour les mycobacteacuteries productrices de pigments (Magee amp Ward 2012)
Elles fixent difficilement la coloration de Gram et bien qursquoelles soient apparenteacutees aux bacteacuteries Gram
+ leur qualification exacte est laquo Bacilles Acido-Alcoolo-Reacutesistants (BAAR) raquo car lors de la coloration
leur enveloppe particuliegravere reacutesiste agrave la deacutecoloration agrave lrsquoalcool et agrave lrsquoacide
Les mycobacteacuteries possegravedent une enveloppe proche de celle des bacteacuteries Gram + mais dont
lrsquoorganisation la richesse et la diversiteacute des acides gras qui la composent sont uniques et
caracteacuteristiques La mycomembrane est constitueacutee drsquoacides gras agrave longue chaine contenant de 60 agrave 90
atomes de carbone les acides mycoliques qui repreacutesentent 30 agrave 40 de sa masse (Brennan amp Nikaido
Chapitre 1
6
1995) Dans sa couche interne les acides mycoliques sont disposeacutes parrallegravelement les uns aux autres
et sont lieacutes au peptidoglycane de faccedilon covalente par lrsquointermeacutediaire de la couche drsquoarabinogalactane
(polymegravere formeacute drsquoarabinose et de galactose) Sa couche externe contient des lipides extractibles (ie
non fixeacutes de faccedilon covalente au peptidoglycane) tels que le treacutehalose monomycolate ou dimycolate
des phospholipides et des glycolipides (Zuber et al 2008) La mycomembrane est recouverte de
glucanes de proteacuteines et drsquoune faible proportion de lipides
Cette enveloppe particuliegravere tregraves riche en lipides confegravere agrave la bacteacuterie ses proprieacuteteacutes
tinctoriales (BAAR) sa grande reacutesistance aux stress chimiques notamment aux antibiotiques et
deacutetergents aux stress physiques (tempeacuterature) ainsi que biologiques (reacutesistance dans les
macrophages de lrsquohocircte et dans les amibes) Crsquoest eacutegalement un facteur important de virulence chez les
mycobacteacuteries pathogegravenes certains composants lipidiques possegravedent en effet une activiteacute
immunogegravene exceptionnelle (Coulombe et al 2009) Ils sont capables de moduler lrsquoimmuniteacute de lrsquohocircte
voire de deacutesorganiser ses meacutecanismes immunitaires de deacutefense et interviennent dans la pathogeacutenie
comme cela a eacuteteacute deacutemontreacute pour M tuberculosis et M avium ssp paratuberculosis (Libero amp Mori
2014 Momotani et al 2012) La paroi est drsquoailleurs une des cibles des antimicrobiens utiliseacutes contre la
tuberculose humaine ou la legravepre Ces derniers agissent notamment en inhibant la biosynthegravese des
moleacutecules qui la constituent Lrsquoutilisation des antigegravenes lipidiques de la paroi constitue eacutegalement un
axe de recherche pour le deacuteveloppement de vaccins (Libero amp Mori 2014)
Dans les matrices environnementales la paroi hydrophobe des mycobacteacuteries leur permet
drsquoadheacuterer fortement aux particules en suspension dans lrsquoeau aux seacutediments aux surfaces favorisant
la formation de biofilms et la survie des bacteacuteries (Primm et al 2004)
Les acides mycoliques de par leur complexiteacute et leur variabiliteacute structurelle sont eacutegalement utiliseacutes
comme marqueurs taxonomiques car ils permettent drsquoidentifier avec speacutecificiteacute le genre et les espegraveces
Chapitre 1
7
Figure 1 Structure et composition de lenveloppe des mycobacteacuteries (drsquoapregraves Marrakchi Laneacuteelle and Daffeacute 2014) AG arabinogalactane PG peptidoglycane TDM treacutehalose dimycolate PL phospholipides TMM treacutehalose monomycolate GL glycolipides
b Caracteacuteristiques culturales pheacutenotypiques et physiologiques
La plupart des mycobacteacuteries sont aeacuterobies strictes mais quelques espegraveces tolegraverent de faibles
pressions drsquooxygegravene notamment M bovis et M tuberculosis Les substrats utiliseacutes comme sources de
carbone et drsquoazote varient consideacuterablement entre les espegraveces et peuvent servir pour leur
identification La majoriteacute des espegraveces croit sur des milieux de culture classiques mais pour certaines
mycobacteacuteries drsquoimportance clinique la culture est ameacutelioreacutee par lrsquoutilisation de milieux agrave lrsquoœuf
(Loumlwenstein-Jensen et Coletsos) apregraves une deacutecontamination chimique ou de milieu Middlebrook ainsi
que par un enrichissement des milieux (en glyceacuterol pour M tuberculosis en mycobactine pour
M avium subsp paratuberculosis en heacutemine pour M haemophilum) Les mycobacteacuteries responsables
de la legravepre sont cultiveacutees sur tissus vivants (coussinets de souris peau de tatou) et se multiplient tregraves
difficilement sur milieux syntheacutetiques
Les mycobacteacuteries ont des vitesses de croissance tregraves variables on distingue les mycobacteacuteries
agrave croissance rapide dont les colonies sont visibles en moins de 7 jours des mycobacteacuteries agrave croissance
lente (gt7 jours) Les bacteacuteries du MTBC sont toutes des mycobacteacuteries agrave croissance lente avec des
temps de geacuteneacuteration eacuteleveacutes de 14 agrave 15h pour M tuberculosis et environ 20h pour M bovis La
tempeacuterature optimale de croissance des bacteacuteries du MTBC est de 37degC Les MNT regroupent des
mycobacteacuteries agrave croissance rapide et agrave croissance lente avec des taux de geacuteneacuteration extrecircment
variables allant de 2h pour les bacteacuteries agrave croissance rapide comme M smegmatis agrave 21h pour celles
agrave croissance lente comme M aurum voire 7 jours pour M avium ssp paratuberculosis dans des
conditions de laboratoire pourtant optimales Les MNT peuvent se multiplier dans un large eacuteventail de
Chapitre 1
8
tempeacuterature allant de 4degC pour M psychrotolerans (Trujillo et al 2004) agrave plus de 45degC voire agrave 52degC
pour M thermotolerans et M phlei (Stahl amp Urbance 1990) avec une tempeacuterature optimale de
croissance entre 28 et 31degC
c Phylogeacutenie du genre Mycobacterium
La phylogeacutenie baseacutee sur le polymorphisme de lrsquoADN codant pour lrsquoARNr 16S permet de
discriminer les espegraveces de mycobacteacuteries et met en eacutevidence trois grands groupes en fonction de leur
vitesse de croissance Le cladogramme preacutesenteacute dans la Figure 2 regroupe distinctement les
mycobacteacuteries agrave croissance lente (haut du cladogramme) les mycobacteacuteries agrave croissance intermeacutediaire
(au milieu) et les mycobacteacuteries agrave croissance rapide (bas du cladogramme) On peut noter de plus que
les mycobacteacuteries du MTBC et le complexe Mac forment deux groupes phylogeacuteneacutetiquement
homogegravenes au sein du groupe des mycobacteacuteries agrave croissance lente
Myc
ob
acteacute
ries
agrave c
rois
san
ce le
nte
Myc
ob
acteacute
ries
agrave
cro
issa
nce
inte
rmeacute
dia
ire
Chapitre 1
9
Figure 2 Arbre phylogeacutenique baseacute sur lrsquoanalyse de lrsquoARNr 16S des principaux complexes et espegraveces de mycobacteacuteries
(drsquoapregraves Magee et Ward 2012)
d Caracteacuteristiques geacuteneacutetiques des mycobacteacuteries
De nombreux geacutenomes complets (annoteacutes ou en cours drsquoanalyse) de mycobacteacuteries sont
actuellement disponibles (NCBI Genome Resources European Nucleotide archive) Mycobacterium
leprae est la mycobacteacuterie qui possegravede le geacutenome le plus petit avec 33 millions de paires de bases
(Mpb) codant pour 1 605 gegravenes et le G+C (pourcentage Guanine+Cytosine) le plus faible (57) Les
mycobacteacuteries environnementales possegravedent les geacutenomes les plus importants de 483 Mpb (codant
4 350 gegravenes) pour M avium k10 agrave 69 Mpb (codant 6 716 gegravenes) pour M smegmatis Les mycobacteacuteries
tuberculeuses M tuberculosis M africanum et M bovis ont un geacutenome de taille intermeacutediaire (de 43
Mpb agrave 45 Mpb) codant pour 3 850 agrave 4 500 gegravenes Les geacutenomes des mycobacteacuteries preacutesentent des
capaciteacutes codantes importantes de lrsquoordre de 90 pour M tuberculosis M bovis M avium ssp
paratuberculosis et M smegmatis (Garnier et al 2003 Cole et al 1998 Li et al 2005)
La comparaison du geacutenome complet de M tuberculosis M bovis M leprae et M avium ssp
paratuberculosis a montreacute une forte conservation des gegravenes impliqueacutes dans le meacutetabolisme
Myc
ob
acteacute
ries
agrave c
rois
san
ce r
apid
e
Chapitre 1
10
eacutenergeacutetique la biosynthegravese des acides amineacutes et des cofacteurs et le meacutetabolisme des nucleacuteotides et
des proteacuteines (Marri et al 2006) Des plasmides ont eacuteteacute mis en eacutevidence chez quelques espegraveces de
mycobacteacuteries environnementales (Rauzier et al 1988 Stinear et al 2004) notamment le plasmide
pMUM001 isoleacute chez M ulcerans responsable de lrsquoulcegravere de Buruli chez lrsquohomme qui code pour un
macrolide aux proprieacuteteacutes cytotoxiques et immunosuppressives la mycolactone (Stinear et al 2004)
Le nombre drsquoopeacuterons ribosomaux 16S dans le geacutenome des mycobacteacuteries a eacuteteacute suggeacutereacute comme
lrsquoun des facteurs expliquant la diffeacuterence des vitesses de croissance observeacutees La croissance lente des
mycobacteacuteries serait lieacutee agrave la preacutesence drsquoun seul opeacuteron ribosomal les espegraveces agrave croissance rapide en
posseacutedant geacuteneacuteralement deux (rrnA et rrnB) (Bercovier et al 1986) Toutefois des exceptions existent
et drsquoautres facteurs tels que la lenteur des eacutechanges agrave travers leur paroi impermeacuteable riche en lipides
et le coucirct eacutenergeacutetique eacuteleveacute de la synthegravese de leurs acides gras agrave longues chaines seraient eacutegalement
impliqueacutes (Primm et al 2004)
Dans le Complexe tuberculosis les diffeacuterentes espegraveces constituent un modegravele drsquoidentiteacute
geacuteneacutetique interspeacutecifique Leurs geacutenomes preacutesentent une identiteacute supeacuterieure agrave 999 au niveau
nucleacuteotidique et des seacutequences drsquoARNr 16S et drsquoITS identiques (Boumlddinghaus et al 1990 Sreevatsan amp
Pan 1997 Huard et al 2006 Cole et al 1998 Garnier et al 2003 Kasai et al 2000) Lrsquoanalyse des
seacutequences de 56 gegravenes de structure de plusieurs centaines drsquoisolats drsquoorigines geacuteographique et
phylogeacuteneacutetique varieacutees suggegravere que le polymorphisme alleacutelique est exceptionnel dans ce Complexe
(Sreevatsan amp Pan 1997 Musser et al 2000) La recombinaison de tregraves petits fragments drsquoADN est
deacutemontreacutee chez M tuberculosis (Namouchi et al 2012) mais le transfert lateacuteral de gegravenes ne semble
pas ecirctre un facteur important dans lrsquoeacutevolution des bacteacuteries du MTBC qui ont une structure de
population fortement clonale (Pepperell et al 2013) contrairement agrave la plupart des autres bacteacuteries
pathogegravenes Malgreacute leur forte identiteacute geacuteneacutetique les espegraveces du Complexe se diffeacuterencient par des
reacutegions polymorphiques ou LSPs (Large Sequence Polymorphisms) lieacutees agrave des deacuteleacutetions et agrave des
reacutearrangements de seacutequences (Brosch et al 2002 Huard et al 2003) Parmi ces LSPs 14 Reacutegions de
Diffeacuterence ou RD de 2 agrave 127 kpb sont recenseacutees dans le MTBC Ces deacuteleacutetions sont consideacutereacutees comme
une adaptation eacutevolutive agrave lrsquoinfection de nouveaux hocirctes plutocirct qursquoagrave une deacuterive geacuteneacutetique (Brodin et
al 2002 Aranaz et al 2003) Cette variabiliteacute geacutenomique permet agrave la fois de diffeacuterencier les espegraveces
entre elles (Huard et al 2003 Huard et al 2006) et de retracer leur eacutevolution phylogeacuteneacutetique
(Rodriguez-Campos et al 2014)
3 Description des diffeacuterents Complexes ou espegraveces de mycobacteacuteries
Alors que les mycobacteacuteries responsables de la tuberculose et de la legravepre sont des pathogegravenes
obligatoires les MNT rassemblent des mycobacteacuteries saprophytes sans pouvoir pathogegravene jusque-lagrave
Chapitre 1
11
aveacutereacute et des mycobacteacuteries pathogegravenes opportunistes responsables de mycobacteacuterioses humaines ou
animales Depuis la fin du 19egraveme siegravecle jusqursquoaux anneacutees 1950 la tuberculose humaine repreacutesentait un
tel problegraveme de santeacute publique que toutes les eacutetudes eacutetaient centreacutees sur elle et son agent
M tuberculosis Les autres mycobacteacuteries isoleacutees dans lrsquoenvironnement ou dans des preacutelegravevements
biologiques humains et animaux eacutetaient consideacutereacutees comme des bacteacuteries saprophytes sans inteacuterecirct
clinique particulier A partir des anneacutees 1950 la communauteacute scientifique a admis la responsabiliteacute
indiscutable de certaines mycobacteacuteries environnementales dans des processus pathologiques
humains et animaux appeleacutes mycobacteacuterioses amorccedilant des eacutetudes les concernant En lrsquoabsence de
contamination inter-humaine deacutemontreacutee il a eacuteteacute admis que lrsquoorigine de ces mycobacteacuteries est
environnementale et que lrsquohomme se contamine agrave partir de lrsquoenvironnement
a Les mycobacteacuteries non pathogegravenes potentiellement opportunistes
Elles regroupent les mycobacteacuteries environnementales qui preacutesentent un pouvoir pathogegravene
pour lrsquohomme et les animaux tregraves variable en fonction des espegraveces (van Ingen et al 2009 Tortoli 2003)
Lrsquoincidence des infections humaines lieacutees agrave des mycobacteacuteries opportunistes semble augmenter dans
les pays industrialiseacutes (Falkinham 2016) peut-ecirctre en lien avec lrsquoameacutelioration des outils pour les isoler
et les identifier Les mycobacteacuterioses qursquoelles engendrent surviennent geacuteneacuteralement sur un terrain
drsquoimmunodeacutepression locale ou geacuteneacuterale Chez lrsquohomme elles sont responsables drsquoun large eacuteventail de
symptocircmes cliniques majoritairement des infections pulmonaires mais eacutegalement des lymphadeacutenites
chez les enfants des infections cutaneacutees (notamment lieacutees agrave M marinum responsable du granulome
des piscines et M ulcerans responsable de lrsquoulcegravere de Buruli) des infections extra-pulmonaires voire
des infections disseacutemineacutees chez les patients seacutevegraverement immunodeacuteprimeacutes (van Ingen 2013 Biet et al
2005 Hoefsloot et al 2013) Environ 25 espegraveces sont retrouveacutees reacuteguliegraverement dans les eacutechantillons
cliniques les autres beaucoup plus rarement (van Ingen 2013) Parmi elles les mycobacteacuteries agrave
croissance lente semblent plus preacutedisposeacutees agrave provoquer des atteintes pulmonaires ou lymphatiques
alors que celles agrave croissance rapide affectent plutocirct la peau les os et les articulations (Tortoli 2003
2009) Les mycobacteacuteries M avium et M intracellulare sont les plus freacutequemment isoleacutees chez
lrsquohomme majoritairement en cas drsquoatteinte pulmonaire (Hoefsloot et al 2013 Whiley et al 2012) La
distribution des espegraveces pathogegravenes isoleacutees chez lrsquohomme diffegravere selon la zone geacuteographique eacutetudieacutee
probablement en lien avec leurs niches eacutecologiques (Hoefsloot et al 2013)
Chez les animaux M avium ssp paratuberculosis (Map) est lrsquoagent de la maladie de Johne ou
paratuberculose chez les ruminants domestiques et sauvages les primates ainsi que drsquoautres espegraveces
(lapins etc) chez qui elle provoque une enteacuterite chronique contagieuse et incurable agrave lrsquoorigine de
pertes eacuteconomiques importantes dans les cheptels infecteacutes Son rocircle est eacutegalement eacutevoqueacute dans la
Chapitre 1
12
maladie de Crohn chez lrsquohomme (Biet et al 2005 Rowe amp Grant 2006 Momotani et al 2012) sans
toutefois que la causaliteacute ait eacuteteacute deacutemontreacutee Mycobacterium avium ssp avium (Maa) est responsable
de la tuberculose aviaire chez les oiseaux et les petits mammifegraveres terrestres chez qui elle provoque
des leacutesions tuberculeuses majoritairement digestives des lymphadeacutenites et des infections disseacutemineacutees
(Biet et al 2005 Pavlik et al 2000) Mycobacterium avium ssp hominissuis (Mah) est responsable de
leacutesions des nœuds lymphatiques et de diffeacuterents organes chez le porc (Pavlik et al 2000)
b Les mycobacteacuteries saprophytes non pathogegravenes
Parmi toutes les mycobacteacuteries environnementales deacutecrites agrave lrsquoheure actuelle certaines sont
retrouveacutees uniquement dans lrsquoenvironnement et nrsquoont jamais eacuteteacute isoleacutees chez lrsquohomme ou lrsquoanimal
comme M arabiense ou M rufum (Tortoli 2014) Ces mycobacteacuteries sont dites saprophytes Parmi
elles certaines ont eacuteteacute isoleacutees dans des sols et des seacutediments fortement pollueacutes par les HAP
(Hydrocarbures Aromatiques Polycycliques) qursquoelles sont potentiellement capables de deacutegrader
(Hennessee et al 2009 Leys et al 2005 Hormisch et al 2004) Elles repreacutesentent donc une voie
inteacuteressante pour la bioremeacutediation de ces sols
c Les mycobacteacuteries pathogegravenes obligatoires
Les mycobacteacuteries responsables de legravepre
Les mycobacteacuteries responsables de la legravepre humaine et murine sont M lepreae M lepromatosis et
M lepreaemurium Elles provoquent des infections dermatologiques et nerveuses chroniques graves
et mutilantes parfois mortelles Mycobacterium leprae touche environ 15 million de personnes dans
le monde et la maladie reste un problegraveme de santeacute publique preacuteoccupant dans les zones endeacutemiques
drsquoAfrique drsquoAsie et drsquoAmeacuterique du sud (Who 2014) Mycobacterium leprae a longtemps eacuteteacute consideacutereacutee
comme un pathogegravene obligatoire de lrsquohomme avec une voie de transmission interhumaine majoritaire
La deacutecouverte de reacuteservoirs animaux (Truman 2005 Meredith et al 2014) et la deacutetection de M leprae
dans le sol la sphaigne ou lrsquoeau de riviegravere (Matsuoka et al 1999 Mostafa et al 1995 Lavania et al
2006) tendent agrave souligner le rocircle potentiel de certains animaux ou de lrsquoenvironnement comme source
de lrsquoinfection chez lrsquohomme Ces 3 espegraveces sont multiplieacutees sur les coussinets de souris (Shepard 1960)
et sur la peau de tatous agrave 9 bandes (Dasypus novemcinctus) (Kirchheimer amp Storrs 1971) car aucun
milieu syntheacutetique ne permet leur croissance optimale
Chapitre 1
13
Les mycobacteacuteries tuberculeuses le Complexe tuberculosis
(MTBC)
Toutes les mycobacteacuteries tuberculeuses sont des pathogegravenes obligatoires Contrairement aux
mycobacteacuteries environnementales leur croissance dans lrsquoenvironnement nrsquoa jamais eacuteteacute deacutemontreacutee Le
complexe tuberculosis comprend agrave ce jour 13 espegraveces qui malgreacute une forte identiteacute geacuteneacutetique
preacutesentent des pheacutenotypes des hocirctes et des pouvoirs pathogegravenes varieacutes
Phylogeacutenie des mycobacteacuteries du MTBC
Lrsquoeacutetude des RD eacutevoqueacutees preacuteceacutedemment et des mutations agrave lrsquoeacutechelle du nucleacuteotide (ou SNP Single
Nucleacuteotide Polymorphism) sur 7 gegravenes (katG gyrA gyrB rpoB mmpL6 oxyR et ADNr 16S) ont permis
de retracer la phylogeacutenie des bacteacuteries du Complexe agrave partir de leur ancecirctre commun (Brosch et al
2002 Rodriguez-Campos et al 2014 Gordon et al 1999 Mahairas et al 1996 Huard et al 2003) les
deacuteleacutetions et mutations jouant un rocircle drsquohorloge moleacuteculaire permettant de replacer lrsquoapparition et
lrsquoeacutevolution drsquoune espegravece dans le temps Le sceacutenario de lrsquoeacutevolution des bacteacuteries du MTBC est
scheacutematiseacute dans la Figure 3 Selon les auteurs lrsquoancecirctre du MTBC appeleacute M protuberculosis aurait
eacutemergeacute il y a entre 15 000 et 40 000 ans en Afrique de lrsquoEst (Gutierrez et al 2005 Wirth et al 2008
Sreevatsan amp Pan 1997) A cocircteacute de M canetti la bacteacuterie du MTBC geacuteneacutetiquement la plus proche de
lrsquoancecirctre deux clades principaux peuvent ecirctre observeacutes Un clade de souches speacutecifiques de lrsquohomme
caracteacuteriseacute par la deacuteleacutetion TbD1 a donneacute les diffeacuterentes ligneacutees modernes de M tuberculosis (Figure
3) Un second clade caracteacuteriseacute par la perte des reacutegions RD7 RD8 RD9 et RD10 regroupe des souches
endeacutemiques drsquoAfrique speacutecifiques de lrsquohomme (M africanum) et drsquoanimaux (le Chimpanzee bacillus
M mungi le Dassie bacillus et M suricattae) ainsi que des souches adapteacutees agrave diffeacuterentes espegraveces de
mammifegraveres (M orygis M microti M pinnipedii M caprae et M bovis)
Chapitre 1
14
Figure 3 Phylogeacutenie des souches du MTBC baseacutee sur les Reacutegions de Diffeacuterences et diffeacuterents Single Nucleotide Polymorphism (Dippenaar et al 2015)
Virulence des mycobacteacuteries du MTBC
Les mycobacteacuteries du MTBC ne possegravedent pas de facteurs de virulence classiques tels que les
toxines par exemple observeacutes chez drsquoautres bacteacuteries pathogegravenes Pour certains auteurs la virulence
des mycobacteacuteries est restreinte agrave lrsquoaptitude du bacille agrave peacuteneacutetrer dans lrsquohocircte agrave y survivre et srsquoy
multiplier (Pollock amp Neill 2002) De faccedilon plus large la notion de virulence chez les membres du MTBC
a eacuteteacute conceptualiseacutee comme lrsquoassociation de i) la capaciteacute de la bacteacuterie agrave faire face agrave la reacuteponse
immunitaire de lrsquohocircte infecteacute ii) la capaciteacute agrave engendrer des leacutesions iii) la capaciteacute agrave survivre en dehors
de son hocircte notamment lors de lrsquoaeacuterosolisation et iiii) la transmission et lrsquoinfection drsquoun nouvel hocircte
(Coscolla amp Gagneux 2014) La virulence implique donc un ensemble de meacutecanismes complexes et
varieacutes faisant intervenir de nombreux gegravenes et proteacuteines permettant notamment au bacille de peacuteneacutetrer
dans les cellules de lrsquohocirctes et drsquoy survivre en bloquant les diffeacuterents meacutecanismes de deacutefense des
macrophages (inhibition de la fusion phagosome-lysosome et de la production de ROS1 et RNS2
deacutesorganisation de lrsquoimmuniteacute de lrsquohocircte inhibition de lrsquoautophagie et de lrsquoapoptose) (Hmama et al
1 ROS Reactive Oxygen Species 2 RNS Reactive Nitrogen Species
Chapitre 1
15
2015) Plus de cent gegravenes seraient impliqueacutes dans la virulence des bacteacuteries du MTBC et lrsquoutilisation
de mutants permet peu agrave peu de connaitre le rocircle de chacun (Forrellad et al 2013) Lrsquoanalyse du
transcriptome (ARN) de M tuberculosis au cours de lrsquoinfection de macrophages et de cellules
dendritiques a reacuteveacuteleacute lrsquoexpression ou la surexpression de nombreux gegravenes codant des proteacuteines
intervenant notamment dans le meacutetabolisme lipidique la synthegravese de lrsquoenveloppe bacteacuterienne
lrsquoapprovisionnement en fer la reacutesistance au stress oxydatif et lrsquoinhibition de lrsquoapoptose (Mukhopadhyay
et al 2012) Les meacutecanismes de survie en dehors de lrsquohocircte sont inconnus agrave ce jour
Il a eacuteteacute montreacute chez M tuberculosis qursquoen fonction des ligneacutees ou des souches la virulence pouvait
varier entrainant une reacuteponse immunitaire reacuteduite ou retardeacutee chez lrsquohocircte une plus grande seacuteveacuteriteacute
de la maladie et une transmission augmenteacutee (Coscolla amp Gagneux 2014) Ce lien souche-virulence est
eacutegalement suspecteacute chez M bovis pour lequel certains geacutenotypes srsquoaccompagnent drsquoun plus grand
nombre drsquoanimaux agrave leacutesions et ou drsquoun score leacutesionnel plus eacuteleveacute (Wright et al 2013 Gavier-Wideacuten et
al 2009 Garbaccio et al 2014) mais des eacutetudes compleacutementaires restent neacutecessaires
Les mycobacteacuteries speacutecifiques de lrsquohomme
Mycobacterium tuberculosis
Mycobacterium tuberculosis est lrsquoagent de la tuberculose humaine maladie retrouveacutee dans
toutes les reacutegions du monde Crsquoest un problegraveme majeur de santeacute publique puisque 2 milliards
drsquoindividus seraient infecteacutes dans le monde En 2013 lrsquoOMS a eacutevalueacute agrave neuf millions le nombre de
nouveaux cas annuels et agrave un million et demi le nombre de deacutecegraves lieacutes agrave la tuberculose (dont 24 de
personnes seacuteropositives) Les reacutegions de lrsquoAsie du sud-est et du Pacifique occidental regroupent 56
des nouveaux cas lrsquoAfrique 25 lrsquoInde 24 et la Chine 11 (WHO 2014) Lrsquoinfection par le HIV est un
facteur preacutedisposant majeur dans la transmission de la maladie et la progression de lrsquoinfection chez
lrsquohomme de par les effets synergiques deacuteleacutetegraveres des 2 micro-organismes sur le systegraveme immunitaire
(Pawlowski et al 2012) Environ 78 des personnes co-infecteacutees vivent en Afrique (WHO 2014) La
tuberculose se manifeste chez lrsquohomme par des granulomes pulmonaires (tuberculose pulmonaire)
mais peut se deacutevelopper dans drsquoautres organes (tuberculose extra-pulmonaire) Elle se transmet
essentiellement par voie respiratoire par lrsquoinhalation de gouttelettes contamineacutees eacutemises par une
personne infecteacutee lors de toux par exemple Le traitement repose sur lrsquoassociation de plusieurs
antibiotiques adapteacutes tels que lrsquoisoniazide la rifampicine le pyrazinamide et lrsquoeacutethambutol
Lrsquoapparition de souches reacutesistantes aux antibiotiques MDR (Multi Drug Resistant) et XDR
(eXtensively Drug Resistant) constitue agrave lrsquoheure actuelle un problegraveme majeur dans la lutte contre la
maladie Bien que lrsquohomme constitue le reacuteservoir majeur de la bacteacuterie M tuberculosis eacuteteacute isoleacutee chez
Chapitre 1
16
un grand nombre drsquoespegraveces animales telles que les bovins domestiques (Romero et al 2011 Prasad et
al 2005 Ameni et al 2011 Pavlik et al 2005 Cadmus et al 2006) les chegravevres (Cadmus Adesokan et
al 2009) les porcs domestiques (Jenkins et al 2011) les chiens (Pavlik et al 2005) les oiseaux et des
animaux de zoos (Michel et al 2003) dont les eacuteleacutephants
Mycobacterium africanum
Elle est principalement retrouveacutee dans les pays drsquoAfrique de lrsquoouest ougrave elle peut ecirctre impliqueacutee
dans la moitieacute des cas de tuberculose humaine agrave cocircteacute de M tuberculosis (de Jong et al 2010) Isoleacutee
et deacutecrite pour la premiegravere fois au Seacuteneacutegal en 1968 elle preacutesente des caracteacuteristiques biochimiques
variables intermeacutediaires entre celles de M tuberculosis et M bovis (Castets amp Sarrat 1968)
Mycobacterium africanum est parfois isoleacutee chez lrsquohomme en dehors de lrsquoAfrique (Schroumlder 1982
Grange amp Yates 1989 Desmond et al 2004 Frottier et al 1990) La proximiteacute phylogeacuteneacutetique de
M africanum avec les autres membres du MTBC infectant des espegraveces animales a souleveacute la question
drsquoun reacuteservoir animal de la bacteacuterie Mais elle a eacuteteacute isoleacutee de faccedilon sporadique sur des singes (Thorel
1980) des bovins (Rahim et al 2007 Cadmus et al 2006) et du lait (Cadmus et al 2010) infirmant
cette hypothegravese
Mycobacterium canetti et smooth tubercle bacilli
Elle est responsable de tuberculose humaine dans une zone geacuteographique confineacutee agrave la corne de
lrsquoAfrique (Djibouti Somalie Kenya) (Pfyffer et al 1998 Miltgen et al 2002) Elle est la plus proche
geacuteneacutetiquement de lrsquoancecirctre des bacteacuteries du MTBC Elle constitue un taxon divergent des autres
membres du MTBC par son pheacutenotype particulier et son geacutenotype qui preacutesente une large diversiteacute
geacuteneacutetique et des signes de transfert horizontal et de recombinaison de gegravenes contrairement aux autres
bacteacuteries du MTBC (Fabre et al 2004 Supply et al 2013 Gutierrez et al 2005) Sa situation
geacuteographique confineacutee la grande diversiteacute geacuteneacutetique des souches rencontreacutees et le manque de
connaissances sur la transmission de la bacteacuterie ont suggeacutereacute lrsquoexistence drsquoun reacuteservoir environnemental
(Fabre et al 2004 Koeck et al 2011) A ce jour cette hypothegravese nrsquoa pas eacuteteacute confirmeacutee La tuberculose
agrave M canetti est consideacutereacutee aujourdrsquohui comme une maladie eacutemergente
Les mycobacteacuteries speacutecifiques des animaux
Mycobacterium bovis
Mycobacterium bovis est lrsquoagent de la tuberculose bovine maladie preacutesente sur tous les
continents Les bovins constituent le reacuteservoir naturel du bacille mais un grand nombre drsquoespegraveces
Chapitre 1
17
animales domestiques et sauvages peuvent ecirctre infecteacutees ainsi que lrsquohomme (Biet et al 2005 Palmer
2013 Grange amp Yates 1994 OrsquoReilly amp Daborn 1995) Sur le plan microbiologique M bovis se
caracteacuterise par une croissance tregraves lente sur milieu de Loumlwenstein-Jensen (6 agrave 10 semaines) par des
colonies petite taille (1 agrave 2 mm) plates lisses laquo dysgoniques raquo et achromogegravenes Sa croissance est
ameacutelioreacutee par lrsquoajout de pyruvate et inhibeacutee par lrsquoajout de glyceacuterol Sur Middlebrook 7H11 les colonies
sont rugueuses et agrave bords eacutechancreacutes (Figure 4) Le pH optimum de culture de M bovis est compris
entre 58 et 69 mais la bacteacuterie survit 20 jours dans du yaourt agrave 4degC (Mitscherlich amp Marth 2012)
Lrsquoidentification pheacutenotypique repose notamment sur lrsquoabsence de production de niacine lrsquoabsence de
nitrate reacuteductase une reacutesistance au pyrazinamide agrave la D-cycloseacuterine (30mgmL-1) et une croissance
favoriseacutee par la preacutesence de pyruvate (Delaune et al 2012)
Figure 4 Aspect des colonies de M bovis sur milieu de Lowenstein-Jensen (agrave gauche) et Middlebrook 7H11 suppleacutementeacute (agrave droite)
Concernant les caracteacuteristiques geacutenomiques de M bovis le seacutequenccedilage complet de la
premiegravere souche (M bovis AF212297 ou ATCC BAA-935 isoleacutee drsquoune leacutesion caseacuteeuse de bovin en
Grande-Bretagne) en 2003 et son annotation ont enrichi les connaissances sur ce bacille Son geacutenome
comprend 4345 Mpb un pourcentage G-C eacuteleveacute de 656 et 3952 gegravenes codant des proteacuteines dont 1
prophage et 42 seacutequences drsquoinsertion Il preacutesente 11 deacuteleacutetions de 1 agrave 127 kb par comparaison avec le
geacutenome de M tuberculosis H37Rv et 1 locus absent chez M tuberculosis appeleacute TbD1 (Garnier et al
2003) Ne posseacutedant pas de gegravenes propres agrave lrsquoespegravece bovis le geacutenome de M bovis a eacuteteacute faccedilonneacute par
lrsquoapparition de ces deacuteleacutetions conditionnant lrsquoexpression des gegravenes Depuis 2003 une quarantaine de
souches isoleacutees sur des animaux domestiques et sauvages ont eacuteteacute entiegraverement seacutequenceacutees (Sites
NCBI et European Nucleotide archive)
La structure de la population de M bovis reacutesulte de lrsquoexpansion de plusieurs complexes clonaux
issus drsquoun mecircme ancecirctre Quatre complexes clonaux majeurs ont eacuteteacute identifieacutes dans le monde pour
Chapitre 1
18
M bovis en se basant sur la preacutesence de deacuteleacutetions chromosomiques caracteacuteristiques dans le geacutenome
de la bacteacuterie etou sur des SNPs ainsi que sur le spoligotype et lrsquoabsence de certains spacers (Smith
2012) le Complexe African 1 est retrouveacute dans plusieurs pays africains (Mali Cameroun Nigeacuteria
Tchad) (Muumlller et al 2009) le Complexe African 2 est essentiellement retrouveacute chez les bovideacutes
drsquoAfrique de lrsquoEst (Berg et al 2011) le Complexe European 1 est retrouveacute en Europe il est dominant au
Royaume-Uni et beaucoup plus rare en Espagne au Portugal et en France On le retrouve eacutegalement
chez les anciens partenaires commerciaux du Royaume-Uni (USA Afrique du Sud Nouvelle Zeacutelande
Australie Canada) et en Ameacuterique du sud La diffusion geacuteographique de ce Complexe est lieacutee aux
eacutechanges commerciaux de bovins (Smith et al 2011) Enfin le Complexe European 2 est majoritaire au
Portugal et en Espagne et assez freacutequent en France et en Italie (Rodriguez-Campos et al 2012) Drsquoautres
complexes clonaux minoritaires existent En France plusieurs complexes clonaux coexistent certains
autochtones eacutevoluent depuis longtemps sur le territoire alors que drsquoautres ont eacuteteacute importeacutes agrave la faveur
des eacutechanges commerciaux de bovins (Hauer et al 2015)
Une souche atteacutenueacutee obtenue agrave partir drsquoun isolat virulent de M bovis a eacuteteacute creacuteeacutee par Calmette
et Gueacuterin apregraves 230 repiquages sur un milieu bilieacute enrichi en glyceacuterol entre 1908 et 1921 le BCG pour
Bacille de Calmette et Gueacuterin Testeacutee sur des modegraveles animaux cette souche atteacutenueacutee immunisait
contre une infection expeacuterimentale agrave M bovis et M tuberculosis trente jours apregraves injection Utiliseacutee
pour la vaccination elle a eacuteteacute distribueacutee dans le monde entier et maintes fois repiqueacutee ce qui a meneacute
agrave une eacutevolution de la souche initiale en plusieurs souches tregraves heacuteteacuterogegravenes La comparaison geacutenomique
de la souche BCG avec M tuberculosis reacutevegravele la perte de 14 reacutegions (RD1 agrave RD14) chez BCG Lrsquoabsence
de la reacutegion RD1 expliquerait en partie lrsquoatteacutenuation du BCG (Pym et al 2002) toutefois comme la
reacuteintroduction de RD1 dans une souche mutante ne suffit pas agrave restaurer totalement la virulence
drsquoautres reacutegions sont impliqueacutees dans lrsquoatteacutenuation (Brosch et al 2007)
Mycobacterium pinnipedii
Deacutecouverte en 1986 cette mycobacteacuterie touche principalement les diffeacuterentes espegraveces de
phoques et les lions de mer sauvages de lrsquoheacutemisphegravere sud ou en captiviteacute Elle a eacuteteacute isoleacutee en Australie
Nouvelle-Zeacutelande Uruguay Argentine et Grande-Bretagne (Cousins et al 2003) Des infections chez
lrsquohomme ont eacuteteacute rapporteacutees principalement chez les soigneurs en contact avec les animaux malades
en captiviteacute (Kiers et al 2008) La transmission indirecte du bacille par inhalation drsquoaeacuterosols lors du
nettoyage des enclos est fortement suspecteacutee
Mycobacterium caprae
Cette mycobacteacuterie du MTBC a eacuteteacute isoleacutee pour la premiegravere fois sur des chegravevres en Espagne
(Aranaz et al 1999 Aranaz et al 2003) Crsquoest lrsquoun des agents de la tuberculose bovine avec M bovis
Chapitre 1
19
et elle induit des leacutesions identiques agrave la tuberculose agrave M bovis chez les animaux infecteacutes La
tuberculose agrave M caprae touche principalement lrsquoEurope ougrave elle a deacutejagrave eacuteteacute deacutetecteacutee en Espagne
(Rodriacuteguez et al 2011) en France (Haddad et al 2001) en Autriche (Prodinger et al 2002) en
Allemagne (Kubica et al 2003) et dans plusieurs pays drsquoEurope centrale (Pavlik et al 2005 Beširović
et al 2012 Cvetnic et al 2007) Le nombre de cas deacutetecteacutes tend agrave augmenter chez les bovins et les
animaux sauvages notamment en Espagne en Allemagne en Autriche (Rodriacuteguez et al 2011 Schoepf
et al 2012) En Europe centrale elle repreacutesente lrsquoagent principal de la tuberculose bovine notamment
en Reacutepublique Tchegraveque Slovaquie Bosnie Herzeacutegovine et Croatie ougrave elle circule entre les bovins et de
nombreuses espegraveces sauvages (Prodinger et al 2005 Beširović et al 2012) Elle a eacutegalement eacuteteacute isoleacutee
pour la premiegravere fois en Chine en 2013 (Zeng et al 2013) Crsquoest eacutegalement un agent zoonotique
important puisque des infections agrave M caprae sont rapporteacutees chez lrsquohomme dans diffeacuterents pays
drsquoEurope ((Kubica et al 2003 Prodinger et al 2014 Prodinger et al 2002 Muumlller et al 2013)
Mycobacterium microti
Cette mycobacteacuterie est lrsquoagent responsable de la tuberculose des petits rongeurs (campagnols
souris musaraignes hellip) chez qui elle a eacuteteacute isoleacutee dans les anneacutees 30 (Wells 1937 Wells amp Robb-Smith
1946) Elle a eacuteteacute isoleacutee chez plusieurs espegraveces de mammifegraveres dont des chats (Gunn-Moore et al 1996
Gunn-Moore et al 2011) des bovins (Jahans et al 2004) des porcs (Taylor et al 2006) des lamas des
blaireaux (Smith et al 2009) des sangliers (Dondo et al 2007) des suricates (Palgrave et al 2012) En
France la deacutetection de M microti chez des animaux domestiques et sauvages a suggeacutereacute que la bacteacuterie
eacutetait endeacutemique dans certaines reacutegions de France (Michelet et al 2015 Laprie et al 2013) Chez
lrsquohomme une trentaine de cas drsquoinfections agrave M microti principalement agrave localisation pulmonaire ont
eacuteteacute reporteacutes dans la litteacuterature entre 2000 et 2010 (Panteix et al 2010) Bien que les personnes
immunodeacuteprimeacutees soient les plus toucheacutees lrsquoinfection se rencontre eacutegalement chez des sujets
immunocompeacutetents (Niemann et al 2000 Frank et al 2009)
Mycobacterium orygis
Isoleacutee pour la premiegravere fois en 1976 sur des oryxes en Afrique de lrsquoest (Lomme et al 1976) elle
a eacuteteacute isoleacutee dans de rares occasions sur drsquoautres espegraveces de ruminants tels que les gazelles les
antilopes (Van Soolingen et al 1994) les buffles africains (van Pittius et al 2012) en Afrique du sud
dans la Peacuteninsule arabe et dans des zoos des Pays Bas et du Royaume Uni
Des infections agrave Mycobacterium orygis ont eacuteteacute rapporteacutees chez une dizaine de patients humains
(Dawson et al 2012 van Ingen et al 2012)
Chapitre 1
20
Autres mycobacteacuteries agrave reacuteservoir animal
Quatre autres espegraveces de mycobacteacuteries animales phylogeacuteneacutetiquement proches de
M africanum West African 2 ont eacuteteacute isoleacutees sur des animaux en Afrique le Chimpazee bacillus isoleacute
sur un chimpanzeacute de Cocircte drsquoIvoire (Coscolla et al 2013) M mungi isoleacutee sur des mangoustes rayeacutees
(Mungos mungo) du Bostwana (Alexander et al 2010) le Dassie bacillus isoleacute sur un daman du Cap
(Procavia capensis) (Wagner et al 1958) et M suricattae isoleacutee sur des suricates (Suricata suricatta) en
Afrique du sud (Parsons et al 2013) Ces espegraveces nrsquoont agrave ce jour jamais eacuteteacute isoleacutees chez lrsquohomme La
deacutecouverte reacutecente de ces espegraveces rattacheacutees au MTBC suggeacutere que ce Complexe est probablement
plus varieacute que ce qui eacutetait imagineacute
B LA TUBERCULOSE UNE MALADIE TRES ANCIENNE MAIS UNE MENACE TOUJOURS ACTUELLE
1 Introduction
La tuberculose est une maladie bacteacuterienne chronique et contagieuse qui affecte lrsquohomme
ainsi que de nombreux animaux domestiques et sauvages Elle est provoqueacutee par les diffeacuterentes
espegraveces de mycobacteacuteries du MTBC Lrsquoobservation de tubercules granulomes inflammatoires formeacutes
de cellules immunitaires et de caseacuteum (zones neacutecrotiques) dans les organes ou les nœuds
lymphatiques des personnes infecteacutees a donneacute son nom agrave la maladie
2 Histoire de la tuberculose
La tuberculose est une maladie tregraves ancienne puisque lrsquoancecirctre du Complexe tuberculosis serait
apparu il y a 40 000 ans (Wirth et al 2008) La preacutesence exclusive de certaines ligneacutees du MTBC sur le
continent africain suggegravere que le Complexe aurait eacutemergeacute en Afrique de lrsquoEst puis aurait co-eacutevolueacute
avec lrsquohomme et se serait reacutepandu dans le monde entier par le biais des migrations humaines en
Europe en Asie et en Afrique continentale Les populations de mycobacteacuteries tuberculeuses eacutetant
infeacuteodeacutees agrave une zone geacuteographique elles se sont adapteacutees agrave la population locale (Gutierrez et al 2005
Djelouadji et al 2011 Wirth et al 2008) Le passage drsquoun mode de vie nomade agrave une vie seacutedentaire
dans des fermes agricoles de plus en plus peupleacutees ainsi que la domestication des animaux ont
probablement contribueacute agrave la transmission de la maladie agrave divers hocirctes animaux et agrave lrsquoadaptation du
bacille permettant lrsquoapparition de M bovis M caprae et des autres membres du Complexe (Wirth et
al 2008) Les plus anciennes traces drsquoADN du Complexe tuberculosis ont eacuteteacute deacutecouvertes sur des
Chapitre 1
21
ossements de bison vieux de 17 000 ans retrouveacutes dans une grotte drsquoAmeacuterique du Nord (Rothschild et
al 2001) Des traces drsquoADN du Complexe tuberculosis de M tuberculosis ou M bovis ont eacuteteacute
retrouveacutees sur des ossements humains datant de la Preacutehistoire ou de lrsquoAntiquiteacute (les plus vieux ont
9000 ans) deacutecouverts dans diffeacuterentes parties du monde (Chili Royaume Uni Russie Israeumll Indoneacutesie
etc) (Djelouadji et al 2011) Appeleacutee laquo phtisie raquo par les Grecs peste blanche ou consomption (suite au
deacutepeacuterissement des malades) les eacutecrits drsquoHippocrate (5egraveme siegravecle avant JC) sont les premiers agrave deacutecrire la
symptomatologie et les diffeacuterentes formes de la tuberculose Fracastor (15egraveme siegravecle) suggegravere le
caractegravere heacutereacuteditaire de la maladie mais place la phtisie dans les maladies contagieuses au mecircme titre
que la veacuterole hypothegravese reprise en 1722 par Marten puis par Villemin en 1865 qui montre que la
maladie est infectieuse contagieuse et inoculable (Dutau 2005) A la fin du 18egraveme siegravecle et au deacutebut du
19egraveme la tuberculose est la premiegravere cause de mortaliteacute en Europe de lrsquoOuest et en Ameacuterique du Nord
La Reacutevolution industrielle et le deacuteveloppement des villes favorisent lrsquoexpansion de la maladie les
conditions de vie insalubres des taudis de banlieues la promiscuiteacute et la pauvreteacute acceacutelegraverent lrsquoendeacutemie
et agrave cette eacutepoque environ une personne sur 4 est atteinte de tuberculose La fin du 19egraveme siegravecle voit se
succeacuteder les deacutecouvertes agrave un rythme effreacuteneacute dont les principales sont la deacutecouverte du bacille en
1882 par Koch la deacutecouverte du bacille bovin en 1896 lrsquoutilisation de la tuberculine dans le diagnostic
humain en 1907 la creacuteation drsquoune souche atteacutenueacutee M bovis BCG en 1921 et le deacutebut de la vaccination
en 1924 La deacutecouverte de la streptomycine en 1944 le premier antibiotique contre la tuberculose
puis de lrsquoisoniazide en 1952 et la rifamycine en 1957 a marqueacute une eacutetape importante dans le traitement
contre la tuberculose (Daniel 2006)
3 La tuberculose bovine
a Introduction
La tuberculose bovine est une maladie bacteacuterienne contagieuse drsquoeacutevolution chronique causeacutee
par Mycobacterium bovis Cette bacteacuterie a pour hocircte originel les bovins mais peut infecter un grand
nombre drsquoespegraveces de mammifegraveres dont lrsquohomme (Gavier-Wideacuten et al 2009 Broughan Crawshaw et
al 2013 Grange amp Yates 1994) ce qui en fait une maladie zoonotique Elle a un tropisme respiratoire
preacutedominant mais peut infecter un grand nombre drsquoautres organes La tuberculose bovine fait partie
des maladies animales agrave deacuteclaration obligatoire aupreacutes de lrsquoOffice International des Epizooties (OIE)
Le passage de lrsquoinfection dans la faune sauvage a compliqueacute le scheacutema drsquoeacuteradication de la
maladie chez les bovins puisque certaines espegraveces sauvages peuvent entretenir la maladie et
contaminer les bovins en retour A lrsquoheure actuelle M bovis fait partie des six organismes pathogegravenes
prioritaires cibleacutes par les programmes de surveillance europeacuteens (Ciliberti et al 2015)
Chapitre 1
22
b Les hocirctes et les communauteacutes drsquohocirctes de Mycobacterium bovis dans le monde
et leurs statuts eacutepideacutemiologiques
Mycobacterium bovis est la mycobacteacuterie du MTBC qui possegravede le plus large spectre drsquohocirctes
incluant lrsquohomme (Grange 2001 OrsquoReilly amp Daborn 1995) Lrsquoinfection a eacuteteacute retrouveacutee chez de
nombreuses espegraveces de mammifegraveres appartenant aux Ordres des marsupiaux des carnivores des
primates des rongeurs des lagomorphes et des artiodactyles (Broughan Downs et al 2013 OrsquoReilly
amp Daborn 1995 Palmer 2013)
La circulation de lrsquoinfection chez des hocirctes appartenant agrave des espegraveces domestiques et sauvages
diffeacuterentes est agrave lrsquoorigine de systegravemes eacutepideacutemiologiques complexes appeleacutes systegraveme multi-hocirctes
(Haydon et al 2002) qui complexifient fortement les scheacutemas drsquoeacuteradication de la maladie Dans ces
systegravemes les diffeacuterents hocirctes peuvent avoir un statut eacutepideacutemiologique diffeacuterent (Haydon et al 2002
Palmer 2013) ou former une communauteacute drsquohocirctes reacuteservoir (Palmer 2013) Les hocirctes de maintenance
ou reacuteservoirs deacutefinissent une population drsquoanimaux dans laquelle lrsquoinfection est entretenue de faccedilon
autonome Ils sont agrave lrsquoorigine de lrsquoinfection des espegraveces cibles Les hocirctes de liaison sont incapables de
maintenir lrsquoinfection indeacutefiniment au sein de leur population agrave moins qursquoils soient reacuteinfecteacutes par une
autre espegravece Ils sont toutefois capables de transmettre lrsquoinfection agrave drsquoautres populations (Corner 2006
Nugent 2011) Et enfin les hocirctes laquo cul-de-sac raquo deacutefinissent les espegraveces qui ne peuvent pas maintenir
ni transmettre la maladie (Nugent 2011) Diffeacuterentes espegraveces ont ainsi eacuteteacute identifieacutees comme des
reacuteservoirs de lrsquoinfection notamment le blaireau (Meles meles) en Angleterre et en Irlande (Delahay et
al 2001) le sanglier (Sus scrofa) dans le centre et le sud de lrsquoEspagne (Naranjo et al 2008) le cerf
eacutelaphe en France (Zanella et al 2008) le cerf de Virginie (Odocoileus virginianus) dans le Michigan
(USA) (Palmer 2013 OrsquoBrien et al 2006) le bison des bois (Bison bison athabascae) et le wapiti (Cervus
elaphus manitobensis) au Canada (Nishi et al 2006 Wobeser 2009) le phalanger renard (Trichosurus
vulpecula) en Nouvelle-Zeacutelande (Morris et al 1994 Nugent 2011) et le buffle (Syncerus caffer) en
Afrique du Sud (Renwick et al 2007) Drsquoautres espegraveces animales ont eacuteteacute identifieacutees comme hocirctes de
liaison comme le furet le cerf et le sanglier en Nouvelle-Zeacutelande
La transmission retour de lrsquoinfection par les espegraveces sauvages reacuteservoir est une menace pour
les bovins et les espegraveces sauvages sensibles vivant agrave proximiteacute La connaissance du statut
eacutepideacutemiologique de chaque espegravece est important pour comprendre le rocircle de la faune sauvage dans la
maintenance et la transmission de lrsquoinfection et adapter les programmes de lutte et drsquoeacuteradication de la
maladie chez les bovins les animaux sauvages et lrsquohomme (Delahay et al 2001 Corner 2006)
Chapitre 1
23
c Aspect zoonotique de Mycobacterium bovis
Plusieurs mycobacteacuteries du MTBC peuvent engendrer une tuberculose zoonotique chez
lrsquohomme Le principal agent infectieux impliqueacute est M bovis (Thoen et al 2006) dans une moindre
mesure M caprae (EFSA 2014) et M microti (Panteix et al 2010) Cette derniegravere a un pouvoir
zoonotique plus faible mais nrsquoest pas neacutegligeable La tuberculose zoonotique agrave M bovis est
majoritairement lieacutee agrave la consommation de produits issus drsquoanimaux infecteacutes principalement le lait cru
et ses deacuteriveacutes (transmission indirecte) (Evans et al 2007) La transmission par aeacuterosols respiratoires est
eacutegalement deacutecrite lors de contacts rapprocheacutes avec des animaux domestiques ou sauvages infecteacutes
(transmission directe) (Cosivi et al 1999 Ayele et al 2004 Kleeberg 1984) Lrsquoinfection est
sporadiquement rapporteacutee chez des eacuteleveurs de bovins et de cerfs (Smith et al 2004 Fanning amp
Edwards 1991) du personnel veacuteteacuterinaire (Shrikrishna et al 2009) et du personnel drsquoabattoir (Liss et al
1994) La transmission interhumaine de M bovis par voie respiratoire existe chez les patients
immunodeacuteprimeacutes ou immunocompeacutetents mais reste rare (Evans et al 2007 Grange amp Yates 1994 de
la Rua-Domenech 2006 Sunder et al 2009 Shrikrishna et al 2009 Rivero et al 2001)
Les signes cliniques chez lrsquohomme sont identiques agrave ceux retrouveacutes lors de tuberculose agrave
M tuberculosis et se traduisent par une tuberculose pulmonaire etou des atteintes extra-pulmonaires
plus freacutequentes qursquoavec M tuberculosis en raison des diffeacuterences dans le mode de transmission
(atteintes osteacuteo-articulaires ganglionnaires uro-geacutenitales disseacutemineacuteeshellip)(Antoine amp Jarlier 2010
Delaune et al 2012 Mignard et al 2006) Le traitement repose sur lrsquoadministration drsquoantituberculeux
classiques agrave lrsquoexception du pyrazinamide pour lequel M bovis est naturellement reacutesistant Des souches
MDR (Multi Drug Resistant) sont reacuteguliegraverement signaleacutees chez lrsquohomme (McLaughlin et al 2012
Cambau et al 2003) De rares souches XDR (Extensively Drug Resistant) sont eacutegalement deacutecrites
majoritairement chez des patients seacuteropositifs (Rivero et al 2001) Dans les pays industrialiseacutes
lrsquoinfection est principalement diagnostiqueacutee chez des personnes acircgeacutees etou immunodeacuteprimeacutees
(diabegravete VIH traitement immunosuppresseur alcoolismehellip) dont les formes extra-pulmonaires
majoritaires eacutevoquent une contamination ancienne par voie digestive (avant la mise en place de la
pasteurisation du lait) et la reacuteactivation ulteacuterieure de la maladie ainsi que chez des personnes neacutees ou
ayant seacutejourneacute dans des pays ougrave la maladie est endeacutemique (Delaune et al 2012)
Les pays ayant mis en place la pasteurisation du lait et des programmes de lutte efficaces contre
la maladie chez les bovins (deacutepistage et abattage) ont fortement reacuteduit lrsquoincidence de la maladie chez
lrsquohomme et les bovins Malgreacute quelques cas reacuteguliegraverement deacutecrits chez lrsquohomme la tuberculose
zoonotique est une zoonose mineure dans les pays deacuteveloppeacutes notamment en Europe (Torgerson amp
Torgerson 2010 Michel et al 2010) En France lrsquoobligation de pasteurisation du lait en 1955 associeacutee
agrave lrsquoabattage systeacutematique des bovins reacuteagissant agrave la tuberculination agrave partir de 1963 ont permis de faire
Chapitre 1
24
baisser lrsquoincidence de lrsquoinfection agrave M bovis chez lrsquohomme de 15 dans les anneacutees 60 agrave 05 en 1995
(Boulahbal et al 1998) Des eacutetudes locales mettent parfois en eacutevidence des preacutevalences plus eacuteleveacutees
de lrsquoordre de 2 agrave Lyon entre 2000 et 2005 (Mignard et al 2006) et de 29 en Aquitaine entre 2004
et 2008 (Aimeacute et al 2012) Au Royaume-Uni par exemple malgreacute lrsquoincidence eacuteleveacutee chez les bovins
(1806 nouveaux foyers entre janvier et avril 2015)(site DEFRA) la preacutevalence de la maladie nrsquoaugmente
pas chez lrsquohomme gracircce aux programmes de lutte mis en place chez lrsquoanimal (Torgerson amp Torgerson
2010 Jalava et al 2007)
La situation est tregraves diffeacuterente dans les pays en voie de deacuteveloppement (PVD) Lrsquoabsence de
programme de lutte contre la tuberculose bovine et la consommation de lait cru dans des pays ougrave la
maladie est endeacutemique chez les bovins notamment dans certains pays drsquoAfrique drsquoAmeacuterique du sud
et drsquoAsie repreacutesentent un risque de santeacute publique important (Cosivi et al 1999 Ayele et al 2004) La
tuberculose zoonotique est souvent fortement relieacutee agrave la preacutevalence de lrsquoinfection bovine dans le pays
ou la reacutegion (Fetene et al 2011) et touche majoritairement les professions en contact avec les bovins
(eacuteleveurs de bovins personnel drsquoabattoir et drsquousines agro-alimentairehellip) et les consommateurs de lait
cru et de fromage frais dont les enfants drsquoautant plus srsquoils sont immunodeacuteprimeacutes Une eacutetude reacutecente
rapporte que la tuberculose zoonotique repreacutesente environ 28 des cas de tuberculose en Afrique
(de 0 agrave 377 selon les pays) 03 des cas sur le continent ameacutericain (de 0 agrave 339 ) et 04 des cas
en Europe (de 0 agrave 211 ) (Muumlller et al 2013) Lrsquoinfection par le HIV la pauvreteacute et le manque drsquoaccegraves
aux soins sont des facteurs de disseacutemination de la tuberculose humaine et zoonotique dans ces pays
(Ayele et al 2004 WHO 2014) Dans les PVD le diagnostic chez lrsquohomme srsquoarrecircte parfois agrave lrsquoobservation
microscopique de BAAR associeacutes agrave M tuberculosis sous-estimant probablement lrsquooccurrence de
M bovis (Cosivi et al 1999 Michel et al 2010) La transmission retour de lrsquoinfection de lrsquohomme aux
bovins pourrait avoir un impact non neacutegligeable dans les PVD mais aucune donneacutee nrsquoest disponible
(de la Rua-Domenech 2006)
d Pathogeacutenie de la tuberculose bovine
La pathogeacutenie de la maladie est multifactorielle et deacutepend de facteurs lieacutes au pouvoir pathogegravene de
M bovis agrave la reacuteceptiviteacute et la sensibiliteacute de lrsquohocircte exposeacute ainsi qursquoagrave la voie drsquoinfection
Facteurs influenccedilant le pouvoir pathogegravene de M bovis voies
drsquoinfection et dose infectante
La voie respiratoire est la voie de contamination majeure chez les bovins (Pritchard 1988
Morris et al 1994 Phillips et al 2003) Des infections expeacuterimentales de M bovis par voie nasale intra
Chapitre 1
25
tracheacuteale ou intra-tonsillaire ainsi que par la mise en contact drsquoanimaux sains avec des animaux
infecteacutes ont confirmeacute lrsquoefficaciteacute de la transmission de M bovis par cette voie (Palmer et al 1999 Dean
et al 2005 Johnson et al 2007 Collins amp Grange 1983 Cassidy et al 1999) La dose infectante par voie
respiratoire est tregraves faible puisque lrsquoinoculation intra-tracheacuteale de 6 agrave 10 bacilles viables agrave des veaux
suffit agrave provoquer des leacutesions pulmonaires etou ganglionnaires (Dean et al 2005 Johnson et al 2007)
Des reacutesultats identiques ont eacuteteacute obtenus chez le blaireau (Corner et al 2007 Corner et al 2011) Chez
le cerf de Virginie lrsquoinoculation intra-tonsillaire de 300 agrave 500 UFC (Uniteacute Formant Colonie) infecte 100
des animaux (Palmer et al 2002 Griffin et al 2006) Lrsquoinoculation oro-pharyngeacutee de 100 UFC agrave des
sangliers deacuteclenche uniquement des leacutesions amygdaliennes alors que 10 000 UFC entrainent des
leacutesions pulmonaires et ganglionnaires seacutevegraveres (Ballesteros et al 2009) Chez toutes ces espegraveces la dose
infectante affecte la seacuteveacuteriteacute et la progression de la maladie (Neill et al 1994 Palmer et al 2002 Corner
et al 2007)
La transmission de la tuberculose bovine peut eacutegalement se faire par voie digestive suite agrave
lrsquoingestion de matrices contamineacutees La dose infectante neacutecessaire serait bien supeacuterieure agrave celle par
voie respiratoire et selon certains auteurs neacutecessiterait plusieurs millions de bacilles (Phillips et al
2003 OrsquoReilly amp Daborn 1995) Pour les veaux Palmer et al (2004) ont estimeacute agrave 5 000 UFC la dose
infectante par cette voie Lrsquoingestion reacutepeacutetitive de faibles doses de M bovis semble eacutegalement
repreacutesenter un facteur de risque comme cela a eacuteteacute deacutemontreacute chez le cochon drsquoInde (Maddock 1934)
La transmission par voie cutaneacutee ou transcutaneacutee ainsi que la transmission de la megravere au veau par
consommation de lait contamineacute ou par leacutechage sont eacutegalement deacutecrites mais restent rares (Phillips et
al 2003 Morris et al 1994) La transmission du bacille par voie transcutaneacutee (morsure) par le biais de
salive infecteacutee est reacuteguliegraverement deacutecrite chez le blaireau en Angleterre (Corner et al 2011)
Facteurs influenccedilant la reacuteceptiviteacute et la sensibiliteacute de lrsquohocircte
De nombreux facteurs intrinsegraveques agrave lrsquoanimal entrent en jeu dans la reacuteceptiviteacute et la sensibiliteacute
de lrsquohocircte agrave la maladie notamment lrsquoespegravece la race lrsquoacircge le sexe et lrsquoeacutetat geacuteneacuteral (Griffin et al 1996
Menzies amp Neill 2000 Humblet et al 2009 Goodchild amp Clifton-Hadley 2001) Certains animaux
preacutesentent une reacutesistance naturelle ou au contraire une sensibiliteacute exacerbeacutee agrave lrsquoinfection ou agrave la
progression de la maladie (Gavier-Wideacuten et al 2009) Des meacutecanismes modulant la reacuteponse
immunitaire et inflammatoire de lrsquohocircte ont pu ecirctre mis en eacutevidence chez le cerf (Mackintosh et al
2000) Les caracteacuteristiques geacuteneacutetiques de lrsquohocircte pourraient eacutegalement jouer un rocircle important dans la
reacutesistance ou la sensibiliteacute agrave lrsquoinfection (Mackintosh et al 2000 Acevedo-Whitehouse et al 2005
Brotherstone et al 2010) Des eacutetudes reacutecentes sur des geacutenomes complets de bovins ont identifieacute des
Chapitre 1
26
loci particuliers impliqueacutes dans la reacutesistance agrave la maladie (Bermingham et al 2014) ouvrant des
perspectives de seacutelection geacutenomique de populations reacutesistantes agrave la tuberculose bovine (Tsairidou et
al 2014)
Le statut immunitaire de lrsquoanimal est un facteur important puisque certains animaux sont
capables drsquoeacuteliminer le bacille gracircce agrave leur immuniteacute inneacutee etou acquise certains sont capables de
controcircler lrsquoinfection plusieurs anneacutees dans les granulomes et drsquoautres deacuteveloppent une tuberculose
eacutevolutive (Pollock amp Neill 2002) La modulation immunitaire expliquerait pourquoi dans un cheptel
bovin malgreacute la contagiositeacute importante de M bovis seuls quelques animaux sont infecteacutes (Cassidy
2006) Les notions de latence et de reacuteactivation de la maladie deacutecrites chez M tuberculosis sont
discuteacutees dans la tuberculose bovine (Cassidy 2006) et commencent agrave ecirctre appreacutehendeacutees (Aacutelvarez et
al 2009)
Les eacutetapes de lrsquoinfection et la reacuteponse immunitaire associeacutee
Lorsque les bacteacuteries peacutenegravetrent dans lrsquoorganisme le plus souvent au niveau des alveacuteoles
pulmonaires elles sont phagocyteacutees par les macrophages Si la dose infectante est importante ou
lrsquoimmuniteacute insuffisante les mycobacteacuteries eacutechappent agrave la digestion et se multiplient permettant la
formation dans le tissu pulmonaire de la leacutesion primaire appeleacutee chancre drsquoinoculation La primo-
infection est caracteacuteriseacutee par le laquo complexe primaire raquo qui associe le chancre drsquoinoculation et les nœuds
lymphatiques infecteacutes par le drainage de la leacutesion primaire (Neill et al 1994) Lrsquoinfection peut ensuite
eacutevoluer selon 3 modes diffeacuterents la gueacuterison avec une eacutelimination des bacilles la stabilisation
caracteacuteriseacutee par une reacuteactivation possible de lrsquoinfection quelques mois ou anneacutees plus tard ou la
geacuteneacuteralisation preacutecoce de lrsquoinfection lieacutee agrave une multiplication active des bacteacuteries et agrave leur
disseacutemination dans lrsquoorganisme Lrsquoinfection et son eacutevolution sont toutefois fortement deacutependantes de
la dose infectante de la voie drsquoentreacutee de la bacteacuterie de la reacuteceptiviteacute et de la sensibiliteacute de lrsquohocircte
eacutevoqueacutees preacuteceacutedemment
Chez lrsquoanimal infecteacute par M bovis la reacuteponse immunitaire agrave meacutediation cellulaire est la premiegravere
agrave se mettre en place dans les stades preacutecoces de lrsquoinfection par la mobilisation de macrophages et de
lymphocytes T (Figure 5) Ces derniers libegraverent diffeacuterentes cytokines dont lrsquointerfeacuteron-γ (IFN-γ) qui
active les macrophages et la formation de granulomes pour isoler et deacutetruire lrsquoagent infectieux
Lrsquoimmuniteacute agrave meacutediation humorale se met en place tardivement et remplace peu agrave peu lrsquoimmuniteacute agrave
meacutediation cellulaire dans les stades plus avanceacutes de la maladie par la production drsquoanticorps par les
lymphocytes B (Pritchard 1988 Ritacco et al 1991 Fifis et al 1994) Les diffeacuterents tests utiliseacutes pour
le deacutepistage de lrsquoinfection chez les bovins reposent principalement sur lrsquoimmuniteacute agrave meacutediation
cellulaire (Figure 5)
Chapitre 1
27
Figure 5 Evolution de la reacuteponse immunitaire chez le bovin infecteacute par M bovis au cours du temps et aux diffeacuterents tests de deacutepistage de la tuberculose bovine (de la Rua-Domenech et al 2006 adapteacute de Vordermeier et al 2004)
Expression clinique de la maladie et leacutesions observeacutees
La tuberculose bovine est une maladie chronique drsquoeacutevolution lente geacuteneacuteralement
asymptomatique dans ses deacutebuts Lorsque la maladie eacutevolue hormis lrsquoamaigrissement possible les
signes cliniques sont tregraves variables et deacutependent des organes ou tissus atteints mais ne sont pas
pathognomoniques (de la Rua-Domenech et al 2006) La localisation des leacutesions observeacutees chez les
animaux deacutepend geacuteneacuteralement de la voie drsquoentreacutee de la bacteacuterie et de lrsquoeacutevolution de la maladie Chez
les bovins la majoriteacute des leacutesions sont retrouveacutees dans la caviteacute thoracique avec une atteinte des lobes
pulmonaires et des nœuds lymphatiques associeacutes (bronchiques et meacutediastinaux) etou des nœuds
lymphatiques ceacutephaliques (reacutetro-pharyngiens principalement) Les leacutesions sur les nœuds lymphatiques
meacutesenteacuteriques la rate et le foie sont plus rares (Neill et al 1994)
Chez le blaireau infecteacute naturellement la seacuteveacuteriteacute de la maladie est tregraves variable drsquoune
infection latente asymptomatique sans leacutesion macroscopique visible agrave une maladie modeacutereacutee avec des
leacutesions pulmonaires ou extra-pulmonaires de petite taille jusqursquoagrave une maladie geacuteneacuteraliseacutee entrainant
la cachexie puis la mort de lrsquoanimal (Murphy et al 2010) Les leacutesions retrouveacutees sont essentiellement
respiratoires probablement lieacutees agrave lrsquoinhalation drsquoaeacuterosols contamineacutes plus rarement digestives et
cutaneacutees suite agrave des morsures de congeacutenegraveres infecteacutes (Corner et al 2011 Payne et al 2012 Gavier-
Widen et al 2001) Toutefois diffeacuterentes eacutetudes ont montreacute que la majoriteacute des blaireaux infecteacutes ne
Chapitre 1
28
preacutesente pas de leacutesion visible (Gallagher amp Clifton-Hadley 2000 Jenkins et al 2008 Payne et al 2012)
Les blaireaux constituent des hocirctes de persitance efficaces puisqursquoils peuvent supporter lrsquoinfection
plusieurs anneacutees et se reproduire tout en excreacutetant la bacteacuterie (Delahay et al 2001)
Chez le sanglier les leacutesions sont majoritairement observeacutees dans les nœuds lymphatiques
ceacutephaliques notamment les mandibulaires dans les poumons et moins freacutequemment au niveau
meacutesenteacuterique (Zanella et al 2008 Martiacuten-Hernando et al 2007 Naranjo et al 2008) Des formes
geacuteneacuteraliseacutees sont eacutegalement deacutecrites (Naranjo et al 2008)
Le cerf est une espegravece deacutecrite comme tregraves sensible agrave lrsquoinfection par M bovis et deacuteveloppe des
leacutesions ouvertes et disseacutemineacutees pouvant ecirctre agrave lrsquoorigine drsquoune excreacutetion bacillaire importante (Lugton
et al 1998 Palmer et al 2004) Les leacutesions sont geacuteneacuteralement localiseacutees au niveau des nœuds
lymphatiques ceacutephaliques et des poumons (Palmer et al 2004 Delahay et al 2002) des nœuds
lymphatiques meacutesenteacuteriques voire geacuteneacuteraliseacutees (Zanella et al 2008)
e Le deacutepistage et le diagnostic de lrsquoinfection chez les animaux
Le deacutepistage de lrsquoinfection chez les bovins vivants repose sur plusieurs modes de surveillance
(Cavalerie et al 2014)
La surveillance systeacutematique agrave lrsquoabattoir concerne tous les bovins abattus pour la consommation
humaine Il repose sur un examen post mortem de lrsquoanimal avec lrsquoinspection drsquoun certain nombre
drsquoorganes cibles de lrsquoinfection (poumons nœuds lymphatiques reacutetro-pharyngiens tracheacuteo-
bronchiques et meacutediastinaux) Dans le cas de leacutesions suspectes visibles celles-ci sont preacuteleveacutees et
transmises agrave un laboratoire agreacuteeacute pour recherche de mycobacteacuteries par PCR et culture
La surveillance programmeacutee en eacutelevage repose sur le deacutepistage de la maladie par prophylaxie Le
rythme des prophylaxies est fixeacute par le preacutefet en fonction de la situation sanitaire du deacutepartement
Dans les deacutepartements ougrave cette surveillance est absente le deacutepistage deacutepend alors uniquement de la
surveillance agrave lrsquoabattoir Pour quelques deacutepartements agrave fort risque elle est annuelle et peut ecirctre
compleacuteteacutee par un deacutepistage lors de mouvements drsquoanimaux Le test de deacutepistage de reacutefeacuterence est
lrsquointradermotuberculination (IDR) Ce test vise agrave mettre en eacutevidence in vivo la reacuteaction
drsquohypersensibiliteacute retardeacutee conseacutecutive agrave lrsquoinjection intradermique de tuberculine (proteacuteines purifieacutees
de surnageant de culture de mycobacteacuteries) chez un bovin infecteacute (Buddle et al 2011 De la Rua-
Domenech et al 2006) Cette reacuteaction se traduit par une tumeacutefaction cutaneacutee douloureuse et chaude
Deux types drsquoIDR sont utiliseacutes lrsquoIntradermotuberculination simple ou IDS qui consiste en lrsquoinjection
intradermique de tuberculine bovine sur le plat de lrsquoencolure et lrsquoIntradermotuberculination
comparative ou IDC qui associe lrsquoinjection intradermique de tuberculine bovine et de tuberculine
aviaire LrsquoIDC est utiliseacutee dans certains deacutepartements dont la Cocircte drsquoOr ougrave de nombreuses
Chapitre 1
29
mycobacteacuteries environnementales sont retrouveacutees pour augmenter la speacutecificiteacute du test et eacutecarter les
reacuteactions croiseacutees chez les bovins sensibiliseacutes agrave une autre mycobacteacuterie (des antigegravenes sont communs
entre les mycobacteacuteries) (De la Rua-Domenech et al 2006 OIE 2009)
LrsquoIDR peut ecirctre compleacuteteacutee par le dosage de lrsquoInterfeacuteron γ (IFNγ) Cette cytokine qui est libeacutereacutee
par les lymphocytes T activeacutes lors drsquoune stimulation antigeacutenique in vivo mais eacutegalement in vitro permet
entre autre lrsquoactivation de la fonction phagocytaire des macrophages La quantiteacute drsquoIFNγ libeacutereacutee lors de
la culture de sang total avec de la tuberculine bovine ou aviaire est suffisante pour ecirctre mesureacutee par
un test ELISA (Gormley et al 2006 de la Rua-Domenech et al 2006) Le dosage de lrsquoIFNγ est pratiqueacute
pour renforcer lrsquoIDR lorsque la contention des animaux est difficile essentiellement en Camargue et
dans les Landes pour les taureaux de combat Il est eacutegalement utiliseacute agrave titre expeacuterimental dans quelques
deacutepartements comme la Dordogne et la Cocircte drsquoOr (Fediaevsky et al 2014) pour renforcer les reacutesultats
drsquoIDR
Drsquoautres tests notamment seacuterologiques sont en cours drsquoeacutevaluation chez les bovins Les essais
reacutealiseacutes jusqursquoagrave maintenant montrent une sensibiliteacute et une speacutecificiteacute infeacuterieures agrave lrsquoIDR ou lrsquoIFNγ
notamment chez les animaux en deacutebut drsquoinfection mais ils repreacutesentent un axe de recherche important
au vu du faible coucirct et de la simpliciteacute de la technique (Waters et al 2011 de la Rua-Domenech et al
2006)
La seacuterologie et lrsquoIFNγ font lrsquoobjet de nombreux deacuteveloppements pour lrsquoameacutelioration de la
surveillance de la maladie chez les animaux sauvages Reacutecemment des techniques ELISA ont eacuteteacute
deacuteveloppeacutees pour de nombreuses espegraveces dont les sangliers les cerfs les bisons (Boadella et al 2011
Shuralev et al 2012 Boadella et al 2012 Lyashchenko et al 2013 Chapinal et al 2012) LrsquoIFNγ est
eacutegalement testeacute pour le deacutepistage chez les espegraveces sauvages notamment chez les cervideacutes drsquoeacutelevage
et les blaireaux (Palmer et al 2010 Dalley et al 2008) La seacuterologie (kit commercial Brock Stat-Pack
Chembio Diagnostic Systems) et le dosage de lrsquoIFNγ (meacutethode ELISA) sont utiliseacutes en routine apregraves
capture et preacutelegravevement sanguin pour le suivi eacutepideacutemiologique de groupes sociaux de blaireaux agrave
Woodchester Park en Angleterre (Drewe et al 2010 King Murphy James Travis Porter Sawyer et al
2015 King Murphy James Travis Porter Hung et al 2015) La combinaison des deux tests est
conseilleacutee pour ameacuteliorer la sensibiliteacute et la speacutecificiteacute du deacutepistage (Drewe et al 2010) Une meacutethode
de deacutepistage non invasive baseacutee sur la collecte puis lrsquoanalyse par qPCR des latrines a eacuteteacute eacutevalueacutee sur
douze groupes sociaux en parallegravele avec la seacuterologie et le dosage de lrsquoIFNγ (King Murphy James Travis
Porter Sawyer et al 2015 King Murphy James Travis Porter Hung et al 2015) Bien que moins
contraignante et moins couteuse que le suivi par capture lrsquoanalyse des latrines pour lrsquoidentification des
groupes sociaux de blaireaux positifs neacutecessite au moins deux campagnes annuelles de preacutelegravevements
(deacutebut et fin drsquoeacuteteacute) avec un nombre drsquoeacutechantillons minimum variable selon les groupes sociaux (de 10
agrave plus de 20) agrave cause de lrsquoexcreacutetion intermittente du bacille par les blaireaux infecteacutes De plus elle ne
Chapitre 1
30
permet pas un deacutepistage individuel
Concernant le diagnostic de la tuberculose bovine lrsquoinfection chez les bovins par M bovis doit
ecirctre eacutetablie par histologie en cas de leacutesion et confirmeacutee par culture selon la reacuteglementation
europeacuteenne Lrsquohistologie est reacutealiseacutee sur des coupes de nœuds lymphatiques et de leacutesions fixeacutees agrave la
formaline La culture repose sur le protocole deacuteveloppeacute et ameacutelioreacute par le Laboratoire National de
Reacutefeacuterence (LNR) devenu la norme NF U47-104 qui associe une phase de broyage-deacutecontamination
des eacutechantillons puis un ensemencement sur deux milieux agrave lrsquoœuf (Loumlwenstein-Jensen et Coletsos) (OIE
2009) Toutefois ces deux techniques preacutesentent des inconveacutenients car lrsquohistologie manque de
speacutecificiteacute (environ 92 ) par rapport agrave la culture (Varello et al 2008) et le reacutesultat de culture est peu
sensible long (entre trois semaines et trois mois) ce qui pose des problegravemes pratiques en cas de
suspicion (restriction des mouvements du cheptel) Bien que pratiqueacutee depuis quelques anneacutees sans
base regraveglementaire il a eacuteteacute deacutemontreacute que la PCR baseacutee sur la deacutetection de lrsquoIS6110 (kit commercial LSI
VetMAXtrade Mycobacterium tuberculosis Complex PCR kit) serait un outil performant de diagnostic de la
tuberculose chez les bovins avec une bonne speacutecificiteacute (97 ) et une sensibiliteacute supeacuterieure agrave la culture
(877 vs 781 ) (Courcoul et al 2014) En apportant un diagnostic en 48 heures la PCR permettrait
une gestion plus rapide des foyers
Le diagnostic dans la faune sauvage repose sur une PCR systeacutematique sur les leacutesions eacutevocatrices
de tuberculose ou un pool de nœuds lymphatiques en lrsquoabsence de leacutesions En cas de reacutesultat non
neacutegatif lrsquoeacutechantillon est mis en culture pour identification de lrsquoagent infectieux (Source Note de service
DGALSDSPA2015-556 du 26062015) Quelle que soit lrsquoespegravece concerneacutee la culture et la PCR sont
reacutealiseacutees par des laboratoires agreacuteeacutes En cas de PCR etou de culture positive les eacutechantillons sont
systeacutematiquement envoyeacutes au Laboratoire National de Reacutefeacuterence pour la tuberculose bovine pour
confirmation et geacutenotypage des souches
f Epideacutemiologie de la tuberculose bovine
Evolution de la tuberculose bovine dans le monde et en
Europe et mesures de lutte
Avant 1920 la tuberculose bovine eacutetait lrsquoune des maladies majeures des animaux domestiques
agrave lrsquoeacutechelle mondiale A lrsquoheure actuelle elle est toujours largement reacutepandue dans le monde (Cosivi et
al 1999) La maladie est plutocirct sporadique et limiteacutee agrave quelques reacutegions dans les pays industrialiseacutes
toucheacutes (sauf en Irlande au Royaume-Uni en Espagne et au Portugal) et endeacutemique dans les pays agrave
faibles revenus drsquoAfrique drsquoAmeacuterique du sud et drsquoAsie (Figure 6)
Chapitre 1
31
Dans les pays industrialiseacutes la mise en place de la pasteurisation du lait dans les anneacutees 1950-
60 et des programmes drsquoeacuteradication de la maladie baseacutes sur le deacutepistage et lrsquoabattage des bovins
infecteacutes ont permis de reacuteduire fortement lrsquoincidence de lrsquoinfection chez les bovins et lrsquohomme
Aujourdrsquohui dans ces pays la tuberculose bovine repreacutesente principalement un enjeu socio-
eacuteconomique lieacute aux restrictions des eacutechanges internationaux drsquoanimaux et aux coucircts des programmes
de lutte contre la maladie chez les animaux le problegraveme sanitaire est devenu mineur Dans les pays agrave
faibles revenus ougrave lrsquoabsence ou quasi-absence de controcircle de la maladie chez les bovins et la
consommation de lait et produits deacuteriveacutes crus les enjeux de la tuberculose bovine sont agrave la fois
sanitaires et socio-eacuteconomiques
Figure 6 Carte de distribution de la tuberculose bovine chez les bovins entre juillet et deacutecembre 2014 (Source WAHID Interface)
En Europe depuis plus drsquoun siegravecle lrsquoeacuteradication de la maladie a eacuteteacute un objectif majeur pour les
organisations agricoles et les autoriteacutes publiques (Reviriego-Gordejo amp Vermeersch 2006) Les mesures
reacuteglementaires eacutelaboreacutees notamment par lrsquoOIE (Office International des Epizooties aujourdrsquohui appeleacute
Organisation Mondiale pour la Santeacute Animale) visent agrave seacutecuriser les eacutechanges commerciaux drsquoanimaux
lrsquoameacutelioration des techniques de deacutepistage de la maladie les programmes drsquoeacuteradication et leurs aides
financiegraveres Les programmes mis en place chez les bovins ont permis de faire fortement diminuer
lrsquoincidence de la maladie dans la plupart des pays de lrsquoUnion Europeacuteenne (de la Rua-Domenech et al
2006) voire de lrsquoeacuteradiquer leur confeacuterant le statut de pays laquo officiellement indemne de tuberculose
bovine raquo (OTF Officially Tuberculosis Free) Le statut OTF est attribueacute agrave un pays lorsque
- le pourcentage de cheptels bovins confirmeacutes comme infecteacutes nrsquoexcegravede pas 01 par an pour
tous les cheptels pendant 6 ans
Chapitre 1
32
- chaque bovin est identifieacute en accord avec la regraveglementation europeacuteenne
- tous les bovins abattus subissent un examen post-mortem officiel (Reviriego-Gordejo amp
Vermeersch 2006)
Ce statut est tregraves important pour lrsquoeacuteconomie drsquoun pays puisqursquoil facilite le commerce national
et international des animaux par un allegravegement des controcircles sanitaires avant lrsquoexportation de leurs
bovins dans un autre pays membre ou avant leur abattage en les exemptant notamment drsquoune
tuberculination (Reviriego-Gordejo amp Vermeersch 2006) Les pays europeacuteens officiellement indemnes
de tuberculose sont lrsquoAllemagne lrsquoAutriche la Belgique le Danemark lrsquoEstonie la Finlande la France
lrsquoEcosse la Lettonie le Liechtenstein le Luxembourg la Norvegravege les Pays-Bas la Pologne la
Reacutepublique Tchegraveque la Slovaquie la Sloveacutenie la Suegravede la Suisse et une partie de lrsquoItalie (Figure 7) Les
pays drsquoEurope les plus toucheacutes sont la Grande-Bretagne lrsquoIrlande et lrsquoEspagne avec des preacutevalences
respectives de 104 44 et 12 en 2012 (EFSA 2014) Toutefois dans certains pays OTF comme la
France la Belgique lrsquoAllemagne lrsquoAutriche et la Pologne lrsquoeacuteradication de la maladie nrsquoest pas complegravete
malgreacute la preacutevention les controcircles et la surveillance engageacutes Dans ces pays lrsquoeacuteradication de la maladie
est fortement compliqueacutee par la circulation de lrsquoinfection dans des reacuteservoirs sauvages source
potentielle de contamination pour les bovins (Corner 2006 Nugent 2011 Gortaacutezar et al 2012)
Figure 7 Statuts des pays europeacuteens vis-agrave-vis de la tuberculose bovine en 2012 et proportion de bovins infecteacutes OTF Officially Tuberculosis Free (officiellement indemne de tuberculose bovine) MS Member State (Etat membre) (Source EFSA 2014)
Chapitre 1
33
Evolution de la situation sanitaire en France
Chez les bovins
Dans les anneacutees 50 environ 25 des cheptels bovins franccedilais eacutetaient infecteacutes par la tuberculose
bovine (Beacutenet et al 2006) Cette preacutevalence importante avait un fort impact eacuteconomique (saisies de
viande agrave lrsquoabattoir diminution de la productiviteacute en viande et en lait) mais aussi sanitaire puisque 2 agrave
11 des cas de tuberculose chez lrsquohomme eacutetaient imputeacutes agrave M bovis Les programmes de lutte mis en
place ont permis de faire diminuer de faccedilon spectaculaire la preacutevalence cheptel de la maladie de 25
en 1955 agrave moins de 01 degraves 1998 permettant agrave la France drsquoacceacuteder au statut OTF en 2000 (Figure 8)
Ce statut a permis drsquoalleacuteger voire de stopper la prophylaxie sur le territoire drsquoalleacuteger les controcircles mis
en place pour les mouvements de bovins dans lrsquoUE et de fournir une garantie sanitaire aux pays
importateurs de bovins franccedilais dans lrsquoUE et en dehors
Les programmes de lutte contre la tuberculose bovine reposent sur une base reacuteglementaire en
eacutevolution constante eacutelaboreacutee par les services veacuteteacuterinaires (maintenant DDPP Direction
Deacutepartementale de la Protection des Populations) en collaboration technique avec les veacuteteacuterinaires
sanitaires les groupements de deacutefense sanitaire (GDS) et les laboratoires drsquoanalyse agreacuteeacutes Ces
programmes sont baseacutes sur la surveillance systeacutematique et geacuteneacuteraliseacutee en abattoir (inspection post-
mortem recherche et identification de mycobacteacuteries par culture et PCR sur les leacutesions suspectes) et
sur la surveillance programmeacutee en eacutelevage (deacutepistage par intradermo-tuberculination et abattage total
ou partiel des troupeaux infecteacutes) Ils assurent eacutegalement la protection des cheptels indemnes par la
maitrise des facteurs de diffusion de la tuberculose bovine tels que lrsquointroduction drsquoun animal le
voisinage avec des cheptels infecteacutes et la reacutesurgence (Fediaevsky et al 2014) Depuis 2013 deux
Cellules Inter Reacutegionales drsquoEpideacutemiologie Veacuteteacuterinaire (CIREV) ont eacuteteacute creacuteeacutees en Bourgogne et en
Aquitaine afin de renforcer les investigations eacutepideacutemiologiques pour adapter de faccedilon optimale les
mesures de lutte contre la maladie (Fediaevsky et al 2013)
Chapitre 1
34
Figure 8 Evolution du taux de preacutevalence annuelle national de la tuberculose bovine chez les bovins (Beacutenet et al 2006)
Depuis 2004 la France connait une situation preacuteoccupante vis-agrave-vis de la tuberculose bovine
car lrsquoaugmentation du taux de preacutevalence met en danger le statut OTF du pays (Figure 9) Cette
augmentation est lieacutee agrave lrsquoapparition de nouveaux foyers de tuberculose concentreacutes dans des zones
geacuteographiques bien localiseacutees la Cocircte drsquoOr la Dordogne et les Pyreacuteneacutees Atlantiques cumulent 75
des foyers et dans une moindre mesure la Charente la Camargue les Hautes Pyreacuteneacutees lrsquoAriegravege les
Landes et la Corse (Figure 10) Dans ces deacutepartements caracteacuteriseacutes par de lrsquoeacutelevage allaitant
preacutedominant la contamination des troupeaux par le voisinage (par contacts directs avec des bovins
voisins ou des animaux sauvages infecteacutes ou par contacts indirects avec du mateacuteriel ou des matrices
contaminants) et lrsquointroduction drsquoun animal infecteacute dans le cheptel (notamment par lrsquoachat drsquoanimaux
le precirct et la pension) repreacutesentaient la moitieacute des cas en 2011 La reacutesurgence drsquoune ancienne infection
(qui persiste agrave bas bruit chez les animaux ou dans des matrices environnementales souilleacutees) et la faune
sauvage eacutetaient agrave lrsquoorigine preacutesumeacutee drsquoenviron 10 des cas respectivement toutefois sans
deacutemonstration scientifique Pour 35 des foyers bovins lrsquoorigine de lrsquoinfection eacutetait inconnue
(Fediaevsky et al 2012) La contamination des bovins via le voisinage la reacutesurgence et les foyers
drsquoorigine inconnue soulegravevent la question du rocircle de la transmission indirecte de M bovis entre la faune
sauvage et les bovins et donc le rocircle potentiel de lrsquoenvironnement comme reacuteservoir du bacille
25
15
10
5
20
1955
1963
1960
1970
1980
1990
2000
Chapitre 1
35
Figure 9 Evolution de la preacutevalence et de lrsquoincidence de la tuberculose bovine en France entre 1995 et 2014 (Cavalerie et al 2014)
La menace de la perte du statut OTF inquiegravete les pouvoirs publics et les instances agricoles car
la France est le plus gros exportateur de bovins vivants de lrsquoUE elle exporte principalement des bovins
maigres en Italie et en Espagne pour lrsquoengraissement (environ 12 million de tecirctes) mais aussi des gros
bovins destineacutes agrave la boucherie (220 000 tecirctes) et des veaux (128 000 tecirctes) (Source France AgriMer
les filiegraveres de lrsquoeacutelevage franccedilais eacutedition 2013) Les conseacutequences eacuteconomiques sur la filiegravere-
agroalimentaire lieacutees au blocage des animaux seraient consideacuterables De plus la gestion sanitaire de la
tuberculose bovine est tregraves coucircteuse puisque environ 20 millions drsquoeuros sont deacutepenseacutes chaque anneacutee
agrave lrsquoeacutechelle nationale pour lrsquoindemnisation des eacuteleveurs et la reacutealisation des analyses veacuteteacuterinaires
(Fediaevsky et al 2014)
Chapitre 1
36
Figure 10 Incidence et distribution de la tuberculose bovine en France dans les anneacutees 50 (en haut) et entre 2000 et 2014 (en bas) (Cavalerie et al 2014)
Chapitre 1
37
Dans la faune sauvage
En France agrave lrsquoheure actuelle la faune sauvage infecteacutee nrsquoest deacutetecteacutee qursquoagrave proximiteacute de foyers
bovins Le 1er foyer de tuberculose bovine dans la faune sauvage a eacuteteacute deacutecouvert en 2001 dans la forecirct
de Brotonne (Seine-Maritime et Eure) sur des cervideacutes puis des sangliers (Hars et al 2010 Zanella et
al 2008) Lrsquoaggravation de la situation sanitaire en 2005-2006 a neacutecessiteacute un abattage total des cerfs
consideacutereacutes comme reacuteservoir primaire de lrsquoinfection ainsi qursquoune diminution de la densiteacute des sangliers
reacuteservoir secondaire (Zanella et al 2008) Drsquoautres deacutepartements comme la Cocircte drsquoOr la Corse les
Pyreacuteneacutees Atlantiques la Dordogne et la Charente notamment ont par la suite deacutetecteacute des animaux
sauvages infecteacutes essentiellement blaireaux sangliers et dans une moindre mesure cervideacutes avec
toutefois des situations eacutepideacutemiologiques tregraves diffeacuterentes selon les reacutegions toucheacutees Ces situations
deacutependent de la population animale impliqueacutee (espegraveces densiteacutes interactionshellip) et de
lrsquoenvironnement (climat paysagehellip) Dans tous les cas le geacutenotypage (spoligotype et VNTR) des
souches incrimineacutees dans la faune sauvage a montreacute qursquoelles eacutetaient identiques agrave celles retrouveacutees
dans les eacutelevages bovins voisins suggeacuterant un lien eacutepideacutemiologique entre cas domestiques et sauvages
(Zanella et al 2008 Payne et al 2012 Richomme et al 2010)
Depuis 2011 un reacuteseau de surveillance eacutepideacutemiologique de la tuberculose bovine dans la faune
sauvage appeleacute Sylvatub a eacuteteacute mis en place en France Les diffeacuterentes modaliteacutes de surveillance ont
pour but la deacutetection de lrsquoinfection chez les animaux sauvages dans les zones agrave risque et preacutesumeacutees
indemnes ainsi que le suivi de son eacutevolution afin de proposer des mesures de lutte aux autoriteacutes
sanitaires (Riviegravere et al 2011) La surveillance est baseacutee sur un classement du territoire national en 3
niveaux de risque Pour les zones situeacutees en risque 1 (risque faible correspondant agrave une absence de
bovins deacutetecteacutes) il est demandeacute aux chasseurs de signaler toute leacutesion suspecte lors de lrsquoexamen initial
de la venaison Concernant les zones ougrave il existe un risque aveacutereacute (niveau 2 ou 3) la surveillance peut
ecirctre agrave la fois eacuteveacutenementielle (ramassage et analyse des cadavres drsquoanimaux trouveacutes morts) etou
programmeacutee (analyse des cerfs et sangliers tueacutes agrave la chasse et les blaireaux pieacutegeacutes) Les reacutesultats des
analyses effectueacutees en 2013-2014 sont preacutesenteacutes dans la Figure 11
Chapitre 1
38
Figure 11 Reacutepartition et reacutesultats des analyses effectueacutees dans le cadre du dispositif Sylvatub chez le grand gibier drsquoaoucirct 2013 agrave aoucirct 2014 et chez les blaireaux en 2014 (Source Sylvatub)
Evolution de la situation sanitaire en Cocircte drsquoOr
Quelques geacuteneacuteraliteacutes sur lrsquoeacutelevage bovin en Cocircte drsquoOr
La Cocircte drsquoOr est un deacutepartement drsquoeacutelevage bovin principalement allaitant (231 270 tecirctes en 2013
dont 74 565 vaches allaitantes et 15 687 vaches laitiegraveres) Lrsquoeacutelevage allaitant est majoritairement
concentreacute dans les zones herbagegraveres de lrsquoouest du deacutepartement et lrsquoeacutelevage laitier sur le plateau de
Langres au nord de Dijon Lrsquoeacutelevage allaitant est extensif (les bovins sont au preacute drsquoavril agrave octobre) lieacute agrave
la production de broutards maigres destineacutes agrave ecirctre engraisseacutes principalement en Italie En Cocircte drsquoOr le
parcellaire des exploitations de bovins allaitant est souvent tregraves morceleacute
Situation eacutepideacutemiologique chez les bovins et mesures de lutte associeacutees
Historiquement la Cocircte drsquoOr est un deacutepartement en prise avec la tuberculose depuis de
nombreuses anneacutees (Figure 10) A la fin des anneacutees 50 le taux de preacutevalence annuel chez les bovins
atteignait 65 Les mesures de lutte collective mises en place ont permis drsquoabaisser ce taux en dessous
du seuil des 10 en 1969 puis de le faire diminuer lentement et reacuteguliegraverement jusqursquoen 1985 ougrave la
maladie semblait eacuteradiqueacutee A la fin des anneacutees 90 les mesures prophylactiques ont cesseacute dans le
Chapitre 1
39
deacutepartement et la vigilance vis-agrave-vis de la maladie a baisseacute (Communication personnelle DDPP 21)
Les premiers signes drsquoune nouvelle eacutepizootie sont apparus en 2002 sur le canton de Pouilly-en-
Auxois (zone Sud) puis en 2003 dans ceux de Venarey-Les-Laumes et Vitteaux (zone Nord) (Source
DDPP21) Localiseacutee agrave quelques eacutelevages agrave son deacutebut la zone de lrsquoinfection srsquoest peu agrave peu eacutelargie au
cours du temps Un plan de prophylaxie restreint aux communes infecteacutees a eacuteteacute mis en place en 2002
puis eacutelargi aux cantons en 2003 avec de plus une surveillance des cheptels en lien eacutepideacutemiologique
avec les troupeaux infecteacutes et ceux des communes dans lesquelles les bovins infecteacutes avaient patucircreacute
Mais ces mesures nrsquoont pas suffi agrave limiter la progression de lrsquoinfection puisque limiteacute agrave moins de 5 par
an jusqursquoen 2006 le nombre de foyers est passeacute de 11 en 2007 agrave 17 en 2009 (Figure 13) Le
renforcement de la lutte comprenant notamment le deacutepistage annuel de tous les bovins de plus de 12
mois dans tout le deacutepartement a permis la deacutecouverte de 45 foyers en 2010 signe que lrsquoinfection
circulait agrave bas bruit De 2011 agrave 2013 le nombre de foyers a eacuteteacute diviseacute par 2 et pour lrsquoanneacutee 2014 10
foyers seulement ont eacuteteacute mis en eacutevidence ce qui semble conforter une maitrise de lrsquoeacutepizootie (Figure
13)
En Cocircte drsquoOr les foyers bovins sont circonscrits agrave une zone geacuteographique bien deacutelimiteacutee dans le
deacutepartement et partagent le mecircme spoligotype M bovis spoligotype SB0134 dans la zone Sud et
M bovis spoligotype SB0120 dans la zone Nord (Figure 12) Depuis 3 ans toutefois les deux zones ont
tendance agrave se teacutelescoper agrave cause notamment de la deacutecouverte drsquoanimaux sauvages infecteacutes avec
M bovis SB0120 en zone Sud Les deux spoligotypes ont deacutejagrave eacuteteacute retrouveacutes simultaneacutement dans des
cheptels infecteacutes Ces observations associeacutees agrave lrsquoabsence drsquoinstallation peacuterenne de la maladie dans des
zones du deacutepartements ougrave les concentrations de bovins sont aussi eacuteleacuteveacutees que dans la zone infecteacutee
soulegravevent la question de lrsquoexistence de facteurs locaux assurant lrsquoentretien et la propagation de
lrsquoinfection entre les eacutelevages bovins Au vu du morcellement et de lrsquoimbrication des pacirctures la
contamination par voisinage est probablement le facteur primordial chez les bovins Une eacutetude reacutecente
(de type cas-teacutemoin) reacutealiseacutee dans des cheptels de Cocircte drsquoOr de Dordogne et des Ardennes a montreacute
que les contacts de voisinage entre troupeaux eacutetaient le facteur de risque principal drsquoinfection des
troupeaux (Marsot et al Article soumis) Toutefois la deacutetection drsquoanimaux sauvages agrave proximiteacute de
certains foyers bovins interroge sur le rocircle potentiel de la faune sauvage infecteacutee dans la contamination
des bovins et dans la recontamination rapide de plusieurs cheptels apregraves abattage total des bovins et
vide sanitaire (Figure 13) (ANSES 2011)
Chapitre 1
40
Figure 12 Localisation du parcellaire des cheptels infecteacutes par la tuberculose bovine entre 2010 et 2015 et spoligotypes incrimineacutes (Source CIREV)
Figure 13 Nombre de nouveaux foyers bovins et de foyers recontamineacutes entre 2002 et 2014 (Donneacutees DDPP21)
0
5
10
15
20
25
30
35
40
45
50
2002 2003 2004 2005 2006 2007 2008 2009 2010 2011 2012 2013 2014
No
mb
re d
e fo
yers
bo
vin
s
Foyers contamineacutes pour la 1egravere fois Foyers recontamineacutes
Chapitre 1
41
Situation eacutepideacutemiologique dans la faune sauvage et mesures de lutte associeacutees
Le premier animal sauvage infecteacute une biche a eacuteteacute deacutecouvert en 2003 agrave proximiteacute de 2 foyers
bovins de 2002 dans la zone Sud Entre 2003 et 2007 la surveillance active de la faune sauvage mise
en place et porteacutee par lrsquoONCFS (Office National de la Chasse et de la Faune Sauvage) et la DDPP et sur
le terrain par les chasseurs les pieacutegeurs et les veacuteteacuterinaires a permis de mettre en eacutevidence lrsquoinfection
chez 1 cervideacute (n=284) 2 sangliers (n= 160) mais aucun blaireau (n=63) Depuis 2007 des sangliers
infecteacutes sont retrouveacutes tous les ans notamment de jeunes animaux avec des leacutesions eacutevolutives
majoritairement dans la zone Sud ougrave les populations sont plus importantes Depuis 2009 lrsquoinfection
est retrouveacutee chez des blaireaux vivant agrave proximiteacute des cheptels infecteacutes majoritairement dans la zone
Nord (Hars Richomme and Boschiroli 2010 DDPP21)(Figure 14) La maladie circule donc dans un
systegraveme multi-hocirctes complexe et unique incluant les bovins les blaireaux les sangliers et les cervideacutes
(Payne 2014)
Figure 14 Nombre de cervideacutes sangliers et blaireaux infecteacutes deacutecouverts en Cocircte drsquoOr entre 2002 et 2014
Dans la crainte de lrsquoeacutetablissement drsquoun reacuteservoir sauvage chez le sanglier comme en Espagne
(Naranjo et al 2008) ou le blaireau au Royaume-Uni (Delahay et al 2001) des mesures de diminution
des densiteacutes ont eacuteteacute mises en place (augmentation des tableaux de chasse pour le sanglier et les
cervideacutes et intensification du pieacutegeage des blaireaux) le ramassage des viscegraveres drsquoanimaux tueacutes agrave la
chasse a eacuteteacute rendu obligatoire et lrsquoagrainage interdit
0
5
10
15
20
25
30
35
No
mb
re d
an
imau
x in
fect
eacutes
Cervideacutes Sangliers Blaireaux
Chapitre 1
42
C ECOLOGIE DE M BOVIS DANS LrsquoENVIRONNEMENT
1 Introduction
Avant les anneacutees 30 il eacutetait admis sine qua none qursquoen tant que bacteacuteries intracellulaires
obligatoires les bacteacuteries tuberculeuses ne survivaient pas en dehors de leur hocircte Toutefois degraves le
deacutebut du 20egraveme siegravecle la deacutetection de Bacillus tuberculosis virulent dans lrsquoeau et les biofilms des
canalisations drsquoeffluents de diffeacuterents sanatoria en Allemagne et au Royaume-Uni (Musehold en 1900
citeacute par Jensen 1954 et Conroy et al en 1921 et 1922 et Cummins et Ackland en 1929 citeacutes par (Williams
amp Hoy 1930) a souleveacute la question du rocircle potentiel de lrsquoenvironnement dans lrsquoeacutepideacutemiologie des
mycobacteacuteries tuberculeuses Concernant la tuberculose bovine les diffeacuterentes recherches meneacutees
pour deacutetecter et caracteacuteriser la persistance de M bovis dans lrsquoenvironnement ont pour objectif
principal de preacuteciser le rocircle de la transmission indirecte dans leacutepideacutemiologie de la maladie La
transmission indirecte semble en effet prendre toute son importance lorsque des reacuteservoirs sauvages
sont identifieacutes etou des systegravemes multi-hocirctes car ils ont la capaciteacute de transmettre lrsquoinfection aux
bovins (Corner 2006 Payne 2014)
Reacutepondre agrave cette question neacutecessite de connaitre les modaliteacutes de transfert des bacteacuteries de
lrsquoanimal ou lrsquohomme infecteacute vers lrsquoenvironnement la dureacutee de survie dans les diffeacuterentes matrices les
strateacutegies de survie de la bacteacuterie ainsi que les facteurs qui affectent sa survie
2 Origines de la contamination environnementale par M bovis
a Lrsquoexcreacutetion animale de Mycobacterium bovis
Lrsquohomme et les animaux infecteacutes ont la capaciteacute drsquoexcreacuteter le bacille via diffeacuterentes voies et
constituent donc une source de contamination potentielle pour lrsquoenvironnement Comme nous lrsquoavons
indiqueacute preacuteceacutedemment les voies drsquoexcreacutetion ainsi que les niveaux drsquoexcreacutetion deacutependent de la
localisation des leacutesions et de leur seacuteveacuteriteacute Les tableaux leacutesionnels deacutecrits preacuteceacutedemment indiquent
que M bovis est susceptible drsquoecirctre excreacuteteacutee principalement par voie orale ou nasale lors drsquoatteinte
pulmonaire etou des nœuds lymphatiques ceacutephaliques par voie digestive via les matiegraveres feacutecales et
par voie urinaire lors drsquoinfection geacuteneacuteraliseacutee (Phillips et al 2003 Corner et al 2012) Lrsquoexcreacutetion feacutecale
de bacteacuteries du MTBC en lrsquoabsence de leacutesions gastro-intestinales semble majoritairement lieacutee au
passage de mucus respiratoire chargeacute en bacilles dans le tube digestif La reacutesistance aux pH acides
(Vandal et al 2008) des bacteacuteries du MTBC leur permet de reacutesister au passage du tractus digestif et
drsquoecirctre excreacuteteacutees sous forme viable dans les matiegraveres feacutecales comme cela a eacuteteacute deacutemontreacute pour M bovis
dans certaines espegraveces animales (Corner Murphy and Gormley 2011 Gallagher and Clifton-Hadley
Chapitre 1
43
2000 Gavier-Widen et al 2001) pour M tuberculosis (El Kheacutechine et al 2009) et M africanum
(Cadmus et al 2009)
Chez les bovins lrsquoexcreacutetion du bacille a eacuteteacute deacutemontreacutee dans le mucus nasal et tracheacuteal et les
fegraveces drsquoanimaux infecteacutes expeacuterimentalement (Neill et al 1988 McCorry et al 2005 Phillips et al 2003
Maddock 1936) Le deacutelai drsquoexcreacutetion apregraves lrsquoinoculation la dureacutee de lrsquoexcreacutetion (plusieurs semaines) et
la quantiteacute de bacilles excreacuteteacutes sont proportionnels agrave la dose infectieuse utiliseacutee (Neill et al 1988)
Lrsquoexcreacutetion feacutecale apparait peu freacutequente et intermittente chez les bovins (Neill et al 1988 Goodchild
amp Clifton-Hadley 2001) Dans les pays industrialiseacutes ougrave le deacutepistage des animaux infecteacutes est en place
lrsquoexcreacutetion de M bovis dans le lait les fegraveces et les urines est consideacutereacutee comme insignifiante chez les
bovins (Morris et al 1994)
Chez le blaireau infecteacute naturellement ou expeacuterimentalement M bovis est excreacuteteacutee par 25 agrave
50 des animaux par voie respiratoire (mucus tracheacuteal et aeacuterosols) urinaire et feacutecale ainsi que par la
suppuration drsquoabcegraves reacutesultant de morsures (Corner et al 2011 Delahay et al 2000 Clifton-Hadley et
al 1993) Lrsquoisolement de M bovis dans des glandes salivaires de blaireaux suggegravere que la salive
constitue eacutegalement une voie possible drsquoexcreacutetion du bacille (Payne 2014) Les blaireaux preacutesentant un
stade avanceacute de la maladie et des leacutesions macroscopiques semblent ecirctre les plus excreacuteteurs en terme
de dureacutee et drsquointensiteacute drsquoexcreacutetion (Corner et al 2012 Gallagher amp Clifton-Hadley 2000 Jenkins et al
2008) toutefois lrsquoexcreacutetion du bacille a eacuteteacute deacutecrite chez des animaux sans leacutesions visibles (Jenkins et al
2008 Gavier-Wideacuten et al 2009) Dans les stades avanceacutes de la maladie les quantiteacutes excreacuteteacutees peuvent
atteindre 2105 UFCmL1 de mucus bronchique 25105 UFCmL1 drsquourine et 75104 UFCg1 de fegraveces
(Gallagher et al 1998) Le bacille est excreacuteteacute par voie feacutecale de 165 agrave 1305 jours avant la mort de
lrsquoanimal (Little et al 1982) Dans cette espegravece lrsquoexcreacutetion est geacuteneacuteralement intermittente mais des
individus laquo super excreacuteteurs raquo ont eacuteteacute deacutecrits et se caracteacuterisent par une excreacutetion persistante dans le
temps ou par lrsquoexcreacutetion du bacille par plusieurs voies simultaneacutement (Delahay et al 2000) Des
blaireaux laquo super excreacuteteurs raquo ont eacuteteacute mis en eacutevidence en Cocircte drsquoOr la voie feacutecale eacutetant toujours
positive chez ces animaux (Payne 2014) Lrsquointensiteacute de lrsquoexcreacutetion semble varier avec lrsquoacircge (les adultes
excregravetent plus que les jeunes) et la saison puisque lrsquoexcreacutetion semble supeacuterieure au printemps et en
eacuteteacute (Delahay et al 2013 King Murphy James Travis Porter Hung et al 2015)
Chez le sanglier lrsquoexcreacutetion de M bovis se produit par voie orale nasale feacutecale et urinaire
(Martiacuten-Hernando et al 2007 Santos Almeida et al 2015 Barasona et al 2015) Lrsquoexcreacutetion salivaire
est encore discuteacutee (Payne 2014) En Espagne lrsquoexcreacutetion par au moins une des voies a eacuteteacute mise en
eacutevidence par PCR sur 30 des sangliers analyseacutes (n = 39) et 25 des 39 animaux excreacutetaient par les trois
voies (Barasona et al 2015) Lrsquoexcreacutetion eacutetait majoritairement associeacutee agrave une maladie geacuteneacuteraliseacutee Au
Portugal sur 39 sangliers adultes ou subadultes infecteacutes analyseacutes 80 excreacutetaient le bacille par au
moins une voie et 13 eacutetaient des laquo super excreacuteteurs raquo (excreacutetant par trois voies diffeacuterentes) Environ
Chapitre 1
44
28 des sangliers excreacutetaient des quantiteacutes de bacilles supeacuterieures agrave 1 000 UFC par g ou mL drsquoexcreacuteta
par au moins une voie (Santos Almeida et al 2015) Chez le sanglier agrave notre connaissance lrsquoexcreacutetion
nrsquoa jamais eacuteteacute deacutecrite en lrsquoabsence de leacutesions
Chez les cervideacutes diffeacuterentes eacutetudes ont montreacute que le bacille eacutetait excreacuteteacute de faccedilon tregraves
sporadique dans les mucus nasal oro-pharyngeacute et tracheacuteal ainsi que dans les matiegraveres feacutecales lors
drsquoinfections naturelles ou expeacuterimentales (Lugton et al 1998 Santos Almeida et al 2015 Palmer et
al 2001) Dans cette espegravece une excreacutetion intermittente apparait degraves les premiers stades de la
maladie la seacuteveacuteriteacute des tableaux leacutesionnels rencontreacutes explique le niveau drsquoexcreacutetion eacuteleveacute (Palmer et
al 2001 Santos Almeida et al 2015)
Les carcasses drsquoanimaux infecteacutes peuvent constituer une source drsquoinfection directe pour les
animaux charognards qui srsquoen nourrissent mais eacutegalement une source de contamination indirecte pour
les bovins qui pacircturent sur le site de deacutecomposition de la carcasse (Morris et al 1994) Concernant les
animaux tueacutes agrave la chasse lrsquoabandon des viscegraveres ou leur ensevelissement repreacutesente eacutegalement un
risque pour les animaux charognards De ce fait en France la collecte et lrsquoeacutelimination des viscegraveres des
espegraveces sensibles sont obligatoires dans les zones toucheacutees par la tuberculose bovine
b Le transfert de Mycobacterium bovis par les effluents drsquoeacutelevage ou urbains
Les eacutetudes sur la contamination potentielle des effluents drsquoeacutelevage avec M bovis sont tregraves
rares probablement agrave cause de la difficulteacute agrave cultiver le bacille dans ces matrices complexes Les
multiples voies drsquoexcreacutetion du bacille et plus particuliegraverement lrsquoexcreacutetion par voie feacutecale mecircme si elle
est intermittente peuvent probablement contribuer agrave la contamination des effluents drsquoeacutelevage bovin
tels que les fumiers lisiers et eaux useacutees Lrsquoeacutepandage de ces effluents sur les pacirctures ou lrsquoirrigation des
cultures peut constituer un vecteur de contamination du sol des cultures et de lrsquoeau profonde ou de
surface et repreacutesenter un risque pour les animaux et lrsquohomme mais aucune donneacutee chiffreacutee nrsquoexiste
pour ces matrices en conditions naturelles Lrsquoabattage et la deacutecoupe de bovins tuberculeux peut
eacutegalement repreacutesenter une source de contamination des rejets drsquoabattoir (Jensen 1954) Au vu des
mesures actuelles de collecte stockage traitement et controcircle des effluents drsquoabattoir cette
contamination semble tregraves peu probable en France Toutefois elle reste complegravetement valable dans
les zones drsquoendeacutemie des pays nrsquoappliquant pas ces mesures et ne disposant pas de structures
drsquoabattage adeacutequates
Il est inteacuteressant de noter que quelques eacutetudes sont disponibles concernant la contamination
drsquoeffluents urbains avec M tuberculosis Jensen (1954) a montreacute que les effluents bruts de sanatoria
de 5 villes des Pays-Bas les effluents purifieacutes les boues digeacutereacutees et les boues seacutecheacutees (preacuteleveacutees 5
semaines apregraves lrsquoeacutepandage) contenaient des bacilles virulents de M tuberculosis Plus reacutecemment la
Chapitre 1
45
mise en culture de 800 eacutechantillons drsquoeau et 700 eacutechantillons de sol preacuteleveacutes dans 3 quartiers de
Teacuteheacuteran (Iran) entre 2012 et 2014 a deacutemontreacute la preacutesence de M tuberculosis dans 71 des eacutechantillons
drsquoeau (88) et 11 eacutechantillons de sol (15) Les eacutechantillons drsquoeau contamineacutes ont eacuteteacute retrouveacutes
principalement dans les rigoles de ciment acheminant les eaux useacutees et pluviales (5671) et dans les
eaux traiteacutees (1571) La bacteacuterie nrsquoa toutefois pas eacuteteacute isoleacutee du reacuteseau drsquoeau potable Environ 10 des
souches isoleacutees eacutetaient multi-reacutesistants aux antibiotiques (Velayati et al 2015)
3 Deacutetection et survie de Mycobacterium bovis dans lrsquoenvironnement
a Deacutetection de Mycobacterium bovis en conditions naturelles
A ce jour peu drsquoeacutetudes concernant la deacutetection de M bovis dans des substrats naturellement
contamineacutes ont eacuteteacute publieacutees La deacutetection de M bovis dans des eacutechantillons environnementaux
naturellement contamineacutes gracircce agrave des meacutethodes baseacutees sur la culture est exceptionnelle A
Woodchester Park dans le Gloucestershire (Sud-Ouest de lrsquoAngleterre) ougrave les densiteacutes de blaireaux
sont eacuteleveacutees et la preacutevalence importante (20 en 2000 et 35 en 2005) un suivi longitudinal du statut
des populations est reacutealiseacute depuis 1980 (Delahay et al 2000) et diffeacuterentes campagnes de
preacutelegravevements de sols de terriers et de latrines ont eacuteteacute reacutealiseacutees Le traitement drsquoeacutechantillons de sol et
latrines provenant de Woodchester Park traiteacute par immunocapture magneacutetique a permis pour la
premiegravere et seule fois lrsquoisolement de M bovis par culture dans des eacutechantillons environnementaux
(Sweeney et al 2006 Sweeney et al 2007) Cette technique est toutefois peu applicable agrave un grand
nombre drsquoeacutechantillons en termes de coucirct et de temps et nrsquoa jamais eacuteteacute reacuteutiliseacutee par la suite
Drsquoautres tentatives de deacutetection de M bovis par diffeacuterentes meacutethodes baseacutees sur la culture
dans des zones endeacutemiques de tuberculose bovine ont eacutechoueacute malgreacute le grand nombre drsquoeacutechantillons
environnementaux preacuteleveacutes (Pillai et al 2000 Fine Bolin et al 2011 Iivanainen et al 1999 Williams
amp Hoy 1930a Little et al 1982) La deacutetection de M bovis dans des eacutechantillons environnementaux par
la culture est compliqueacutee par la nature des eacutechantillons analyseacutes en effet ces derniers (sol seacutediments
eau stagnante matiegraveres feacutecales etc) contiennent des micro-organismes tregraves divers et abondants La
croissance de M bovis eacutetant tregraves lente les organismes agrave croissance rapide contaminent rapidement les
milieux de culture compliquant voire empecircchant lrsquoisolement de la bacteacuterie Des traitements de
deacutecontamination preacuteliminaires sont indispensables pour eacuteliminer cette microflore mais la
deacutecontamination reacuteduit en geacuteneacuteral fortement la viabiliteacute de M bovis reacuteduisant aussi la sensibiliteacute des
meacutethodes baseacutees sur la culture (Fine Bolin et al 2011) Des microorganismes voisins des
mycobacteacuteries et preacutesents dans les sols peuvent eacutegalement reacutesister aux proceacutedeacutes de deacutecontamination
et ecirctre retrouveacutes dans les isolements De plus lrsquointermittence et le faible niveau drsquoexcreacutetion chez les
Chapitre 1
46
animaux infecteacutes associeacutes au volume limiteacute des preacutelegravevements analyseacutes diminuent les chances de
deacutetection de M bovis dans lrsquoenvironnement (Fine OrsquoBrien et al 2011) Lrsquoameacutelioration des protocoles
de traitement des eacutechantillons avant la culture est fondamentale pour lrsquoisolement des souches
environnementales
Le deacuteveloppement des outils moleacuteculaires a permis de contourner les inconveacutenients lieacutes aux
techniques baseacutees sur la culture deacutecrits preacuteceacutedemment En Irlande lrsquoanalyse de preacutelegravevements reacutealiseacutes
en 2000 dans un eacutelevage bovin toucheacute plusieurs fois par la tuberculose a mis en eacutevidence par des
meacutethodes moleacuteculaires la preacutesence de 1 times 103 agrave 25 times 103 copies de gegravenes cibles de M bovis (mpb64 et
mpb70) dans des pacirctures et des sols drsquoentreacutee de terriers de blaireaux adjacents aux pacirctures (Young et
al 2005) La reacuteiteacuteration des analyses 30 mois plus tard en novembre 2002 a donneacute le mecircme reacutesultat
sur les terriers bien qursquoils soient resteacutes inhabiteacutes Le rocircle des blaireaux dans la contamination des bovins
est fortement suspecteacute dans cette ferme En Angleterre lrsquoanalyse moleacuteculaire de preacutelegravevements
environnementaux (terriers et latrines) reacutealiseacutes en 2003 autour ou dans les pacirctures drsquoeacutelevages bovins
infecteacutes dans six zones endeacutemiques de tuberculose bovine a mis en eacutevidence M bovis dans les sols
de terriers de blaireaux de 78 des fermes analyseacutees (n=60) Dans chaque ferme 47 des terriers et
29 des latrines analyseacutes eacutetaient positifs en PCR une forte proportion eacutetait accessible aux bovins
(Courtenay et al 2006)
Entre 2004 et 2013 lrsquoanalyse par deacutetection moleacuteculaire de latrines appartenantt agrave des groupes sociaux
de blaireaux infecteacutes de Woodchester Park a montreacute que selon les eacutetudes 13 agrave 100 des latrines eacutetaient
contamineacutees avec des quantiteacutes pouvant atteindre 11 times 108 copies de gegravenes par g de latrine
(Courtenay et al 2006 Travis et al 2011 Sweeney et al 2006) Lrsquoamplification de seacutequences ciblant
lrsquoARNr 16S dans certains eacutechantillons des eacutetudes preacutesenteacutees ci-dessus suggegravere que les bacilles deacutetecteacutes
eacutetaient viables (Young et al 2005 Courtenay et al 2006) toutefois la seacutequence de lrsquoARNr 16S cibleacutee
eacutetant speacutecifique de toutes les mycobacteacuteries agrave croissance lente les reacutesultats sont agrave consideacuterer avec
prudence Dans ces eacutetudes le nombre de blaireaux excreacuteteurs et leur niveau drsquoexcreacutetion sont
fortement correacuteleacutes agrave la contamination des terriers et des latrines
Dans le sud du Portugal une eacutetude reacutecente dans une zone endeacutemique a mis en eacutevidence par PCR ciblant
lrsquoIS6110 des bacteacuteries du MTBC dans 38 des 248 eacutechantillons preacuteleveacutes dans des zones drsquoagreacutegation
drsquoanimaux domestiques et sauvages zones drsquoabreuvement utiliseacutees par les bovins et la faune sauvage
(seacutediments et eau drsquoeacutetangs 1358 et 357 et de sources saisonniegraveres 1161) sol des zones de
nourrissage des animaux (528) du sol de boutis de sangliers (416) ainsi que dans le sol des zones de
nourrissage des vautours (119) et dans leurs fegraveces (19) (Santos Santos et al 2015) Lrsquoutilisation drsquoune
seconde cible PCR (RD12) a confirmeacute 4 eacutechantillons comme positifs pour M bovisM caprae (2
eacutechantillons de seacutediments drsquoeacutetang 1 de source et 1 de sol de nourrissage) sans pouvoir diffeacuterencier les
deux espegraveces Le plus grand nombre drsquoeacutechantillons positifs a eacuteteacute retrouveacute au printemps (tempeacuterature
Chapitre 1
47
moyenne de 16degC)
La PCR permet drsquoobtenir un reacutesultat rapide avec une speacutecificiteacute acceptable Lrsquoanalyse
drsquoeacutechantillons de sol drsquoeau de foin et de maiumls inoculeacutes avec M bovis par une meacutethode baseacutee sur la
culture et la PCR a montreacute que cette derniegravere permet de deacutetecter lrsquoADN de M bovis dans lrsquoeacutechantillon
sur une plus longue peacuteriode apregraves la contamination que la culture indeacutependamment du substrat
(Adams et al 2013) Toutefois les seuils de deacutetection eacuteleveacutes des systegravemes speacutecifiques de M bovis (de
4 times 104 agrave 5 times 105 par g ou mL drsquoeacutechantillon Tableau 3) limitent leur utilisation dans le screening
drsquoeacutechantillons environnementaux pour lesquels la charge bacteacuterienne est probablement infeacuterieure au
seuil de deacutetection Enfin la technique PCR ne donne aucune information sur la viabiliteacute et la virulence
des bacteacuteries deacutetecteacutees
b Dureacutees de survie de Mycobacterium bovis en conditions expeacuterimentales
Les eacutetudes expeacuterimentales sur la survie environnementale de M bovis ont deacutebuteacute dans les
anneacutees 1930 en Angleterre dans lrsquoobjectif de preacuteciser le rocircle de lrsquoenvironnement dans lrsquoeacutepideacutemiologie
de la tuberculose bovine et drsquoameacuteliorer les programmes drsquoeacuteradication de la maladie (Maddock 1933
Maddock 1934 Maddock 1936 Williams amp Hoy 1930) Mycobacterium bovis et M bovis BCG sont
capables de survivre dans les fegraveces les effluents le sol et les divers excreacutetas animaux de quelques jours
agrave quelques mois voire jusqursquoagrave deux ans () Les nombreuses conditions drsquoinoculation (concentration de
lrsquoinoculum souche utiliseacutee M bovis ou M bovis BCG origine de lrsquoinoculum culture ou broyat de
leacutesions) la varieacuteteacute des matrices inoculeacutees (matiegraveres feacutecales urine mucus organes sol eau etc) et
des conditions environnementales testeacutees (conditions controcircleacutees de laboratoire ou exposition aux
conditions climatiques) dans ces diffeacuterentes eacutetudes expliquent vraisemblablement de tels eacutecarts de
survie
La compilation de toutes ces donneacutees indique que la tempeacuterature lrsquoexposition aux UV et
lrsquohumiditeacute de la matrice semblent ecirctre les trois facteurs primordiaux intervenant dans la persistance
de M bovis quel que soit le substrat analyseacute Il est montreacute que dans lrsquoheacutemisphegravere Nord la dureacutee de
survie de M bovis est plus longue en automne hiver et printemps (Williams amp Hoy 1930 Tanner amp
Michel 1999 Fine Bolin et al 2011 Duffield amp Young 1985 Jackson et al 1995) saisons caracteacuteriseacutees
par des tempeacuteratures plus basses qursquoen eacuteteacute un ensoleillement minimal et une eacutevapotranspiration par
le couvert veacutegeacutetal limiteacutee Toutefois Fine et al (2011) ont montreacute que M bovis pouvait survivre jusqursquoagrave
48 jours dans lrsquoeau en eacuteteacute malgreacute les tempeacuteratures eacuteleveacutees et lrsquoensoleillement Les eacutetudes reacutealiseacutees en
Afrique du Sud et en Nouvelle-Zeacutelande ougrave les tempeacuteratures sont eacuteleveacutees sur de longues dureacutees avec
un ensoleillement important ont montreacute que la survie de M bovis est ameacutelioreacutee lorsque le sol contient
une humiditeacute suffisante sous une exposition ombrageacutee (Duffield amp Young 1985 Tanner amp Michel 1999)
Chapitre 1
48
Lrsquoexposition des bacilles aux UV diminue en effet fortement leur dureacutee de persistance dans
lrsquoenvironnement (Williams amp Hoy 1930a Genov 1965 Fine Bolin et al 2011) et la dureacutee de survie est
tregraves prolongeacutee lorsque les eacutechantillons sont enterreacutes agrave quelques centimegravetres sous la surface du sol
(Genov 1965) La survie de M bovis est optimale dans les matiegraveres feacutecales ougrave le bacille est proteacutegeacute
dans un environnement humide agrave lrsquoabri des radiations solaires et minimale lorsqursquoil est pulveacuteriseacute sur
les veacutegeacutetaux et soumis aux UV et agrave la dessiccation (Maddock 1933 Van Donsel amp Larkin 1977)
49
Reacutefeacuterences BCG M bovis Matrices inoculeacutees Concentration de
lrsquoinoculum Conditions
Meacutethodes de deacutetection
de M bovis Reacutesultats de survie (virulence) de M bovis
(Williams amp Hoy
1930)
X Fegraveces de bovin
Inconnue Expeacuterimentales
(tempeacuterature
fraicircche obscuriteacute)
Injection sous-
cutaneacutee agrave des cochons
drsquoInde (cuisse)
12 mois dans fegraveces naturellement infecteacutees
5104 agrave 5107 UFC 2 ans dans fegraveces artificiellement infecteacutees
X Lisier de bovin 5000 UFCcm3
Expeacuterimentales
(tempeacuterature
fraicircche obscuriteacute)
Injection sous-
cutaneacutee agrave des cochons
drsquoInde (cuisse)
Survie 4 mois avec diminution de la virulence du
bacille au cours du temps
X Fegraveces de bovin 5104 agrave 5107 UFC Environnementales
(pacircture)
Injection sous-cutaneacutee
agrave des cochons drsquoInde
(cuisse)
5 mois en hiver
4 mois en automne
2 mois au printemps eacuteteacute
4 mois si protection UV en eacuteteacute
6 mois si protection insectes-vers en automne
(Maddock 1933)
X
Sol
Sol + Fegraveces de bovin
Bouse
Inconnue Environnementales
(pacircture)
Injection sous-cutaneacutee
agrave des cochons drsquoInde
(cuisse)
178 j dans toutes les matrices
X Herbe
12105 01 m2 Environnementales
(pacircture)
Injection sous-
cutaneacutee agrave des cochons
drsquoInde (cuisse)
14 jours
12106 01 m2 28 jours
120106 01 m2 49 jours
(Maddock 1934) X Herbe
Inoculation
mensuelle de 5 agrave
25106 01 m2
reacutepeacuteteacutee
Environnementales
et expeacuterimentales
Ingestion drsquoherbe
contamineacutee
1422 cochons drsquoInde avec leacutesions
Tableau 1 Dureacutees de survie de M bovis dans diffeacuterentes matrices environnementales obtenues en conditions expeacuterimentales naturelles ou de laboratoire
50
(Maddock 1936)
X
Veaux infecteacutes
(excreacutetion prouveacutee
par injection agrave des
cochons drsquoInde)
Inconnue Environnementales
Pacircture seacutepareacutee en 3 2
veaux mis 1 mois apregraves
le retrait des veaux
infecteacutes 2 veaux 1 mois
plus tard 2 veaux 2
mois plus tard (laisseacutes 3
sem)
Aucun veau IDR +
Aucune leacutesion tuberculeuse agrave lrsquoautopsie
X
Vache infecteacutee
(excreacutetion prouveacutee
par injection agrave des
cochons drsquoInde)
Pacircture seacutepareacutee en 2 3
veaux mis 1 mois et 12
apregraves le retrait de la
vache infecteacutee (laisseacutes
53j) 3 veaux mis 1 mois
plus tard (laisseacutes 24j)
Abattage et autopsie 1 an plus tard aucune leacutesion
Bouses infectieuses 1 mois et frac12 apregraves retrait de la
vache infecteacutee puis neacutegatif 1 2 et 3 mois plus tard
Sol et herbe sous bouses neacutegatifs 3 4 et 5 mois apregraves
retrait de la vache infecteacutee
(Genov 1965) X
Fegraveces de bovin
Sang
Urine
Environnementales Culture
150 agrave 332 jours agrave 12-24degC sans drsquoUV
18 agrave 31 jours agrave 24-34degC avec UV
2 ans si enterreacute dans sol agrave 5 cm
11 mois si enterreacute agrave 1 cm
452 et 469 jours dans eaubouse et eauurine
(Van Donsel amp
Larkin 1977) X
Effluents animaux
Boues drsquoeacutepuration Environnementales
Abattage 90 en 11 jours pour sols arroseacutes avec
effluents 8 jours avec boues
Isolement apregraves 4 agrave 6 jours sur des radis et jusqursquoagrave
35 jours sur des laitues
Anon 1979 citeacute par
(Morris et al 1994)
Excreacutetas de
blaireaux conserveacutes
dans un terrier
Environnementales Culture
Urine 3 jours en eacuteteacute 28 jours en hiver
Mucus bronchique 7 et 70 jours
Fegraveces naturellement infecteacutes 14 et 28 jours
(Little et al 1982)
Sol foin litiegravere et
eau de blaireaux
inoculeacutes
Inconnue
Expeacuterimentales
Culture 1 culture positive (eau drsquoun abreuvoir)
51
(Duffield amp Young
1985)
Sol sec steacuterile ou non
Sol 30 humiditeacute
steacuterile ou non
Fegraveces
Culture
lt8 semaines dans sols secs ou humides non steacuteriles
avec 80 ombre (34degc) obscuriteacute (32degC) et dans
labo (23degC)
lt8 semaines dans sol humide steacuterile avec 80
ombre et obscuriteacute
Pas de survie plein soleil (43degC) dans sols et fegraveces
(Tanner amp Michel
1999)
Leacutesions pulmonaires
ou ganglionnaires de
buffle
Fegraveces de bovin
inoculeacutees
8107 UFC 50g
fegraveces Culture
Poumon 42 jours en hiver agrave lrsquoombre dans sol
humide ou sec (20degC)
Fegraveces 28 jours en hiver au soleil dans sol humide
ou sec
(Jackson et al
1995) X Rubans de coton
Environ 106 UFC
ruban
Environnementales
(rubans deacuteposeacutes
dans terriers de
possum et sols de
pacirctures ou de forecirct)
Culture
2 jours sur les sols de pacirctures
14 jours sur les sols forestiers et dans les terriers de
possum
(Scanlon amp Quinn
2000b) X Lisier bovin steacuteriliseacute 65 times 103 UFCmL Expeacuterimentales Culture 171 jours (concentration finale 2 UFCmL)
(Young et al 2005) X Sol steacuterile et non
steacuterile 108 UFC g sol
Expeacuterimentales
(4 15 25 37degC)
PCR pour les sols non
steacuteriles
Culture pour les sols
steacuteriles
Survie agrave 60 jours optimale agrave 37degC agrave -20kPa
drsquohumiditeacute dans le sol steacuterile
Deacutetection drsquoARNm pendant 15 mois (sol steacuterile)
(Palmer amp Whipple
2006) X
Betterave pommes
carottes pommes de
terre maiumls foin
11 times 106
UFC10g
Expeacuterimentales
(-20 8 et 23degC)
Culture
112 jours agrave -20degC pour tous les substrats
112 jours agrave 8degC pour pommes maiumls foin et pdeterre
112 jours agrave 23degC pour pommes maiumls et pdeterre
(Fine Bolin et al
2011) X Sol foin maiumls eau
50 times 104
UFCeacutechantillon Environnementales Culture
88 jours dans le sol en hiverprintemps
58 jours dans lrsquoeau 43 jours dans le foin et 37 jours
dans le maiumls en automnehiver
52
(Ghodbane et al
2014) X Sol steacuterile 108 UFC g sol
Expeacuterimentales
(18 agrave 24degC
obscuriteacute)
Culture
Survie 12 mois (concentration finale 150 UFCg
sol)
Virulence conserveacutee sur souris
Chapitre 1
53
Dans les fegraveces drsquoanimaux M bovis survit jusqursquo agrave 2 ans dans les bouses de bovins en conditions
de laboratoire (Williams amp Hoy 1930) et de 14 jours agrave 6 mois lorsque les fegraveces sont exposeacutees dans les
pacirctures (Williams amp Hoy 1930a Genov 1965 Morris et al 1994 Tanner amp Michel 1999) Dans les
effluents drsquoeacutelevages contamineacutes il a eacuteteacute montreacute que M bovis survivait de 4 mois dans du lisier en
conditions de laboratoire (Williams amp Hoy 1930a) et jusqursquoagrave 171 jours dans du lisier steacuteriliseacute conserveacute
agrave tempeacuterature ambiante et agrave lrsquoobscuriteacute (Scanlon amp Quinn 2000b) Lrsquoeacutepandage drsquoeffluents ou de boues
inoculeacutees avec M bovis BCG sur des cultures de radis et laitue a montreacute la reacuteduction de 90 des bacilles
en respectivement 11 et 8 jours M bovis a toutefois eacuteteacute isoleacutee de faccedilon sporadique 35 jours apregraves
arrosage (Van Donsel amp Larkin 1977)
Dans le sol les dureacutees de survie les plus longues pour M bovis et M bovis BCG ont eacuteteacute
observeacutees dans du sol steacuterile (63 de sable pH 65-68) et en conditions de laboratoire (Young et al
2005 Ghodbane et al 2014) Young et al (2005) ont amplifieacute lrsquoARNr 16S de M bovis BCG jusqursquoagrave 15
mois apregraves inoculation du sol alors que Ghodbane et al (2014) ont cultiveacute M bovis apregraves 12 mois de
seacutejour dans le sol (pH 83) Dans les conditions environnementales Fine et al (2011) ont confirmeacute
lrsquoeffet de la saison sur la survie de M bovis dans le sol sur du maiumls et du foin avec une survie maximale
en hiverprintemps (feacutevrier-mai tempeacuterature moyenne 29degC radiations solaires 800 Wm2) une
survie de 34 jours en automnehiver (novembre-janvier tempeacuterature moyenne 04degC radiations
solaires 243 Wm2) et une survie minimale de 12 jours pendant la peacuteriode printempseacuteteacute (mai-aoucirct
tempeacuterature moyenne 217degC radiations solaires 1500 Wm2) De faccedilon contradictoire lrsquoeacutetude de
Young et al (2005) a mis en eacutevidence une dureacutee de survie optimale (60 jours) de M bovis BCG agrave 37degC
et minimale (16 jours) agrave 4degC
La survie de M bovis a eacutegalement eacuteteacute mise en eacutevidence par meacutethode culturale sur diffeacuterents
aliments du beacutetail (Palmer amp Whipple 2006 Fine Bolin et al 2011) Apregraves 112 jours la bacteacuterie est
cultiveacutee sur tous les substrats inoculeacutes conserveacutes agrave -20degC (betteraves pommes carottes pommes de
terre maiumls et foin) sur les pommes le maiumls le foin et les pommes de terre conserveacutes agrave 8degC et
uniquement sur les pommes le maiumls et les pommes de terre conserveacutes agrave 23degC (Palmer amp Whipple
2006) Fine et al (2011) ont eacutegalement montreacute que M bovis survivait plus longuement en
automnehiver sur du maiumls (43 jours) et du foin (37 jours) qursquoagrave la peacuteriode printempseacuteteacute (survie
infeacuterieure agrave 3 jours)
La bacteacuterie peut eacutegalement survivre entre 2 et 6 semaines dans des leacutesions tuberculeuses
macroscopiques de poumons et nœuds lymphatiques de buffle (Tanner amp Michel 1999) la survie
optimale eacutetant observeacutee en hiver dans des situations ombrageacutees et humides Des reacutesultats identiques
sont observeacutes pour des eacutechantillons drsquourine (3 jours en eacuteteacute vs 28 jours en hiver) et des seacutecreacutetions
bronchiques (7 jours vs 70 jours) (Morris et al 1994)
Les diffeacuterentes eacutetudes preacutesenteacutees ci-dessus deacutemontrent que M bovis survit un temps variable dans
Chapitre 1
54
lrsquoenvironnement une fois excreacuteteacute par son hocircte Ce temps est fonction de lrsquoexcreacuteta et du substrat qui le
reccediloit de sa charge bacteacuterienne et des conditions climatiques Il existe donc une peacuteriode appeleacutee
fenecirctre temporelle critique pendant laquelle les animaux qui interagissent avec les matrices
contamineacutees par des bacilles viables preacutesentent un risque accru drsquoinfection Comme nous lrsquoavons vu
preacuteceacutedemment la fenecirctre temporelle critique est fortement deacutependante du pays ou de la reacutegion
eacutetudieacutes et des conditions climatiques
Tregraves peu de donneacutees existent sur la conservation de la virulence de M bovis apregraves un passage
en dehors de son hocircte La virulence du bacille a eacuteteacute confirmeacutee par injection agrave des cobayes dans des
fegraveces de bovins naturellement contamineacutees apregraves 178 jours drsquoexposition dans une pacircture et apregraves 12
mois drsquoincubation en conditions de laboratoire (Maddock 1933) Lrsquoeacutetude reacutecente de Ghodbane et al
(2014) a deacutemontreacute que M tuberculosis M canetti et M bovis conservaient leur virulence apregraves un
seacutejour de 12 mois dans le sol En effet lrsquoinjection intra-peacuteritoneacuteale de suspensions bacteacuteriennes agrave des
souris a entraineacute lrsquoapparition de leacutesions avec M canetti (nodules sur les poumons le foie et la rate) et
M tuberculosis (nodules sur les poumons et la rate) Malgreacute lrsquoabsence de leacutesions lors de lrsquoinjection de
M bovis la bacteacuterie a pu ecirctre isoleacutee en quantiteacute importante des 3 organes Toutefois les injections
eacutetaient reacutealiseacutees avec les colonies de mycobacteacuteries obtenues apregraves culture sur milieu geacuteloseacute induisant
un biais important De plus lrsquoingestion par des souris de sol inoculeacute par M tuberculosis apregraves 12 mois
drsquoincubation a provoqueacute des leacutesions pulmonaires (1 souris5) et des leacutesions de la rate (25) mais la
bacteacuterie a pu ecirctre cultiveacutee agrave partir des poumons de la rate de lrsquoestomac et de lrsquointestin chez toutes les
souris
4 Strateacutegies de survie des mycobacteacuteries dans lrsquoenvironnement
Lrsquoenveloppe particuliegravere des mycobacteacuteries deacutecrite preacuteceacutedemment est probablement un des
facteurs majeurs de la reacutesistance de M bovis en dehors de son hocircte dans des conditions
environnementales deacutefavorables Drsquoautres strateacutegies de survie sont probablement mises en jeu par
M bovis pour persister dans lrsquoenvironnement mais sont totalement inconnues agrave ce jour Les
mycobacteacuteries pathogegravenes et opportunistes ont deacuteveloppeacute diffeacuterents meacutecanismes pour survivre agrave
lrsquointeacuterieur des cellules de lrsquohocircte strateacutegies qui peuvent eacutegalement ecirctre utiliseacutees en dehors de lrsquohocircte
Les meacutecanismes moleacuteculaires de certains processus sont eacutegalement en cours de compreacutehension Les
principales strateacutegies de survie sont preacutesenteacutees dans la partie suivante
a Lrsquoeacutetat de dormance
La dormance est deacutefinie chez les bacteacuteries non sporuleacutees comme un eacutetat physiologique
Chapitre 1
55
reacuteversible permettant la survie de la bacteacuterie pendant une longue peacuteriode sans croissance ni reacuteplication
(Kaprelyants et al 1993) Le programme geacuteneacutetique de la bacteacuterie lui permet drsquoentrer en dormance (en
anglais Non-Replicating Persistence) lorsque les conditions exteacuterieures deviennent deacutefavorables
(notamment en conditions oligotrophiques) puis de retrouver sa capaciteacute agrave se diviser lorsque les
conditions redeviennent favorables La dormance est deacutecrite chez plusieurs espegraveces de mycobacteacuteries
Lrsquoentreacutee en dormance de M smegmatis (Dick et al 1998) M bovis (Lim et al 1999 Boon et al 2001)
et M tuberculosis (Wayne amp Hayes 1996) en culture est provoqueacutee par la rareacutefaction de lrsquooxygegravene
Lrsquoeacutetat de dormance est suggeacutereacute chez Map et pourrait expliquer sa longue survie dans le fumier le lisier
et le sol (Whittington et al 2004) Lrsquoeacutetat de dormance est eacutegalement deacutecrit chez M avium (Archuleta
et al 2005)
Chez les mycobacteacuteries lrsquoentreacutee en eacutetat de dormance est controcircleacutee par le facteur de
transcription DosR (Dormancy Survival Regulator) (Boon amp Dick 2002) Composeacute de 48 gegravenes
(Schnappinger et al 2003 Flores Valdez amp Schoolnik 2010) son expression est activeacutee in vitro par
lrsquooxyde nitrique (NO) (Voskuil et al 2003) le monoxyde de carbone (CO) (Shiloh et al 2008) le deacuteficit
en nutriments (Rodriguez et al 2008) et lrsquohypoxie (Rodriguez et al 2008) Lrsquoexpression de ce reacutegulon a
eacuteteacute deacutemontreacutee lors drsquoinfection expeacuterimentale de macrophages (Schnappinger et al 2003 Rodriguez et
al 2008) et drsquoanimaux modegraveles (Voskuil et al 2003) pour M tuberculosis et dans des nœuds
lymphatiques de bovins positifs en IDR pour M bovis (Jimeacutenez et al 2013) Ce reacutegulon permettrait non
seulement la survie de la bacteacuterie pendant la phase de dormance mais aussi la transition de la
respiration aeacuterobie agrave lrsquoanaeacuterobiose et inversement (Leistikow et al 2010) A lrsquoheure actuelle le rocircle de
certaines proteacuteines codantes et drsquoARN non codants de ce reacutegulon demeure toujours inconnu Une
eacutetude geacutenomique comparative a permis de constater que le reacutegulon de la dormance eacutetait conserveacute
chez les diffeacuterentes espegraveces de mycobacteacuteries y compris chez les mycobacteacuteries environnementales
(Gerasimova et al 2011) Cette donneacutee suggegravere que la dormance est une reacuteponse adaptative
probablement tregraves ancienne chez les mycobacteacuteries pour survivre dans des environnements ougrave les
concentrations en oxygegravene sont fluctuantes (Boon amp Dick 2012) et que cette adaptation a eacuteteacute mise agrave
profit par les mycobacteacuteries pour survivre dans les cellules immunitaires des hocirctes qursquoelles infectent
Aucune eacutetude nrsquoest agrave ce jour disponible concernant lrsquoentreacutee en dormance potentielle de
M bovis dans lrsquoenvironnement Le sol constitue toutefois un environnement hostile ougrave facteurs
biotiques et abiotiques pourraient deacuteclencher le passage de la bacteacuterie drsquoun eacutetat actif agrave un eacutetat
dormant (hypoxie carence en nutriments tempeacuterature hellip)
b La sporulation
Lrsquoeacutevocation de la sporulation chez les mycobacteacuteries est tregraves controverseacutee Ghosh et al (Ghosh
Chapitre 1
56
et al 2009) ont deacutecrit la sporulation expeacuterimentale de M marinum sous forme drsquoendospores dans des
cultures liquides acircgeacutees de 2 mois et dans des cultures solides de M bovis BCG de 8 mois Malgreacute des
veacuterifications pheacutenotypiques biochimiques et geacutenomiques deacutemontrant que ces spores eacutetaient bien
issues de M marinum ces reacutesultats ont eacuteteacute contesteacutes par Traag et al (Traag et al 2010) qui ont eacutechoueacute
agrave les reproduire Lamont et al (Lamont et al 2012) ont pour la premiegravere fois mis en eacutevidence
lrsquoapparition de morphotype spore-like dans des cultures liquides de Map drsquoorigine bovine (K-10) ovine
(7565) et humaine acircgeacutees drsquoun an en conditions oligotrophiques Le seacutequenccedilage de lrsquoARNr 16S et
lrsquoamplification de lrsquoIS900 speacutecifique de Map ont confirmeacute que les colonies bacteacuteriennes issues des
spores correspondaient bien agrave Map Le chauffage des spores obtenues agrave 70degC pendant 30 minutes ne
semble pas affecter sa virulence sur les macrophages Des eacutetudes compleacutementaires sont neacutecessaires
pour confirmer ces observations A notre connaissance la sporulation nrsquoa jamais eacuteteacute eacutevoqueacutee pour les
mycobacteacuteries du MTBC
c Interactions des mycobacteacuteries avec la faune du sol
La microfaune et plus particuliegraverement les amibes libres sont largement preacutesentes dans
lrsquoenvironnement et colonisent les eacutecosystegravemes aquatiques le sol (Khan 2006 Rodriacuteguez-Zaragoza
1994) et lrsquoair Elles ont eacuteteacute eacutegalement isoleacutees chez lrsquohomme Elles vivent preacutefeacuterentiellement dans les
biofilms et aux interfaces eau-sol eau-air et eau-plantes (Rodriacuteguez-Zaragoza 1994) ougrave elles
laquo broutent raquo les bacteacuteries les champignons et les algues dont elles reacutegulent les communauteacutes (Huws
et al 2005) Les mycobacteacuteries environnementales sont largement preacutesentes dans les milieux
aquatiques (Primm et al 2004) ougrave elles peuvent interagir avec les amibes De nombreuses eacutetudes
rapportent la capaciteacute des mycobacteacuteries environnementales agrave survivre et se multiplier dans les
trophozoiumltes etou les kystes drsquoamibes (Adekambi et al 2006 Drancourt et al 2007 Ben Salah amp
Drancourt 2010 Krishna-Prasad amp Gupta 1978 Mura amp Bull 2006) Les amibes pourraient mecircme
constituer un veacuteritable reacuteservoir pour M xenopi (Drancourt et al 2007 Thomas et al 2006)
La plupart des mycobacteacuteries pathogegravenes y compris M bovis sont capables de survivre dans
les amibes voire de srsquoy multiplier (Thomas amp McDonnell 2007 Mba Medie et al 2011 Greub amp Raoult
2004 Cirillo et al 1997 Adekambi et al 2006) La survie de M bovis a eacuteteacute confirmeacutee dans les
trophozoiumltes et les kystes drsquoAcanthamoeba castellanii jusqursquoagrave 5 mois apregraves infection en conditions
expeacuterimentales (Taylor et al 2003 Mardare et al 2013) La virulence vis-agrave-vis des macrophages
humains et des cellules intestinales de souris semble ecirctre exacerbeacutee par la reacuteplication intra-amibienne
(Cirillo et al 1997) En conditions naturelles lrsquoobservation de protozoaires isoleacutes de latrines de
blaireaux infecteacutes a mis en eacutevidence des BAAR sans pouvoir identifier preacuteciseacutement M bovis (Mardare
et al 2013) Des eacutetudes compleacutementaires sont neacutecessaires pour preacuteciser le rocircle de reacuteservoir eacuteventuel
Chapitre 1
57
des amibes pour M bovis Les interactions des mycobacteacuteries pathogegravenes avec les amibes pourraient
ecirctre du mecircme ordre que celles avec les macrophages de lrsquohocircte infecteacute notamment au niveau des
meacutecanismes de peacuteneacutetration (Danelishvili et al 2007) et de lrsquoinhibition de la fusion entre le phagosome
et le lysosome suggeacuterant qursquoune relation eacutevolutive des mycobacteacuteries avec les amibes auraient permis
aux mycobacteacuteries lrsquoacquisition de ces caracteacuteristiques de survie (Cirillo et al 1997)
La localisation intra-amibienne des mycobacteacuteries les protegravege des conditions
environnementales deacutefavorables des biocides tels que le chlore (Adekambi et al 2006 Whan et al
2006) des antibiotiques (Miltner amp Bermudez 2000) des diffeacuterents traitements comme lrsquoozonation de
lrsquoeau (Thomas et al 2008) et favorise leur disseacutemination et leur implication potentielle dans des
maladies humaines (Greub amp Raoult 2004) Lors de la peacuteneacutetration dans lrsquohocircte lrsquoamibe joue un rocircle de
laquo cheval de Troie raquo (Barker amp Brown 1994) proteacutegeant les bacteacuteries contre les premiegraveres deacutefenses de
lrsquohocircte
Lrsquoutilisation des amibes en co-cultures pour isoler des bacteacuteries reacutesistantes aux amibes dans
des eacutechantillons environnementaux et meacutedicaux a permis drsquoisoler des mycobacteacuteries drsquoeacutecosystegravemes
aquatiques (Thomas et al 2006 Thomas amp McDonnell 2007 Pagnier et al 2008) et de sols pollueacutes
(Wang et al 2006)
Les eacutetudes relatant les interactions entre M bovis et la meacuteso- et macrofaune sont rares Une
eacutetude reacutealiseacutee dans des eacutelevages bovins et caprins infecteacutes par Map a mis en eacutevidence la contamination
de 31 des vers de terre preacuteleveacutes sur les pacirctures freacutequenteacutees par les animaux (Fischer et al 2003)
Dans cette eacutetude Map a eacuteteacute retrouveacutee dans le corps et les fegraveces de vers de terre 1 et 2 jours apregraves
lrsquoingestion de bouses infecteacutees expeacuterimentalement suggeacuterant qursquoils peuvent probablement jouer un
rocircle de vecteurs dans la disseacutemination des mycobacteacuteries
Les eacutetudes disponibles bien que tregraves rares suggegraverent que la meacuteso- et macrofaune du sol
peuvent heacuteberger et probablement jouer un rocircle de vecteur des mycobacteacuteries Toutefois des eacutetudes
compleacutementaires sont indispensables concernant M bovis
d Interactions des mycobacteacuteries avec les diptegraveres
Le portage digestif et feacutecal de M bovis par des mouches communes (Musca domestica) en
contact avec des viscegraveres infecteacutes de bovins est deacutecrit La bacteacuterie survit de 12 jours sur leurs corps et
15 jours dans le tractus digestif de mouches mortes (Schlee 1957 citeacute par Kazda et al 2009)
e La formation de biofilms
Lrsquoaptitude agrave former des biofilms est connue pour de nombreuses espegraveces de mycobacteacuteries
Chapitre 1
58
environnementales agrave croissance rapide comme M smegmatis M fortuitum ou M abcessus ou agrave
croissance lente notamment M avium et M intracellulare (Falkinham et al 2001 Beumer et al 2010
Torvinen et al 2007) Lrsquoorganisation des mycobacteacuteries en biofilm leur confegravere une reacutesistance accrue
vis-agrave-vis des conditions deacutefavorables dans lrsquoenvironnement mais eacutegalement chez lrsquohocircte humain ougrave
elles peuvent coloniser les catheacuteters et provoquer des infections sanguines tregraves graves (El Helou et al
2013) Concernant les mycobacteacuteries du MTBC la formation de biofilms est deacutecrite pour
M tuberculosis dans des conditions expeacuterimentales tregraves particuliegraveres tregraves eacuteloigneacutees des conditions
naturelles (Kulka et al 2012) La formation drsquoagreacutegats (clumping ou cording) observeacutes lors de la culture
des bacteacuteries du MTBC en milieu liquide gracircce agrave une matrice extracellulaire riche en acides mycoliques
peut ecirctre assimileacutee agrave une forme organiseacutee de reacutesistance des bacteacuteries puisque sous cette forme les
bacteacuteries survivent agrave des concentrations tregraves eacuteleveacutees en antibiotique (Ojha et al 2008) Il a eacuteteacute montreacute
chez M tuberculosis que la formation de biofilms mettait en jeu des processus geacuteneacutetiques et
physiologiques diffeacuterents de la croissance planctonique (Ojha et al 2008) Toutefois comme il nrsquoa
jamais eacuteteacute deacutemontreacute que ces bacteacuteries se multiplient dans lrsquoenvironnement il semble peu probable
que les biofilms constituent une forme de reacutesistance pour M bovis
5 Facteurs de risque de contamination de lrsquoenvironnement et de transmission de Mycobacterium bovis aux animaux par lrsquoenvironnement contamineacute
Plusieurs eacutetudes portant sur les interactions entre bovins et faune sauvage ont montreacute que les
contacts directs eacutetaient tregraves peu freacutequents entre les bovins et les blaireaux (Benham amp Broom 1989
Drewe et al 2013 Boumlhm et al 2009) ainsi qursquoentre les cerfs les sangliers et les bovins (Cooper et al
2010 Kukielka et al 2013) La transmission indirecte de M bovis semble donc ecirctre le mode de
transmission preacutepondeacuterant entre la faune sauvage et les bovins voire entre les diffeacuterentes espegraveces
sauvages (Cowie et al 2015 Carrasco-Garcia et al 2015) Diffeacuterents facteurs de risque jouent un rocircle
sur ce mode de transmission qui neacutecessite un recouvrement des domaines vitaux des animaux
sauvages infecteacutes et excreacuteteurs avec les sites drsquoeacutelevage bovin (pacirctures et bacirctiments) Ces principaux
facteurs de risque sont deacutetailleacutes ci-dessous
a Risques lieacutes agrave lrsquoeacutecologie des espegraveces sauvages sensibles agrave Mycobacterium bovis
Concernant le blaireau et le sanglier les pacirctures peuvent constituer une grande partie de leur
domaine vital (Hutchings amp Harris 1999 Payne 2014) Elles sont utiliseacutees pour la recherche de
Chapitre 1
59
nourriture essentiellement des lombrics qui repreacutesentent une part importante de leur reacutegime
alimentaire (Tolhurst et al 2009 Cleary et al 2011 Baubet et al 2003) Une eacutetude reacutealiseacutee en Cocircte
drsquoOr a montreacute que la freacutequentation des pacirctures par les blaireaux est optimale au printemps peacuteriode
de mise au preacute des bovins par rapport agrave lrsquoautomne et agrave lrsquohiver Pour les sangliers la freacutequentation des
pacirctures est plus importante au printemps et en eacuteteacute (Payne 2014)
Concernant plus preacuteciseacutement les blaireaux ces derniers urinent et deacutefegravequent dans des latrines
pour deacutelimiter leurs territoires (Hutchings et al 2001) mais ils urinent eacutegalement sur les pacirctures
preacutefeacuterentiellement au niveau des lisiegraveres de forecirct des clocirctures et des haies qursquoils croisent au greacute de
leurs deacuteplacements dans un but de marquage (White et al 1993 Drewe et al 2013) Dans les pacirctures
il a eacuteteacute montreacute que le taux de contact entre bovins et latrines est eacuteleveacute (Hutchings amp Harris 1997 Drewe
et al 2013) Dans lrsquoeacutetude de Hutchings et Harris (1997) 48 des 50 vaches du troupeau ont cumuleacute 416
contacts avec 4 latrines preacutesentes dans la pacircture eacutetudieacutee et 33 animaux ont brouteacute sur les latrines (suivi
sur 1776 heures) Pour Drewe et al (2013) 1716 contacts avec 13 latrines ont eacuteteacute enregistreacutes pour les
33 bovins drsquoun troupeau sur les cinq mois de lrsquoeacutetude Toutefois le rang hieacuterarchique des bovins du
troupeau joue un rocircle important puisque les animaux domineacutes sont les plus enclins agrave brouter au
voisinage des latrines Enfin les bovins consomment lrsquoherbe au voisinage des latrines de faccedilon plus
importante lorsque lrsquoherbe de la pacircture est rase (en cas de surpacircturage ou de fauchage) (Hutchings amp
Harris 1997) Compte tenu du grand nombre de bacilles qui peuvent ecirctre excreacuteteacutes par la voie urinaire
du fort taux de contact bovinslatrines et de la consommation de lrsquoherbe sur les latrines par certains
bovins il semblerait que les latrines et plus particuliegraverement les zones de marquage urinaire dans les
pacirctures repreacutesentent un risque important de transmission indirecte de la maladie par inhalation ou
ingestion (Hutchings amp Harris 1999)
Bien qursquoil nrsquoexiste aucune eacutetude sur le sujet la contamination des pacirctures par des blaireaux
preacutesentant des eacutecoulements purulents de morsures a eacutegalement eacuteteacute envisageacutee (Gallagher amp Clifton-
Hadley 2000)
Les blaireaux vivent dans des terriers qursquoils creusent dans le sol en milieu forestier ou en
dehors notamment dans les pacirctures La contamination des sols drsquoentreacutee de terrier par M bovis
repreacutesente eacutegalement un risque pour les bovins qui peuvent se contaminer en explorant les entreacutees de
terrier localiseacutees dans les pacirctures par ingestion ou inhalation de la litiegravere expulseacutee des tunnels
(Benham 1985) De plus le sol excaveacute des tunnels par les blaireaux est attirant pour les bovins qui
peuvent en ingeacuterer pour combler des carences mineacuterales (Dewes 1996) ou srsquoy frotter la tecircte creacuteant
des aeacuterosols de poussiegravere potentiellement infectant (Phillips et al 2003) La contamination des sols de
terriers repreacutesente eacutegalement un risque important de transmission de lrsquoinfection dans le groupe social
de blaireaux qui partage le terrier Le sol du terrier constitue un biotope adeacutequat pour la survie de
M bovis agrave lrsquoabri des UV sans variation extrecircme de tempeacuterature et avec une humiditeacute relative
Chapitre 1
60
importante (Moore amp Roper 2003) Lrsquoentretien de la maladie dans un groupe social par le terrier
contamineacute a deacutejagrave eacuteteacute eacutevoqueacute (Rogers et al 2003)
La freacutequentation des pacirctures par les sangliers pour la recherche de nourriture constitue
eacutegalement un facteur de risque dans la transmission indirecte de la bacteacuterie aux bovins Lrsquoexploration
du sol avec leurs museaux agrave la recherche de lombrics larves et autre nourriture ainsi que les reliquats
de sol deacuteplaceacutes peuvent constituer un facteur de contamination potentiel de lrsquoenvironnement par la
deacuteposition drsquoexcreacutetas oro-nasaux infecteacutes
Les cervideacutes peuvent eacutegalement utiliser les preacutes pour se nourrir et ecirctre une source de
contamination indirecte pour les bovins (Brook amp McLachlan 2009)
b Risques lieacutes aux pratiques drsquoeacutelevage
Le seacutejour prolongeacute des bovins au preacute est susceptible drsquoaugmenter les contacts directs et
indirects avec la faune sauvage infecteacutee Lrsquoeacutelevage allaitant est caracteacuteriseacute par une longue peacuteriode de
pacircturage entre avril et octobre puis une hivernation en bacirctiments au moment des vecirclages Certaines
races rustiques notamment la race salers qui ont repeupleacute une partie des cheptels bovins infecteacutes
apregraves abattage total passent toute lrsquoanneacutee au pacircturage Ces longues peacuteriodes de pacircturage constituent
un facteur de risque non neacutegligeable pour les bovins qui partagent leur habitat avec des espegraveces
sauvages infecteacutees Le surpacircturage est eacutegalement un facteur de risque pour les bovins qui sont ameneacutes
agrave brouter les latrines de blaireaux et agrave ingeacuterer des quantiteacutes de sol potentiellement contamineacute plus
importantes (Hutchings amp Harris 1999)
Les zones drsquoabreuvement et de nourrissage des bovins dans la pacircture constituent des zones
attractives pour la faune sauvage notamment lrsquohiver lorsque les ressources alimentaires font deacutefaut
Le partage de fourrage entre bovins et cervideacutes est probablement une source de contamination des
bovins dans le Michigan (OrsquoBrien et al 2006 Brook amp McLachlan 2009 Brook et al 2012) drsquoautant plus
que la transmission indirecte de la maladie entre des cerfs et des veaux par le partage du foin a eacuteteacute
prouveacutee expeacuterimentalement (Palmer et al 2010 Palmer et al 2004) La freacutequentation des auges de
bovins et des zones de nourrissage au sol par les blaireaux qui viennent manger les reliquats
alimentaires est reacuteguliegraverement deacutecrite au Royaume-Uni ainsi que celle des reacuteserves de grains
drsquoensilage et de foin dans lesquels ils peuvent uriner et deacutefeacutequer (Garnett et al 2002 Tolhurst et al
2009 Ward et al 2010) Les pierres agrave sel souvent poseacutees au sol dans les preacutes attirent particuliegraverement
les cervideacutes (Payne et al 2015 Brook amp McLachlan 2009) Les abreuvoirs et les zones drsquoabreuvement
(trous drsquoeau ruisseau source) constituent eacutegalement un facteur de risque lorsqursquoils sont utiliseacutes par
les bovins et la faune sauvage la survie de la bacteacuterie excreacuteteacutee eacutetant optimale dans les zones humides
Le comportement de baignade des sangliers dans les trous drsquoeau et les souilles entraine une forte
Chapitre 1
61
freacutequentation de ces zones notamment en eacuteteacute (Payne 2014)
Finalement lrsquoeacutepandage de fumiers frais augmenterait le risque drsquoinfection drsquoun troupeau par
rapport agrave lrsquoeacutepandage drsquoengrais ou de fumiers non frais (stockeacutes au moins deux mois) les animaux se
contaminant par inhalation ou ingestion de substrats contamineacutes dans les deux mois suivant lrsquoeacutepandage
(Griffin et al 1996 Humblet et al 2009) Certaines preacuteconisations conseillent un temps de stockage
minimum des fumiers avant eacutepandage (au moins 6 mois suite aux reacutesultats de Scanlon et Quinn 2000a
et 2000b) ou leur traitement chimique (Scanlon amp Quinn 2000b Scanlon amp Quinn 2000a) un
compostage (McCallan et al 2014) voire lrsquointerdiction drsquoeacutepandage de fumiers pour les cheptels infecteacutes
(Phillips et al 2003)
c Risques lieacutes aux pratiques cyneacutegeacutetiques
Les zones drsquoagreacutegation des animaux sauvages telles que les zones drsquoagrainage ou de
nourrissage agrave points fixes et la reacutealisation de trous drsquoeau artificiels pour lrsquoabreuvement et la baignade
des animaux sauvages sont des zones agrave risque qui augmentent les interactions directes et indirectes
entre les animaux et donc les risques de transmission de M bovis entre eux (Corner 2006 Vicente et
al 2007 Delahay et al 2002 Coleman amp Cooke 2001) En Espagne lrsquoaugmentation des densiteacutes
drsquoonguleacutes sauvages (cervideacutes et sangliers) dans le Sud du pays a entraineacute le deacuteveloppement drsquoactiviteacutes
de chasse agrave but commercial amenant agrave la reacutealisation de parcs gigantesques pour contenir les animaux
sauvages Dans ces zones lrsquoagreacutegation des animaux sauvages de diffeacuterentes espegraveces (sangliers
cervideacutes cochons sauvages et bovins) autour des trous drsquoeau et des zones de nourrissage a favoriseacute la
transmission intra- et interspeacutecifique de la maladie (Vicente et al 2007 Cowie et al 2015) Chez le
sanglier la preacutevalence varie de 52 agrave 70 voire presque 100 dans certaines populations (Gortaacutezar et
al 2008 Naranjo et al 2008) Cowie et al (2015) ont montreacute que 40 agrave 66 des interactions indirectes
avaient lieu dans une fenecirctre temporelle de 3 jours en saison segraveche et 72 agrave 100 dans une fenecirctre
temporelle de 12 jours en saison humide Or dans ces zones la survie de M bovis a eacuteteacute estimeacutee agrave 3
jours pendant la saison segraveche et agrave 12 jours pendant la saison humide (Kukielka et al 2013) suggeacuterant
que si le pathogegravene est preacutesent dans une population lrsquoagreacutegation est un facteur de maintenance et de
disseacutemination de lrsquoinfection dans et entre les espegraveces De plus la preacutesence drsquoeacutelevages bovins agrave
proximiteacute de ces parcs de chasse entraine une freacutequentation importante des structures drsquoeacutelevage par
les animaux sauvages et un risque accru drsquoinfection pour les bovins des eacutelevages visiteacutes (Carrasco-
Garcia et al 2015)
Le nourrissage des cervideacutes dans le Michigan dans le but de maintenir les populations pour la
chasse est la cause du maintien de lrsquoinfection dans les populations sauvages qui se contaminent entre
eux via les aliments distribueacutes (OrsquoBrien et al 2006)
Chapitre 1
62
Comme nous lrsquoavons deacutejagrave dit preacuteceacutedemment les viscegraveres drsquoanimaux infecteacutes laisseacutes agrave la
disposition des animaux sauvages par les chasseurs peuvent repreacutesenter un risque important pour les
animaux charognards (Morris et al 1994)
d Risques lieacutes aux facteurs paysagers climatiques et peacutedologiques
La structure du paysage est un facteur qui influence le recouvrement des domaines vitaux des
animaux sauvages et des bovins La localisation des pacirctures ou des bacirctiments drsquoeacutelevage en lisiegravere de
forecirct augmente les risques drsquointeractions entre animaux sauvages et domestiques (Ward et al 2010)
lrsquointerface pacircture-forecirct repreacutesenteacutee par les lisiegraveres eacutetant un eacutecotone agrave risque pour lrsquoeacutemergence des
agents pathogegravenes multi-hocirctes (Boyard et al 2008) La fragmentation du paysage (preacutes champs forecircts
bosquets) augmente le deacutepocirct drsquourine par les blaireaux aux zones frontiegraveres entre deux milieux
augmentant le risque de contamination de ces sites (White et al 1993 Humblet et al 2009) Les lisiegraveres
sont tregraves freacutequenteacutees par les sangliers et les blaireaux (Payne 2014) et constituent une zone privileacutegieacutee
par les blaireaux pour eacutetablir leurs terriers et marquer leurs territoires par des mictions ou des latrines
(Hutchings et al 2002)
Comme nous lrsquoavons vu preacuteceacutedemment les conditions climatiques sont un facteur essentiel
dans la dureacutee de survie de M bovis dans les diffeacuterents substrats environnementaux contamineacutes et in
fine dans la transmission indirecte de la maladie aux animaux qui interagissent avec ces substrats Les
saisons influencent eacutegalement la freacutequentation des pacirctures par les animaux sauvages avec un risque
accru au printemps et en eacuteteacute vis-agrave-vis des blaireaux et des sangliers qui vermillent dans les pacirctures
notamment lors des nuits humides ougrave les vers de terre remontent en surface appeleacutees laquo nuits agrave
lombrics raquo (Kruuk 1978 Cleary et al 2011 Baubet et al 2003) Lors de temps plus sec voire de
seacutecheresse les points drsquoeau destineacutes aux bovins constituent des zones attractives pour la faune sauvage
(Vicente et al 2007 Santos Santos et al 2015 Barasona et al 2014) favorisant lrsquoagreacutegation des
animaux domestiques et sauvages
Le rocircle des facteurs peacutedologiques dans la survie de M bovis est peu connu agrave ce jour La texture
du sol qui conditionne la structure la porositeacute et le reacutegime hydrique du sol le pH et la matiegravere
organique disponible sont des facteurs importants dans la survie des bacteacuteries A notre connaissance
une seule eacutetude statistique reacutealiseacutee dans le Michigan a montreacute que la nature du sol et plus
preacuteciseacutement le pourcentage de sable eacutetait associeacute agrave un risque drsquoinfection plus eacuteleveacute chez les bovins
(Walter et al 2014) Une augmentation drsquo1 de la quantiteacute de sable dans le sol augmentait de 36
le risque pour des eacutelevages bovins drsquoecirctre infecteacutes Hormis cette eacutetude qui relie indirectement infection
animale et survie de la bacteacuterie dans le sol aucune eacutetude nrsquoest actuellement disponible sur le lien
direct entre nature du sol et survie de M bovis
Chapitre 1
63
A la lumiegravere des informations preacutesenteacutees dans ce chapitre il apparait qursquoune fois excreacuteteacutee en
dehors de son hocircte M bovis peut survivre longuement dans lrsquoenvironnement si les conditions sont
adeacutequates par la mise en jeu de strateacutegies de survie encore inconnues agrave lrsquoheure actuelle Les
connaissances sur les scheacutemas de circulation de la bacteacuterie entre les animaux infecteacutes et
lrsquoenvironnement (et vice et versa) srsquoameacuteliorent et permettent drsquoadapter les programmes de lutte contre
la maladie mais elles sont encore insuffisantes pour permettre la maitrise de tous les facteurs en jeu
Ces diffeacuterents facteurs sont scheacutematiseacutes dans la Figure 15
Figure 15 Repreacutesentation scheacutematique de la circulation de M bovis dans lrsquoenvironnement agricole (modifieacute drsquoapregraves Elliott et al 2014)
Chapitre 1
64
D PRINCIPAUX OUTILS DE DETECTION DE PHENOTYPAGE ET DE GENOTYPAGE APPLICABLES AU MTBC ET PLUS PARTICULIEREMENT A MYCOBACTERIUM BOVIS
Depuis la deacutecouverte du bacille de la tuberculose de nombreux outils ont eacuteteacute deacuteveloppeacutes pour
deacutetecter et discriminer les diffeacuterentes mycobacteacuteries du MTBC Ces meacutethodes microscopiques
pheacutenotypiques et biochimiques essentiellement deacuteveloppeacutees pour le diagnostic meacutedical ont eacuteteacute peu
agrave peu ameacutelioreacutees et associeacutees agrave de nouvelles techniques de deacutetection de caracteacuterisation et
drsquoidentification des bacilles telles que lrsquoamplification geacutenique et le seacutequenccedilage notamment ameacuteliorant
la rapiditeacute du diagnostic la discrimination des espegraveces la pertinence de lrsquoantibiotheacuterapie et permettant
le geacutenotypage des souches agrave des fins eacutepideacutemiologiques Adapteacutees agrave des eacutechantillons biologiques
divers tels que les crachats les liquides de lavage broncho-alveacuteolaire les tissus de biopsies chez
lrsquohomme ou les broyats de nœuds lymphatiques chez les bovins et les animaux sauvages ces outils ne
sont pas tous applicables aux eacutechantillons environnementaux heacutebergeant des communauteacutes
microbiennes tregraves complexes
1 La deacutetection des mycobacteacuteries par microscopie
La microscopie optique
Lrsquoexamen microscopique de frottis ou drsquoeacutetalement drsquoeacutechantillons biologiques est la premiegravere
eacutetape dans le diagnostic bacteacuteriologique de la tuberculose humaine Elle est parfois la seule meacutethode
de diagnostic dans les pays en voie de deacuteveloppement Les proprieacuteteacutes tinctoriales particuliegraveres de la
mycomembrane des mycobacteacuteries sont exploiteacutees dans les colorations utiliseacutees La coloration de
reacutefeacuterence est la coloration de Ziehl et Neelsen Cette coloration agrave chaud est baseacutee sur la reacutesistance de
la paroi des mycobacteacuteries agrave la deacutecoloration agrave lrsquoacide et lrsquoalcool apregraves traitement agrave la fuschine Les
mycobacteacuteries apparaissent comme des bacirctonnets rouges sur un fond bleu Des meacutethodes agrave froid
baseacutees sur les mecircmes reacuteactifs existent eacutegalement (meacutethode de Kinyoun) La coloration agrave lrsquoauramine
un fluorochrome permet drsquoobserver en microscopie agrave fluorescence les bacilles en vert-jaune brillant
sur un fond jaune-orangeacute Cette coloration est plus sensible que la coloration de ZN (Steingart et al
2006)
Ces meacutethodes de coloration ne permettent pas de discriminer les mycobacteacuteries entre elles et
ne sont pas speacutecifiques du genre Mycobacterium puisqursquoelles peuvent colorer drsquoautres actinobacteacuteries
comme les genres bacteacuteriens Nocardia Rhodococcus et Corynebacterium (Shinnick amp Good 1994)
Lrsquoutilisation de la microscopie optique nrsquoest pas deacutecrite pour lrsquoanalyse des eacutechantillons
environnementaux drsquoune part car le seuil de deacutetection est eacuteleveacute (de lrsquoordre de 104 BAARmL) et drsquoautre
Chapitre 1
65
part car la coloration est difficile agrave mettre en œuvre sur des matrices telles que du sol ou des matiegraveres
feacutecales
Lrsquohybridation avec des sondes nucleacuteotidiques FISH (Fluorescence in situ hybridization)
Cette meacutethode est baseacutee sur lrsquohybridation speacutecifique drsquoune sonde marqueacutee avec un
fluorochrome sur une reacutegion cible de lrsquoADN bacteacuterien Les sondes utiliseacutees ADN ARNm ou
oligonucleacuteotides syntheacutetiques (PNA peptide nucleic acid) drsquoenviron 20 bases peacutenegravetrent agrave lrsquointeacuterieur
des cellules preacutealablement permeacuteabiliseacutees ougrave elles srsquohybrident speacutecifiquement avec leur ADN cible
Lrsquoobservation se fait agrave lrsquoaide drsquoun microscope agrave eacutepifluorescence
La meacutethode FISH a surtout eacuteteacute deacuteveloppeacutee pour la deacutetection des mycobacteacuteries dans les
crachats dans des cultures liquides chez des animaux infecteacutes expeacuterimentalement (Hongmanee et al
2001 Stender Mollerup et al 1999 Stender Lund et al 1999) et dans des eacutechantillons biologiques
(Stender Mollerup et al 1999 Lefmann et al 2006 Rodriguez-Nuntildeez et al 2012) La meacutethode FISH
est peu utiliseacutee pour la recherche de mycobacteacuteries dans des eacutechantillons environnementaux mais elle
a permis de mettre en eacutevidence Maa et Map dans de lrsquoeau et des biofilms de canalisations (Lehtola et
al 2006 Torvinen et al 2007) Elle parait toutefois difficilement applicable agrave des matrices telles que le
sol agrave cause de lrsquoauto-fluorescence geacuteneacutereacutee par les particules de sol et du risque de non-speacutecificiteacute des
sondes
2 Les meacutethodes de culture des mycobacteacuteries
Le diagnostic de la tuberculose est indissociable de la culture car historiquement crsquoest la
meacutethode la plus utiliseacutee Elle est sensible et permet drsquoisoler les souches de les caracteacuteriser et de tester
leur sensibiliteacute aux antibiotiques La recherche de mycobacteacuteries par des meacutethodes baseacutees sur la
culture est tregraves standardiseacutee en diagnostic humain et veacuteteacuterinaire tout comme les traitements
preacuteliminaires indispensables A lrsquoopposeacute aucune meacutethode standardiseacutee nrsquoest disponible pour la
recherche de mycobacteacuteries dans lrsquoenvironnement De nombreux protocoles emprunteacutes agrave la
microbiologie clinique ont eacuteteacute adapteacutes pour la recherche de mycobacteacuteries environnementales
notamment dans lrsquoeau (Narang et al 2009 Radomski et al 2011 Norby et al 2007 Pickup et al 2006
Whiley et al 2012 Le Dantec et al 2002 Bland et al 2005) Les meacutethodes utiliseacutees pour cultiver les
mycobacteacuteries pathogegravenes sont moins nombreuses et sont deacutecrites ci-dessous
a Les diffeacuterents traitements preacutealables agrave la mise en culture des eacutechantillons
De nombreuses meacutethodes de deacutecontamination des eacutechantillons biologiques ont eacuteteacute deacutecrites
dans le diagnostic meacutedical de la tuberculose Le choix de la meacutethode agrave utiliser deacutepend essentiellement
Chapitre 1
66
du type drsquoeacutechantillon analyseacute (tissu crachat lait fegraveces eau solhellip) des milieux de culture employeacutes et
de sa compatibiliteacute avec les meacutethodes moleacuteculaires Les meacutethodes les plus utiliseacutees en microbiologie
humaine meacutethodes de Petroff Taquet-Tison et Kubica utilisent la soude comme agent deacutecontaminant
principal (Petroff 1915 Taquet amp Tison 1961 Kubica et al 1963) Lrsquoutilisation drsquoacides comme agent
deacutecontaminant tels que lrsquoacide oxalique (David et al 1989) ou lrsquoacide sulfurique (Hohn amp Lowenstein
1926) ou drsquoammoniums quaternaires comme le chlorure de ceacutetyl pyridinium (Smithwick et al 1975)
ou le chlorure de benzalkonium (Krasnow amp Wayne 1966) est reacuteserveacutee aux eacutechantillons tregraves contamineacutes
(urines fegraveces) ou aux biopsies (cutaneacutees coliques pulmonaires etc) Ces meacutethodes ne sont pas toutes
compatibles avec la culture en milieu liquide et lrsquoamplification geacutenique
Dans le domaine veacuteteacuterinaire les tissus animaux (broyats de ganglions ou de tissus) sont
deacutecontamineacutes selon les preacuteconisations de lrsquoANSES et suivent la norme AFNOR NF U47-104 baseacutee sur
lrsquoutilisation de lrsquoacide sulfurique et de la soude agrave 4 Malgreacute la reacutesistance des mycobacteacuteries aux agents
chimiques la deacutecontamination reacuteduit fortement la population initiale preacutesente dans lrsquoeacutechantillon et
abaisse la sensibiliteacute de la culture (Fine Bolin et al 2011)
Concernant lrsquoisolement de bacteacuteries du MTBC dans les matrices environnementales ou les
excreacutetas animaux les principales meacutethodes utiliseacutees deacutecoulent des meacutethodes citeacutees preacuteceacutedemment
La meacutethode drsquoimmunocapture magneacutetique utiliseacutee par Sweeney (2006 et 2007) a permis lrsquoisolement
de M bovis dans des sols de terriers et des latrines contamineacutes naturellement Cette meacutethode est
baseacutee sur lrsquoutilisation de microbilles magneacutetiques recouvertes avec un ligand speacutecifique drsquoun antigegravene
de lrsquoenveloppe de M bovis Son avantage est qursquoelle permet la capture et la concentration de la bacteacuterie
cible dans un eacutechantillon complexe sans alteacuterer sa viabiliteacute contrairement agrave la deacutecontamination
chimique Cette meacutethode a eacutegalement eacuteteacute testeacutee avec succegraves sur des broyats de nœuds lymphatiques
de bovins (Stewart et al 2012 Stewart et al 2013 Garbaccio amp Cataldi 2010)
b La culture sur milieu solide
Les milieux de culture solides agrave lrsquoœuf (milieux de Loumlwenstein-Jensen et milieu de Coletsos) sont
les plus utiliseacutes en France dans le diagnostic bacteacuteriologique chez lrsquohomme et lrsquoanimal Ils sont sensibles
et reacutepondent aux exigences culturales de la plupart des mycobacteacuteries tuberculeuses et non
tuberculeuses Le milieu de Coletsos est particuliegraverement indiqueacute pour la culture des mycobacteacuteries
exigeantes telles que M bovis et M africanum
Les milieux geacuteloseacutes Middlebrook 7H10 et 7H11 sont des milieux semi-syntheacutetiques non utiliseacutes
en diagnostic de routine en France mais tregraves utiliseacutes aux Etats-Unis Enrichis avec de lrsquoOADC meacutelange
drsquoacide oleacuteique drsquoalbumine bovine de dextrose de chlorure de sodium et de catalase ils permettent
une croissance plus rapide de M tuberculosis (Mejia et al 1999) et de M bovis (Idigoras et al 1995)
Chapitre 1
67
que les milieux agrave base drsquoœuf Lrsquoajout de seacuterum bovin et de sang de mouton lyseacute au milieu 7H11 enrichi
en OADC ameacuteliore fortement la croissance de M bovis agrave partir drsquoeacutechantillons animaux et raccourcit le
deacutelai drsquoobservation des colonies (Gallagher amp Horwill 1977) Ce milieu a eacuteteacute utiliseacute pour la mise en
culture drsquoeacutechantillons environnementaux tels que le sol ou les fegraveces apregraves un traitement preacutealable de
lrsquoeacutechantillon (Sweeney et al 2006 Fine OrsquoBrien et al 2011) mais son manque de seacutelectiviteacute neacutecessite
lrsquoajout drsquoun cocktail drsquoantibiotiques souvent composeacute de lrsquoassociation drsquoune carboxypeacutenicilline de
trimeacutethoprime et de polymyxine B et drsquoun antifongique (amphoteacutericine B) (Sweeney et al 2007 Fine
OrsquoBrien et al 2011) Lrsquoincubation des milieux solides ensemenceacutes est reacutealiseacutee agrave 37degC pendant au moins
8 semaines
c La culture en milieu liquide
Les milieux liquides ont eacuteteacute deacuteveloppeacutes pour pallier la croissance lente des mycobacteacuteries
drsquointeacuterecirct clinique sur les milieux solides afin de confirmer preacutecocement le diagnostic Diffeacuterents
systegravemes manuels ou automatiseacutes existent agrave lrsquoheure actuelle Ils sont essentiellement utiliseacutes dans le
diagnostic chez lrsquohomme plus rarement chez lrsquoanimal
Le systegraveme Bactectrade 460 (Becton Dickinson) baseacute sur la respiromeacutetrie radiomeacutetrique nrsquoest plus
utiliseacute en France mais il lrsquoa eacuteteacute pendant une vingtaine drsquoanneacutees Il eacutetait baseacute sur la mesure du dioxyde
de carbone marqueacute au 14C rejeteacute par les mycobacteacuteries en croissance dans un milieu contenant de
lrsquoacide palmitique marqueacute au 14C La libeacuteration de 14CO2 eacutetait mesureacutee reacuteguliegraverement de faccedilon manuelle
gracircce agrave un appareil adapteacute La contrainte drsquoutilisation de stockage et de recyclage de produits
radioactifs ainsi que lrsquoabsence drsquoautomatisation ont conduit agrave lrsquoarrecirct de sa commercialisation Son
utilisation a eacuteteacute deacutecrite pour la deacutetection de M bovis sur des tissus animaux et des fegraveces (Corner et al
2012 Thacker et al 2015)
Pour eacuteviter les inconveacutenients lieacutes agrave la radioactiviteacute des meacutethodes non radiomeacutetriques ont eacuteteacute
deacuteveloppeacutees La croissance bacteacuterienne en milieu liquide est mise en eacutevidence soit par la
consommation de lrsquooxygegravene soit par le deacutegagement de CO2 dans le tube La mise en eacutevidence de la
diminution de la pression partielle drsquooxygegravene dissous dans le milieu de culture lors de la croissance
bacteacuterienne est utiliseacutee dans les tubes BBLtrade MGITtrade de Becton Dickinson Lorsque lrsquoO2 diminue les sels
de rutheacutenium preacutesents au fond du tube eacutemettent une fluorescence Lrsquoutilisation de systegravemes
automatiseacutes tels que les systegravemes Bactectrade MGITtrade assurent une lecture en continue de la fluorescence
eacutemise Le systegraveme Bactectrade MGITtrade a eacuteteacute utiliseacutee pour la recherche de M bovis dans des broyats de
nœuds lymphatiques de bovins (Hines et al 2006 Stewart et al 2013) des tissus drsquoanimaux sauvages
(Murphy et al 2010 Witmer et al 2010) et dans du fromage frais (Harris et al 2007) Lrsquoaugmentation
du CO2 dans le milieu de culture lors de la croissance bacteacuterienne est agrave la base du systegraveme BacTALERT
Chapitre 1
68
3D de Biomeacuterieux Cette meacutethode colorimeacutetrique est baseacutee sur la diminution du pH suite agrave la libeacuteration
du CO2 Lrsquoautomate eacutevalue en continu le virage de couleur drsquoune pastille fixeacutee au fond du tube Les
systegravemes deacutecrits permettent eacutegalement la recherche de reacutesistance agrave diffeacuterents antibiotiques
(Piersimoni et al 2006) A notre connaissance cette technique nrsquoa jamais eacuteteacute utiliseacutee pour la culture
drsquoeacutechantillons environnementaux
Ces diffeacuterents systegravemes utilisent comme milieu liquide le Middlebrook 7H9 suppleacutementeacute en
OADC ou en ADC (Albumine bovine dextrose catalase) pour ameacuteliorer la croissance des mycobacteacuteries
et rendu plus seacutelectif par lrsquoajout drsquoun meacutelange drsquoantibiotiques et drsquoantifongiques pour eacuteviter la
croissance de bacteacuteries indeacutesirables Le plus utiliseacute est le PANTAtrade (polymyxine B amphoteacutericine acide
nalidixique trimeacutethoprime azlocilline) Il peut ecirctre compleacuteteacute avec de la vancomycine (effet contre les
bacteacuteries gram +) Lrsquoavantage majeur de ces systegravemes en milieu liquide est la diminution du deacutelai moyen
de deacutetection des mycobacteacuteries Avec le systegraveme Bactectrade M tuberculosis est deacutetecteacutee entre 8 et 30
jours en fonction de la charge bacteacuterienne initiale de lrsquoeacutechantillon La deacutetection de M bovis est reacutealiseacutee
en 15 jours contre environ 45 jours sur milieu solide Lorsque la culture est deacuteclareacutee positive la
preacutesence de BAAR doit ecirctre veacuterifieacutee par coloration sub-culture sur milieu de Loumlwenstein-Jensen
identification biochimique ou moleacuteculaire Lrsquoassociation de la culture en milieu liquide et solide donne
les meilleurs reacutesultats avec une limite de deacutetection de 10 agrave 102 bacillesmL drsquoeacutechantillon (Guillet-Caruba
et al 2014)
NB A la fin du 19egraveme siegravecle et au deacutebut du 20egraveme la discrimination des espegraveces M bovis M tuberculosis
et M avium se faisait principalement par inoculation de lrsquoisolat bacteacuterien au cobaye au lapin et au
poulet Des leacutesions pulmonaires heacutepatiques et spleacuteniques importantes chez le cobaye et le lapin
identifiaient M bovis des leacutesions retrouveacutees uniquement chez le cobaye ou le poulet identifiaient
respectivement M tuberculosis et M avium (Kazda et al 2009) Cette meacutethode eacutetait utiliseacutee pour
lrsquoidentification de M bovis dans les tissus animaux et les eacutechantillons environnementaux comme le sol
et lrsquoeau apregraves deacutecontamination avec un meacutelange de javel et de soude (Maddock 1933 Maddock 1934
Maddock 1936 Williams amp Hoy 1930)
3 Meacutethodes de deacutetection et drsquoidentification des bacteacuteries du MTBC
Pour des raisons eacutepideacutemiologiques sanitaires (isolement du malade pour eacuteviter la
transmission) et theacuterapeutiques (mise en place drsquoune antibiotheacuterapie adeacutequate) il est primordial de
diffeacuterencier les membres du MTBC des autres mycobacteacuteries et drsquoidentifier preacuteciseacutement lrsquoespegravece du
MTBC en cause
Chapitre 1
69
a Caracteacuterisation pheacutenotypique et biochimique des mycobacteacuteries du MTBC
Depuis la deacutecouverte du bacille de la tuberculose jusqursquoau deacuteveloppement des outils
modernes les diffeacuterentes espegraveces de mycobacteacuteries ont eacuteteacute identifieacutees gracircce agrave leurs caracteacuteristiques
pheacutenotypiques et biochimiques Certaines de ces caracteacuteristiques sont encore exploiteacutees agrave lrsquoheure
actuelle en compleacutement des meacutethodes de biologie moleacuteculaire plus rapides Lrsquoaspect des colonies sur
un milieu solide leur deacutelai drsquoapparition leur nombre leur aspect rugueux ou lisse et la preacutesence drsquoune
pigmentation ou non permet drsquoorienter lrsquoidentification vers une mycobacteacuterie du MTBC ou une
NTM Dans le MTBC par exemple M tuberculosis apparait en 20 jours sur milieux agrave lrsquoœuf sous forme
de colonies ivoire en chou-fleur drsquoaspect rugueux Les colonies de M bovis apparaissent tardivement
(6 agrave 10 semaines) elles sont non pigmenteacutees petites fines et lisses (Carbonnelle et al 2003) Les
principaux tests biochimiques utiliseacutes pour diffeacuterencier les diffeacuterentes espegraveces du MTBC sont le test agrave
la niacine la reacuteduction des nitrates lrsquohydrolyse du Tween la sensibiliteacute au TCH (acide thiophegravene-2-
carboxylique) agrave la cycloseacuterine et au pyrazinamide Les reacutesultats des observations pheacutenotypiques et
biochimiques permettant de diffeacuterencier les principales espegraveces du MTBC sont preacutesenteacutes dans le
Tableau 2 Les tests biochimiques neacutecessitent des cultures riches et ne sont donc pas reacutealisables avant
la croissance avanceacutee de la bacteacuterie Chronophages et fastidieux ces tests sont remplaceacutes par drsquoautres
techniques plus rapides
Tableau 2 Identification des principales mycobacteacuteries du MTBC par des observations pheacutenotypiques et des tests biochimiques
M tuberculosis M africanum M bovis M bovis BCG
Aspect des colonies Rugueuses Rugueuses Lisses Rugueuses
Pigmentation Non pigmenteacute Non pigmenteacute Non pigmenteacute Non pigmenteacute
Croissance Lente Lente Lente Lente
Niacin test + - - -
Reacuteduction nitrates + +- - -
TCH R S S S
Cycloseacuterine S S R R
Pyrazinamide S S R R
+ test positif - test neacutegatif R reacutesistant S sensible
b Lrsquoidentification des mycobacteacuteries par recherche de biomarqueurs
Diffeacuterentes techniques de recherche de biomarqueurs potentiels de M tuberculosis par
chromatographie ont eacuteteacute deacuteveloppeacutees reacutecemment La chromatographie en phase liquide (HPLC High-
Chapitre 1
70
Performance Liquid Chromatography) a eacuteteacute deacuteveloppeacutee pour lrsquoanalyse des acides mycoliques (Butler amp
Guthertz 2001) La chromatographie en phase gazeuse coupleacutee agrave de la spectromeacutetrie de masse (GC-
MS) est utiliseacutee pour identifier des acides gras speacutecifiques de la paroi de M tuberculosis comme lrsquoacide
tuberculosteacutearique ou lrsquoacide hexacosanoiumlque dans des crachats (Cha et al 2009 Kaal et al 2009)
Lrsquoidentification de biomarqueurs speacutecifiques permet de diffeacuterencier M tuberculosis des NTM en GC-
MS en diagnostic direct sur des crachats (Dang et al 2013)
Depuis quelques anneacutees la spectromeacutetrie de masse de type MALDI-TOF (matrix-assisted laser
desorption ionization-time of flight) est utiliseacutee pour lrsquoidentification rapide des mycobacteacuteries du
complexe tuberculosis et de la majoriteacute des autres mycobacteacuteries agrave partir de cultures en milieu solide
et liquide (El Kheacutechine et al 2011 Bille et al 2012 Lotz et al 2010 Biswas amp Rolain 2013) Cette
technique permet drsquoeacutetablir un spectre de masse des diffeacuterentes proteacuteines constituant la mycobacteacuterie
(profil peptidique obtenu apregraves digestion trypsique des proteacuteines totales) La comparaison du profil
proteacuteique obtenu avec des bases de donneacutees internationales de plus en plus eacutetoffeacutees permet
drsquoidentifier un grand nombre drsquoespegraveces de mycobacteacuteries Elle peut ecirctre eacutegalement utiliseacutee comme
meacutethode de spoligotypage (Shitikov et al 2012) Cette meacutethode est simple rapide reproductible et
neacutecessite des consommables peu coucircteux (hors achat et entretien de lrsquoappareil) La qualiteacute du profil
proteacuteique obtenu est toutefois fortement deacutependante de la qualiteacute du protocole drsquoextraction des
proteacuteines (Saleeb et al 2011) La technique MALDI-TOF est probablement une technique drsquoavenir pour
lrsquoidentification rapide des mycobacteacuteries agrave croissance lente (le deacutelai de croissance reste toutefois le
facteur limitant) et plus geacuteneacuteralement des microorganismes pathogegravenes drsquointeacuterecirct clinique dans les
eacutechantillons A lrsquoheure actuelle elle ne permet pas de diffeacuterencier les diffeacuterentes espegraveces du MTBC
Elle nrsquoest pour lrsquoinstant applicable qursquoagrave des isolats bacteacuteriens issus de culture et non agrave des eacutechantillons
complexes
c Lrsquoidentification des mycobacteacuteries par des meacutethodes drsquohybridation coupleacutees ou
non agrave la PCR
La technique baseacutee sur lrsquohybridation de lrsquoADN est une meacutethode sensible rapide et simple
drsquoemploi couramment utiliseacutee en bacteacuteriologie humaine Elle est reacutealiseacutee sur une culture liquide ou
solide La technique est baseacutee sur lrsquohybridation drsquoune sonde moleacuteculaire conjugueacutee agrave un marqueur
chimioluminescent compleacutementaire de lrsquoARN ribosomal de la bacteacuterie cible Le kit commercial
AccuProbe (Gen Probe) fournit diffeacuterentes sondes permettant drsquoidentifier les Complexes tuberculosis
avium gordonae kansaii (sans discrimination des espegraveces) ainsi que les espegraveces M avium et
M intracellulare
Le systegraveme le plus utiliseacute agrave lrsquoheure actuelle est le GenXpert MTBRIF (Cepheid) utilisant la PCR
Chapitre 1
71
en temps reacuteel multi-colorimeacutetrique Il utilise 5 sondes Beacon (sondes avec une structure en eacutepingle agrave
cheveux qui fluorescent lors de lrsquohybridation) speacutecifiques du locus rpoB (reacutegion de 81 pb deacuteterminant
la reacutesistance agrave la rifampicine) des membres du MTBC Lrsquointeacuterecirct de cette technique est agrave la fois pratique
puisque lrsquoopeacuterateur deacutepose lrsquoeacutechantillon deacutecontamineacute dans une cassette contenant tous les reacuteactifs
lyophiliseacutes lrsquoappareil gegravere toutes les autres eacutetapes et diagnostique car elle couple la deacutetection des
bacteacuteries du MTBC et la mise en eacutevidence drsquoune eacuteventuelle reacutesistance agrave la rifampicine antituberculeux
de premiegravere ligne chez lrsquohomme
Drsquoautres systegravemes tels que lrsquoINNO-Lipareg Mycobacteria v2 (Fujirebio) le GenoType MTBC et le
GenoType Mycobacterium CMAS (Hain Lifescience) sont baseacutes sur lrsquohybridation drsquoune sonde
speacutecifique drsquoun Complexe ou drsquoune espegravece sur la reacutegion intergeacutenique variable 16S-23S apregraves
amplification PCR Ils permettent drsquoidentifier les Complexes tuberculosis et avium ainsi que de 16 agrave 40
espegraveces de NTM
Ces techniques sont essentiellement utiliseacutees dans le diagnostic humain mais elles sont parfois
deacutecrites pour lrsquoidentification du Complexes tuberculosis en diagnostic animal (Oloya et al 2007 Pavlik
et al 2005 Bollo et al 1998 Lieacutebana et al 1995) Bien que deacutejagrave utiliseacutees sur des eacutechantillons
environnementaux comme lrsquoeau (Aronson et al 1999) elles sont agrave utiliser avec preacutecaution car des
erreurs drsquoidentification ont eacuteteacute rapporteacutees (croisement entre des mycobacteacuteries saprophytes et
pathogegravenes) (Turenne 2004) en outre elles ne permettent pas la discrimination des diffeacuterentes
espegraveces du MTBC
d Le seacutequenccedilage
Lrsquooutil geacutenotypique de reacutefeacuterence pour lrsquoidentification des diffeacuterentes espegraveces de bacteacuteries est
le seacutequenccedilage du gegravene codant lrsquoARN de la sous-uniteacute 16S du ribosome (gegravene rrs) De nombreux couples
drsquoamorces ont eacuteteacute proposeacutes dans la litteacuterature pour amplifier entiegraverement ou partiellement le gegravene rrs
avant seacutequenccedilage mais lrsquoidentification des mycobacteacuteries par cette meacutethode nrsquoest pas toujours
possible agrave cause de la forte conservation de cette seacutequence Afin drsquoameacuteliorer la discrimination de ces
espegraveces le seacutequenccedilage total ou partiel drsquoautres gegravenes cibles a eacuteteacute deacuteveloppeacute tels les gegravenes de meacutenage
ciblant rpoB codant la sous-uniteacute β de lrsquoARN polymeacuterase (Adeacutekambi amp Drancourt 2004 Lee et al 2000)
hsp65 codant une proteacuteine de reacuteponse au stress thermique de 65 kDa (McNabb et al 2004 Ringuet et
al 1999 Telenti et al 1993) gyrA et gyrB codant les sous-uniteacutes α et β de lrsquoADN gyrase (enzyme
intervenant dans lrsquoenroulement et le deacuteroulement de lrsquoheacutelice drsquoADN) recA intervenant dans la
recombinaison homologue de lrsquoADN et dans sa reacuteparation (Blackwood et al 2000) sodA codant la super
oxyde dismutase (Zolg amp Philippi-Schulz 1994) secA1 codant la proteacuteine SecA1 intervenant dans le
seacutecreacutetion des proteacuteines agrave travers la membrane plasmique (Zelazny et al 2005) ou la reacutegion inter-
Chapitre 1
72
geacutenique encadreacutee par les ARNr 16S et 23S (appeleacutee ITS Internal Transcribed Spacer) (Roth et al 1998)
Malgreacute une ameacutelioration de la discrimination par lrsquoanalyse combineacutee de ces diffeacuterentes cibles le
seacutequenccedilage ne permet pas drsquoidentifier toutes les diffeacuterentes bacteacuteries du MTBC A cause de la forte
conservation des seacutequences dans le MTBC le seacutequenccedilage drsquoun grand nombre de gegravenes est neacutecessaire
pour discriminer les espegraveces (Kirschner et al 1993 Boumlddinghaus et al 1990) ce qui nrsquoest pas reacutealisable
en routine
Depuis lrsquoidentification de la plupart des gegravenes et des mutations responsables de la reacutesistance
des mycobacteacuteries aux antituberculeux le seacutequenccedilage est un des moyens utiliseacute pour identifier les
souches reacutesistantes La reacutesistance agrave la rifampicine par exemple est lieacutee agrave plusieurs mutations sur le gegravene
rpoB modifiant la structure de la sous-uniteacute β de lrsquoARN polymeacuterase et empecircchant la fixation de la
rifampicine sur lrsquoARN polymeacuterase conduisant agrave son inefficaciteacute (Zhang amp Yew 2009) Ces mutations
peuvent ecirctre deacutetecteacutees par seacutequenccedilage du gegravene amplifieacute par PCR
e Deacutetection et geacutenotypage des mycobacteacuteries du MTBC les marqueurs utiliseacutes
Les seacutequences drsquoinsertion
Les seacutequences drsquoinsertion (IS) correspondent agrave de petits eacuteleacutements geacuteneacutetiques transposables
Elles contiennent uniquement les informations geacuteneacutetiques neacutecessaires agrave leur transposition dans le
geacutenome de la bacteacuterie (transposase) On deacutenombre 42 IS dans le premier geacutenome annoteacute de M bovis
AF212297 (Garnier et al 2003) La seacutequence drsquoinsertion IS6110 speacutecifique du MTBC est la cible
historique utiliseacutee pour la deacutetection moleacuteculaire du Complexe tuberculosis en PCR (Thierry Cave et al
1990 Thierry Brisson-Noeumll et al 1990 Dziadek et al 2001) La meacutethode de reacutefeacuterence pour le
geacutenotypage de M tuberculosis est la RFLP-IS6110 (Restriction Fragment Lenght Polymorphism sur
lrsquoIS6110) Crsquoest la seacutequence drsquoinsertion la plus abondante dans le geacutenome de M tuberculosis qui en
contient de 1 agrave plus de 25 copies (Thierry Cave et al 1990 Thierry Brisson-Noeumll et al 1990) Elle
jouerait un rocircle dans lrsquoadaptation des bacteacuteries du MTBC agrave leur hocircte et dans leur virulence (Otal et al
2008 Safi et al 2004)
De nombreux systegravemes drsquoamorces ont eacuteteacute deacuteveloppeacutes pour la deacutetection de lrsquoIS6110 par PCR
dans les eacutechantillons biologiques (Dziadek et al 2001 El Kheacutechine et al 2009 Lieacutebana et al 1996
Savelkoul et al 2006) Diffeacuterents kits commerciaux utiliseacutes agrave lrsquoheure actuelle sont baseacutes sur la deacutetection
de lrsquoIS6110 notamment dans le diagnostic animal (Courcoul et al 2014) Cette cible a deacutejagrave eacuteteacute utiliseacutee
pour la deacutetection de M bovis dans des eacutechantillons inoculeacutes en PCR nicheacutee (Adams et al 2013) ainsi
que pour la deacutetection moleacuteculaire du MTBC dans du sol de lrsquoeau des seacutediments et des matiegraveres feacutecales
drsquoanimaux (Santos Santos et al 2015) (Tableau 3)
Chapitre 1
73
LrsquoIS6110 est eacutegalement utiliseacutee pour le typage des bacteacuteries du MTBC La comparaison des
souches repose sur le nombre drsquoIS6110 et leurs positions sur le chromosome apregraves extraction de lrsquoADN
digestion par une endonucleacutease (PvuII qui coupe lrsquoIS6110 sur un seul site) eacutelectrophoregravese transfert
sur membrane et hybridation (Van Soolingen et al 1991) Chaque fragment contenant une copie de
lrsquoIS6110 produit une bande (Figure 16) Cette meacutethode standardiseacutee beacuteneacuteficie de banques de donneacutees
de profils drsquohybridation internationales qui permettent des investigations eacutepideacutemiologiques Cette
meacutethode est beaucoup moins discriminante sur les souches agrave faible nombre drsquoIS (Allix et al 2006)
comme M bovis qui nrsquoen contient qursquoune agrave cinq copies et M bovis BCG qui nrsquoen contient qursquoune seule
copie (Van Soolingen et al 1991 Skuce et al 1994) Le reacutesultat est obtenu en une dizaine de jours
neacutecessite un volume important de culture et des manipulations fastidieuses De ce fait il a eacuteteacute
remplaceacute par des meacutethodes plus rapides indeacutependantes de la culture le spoligotypage et le typage
MLVA
Figure 16 Profils de restriction disolats de M tuberculosis obtenus suite agrave une RFLP-IS6110 Les isolats 3 5 6 9 et 10 ont le mecircme profil et posseacutedaient un lien eacutepideacutemiologique S repreacutesente le marqueur de poids moleacuteculaire (Source web site CDC)
Les courtes seacutequences drsquoADN reacutepeacuteteacutees
Le locus DR
Les souches du Complexe tuberculosis contiennent une reacutegion chromosomique particuliegravere
unique et figeacutee caracteacuteriseacutee par des seacutequences reacutepeacuteteacutees de 36 pb appeleacutee locus DR (Direct Repeat)
qui appartient agrave la famille des ADN reacutepeacutetitifs CRISPR (Clustered Regularly Interspersed Short
Chapitre 1
74
Palindromic Repeats) Ces DR sont seacutepareacutes par des espaceurs de 35 agrave 41 pb dont les seacutequences sont
uniques et polymorphes Ce polymorphisme associeacute agrave la deacuteleacutetion de certains espaceurs par des
reacutearrangements geacuteneacutetiques est exploiteacute dans le spoligotypage (en anglais spoligotyping
SPacer OLIGOnucleacuteotide TYPING) (Kamerbeek et al 1997) Par exemple les espaceurs 3 9 16 et 39 agrave
43 sont absents chez M bovis M caprae M microti et M pinnipedii (Smith et al 2006) comme
preacutesenteacute dans la Figure 17 Le geacutenotypage par spoligotypage est baseacute sur lrsquoamplification par PCR du
locus DR le marquage des amplicons et leur hybridation par des oligonucleacuteotides fixeacutes agrave une
membrane (spoligotypage 2D) Le profil final est deacutetecteacute par chimioluminescence (Kamerbeek et al
1997) Dans la technologie Luminex les oligonucleacuteotides sont fixeacutes sur des microbilles agrave signature
spectrale diffeacuterente (spoligotypage 3D) (Zhang et al 2010) Le spolygotypage standard analyse 43
espaceurs et permet drsquoobtenir des donneacutees eacutepideacutemiologiques et phylogeacuteneacutetiques deacuteterminantes (Sola
et al 2001) Il est largement utiliseacute pour typer les souches de M bovis notamment en France (Haddad
et al 2001) Cette meacutethode est reacutealisable sur de faibles quantiteacutes drsquoADN mecircme impur et des bacteacuteries
non cultivables Crsquoest une technique robuste rapide simple de reacutealisation dont le reacutesultat est facile agrave
interpreacuteter car il fournit un code binaire (pour chaque espaceur preacutesent 1 ou absent 0) faisant lrsquoobjet
drsquoune convention internationale (Dale et al 2001) Toutefois le spoligotypage ne preacutesente pas un
pouvoir de reacutesolution suffisant pour la diffeacuterenciation de certains groupes clonaux car il ne cible qursquoun
seul locus sur lrsquoensemble du geacutenome
Chapitre 1
75
C
Figure 17 Principe du spoligotypage des bacteacuteries du MTBC dapregraves Kamerbeek et al (1997) A Repreacutesentation du locus DR de M tuberculosis H37Rv et M bovis BCG B Amplifications des espaceurs par un couple damorces (a et b) et amplicons obtenus C Profils drsquohybridation sur membrane (un carreacute noir signale la preacutesence de lrsquoespaceur les parties blanches leur absence)
76
Gegravene ou locus
cible Espegraveces cibles PCR
Taille du
produit PCR
(pb)
Limites de deacutetection
(bacteacuteries g-1 sol
ou mL-1)
Matrices analyseacutees
Quantification si
deacutetection positive (nb
gegravenes ou UFC g-1 sol
ou mL-1)
Reacutefeacuterence Utiliseacute par
mpb64 MTBC PCR classique 543 101-102 Sols (terriers et preacutes
adjacents) 10 - 25 times 103 (Gormley 1999) (Young et al 2005)
mpb70 MTBC PCR classique 471 101-102 Sols (terriers et preacutes
adjacents) 10 - 25 times 103 (Gormley 1999)
(Young et al 2005)
(Courtenay et al 2006)
Gegravene codant
lrsquoARNr 16S
Mycobacteacuteries
agrave croissance
lente
RT-PCR 605 102-103 Sols (terriers et preacutes
adjacents) -
(Young et al
2005)
(Young et al 2005)
(Courtenay et al 2006)
IS6110 MTBC PCR nicheacutee PCR1 252
PCR2 116 -
Sol Eau
Foin Maiumls -
(Adams et al
2013) (Adams et al 2013)
IS6110 MTBC PCR nicheacutee 110
Sol 40 times 104
Eau 50 times 105
Sol
Seacutediment
Eau et fegraveces
(Soo et al 2006) (Santos Santos et al 2015)
RD12 M bovis
M caprae PCR classique 306
Sol 10 times 106
Eau 50 times 105
Sol
Seacutediment
Eau et fegraveces
(Warren et al
2006) (Santos Santos et al 2015)
RD12
M microti
M tuberculosis
M africanum
M pinnipedii
PCR semi-
nicheacutee 369
Sol 40 times 104
Eau 50 times 105
Sol
Seacutediment
Eau et fegraveces
(Warren et al
2006) (Santos Santos et al 2015)
RD4 (zone
flanquante) M bovis qPCR 142
42 times 105
(Pontiroli et al 2011)
10 times 105
(Pontiroli et al 2011)
Sols de terriers
Latrines
68 times 104 agrave 54 times 106
(Sweeney et al 2007)
20 times 104 agrave 10 times 108
(Travis et al 2011)
(Sweeney et al
2007)
(Sweeney et al 2007)
(King Murphy James Travis
Porter Sawyer et al 2015)
(King Murphy James Travis
Porter Hung et al 2015)
(Pontiroli et al 2011)
(Roug et al 2014)
Rv1510 (RD4) MTBC sauf
M bovis
PCR classique 1 033 - Sols de terriers
Latrines
- (Huard et al
2003)
(Courtenay et al 2006)
(Sweeney et al 2006)
(Sweeney et al 2007)
Tableau 3 Cibles moleacuteculaires deacutecrites dans la litteacuterature pour la deacutetection du Complexe tuberculosis ou de M bovis dans les eacutechantillons environnementaux
Chapitre 1
77
Les mini-satellites
Les mini-satellites correspondent agrave de courtes seacutequences drsquoADN reacutepeacuteteacutees (de 10 agrave 1 000 pb) et
conseacutecutives Pour pallier le manque de discrimination du spoligotypage un typage sur diffeacuterents loci
a eacuteteacute deacuteveloppeacute le MLVA (Multi Locus VNTR Analysis) baseacute sur lrsquoanalyse drsquoeacuteleacutements situeacutes sur des
seacutequences intergeacuteniques (les MIRU Mycobacterial interspersed Repetitive Unit) composeacutes de
seacutequences reacutepeacuteteacutees en tandem dont le nombre et la seacutequence sont variables (notamment les VNTR
Variable Number Tandem Repeat et les ETR Exact Tandem Repeat) (Supply et al 2001 Supply et al
2006) Lrsquoeacutetude de loci disperseacutes dans le geacutenome apporte une discrimination robuste des souches Le
geacutenotypage des souches de M tuberculosis ou M bovis par la meacutethode MIRU-VNTR repose sur la
recherche du nombre drsquoexemplaires des seacutequences reacutepeacuteteacutees de chaque locus eacutetudieacute Plus le nombre
de loci analyseacutes est important plus la discrimination est fine ainsi le MIRU-VNTR peut porter sur 12
15 ou 24 loci pour le typage des souches de M tuberculosis selon lrsquoobjectif envisageacute (eacutepideacutemiologie ou
phylogeacutenie) (Supply et al 2001 Supply et al 2006) Concernant le typage des souches animales de
M bovis diffeacuterentes combinaisons de marqueurs VNTR ont eacuteteacute eacutevalueacutees (Roring et al 2002 Allix et al
2006 Skuce et al 2002 Boniotti et al 2009) et ont montreacute que lrsquoanalyse de 5 agrave 8 loci suffisait pour
obtenir un pouvoir de discrimination supeacuterieur agrave 90 En France 8 loci ont eacuteteacute seacutelectionneacutes sur leur
polymorphisme et leur pouvoir de discrimination et utiliseacutes pour typer plus de mille isolats cliniques
recueillis entre 1978 et 2011 (Hauer et al 2015) Dans cette eacutetude les reacutesultats de geacutenotypage se
preacutesentent sous forme drsquoun code agrave 8 chiffres (Tableau 4) Pour un mecircme spoligotype SB0120 le
geacutenotypage MIRU-VNTR permet de diffeacuterencier les souches retrouveacutees en Cocircte drsquoOr de celles de
Dordogne Ces reacutesultats ont eacutegalement permis de deacutemontrer lrsquohomogeacuteneacuteiteacute des profils geacuteneacutetiques
retrouveacutes chez les animaux domestiques et sauvages dans une zone geacuteographique confirmant
lrsquohypothegravese de la transmission interspeacutecifique de lrsquoinfection
Tableau 4 Spoligotypes et geacutenotypes MIRU-VNTR des souches de M bovis isoleacutees chez les bovins et la faune sauvage en Cocircte dOr et en Dordogne (drsquoapregraves Hauer et al 2015)
Spoligotype Geacutenotype MIRU-VNTR (8 loci cibleacutes)
Origine des isolats 2165 2461 0577 580 2163a 2163b 4052 3232
SB0120
Cocircte drsquoOr 5 5 4 3 11 4 5 6
Bovins blaireaux
sangliers
SB0120
Dordogne 5 3 5 3 9 4 5 6
Bovins blaireaux
sangliers
SB0134
Cocircte drsquoOr 6 4 5 3 6 4 3 6
Bovins blaireaux
sangliers cerfs
Chapitre 1
78
La combinaison du spoligotypage et de la MLVA optimise la discrimination des souches pour
les suivis eacutepideacutemiologiques et apporte des informations phylogeacuteneacutetiques inteacuteressantes Elle permet
eacutegalement de suivre lrsquoeacutemergence de variants clonaux chez un animal ou dans un troupeau infecteacute en
fonction du temps (Navarro et al 2015 Rodriguez-Campos et al 2014) Toutefois les loci agrave analyser
doivent ecirctre adapteacutes en fonction des complexes clonaux rencontreacutes dans la reacutegion ou le pays eacutetudieacutes
(Boniotti et al 2009 Hauer et al 2015) Diffeacuterentes bases de donneacutees internationales sont disponibles
pour la comparaison des reacutesultats de spoligotypage et de MIRU-VNTR comme SpolDB4 SITVITWEB
MIRU-VNTRplus et Mbovisorg (regroupe les spolygotypes des espegraveces deacuteleacuteteacutees pour RD9) (Couvin amp
Rastogi 2014 Smith amp Upton 2012)
Reacutegions de Diffeacuterence (RD) et Single Nucleotide
Polymorphism (SNP)
Les Reacutegions de Diffeacuterences (RD1 agrave RD14) identifieacutees chez les bacteacuteries du MTBC correspondent
agrave des deacuteleacutetions et des reacutearrangements geacuteneacutetiques irreacuteversibles et discriminants Ces RD sont
speacutecifiques drsquoune espegravece ou communes agrave plusieurs espegraveces Le plus grand nombre de RD est observeacute
dans le geacutenome de M bovis BCG qui preacutesente 14 RD par comparaison agrave M tuberculosis H37Rv (Gordon
et al 1999 Brosch et al 2002) En parallegravele six seacutequences deacuteleacuteteacutees chez M tuberculosis H37Rv (les
RvD) et une seacutequence absente de toutes les souches de M tuberculosis (TbD1) sont preacutesentes chez les
autres bacteacuteries du MTBC (Gordon et al 1999 Brosch et al 2002) Lrsquoabsence de RD4 est speacutecifique agrave
M bovis et M bovis BCG et permet de les diffeacuterencier des autres membres du MTBC (Huard et al 2003
Huard et al 2006) La RD4 a eacuteteacute utiliseacutee dans diffeacuterentes eacutetudes pour la deacutetection de M bovis dans les
eacutechantillons environnementaux (Courtenay et al 2006 Sweeney et al 2007 King Murphy James
Travis Porter Sawyer et al 2015 Travis et al 2011 Roug et al 2014 Pontiroli et al 2011) Dans une
eacutetude reacutecente Santos et al (2015) ont associeacute la deacutetection du MTBC par lrsquoIS6110 agrave une identification
de lrsquoespegravece par amplification de deux reacutegions de la RD12 pour diffeacuterencier M bovisM caprae des
autres espegraveces du Complexe (Tableau 3)
Lrsquointeacuterecirct des RD est bien plus phylogeacuteneacutetique que geacutenotypique puisque lrsquoeacutetude de leur
apparition successive au cours de lrsquoeacutevolution a permis drsquoeacutetablir preacuteciseacutement la phylogeacutenie des
diffeacuterentes espegraveces comme preacutesenteacute preacuteceacutedemment dans la Figure 3 (Huard et al 2006 Rodriguez-
Campos et al 2014 Dippenaar et al 2015)
Les Single Nucleotide Polymorphism (SNPs) correspondent agrave des mutations ponctuelles drsquoune
base dans un geacutenome Ils peuvent ecirctre synonymes (le changement de base nrsquoaffecte pas la seacutequence
proteacuteique correspondante) ou non synonymes (la seacutequence proteacuteique est affecteacutee) et repreacutesentent
Chapitre 1
79
une variabiliteacute du geacutenome utile pour la caracteacuterisation de la variabiliteacute drsquoun groupe clonal et lrsquoeacutetude
des relations phylogeacuteneacutetiques entre les souches (Mathema et al 2006)
f Identification par seacutequenccedilage complet des geacutenomes
Le seacutequenccedilage complet des geacutenomes (WGS Whole Genome Sequencing) beacuteneacuteficie depuis
quelques anneacutees de nouvelles technologies (NGS Next Generation Sequencing) plus rapides et moins
coucircteuses que la meacutethode de Sanger Diffeacuterentes technologies coexistent agrave lrsquoheure actuelle dont les
plus utiliseacutees sont le pyroseacutequenccedilage 454 (toutefois en fin drsquoutilisation) la technique Illumina et la
technique Ion PGM (Ion Personal Genome Machine) (Desikan amp Narayanan 2015) Ces techniques ont
reacutevolutionneacute le geacutenotypage en fournissant le plus haut niveau de discrimination geacuteneacutetique possible
Ces techniques parfois coupleacutees au typage des SNPs permettent de suivre la dynamique de
transmission drsquoune maladie de caracteacuteriser lrsquoantibioreacutesistance drsquoune souche et drsquoeacutevaluer la divergence
geacuteneacutetique de souches qui ont les mecircmes profils de restriction ou de geacutenotypage (Schuumlrch amp van
Soolingen 2012) Une eacutetude irlandaise sur des souches de M bovis isoleacutees de bovins et de blaireaux
vivant dans les mecircmes zones a permis gracircce au WGS et agrave lrsquoanalyse de 39 SNPs de deacutemontrer la
circulation et la persistance locales drsquoune souche entre animaux sauvages et domestiques (Biek et al
2012) Une eacutetude reacutecente portant sur le seacutequenccedilage complet de 82 souches de M bovis isoleacutees en
France entre 1978 et 2011 a permis de deacutemontrer la congruence entre le typage par
spoligotypageMIRU-VNTR et le typage des souches par analyse des SNP (Hauer 2015) suggeacuterant que
certains SNP pourraient ecirctre utiliseacutes comme cibles dans le geacutenotypage de M bovis Dans un futur
proche le WGS sera probablement lrsquooutil qui offrira lrsquoinformation la plus complegravete sur le geacutenotype des
souches les liens phylogeacuteneacutetiques entre souches et qui permettra de deacutecoder les scheacutemas de
transmission de lrsquoinfection entre animaux (Biek et al 2012 Pepperell et al 2013 Rodriguez-Campos et
al 2014)
Les puces (microarrays) agrave ADN ou ARN sont utiliseacutees pour la comparaison de geacutenomes complets
des espegraveces du MTBC et ont notamment apporteacute des informations sur lrsquoexpression de gegravenes de la
virulence chez M bovis (Voskuil et al 2004) ainsi que sur lrsquoidentification de seacutequences polymorphiques
distinguant les principaux complexes clonaux (Berg et al 2011 Muumlller et al 2009 Smith et al 2011)
Ces techniques appartiennent toutefois au domaine de la recherche et ne sont pas utiliseacutees en routine
g Meacutethode speacutecifique drsquoidentification des mycobacteacuteries animales utiliseacutee par le
Laboratoire National de reacutefeacuterence de la tuberculose bovine LNR TB
Lrsquoidentification des mycobacteacuteries isoleacutees sur des animaux est reacutealiseacutee au LNR TB de Maisons-
Chapitre 1
80
Alfort En premiegravere intention quatre systegravemes de deacutetection moleacuteculaires ciblant des seacutequences
drsquoinsertion (IS6110 IS1081 IS1245) et le gegravene atpE sont utiliseacutes en qPCR pour orienter lrsquoidentification
(Tableau 5)
Tableau 5 Systegravemes de deacutetection utiliseacutes en qPCR en premiegravere intention pour lidentification dune mycobacteacuterie dorigine animale et reacutesultats obtenus + amplification de la cible - absence drsquoamplification de la cible
ARNr 16S IS1245 IS901 IS6110 IS1081 IS1561rsquo RD4 RD7 atpE
Complexe tuberculosis
M bovis M tuberculosis
M microti M caprae
+ + + +
- - - -
- - - -
+ + + +
+ + + +
+ + - +
- + + +
- + + -
- - - -
Complexe avium + + +- - - - - + +
Autres mycobacteacuteries
+ - - - - - - + +
(Slana et
al 2010) (Slana et al
2010) (Courcoul et
al 2014) (cette thegravese)
(Radomski et al 2013)
Lrsquoidentification des espegraveces du complexe tuberculosis (IS6110 et IS1081 positives) est ensuite
reacutealiseacutee par geacutenotypage gracircce agrave la technique de spoligotypage (Zhang et al 2010) Lrsquoamplification ou
non des IS1245 et IS901 permet drsquoidentifier les espegraveces Mah (IS1245 et IS901 neacutegatives) Maa (IS1245
et IS901 positives) et Mah (IS1245 positive et IS901 neacutegative) (Slana et al 2010) Lrsquoidentification des
autres mycobacteacuteries repose sur le seacutequenccedilage de lrsquohsp65 puis de la comparaison par BLAST avec une
base de donneacutees (Telenti et al 1993)
De par le grand nombre drsquoespegraveces de mycobacteacuteries et la forte identiteacute des espegraveces dans le
Complexe MTBC la deacutetection et le geacutenotypage des souches du MTBC reposent obligatoirement sur
lrsquoutilisation conjointe de diffeacuterentes techniques En fonction des eacutechantillons disponibles et des
objectifs souhaiteacutes qursquoils soient diagnostiques eacutepideacutemiologiques ou phylogeacuteneacutetiques le choix des
techniques agrave utiliser doit ecirctre reacutefleacutechi et pertinent
82
Objectifs de la Thegravese
Depuis 2004 la situation de la Cocircte drsquoOr vis-agrave-vis de la tuberculose bovine est preacuteoccupante
Malgreacute la mise en place de moyens de surveillance et de lutte en premier lieu dans les cheptels bovins
puis dans la faune sauvage lrsquoeacuteradication de la maladie semble difficile Depuis 2011 on observe une
diminution du nombre des foyers bovins et une stabiliteacute du nombre de cas observeacutes dans la faune
sauvage avec une concentration spatiale des foyers dans des petites valleacutees Dans ces valleacutees ougrave
cohabitent faune sauvage et bovins la transmission indirecte du bacille entre animaux sauvages et
domestiques par le biais de lrsquoenvironnement est fortement suspecteacutee pour plusieurs cas bovins Bien
que plusieurs eacutetudes aient deacutemontreacute la preacutesence de M bovis dans diffeacuterentes matrices
environnementales naturelles sur les lieux de vie drsquoanimaux domestiques et sauvages infecteacutes
notamment en Angleterre et en Espagne aucune donneacutee nrsquoest disponible sur la contamination
environnementale potentielle par M bovis dans les zones endeacutemiques de tuberculose bovine en
France et plus particuliegraverement en Cocircte drsquoOr
Pour reacutepondre agrave cette question le premier objectif du travail de thegravese a eacuteteacute de deacutevelopper et de valider
sur des eacutechantillons inoculeacutes artificiellement des outils moleacuteculaires speacutecifiques et sensibles pour la
deacutetection des souches du MTBC et de M bovis dans les substrats environnementaux naturellement
contamineacutes tels que le sol lrsquoeau et les seacutediments et les matiegraveres feacutecales animales Nous avons
eacutegalement deacuteveloppeacute un protocole drsquoextraction-deacutecontamination des mycobacteacuteries du sol coupleacute agrave
une technique originale de suivi de croissance des mycobacteacuteries en milieu liquide dans lrsquoobjectif
drsquoisoler les souches environnementales de M bovis (Chapitre 2)
Les outils moleacuteculaires deacuteveloppeacutes ont eacuteteacute utiliseacutes pour la recherche de M bovis dans un grand nombre
drsquoeacutechantillons environnementaux (sol eau seacutediments fegraveces animales et membres de la micro- ou
meacuteso-faune des sols) preacuteleveacutes dans la zone endeacutemique de Cocircte drsquoOr et plus preacuteciseacutement dans des
zones partageacutees par la faune domestique et sauvage infecteacutee afin de preacuteciser le niveau de
contamination des lieux de vie des bovins et des animaux sauvages (Chapitre 3)
Face aux eacutetudes parfois controverseacutees sur le rocircle des facteurs abiotiques dans la persistance de
M bovis dans le sol nous avons rechercheacute lrsquoeffet de la nature du sol et de la tempeacuterature sur la dureacutee
de survie de M bovis SB0120 en conditions de laboratoire (Chapitre 4)
Nous avons enfin eacutetudieacute le rocircle de la faune du sol et plus particuliegraverement des lombrics tregraves abondants
dans les pacirctures et gros consommateurs de matiegravere organique dans la circulation et la disseacutemination
de M bovis dans les sols (Chapitre 5)
CHAPITRE 2
Deacuteveloppement et optimisation drsquooutils pour la deacutetection
de Mycobacterium bovis dans les eacutechantillons
environnementaux
PARTIE CONFIDENTIELLE agrave lrsquoexception des meacutethodes de deacutetection moleacuteculaire
86
CHAPITRE 2 - Deacuteveloppement et optimisation drsquooutils pour la
deacutetection de Mycobacterium bovis dans les eacutechantillons
environnementaux
A DEVELOPPEMENT DrsquoOUTILS MOLECULAIRES ADAPTES AUX MATRICES ENVIRONNEMENTALES
Les diffeacuterents systegravemes moleacuteculaires utiliseacutes pour la deacutetection de M bovis dans
lrsquoenvironnement citeacutes dans la litteacuterature sont syntheacutetiseacutes dans le Tableau 3 En consideacuterant la diversiteacute
et lrsquoabondance des mycobacteacuteries potentiellement preacutesentes dans lrsquoenvironnement nous avons
eacutelimineacute les systegravemes ciblant toutes les espegraveces du MTBC notamment lrsquoIS6110 car bien que tregraves
sensible et speacutecifique sur les eacutechantillons biologiques et tregraves utiliseacutee en diagnostic humain et
veacuteteacuterinaire nous avons montreacute qursquoelle nrsquoeacutetait pas utilisable en diagnostic environnemental En effet
nous avons effectueacute des essais preacuteliminaires avec le kit LSI VetMAXtrade Mycobacterium tuberculosis PCR
Kit (Life Technologies) utiliseacute en diagnostic veacuteteacuterinaire sur 25 eacutechantillons de sol provenant de zones
geacuteographiques non impacteacutees par la tuberculose bovine Pour tous ces sols lrsquoIS6110 a donneacute un signal
moleacuteculaire quantifiable teacutemoignant de lrsquoabsence de speacutecificiteacute de ce systegraveme pour la matrice sol
De plus M microti membre du MTBC agent de la tuberculose des petits rongeurs est susceptible
drsquoecirctre retrouveacute dans le sol (Courtenay et al 2006) et a deacutejagrave eacuteteacute isoleacute sur des feacutelins et des animaux
sauvages en Cocircte drsquoOr (Laprie et al 2013 Michelet et al 2015) Nous avons donc deacuteveloppeacute des
systegravemes moleacuteculaires qui nrsquoamplifient pas M microti
Enfin M bovis est une mycobacteacuterie pathogegravene de classe III qui neacutecessite une manipulation dans un
laboratoire de confinement L3 Pour des raisons techniques et pour srsquoaffranchir de la contrainte de
confinement dans la mise au point de la technique de PCR nous avons utiliseacute une des souches
atteacutenueacutees de M bovis et plus preacuteciseacutement M bovis BCG Pasteur 1173P2 manipulable dans un
laboratoire de type L2 disponible agrave lrsquoINRA de Dijon Nous avons donc choisi des cibles moleacuteculaires
eacutegalement preacutesentes chez M bovis BCG
Pour augmenter la speacutecificiteacute de la deacutetection moleacuteculaire dans les matrices environnementales
complexes nous avons choisi drsquoutiliser la PCR quantitative avec une sonde TaqManreg qui est formeacutee
par un nucleacuteotide lieacute de faccedilon covalente agrave un fluorophore (reporter) en 5rsquo et agrave un deacutesactivateur
(quencher) en 3rsquo Le clivage de la sonde par la Taq polymeacuterase apregraves lrsquohybridation agrave la seacutequence drsquoADN
compleacutementaire eacuteloigne le quencher du fluorophore qui eacutemet une fluorescence mesureacutee en temps
reacuteel dans un thermocycleur adapteacute
Chapitre 2
87
Le systegraveme moleacuteculaire deacutecrit preacuteceacutedemment par Sweeney et al (2007) ciblant les marges de
la reacutegion RD4 deacuteleacuteteacutee uniquement chez M bovis et M bovis BCG nous a particuliegraverement inteacuteresseacutes
Des amorces internes agrave la seacutequence RD4 ont eacuteteacute utiliseacutees en PCR pour la discrimination de M bovis
avec les autres membres du MTBC (pas de signal pour M bovis mais un signal pour les autres membres
du MTBC) (Huard et al 2003 Courtenay et al 2006) Des amorces de PCR classique ont eacutegalement eacuteteacute
dessineacutees sur les zones flanquantes de la reacutegion RD4 (Brosch et al 2002) Ces amorces ont eacuteteacute adapteacutees
agrave la technologie TaqManreg avec une amorce et une forward localiseacutees sur les marges de la seacutequence
deacuteleacuteteacutee et une sonde agrave cheval sur la zone de deacuteleacutetion (Sweeney et al 2007) La speacutecificiteacute et la
validation de ce systegraveme sur diffeacuterents eacutechantillons environnementaux tels que des sols de terriers
des fegraveces de blaireaux et de buffles africains ont eacuteteacute deacutemontreacutees dans plusieurs eacutetudes (Sweeney et al
2007 Roug et al 2014 Travis et al 2011 Pontiroli et al 2013 King et al 2015) Ce systegraveme de deacutetection
nous paraissant a priori speacutecifique pour la discrimination de M bovis dans le MTBC et adapteacute aux
matrices complexes nous lrsquoavons inclus dans lrsquoeacutetude Toutefois lrsquoamorce Reverse du systegraveme de
deacutetection a eacuteteacute modifieacutee par nos soins afin de reacuteduire lrsquoamplicon obtenu de 146 agrave 116 bases taille plus
adapteacutee agrave la technologie TaqManreg
Face au manque de systegravemes discriminants disponibles dans la litteacuterature nous avons dessineacute
deux nouveaux systegravemes Nous avons choisi de cibler la seacutequence drsquoinsertion IS1561rsquo (Rv3349c) car elle
est preacutesente chez tous les membres du MTBC sauf M microti (Gordon et al 1999 Huard et al 2003
Brodin et al 2002) Elle appartient agrave la famille des ISL3 qui regroupe des copies drsquoIS deacutefectueuses chez
M tuberculosis elles sont alors annoteacutees avec une apostrophe
La troisiegraveme cible a eacuteteacute choisie dans une reacutegion chromosomique de M tuberculosis appeleacutee
RD1 preacutesente chez les souches virulentes de M tuberculosis et M bovis mais partiellement deacuteleacuteteacutee
chez M bovis BCG (Mahairas et al 1996) et M microti (Brodin et al 2002) Cette reacutegion contient
notamment un cluster de 6 gegravenes (Rv3871 agrave Rv3876) qui fait partie des gegravenes codant les proteacuteines
antigeacuteniques ESAT-6 (Early Secreted Antigenic Target 6kD) et CFP-10 (Culture Filtrat Protein 10kD)
responsables de la reacuteponse immunitaire de lrsquohocircte M bovis BCG et M microti sont les deux seuls
membres du MTBC agrave preacutesenter une deacuteleacutetion de ce cluster deacuteleacutetion probablement en partie
responsable de leur absence de virulence et de leur utilisation en vaccination (Pym et al 2002) Chez
M bovis BCG la reacutegion aval de ce cluster est deacuteleacuteteacutee la reacutegion deacuteleacuteteacutee est appeleacutee RD1bcg (Figure 18)
Chez M microti crsquoest la reacutegion amont du cluster qui est absente la reacutegion deacuteleacuteteacutee est appeleacutee RD1mic
Nous avons plus particuliegraverement cibleacute le locus Rv3866 preacutesent chez M tuberculosis et dans toutes les
souches de M bovis et M bovis BCG mais absente chez M microti (Brodin et al 2002) M avium et
M smegmatis (Mahairas et al 1996)
Chapitre 2
88
Figure 18 Diffeacuterences geacutenomiques de la reacutegion RD1 entre M bovis BCG (A) M microti (C) par comparaison agrave M tuberculosis H37Rv (B) drsquoapregraves Brodin et al 2002 et localisation du locus Rv3866 RD1mic correspond agrave la seacutequence de M tuberculosis deacuteleacuteteacutee chez M microti RD1bcg correspond agrave la seacutequence de M tuberculosis deacuteleacuteteacutee chez M bovis BCG
Les seacutequences drsquointeacuterecirct ont eacuteteacute rechercheacutees pour les membres du MTBC et certaines MNT sur
la base de donneacutees GenBank Un alignement de ces seacutequences a eacuteteacute reacutealiseacute gracircce au portail drsquoanalyses
bioinformatiques MOBYLE PASTEUR pour rechercher les reacutegions conserveacutees et les reacutegions de
diffeacuterence Le dessin des oligonucleacuteotides a eacuteteacute reacutealiseacute grace au logiciel en ligne Primer3
(httpbioinfouteeprimer3-040) Les systegravemes de deacutetection ont eacuteteacute testeacutes in silico gracircce agrave lrsquooutil
BLAST (Basic Local Alignment Search Tool) afin drsquoeacutevaluer la speacutecificiteacute de nos systegravemes pour chaque
cible par rapport agrave une base de donneacutees de geacutenomes seacutequenceacutes
La validation (speacutecificiteacute et seuil de deacutetection) de ces trois systegravemes ainsi que leur utilisation
dans le screening drsquoun nombre important drsquoeacutechantillons environnementaux sont preacutesenteacutees dans le
Chapitre 3 dans lrsquoarticle intituleacute laquo First molecular detection of Mycobacterium bovis in environmental
samples from a French region with endemic bovine tuberculosis raquo Les reacutesultats des analyses reacutealiseacutees
en 2013 sont preacutesenteacutes dans un article publieacute en 2014 dans la revue de lrsquoAEEMA (association pour
leacutetude de leacutepideacutemiologie des maladies animales) en Annexe 1
A
B
C
Chapitre 2
89
CHAPITRE 3
Etude de la preacutevalence environnementale de
Mycobacterium bovis dans la zone endeacutemique de
tuberculose bovine de Cocircte drsquoOr et reacutesultats obtenus
en Charente
Elodie Barbier Maria Laura Boschiroli Eric Gueneau Murielle Rochelet Ariane Payne Krystel
de Cruz AnneLaure Blieux Ceacuteline Fossot Alain Hartmann
First molecular detection of Mycobacterium bovis in environmental samples from a French
region with endemic bovine tuberculosis
(Article accepteacute en cours de publication Journal of Applied Microbiology)
93
CHAPITRE 3 - Etude de la preacutevalence environnementale de
Mycobacterium bovis dans la zone endeacutemique de tuberculose
bovine de Cocircte drsquoOr et reacutesultats obtenus en Charente
A AVANT-PROPOS
Dans la synthegravese bibliographique preacutesenteacutee dans le chapitre 1 nous avons deacutecrit la situation
eacutepideacutemiologique complexe de la tuberculose bovine en Cocircte drsquoOr Dans ce deacutepartement
Mycobacterium bovis circule dans un systegraveme multi-hocirctes complexe incluant les bovins les blaireaux
les cervideacutes et les sangliers sur une zone geacuteographique limiteacutee (Payne 2014) La faune sauvage infecteacutee
et potentiellement excreacutetrice est suspecteacutee drsquoecirctre agrave lrsquoorigine de la contamination ou recontamination
de certains foyers bovins dans cette zone (ANSES 2011 Courcoul Moutou amp Vialard 2013) Le mode
de transmission indirecte est suspecteacute car les contacts directs entre bovins et animaux sauvages sont
rarement deacutecrits (Drewe OrsquoConnor Weber McDonald amp Delahay 2013 Kukielka et al 2013) Ce mode
de transmission repose sur la contamination de matrices environnementales par des mycobacteacuteries
viables et virulentes agrave la faveur de lrsquoexcreacutetion animale puis sur lrsquointeraction drsquoanimaux avec ces
matrices Deacutemontreacutee par des meacutethodes moleacuteculaires etou culturales en Angleterre et en Espagne sur
des sites partageacutes par les espegraveces sauvages et les bovins (King et al 2015 Santos et al 2015 Sweeney
et al 2006 2007) la contamination environnementale par M bovis nrsquoa pas encore fait lrsquoobjet drsquoeacutetudes
dans les zones endeacutemiques franccedilaises
Afin drsquoeacutevaluer la preacutevalence et lrsquoexistence de reacuteservoirs potentiels de M bovis dans
lrsquoenvironnement des bovins et des animaux sauvages infecteacutes en Cocircte drsquoOr nous avons deacuteveloppeacute et
valideacute des systegravemes de deacutetection moleacuteculaire ou modifieacute des systegravemes existants adapteacutes aux matrices
environnementales complexes pour deacutetecter les espegraveces du MTBC et plus speacutecifiquement M bovis
Ces systegravemes ont eacuteteacute utiliseacutes pour analyser plus de 400 eacutechantillons environnementaux (sol eau
seacutediments microfaune fegraveces) preacuteleveacutes en Cocircte drsquoOr Notre strateacutegie drsquoeacutechantillonnage a cibleacute des
pacirctures suspectes sur lesquelles des bovins infecteacutes ont transiteacute et autour desquelles des animaux
sauvages infecteacutes ont eacuteteacute deacutetecteacutes et plus particuliegraverement les zones pouvant ecirctre partageacutees par les
diffeacuterentes espegraveces notamment les zones drsquoabreuvement Nous avons eacutegalement analyseacute des
eacutechantillons provenant de sites freacutequenteacutes exclusivement par la faune sauvage notamment des sols
de terriers et des latrines de blaireaux localiseacutes agrave proximiteacute du site de pieacutegeage de blaireaux infecteacutes
ainsi que des seacutediments preacuteleveacutes dans les trous drsquoeau ou souilles freacutequenteacutes par les sangliers et
cervideacutes
Chapitre 3
95
B FIRST MOLECULAR DETECTION OF MYCOBACTERIUM BOVIS IN ENVIRONMENTAL SAMPLES FROM A FRENCH REGION WITH ENDEMIC BOVINE TUBERCULOSIS
E Barbier12 ML Boschiroli3 E Gueneau4 M Rochelet2 A Payne1 Krystel de Cruz3 AL Blieux5 C
Fossot6 A Hartmann1
1 INRA UMR 1347 Agroeacutecologie Dijon Cedex France
2 Universiteacute de Bourgogne Franche Comteacute UMR 1347 Agroeacutecologie Dijon Cedex France
3 Universiteacute Paris-Est Laboratoire National de Reacutefeacuterence de la Tuberculose Uniteacute de Zoonoses
Bacteacuteriennes Laboratoire de Santeacute Animale ANSES Maisons-Alfort Cedex France
4 Laboratoire Deacutepartemental de la Cocircte drsquoOr Dijon France
5 Welience SATT Grand-Est AgrOnov Breteniegravere France
6 Phytocontrol Paris MIN de Rungis Rungis Cedex France
Running head
Environmental detection of Mycobacterium bovis
Corresponding author
Alain Hartmann UMR 1347 Agroeacutecologie INRA 17 rue Sully BP 86510 21065 Dijon Cedex France E-
mail alainhartmanndijoninrafr
Chapitre 3
96
ABSTRACT
Aims
The aim of the study was to determine the prevalence of Mycobacterium bovis (the causative agent of
bovine tuberculosis bTB) in environmental matrices within a French region (Cocircte drsquoOr) affected by this
zoonotic disease
Methods and Results
We report here the development and the use of molecular detection assays based on qPCR (double
fluorescent dye labelled probe) to monitor the occurrence of Mycobacterium tuberculosis complex
(MTBC) or M bovis in environmental samples collected in pastures where infected cattle and wildlife
had been reported Three qPCR assays targeting members of the MTBC (IS1561rsquo and Rv3866 loci) or M
bovis (RD4 locus) were developed or refined from existing assays These tools were validated using
M bovis spiked soil water and feces samples Environmental samples were detected positive for the
presence of MTBC strains and M bovis in the environment of bTB-infected farms in the Cocircte drsquoOr
region
Conclusions
The development of molecular assays permitted testing of several types of environmental samples
including spring water sediment samples and soils from badger setts entrance located in the vicinity
of these farms which were repeatedly contaminated with M bovis (up to 87 times 103 gene copies per g
of badger sett soil) For the first time direct spoligotyping of soil DNA enabled identification of M bovis
genotypes from environmental matrices
Significance and Impact of the Study
All together these results suggest that M bovis occurs at low levels in environmental matrices in Cocircte
drsquoOr within the bTB-infected area Drinking contaminated water or inhaling contaminated bioaerosols
might explain cattle infection in some cases
Keywords
Mycobacterium bovis environment quantitative real-time PCR indirect transmission
Chapitre 3
97
INTRODUCTION
Bovine tuberculosis (bTB) is a worldwide spread zoonosis caused by Mycobacterium bovis a
pathogenic mycobacteria that belongs to the Mycobacterium tuberculosis complex (MTBC) This
disease affects cattle that are the natural host of M bovis but also a wide range of domestic or wild
mammals including humans (Morris et al 1994 Biet et al 2005) Infected wildlife can act as a bTB
reservoir maintaining and spreading M bovis infection in cattle Many studies including French ones
showed the circulation of the bacteria within multi-host systems including cattle and various wild
species as badgers (Meles meles) wild boar (Sus scrofa) or red deer (Cervus elaphus) (Little et al 1982
Aranaz et al 2004 Gortaacutezar et al 2008 Zanella et al 2008 Richomme et al 2010 Payne et al 2012)
The zoonotic nature of bTB may represent a public health threat Moreover animal disease leads to
important economic losses mainly due to cattle testing slaughtering campaigns and wildlife screening
trials
Infected animals can excrete M bovis via sputum feces urine and any type of aerosols (Little
et al 1982 Phillips et al 2003 Palmer et al 2004 Payne 2014) and thus contaminate their environment
(Phillips et al 2003) This may contribute to contamination of cattle pastures through deposition of
M bovis on cattle feedstuffs and into drinking water Indeed many studies confirmed the presence and
the persistence of M bovis in environmental substrates such as soil feces hay corn water (Young et
al 2005 Courtenay et al 2006 Sweeney et al 2006 Fine et al 2011 Ghodbane et al 2014) despite
many technical difficulties in isolating this organism from environmental substrates Altogether these
data strongly suggest that the environment may constitute a reservoir of M bovis from which cattle
might get infected However there is still very little direct evidence concerning these potential M bovis
environmental reservoirs and the way that animals might get infected through them
The European Commission declared that France was bTB-free in 2001 However the disease is
still a persistent problem in several regions including Cocircte drsquoOr (Middle East of France Figure 1)
Despite a very low national prevalence in cattle (0075 since 2012) complete eradication of the
disease seems difficult to achieve (Fediaevsky et al 2014) In Cocircte drsquoOr M bovis circulates in a multi-
host system including badgers wild boar and deer within a restricted area of the region (Payne 2014
Hauer et al 2015) As shown in Figure 1 two genotypes are found in this area M bovis spoligotype
SB0120 VNTR profile 5 5 4 3 11 4 5 6 in the northern bTB endemic area and M bovis spoligotype SB0134
VNTR profile 6 4 5 3 6 4 3 6 in the southern area Between 2002 and 2014 196 cattle breakdowns have
been registered in Cocircte drsquoOr (Cote drsquoOr Regional Veterinary Service personal communication) Wild
animals in the vicinity of infected cattle herds were found to be infected by M bovis with the same
genotypes (Hauer et al 2015)
Direct cross contamination between cattle has historically been considered as the main
Chapitre 3
98
transmission route of M bovis (Morris et al 1994 Phillips et al 2003) but environmental
contamination is suspected in many herd infections when cattle and wildlife share the same resources
(Garnett et al 2002 Palmer et al 2004) However this hypothesis has never been extensively explored
in France This is probably due to the lack of adequate detection tools to monitor M bovis under natural
environmental conditions Indeed most of the studies report the detection of M bovis from
experimentally inoculated environmental samples (Fine et al 2011 Adams et al 2013) Although the
culture method is the gold standard for direct diagnosis of M bovis in animals it is not well suited to
studying environmental samples due to the abundance of soil microbial communities and the slow
growth rate of M bovis To circumvent these drawbacks the use of highly selective media containing
antibiotics and antifungals in combination with sample decontamination is required in order to prevent
competition of fast growing organisms However such pretreatments strongly reduce M bovis viability
(Thornton et al 1998 Fine 2006) On the other hand molecular detection tools such as semi-
quantitative PCR have been used to detect M bovis in soils water and badger feces Following DNA
extraction detection of MTBC members or M bovis was done using primers targeting mpb64 mpb70
hsp65 16SrRNA genes or IS6110 insertion sequence (Young et al 2005 Courtenay et al 2006
Sweeney et al 2006 Adams et al 2013 Santos et al 2015) More recently a M bovis specific qPCR
(based on double fluorescent dye probe detection) was developed to target the RD4 deletion flanking
sequences (Sweeney et al 2007) This molecular system has been validated in several studies to test
environmental substrates such as soil badger feces and cow slurry (Pontiroli et al 2011 Travis et al
2011 King et al 2015a King et al 2015b) This RD4 based qPCR assay allowed to detect the occurrence
of M bovis within environmental matrices in UK (Sweeney et al 2007 King et al 2015a King et al
2015b) Finally Santos et al (Santos et al 2015) successfully used a hemi-nested PCR targeting the
RD12 locus which is common to M bovis and M caprae to monitor these species in environmental
substrates from bTB endemic regions located in the Iberian Peninsula
In this context the objectives of this study were firstly to develop new sensitive qPCR based
tools and to refine an existing one (based on RD4 deletion sequence) for the specific detection of MTBC
or M bovis in environmental matrices The sensitivity and specificity of qPCR based detection systems
were evaluated on a collection of mycobacterial strain DNAs but also on DNAs extracted from samples
(soil water and feces) spiked with known amounts of M bovis Secondly we aimed to screen large
numbers of environmental samples (soil sediments water feces) originating from both bTB endemic
or bTB free areas to assess the presence of MTBC and more specifically the presence of M bovis in
these areas
Chapitre 3
99
Figure 1 Distribution of bTB cumulative outbreaks in cattle in France (picture in the insert above) and per municipality in Cocircte drsquoOr from 2005 to 2013 Geographical location of the sampled areas are indicated by black stars and the location of the six positive areas are indicated by white stars and named A B C D E and F respectively
MATERIALS AND METHODS
Bacterial strains
A collection of bacterial strains belonging to the species M bovis M bovis BCG vaccine strain
and other mycobacterial species (Table 1) was used to test the specificity of the primers to detect
M bovis Crude thermo-lysates of strains corresponding to 20 bacterial spoligotypes of the M bovis
species 4 other species of the MTBC 2 sub-species of the Mycobacterium avium Complex (MAC) and
20 non tuberculous mycobacteria (NTM) Field strains belong to the National Reference Laboratory
collection and were previously identified by several genotyping methods using four qPCR assays done
in parallel and targeting IS6110 IS1081 IS1245 and atpE Namely MTBC members are positive for
Chapitre 3
100
IS6110 and IS1081 targets (Courcoul et al 2014) MAC members are positive for the IS1245 target
(Slana et al 2010) and finally NTM are positive for the atpE target (Radomski et al 2013) Furthermore
spoligotyping was used to discriminate the different MTBC species (Zhang et al 2010) IS901 was used
to discriminate between MAC members (Slana et al 2010) and hsp65 sequencing was used to
discriminate between other mycobacteria species (Telenti et al 1993) Attenuated M bovis BCG strain
Pasteur 1173P2 was used as target organism to validate the specificity and detection limits of the qPCR
assay in soil water and feces microcosms
Origin of environmental samples
Within the bTB endemic zone in Cocircte drsquoOr a total number of 356 environmental samples were
collected from 15 areas (an area corresponds to a municipality territory including at least one bTB
infected cattle farm) between April 2013 and December 2014 In those areas typical landscape is
observed ie limestone plateau with forest or farm fields overhang small hills The hillslopes are
covered with hydromorphic soils containing marl and claylimestone Alluvial valleys set at the bottom
of the slopes Cattle usually graze on those pastures during summer and spring Samples of soil
sediment and water were collected in areas frequented by both wildlife and cattle such as feeding
areas waterholes naturally occurring springs drinking troughs with open access to livestock and
wildlife Knowing that bTB infected badgers were discovered in the vicinity of the pastures grazed by
infected cattle (Cote drsquoOr Regional Veterinary Service personal communication) soils from badger
setts and latrines were also collected
Twenty-four agricultural soils were randomly chosen as control soils in French bTB-free areas
among a panel of soils from the French soil quality monitoring network (Reacuteseau de Mesures de la
Qualiteacute des Sols Infosol INRA Orleacuteans) Among them 11 french soils were collected from Brittany
(North West of France) Yvelines (Middle north of France) Haut Rhin (North east of France) while 13
soils were collected in the bTB-free area of Cocircte drsquoOr
In order to evaluate the detection limit of the qPCR assay we used two agricultural soils from two bTB-
free areas of Cocircte drsquoOr (Soil S1 and soil S2) one pond water and badger feces from animals tested as
bTB-free from non-infected areas Both soils were sampled in a bTB-free cattle pasture in November
2013 They were chosen for their contrasted structural and physico-chemical properties (Table 2)
DNA extraction and purification
DNAs from all soil sediment and latrine samples were extracted from 2 g-samples as following
Briefly 8 ml of lysis buffer (01 mol l-1 Tris pH 80 01 mol l-1 EDTA pH 80 01 mol l-1 NaCl and 2 (wv)
SDS) 4 g of silica beads (100 μm diameter) 5 g of ceramic beads (14 mm diameter) and 8 glass beads
(4 mm diameter) were added to each sample placed in a 15 ml conical centrifugation tube (Falcon
Chapitre 3
101
Dominique Dutscher France) Samples were disrupted for 3 times 30 s at 4 m s-1 in a FastPrep-24 Instrument
(MP Biomedicals Europe France) incubated for 30 min at 70degC and centrifuged for 5 min at 7000 g at
room temperature in a Sigma 3-18K centrifuge (Sigma-Aldrich Germany) Supernatants were
incubated on ice for 10 min with 110 volume of 3 mol l-1 potassium acetate pH 55 and then
centrifuged for 5 min at 14000 g One volume of ice-cold isopropanol was added to the supernatant
for DNA precipitation overnight at minus 20degC DNA was collected by centrifugation (30 min at 14000 g
4degC) and DNA pellets were washed with ice-cold ethanol (70 ) and dissolved in 100 μl of water Crude
DNA extracts (100 μl) were loaded on a Microbiospin column (Biorad France) filled with 100 mg of
polyvinyl polypyrrolidone (Sigma Chemical Co France) After centrifugation (4 min at 1000 g 4degC)
eluted DNA was further purified using the Geneclean Turbo Kit (Qiagen France) according to the
manufacturers instructions For water samples the microbial biomass was concentrated by filtering
from 100 ml to a maximum of 1 l of water sample (depending on water turbidity) through a 045 microm
Millipore HAWP filter (47 mm Merk Millipore Germany) Filters were then ground in liquid nitrogen
and DNA was extracted as described above Nematodes were extracted from soil using an Oostenbrink
elutriator (Oostenbrink 1960) DNA extraction was achieved as described above
Purified DNA concentrations were estimated using a NanoDropreg 2000 spectrophotometer
(Thermo Scientific France) at 260 nm wavelength
Choice of gene targets design of primers and probes for qPCR assays
A first qPCR detection system assay targeting the insertion sequence IS1561rsquo was designed
This insertion sequence is present in all the MTBC members except in M microti (Huard et al 2003
Huard et al 2006) The primers and double dye probe sequences are given in Table 3 IS1561rsquo contains
a defective transposase and belongs to the ISL3 family (Gordon et al 1999) In order to confirm the
specificity of the detection for MTBC a nested PCR protocol was additionally developed the first step
was a classical PCR using primers targeting a sequence of 222-bp sequence in the IS1561rsquo surrounding
the 76-bp qPCR product (Table 3) the second step was a qPCR assay targeting the 76-bp fragment To
confirm the results obtained with IS1561rsquo qPCR assay two supplementary qPCR targets were used The
first one was the Rv3866 locus located in the RD1 locus (Brodin et al 2002) The second one was the
RD4 flanking region (the RD4 locus is specifically deleted in M bovis) To improve the qPCR efficiency
the RD4 based assay was derived from a previously reported qPCR assay (Sweeney et al 2007) by
modifying the reverse primer to reduce the size of the qPCR product from 142 to 116 bp (Table 3) The
RD4 based qPCR assay designed by Sweeney et al (Sweeney et al 2007) has previously been validated
for environmental matrices in several studies (Pontiroli et al 2011 Travis et al 2011 King et al 2015a
King et al 2015b) Primers and probe were designed from alignments of mycobacterial genes from
GenBank (httpwwwncbinlmnihgovgenbank) such as M bovis AF212797 four strains of M bovis
Chapitre 3
102
BCG M tuberculosis H37Rv M africanum GM041182 and Mycobacterium canettii CIPT 140010059
(respective accession numbers NC_0029453 NC_0087691 NC_0168041 NC_0122071
GCA_0004632151 NC_0009623 NC_0157581 NC_0158481) using Primer3web
(httpprimer3utee) The specificity of the assay was tested in silico by BLAST analysis
(httpblastncbinlmnihgovBlastcgi) Primers and probes were synthesized by Eurogentec (Seraing
Belgium)
Quantitative real time PCR (qPCR) and nested PCR conditions
For the three qPCR assays reactions were carried out in a 25 microl reaction mix containing 125 microl
of Absolute QPCR ROX Mix (Thermo Scientific France) at a final concentration of 1X 1 microl of T4
bacteriophage gene 32 product (final concentration 25 ng microl-1 MP Biomedicals Europe France) 1 microl of
each primer (final concentration 400 nmol l-1) 05 microl of each probe (final concentration 200 nmol l-1)
4 microl of ultrapure water and 5 microl of diluted DNAs for the environmental samples screening Absolute
quantification was achieved using standard DNA dilutions A recombinant plasmid containing one copy
of the IS1561rsquo fragment of M bovis BCG str Pasteur 1173P2 cloned in the pCRregII-TOPOreg vector
(Invitrogen France) was used as a standard A calibration curve was obtained by amplification of serial
dilutions of the plasmid ranging from 1 to 106 copies per PCR reaction All the environmental
metagenomic DNA samples were screened with the IS1561rsquo based qPCR assay and final results were
expressed as the gene copy number per g of soil feces or water qPCR assays were performed in a VIIA
7 Real-Time PCR System (Thermo Fischer Scientific France) Initial denaturation was made at 95degC for
15 min followed by 45 cycles with denaturation at 95degC for 15 s and annealing and elongation at 60degC
for 1 min
Positive DNA extracts were then tested by the nested IS1561rsquo PCR protocol as follows the first PCR was
achieved in a 25 microl reaction mixture containing 1725 microl of ultrapure water for molecular biology 25
microl of PCR Buffer 10X with MgCl2 02 microl of dNTP mix at 25 mmol l-1 each 005 microl of Taq DNA polymerase
15 U microl-1 (Taq Core Kit 25 MP Biomedicals Europe France) 125 microl of each primers (final concentration
500 nmol l-1) and 25 microl of environmental DNA The nested PCR was achieved in a Veriti Thermal Cycler
(Applied Biosystems France) with a first step of denaturation at 94degC for 5 min then 30 cycles for 30
s at 94degC for denaturation 30 s at 60degC for annealing and 30 s at 72degC for elongation and 5 min at 72degC
for final elongation The qPCR was then achieved in the same conditions as described above using as a
target a 5 microl aliquot of a 100-fold dilution of the first PCR product
Preliminary screening of all environmental samples was done using the IS1561rsquo based qPCR
assay targeting MTBC species The IS1561rsquo positive samples were then further analysed using the
Rv3866 based qPCR to confirm the presence of MTBC and the RD4 based qPCR assay to confirm the
presence of M bovis All qPCR assays were done in triplicate
Chapitre 3
103
Determination of qPCR assays analytical sensitivity in spiked soil feces or water microcosms
Microcosms consisting of 2 g of fresh soil S1 and S2 sieved to 4 mm 2 g of fresh badger feces
or 100 mL of pond water were spiked in triplicate with 100 microl of 10-fold serially diluted suspensions of
M bovis BCG strain Pasteur to reach M bovis concentrations ranging from 25 to 25 times 106 CFU per
gram of soil or feces and 50 to 50 times 106 CFU per 100 mL of water Briefly inoculum suspensions were
realized as following a thirty days M bovis BCG liquid culture was vortexed for 15 s with glass beads (4
mm diameter) in order to disrupt aggregates Optical bacterial density was then measured at 600 nm
(Biophotometre Eppendorf France) to roughly estimate mycobacterial concentration Appropriate 10-
fold dilutions (10-1 to 10-6) were prepared in sterile water M bovis concentration was estimated by
spreading in triplicate 100 microL-aliquot of 10-4 10-5 and 10-6 diluted suspension onto Middlebrook 7H11
agar enriched with OADC 10 (Sigma-Aldrich France) Plates were incubated at 37degC for two months
and then M bovis colonies were counted Results were reported as CFU per ml of suspension
The spiked microcosms were incubated at room temperature for 3-4 hours prior to DNA
extraction as described in the section 3 To assess the reproducibility all qPCR assays were done in
triplicate and performed as previously using the three detection systems targeting IS1561rsquo Rv3866 and
RD4 Non-inoculated soil feces or water used to prepare the microcosms proved to be negative with
the three qPCR systems in preliminary experiments
Estimation of DNA quality for PCR amplification
Efficiency of qPCR might be impaired by the presence of PCR inhibitors in DNA extracts The
quality of DNA extracted from soil water and feces was assessed by measuring the capacity to amplify
a control DNA target in the presence of various dilutions of soil DNA extracts Briefly qPCR reactions
were done in a final volume of 25 microl containing 25 μl of 106 copies of the circularized pGEM-T Easy
plasmid DNA and 5 μl of water (control) or diluted soil DNA (5- 10- 20-fold dilutions) 1X ABsolute
qPCR SYBR Green ROX Mix (final concentration 500 nmol l-1 Thermo Scientific France) 25 ng microl-1 of T4
bacteriophage gene 32 product qPCR primers were SP6 (5rsquo - ATTTAGGTGACACTATAG - 3rsquo) and T7 (5rsquo -
TAATACGACTCACTATAGGG - 3rsquo) universal primers (final concentration 500 nmol l-1) targeting the
polylinker of the plasmid qPCR and cycling conditions were as following initial denaturation at 95degC
for 15 min followed by 40 cycles with denaturation at 95degC for 15 s annealing at 55degC and elongation
at 72degC for 30 s each The PCR product specificity was assessed by the melting curve analysis A 10-fold
reduction of the copy number amplified by qPCR in the presence of diluted soil DNA with respect to
the copy number obtained in the control reaction (water) was considered as 10 amplification
efficiency or conversely as 90 inhibition Although inhibition tests and detection of specific targets
were conducted with different mixes and primers the DNA polymerases from the ABsolute qPCR SYBR
Chapitre 3
104
Green ROX Mix (used for inhibition testing) and the ABsolute QPCR ROX Mix (used for mycobacterial
detection in environmental samples) are the same (Thermo-Start DNA polymerase) Thus the quality
of DNAs extracted from environmental samples was estimated by this method that is compatible with
acceptable experimental costs associated with the high number of analysed samples
Direct M bovis spoligotyping from environmental DNA
Environmental DNA samples that proved to be positive for the presence of MTBC were
submitted for spoligotyping using the Luminex method described previously by Zhang et al (Zhang et
al 2010) Spoligotypes were named according to an agreed international convention
(wwwmbovisorg)
RESULTS
Validation of the qPCR assays on pure bacterial strains
The results of the specificity test for the three qPCR assays are shown in Table 1 Using IS1561rsquo-
and Rv3866-based qPCR assays all spoligotypes of M bovis were detected with cycle threshold (Ct)
values below 25 cycles as well as all strains from the other species belonging to the MTBC (M caprae
M tuberculosis M pinnipedi) except the M microti species Using the RD4-based qPCR assay only
M bovis strains were detected with Ct values below 25 cycles For the three qPCR assays the 22 strains
of the NTM and MAC members were not detected or gave Ct values above 35 cycles Strains with Ct
values above 35 cycles when using high DNA concentration from pure culture are unlikely to give any
signal when testing soil metagenomic DNA where targets are highly diluted These results indicated
that IS1561rsquo- and Rv3866-based qPCR assays are highly specific for MTBC members and exclude
M microti whereas the RD4-based qPCR assay is specific for M bovis species
The calibration curve used for absolute quantification is presented in Figure 2 for the IS1561rsquo-
based qPCR assay A linear regression was established between the log (gene copy number) and the Ct
value over a wide range of target concentrations from 10 to 106 gene copies per PCR (r2= 09989) Below
10 target gene copies per reaction the detection was still possible but results were not reproducible
The quantification limit of the qPCR detection system was estimated to be 10 gene copies per PCR
reaction thus indicating a high sensitivity Similar calibration curves and detection thresholds were
obtained for the two other qPCR assays targeting Rv3866 and RD4 (data not shown)
Chapitre 3
105
Table 1 Strains used for testing oligonucleotides specificity Results are given with Ct (Cycle Threshold) ND (not detected) means that no signal was observed after 40 cycles
Strains Ct
IS1561rsquo
Ct
Rv3866 Ct RD4
Mycobacterium bovis BCG
BCG str Pasteur 1173P2
BCG str Danish 1331
lt25 lt25 lt25
Mycobacterium bovis
SB0089 SB0120 SB0121 SB0130
SB0134 SB0162 SB0166 SB0821
SB0823 SB0825 SB0826 SB0827
SB0878 SB0885 SB0912 SB0948
SB0999 SB1081
lt25 lt25 lt25
Others MTBC members
Mycobacterium microti ND ND ND
Mycobacterium tuberculosis lt25 lt25 ND
Mycobacterium caprae lt25 lt25 ND
Mycobacterium pinnipedi lt25 lt25 ND
Non tuberculous mycobacteria
Mycobacterium avium ND ND ND
Mycobacterium intracellulare ND ND ND
Mycobacterium aichiense ND ND ND
Mycobacterium arupense ND gt35 gt35
Mycobacterium celatum gt35 ND ND
Mycobacterium chelonae ND ND ND
Mycobacterium chitae ND ND ND
Mycobacterium confluentis gt35 ND gt35
Mycobacterium elephantis ND ND ND
Mycobacterium fortuitum ND ND ND
Mycobacterium intermedium ND ND ND
Mycobacterium kansai ND gt35 ND
Mycobacterium non chromogenicum ND ND ND
Mycobacterium parafinicum gt35 ND ND
Mycobacterium porcinum ND ND ND
Mycobacterium rufum ND ND ND
Mycobacterium rutilum ND ND ND
Mycobacterium terrae ND ND ND
Mycobacterium smegmatis ND gt35 ND
Mycobacterium szulgai ND gt35 ND
Mycobacterium thermoresitibile ND ND ND
Mycobacterium vaccae ND ND ND
Mycobacterium xenopi gt35 gt35 gt35
Chapitre 3
106
Detection threshold of qPCR assays in M bovis BCG spiked environmental matrices
The detection thresholds of the three qPCR assays were evaluated in complex environmental
matrices (soil feces and water) spiked with 25 to 25 times 106 CFU per g of soil or feces and 50 to 50 times
106 CFU per 100 ml of water The quality of DNAs extracted from microcosms was assessed as described
in the MampM section ldquoEstimation of DNA quality for PCR amplificationrdquo The results showed that 20-fold
dilutions of the DNA extracts did not inhibit the qPCR assay and were thus used for all qPCR assays
performed on spiked microcosms Results of M bovis detection in spiked microcosms are given in Table
4 for IS1561rsquo-based qPCR assay No signal was detected in control samples (not inoculated)
For M bovis cell densities above 250 CFU per g of soil 2500 CFU per g of feces and 5 CFU per
g of water (or 500 CFU per 100 ml of water) the detection by the qPCR assay yielded eight to nine
positive reactions out of nine repeats which allowed determination of the numbers of M bovis gene
copy number In contrast although M bovis was sometimes detected in spiked material with lower
densities than those described above quantification was unreliable A strong discrepancy between the
results obtained by the culture-based method and those obtained with the qPCR assay was observed
(10- to 100-fold higher for the qPCR assay) All of the microcosms were analysed with the two
complementary detection systems (targeting the Rv3866 locus and the flanking region of the RD4
deletion) and similar results were obtained (data not shown) Despite soils S1 and S2 being different
based on their physico-chemical characteristics detection limits were equivalent with the three qPCR
systems on these two soils
Extrapolating these results for natural environmental samples suggest that the three qPCR
assays allow a reliable quantification of MTBC (IS1561rsquo and Rv3866) or M bovis (RD4) targets in
environmental samples for Ct values below 39 Above this threshold detection may occur but lacks
reproducibility The word ldquotracesrdquo was then chosen in all further experiments in order to indicate that
MTBC or M bovis targets are present in very low quantities
Chapitre 3
107
Figure 2 Standard curve used for absolute quantification of M bovis in samples with IS1561rsquo qPCR assay (Mean values of ten standard curves) Error bars represent the 95 confidence interval associated to the values
Table 2 Properties of the two soils used for the qPCR sensitivity detection of Mycobacterium bovis
Table 3 Sets of primers tested in this study F forward primer R reverse primer P Double dye probe
Target locus Nucleotide sequence Location Size (bp)
IS1561rsquo F 5rsquo GATCCAGGCCGAGAGAATCTG 3rsquo
R 5rsquo GGACAAAAGCTTCGCCAAAA 3rsquo
P 5rsquo FAM ACGGCGTTGATCCGATTCCGC TAMRA 3rsquo
3742298-3742319
3742354-3782374
3742332-3742353
76
Nested IS1561rsquo F 5rsquo AGGAAGCGGTGTAGGTGATG 3rsquo
R 5rsquo AAGATCGACCCGCAGTG 3rsquo
3742212-3742232
3742416-3742434 222
RD4 F 5rsquo GCAGAAGCGCAACACTCTTG 3rsquo
R 5rsquo ATCTTAGCTGGTCAATAGCCATTTTT 3rsquo
P 5rsquo FAM AGTGGCCTACAACGGCGCTCTCC 3rsquo
1692712-1692736
1692802-1692827
1692743-1692768
116
Rv3866 F 5rsquo ACCCGCCAACGTCGAA 3rsquo
R 5rsquo CAATTGACGCAATACGGTATCG 3rsquo
P 5rsquo FAM CGAGCTAGCCGAATGCACAACCG 3rsquo
4278658-4278673
4278721-4278743
4278693-4278716
86
Location is given by reference to the M bovis AF212297 genome sequence accession ndeg NC_0029453
y = -33818x + 4394Rsup2 = 09989
15
20
25
30
35
40
45
1 2 3 4 5 6
Cyc
le T
hre
sho
ld C
t
DNA concentrations [log(IS1561 copies5microL-1)]
Site Clay Loam Sand Total
Carbon
Total Nitrogen
pH H2O
Organic matter
Land use
S1 397 485 118 361 038 775 0625 Pasture S2 192 463 345 292 0269 546 0506 Pasture
Chapitre 3
108
Molecular detection of M bovis in environmental samples
Our molecular detection systems were first evaluated on twenty-four soils from bTB-free zones
in France Whatever the detection system no signal was observed showing a good specificity of the
IS1561rsquo Rv3866 and RD4 qPCR systems
Between April 2013 and December 2014 63 samples of pasture soil 150 samples of setts soil 21
latrines samples 67 water samples from naturally occurring springs (and subsequent flowing water on
soil surface) within cattle pastures and 55 samples of spring sediments within 15 areas located in the
bTB-endemic zone in Cocircte drsquoOr were analysed Some spots were sampled several times (monthly from
January to June and in November) to assess the persistence of the MTBC signals Altogether 356 DNA
samples were primarily screened using the qPCR assay targeting IS1561rsquo locus Molecular signals
corresponding to MTBC were found in six areas out of 15 analysed and only positive results are
presented in Table 5 Apparent gene copy numbers were the average value from n = 2 to n = 3 repeats
These results indicate a MTBC contamination of the environment in 40 of the studied areas
Among the 36 positive samples 19 were found to be positive for MTBC detection with low
signals (indicated as ldquoTracesrdquo in Table 5) These samples include 7 badger sett soils 3 badger latrines
1 sediment sample and 8 water samples from naturally occurring springs in pastures More
interestingly high positive signals for MTBC were observed for the other 17 positive samples with 58
to 8 700 gene copies per g of soil It is worth noting that these samples only included badger sett soils
in area D and one nematodes sample in area C while no signal was recorded for the pasture soils
In order to confirm these positive results some of them were subjected to the nested IS1561rsquo
qPCR assay Eleven samples including eight badger sett soils two samples of water from springs and
one sample of nematodes isolated from spring sediments were confirmed to be positive for MTBC
detection with this nested qPCR assay Badger sett soils from area D were sampled five times over an
eight month-period The occurrence of MTBC signal was discontinuous in the seven soils from badger
setts that were sampled However MTBC signals were detected in several badger sett soils (badger sett
soils 1 4 and 5 in Table 5) following a seven to eight months period after M bovis was detected for the
first time The IS1561rsquo gene copy numbers for the positive badger sett soils ranged levels from
254 times 102 to 505 times 104 per g of soil The MTBC signal remained at high levels throughout the eight
month-period although the badgers moved out one month after the first sampling of the sett (Table
5) Furthermore confirmation of M bovis detection using the RD4 qPCR detection system was achieved
for a few IS1561rsquo positive samples Only three out of eight tested samples proved to be positive with
RD4-based detection system (see Table 5) This result definitely demonstrates the occurrence of M
bovis DNA in these samples
Spoligotyping was performed on positive DNA samples and the results are shown in Table 6 A
badger sett soil DNA was fully spoligotyped and found to belong to the SB0120 M bovis spoligotype
Chapitre 3
109
which is responsible for cattle and wildlife infection in this area of Cocircte drsquoOr Partial M bovis
spoligotypes were obtained for three badger sett soils DNAs The two other partial spoligotypes did
not allow identification of the species within the MTBC
Table 4 Evaluation of the detection of the IS1561rsquo qPCR assay with DNA extracted from control and spiked test matrices
Traces means that IS1561rsquo signal was detected but that quantification was unreliable (Ct value above 39)
Number of M bovis spiked per g of dry soil or per 100 mL of water
Number of positive PCR 9
reactions
Mean Ct Standard deviation Ct
Mean IS1561rsquo genome-equivalent per g of soil or feces or 100 mL of water
Soil S1
Control soil 0 ND ND ND
25 1 396 ND Traces 25101 1 394 ND Traces 25102 8 389 1 (654 plusmn 366) times103
25103 9 345 05 (217 plusmn 086) times105
25104 9 302 02 (218 plusmn 026) times106
25105 9 276 05 (132 plusmn 039) times107
Soil S2
Control soil 0 ND ND ND 25 0 ND ND ND 25101 5 404 06 Traces 25102 9 381 07 (800 plusmn 319) times103
25103 9 342 04 (141 plusmn 049) times105
25104 9 296 05 (374 plusmn 161) times106
25105 9 267 07 (265 plusmn 185) times107
Water Control water 0 ND ND ND 5101 1 401 ND Traces 5102 9 381 11 (220 plusmn 145) times104
5103 9 351 05 (148 plusmn 058) times105
5104 9 31 16 (320 plusmn 134) times106
5105 9 259 03 (819 plusmn 209) times107
5106 9 228 03 (698 plusmn 131) times108
Badger feces
Control feces 0 ND ND ND 25101 0 ND ND ND 25102 1 399 ND Traces 25103 9 379 105 (232 plusmn 077) times104
25104 9 347 159 (212 plusmn 187) times105
25105 9 312 203 (244 plusmn 286) times106
25106 9 261 059 (336 plusmn 107) times107
Chapitre 3
110
Table 5 Molecular detection of MTBC or M bovis targets in positive environmental samples from 6 areas in Cocircte drsquoOr Only positive samples are presented MTBC gene copy numbers were computed from calibration curve and amounts of DNA extracted from the various samples (n=2 ou n=3)
Area
Type of sample Sampling date Mean MTBC gene copy numbers g-1 of soil or feces (IS1561rsquo qPCR assay)
Confirmation after IS1561rsquo-nested PCR
Confirmation of presence of M bovis after RD4 qPCR assay
A Badger sett soil June 2013 Traces Negative NT dagger
B Spring water 1 January 2014 February 2014 March 2014
Traces
Traces
Traces
Negative Negative Negative
Negative Negative Negative
C Spring water 1 Spring water 2 Sediments Soil nematodes
February 2014 February 2014 February 2014 February 2014
Traces Traces Traces 582101
Positive Negative Negative Positive
NT
NT
NT
NT
D Badger sett soil 1 March 2014 November 2014
193103 577103
Positive NT
Positive NT
Badger sett soil 2 March 2014 May 2014 June 2014
870103 502102 143103
Positive Positive NT
Positive NT
NT Badger sett soil 3 March 2014
May 2014 June 2014
441102
741103 282103
Positive Positive NT
Negative NT
NT Badger sett soil 4 April 2014
May 2014 June 2014 November 2014
446103
236103 502102 607103
Positive NT
NT
NT
Negative NT
NT
NT Badger sett soil 5 April 2014
May 2014 June 2014 November 2014
248103
254102
202103
360103
Positive NT NT
NT
Positive NT
NT
NT Badger sett soil 6 November 2014 Traces NT NT Badger sett soil 7 November 2014 Traces NT NT Latrine 1 March 2014 Traces Negative NT Latrine 2 March 2014 Traces Negative NT
E Spring water 1 March 2014 March 2014 March 2014
Traces Negative NT Spring water 2 Traces Positive NT Spring water 3 Traces Negative NT Badger sett soil 1 March 2014
March 2014 Traces Negative NT
Badger sett soil 2 Traces Negative NT Latrine 1 March 2014 Traces Negative NT
F Badger sett soil 1 June 2014 June 2014
Traces Negative NT Badger sett soil 2 Traces Positive NT
Traces MTBC targets were present but MTBC genome equivalent were not calculated since Ct were close to the detection limit of the method dagger NT not tested
Chapitre 3
111
Table 6 Spoligotyping results of DNA positive samples extracted from environmental matrices Black squares represent positive hybridization signals and dots represent lack of hybridization
Figure 3 Spatial location of MTBC positive water and sediment samples in a pasture (site C) along the run off of one naturally occurring springs Star 1 indicates the sampling zone of positive spring sediment and nematodes sample Stars 2 and 3 indicate the 2 sampling sites of positive source water Contour lines are superimposed on aerial photography (450 m and 425 m contour lines are visible 5 m elevation interval between 2 consecutive lines)
Area Type of sample
Sampling date Hybridization pattern Spoligotype
D Badger sett soil 2
March 2014 M bovis SB0120
D Badger sett soil 1
March 2014 Partial M bovis spoligotype
D Badger sett soil 3
March 2014 Partial M bovis spoligotype
D Badger sett soil 5
November 2014
Partial M bovis spoligotype
C Water February 2014
Partial Mycobacterium spoligotype
Chapitre 3
112
DISCUSSION Mycobacterium bovis is described as an obligate intracellular microorganism transmitted by
direct contact between mammalian hosts However direct contacts between species seem to be scarce
and indirect transmission involving environmental matrices is likely to occur (Boumlhm et al 2009
Kukielka et al 2013) Many studies have clearly demonstrated the presence and the persistence of
M bovis in naturally infected environmental substrates in bTB endemic regions of United Kingdom
(Young et al 2005 Courtenay et al 2006 Sweeney et al 2006 Sweeney et al 2007 King et al 2015a
King et al 2015b) Evaluation of the environment as a reservoir of M bovis could help to understand
its epidemiological significance and its role in the indirect transmission of the disease among livestock
and wildlife To our knowledge the results from our study demonstrate for the first time the
contamination of environmental matrices with MTBC and more specifically M bovis in Cocircte drsquoOr which
is a region where bTB is endemic in cattle and wildlife
qPCR detection is a method of choice for M bovis environmental monitoring since it is a faster
alternative to culture based methods that are hampered by the slow-growing rate of M bovis and the
high number and diversity of microbial communities Moreover PCR is able to detect M bovis DNA in
the environment over a long period following initial contamination with a relatively high sensitivity
(Adams et al 2013)
The detection of M bovis in environmental samples (soil water and feces) that contain highly
diverse microbial communities including numerous described and potentially non-described NTM
(Falkinham 2009) was still a challenge The two qPCR assays developed in this study (IS1561rsquo and
Rv3866 targets) proved to be specific for the MTBC excluding M microti These results are in agreement
with the fact that IS1561rsquo is present in the genomes of all the members of the MTBC except in M microti
(Gordon et al 1999 Brodin et al 2002 Huard et al 2003) Itrsquos worth noting that M microti is quite
widespread in animals in France (Michelet et al 2015) and thus may also occur in environmental
matrices (Courtenay et al 2006) The refined RD4 qPCR assay was shown to be highly specific for
M bovis species in agreement with previous results (King et al 2015a) Although some strains gave Ct
gt 35 when concentrated in pure culture with these molecular assays these targets would not be
detected in natural samples due to extreme dilution in highly complex environmental DNAs
The DNA extraction method used in this study proved to be highly efficient in soils harboring
different physico-chemical characteristics (especially in clayey soils) (Dequiedt et al 2011 Locatelli et
al 2013) The large mass of samples (2 g) allowed a better representativeness of sampling to overcome
the potential heterogeneity of substrates ie a physical heterogeneity as well as a heterogeneous
distribution of mycobacteria in environmental samples The use of an amplification facilitator (T4
bacteriophage gene 32 product) allowed minimizing PCR inhibition by co-extracted contaminants
(Matheson et al 2010 Opel et al 2010) and significantly improving M bovis detection In previous
Chapitre 3
113
published studies (Travis et al 2011 Pontiroli et al 2011 Sweeney et al 2007) DNA extraction was
usually achieved from smaller masses (01 to 05 g) of soil or feces using commercial DNA extraction kit
in order to limit PCR inhibition
The three qPCR assays developed in this study using 20-fold dilutions of DNA extracts of spiked
samples allowed a reliable detection of spiked M bovis (BCG strain) at levels as low as 25 times 102 CFU to
25 times 103 CFU g-1 of sample providing an improved detection limit compared to a recent study where
detection limit was 40 times 104 CFU g-1 soil and 50 times 105 CFU ml-1 water (Santos et al 2015) In other
previous studies detection limits were estimated to 85 times 103 cells g1 for soil and 425 times 105 cells g1 for
fecal samples using the original RD4 qPCR assay (Pontiroli et al 2011 Travis et al 2011) As cell numbers
were estimated by fluorescent dye assisted flow cytometry comparison with our results is difficult
Finally King et al (King et al 2015a King et al 2015b) were able to detect 103 M bovis genome
equivalents per g-1 for badger feces
In our study the detection limits are equivalent for the three qPCR systems developed or refined
whatever the physico-chemical characteristics of the soils
The 10- to 100-fold discrepancy between bacterial numbers obtained using culture based methods and
qPCR based methods that we observed could be explained by the underestimation of cultivable
bacteria due to cell aggregation (linked to a strong hydrophobicity of mycobacteria cell wall) in
inoculum as well as the occurrence of viable but not cultivable cells (VBNC) and cell mortality (with
persistence of DNA) Similar discrepancies have already been observed in previous studies (eg Betelli
et al 2013)
Though similar detection thresholds were observed for the three systems in spiked samples
RD4 and RV3866 based qPCR assays seemed less sensitive than IS1561rsquo-based detection system in
environmental samples (range of 2 Ct between IS1561rsquo and the 2 other systems) However the
occurrence of RD4 based positive signals in several samples gave an additional proof of the specific
detection of M bovis Furthermore the direct spolygotyping of DNA extracted from environmental
matrices confirmed for the first time the presence of M bovis in several environmental samples in
France
All 63-pasture soil samples from the bTB endemic area in Cocircte drsquoOr were found to be negative
for the presence of MTBC (below the detection threshold) Failure to detect MTBC in pasture soil
samples can be related to the low prevalence of bTB in cattle in the sampling areas (herd prevalence
reached 08 in 2014 Cote drsquoOr Regional Veterinary Service) and the low prevalence of bTB infection
in badgers which territories overlap the sampled pastures (Payne et al 2012) This result might reflect
the high dilution rate of M bovis targets in soil DNA extracts which contains high numbers of other
microorganisms (abundant soil microbial communities containing up to 1010 genomes per g of soil)
The discontinuous (spatial and temporal) distribution of M bovis in soil pasture because of
Chapitre 3
114
intermittent andor low shedding (Neill et al 1988 Clifton-Hadley et al 1993) might also limit the
detection of M bovis in soil samples Despite the high number of soil sample analyzed financial and
time constraints have limited more intensive sampling efforts that could have overcome this limit
MTBC signal was detected in 101 (7 positive out of 67) of the water samples taken from
naturally occurring springs with the IS1561rsquo qPCR assay As the level of detection was close to the
detection threshold we were unable to quantify MTBC signals Interestingly one sample of nematodes
originated from sediments located in close vicinity of one of the positive sources was also found
positive for the presence of MTBC Although nematodes are known to host bacterial pathogen no data
is available concerning MTBC This intriguing result would require further investigations These water
springs (and accompanying flowing water) are located in pastures where both cattle and wildlife have
access for drinking In several pastures (four sites) water was sampled along the slope following the
gravitational water flow In two of these pastures we repeatedly found (two to three) positive samples
indicating multiple point contamination of spring and flowing water along these slopes (see Figure 3
for localization of positive samples in one of these pastures) This result is an important new finding of
our study that can support several hypotheses on the role of water in the dissemination of MTBC in
the environment and in animal contamination We can hypothesize from these results that bioaerosols
formation from contaminated water and subsequent inhalation by animals (cattle and wildlife) might
contribute to animal infection with MTBC members especially as the infective dose through this
transmission route is very low (lt 10 CFU) (Dean et al 2005 Corner et al 2007) Infection of animals
might also occur through oral route by repeatedly drinking contaminated water
A special effort was made to monitor the contamination of badger sett soils since bTB infected
badgers were detected in our study area Between 2009 and 2011 878 badgers had been trapped and
analyzed in the infected area of Cocircte drsquoOr (Payne et al 2012) Macroscopic lesions were present in 19
of them despite culture-based method detecting 56 of positive badgers Only one badger showed
widespread tuberculosis Moreover in this infected area numerous badger setts were located either
within pastures or in close vicinity (forest-pasture interface) Our study demonstrated an average
prevalence of MTBC of 73 (11 out of 150) in soils from badger setts with the highest gene copy
number up to 37 times 104 gene copies g-1 of soil observed among environmental samples analysed in our
study The prevalence of MTBC molecular signature in latrines was estimated to 7 (3 positive out of
21) Our results demonstrate that infected badgers might contaminate their close environment for the
first time in France Indeed infected badgers have been shown to excrete M bovis from the respiratory
digestive and urinary tracts and in bite wounds exudates (Gavier-Widen et al 2001 Corner et al 2012)
Our results are in agreement with previous findings from several studies in UK (Young et al 2005
Courtenay et al 2006 Sweeney et al 2007 King et al 2015a) that recovered M bovis from badger
setts latrines and adjacent fields with gene copy numbers ranging from 1 times 103 to 54 times 106 per g of
Chapitre 3
115
soil In Woodchester Park in UK where badgers populations have been extensively studied for more
than 20 years bTB prevalence was estimated around 30 in 2005 which might explain high numbers
of positive setts and latrines observed in this area (Courtenay et al 2006 King et al 2015a) As
suggested by Courtenay et al (Courtenay et al 2006) the low bTB prevalence of infected sett soils
observed in our study may be linked to the low prevalence and the low density of badgers excreting
the bacteria within their social group
Finally in soils of 7 badger setts we were able to detect M bovis signatures (RD4 positive qPCR)
either continuously (4 setts) or discontinuously (3 setts) The presence of M bovis was confirmed by
direct soil DNA spolygotyping (see Table 6) in soils of the setts The presence of M bovis molecular
signatures was proved over an eight month-period (despite the badgers moved out) These results are
in agreement with a previous study where positive molecular detection of M bovis was demonstrated
over a period of 15 months despite the badgers having moved out (Young et al 2005) To our
knowledge this is the first time that spoligotype SB0120 was observed by direct spoligotyping of soil
DNA this spoligotype being identical to the spoligotype of strains isolated from cattle and wildlife in
these sites (Payne et al 2012)
Although the molecular detection of MTBC or M bovis by specific qPCR assay doesnrsquot give any
information about mycobacterial viability Young et al (Young et al 2005) demonstrated that MTBC DNA
did not persist in soil for more than ten days suggesting that DNA detected in our environmental
samples could belong to viable cells
A remaining challenge is to develop adequate methods to cultivate M bovis from
environmental matrices Indeed culture based methods might also be impaired by the occurrence of
VBNC state or dormancy in M bovis present in environmental matrices as previously suggested by
Whittington et al (Whittington et al 2004) for Mycobacterium avium subsp paratuberculosis Further
work is needed to assess if VBNC or dormancy occur in M bovis under environmental conditions
Although we concentrated our sampling efforts on badger environment it is however
acknowledged that in Cocircte drsquoOr M bovis circulates in a multi-host system that includes also wild boar
and deer We can thus hypothesize that these wild animals can also contribute to the shedding of
M bovis in the environment
Further research is needed to understand the ecology of M bovis in the environment and its
epidemiological role in indirect transmission This knowledge would help to improve herd
management in reducing indirect contacts between livestock and wildlife in controlling water access
in areas considered as ldquoat riskrdquo and in preventing contacts with badger setts for example Biosecurity
should thus be an integral part of the long-term strategy for the management of bovine tuberculosis in
Cocircte drsquoOr
Chapitre 3
116
Acknowledgements
The authors thank F Chevalier and M Beacuteral from the CIREV (Cellule Inter-Reacutegionale
drsquoEpideacutemiologie Veacuteteacuterinaire) AE Taconet F Bartheacuteleacutemy and T Grosjean from the Cote drsquoOr Regional
Veterinary Service (DDPP21) We are grateful to the cattle breeders trappers and hunters who helped
us during this study We wish also to thank C Ferrier from ONCFS (National Game Hunting and Wildlife
Agency) Dominique Meunier and Luc Biju-Duval from INRA Dijon Ceacuteline Richomme from ANSES The
work was supported by the Conseil Reacutegional de Bourgogne and Conseil Geacuteneacuteral de Cocircte drsquoOr Interbev
GDS Cocircte drsquoOr
Conflict of interest
No conflict of interest declared
References
Adams AP Bolin SR Fine AE Bolin CA and Kaneene JB (2013) Comparison of PCR versus culture for detection of Mycobacterium bovis after experimental inoculation of various matrices held under environmental conditions for extended periods Appl Environ Microbiol 79 6501-6506
Aranaz A De Juan L Montero N Saacutenchez C Galka M Delso C Alvarez J Romero B Bezos J Vela AI Briones V Mateos A and Domiacutenguez L (2004) Bovine tuberculosis (Mycobacterium bovis) in wildlife in Spain J Clin Microbiol 42 2602-2608
Betelli L Duquenne P Grenouillet F Simon X Scherer E Gehin E and Hartmann A (2013) Development and evaluation of a method for the quantification of airborne Thermoactinomyces vulgaris by real-time PCR J Microbiol Methods 92 25-32
Biet F Boschiroli ML Thorel MF and Guilloteau LA (2005) Zoonotic aspects of Mycobacterium bovis and Mycobacterium avium-intracellulare complex (MAC) Vet Res 36 411-436
Boumlhm M Hutchings MR and White PCL (2009) Contact networks in a wildlife-livestock host community identifying high-risk individuals in the transmission of bovine TB among badgers and cattle PLoS One 4 e5016
Brodin P Eiglmeier K Marmiesse M Billault A Garnier T Niemann S Cole ST and Brosch R (2002) Bacterial artificial chromosome-based comparative genomic analysis identifies Mycobacterium microti as a natural ESAT-6 deletion mutant Infect Immun 70 5568-5578
Clifton-Hadley RS Wilesmith JW and Stuart Fa (1993) Mycobacterium bovis in the European badger (Meles meles) epidemiological findings in tuberculous badgers from a naturally infected population Epidemiol Infect 111 9-19
Corner LA OMeara D Costello E Lesellier S and Gormley E (2012) The distribution of Mycobacterium bovis infection in naturally infected badgers Vet J 194 166-172
Corner LA Costello E Lesellier S OMeara D Sleeman DP and Gormley E (2007) Experimental tuberculosis in the European badger (Meles meles) after endobronchial
Chapitre 3
117
inoculation of Mycobacterium bovis I Pathology and bacteriology Res Vet Sci 83 53-62
Courcoul A Moyen J-L Brugegravere L Faye S Heacutenault S Gares H and Boschiroli M-L (2014) Estimation of sensitivity and specificity of bacteriology histopathology and PCR for the confirmatory diagnosis of bovine tuberculosis using latent class analysis PLoS One 9 e90334
Courtenay O Reilly LA Sweeney FP Hibberd V Bryan S Ul-Hassan A Newman C Macdonald DW Delahay RJ Wilson GJ and Wellington EMH (2006) Is Mycobacterium bovis in the environment important for the persistence of bovine tuberculosis Biol Lett 71 1946-1952
Dean GS Rhodes SG Coad M Adam O Cockle PJ Clifford DJ Hewinson RG and Vordermeier HM (2005) Minimum infective dose of Mycobacterium bovis in cattle Infect Immun 73 6467-6471
Dequiedt S Saby NPa Lelievre M Jolivet C Thioulouse J Toutain B Arrouays D Bispo a Lemanceau P and Ranjard L (2011) Biogeographical patterns of soil molecular microbial biomass as influenced by soil characteristics and management Global Ecol Biogeogr 20 641-652
Falkinham JO (2009) Surrounded by mycobacteria nontuberculous mycobacteria in the human environment J Appl Microbiol 107 356-367
Fediaevsky A Courcoul A Boschiroli M-L and Reveillaud E (2014) Tuberculose bovine en France reacutesultats dune strateacutegie plus offensive Bull Epid Santeacute Anim Alim 64 4-11
Fine AE (2006) The role of indirect transmission in the epidemiology of bovine tuberculosis in cattle and white-tailed deer in Michigan Michigan Bovine Tuberculosis Bibliography and DatabasePaper httpdigitalcommonsunledumichbovinetb33
Fine AE Bolin Ca Gardiner JC and Kaneene JB (2011) A study of the persistence of Mycobacterium bovis in the environment under natural weather conditions in Michigan USA Vet Med Int 2011
Garnett BT Delahay RJ and Roper TJ (2002) Use of cattle farm resources by badgers (Meles meles) and risk of bovine tuberculosis (Mycobacterium bovis) transmission to cattle Proc R Soc Lond B Biol Sci 269 1487-1491
Gavier-Widen D Chambers MA Palmer N Newell DG and Hewinson RG (2001) Pathology of natural Mycobacterium bovis infection in European badgers (Meles meles) and its relationship with bacterial excretion Vet Rec 148 299-304
Ghodbane R Mba Medie F Lepidi H Nappez C and Drancourt M (2014) Long-term survival of tuberculosis complex mycobacteria in soil Microbiology 160 496-501
Gordon SV Heym B Parkhill J Barrell B and Cole ST (1999) New insertion sequences and a novel repeated sequence in the genome of Mycobacterium tuberculosis H37Rv Microbiology 145 881-892
Gortaacutezar C Torres MJ Vicente J Acevedo P Reglero M de la Fuente J Negro JJ and Aznar-Martiacuten J (2008) Bovine tuberculosis in Dontildeana Biosphere Reserve The role of wild ungulates as disease reservoirs in the last Iberian lynx strongholds PLoS One 3 e2776
Chapitre 3
118
Hauer A Cruz KD Cochard T Godreuil S Karaoui C Henault S Bulach T Banuls AL Biet F and Boschiroli M-L (2015) Genetic evolution of Mycobacterium bovis causing tuberculosis in livestock and wildlife in France since 1978 PLoS One 10 e0117103
Huard RC de Oliveira Lazzarini LC Butler WR van Soolingen D and Ho JL (2003) PCR-based method to differentiate the subspecies of the Mycobacterium tuberculosis complex on the basis of genomic deletions J Clin Microbiol 41 1637-1650
Huard RC Fabre M de Haas P Lazzarini LCO van Soolingen D Cousins D and Ho JL (2006) Novel genetic polymorphisms that further delineate the phylogeny of the Mycobacterium tuberculosis complex J Bacteriol 188 4271-4287
King HC Murphy A James P Travis E Porter D Hung Y-J Sawyer J Cork J Delahay RJ Gaze W Courtenay O and Wellington EM (2015a) The variability and seasonality of the environmental reservoir of Mycobacterium bovis shed by wild European badgers Sci Rep 5 1-7
King HC Murphy A James P Travis E Porter D Sawyer J Cork J Delahay RJ Gaze W Courtenay O and Wellington EM (2015b) Performance of a noninvasive test for detecting Mycobacterium bovis shedding in European badger (Meles meles) populations J Clin Microbiol 53 2316-2323
Kukielka E Barasona JA Cowie CE Drewe JA Gortazar C Cotarelo I and Vicente J (2013) Spatial and temporal interactions between livestock and wildlife in South Central Spain assessed by camera traps Prev Vet Med 112 213-221
Little TW Naylor PF and Wilesmith JW (1982) Laboratory study of Mycobacterium bovis infection in badgers and calves Vet Rec 111 550-557
Locatelli A Depret G Jolivet C Henry S Dequiedt S Piveteau P and Hartmann A (2013) Nation-wide study of the occurrence of Listeria monocytogenes in French soils using culture-based and molecular detection methods J Microbiol Methods 93 242-250
Matheson CD Gurney C Esau N and Lehto R (2010) Assessing PCR inhibition from humic substances Open Enzym Inhib J 3 38-45
Michelet L de Cruz K Zanella G Aaziz R Bulach T Karoui C Heacutenault S Joncour G and Boschiroli ML (2015) Infection with Mycobacterium microti in animals in France J Clin Microbiol 53 981-985
Morris RS Pfeiffer DU and Jackson R (1994) The epidemiology of Mycobacterium bovis infections Vet Microbiol 40 153-177
Neill S Hanna J OBrien J and McCracken R (1988) Excretion of Mycobacterium bovis by experimentally infected cattle Vet Rec 123 340-343
Oostenbrink M (1960) Estimating nematode populations by some selected methods Nematology 6(8) 165-2015
Opel KL Chung D and McCord BR (2010) A study of PCR inhibition mechanisms using real time PCR J Forensic Sci 55 25-33
Palmer MV Waters WR and Whipple DL (2004) Investigation of the transmission of
Chapitre 3
119
Mycobacterium bovis from deer to cattle through indirect contact Am J Vet Res 65 1483-1489
Payne A (2014) Rocircle de la faune sauvage dans le systegraveme multi-hocirctes de Mycobacterium bovis et risque de transmission entre faune sauvage et bovins eacutetude expeacuterimentale en Cocircte drsquoOr In Epidemiologie Universiteacute Claude Bernard Lyon 1 httpwwwthesesfr2014LYO10040
Payne A Boschiroli ML Gueneau E Moyen JL Rambaud T Dufour B Gilot-Fromont E and Hars J (2012) Bovine tuberculosis in ldquoEurasianrdquo badgers (Meles meles) in France Eur J Wildlife Res 59 331-339
Phillips CJC Foster CRW Morris PA and Teverson R (2003) The transmission of Mycobacterium bovis infection to cattle Res Vet Sci 74 1-15
Pontiroli A Travis ER Sweeney FP Porter D Gaze WH Mason S Hibberd V Holden J Courtenay O and Wellington EMH (2011) Pathogen quantitation in complex matrices a multi-operator comparison of DNA extraction methods with a novel assessment of PCR inhibition PLoS One 6 e17916
Radomski N Roguet A Lucas FS Veyrier FJ Cambau E Accrombessi H Moilleron R Behr MA and Moulin L (2013) atpE gene as a new useful specific molecular target to quantify Mycobacterium in environmental samples BMC Microbiol 13 277
Richomme C Boschiroli ML Hars J Casabianca F and Ducrot C (2010) Bovine tuberculosis in livestock and wild boar on the Mediterranean island Corsica J Wildl Dis 46 627-631
Santos N Santos C Valente T Gortazar C Almeida V and Correia-Neves M (2015) Widespread environmental contamination with Mycobacterium tuberculosis Complex revealed by a molecular detection protocol PLoS One 10 e0142079
Slana I Kaevska M Kralik P Horvathova A and Pavlik I (2010) Distribution of Mycobacterium avium subsp avium and M a hominissuis in artificially infected pigs studied by culture and IS901 and IS1245 quantitative real time PCR Vet Microbiol 144 437-443
Sweeney FP Courtenay O Hibberd V Hewinson RG Reilly LA Gaze WH and Wellington EMH (2007) Environmental monitoring of Mycobacterium bovis in badger feces and badger sett soil by Real-Time PCR as confirmed by immunofluorescence immunocapture and cultivation Appl Environ Microbiol 73 7471-7473
Sweeney FP Courtenay O Ul-Hassan A Hibberd V Reilly LA and Wellington EMH (2006) Immunomagnetic recovery of Mycobacterium bovis from naturally infected environmental samples Lett Appl Microbiol 43 364-369
Telenti A Marchesi F Balz M Bally F Bottger EC and Bodmer T (1993) Rapid identification of mycobacteria to the species level by polymerase chain reaction and restriction enzyme analysis J Clin Microbiol 31 175-178
Thornton CG MacLellan KM Brink TL Lockwood DE Romagnoli M Turner J Merz WG Schwalbe RS Moody M Lue Y and Passen S (1998) Novel method for processing respiratory specimens for detection of mycobacteria by using C-18-carboxypropylbetaine Blinded study J Clin Microbiol 36 1996-2003
Travis ER Gaze WH Pontiroli A Sweeney FP Porter D Mason S Keeling MJC Jones RM
Chapitre 3
120
Sawyer J Aranaz A Rizaldos EC Cork J Delahay RJ Wilson GJ Hewinson RG Courtenay O and Wellington EMH (2011) An inter-laboratory validation of a real time PCR assay to measure host excretion of bacterial pathogens particularly of Mycobacterium bovis PLoS One 6 e27369
Whittington R Marshall D Nicholls PJ Marsh IB and Reddacliff LA (2004) Survival and dormancy of Mycobacterium avium subsp paratuberculosis in the environment Appl Environ Microbiol 70 2989-3004
Young JS Gormley E and Wellington EM (2005) Molecular detection of Mycobacterium bovis and Mycobacterium bovis BCG (Pasteur) in soil Appl Environ Microbiol 71 1946-1952
Zanella G Durand B Hars J Moutou F Garin-Bastuji B Duvauchelle A Ferme M Karoul C and Boschiroli ML (2008) Mycobacterium bovis in wildlife in France J Wildl Dis 44 99-108
Zhang J Abadia E Refregier G Tafaj S Boschiroli ML Guillard B Andremont A Ruimy R and Sola C (2010) Mycobacterium tuberculosis complex CRISPR genotyping improving efficiency throughput and discriminative power of spoligotyping with new spacers and a microbead-based hybridization assay J Med Microbiol 59 285-294
Chapitre 3
121
C CAMPAGNES DE PRELEVEMENTS SUPPLEMENTAIRES
Entre deacutecembre 2014 et mars 2015 deux campagnes de preacutelegravevement drsquoeacutechantillons
environnementaux suppleacutementaires ont eacuteteacute reacutealiseacutees dans la zone endeacutemique de Cocircte drsquoOr ainsi
qursquoune campagne en Charente ougrave des foyers bovins et sauvages sont recenseacutes respectivement depuis
2006 et 2009
1 Fin des preacutelegravevements en Cocircte drsquoOr
Les preacutelegravevements ont concerneacute drsquoune part une forecirct domaniale agrave cheval sur deux communes
de la zone Sud (eacutetoiles blanches sur la Figure 19) dans laquelle nous nous sommes focaliseacutes sur les
trous drsquoeau freacutequenteacutes par les sangliers et les cervideacutes Cinq trous drsquoeau importants et freacutequenteacutes ont
eacuteteacute cibleacutes et sur chacun trois preacutelegravevements de solsseacutediments et deux preacutelegravevements drsquoeau ont eacuteteacute
reacutealiseacutes en deacutecembre 2014 Drsquoautre part des preacutelegravevements ont eacuteteacute effectueacutes dans les terriers de
blaireaux infecteacutes de la commune D citeacutee dans lrsquoarticle preacuteceacutedent que nous suivons depuis mars 2014
et dans laquelle 1 agrave 3 blaireaux infecteacutes sont pieacutegeacutes tous les ans ainsi que dans des terriers freacutequenteacutes
drsquoune commune limitrophe (commune G) sur laquelle un blaireau infecteacute a eacuteteacute pieacutegeacute en 2014 Quatorze
sols de terriers (5 sur la commune D et 9 sur la commune G) et cinq latrines proches des terriers (3 sur
D et 2 sur G) ont eacuteteacute preacuteleveacutes en mars 2015 Ces deux sites sont repreacutesenteacutes par les eacutetoiles noires sur
la Figure 19 Tous ces eacutechantillons ont eacuteteacute analyseacutes selon les techniques deacutecrites dans lrsquoarticle
preacuteceacutedent (section Mateacuteriel et meacutethodes)
Les vingt-cinq eacutechantillons preacuteleveacutes dans la forecirct domaniale de la zone Sud en deacutecembre 2014
nrsquoont donneacute aucun signal moleacuteculaire avec le systegraveme de deacutetection ciblant lrsquoIS1561rsquo Dans cette zone
forestiegravere aucun animal sauvage infecteacute nrsquoa eacuteteacute deacutetecteacute depuis 2013
Concernant les terriers et latrines de la commune D les cinq eacutechantillons de sols de terriers
analyseacutes et une des deux latrines ont donneacute un signal moleacuteculaire positif pour le MTBC Pour lrsquoun des
sols la quantification nrsquoa pas eacuteteacute possible avec la qPCR (Ct gt 39 traces) Pour les quatre autres sols
des valeurs de 10 times 104 agrave 46 times 104 copies de gegravenes ont eacuteteacute obtenues et de 12 times 104 copies de gegravenes
pour la latrine Trois des eacutechantillons de sol ont donneacute un reacutesultat positif avec la PCR nicheacutee ciblant
lrsquoIS1561rsquo Dans ces terriers les preacutelegravevements reacutealiseacutes en mars avril mai juin et novembre 2014 puis
mars 2015 ont montreacute une persistance du signal moleacuteculaire au cours du temps Pour quatre des
terriers le signal a eacuteteacute deacutetecteacute agrave chaque campagne de preacutelegravevements alors que pour trois autres la
deacutetection a eacuteteacute intermittente Il est inteacuteressant de noter que les valeurs obtenues en mars 2015 sont
supeacuterieures agrave celles obtenues en 2014 (voir article preacuteceacutedent) Plusieurs hypothegraveses sont eacutemises pour
Chapitre 3
122
expliquer ce reacutesultat La premiegravere est lieacutee au fait que les terriers eacutetaient habiteacutes lors de nos
preacutelegravevements en mars 2015 alors qursquoils ne lrsquoeacutetaient plus apregraves les preacutelegravevements de mars 2014
La surveillance videacuteo reacutealiseacutee sur ces terriers a montreacute que les terriers ont agrave nouveau eacuteteacute
occupeacutes agrave partir de septembre 2014 Il est possible que cette occupation ait geacuteneacutereacute lrsquoexteacuteriorisation de
sol profond et de litiegravere potentiellement plus contamineacutes et que nous ayons preacuteleveacute ce sol drsquoorigine
profonde La seconde hypothegravese est lieacutee au fait que nous avons preacuteleveacute plus profondeacutement dans le
terrier que dans les campagnes de preacutelegravevements preacuteceacutedentes (utilisation drsquoun aspirateur) Enfin il est
eacutegalement possible que le groupe de blaireaux peuplant ce terrier possegravede un ou plusieurs animaux
infecteacutes et excreacuteteurs agrave lrsquoorigine drsquoune contamination plus importante du sol du terrier Ces terriers sont
situeacutes dans le bosquet drsquoun preacute qui fournit aux bovins de lrsquoombre et de la fraicheur en eacuteteacute et un abri en
cas de pluie ou de vent La surveillance videacuteo de cette zone nous a permis drsquoobserver que plusieurs
veaux ont reacuteguliegraverement exploreacute les entreacutees de gueules et consommeacute les brins drsquoherbe autour de ces
entreacutees (surveillance videacuteo)
Sur les neuf eacutechantillons de sol preacuteleveacutes dans la commune G deux ont donneacute un signal positif
pour le MTBC Pour lrsquoun drsquoeux seules des traces de M bovis ont eacuteteacute deacutetecteacutees pour lrsquoautre le nombre
de copies de gegravenes a eacuteteacute estimeacute agrave 12 times 104 La PCR nicheacutee srsquoest reacuteveacuteleacutee neacutegative pour les deux
eacutechantillons
Chapitre 3
123
Figure 19 Localisation des communes de Cocircte drsquoOr sur lesquelles de foyers bovins et des animaux sauvages infecteacutes ont eacuteteacute deacutetecteacutes et localisation des preacutelegravevements environnementaux effectueacutes en 2015 (les preacutelegravevements positifs sont repreacutesenteacutes par des eacutetoiles noires) (Reacutealisation Marina Beacuteral CIREV Bourgogne)
2 Preacutelegravevements en Charente
Depuis 2006 25 foyers bovins ont eacuteteacute deacutetecteacutes en Charente avec 12 foyers pour la seule anneacutee
2014 et la deacutecouverte de foyers anciens qui eacutevoluaient agrave bas bruit (communication personnelle
DDPP16) Depuis 2009 lrsquoinfection est mise en eacutevidence chez les blaireaux (6 positifs en 2013 et 11 en
2014) et les sangliers (8 positifs en 2014 donneacutees Sylvatub) et un chevreuil en 2015 (Communication
personnelle Sylvatub) autour des foyers bovins avec les mecircmes spoligotypes Le systegraveme multi-hocirctes
observeacute diffegravere donc de celui de la Cocircte drsquoOr puisqursquoagrave lrsquoheure actuelle lrsquoinfection nrsquoa pas eacuteteacute observeacutee
chez les cerfs dont les densiteacutes sont beaucoup moins importantes qursquoen Cocircte drsquoOr Les foyers bovins
Chapitre 3
124
et sauvages sont majoritairement localiseacutes dans le sud du deacutepartement dans une zone limitrophe de
la Dordogne deacutepartement fortement infecteacute depuis 2005 (Figure 20)
A la demande de la DDPP16 nous sommes alleacutes preacutelever des eacutechantillons environnementaux
dans le Sud du deacutepartement dans sept communes fortement toucheacutees par la maladie (Figure 20) Entre
avril et mai 2015 55 eacutechantillons environnementaux (50 eacutechantillons de sol de terrier de blaireau 3
sols de souille 1 latrine de blaireau et 1 fegraveces de sanglier) ont eacuteteacute preacuteleveacutes sur 7 communes de
Charente Ces communes font lrsquoobjet de la deacuteclaration drsquoun ou plusieurs foyers bovins de tuberculose
bovine etou drsquoanimaux sauvages infecteacutes (Figure 20) De plus 3 eacutechantillons de matiegraveres feacutecales
preacuteleveacutes pendant lrsquoautopsie de 3 blaireaux (BLAIR 7057 BLAIR 7058 et BLAIR 7061) ont eacuteteacute analyseacutes
Six eacutechantillons positifs ont eacuteteacute mis en eacutevidence sur les 55 analyseacutes La qPCR ciblant lrsquoIS1561rsquo a permis
de deacutetecter et de quantifier M bovis dans quatre ADN de sol de terrier avec un nombre de copies de
gegravenes variant de 11 times 103 agrave 54 times 103 par g de sol Deux eacutechantillons suppleacutementaires (1 sol de souille
et 1 sol de terrier) ont donneacute un signal non quantifiable avec la qPCR (traces) Tous les eacutechantillons
positifs ont eacuteteacute preacuteleveacutes sur une mecircme commune sur 2 lieux-dits diffeacuterents Maine-Joli (4 sols de
terrier) et la Tuilerie (1 sol de terrier et 1 sol de souille) A notre connaissance 2 blaireaux et 3 sangliers
infecteacutes ont eacuteteacute deacutecouverts agrave proximiteacute du lieu-dit la Tuilerie deacutebut 2015 (Source DDPP16) Lors de
notre campagne de preacutelegravevement tous les terriers deacutetecteacutes positifs eacutetaient freacutequenteacutes
Les ADN des eacutechantillons positifs ont eacuteteacute envoyeacutes agrave lrsquoANSES pour geacutenotypage Aucun ADN nrsquoa toutefois
pu ecirctre spoligotypeacute
Les fegraveces de blaireaux (BLAIR 7057 BLAIR 7058 et BLAIR 7061) preacuteleveacutes lors des autopsies nrsquoont donneacute
aucun signal avec la qPCR
Chapitre 3
125
Figure 20 Localisation des communes de Charente sur lesquelles de foyers bovins et des animaux sauvages infecteacutes ont eacuteteacute deacutetecteacutes et localisation des preacutelegravevements environnementaux effectueacutes en 2015 (les preacutelegravevements positifs sont repreacutesenteacutes par des eacutetoiles noires) (Reacutealisation Marina Beacuteral CIREV Bourgogne)
3 Compleacutements paysage geacuteomorphologie et peacutedologie
La zone endeacutemique Nord de tuberculose bovine en Cocircte drsquoOr que nous avons principalement
eacutetudieacutee est situeacutee sur un territoire appeleacute lrsquoAuxois caracteacuteriseacute par un paysage bocager ponctueacute de
buttes et de plateaux calcaires ougrave affleure un calcaire dur et karstique Les pacirctures suspectes
preacutesentent un contexte paysager hydrologique et geacuteomorphologique commun Le parcellaire suspect
est localiseacute dans trois petites valleacutees parallegraveles au fond desquelles circule un cours drsquoeau en direction
du Nord-Ouest (Figure 21) Chaque cours drsquoeau est alimenteacute par des sources naissant entre les couches
calcaires et marneuses du sous-sol dans les pentes Ces reacutesurgences drsquoeau proviennent de lrsquoinfiltration
des preacutecipitations dans les plateaux calcaires agrave lrsquoorigine du deacuteveloppement drsquoun reacuteseau karstique
drainant drsquoimportants eacutecoulements souterrains et la reacutesurgence drsquoeau en surface agrave cause des marnes
impermeacuteables du sous-sol Les pacirctures sont situeacutees sur les pentes douces marneuses des buttes ainsi
que dans les valleacutees alluviales humides Au-dessus des pacirctures un bourrelet boiseacute marque la rupture
Chapitre 3
126
de pente il preacutesente en geacuteneacuteral des affleurements plus ou moins importants de calcaire dur (Figure
22) Les pacirctures sont majoritairement situeacutees sur la mecircme formation geacuteologique des marnes du Lias
ou du Jurassique infeacuterieur correspondant agrave des formations peacutedologiques particuliegraveres caracteacuteriseacutees
par des sols de pentes argilo-limoneux localement hydromorphes (sources et laquo mouilles raquo)
Il est inteacuteressant drsquoobserver que les cantons toucheacutes par la tuberculose bovine en Cocircte drsquoOr
possegravedent une densiteacute de bovins allaitant quasi-identique agrave celle des cantons indemnes localiseacutes dans
le sud-ouest du deacutepartement et limitrophe de la zone drsquoendeacutemie de bTB (Figure 23) Ces deux groupes
de cantons sont localiseacutes sur des formations peacutedologiques diffeacuterentes suggeacuterant que le type de sol des
pacirctures pourrait ecirctre un facteur de risque dans le maintien de la maladie et que la densiteacute des bovins
ne semble pas ecirctre un facteur majeur dans la distribution des foyers de bTB Lrsquoobservation de la
distribution du parcellaire des exploitations foyers de tuberculose bovine sur la carte peacutedologique du
deacutepartement montre que 74 des parcelles de ces exploitations sont localiseacutees sur cette uniteacute
paysagegravere (sols argilo-limoneux et hydromorphes des pentes 48 des parcelles sols calcaires humides
sur alluvions des valleacutees 14 et sols argileux des plateaux calcaires 12 ) (Figure 24Figure 24)
Une enquecircte (Etude PatRisk) reacutealiseacutee dans le cadre drsquoun Master drsquoEpideacutemiologie dans neuf
eacutelevages bovins deacuteclareacutes infecteacutes entre 2010 et 2014 a chercheacute agrave identifier les pacirctures laquo agrave risque raquo de
tuberculose et agrave les caracteacuteriser par des variables concernant les pratiques drsquoeacutelevage le voisinage la
faune sauvage et lrsquoenvironnement (pente type de sol relief exposition humiditeacute) Lrsquoanalyse
multivarieacutee a mis en eacutevidence deux expositions indicatrices de risque la pratique du fauchage avant
la mise agrave lrsquoherbe et la preacutesence de zones humides localiseacutees sur la pacircture (Bouchez-Zacria 2015) Ces
reacutesultats sont en accord avec le fait que la survie de M bovis est favoriseacutee par la preacutesence drsquohumiditeacute
et qursquoune faible hauteur drsquoherbe augmente la possibiliteacute de contact avec les mycobacteacuteries du sol Bien
que des eacutetudes plus preacutecises soient recommandeacutees il semble toutefois que les sols argilo-limoneux
hydromorphes des pacirctures suspectes constituent un facteur de risque agrave prendre en compte dans
lrsquoeacutepideacutemiologie de la maladie
Chapitre 3
127
Figure 21 Orographie et hydrographie de lrsquoAuxois Les valleacutees supportant les parcelles suspectes sont indiqueacutees par une flegraveche noire (Source BD Carthage et BD alti - IGN)
Figure 22 Localisation de lrsquoAuxois en Cocircte drsquoOr structure paysagegravere et geacuteomorphologie de ce territoire
Chapitre 3
128
Figure 23 Nombre de vaches allaitantes par canton dans le deacutepartement de la Cocircte drsquoOr (agrave gauche) et parcellaire des foyers bovins de 2010 agrave 2015 (agrave droite) (Source Recensement agricole 2010 ndash Agreste)
Chapitre 3
129
Figure 24 Reacutepartition spatiale des parcelles des eacutelevages bovins infecteacutes de 2010 agrave 2014 en Cocircte drsquoOr et repreacutesentation des couches peacutedologiques (Reacutealisation Marina Beacuteral CIREV Bourgogne)
Chapitre 3
130
D BILAN GLOBAL
Nous avons deacuteveloppeacute deux systegravemes de deacutetection moleacuteculaire adapteacutes aux matrices
environnementales ciblant les espegraveces du MTBC (IS1561rsquo et Rv3866) et modifieacute un systegraveme existant
speacutecifique de M bovis (RD4) Nous avons valideacute ces systegravemes sur trois matrices diffeacuterentes (sol eau
fegraveces) inoculeacutees avec M bovis BCG en deacutemontrant leur speacutecificiteacute soit pour le MTBC (excluant
M microti potentiellement preacutesent dans lrsquoenvironnement) soit pour M bovis Leur sensibiliteacute est
bonne puisque les seuils de deacutetection obtenus sont infeacuterieurs aux seuils deacutecrits dans la litteacuterature (en
comparant avec une meacutethode de deacutenombrement identique agrave la nocirctre) Le systegraveme RD4 est agrave ce jour le
seul systegraveme de deacutetection speacutecifique de M bovis dans les eacutechantillons environnementaux toutefois
drsquoapregraves notre expeacuterience il preacutesente une sensibiliteacute moins importante que les deux autres systegravemes
malgreacute des seuils de deacutetection eacutequivalents sur les eacutechantillons inoculeacutes au laboratoire
La deacutetection du MTBC par lrsquoamplification de lrsquoIS1561rsquo a eacuteteacute effective dans 42 eacutechantillons (sur
plus de 400) la PCR nicheacutee ciblant la mecircme seacutequence a mis en eacutevidence 14 eacutechantillons positifs et le
systegraveme RD4 trois eacutechantillons positifs Les niveaux de deacutetection varient fortement en fonction des
eacutechantillons lrsquoeffet de dilution reste tregraves important dans des matrices comportant des communauteacutes
microbiennes abondantes et diversifieacutees (quantiteacute relative drsquoADN cible faible par rapport agrave des
quantiteacutes eacuteleveacutees drsquoADN non cible) Lrsquoutilisation de la PCR quantitative a permis dans certains cas de
quantifier les cibles preacutesentes dans les eacutechantillons environnementaux Les sols de terriers preacutesentent
les concentrations en mycobacteacuteries du MTBC les plus eacuteleveacutees pouvant atteindre 46 times 104 copies du
gegravene cibleacute ce qui correspond agrave des quantiteacutes de cellules eacuteleveacutees sachant que le nombre de copies du
gegravene cible par geacutenome est de 1
Parmi les diffeacuterents sites preacuteleveacutes la contamination a eacuteteacute retrouveacutee exclusivement dans deux
eacutecosystegravemes particuliers les zones humides de pacirctures (sources et laquo mouilles raquo) et les sols de terrier
et latrines de blaireaux La contamination apparait le plus souvent localiseacutee agrave une zone geacuteographique
et ponctuelle mais un signal moleacuteculaire positif a eacuteteacute retrouveacute pendant un an dans les sols de terriers
de blaireaux localiseacutes sur une pacircture
Ces reacutesultats correacuteleacutes au contexte paysager et peacutedologique des parcelles suspectes nous ont
ameneacute agrave penser que certaines formations peacutedologiques seraient plus susceptibles de favoriser la
persistance de M bovis Nous avons donc eacutevalueacute lrsquoimpact de la nature du sol retrouveacute dans ces
parcelles sur la survie de M bovis Nous avons eacutegalement eacutevalueacute lrsquoimpact de la tempeacuterature sur la
dureacutee de survie de la bacteacuterie dans le sol
CHAPITRE 4
Impact de la tempeacuterature et de la nature du sol sur la
survie de Mycobacterium bovis SB0120
Elodie Barbier Murielle Rochelet Laurent Gal Maria Laura Boschiroli Alain Hartmann
Experimental investigation of the impact of the temperature and the soil type on Mycobacterium
bovis survival in the environment
(Article en cours de soumission)
133
CHAPITRE 4 - Impact de la tempeacuterature et de la nature du sol sur la
survie de Mycobacterium bovis SB0120
A AVANT-PROPOS
La deacutetection moleacuteculaire de Mycobacterium bovis dans les eacutecosystegravemes agricoles de la zone
endeacutemique de Cocircte drsquoOr a confirmeacute que les animaux infecteacutes et excreacuteteurs ont la capaciteacute de
contaminer les sols et lrsquoeau accessibles agrave la faune domestique et sauvage (Chapitre 2) La bacteacuterie ne
se multipliant pas dans lrsquoenvironnement sa survie dans les substrats environnementaux conditionne la
transmission indirecte du bacille entre un animal infecteacute excreacuteteur et un animal sain par le biais de ces
substrats
Des eacutetudes expeacuterimentales sur divers substrats environnementaux notamment du sol
inoculeacutes avec M bovis ou M bovis BCG ont suggeacutereacute que les principaux facteurs influenccedilant la survie
des bacteacuteries sont les facteurs climatiques tels que tempeacuterature humiditeacute et exposition aux UV (Fine
et al 2011 Duffield amp Young 1985 Jackson et al 1995 Young et al 2005) Bien que quelques eacutetudes
aient rechercheacute par la culture ou par des approches statistiques une eacuteventuelle association de la
contamination environnementale ou animale par des mycobacteacuteries avec la nature du sol les reacutesultats
sont difficilement comparables et extrapolables (Walter et al 2012 Dhand et al 2009 Reviriego et al
2000 Johnson-Ifearulundu amp Kaneene 1999 Ward amp Perez 2004)
Face agrave ce manque de connaissances nous avons donc souhaiteacute eacutevaluer la dureacutee de la survie de
M bovis dans deux sols de Cocircte drsquoOr preacutesentant des caracteacuteristiques physico-chimiques contrasteacutees et
ce agrave deux tempeacuteratures diffeacuterentes afin de rechercher un eacuteventuel impact de la tempeacuterature et de la
nature du sol sur la persistance de la bacteacuterie Pour cette eacutetude nous avons utiliseacute une souche
pathogegravene de M bovis (geacutenotype SB0120) ce geacutenotype est retrouveacute majoritairement dans la zone
Nord chez les bovins et dans la faune sauvage Deux meacutethodes de suivi de la population bacteacuterienne
ont eacuteteacute utiliseacutees la culture sur milieu geacuteloseacute reacutealiseacutee agrave partir de sols irradieacutes et la qPCR pour le suivi
des sols irradieacutes et biotiques
135
B EXPERIMENTAL INVESTIGATION OF THE IMPACT OF THE TEMPERATURE AND THE SOIL TYPE ON MYCOBACTERIUM BOVIS SURVIVAL IN THE ENVIRONMENT
Elodie Barbierab Murielle Rocheletab Laurent Galac Maria Laura Boschirolid Alain Hartmanna
a INRA UMR 1347 Agroeacutecologie Dijon Cedex France
b Universiteacute de Bourgogne Franche Comteacute Dijon Cedex France
c AgroSup Dijon UMR1347 Agroeacutecologie Dijon France
d Universiteacute Paris-Est Laboratoire National de Reacutefeacuterence de la Tuberculose Uniteacute des Zoonoses Bacteacuteriennes Laboratoire de
Santeacute Animale ANSES Maisons-Alfort Cedex France
Key words Mycobacterium bovis soil temperature culture quantitative PCR
Corresponding Author Alain Hartmann UMR 1347 Agroeacutecologie INRA 17 rue Sully BP 86510 21065
Dijon Cedex France E-mail alainhartmanndijoninrafr
INRA UMR1347 Agroeacutecologie BP 86510 F-21000 Dijon France
Chapitre 4
136
Abstract
Mycobacterium bovis the causative agent of the bovine tuberculosis (bTB) mainly affect cattle its
natural reservoir but also a wide range of domestic and wild mammals Besides direct transmission via
contaminated aerosols indirect transmission of the disease between wildlife and livestock is thought
to occur by inhalation or ingestion of environmental substrates contaminated by M bovis through
infected animal shedding We monitored the survival of M bovis in two soils chosen for their contrasted
physico-chemical properties at two temperatures The population of M bovis spiked in disinfected soils
was enumerated by a culture-based method after 14 30 60 90 120 and 150 days of incubation at 4degC
and 22degC The fate of M bovis spiked in disinfected or biotic soils was monitored using a qPCR based
assay targeting the IS1561rsquo locus The analysis of survival profiles using culture-based method showed
that M bovis survived longer at cooler temperature but impact of soil characteristics on M bovis
persistence was not obvious Futhermore qPCR based assay detected M bovis longer than the culture
based method with higher gene copy numbers in sterile soils than in biotic ones Impact of soil type on
M bovis persistence need to be further studied in order to shed light on the risk and mechanisms of
indirect transmission of the disease
Chapitre 4
137
INTRODUCTION
Mycobacterium bovis is a pathogenic mycobacteria responsible for bovine tuberculosis (bTB)
Tuberculosis caused by M bovis is a primarily respiratory disease that may also affect various organs in
animals (Morris et al 1994) Even if bTB mainly concerns cattle a wide range of domestic and wild
mammals may be infected as well (Biet et al 2005) On the other hand bTB a zoonotic disease
impacting human health (Peacuterez-Lago et al 2013) Although cattle is considered as the principal host of
M bovis wild fauna such as badgers (Meles meles) wild boar (Sus scrofa) and red deer (Cervus elaphus)
has recently been identified as potential wildlife reservoirs (Palmer 2013) Though the transmission of
M bovis among cattle usually occurs via the inhalation of contaminated aerosols released by infected
animals andor the close contacts (Morris et al 1994 Phillips et al 2003) the indirect transmission
either by inhalation of environmental bioaerosols or ingestion of infected environmental matrices may
be another potential routes of animal infection (Vicente et al 2007 Palmer et al 2004 Phillips et al
2003) Indirect transmission through feed or water has already been demonstrated between deer deer
and cattle (Palmer et al 2004 Palmer et al 2001) and badgers and cattle (Little et al 1982) in laboratory
experiments
Investigating this type of indirect transmission is challenging because it results at least from the
combination of three essential factors iei) the environmental contamination by shedding from
infected animals ii) the persistence of the bacteria under a virulent state in environmental matrices
and finally iii) the interaction between a new susceptible host with the contaminated matrices The
shedding step by animals has already been demonstrated for many species via oro-nasal mucus
sputum urine feces and wound discharges depending on the animal species (Corner et al 2012
Palmer et al 2004 Barasona et al 2015) Secondly a few previous experimental studies using various
environmental substrates spiked with M bovis showed that the bacteria was able to survive for a long
time period outside of its host (Williams amp Hoy 1930 Duffield amp Young 1985 Young et al 2005 Fine et
al 2011 Ghodbane et al 2014) For example using a culture-based method Ghodbane et al (2014)
recovered M bovis bacteria from sterile soils incubated in controlled laboratory conditions 12 months
after their inoculation whereas an incubation under the natural weather conditions of Michigan USA
allowed the persistence of the bacteria in sterile soils for 88 days (Fine et al 2011) Moreover several
studies have identified several climatic factors such as a cool temperature an adequate moisture and
a protection against solar radiation (ultra violet) as positive factors for the persistence of M bovis in
environmental matrices like feces (Williams amp Hoy 1930 Tanner amp Michel 1999) food supply such as
corn hay apples (Palmer amp Whipple 2006 Fine et al 2011) water (Little et al 1982 Fine et al 2011)
and soil (Duffield amp Young 1985 Fine et al 2011) (Duffield amp Young 1985 Williams amp Hoy 1930 Tanner
amp Michel 1999 Jackson et al 1995) When Fine et al (2011) compared the persistence of M bovis in
Chapitre 4
138
four substrates (corn hay water and soil) the longest survival was found in cool and moist soil (88
days in winterspring) suggesting that soil probably ensured better conditions of persistence for
M bovis than the other matrices tested Moreover M bovis was often detected in soils especially in
badger sett soil or in pasture soil (Courtenay et al 2006 Santos et al 2015 Sweeney et al 2007
2006 Young et al 2005 Barbier et al manuscript submitted)
More generally low pH and high iron content are often described as major factors driving
mycobacterial survival in soils especially for Mycobacterium avium ssp paratuberculosis (Johnson-
Ifearulundu amp Kaneene 1999 Ward amp Perez 2004) Concerning M bovis one statistical study
established a relationship between the occurrence of bTB in animals and soil parameters by showing
that every a 1 increase in the sandy content of soil led to an increase in odds of bTB infection by 4
(Walter et al 2014) suggesting that soil type may promote or hinder M bovis environmental
persistence
To our knowledge no comprehensive study focuses on the influence of soil biotic and abiotic
parameters on the persistence M bovis With the aim to further assess the role of the soil
characteristics on the persistence of Mbovis and thus to better understand the impact of the
environment in the epidemiology of the bTB disease the influence of both the physicochemical
characteristics of the soil and the temperature was studied under controlled laboratory conditions For
this purpose two different soils selected for their contrasted properties were spiked with known
concentrations of M bovis and incubated at two different temperatures to mimic seasonal
temperatures The persistence of M bovis over time was monitored using both a culture-based
method for the detection of viable M bovis and a molecular detection method based on a qPCR assay
MATERIAL AND METHODS
All experiments were conducted in a biosafety level 3 laboratory unless otherwise stated
Mycobacterium bovis cultivation and inoculum preparation
The strain Mycobacterium bovis SB0120 VNTR 5 5 4 3 11 4 5 6 that infects cattle and wild
animals such as badgers and wild boar in France was provided by the National Reference Laboratory of
bovine tuberculosis (Maisons-Alfort France) and grown aerobically at 37degC in Middlebrook 7H9
supplemented with 10 Middlebrook ADC Growth Supplement (Sigma-Aldrich France) for thirty days
The M bovis inoculum was prepared as following a 20 mL stationary phase culture was pelleted and
once the supernatant removed the cell aggregates were disrupted by vortexing with glass beads for 15
seconds The pellet was suspended in 1 mL of sterile water constituting the stock solution and serial
10-fold dilutions were subsequently plated in triplicate onto Middlebrook 7H11 supplemented with
10 heat inactivated bovine serum (Dominique Dutscher France) and 10 Middlebrook OADC
Chapitre 4
139
Growth Supplement (Sigma-Aldrich France) for enumeration Colonies were counted following
incubation for two months at 37degC The inoculum for the culture-based assay and the qPCR-based assay
were respectively prepared by diluting 10-fold and 1000-fold the M bovis stock solution
Soil characterization and microcosm preparation
Two different soils were collected in two cattle-grazed pastures at 0-30 cm depth in Cocircte drsquoOr
(a department located in the Middle East of France) The soil A is a clay loamy soil (calcisol) and soil B
is a loamy sandy soil Soils were prepared as following soil samples were gently air dried and sieved to
recover 4 mm aggregates The moisture contents of soils were determined by drying for 24 h at 105degC
and their physico-chemical parameters were determined by the Soil Analysis Laboratory of Arras
France Fresh sieved soil was used to prepare the microcosms
Vials of 15 mL (Falcon Dominique Dutscher France) were filled with 2 g of the soils A and B Half of the
soil vials were sterilized by gamma-irradiation up to an internal minimum dose of 35 KGray (Ionisos
France) All the microcosm soils (biotic and sterile) were then adjusted to 90 of the water holding
capacity We verified that neither soil A nor B were positive for M bovis with the culture-based method
(sterile soils) and with the qPCR assay (sterile and biotic soils) described later All the soil microcosms
were spiked with 100 microL of the M bovis inoculum 10- or 1000-fold diluted (for sterile and biotic soil
respectively) and stored in the dark in a biosafety level 3 laboratory until their analysis Sterile spiked
soils were incubated at 4degC and 22degC while biotic spiked soils were only incubated at 22degC Vials were
opened in a microbiological safety cabinet every fifteen days to regenerate their atmosphere
Schedule of microcosm sampling
In order to monitor M bovis cell numbers over time both plate enumeration and qPCR assay
were performed on irradiated soils while qPCR assay only was carried out for biotic soils At each
sampling date ie day 0 14 30 60 90 120 and 150 three mesocosms of each biotic and sterile soils
were analysed with the qPCR assay after DNA extractionpurification and four mesocosms of each
sterile soil at each temperature were analysed using culture-based method
DNA extraction and purification
DNAs from the soil samples were extracted as follows 8 mL of lysis buffer (100 mM Tris pH 80
100 mM EDTA pH 80 100 mM NaCl and 2 (wv) SDS) 4 g of silica beads (100 μm diameter) 5 g of
ceramic beads (14 mm diameter) and 8 glass beads (4 mm diameter) were added to each vial of soil
However as DNA extraction could not be achieved within the biosafety level 3 laboratory for practical
reason soil was suspended in 8 mL of lysis buffer and heated 2 hours at 80degC to kill viable bacteria The
following steps of DNA extraction were then done in a biosafety level 2 laboratory Samples were
Chapitre 4
140
disrupted for 3 times 30 s at 4 ms in a FastPrepreg-24 Instrument (MP Biomedicals Europe France)
incubated for 30 min at 70degC and centrifuged for 5 min at 7000 timesg at room temperature Supernatants
were incubated on ice for 10 min with 110 volume of 3 M potassium acetate pH 55 and then
centrifuged for 5 min at 14000 times g One volume of ice-cold isopropanol was added to the supernatant
for DNA precipitation overnight at minus 20degC DNA was collected by centrifugation (30 min at 14000 times g)
and DNA pellets were washed with ice-cold ethanol (70 ) and dissolved in 100 μL of water Crude DNA
extracts (100 μL) were loaded on Microbiospintrade columns (Biorad France) filled with 100 mg of
polyvinyl polypyrrolidone (PVPP Sigma Chemical Co France) After centrifugation (4 min at 1000 times g
4 degC) eluted DNA was further purified using the Geneclean Turbo Kit (Qiagen France) according to the
manufacturers instructions Purified DNA concentrations were estimated by spectrophotometry (λ =
260 nm) using a NanoDropreg 2000 spectrophotometer (Thermo Scientific France)
Estimation of DNA quality for PCR amplification
The quality of the purified DNAs extracted from the two soils was assessed by measuring the
capacity to amplify a control DNA target in the presence of various dilutions of soil DNA extracts Briefly
qPCR reactions were done in a final volume of 25 microL containing 25 μl of 106 copies of the circularized
pGEM-T Easy plasmid DNA and 5 μl of water (control) or diluted soil DNA (10- and 20-fold dilutions)
1X Absolute qPCR SYBR Green ROX (500 nM) Mix (Thermo Scientific) 25ngmicroL of T4 bacteriophage
gene 32 product qPCR primers were SP6 (5rsquondashATTTAGGTGACACTATAG-3rsquo) and T7 (5rsquo-
TAATACGACTCACTATAGGG-3rsquo) universal primers (final concentration 500 nM) targeting the polylinker
of the plasmid qPCR and cycling conditions were as following initial denaturation at 95degC for 15 min
followed by 40 cycles with denaturation at 95degC for 15 s annealing at 55degC and elongation at 72degC for
30 s each The analysis of the melting curves allowed to assess PCR specificity A tenfold reduction of
the copy number amplified by qPCR in the presence of diluted soil DNA with respect to the copy
number obtained in the control reaction (water) was considered as 10 amplification efficiency or
conversely as 90 inhibition of the PCR
Cuture-based method
The sterile soil microcosms were suspended in 9 mL of Middlebrook 7H9 supplemented with
ADC by vortexing the vials for 2 min Tenfold dilutions of soil suspensions were achieved and 100 microL
were plated in triplicate onto Middlebrook 7H11 agar enriched as described above and supplemented
with antibiotics and antifungal (100 mgL-1Ticarcillin 10 mgL-1 Trimethoprim 200 000 UIL-1 Polymyxin
B and 100 mgL-1 Cycloheximid Sigma-Aldrich France) for enumeration Plates were then incubated in
gas-permeable bags at 37degC for 2 months and checked every 15 days
Chapitre 4
141
Mycobacterium tuberculosis complex quantitative PCR (qPCR)
Mycobacterium bovis was detected using the Mycobacterium tuberculosis complex (MTBC)
specific qPCR targeting the IS1561rsquo locus (Forward 5rsquo- GATCCAGGCCGAGAGAATCTG -3rsquo Reverse 5rsquo-
GGACAAAAGCTTCGCCAAAA - 3rsquo and probe 5rsquo - FAM ACGGCGTTGATCCGATTCCGC TAMRA - 3rsquo) as
previously described (Barbier et al manuscript submitted for publication)
qPCR reactions were carried out in triplicate with a 25 microL reaction mix containing 125 microL of ABsolutetrade
QPCR ROX Mix (Thermo Scientific France) 1 microL (final concentration 25 ngmicroL-1) of T4 bacteriophage
gene 32 product (MP Biomedicals Europe France) 1 microL of each primer (final concentration 400 nmolL-
1) 05 microL of each probe (final concentration 200 nmol L-1) 4 microL of ultrapure water and 5 microL of 20-fold
diluted DNA Absolute quantification was achieved using standard DNA dilutions A recombinant
plasmid containing one copy of the IS1561rsquo fragment of M bovis BCG strain Pasteur 1173P2 cloned in
the pCRregII-TOPOreg vector (Invitrogen France) was used as a standard A calibration curve was obtained
by amplification of serial dilutions of the plasmid ranging from 1 to 106 copies per PCR reaction All
DNAs extracted from the soil samples were screened with the IS1561rsquo-based qPCR assay and results
were expressed as the number of M bovis gene copy number per g of soil qPCR assay was performed
in a VIIA 7 Real-Time PCR System (Thermo Fischer Scientific France) Initial denaturation was done at
95degC for 15 min followed by fourty-five cycles with denaturation at 95degC for 15 s annealing and
elongation at 60degC for 1 min Detection limits of this molecular system was previously determined in
soil (Barbier et al manuscript submitted for publication)
RESULTS
Physico-chemical characteristics of soils A and B
The physico-chemical characteristics of soils A and B are summarized in Table 1 The soil A is a
clay loamy soil with a slightly basic pH (775) and a high content of clay (clay account for 397 in
particle-size distribution and sand for only 118 ) while the soil B is a loamy sandy soil with an acidic
pH (546) and a high content of sand (sand account for 345 in particle-size distribution and clay for
only 192 ) Silt content was roughly equivalent in the two soils (485 g kg-1 for soil A vs 463 g kg-1 for
soil B) We chose to adjust soil moisture to 90 of their water field holding capacity leading to a
moisture content of 31 for soil A and 21 for soil B Finally the C-to-N ratio was 95 for soil A and
109 for soil B
Chapitre 4
142
Table 1 Physico-chemical parameters of soils A and B
Physical parameters Soil A Soil B Unit
Clay (lt 2 microm) 397 192 g kg-1
Silt 485 463 g kg-1
Sand 118 345 g kg-1
Water field capacity 352 247 g kg-1
Chemical parameters -
pH 775 546
Total C 361 292 g kg-1
Total N 38 269 g kg-1
C- to N- ratio 95 109 -
Organic matter 625 506 g kg-1
P 0031 0025 g kg-1
K 0158 00877 g kg-1
Ca 717 11 g kg-1
Na 00115 00129 g kg-1
Mg 00541 00789 g kg-1
Fe 0526 0227 g kg-1
CECa 183 108 cmol+kg-1b
aCEC Cation Exchange Capacity bcmol+kg-1 centimoles of positive cations per kg of soil
Effect of temperature and soil type on M bovis survival time measured by culture-based method
Theoretical detection limit of our culture-based method was estimated at 15 times 103 CFU g-1 soil
Total cultivable M bovis (CFU g-1 soil) were enumerated by the culture-based method described above
for each sterile soil incubated at 4degC and 22degC at day 0 14 30 60 90 120 and 150 At day 0 number
of M bovis was estimated at 15 (plusmn 06) times 107 CFU g-1 for soil A and 17 (plusmn 07) times 107 CFU g-1 for soil B by
the culture-based method
Two different patterns of mycobacterial survival were observed depending on the temperature
(Figure 1a and 1b) At 22degC M bovis cells decreased over the course of the experiment in the two
sterile soils A and B (Figure 1a) No more cultivable bacteria growth was observed from day 90 in soil B
and day 120 in soil A to the end of the experiment at day 150 Although M bovis SB0120 was detected
by culture for one month longer in soil A than in soil B no statistically significant difference of CFU
numbers was observed between the two soils at 22degC (Student-Newman-Keuls Test p = 0559) thus
no significant difference in M bovis survival was observed due to soil type
At 4degC M bovis cultivable cells were recovered in high concentrations in the two soils at every
sampling date (over the five months of the experiment) The M bovis CFU numbers remained stable
during the first 30 days in the two soils as indicated in Figure 1b We observed a slight decrease of the
Chapitre 4
143
CFU numbers in the two soils starting from day 60 to the end of the study In both soils M bovis CFU
numbers decreased significantly from day 0 to day 150 (Student-Newman-Keuls Test plt0001)
reaching 87 (plusmn 25) times 10 5 CFU g- 1 and 22 (plusmn 11) times 10 5 CFU g- 1 in soil A and B respectively
Effect of soil type and endogenous microbiota on M bovis survival time measured by qPCR assay
Survival of M bovis when biotic or sterile soils A and B were incubated at 22degC was monitored
using the qPCR based assay The M bovis gene copy numbers were estimated with IS1561rsquoqPCR-based
method for each sterile and biotic soils incubated at 22degC at day 0 14 30 60 90 120 and 150 (see
Figure 2) At day 0 number of M bovis gene copies per g of soil yielded 36 (plusmn 12) times 105 and 60 (plusmn 19)
times 105 respectively for sterile soils A and B and 12 (plusmn 07) times 105 and 25 (plusmn 15) times 105 respectively for
biotic soils A and B by the qPCR-based method
Overall high amounts of M bovis gene copies were detected in the four soils at the eight sampling
dates
A statistically significant interaction between time and soil condition (biotic vs sterile) was
observed when considering the two soils independently and dependently (Student-Newman-Keuls
Test plt0001) with higher amounts of gene copies recovered in sterile soils than in biotic ones from
day 0 to day 150 No significant differences were observed when comparing the gene copy numbers in
the two sterile soils or in the two biotic soils at each sampling date
Even though the M bovis gene copy numbers slightly oscillated in the four soils over the course of the
experiment a slight but significant decrease of these numbers was observed between day 0 and day
150 for each soil The highest decreases of M bovis were observed in the biotic soils with a rough loss
of 1 log for soil B and 075 log for soil A between day 0 and day 150 (Figure 2)
Finally qPCR assay allowed a longer M bovis detection in sterile soils A and B incubated at 22degC
than culture-based assay Indeed M bovis DNA was detected by qPCR for up to 150 days in the two
sterile soil samples while bacterial culture detected viable M bovis cells up to 60 days in soil D and 90
days in soil C
Chapitre 4
144
Figure 1 Detection of M bovis SB0120 by culture-based method in sterile soils A and B after incubation at 22degC (figure above) and 4degC (figure below) Error bars represent the standard deviation values on 8 replicates
Figure 2 Detection of M bovis gene copy numbers by IS1561rsquo qPCR-based assay on sterile and biotic soils A and B after incubation at 22degC Error bars represent standard deviation values on 9 replicates
10E+00
10E+01
10E+02
10E+03
10E+04
10E+05
10E+06
10E+07
10E+08
0 14 30 60 90 120 150
M b
ovi
s C
FU n
um
ber
per
g o
f
ster
ile s
oil
Sampling day
22degC
Soil A
Soil B
10E+00
10E+01
10E+02
10E+03
10E+04
10E+05
10E+06
10E+07
10E+08
0 14 30 60 90 120 150
M b
ovi
s C
FU n
um
ber
per
g o
f st
erile
so
il
Sampling day
4degC
Soil A
Soil B
10E+00
10E+01
10E+02
10E+03
10E+04
10E+05
10E+06
0 14 30 60 90 120 150
M b
ovi
s IS
15
61
co
py
nu
mb
er p
er g
of
ster
ile s
oil
Sampling day
Sterile soil A Sterile soil B Soil A Soil B
Chapitre 4
145
DISCUSSION
Contamination of the environment with M bovis is considered as essential for the persistence
of bovine tuberculosis in animals and its interspecific indirect transmission The survival of this
mycobacteria in soil has been documented (Duffield amp Young 1985 Jackson et al 1995 Fine et al 2011
Young et al 2005) but limited information is available on the soils used and the edaphic factors that
may affect M bovis persistence are poorly understood These experiments have dealt with controlled
or environmental parameters that promote or hinder M bovis persistence in soil such as temperature
moisture or UV exposition but to our knowledge impact of the soil type has not been thoroughly
addressed This study was designed to evaluate temperature and soil type impact on M bovis survival
In this study we demonstrated that M bovis SB0120 may survive several months in soil under
different laboratory conditions in two contrasted soils and that cool temperature (4degC) strongly
promotes M bovis persistence whatever the soil type while warmer temperature (22degC) shortens its
survival time Our results are in contradiction with a previous laboratory study on M bovis persistence
(Young et al 2005) where mycobacterial survival was longer at 25degC than at 4degC in a sterile sandy soil
(pH 65-68 60 days vs 16 days) but in agreement with previous field studies where lower temperature
increased M bovis survival time up to 88 days (Fine et al 2011 Jackson et al 1995)
We also pointed out that soil type may impact on M bovis survival time Indeed despite that
no statistical difference was observed the soil A that exhibits high clay and loam content and slightly
basic pH allowed persistence of cultivable M bovis for up to three months (90 days) while this
persistence was only two months in the acidic sandy loamy soil B when soils were incubated at 22degC
On the contrary a difference of M bovis survival was observed at 4degC between the two soils with a
better survival in soil A than in soil B suggesting an impact of soil type on M bovis survival at cool
temperature However we were hampered by the detection limit of the culture-based method in the
experiment on sterile soil at 22degC where CFU numbers and detection limit ovelaped after day 60
Enrichment broth are very useful to recover low number of bacteria in complex matrices such as
Salmonella typhimurium in food or Listeria monocytogenes in soil (Locatelli et al 2013 Zheng et al
2013) but mycobacterial enrichment broths are not yet available for mycobacteria
In our study clay soil with slightly basic pH seemed to promote M bovis SB0120 survival
although mycobacterial culture detection or persistence are usually associated with sandy soils
harbouring acidic pH as previously suggested by positive culture or statistical studies (Walter et al
2014 Johnson-Ifearulundu amp Kaneene 1999 Reviriego et al 2000 Santos et al 2015) suggesting an
improved survival of the mycobacteria such as M avium ssp paratuberculosis and M bovis in these
soils However a one year persistence of cultivable M bovis was demonstrated in a sandy soil with a
basic pH (83) spiked with 108 CFU g- 1 (Ghodbane et al 2014) suggesting that M bovis may survive a
Chapitre 4
146
long time in soils harbouring a wide range of pH Moreover iron content of soils A and B seemed to be
significantly higher than soils described in previous studies
In this study qPCR was able to detect the presence of M bovis DNA over a longer period of
time than bacterial culture in soils samples incubated at 22degC whatever the soil type As one previous
study reported that DNA of dead M bovis cells was no more amplified 10 days after cell death (Young
et al 2005) we hypothesized that our qPCR assay detected live M bovis up to 150 days after
inoculation of soils Viable M bovis cells have already been detected in sterile and biotic soil over a 15
month period targeting the 16S rRNA encoding gene (Young et al 2005) Physiological state of
pathogenic mycobacteria such as M bovis in soil is unknown but dormancy or a viable but non
cultivable state (VBNC) have been suggested to explain their survival in environmental matrices
(Whittington et al 2004 Adams et al 2013) Further analysis of the expression of dormancy-related
genes (Aacutelvarez et al 2009) under soil conditions might allow to support this hypothesis Further
researches are critical to ascertain the long bacterial survival in soil and to understand the genetic
mechanisms involved in soil persistence
Our laboratory conditions were far from field conditions where temperature fluctuations
moisture variations and UV radiation are detrimental to the mycobacteria Mycobacterium bovis
survival time obtained in this study is probably longer than survival time in environmental conditions
As the gap of knowledge is important about how M bovis survive in soils laboratory studies have to
be set out in order to determine the main factors involved in M bovis survival However experiments
dealing with the exposition of M bovis naturally or artificially contaminated samples to environmental
conditions must be carried out in specific open air enclosure with high levels of protection to exclude
mycobacterial dissemination (Fine et al 2011) or require climate chambers with a L3 biosafety level
Significant differences in the recovery of M bovis DNA between biotic and sterile soils were
observed with the qPCR-based assay from the first day of the experiment indicating that qPCR
detection is hampered when soil metagenomic DNA is concentrated although no inhibition was
observed
In our experiment the use two soils gave an insight on the impact of soil type on M bovis
survival However extensive studies on larger number of well-characterised and contrasted soils would
be required to target the key edaphic factors that promote M bovis survival in soil Further work is
needed to identify those key factors and determine if the population detected with qPCR consists of
non-culturable or dormant cells capable of resuscitation under specific growth conditions or following
inhalation or ingestion by animals
Chapitre 4
147
Funding information
The work was supported by the Regional Council of Burgundy (Elodie Barbier PhD fellowship) the Cocircte
drsquoOr General Council Interbev and the Cocircte drsquoOr GDS (Groupement de Deacutefense Sanitaire Livestock
health protection group) The funders had no role in study design data collection and interpretation
or the decision to submit the work for publication
Conflict of interest
The authors have no conflict of interest to declare
Acknowledgements
The authors would like to thank Jean-Marie Duez Julien Bador and Catherine Neuwirth from the
Plateau Technique de Biologie of the Hospital of Dijon
REFERENCES
Adams AP et al 2013 Comparison of PCR versus culture for detection of Mycobacterium bovis after experimental inoculation of various matrices held under environmental conditions for extended periods Applied and Environmental Microbiology 79(20) pp6501ndash6506
Aacutelvarez AH Estrada-Chaacutevez C amp Flores-Valdez MA 2009 Molecular findings and approaches spotlighting Mycobacterium bovis persistence in cattle Veterinary Research 40(3) pp1ndash16
Barasona JA et al 2015 DNA detection reveals Mycobacterium tuberculosis complex shedding routes in its wildlife reservoir the Eurasian wild boar Transboundary and Emerging Diseases (DECEMBER)
Biet F et al 2005 Zoonotic aspects of Mycobacterium bovis and Mycobacterium avium-intracellulare complex (MAC) Veterinary Research 36 pp411ndash436
Corner LAL et al 2012 The distribution of Mycobacterium bovis infection in naturally infected badgers Veterinary journal (London England 1997) 194(2) pp166ndash72
Courtenay O et al 2006 Is Mycobacterium bovis in the environment important for the persistence of bovine tuberculosis Biology letters 2(3) pp460ndash2
Dhand NK et al 2009 Association of farm soil characteristics with ovine Johnersquos disease in Australia Preventive Veterinary Medicine 89 pp110ndash120
Duffield BJ amp Young DA 1985 Survival of Mycobacterium bovis in defined environmental conditions Veterinary Microbiology 10 pp193ndash197
Fine AE et al 2011 A study of the persistence of Mycobacterium bovis in the environment under natural weather conditions in Michigan USA Veterinary medicine international 2011 p765430
Chapitre 4
148
Ghodbane R et al 2014 Long-term survival of tuberculosis complex mycobacteria in soil Microbiology (Reading England) 160(Pt 3) pp496ndash501
Jackson R De Lisle GW amp Morris RS 1995 A study of the environmental survival of Mycobacterium bovis on a farm in New Zealand New Zealand Veterinary Journal 43(7) pp346ndash352
Johnson-Ifearulundu Y amp Kaneene JB 1999 Distribution and environmental risk factors for paratuberculosis in dairy cattle herds in Michigan American journal of veterinary research 60(5) pp589ndash96
Little TW Naylor PF amp Wilesmith JW 1982 Laboratory study of Mycobacterium bovis infection in badgers and calves The Veterinary record 111(24) pp550ndash7
Locatelli A et al 2013 Nation-wide study of the occurrence of Listeria monocytogenes in French soils using culture-based and molecular detection methods Journal of microbiological methods 93(3) pp242ndash50
Morris RS Pfeiffer DU amp Jackson R 1994 The epidemiology of Mycobacterium bovis infections Veterinary microbiology 40(1-2) pp153ndash77
Palmer M V 2013 Mycobacterium bovis Characteristics of wildlife reservoir hosts Transboundary and Emerging Diseases 60(SUPPL1) pp1ndash13
Palmer M V Waters WR amp Whipple DL 2004 Investigation of the transmission of Mycobacterium bovis from deer to cattle through indirect contact American Journal of Veterinary Research 65(11) pp1483ndash1489
Palmer M V amp Whipple DL 2006 Survival of Mycobacterium bovis on feedstuffs commonly used as supplemental feed for white-tailed deer (Odocoileus virginianus) Journal of wildlife diseases 42(4) pp853ndash8
Palmer M V Whipple DL amp Waters WR 2001 Experimental deer-to-deer transmission of Mycobacterium bovis American journal of veterinary research 62(5) pp692ndash6
Peacuterez-Lago L Navarro Y amp Garciacutea-de-Viedma D 2013 Current knowledge and pending challenges in zoonosis caused by Mycobacterium bovis A review Research in Veterinary Science 97 ppS94ndashS100
Phillips CJC et al 2003 The transmission of Mycobacterium bovis infection to cattle Research in veterinary science 74(1) pp1ndash15
Reviriego FJ Moreno M a amp Domiacutenguez L 2000 Soil type as a putative risk factor of ovine and caprine paratuberculosis seropositivity in Spain Preventive Veterinary Medicine 43 pp43ndash51
Santos N et al 2015 Widespread environmental contamination with Mycobacterium tuberculosis Complex revealed by a molecular detection protocol Plos One 10(11) pe0142079
Sweeney FP et al 2007 Environmental monitoring of Mycobacterium bovis in badger feces and badger sett soil by real-time PCR as confirmed by immunofluorescence immunocapture and cultivation Applied and environmental microbiology 73(22) pp7471ndash3
Chapitre 4
149
Sweeney FP et al 2006 Immunomagnetic recovery of Mycobacterium bovis from naturally infected environmental samples Letters in applied microbiology 43(4) pp364ndash9
Tanner M amp Michel AL 1999 Investigation of the viability of M bovis under different environmental conditions in the Kruger National Park Journal of Veterinary Research (66) pp185ndash190
Vicente J et al 2007 Risk factors associated with the prevalence of tuberculosis-like lesions in fenced wild boar and red deer in south central Spain Veterinary Research 38(3) pp451ndash464
Walter WD et al 2014 Linking bovine tuberculosis on cattle farms to white-tailed deer and environmental variables using Bayesian hierarchical analysis PloS one 9(3) pe90925
Walter WD et al 2012 On-farm mitigation of transmission of tuberculosis from white-tailed deer to cattle literature review and recommendations Veterinary medicine international 2012 p616318
Ward MP amp Perez AM 2004 Association between soil type and paratuberculosis in cattle herds American Journal of Veterinary Research 65(1) pp10ndash14
Whittington R et al 2004 Survival and dormancy of Mycobacterium avium subsp paratuberculosis in the environment Applied and environmental microbiology 70(5) pp2989ndash3004
Williams RS amp Hoy WA 1930 The viability of B tuberculosis (bovinus) on pasture land in stored faeces and in liquid manure The Journal of hygiene 30(4) pp413ndash419
Young JS Gormley E amp Wellington EMH 2005 Molecular detection of Mycobacterium bovis and Mycobacterium bovis BCG (Pasteur) in soil Applied and environmental microbiology 71(4) pp1946ndash1952
Zheng Q et al 2013 Comparison of enrichment broths for the recovery of healthy and heat-injured Salmonella typhimurium on raw duck wings Journal of food protection 76(11) pp1963ndash8
Chapitre 4
150
C BILAN
Les reacutesultats preacutesenteacutes dans ce chapitre deacutemontrent la capaciteacute de M bovis SB0120 agrave survivre
plusieurs mois dans les deux sols de Cocircte drsquoOr sous une forme viable cette dureacutee eacutetant impacteacutee par
un facteur abiotique particulier la tempeacuterature Une survie drsquoau moins cinq mois est observeacutee agrave basse
tempeacuterature (4degC) dans les deux sols avec une tregraves faible diminution de la population bacteacuterienne
initialement inoculeacutee alors qursquoelle nrsquoest que de deux agrave trois mois agrave 22degC Bien que M bovis soit cultiveacutee
un mois de plus dans le sol C par rapport au sol D nous nrsquoavons pas mis en eacutevidence drsquoeffet de la nature
du sol sur la survie de la bacteacuterie agrave 22degC Toutefois agrave cause de contraintes techniques notre analyse
nrsquoa porteacute que sur deux sols limitant fortement lrsquointerpreacutetation des reacutesultats Lrsquoanalyse des sols inoculeacutes
et incubeacutes agrave 22degC par qPCR montre un tout autre profil de survie puisque M bovis est deacutetecteacutee en
quantiteacute importante pendant les cinq mois de lrsquoeacutetude sur les deux sols et les deux conditions (biotique
vs steacuterile)
Les reacutesultats divergents obtenus par la culture et la qPCR sur les sols incubeacutes agrave 22degC amegravenent
agrave faire diffeacuterentes hypothegraveses pour expliquer lrsquoabsence de croissance des bacteacuteries puisque la qPCR
deacutetecte lrsquoADN des cellules vivantes et mortes ainsi que lrsquoADN libre Nous pouvons donc envisager que
lrsquoADN deacutetecteacute entre 60 et 150 jours appartienne agrave des cellules viables non cultivables ou agrave des cellules
mortes entiegraveres ou lyseacutees Des eacutetudes compleacutementaires sont indispensables pour preacuteciser lrsquoeacutetat
physiologique des bacteacuteries devenues incultivables apregraves une incubation dans le sol
En extrapolant les reacutesultats obtenus aux conditions de tempeacuterature rencontreacutees en Cocircte drsquoOr
nous pouvons suggeacuterer une fenecirctre temporelle pendant laquelle la survie de M bovis dans le sol est
maximale La Figure 25 repreacutesente les tempeacuteratures minimales et maximales releveacutees agrave Dijon entre
janvier 2012 et deacutecembre 2015 Nous estimons que les tempeacuteratures rencontreacutees dans lrsquoAuxois sont
probablement plus proches des tempeacuteratures minimales observeacutees dans la plaine de Dijon-Longvic ougrave
se trouve la station meacuteteacuteo (anthropisation minimale altitude humiditeacute veacutegeacutetation) que des
maximales Ainsi la peacuteriode la plus agrave risque va de novembre agrave avril (tempeacuteratures minimales de
lrsquoair lt 5degC) et correspond agrave la fin de lrsquoautomne lrsquohiver et le deacutebut du printemps Durant le reste de
lrsquoanneacutee et en particulier en eacuteteacute (tempeacuteratures maximales de lrsquoair gt 22degC) les risques de survie de M
bovis semblent moins importants Ces conclusions restent des hypothegraveses dans la mesure ougrave la
tempeacuterature reacuteelle des sols nrsquoa pas eacuteteacute mesureacutee et la variation des autres facteurs climatiques nrsquoa pas
eacuteteacute prise en compte Cependant on peut penser qursquoen eacuteteacute (sous forte insolation) la tempeacuterature des
sols peut augmenter rapidement dans les quelques premiers centimegravetres de sol limitant la dureacutee de
survie de la bacteacuterie
Chapitre 4
151
Figure 25 Tempeacuteratures minimales et maximales observeacutees agrave Dijon-Longvic de 2012 agrave 2015 (Source Meacuteteacuteo France)
-5
0
5
10
15
20
25
30
35
1 2 3 4 5 6 7 8 9 10 11 12
Tmin2012
Tmin2013
Tmin2014
Tmin2015
Tmax2012
Tmax2013
Tmax2014
Tmax2015
4degC
22degC
CHAPITRE 5
Rocircle de la faune du sol dans la circulation de
Mycobacterium bovis eacutetude de la disseacutemination de
M bovis par les lombrics
E Barbier B Chantemesse M Rochelet L Fayolle L Bollache M L Boschiroli A Hartmann
Rapid dissemination of Mycobacterium bovis from cattle dung to soil by the earthworm
Lumbricus terrestris
(Article accepteacute en cours de publication Veterinary Microbiology)
155
CHAPITRE 5 - Rocircle de la faune du sol dans la circulation de
Mycobacterium bovis eacutetude de sa disseacutemination par les lombrics
A AVANT-PROPOS
Le rocircle de la faune et de la microfaune du sol dans la survie et la disseacutemination de
Mycobacterium bovis dans lrsquoenvironnement est inconnu aujourdrsquohui Comme nous lrsquoavons eacutevoqueacute dans
la synthegravese bibliographique les protozoaires plus particuliegraverement les amibes jouent un rocircle
vraisemblable dans la survie des mycobacteacuteries environnementales (Adekambi et al 2006 Mba Medie
et al 2011 Mura amp Bull 2006) voire dans la virulence des mycobacteacuteries opportunistes (Cirillo et al
1997) In vitro la survie intra-amibienne de M bovis pendant plusieurs mois a eacuteteacute deacutemontreacutee mais
aucune eacutetude de terrain eacutevaluant le rocircle potentiel des amibes comme reacuteservoir de M bovis nrsquoest
disponible Concernant la meacutesofaune les lombrics ont eacuteteacute identifieacutes comme vecteur potentiel dans la
transmission de M avium ssp paratuberculosis dans des eacutelevages bovins et caprins infecteacutes (Fischer
et al 2003) Veacuteritables ingeacutenieurs du sol ces animaux ont un rocircle important dans la circulation des
microorganismes en les transfeacuterant et les seacutelectionnant depuis la matiegravere organique qursquoils ingegraverent et
dont ils sont de gros consommateurs vers le sol qui les entoure par le biais de leurs turricules
Afin drsquoeacutevaluer le rocircle potentiel des lombrics dans la disseacutemination de M bovis dans le sol suite
agrave lrsquoingestion de matiegraveres feacutecales animales contamineacutees nous avons eacutetudieacute le transfert de M bovis BCG
depuis les fegraveces inoculeacutes vers le sol environnant par une espegravece de lombric aneacutecique Lumbricus
terrestris La population bacteacuterienne de M bovis a eacuteteacute deacutetecteacutee et quantifieacutee par qPCR dans le sol
environnant les turricules et le tube digestif des lombrics
Chapitre 5
157
B RAPID DISSEMINATION OF M BOVIS FROM CATTLE DUNG TO SOIL BY THE EARTHWORM LUMBRICUS TERRESTRIS
Elodie Barbierab Benoit Chantemesseab Murielle Rocheletab Leacuteon Fayolle a Loiumlc Bollachebc
Maria Laura Boschiroli d Alain Hartmann a
a INRA UMR 1347 Agroeacutecologie Dijon Cedex France
b Universiteacute de Bourgogne Franche Comteacute Dijon Cedex France
c Laboratoire Chrono-environnement UMR CNRS 6249 Besanccedilon France
d Universiteacute Paris-Est Laboratoire National de Reacutefeacuterence de la Tuberculose Uniteacute de Zoonoses Bacteacuteriennes Laboratoire de
Santeacute Animale ANSES Maisons-Alfort Cedex France
Key words Mycobacterium bovis spreading earthworms Lumbricus terrestris cattle soil quantitative
PCR
Running title Environmental dissemination of Mycobacterium bovis by earthworms
Corresponding Author Alain Hartmann UMR 1347 Agroeacutecologie INRA 17 rue Sully BP 86510 21065
Dijon Cedex France E-mail alainhartmanndijoninrafr
Chapitre 5
158
Abstract
Indirect transmission of the bovine tuberculosis (bTB) between wildlife and livestock is thought to occur
by inhalation or ingestion of environmental substrates contaminated through animal shedding The
role of the soil fauna such as earthworms in the circulation of Mycobacterium bovis from
contaminated animal feces is of interest in the epidemiology of bTB The objective of this study was to
assess the impact of earthworm activity on M bovis transfer from animal dung to castings and the
surrounding soil For this purpose microcosms of soil containing the anecic eathworms Lumbricus
terrestris were prepared and covered with cattle feces spiked with the M bovis BCG strain Pasteur to
carry out two separate experiments The dissemination the gut carriage and the excretion of M bovis
were all monitored using a specific qPCR-based assay Our results showed that the earthworm L
terrestris was able to rapidly disseminate M bovis from the contaminated cattle feces to the
surrounding soil through casting egestion Moreover contaminated earthworms were shown to shed
the bacteria for 4 days when transferred in a bTB-free soil This study highlights for the first time the
possible role of earthworms in the dissemination and the persistence of M bovis in soils in bTB endemic
areas
Chapitre 5
159
INTRODUCTION
Mycobacterium bovis is a pathogenic mycobacteria that can infect and cause bovine
tuberculosis (bTB) in cattle and in a wide range of domestic or wild mammals as well as in humans
(Biet et al 2005) bTB is a primarily respiratory disease but it may affect various organs in animals
(Morris et al 1994) Although cattle are considered as the principal host of M bovis wild fauna such
as badgers (Meles meles) wild boar (Sus scrofa) and red deer (Cervus elaphus) has also been identified
as potential wildlife reservoirs (Palmer 2013) Transmission of bTB between cattle mainly occurs by
inhalation of contaminated aerosols during close contacts with infected animals (Morris et al 1994
Phillips et al 2003) An inoculum of less than ten bacilli is enough to cause respiratory disease in cattle
(Dean et al 2005) Moreover indirect transmission by inhalation of environmental bioaerosols or
ingestion of infected soil and water could also be a potential way of animal contamination (Palmer et
al 2004 Vicente et al 2007)
Infected animals were shown to shed M bovis via oro-nasal mucus sputum urine feces and
wound discharges (Corner et al 2012 Palmer et al 2004) that may contaminate environment
especially pastures shared by cattle and wild animals Thus investigation of soil as a potential reservoir
of M bovis is an important step towards understanding the epidemiology of the bacteria To reach this
goal deciphering the ecological factors that govern the survival and the spread of M bovis is first
required
Among soil fauna earthworms constitute the largest animal biomass in most terrestrial
ecosystems especially in grasslands (Lavelle and Spain 2001) Earthworms play a role of engineers in
soil processes impacting on soil functions and ecosystem services (Blouin et al 2013) affecting soil
structure water movement organic matter decomposition nutrients releasing plant growth and
pollution remediation Earthworms are usually classified in three ecological types according to their
feeding habits i) the epigeic ones are restricted to the surface organic layers where they feed
themselves ii) the endogeic ones live in the subsurface where they burrow lateral tunnels to ingest soil
and organic matter and iii) the anecic ones feed preferentially on surface litter but live in burrows in
the soil (Lee 1985) Earthworms ingest soil particles mixed with organic matter that are mechanically
and chemically digested by both gut fluid and microbiota (Curry and Schmidt 2007) and release
castings (undigested soil egested on or under the soil surface) Earthworm gut microbiota acts as a
selective filter for soil microorganisms shapes microbial communities of soil and either up- or down-
regulates microbial populations (Byzov et al 2007 Egert et al 2004 Goacutemez-Brandoacuten et al 2011) It
has been shown that earthworms may have microbial beneficial activities especially in the transport of
bacterial inocula through soil (Thorpe et al 1996) but earthworms may also act as pests by promoting
the dissemination of plant pathogens such as Phytophtora cactorum and Fusarium reticulatum
Chapitre 5
160
(Montecchio et al 2015) More interestingly a few studies have described that human and animal
pathogenic bacteria viruses and parasites especially Escherichia coli O157H7 foot-and-mouth disease
virus and Cestoda tapeworms may be carried and spread by earthworms (Edwards and Bohlen 1996
Prysor Williams et al 2006)
Although animal dung is a very attractive food for earthworms (Lowe and Butt 2005) few
studies have evaluated the role of earthworms in the survival and the dissemination of potential
pathogenic mycobacteria from animal dung or manure to surrounding soil (Fischer et al 2003) In
addition since earthworms represent an important part of the diet of various animal species including
badgers and wild boar (Granval and Muys 1995 Lee 1985) the consumption of contaminated
earthworms (gut carriage) might be a source of contamination for wild fauna
With the aim to investigate the above hypothesis this current study was first devoted to assess
the impact of earthworm activity on M bovis transfer from animal dung to their released castings and
the surrounding soil Then the M bovis gut carriage and the casting contamination following ingestion
of infected feces were investigated
MATERIAL AND METHODS
Mycobacterium bovis strain preparation
Mycobacterium bovis BCG strain Pasteur 1173P2 was grown aerobically at 37degC in Middlebrook
7H9 broth supplemented with 10 Middlebrook ADC Growth Supplement (Sigma-Aldrich France) The
M bovis inoculum was prepared as following a 10 mL stationary phase culture was pelleted and once
the supernatant removed the cell aggregates were disrupted by vortexing with glass beads for 15
seconds The pellet was suspended in 1 mL of sterile water and serial 10-fold dilutions of the suspension
were spiked on modified Middlebrook 7H11 plates (Middlebrook 7H11 supplemented with 10 heat
inactivated bovine serum (Dominique Dutscher France) and 10 Middlebrook OADC Growth
Supplement (Sigma-Aldrich France) for enumeration The plates were always incubated at 37degC for 2
months A 10-fold dilution of the prepared M bovis suspension was also directly quantified with the
RD4 qPCR assay described in the section ldquoMycobacterium bovis quantitative PCR (qPCR)rdquo
Soil cattle feces and earthworms characteristics
A bTB-free pasture (that has never been grazed by bTB infected cattle) located in Burgundy
(Middle East of France) was selected to sample the soil used in this study This latter is a clayey loamy
soil with a pH of 775 Organic carbon and nitrogen contents were 361 and 38 gkg-1 respectively The
soil was sieved to 4 mm and stored at 4degC until its use
Fresh cattle feces were collected from a bTB-free Charolais cattle in a bTB-free area of Burgundy and
Chapitre 5
161
immediately used
The adult anecic earthworms (Lumbricus terrestris) were provided by LombrirsquoCarraz (France)
and kept at 4degC in large buckets containing soil and organic matter until starting the study Soil
earthworms and cattle feces used in this work were previously proved to be negative for the presence
of M bovis using the qPCR-based assay
DNA extraction and purification
DNAs from all the samples collected were extracted as described previously (Dequiedt et al
2011) Briefly 8 mL of lysis buffer (100 mM Tris pH 80 100 mM EDTA pH 80 100 mM NaCl and 2
(wv) SDS) 4 g of silica beads (100 μm diameter) 5 g of ceramic beads (14 mm diameter) and 8 glass
beads (4 mm diameter) were added to 2 g of sample in a 15 ml sterile vial - whereas 1 mL of lysis
buffer 05 g of silica beads 0625 g of ceramic beads 1 glass beads were added to 250 mg-samples
Samples were disrupted for 3 times 30 s at 4 ms in a FastPrepreg-24 Instrument (MP Biomedicals Europe
Illkirch France) incubated for 30 min at 70 degC and centrifuged for 5 min at 7000 timesg at room
temperature Supernatants were incubated on ice for 10 min with 110 volume of 3 M potassium
acetate pH 55 and then centrifuged for 5 min at 14000 times g One volume of ice-cold isopropanol was
added to the supernatant for DNA precipitation overnight at minus 20 degC DNA was collected by
centrifugation (30 min at 14000 times g) and DNA pellets were washed with ice-cold ethanol (70) and
dissolved in 100 μl of water Crude DNA extracts (100 μl) were loaded on Microbiospintrade columns
(Biorad Marnes-La-Coquette France) filled with 100 mg of polyvinyl polypyrrolidone (PVPP Sigma
Chemical Co France) After centrifugation (4 min 1000 times g 4 degC) eluted DNA was further purified
using the Geneclean Turbo Kit (Qiagen Courtaboeuf France) according to the manufacturers
instructions Purified DNA concentrations were estimated by spectrophotometry (λ = 260 nm) using a
NanoDropreg 2000 spectrophotometer (Thermo Scientific Illkirch France)
Mycobacterium bovis quantitative PCR (qPCR)
Mycobacterium bovis-specific qPCR was set up targeting the RD4 flanking region using
previously described forward RD4 flanking primer (5rsquo-TGTGAATTCATACAAGCCGTAGTCG-3rsquo) and probe
(5rsquo-6-FAM-AGCGCAACACTCTTGGAGTGGCCTAC-3rsquo TAMRA) (Sweeney et al 2007) In order to improve
qPCR efficiency the reverse RD4 flanking primer based assay was modified (5rsquo-
ATCTTAGCTGGTCAATAGCCATTTTT-3rsquo) to reduce the size of the qPCR product to 116 bp
qPCR reactions were carried out in triplicate with a 25 microL reaction mix containing 125 microL of
ABsolutetrade QPCR ROX Mix (Thermo Scientific France) 1 microL (final concentration 25 ngmicroL-1) of T4
bacteriophage gene 32 product (MP Biomedicals Europe France) 1 microL of each primer (final
concentration 400 nmolL-1) 05 microL of each probe (final concentration 200 nmol L-1) 4 microL of ultrapure
Chapitre 5
162
water and 5 microL of pure or diluted (10- or 20-fold) DNA Absolute quantification was achieved using
standard DNA dilutions A recombinant plasmid containing one copy of the RD4 fragment of M bovis
BCG strain Pasteur 1173P2 cloned in the pCRregII-TOPOreg vector (Invitrogen France) was used as a
standard A calibration curve was obtained by amplification of serial dilutions of the plasmid ranging
from 1 to 106 copies per PCR reaction All DNA samples extracted from the soil castings and feces
samples were screened with the RD4-based qPCR assay and results were expressed as the number of
M bovis gene copies per g of soil castings or feces We previously assessed that when Cycle threshold
(Ct) was above 39 detection was possible but unreliable (unpublished results) The word ldquotracesrdquo was
then chosen in all further experiments in order to indicate that M bovis targets are present in very low
quantities (below 250 CFUg-1 of soil) The qPCR assay was performed in a VIIA 7 Real-Time PCR System
(Thermo Fischer Scientific France) Initial denaturation was done at 95degC for 15 min followed by 45
cycles with denaturation at 95degC for 15 s annealing and elongation at 60degC for 1 min
Mesocosms preparation
Plastic jars (15 L) were filled with 1 kg of the clay-loamy soil Sterile water was added to each
soil sample to reach final soil moisture of 80 of the water-holding capacity (WHC) Once the soil was
packed down to obtain a 1 gcm-3 soil density 6 earthworms Lumbricus terrestris were placed into each
jar A 4-days delay was necessary to acclimatize the earthworms Nine control mesocosms where no
earthworms were added were processed and analysed
Evaluation of earthworm dissemination of M bovis to soil mesocosms
Aliquots of 50 g of cattle feces previously spiked with 1 mL of M bovis BCG suspension were
layered on the soil surface of each mesocosm Jars were covered with pierced fitted lids and kept in a
dark room at 20degC in a L2 confinement facility (day 0)
In order to sample fresh earthworm castings all the castings were removed 48 h before each
sampling date A sampling schedule ie 2 4 7 10 14 21 30 45 and 60 days was followed For each
sampling date 2 mesocosms with earthworms and one control mesocosm were analysed as following
First earthworm castings were collected at the surface of each mesocosm containing earthworms
pooled homogenized and then aliquoted in triplicate in 2 g-samples Secondly the cattle feces and
earthworms were removed out of the mesocosms and the remaining soil was mixed and aliquoted in
triplicate in 2 g-samples Finally the six earthworms contained in each mesocosm were rinsed twice
with sterile water dried with a paper towel and individually transferred into a 60 mL empty vessel
After a 48h incubation at 20degC in a dark room to release the gut content one 250 mg-aliquot of fecal
samples was sampled for each earthworm In the control mesocosm the cattle feces were removed
from the top of mesocosm the soil was then mixed sampled and analysed as described above All
Chapitre 5
163
samples were frozen at -20degC until their DNA extraction and RD4 qPCR analysis Gene copy numbers
found in each sample were box-cox transformed before analysis to normalize the data Generalized
Linear Models (GLMs) were used to analyze the effects of sampling date (number of days after the
M bovis spiked cattle feces was layered onto the soil) and treatment (feces castings and soil) on the
number of CFU Tests were performed using JMP 70 Software (SAS Institute Inc) Results were
considered to be significantly different for P values lt 005
Gut carriage and excretion of M bovis by earthworms
Five mesocosms each containing 6 earthworms were prepared as described in the section
ldquomesocosm preparationrdquo and were then covered with 50 g of cattle feces spiked with 1 mL of M bovis
BCG suspension After 7 days of incubation each earthworm was rinsed twice with sterile water dried
with a paper towel To estimate M bovis carriage on earthworm tegument 6 earthworms out of the
30 were deposited in an individual Petri plate containing modified Middlebrook 7H11 media
supplemented with antibiotics and antifungal (100 mgL-1Ticarcillin 10 mgL-1 Trimethoprim 200 000
UIL-1 Polymyxin B and 100 mgL-1 Cycloheximid Sigma-Aldrich France) After gently moving it on the
plate surface for few minutes the earthworms were individually transferred in a 200 mL vessel
containing 100 g of M bovis-free fresh soil as well as the 24 earthworms This step corresponded to
the starting point of the experiment (day 0)
At each sampling date ie day 1 2 3 4 and 5 the castings from series of 6 randomly selected
remaining vessels were collected pooled homogenized and one aliquot of 2 g was sampled Then the
earthworms from 1 series of 6 vessels were removed rinsed twice with sterile water dried with a paper
towel and individually transferred in a 60 mL empty vessel After 48h at 20degC in a dark room the
animals have released their gut contents and a 250 mg-aliquot of fecal sample was recovered from
each vessel containing earthworms All of the samples were stored at -20degC until further analysis
RESULTS
Earthworm dissemination of M bovis from cattle feces spiked with the mycobacteria to soil
mesocosms
For this purpose the protocol sketched in Figure 1 was carried out over a 60 days-period and
the results are summarized in Figure 2 Both the culture-based method and the RD4 qPCR assay were
used to estimate the concentration of the Mycobacterium bovis BCG suspension used to spike the cattle
feces Different values of 125 times 107 CFU per mL and of (65 plusmn 05) times 108 gene copies per mL were
determined with the culture and the molecular methods respectively However given that only the
RD4 qPCR assay was carried out to quantify M bovis throughout this experiment the M bovis genome
Chapitre 5
164
equivalent number contained in the 50 g of cattle feces was estimated to (65 plusmn 05) times 108 at the
beginning of the experiment (day 0 dashed line in Figure 2)
Figure 1 Schematic representation of the procedure followed for the evaluation of M bovis dissemination in
soil by earthworms
Overall M bovis was repeatedly detected in all of the analysed samples ie earthworm castings
fecal samples and soils from 2 days after depositing the cattle feces spiked with M bovis on the surface
of the mesocosms Gene copy number was significantly influenced by the sampling date (df= 8 sup2 =
14508 Plt00001) treatment (df= 2 sup2 = 22430 Plt00001) and interaction between sampling date
and treatment (df= 16 sup2 = 14464 Plt00001) However at the end of the experiment (day 60)
M bovis gene copy numbers were not statistically different (Average and P value) whatever the kind of
samples More precisely the earthworm fecal samples (open circle Figure 2) showed the highest
contamination (up to (57 plusmn 03) times 106 M bovis gene copies per g) at day 2 while the M bovis gene copy
numbers in the fecal samples decreased (20-fold factor) after day 4 to reach a final value of (25 plusmn 22)
times 105 gene copy number per g On the other hand the M bovis gene copy numbers per g of earthworm
castings were up to around 10-fold lower than those per g of earthworm fecal samples until day 45
after which similar M bovis gene concentrations were measured (full circle Figure 2) Although soil
samples from mesocosms also led to a M bovis positive signal from day 2 (44 plusmn 42) times 104 gene copy
number per g) it is worth noting that only 4 out of the 6 analyzed samples were positive at day 2 (full
square Figure 2) Afterwards (ie from day 4) all of the soil samples gave a positive signal for M bovis
with a strong increase (10-fold) of the M bovis gene copy number per g of soil between day 2 and day
(D0 + 2 4 7 10 14 21 30 45)
Mesocosm Deposition of M bovis spiked feces
D0
CASTING POOL
FECAL SAMPLE
48h00
Earthworm
Fresh soil
M bovis spiked cattle feces
SOIL
Sampling day
Chapitre 5
165
7 followed by minor fluctuations until day 60 For the control mesocosms (ie without any
earthworms) no M bovis signal were measured in the soil samples throughout the experiment until
day 60 where a positive signal value of (35 plusmn 19) times 104 M bovis per g of soil was obtained (cross symbol
on Figure 2) thus indicating a very slow spread of M bovis in the bulk soil in the absence of earthworms
Finally it should be noticed that the cattle feces remained undisturbed in the control mesocosms over
the course of the experiment whereas the earthworms totally ingested the spiked cattle feces in all of
the mesocosms containing these animals during the first week of the experiment
Figure 2 Concentration of M bovis DNA determined by RD4 qPCR in soil earthworm castings and fecal samples recovered at each sampling date Concentrations of M bovis DNA in control mesocosms are represented with crosses (times) The dashed line corresponds to the concentration of M bovis spiked in 50 g of cattle feces Error bars represent the standard deviation values of 3 replicates
3
4
5
6
7
8
0 5 10 15 20 25 30 35 40 45 50 55 60
Lo
g1
0(M
b
ovi
sg
ene
cop
y n
um
ber
) in
1 g
of
sam
ple
Days
Chapitre 5
166
Table 1 RD4-based qPCR detection of M bovis gene copy number contained in the earthworm castings recovered according to the protocol of Figure 2
Sampling day Number of analysed
casting poolsa Number of positive
casting poolsb M bovis gene copy number per gc
1 5 5 (196 plusmn 136) times 105
2 4 4 (104 plusmn 060) times 105
3 3 2 Tracesd
4 2 1 Traces
5 1 0 Negative a
From 6 vessels b
Threshold cycle (Ct) between 20 and 39 c Mean value calculated with the signal obtained for the positive earthworm casting pools
d Detected but not quantifiable (below 250 CFUg-1 of soil)
Evaluation of the carriage and the excretion of M bovis by earthworms
In order to check whether earthworms carry M bovis in their gut and are able to excrete the
bacteria the following experiment was realized (see detailed protocol in Figure 3) Earthworms were
artificially contaminated by feeding for 7 days from cattle feces spiked with M bovis then their castings
and their fecal samples were analysed for a 5-days period First no growth of M bovis was observed
from earthworm tegument surfaces suggesting the absence or the low carriage of M bovis by
earthworm teguments The results shown in Table 1 indicated that all of the earthworms produced
castings highly contaminated with M bovis 1 and 2 days after their introduction in the fresh M bovis
free soil (up to around 105 M bovis gene copy number per g) Then the percentage of positive M bovis
casting pools gradually decreased (66 and 50 for sampling day 3 and 4 respectively) until the
sampling at day 5 for which no M bovis molecular signal was detected Although positive signals were
obtained for the sampling days 3 and 4 only traces of M bovis were detected (Ct above 39) With
values ranging from 1 to 5 105 M bovis gene copy number per g of fecal sample (Table 2) the RD4
qPCR assay yielded M bovis positive signals for the sampling days 1 to 4 Nevertheless for each of
these sampling days some earthworms (17-50 ) led to negative M bovis qPCR signal thus indicating
the absence or a very low amount of M bovis in the earthworm feces (below the detection limit of the
qPCR) Finally all of the six earthworm fecal samples analysed at day 5 were M bovis free
Chapitre 5
167
Figure 3 Schematic representation of the procedure followed for the estimation of M bovis gut carriage and excretion by earthworms
Table 2 RD4-based qPCR detection of M bovis gene copy number contained in the earthworm fecal samples recovered according to the protocol of Figure 3
Sampling day Number of positive
fecal samplesa (n = 6) M bovis gene copy number per gb
1 5 (123 plusmn 082) times 105
2 4 (228 plusmn 214) times 105
3 3 (542 plusmn 049) times 105
4 4 (213 plusmn 164) times 105
5 0 Negative
a Threshold cycle (Ct) between 20 and 39 b Mean value calculated with the signal obtained for the positive earthworm fecal samples
Sampling day (D0 + 1 2 3 4 5)Mesocosm
(n = 5)
Deposition of M bovisspiked feces (n = 5 )
Investigation of M bovis tegumentcarriage on 6 earthworms
D0
7 days
Transfer of each earthworm
5 series of 6 vessels(n = 30)
FECAL SAMPLE
CASTING POOL
n = 6
48h00
n = number of remainingseries of 6 vessels
Earthworm
Fresh soil
M bovis spiked cattle feces
Chapitre 5
168
DISCUSSION
The results shown in Figure 1 evidenced for the first time that earthworms were able to widely
transfer M bovis from cattle feces to their castings after ingestion of cattle feces spiked with the
bacteria Owing to the concomitant ingestion of M bovis-free soil and dung remains and thus the
dilution of M bovis cells in the gut content the number of M bovis cells in castings gradually decreased
from day 4 until the end of the experiment These results were in good agreement with earlier reported
laboratory trials with Mycobacterium avium subsp paratuberculosis that is responsible for the Johnersquos
disease in cattle and goat (Fischer et al 2003) Prysor Williams et al (Prysor Williams et al 2006) have
also reported the excretion of the human food-borne pathogen Escherichia coli O157H7 in castings
after ingestion of spiked manure by earthworms
On the other hand this study demonstrated that when earthworms were maintained in the
M bovis infected environment they were able to shed M bovis in their castings for at least 60 days
(Tableau 1) However when transferred in a M bovis-free soil earthworms only shed the bacteria up
to 4 days This data suggests that M bovis is unable to colonize the gut of the earthworm L terrestris
over extended periods Moreover L terrestris may ingest 490 to 3500 mg dry mass g-1 fresh mass of
arable soil per day (Curry and Schmidt 2007) and has a mean gut transit time of about 12 hours (Taylor
and Taylor 2014) leading to the rapid egestion of the bacteria from earthworm guts through castings
Furthermore earthworms were able to transfer M bovis from cattle feces to the bulk soil by
releasing castings in the tunnels they burrow Our results showed that M bovis was still detected in
mixed soil over the course of the experiment (60 days) highlighting a long persistence of M bovis in
soil Rapid dispersal of the bacteria in soil can be fully imputed to earthworm activity since soil from
control mesocosms without earthworms remained free from M bovis-except at day 60 Moreover
bacteria movement was mainly due to M bovis earthworm excretion since no M bovis tegument
carriage was observed The M bovis signal observed in control mesocosms at day 60 is probably due
to the moving of M bovis cells during moisture transfer from dung to soil surface and to the presence
of remaining dung particles in soil (incomplete removal of contaminated dung before soil mixing) As
M bovis is sensitive to solar radiation (UV) and soil dryness the burying of M bovis in deep soil by
earthworms may contribute to enhance its survival in soil environment Indeed survival of M bovis in
soil may be enhanced up to 2 years when the bacteria is buried to a depth of 5 cm (Genov 1965) Our
experimental design based on 1 kg-soil mesocosms did not allow estimating the real spatial distribution
of the bacilli in soil by earthworms It has been shown that earthworms are potentially able to move
several meters apart from the area where they graze organic matter (Mather and Christensen 1988)
thus disseminating soil-borne plan pathogen through large surfaces of soil (Thorpe et al 1996)
Concerning potential spread of animal or human pathogens earthworms were demonstrated in
Chapitre 5
169
experimental boxes to significantly aid vertical and lateral movement of ingested bacteria both up to
40 cm from the inoculation point (Prysor Williams et al 2006)
Finally we showed that earthworms may carry M bovis for many days in their guts even when
transferred in a M bovis-free environment Earthworms are known to host animal parasites and
pathogen such as protozoan tapeworms (Cestoda) and nematodes they can transmit to birds and
mammals when earthworms are ingested (Edwards and Bohlen 1996) However to our knowledge
the fact that earthworms may act as a source of contamination for animals feeding from earthworms
has never been studied Earthworms that constitute a nutritious food resource are an important part
of the diet of badgers and wild boar (Baubet et al 2003 Cleary et al 2011) that often forage in cattle
pastures hosting high earthworm populations (Muldowney et al 2003) Badgers and wild boar are
able to eat up to 200 and 120 earthworms respectively in a night especially during worm nights when
humid and mild weather conditions are optimal for earthworms to ascend to the surface (Baubet et
al 2003 Kruuk and Parish 1981) In endemic area of bTB where fecal M bovis shedding may occur in
infected animals it would be interesting to evaluate M bovis carriage in earthworm gut in the vicinity
of infected animal dung Further studies are needed to assess the role of earthworms harbouring
M bovis gut carriage in the contamination of bTB sensible earthworm feeders
One field study reports the isolation of non-tuberculous mycobacteria such as Mycobacterium
avium complex members (M avium ssp paratuberculosis and M avium ssp hominisuis) as well as M
abcessus M scrofulaceum and M gastri) from 9 out of 109 earthworm feces in sites where cattle
goats and swine were severely infected with the Johnersquos disease or mycobacteriosis (Fischer et al
2003) However this study failed to isolate M avium ssp avium from earthworms feces in aviaries
infected with avian tuberculosis
Cattle feces were spiked with M bovis at a final concentration of 125 times 107 CFU per 50 g of
feces ie 25 times 105 CFU per g (estimated by culture-based enumeration of the inoculum) M bovis
concentration in spiked cattle feces was also measured by the qPCR-based method We observed
numbers of M bovis 52-fold higher when using the molecular detection method than with the culture-
based method This discrepancy is probably due first to the strong aggregation of mycobacteria cells in
the inoculum despite physical disruption of cell aggregates and second to the presence of dead or
viable but non-cultivable cells of M bovis non detected by culture-based method The amount of
M bovis spiked in cattle feces is equivalent or lower than the number of M bovis cells recorded per g
of urine or feces shed by infected badgers (Gallagher and Clifton-Hadley 2000 Sweeney et al 2007)
leading to the presence of millions of bacteria in a latrine (burrowed hole in the soil where badgers
defecate and urinate) But it is probably larger than M bovis cells deposited in infected cattle feces
(Neill et al 1988) For practical reasons we used cattle feces in our study but it could be interesting to
compare if dung from other animals than cattle would be as attractive for earthworms
Chapitre 5
170
Our study was based on a qPCR assay method detecting M bovis DNA in soil castings and feces
samples This molecular tool does not give any information on the viability of the M bovis cells egested
by the earthworms However as mycobacteria harbour a resistant waxy cell envelope (Brennan and
Nikaido 1995) and as Mycobacterium avium ssp paratuberculosis and other environmental
mycobacteria have been showed to largely survive through the gut passage (Fischer et al 2003) these
data strongly suggest that M bovis could resist through the passage of earthworm guts and remain
alive in castings Further research is needed to clarify if tuberculous mycobacteria such as M bovis are
digested or survive during transit through the earthworm gut and if their virulence is conserved
Evidencing M bovis presence with culture-based method in environmental matrices such as soil
castings or animal feces is still a challenge As M bovis is a slow growing mycobacteria environmental
samples have to be strongly decontaminated prior to culturing to overcome contamination with fast
growing microorganisms leading to a decrease in the sensitivity of the method
Surface and subsurface casting shedding of M bovis is all the more relevant because this
pathogen bacteria may survive in soil and feces for long periods (Young et al 2005) Castings are then
within the reach of animals especially cattle that may be infected by inhaling soil aerosols or by
ingesting contaminated casting soil during their exploratory foraging or feeding activities While
Mycobacterium bovis infective dose by the respiratory route is very low since only ten viable bacilli are
able to cause respiratory bovine tuberculosis (Dean et al 2005) 5000 bacilli would be required to
initiate infection through the oral route in cattle (Palmer et al 2004) As cattle usually avoid grazing
close to the feces of other cattle or wild animals (Hutchings and Harris 1997) further dissemination of
M bovis by earthworms away from infected feces might induce contamination risk for cattle grazing in
this area
Further work is still needed to study the occurrence of M bovis in earthworms under natural
field conditions in bTB-contaminated area Cultivation of environmental M bovis is also required to
characterize properly these environmental strains Finally virulence of M bovis contained in
earthworm castings remains to be deciphered
Funding information
The work was supported by the Regional Council of Burgundy (Elodie Barbier PhD fellowship) the Cocircte
drsquoOr General Council Interbev and the Cocircte drsquoOr GDS (Groupement de Deacutefense Sanitaire Livestock
health protection group) The funders had no role in study design data collection and interpretation
or the decision to submit the work for publication
Conflict of interest
The authors have no conflict of interest to declare
Chapitre 5
171
Acknowledgements
The authors would like to thank AE Taconet F Bartheacuteleacutemy and T Grosjean from the Cocircte drsquoOr Regional
Veterinary Service (DDPP21) F Chevalier and M Beacuteral from the CIREV (Cellule Inter-Reacutegionale
drsquoEpideacutemiologie Veacuteteacuterinaire)
References
Baubet E Ropert-Coudert Y Brandt S 2003 Seasonal and annual variations in earthworm consumption by wild boar (Sus scrofa scrofa L) Wildl Res 30 179 doi101071WR00113
Biet F Boschiroli ML Thorel MF Guilloteau LA 2005 Zoonotic aspects of Mycobacterium bovis and Mycobacterium avium-intracellulare complex (MAC) Vet Res doi101051vetres
Blouin M Hodson ME Delgado E a Baker G Brussaard L Butt KR Dai J Dendooven L Peres G Tondoh JE Cluzeau D Brun JJ 2013 A review of earthworm impact on soil function and ecosystem services Eur J Soil Sci 64 161ndash182 doi101111ejss12025
Brennan PJ Nikaido H 1995 The envelope of mycobacteria Annu Rev Biochem 64 29ndash63 doi101146annurevbiochem64129
Byzov BA Khomyakov N V Kharin SA Kurakov A V 2007 Fate of soil bacteria and fungi in the gut of earthworms Eur J Soil Biol 43 149ndash156 doi101016jejsobi200708012
Cleary GP Corner LAL OrsquoKeeffe J Marples NM 2011 Diet of the European badger (Meles meles) in the Republic of Ireland A comparison of results from an analysis of stomach contents and rectal faeces Mamm Biol 76 470ndash475 doi101016jmambio201010012
Corner LAL OrsquoMeara D Costello E Lesellier S Gormley E 2012 The distribution of Mycobacterium bovis infection in naturally infected badgers Vet J 194 166ndash72 doi101016jtvjl201203013
Curry JP Schmidt O 2007 The feeding ecology of earthworms - A review Pedobiologia (Jena) 50 463ndash477 doi101016jpedobi200609001
Dean GS Rhodes SG Coad M Adam O Cockle PJ Clifford DJ Hewinson RG Vordermeier HM 2005 Minimum infective dose of Mycobacterium bovis in cattle Infect Immun 73 6467ndash6471 doi101128IAI73106467
Edwards CA Bohlen PJ 1996 Biology and ecology of earthworms Chapman and Hall London
Egert M Marhan S Wagner B Scheu S Friedrich MW 2004 Molecular profiling of 16S rRNA genes reveals diet-related differences of microbial communities in soil gut and casts of Lumbricus terrestris L (Oligochaeta Lumbricidae) FEMS Microbiol Ecol 48 187ndash197 doi101016jfemsec200401007
Fischer OA Matlova L Bartl J Dvorska L Svastova P Du Maine R Melicharek I Bartos M Pavlik I 2003 Earthworms (Oligochaeta Lumbricidae) and mycobacteria Vet Microbiol 91 325ndash338 doi101016S0378-1135(02)00302-4
Chapitre 5
172
Gallagher J Clifton-Hadley RS 2000 Tuberculosis in badgers a review of the disease and its significance for other animals Res Vet Sci 69 203ndash17 doi101053rvsc20000422
Genov I 1965 The effects of certain physical and chemical agents on Mycobacterium tuberculosis Veteriaacuternı Med Nauk Sofia 2 97ndash107
Goacutemez-Brandoacuten M Aira M Lores M Domiacutenguez J 2011 Epigeic Earthworms exert a bottleneck effect on microbial communities through gut associated processes PLoS One 6 doi101371journalpone0024786
Granval P Muys B 1995 Predation on earthworms by terrestrial vertebrates in XII Congress ldquoThe Game and the Manrdquo Sofia Bulgaria p 480
Hutchings MR Harris S 1997 Effects of farm management practices on cattle grazing behaviour and the potential for transmission of bovine tuberculosis from badgers to cattle Vet J 153 149ndash62
Kruuk H Parish T 1981 Feeding specialization of the European badger Meles meles in Scotland UK J Anim Ecol 50 773ndash788 doi1023074136
Lavelle P Spain A 2001 Soil ecology Kluwer Academic Publishers Dordrecht
Lee KE 1985 Earthworms their ecology and relationship with soils and land use Academic P ed
Lowe CN Butt KR 2005 Culture techniques for soil dwelling earthworms A review Pedobiologia (Jena) 49 401ndash413 doi101016jpedobi200504005
Mather JG Christensen O 1988 Surface movements of earthworms in agricultural land Pedobiologia (Jena) 32 399ndash405
Montecchio L Scattolin L Squartini A Butt K 2015 Potential spread of forest soil-borne fungi through earthworm consumption and casting iForest 8 295ndash301 doi103832ifor1217-008
Morris RS Pfeiffer DU Jackson R 1994 The epidemiology of Mycobacterium bovis infections Vet Microbiol 40 153ndash77
Muldowney J Curry J Okeeffe J Schmidt O 2003 Relationships between earthworm populations grassland management and badger densities in County Kilkenny Ireland Pedobiologia (Jena) 47 913ndash919 doi101016S0031-4056(04)70289-9
Neill SD Hanna J OrsquoBrien J McCracken R 1988 Excretion of Mycobacterium bovis by experimentally infected cattle Vet Rec 123 340ndash343 doi101136vr12313340
Palmer M V 2013 Mycobacterium bovis Characteristics of wildlife reservoir hosts Transbound Emerg Dis 60 1ndash13 doi101111tbed12115
Palmer M V Waters WR Whipple DL 2004 Investigation of the transmission of Mycobacterium bovis from deer to cattle through indirect contact Am J Vet Res 65 1483ndash1489 doi102460ajvr2004651483
Phillips CJC Foster CRW Morris PA Teverson R 2003 The transmission of Mycobacterium bovis infection to cattle Res Vet Sci 74 1ndash15
Chapitre 5
173
Prysor Williams A Roberts P Avery LM Killham K Jones DL 2006 Earthworms as vectors of Escherichia coli O157H7 in soil and vermicomposts FEMS Microbiol Ecol 58 54ndash64 doi101111j1574-6941200600142x
Sweeney FP Courtenay O Hibberd V Hewinson RG Reilly L a Gaze WH Wellington EMH 2007 Environmental monitoring of Mycobacterium bovis in badger feces and badger sett soil by real-time PCR as confirmed by immunofluorescence immunocapture and cultivation Appl Environ Microbiol 73 7471ndash3 doi101128AEM00978-07
Taylor AR Taylor AFS 2014 Assessing daily egestion rates in earthworms using fungal spores as a natural soil marker to estimate gut transit time Biol Fertil Soils 50 179ndash183 doi101007s00374-013-0823-5
Thorpe IS Prosser JI Glover L a Killham K 1996 The role of the earthworm Lumbricus terrestris in the transport of bacterial inocula through soil Biol Fertil Soils 23 132ndash139
Vicente J Houmlfle U Garrido JM Fernaacutedez-De-Mera IG Acevedo P Juste R Barral M Gortazar C 2007 Risk factors associated with the prevalence of tuberculosis-like lesions in fenced wild boar and red deer in south central Spain Vet Res 38 451ndash464 doi101051vetres2007002
Young JS Gormley E Wellington EMH 2005 Molecular detection of Mycobacterium bovis and Mycobacterium bovis BCG (Pasteur) in soil Appl Environ Microbiol 71 1946ndash1952 doi101128AEM7141946
Chapitre 5
174
C COMPLEMENT PRELEVEMENT ET ANALYSE DE LOMBRICS DANS UNE PATURE SUSPECTE
Afin drsquoeacutevaluer la contamination naturelle eacuteventuelle des lombrics par M bovis nous avons
analyseacute 90 lombrics preacuteleveacutes en feacutevrier 2014 agrave proximiteacute drsquoune source permanente drsquoune des pacirctures
suspectes de lrsquoAuxois ougrave des bovins infecteacutes ont pacirctureacute lrsquoeacuteteacute preacuteceacutedent (Site C de lrsquoeacutetude
environnementale Chapitre 2) Les lombrics ont eacuteteacute preacuteleveacutes le mecircme jour que les trois eacutechantillons
positifs de ce site (eau seacutediments neacutematodes) Sur sept points de la pacircture choisis au hasard une
pelleteacutee drsquoenviron 20 kg de sol a eacuteteacute retourneacutee Les vers ont eacuteteacute seacutepareacutes du sol rinceacutes avec de lrsquoeau et
transfeacutereacutes individuellement dans des pots en plastique de 60 mL Les pots ont eacuteteacute stockeacutes 48 h agrave 20degC
dans lrsquoobscuriteacute afin de laisser les vers se vider Pour chaque point de la pacircture preacuteleveacute les fegraveces des
vers ont eacuteteacute pooleacutees et homogeacuteneacuteiseacutees puis 2 g ont eacuteteacute preacuteleveacutes et congeleacutes jusqursquoagrave la reacutealisation de
lrsquoanalyse Apregraves extraction de lrsquoADN chaque eacutechantillon a eacuteteacute analyseacute en qPCR avec le systegraveme de
deacutetection RD4 Les protocoles utiliseacutes sont deacutecrits dans le Mateacuteriel et Meacutethodes de lrsquoarticle preacuteceacutedent
Lrsquoidentification pheacutenotypique des 90 vers collecteacutes dans la pacircture a permis de mettre en
eacutevidence cinq genres de lombric diffeacuterents Lumbricus terrestris Lumbricus castaneus Nicodrilus sp
Octolosium cyaneum Allolobophora sp Aporrectodea caliginosa
Aucun des sept pools de fegraveces analyseacutes nrsquoa donneacute de signal positif en qPCR avec le systegraveme RD4
Chapitre 5
175
D BILAN
Les lombrics sont de gros consommateurs de matiegravere organique naturellement chargeacutee drsquoune
flore abondante et varieacutee et potentiellement drsquoune flore bacteacuterienne pathogegravene Toutefois il existe
tregraves peu de donneacutees sur la disseacutemination des bacteacuteries pathogegravenes pour lrsquohomme et les animaux par
les lombrics dans le sol ainsi que sur les risques lieacutes au portage intestinal de bacteacuteries pathogegravenes par
les lombrics pour les animaux qui les consomment
Lrsquoeacutetude deacutecrite dans lrsquoarticle preacuteceacutedent montre qursquoapregraves avoir ingeacutereacute des matiegraveres feacutecales
fraiches inoculeacutees les lombrics ont la capaciteacute de disseacuteminer rapidement M bovis dans le sol
environnant par le biais des turricules qursquoils eacutemettent agrave la surface du sol et en profondeur dans les
tunnels qursquoils creusent Lrsquoenfouissement de la bacteacuterie dans le sol permet probablement drsquoameacuteliorer la
persistance de la bacteacuterie en la proteacutegeant des UV et de la deacuteshydratation Drsquoautre part le transfert
de la bacteacuterie agrave quelques cm des deacutejections animales contamineacutees par le biais des turricules de surface
la rend plus accessible aux animaux (qui en geacuteneacuteral eacutevitent les fegraveces des autres animaux) Ainsi en
fonction des comportements exploratoires ou alimentaires des diffeacuterentes espegraveces freacutequentant les
pacirctures il existe un risque potentiel de contamination par du sol ou des turricules contamineacutes
(inhalation de bioaeacuterosols ou ingestion)
Nous avons eacutegalement montreacute que le tube digestif des lombrics ne constituait pas un reacuteservoir
peacuterenne de M bovis la bacteacuterie nrsquoeacutetait deacutetecteacutee que transitoirement (4 jours) apregraves avoir remis les
lombrics dans un sol sain Le portage digestif pendant les 4 premiers jours constitue toutefois un risque
pour les animaux qui consomment les lombrics en grande quantiteacute notamment les blaireaux et les
sangliers
Cette eacutetude soulegraveve les mecircmes inconnues que les eacutetudes preacuteceacutedentes principalement sur la
viabiliteacute lrsquoeacutetat physiologique et la conservation de la virulence de M bovis apregraves un transfert dans le
sol ou suite au passage de la bacteacuterie dans le tube digestif des lombrics Ces facteurs sont
indispensables pour permettre lrsquoinfection drsquoanimaux via des matrices environnementales contamineacutees
Les moyens drsquoeacutevaluation de ces diffeacuterents facteurs seront discuteacutes dans la partie Perspectives du
manuscrit
Chapitre 5
176
Discussion geacuteneacuterale
Discussion geacuteneacuterale
179
Depuis 2004 face agrave la recrudescence de la tuberculose bovine en Cocircte drsquoOr la mise en place
drsquoun programme de lutte sanitaire reacuteguliegraverement ameacutelioreacute contre la maladie a permis de controcircler et
drsquoameacuteliorer la situation dans lrsquoeacutelevage bovin depuis 2010 Dans la zone endeacutemique du deacutepartement la
circulation de Mycobacterium bovis dans un systegraveme multi-hocircte speacutecifiques impliquant les bovins et
des espegraveces sauvages complique fortement la lutte contre la maladie notamment par la difficulteacute agrave
geacuterer lrsquoinfection dans la faune sauvage malgreacute les mesures de controcircle entreprises jusqursquoagrave maintenant
La transmission indirecte de la maladie entre animaux sauvages et bovins par le biais de substrats
environnementaux contamineacutes est fortement suspecteacutee dans les zones ougrave bovins et faune sauvage
infecteacutes partagent les mecircmes territoires et ressources La transmission environnementale a deacutejagrave eacuteteacute
deacutemontreacutee en conditions expeacuterimentales (Little et al 1982 Palmer et al 2004) mais elle est tregraves
difficile agrave deacutemontrer en conditions naturelles et passe notamment par lrsquoeacutevaluation du niveau
drsquoexcreacutetion par les animaux infecteacutes du rocircle de lrsquoenvironnement comme reacuteservoir de la bacteacuterie de sa
survie dans un eacutetat viable et du maintien de sa virulence
Dans ce contexte les objectifs de ce travail eacutetaient drsquoune part drsquoeacutevaluer la contamination
environnementale en Cocircte drsquoOr sur des agroeacutecosystegravemes freacutequenteacutes par des bovins etou des animaux
sauvages potentiellement infecteacutes dans le but de mettre en eacutevidence des reacuteservoirs
environnementaux eacuteventuels Drsquoautre part nous avons chercheacute agrave preacuteciser les facteurs impliqueacutes dans
la persistance de M bovis dans le sol et sa disseacutemination par la micro- et meacutesofaune du sol
1 Des meacutethodes deacuteveloppeacutees (ou adapteacutees) puis valideacutees dans des matrices environnementales
Notre objectif initial pour lrsquoanalyse des eacutechantillons environnementaux eacutetait drsquoutiliser en
parallegravele la PCR quantitative apregraves extractionpurification de lrsquoADN pour la deacutetection et la quantification
de M bovis et une meacutethode baseacutee sur la culture afin drsquoisoler les souches environnementales pour les
caracteacuteriser Cependant les protocoles de preacutetraitement et de culture tireacutes de la litteacuterature
(immunocapture et protocoles de deacutecontamination) que nous avons utiliseacutes en premiegravere intention sur
les eacutechantillons environnementaux nrsquoont pas donneacute les reacutesultats escompteacutes du fait de la croissance tregraves
lente de M bovis et de la preacutesence dans les eacutechantillons de tregraves nombreux microorganismes
interfeacuterents agrave croissance rapide qui ont gecircneacute la culture (envahissement rapide des boicirctes en une
semaine) La culture de M bovis agrave partir drsquoeacutechantillons environnementaux reste agrave lrsquoheure actuelle un
challenge (Fine OrsquoBrien et al 2011 Fine Bolin et al 2011 Young et al 2005 King et al 2015)
notamment agrave cause de lrsquoabsence de milieux suffisamment seacutelectifs et de bouillons drsquoenrichissement
Discussion geacuteneacuterale
180
adapteacutes comme il en existe notamment pour lrsquoisolement de bacteacuteries pathogegravenes telluriques telles
que Listeria monocytogenes ou Pseudomonas aeruginosa En lrsquoabsence de meacutethodes disponibles nous
avons donc mis au point deux protocoles tregraves innovants agrave la fin de nos travaux un protocole
drsquoextractiondeacutecontamination des mycobacteacuteries dans le sol et une meacutethode de deacutetection des
mycobacteacuteries en culture liquide Ces deux meacutethodes sont en cours drsquoeacutevaluation par la SATT (Socieacuteteacute
drsquoAcceacuteleacuteration de Transfert de Technologies) en vue drsquoune valorisation sous forme de savoir-faire ou de
brevet Des eacutetudes compleacutementaires seront neacutecessaires pour deacutevelopper ces outils innovants
La recherche de reacuteservoirs environnementaux potentiels de M bovis en Cocircte drsquoOr a donc eacuteteacute
baseacutee sur lrsquoutilisation de meacutethodes moleacuteculaires adapteacutees aux matrices environnementales
Contrairement aux meacutethodes culturales la qPCR permet une deacutetection et une quantification rapide
des ADN cibles sans ecirctre gecircneacutee par la preacutesence de contaminants Face au grand nombre de
mycobacteacuteries potentiellement preacutesentes dans les sols de Cocircte drsquoOr dont M microti (Michelet et al
2015) nous avons choisi drsquoutiliser trois systegravemes ciblant trois seacutequences diffeacuterentes dans le geacutenome
des espegraveces du MTBC ou de M bovis que nous avons deacuteveloppeacutes et valideacutes pour leur speacutecificiteacute et leur
sensibiliteacute Lrsquoutilisation conjointe de ces trois systegravemes nous a permis drsquoaugmenter la speacutecificiteacute de la
deacutetection Toutefois le faible niveau de deacutetection observeacute dans certains eacutechantillons ne nous a pas
permis de quantifier le MTBC ou M bovis dans tous les eacutechantillons
2 Contamination environnementale les zones humides les eaux de ruissellement et les sols de terriers sont potentiellement agrave risque
Les systegravemes moleacuteculaires deacuteveloppeacutes ont eacuteteacute utiliseacutes pour lrsquoanalyse de plus de 400
eacutechantillons environnementaux varieacutes preacuteleveacutes dans les agroeacutecosystegravemes de la zone endeacutemique de
Cocircte drsquoOr et de Charente La contamination environnementale a eacuteteacute deacutetecteacutee de faccedilon reacutepeacuteteacutee dans
deux types drsquoeacutecosystegravemes bien particuliers les zones humides preacutesentes sur les pacirctures (sources et
laquo mouilles raquo) les terriers et latrines de blaireaux
Concernant les zones humides nous avons mis en eacutevidence une contamination dans lrsquoeau
circulante et les seacutediments de deux sources naturelles et permanentes situeacutees sur des pacirctures utiliseacutees
par deux cheptels diffeacuterents dans deux communes diffeacuterentes Cette faible contamination de lrsquoeau
etou des seacutediments retrouveacutee agrave lrsquoendroit de la reacutesurgence ainsi que sur diffeacuterents points en aval est
une donneacutee nouvelle puisqursquoagrave ce jour M bovis nrsquoavait eacuteteacute isoleacutee que dans lrsquoeau stagnante et ses
seacutediments (Santos et al 2015) Lors de nos campagnes de preacutelegravevements nous avons constateacute que ces
reacutesurgences parfois capteacutees dans des abreuvoirs eacutetaient parfois le seul moyen drsquoabreuvement des
bovins dans les pacirctures Ces zones humides semblent tregraves attractives pour les bovins au vu des indices
de preacutesence constateacutes (matiegraveres feacutecales et nombreuses empreintes) La persistance de M bovis eacutetant
Discussion geacuteneacuterale
181
favoriseacutee dans les matrices humides riches en matiegravere organique et dans lrsquoeau (Young et al 2005 Fine
Bolin et al 2011 Duffield amp Young 1985 Wint et al 2002) ces zones constituent un milieu favorable
agrave la survie de la bacteacuterie Dans notre eacutetude la mise en eacutevidence de la contamination des zones humides
dans des pacirctures suspectes corrobore le reacutesultat de lrsquoeacutetude PatRisk dans laquelle une association
significative a eacuteteacute eacutetablie entre la preacutesence de mouilles et le statut laquo agrave risque drsquoune pacircture raquo (Bouchez-
Zacria 2015)
Lrsquoorigine de la contamination de ces sources et laquo mouilles raquo est inconnue toutefois elles sont
accessibles aux bovins et animaux sauvages infecteacutes qui peuvent potentiellement excreacuteter la bacteacuterie
lors de lrsquoabreuvement ou du fouissement Vu le contexte geacuteomorphologique il est eacutegalement
envisageable que les infiltrations drsquoeau dans les plateaux calcaires coiffant les valleacutees entrainent avec
elles des matrices environnementales fortement contamineacutees (latrines sols) qui ressortent au niveau
des pacirctures avec toutefois un effet de dilution tregraves important
Des mesures de bioseacutecuriteacute visant agrave limiter les risques lieacutes aux zones humides sont deacutejagrave
preacuteconiseacutees par la DDPP 21 et le GDS 21 agrave lrsquoeacutechelle de la pacircture Lrsquoameacutenagement drsquoabreuvoirs peu
accessibles aux animaux sauvages et la condamnation par des clocirctures de lrsquoaccegraves des bovins aux
laquo mouilles raquo propices agrave la survie de la bacteacuterie sont fortement conseilleacutes aux eacuteleveurs des zones agrave
risque pour limiter le risque de transmission indirecte de la maladie Le drainage des pacirctures est une
solution envisageacutee pour asseacutecher les zones humides et canaliser les sources dans les pacirctures agrave risque
mais il est strictement reacuteglementeacute et soumis agrave autorisation avec enquecircte publique sous le controcircle de
la Police de lrsquoeau (Direction Deacutepartementale des Territoires) La correacutelation de nos reacutesultats sur la
contamination des zones humides avec le bilan des foyers bovins et des cas sauvages ainsi que les
reacutesultats de lrsquoeacutetude PatRisk ont initieacute une reacuteflexion entre la DDPP21 la CIREV le GDS et le SIRTAVA
(Syndicat Intercommunal pour la Reacutealisation des Travaux drsquoAmeacutenagement de la Valleacutee de lrsquoArmanccedilon)
pour eacutevaluer la faisabiliteacute de travaux de drainage dans les pacirctures agrave risque dans le respect de la
regraveglementation Un problegraveme se pose toutefois pour les pacirctures preacutesentant de tregraves nombreuses
reacutesurgences temporaires sur lesquelles le drainage nrsquoest pas envisageable Lrsquoutilisation de ces pacirctures
pour drsquoautres productions animales (ovins chevaux) ou veacutegeacutetales (production de foin la mise en
culture nrsquoest pas toujours possible agrave cause des pentes) est une solution
Notre eacutetude a eacutegalement mis en eacutevidence la contamination des sols de terriers de blaireaux
par M bovis Le geacutenotype retrouveacute dans les sols de terriers est identique agrave celui retrouveacute chez les
blaireaux infecteacutes pieacutegeacutes agrave proximiteacute confirmant que cette espegravece a la capaciteacute de contaminer son
milieu de vie (Young et al 2005 Courtenay et al 2006 Sweeney et al 2006 Sweeney et al 2007) Dans
cette espegravece des animaux excreacuteteurs et super excreacuteteurs ont eacuteteacute mis en eacutevidence en Cocircte drsquoOr (Payne
2014) expliquant les quantiteacutes importantes de bacilles retrouveacutees dans les sols de terrier (Young et al
Discussion geacuteneacuterale
182
2005 Courtenay et al 2006) et les latrines dans notre eacutetude La deacutetection drsquoun signal moleacuteculaire
pendant 12 mois dans plusieurs entreacutees drsquoun mecircme terrier suggegravere fortement que les sols de terriers
peuvent constituer un reacuteservoir environnemental pour M bovis La survie des mycobacteacuteries est
favoriseacutee dans ces structures complexes et profondes ougrave regravegne un microclimat propice (Moore amp Roper
2003) tant au niveau de la stabiliteacute de la tempeacuterature de lrsquohumiditeacute optimale que de la protection
contre les UV Ce reacuteservoir potentiel repreacutesente un risque pour le maintien de la maladie dans le groupe
social de blaireaux qui partage le terrier notamment pour les blairotins qui y naissent pendant lrsquohiver
mais aussi entre les groupes sociaux qui peuvent reacuteguliegraverement changer de terriers Lorsqursquoils sont
localiseacutes dans les pacirctures et sont accessibles aux bovins le sol des terriers contamineacutes repreacutesenterait
un risque potentiel pour les bovins qui les explorent notamment un risque drsquoinhalation de bioaeacuterosols
contamineacutes geacuteneacutereacutes agrave partir de sol Enfin les sols et latrines contamineacutes souvent en limite des plateaux
calcaires seraient susceptibles drsquoecirctre agrave lrsquoorigine de la contamination des pacirctures par le biais des
infiltrations drsquoeau des pheacutenomegravenes de reacutesurgence drsquoeau et de ruissellement
Les mesures de bioseacutecuriteacute preacuteconisant la clocircture des terriers situeacutes dans les pacirctures semblent
essentielles agrave respecter par les eacuteleveurs si lrsquoon considegravere la longue persistance de la bacteacuterie dans ce
biotope particulier Le repeuplement des terriers contamineacutes ne fait lrsquoobjet drsquoaucun controcircle agrave lrsquoheure
actuelle Sous le controcircle du CSRPN (Conseil Scientifique Reacutegional du Patrimoine Naturel) une eacutetude
est actuellement en cours en Cocircte drsquoOr pour lrsquoeacutetude de faisabiliteacute drsquoun protocole de deacutepeuplement des
terriers (pieacutegeage de tous les individus du groupe) associeacute agrave un traitement des tunnels par des reacutepulsifs
permanents pour eacuteviter la recolonisation immeacutediate
La preacutevalence de M bovis dans lrsquoenvironnement des animaux infecteacutes en Cocircte drsquoOr est
probablement sous-estimeacutee dans notre eacutetude agrave cause de limites temporelles spatiales et
meacutethodologiques auxquelles nous avons fait face En premier lieu il existe un deacutelai incompressible de
plusieurs semaines agrave plusieurs mois entre les trois eacutevegravenements suivants i) la contamination
environnementale par un animal excreacuteteur (bovin ou faune sauvage) ii) le diagnostic de lrsquoinfection chez
cet animal (apregraves prophylaxie et abattage pour les bovins en hiver et pieacutegeage pour les blaireaux au
printemps et en eacuteteacute) et iii) le preacutelegravevement et lrsquoanalyse drsquoeacutechantillons environnementaux dans les zones
de vie de ces animaux Cette limite temporelle est eacutegalement eacutevoqueacutee par drsquoautres auteurs (Fine
OrsquoBrien et al 2011) Il serait ideacuteal drsquoeffectuer les preacutelegravevements environnementaux dans un deacutelai le plus
court possible apregraves lrsquoexcreacutetion du bacille par un animal infecteacute toutefois ce deacutelai deacutepend du rythme
des prophylaxies et de la freacutequence du pieacutegeage des blaireaux De plus sur le terrain malgreacute notre
strateacutegie drsquoeacutechantillonnage ciblant les zones partageacutees preacutefeacuterentiellement par la faune domestique et
sauvage les surfaces agrave preacutelever restent importantes Lrsquoanalyse de la totaliteacute des surfaces cibleacutees est
humainement techniquement et eacuteconomiquement irreacutealisable le nombre de preacutelegravevements a donc ducirc
Discussion geacuteneacuterale
183
ecirctre optimiseacute pour aboutir agrave une analyse pertinente Les reacutesultats obtenus ne donnent de ce fait qursquoune
image partielle de la contamination reacuteelle des zones preacuteleveacutees Enfin la charge bacteacuterienne de certains
preacutelegravevements environnementaux nrsquoa pu ecirctre quantifieacutee signifiant que les quantiteacutes de cibles preacutesentes
eacutetaient proches des limites de deacutetection de nos systegravemes moleacuteculaires Pour ces diffeacuterentes raisons
nous ne pouvons donc pas exclure une contamination environnementale plus large avec des quantiteacutes
tregraves faibles de M bovis
3 Survie et circulation de M bovis dans lrsquoenvironnement
La survie des bacteacuteries dans le sol deacutepend des facteurs biotiques et abiotiques qui caracteacuterisent
ces sols Les diffeacuterentes eacutetudes sur la survie de M bovis ont essentiellement consideacutereacute les facteurs
abiotiques climatiques telles que la tempeacuterature lrsquohumiditeacute et lrsquoexposition aux UV dans un seul type
de sol (Young et al 2005 Jackson et al 1995 Duffield amp Young 1985) Nous avons montreacute que M bovis
SB0120 eacutetait cultivable au moins cinq mois agrave une tempeacuterature basse (4degC) avec une mortaliteacute
bacteacuterienne minime et jusqursquoagrave trois mois agrave 22degC Ces dureacutees de survie sont supeacuterieures agrave celles
rapporteacutees dans de preacuteceacutedentes eacutetudes (Fine Bolin et al 2011 Jackson et al 1995 Duffield amp Young
1985) Les sols retrouveacutes dans lrsquoAuxois ont agrave la fois des caracteacuteristiques diffeacuterentes des sols
habituellement deacutecrits comme favorables agrave la survie des mycobacteacuteries (sols sableux agrave pH acide) et
des caracteacuteristiques communes leur richesse en matiegravere organique et en fer Bien que nous nrsquoayons
pas pu mettre en eacutevidence un effet de la nature du sol dans notre eacutetude sur la survie de M bovis il
nous parait probable que les sols argilo-limoneux peacuterennisent la maladie dans les valleacutees de lrsquoAuxois
Notre expeacuterience limiteacutee par des contraintes techniques lieacutees agrave lrsquoutilisation du laboratoire de
confinement L3 du CHU de Dijon aurait neacutecessiteacute lrsquoutilisation drsquoun plus grand nombre de sols de points
de preacutelegravevements et de reacutepeacutetitions biologiques La concentration des foyers bovins (notamment des
recontaminations de cheptels) dans une zone limiteacutee du deacutepartement en Auxois interroge eacutegalement
sur le rocircle du contexte paysager et geacuteomorphologique commun des pacirctures suspectes dans la survie
de M bovis
Ces dureacutees de survie de M bovis dans les sols doivent ecirctre consideacutereacutees avec preacutecaution car
obtenues dans des conditions expeacuterimentales controcircleacutees qui ne miment que partiellement les
conditions environnementales reacuteelles (sols deacutebarrasseacutees de leur flore endogegravene agrave une tempeacuterature
drsquoincubation constante une humiditeacute optimale et une protection contre les UV) Il est probable qursquoen
conditions naturelles la survie de M bovis soit moins longue agrave cause des variations nyctheacutemeacuterales de
tempeacuteratures des variations des preacutecipitations des UV ainsi que de lrsquoeffet barriegravere potentiel de la flore
endogegravene des matrices environnementales La correacutelation de ces donneacutees avec les courbes de
Discussion geacuteneacuterale
184
tempeacuteratures permettent de montrer que la peacuteriode de novembre agrave avril ougrave les tempeacuteratures sont les
plus basses est compatible avec une survie optimale de M bovis dans les sols
Les reacutesultats obtenus dans notre eacutetude de survie ont souleveacute la question de la persistance de
M bovis dans le sol dans un eacutetat viable mais non cultivable (VNC) puisque la bacteacuterie nrsquoest plus
cultivable mais toujours deacutetecteacutee par qPCR Lrsquoeacutetat VNC de M bovis a deacutejagrave eacuteteacute suggeacutereacute dans des eacutetudes
de survie dans du sol (Adams et al 2013 Young et al 2005) Les mycobacteacuteries possegravedent une capaciteacute
particuliegravere agrave srsquoadapter et agrave survivre dans des conditions deacutefavorables en lrsquoabsence de nutriments
drsquooxygegravene sous diffeacuterents stress exogegravenes notamment dans les phagosomes de lrsquohocircte pour les
mycobacteacuteries pathogegravenes (Gengenbacher amp Kaufmann 2012 Boon amp Dick 2012 Voskuil et al 2003)
Les mycobacteacuteries disposent de diffeacuterents systegravemes agrave deux composants (permettant une transduction
de signal) leur permettant la perception sensorielle des conditions environnementales et en particulier
de la composition du milieu extracellulaire (Cook et al 2009 Cole et al 1998) Dans les substrats
environnementaux la bacteacuterie conserve probablement tout son potentiel de perception du milieu qui
lrsquoentoure et ses capaciteacutes drsquoadaptation mais elle ne peut pas interagir avec lui comme avec les cellules
de lrsquohocircte A lrsquoheure actuelle les meacutecanismes drsquoadaptation mis en jeu par M bovis pour survivre lors de
son excreacutetion par lrsquohocircte dans lrsquoenvironnement sont inconnus
Nous avons montreacute pour la premiegravere fois que les lombrics veacuteritables ingeacutenieurs du sol ont la
capaciteacute de disseacuteminer la bacteacuterie dans leur environnement par le biais des tunnels qursquoils creusent et
des turricules qursquoils eacutemettent agrave et sous la surface du sol Lrsquoenfouissement de la bacteacuterie agrave quelques
centimegravetres sous le sol favorise probablement sa survie Les lombrics peuvent eacutegalement jouer un rocircle
de vecteur quand ils portent du sol contamineacute dans leur tube digestif et qursquoils sont ingeacutereacutes par des
blaireaux ou des sangliers Cette eacutetude soulegraveve la probleacutematique du rocircle de la faune du sol dans la
survie et la circulation de M bovis dans lrsquoenvironnement et le risque que des reacuteservoirs cacheacutes de la
bacteacuterie entretiennent agrave bas bruit une contamination de lrsquoenvironnement
Perspectives
Perspectives
187
Lrsquoeacutevaluation de la contamination environnementale reacutealiseacutee en Cocircte drsquoOr et dans une moindre
mesure en Charente (terriers de blaireaux uniquement) nrsquoest pas extrapolable aux autres reacutegions
franccedilaises toucheacutees par la tuberculose A ce jour aucune investigation environnementale aussi
complegravete nrsquoa eacuteteacute meneacutee dans des deacutepartements fortement infecteacutes comme la Dordogne et les
Pyreacuteneacutees Atlantiques ougrave bovins et faune sauvage cohabitent Il serait donc inteacuteressant drsquoeacutevaluer le
niveau de contamination environnementale des zones partageacutees par les bovins et la faune sauvage afin
de veacuterifier si les zones humides et les terriers sont eacutegalement agrave risque dans ces deacutepartements De plus
les reacutesultats obtenus dans la zone endeacutemique de Cocircte drsquoOr ont suggeacutereacute le rocircle possible du contexte
geacuteomorphologique hydrologique et peacutedologique de lrsquoAuxois dans la contamination environnementale
et la persistance de M bovis Pour veacuterifier cette hypothegravese il serait inteacuteressant de comparer le contexte
des pacirctures suspectes de lrsquoAuxois avec les pacirctures suspectes des deacutepartements les plus toucheacutes par la
tuberculose bovine La caracteacuterisation physico-chimique du sol de ces pacirctures permettrait peut-ecirctre de
mettre en relation drsquoeacuteventuelles proprieacuteteacutes communes entre les sols incrimineacutes et la persistance de la
bacteacuterie La mise en eacutevidence de facteurs de risque paysagers hydrographiques etou peacutedologiques
permettrait eacuteventuellement de mettre au point un outil de preacutevision des risques en fonction du
contexte rencontreacute (pacirctures agrave risque) drsquoadapter au mieux la gestion sanitaire et drsquoameacuteliorer la
bioseacutecuriteacute dans les eacutelevages Si le rocircle des zones humides (laquomouilles raquo) et des sources dans drsquoautres
reacutegions est confirmeacute il conviendra de limiter lrsquoaccegraves des bovins agrave ces zones pour limiter la
contamination En Cocircte drsquoOr des approches de ce type (drainage et clocircture des zones humides) pour
empecirccher lrsquoaccegraves des animaux sont en cours en collaboration avec les eacuteleveurs les agences de bassin
lrsquoONEMA ainsi que les gestionnaires des zones humides agrave lrsquoeacutechelle du bassin versant de la Brenne
La dureacutee de survie deacutemontreacutee de M bovis dans les sols remet partiellement en cause la dureacutee
du deacutelai de carence de 2 mois avant reacuteinstallation drsquoanimaux suite agrave la peacuteriode de vide sanitaire dans
les exploitations qui ont eacuteteacute foyer de tuberculose Nos observations devront ecirctre discuteacutees dans le
cadre de lrsquooptimisation de ces pratiques
Sur le plan expeacuterimental notre expeacuterience de survie sur deux sols eacutetant insuffisante une eacutetude
de la survie de M bovis sur un grand nombre de sols aux caracteacuteristiques physico-chimiques
contrasteacutees permettrait drsquoidentifier les facteurs physico-chimiques du sol associeacutes agrave une survie
optimale de la bacteacuterie comme cela a pu ecirctre fait au sein de notre eacutequipe pour Listeria monocytogenes
(Locatelli et al 2013) Lrsquoidentification des facteurs eacutedaphiques favorisant la persistance de M bovis
permettra de mettre en eacutevidence les sols agrave risque
La recherche de M bovis dans des matrices environnementales se heurte aux limites des
techniques utiliseacutees Concernant la qPCR bien qursquoelle reste une technique de choix des efforts sont
encore neacutecessaires pour abaisser les seuils de deacutetection dans les matrices complexes faiblement
Perspectives
188
chargeacutees en mycobacteacuteries Drsquoautres techniques PCR pourraient ecirctre envisageacutees notamment la PCR
digitale (ou PCR numeacuterique) qui permet de gagner en sensibiliteacute et de limiter lrsquoimpact des inhibiteurs
mais dont lrsquoutilisation est encore limiteacutee par le coucirct De plus la qPCR deacutetecte lrsquoADN bacteacuterien sans
donner aucune indication sur la viabiliteacute de la cellule Devant les difficulteacutes agrave isoler M bovis par la
culture agrave partir de matrices environnementales complexes agrave cause notamment de sa croissance tregraves
lente une approche transcriptomique permettrait de preacuteciser lrsquoeacutetat viable de M bovis Cette approche
pourrait eacutegalement apporter des connaissances encore totalement inconnues agrave lrsquoheure actuelle sur les
meacutecanismes de survie mis en jeu par la bacteacuterie une fois excreacuteteacutee par son hocircte dans lrsquoenvironnement
et mettre en eacutevidence les gegravenes impliqueacutes dans son adaptation aux conditions environnementales et
donc sa persistance Dans lrsquoobjectif drsquoeacuteclaircir ces meacutecanismes qui semblent fondamentaux pour la
compreacutehension de lrsquoadaptation et de la survie de M bovis dans lrsquoenvironnement lrsquoutilisation drsquoune
approche transcriptomique permettrait drsquoidentifier les modifications transcriptionnelles agrave lrsquoeacutechelle du
geacutenome complet par une approche de seacutequenccedilage des ARN (RNA seq) et de certains gegravenes par RT-PCR
lors de lrsquointroduction de M bovis dans le sol La comparaison de ces donneacutees avec les reacutesultats obtenus
in vitro et in vivo dans diffeacuterentes conditions de stress (Rodriguez et al 2008 Jang et al 2009)
permettraient de veacuterifier si les mecircmes meacutecanismes sont impliqueacutes Les gegravenes du reacutegulon de la
dormance preacutesents chez M bovis sont surexprimeacutes dans les macrophages in vitro et in vivo (Rodriguez
et al 2008) il serait particuliegraverement inteacuteressant drsquoeacutevaluer leur niveau drsquoexpression lors de
lrsquointroduction de la bacteacuterie dans le sol Comme il est deacutelicat drsquoextraire lrsquoARN de matrices complexes
comme le sol lrsquoutilisation drsquoextraits aqueux de sol pourrait faciliter lrsquoeacutetude (Piveteau et al 2011) Pour
aller plus loin il serait inteacuteressant de comparer les transcriptomes de M bovis lorsque les conditions
abiotiques (type de sol tempeacuterature humiditeacute etc) et biotiques (comparaison absencepreacutesence du
microbiote endogegravene du sol) varient pour cerner plus finement les meacutecanismes drsquoadaptation deacuteployeacutes
face aux diffeacuterents stress
Les meacutethodes de culture de M bovis agrave partir de matrices environnementales complexes
comme le sol sont encore agrave ameacuteliorer Lrsquoisolement de souches environnementales de M bovis
permettrait gracircce aux nouvelles techniques de seacutequenccedilage (NGS) de faire de la geacutenomique
comparative sur geacutenome complet avec les mecircmes souches isoleacutees chez les animaux et drsquoameacuteliorer les
connaissances sur les meacutecanismes geacuteneacutetiques mis en jeu dans lrsquoadaptation et la persistance de la
bacteacuterie dans lrsquoenvironnement
Concernant la faune la persistance et la circulation des mycobacteacuteries dans lrsquoeau et le sol
pourraient ecirctre favoriseacutees par la micro- et la meacutesofaune preacutesentes dans ces matrices Les protozoaires
et particuliegraverement les amibes ont fait lrsquoobjet drsquoeacutetudes montrant la survie dans leurs trophozoiumltes et
leurs cystes de nombreuses espegraveces dont M bovis (Taylor et al 2003 Mba Medie et al 2011) Dans
notre eacutetude un des eacutechantillons de neacutematodes preacuteleveacute dans les seacutediments drsquoune source a donneacute un
Perspectives
189
signal moleacuteculaire reacuteveacutelant un lien potentiel entre neacutematodes et persistance des mycobacteacuteries Pour
des raisons de temps et de compeacutetences nous nrsquoavons pas exploreacute minutieusement ces eacuteventuels
reacuteservoirs cacheacutes de M bovis Devant lrsquoabondance et la diversiteacute de ces animaux dans les matrices que
nous analysons des recherches compleacutementaires nous semblent neacutecessaires pour preacuteciser leur rocircle
potentiel dans la persistance de M bovis
Les diffeacuterents reacutesultats obtenus pendant ce travail et les implications qui en ressortent ont eacuteteacute
reacuteguliegraverement partageacutes avec les instances sanitaires et agricoles de Cocircte drsquoOr afin drsquoenrichir les
connaissances sur lrsquoeacutepideacutemiologie tregraves complexe de cette maladie et de participer agrave lrsquoameacutelioration des
mesures de bioseacutecuriteacute qui en deacutecoulent Drsquoautres deacutepartements toucheacutes par la tuberculose bovine
inteacuteresseacutes par nos travaux nous ont fait parvenir des demandes drsquoanalyse environnementale dans des
zones ougrave la transmission indirecte de la maladie est fortement suspecteacutee
Les deux protocoles innovants pour lrsquoextraction des mycobacteacuteries du sol et le suivi de leur
croissance en milieu liquide deacuteveloppeacutes dans ce travail de thegravese font lrsquoobjet drsquoune eacutetude de valorisation
(protection intellectuelle et eacutetude de marcheacute) en collaboration avec la SATT Grand Est
Enfin les diffeacuterentes ideacutees et perspectives deacuteveloppeacutees dans cette thegravese ont eacuteteacute reprises en
partie dans le cadre de lrsquoappel drsquooffre Horizon 2020 agrave travers le deacutepocirct drsquoun projet europeacuteen
multipartenaire (scientifiques et entreprises) qui est coordonneacute par lrsquoINRA de Tours Nouzilly et qui
implique de nombreux partenaires en Allemagne (Lionex PME speacutecialiseacutee dans les outils de deacutetection
seacuterologique speacutecifiques) en Hollande (WUR) en UK (APHA) en Irlande (UCD) en Italie (Universiteacute de
Naples) Ce projet intituleacute DEPICT vise entre autre agrave une meilleure compreacutehension des meacutecanismes
infectieux de la reacuteaction inflammatoire et immunologique des animaux mais eacutegalement agrave deacuteterminer
le rocircle de lrsquoenvironnement dans la survie des pathogegravenes comme M bovis et Map entre autres La
diversiteacute et la persistance de M bovis et Map seront eacutetudieacutees dans lrsquoenvironnement afin de deacuteterminer
les facteurs influenccedilant la survie de ces pathogegravenes en particulier dans les sols Au travers de ce
programme et drsquoautres collaborations agrave venir avec lrsquoINRA de Tours le travail deacutebuteacute au cours de cette
thegravese sur M bovis sera eacutelargi agrave Map mycobacteacuterie eacutegalement difficilement cultivable et revecirctant une
grande importance pour la santeacute animale et la santeacute humaine (elle serait impliqueacutee dans la maladie de
Crohn) Les nouveaux outils (culture et deacutetection) deacuteveloppeacutes au cours de cette thegravese seront testeacutes sur
Map pour permettre lrsquoisolement drsquoun plus grand nombre de souches environnementales
Une des questions en suspens reste la deacutemonstration de la contamination de bovins agrave partir
de M bovis preacutesente dans des matrices environnementales Des expeacuteriences de contamination
expeacuterimentale en animalerie P3 agrave partir de sol ou drsquoeau contamineacutes pourraient permettre cette
deacutemonstration Lrsquoutilisation du modegravele cobaye espegravece tregraves sensible agrave lrsquoinfection par M bovis pour
reacutealiser des expeacuteriences preacuteliminaires pourrait ecirctre envisageacutee avant de concevoir des expeacuteriences sur
bovins Bien que ce modegravele animal soit assez eacuteloigneacute du modegravele bovin en ce qui concerne lrsquoeacutetude de la
Perspectives
190
transmission indirecte de M bovis (les cobayes geacutenegraverent par exemple beaucoup moins drsquoaeacuterosols que
les bovins quand ils boivent et ils ne ruminent pas) il constitue un modegravele plus facilement manipulable
et moins coucircteux Il permettrait drsquoaffiner les donneacutees sur les voies de transmission environnementales
potentielles avant de passer sur un modegravele bovin beaucoup plus contraignant de par la taille des
animaux et plus coucircteux
Reacutefeacuterences bibliographiques
Reacutefeacuterences bibliographiques
192
Acevedo-Whitehouse K Vicente J Gortaacutezar C Houmlfle U Fernaacutendez-de-Mera IG Amos W 2005 Genetic resistance to bovine tuberculosis in the Iberian wild boar Mol Ecol 14 3209ndash17
Adams AP Bolin SR Fine AE Bolin CA Kaneene JB 2013 Comparison of PCR versus culture for detection of Mycobacterium bovis after experimental inoculation of various matrices held under environmental conditions for extended periods Appl Environ Microbiol 79 6501ndash6506
Adekambi T Ben Salah S Khlif M Raoult D Drancourt M 2006 Survival of environmental mycobacteria in Acanthamoeba polyphaga Appl Environ Microbiol 72 5974ndash5981
Adeacutekambi T Drancourt M 2004 Dissection of phylogenetic relationships among 19 rapidly growing Mycobacterium species by 16S rRNA hsp65 sodA recA and rpoB gene sequencing Int J Syst Evol Microbiol 54 2095ndash105
Aimeacute B Lequen L Balageas A Haddad N Maugein J 2012 M bovis and M caprae infections in Aquitaine A clinico-epidemiologic study of 15 patients Pathol Biol (Paris) 60 156ndash159
Alexander K A Laver PN Michel AL Williams M van Helden PD Warren RM van Pittius NCG 2010 Novel Mycobacterium tuberculosis complex pathogen M Mungi Emerg Infect Dis 16 1296ndash1299
Allix C Walravens K Saegerman C Godfroid J Supply P Fauville-Dufaux M 2006 Evaluation of the epidemiological relevance of variable-number tandem-repeat genotyping of Mycobacterium bovis and comparison of the method with IS6110 restriction fragment length polymorphism analysis and spoligotyping J Clin Microbiol 44 1951ndash1962
Aacutelvarez AH Estrada-Chaacutevez C Flores-Valdez MA 2009 Molecular findings and approaches spotlighting Mycobacterium bovis persistence in cattle Vet Res 40 1ndash16
Ameni G Vordermeier M Firdessa R Aseffa A Hewinson G Gordon S V Berg S 2011 Mycobacterium tuberculosis infection in grazing cattle in central Ethiopia Vet J 188 359ndash361
ANSES 2011 Tuberculose bovine et faune sauvageRapport Avril 2011
Antoine D Jarlier V 2010 La tuberculose humaine agrave Mycobacterium bovis en France Bull eacutepideacutemiologique santeacute Anim Aliment Septembre
Aranaz A Cousins D Mateos A Domınguez L 2003 Elevation of Mycobacterium tuberculosis subsp caprae (Aranaz et al 1999) to species rank as Mycobacterium caprae comb nov sp Int J Syst Evol Microbiol 53 1785ndash1789
Aranaz A Lieacutebana E Goacutemez-Mampaso Enrique Galaacuten JC Cousins D Ortega A Blaacutezquez J Baquero F Mateos A Suacutearez G Domiacutenguez L 1999 Mycobacterium tuberculosis subsp caprae subsp nov a taxonomic study of a new member of the Mycobacterium tuberculosis complex isolated from goats in Spain Int J Syst Bacteriol 49 1263ndash1273
Archuleta RJ Yvonne Hoppes P Primm TP 2005 Mycobacterium avium enters a state of metabolic dormancy in response to starvation Tuberculosis (Edinb) 85 147ndash58
Aronson T Holtzman A Glover N Boian M Froman S Berlin OGW Hill H Stelma G J 1999 Comparison of large restriction fragments of Mycobacterium avium isolates recovered from AIDS and non-AIDS patients with those of isolates from potable water J Clin Microbiol 37 1008ndash1012
Ayele WY Neill SD Zinsstag J Weiss MG Pavlik I 2004 Bovine tuberculosis an old disease but a new threat to Africa Int J Tuberc Lung Dis 8 924ndash937
Ballesteros C Garrido JM Vicente J Romero B Galindo RC Minguijoacuten E Villar M Martiacuten-Hernando
Reacutefeacuterences bibliographiques
193
MP Sevilla I Juste R Aranaz A de la Fuente J Gortaacutezar C 2009 First data on Eurasian wild boar response to oral immunization with BCG and challenge with a Mycobacterium bovis field strain Vaccine 27 6662ndash6668
Barasona JA Latham MC Acevedo P Armenteros JA Latham ADM Gortazar C Carro F Soriguer RC Vicente J 2014 Spatiotemporal interactions between wild boar and cattle Implications for cross-species disease transmission Vet Res 45 1ndash11
Barasona JA Torres MJ Aznar J Gortaacutezar C Vicente J 2015 DNA detection reveals Mycobacterium tuberculosis complex shedding routes in its wildlife reservoir the Eurasian wild boar Transbound Emerg Dis (December) doi101111tbed12458
Barker J Brown MRW 1994 Trojan Horses of the microbial world protozoa and the survival of bacterial pathogens in the environment Microbiology 140 1253ndash1259
Baubet E Ropert-Coudert Y Brandt S 2003 Seasonal and annual variations in earthworm consumption by wild boar (Sus scrofa scrofa L) Wildl Res 30 179
Ben Salah I Drancourt M 2010 Surviving within the amoebal exocyst the Mycobacterium avium complex paradigm BMC Microbiol 10 99
Beacutenet J Boschiroli ML Dufour B Garin-Bastuji B 2006 Lutte contre la tuberculose bovine en France de 1954 agrave 2005 analyse de la pertinence eacutepideacutemiologique de lrsquoeacutevolution de la reacuteglementation Epideacutemiologie santeacute Anim 50 127ndash143
Benham PF 1985 The behaviour of badgers and cattle and some factors that affect the chance of contact between the species Appl Anim Behav Sci 14 390ndash391
Benham PF Broom DM 1989 Interactions between cattle and badgers at pasture with reference to bovine tuberculosis transmission Br Vet J 145 226ndash241
Bercovier H Kafri O Sela S 1986 Mycobacteria possess a surprisingly small number of ribosomal RNA genes in relation to the size of their genome Biochem Biophys Res Commun 136 1136ndash1141
Berg S Garcia-Pelayo MC Muumlller B Hailu E Asiimwe B Kremer K Dale J Boniotti MB Rodriguez S Hilty M Rigouts L Firdessa R MacHado A Mucavele C Ngandolo BNR Bruchfeld J Boschiroli ML Muumlller A Sahraoui N Pacciarini M Cadmus SI Joloba M Van Soolingen D Michel AL Djoslashnne B Aranaz A Zinsstag J Van Helden P Portaels F Kazwala R Kaumlllenius G Hewinson RG Aseffa A Gordon S V Smith NH 2011 African 2 a clonal complex of Mycobacterium bovis epidemiologically important in East Africa J Bacteriol 193 670ndash678
Bermingham ML Bishop SC Woolliams JA Pong-Wong R Allen AR McBride SH Ryder JJ Wright DM Skuce RA McDowell SW Glass EJ 2014 Genome-wide association study identifies novel loci associated with resistance to bovine tuberculosis Heredity (Edinb) 112 543ndash551
Beširović H Alić A Špičić S Cvetnić Ž Prašović S Velić L 2012 Bovine tuberculosis in Bosnia and Herzegovina caused by Mycobacterium caprae Vet Arh 82 341ndash349
Beumer A King D Donohue M Mistry J Covert T Pfaller S 2010 Detection of Mycobacterium avium subsp paratuberculosis in drinking water and biofilms by quantitative PCR Appl Environ Microbiol 76 7367ndash7370
Biek R OrsquoHare A Wright D Mallon T McCormick C Orton RJ McDowell S Trewby H Skuce RA Kao RR 2012 Whole genome sequencing reveals local transmission patterns of Mycobacterium bovis in sympatric cattle and badger populations PLoS Pathog 8 e1003008
Biet F Boschiroli ML Thorel MF Guilloteau LA 2005 Zoonotic aspects of Mycobacterium bovis and
Reacutefeacuterences bibliographiques
194
Mycobacterium avium-intracellulare complex (MAC) Vet Res36 411-436
Bille E Dauphin B Leto J Bougnoux M-E Beretti J-L Lotz A Suarez S Meyer J Join-Lambert O Descamps P Grall N Mory F Dubreuil L Berche P Nassif X Ferroni A 2012 MALDI-TOF MS Andromas strategy for the routine identification of bacteria mycobacteria yeasts Aspergillus spp and positive blood cultures Clin Microbiol Infect 18 1117ndash1125
Biswas S Rolain JM 2013 Use of MALDI-TOF mass spectrometry for identification of bacteria that are difficult to culture J Microbiol Methods 92 14ndash24
Blackwood KS He C Gunton J Turenne CY Wolfe J Kabani A M 2000 Evaluation of recA sequences for identification of Mycobacterium species J Clin Microbiol 38 2846ndash2852
Bland CS Ireland JM Lozano E Alvarez ME Primm TP 2005 Mycobacterial ecology of the Rio Grande Appl Environ Microbiol 71 5719ndash5727
Boadella M Barasona JA Diaz-Sanchez S Lyashchenko KP Greenwald R Esfandiari J Gortaacutezar C 2012 Performance of immunochromatographic and ELISA tests for detecting fallow deer infected with Mycobacterium bovis Prev Vet Med 104 160ndash164
Boadella M Lyashchenko K Greenwald R Esfandiari J Jaroso R Carta T Garrido JM Vicente J de la Fuente J Gortaacutezar C 2011 Serologic tests for detecting antibodies against Mycobacterium bovis and Mycobacterium avium subspecies paratuberculosis in Eurasian wild boar (Sus scrofa scrofa) J Vet Diagnostic Investig 23 77ndash83
Boumlddinghaus B Rogall T Flohr T Bloumlcker H Boumlttger EC 1990 Detection and identification of mycobacteria by amplification of rRNA J Clin Microbiol 28 1751ndash1759
Boumlhm M Hutchings MR White PCL 2009 Contact networks in a wildlife-livestock host community identifying high-risk individuals in the transmission of bovine TB among badgers and cattle PLoS One 4 e5016
Bollo E Guarda F Capucchio MT Galietti F 1998 Direct detection of Mycobacterium tuberculosis Complex and M avium Complex in tissue specimens from cattle through identification of specific rRNA sequences J Vet Med 45 395ndash400
Boniotti MB Goria M Loda D Garrone A Benedetto A Mondo A Tisato E Zanoni M Zoppi S Dondo A Tagliabue S Bonora S Zanardi G Pacciarini ML 2009 Molecular typing of Mycobacterium bovis strains isolated in Italy from 2000 to 2006 and evaluation of variable-number tandem repeats for geographically optimized genotyping J Clin Microbiol 47 636ndash644
Boon C Dick T 2012 How Mycobacterium tuberculosis goes to sleep the dormancy survival regulator DosR a decade later Future Microbiol 7 513ndash518
Boon C Dick T 2002 Mycobacterium bovis BCG response regulator essential for hypoxic dormancy J Bacteriol 184 6760ndash6767
Boon C Li R Qi R Dick T 2001 Proteins of Mycobacterium bovis BCG induced in the Wayne dormancy model J Bacteriol 183 2672ndash2676
Boulahbal F Robert J Trystram D Benoist A C Vincent V Jarlier V 1998 La tuberculose humaine agrave Mycobacterium bovis en France durant lrsquoanneacutee 1995 Bull eacutepideacutemiologique Hebd 48
Boyard C Vourcrsquoh G Barnouin J 2008 The relationships between Ixodes ricinus and small mammal species at the woodland-pasture interface Exp Appl Acarol 44 61ndash76
Brennan PJ Nikaido H 1995 The envelope of mycobacteria Annu Rev Biochem 64 29ndash63
Reacutefeacuterences bibliographiques
195
Brodin P Eiglmeier K Marmiesse M Billault A Garnier T Niemann S Cole ST Brosch R 2002 Bacterial artificial chromosome-based comparative genomic analysis identifies Mycobacterium microti as a natural ESAT-6 deletion mutant Infect70 5568ndash5578
Brook RK McLachlan SM 2009 Transdisciplinary habitat models for elk and cattle as a proxy for bovine tuberculosis transmission risk Prev Vet Med 91 197ndash208
Brook RK Wal E Vander van Beest FM McLachlan SM 2012 Evaluating use of cattle winter feeding areas by elk and white-tailed deer Implications for managing bovine tuberculosis transmission risk from the ground up Prev Vet Med 1ndash11
Brosch R Gordon SV Marmiesse M Brodin P Buchrieser C Eiglmeier K Garnier T Gutierrez C Hewinson G Kremer K Parsons LM Pym A Samper S van Soolingen D Cole S 2002 A new evolutionary scenario for the Mycobacterium tuberculosis complex Proc Natl Acad Sci U S A 99 3684ndash3689
Brosch R Gordon S V Garnier T Eiglmeier K Frigui W Valenti P Dos Santos S Duthoy S Lacroix C Garcia-Pelayo C Inwald JK Golby P Garcia JN Hewinson RG Behr M A Quail M A Churcher C Barrell BG Parkhill J Cole ST 2007 Genome plasticity of BCG and impact on vaccine efficacy Proc Natl Acad Sci U S A 104 5596ndash5601
Brotherstone S White IMS Coffey M Downs SH Mitchell AP Clifton-Hadley RS More SJ Good M Woolliams JA 2010 Evidence of genetic resistance of cattle to infection with Mycobacterium bovis J Dairy Sci 93 1234ndash1242
Broughan JM Crawshaw TR Downs SH Brewer J Clifton-Hadley RS 2013a Mycobacterium bovis infections in domesticated non-bovine mammalian species Part 2 A review of diagnostic methods Vet J 198 346ndash351
Broughan JM Downs SH Crawshaw TR Upton PA Brewer J Clifton-Hadley RS 2013b Mycobacterium bovis infections in domesticated non-bovine mammalian species Part 1 Review of epidemiology and laboratory submissions in Great Britain 2004-2010 Vet J 198 339ndash345
Buddle BM Wedlock DN Denis M Vordermeier HM Hewinson RG 2011 Update on vaccination of cattle and wildlife populations against tuberculosis Vet Microbiol 151 14ndash22
Butler WR Guthertz LS 2001 Mycolic acid analysis by High-Performance Liquid Chromatography for identification of Mycobacterium species Clin Microbiol Rev 14 704ndash726
Cadmus SI Adesokan HK Jenkins AO van Soolingen D 2009a Mycobacterium bovis and M tuberculosis in goats Nigeria Emerg Infect Dis 15 2066ndash2067
Cadmus SI Jenkins AO Godfroid J Osinusi K Adewole IF Murphy RL Taiwo BO 2009b Mycobacterium tuberculosis and Mycobacterium africanum in stools from children attending an immunization clinic in Ibadan Nigeria Int J Infect Dis 13 740ndash744
Cadmus SI Palmer S Okker M Dale J Gover K Smith N Jahans K Hewinson RG Gordon S V 2006 Molecular analysis of human and bovine tubercle bacilli from a local setting in Nigeria J Clin Microbiol 44 29ndash34
Cadmus SI Yakubu MK Magaji AA Jenkins AO van Soolingen D 2010 Mycobacterium bovis but also M africanum present in raw milk of pastoral cattle in north-central Nigeria Trop Anim Health Prod 42 1047ndash1048
Cambau E Lamaitre N Sougakoff W Jarlier V 2003 Reacutesistance aux antituberculeux Antibiotiques 5 29ndash37
Reacutefeacuterences bibliographiques
196
Carbonnelle B Dailloux M Lebrun L Maugein J Pernot C 2003 Cahier de biologie meacutedicale Ndeg29Mycobacteacuteries mycobacteacuterioses
Carrasco-Garcia R Barasona JA Gort Montoro V Sanchez-Vizcaino JM Vicente J 2015 Wildlife and livestock use of extensive farm resources in South Central Spain implications for disease transmission Eur J Wildl Res62(1) 65-78
Cassidy JP 2006 The pathogenesis and pathology of bovine tuberculosis with insights from studies of tuberculosis in humans and laboratory animal models Vet Microbiol 112 151ndash161
Cassidy JP Bryson DG Pollock JM Evans RT Forster F Neill SD 1999 Lesions in cattle exposed to Mycobacterium bovis-inoculated calves J Comp Pathol 121 321ndash37
Castets M Sarrat H 1968 Bacteriologic aspects of mycobacteria isolated at Dakar in 1967 Bull Soc Med Afr Noire Lang Fr 13 463ndash469
Cavalerie L Courcoul A Boschiroli ML Reacuteveillaud E Gay P 2014 Tuberculose bovine en France en 2014 une situation stable Bull eacutepideacutemiologique santeacute Anim Aliment 4ndash11
Cha D Cheng D Liu M Zeng Z Hu X Guan W 2009 Analysis of fatty acids in sputum from patients with pulmonary tuberculosis using gas chromatography-mass spectrometry preceded by solid-phase microextraction and post-derivatization on the fiber J Chromatogr A 1216 1450ndash1457
Chapinal N Elkin BT Joly DO Schumaker BA Stephen C 2012 Agreement between the caudal fold test and serological tests for the detection of Mycobacterium bovis infection in bison Prev Vet Med 105 326ndash330
Ciliberti A Gavier-Wideacuten D Yon L Hutchings MR Artois M 2015 Prioritisation of wildlife pathogens to be targeted in European surveillance programmes Expert-based risk analysis focus on ruminants Prev Vet Med 118 271ndash284
Cirillo JD Falkow S Tompkins LS Bermudez LE 1997 Interaction of Mycobacterium avium with environmental amoebae enhances virulence Infect Immun 65 3759ndash3767
Cleary GP Corner LAL OrsquoKeeffe J Marples NM 2011 Diet of the European badger (Meles meles) in the Republic of Ireland A comparison of results from an analysis of stomach contents and rectal faeces Mamm Biol 76 470ndash475
Clifton-Hadley RS Wilesmith JW Stuart F A 1993 Mycobacterium bovis in the European badger (Meles meles) epidemiological findings in tuberculous badgers from a naturally infected population Epidemiol Infect 111 9ndash19
Cole ST Brosch R Parkhill J Garnier T Churcher C Harris D Gordon S V Eiglmeier K Gas S Barry CE Tekaia F Badcock K Basham D Brown D Chillingworth T Connor R Davies R Devlin K Feltwell T Gentles S Hamlin N Holroyd S Hornsby T Jagels K Krogh A McLean J Moule S Murphy L Oliver K Osborne J Quail MA Rajandream MA Rogers J Rutter S Seeger K Skelton J Squares R Squares S Sulston JE Taylor K Whitehead S Barrell BG 1998 Deciphering the biology of Mycobacterium tuberculosis from the complete genome sequence Nature 396 190
Coleman JD Cooke MM 2001 Mycobacterium bovis infection in wildlife in New Zealand Tuberculosis (Edinb) 81 191ndash202
Collins CH Grange JM 1983 The bovine tubercle bacillus J Appl Bacteriol 55 13ndash29
Cooper SM Scott HM de la Garza GR Deck AL Cathey JC 2010 Distribution and interspecies contact of feral Swine and cattle on rangeland in South Texas implications for disease transmission J Wildl Dis 46 152ndash164
Reacutefeacuterences bibliographiques
197
Corner LAL 2006 The role of wild animal populations in the epidemiology of tuberculosis in domestic animals How to assess the risk Vet Microbiol 112 303ndash312
Corner LAL Costello E Lesellier S OrsquoMeara D Sleeman DP Gormley E 2007 Experimental tuberculosis in the European badger (Meles meles) after endobronchial inoculation of Mycobacterium bovis I Pathology and bacteriology Res Vet Sci 83 53ndash62
Corner LAL Murphy D Gormley E 2011 Mycobacterium bovis infection in the Eurasian badger (Meles meles) the disease pathogenesis epidemiology and control J Comp Pathol 144 1ndash24
Corner LAL OrsquoMeara D Costello E Lesellier S Gormley E 2012 The distribution of Mycobacterium bovis infection in naturally infected badgers Vet J 194 166ndash172
Coscolla M Gagneux S 2014 Consequences of genomic diversity in Mycobacterium tuberculosis Semin Immunol 26 431ndash444
Coscolla M Lewin A Metzger S Maetz-Rennsing K Calvignac-Spencer S Nitsche A Dabrowski PW Radonic A Niemann S Parkhill J Couacy-Hymann E Feldman J Comas I Boesch C Gagneux S Leendertz FH 2013 Novel Mycobacterium tuberculosis complex isolate from a wild chimpanzee Emerg Infect Dis 19 969ndash976
Cosivi O Grange JM Daborn CJ Raviglione MC Fujikura T Cousins D Robinson R A Huchzermeyer HF de Kantor I Meslin FX 1999 Zoonotic tuberculosis due to Mycobacterium bovis in developing countries Emerg Infect Dis 4 59ndash70
Coulombe F Divangahi M Veyrier F de Leacuteseacuteleuc L Gleason JL Yang Y Kelliher MA Pandey AK Sassetti CM Reed MB Behr M 2009 Increased NOD2-mediated recognition of N-glycolyl muramyl dipeptide J Exp Med 206 1709ndash1716
Courcoul A Moyen J-L Brugegravere L Faye S Heacutenault S Gares H Boschiroli ML 2014 Estimation of sensitivity and specificity of bacteriology histopathology and PCR for the confirmatory diagnosis of bovine tuberculosis using latent class analysis PLoS One 9 e90334
Courtenay O Reilly L A Sweeney FP Hibberd V Bryan S Ul-Hassan A Newman C Macdonald DW Delahay RJ Wilson GJ Wellington EMH 2006 Is Mycobacterium bovis in the environment important for the persistence of bovine tuberculosis Biol Lett 2 460ndash462
Cousins D V Bastida R Cataldi A Quse V Redrobe S Dow S Duignan P Murray A Dupont C Ahmed N Collins DM Butler WR Dawson D Rodriacuteguez D Loureiro J Romano MI Alito A Zumarraga M Bernardelli A 2003 Tuberculosis in seals caused by a novel member of the Mycobacterium tuberculosis complex Mycobacterium pinnipedii sp nov Int J Syst Evol Microbiol 53 1305ndash1314
Couvin D Rastogi N 2014 La mise en place de bases de donneacutees des geacutenotypes circulants du complexe Mycobacterium tuberculosis et des outils web permettant de mieux surveiller comprendre et controcircler lrsquoeacutepideacutemie de la tuberculose dans le monde EuroReference Cahier num 12
Cowie CE Hutchings MR Barasona JA Gortaacutezar C Vicente J White PCL 2015 Interactions between four species in a complex wildlife livestock disease community implications for Mycobacterium bovis maintenance and transmission Eur J Wildl Res62(1) 51-64
Cvetnic Z Katalinic-Jankovic V Sostaric B Spicic S Obrovac M Marjanovic S Benic M Kirin BK Vickovic I 2007 Mycobacterium caprae in cattle and humans in Croatia Int J Tuberc Lung Dis 11 652ndash658
Dale JW Brittain D Cataldi AA Cousins D Crawford JT Driscoll J Heersma H Lillebaek T Quitugua T Rastogi N Skuce RA Sola C Van Soolingen D Vincent V 2001 Spacer oligonucleotide typing of bacteria of the Mycobacterium tuberculosis complex Recommendations for standardised nomenclature
Reacutefeacuterences bibliographiques
198
Int J Tuberc Lung Dis 5 216ndash219
Dalley D Daveacute D Lesellier S Palmer S Crawshaw T Hewinson RG Chambers M 2008 Development and evaluation of a gamma-interferon assay for tuberculosis in badgers (Meles meles) Tuberculosis 88 235ndash243
Danelishvili L Wu M Stang B Harriff M Cirillo SLG Cirillo JD Bildfell R Arbogast B Bermudez LE 2007 Identification of Mycobacterium avium pathogenicity island important for macrophage and amoeba infection Proc Natl Acad Sci U S A 104 11038ndash11043
Dang NA Kuijper S Walters E Claassens M van Soolingen D Vivo-Truyols G Janssen H-G Kolk AHJ 2013 Validation of biomarkers for distinguishing Mycobacterium tuberculosis from non-tuberculous mycobacteria using gas chromatography-mass spectrometry and chemometrics PLoS One 8 e76263
Daniel TM 2006 The history of tuberculosis Respir Med 100 1862ndash1870
David H Levy Frebaut U Thorel MF 1989 Meacutethodes de laboratoire pour mycobacteacuteriologie clinique Institut Pasteur p 35-36
Dawson KL Bell A Kawakami RP Coley K Yates G Collins DM 2012 Transmission of Mycobacterium orygis (M tuberculosis complex species) from a tuberculosis patient to a dairy cow in New Zealand J Clin Microbiol 50 3136ndash3138
de Jong BC Antonio M Gagneux S 2010 Mycobacterium africanum-review of an important cause of human tuberculosis in West Africa PLoS Negl Trop Dis 4(9) e744
de la Rua-Domenech R 2006 Human Mycobacterium bovis infection in the United Kingdom Incidence risks control measures and review of the zoonotic aspects of bovine tuberculosis Tuberculosis (Edinb) 86 77ndash109
de la Rua-Domenech R Goodchild AT Vordermeier HM Hewinson RG Christiansen KH Clifton-Hadley RS 2006 Ante mortem diagnosis of tuberculosis in cattle a review of the tuberculin tests gamma-interferon assay and other ancillary diagnostic techniques Res Vet Sci 81 190ndash210
Dean GS Rhodes SG Coad M Adam O Cockle PJ Clifford DJ Hewinson RG Vordermeier HM 2005 Minimum infective dose of Mycobacterium bovis in cattle Infect Immun 73 6467ndash6471
Delahay RJ Cheeseman CL Clifton-Hadley RS 2001 Wildlife disease reservoirs the epidemiology of Mycobacterium bovis infection in the European badger (Meles meles) and other British mammals Tuberculosis (Edinb) 81 43ndash49
Delahay RJ De Leeuw ANS Barlow AM Clifton-Hadley RS Cheeseman CL 2002 The status of Mycobacterium bovis Infection in UK Wild Mammals A Review Vet J 164 90ndash105
Delahay RJ Langton S Smith GC Clifton-Hadley RS Cheeseman CL 2000 The spatio-temporal distribution of Mycobacterium bovis (bovine tuberculosis) infection in a high-density badger population J Anim Ecol 69 428ndash441
Delahay RJ Walker N Smith G Wilkinson D Clifton-Hadley R Cheeseman C Tomlinson A Chambers M 2013 Long-term temporal trends and estimated transmission rates for Mycobacterium bovis infection in an undisturbed high-density badger (Meles meles) population Epidemiol Infect 141 1445ndash1456
Delaune D Janvier F Rapp C Geacuterocircme P Fabre M Soler C Meacuterens A 2012 Actualiteacutes de lrsquoinfection agrave Mycobacterium bovis en France agrave propos de 4 cas Ann Biol Clin (Paris) 70 231ndash236
Desikan S Narayanan S 2015 Genetic markers genotyping methods amp next generation sequencing in Mycobacterium tuberculosis Indian J Med Res 141 761ndash774
Reacutefeacuterences bibliographiques
199
Desmond E Ahmed AT Probert WS Ely J Jang Y Sanders C A Lin SY Flood J 2004 Mycobacterium africanum Cases California Emerg Infect Dis 10 921ndash923
Dewes HF 1996 The rate of soil ingestion by dairy cows and the effect on available copper calcium sodium and magnesium N Z Vet J 44 199ndash200
Dick T Lee BH Murugasu-Oei B 1998 Oxygen depletion induced dormancy in Mycobacterium smegmatis FEMS Microbiol Lett 163 159ndash164
Dippenaar A Parsons SDC Sampson SL van der Merwe RG Drewe JA Abdallah AM Siame KK Gey van Pittius NC van Helden PD Pain A Warren RM 2015 Whole genome sequence analysis of Mycobacterium suricattae Tuberculosis 95(6) 682-688
Djelouadji Z Raoult D Drancourt M 2011 Palaeogenomics of Mycobacterium tuberculosis Epidemic bursts with a degrading genome Lancet Infect Dis 11 641ndash650
Dondo A Zoppi S Rossi F Chiavacci L Barbaro A Garrone A Benedetto A Goria M 2007 Mycobacteriosis in Wild Boar Results of 2000-2006 Activity in North-Western Italy Epideacutemiologie santeacute Anim 51 35ndash42
Drancourt M Adeacutekambi T Raoult D 2007 Interactions between Mycobacterium xenopi amoeba and human cells J Hosp Infect 65 138ndash142
Drewe JA OrsquoConnor HM Weber N McDonald RA Delahay RJ 2013 Patterns of direct and indirect contact between cattle and badgers naturally infected with tuberculosis Epidemiol Infect 141 1467ndash1475
Drewe JA Tomlinson AJ Walker NJ Delahay RJ 2010 Diagnostic accuracy and optimal use of three tests for tuberculosis in live badgers PLoS One 5 e11196
Duffield BJ Young DA 1985 Survival of Mycobacterium bovis in defined environmental conditions Vet Microbiol 10 193ndash197
Dutau G 2005 Petite histoire illustreacutee de la tuberculose The history of tuberculosis Arch Pediatr 12 88ndash95
Dziadek J Sajduda A Boruń M 2001 Specificity of insertion sequence-based PCR assays for Mycobacterium tuberculosis complex Int J Tuberc Lung Dis 5 569ndash574
EFSA 2014 The European Union summary report on trends ans sources of zoonotic agents and food-borne outbreaks in 2012 EFSA J 12 169
El Helou G Viola GM Hachem R Han XY Raad II 2013 Rapidly growing mycobacterial bloodstream infections Lancet Infect Dis 13 166ndash174
El Kheacutechine A Couderc C Flaudrops C Raoult D Drancourt M 2011 Matrix-assisted laser desorptionionization time-of-flight mass spectrometry identification of mycobacteria in routine clinical practice PLoS One 6 e24720
El Kheacutechine A Henry M Raoult D Drancourt M 2009 Detection of Mycobacterium tuberculosis complex organisms in the stools of patients with pulmonary tuberculosis Microbiology 155 2384ndash2389
Elliott GN Hough RL Avery LM Maltin C A Campbell CD 2014 Environmental risk factors in the incidence of Johnersquos disease Crit Rev Microbiol 7828 1ndash20
Evans JT Smith EG Banerjee A Smith RMM Dale J Innes JA Hunt D Tweddell A Wood A Anderson C Hewinson RG Smith NH Hawkey PM Sonnenberg P 2007 Cluster of human tuberculosis caused by Mycobacterium bovis evidence for person-to-person transmission in the UK Lancet 369 1270ndash1276
Reacutefeacuterences bibliographiques
200
Fabre M Koeck JL Le Flegraveche P Simon F Herveacute V Vergnaud G Pourcel C 2004 High genetic diversity revealed by variable-number tandem repeat genotyping and analysis of hsp65 gene polymorphism in a large collection of ldquoMycobacterium canettiirdquo strains indicates that the M tuberculosis complex is a recently emerged clone of M can J Clin Microbiol 42 3248ndash3255
Falkinham JO 2016 Nontuberculous mycobacteria community and nosocomial waterborne opportunistic pathogens Clin Microbiol Newsl 38 1ndash7
Falkinham JO Norton CD Mark W 2001 Factors influencing numbers of Mycobacterium avium Mycobacterium intracellulare and other Mycobacteria in drinking water distribution systems Appl Environ Microbiol 67 1225ndash1231
Fanning A Edwards S 1991 Mycobacterium bovis infection in human beings in contact with elk (Cervus elaphus) in Alberta Canada Lancet 338 1253ndash1255
Fediaevsky A Beacutenet J-J Boschiroli ML Riviegravere J Hars J 2012 La tuberculose bovine en France en 2011 poursuite de la reacuteduction du nombre de foyers Bull eacutepideacutemiologique santeacute Anim Aliment Novembre 4ndash12
Fediaevsky A Courcoul A Boschiroli ML Reveillaud E 2014 Tuberculose bovine en France reacutesultats drsquoune strateacutegie plus offensive Bull eacutepideacutemiologique santeacute Anim Aliment 64 4ndash11
Fediaevsky A Courcoul A Boschiroli ML Reveillaud E 2013 Tuberculose bovine en France des signaux favorables mais une situation toujours complexe dans certaines zones Bull Epideacutemiologique - Santeacute Anim Aliment 59 4ndash10
Fetene T Kebede N Alem G 2011 Tuberculosis infection in animal and human populations in three districts of Western Gojam Ethiopia Zoonoses Public Health 58 47ndash53
Fifis T Corner LAL Rothel JS Wood PR 1994 Cellular and humoral immune responses of cattle to purified Mycobacterium bovis antigens Scand J Immunol 39 267ndash274
Fine AE Bolin CA Gardiner JC Kaneene JB 2011a A study of the persistence of Mycobacterium bovis in the environment under natural weather conditions in Michigan USA Vet Med Int 2011 ID 765430 1-12
Fine AE OrsquoBrien DJ Winterstein SR Kaneene JB 2011b An effort to isolate Mycobacterium bovis from environmental substrates during investigations of bovine tuberculosis transmission sites (cattle farms and wildlife areas) in Michigan USA ISRN Vet Sci 2011 ID 787181 1-11
Fischer OA Matlova L Bartl J Dvorska L Svastova P Du Maine R Melicharek I Bartos M Pavlik I 2003 Earthworms (Oligochaeta Lumbricidae) and mycobacteria Vet Microbiol 91 325ndash338
Flores Valdez MA Schoolnik GK 2010 DosR-regulon genes induction in Mycobacterium bovis BCG under aerobic conditions Tuberculosis (Edinb) 90 197ndash200
Forrellad M Klepp L Gioffreacute A 2013 Virulence factors of the Mycobacterium tuberculosis complex Virulence 4 3ndash66
Frank W Reisinger EC Brandt-Hamerla W Schwede I Handrick W 2009 Mycobacterium microti--pulmonary tuberculosis in an immunocompetent patient Wien Klin Wochenschr 121 282ndash6
Frottier J Eliaszewicz M Arlet V Gaudillat C 1990 Infections caused by Mycobacterium africanum Bull Acad Natl Med 174 29ndash35
Gallagher J Clifton-Hadley RS 2000 Tuberculosis in badgers a review of the disease and its significance for other animals Res Vet Sci 69 203ndash217
Reacutefeacuterences bibliographiques
201
Gallagher J Horwill DM 1977 A selective oleic acid albumin agar medium for the cultivation of Mycobacterium bovis J Hyg (Lond) 79 155ndash160
Gallagher J Monies R Gavier-Widen M Rule B 1998 Role of infected non-diseased badgers in the pathogenesis of tuberculosis in the badger Vet Rec 142 710ndash714
Garbaccio S Macias A Shimizu E Paolicchi F Pezzone N Magnano G Zapata L Abdala A Tarabla H Peyru M Caimi K Zumaacuterraga M Canal A Cataldi A 2014 Association between spoligotype-VNTR types and virulence of Mycobacterium bovis in cattle Virulence 5 297ndash302
Garbaccio SG Cataldi A 2010 Evaluation of an immunomagnetic capture method followed by PCR to detect Mycobacterium bovis in tissue samples from cattle Rev Argent Microbiol 42 247ndash53
Garnett BT Delahay RJ Roper TJ 2002 Use of cattle farm resources by badgers (Meles meles) and risk of bovine tuberculosis (Mycobacterium bovis) transmission to cattle Proc Biol Sci 269 1487ndash1491
Garnier T Eiglmeier K Camus J-C Medina N Mansoor H Pryor M Duthoy S Grondin S Lacroix C Monsempe C Simon S Harris B Atkin R Doggett J Mayes R Keating L Wheeler PR Parkhill J Barrell BG Cole ST Gordon S V Hewinson RG 2003 The complete genome sequence of Mycobacterium bovis Proc Natl Acad Sci U S A 100 7877ndash7882
Gavier-Widen D Chambers MA Palmer N Newell DG Hewinson RG 2001 Pathology of natural Mycobacterium bovis infection in European badgers (Meles meles) and its relationship with bacterial excretion Vet Rec 148 299ndash304
Gavier-Wideacuten D Cooke MM Gallagher J Chambers MA Gortaacutezar C 2009 A review of infection of wildlife hosts with Mycobacterium bovis and the diagnostic difficulties of the ldquono visible lesionrdquo presentation N Z Vet J 57 122ndash131
Genov I 1965 The effects of certain physical and chemical agents on Mycobacterium tuberculosis Veteriaacuternı Med Nauk Sofia 2 97ndash107
Gerasimova A Kazakov AE Arkin AP Dubchak I Gelfand MS 2011 Comparative genomics of the dormancy regulons in mycobacteria J Bacteriol 193 3446ndash3452
Ghodbane R Mba Medie F Lepidi H Nappez C Drancourt M 2014 Long-term survival of tuberculosis complex mycobacteria in soil Microbiology 160 496ndash501
Ghosh J Larsson P Singh B Pettersson BMF Islam NM Sarkar SN Dasgupta S Kirsebom LA 2009 Sporulation in mycobacteria Proc Natl Acad Sci U S A 106 10781ndash10786
Goodchild A V Clifton-Hadley RS 2001 Cattle-to-cattle transmission of Mycobacterium bovis Tuberculosis (Edinb) 81 23ndash41
Gordon S V Heym B Parkhill J Barrell B Cole ST 1999 New insertion sequences and a novel repeated sequence in the genome of Mycobacterium tuberculosis H37Rv Microbiology 145 Pt 4 881ndash892
Gormley E 1999 Detection of Mycobacterium bovis lymphocyte stimulating antigens in culture filtrates of a recombinant Mycobacterium smegmatis cosmid library Vaccine 17 2792ndash2801
Gormley E Doyle MB Fitzsimons T McGill K Collins JD 2006 Diagnosis of Mycobacterium bovis infection in cattle by use of the gamma-interferon (Bovigam) assay Vet Microbiol 112 171ndash179
Gortaacutezar C Delahay RJ McDonald RA Boadella M Wilson GJ Gavier-Wideacuten D Acevedo P 2012 The status of tuberculosis in European wild mammals Mamm Rev 42 193ndash206
Gortaacutezar C Torres MJ Vicente J Acevedo P Reglero M de la Fuente J Negro JJ Aznar-Martiacuten J
Reacutefeacuterences bibliographiques
202
2008 Bovine tuberculosis in Dontildeana Biosphere Reserve The role of wild ungulates as disease reservoirs in the last Iberian lynx strongholds PLoS One 3(7) e2776
Grange JM 2001 Mycobacterium bovis infection in human beings Tuberculosis (Edinb) 81 71ndash77
Grange JM Yates MD 1994 Zoonotic aspects of Mycobacterium bovis infection Vet Microbiol 40 137ndash151
Grange JM Yates MD 1989 Incidence and nature of human tuberculosis due to Mycobacterium africanum in South-East England 1977-1987 Epidemiol Infect 103 127ndash132
Greub G Raoult D 2004 Microorganisms resistant to free-living amoebae Clin Microbiol Rev 17 413ndash433
Griffin JFT Rodgers CR Liggett S Mackintosh CG 2006 Tuberculosis in ruminants Characteristics of intra-tonsilar Mycobacterium bovis infection models in cattle and deer Tuberculosis 86 404ndash418
Griffin JM Martin SW Thorburn MA Eves JA Hammond RF 1996 A case-control study on the association of selected risk factors with the occurrence of bovine tuberculosis in the Republic of Ireland Prev Vet Med 27 217ndash229
Guillet-Caruba C Martinez V Doucet-Populaire F 2014 The new tools of microbiological diagnosis of tuberculosis La Rev meacutedecine interne 35 794ndash800
Gunn-Moore DA McFarland SE Brewer JI Crawshaw TR Clifton-Hadley RS Kovalik M Shaw DJ 2011 Mycobacterial disease in cats in Great Britain I Culture results geographical distribution and clinical presentation of 339 cases J Feline Med Surg 13 934ndash944
Gunn-Moore DA Jenkins PA Lucke VM 1996 Feline tuberculosis a literature review and discussion of 19 cases caused by an unusual mycobacterial variant Vet Rec 138 53ndash58
Gutierrez MC Brisse S Brosch R Fabre M Omaiumls B Marmiesse M Supply P Vincent V 2005 Ancient origin and gene mosaicism of the progenitor of Mycobacterium tuberculosis PLoS Pathog 1(1) e5
Haddad N Ostyn A Karoui C Masselot M Thorel MF Hughes SL Inwald J Hewinson RG Durand B 2001 Spoligotype diversity of Mycobacterium bovis strains isolated in France from 1979 to 2000 J Clin Microbiol 39 3623ndash3632
Harris NB Payeur J Bravo D Osorio R Stuber T Farrell D Paulson D Treviso S Mikolon A Rodriguez-Lainz A Cernek-Hoskins S Rast R Ginsberg M Kinde H 2007 Recovery of Mycobacterium bovis from soft fresh cheese originating in Mexico Appl Environ Microbiol 73 1025ndash1028
Hars J Richomme C Boschiroli ML 2010 La tuberculose bovine dans la faune sauvage en France Bull Epideacutemiologique - Santeacute Anim Aliment 38 28ndash31
Hauer A Cruz K De Cochard T Godreuil S Karaoui C Henault S Bulach T Banuls A Biet F Boschiroli ML 2015 Genetic evolution of Mycobacterium bovis causing tuberculosis in livestock and wildlife in France since 1978 PLoS One 10(2) e 0117103
Haydon DT Cleaveland S Taylor LH Laurenson MK 2002 Identifying reservoirs of infection A conceptual and practical challenge Emerg Infect Dis 8 1468ndash1473
Hennessee CT Seo JS Alvarez AM Li QX 2009 Polycyclic aromatic hydrocarbon-degrading species isolated from Hawaiian soils Mycobacterium crocinum sp nov Mycobacterium pallens sp nov Mycobacterium rutilum sp nov Mycobacterium rufum sp nov and Mycobacterium aromaticivorans sp nov Int J Syst Evol Microbiol 59 378ndash387
Hines N Payeur JB Hoffman LJ 2006 Comparison of the recovery of Mycobacterium bovis isolates using
Reacutefeacuterences bibliographiques
203
the BACTEC MGIT 960 system BACTEC 460 system and Middlebrook 7H10 and 7H11 solid media J Vet Diagn Invest 18 243ndash250
Hmama Z Pena-Diaz S Sunil J Av-Gay Y 2015 Immunoevasion and immunosuppression of the macrophage by Mycobacterium tuberculosis Immunol Rev 264 220ndash232
Hoefsloot W Van Ingen J Andrejak C Aumlngeby K Bauriaud R Bemer P Beylis N Boeree MJ Cacho J Chihota V Chimara E Churchyard G Cias R Daza R Daley CL Dekhuijzen PNR Domingo D Drobniewski F Esteban J Fauville-Dufaux M Folkvardsen DB Gibbons N Goacutemez-Mampaso E Gonzalez R Hoffmann H Hsueh PR Indra A Jagielski T Jamieson F Jankovic M Jong E Keane J Koh WJ Lange B Leao S Macedo R Mannsaringker T Marras TK Maugein J Milburn HJ Mlinkoacute T Morcillo N Morimoto K Papaventsis D Palenque E Paez-Pentildea M Piersimoni C Polanovaacute M Rastogi N Richter E Ruiz-Serrano MJ Silva A Da Silva MP Simsek H Van Soolingen D Szaboacute N Thomson R Fernandez TT Tortoli E Totten SE Tyrrell G Vasankari T Villar M Walkiewicz R Winthrop KL Wagner D 2013 The geographic diversity of nontuberculous mycobacteria isolated from pulmonary samples An NTM-NET collaborative study Eur Respir J 42 1604ndash1613
Hohn A Lowenstein 1926 Die Kulture des tuberklebacillus zur diagnose der tuberculose Zeut fur BaktI Orig 98 460ndash477
Hongmanee P Stender H Rasmussen OF 2001 Evaluation of a fluorescence in situ hybridization assay for differentiation between tuberculous and nontuberculous Mycobacterium species in smears of Lowenstein-Jensen and Mycobacteria Growth Indicator Tube cultures using peptide nucleic acid prob J Clin Microbiol 39 1032ndash1035
Hormisch D Brost I Kohring G Giffhorn F Kroppenstedt RM Stackebrandt E Faumlrber P 2004 Mycobacterium fluoranthenivorans sp nov a Fluoranthene and Aflatoxin B 1 Degrading Bacterium from Contaminated Soil of a Former Coal Gas Plant Syst Appl Microbiol 27 653ndash660
Huard RC Claudio L Lazzarini DO Butler WR Soolingen D Van Ho JL 2003 PCR-based method to differentiate the subspecies of the Mycobacterium tuberculosis complex on the basis of genomic deletions J Clin Microbiol 41 1637ndash1650
Huard RC Fabre M de Haas P Lazzarini LCO van Soolingen D Cousins D Ho JL 2006 Novel genetic polymorphisms that further delineate the phylogeny of the Mycobacterium tuberculosis complex J Bacteriol 188 4271ndash4287
Humblet MF Boschiroli ML Saegerman C 2009 Classification of worldwide bovine tuberculosis risk factors in cattle A stratified approach Vet Res 40(5) 1-24
Hutchings MR Harris S 1999 Quantifying the risks of TB infection to cattle posed by badger excreta Epidemiol Infect 122 167ndash173
Hutchings MR Harris S 1997 Effects of farm management practices on cattle grazing behaviour and the potential for transmission of bovine tuberculosis from badgers to cattle Vet J 153 149ndash162
Hutchings MR Service KM Harris S 2002 Is population density correlated with faecal and urine scent marking in European badgers (Meles meles) in the UK Mamm Biol 67 286ndash293
Hutchings MR Service KM Harris S 2001 Defecation and urination patterns of badgers Meles meles at low density in south west England- Acta Theriol (Warsz) 46 87ndash96
Huws SA McBain AJ Gilbert P 2005 Protozoan grazing and its impact upon population dynamics in biofilm communities J Appl Microbiol 98 238ndash244
Idigoras P Peacuterez-Trallero E Alcorta M Gutieacuterrez C Muntildeoz-Baroja I 1995 Rapid detection of tuberculous
Reacutefeacuterences bibliographiques
204
and non-tuberculous mycobacteria by microscopic observation of growth on Middlebrook 7H11 agar Eur J Clin Microbiol Infect Dis 14 6ndash10
Iivanainen E Martikainen PJ Vaumlaumlnaumlnen PK Vaananen PK Katila M 1999 Environmental factors affecting the occurrence of mycobacteria in brook waters J Appl Microbiol 86 673ndash681
Jackson R De Lisle GW Morris RS 1995 A study of the environmental survival of Mycobacterium bovis on a farm in New Zealand N Z Vet J 43 346ndash352
Jahans K Palmer S Inwald J Brown J Abayakoon S 2004 Isolation of Mycobacterium microti from a male Charolais-Hereford cross Vet Rec 155 373ndash374
Jalava K Jones JA Goodchild T Clifton-Hadley R Mitchell A Story A Watson JM 2007 No increase in human cases of Mycobacterium bovis disease despite resurgence of infections in cattle in the United Kingdom Epidemiol Infect 135 40ndash45
Jenkins AO Cadmus SI Venter EH Pourcel C Hauk Y Vergnaud G Godfroid J 2011 Molecular epidemiology of human and animal tuberculosis in Ibadan Southwestern Nigeria Vet Microbiol 151 139ndash147
Jenkins HE Morrison WI Cox DR Donnelly CA Johnston WT Bourne FJ Clifton-Hadley RS Gettinby G McInerney JP Watkins GH Woodroffe R 2008 The prevalence distribution and severity of detectable pathological lesions in badgers naturally infected with Mycobacterium bovis Epidemiol Infect 136 1350ndash1361
Jensen KE 1954 Presence and destruction of tubercle bacilli in sewage Bull World Health Organ 10 171ndash179
Jimeacutenez BA Hinojoza-Loza E Flores-Valdez MA Prado-Montes de Oca E Allen K Estrada-Chaacutevez C Herrera-Rodriacuteguez S Flores-Fernaacutendez JM Martiacutenez-Velaacutezquez M Hernaacutendez-Gutieacuterrez R Alvarez AH 2013 Expression of non-replicating persistence associated genes of Mycobacterium bovis in lymph nodes from skin test-reactor cattle Microb Pathog 61-62 23ndash28
Johnson L Dean G Rhodes S Hewinson G Vordermeier M Wangoo A 2007 Low-dose Mycobacterium bovis infection in cattle results in pathology indistinguishable from that of high-dose infection Tuberculosis (Edinb) 87 71ndash76
Kaal E Kolk AHJ Kuijper S Janssen H-G 2009 A fast method for the identification of Mycobacterium tuberculosis in sputum and cultures based on thermally assisted hydrolysis and methylation followed by gas chromatography-mass spectrometry J Chromatogr A 1216 6319ndash6325
Kamerbeek J Schouls L Kolk A van Agterveld M van Soolingen D Kuijper S Bunschoten A Molhuizen H Shaw R Goyal M van Embden J 1997 Simultaneous detection and strain differentiation of Mycobacterium tuberculosis for diagnosis and epidemiology J Clin Microbiol 35 907ndash914
Kaprelyants AS Gottschal JC Kell DB 1993 Dormancy in non-sporulating bacteria FEMS Microbiol Rev 104 271ndash285
Kasai H Ezaki T Harayama S 2000 Differentiation of phylogenetically related slowly growing mycobacteria by their gyrB sequences J Clin Microbiol 38(1) 301
Kazda J Pavlik I Falkinham JO Hruska K 2009 The Ecology of Mycobacteria Impact on Animalrsquos and Humans Health Springer Netherlands Dordrecht
Khan NA 2006 Acanthamoeba biology and increasing importance in human health FEMS Microbiol Rev 30 564ndash595
Kiers A Klarenbeek A Mendelts B Van Soolingen D Koeumlter G 2008 Transmission of Mycobacterium
Reacutefeacuterences bibliographiques
205
pinnipedii to humans in a zoo with marine mammals Int J Tuberc Lung Dis 12 1469ndash1473
King HC Murphy A James P Travis E Porter D Hung Y-J Sawyer J Cork J Delahay RJ Gaze W Courtenay O Wellington EM 2015a The variability and seasonality of the environmental reservoir of Mycobacterium bovis shed by wild European badgers Sci Rep 5 12318
King HC Murphy A James P Travis E Porter D Sawyer J Cork J Delahay RJ Gaze W Courtenay O Wellington EM 2015b Performance of a non-invasive test for detecting Mycobacterium bovis shedding in European badger (Meles meles) populations J Clin Microbiol 53(7) 2316-2323
Kirchheimer WF Storrs EE 1971 Attempts to establish the armadillo (Dasypus novemcinctus Linn) as a model for the study of leprosy I Report of lepromatoid leprosy in an experimentally infected armadillo Int J Lepr Other Mycobact Dis 39 693ndash702
Kirschner P Springer B Vogel U Meier A Wrede A Kiekenbeck M Bange F Bottger EC 1993 Genotypic identification of Mycobacteria by nucleic acid sequence determination report of a 2-year experience in a clinical laboratory J Clin Microbiol 31 2882ndash2889
Kleeberg HH 1984 Tuberculose humaine drsquoorigine bovine et santeacute publique Rev Sci Tech 3 33ndash54
Koeck JL Fabre M Simon F Daffeacute M Garnotel Eacute Matan A B Geacuterocircme P Bernatas JJ Buisson Y Pourcel C 2011 Clinical characteristics of the smooth tubercle bacilli ldquoMycobacterium canettiirdquo infection suggest the existence of an environmental reservoir Clin Microbiol Infect 17 1013ndash1019
Krasnow I Wayne LG 1966 Sputum Digestion I The mortality rate of tubercle bacilli in various digestion systems Am J Clin Pathol 45 352ndash355
Krishna-Prasad BN Gupta SK 1978 Preliminary report on engulfment and retention of mycobacteria by trophozoites of axenically grown Acanthamoeba castellanii Douglas Curr Sci 47 245ndash247
Kruuk H 1978 Foraging and spatial organisation of the European badger Meles meles L Behav Ecol Sociobiol 4 75ndash89
Kubica GP Dye WE Cohn ML Middlebrook GSD and D with N-A-L-CH for C of M 1963 Sputum digestion and decontamination with N-acetyl-L-cysteinemdashsodium hydroxide for culture of mycobacteria Am Rev Respir Dis 87 775ndash779
Kubica T Rusch-Gerdes S Niemann S 2003 Mycobacterium bovis subsp caprae Caused One-Third of Human M bovis-Associated Tuberculosis Cases Reported in Germany between 1999 and 2001 J Clin Microbiol 41 3070ndash3077
Kukielka E Barasona JA Cowie CE Drewe J a Gortaacutezar C Cotarelo I Vicente J 2013 Spatial and temporal interactions between livestock and wildlife in South Central Spain assessed by camera traps Prev Vet Med 112 213ndash221
Kulka K Hatfull G Ojha AK 2012 Growth of Mycobacterium tuberculosis biofilms J Vis Exp (60) e3820
Lamont EA Bannantine JP Armieacuten A Ariyakumar DS Sreevatsan S 2012 Identification and characterization of a spore-like morphotype in chronically starved Mycobacterium avium subsp paratuberculosis cultures PLoS One 7 e30648
Laprie C Duboy J Malik R Fyfe J 2013 Feline cutaneous mycobacteriosis a review of clinical pathological and molecular characterization of one case of Mycobacterium microti skin infection and nine cases of feline leprosy syndrome from France and New Caledonia Vet Dermatol 24(6) 561ndash9 e133ndash4
Lavania M Katoch K Sachan P Dubey A Kapoor S Kashyap M Chauhan DS Singh HB Sharma VD Jadhav RS Katoch VM 2006 Detection of Mycobacterium leprae DNA from soil samples by PCR
Reacutefeacuterences bibliographiques
206
targeting RLEP sequences J Commun Dis 38 269ndash273
Le Dantec C Duguet J-P Montiel A Dumoutier N Dubrou S Vincent V 2002 Occurrence of Mycobacteria in Water Treatment Lines and in Water Distribution Systems Appl Environ Microbiol 68 5318ndash5325
Lee H Park H Cho S Bai G Kim S 2000 Species identification of Mycobacteria by PCR-Restriction Fragment Length Polymorphism of the rpoB gene J Clin Microbiol 38 2966ndash2971
Lefmann M Schweickert B Buchholz P Goumlbel UB Ulrichs T Seiler P Theegarten D Moter A 2006 Evaluation of peptide nucleic acid-fluorescence in situ hybridization for identification of clinically relevant mycobacteria in clinical specimens and tissue sections J Clin Microbiol 44 3760ndash3767
Lehtola MJ Torvinen E Miettinen IT Keevil CW 2006 Fluorescence in situ hybridization using peptide nucleic acid probes for rapid detection of Mycobacterium avium subsp avium and Mycobacterium avium subsp paratuberculosis in potable-water biofilms Appl Environ Microbiol 72 848ndash853
Leistikow RL Morton RA Bartek IL Frimpong I Wagner K Voskuil MI 2010 The Mycobacterium tuberculosis dosR regulon assists in metabolic homeostasis and enables rapid recovery from nonrespiring dormancy J Bacteriol 192 1662ndash1670
Leys NM Ryngaert A Bastiaens L Wattiau P Top EM Verstraete W Springael D 2005 Occurrence and community composition of fast-growing Mycobacterium in soils contaminated with polycyclic aromatic hydrocarbons FEMS Microbiol Ecol 51 375ndash388
Li L Bannantine JP Zhang Q Amonsin A May BJ Alt D Banerji N Kanjilal S Kapur V 2005 The complete genome sequence of Mycobacterium avium subspecies paratuberculosis Proc Natl Acad Sci U S A 102 12344ndash12349
Libero G De Mori L 2014 The T-cell response to lipid antigens of Mycobacterium tuberculosis Front Immunol 5 1ndash9
Lieacutebana E Aranaz A Francis B Cousins D 1996 Assessment of genetic markers for species differentiation within the Mycobacterium tuberculosis complex J Clin Microbiol 34 933ndash938
Lieacutebana E Aranaz A Mateos A Vilafranca M Gomez-Mampaso E Tercero JC Alemany J Suarez G Domingo M Dominguez L 1995 Simple and rapid detection of Mycobacterium tuberculosis complex organisms in bovine tissue samples by PCR J Clin Microbiol 33 33ndash36
Lim A Eleuterio M Hutter B Murugasu-Oei B Dick T 1999 Oxygen depletion-induced dormancy in Mycobacterium bovis BCG J Bacteriol 181 2252ndash2256
Liss GM Wong L Kittle DC Simor A Naus M Martiquet P Misener CR 1994 Occupational exposure to Mycobacterium bovis infection in deer and elk in Ontario Can J Public Heal 85 326ndash329
Little TW Naylor PF Wilesmith JW 1982 Laboratory study of Mycobacterium bovis infection in badgers and calves Vet Rec 111 550ndash557
Lomme JR Thoen CO Himes EM Vinson JW King RE 1976 Mycobacterium tuberculosis infection in two East African oryxes J Am Vet Med Assoc 169 912ndash914
Lotz A Ferroni A Beretti J-L Dauphin B Carbonnelle E Guet-Revillet H Veziris N Heym B Jarlier V Gaillard J-L Pierre-Audigier C Frapy E Berche P Nassif X Bille E 2010 Rapid identification of mycobacterial whole cells in solid and liquid culture media by matrix-assisted laser desorption ionization-time of flight mass spectrometry J Clin Microbiol 48 4481ndash4486
Ludwig W Euzeacuteby J Schumann P Busse H-J Trujillo ME Kaumlmpfer P Whitman WB 2008 Road map
Reacutefeacuterences bibliographiques
207
of the phylum Actinobacteria Road Map of the phylum Actinobacteria Bergeys Manual of Systematic Bacteriology second Edition Volume five Springer Editions
Lugton IW Wilson PR Morris RS Nugent G 1998 Epidemiology and pathogenesis of Mycobacterium bovis infection of red deer (Cervus elaphus) in New Zealand N Z Vet J 46 147ndash156
Lyashchenko KP Greenwald R Esfandiari J OrsquoBrien DJ Schmitt SM Palmer M V Waters WR 2013 Rapid detection of serum antibody by dual-path platform VetTB assay in white-tailed deer infected with Mycobacterium bovis Clin Vaccine Immunol 20 907ndash911
Mackintosh CG Qureshi T Waldrup K Labes RE Dodds KG Griffin JFT 2000 Genetic resistance to experimental infection with Mycobacterium bovis in red deer (Cervus elaphus) Infect Immun 68 1620ndash1625
Maddock EC 1936 Experiments on the infectivity for healthy calves of bovine tubercle bacilli discharged in dung upon pasture Part I From tubercular calves fed with emulsions of Tubercle bacilli 1934-5 Part II From tubercular cows passing Tubercle bacilli in their dun J Hyg (Lond) 36 594ndash601
Maddock EC 1934 Further studies on the survival time of the bovine tubercle bacillus in soil soil and dung in dung and on grass with experiments on feeding guinea-pigs and calves on grass artificially infected with bovine tubercle bacilli J Hyg (Lond) 34 372ndash379
Maddock EC 1933 Studies on the survival time of the bovine tubercle bacillus in soil soil and dung in dung and on grass with experiments on the preliminary treatment of infected organic matter and the cultivation of the organism J Hyg (Lond) 33 103ndash117
Magee JG Ward AC 2012 Mycobacteriaceae Bergeys Manual of Systematic Bacteriology second Edition Volume five Springer Editions
Mahairas GG Sabo PJ Hickey MJ Singh DC Stover CK Mahairas GG Sabo PJ Hickey MJ Singh DC Stover CK 1996 Molecular analysis of genetic differences between Mycobacterium bovis BCG and virulent M bovis J Bacteriol 178 1274ndash1282
Mardare C Delahay RJ Dale JW 2013 Environmental amoebae do not support the long-term survival of virulent mycobacteria J Appl Microbiol 114 1388ndash1394
Marrakchi H Laneacuteelle MA Daffeacute M 2014 Mycolic acids Structures biosynthesis and beyond Chem Biol 21 67ndash85
Marri PR Bannantine JP Golding GB 2006 Comparative genomics of metabolic pathways in Mycobacterium species gene duplication gene decay and lateral gene transfer FEMS Microbiol Rev 30 906ndash925
Martiacuten-Hernando MP Houmlfle U Vicente J Ruiz-Fons F Vidal D Barral M Garrido JM de la Fuente J Gort 2007 Lesions associated with Mycobacterium tuberculosis complex infection in the European wild boar Tuberculosis 87 360ndash367
Mathema B Kurepina NE Bifani PJ Kreiswirth BN 2006 Molecular epidemiology of tuberculosis Current insights Clin Microbiol Rev 19 658ndash685
Matsuoka M Izumi S Budiawan T Nakata N Saeki K 1999 Mycobacterium leprae DNA in daily using water as a possible source of leprosy infection Indian J Lepr 71 61ndash67
Mba Medie F Ben Salah I Henrissat B Raoult D Drancourt M 2011 Mycobacterium tuberculosis complex mycobacteria as amoeba-resistant organisms PLoS One 6 e20499
McCallan L Mcnair J Skuce R 2014 A review of the potential role of cattle slurry in the spread of bovine
Reacutefeacuterences bibliographiques
208
tuberculosis 1ndash48
McCorry T Whelan AO Welsh MD McNair J Walton E Bryson DG Hewinson RG Vordermeier HM Pollock JM 2005 Shedding of Mycobacterium bovis in the nasal mucus of cattle infected experimentally with tuberculosis by the intranasal and intratracheal routes Vet Rec 157 613ndash618
McLaughlin AM Gibbons N Fitzgibbon M Power JT Foley SC Hayes JP Rogers T Keane J 2012 Primary isoniazid resistance in Mycobacterium bovis disease A prospect of concern Am J Respir Crit Care Med 186 110ndash111
McNabb A Eisler D Adie K Amos M Rodrigues M Stephens G Black WA Isaac-Renton J 2004 Assessment of partial sequencing of the 65-kilodalton heat shock protein gene (hsp65) for routine identification of Mycobacterium species isolated from clinical sources J Clin Microbiol 42 3000ndash3011
Mejia GI Castrillon L Trujillo H Robledo J A 1999 Microcolony detection in 7H11 thin layer culture is an alternative for rapid diagnosis of Mycobacterium tuberculosis infection Int J Tuberc Lung Dis 3 138ndash142
Menzies FD Neill SD 2000 Cattle-to-Cattle Transmission of Bovine Tuberculosis Vet J 160 92ndash106
Meredith A Del Pozo J Smith S Milne E Stevenson K McLuckie J 2014 Leprosy in red squirrels in Scotland Vet Rec 175 285ndash286
Michel AL Muumlller B van Helden PD 2010 Mycobacterium bovis at the animal-human interface a problem or not Vet Microbiol 140 371ndash381
Michel AL Venter L Espie IW Coetzee ML 2003 Mycobacterium tuberculosis infections in eight species at the national zoological gardens of South Africa 1991-2001 J Zoo Wildl Med 34 364ndash370
Michelet L de Cruz K Zanella G Aaziz R Bulach T Karoui C Heacutenault S Joncour G Boschiroli ML 2015 Infection with Mycobacterium microti in animals in France J Clin Microbiol 53 981ndash985
Mignard S Pichat C Carret G 2006 Mycobacterium bovis infection Lyon France Emerg Infect Dis 12 1431ndash1433
Miltgen J Morillon M Koeck J-L Varnerot A Briant J-F Nguyen G Verrot D Bonnet D Vincent V 2002 Two cases of pulmonary tuberculosis caused by Mycobacterium tuberculosis subsp canetti Emerg Infect Dis 8 1350ndash1352
Miltner EC Bermudez LE 2000 Mycobacterium avium grown in Acanthamoeba castellanii is protected from the effects of antimicrobials Antimicrob Agents Chemother 44 1990ndash1994
Mitscherlich E Marth EH 2012 Microbial survival in the environment Bacteria and rickettsiae important in human and animal health Springer S ed
Momotani E Ozaki H Hori M Yamamoto S Kuribayashi T Eda S Ikegami M 2012 Mycobacterium avium subsp paratuberculosis lipophilic antigen causes Crohnrsquos disease-type necrotizing colitis in mice Springerplus 1 1ndash10
Moore J Roper T 2003 Temperature and humidity in badger Meles meles setts Mamm Rev 33 308ndash313
Morris RS Pfeiffer DU Jackson R 1994 The epidemiology of Mycobacterium bovis infections Vet Microbiol 40 153ndash177
Mostafa HM Kazda J Irgens LM Luesse HG 1995 Acid-fast bacilli from former leprosy regions in coastal Norway showing PCR positivity for Mycobacterium leprae Int J Lepr other Mycobact Dis 63 97ndash99
Mukhopadhyay S Nair S Ghosh S 2012 Pathogenesis in tuberculosis Transcriptomic approaches to
Reacutefeacuterences bibliographiques
209
unraveling virulence mechanisms and finding new drug targets FEMS Microbiol Rev 36 463ndash485
Muumlller B Duumlrr S Alonso S Hattendorf J Laisse CJM Parsons SDC van Helden PD Zinsstag J 2013 Zoonotic Mycobacterium bovis ndashinduced tuberculosis in humans Emerg Infect Dis 19 899ndash908
Muumlller B Hilty M Berg S Garcia-Pelayo MC Dale J Boschiroli ML Cadmus SI Ngandolo BNR Godreuil S Diguimbaye-Djaibeacute C Kazwala R Bonfoh B Njanpop-Lafourcade BM Sahraoui N Guetarni D Aseffa A Mekonnen MH Razanamparany VR Ramarokoto H Djoslashnne B Oloya J Machado A Mucavele C Skjerve E Portaels F Rigouts L Michel A Muumlller A Kaumlllenius G Van Helden PD Hewinson RG Zinsstag J Gordon S V Smith NH 2009 African 1 an epidemiologically important clonal complex of Mycobacterium bovis dominant in Mali Nigeria Cameroon and Chad J Bacteriol 191 1951ndash1960
Mura M Bull T 2006 Replication and long-term persistence of bovine and human strains of Mycobacterium avium subsp paratuberculosis within Acanthamoeba polyphaga Appl Environ Microbiol 72 854ndash859
Murphy D Gormley E Costello E OrsquoMeara D Corner LAL 2010 The prevalence and distribution of Mycobacterium bovis infection in European badgers (Meles meles) as determined by enhanced post mortem examination and bacteriological culture Res Vet Sci 88 1ndash5
Musser JM Amin A Ramaswamy S 2000 Negligible genetic diversity of Mycobacterium tuberculosis host immune system protein targets Evidence of limited selective pressure Genetics 155 7ndash16
Namouchi A Didelot X Schoumlck U Gicquel B Rocha EPC 2012 After the bottleneck Genome-wide diversification of the Mycobacterium tuberculosis complex by mutation recombination and natural selection Genome Res 22 721ndash734
Narang R Narang P Mendiratta DK 2009 Isolation and identification of nontuberculous mycobacteria from water and soil in central India Indian J Med Microbiol 27 247ndash250
Naranjo V Gortaacutezar C Vicente J de la Fuente J 2008 Evidence of the role of European wild boar as a reservoir of Mycobacterium tuberculosis complex Vet Microbiol 127 1ndash9
Navarro Y Romero B Bouza E Domiacutenguez L Juan L de Garciacutea-de-Viedma D 2015 Detailed chronological analysis of microevolution events in herds infected persistently by Mycobacterium bovis Vet Microbiol 183 97-102
Neill SD Hanna J OrsquoBrien J McCracken R 1988 Excretion of Mycobacterium bovis by experimentally infected cattle Vet Rec 123 340ndash343
Neill SD Pollock JM Bryson DB Hanna J 1994 Pathogenesis of Mycobacterium bovis infection in cattle Vet Microbiol 40 41ndash52
Niemann S Richter E Daluumlgge-Tamm H Schlesinger H Graupner D Koumlnigstein B Gurath G Greinert U Ruumlsch-Gerdes S 2000 Two cases of Mycobacterium microti-derived tuberculosis in HIV-negative immunocompetent patients Emerg Infect Dis 6 539ndash542
Nishi JS Shury T Elkin BT 2006 Wildlife reservoirs for bovine tuberculosis (Mycobacterium bovis) in Canada Strategies for management and research Vet Microbiol 112 325ndash338
Norby B Fosgate GT Manning EJB Collins MT Roussel AJ 2007 Environmental mycobacteria in soil and water on beef ranches Association between presence of cultivable mycobacteria and soil and water physicochemical characteristics Vet Microbiol 124 153ndash159
Nugent G 2011 Maintenance spillover and spillback transmission of bovine tuberculosis in multi-host wildlife complexes a New Zealand case study Vet Microbiol 151 34ndash42
Reacutefeacuterences bibliographiques
210
OrsquoBrien DJ Schmitt SM Fitzgerald SD Berry DE Hickling GJ 2006 Managing the wildlife reservoir of Mycobacterium bovis the Michigan USA experience Vet Microbiol 112 313ndash323
OrsquoReilly LM Daborn CJ 1995 The epidemiology of Mycobacterium bovis infections in animals and man a review Tuber Lung Dis 76 Suppl 1 1ndash46
OIE 2009 Bovine Tuberculosis Terrestrial Manual 2009Chap 247
Ojha AK Baughn AD Sambandan D Hsu T Trivelli X Guerardel Y Alahari A Kremer L Jacobs WR Hatfull GF 2008 Growth of Mycobacterium tuberculosis biofilms containing free mycolic acids and harbouring drug-tolerant bacteria Mol Microbiol 69 164ndash174
Oloya J Kazwala R Lund A Opuda-Asibo J Demelash B Skjerve E Johansen TB Djoslashnne B 2007 Characterisation of mycobacteria isolated from slaughter cattle in pastoral regions of Uganda BMC Microbiol 7 95
Otal I Isabel O Gomez AB Kremer K de Haas P Garciacutea MJ Martiacuten C van Soolingen D 2008 Mapping of IS6110 insertion sites in Mycobacterium bovis isolates in relation to adaptation from the animal to human host Vet Microbiol 129 333ndash341
Pagnier I Raoult D La Scola B 2008 Isolation and identification of amoeba-resisting bacteria from water in human environment by using an Acanthamoeba polyphaga co-culture procedure Environ Microbiol 10 1135ndash1144
Palgrave CJ Benato L Eatwell K Laurenson IF Smith NH 2012 Mycobacterium microti infection in two meerkats (Suricata suricatta) J Comp Pathol 146 278ndash282
Palmer M V 2013 Mycobacterium bovis Characteristics of wildlife reservoir hosts Transbound Emerg Dis 60 1ndash13
Palmer M V C Thacker T Waters WR Robbe-Austerman S Harris BN 2010 Investigations on deer to deer and deer to cattle transmission of the vaccine Mycobacterium bovis bacillus Calmette-Gueacuterin (BCG) J Vaccines Vaccin 01 1ndash5
Palmer M V Waters WR Whipple DL 2004 Investigation of the transmission of Mycobacterium bovis from deer to cattle through indirect contact Am J Vet Res 65 1483ndash1489
Palmer M V Waters WR Whipple DL 2002 Lesion development in white-tailed Deer (Odocoileus virginianus) experimentally infected with Mycobacterium bovis Vet Pathol 39 334ndash340
Palmer M V Whipple DL 2006 Survival of Mycobacterium bovis on feedstuffs commonly used as supplemental feed for white-tailed deer (Odocoileus virginianus) J Wildl Dis 42 853ndash8
Palmer M V Whipple DL Rhyan JC Bolin CA Saari DA 1999 Granuloma development in cattle after intratonsilar inoculation with Mycobacterium bovis Am J Vet Res 60 310ndash315
Palmer M V Whipple DL Waters WR 2001 Experimental deer-to-deer transmission of Mycobacterium bovis Am J Vet Res 62 692ndash696
Panteix G Gutierrez MC Boschiroli ML Rouviere M Plaidy A Pressac D Porcheret H Chyderiotis G Ponsada M Van Oortegem K Salloum S Cabuzel S Bantildeuls AL Van de Perre P Godreuil S 2010 Pulmonary tuberculosis due to Mycobacterium microti a study of six recent cases in France J Med Microbiol 59 984ndash989
Parsons SDC Drewe JA van Pittius NCG Warren RM van Helden PD 2013 Novel cause of tuberculosis in meerkats South Africa Emerg Infect Dis 19 2004ndash2007
Reacutefeacuterences bibliographiques
211
Pavlik I Svastova P Bartl J Dvorska L Rychlik I 2000 Relationship between IS901 in the Mycobacterium avium complex strains isolated from birds animals humans and the environment and virulence for poultry Clin Diagn Lab Immunol 7 212ndash217
Pavlik I Trcka I Parmova I Svobodova J Melicharek I Nagy G Cvetnic Z Ocepek M Pate M Lipiec M 2005 Detection of bovine and human tuberculosis in cattle and other animals in six Central European countries during the years 2000 ndash 2004 Vet Med-Czech 2005 291ndash299
Pawlowski A Jansson M Skoumlld M Rottenberg ME Kaumlllenius G 2012 Tuberculosis and HIV co-infection PLoS Pathog 8(2) e1002464
Payne A 2014 Rocircle de la faune sauvage dans le systegraveme multi-hocirctes de Mycobacterium bovis et risque de transmission entre faune sauvage et bovins Etude expeacuterimentale en Cocircte drsquoOr Thegravese Universiteacute Lyon
Payne A Boschiroli ML Gueneau E Moyen JL Rambaud T Dufour B Gilot-Fromont E Hars J 2012 Bovine tuberculosis in ldquoEurasianrdquo badgers (Meles meles) in France Eur J Wildl Res 59 331ndash339
Payne A Chappa S Hars J Dufour B Gilot-Fromont E 2015 Wildlife visits to farm facilities assessed by camera traps in a bovine tuberculosis-infected area in France Eur J Wildl Res 62(1) 33-42
Pepperell CS Casto AM Kitchen A Granka JM Cornejo OE Holmes EC Holmes EC Birren B Galagan J Feldman MW 2013 The role of selection in shaping diversity of natural M tuberculosis populations PLoS Pathog 9 e1003543
Petroff SA 1915 Some cultural studies on the tubercle bacillus Bull Johns Hopkins Hosp 26 276ndash279
Pfyffer GE Auckenthaler R Van Embden JD a Van Soolingen D 1998 Mycobacterium canettii the smooth variant of M tuberculosis isolated from a Swiss patient exposed in Africa Emerg Infect Dis 4 631ndash634
Phillips CJC Foster CRW Morris PA Teverson R 2003 The transmission of Mycobacterium bovis infection to cattle Res Vet Sci 74 1ndash15
Pickup RW Rhodes G Bull TJ Arnott S Sidi-Boumedine K Hurley M Hermon-Taylor J 2006 Mycobacterium avium subsp paratuberculosis in lake catchments in river water abstracted for domestic use and in effluent from domestic sewage treatment works diverse opportunities for environmental cycling and human exposure Appl Environ Microbiol 72 4067ndash4077
Piersimoni C Olivieri A Benacchio L Scarparo C 2006 Current perspectives on drug susceptibility testing of Mycobacterium tuberculosis complex the automated nonradiometric systems J Clin Microbiol 44 20ndash28
Pillai SD Widmer KW Ivey LJ Coker KC Newman E Lingsweiler S Baca D Kelley M Davis DS Silvy NJ Adams LG 2000 Failure to identify non-bovine reservoirs of Mycobacterium bovis in a region with a history of infected dairy-cattle herds Prev Vet Med 43 53ndash62
Pollock JM Neill SD 2002 Mycobacterium bovis infection and tuberculosis in cattle Vet J 163 115ndash127
Pontiroli A Travis ER Sweeney FP Porter D Gaze WH Mason S Hibberd V Holden J Courtenay O Wellington EMH 2011 Pathogen quantitation in complex matrices a multi-operator comparison of DNA extraction methods with a novel assessment of PCR inhibition PLoS One 6 e17916
Prasad HK Singhal A Mishra A Shah NP Katoch VM Thakral SS Singh D V Chumber S Bal S Aggarwal S Padma M V Kumar S Singh MK Acharya SK 2005 Bovine tuberculosis in India potential basis for zoonosis Tuberculosis (Edinb) 85 421ndash428
Primm TP Lucero CA Falkinham JO 2004 Health impacts of environmental mycobacteria Clin Microbiol Rev 17 98ndash106
Reacutefeacuterences bibliographiques
212
Pritchard DG 1988 A Century of Bovine Tuberculosis 1888-1988 Conquest and Controversy J Comp Pathol 99 357ndash399
Prodinger WM Brandsta A Naumann L Pacciarini M Kubica T Boschiroli ML Aranaz A Cvetnic Z Ocepek M Skrypnyk A Erler W Niemann S Pavlik I Moser I 2005 Characterization of Mycobacterium caprae isolates from Europe by Mycobacterial Interspersed Repetitive Unit Genotyping J Clin Microbiol 43 4984ndash4992
Prodinger WM Eigentler A Allerberger F Schonbauer M Glawischnig W 2002 Infection of Red Deer Cattle and Humans with Mycobacterium bovis subsp caprae in Western Austria J Clin Microbiol 40 2270ndash2272
Prodinger WM Indra A Koksalan OK Kilicaslan Z Richter E 2014 Mycobacterium caprae infection in humans Expert Rev Anti Infect Ther12(12) 1501-1513
Pym AS Brodin P Brosch R Huerre M Cole ST 2002 Loss of RD1 contributed to the attenuation of the live tuberculosis vaccines Mycobacterium bovis BCG and Mycobacterium microti Mol Microbiol 46 709ndash717
Radomski N Betelli L Moilleron R Haenn S Moulin L Cambau E Rocher V Gonccedilalves A Lucas FS 2011 Mycobacterium behavior in wastewater treatment plant a bacterial model distinct from Escherichia coli and Enterococci Environ Sci Technol 45 5380ndash5386
Radomski N Roguet A Lucas FS Veyrier FJ Cambau E Accrombessi H Moilleron R Behr M Moulin L 2013 atpE gene as a new useful specific molecular target to quantify Mycobacterium in environmental samples BMC Microbiol 13 277
Radomski N Thibault VC Karoui C de Cruz K Cochard T Gutierrez C Supply P Biet F Boschiroli ML 2010 Determination of genotypic diversity of Mycobacterium avium subspecies from human and animal origins by Mycobacterial Interspersed Repetitive-Unit-Variable-Number Tandem-Repeat and IS1311 Restriction Fragment Length Polymorphism typing methods J Clin Microbiol 48 1026ndash1034
Rahim Z Moumlllers M Te Koppele-Vije A De Beer J Zaman K Matin MA Kamal M Raquib R Van Soolingen D Baqi M a Heilmann FGC Van Der Zanden AGM 2007 Characterization of Mycobacterium africanum subtype i among cows in a dairy farm in Bangladesh using spoligotyping Southeast Asian J Trop Med Public Health 38 706ndash713
Rauzier J Moniz-Pereira J Gicquel-Sanzey B 1988 Complete nucleotide sequence of pAL5OOO a plasmid from Mycobacterium fortuitum Gene 71 315ndash321
Renwick AR White PCL Bengis RG 2007 Bovine tuberculosis in southern African wildlife a multi-species host-pathogen system Epidemiol Infect 135 529ndash540
Reviriego-Gordejo FJ Vermeersch JP 2006 Towards eradication of bovine tuberculosis in the European Union Vet Microbiol 112 101ndash109
Richomme C Boschiroli ML Hars J Casabianca F Ducrot C 2010 Bovine tuberculosis in livestock and wild boar on the Mediterranean island Corsica J Wildl Dis 46 627ndash631
Ringuet H Akoua-Koffi C Honore S Varnerot A Vincent V Berche P Gaillard JL Pierre-Audigier C 1999 hsp65 sequencing for identification of rapidly growing mycobacteria J Clin Microbiol 37 852ndash857
Ritacco V Loacutepez B De Kantor IN Barrera L Errico F Nader A 1991 Reciprocal cellular and humoral immune responses in bovine tuberculosis Res Vet Sci 50 365ndash367
Rivero A Maacuterquez M Santos J Pinedo A Saacutenchez MA Esteve A Samper S Martiacuten C 2001 High rate of tuberculosis reinfection during a nosocomial outbreak of multidrug-resistant tuberculosis caused by
Reacutefeacuterences bibliographiques
213
Mycobacterium bovis strain B Clin Infect Dis 32 159ndash161
Riviegravere J Fediaevsky A Hars J Richomme C Calavas D Hendrikx P 2011 SYLVATUB Dispositif national de surveillance de la tuberculose bovine dans la faune sauvage Organisation institutionnelle et reacuteseau dacteurs 2011ndash2012
Rodriguez JG Burbano CS Nuntildeez C Gonzaacutelez CE Zambrano MM Garciacutea MJ Del Portillo P 2008 Rv3134cdevRdevS operon of Mycobacterium bovis BCG is differentially transcribed under ldquoin vitrordquo stress conditions Tuberculosis 88 273ndash282
Rodriacuteguez S Bezos J Romero B de Juan L Aacutelvarez J Castellanos E Moya N Lozano F Javed MT Saacuteez-Llorente JL Lieacutebana E Mateos A Domiacutenguez L Aranaz A 2011 Mycobacterium caprae infection in livestock and wildlife Spain Emerg Infect Dis 17 532ndash535
Rodriguez-Campos S Schuumlrch AC Dale J Lohan AJ Cunha M V Botelho A Cruz K Boschiroli ML Boniotti MB Pacciarini M Garcia-Pelayo MC Romero B de Juan L Domiacutenguez L Gordon S V van Soolingen D Loftus B Berg S Hewinson RG Aranaz A Smith NH 2012 European 2 - A clonal complex of Mycobacterium bovis dominant in the Iberian Peninsula Infect Genet Evol 12 866ndash872
Rodriguez-Campos S Smith NH Boniotti MB Aranaz A 2014 Overview and phylogeny of Mycobacterium tuberculosis complex organisms Implications for diagnostics and legislation of bovine tuberculosis Res Vet Sci 97 Suppl S5ndashS19
Rodriguez-Nuntildeez J Avelar FJ Marquez F Rivas-Santiago B Quintildeones C Guerrero-Barrera AL 2012 Mycobacterium tuberculosis complex detected by modified fluorescent in situ hybridization in lymph nodes of clinical samples J Infect Dev Ctries 6(1) 58ndash66
Rodriacuteguez-Zaragoza S 1994 Ecology of free-living Amoebae Crit Rev Microbiol 20(3) 225-241
Rogers LM Forrester GJ Wilson GJ Yarnell RW Cheeseman CL 2003 The role of setts in badger (Meles meles) group size breeding success and status of TB (Mycobacterium bovis) J Zool 260 209ndash215
Romero B Rodriguez S Bezos J Diaz R Francisca Copano M Merediz I Minguez O Marques S Palacios JJ De Viedma DG Saez JL Mateos A Aranaz A Dominguez L De Juan L 2011 Humans as source of Mycobacterium tuberculosis infection in cattle Spain Emerg Infect Dis 17 2393ndash2395
Roring S Scott A Brittain D Walker I Hewinson G Neill SD Skuce R 2002 Development of variable-number tandem repeat typing of Mycobacterium bovis comparison of results with those obtained by using existing exact tandem repeats and spoligotyping J Clin Microbiol 40 2126ndash2133
Roth A Fischer M Hamid ME Michalke S Ludwig W Mauch H 1998 Differentiation of phylogenetically related slowly growing mycobacteria based on 16S-23S rRNA gene internal transcribed spacer sequences J Clin Microbiol 36 139ndash147
Rothschild BM Martin LD Lev G Bercovier H Bar-Gal GK Greenblatt C Donoghue H Spigelman M Brittain D 2001 Mycobacterium tuberculosis complex DNA from an extinct bison dated 17000 years before the present Clin Infect Dis 33 305ndash311
Roug A Geoghegan C Wellington E Miller W A Travis E Porter D Cooper D Clifford DL Mazet J AK Parsons S 2014 Utility of a fecal real-time PCR protocol for detection of Mycobacterium bovis infection in African buffalo (Syncerus caffer) J Wildl Dis 50 140ndash142
Rowe MT Grant IR 2006 Mycobacterium avium ssp paratuberculosis and its potential survival tactics Lett Appl Microbiol 42 305ndash11
Safi H Barnes PF Lakey DL Shams H Samten B Vankayalapati R Howard ST 2004 IS6110 functions as a mobile monocyte-activated promoter in Mycobacterium tuberculosis Mol Microbiol 52 999ndash1012
Reacutefeacuterences bibliographiques
214
Saleeb PG Drake SK Murray PR Zelazny AM 2011 Identification of mycobacteria in solid-culture media by matrix-assisted laser desorption ionization-time of flight mass spectrometry J Clin Microbiol 49 1790ndash1794
Santos N Almeida V Gortaacutezar C Correia-Neves M 2015a Patterns of Mycobacterium tuberculosis-complex excretion and characterization of super-shedders in naturally-infected wild boar and red deer Vet Res 46 129
Santos N Santos C Valente T Gortaacutezar C Almeida V Correia-Neves M 2015b Widespread environmental contamination with Mycobacterium tuberculosis Complex revealed by a molecular detection protocol PLoS One 10 e0142079
Savelkoul PHM Catsburg A Mulder S Oostendorp L Schirm J Wilke H van der Zanden AGM Noordhoek GT 2006 Detection of Mycobacterium tuberculosis complex with Real Time PCR Comparison of different primer-probe sets based on the IS6110 element J Microbiol Methods 66 177ndash180
Scanlon MP Quinn PJ 2000a The survival of Mycobacterium bovis in sterilized cattle slurry and its relevance to the persistence of this pathogen in the environment Ir Vet J 53 412ndash415
Scanlon MP Quinn PJ 2000b Inactivation of Mycobacterium bovis in cattle slurry by five volatile chemicals J Appl Microbiol 89 854ndash861
Schnappinger D Ehrt S Voskuil MI Liu Y Mangan JA Monahan IM Dolganov G Efron B Butcher PD Nathan C Schoolnik GK 2003 Transcriptional adaptation of Mycobacterium tuberculosis within macrophages insights into the phagosomal environment J Exp Med 198 693ndash704
Schoepf K Prodinger WM Glawischnig W Hofer E Revilla-Fernandez S Hofrichter J Fritz J Koumlfer J Schmoll F 2012 A two-yearsrsquo survey on the prevalence of tuberculosis caused by Mycobacterium caprae in red deer (Cervus elaphus) in the Tyrol Austria ISRN Vet Sci 2012 245138
Schroumlder KH 1982 Isolation of M africanum from German patients Bull Int Union Tuberc 57 249ndash53
Schuumlrch AC van Soolingen D 2012 DNA fingerprinting of Mycobacterium tuberculosis from phage typing to whole-genome sequencing Infection Genetics and Evolution 12 602-609
Shepard CC 1960 The experimental disease that follows the injection of human leprosy bacilli into foot-pads of mice J Exp Med 112 445ndash454
Shiloh MU Manzanillo P Cox JS 2008 Mycobacterium tuberculosis Senses Host-Derived Carbon Monoxide during Macrophage Infection Cell Host Microbe 3 323ndash330
Shinnick TM Good RC 1994 Mycobacterial taxonomy Eur J Clin Microbiol Infect Dis 13 884ndash901
Shitikov E Ilina E Chernousova L Borovskaya A Rukin I Afanasrsquoev M Smirnova T Vorobyeva A Larionova E Andreevskaya S Kostrzewa M Govorun V 2012 Mass spectrometry based methods for the discrimination and typing of mycobacteria Infect Genet Evol 12 838ndash845
Shrikrishna D de la Rua-Domenech R Smith NH Colloff A Coutts I 2009 Human and canine pulmonary Mycobacterium bovis infection in the same household re-emergence of an old zoonotic threat Thorax 64 89ndash91
Shuralev E Quinn P Doyle M Duignan A Kwok HF Bezos J Olwill SA Gormley E Aranaz A Good M Davis WC Clarke J Whelan C 2012 Application of the Enfer chemiluminescent multiplex ELISA system for the detection of Mycobacterium bovis infection in goats Vet Microbiol 154 292ndash297
Skuce RA McCorry TP McCarroll JF Roring SMM Scott AN Brittain D Hughes SL Hewinson RG Neill SD 2002 Discrimination of Mycobacterium tuberculosis complex bacteria using novel VNTR-PCR
Reacutefeacuterences bibliographiques
215
targets Microbiology 148 519ndash528
Skuce RA Brittain D Hughes MS Beck LA Neill SD 1994 Genomic fingerprinting of Mycobacterium bovis from cattle by restriction fragment length polymorphism analysis J Clin Microbiol 32 2387ndash2392
Slana I Kaevska M Kralik P Horvathova A Pavlik I 2010 Distribution of Mycobacterium avium subsp avium and M a hominissuis in artificially infected pigs studied by culture and IS901 and IS1245 quantitative real time PCR Vet Microbiol 144 437ndash443
Smith NH 2012 The global distribution and phylogeography of Mycobacterium bovis clonal complexes Infect Genet Evol 12 857ndash865
Smith NH Berg S Dale J Allen AR Rodriguez S Romero B Matos F Ghebremichael S Karoui C Donati C Machado A da C Mucavele C Kazwala RR Hilty M Cadmus SI Ngandolo BNR Habtamu M Oloya J Muller A Milian-Suazo F Andrievskaia O Projahn M Barandiaraacuten S Maciacuteas A Muumlller B Zanini MS Ikuta CY Rodriguez CAR Pinheiro SR Figueroa A Cho S-N Mosavari N Chuang P-C Jou R Zinsstag J van Soolingen D Costello E Aseffa A Proantildeo-Perez F Portaels F Rigouts L Cataldi AA Collins DM Boschiroli ML Hewinson RG Ferreira Neto JS Surujballi O Tadyon K Botelho A Zaacuterraga AM Buller N Skuce R Michel A Aranaz A Gordon S V Jeon B-Y Kaumlllenius G Niemann S Boniotti MB van Helden PD Harris B Zumaacuterraga MJ Kremer K 2011 European 1 a globally important clonal complex of Mycobacterium bovis Infect Genet Evol 11 1340ndash1351
Smith NH Crawshaw T Parry J Birtles RJ 2009 Mycobacterium microti More diverse than previously thought J Clin Microbiol 47 2551ndash2559
Smith NH Kremer K Inwald J Dale J Driscoll JR Gordon S V Van Soolingen D Glyn Hewinson R Maynard Smith J 2006 Ecotypes of the Mycobacterium tuberculosis complex J Theor Biol 239 220ndash225
Smith NH Upton P 2012 Naming spoligotype patterns for the RD9-deleted lineage of the Mycobacterium tuberculosis complex wwwMbovisorg Infect Genet Evol 12 873ndash876
Smith RMM Drobniewski F Gibson A Montague JDE Logan MN Hunt D Hewinson G Salmon RL OrsquoNeill B 2004 Mycobacterium bovis infection United Kingdom Emerg Infect Dis 10 539ndash541
Smithwick RW Stratigos CB David HL 1975 Use of cetylpyridinium chloride and sodium chloride for the decontamination of sputum specimens that are transported to the laboratory for the isolation of Mycobacterium tuberculosis J Clin Microbiol 1 411ndash413
Soini H Musser JM 2001 Molecular diagnosis of mycobacteria Clin Chem 47 809ndash814
Sola C Filliol I Gutierrez MC Mokrousov I Vincent V Rastogi N 2001 Spoligotype database of Mycobacterium tuberculosis biogeographic distribution of shared types and epidemiologic and phylogenetic perspectives Emerg Infect Dis 7 390ndash396
Soo P-C Horng Y-T Hsueh P-R Shen B-J Wang J-Y Tu H-H Wei J-R Hsieh S-C Huang C-C Lai H-C 2006 Direct and simultaneous identification of Mycobacterium tuberculosis complex (MTBC) and Mycobacterium tuberculosis (MTB) by rapid multiplex nested PCR-ICT assay J Microbiol Methods 66 440ndash448
Sreevatsan S Pan X 1997 Restricted structural gene polymorphism in the Mycobacterium tuberculosis complex indicates evolutionarily recent global dissemination ProcNatl Acad Sci USA 94 9869ndash9874
Stahl DA Urbance JW 1990 The division between fast- and slow-growing species corresponds to natural relationships among the mycobacteria J Bacteriol 172 116ndash124
Reacutefeacuterences bibliographiques
216
Steingart KR Ng V Henry M Hopewell PC Ramsay A Cunningham J Urbanczik R Perkins MD Aziz MA Pai M 2006 Sputum processing methods to improve the sensitivity of smear microscopy for tuberculosis a systematic review Lancet Infect Dis 6 664ndash674
Stender H Lund K Petersen KH Rasmussen OF Hongmanee P Miorner H Godtfredsen SE 1999a Fluorescence In Situ Hybridization assay using peptide nucleic acid probes for differentiation between tuberculous and nontuberculous Mycobacterium species in smears of Mycobacterium cultures J Clin Microbiol 37 2760ndash2765
Stender H Mollerup TA Lund K Petersen KH Hongmanee P Godtfredsen SE 1999b Direct detection and identification of Mycobacterium tuberculosis in smear-positive sputum samples by fluorescence in situ hybridization (FISH) using peptide nucleic acid (PNA) probes Int J Tuberc lung Dis 3 830ndash837
Stewart LD McNair J McCallan L Gordon A Grant IR 2013 Improved detection of Mycobacterium bovis infection in bovine lymph node tissue using immunomagnetic separation (IMS)-based methods PLoS One 8 e58374
Stewart LD McNair J McCallan L Thompson S Kulakov LA Grant IR 2012 Production and evaluation of antibodies and phage display-derived peptide ligands for immunomagnetic separation of Mycobacterium bovis J Clin Microbiol 50 1598ndash1605
Stinear TP Mve-Obiang A Small PLC Frigui W Pryor MJ Brosch R Jenkin GA Johnson PDR Davies JK Lee RE Adusumilli S Garnier T Haydock SF Leadlay PF Cole ST 2004 Giant plasmid-encoded polyketide synthases produce the macrolide toxin of Mycobacterium ulcerans Proc Natl Acad Sci U S A 101 1345ndash1349
Sunder S Lanotte P Godreuil S Martin C Boschiroli ML Besnier JM 2009 Human-to-human transmission of tuberculosis caused by Mycobacterium bovis in immunocompetent patients J Clin Microbiol 47 1249ndash1251
Supply P Allix C Lesjean S Cardoso-Oelemann M Rusch-Gerdes S Willery E Savine E de Haas P van Deutekom H Roring S Bifani P Kurepina N Kreiswirth B Sola C Rastogi N Vatin V Gutierrez MC Fauville M Niemann S Skuce R Kremer K Locht C van Soolingen D 2006 Proposal for standardization of optimized Mycobacterial Interspersed Repetitive Unit-Variable-Number Tandem Repeat typing of Mycobacterium tuberculosis J Clin Microbiol 44 4498ndash4510
Supply P Lesjean S Savine E Kremer K Locht C 2001 Automated high-throughput genotyping for study of global epidemiology of Mycobacterium tuberculosis based on Mycobacterial Interspersed Repetitive Units J Clin 39 3563ndash3571
Supply P Marceau M Mangenot S Roche D Rouanet C Khanna V Majlessi L Criscuolo A Tap J Pawlik A Fiette L Orgeur M Fabre M Parmentier C Frigui W Simeone R Boritsch EC Debrie A-S Willery E Walker D Quail M a Ma L Bouchier C Salvignol G Sayes F Cascioferro A Seemann T Barbe V Locht C Gutierrez M-C Leclerc C Bentley SD Stinear TP Brisse S Meacutedigue C Parkhill J Cruveiller S Brosch R 2013 Genomic analysis of smooth tubercle bacilli provides insights into ancestry and pathoadaptation of Mycobacterium tuberculosis Nat Genet 45 172ndash179
Sweeney FP Courtenay O Hibberd V Hewinson RG Reilly LA Gaze WH Wellington EMH 2007 Environmental monitoring of Mycobacterium bovis in badger feces and badger sett soil by real-time PCR as confirmed by immunofluorescence immunocapture and cultivation Appl Environ Microbiol 73 7471ndash7473
Sweeney FP Courtenay O Ul-Hassan A Hibberd V Reilly LA Wellington EMH 2006 Immunomagnetic recovery of Mycobacterium bovis from naturally infected environmental samples Lett Appl Microbiol 43 364ndash369
Reacutefeacuterences bibliographiques
217
Tanner M Michel AL 1999 Investigation of the viability of M bovis under different environmental conditions in the Kruger National Park J Vet Res 185ndash190
Taquet A Tison F 1961 Nouvelle technique drsquoisolement des mycobacteacuteries par le lauryl sulfate de sodium Ann Inst Pasteur 100 676ndash680
Taylor C Jahans K Palmer S Okker M Brown J Steer K 2006 Mycobacterium microti isolated from two pigs Vet Rec 159 59ndash60
Taylor SJ Ahonen LJ de Leij FAAM Dale JW 2003 Infection of Acanthamoeba castellanii with Mycobacterium bovis BCG and survival of M bovis within the amoeba Appl Environ Microbiol 69 4316ndash4319
Telenti A Marchesi F Balz M Bally F Botrger EC Bodmer T 1993 Rapid identification of mycobacteria to the species level by Polymerase Chain Reaction and Restriction Enzyme Analysis J Clin Microbiol 31 175ndash178
Thacker TC Palmer M V Robbe-Austerman S Stuber TP Waters WR 2015 Anatomical distribution of Mycobacterium bovis genotypes in experimentally infected white-tailed deer Vet Microbiol180(1-2) 75-81
Thierry D Brisson-Noeumll A Vincent-Leacutevy-Freacutebault V Nguyen S Guesdon JL Gicquel B 1990a Characterization of a Mycobacterium tuberculosis insertion sequence IS6110 and its application in diagnosis J Clin Microbiol 28 2668ndash2673
Thierry D Cave MD Eisenach KD Crawford JT Bates JH Gicquel B Guesdon JL 1990b IS6110 an IS-like element of Mycobacterium tuberculosis complex Nucleic Acids Res 18 188
Thoen C LoBue P De Kantor I 2006 The importance of Mycobacterium bovis as a zoonosis Vet Microbiol 112 339ndash345
Thomas V Blanc D Bille J Greub G 2006 Biodiversity of amoebae resistant micro-organisms in an hospital water network Appl Environ Microbiol 72 2428ndash2438
Thomas V Loret J-F Jousset M Greub G 2008 Biodiversity of amoebae and amoebae-resisting bacteria in a drinking water treatment plant Environ Microbiol 10 2728ndash2745
Thomas V McDonnell G 2007 Relationship between mycobacteria and amoebae ecological and epidemiological concerns Lett Appl Microbiol 45 349ndash357
Thorel MF 1980 Isolation of Mycobacterium africanum from monkeys Tubercle 61 101ndash104
Tolhurst BA Delahay RJ Walker NJ Ward AI Roper TJ 2009 Behaviour of badgers (Meles meles) in farm buildings Opportunities for the transmission of Mycobacterium bovis to cattle Appl Anim Behav Sci 117 103ndash113
Torgerson PR Torgerson DJ 2010 Public health and bovine tuberculosis whatrsquos all the fuss about Trends Microbiol 18 67ndash72
Tortoli E 2014 Microbiological features and clinical relevance of new species of the genus Mycobacterium Clin Microbiol Rev 27 727ndash752
Tortoli E 2009 Clinical manifestations of nontuberculous mycobacteria Clin Microbiol Infect 15 906ndash910
Tortoli E 2003 Impact of genotypic studies on mycobacterial taxonomy the new mycobacteria of the 1990s Clin Microbiol Rev 16 319ndash354
Reacutefeacuterences bibliographiques
218
Torvinen E Lehtola MJ Martikainen PJ Miettinen IT 2007 Survival of Mycobacterium avium in drinking water biofilms as affected by water flow velocity availability of phosphorus and temperature Appl Environ Microbiol 73 6201ndash6207
Traag BA Driks A Stragier P Bitter W Broussard G Hatfull G Chu F Adams KN Ramakrishnan L Losick R 2010 Do mycobacteria produce endospores Proc Natl Acad Sci 107 878ndash881
Travis ER Gaze WH Pontiroli A Sweeney FP Porter D Mason S Keeling MJC Jones RM Sawyer J Aranaz A Rizaldos EC Cork J Delahay RJ Wilson GJ Hewinson RG Courtenay O Wellington EMH 2011 An inter-laboratory validation of a real time PCR assay to measure host excretion of bacterial pathogens particularly of Mycobacterium bovis PLoS One 6 e27369
Trujillo ME Velaacutezquez E Kroppenstedt RM Schumann P Rivas R Mateos PF Martiacutenez-Molina E 2004 Mycobacterium psychrotolerans sp nov isolated from pond water near a uranium mine Int J Syst Evol Microbiol 54 1459ndash1463
Truman R 2005 Leprosy in wild armadillos Lepr Rev 76 198ndash208
Tsairidou S Woolliams JA Allen AR Skuce RA McBride SH Wright DM Bermingham ML Pong-Wong R Matika O McDowell SWJ Glass EJ Bishop SC 2014 Genomic prediction for tuberculosis resistance in dairy cattle PLoS One 9(5) e96728
Turenne CY 2004 Mycobacterium saskatchewanense sp nov a novel slowly growing scotochromogenic species from human clinical isolates related to Mycobacterium interjectum and Accuprobe-positive for Mycobacterium avium complex Int J Syst Evol Microbiol 54 659ndash667
Van Donsel DJ Larkin EP 1977 Persistence of Mycobacterium bovis BCG in soil and on vegetables spray irrigated with sewage effluent and sludge J Food Prot 40 160ndash163
Van Ingen J Bendien SA de Lange WCM Hoefsloot W Dekhuijzen PNR Boeree MJ van Soolingen D 2009 Clinical relevance of non-tuberculous mycobacteria isolated in the Nijmegen-Arnhem region The Netherlands Thorax 64 502ndash506
Van Ingen J Rahim Z Mulder A Boeree MJ Simeone R Brosch R van Soolingen D 2012 Characterization of Mycobacterium orygis as M tuberculosis complex subspecies Emerg Infect Dis 18 653ndash655
Van Ingen J Van 2013 Diagnosis of nontuberculous mycobacterial infections Semin Respir Crit Care Med 34 103ndash109
Van Pittius NCG Perrett KD Michel AL Keet DF Hlokwe T Streicher EM Warren RM van Helden PD 2012 Infection of African Buffalo (Syncerus Caffer) by Oryx Bacillus A rare member of the antelope clade of the Mycobacterium Tuberculosis Complex J Wildl Dis 48 849ndash857
Van Soolingen D De Haas PEW Haagsma J Eger T Hermans PWM Ritacco V Alito A Van Embden JDA 1994 Use of various genetic markers in differentiation of Mycobacterium bovis strains from animals and humans and for studying epidemiology of bovine tuberculosis J Clin Microbiol 32 2425ndash2433
Van Soolingen D Hermans PWM De Haas PEW Soll DR Van Embden JDA 1991 Occurrence and stability of insertion sequences in Mycobacterium tuberculosis complex strains Evaluation of an insertion sequence-dependent DNA polymorphism as a tool in the epidemiology of tuberculosis J Clin Microbiol 29 2578ndash2586
Vandal OH Pierini LM Schnappinger D Nathan CF Ehrt S 2008 A membrane protein preserves intrabacterial pH in intraphagosomal Mycobacterium tuberculosis Nat Med 14 849ndash854
Varello K Pezzolato M Mascarino D Ingravalle F Caramelli M Bozzetta E 2008 Comparison of
Reacutefeacuterences bibliographiques
219
histologic techniques for the diagnosis of bovine tuberculosis in the framework of eradication programs J Vet Diagnostic Investig 20 164ndash169
Velayati AA Farnia P Mozafari M Malekshahian D Farahbod AM Seif S Rahideh S Mirsaeidi M 2015 Identification and genotyping of Mycobacterium tuberculosis isolated from water and soil samples of a metropolitan city CHEST J 147 1094
Vicente J Houmlfle U Garrido JM Fernaacutedez-De-Mera IG Acevedo P Juste R Barral M Gortaacutezar C 2007 Risk factors associated with the prevalence of tuberculosis-like lesions in fenced wild boar and red deer in south central Spain Vet Res 38 451ndash464
Voskuil MI Schnappinger D Rutherford R Liu Y Schoolnik GK 2004 Regulation of the Mycobacterium tuberculosis PEPPE genes Tuberculosis (Edinb) 84 256ndash262
Voskuil MI Schnappinger D Visconti KC Harrell MI Dolganov GM Sherman DR Schoolnik GK 2003 Inhibition of respiration by nitric oxide induces a Mycobacterium tuberculosis dormancy program J Exp Med 198 705ndash713
Wagner JC Buchanan G Bokkenheuser V Leviseur S 1958 An acid-fast bacillus isolated from the lungs of the Cape Hyrax Procavia capensis (Pallas) Nature 181 284ndash285
Walter WD Smith R Vanderklok M VerCauteren KC 2014 Linking bovine tuberculosis on cattle farms to white-tailed deer and environmental variables using Bayesian hierarchical analysis PLoS One 9 e90925
Wang Y Ogawa M Fukuda K Miyamoto H Taniguchi H 2006 Isolation and identification of mycobacteria from soils at an illegal dumping site and landfills in Japan Microbiol Immunol 50 513ndash524
Ward AI Judge J Delahay RJ 2010 Farm husbandry and badger behaviour Opportunities to manage badger to cattle transmission of Mycobacterium bovis Prev Vet Med 93 2ndash10
Warren RM Gey Van Pittius NC Barnard M Hesseling A Engelke E De Kock M Gutierrez MC Chege GK Victor TC Hoal EG Van Helden PD 2006 Differentiation of Mycobacterium tuberculosis complex by PCR amplification of genomic regions of difference Int J Tuberc Lung Dis 10 818ndash822
Waters WR Buddle BM Vordermeier HM Gormley E Palmer M V Thacker TC Bannantine JP Stabel JR Linscott R Martel E Milian F Foshaug W Lawrence JC 2011 Development and evaluation of an enzyme-linked immunosorbent assay for use in the detection of bovine tuberculosis in cattle Clin Vaccine Immunol 18 1882ndash1888
Wayne LG Hayes LG 1996 An in vitro model for sequential study of shiftdown of Mycobacterium tuberculosis through two stages of nonreplicating persistence Infect Immun 64 2062ndash2069
Wells AQ 1937 Tuberculosis in wild voles Lancet 229 1221
Wells AQ Robb-Smith AHT 1946 Murine type of tubercle bacillus (the vole acid-fast bacillus) Spec Rep Ser Med Res Cou London 259
Whan L Grant IR Rowe MT 2006 Interaction between Mycobacterium avium subsp paratuberculosis and environmental protozoa BMC Microbiol 6 63
Whiley H Keegan A Giglio S Bentham R 2012 Mycobacterium avium complex - the role of potable water in disease transmission J Appl Microbiol 113 223ndash32
White PCL Brown JA Harris S 1993 Badgers (Meles meles) cattle and bovine tuberculosis (Mycobacterium bovis) a hypothesis to explain the influence of habitat on the risk of disease transmission in southwest England Proc R Soc London B 253 277ndash284
Reacutefeacuterences bibliographiques
220
Whittington R Marshall D Nicholls PJ Marsh IB Reddacliff LA 2004 Survival and dormancy of Mycobacterium avium subsp paratuberculosis in the environment Appl Environ Microbiol 70 2989ndash3004
WHO 2014 Global leprosy update 2013 Wkly Epidemiol Rec 89 389ndash400
WHO 2014 Global Tuberculosis Report 2014
Williams RS Hoy WA 1930 The viability of B tuberculosis (bovinus) on pasture land in stored faeces and in liquid manure J Hyg (Lond) 30 413ndash419
Wirth T Hildebrand F Allix-Beacuteguec C Woumllbeling F Kubica T Kremer K Van Soolingen D Ruumlsch-Gerdes S Locht C Brisse S Meyer A Supply P Niemann S 2008 Origin spread and demography of the Mycobacterium tuberculosis complex PLoS Pathog 4(9) e1000160
Witmer G Fine AE Gionfriddo J Pipas M Shively K Piccolo K Burke P 2010 Epizootiologic survey of Mycobacterium bovis in wildlife and farm environments in northern Michigan J Wildl Dis 46 368ndash378
Wobeser G 2009 Bovine tuberculosis in Canadian wildlife an updated history Can Vet J 50 1169ndash1176
Wright DM Allen AR Mallon TR McDowell SWJ Bishop SC Glass EJ Bermingham ML Woolliams JA Skuce RA 2013 Field-isolated genotypes of Mycobacterium bovis vary in virulence and influence case pathology but do not affect outbreak size PLoS One 8(9) e74503
Young JS Gormley E Wellington EMH 2005 Molecular detection of Mycobacterium bovis and Mycobacterium bovis BCG (Pasteur) in soil Appl Environ Microbiol 71 1946ndash1952
Zanella G Durand B Hars J Moutou F Garin-Bastuji B Duvauchelle A Fermeacute M Karoui C Boschiroli ML 2008 Mycobacterium bovis in wildlife in France J Wildl Dis 44 99ndash108
Zelazny AM Calhoun LB L Shea YR Fischer SH 2005 Identification of Mycobacterium species by secA1 sequences J Clin Microbiol 43 1051ndash1058
Zeng W Zhang Y Zhao X Huang G Jiang Y Dong H Li X Wan K He C 2013 Occurrence of non-tuberculous mycobacteria species in livestock from northern China and first isolation of Mycobacterium caprae Epidemiol Infect 141 1545ndash1551
Zhang J Abadia E Refregier G Tafaj S Boschiroli ML Guillard B Andremont A Ruimy R Sola C 2010 Mycobacterium tuberculosis complex CRISPR genotyping Improving efficiency throughput and discriminative power of ldquospoligotypingrdquo with new spacers and a microbead-based hybridization assay J Med Microbiol 59 285ndash294
Zhang Y Yew W 2009 Mechanisms of drug resistance in Mycobacterium tuberculosis Int J Tuberc Lung Dis 13 1320ndash1330
Zolg JW Philippi-Schulz S 1994 The superoxide dismutase gene a target for detection and identification of mycobacteria by PCR J Clin Microbiol 32 2801ndash2812
Zuber B Chami M Houssin C Dubochet J Griffiths G Daffeacute M 2008 Direct visualization of the outer membrane of mycobacteria and corynebacteria in their native state J Bacteriol 190 5672ndash5680
Annexes
223
Annexe 1 Deacutetection de Mycobacterium bovis dans lrsquoenvironnement Premiers
reacutesultats des recherches meneacutees en Cocircte-drsquoOr
Deacutetection de Mycobacterium bovis dans lrsquoenvironnement
Premiers reacutesultats des recherches meneacutees en Cocircte-drsquoOr
Elodie Barbier1 et Alain Hartmann1
RESUME
Depuis les anneacutees 2000 en Cocircte-drsquoOr on assiste agrave une recrudescence des cas de tuberculose agrave
Mycobacterium bovis dans les cheptels bovins associeacutee agrave une contamination de la faune sauvage agrave
proximiteacute des eacutelevages contamineacutes Dans certaines zones du deacutepartement la faune sauvage infecteacutee
serait impliqueacutee dans le retour de la maladie aux bovins La transmission indirecte de la maladie par
lrsquoenvironnement semblerait une voie importante de la contamination entre faune sauvage et
domestique
Afin drsquoeacutevaluer la preacutevalence de M bovis dans lrsquoenvironnement et drsquoeacutevaluer le rocircle de lrsquoenvironnement
dans la transmission indirecte de la maladie nous avons deacuteveloppeacute des outils moleacuteculaires de
deacutetection adapteacutes aux matrices environnementales complexes (sol eau matiegraveres feacutecales) Ces outils agrave
la fois speacutecifiques et sensibles nous ont permis drsquoanalyser de nombreux preacutelegravevements reacutealiseacutes en 2013
Seuls deux eacutechantillons de sol se sont reacuteveacuteleacutes faiblement positifs
Mots-cleacutes tuberculose bovine bovins faune sauvage environnement PCR en temps reacuteel
SUMMARY
Since 2000 an increasing number of cases of bovine tuberculosis were found in cattle in Cocircte-drsquoOr and
in wild animals infected by Mycobacterium bovis in the vicinity of contaminated farms In some areas
wildlife could be considered as the likely source of cattle contamination We assume that
contamination between wild and domestic animals may occur indirectly by environmental factors (soil
water feces)
To quantify the prevalence of M bovis in the environment and to evaluate its role in indirect
transmission of the disease between species we developed molecular detection tools These are
adapted to complex environmental matrices (soil water faces) and are both specific and sensitive
They allowed us to analyze numerous samples collected in 2013 Two samples gave slight positive
results
Keywords Bovine tuberculosis Cattle Wildlife Environment Real time PCR
Texte de la confeacuterence preacutesenteacutee au cours de la Journeacutee scientifique AEEMA-RFSA 18 mars 2014
1 INRA UMR Agroeacutecologie 16 rue de Sully BP 86510 21065 Dijon France
Eacutepideacutemiol et santeacute anim 2014 65 Deacutetection de Mycobacterium bovis dans lrsquoenvironnement Premiers reacutesultats des recherches meneacutees en Cocircte-drsquoOr
224
I - Introduction
Deacuteclareacutee indemne de tuberculose bovine en
2000 la France connait depuis 2002 une
reacuteeacutemergence de lrsquoinfection dans certains
deacutepartements notamment en Cocircte-drsquoOr Entre
2002 et aujourdrsquohui 178 foyers ont eacuteteacute
successivement deacuteclareacutes dans ce deacutepartement
Pour la saison 2012-2013 la moitieacute des
cheptels infecteacutes avaient deacutejagrave eacuteteacute deacuteclareacutes
foyer une deux voire trois fois auparavant La
reacutepartition des cas bovins sur le deacutepartement
nrsquoest pas aleacuteatoire mais concentreacutee sur une
zone assez reacuteduite
Deux spoligotypes principaux sont retrouveacutes
chez les bovins SB0120 (VNTR 5544) appeleacute
plus simplement BCG-like dans la reacutegion de
Veacutenarey-Les-Laumes et Vitteaux et SB0134
(VNTR 5355) appeleacute GB35 dans la reacutegion de
Pouilly-en-Auxois
Parallegravelement agrave lrsquoinfection bovine et agrave
proximiteacute drsquoun cheptel foyer lrsquoinfection a eacuteteacute
deacutetecteacutee pour la premiegravere fois chez un animal
sauvage en 2003 chez un cerf Entre 2003 et
2007 seulement deux sangliers en plus du
cervideacute ont eacuteteacute trouveacutes infecteacutes Depuis 2007
et chaque anneacutee la maladie est reacuteguliegraverement
retrouveacutee chez des blaireaux et des sangliers
moins reacuteguliegraverement chez les cerfs Les
animaux sauvages infecteacutes sont
majoritairement trouveacutes agrave proximiteacute de
cheptels bovins infecteacutes
Lrsquohypothegravese drsquoun cycle eacutepideacutemiologique de
transmission de la maladie entre bovins et
faune sauvage est maintenant reconnue
[Richomme et al 2010] mecircme si celui-ci est
encore mal connu En France les exemples de
la forecirct de Brotonne [Zanella et al 2008] de la
Cocircte-drsquoOr de la Dordogne [Payne et al 2012]
de la Corse [Richomme et al 2010] et ceux de
lrsquoEspagne [Aranaz et al 2004] ont montreacute que
les spoligotypes et les profils VNTR (Variable
Number Tandem Repeat) des souches isoleacutees
chez les animaux sauvages tueacutes ou pieacutegeacutes agrave
proximiteacute des cheptels bovins infecteacutes sont
exactement les mecircmes que ceux des souches
bovines Cette identiteacute des souches reacutesulterait
drsquoune transmission directe ou indirecte entre
faune domestique et sauvage et vice et versa
Bien que quelques contacts directs soient
possibles entre faune sauvage et bovins [Boumlhm
et al 2009] il semble probable que la
transmission indirecte de M bovis soit la voie
de transmission preacutepondeacuterante entre bovin et
faune sauvage et inversement Il semble eacutetabli
que la transmission indirecte de M bovis soit la
principale voie de transmission entre bovins et
faune sauvage et inversement La transmission
indirecte ferait intervenir une excreacutetion de M
bovis par les animaux infecteacutes au niveau
respiratoire salivaire urinaire etou feacutecal les
animaux excreacuteteurs deacuteposeraient donc le
bacille dans lrsquoenvironnement par le biais de
leurs excreacutetats lrsquoenvironnement deviendrait
alors un reacuteservoir potentiel pour le bacille La
contamination drsquoanimaux sains pourrait se
produire par ingestion ou inhalation drsquoaeacuterosols
produits agrave partir des matrices
environnementales souilleacutees Par exemple la
formation de bioaeacuterosols contamineacutes par des
souches bacteacuteriennes drsquoactinomycegravetes
thermotoleacuterants est maintenant bien
documenteacutee agrave partir de matrices solides ou
liquides [Betteli et al 2013] Dans
lrsquoenvironnement des bovins ces bioaeacuterosols
pourraient ecirctre formeacutes agrave partir drsquoeau ou de
meacutelanges eau-seacutediments dans les zones
drsquoabreuvement ou agrave partir de particules fines
de sol (argiles)
Lrsquoexcreacutetion respiratoire salivaire urinaire et
feacutecale de M bovis par des blaireaux
tuberculeux a eacuteteacute documenteacutee en Angleterre
[Gallagher et al 1998 Gavier-Widen et al
2001 Corner et al 2011] De plus
Mycobacterium bovis a la capaciteacute de survivre
en dehors de ses hocirctes pendant des dureacutees
Eacutepideacutemiol et santeacute anim 2014 65 Deacutetection de Mycobacterium bovis dans lrsquoenvironnement Premiers reacutesultats des recherches meneacutees en Cocircte-drsquoOr
225
assez longues Cette dureacutee varie en fonction de
la nature du support contamineacute (eau sol foin
maiumls etc) de la tempeacuterature de lrsquohumiditeacute et
de lrsquoexposition aux UV [Courtenay et al 2006
Fine et al 2011 Ghodbane et al 2014] La
survie de M bovis dans des microcosmes de
sols contamineacutes soumis aux facteurs
climatiques a mecircme eacuteteacute deacutemontreacutee aux Etats-
Unis pour des dureacutees allant jusqursquoagrave environ 88
jours [Fine et al 2011]
La recherche de M bovis par une meacutethode
culturale dans lrsquoenvironnement drsquoanimaux
infecteacutes est souvent infructueuse [Pillai et al
2000 Michel et al 2007 Fine et al 2011] car
elle se heurte agrave plusieurs difficulteacutes i) la vitesse
de croissance tregraves lente de M bovis de lrsquoordre
de 45 jours qui impose un traitement preacutealable
des eacutechantillons de faccedilon agrave deacutetruire les micro-
organismes agrave croissance plus rapide ii) la
richesse des matrices analyseacutees (sol
seacutediments eau matiegraveres feacutecales etc) en
bacteacuteries saprophytes et en champignons tregraves
diversifieacutes qui rendent la deacutecontamination des
eacutechantillons tregraves difficile La deacutecontamination a
toutefois un effet non neacutegligeable sur la
viabiliteacute de M bovis et interfegravere avec la
sensibiliteacute de deacutetection par la mise en culture
[Fine 2006] Lrsquoimmunocapture magneacutetique a
eacuteteacute testeacutee avec succegraves en Angleterre pour isoler
et concentrer le bacille avant la mise en culture
[Sweeney et al 2006 et 2007]
La technique de PCR (Polymerase Chain
Reaction) est une technique indeacutependante de
la croissance de la bacteacuterie Quelques eacutetudes
ont permis la deacutetection par PCR du bacille dans
des sols et des latrines de blaireaux au
Royaume-Uni et en Irlande agrave proximiteacute de
cheptels bovins infecteacutes [Young et al 2005
Courtenay et al 2006 Sweeney et al 2006 et
Les objectifs de ce travail visaient en premier
lieu agrave mettre au point un systegraveme de deacutetection
moleacuteculaire agrave la fois speacutecifique du bacille et
assez sensible pour deacutetecter de faibles
concentrations dans les diffeacuterentes matrices
environnementales suspecteacutees pour leur rocircle
de reacuteservoir potentiel Ces nouveaux outils
moleacuteculaires nous ont permis drsquoanalyser les
eacutechantillons preacuteleveacutes dans des parcelles
partageacutees par les bovins et la faune sauvage
Ces parcelles ont eacuteteacute choisies car une
transmission indirecte de lrsquoinfection entre
animaux sauvages et domestiques eacutetait
suspecteacutee (origine de lrsquoinfection bovine
indeacutetermineacutee suite agrave lrsquoenquecircte
eacutepideacutemiologique et preacutesence drsquoanimaux
sauvages infecteacutes agrave proximiteacute) Cette approche
permettra drsquoeacutevaluer lrsquoimportance de la
transmission indirecte du bacille entre les
espegraveces et drsquoappreacutecier le rocircle joueacute par
lrsquoenvironnement
Cet article ne traite que de la deacutetection
moleacuteculaire la meacutethode fondeacutee sur la culture
eacutetant toujours en cours de validation
II - Mateacuteriel et meacutethodes
Souches bacteacuteriennes utiliseacutees
La souche vaccinale Mycobacterium bovis BCG
strain 1331 (SSI Copenhague Danemark) a eacuteteacute
utiliseacutee pour lrsquoeacutevaluation de la sensibiliteacute de la
PCR en temps reacuteel (qPCR) sur souche pure et
sur microcosmes de sol inoculeacute ainsi que pour
la reacutealisation des gammes eacutetalon de qPCR (PCR
quantitative) Cette souche a eacuteteacute cultiveacutee sur
geacutelose Coletsos pendant 30 jours puis remise
en suspension dans du milieu Middlebrook 7H9
enrichi en ADC (Sigma Aldrich) Apregraves mesure
de la densiteacute optique agrave 600nm la suspension a
eacuteteacute dilueacutee en cascade au 110 pour inoculer les
microcosmes de sol et drsquoeau Un
Eacutepideacutemiol et santeacute anim 2014 65 Deacutetection de Mycobacterium bovis dans lrsquoenvironnement Premiers reacutesultats des recherches meneacutees en Cocircte-drsquoOr
226
deacutenombrement des dilutions est ensuite
reacutealiseacute sur milieu Middlebrook 7H11 enrichi en
OADC (Sigma Aldrich)
Les ADN de 20 spoligotypes de M bovis et de
30 souches de mycobacteacuteries issus drsquoisolats
cliniques drsquoanimaux infecteacutes ont eacuteteacute fournis par
lrsquoANSES Maisons-Alfort pour les tests de
speacutecificiteacute (inclusion - exclusion) de la PCR en
temps reacuteel (tableau 1)
Extraction de lrsquoADN et purification
Lrsquoextraction drsquoADN et la purification des
microcosmes de sol drsquoeau et des eacutechantillons
environnementaux a eacuteteacute reacutealiseacutee selon le
protocole modifieacute deacutecrit par Ranjard et al
[2003]
Les eacutechantillons drsquoeau ont eacuteteacute filtreacutes sur des
membranes drsquoester de cellulose de 045 microm de
porositeacute Les filtres ont eacuteteacute broyeacutes avec de
lrsquoazote liquide puis lrsquoADN extrait selon la
meacutethode preacuteceacutedemment citeacutee agrave lrsquoexception de
la purification sur colonnes PVPP
(polyvinylpolypyrrolidone insoluble)
Les ADN extraits ont eacuteteacute quantifieacutes avec un
spectrophotomegravetre (NanoDropreg 2000 Thermo
Scientific) et dilueacutes au 15 110 et 120 pour la
PCR en temps reacuteel
Deacuteveloppement de trois systegravemes de
deacutetection moleacuteculaire
La bibliographie nous a permis de seacutelectionner
trois cibles geacuteneacutetiques situeacutees sur des locus
diffeacuterents tregraves speacutecifiques de M bovis [Huard
et al 2002 Brodin et al 2002] Les geacutenomes
entiegraverement seacutequenceacutes des mycobacteacuteries du
MTBC (complexe M tuberculosis comprenant
M bovis) et des MNT (mycobacteacuteries non
tuberculeuses) sur le site NCBI nous ont permis
drsquoaligner les seacutequences de ces trois gegravenes agrave la
recherche de zones tregraves conserveacutees dans le
MTBC Chaque systegraveme de deacutetection
moleacuteculaire en PCR temps reacuteel creacuteeacute comprend
une amorce Forward une amorce Reverse et
une sonde oligonucleacuteotidique marqueacutee par
deux fluorochromes (sonde TaqMan) choisies
agrave lrsquoaide du logiciel Primer3 (Simgenecom) Pour
chaque systegraveme nous avons eacutegalement conccedilu
des amorces de PCR agrave lrsquoexteacuterieur des seacutequences
preacuteceacutedentes de faccedilon agrave reacutealiser des PCR
nicheacutees (utiliseacutees uniquement en confirmation
des PCR en temps reacuteel positives)
Speacutecificiteacute et sensibiliteacute des systegravemes de
deacutetection sur souches pures sol et eau
La speacutecificiteacute des trois systegravemes de deacutetection a
eacuteteacute testeacutee par PCR en temps reacuteel sur lrsquoADN de
19 spoligotypes diffeacuterents de M bovis quatre
souches de mycobacteacuteries appartenant au
MTBC deux au Complexe avium et 20 MNT
(tableau 1) Nous avons eacutegalement veacuterifieacute que
la PCR en temps reacuteel ne donnait pas de signal
sur des ADN extraits de 22 sols provenant du
Morbihan des Yvelines du Haut-Rhin et de
Cocircte-drsquoOr preacuteleveacutes en dehors des zones ougrave
seacutevit la tuberculose bovine Ces sols preacutesentent
des caracteacuteristiques physico-chimiques varieacutees
Pour deacuteterminer la sensibiliteacute de deacutetection de
la PCR en temps reacuteel de chacun des systegravemes
la quantification a eacuteteacute reacutealiseacutee sur des dilutions
du gegravene cible cloneacute dans un plasmide (gamme
allant de 1 copie agrave 106 copies du gegravene rechercheacute
dans 5 microl) La limite de deacutetection de la PCR en
temps reacuteel dans le sol a eacuteteacute eacutevalueacutee par
lrsquoinoculation de suspension bacteacuterienne de M
bovis BCG dans un sol dont les caracteacuteristiques
physico-chimiques sont identiques aux sols
incrimineacutes en Cocircte-drsquoOr Chaque microcosme
de sol de 2 g a eacuteteacute inoculeacute avec des suspensions
bacteacuteriennes pour obtenir une concentration
finale en bacteacuteries variant de 10 agrave 105 CFU par
g de sol Des aliquotes de 100 ml drsquoeau non
steacuterile issue drsquoun ruisseau hors zone de
tuberculose ont eacuteteacute inoculeacutes avec ces mecircmes
concentrations
Eacutepideacutemiol et santeacute anim 2014 65 Deacutetection de Mycobacterium bovis dans lrsquoenvironnement Premiers reacutesultats des recherches meneacutees en Cocircte-drsquoOr
227
Tableau 1
Souches bacteacuteriennes et spoligotypes utiliseacutes pour eacutevaluer la speacutecificiteacute des trois systegravemes
de deacutetection moleacuteculaire
M bovis (spoligotypes) Mycobacteacuteries du MTBC Mycobacteacuteries atypiques Mycobacteacuteries du
Complexe avium
GB35 M microti M chitae Mavium
BCG M tuberculosis M elephantis M intracellulare
F1 M caprae M confluentis
F2 M pinnipedi M fortuitum
F5 M rufum
F6 M non chromogenicum
F7 M aichiense
F23 M thermoresitibile
F41 M xenopi
F57 M kansai
F61 M vaccae
F105 M arupense
F151 M chelonae
GB21 M porcinum
GB54 M parafinicum
SB0166 M celatum
SB0885 M intermedium
SB0999 M rutilum
SB1081 M smegmatis
M szulgai
Choix des sites de preacutelegravevements
Les eacutechantillons environnementaux ont eacuteteacute
preacuteleveacutes sur huit communes de Cocircte-drsquoOr
situeacutees dans la zone Nord entre mai et juillet
2013 Nous avons cibleacute avec lrsquoaide de la DDPP21
des parcelles
Sur lesquelles un ou plusieurs bovins
deacutecouverts infecteacutes lors de la prophylaxie
2012-2013 ont pacirctureacute en 2012 et pour
lesquels lrsquoorigine de la contamination reste
inexpliqueacutee
Autour desquelles la surveillance de la
faune sauvage a mis en eacutevidence lrsquoinfection
agrave M bovis chez des blaireaux pieacutegeacutes ou des
sangliers tueacutes agrave la chasse
Sur les pacirctures nous avons privileacutegieacute les zones
susceptibles drsquoecirctre freacutequenteacutees par les bovins
et la faune sauvage zones drsquoabreuvement
(trous drsquoeau sources abreuvoirs) zones de
nourrissage au sol De lrsquoeau du sol et des
seacutediments ont eacuteteacute reacutecolteacutes Autour des
parcelles les terriers et matiegraveres feacutecales de
blaireaux (latrines) ainsi que les zones
drsquoabreuvement de la faune sauvage ont eacuteteacute
preacuteleveacutes
Eacutepideacutemiol et santeacute anim 2014 65 Deacutetection de Mycobacterium bovis dans lrsquoenvironnement Premiers reacutesultats des recherches meneacutees en Cocircte-drsquoOr
228
III - Reacutesultats
Les systegravemes de deacutetection conccedilus sont
speacutecifiques des espegraveces appartenant au MTBC
in silico
Les gegravenes ou locus cibles ont eacuteteacute choisis soit
pour leur speacutecificiteacute agrave M bovis soit pour leur
absence chez M microti seule mycobacteacuterie
du complexe MTBC agrave avoir eacuteteacute deacutecrite dans
lrsquoenvironnement avec M bovis Les systegravemes de
deacutetection ciblant lrsquoIS1561 et la reacutegion Rv3866
ont eacuteteacute choisis car ces locus sont preacutesents chez
tous les membres du MTBC mais absents chez
M microti chez les mycobacteacuteries du complexe
avium ainsi que chez les MNT [Huard et al
2002 Brodin et al 2002] La reacutegion RD4
preacutesente chez tous les membres du MTBC est
absente chez M bovis [Huard et al 2002]
Deux systegravemes de deacutetection moleacuteculaire
ciblant lrsquoIS1561 et la reacutegion Rv3866 ont eacuteteacute
reacutealiseacutes agrave lrsquoaide du logiciel Primer3 Le troisiegraveme
systegraveme ciblant la zone encadrant la RD4 utilise
une amorce Forward et une sonde deacutejagrave deacutecrite
[Sweeney et al 2007] seule lrsquoamorce Reverse
a eacuteteacute redessineacutee afin drsquoobtenir un produit PCR
plus petit La sonde utiliseacutee ne peut se fixer que
sur lrsquoADN des souches preacutesentant la deacuteleacutetion La
speacutecificiteacute des systegravemes de deacutetection a eacuteteacute
deacutemontreacutee in silico par alignement de
seacutequences agrave lrsquoaide du programme BLASTreg
Aucune deacutetection croiseacutee avec drsquoautres
espegraveces que celles appartenant au MTBC nrsquoa
eacuteteacute mise en eacutevidence in silico pour les systegravemes
de deacutetection utiliseacutes
Speacutecificiteacute et sensibiliteacute des systegravemes de
deacutetection sur souches pures et microcosmes
de sol inoculeacutes
Sur les 18 spoligotypes de M bovis testeacutes tous
ont donneacute un signal fort et conserveacute lors de la
PCR en temps reacuteel (Cycle threshold = 20)
Dans le complexe MTBC nous avons testeacute nos
systegravemes sur M microti M tuberculosis M
caprae et M pinnipedii Le systegraveme RD4
preacutesente la plus grande speacutecificiteacute puisqursquoil
nrsquoamplifie que M bovis parmi les membres du
complexe testeacutes LrsquoIS1561 et la reacutegion Rv3866
donnent un signal identique pour M bovis M
caprae M tuberculosis et M pinnipedii mais
ces systegravemes nrsquoamplifient pas M microti
Mecircme srsquoil a eacuteteacute montreacute que M tuberculosis
peut survivre expeacuterimentalement dans le sol
(Ghodbane et al 2014) seuls M microti et M
bovis y ont eacuteteacute deacutejagrave eacuteteacute deacutecrits [Courtenay et
al 2006 Sweeney et al 2006 et 2007]
Sur les 20 souches de MNT testeacutees cinq ont
donneacute un tregraves faible signal (Ct gt 38) avec les
trois systegravemes de deacutetection Pour le complexe
avium seul M avium a eacuteteacute deacutetecteacute avec un
Ctgt38
Aucun des 22 sols testeacutes nrsquoa donneacute de signal en
PCR en temps reacuteel quel que soit le systegraveme de
deacutetection utiliseacute ce qui indique une tregraves bonne
speacutecificiteacute de ces systegravemes de deacutetection sur
une large gamme de sols aux caracteacuteristiques
physico-chimiques diffeacuterentes et contenant des
communauteacutes microbiennes varieacutees
En ce qui concerne la sensibiliteacute des systegravemes
la limite de deacutetection des cibles se situe entre 1
et 10 copies lorsqursquoon utilise des solutions de
plasmide dilueacutees La reacutepeacutetabiliteacute de la
quantification des dilutions de plasmides est
tregraves bonne entre 10 et 106 copies de gegravenes avec
les trois systegravemes En revanche en dessous de
10 copies la deacutetection est moins reacutepeacutetable
(non deacutetection dans certaines reacutepeacutetitions)
Sur les microcosmes de sol et drsquoeau inoculeacutes la
limite de deacutetection des cibles se situe agrave 103
copies du gegravene par g de sol et agrave 10 copies du
gegravene par ml drsquoeau
Le systegraveme ciblant lrsquoIS1561 semble ecirctre tregraves
leacutegegraverement plus sensible que les deux autres
Nous lrsquoavons donc utiliseacute en 1egravere intention pour
Eacutepideacutemiol et santeacute anim 2014 65 Deacutetection de Mycobacterium bovis dans lrsquoenvironnement Premiers reacutesultats des recherches meneacutees en Cocircte-drsquoOr
229
cribler lrsquoensemble des eacutechantillons
environnementaux
Premiers reacutesultats sur les eacutechantillons
environnementaux
Sur les huit communes cibleacutees 150
eacutechantillons de sol 18 eacutechantillons de
seacutediments cinq eacutechantillons drsquoeau et huit
eacutechantillons de fegraveces de blaireaux ont eacuteteacute
preacuteleveacutes Parmi les 150 sols preacuteleveacutes 28 lrsquoont
eacuteteacute dans des terriers de blaireaux et de renards
Parmi les terriers de blaireaux preacuteleveacutes
certains eacutetaient potentiellement des terriers de
blaireaux infecteacutes par M bovis car des
animaux pieacutegeacutes agrave proximiteacute se sont reacuteveacuteleacutes ecirctre
porteurs de M bovis Cependant en 2013 nous
nrsquoavons pu obtenir aucune certitude sur les
terriers reacuteellement occupeacutes par les blaireaux
porteurs de M bovis (pieacutegeage des blaireaux en
couleacutee et non pas en gueule de terrier)
Sur les 181 eacutechantillons analyseacutes seuls deux
eacutechantillons de sol dans deux communes
diffeacuterentes ont donneacute un signal en PCR en
temps reacuteel avec lrsquoIS1561 agrave lrsquoune des dilutions
seulement Ce signal est toutefois faible et nrsquoa
pas pu ecirctre reproduit avec les autres systegravemes
de deacutetection Un des eacutechantillons positifs
provient drsquoun terrier agrave proximiteacute duquel un
blaireau infecteacute a eacuteteacute pieacutegeacute Le second
eacutechantillon de sol a eacuteteacute preacuteleveacute dans une latrine
de blaireau creuseacutee sous une clocircture de preacute
Lrsquoanalyse des fegraveces de cette latrine nrsquoa toutefois
pas donneacute de signal
La campagne de preacutelegravevement 2014 sera
reacutealiseacutee pour augmenter le nombre
drsquoeacutechantillons preacuteleveacutes sur une zone plus
restreinte en particulier ougrave des contaminations
reacutecurrentes des cheptels bovins ont eacuteteacute et sont
observeacutees
IV - Discussion
Mycobacterium bovis lrsquoagent de la tuberculose
bovine est une bacteacuterie pathogegravene qui infecte
les bovins et certains mammifegraveres sauvages en
France La maicirctrise de la maladie chez les
bovins dans certains deacutepartements franccedilais
semble ecirctre entraveacutee par la proximiteacute
drsquoanimaux sauvages infecteacutes La transmission
indirecte du bacille entre faune sauvage et
domestique par lrsquointermeacutediaire de
lrsquoenvironnement permettrait drsquoexpliquer
certains cas de re-contaminations chez les
bovins Lrsquoenvironnement pourrait servir de
reacuteservoir de la bacteacuterie par le biais de
lrsquoexcreacutetion animale il pourrait en retour
infecter des animaux sains soit par inhalation
drsquoaeacuterosols contamineacutes soit par ingestion
(abreuvement) Quelques eacutetudes en
Angleterre et en Irlande ont mis en eacutevidence la
preacutesence de M bovis dans des sols de preacutes des
sols de terriers et des latrines de blaireaux agrave
proximiteacute de cheptels bovins infecteacutes
[Courtenay et al 2006 Sweeney et al 2006
et 2007 Young et al 2005 Travis et al 2011]
Des eacutetudes expeacuterimentales ont montreacute que M
bovis pouvait survivre plusieurs mois dans le sol
[Maddock 1933 Young et al 2005 Fine et al
2011 Ghodbane et al 2014] tout en restant
virulent [Maddock 1933 Ghodbane et al
2014] La deacutetection du bacille dans
lrsquoenvironnement freacutequenteacute par les animaux
domestiques ou sauvages infecteacutes nrsquoa jamais
eacuteteacute deacutecrite en France Dans certaines zones de
Cocircte-drsquoOr la transmission indirecte du bacille
aux bovins par lrsquoenvironnement est fortement
suspecteacutee
Dans cette eacutetude nous avons deacuteveloppeacute trois
systegravemes de deacutetection moleacuteculaire afin
drsquooptimiser la mise en eacutevidence de M bovis
Eacutepideacutemiol et santeacute anim 2014 65 Deacutetection de Mycobacterium bovis dans lrsquoenvironnement Premiers reacutesultats des recherches meneacutees en Cocircte-drsquoOr
230
dans des eacutechantillons environnementaux
complexes afin de srsquoaffranchir de la culture En
effet lrsquoisolement de M bovis dans les matrices
environnementales par la culture est long car
drsquoune part un traitement preacutealable est
neacutecessaire pour srsquoaffranchir de la flore
microbienne de contamination et drsquoautre part
le bacille a une croissance tregraves lente
Contrairement agrave la culture la technique de PCR
en temps reacuteel permet une deacutetection rapide du
bacille sous reacuteserve que le ou les gegravenes cibleacutes
soient tregraves speacutecifiques de faccedilon agrave deacutetecter et
quantifier une espegravece bacteacuterienne particuliegravere
dans un meacutelange drsquoADN complexe Mais cette
technique rencontre neacuteanmoins quelques
limites techniques
Les eacutechantillons environnementaux
contiennent une flore microbienne et animale
abondante et varieacutee lrsquoADN extrait contient une
grande diversiteacute de seacutequences geacuteneacutetiques agrave
lrsquoorigine de faux positifs Ainsi des espegraveces
proches de M bovis notamment les
mycobacteacuteries environnementales ainsi que
des espegraveces encore inconnues peuvent ecirctre
preacutesentes dans les eacutechantillons Il est donc
neacutecessaire de srsquoassurer de la speacutecificiteacute des
systegravemes de deacutetection La deacutetection du bacille
par la recherche associeacutee de trois cibles
geacuteneacutetiques avec le systegraveme TaqMan nous a
permis de deacutevelopper une meacutethode de
deacutetection moleacuteculaire speacutecifique et sensible de
M bovis dans des matrices complexes
Lrsquoextraction drsquoADN de matrices environ-
nementales srsquoaccompagne drsquoune co-extraction
drsquoinhibiteurs potentiels de la PCR (acides
humiques polypheacutenols polysaccharides et
meacutetaux) Lrsquoacide humique par exemple inhibe
lrsquoactiviteacute de la Taq polymeacuterase [Kermekchiev et
al 2009] et se lie sur des sites speacutecifiques de
lrsquoADN empecircchant son amplification [Opel et al
2010] La dilution des eacutechantillons drsquoADN
utiliseacutes en PCR en temps reacuteel permet la dilution
des inhibiteurs et limite leur action Toutefois
lorsque la cible rechercheacutee est preacutesente en
petit nombre de copies la dilution de lrsquoADN
rend la deacutetection moins efficace Crsquoest
pourquoi nous analysons lrsquoADN extrait pur et
dilueacute au 15 au 110 et au 120 en PCR en
temps reacuteel de faccedilon agrave rechercher la quantiteacute
drsquoinhibiteurs minimale permettant drsquoamplifier
la cible
Les limites de deacutetection de la PCR se situent au
mieux autour de 1000 copies de gegravenes par g de
sol Les seuils de deacutetection des trois systegravemes
que nous avons creacuteeacutes se situent au-dessus de
1000 copies du gegravene par g de sol ce qui
correspond agrave des valeurs en adeacutequation avec
drsquoautres reacutesultats obtenus sur la deacutetection de
bacteacuteries dans les sols [Ibekwe et Grieve
2003]
En 2013 seulement deux des eacutechantillons
analyseacutes ont donneacute un signal faible avec un des
systegravemes de deacutetection moleacuteculaire Il srsquoagissait
dans les deux cas drsquoeacutechantillons de sol un sol
de terrier probablement habiteacute par un blaireau
infecteacute et un sol de latrine Nous nrsquoavons pu
reacutepeacuteter les reacutesultats probablement agrave cause de
la faible charge en M bovis des eacutechantillons
analyseacutes en dessous de la limite de deacutetection
de nos outils Il reste difficile drsquoisoler M bovis
de lrsquoenvironnement malgreacute la sensibiliteacute et la
speacutecificiteacute des techniques utiliseacutees Plusieurs
eacutetudes malgreacute lrsquoabondance de preacutelegravevements
reacutealiseacutes ont eacuteteacute infructueuses [Pillai et al
2000 Michel et al 2007 Fine et al 2011]
Cette difficulteacute reacutesulte des limites techniques
preacuteceacutedemment citeacutees mais eacutegalement du
caractegravere variable et intermittent de lrsquoexcreacutetion
de M bovis par les animaux infecteacutes
Lrsquoexcreacutetion de la bacteacuterie par la voie
respiratoire les urines les matiegraveres feacutecales ou
les plaies de morsure nrsquoest pas systeacutematique
chez les blaireaux infecteacutes Le niveau
drsquoexcreacutetion deacutepend de la localisation et de
lrsquointensiteacute des leacutesions Il semble que lrsquoexcreacutetion
la plus importante soit trouveacutee chez des
animaux agrave un stade avanceacute de la maladie
[Gallagher et al 1998 Corner et al 2012] En
Eacutepideacutemiol et santeacute anim 2014 65 Deacutetection de Mycobacterium bovis dans lrsquoenvironnement Premiers reacutesultats des recherches meneacutees en Cocircte-drsquoOr
231
Cocircte-drsquoOr une eacutetude meneacutee entre 2009 et
2011 a montreacute que seuls 17 des blaireaux
pieacutegeacutes et analyseacutes (19 sur 1 146) preacutesentaient
des leacutesions visibles alors que la bacteacuterie eacutetait
retrouveacutee sur 56 des blaireaux (49 sur 878)
dans la zone de tuberculose bovine [Payne et
al 2012] Lrsquoabsence de leacutesion visible sur la
majoriteacute des blaireaux analyseacutes pourrait
expliquer une absence drsquoexcreacutetion ou une
faible excreacutetion de M bovis dans
lrsquoenvironnement
Une autre difficulteacute reacuteside dans la reacutealisation
spatiale et temporelle de lrsquoeacutechantillonnage Les
bacilles sont excreacuteteacutes par les animaux sur de
petites zones bien localiseacutees Hormis les
latrines de blaireaux bien visibles les
preacutelegravevements agrave lrsquoeacutechelle drsquoune parcelle drsquoun
terrier drsquoune source sont reacutealiseacutes de maniegravere
ponctuelle et au hasard ce qui diminue la
probabiliteacute de deacutetection de cette bacteacuterie De
plus nous avons toujours un temps de
deacutecalage entre le deacutepistage des bovins ou des
animaux sauvages infecteacutes et la date des
preacutelegravevements car la certitude du diagnostic
animal neacutecessite plusieurs semaines
V - Conclusion et perspectives
Nous avons deacuteveloppeacute des outils de PCR en
temps reacuteel agrave la fois speacutecifiques et sensibles
pour la deacutetection de M bovis dans les matrices
environnementales Malgreacute le nombre
important drsquoeacutechantillons analyseacutes seuls deux
eacutechantillons peu chargeacutes en M bovis ont pu
ecirctre mis en eacutevidence Ces outils de PCR en
temps reacuteel vont encore ecirctre optimiseacutes en 2014
et permettre lrsquoanalyse des nombreux
preacutelegravevements preacutevus dans des zones plus
cibleacutees de Cocircte-drsquoOr dans lesquelles bovins et
faune sauvage infecteacutes ont eacuteteacute retrouveacutes
Plusieurs meacutethodes de culture sont en cours
drsquoessai et seront utiliseacutees sur les eacutechantillons
preacuteleveacutes afin de montrer la viabiliteacute des
bacteacuteries dans lrsquoenvironnement
Eacutepideacutemiol et santeacute anim 2014 65 Deacutetection de Mycobacterium bovis dans lrsquoenvironnement Premiers reacutesultats des recherches meneacutees en Cocircte-drsquoOr
232
Bibliographie
Aranaz A De Juan L Montero N Saacutenchez C Galka M Delso C Alvarez J Romero B Bezos J Vela A I Briones V Mateos A Domiacutenguez L - Bovine tuberculosis (Mycobacterium bovis) in wildlife in Spain Journal of clinical microbiology 2004 42 2602-2608
Betelli L Duquenne P Grenouillet F Simon X Scherer E Geacutehin E Hartmann A - Development and evaluation of a method for the quantification of airborne Thermoactinomyces vulgaris by real-time PCR Journal of microbiological methods 2013 92 25-32
Boumlhm M Hutchings MR White PCL - Contact networks in a wildlife-livestock host community identifying high-risk individuals in the transmission of bovine TB among badgers and cattle PloS One 2009 4(4) e5016
Brodin P Eiglmeier K Marmiesse M Garnier T Niemann S Cole S T Brosch R Billault A - Bacterial Artificial Chromosome-Based Comparative Genomic Analysis Identifies Mycobacterium microti as a Natural ESAT-6 Deletion Mutant Bacterial Artificial Chromosome-Based Comparative Genomic Analysis Identifies Mycobacterium microti as a Natural ESAT-6 Infection and immunity 2002 70 10 5568-5578
Colinon C Deredjian A Hien E Brothier E Bouziri L Cournoyer B Hartman A Henry S Jolivet C Ranjard L Nazaret S - Detection and enumeration of Pseudomonas aeruginosa in soil and manure assessed by an ecfX qPCR assay Journal of applied microbiology 2013 114 1734-1749
Corner L A L Murphy D Gormley E - Mycobacterium bovis infection in the Eurasian badger (Meles meles) the disease pathogenesis epidemiology and control Journal of comparative pathology 2011 144 1-24
Corner L A L OrsquoMeara D Costello E Lesellier S Gormley E - The distribution of Mycobacterium bovis infection in naturally infected badgers Veterinary journal (London England 1997) 2012 194 166-172
Courtenay O Reilly L A Sweeney F P Hibberd V Bryan S Ul-Hassan A Newman C Macdonald D W Delahay R J Wilson G J Wellington E M H - Is Mycobacterium bovis in the environment important for the persistence of bovine tuberculosis Biology letters 2006 2 460-462
Fine AE - The Role of Indirect Transmission in the Epidemiology of Bovine Tuberculosis in Cattle and White-Tailed Deer in Michigan 2006 Dissertation for the degree of Doctor of Philosophy Michigan State University
Fine A E Bolin C A Gardiner J C Kaneene J B - A Study of the Persistence of Mycobacterium bovis in the Environment under Natural Weather Conditions in Michigan USA Veterinary medicine international 2011 2011 765430
Gallagher J Clifton-Hadley R S - Tuberculosis in badgers a review of the disease and its significance for other animals Research in veterinary science 2000 69 203-217
Gallagher J Monies R Gavier-Widen M Rule B - Role of infected non-diseased badgers in the pathogenesis of tuberculosis in the badger Veterinary Record 1998 142 710-714
Gavier-Widen D Chambers M A Palmer N Newell DG Hewinson R G - Pathology of natural Mycobacterium bovis infection in European badgers (Meles meles) and its relationship with bacterial excretion Veterinary Record 2001 148(10) 299-304
Ghodbane R Mba Medie F Lepidi H Nappez C Drancourt M - Long-term survival of tuberculosis complex mycobacteria in soil Microbiology (Reading England) 2014 160(Pt 3) 496-501
Goodchild A V Clifton-Hadley R S - Cattle-to-cattle transmission of Mycobacterium bovis Tuberculosis (Edinburgh Scotland) 2001 81 23-41
Huard R C Claudio L Lazzarini D O Butler W R Soolingen D Van Ho J L - PCR-Based Method To Differentiate the Subspecies of the Mycobacterium tuberculosis Complex on the Basis of Genomic Deletions Journal of clinical microbiology 2003 41 1637-1650
Ibekwe AM Grieve CM - Detection and quantification of Escherichia coli O157H7 in environmental samples by real-time PCR Journal of Applied Microbiology 2003 94(3) 421-431
Kermekchiev M B Kirilova L I Vail E E Barnes W M - Mutants of Taq DNA polymerase resistant to PCR inhibitors allow DNA amplification from whole blood and crude soil samples Nucleic acids research 2009 37 e40
Maddock EC - Studies on the survival time of the bovine tubercle bacillus in soil soil and dung in dung and on grass with experiments on the
Eacutepideacutemiol et santeacute anim 2014 65 Deacutetection de Mycobacterium bovis dans lrsquoenvironnement Premiers reacutesultats des recherches meneacutees en Cocircte-drsquoOr
233
preliminary treatment of infected organic matter and the cultivation of the organism The Journal of Hygiene 1933 33(1) 103-117
Michel AL de Klerk LM Gey van Pittius NC Warren RM van Helden PD - Bovine tuberculosis in African buffaloes observations regarding Mycobacterium bovis shedding into water and exposure to environmental mycobacteria BMC Veterinary Research 2007 3 23
Neill S Hanna J OrsquoBrien J McCracken R - Excretion of Mycobacterium bovis by experimentally infected cattle Veterinary Record 1988 123 340-343
Opel K L Chung D McCord B R - A study of PCR inhibition mechanisms using real time PCR Journal of forensic sciences 2010 55 25-33
Palmer M V Whipple D L Waters W R Mitchell V Diana L Ray W Transmission E D - Experimental Deer-To-Deer Transmission of Mycobacterium bovis Experimental deer-to-deer transmission of Mycobacterium bovis American journal of veterinary research 2001 62 692-696
Payne A Boschiroli M L Gueneau E Moyen J L Rambaud T Dufour B Gilot-Fromont E Hars J - Bovine tuberculosis in ldquoEurasianrdquo badgers (Meles meles) in France European Journal of Wildlife Research 2012 59 331-339
Pillai S D Widmer K W Ivey L J Coker K C Newman E Lingsweiler S Baca D Kelley M Davis D S Silvy N J Adams L G - Failure to identify non-bovine reservoirs of Mycobacterium bovis in a region with a history of infected dairy-cattle herds Preventive veterinary medicine 2000 43 53-62
Ranjard L Lejon DPH Mougel C Schehrer L Merdinoglu D Chaussod R - Sampling strategy in molecular microbial ecology influence of soil sample size on DNA fingerprinting analysis of fungal and bacterial communities Environmental Microbiology 2003 5(11) 1111-1120
Richomme C Boschiroli M L Hars J Casabianca F Ducrot C - Bovine tuberculosis in livestock and wild boar on the Mediterranean island Corsica Journal of wildlife diseases 2010 46 627-631
Sweeney F P Courtenay O Ul-Hassan A Hibberd V Reilly L A Wellington E M H - Immunomagnetic recovery of Mycobacterium bovis from naturally infected environmental
samples Letters in applied microbiology 2006 43 364-369
Sweeney F P Courtenay O Hibberd V Hewinson R G Reilly L A Gaze W H Wellington E M H - Environmental monitoring of Mycobacterium bovis in badger feces and badger sett soil by real-time PCR as confirmed by immunofluorescence immunocapture and cultivation Applied and environmental microbiology 2007 73 7471-7473
Travis ER Gaze WH Pontiroli A Sweeney FP Porter D Mason S Keeling MJC Jones RM Sawyer J Aranaz A Rizaldos EC Cork J Delahay RJ Wilson GJ Hewinson RG Courtenay O Wellington EMH - An inter-laboratory validation of a real time PCR assay to measure host excretion of bacterial pathogens particularly of Mycobacterium bovis PloS One 2011 6(11) e27369
Walter W D Anderson C W Smith R Vanderklok M Averill J J Vercauteren K C - On-farm mitigation of transmission of tuberculosis from white-tailed deer to cattle literature review and recommendations Veterinary medicine international 2012 2012 616318
Young J S Gormley E amp Wellington E M H - Molecular Detection of Mycobacterium bovis and Mycobacterium bovis BCG ( Pasteur ) in Soil Applied and environmental microbiology 2005 71 1946-1952
Zanella G Durand B Hars J Moutou F Garin-Bastuji B Duvauchelle A Fermeacute M Karoui C Boschiroli M L - Mycobacterium bovis in wildlife in France Journal of wildlife diseases 2008 44 99-108
235
Annexe 2 Preacuteparation du Middlebrook 7H9
Middlebrook 7H9 suppleacutementeacute pour la culture liquide de Mycobacterium bovis
Preacuteparation de la base 7H9 (pour 500 mL de milieu)
1 Diluer 235 g de Middlebrook 7H9 (Fluka analytical) dans 450 mL drsquoeau osmoseacutee 2 Autoclaver 15 min agrave 120degC
Preacuteparation de lrsquoenrichissement ADC (pour 50 mL)
1 Peser 25 g de BSA 1 g de Dextrose et 0005 g de catalase bovine 2 Ajouter 50 mL drsquoeau osmoseacutee 3 Agiter environ 30 min (agitation magneacutetique modeacutereacutee pour eacuteviter la formation de mousse) 4 Filtrer lrsquoenrichissement sur une membrane filtrante de porositeacute 022 microM (Conservation 1 mois agrave 4degC)
Preacuteparation du Middlebrook 7H9 enrichi
1 Apregraves lrsquoautoclavage une fois le milieu refroidi ajouter 50 mL drsquoADC (conservation 1 mois agrave 4degC)
237
Annexe 3 Preacuteparation du Middlebrook 7H11
Middlebrook 7H11 suppleacutementeacute pour la culture de Mycobacterium bovis
Preacuteparation des solutions stock (tous les produits ont eacuteteacute fournis par le laboratoire Sigma-Aldrich)
Vert malachite 06 dissoudre 03 g dans 50 mL drsquoeau osmoseacutee
TicarcillineTrimeacutethoprime peser 1 g ticarcilline et 01 g de trimeacutethoprime lactate ajouter 10 mL drsquoeau osmoseacutee steacuterile Vortexer jusqursquoagrave dissolution reacutepartir en Eppendorf de 1 mL congeler (conservation 6 mois agrave -20degC)
Cycloheximide peser 500 mg et dissoudre dans 10 mL drsquoeacutethanol absolu (conservation 1 an agrave 4degC)
Polymyxine B dilution dans eau osmoseacutee steacuterile pour concentration finale de 100 000 UI par mL
Sang mouton lyseacute Meacutelanger 25 mL de sang avec 25 mL drsquoeau osmoseacutee steacuterile bain-marie 10 min agrave 60degC puis utilisation immeacutediate
Preacuteparation de la base 7H11 (pour 1 L de milieu)
1 Diluer 189 g de Middlebrook 7H11 (Fluka analytical) dans 781 mL drsquoeau 2 Ajouter 1 mL de Vert de malachite 06 3 Ajouter un barreau aimanteacute dans la bouteille 4 Autoclaver 15 min agrave 120degC
Preacuteparation de lrsquoenrichissement OADC (pour 100mL)
1 Peser 5 g de BSA 2 g de Dextrose 085 g de NaCl et 0005g de catalase bovine 2 Ajouter 100 mL drsquoeau osmoseacutee 3 Agiter environ 30 min (agitation magneacutetique modeacutereacutee pour eacuteviter la formation de mousse) 4 Ajouter 125 microL drsquoacide oleacuteique 5 Filtrer lrsquoenrichissement sur une membrane filtrante de porositeacute 022 microM (Conservation 1 mois agrave 4degC)
Preacuteparation du Middlebrook 7H11 enrichi
1 Apregraves lrsquoautoclavage porter le milieu agrave 45-50degC 2 Ajouter 100 mL de seacuterum bovin inactiveacute (agitation magneacutetique) 3 Ajouter 100 mL drsquoOADC (agitation magneacutetique) 4 Ajouter 5 mL de sang de mouton lyseacute 5 Ajouter 1 mL de TicarcillineTrimeacutethoprime 2 mL de Polymyxine B 2 mL de cycloheximide
(agitation magneacutetique) (facultatif si ensemencement de cultures pures)(Concentrations finales respectives 100 mgL 10 mgL 200 000 UIL et 100 mgL)
6 Reacutepartition de 25 mL de milieu par boite (Conservation 1 mois agrave 4degC)
Reacutesumeacute La tuberculose bovine est une maladie infectieuse contagieuse causeacutee par Mycobacterium bovis Cette maladie touche les bovins
et de nombreuses espegraveces de mammifegraveres domestiques et sauvages ainsi que lrsquohomme La circulation de la bacteacuterie dans des systegravemes
multi-hocirctes varieacutes favorise lrsquoentretien de la maladie et la contamination des bovins vivant agrave proximiteacute des animaux sauvages infecteacutes En
marge de la transmission directe de M bovis par voie respiratoire la transmission indirecte aux bovins lieacutee agrave lrsquoinhalation ou agrave lrsquoingestion de
matrices environnementales contamineacutees par un animal infecteacute excreacuteteur est suspecteacutee dans plusieurs reacutegions du monde Lrsquoexistence de
reacuteservoirs environnementaux ougrave le bacille M bovis est capable de persister pourrait donc ecirctre un facteur important de la reacuteeacutemergence puis
du maintien de la maladie dans les systegravemes multi-hocirctes
En Cocircte drsquoOr deacutepartement fortement toucheacute par la tuberculose bovine depuis 2004 la transmission indirecte de la bacteacuterie entre la faune
sauvage infecteacutee et les bovins est suspecteacutee dans plusieurs eacutelevages Pour eacutevaluer la preacutesence et la survie de cette bacteacuterie dans
lrsquoenvironnement nous avons analyseacute un grand nombre drsquoeacutechantillons preacuteleveacutes dans des zones partageacutees par les bovins etou la faune sauvage
infecteacutes dans le but de deacuteterminer la distribution environnementale de M bovis Pour ce faire nous avons deacuteveloppeacute ou modifieacute des systegravemes
de deacutetection moleacuteculaire adapteacutes aux matrices environnementales complexes Nous avons eacutegalement eacutevalueacute lrsquoimpact de la tempeacuterature et
des proprieacuteteacutes physico-chimiques de deux sols sur la survie de M bovis ainsi que le rocircle de la meacutesofaune du sol (lombrics en particulier)
dans la disseacutemination de la bacteacuterie agrave partir de matiegravere organique contamineacutee Lrsquoeacutetude environnementale a mis plus particuliegraverement en
eacutevidence la contamination de deux biotopes les zones humides des pacirctures et les sols de terriers de blaireaux De plus les eacutetudes
expeacuterimentales ont montreacute que M bovis pouvait survivre plusieurs mois dans le sol agrave 4degC et que les lombrics pouvaient disseacuteminer la bacteacuterie
dans le sol voire jouer un rocircle potentiel de vecteur pour les animaux qui les consomment Ces reacutesultats apportent de nouvelles connaissances
sur la persistance et la circulation de M bovis dans lrsquoenvironnement en Cocircte drsquoOr et permettront de proposer des ameacuteliorations aux mesures
de bioseacutecuriteacute deacutejagrave existantes dans les eacutelevages bovins
Mots Cleacutes Mycobacterium bovis environnement sol eau fegraveces bovins faune sauvage qPCR culture
Abstract Bovine tuberculosis is an infectious disease caused by Mycobacterium bovis This disease affects multiple species of domestic
livestock notably cattle wild mammals and humans The circulation of the bacteria within complex multi-host systems promotes the
maintenance of the disease and the reinfection of cattle Besides the classical direct transmission of the bacteria through the respiratory
route indirect transmission through inhalation or ingestion of environmental substrates contaminated by the excretions of infected animals
is suspect in several countries Environmental contamination with M bovis appears to be a crucial factor in the persistence of the infection in
multi-host systems
In Cocircte dOr a French department impacted by bovine tuberculosis since 2004 the indirect transmission of the bacteria from infected wildlife
to cattle is now suspect in several cases To assess this mode of transmission we evaluated environmental contamination with M bovis on a
large number of samples collected in areas shared by cattle and infected wildlife For this purpose we both developed and modified molecular
detection systems adapted for complex environmental substrates We also assessed the impact of temperature and soil physicochemical
properties on the survival of M bovis as well as the role of earthworms in the dissemination of the bacteria from contaminated organic
material The environmental study showed contamination of two habitats in particular wetland pastures and soil badger setts Moreover
results indicated that M bovis can survive in soil for several months at 4degC and that worms may disseminate the bacteria in the soil and even
serve as a vector for the mammals that consume them This study increases our understanding of the persistence and the circulation of
M bovis in the environment in Cocircte dOr In addition these results will be used to improve the existing biosecurity measures on cattle farms
in the region
Key words Mycobacterium bovis environment soil water feces cattle wildlife qPCR culture

Merci agrave Franck Biet et Thierry Cochard de lrsquoINRA de Tours qui nous ont fourni les souches de
Map et M smegmatis
Des eacutenormes remerciements pour les lieutenants de louveterie les pieacutegeurs et les chasseurs
qui mrsquoont accompagneacutee dans mes preacutelegravevements sur le terrain et sans qui je nrsquoaurai pas pu preacutelever
tous ces eacutechantillons Jeannick Gautherot Dominique Rigaud Yannick Thibert Georges Balluet Jean-
Luc Moindrot Marcel Guillier qui connaissent les bois comme leur poche et font un travail
consideacuterable
Je remercie les eacuteleveurs qui ont participeacute au travail en nous accompagnant sur leurs pacirctures
Merci agrave Leacuteon Fayolle pour son expertise sur les lombrics et Dominique Meunier pour son
expertise sur les formations peacutedologiques de lrsquoAuxois merci agrave Luc Biju-Duval pour les cartes reacutealiseacutees
Un eacutenorme merci agrave tous les membres du pocircle MERS qui ont fait de ces trois anneacutees un moment
extrecircmement riche en eacutechanges scientifiques humains amicaux et sportifshellipMerci agrave Dominique
Pascal Laurent Jean-Paul Seacutebastien Un merci particulier aux filles de lrsquoeacutequipe Geacuteraldine Ceacutecile
Laetitia pour leur gentillesse leurs compeacutetences et tous les bons moments passeacutes agrave discuterhellipet agrave
marcher Des remerciements tout particuliers agrave Murielle Rochelet pour ses conseils hyper-aviseacutes sa
rigueur son esprit drsquoinnovation son anglais parfait et encore plein plein drsquoautres choses Merci
infiniment pour tout le temps que tu as passeacute agrave me relire et me corrigerhellip
Un grand mais alors tregraves grand merci agrave Benoicirct sans qui lrsquoexpeacuterience lombric nrsquoaurait pas eacuteteacute
aussihellipexceptionnelle Bon courage pour ta thegravese Merci agrave Cleacutement pour les kilos de sols de terriers
ramasseacutes peseacutes analyseacutes lors de ces deux stages Un grand merci agrave Pierre pour sa gentillesse rare son
travail tregraves seacuterieux et son aide inestimable pour la mise en page la relecture et lrsquoimpression de ce
manuscrit
Merci eacutegalement agrave mes collegravegues du Master AE Caroline Alexandre Fam-Ky Justin Marion
et tous les autres qui mrsquoont parfois (souvent) aideacutee agrave supporter de longues heures de cours
Merci agrave tous ceux que jrsquoai croiseacute agrave lrsquoINRA et ailleurs qui ont fait de ces trois ans une expeacuterience
humaine et scientifique inoubliable Ariane Emilie Kadiya Emeline Ceacuteline Heacutelegravene Charline Anne-
Laure Aude Gilles David Marion Nicolas Romain Fabrice Chloeacute Aymeacute Laure Luiz Marie-
Christine Nadine Estelle Amy Amandinehellip et tous ceux que jrsquoai sucircrement oublieacute
Enfin pour finir un grand merci agrave Steacutephane Lola Lucien et Antonin qui ont supporteacute mes hauts
et mes bas pendant ces trois anneacutees agrave ma famille et ma belle-famille qui ont toujours eacuteteacute lagravehellip
Reacutesumeacute
La tuberculose bovine est une maladie infectieuse contagieuse causeacutee par Mycobacterium
bovis Cette maladie touche les bovins et de nombreuses espegraveces de mammifegraveres domestiques et
sauvages ainsi que lrsquohomme La circulation de la bacteacuterie dans des systegravemes multi-hocirctes varieacutes favorise
lrsquoentretien de la maladie et la contamination des bovins vivant agrave proximiteacute des animaux sauvages
infecteacutes En marge de la transmission directe de M bovis par voie respiratoire la transmission indirecte
aux bovins lieacutee agrave lrsquoinhalation ou agrave lrsquoingestion de matrices environnementales contamineacutees par un
animal infecteacute excreacuteteur est suspecteacutee dans plusieurs reacutegions du monde Lrsquoexistence de reacuteservoirs
environnementaux ougrave le bacille M bovis est capable de persister pourrait donc ecirctre un facteur
important de la reacuteeacutemergence puis du maintien de la maladie dans les systegravemes multi-hocirctes
En Cocircte drsquoOr deacutepartement fortement toucheacute par la tuberculose bovine depuis 2004 la
transmission indirecte de la bacteacuterie entre la faune sauvage infecteacutee et les bovins est suspecteacutee dans
plusieurs eacutelevages Pour eacutevaluer la preacutesence et la survie de cette bacteacuterie dans lrsquoenvironnement nous
avons analyseacute un grand nombre drsquoeacutechantillons preacuteleveacutes dans des zones partageacutees par les bovins etou
la faune sauvage infecteacutes dans le but de deacuteterminer la distribution environnementale de M bovis Pour
ce faire nous avons deacuteveloppeacute ou modifieacute des systegravemes de deacutetection moleacuteculaire adapteacutes aux matrices
environnementales complexes Nous avons eacutegalement eacutevalueacute lrsquoimpact de la tempeacuterature et des
proprieacuteteacutes physico-chimiques de deux sols sur la survie de M bovis ainsi que le rocircle de la meacutesofaune
du sol (lombrics en particulier) dans la disseacutemination de la bacteacuterie agrave partir de matiegravere organique
contamineacutee Lrsquoeacutetude environnementale a mis plus particuliegraverement en eacutevidence la contamination de
deux biotopes les zones humides des pacirctures et les sols de terriers de blaireaux De plus les eacutetudes
expeacuterimentales ont montreacute que M bovis pouvait survivre plusieurs mois dans le sol agrave 4degC et que les
lombrics pouvaient disseacuteminer la bacteacuterie dans le sol voire jouer un rocircle potentiel de vecteur pour les
animaux qui les consomment Ces reacutesultats apportent de nouvelles connaissances sur la persistance et
la circulation de M bovis dans lrsquoenvironnement en Cocircte drsquoOr et permettront de proposer des
ameacuteliorations aux mesures de bioseacutecuriteacute deacutejagrave existantes dans les eacutelevages bovins
Mots cleacutes Mycobacterium bovis environnement sol eau fegraveces bovins faune sauvage qPCR culture
Abstract
Bovine tuberculosis is an infectious disease caused by Mycobacterium bovis This disease
affects cattle and many species of domestic and wild mammals and humans The circulation of the
bacteria in various multi-host systems promotes the maintenance of the disease and the contamination
of cattle in the vicinity Beside direct transmission of the bacteria through the respiratory route indirect
transmission through inhalation or ingestion of environmental matrices contaminated by an infected
animal excretory is suspected in several countries Environmental contamination with M bovis appears
to be a crucial factor in the persistence of the infection in multi-host systems
In Cocircte dOr a French department affected by bovine tuberculosis since 2004 the indirect
transmission of the bacteria from infected wildlife to cattle is suspected in several cases To assess this
type of transmission of the bacillus we evaluated the environmental contamination with M bovis on
a large number of samples taken in areas shared by cattle and or wildlife infected For this purpose
we developed or modified molecular detection systems adapted for environmental complex matrices
We also assessed the impact of physicochemical properties of both soil and temperature on survival of
M bovis and the role of earthworms in the spread of the bacteria from contaminated organic material
The environmental study showed the contamination of two media in particular wetlands pastures and
soil badger setts Moreover experimental studies have shown that M bovis can survive in soil for
several months at 4 deg C and the worms could spread the bacteria in the soil or even play a potential
role for vector animals that consume them These results will propose improvements to existing
biosecurity measures on cattle farms and provide new knowledge about the persistence and circulation
of M bovis in the environment in Cocircte dOr
Key words Mycobacterium bovis environment soil water feces cattle wildlife qPCR culture
Table des matiegraveres
INTRODUCTION GENERALE 1
CHAPITRE 1 - CONTEXTE BIBLIOGRAPHIQUE 4
A LES MYCOBACTERIES DIVERSITE PHYLOGENIE SPECTRE DrsquoHOcircTES ET VIRULENCE 4
1 Le genre Mycobacterium taxonomie et meacutethodes de classification 4
2 Caracteacuteristiques geacuteneacuterales des mycobacteacuteries 5
a Morphologie des mycobacteacuteries 5
b Caracteacuteristiques culturales pheacutenotypiques et physiologiques 7
c Phylogeacutenie du genre Mycobacterium 8
d Caracteacuteristiques geacuteneacutetiques des mycobacteacuteries 9
3 Description des diffeacuterents Complexes ou espegraveces de mycobacteacuteries 10
a Les mycobacteacuteries non pathogegravenes potentiellement opportunistes 11
b Les mycobacteacuteries saprophytes non pathogegravenes 12
c Les mycobacteacuteries pathogegravenes obligatoires 12
Les mycobacteacuteries responsables de legravepre 12
Les mycobacteacuteries tuberculeuses le Complexe tuberculosis (MTBC) 13
B LA TUBERCULOSE UNE MALADIE TRES ANCIENNE MAIS UNE MENACE TOUJOURS ACTUELLE 20
1 Introduction 20
2 Histoire de la tuberculose 20
3 La tuberculose bovine 21
a Introduction 21
b Les hocirctes et les communauteacutes drsquohocirctes de Mycobacterium bovis dans le monde et leurs statuts
eacutepideacutemiologiques 22
c Aspect zoonotique de Mycobacterium bovis 23
d Pathogeacutenie de la tuberculose bovine 24
Facteurs influenccedilant le pouvoir pathogegravene de M bovis voies drsquoinfection et dose infectante 24
Facteurs influenccedilant la reacuteceptiviteacute et la sensibiliteacute de lrsquohocircte 25
Les eacutetapes de lrsquoinfection et la reacuteponse immunitaire associeacutee 26
Expression clinique de la maladie et leacutesions observeacutees 27
e Le deacutepistage et le diagnostic de lrsquoinfection chez les animaux 28
f Epideacutemiologie de la tuberculose bovine 30
Evolution de la tuberculose bovine dans le monde et en Europe et mesures de lutte 30
Evolution de la situation sanitaire en France 33
Evolution de la situation sanitaire en Cocircte drsquoOr 38
C ECOLOGIE DE M BOVIS DANS LrsquoENVIRONNEMENT 42
1 Introduction 42
2 Origines de la contamination environnementale par M bovis 42
a Lrsquoexcreacutetion animale de Mycobacterium bovis 42
b Le transfert de Mycobacterium bovis par les effluents drsquoeacutelevage ou urbains 44
3 Deacutetection et survie de Mycobacterium bovis dans lrsquoenvironnement 45
a Deacutetection de Mycobacterium bovis en conditions naturelles 45
b Dureacutees de survie de Mycobacterium bovis en conditions expeacuterimentales 47
4 Strateacutegies de survie des mycobacteacuteries dans lrsquoenvironnement 54
a Lrsquoeacutetat de dormance 54
b La sporulation 55
c Interactions des mycobacteacuteries avec la faune du sol 56
d Interactions des mycobacteacuteries avec les diptegraveres 57
e La formation de biofilms 57
5 Facteurs de risque de contamination de lrsquoenvironnement et de transmission de Mycobacterium
bovis aux animaux par lrsquoenvironnement contamineacute 58
a Risques lieacutes agrave lrsquoeacutecologie des espegraveces sauvages sensibles agrave Mycobacterium bovis 58
b Risques lieacutes aux pratiques drsquoeacutelevage 60
c Risques lieacutes aux pratiques cyneacutegeacutetiques 61
d Risques lieacutes aux facteurs paysagers climatiques et peacutedologiques 62
D PRINCIPAUX OUTILS DE DETECTION DE PHENOTYPAGE ET DE GENOTYPAGE APPLICABLES AU MTBC ET
PLUS PARTICULIEREMENT A MYCOBACTERIUM BOVIS 64
1 La deacutetection des mycobacteacuteries par microscopie 64
2 Les meacutethodes de culture des mycobacteacuteries 65
a Les diffeacuterents traitements preacutealables agrave la mise en culture des eacutechantillons 65
b La culture sur milieu solide 66
c La culture en milieu liquide 67
3 Meacutethodes de deacutetection et drsquoidentification des bacteacuteries du MTBC 68
a Caracteacuterisation pheacutenotypique et biochimique des mycobacteacuteries du MTBC 69
b Lrsquoidentification des mycobacteacuteries par recherche de biomarqueurs 69
c Lrsquoidentification des mycobacteacuteries par des meacutethodes drsquohybridation coupleacutees ou non agrave la PCR 70
d Le seacutequenccedilage 71
e Deacutetection et geacutenotypage des mycobacteacuteries du MTBC les marqueurs utiliseacutes 72
Les seacutequences drsquoinsertion 72
Les courtes seacutequences drsquoADN reacutepeacuteteacutees 73
Reacutegions de Diffeacuterence (RD) et Single Nucleotide Polymorphism (SNP) 78
f Identification par seacutequenccedilage complet des geacutenomes 79
g Meacutethode speacutecifique drsquoidentification des mycobacteacuteries animales utiliseacutee par le Laboratoire National de
reacutefeacuterence de la tuberculose bovine LNR TB 79
OBJECTIFS DE LA THESE 82
CHAPITRE 2 - DEVELOPPEMENT ET OPTIMISATION DrsquoOUTILS POUR LA DETECTION DE MYCOBACTERIUM
BOVIS DANS LES ECHANTILLONS ENVIRONNEMENTAUX 86
A DEVELOPPEMENT DrsquoOUTILS MOLECULAIRES ADAPTES AUX MATRICES ENVIRONNEMENTALES 86
CHAPITRE 3 - ETUDE DE LA PREVALENCE ENVIRONNEMENTALE DE MYCOBACTERIUM BOVIS DANS LA ZONE
ENDEMIQUE DE TUBERCULOSE BOVINE DE COTE DrsquoOR ET RESULTATS OBTENUS EN CHARENTE 93
A AVANT-PROPOS 93
B FIRST MOLECULAR DETECTION OF MYCOBACTERIUM BOVIS IN ENVIRONMENTAL SAMPLES FROM A
FRENCH REGION WITH ENDEMIC BOVINE TUBERCULOSIS 95
C CAMPAGNES DE PRELEVEMENTS SUPPLEMENTAIRES 121
1 Fin des preacutelegravevements en Cocircte drsquoOr 121
2 Preacutelegravevements en Charente 123
3 Compleacutements paysage geacuteomorphologie et peacutedologie 125
D BILAN GLOBAL 130
CHAPITRE 4 - IMPACT DE LA TEMPERATURE ET DE LA NATURE DU SOL SUR LA SURVIE DE MYCOBACTERIUM
BOVIS SB0120 133
A AVANT-PROPOS 133
B EXPERIMENTAL INVESTIGATION OF THE IMPACT OF THE TEMPERATURE AND THE SOIL TYPE ON
MYCOBACTERIUM BOVIS SURVIVAL IN THE ENVIRONMENT 135
C BILAN 150
CHAPITRE 5 - ROLE DE LA FAUNE DU SOL DANS LA CIRCULATION DE MYCOBACTERIUM BOVIS ETUDE DE SA
DISSEMINATION PAR LES LOMBRICS 155
A AVANT-PROPOS 155
B RAPID DISSEMINATION OF M BOVIS FROM CATTLE DUNG TO SOIL BY THE EARTHWORM LUMBRICUS
TERRESTRIS 157
C COMPLEMENT PRELEVEMENT ET ANALYSE DE LOMBRICS DANS UNE PATURE SUSPECTE 174
D BILAN 175
DISCUSSION GENERALE 177
1 Des meacutethodes deacuteveloppeacutees (ou adapteacutees) puis valideacutees dans des matrices environnementales 179
2 Contamination environnementale les zones humides les eaux de ruissellement et les sols de
terriers sont potentiellement agrave risque 180
3 Survie et circulation de M bovis dans lrsquoenvironnement 183
PERSPECTIVES 185
REFERENCES BIBLIOGRAPHIQUES 191
ANNEXES 221
Liste des Figures
Figure 1 Structure et composition de lenveloppe des mycobacteacuteries (drsquoapregraves Marrakchi Laneacuteelle and Daffeacute 2014) AG arabinogalactane PG peptidoglycane TDM treacutehalose dimycolate PL phospholipides TMM treacutehalose monomycolate GL glycolipides 7
Figure 2 Arbre phylogeacutenique baseacute sur lrsquoanalyse de lrsquoARNr 16S des principaux complexes et espegraveces de mycobacteacuteries (drsquoapregraves Magee et Ward 2012) 9
Figure 3 Phylogeacutenie des souches du MTBC baseacutee sur les Reacutegions de Diffeacuterences et diffeacuterents Single Nucleotide Polymorphism (Dippenaar et al 2015) 14
Figure 4 Aspect des colonies de M bovis sur milieu de Lowenstein-Jensen (agrave gauche) et Middlebrook 7H11 suppleacutementeacute (agrave droite) 17
Figure 5 Evolution de la reacuteponse immunitaire chez le bovin infecteacute par M bovis au cours du temps et aux diffeacuterents tests de deacutepistage de la tuberculose bovine (de la Rua-Domenech et al 2006 adapteacute de Vordermeier et al 2004) 27
Figure 6 Carte de distribution de la tuberculose bovine chez les bovins entre juillet et deacutecembre 2014 (Source WAHID Interface) 31
Figure 7 Statuts des pays europeacuteens vis-agrave-vis de la tuberculose bovine en 2012 et proportion de bovins infecteacutes OTF Officially Tuberculosis Free (officiellement indemne de tuberculose bovine) MS Member State (Etat membre) (Source EFSA 2014) 32
Figure 8 Evolution du taux de preacutevalence annuelle national de la tuberculose bovine chez les bovins (Beacutenet et al 2006) 34
Figure 9 Evolution de la preacutevalence et de lrsquoincidence de la tuberculose bovine en France entre 1995 et 2003 (Cavalerie et al 2014) 35
Figure 10 Incidence et distribution de la tuberculose bovine en France dans les anneacutees 50 (en haut) et entre 2000 et 2013 (en bas) (Cavalerie et al 2014) 36
Figure 11 Reacutepartition et reacutesultats des analyses effectueacutees dans le cadre du dispositif Sylvatub chez le grand gibier drsquoaoucirct 2013 agrave aoucirct 2014 et chez les blaireaux en 2014 (Source Sylvatub) 38
Figure 12 Localisation du parcellaire des cheptels infecteacutes par la tuberculose bovine entre 2010 et 2015 et spoligotypes incrimineacutes (Source CIREV) 40
Figure 13 Nombre de nouveaux foyers bovins et de foyers recontamineacutes entre 2002 et 2014 (Donneacutees DDPP21) 40
Figure 14 Nombre de cervideacutes sangliers et blaireaux infecteacutes deacutecouverts en Cocircte drsquoOr entre 2002 et 2014 41
Figure 15 Repreacutesentation scheacutematique de la circulation de M bovis dans lrsquoenvironnement agricole (modifieacute drsquoapregraves Elliott et al 2014) 63
Figure 16 Profils de restriction disolats de M tuberculosis obtenus suite agrave une RFLP-IS6110 Les isolats 3 5 6 9 et 10 ont le mecircme profil et posseacutedaient un lien eacutepideacutemiologique S repreacutesente le marqueur de poids moleacuteculaire (Source web site CDC) 73
Figure 17 Principe du spoligotypage des bacteacuteries du MTBC dapregraves Kamerbeek et al (1997) A Repreacutesentation du locus DR de M tuberculosis H37Rv et M bovis BCG B Amplifications des espaceurs par un couple damorces (a et b) et amplicons obtenus C Profils drsquohybridation sur membrane (un carreacute noir signale la preacutesence de lrsquoespaceur les parties blanches leur absence) 75
Figure 18 Diffeacuterences geacutenomiques de la reacutegion RD1 entre M bovis BCG (A) M microti (C) par comparaison agrave M tuberculosis H37Rv (B) drsquoapregraves Brodin et al 2002 et localisation du locus Rv3866 RD1mic correspond agrave la seacutequence de M tuberculosis deacuteleacuteteacutee chez M microti RD1bcg correspond agrave la seacutequence de M tuberculosis deacuteleacuteteacutee chez M bovis BCG 88
Figure 27 Localisation des communes de Cocircte drsquoOr sur lesquelles de foyers bovins et des animaux sauvages infecteacutes ont eacuteteacute deacutetecteacutes et localisation des preacutelegravevements environnementaux effectueacutes en 2015 (les preacutelegravevements positifs sont repreacutesenteacutes par des eacutetoiles noires) (Reacutealisation Marina Beacuteral CIREV Bourgogne) 123
Figure 28 Localisation des communes de Charente sur lesquelles de foyers bovins et des animaux sauvages infecteacutes ont eacuteteacute deacutetecteacutes et localisation des preacutelegravevements environnementaux effectueacutes en 2015 (les preacutelegravevements positifs sont repreacutesenteacutes par des eacutetoiles noires) (Reacutealisation Marina Beacuteral CIREV Bourgogne) 125
Figure 29 Orographie et hydrographie de lrsquoAuxois Les valleacutees supportant les parcelles suspectes sont indiqueacutees par une flegraveche noire (Source BD Carthage et BD alti - IGN) 127
Figure 30 Localisation de lrsquoAuxois en Cocircte drsquoOr structure paysagegravere et geacuteomorphologie de ce territoire 127
Figure 31 Nombre de vaches allaitantes par canton dans le deacutepartement de la Cocircte drsquoOr (agrave gauche) et parcellaire des foyers bovins de 2010 agrave 2015 (agrave droite) (Source Recensement agricole 2010 ndash Agreste) 128
Figure 32 Reacutepartition spatiale des parcelles des eacutelevages bovins infecteacutes de 2010 agrave 2014 en Cocircte drsquoOr et repreacutesentation des couches peacutedologiques (Reacutealisation Marina Beacuteral CIREV Bourgogne) 129
Figure 33 Tempeacuteratures minimales et maximales observeacutees agrave Dijon-Longvic de 2012 agrave 2015 (Source Meacuteteacuteo France) 151
Liste des Tableaux
Tableau 1 Dureacutees de survie de M bovis dans diffeacuterentes matrices environnementales obtenues en conditions expeacuterimentales naturelles ou de laboratoire 49
Tableau 2 Identification des principales mycobacteacuteries du MTBC par des observations pheacutenotypiques et des tests biochimiques 69
Tableau 3 Cibles moleacuteculaires deacutecrites dans la litteacuterature pour la deacutetection du Complexe tuberculosis ou de M bovis dans les eacutechantillons environnementaux 76
Tableau 4 Spoligotypes et geacutenotypes MIRU-VNTR des souches de M bovis isoleacutees chez les bovins et la faune sauvage en Cocircte dOr et en Dordogne (drsquoapregraves Hauer et al 2015) 77
Tableau 5 Systegravemes de deacutetection utiliseacutes en qPCR en premiegravere intention pour lidentification dune mycobacteacuterie dorigine animale et reacutesultats obtenus 80
Liste des Annexes
Annexe 1 Deacutetection de Mycobacterium bovis dans lrsquoenvironnement Premiers reacutesultats des recherches meneacutees en Cocircte-drsquoOr 223
Annexe 2 Preacuteparation du Middlebrook 7H9 235
Annexe 3 Preacuteparation du Middlebrook 7H11 237
Liste des Abreacuteviations
ADC Albumine Dextrose Catalase
ADNDNA Acide deacutesoxyribonucleacuteique
AEEMA Association pour lEtude de lEpideacutemiologie des Maladies Animales
AFNOR Agence Franccedilaise de NORmalisation
ANSES Agence nationale de seacutecuriteacute sanitaire de lrsquoalimentation de lrsquoenvironnement et du travail
ARNRNA Acide ribonucleacuteique
ARNr ARN ribosomique
BAAR Bacille acido-alcoolo reacutesistant
BCG Bacille de Calmette et Gueacuterin
BLAST Basic Local Alignment Search Tool
bTB bovine Tuberculosis
CDC Centers of Disease Control and Prevention
CFP-10 Culture Filtrat Protein 10 kD
CIREV Cellules Inter Reacutegionales dEpideacutemiologie Veacuteteacuterinaire
CRISPR Clustered Regularly Interspersed Short Palindromic Repeats
Ct Cycle Threshold
DDPP Direction Deacutepartementale de la Protection des Populations
DEFRA Department for Environment Food and Rural Affairs
DGAL Direction Geacuteneacuterale de lALimentation
DosR Dormancy Survival Regulator
DR Direct Repeat
EFSA European Food Safety Authority
ELISA Enzyme-Linked ImmunoSorbent Assay
ESAT-6 Early Secreted Antigenic Target 6kD
ETR Exact Tandem Repeat
Fbp Fibronectin binding protein
FISH Fluorescence In Situ Hybridization
GC-MS Gas Chromatography-Mass Spectrometry
GDS Groupements de Deacutefense Sanitaire
HAP Hydrocarbures Aromatiques Polycycliques
HIV Human Immunodeficiency Virus
HPLC High Performance Liquid Chromatography
IDC Intradermotuberculination comparative
IDR Intradermotuberculination
IDS Intradermotuberculination simple
IFN-γ Interfeacuteron γ
INRA Institut National de la Recherche Agronomique
Ion PGM Ion Personal Genome Machine
IS Insertion Sequence
ITS Internal Transcribed Spacer
kDkDa kilo Daltons
LNR Laboratoire National de Reacutefeacuterence
LSP Large Sequence Polymorphism
Maa Mycobacterium avium subspecies avium
MAC Mycobacterium avium complex
MALDI-TOF Matrix-Assisted Laser Desorption Ionization-Time Of Flight
Map Mycobacterium avium subspecies paratuberculosis
MDR Multi Drug Resistant
MIRU Mycobacterial Interspersed Repetitive Unit
MLVA Multi Locus VNTR Analysis
MNT Mycobacteacuteries Non Tuberculeuses
MS Member State
MTBC Mycobacterium tuberculosis complex
NCBI National Center for Biotechnology Information
NF Norme Franccedilaise
NGS Next Generation Sequencing
OADC Oleic acid Albumine Dextrose Catalase
OIE Office International des Epizooties
OMS Organisation Mondiale de la Santeacute
ONCFS Office National de la Chasse et de la Faune Sauvage
ONEMA Office National de lEau et des Milieux Aquatiques
OTF Officialy Tuberculosis Free
pb paire de bases
PCR Polymerase chain reaction
PNA Peptide Nucleic Acid
qPCR quantitative Polymerase Chain Reaction
RD Reacutegions de Diffeacuterence
RFLP Restriction Fragment Lenght Polymorphism
RNS Reactive Nitrogen Species
ROS Reactive Oxygen Species
RT-PCR Reverse-Transcription Polymerase Chain Reaction
SNP Single Nucleotide Polymorphism
ssp subspecies
TDM Treacutehalose dimycolate
TMM Treacutehalose monomycolate
UFCCFU Uniteacute Formant Colonie
VBNC Viable But Non Cultivable
VNTR Variable Number Tandem Repeat
WAHID World Animal Health Information Database
WGS Whole Genome Sequencing
WHO World Health Organization
XDR eXtensively Drug Resistant
1
Introduction geacuteneacuterale
La tuberculose bovine est une maladie drsquoimportance sanitaire majeure en santeacute animale agrave
travers le monde Son agent principal Mycobacterium bovis est un membre du Complexe
Mycobacterium tuberculosis qui regroupe les bacteacuteries tuberculeuses infectant lrsquohomme et les
mammifegraveres Les bovins constituent le reacuteservoir originel de M bovis mais de nombreuses espegraveces de
mammifegraveres peuvent eacutegalement ecirctre infecteacutees dont lrsquohomme Les enjeux sanitaires eacuteconomiques et
sociaux induits par cette maladie ont conduit agrave la mise en place de programmes visant lrsquoeacuteradication de
cette maladie chez les bovins agrave travers le monde Dans les pays deacuteveloppeacutes ces mesures associeacutees agrave
la pasteurisation du lait ont fortement reacuteduit lrsquoincidence de la tuberculose bovine voire eacuteradiqueacute
lrsquoinfection chez les bovins Toutefois dans quelques pays lrsquoinfection persiste dans les cheptels bovins
malgreacute des actions de preacutevention de controcircle et de surveillance Dans les pays en deacuteveloppement ougrave
les controcircles organiseacutes sont quasi- voire totalement absents la preacutevalence et la propagation de la
tuberculose bovine sont inquieacutetantes et repreacutesentent un risque sanitaire et eacuteconomique vis-agrave-vis des
populations sensibles En conseacutequence la tuberculose bovine attire chaque anneacutee un peu plus
lrsquoattention de la communauteacute internationale en raison du nombre croissant de troupeaux infecteacutes et
des effets sur les productions animales ainsi que du risque que la maladie fait planer sur la santeacute
publique
Le passage de lrsquoinfection du reacuteservoir bovin agrave diffeacuterentes populations sauvages qui
entretiennent la maladie et peuvent la transmettre en retour aux bovins a fortement compliqueacute les
scheacutemas drsquoeacuteradication de la maladie dans plusieurs reacutegions du monde et explique lrsquoeacutechec partiel des
programmes de lutte mis en place Alors que la transmission directe du bacille par voie respiratoire
semble ecirctre le mode de transmission preacutepondeacuterant au sein drsquoune mecircme espegravece la transmission
indirecte de M bovis est fortement suspecteacutee entre espegraveces partageant les mecircmes territoires et
ressources compliquant encore lrsquoeacutepideacutemiologie de cette maladie
En France une recrudescence du nombre de cheptels bovins infecteacutes dans certains
deacutepartements dont la Cocircte drsquoOr menace son statut de pays laquo officiellement indemne de tuberculose
bovine raquo obtenu en 2001 Dans ce deacutepartement ougrave plus de 190 foyers bovins ont eacuteteacute deacutetecteacutes depuis
2003 M bovis circule dans un systegraveme multi-hocirctes incluant les bovins les cervideacutes les blaireaux et les
sangliers dans une zone geacuteographique localiseacutee Face agrave des contaminations ou des recontaminations
inexpliqueacutees de cheptels avec des souches identiques agrave celles retrouveacutees dans la faune sauvage
infecteacutee deacutetecteacutee agrave proximiteacute dans des eacutelevages le rocircle de lrsquoenvironnement a eacuteteacute suggeacutereacute dans la
transmission indirecte du bacille
Dans ce contexte mes travaux de thegravese visent agrave eacutevaluer la contamination environnementale
2
des zones partageacutees par les bovins et la faune sauvage infecteacutes gracircce agrave des meacutethodes adapteacutees
utilisables sur un grand nombre de preacutelegravevements et agrave rechercher les facteurs favorisant la survie et la
circulation de M bovis dans lrsquoenvironnement Le premier chapitre de ce manuscrit est une synthegravese
des connaissances relatives aux mycobacteacuteries en geacuteneacuteral et aux mycobacteacuteries tuberculeuses en
particulier notamment en terme drsquoeacutecologie de deacutetection et drsquoidentification Le deuxiegraveme chapitre
deacutetaille le deacuteveloppement des meacutethodes moleacuteculaires utiliseacutees pour le diagnostic environnemental
ainsi que le deacuteveloppement de deux nouveaux protocoles pour isoler les mycobacteacuteries du sol et suivre
leur croissance en milieu liquide Le troisiegraveme chapitre preacutesente les modaliteacutes drsquoeacutetude et les reacutesultats
obtenus entre 2013 et 2015 dans lrsquoeacutevaluation de la contamination environnementale en Cocircte drsquoOr Les
reacutesultats drsquoune eacutetude reacutealiseacutee en Charente sont eacutegalement preacutesenteacutes Le quatriegraveme chapitre traite de
lrsquoinfluence de la tempeacuterature et du type de sol sur la dureacutee de survie de M bovis Enfin le cinquiegraveme
chapitre srsquointeacuteresse au rocircle des lombrics sur la disseacutemination de M bovis dans le sol Pour terminer
une discussion geacuteneacuterale syntheacutetise lrsquoensemble des reacutesultats obtenus lors de ce travail et ouvre de
nombreuses perspectives de recherche
Cette thegravese financeacutee par la Reacutegion Bourgogne entre dans le dispositif Jeunes Chercheurs
Entrepreneurs (JCE) Le travail scientifique de recherche srsquoaccompagne de la reacutealisation drsquoun Master
des Entreprises (Master 2) dans lrsquoobjectif drsquoouvrir les doctorants au monde de lrsquoentreprise et au
deacuteveloppement de projets innovants Ce Master a eacuteteacute lrsquooccasion pour moi drsquoacqueacuterir des compeacutetences
compleacutementaires dans le domaine de lrsquoadministration et de la gestion des entreprises notamment
dans la gestion de projets Mon investissement personnel important en parallegravele de mon travail de
recherche mrsquoa permis drsquoobtenir le Master avec la mention Bien
Dans le cadre de la thegravese JCE lrsquoentreprise LSI (speacutecialiseacutee dans le diagnostic veacuteteacuterinaire) a
accompagneacute le deacutemarrage de la thegravese et mrsquoa permis drsquoeacutechanger avec des industriels autour du
deacuteveloppement de systegravemes de deacutetection baseacutes sur la qPCR (IS1561rsquo et autres systegravemes) Suite au
rachat de lrsquoentreprise par Life Science puis Thermo Fisher le transfert de technologie nrsquoa pas eacuteteacute
poursuivi Cependant ces systegravemes de qPCR sont utiliseacutes en routine par le Laboratoire National de
Reacutefeacuterence pour la tuberculose bovine encadreacute par Maria Laura Boschiroli pour caracteacuteriser les souches
de mycobacteacuteries issues drsquoeacutechantillons animaux Drsquoautre part des travaux et reacutesultats obtenus lors de
ma thegravese vont faire lrsquoobjet drsquoune valorisation sous forme drsquoun brevet en cours de deacutepocirct pour la
deacutetection des mycobacteacuteries et drsquoun savoir-faire concernant lrsquoextraction des mycobacteacuteries du sol par
des meacutethodes chimiques en vue de leur mise en culture Jrsquoai donc au cours de mes travaux de recherche
toujours conserveacute lrsquoesprit drsquoinnovation et de transfert potentiel de technologie
CHAPITRE 1
Contexte bibliographique
Chapitre 1
4
CHAPITRE 1 - Contexte bibliographique
A LES MYCOBACTERIES DIVERSITE PHYLOGENIE SPECTRE DrsquoHOcircTES ET VIRULENCE
1 Le genre Mycobacterium taxonomie et meacutethodes de classification
Les connaissances sur les mycobacteacuteries commencent en 1882 date agrave laquelle Robert Koch
deacutecouvre le bacille responsable de la tuberculose humaine alors baptiseacute Bacterium tuberculosis et
partage son postulat sur lrsquoeacutetiologie infectieuse de la maladie La coloration de Ziehl et Neelsen
deacuteveloppeacutee en 1883 pour la recherche des bacilles acido-alcoolo-reacutesistants permet agrave Smith en 1892 de
mettre en eacutevidence un bacille tregraves proche de Bacterium tuberculosis dans des leacutesions tuberculeuses de
bovins Mycobacterium bovis avec toutefois des diffeacuterences pheacutenotypiques et pathogeacuteniques qui
permettent de distinguer les deux bacilles Le bacille de la tuberculose aviaire M avium ssp avium est
isoleacute en 1894 sur des oiseaux puis M avium ssp paratuberculosis en 1895 sur des leacutesions digestives
de bovins La deacutecouverte des premiegraveres mycobacteacuteries non tuberculeuses a suivi celle de Bacterium
tuberculosis notamment gracircce agrave la coloration Mycobacterium smegmatis a eacuteteacute isoleacutee en 1889 dans
des preacutelegravevements biologiques de patients atteints de syphilis puis dans du beurre par lrsquoeacutequipe de Koch
dans les anneacutees 1896 Les premiegraveres recherches meneacutees dans le sol les plantes puis lrsquoeau ont montreacute
que ces substrats pouvaient ecirctre des sources de bacteacuteries morphologiquement tregraves proches du bacille
de Koch Le genre Mycobacterium (mứkecircs champignon et baktecircrίa bacircton) est proposeacute en 1896 par
Lehmann et Neumann Dans les anneacutees 50 environ 20 espegraveces de mycobacteacuteries eacutetaient deacutecrites Le
deacuteveloppement des meacutethodes de culture puis des outils moleacuteculaires dans les anneacutees 1990 ont permis
drsquoisoler de nouvelles espegraveces dans des niches environnementales varieacutees ainsi que chez lrsquohomme et
les animaux A lrsquoheure actuelle 172 espegraveces de mycobacteacuteries sont identifieacutees (Site bacterionetorg)
La diversiteacute pheacutenotypique physiologique eacutecologique et pathogeacutenique des espegraveces placeacutees au
fur et agrave mesure de leur deacutecouverte dans ce genre a meneacute agrave lrsquoeacutelaboration de plusieurs classifications
Au sein du genre Mycobacterium les espegraveces sont geacuteneacuteralement classeacutees en deux grands groupes un
groupe qui rassemble les mycobacteacuteries pathogegravenes obligatoires que sont les mycobacteacuteries
tuberculeuses et les mycobacteacuteries responsables de la legravepre et un second groupe qui rassemble les
mycobacteacuteries pathogegravenes opportunistes et saprophytes appeleacute mycobacteacuteries atypiques ou
mycobacteacuteries environnementales ou mycobacteacuteries non tuberculeuses (MNT)
Dans ces groupes des complexes drsquoespegraveces ont eacuteteacute deacutefinis sur des bases drsquoidentiteacute geacuteneacutetique Le
Complexe tuberculosis (MTBC) rassemble les mycobacteacuteries tuberculeuses notamment
Chapitre 1
5
M tuberculosis M africanum M canetti M bovis M caprae M microti M pinnipedii Parmi les
nombreuses MNT diffeacuterents complexes sont observeacutes dont le plus important est le Complexe
Mycobacterium avium (MAC) qui regroupe quatre sous-espegraveces distinctes dont trois drsquointeacuterecirct en santeacute
humaine et animale (M avium ssp avium M avium ssp paratuberculosis et M avium ssp hominisuis)
(Radomski et al 2010) ainsi qursquoune dizaine drsquoespegraveces dont M intracellulare
Plus reacutecemment la phylogeacutenie moleacuteculaire de ce genre baseacutee notamment sur lrsquoanalyse des gegravenes de
lrsquoARNr 16S et 23S ainsi que des reacutearrangements de gegravenes a permis drsquoaffiner la position taxonomique
du genre Mycobacterium Drsquoapregraves le Bergeyrsquos manual (Ludwig et al 2008) le genre Mycobacterium
appartient
au Phylum des Actinobacteria (bacteacuteries Gram positif agrave geacutenome riche en acides nucleacuteiques
G et C ()
o agrave la Classe des Actinobacteria
o agrave lrsquoOrdre des Corynebacteriales qui regroupe diffeacuterentes familles de bacteacuteries
caracteacuteriseacutees par la preacutesence drsquoacides mycoliques dans leur paroi et des
proprieacuteteacutes tinctoriales particuliegraveres lieacutees agrave la reacutesistance de leur paroi agrave la
deacutecoloration par un meacutelange alcool-acide (ou BAAR)
agrave la Famille des Mycobacteriaceae qui ne comprend que le genre
Mycobacterium caracteacuteriseacute par une enveloppe riche en acides
mycoliques contenant un grand nombre drsquoatomes de carbone ainsi
que des quinones particuliegraveres
2 Caracteacuteristiques geacuteneacuterales des mycobacteacuteries
a Morphologie des mycobacteacuteries
Les mycobacteacuteries se preacutesentent sous forme de bacirctonnets droits ou leacutegegraverement incurveacutes (02-
06 x 10-10microm) non mobiles et ne sporulant pas Les colonies observeacutees sont de couleur blanche agrave
cregraveme voire jaune agrave orangeacutee pour les mycobacteacuteries productrices de pigments (Magee amp Ward 2012)
Elles fixent difficilement la coloration de Gram et bien qursquoelles soient apparenteacutees aux bacteacuteries Gram
+ leur qualification exacte est laquo Bacilles Acido-Alcoolo-Reacutesistants (BAAR) raquo car lors de la coloration
leur enveloppe particuliegravere reacutesiste agrave la deacutecoloration agrave lrsquoalcool et agrave lrsquoacide
Les mycobacteacuteries possegravedent une enveloppe proche de celle des bacteacuteries Gram + mais dont
lrsquoorganisation la richesse et la diversiteacute des acides gras qui la composent sont uniques et
caracteacuteristiques La mycomembrane est constitueacutee drsquoacides gras agrave longue chaine contenant de 60 agrave 90
atomes de carbone les acides mycoliques qui repreacutesentent 30 agrave 40 de sa masse (Brennan amp Nikaido
Chapitre 1
6
1995) Dans sa couche interne les acides mycoliques sont disposeacutes parrallegravelement les uns aux autres
et sont lieacutes au peptidoglycane de faccedilon covalente par lrsquointermeacutediaire de la couche drsquoarabinogalactane
(polymegravere formeacute drsquoarabinose et de galactose) Sa couche externe contient des lipides extractibles (ie
non fixeacutes de faccedilon covalente au peptidoglycane) tels que le treacutehalose monomycolate ou dimycolate
des phospholipides et des glycolipides (Zuber et al 2008) La mycomembrane est recouverte de
glucanes de proteacuteines et drsquoune faible proportion de lipides
Cette enveloppe particuliegravere tregraves riche en lipides confegravere agrave la bacteacuterie ses proprieacuteteacutes
tinctoriales (BAAR) sa grande reacutesistance aux stress chimiques notamment aux antibiotiques et
deacutetergents aux stress physiques (tempeacuterature) ainsi que biologiques (reacutesistance dans les
macrophages de lrsquohocircte et dans les amibes) Crsquoest eacutegalement un facteur important de virulence chez les
mycobacteacuteries pathogegravenes certains composants lipidiques possegravedent en effet une activiteacute
immunogegravene exceptionnelle (Coulombe et al 2009) Ils sont capables de moduler lrsquoimmuniteacute de lrsquohocircte
voire de deacutesorganiser ses meacutecanismes immunitaires de deacutefense et interviennent dans la pathogeacutenie
comme cela a eacuteteacute deacutemontreacute pour M tuberculosis et M avium ssp paratuberculosis (Libero amp Mori
2014 Momotani et al 2012) La paroi est drsquoailleurs une des cibles des antimicrobiens utiliseacutes contre la
tuberculose humaine ou la legravepre Ces derniers agissent notamment en inhibant la biosynthegravese des
moleacutecules qui la constituent Lrsquoutilisation des antigegravenes lipidiques de la paroi constitue eacutegalement un
axe de recherche pour le deacuteveloppement de vaccins (Libero amp Mori 2014)
Dans les matrices environnementales la paroi hydrophobe des mycobacteacuteries leur permet
drsquoadheacuterer fortement aux particules en suspension dans lrsquoeau aux seacutediments aux surfaces favorisant
la formation de biofilms et la survie des bacteacuteries (Primm et al 2004)
Les acides mycoliques de par leur complexiteacute et leur variabiliteacute structurelle sont eacutegalement utiliseacutes
comme marqueurs taxonomiques car ils permettent drsquoidentifier avec speacutecificiteacute le genre et les espegraveces
Chapitre 1
7
Figure 1 Structure et composition de lenveloppe des mycobacteacuteries (drsquoapregraves Marrakchi Laneacuteelle and Daffeacute 2014) AG arabinogalactane PG peptidoglycane TDM treacutehalose dimycolate PL phospholipides TMM treacutehalose monomycolate GL glycolipides
b Caracteacuteristiques culturales pheacutenotypiques et physiologiques
La plupart des mycobacteacuteries sont aeacuterobies strictes mais quelques espegraveces tolegraverent de faibles
pressions drsquooxygegravene notamment M bovis et M tuberculosis Les substrats utiliseacutes comme sources de
carbone et drsquoazote varient consideacuterablement entre les espegraveces et peuvent servir pour leur
identification La majoriteacute des espegraveces croit sur des milieux de culture classiques mais pour certaines
mycobacteacuteries drsquoimportance clinique la culture est ameacutelioreacutee par lrsquoutilisation de milieux agrave lrsquoœuf
(Loumlwenstein-Jensen et Coletsos) apregraves une deacutecontamination chimique ou de milieu Middlebrook ainsi
que par un enrichissement des milieux (en glyceacuterol pour M tuberculosis en mycobactine pour
M avium subsp paratuberculosis en heacutemine pour M haemophilum) Les mycobacteacuteries responsables
de la legravepre sont cultiveacutees sur tissus vivants (coussinets de souris peau de tatou) et se multiplient tregraves
difficilement sur milieux syntheacutetiques
Les mycobacteacuteries ont des vitesses de croissance tregraves variables on distingue les mycobacteacuteries
agrave croissance rapide dont les colonies sont visibles en moins de 7 jours des mycobacteacuteries agrave croissance
lente (gt7 jours) Les bacteacuteries du MTBC sont toutes des mycobacteacuteries agrave croissance lente avec des
temps de geacuteneacuteration eacuteleveacutes de 14 agrave 15h pour M tuberculosis et environ 20h pour M bovis La
tempeacuterature optimale de croissance des bacteacuteries du MTBC est de 37degC Les MNT regroupent des
mycobacteacuteries agrave croissance rapide et agrave croissance lente avec des taux de geacuteneacuteration extrecircment
variables allant de 2h pour les bacteacuteries agrave croissance rapide comme M smegmatis agrave 21h pour celles
agrave croissance lente comme M aurum voire 7 jours pour M avium ssp paratuberculosis dans des
conditions de laboratoire pourtant optimales Les MNT peuvent se multiplier dans un large eacuteventail de
Chapitre 1
8
tempeacuterature allant de 4degC pour M psychrotolerans (Trujillo et al 2004) agrave plus de 45degC voire agrave 52degC
pour M thermotolerans et M phlei (Stahl amp Urbance 1990) avec une tempeacuterature optimale de
croissance entre 28 et 31degC
c Phylogeacutenie du genre Mycobacterium
La phylogeacutenie baseacutee sur le polymorphisme de lrsquoADN codant pour lrsquoARNr 16S permet de
discriminer les espegraveces de mycobacteacuteries et met en eacutevidence trois grands groupes en fonction de leur
vitesse de croissance Le cladogramme preacutesenteacute dans la Figure 2 regroupe distinctement les
mycobacteacuteries agrave croissance lente (haut du cladogramme) les mycobacteacuteries agrave croissance intermeacutediaire
(au milieu) et les mycobacteacuteries agrave croissance rapide (bas du cladogramme) On peut noter de plus que
les mycobacteacuteries du MTBC et le complexe Mac forment deux groupes phylogeacuteneacutetiquement
homogegravenes au sein du groupe des mycobacteacuteries agrave croissance lente
Myc
ob
acteacute
ries
agrave c
rois
san
ce le
nte
Myc
ob
acteacute
ries
agrave
cro
issa
nce
inte
rmeacute
dia
ire
Chapitre 1
9
Figure 2 Arbre phylogeacutenique baseacute sur lrsquoanalyse de lrsquoARNr 16S des principaux complexes et espegraveces de mycobacteacuteries
(drsquoapregraves Magee et Ward 2012)
d Caracteacuteristiques geacuteneacutetiques des mycobacteacuteries
De nombreux geacutenomes complets (annoteacutes ou en cours drsquoanalyse) de mycobacteacuteries sont
actuellement disponibles (NCBI Genome Resources European Nucleotide archive) Mycobacterium
leprae est la mycobacteacuterie qui possegravede le geacutenome le plus petit avec 33 millions de paires de bases
(Mpb) codant pour 1 605 gegravenes et le G+C (pourcentage Guanine+Cytosine) le plus faible (57) Les
mycobacteacuteries environnementales possegravedent les geacutenomes les plus importants de 483 Mpb (codant
4 350 gegravenes) pour M avium k10 agrave 69 Mpb (codant 6 716 gegravenes) pour M smegmatis Les mycobacteacuteries
tuberculeuses M tuberculosis M africanum et M bovis ont un geacutenome de taille intermeacutediaire (de 43
Mpb agrave 45 Mpb) codant pour 3 850 agrave 4 500 gegravenes Les geacutenomes des mycobacteacuteries preacutesentent des
capaciteacutes codantes importantes de lrsquoordre de 90 pour M tuberculosis M bovis M avium ssp
paratuberculosis et M smegmatis (Garnier et al 2003 Cole et al 1998 Li et al 2005)
La comparaison du geacutenome complet de M tuberculosis M bovis M leprae et M avium ssp
paratuberculosis a montreacute une forte conservation des gegravenes impliqueacutes dans le meacutetabolisme
Myc
ob
acteacute
ries
agrave c
rois
san
ce r
apid
e
Chapitre 1
10
eacutenergeacutetique la biosynthegravese des acides amineacutes et des cofacteurs et le meacutetabolisme des nucleacuteotides et
des proteacuteines (Marri et al 2006) Des plasmides ont eacuteteacute mis en eacutevidence chez quelques espegraveces de
mycobacteacuteries environnementales (Rauzier et al 1988 Stinear et al 2004) notamment le plasmide
pMUM001 isoleacute chez M ulcerans responsable de lrsquoulcegravere de Buruli chez lrsquohomme qui code pour un
macrolide aux proprieacuteteacutes cytotoxiques et immunosuppressives la mycolactone (Stinear et al 2004)
Le nombre drsquoopeacuterons ribosomaux 16S dans le geacutenome des mycobacteacuteries a eacuteteacute suggeacutereacute comme
lrsquoun des facteurs expliquant la diffeacuterence des vitesses de croissance observeacutees La croissance lente des
mycobacteacuteries serait lieacutee agrave la preacutesence drsquoun seul opeacuteron ribosomal les espegraveces agrave croissance rapide en
posseacutedant geacuteneacuteralement deux (rrnA et rrnB) (Bercovier et al 1986) Toutefois des exceptions existent
et drsquoautres facteurs tels que la lenteur des eacutechanges agrave travers leur paroi impermeacuteable riche en lipides
et le coucirct eacutenergeacutetique eacuteleveacute de la synthegravese de leurs acides gras agrave longues chaines seraient eacutegalement
impliqueacutes (Primm et al 2004)
Dans le Complexe tuberculosis les diffeacuterentes espegraveces constituent un modegravele drsquoidentiteacute
geacuteneacutetique interspeacutecifique Leurs geacutenomes preacutesentent une identiteacute supeacuterieure agrave 999 au niveau
nucleacuteotidique et des seacutequences drsquoARNr 16S et drsquoITS identiques (Boumlddinghaus et al 1990 Sreevatsan amp
Pan 1997 Huard et al 2006 Cole et al 1998 Garnier et al 2003 Kasai et al 2000) Lrsquoanalyse des
seacutequences de 56 gegravenes de structure de plusieurs centaines drsquoisolats drsquoorigines geacuteographique et
phylogeacuteneacutetique varieacutees suggegravere que le polymorphisme alleacutelique est exceptionnel dans ce Complexe
(Sreevatsan amp Pan 1997 Musser et al 2000) La recombinaison de tregraves petits fragments drsquoADN est
deacutemontreacutee chez M tuberculosis (Namouchi et al 2012) mais le transfert lateacuteral de gegravenes ne semble
pas ecirctre un facteur important dans lrsquoeacutevolution des bacteacuteries du MTBC qui ont une structure de
population fortement clonale (Pepperell et al 2013) contrairement agrave la plupart des autres bacteacuteries
pathogegravenes Malgreacute leur forte identiteacute geacuteneacutetique les espegraveces du Complexe se diffeacuterencient par des
reacutegions polymorphiques ou LSPs (Large Sequence Polymorphisms) lieacutees agrave des deacuteleacutetions et agrave des
reacutearrangements de seacutequences (Brosch et al 2002 Huard et al 2003) Parmi ces LSPs 14 Reacutegions de
Diffeacuterence ou RD de 2 agrave 127 kpb sont recenseacutees dans le MTBC Ces deacuteleacutetions sont consideacutereacutees comme
une adaptation eacutevolutive agrave lrsquoinfection de nouveaux hocirctes plutocirct qursquoagrave une deacuterive geacuteneacutetique (Brodin et
al 2002 Aranaz et al 2003) Cette variabiliteacute geacutenomique permet agrave la fois de diffeacuterencier les espegraveces
entre elles (Huard et al 2003 Huard et al 2006) et de retracer leur eacutevolution phylogeacuteneacutetique
(Rodriguez-Campos et al 2014)
3 Description des diffeacuterents Complexes ou espegraveces de mycobacteacuteries
Alors que les mycobacteacuteries responsables de la tuberculose et de la legravepre sont des pathogegravenes
obligatoires les MNT rassemblent des mycobacteacuteries saprophytes sans pouvoir pathogegravene jusque-lagrave
Chapitre 1
11
aveacutereacute et des mycobacteacuteries pathogegravenes opportunistes responsables de mycobacteacuterioses humaines ou
animales Depuis la fin du 19egraveme siegravecle jusqursquoaux anneacutees 1950 la tuberculose humaine repreacutesentait un
tel problegraveme de santeacute publique que toutes les eacutetudes eacutetaient centreacutees sur elle et son agent
M tuberculosis Les autres mycobacteacuteries isoleacutees dans lrsquoenvironnement ou dans des preacutelegravevements
biologiques humains et animaux eacutetaient consideacutereacutees comme des bacteacuteries saprophytes sans inteacuterecirct
clinique particulier A partir des anneacutees 1950 la communauteacute scientifique a admis la responsabiliteacute
indiscutable de certaines mycobacteacuteries environnementales dans des processus pathologiques
humains et animaux appeleacutes mycobacteacuterioses amorccedilant des eacutetudes les concernant En lrsquoabsence de
contamination inter-humaine deacutemontreacutee il a eacuteteacute admis que lrsquoorigine de ces mycobacteacuteries est
environnementale et que lrsquohomme se contamine agrave partir de lrsquoenvironnement
a Les mycobacteacuteries non pathogegravenes potentiellement opportunistes
Elles regroupent les mycobacteacuteries environnementales qui preacutesentent un pouvoir pathogegravene
pour lrsquohomme et les animaux tregraves variable en fonction des espegraveces (van Ingen et al 2009 Tortoli 2003)
Lrsquoincidence des infections humaines lieacutees agrave des mycobacteacuteries opportunistes semble augmenter dans
les pays industrialiseacutes (Falkinham 2016) peut-ecirctre en lien avec lrsquoameacutelioration des outils pour les isoler
et les identifier Les mycobacteacuterioses qursquoelles engendrent surviennent geacuteneacuteralement sur un terrain
drsquoimmunodeacutepression locale ou geacuteneacuterale Chez lrsquohomme elles sont responsables drsquoun large eacuteventail de
symptocircmes cliniques majoritairement des infections pulmonaires mais eacutegalement des lymphadeacutenites
chez les enfants des infections cutaneacutees (notamment lieacutees agrave M marinum responsable du granulome
des piscines et M ulcerans responsable de lrsquoulcegravere de Buruli) des infections extra-pulmonaires voire
des infections disseacutemineacutees chez les patients seacutevegraverement immunodeacuteprimeacutes (van Ingen 2013 Biet et al
2005 Hoefsloot et al 2013) Environ 25 espegraveces sont retrouveacutees reacuteguliegraverement dans les eacutechantillons
cliniques les autres beaucoup plus rarement (van Ingen 2013) Parmi elles les mycobacteacuteries agrave
croissance lente semblent plus preacutedisposeacutees agrave provoquer des atteintes pulmonaires ou lymphatiques
alors que celles agrave croissance rapide affectent plutocirct la peau les os et les articulations (Tortoli 2003
2009) Les mycobacteacuteries M avium et M intracellulare sont les plus freacutequemment isoleacutees chez
lrsquohomme majoritairement en cas drsquoatteinte pulmonaire (Hoefsloot et al 2013 Whiley et al 2012) La
distribution des espegraveces pathogegravenes isoleacutees chez lrsquohomme diffegravere selon la zone geacuteographique eacutetudieacutee
probablement en lien avec leurs niches eacutecologiques (Hoefsloot et al 2013)
Chez les animaux M avium ssp paratuberculosis (Map) est lrsquoagent de la maladie de Johne ou
paratuberculose chez les ruminants domestiques et sauvages les primates ainsi que drsquoautres espegraveces
(lapins etc) chez qui elle provoque une enteacuterite chronique contagieuse et incurable agrave lrsquoorigine de
pertes eacuteconomiques importantes dans les cheptels infecteacutes Son rocircle est eacutegalement eacutevoqueacute dans la
Chapitre 1
12
maladie de Crohn chez lrsquohomme (Biet et al 2005 Rowe amp Grant 2006 Momotani et al 2012) sans
toutefois que la causaliteacute ait eacuteteacute deacutemontreacutee Mycobacterium avium ssp avium (Maa) est responsable
de la tuberculose aviaire chez les oiseaux et les petits mammifegraveres terrestres chez qui elle provoque
des leacutesions tuberculeuses majoritairement digestives des lymphadeacutenites et des infections disseacutemineacutees
(Biet et al 2005 Pavlik et al 2000) Mycobacterium avium ssp hominissuis (Mah) est responsable de
leacutesions des nœuds lymphatiques et de diffeacuterents organes chez le porc (Pavlik et al 2000)
b Les mycobacteacuteries saprophytes non pathogegravenes
Parmi toutes les mycobacteacuteries environnementales deacutecrites agrave lrsquoheure actuelle certaines sont
retrouveacutees uniquement dans lrsquoenvironnement et nrsquoont jamais eacuteteacute isoleacutees chez lrsquohomme ou lrsquoanimal
comme M arabiense ou M rufum (Tortoli 2014) Ces mycobacteacuteries sont dites saprophytes Parmi
elles certaines ont eacuteteacute isoleacutees dans des sols et des seacutediments fortement pollueacutes par les HAP
(Hydrocarbures Aromatiques Polycycliques) qursquoelles sont potentiellement capables de deacutegrader
(Hennessee et al 2009 Leys et al 2005 Hormisch et al 2004) Elles repreacutesentent donc une voie
inteacuteressante pour la bioremeacutediation de ces sols
c Les mycobacteacuteries pathogegravenes obligatoires
Les mycobacteacuteries responsables de legravepre
Les mycobacteacuteries responsables de la legravepre humaine et murine sont M lepreae M lepromatosis et
M lepreaemurium Elles provoquent des infections dermatologiques et nerveuses chroniques graves
et mutilantes parfois mortelles Mycobacterium leprae touche environ 15 million de personnes dans
le monde et la maladie reste un problegraveme de santeacute publique preacuteoccupant dans les zones endeacutemiques
drsquoAfrique drsquoAsie et drsquoAmeacuterique du sud (Who 2014) Mycobacterium leprae a longtemps eacuteteacute consideacutereacutee
comme un pathogegravene obligatoire de lrsquohomme avec une voie de transmission interhumaine majoritaire
La deacutecouverte de reacuteservoirs animaux (Truman 2005 Meredith et al 2014) et la deacutetection de M leprae
dans le sol la sphaigne ou lrsquoeau de riviegravere (Matsuoka et al 1999 Mostafa et al 1995 Lavania et al
2006) tendent agrave souligner le rocircle potentiel de certains animaux ou de lrsquoenvironnement comme source
de lrsquoinfection chez lrsquohomme Ces 3 espegraveces sont multiplieacutees sur les coussinets de souris (Shepard 1960)
et sur la peau de tatous agrave 9 bandes (Dasypus novemcinctus) (Kirchheimer amp Storrs 1971) car aucun
milieu syntheacutetique ne permet leur croissance optimale
Chapitre 1
13
Les mycobacteacuteries tuberculeuses le Complexe tuberculosis
(MTBC)
Toutes les mycobacteacuteries tuberculeuses sont des pathogegravenes obligatoires Contrairement aux
mycobacteacuteries environnementales leur croissance dans lrsquoenvironnement nrsquoa jamais eacuteteacute deacutemontreacutee Le
complexe tuberculosis comprend agrave ce jour 13 espegraveces qui malgreacute une forte identiteacute geacuteneacutetique
preacutesentent des pheacutenotypes des hocirctes et des pouvoirs pathogegravenes varieacutes
Phylogeacutenie des mycobacteacuteries du MTBC
Lrsquoeacutetude des RD eacutevoqueacutees preacuteceacutedemment et des mutations agrave lrsquoeacutechelle du nucleacuteotide (ou SNP Single
Nucleacuteotide Polymorphism) sur 7 gegravenes (katG gyrA gyrB rpoB mmpL6 oxyR et ADNr 16S) ont permis
de retracer la phylogeacutenie des bacteacuteries du Complexe agrave partir de leur ancecirctre commun (Brosch et al
2002 Rodriguez-Campos et al 2014 Gordon et al 1999 Mahairas et al 1996 Huard et al 2003) les
deacuteleacutetions et mutations jouant un rocircle drsquohorloge moleacuteculaire permettant de replacer lrsquoapparition et
lrsquoeacutevolution drsquoune espegravece dans le temps Le sceacutenario de lrsquoeacutevolution des bacteacuteries du MTBC est
scheacutematiseacute dans la Figure 3 Selon les auteurs lrsquoancecirctre du MTBC appeleacute M protuberculosis aurait
eacutemergeacute il y a entre 15 000 et 40 000 ans en Afrique de lrsquoEst (Gutierrez et al 2005 Wirth et al 2008
Sreevatsan amp Pan 1997) A cocircteacute de M canetti la bacteacuterie du MTBC geacuteneacutetiquement la plus proche de
lrsquoancecirctre deux clades principaux peuvent ecirctre observeacutes Un clade de souches speacutecifiques de lrsquohomme
caracteacuteriseacute par la deacuteleacutetion TbD1 a donneacute les diffeacuterentes ligneacutees modernes de M tuberculosis (Figure
3) Un second clade caracteacuteriseacute par la perte des reacutegions RD7 RD8 RD9 et RD10 regroupe des souches
endeacutemiques drsquoAfrique speacutecifiques de lrsquohomme (M africanum) et drsquoanimaux (le Chimpanzee bacillus
M mungi le Dassie bacillus et M suricattae) ainsi que des souches adapteacutees agrave diffeacuterentes espegraveces de
mammifegraveres (M orygis M microti M pinnipedii M caprae et M bovis)
Chapitre 1
14
Figure 3 Phylogeacutenie des souches du MTBC baseacutee sur les Reacutegions de Diffeacuterences et diffeacuterents Single Nucleotide Polymorphism (Dippenaar et al 2015)
Virulence des mycobacteacuteries du MTBC
Les mycobacteacuteries du MTBC ne possegravedent pas de facteurs de virulence classiques tels que les
toxines par exemple observeacutes chez drsquoautres bacteacuteries pathogegravenes Pour certains auteurs la virulence
des mycobacteacuteries est restreinte agrave lrsquoaptitude du bacille agrave peacuteneacutetrer dans lrsquohocircte agrave y survivre et srsquoy
multiplier (Pollock amp Neill 2002) De faccedilon plus large la notion de virulence chez les membres du MTBC
a eacuteteacute conceptualiseacutee comme lrsquoassociation de i) la capaciteacute de la bacteacuterie agrave faire face agrave la reacuteponse
immunitaire de lrsquohocircte infecteacute ii) la capaciteacute agrave engendrer des leacutesions iii) la capaciteacute agrave survivre en dehors
de son hocircte notamment lors de lrsquoaeacuterosolisation et iiii) la transmission et lrsquoinfection drsquoun nouvel hocircte
(Coscolla amp Gagneux 2014) La virulence implique donc un ensemble de meacutecanismes complexes et
varieacutes faisant intervenir de nombreux gegravenes et proteacuteines permettant notamment au bacille de peacuteneacutetrer
dans les cellules de lrsquohocirctes et drsquoy survivre en bloquant les diffeacuterents meacutecanismes de deacutefense des
macrophages (inhibition de la fusion phagosome-lysosome et de la production de ROS1 et RNS2
deacutesorganisation de lrsquoimmuniteacute de lrsquohocircte inhibition de lrsquoautophagie et de lrsquoapoptose) (Hmama et al
1 ROS Reactive Oxygen Species 2 RNS Reactive Nitrogen Species
Chapitre 1
15
2015) Plus de cent gegravenes seraient impliqueacutes dans la virulence des bacteacuteries du MTBC et lrsquoutilisation
de mutants permet peu agrave peu de connaitre le rocircle de chacun (Forrellad et al 2013) Lrsquoanalyse du
transcriptome (ARN) de M tuberculosis au cours de lrsquoinfection de macrophages et de cellules
dendritiques a reacuteveacuteleacute lrsquoexpression ou la surexpression de nombreux gegravenes codant des proteacuteines
intervenant notamment dans le meacutetabolisme lipidique la synthegravese de lrsquoenveloppe bacteacuterienne
lrsquoapprovisionnement en fer la reacutesistance au stress oxydatif et lrsquoinhibition de lrsquoapoptose (Mukhopadhyay
et al 2012) Les meacutecanismes de survie en dehors de lrsquohocircte sont inconnus agrave ce jour
Il a eacuteteacute montreacute chez M tuberculosis qursquoen fonction des ligneacutees ou des souches la virulence pouvait
varier entrainant une reacuteponse immunitaire reacuteduite ou retardeacutee chez lrsquohocircte une plus grande seacuteveacuteriteacute
de la maladie et une transmission augmenteacutee (Coscolla amp Gagneux 2014) Ce lien souche-virulence est
eacutegalement suspecteacute chez M bovis pour lequel certains geacutenotypes srsquoaccompagnent drsquoun plus grand
nombre drsquoanimaux agrave leacutesions et ou drsquoun score leacutesionnel plus eacuteleveacute (Wright et al 2013 Gavier-Wideacuten et
al 2009 Garbaccio et al 2014) mais des eacutetudes compleacutementaires restent neacutecessaires
Les mycobacteacuteries speacutecifiques de lrsquohomme
Mycobacterium tuberculosis
Mycobacterium tuberculosis est lrsquoagent de la tuberculose humaine maladie retrouveacutee dans
toutes les reacutegions du monde Crsquoest un problegraveme majeur de santeacute publique puisque 2 milliards
drsquoindividus seraient infecteacutes dans le monde En 2013 lrsquoOMS a eacutevalueacute agrave neuf millions le nombre de
nouveaux cas annuels et agrave un million et demi le nombre de deacutecegraves lieacutes agrave la tuberculose (dont 24 de
personnes seacuteropositives) Les reacutegions de lrsquoAsie du sud-est et du Pacifique occidental regroupent 56
des nouveaux cas lrsquoAfrique 25 lrsquoInde 24 et la Chine 11 (WHO 2014) Lrsquoinfection par le HIV est un
facteur preacutedisposant majeur dans la transmission de la maladie et la progression de lrsquoinfection chez
lrsquohomme de par les effets synergiques deacuteleacutetegraveres des 2 micro-organismes sur le systegraveme immunitaire
(Pawlowski et al 2012) Environ 78 des personnes co-infecteacutees vivent en Afrique (WHO 2014) La
tuberculose se manifeste chez lrsquohomme par des granulomes pulmonaires (tuberculose pulmonaire)
mais peut se deacutevelopper dans drsquoautres organes (tuberculose extra-pulmonaire) Elle se transmet
essentiellement par voie respiratoire par lrsquoinhalation de gouttelettes contamineacutees eacutemises par une
personne infecteacutee lors de toux par exemple Le traitement repose sur lrsquoassociation de plusieurs
antibiotiques adapteacutes tels que lrsquoisoniazide la rifampicine le pyrazinamide et lrsquoeacutethambutol
Lrsquoapparition de souches reacutesistantes aux antibiotiques MDR (Multi Drug Resistant) et XDR
(eXtensively Drug Resistant) constitue agrave lrsquoheure actuelle un problegraveme majeur dans la lutte contre la
maladie Bien que lrsquohomme constitue le reacuteservoir majeur de la bacteacuterie M tuberculosis eacuteteacute isoleacutee chez
Chapitre 1
16
un grand nombre drsquoespegraveces animales telles que les bovins domestiques (Romero et al 2011 Prasad et
al 2005 Ameni et al 2011 Pavlik et al 2005 Cadmus et al 2006) les chegravevres (Cadmus Adesokan et
al 2009) les porcs domestiques (Jenkins et al 2011) les chiens (Pavlik et al 2005) les oiseaux et des
animaux de zoos (Michel et al 2003) dont les eacuteleacutephants
Mycobacterium africanum
Elle est principalement retrouveacutee dans les pays drsquoAfrique de lrsquoouest ougrave elle peut ecirctre impliqueacutee
dans la moitieacute des cas de tuberculose humaine agrave cocircteacute de M tuberculosis (de Jong et al 2010) Isoleacutee
et deacutecrite pour la premiegravere fois au Seacuteneacutegal en 1968 elle preacutesente des caracteacuteristiques biochimiques
variables intermeacutediaires entre celles de M tuberculosis et M bovis (Castets amp Sarrat 1968)
Mycobacterium africanum est parfois isoleacutee chez lrsquohomme en dehors de lrsquoAfrique (Schroumlder 1982
Grange amp Yates 1989 Desmond et al 2004 Frottier et al 1990) La proximiteacute phylogeacuteneacutetique de
M africanum avec les autres membres du MTBC infectant des espegraveces animales a souleveacute la question
drsquoun reacuteservoir animal de la bacteacuterie Mais elle a eacuteteacute isoleacutee de faccedilon sporadique sur des singes (Thorel
1980) des bovins (Rahim et al 2007 Cadmus et al 2006) et du lait (Cadmus et al 2010) infirmant
cette hypothegravese
Mycobacterium canetti et smooth tubercle bacilli
Elle est responsable de tuberculose humaine dans une zone geacuteographique confineacutee agrave la corne de
lrsquoAfrique (Djibouti Somalie Kenya) (Pfyffer et al 1998 Miltgen et al 2002) Elle est la plus proche
geacuteneacutetiquement de lrsquoancecirctre des bacteacuteries du MTBC Elle constitue un taxon divergent des autres
membres du MTBC par son pheacutenotype particulier et son geacutenotype qui preacutesente une large diversiteacute
geacuteneacutetique et des signes de transfert horizontal et de recombinaison de gegravenes contrairement aux autres
bacteacuteries du MTBC (Fabre et al 2004 Supply et al 2013 Gutierrez et al 2005) Sa situation
geacuteographique confineacutee la grande diversiteacute geacuteneacutetique des souches rencontreacutees et le manque de
connaissances sur la transmission de la bacteacuterie ont suggeacutereacute lrsquoexistence drsquoun reacuteservoir environnemental
(Fabre et al 2004 Koeck et al 2011) A ce jour cette hypothegravese nrsquoa pas eacuteteacute confirmeacutee La tuberculose
agrave M canetti est consideacutereacutee aujourdrsquohui comme une maladie eacutemergente
Les mycobacteacuteries speacutecifiques des animaux
Mycobacterium bovis
Mycobacterium bovis est lrsquoagent de la tuberculose bovine maladie preacutesente sur tous les
continents Les bovins constituent le reacuteservoir naturel du bacille mais un grand nombre drsquoespegraveces
Chapitre 1
17
animales domestiques et sauvages peuvent ecirctre infecteacutees ainsi que lrsquohomme (Biet et al 2005 Palmer
2013 Grange amp Yates 1994 OrsquoReilly amp Daborn 1995) Sur le plan microbiologique M bovis se
caracteacuterise par une croissance tregraves lente sur milieu de Loumlwenstein-Jensen (6 agrave 10 semaines) par des
colonies petite taille (1 agrave 2 mm) plates lisses laquo dysgoniques raquo et achromogegravenes Sa croissance est
ameacutelioreacutee par lrsquoajout de pyruvate et inhibeacutee par lrsquoajout de glyceacuterol Sur Middlebrook 7H11 les colonies
sont rugueuses et agrave bords eacutechancreacutes (Figure 4) Le pH optimum de culture de M bovis est compris
entre 58 et 69 mais la bacteacuterie survit 20 jours dans du yaourt agrave 4degC (Mitscherlich amp Marth 2012)
Lrsquoidentification pheacutenotypique repose notamment sur lrsquoabsence de production de niacine lrsquoabsence de
nitrate reacuteductase une reacutesistance au pyrazinamide agrave la D-cycloseacuterine (30mgmL-1) et une croissance
favoriseacutee par la preacutesence de pyruvate (Delaune et al 2012)
Figure 4 Aspect des colonies de M bovis sur milieu de Lowenstein-Jensen (agrave gauche) et Middlebrook 7H11 suppleacutementeacute (agrave droite)
Concernant les caracteacuteristiques geacutenomiques de M bovis le seacutequenccedilage complet de la
premiegravere souche (M bovis AF212297 ou ATCC BAA-935 isoleacutee drsquoune leacutesion caseacuteeuse de bovin en
Grande-Bretagne) en 2003 et son annotation ont enrichi les connaissances sur ce bacille Son geacutenome
comprend 4345 Mpb un pourcentage G-C eacuteleveacute de 656 et 3952 gegravenes codant des proteacuteines dont 1
prophage et 42 seacutequences drsquoinsertion Il preacutesente 11 deacuteleacutetions de 1 agrave 127 kb par comparaison avec le
geacutenome de M tuberculosis H37Rv et 1 locus absent chez M tuberculosis appeleacute TbD1 (Garnier et al
2003) Ne posseacutedant pas de gegravenes propres agrave lrsquoespegravece bovis le geacutenome de M bovis a eacuteteacute faccedilonneacute par
lrsquoapparition de ces deacuteleacutetions conditionnant lrsquoexpression des gegravenes Depuis 2003 une quarantaine de
souches isoleacutees sur des animaux domestiques et sauvages ont eacuteteacute entiegraverement seacutequenceacutees (Sites
NCBI et European Nucleotide archive)
La structure de la population de M bovis reacutesulte de lrsquoexpansion de plusieurs complexes clonaux
issus drsquoun mecircme ancecirctre Quatre complexes clonaux majeurs ont eacuteteacute identifieacutes dans le monde pour
Chapitre 1
18
M bovis en se basant sur la preacutesence de deacuteleacutetions chromosomiques caracteacuteristiques dans le geacutenome
de la bacteacuterie etou sur des SNPs ainsi que sur le spoligotype et lrsquoabsence de certains spacers (Smith
2012) le Complexe African 1 est retrouveacute dans plusieurs pays africains (Mali Cameroun Nigeacuteria
Tchad) (Muumlller et al 2009) le Complexe African 2 est essentiellement retrouveacute chez les bovideacutes
drsquoAfrique de lrsquoEst (Berg et al 2011) le Complexe European 1 est retrouveacute en Europe il est dominant au
Royaume-Uni et beaucoup plus rare en Espagne au Portugal et en France On le retrouve eacutegalement
chez les anciens partenaires commerciaux du Royaume-Uni (USA Afrique du Sud Nouvelle Zeacutelande
Australie Canada) et en Ameacuterique du sud La diffusion geacuteographique de ce Complexe est lieacutee aux
eacutechanges commerciaux de bovins (Smith et al 2011) Enfin le Complexe European 2 est majoritaire au
Portugal et en Espagne et assez freacutequent en France et en Italie (Rodriguez-Campos et al 2012) Drsquoautres
complexes clonaux minoritaires existent En France plusieurs complexes clonaux coexistent certains
autochtones eacutevoluent depuis longtemps sur le territoire alors que drsquoautres ont eacuteteacute importeacutes agrave la faveur
des eacutechanges commerciaux de bovins (Hauer et al 2015)
Une souche atteacutenueacutee obtenue agrave partir drsquoun isolat virulent de M bovis a eacuteteacute creacuteeacutee par Calmette
et Gueacuterin apregraves 230 repiquages sur un milieu bilieacute enrichi en glyceacuterol entre 1908 et 1921 le BCG pour
Bacille de Calmette et Gueacuterin Testeacutee sur des modegraveles animaux cette souche atteacutenueacutee immunisait
contre une infection expeacuterimentale agrave M bovis et M tuberculosis trente jours apregraves injection Utiliseacutee
pour la vaccination elle a eacuteteacute distribueacutee dans le monde entier et maintes fois repiqueacutee ce qui a meneacute
agrave une eacutevolution de la souche initiale en plusieurs souches tregraves heacuteteacuterogegravenes La comparaison geacutenomique
de la souche BCG avec M tuberculosis reacutevegravele la perte de 14 reacutegions (RD1 agrave RD14) chez BCG Lrsquoabsence
de la reacutegion RD1 expliquerait en partie lrsquoatteacutenuation du BCG (Pym et al 2002) toutefois comme la
reacuteintroduction de RD1 dans une souche mutante ne suffit pas agrave restaurer totalement la virulence
drsquoautres reacutegions sont impliqueacutees dans lrsquoatteacutenuation (Brosch et al 2007)
Mycobacterium pinnipedii
Deacutecouverte en 1986 cette mycobacteacuterie touche principalement les diffeacuterentes espegraveces de
phoques et les lions de mer sauvages de lrsquoheacutemisphegravere sud ou en captiviteacute Elle a eacuteteacute isoleacutee en Australie
Nouvelle-Zeacutelande Uruguay Argentine et Grande-Bretagne (Cousins et al 2003) Des infections chez
lrsquohomme ont eacuteteacute rapporteacutees principalement chez les soigneurs en contact avec les animaux malades
en captiviteacute (Kiers et al 2008) La transmission indirecte du bacille par inhalation drsquoaeacuterosols lors du
nettoyage des enclos est fortement suspecteacutee
Mycobacterium caprae
Cette mycobacteacuterie du MTBC a eacuteteacute isoleacutee pour la premiegravere fois sur des chegravevres en Espagne
(Aranaz et al 1999 Aranaz et al 2003) Crsquoest lrsquoun des agents de la tuberculose bovine avec M bovis
Chapitre 1
19
et elle induit des leacutesions identiques agrave la tuberculose agrave M bovis chez les animaux infecteacutes La
tuberculose agrave M caprae touche principalement lrsquoEurope ougrave elle a deacutejagrave eacuteteacute deacutetecteacutee en Espagne
(Rodriacuteguez et al 2011) en France (Haddad et al 2001) en Autriche (Prodinger et al 2002) en
Allemagne (Kubica et al 2003) et dans plusieurs pays drsquoEurope centrale (Pavlik et al 2005 Beširović
et al 2012 Cvetnic et al 2007) Le nombre de cas deacutetecteacutes tend agrave augmenter chez les bovins et les
animaux sauvages notamment en Espagne en Allemagne en Autriche (Rodriacuteguez et al 2011 Schoepf
et al 2012) En Europe centrale elle repreacutesente lrsquoagent principal de la tuberculose bovine notamment
en Reacutepublique Tchegraveque Slovaquie Bosnie Herzeacutegovine et Croatie ougrave elle circule entre les bovins et de
nombreuses espegraveces sauvages (Prodinger et al 2005 Beširović et al 2012) Elle a eacutegalement eacuteteacute isoleacutee
pour la premiegravere fois en Chine en 2013 (Zeng et al 2013) Crsquoest eacutegalement un agent zoonotique
important puisque des infections agrave M caprae sont rapporteacutees chez lrsquohomme dans diffeacuterents pays
drsquoEurope ((Kubica et al 2003 Prodinger et al 2014 Prodinger et al 2002 Muumlller et al 2013)
Mycobacterium microti
Cette mycobacteacuterie est lrsquoagent responsable de la tuberculose des petits rongeurs (campagnols
souris musaraignes hellip) chez qui elle a eacuteteacute isoleacutee dans les anneacutees 30 (Wells 1937 Wells amp Robb-Smith
1946) Elle a eacuteteacute isoleacutee chez plusieurs espegraveces de mammifegraveres dont des chats (Gunn-Moore et al 1996
Gunn-Moore et al 2011) des bovins (Jahans et al 2004) des porcs (Taylor et al 2006) des lamas des
blaireaux (Smith et al 2009) des sangliers (Dondo et al 2007) des suricates (Palgrave et al 2012) En
France la deacutetection de M microti chez des animaux domestiques et sauvages a suggeacutereacute que la bacteacuterie
eacutetait endeacutemique dans certaines reacutegions de France (Michelet et al 2015 Laprie et al 2013) Chez
lrsquohomme une trentaine de cas drsquoinfections agrave M microti principalement agrave localisation pulmonaire ont
eacuteteacute reporteacutes dans la litteacuterature entre 2000 et 2010 (Panteix et al 2010) Bien que les personnes
immunodeacuteprimeacutees soient les plus toucheacutees lrsquoinfection se rencontre eacutegalement chez des sujets
immunocompeacutetents (Niemann et al 2000 Frank et al 2009)
Mycobacterium orygis
Isoleacutee pour la premiegravere fois en 1976 sur des oryxes en Afrique de lrsquoest (Lomme et al 1976) elle
a eacuteteacute isoleacutee dans de rares occasions sur drsquoautres espegraveces de ruminants tels que les gazelles les
antilopes (Van Soolingen et al 1994) les buffles africains (van Pittius et al 2012) en Afrique du sud
dans la Peacuteninsule arabe et dans des zoos des Pays Bas et du Royaume Uni
Des infections agrave Mycobacterium orygis ont eacuteteacute rapporteacutees chez une dizaine de patients humains
(Dawson et al 2012 van Ingen et al 2012)
Chapitre 1
20
Autres mycobacteacuteries agrave reacuteservoir animal
Quatre autres espegraveces de mycobacteacuteries animales phylogeacuteneacutetiquement proches de
M africanum West African 2 ont eacuteteacute isoleacutees sur des animaux en Afrique le Chimpazee bacillus isoleacute
sur un chimpanzeacute de Cocircte drsquoIvoire (Coscolla et al 2013) M mungi isoleacutee sur des mangoustes rayeacutees
(Mungos mungo) du Bostwana (Alexander et al 2010) le Dassie bacillus isoleacute sur un daman du Cap
(Procavia capensis) (Wagner et al 1958) et M suricattae isoleacutee sur des suricates (Suricata suricatta) en
Afrique du sud (Parsons et al 2013) Ces espegraveces nrsquoont agrave ce jour jamais eacuteteacute isoleacutees chez lrsquohomme La
deacutecouverte reacutecente de ces espegraveces rattacheacutees au MTBC suggeacutere que ce Complexe est probablement
plus varieacute que ce qui eacutetait imagineacute
B LA TUBERCULOSE UNE MALADIE TRES ANCIENNE MAIS UNE MENACE TOUJOURS ACTUELLE
1 Introduction
La tuberculose est une maladie bacteacuterienne chronique et contagieuse qui affecte lrsquohomme
ainsi que de nombreux animaux domestiques et sauvages Elle est provoqueacutee par les diffeacuterentes
espegraveces de mycobacteacuteries du MTBC Lrsquoobservation de tubercules granulomes inflammatoires formeacutes
de cellules immunitaires et de caseacuteum (zones neacutecrotiques) dans les organes ou les nœuds
lymphatiques des personnes infecteacutees a donneacute son nom agrave la maladie
2 Histoire de la tuberculose
La tuberculose est une maladie tregraves ancienne puisque lrsquoancecirctre du Complexe tuberculosis serait
apparu il y a 40 000 ans (Wirth et al 2008) La preacutesence exclusive de certaines ligneacutees du MTBC sur le
continent africain suggegravere que le Complexe aurait eacutemergeacute en Afrique de lrsquoEst puis aurait co-eacutevolueacute
avec lrsquohomme et se serait reacutepandu dans le monde entier par le biais des migrations humaines en
Europe en Asie et en Afrique continentale Les populations de mycobacteacuteries tuberculeuses eacutetant
infeacuteodeacutees agrave une zone geacuteographique elles se sont adapteacutees agrave la population locale (Gutierrez et al 2005
Djelouadji et al 2011 Wirth et al 2008) Le passage drsquoun mode de vie nomade agrave une vie seacutedentaire
dans des fermes agricoles de plus en plus peupleacutees ainsi que la domestication des animaux ont
probablement contribueacute agrave la transmission de la maladie agrave divers hocirctes animaux et agrave lrsquoadaptation du
bacille permettant lrsquoapparition de M bovis M caprae et des autres membres du Complexe (Wirth et
al 2008) Les plus anciennes traces drsquoADN du Complexe tuberculosis ont eacuteteacute deacutecouvertes sur des
Chapitre 1
21
ossements de bison vieux de 17 000 ans retrouveacutes dans une grotte drsquoAmeacuterique du Nord (Rothschild et
al 2001) Des traces drsquoADN du Complexe tuberculosis de M tuberculosis ou M bovis ont eacuteteacute
retrouveacutees sur des ossements humains datant de la Preacutehistoire ou de lrsquoAntiquiteacute (les plus vieux ont
9000 ans) deacutecouverts dans diffeacuterentes parties du monde (Chili Royaume Uni Russie Israeumll Indoneacutesie
etc) (Djelouadji et al 2011) Appeleacutee laquo phtisie raquo par les Grecs peste blanche ou consomption (suite au
deacutepeacuterissement des malades) les eacutecrits drsquoHippocrate (5egraveme siegravecle avant JC) sont les premiers agrave deacutecrire la
symptomatologie et les diffeacuterentes formes de la tuberculose Fracastor (15egraveme siegravecle) suggegravere le
caractegravere heacutereacuteditaire de la maladie mais place la phtisie dans les maladies contagieuses au mecircme titre
que la veacuterole hypothegravese reprise en 1722 par Marten puis par Villemin en 1865 qui montre que la
maladie est infectieuse contagieuse et inoculable (Dutau 2005) A la fin du 18egraveme siegravecle et au deacutebut du
19egraveme la tuberculose est la premiegravere cause de mortaliteacute en Europe de lrsquoOuest et en Ameacuterique du Nord
La Reacutevolution industrielle et le deacuteveloppement des villes favorisent lrsquoexpansion de la maladie les
conditions de vie insalubres des taudis de banlieues la promiscuiteacute et la pauvreteacute acceacutelegraverent lrsquoendeacutemie
et agrave cette eacutepoque environ une personne sur 4 est atteinte de tuberculose La fin du 19egraveme siegravecle voit se
succeacuteder les deacutecouvertes agrave un rythme effreacuteneacute dont les principales sont la deacutecouverte du bacille en
1882 par Koch la deacutecouverte du bacille bovin en 1896 lrsquoutilisation de la tuberculine dans le diagnostic
humain en 1907 la creacuteation drsquoune souche atteacutenueacutee M bovis BCG en 1921 et le deacutebut de la vaccination
en 1924 La deacutecouverte de la streptomycine en 1944 le premier antibiotique contre la tuberculose
puis de lrsquoisoniazide en 1952 et la rifamycine en 1957 a marqueacute une eacutetape importante dans le traitement
contre la tuberculose (Daniel 2006)
3 La tuberculose bovine
a Introduction
La tuberculose bovine est une maladie bacteacuterienne contagieuse drsquoeacutevolution chronique causeacutee
par Mycobacterium bovis Cette bacteacuterie a pour hocircte originel les bovins mais peut infecter un grand
nombre drsquoespegraveces de mammifegraveres dont lrsquohomme (Gavier-Wideacuten et al 2009 Broughan Crawshaw et
al 2013 Grange amp Yates 1994) ce qui en fait une maladie zoonotique Elle a un tropisme respiratoire
preacutedominant mais peut infecter un grand nombre drsquoautres organes La tuberculose bovine fait partie
des maladies animales agrave deacuteclaration obligatoire aupreacutes de lrsquoOffice International des Epizooties (OIE)
Le passage de lrsquoinfection dans la faune sauvage a compliqueacute le scheacutema drsquoeacuteradication de la
maladie chez les bovins puisque certaines espegraveces sauvages peuvent entretenir la maladie et
contaminer les bovins en retour A lrsquoheure actuelle M bovis fait partie des six organismes pathogegravenes
prioritaires cibleacutes par les programmes de surveillance europeacuteens (Ciliberti et al 2015)
Chapitre 1
22
b Les hocirctes et les communauteacutes drsquohocirctes de Mycobacterium bovis dans le monde
et leurs statuts eacutepideacutemiologiques
Mycobacterium bovis est la mycobacteacuterie du MTBC qui possegravede le plus large spectre drsquohocirctes
incluant lrsquohomme (Grange 2001 OrsquoReilly amp Daborn 1995) Lrsquoinfection a eacuteteacute retrouveacutee chez de
nombreuses espegraveces de mammifegraveres appartenant aux Ordres des marsupiaux des carnivores des
primates des rongeurs des lagomorphes et des artiodactyles (Broughan Downs et al 2013 OrsquoReilly
amp Daborn 1995 Palmer 2013)
La circulation de lrsquoinfection chez des hocirctes appartenant agrave des espegraveces domestiques et sauvages
diffeacuterentes est agrave lrsquoorigine de systegravemes eacutepideacutemiologiques complexes appeleacutes systegraveme multi-hocirctes
(Haydon et al 2002) qui complexifient fortement les scheacutemas drsquoeacuteradication de la maladie Dans ces
systegravemes les diffeacuterents hocirctes peuvent avoir un statut eacutepideacutemiologique diffeacuterent (Haydon et al 2002
Palmer 2013) ou former une communauteacute drsquohocirctes reacuteservoir (Palmer 2013) Les hocirctes de maintenance
ou reacuteservoirs deacutefinissent une population drsquoanimaux dans laquelle lrsquoinfection est entretenue de faccedilon
autonome Ils sont agrave lrsquoorigine de lrsquoinfection des espegraveces cibles Les hocirctes de liaison sont incapables de
maintenir lrsquoinfection indeacutefiniment au sein de leur population agrave moins qursquoils soient reacuteinfecteacutes par une
autre espegravece Ils sont toutefois capables de transmettre lrsquoinfection agrave drsquoautres populations (Corner 2006
Nugent 2011) Et enfin les hocirctes laquo cul-de-sac raquo deacutefinissent les espegraveces qui ne peuvent pas maintenir
ni transmettre la maladie (Nugent 2011) Diffeacuterentes espegraveces ont ainsi eacuteteacute identifieacutees comme des
reacuteservoirs de lrsquoinfection notamment le blaireau (Meles meles) en Angleterre et en Irlande (Delahay et
al 2001) le sanglier (Sus scrofa) dans le centre et le sud de lrsquoEspagne (Naranjo et al 2008) le cerf
eacutelaphe en France (Zanella et al 2008) le cerf de Virginie (Odocoileus virginianus) dans le Michigan
(USA) (Palmer 2013 OrsquoBrien et al 2006) le bison des bois (Bison bison athabascae) et le wapiti (Cervus
elaphus manitobensis) au Canada (Nishi et al 2006 Wobeser 2009) le phalanger renard (Trichosurus
vulpecula) en Nouvelle-Zeacutelande (Morris et al 1994 Nugent 2011) et le buffle (Syncerus caffer) en
Afrique du Sud (Renwick et al 2007) Drsquoautres espegraveces animales ont eacuteteacute identifieacutees comme hocirctes de
liaison comme le furet le cerf et le sanglier en Nouvelle-Zeacutelande
La transmission retour de lrsquoinfection par les espegraveces sauvages reacuteservoir est une menace pour
les bovins et les espegraveces sauvages sensibles vivant agrave proximiteacute La connaissance du statut
eacutepideacutemiologique de chaque espegravece est important pour comprendre le rocircle de la faune sauvage dans la
maintenance et la transmission de lrsquoinfection et adapter les programmes de lutte et drsquoeacuteradication de la
maladie chez les bovins les animaux sauvages et lrsquohomme (Delahay et al 2001 Corner 2006)
Chapitre 1
23
c Aspect zoonotique de Mycobacterium bovis
Plusieurs mycobacteacuteries du MTBC peuvent engendrer une tuberculose zoonotique chez
lrsquohomme Le principal agent infectieux impliqueacute est M bovis (Thoen et al 2006) dans une moindre
mesure M caprae (EFSA 2014) et M microti (Panteix et al 2010) Cette derniegravere a un pouvoir
zoonotique plus faible mais nrsquoest pas neacutegligeable La tuberculose zoonotique agrave M bovis est
majoritairement lieacutee agrave la consommation de produits issus drsquoanimaux infecteacutes principalement le lait cru
et ses deacuteriveacutes (transmission indirecte) (Evans et al 2007) La transmission par aeacuterosols respiratoires est
eacutegalement deacutecrite lors de contacts rapprocheacutes avec des animaux domestiques ou sauvages infecteacutes
(transmission directe) (Cosivi et al 1999 Ayele et al 2004 Kleeberg 1984) Lrsquoinfection est
sporadiquement rapporteacutee chez des eacuteleveurs de bovins et de cerfs (Smith et al 2004 Fanning amp
Edwards 1991) du personnel veacuteteacuterinaire (Shrikrishna et al 2009) et du personnel drsquoabattoir (Liss et al
1994) La transmission interhumaine de M bovis par voie respiratoire existe chez les patients
immunodeacuteprimeacutes ou immunocompeacutetents mais reste rare (Evans et al 2007 Grange amp Yates 1994 de
la Rua-Domenech 2006 Sunder et al 2009 Shrikrishna et al 2009 Rivero et al 2001)
Les signes cliniques chez lrsquohomme sont identiques agrave ceux retrouveacutes lors de tuberculose agrave
M tuberculosis et se traduisent par une tuberculose pulmonaire etou des atteintes extra-pulmonaires
plus freacutequentes qursquoavec M tuberculosis en raison des diffeacuterences dans le mode de transmission
(atteintes osteacuteo-articulaires ganglionnaires uro-geacutenitales disseacutemineacuteeshellip)(Antoine amp Jarlier 2010
Delaune et al 2012 Mignard et al 2006) Le traitement repose sur lrsquoadministration drsquoantituberculeux
classiques agrave lrsquoexception du pyrazinamide pour lequel M bovis est naturellement reacutesistant Des souches
MDR (Multi Drug Resistant) sont reacuteguliegraverement signaleacutees chez lrsquohomme (McLaughlin et al 2012
Cambau et al 2003) De rares souches XDR (Extensively Drug Resistant) sont eacutegalement deacutecrites
majoritairement chez des patients seacuteropositifs (Rivero et al 2001) Dans les pays industrialiseacutes
lrsquoinfection est principalement diagnostiqueacutee chez des personnes acircgeacutees etou immunodeacuteprimeacutees
(diabegravete VIH traitement immunosuppresseur alcoolismehellip) dont les formes extra-pulmonaires
majoritaires eacutevoquent une contamination ancienne par voie digestive (avant la mise en place de la
pasteurisation du lait) et la reacuteactivation ulteacuterieure de la maladie ainsi que chez des personnes neacutees ou
ayant seacutejourneacute dans des pays ougrave la maladie est endeacutemique (Delaune et al 2012)
Les pays ayant mis en place la pasteurisation du lait et des programmes de lutte efficaces contre
la maladie chez les bovins (deacutepistage et abattage) ont fortement reacuteduit lrsquoincidence de la maladie chez
lrsquohomme et les bovins Malgreacute quelques cas reacuteguliegraverement deacutecrits chez lrsquohomme la tuberculose
zoonotique est une zoonose mineure dans les pays deacuteveloppeacutes notamment en Europe (Torgerson amp
Torgerson 2010 Michel et al 2010) En France lrsquoobligation de pasteurisation du lait en 1955 associeacutee
agrave lrsquoabattage systeacutematique des bovins reacuteagissant agrave la tuberculination agrave partir de 1963 ont permis de faire
Chapitre 1
24
baisser lrsquoincidence de lrsquoinfection agrave M bovis chez lrsquohomme de 15 dans les anneacutees 60 agrave 05 en 1995
(Boulahbal et al 1998) Des eacutetudes locales mettent parfois en eacutevidence des preacutevalences plus eacuteleveacutees
de lrsquoordre de 2 agrave Lyon entre 2000 et 2005 (Mignard et al 2006) et de 29 en Aquitaine entre 2004
et 2008 (Aimeacute et al 2012) Au Royaume-Uni par exemple malgreacute lrsquoincidence eacuteleveacutee chez les bovins
(1806 nouveaux foyers entre janvier et avril 2015)(site DEFRA) la preacutevalence de la maladie nrsquoaugmente
pas chez lrsquohomme gracircce aux programmes de lutte mis en place chez lrsquoanimal (Torgerson amp Torgerson
2010 Jalava et al 2007)
La situation est tregraves diffeacuterente dans les pays en voie de deacuteveloppement (PVD) Lrsquoabsence de
programme de lutte contre la tuberculose bovine et la consommation de lait cru dans des pays ougrave la
maladie est endeacutemique chez les bovins notamment dans certains pays drsquoAfrique drsquoAmeacuterique du sud
et drsquoAsie repreacutesentent un risque de santeacute publique important (Cosivi et al 1999 Ayele et al 2004) La
tuberculose zoonotique est souvent fortement relieacutee agrave la preacutevalence de lrsquoinfection bovine dans le pays
ou la reacutegion (Fetene et al 2011) et touche majoritairement les professions en contact avec les bovins
(eacuteleveurs de bovins personnel drsquoabattoir et drsquousines agro-alimentairehellip) et les consommateurs de lait
cru et de fromage frais dont les enfants drsquoautant plus srsquoils sont immunodeacuteprimeacutes Une eacutetude reacutecente
rapporte que la tuberculose zoonotique repreacutesente environ 28 des cas de tuberculose en Afrique
(de 0 agrave 377 selon les pays) 03 des cas sur le continent ameacutericain (de 0 agrave 339 ) et 04 des cas
en Europe (de 0 agrave 211 ) (Muumlller et al 2013) Lrsquoinfection par le HIV la pauvreteacute et le manque drsquoaccegraves
aux soins sont des facteurs de disseacutemination de la tuberculose humaine et zoonotique dans ces pays
(Ayele et al 2004 WHO 2014) Dans les PVD le diagnostic chez lrsquohomme srsquoarrecircte parfois agrave lrsquoobservation
microscopique de BAAR associeacutes agrave M tuberculosis sous-estimant probablement lrsquooccurrence de
M bovis (Cosivi et al 1999 Michel et al 2010) La transmission retour de lrsquoinfection de lrsquohomme aux
bovins pourrait avoir un impact non neacutegligeable dans les PVD mais aucune donneacutee nrsquoest disponible
(de la Rua-Domenech 2006)
d Pathogeacutenie de la tuberculose bovine
La pathogeacutenie de la maladie est multifactorielle et deacutepend de facteurs lieacutes au pouvoir pathogegravene de
M bovis agrave la reacuteceptiviteacute et la sensibiliteacute de lrsquohocircte exposeacute ainsi qursquoagrave la voie drsquoinfection
Facteurs influenccedilant le pouvoir pathogegravene de M bovis voies
drsquoinfection et dose infectante
La voie respiratoire est la voie de contamination majeure chez les bovins (Pritchard 1988
Morris et al 1994 Phillips et al 2003) Des infections expeacuterimentales de M bovis par voie nasale intra
Chapitre 1
25
tracheacuteale ou intra-tonsillaire ainsi que par la mise en contact drsquoanimaux sains avec des animaux
infecteacutes ont confirmeacute lrsquoefficaciteacute de la transmission de M bovis par cette voie (Palmer et al 1999 Dean
et al 2005 Johnson et al 2007 Collins amp Grange 1983 Cassidy et al 1999) La dose infectante par voie
respiratoire est tregraves faible puisque lrsquoinoculation intra-tracheacuteale de 6 agrave 10 bacilles viables agrave des veaux
suffit agrave provoquer des leacutesions pulmonaires etou ganglionnaires (Dean et al 2005 Johnson et al 2007)
Des reacutesultats identiques ont eacuteteacute obtenus chez le blaireau (Corner et al 2007 Corner et al 2011) Chez
le cerf de Virginie lrsquoinoculation intra-tonsillaire de 300 agrave 500 UFC (Uniteacute Formant Colonie) infecte 100
des animaux (Palmer et al 2002 Griffin et al 2006) Lrsquoinoculation oro-pharyngeacutee de 100 UFC agrave des
sangliers deacuteclenche uniquement des leacutesions amygdaliennes alors que 10 000 UFC entrainent des
leacutesions pulmonaires et ganglionnaires seacutevegraveres (Ballesteros et al 2009) Chez toutes ces espegraveces la dose
infectante affecte la seacuteveacuteriteacute et la progression de la maladie (Neill et al 1994 Palmer et al 2002 Corner
et al 2007)
La transmission de la tuberculose bovine peut eacutegalement se faire par voie digestive suite agrave
lrsquoingestion de matrices contamineacutees La dose infectante neacutecessaire serait bien supeacuterieure agrave celle par
voie respiratoire et selon certains auteurs neacutecessiterait plusieurs millions de bacilles (Phillips et al
2003 OrsquoReilly amp Daborn 1995) Pour les veaux Palmer et al (2004) ont estimeacute agrave 5 000 UFC la dose
infectante par cette voie Lrsquoingestion reacutepeacutetitive de faibles doses de M bovis semble eacutegalement
repreacutesenter un facteur de risque comme cela a eacuteteacute deacutemontreacute chez le cochon drsquoInde (Maddock 1934)
La transmission par voie cutaneacutee ou transcutaneacutee ainsi que la transmission de la megravere au veau par
consommation de lait contamineacute ou par leacutechage sont eacutegalement deacutecrites mais restent rares (Phillips et
al 2003 Morris et al 1994) La transmission du bacille par voie transcutaneacutee (morsure) par le biais de
salive infecteacutee est reacuteguliegraverement deacutecrite chez le blaireau en Angleterre (Corner et al 2011)
Facteurs influenccedilant la reacuteceptiviteacute et la sensibiliteacute de lrsquohocircte
De nombreux facteurs intrinsegraveques agrave lrsquoanimal entrent en jeu dans la reacuteceptiviteacute et la sensibiliteacute
de lrsquohocircte agrave la maladie notamment lrsquoespegravece la race lrsquoacircge le sexe et lrsquoeacutetat geacuteneacuteral (Griffin et al 1996
Menzies amp Neill 2000 Humblet et al 2009 Goodchild amp Clifton-Hadley 2001) Certains animaux
preacutesentent une reacutesistance naturelle ou au contraire une sensibiliteacute exacerbeacutee agrave lrsquoinfection ou agrave la
progression de la maladie (Gavier-Wideacuten et al 2009) Des meacutecanismes modulant la reacuteponse
immunitaire et inflammatoire de lrsquohocircte ont pu ecirctre mis en eacutevidence chez le cerf (Mackintosh et al
2000) Les caracteacuteristiques geacuteneacutetiques de lrsquohocircte pourraient eacutegalement jouer un rocircle important dans la
reacutesistance ou la sensibiliteacute agrave lrsquoinfection (Mackintosh et al 2000 Acevedo-Whitehouse et al 2005
Brotherstone et al 2010) Des eacutetudes reacutecentes sur des geacutenomes complets de bovins ont identifieacute des
Chapitre 1
26
loci particuliers impliqueacutes dans la reacutesistance agrave la maladie (Bermingham et al 2014) ouvrant des
perspectives de seacutelection geacutenomique de populations reacutesistantes agrave la tuberculose bovine (Tsairidou et
al 2014)
Le statut immunitaire de lrsquoanimal est un facteur important puisque certains animaux sont
capables drsquoeacuteliminer le bacille gracircce agrave leur immuniteacute inneacutee etou acquise certains sont capables de
controcircler lrsquoinfection plusieurs anneacutees dans les granulomes et drsquoautres deacuteveloppent une tuberculose
eacutevolutive (Pollock amp Neill 2002) La modulation immunitaire expliquerait pourquoi dans un cheptel
bovin malgreacute la contagiositeacute importante de M bovis seuls quelques animaux sont infecteacutes (Cassidy
2006) Les notions de latence et de reacuteactivation de la maladie deacutecrites chez M tuberculosis sont
discuteacutees dans la tuberculose bovine (Cassidy 2006) et commencent agrave ecirctre appreacutehendeacutees (Aacutelvarez et
al 2009)
Les eacutetapes de lrsquoinfection et la reacuteponse immunitaire associeacutee
Lorsque les bacteacuteries peacutenegravetrent dans lrsquoorganisme le plus souvent au niveau des alveacuteoles
pulmonaires elles sont phagocyteacutees par les macrophages Si la dose infectante est importante ou
lrsquoimmuniteacute insuffisante les mycobacteacuteries eacutechappent agrave la digestion et se multiplient permettant la
formation dans le tissu pulmonaire de la leacutesion primaire appeleacutee chancre drsquoinoculation La primo-
infection est caracteacuteriseacutee par le laquo complexe primaire raquo qui associe le chancre drsquoinoculation et les nœuds
lymphatiques infecteacutes par le drainage de la leacutesion primaire (Neill et al 1994) Lrsquoinfection peut ensuite
eacutevoluer selon 3 modes diffeacuterents la gueacuterison avec une eacutelimination des bacilles la stabilisation
caracteacuteriseacutee par une reacuteactivation possible de lrsquoinfection quelques mois ou anneacutees plus tard ou la
geacuteneacuteralisation preacutecoce de lrsquoinfection lieacutee agrave une multiplication active des bacteacuteries et agrave leur
disseacutemination dans lrsquoorganisme Lrsquoinfection et son eacutevolution sont toutefois fortement deacutependantes de
la dose infectante de la voie drsquoentreacutee de la bacteacuterie de la reacuteceptiviteacute et de la sensibiliteacute de lrsquohocircte
eacutevoqueacutees preacuteceacutedemment
Chez lrsquoanimal infecteacute par M bovis la reacuteponse immunitaire agrave meacutediation cellulaire est la premiegravere
agrave se mettre en place dans les stades preacutecoces de lrsquoinfection par la mobilisation de macrophages et de
lymphocytes T (Figure 5) Ces derniers libegraverent diffeacuterentes cytokines dont lrsquointerfeacuteron-γ (IFN-γ) qui
active les macrophages et la formation de granulomes pour isoler et deacutetruire lrsquoagent infectieux
Lrsquoimmuniteacute agrave meacutediation humorale se met en place tardivement et remplace peu agrave peu lrsquoimmuniteacute agrave
meacutediation cellulaire dans les stades plus avanceacutes de la maladie par la production drsquoanticorps par les
lymphocytes B (Pritchard 1988 Ritacco et al 1991 Fifis et al 1994) Les diffeacuterents tests utiliseacutes pour
le deacutepistage de lrsquoinfection chez les bovins reposent principalement sur lrsquoimmuniteacute agrave meacutediation
cellulaire (Figure 5)
Chapitre 1
27
Figure 5 Evolution de la reacuteponse immunitaire chez le bovin infecteacute par M bovis au cours du temps et aux diffeacuterents tests de deacutepistage de la tuberculose bovine (de la Rua-Domenech et al 2006 adapteacute de Vordermeier et al 2004)
Expression clinique de la maladie et leacutesions observeacutees
La tuberculose bovine est une maladie chronique drsquoeacutevolution lente geacuteneacuteralement
asymptomatique dans ses deacutebuts Lorsque la maladie eacutevolue hormis lrsquoamaigrissement possible les
signes cliniques sont tregraves variables et deacutependent des organes ou tissus atteints mais ne sont pas
pathognomoniques (de la Rua-Domenech et al 2006) La localisation des leacutesions observeacutees chez les
animaux deacutepend geacuteneacuteralement de la voie drsquoentreacutee de la bacteacuterie et de lrsquoeacutevolution de la maladie Chez
les bovins la majoriteacute des leacutesions sont retrouveacutees dans la caviteacute thoracique avec une atteinte des lobes
pulmonaires et des nœuds lymphatiques associeacutes (bronchiques et meacutediastinaux) etou des nœuds
lymphatiques ceacutephaliques (reacutetro-pharyngiens principalement) Les leacutesions sur les nœuds lymphatiques
meacutesenteacuteriques la rate et le foie sont plus rares (Neill et al 1994)
Chez le blaireau infecteacute naturellement la seacuteveacuteriteacute de la maladie est tregraves variable drsquoune
infection latente asymptomatique sans leacutesion macroscopique visible agrave une maladie modeacutereacutee avec des
leacutesions pulmonaires ou extra-pulmonaires de petite taille jusqursquoagrave une maladie geacuteneacuteraliseacutee entrainant
la cachexie puis la mort de lrsquoanimal (Murphy et al 2010) Les leacutesions retrouveacutees sont essentiellement
respiratoires probablement lieacutees agrave lrsquoinhalation drsquoaeacuterosols contamineacutes plus rarement digestives et
cutaneacutees suite agrave des morsures de congeacutenegraveres infecteacutes (Corner et al 2011 Payne et al 2012 Gavier-
Widen et al 2001) Toutefois diffeacuterentes eacutetudes ont montreacute que la majoriteacute des blaireaux infecteacutes ne
Chapitre 1
28
preacutesente pas de leacutesion visible (Gallagher amp Clifton-Hadley 2000 Jenkins et al 2008 Payne et al 2012)
Les blaireaux constituent des hocirctes de persitance efficaces puisqursquoils peuvent supporter lrsquoinfection
plusieurs anneacutees et se reproduire tout en excreacutetant la bacteacuterie (Delahay et al 2001)
Chez le sanglier les leacutesions sont majoritairement observeacutees dans les nœuds lymphatiques
ceacutephaliques notamment les mandibulaires dans les poumons et moins freacutequemment au niveau
meacutesenteacuterique (Zanella et al 2008 Martiacuten-Hernando et al 2007 Naranjo et al 2008) Des formes
geacuteneacuteraliseacutees sont eacutegalement deacutecrites (Naranjo et al 2008)
Le cerf est une espegravece deacutecrite comme tregraves sensible agrave lrsquoinfection par M bovis et deacuteveloppe des
leacutesions ouvertes et disseacutemineacutees pouvant ecirctre agrave lrsquoorigine drsquoune excreacutetion bacillaire importante (Lugton
et al 1998 Palmer et al 2004) Les leacutesions sont geacuteneacuteralement localiseacutees au niveau des nœuds
lymphatiques ceacutephaliques et des poumons (Palmer et al 2004 Delahay et al 2002) des nœuds
lymphatiques meacutesenteacuteriques voire geacuteneacuteraliseacutees (Zanella et al 2008)
e Le deacutepistage et le diagnostic de lrsquoinfection chez les animaux
Le deacutepistage de lrsquoinfection chez les bovins vivants repose sur plusieurs modes de surveillance
(Cavalerie et al 2014)
La surveillance systeacutematique agrave lrsquoabattoir concerne tous les bovins abattus pour la consommation
humaine Il repose sur un examen post mortem de lrsquoanimal avec lrsquoinspection drsquoun certain nombre
drsquoorganes cibles de lrsquoinfection (poumons nœuds lymphatiques reacutetro-pharyngiens tracheacuteo-
bronchiques et meacutediastinaux) Dans le cas de leacutesions suspectes visibles celles-ci sont preacuteleveacutees et
transmises agrave un laboratoire agreacuteeacute pour recherche de mycobacteacuteries par PCR et culture
La surveillance programmeacutee en eacutelevage repose sur le deacutepistage de la maladie par prophylaxie Le
rythme des prophylaxies est fixeacute par le preacutefet en fonction de la situation sanitaire du deacutepartement
Dans les deacutepartements ougrave cette surveillance est absente le deacutepistage deacutepend alors uniquement de la
surveillance agrave lrsquoabattoir Pour quelques deacutepartements agrave fort risque elle est annuelle et peut ecirctre
compleacuteteacutee par un deacutepistage lors de mouvements drsquoanimaux Le test de deacutepistage de reacutefeacuterence est
lrsquointradermotuberculination (IDR) Ce test vise agrave mettre en eacutevidence in vivo la reacuteaction
drsquohypersensibiliteacute retardeacutee conseacutecutive agrave lrsquoinjection intradermique de tuberculine (proteacuteines purifieacutees
de surnageant de culture de mycobacteacuteries) chez un bovin infecteacute (Buddle et al 2011 De la Rua-
Domenech et al 2006) Cette reacuteaction se traduit par une tumeacutefaction cutaneacutee douloureuse et chaude
Deux types drsquoIDR sont utiliseacutes lrsquoIntradermotuberculination simple ou IDS qui consiste en lrsquoinjection
intradermique de tuberculine bovine sur le plat de lrsquoencolure et lrsquoIntradermotuberculination
comparative ou IDC qui associe lrsquoinjection intradermique de tuberculine bovine et de tuberculine
aviaire LrsquoIDC est utiliseacutee dans certains deacutepartements dont la Cocircte drsquoOr ougrave de nombreuses
Chapitre 1
29
mycobacteacuteries environnementales sont retrouveacutees pour augmenter la speacutecificiteacute du test et eacutecarter les
reacuteactions croiseacutees chez les bovins sensibiliseacutes agrave une autre mycobacteacuterie (des antigegravenes sont communs
entre les mycobacteacuteries) (De la Rua-Domenech et al 2006 OIE 2009)
LrsquoIDR peut ecirctre compleacuteteacutee par le dosage de lrsquoInterfeacuteron γ (IFNγ) Cette cytokine qui est libeacutereacutee
par les lymphocytes T activeacutes lors drsquoune stimulation antigeacutenique in vivo mais eacutegalement in vitro permet
entre autre lrsquoactivation de la fonction phagocytaire des macrophages La quantiteacute drsquoIFNγ libeacutereacutee lors de
la culture de sang total avec de la tuberculine bovine ou aviaire est suffisante pour ecirctre mesureacutee par
un test ELISA (Gormley et al 2006 de la Rua-Domenech et al 2006) Le dosage de lrsquoIFNγ est pratiqueacute
pour renforcer lrsquoIDR lorsque la contention des animaux est difficile essentiellement en Camargue et
dans les Landes pour les taureaux de combat Il est eacutegalement utiliseacute agrave titre expeacuterimental dans quelques
deacutepartements comme la Dordogne et la Cocircte drsquoOr (Fediaevsky et al 2014) pour renforcer les reacutesultats
drsquoIDR
Drsquoautres tests notamment seacuterologiques sont en cours drsquoeacutevaluation chez les bovins Les essais
reacutealiseacutes jusqursquoagrave maintenant montrent une sensibiliteacute et une speacutecificiteacute infeacuterieures agrave lrsquoIDR ou lrsquoIFNγ
notamment chez les animaux en deacutebut drsquoinfection mais ils repreacutesentent un axe de recherche important
au vu du faible coucirct et de la simpliciteacute de la technique (Waters et al 2011 de la Rua-Domenech et al
2006)
La seacuterologie et lrsquoIFNγ font lrsquoobjet de nombreux deacuteveloppements pour lrsquoameacutelioration de la
surveillance de la maladie chez les animaux sauvages Reacutecemment des techniques ELISA ont eacuteteacute
deacuteveloppeacutees pour de nombreuses espegraveces dont les sangliers les cerfs les bisons (Boadella et al 2011
Shuralev et al 2012 Boadella et al 2012 Lyashchenko et al 2013 Chapinal et al 2012) LrsquoIFNγ est
eacutegalement testeacute pour le deacutepistage chez les espegraveces sauvages notamment chez les cervideacutes drsquoeacutelevage
et les blaireaux (Palmer et al 2010 Dalley et al 2008) La seacuterologie (kit commercial Brock Stat-Pack
Chembio Diagnostic Systems) et le dosage de lrsquoIFNγ (meacutethode ELISA) sont utiliseacutes en routine apregraves
capture et preacutelegravevement sanguin pour le suivi eacutepideacutemiologique de groupes sociaux de blaireaux agrave
Woodchester Park en Angleterre (Drewe et al 2010 King Murphy James Travis Porter Sawyer et al
2015 King Murphy James Travis Porter Hung et al 2015) La combinaison des deux tests est
conseilleacutee pour ameacuteliorer la sensibiliteacute et la speacutecificiteacute du deacutepistage (Drewe et al 2010) Une meacutethode
de deacutepistage non invasive baseacutee sur la collecte puis lrsquoanalyse par qPCR des latrines a eacuteteacute eacutevalueacutee sur
douze groupes sociaux en parallegravele avec la seacuterologie et le dosage de lrsquoIFNγ (King Murphy James Travis
Porter Sawyer et al 2015 King Murphy James Travis Porter Hung et al 2015) Bien que moins
contraignante et moins couteuse que le suivi par capture lrsquoanalyse des latrines pour lrsquoidentification des
groupes sociaux de blaireaux positifs neacutecessite au moins deux campagnes annuelles de preacutelegravevements
(deacutebut et fin drsquoeacuteteacute) avec un nombre drsquoeacutechantillons minimum variable selon les groupes sociaux (de 10
agrave plus de 20) agrave cause de lrsquoexcreacutetion intermittente du bacille par les blaireaux infecteacutes De plus elle ne
Chapitre 1
30
permet pas un deacutepistage individuel
Concernant le diagnostic de la tuberculose bovine lrsquoinfection chez les bovins par M bovis doit
ecirctre eacutetablie par histologie en cas de leacutesion et confirmeacutee par culture selon la reacuteglementation
europeacuteenne Lrsquohistologie est reacutealiseacutee sur des coupes de nœuds lymphatiques et de leacutesions fixeacutees agrave la
formaline La culture repose sur le protocole deacuteveloppeacute et ameacutelioreacute par le Laboratoire National de
Reacutefeacuterence (LNR) devenu la norme NF U47-104 qui associe une phase de broyage-deacutecontamination
des eacutechantillons puis un ensemencement sur deux milieux agrave lrsquoœuf (Loumlwenstein-Jensen et Coletsos) (OIE
2009) Toutefois ces deux techniques preacutesentent des inconveacutenients car lrsquohistologie manque de
speacutecificiteacute (environ 92 ) par rapport agrave la culture (Varello et al 2008) et le reacutesultat de culture est peu
sensible long (entre trois semaines et trois mois) ce qui pose des problegravemes pratiques en cas de
suspicion (restriction des mouvements du cheptel) Bien que pratiqueacutee depuis quelques anneacutees sans
base regraveglementaire il a eacuteteacute deacutemontreacute que la PCR baseacutee sur la deacutetection de lrsquoIS6110 (kit commercial LSI
VetMAXtrade Mycobacterium tuberculosis Complex PCR kit) serait un outil performant de diagnostic de la
tuberculose chez les bovins avec une bonne speacutecificiteacute (97 ) et une sensibiliteacute supeacuterieure agrave la culture
(877 vs 781 ) (Courcoul et al 2014) En apportant un diagnostic en 48 heures la PCR permettrait
une gestion plus rapide des foyers
Le diagnostic dans la faune sauvage repose sur une PCR systeacutematique sur les leacutesions eacutevocatrices
de tuberculose ou un pool de nœuds lymphatiques en lrsquoabsence de leacutesions En cas de reacutesultat non
neacutegatif lrsquoeacutechantillon est mis en culture pour identification de lrsquoagent infectieux (Source Note de service
DGALSDSPA2015-556 du 26062015) Quelle que soit lrsquoespegravece concerneacutee la culture et la PCR sont
reacutealiseacutees par des laboratoires agreacuteeacutes En cas de PCR etou de culture positive les eacutechantillons sont
systeacutematiquement envoyeacutes au Laboratoire National de Reacutefeacuterence pour la tuberculose bovine pour
confirmation et geacutenotypage des souches
f Epideacutemiologie de la tuberculose bovine
Evolution de la tuberculose bovine dans le monde et en
Europe et mesures de lutte
Avant 1920 la tuberculose bovine eacutetait lrsquoune des maladies majeures des animaux domestiques
agrave lrsquoeacutechelle mondiale A lrsquoheure actuelle elle est toujours largement reacutepandue dans le monde (Cosivi et
al 1999) La maladie est plutocirct sporadique et limiteacutee agrave quelques reacutegions dans les pays industrialiseacutes
toucheacutes (sauf en Irlande au Royaume-Uni en Espagne et au Portugal) et endeacutemique dans les pays agrave
faibles revenus drsquoAfrique drsquoAmeacuterique du sud et drsquoAsie (Figure 6)
Chapitre 1
31
Dans les pays industrialiseacutes la mise en place de la pasteurisation du lait dans les anneacutees 1950-
60 et des programmes drsquoeacuteradication de la maladie baseacutes sur le deacutepistage et lrsquoabattage des bovins
infecteacutes ont permis de reacuteduire fortement lrsquoincidence de lrsquoinfection chez les bovins et lrsquohomme
Aujourdrsquohui dans ces pays la tuberculose bovine repreacutesente principalement un enjeu socio-
eacuteconomique lieacute aux restrictions des eacutechanges internationaux drsquoanimaux et aux coucircts des programmes
de lutte contre la maladie chez les animaux le problegraveme sanitaire est devenu mineur Dans les pays agrave
faibles revenus ougrave lrsquoabsence ou quasi-absence de controcircle de la maladie chez les bovins et la
consommation de lait et produits deacuteriveacutes crus les enjeux de la tuberculose bovine sont agrave la fois
sanitaires et socio-eacuteconomiques
Figure 6 Carte de distribution de la tuberculose bovine chez les bovins entre juillet et deacutecembre 2014 (Source WAHID Interface)
En Europe depuis plus drsquoun siegravecle lrsquoeacuteradication de la maladie a eacuteteacute un objectif majeur pour les
organisations agricoles et les autoriteacutes publiques (Reviriego-Gordejo amp Vermeersch 2006) Les mesures
reacuteglementaires eacutelaboreacutees notamment par lrsquoOIE (Office International des Epizooties aujourdrsquohui appeleacute
Organisation Mondiale pour la Santeacute Animale) visent agrave seacutecuriser les eacutechanges commerciaux drsquoanimaux
lrsquoameacutelioration des techniques de deacutepistage de la maladie les programmes drsquoeacuteradication et leurs aides
financiegraveres Les programmes mis en place chez les bovins ont permis de faire fortement diminuer
lrsquoincidence de la maladie dans la plupart des pays de lrsquoUnion Europeacuteenne (de la Rua-Domenech et al
2006) voire de lrsquoeacuteradiquer leur confeacuterant le statut de pays laquo officiellement indemne de tuberculose
bovine raquo (OTF Officially Tuberculosis Free) Le statut OTF est attribueacute agrave un pays lorsque
- le pourcentage de cheptels bovins confirmeacutes comme infecteacutes nrsquoexcegravede pas 01 par an pour
tous les cheptels pendant 6 ans
Chapitre 1
32
- chaque bovin est identifieacute en accord avec la regraveglementation europeacuteenne
- tous les bovins abattus subissent un examen post-mortem officiel (Reviriego-Gordejo amp
Vermeersch 2006)
Ce statut est tregraves important pour lrsquoeacuteconomie drsquoun pays puisqursquoil facilite le commerce national
et international des animaux par un allegravegement des controcircles sanitaires avant lrsquoexportation de leurs
bovins dans un autre pays membre ou avant leur abattage en les exemptant notamment drsquoune
tuberculination (Reviriego-Gordejo amp Vermeersch 2006) Les pays europeacuteens officiellement indemnes
de tuberculose sont lrsquoAllemagne lrsquoAutriche la Belgique le Danemark lrsquoEstonie la Finlande la France
lrsquoEcosse la Lettonie le Liechtenstein le Luxembourg la Norvegravege les Pays-Bas la Pologne la
Reacutepublique Tchegraveque la Slovaquie la Sloveacutenie la Suegravede la Suisse et une partie de lrsquoItalie (Figure 7) Les
pays drsquoEurope les plus toucheacutes sont la Grande-Bretagne lrsquoIrlande et lrsquoEspagne avec des preacutevalences
respectives de 104 44 et 12 en 2012 (EFSA 2014) Toutefois dans certains pays OTF comme la
France la Belgique lrsquoAllemagne lrsquoAutriche et la Pologne lrsquoeacuteradication de la maladie nrsquoest pas complegravete
malgreacute la preacutevention les controcircles et la surveillance engageacutes Dans ces pays lrsquoeacuteradication de la maladie
est fortement compliqueacutee par la circulation de lrsquoinfection dans des reacuteservoirs sauvages source
potentielle de contamination pour les bovins (Corner 2006 Nugent 2011 Gortaacutezar et al 2012)
Figure 7 Statuts des pays europeacuteens vis-agrave-vis de la tuberculose bovine en 2012 et proportion de bovins infecteacutes OTF Officially Tuberculosis Free (officiellement indemne de tuberculose bovine) MS Member State (Etat membre) (Source EFSA 2014)
Chapitre 1
33
Evolution de la situation sanitaire en France
Chez les bovins
Dans les anneacutees 50 environ 25 des cheptels bovins franccedilais eacutetaient infecteacutes par la tuberculose
bovine (Beacutenet et al 2006) Cette preacutevalence importante avait un fort impact eacuteconomique (saisies de
viande agrave lrsquoabattoir diminution de la productiviteacute en viande et en lait) mais aussi sanitaire puisque 2 agrave
11 des cas de tuberculose chez lrsquohomme eacutetaient imputeacutes agrave M bovis Les programmes de lutte mis en
place ont permis de faire diminuer de faccedilon spectaculaire la preacutevalence cheptel de la maladie de 25
en 1955 agrave moins de 01 degraves 1998 permettant agrave la France drsquoacceacuteder au statut OTF en 2000 (Figure 8)
Ce statut a permis drsquoalleacuteger voire de stopper la prophylaxie sur le territoire drsquoalleacuteger les controcircles mis
en place pour les mouvements de bovins dans lrsquoUE et de fournir une garantie sanitaire aux pays
importateurs de bovins franccedilais dans lrsquoUE et en dehors
Les programmes de lutte contre la tuberculose bovine reposent sur une base reacuteglementaire en
eacutevolution constante eacutelaboreacutee par les services veacuteteacuterinaires (maintenant DDPP Direction
Deacutepartementale de la Protection des Populations) en collaboration technique avec les veacuteteacuterinaires
sanitaires les groupements de deacutefense sanitaire (GDS) et les laboratoires drsquoanalyse agreacuteeacutes Ces
programmes sont baseacutes sur la surveillance systeacutematique et geacuteneacuteraliseacutee en abattoir (inspection post-
mortem recherche et identification de mycobacteacuteries par culture et PCR sur les leacutesions suspectes) et
sur la surveillance programmeacutee en eacutelevage (deacutepistage par intradermo-tuberculination et abattage total
ou partiel des troupeaux infecteacutes) Ils assurent eacutegalement la protection des cheptels indemnes par la
maitrise des facteurs de diffusion de la tuberculose bovine tels que lrsquointroduction drsquoun animal le
voisinage avec des cheptels infecteacutes et la reacutesurgence (Fediaevsky et al 2014) Depuis 2013 deux
Cellules Inter Reacutegionales drsquoEpideacutemiologie Veacuteteacuterinaire (CIREV) ont eacuteteacute creacuteeacutees en Bourgogne et en
Aquitaine afin de renforcer les investigations eacutepideacutemiologiques pour adapter de faccedilon optimale les
mesures de lutte contre la maladie (Fediaevsky et al 2013)
Chapitre 1
34
Figure 8 Evolution du taux de preacutevalence annuelle national de la tuberculose bovine chez les bovins (Beacutenet et al 2006)
Depuis 2004 la France connait une situation preacuteoccupante vis-agrave-vis de la tuberculose bovine
car lrsquoaugmentation du taux de preacutevalence met en danger le statut OTF du pays (Figure 9) Cette
augmentation est lieacutee agrave lrsquoapparition de nouveaux foyers de tuberculose concentreacutes dans des zones
geacuteographiques bien localiseacutees la Cocircte drsquoOr la Dordogne et les Pyreacuteneacutees Atlantiques cumulent 75
des foyers et dans une moindre mesure la Charente la Camargue les Hautes Pyreacuteneacutees lrsquoAriegravege les
Landes et la Corse (Figure 10) Dans ces deacutepartements caracteacuteriseacutes par de lrsquoeacutelevage allaitant
preacutedominant la contamination des troupeaux par le voisinage (par contacts directs avec des bovins
voisins ou des animaux sauvages infecteacutes ou par contacts indirects avec du mateacuteriel ou des matrices
contaminants) et lrsquointroduction drsquoun animal infecteacute dans le cheptel (notamment par lrsquoachat drsquoanimaux
le precirct et la pension) repreacutesentaient la moitieacute des cas en 2011 La reacutesurgence drsquoune ancienne infection
(qui persiste agrave bas bruit chez les animaux ou dans des matrices environnementales souilleacutees) et la faune
sauvage eacutetaient agrave lrsquoorigine preacutesumeacutee drsquoenviron 10 des cas respectivement toutefois sans
deacutemonstration scientifique Pour 35 des foyers bovins lrsquoorigine de lrsquoinfection eacutetait inconnue
(Fediaevsky et al 2012) La contamination des bovins via le voisinage la reacutesurgence et les foyers
drsquoorigine inconnue soulegravevent la question du rocircle de la transmission indirecte de M bovis entre la faune
sauvage et les bovins et donc le rocircle potentiel de lrsquoenvironnement comme reacuteservoir du bacille
25
15
10
5
20
1955
1963
1960
1970
1980
1990
2000
Chapitre 1
35
Figure 9 Evolution de la preacutevalence et de lrsquoincidence de la tuberculose bovine en France entre 1995 et 2014 (Cavalerie et al 2014)
La menace de la perte du statut OTF inquiegravete les pouvoirs publics et les instances agricoles car
la France est le plus gros exportateur de bovins vivants de lrsquoUE elle exporte principalement des bovins
maigres en Italie et en Espagne pour lrsquoengraissement (environ 12 million de tecirctes) mais aussi des gros
bovins destineacutes agrave la boucherie (220 000 tecirctes) et des veaux (128 000 tecirctes) (Source France AgriMer
les filiegraveres de lrsquoeacutelevage franccedilais eacutedition 2013) Les conseacutequences eacuteconomiques sur la filiegravere-
agroalimentaire lieacutees au blocage des animaux seraient consideacuterables De plus la gestion sanitaire de la
tuberculose bovine est tregraves coucircteuse puisque environ 20 millions drsquoeuros sont deacutepenseacutes chaque anneacutee
agrave lrsquoeacutechelle nationale pour lrsquoindemnisation des eacuteleveurs et la reacutealisation des analyses veacuteteacuterinaires
(Fediaevsky et al 2014)
Chapitre 1
36
Figure 10 Incidence et distribution de la tuberculose bovine en France dans les anneacutees 50 (en haut) et entre 2000 et 2014 (en bas) (Cavalerie et al 2014)
Chapitre 1
37
Dans la faune sauvage
En France agrave lrsquoheure actuelle la faune sauvage infecteacutee nrsquoest deacutetecteacutee qursquoagrave proximiteacute de foyers
bovins Le 1er foyer de tuberculose bovine dans la faune sauvage a eacuteteacute deacutecouvert en 2001 dans la forecirct
de Brotonne (Seine-Maritime et Eure) sur des cervideacutes puis des sangliers (Hars et al 2010 Zanella et
al 2008) Lrsquoaggravation de la situation sanitaire en 2005-2006 a neacutecessiteacute un abattage total des cerfs
consideacutereacutes comme reacuteservoir primaire de lrsquoinfection ainsi qursquoune diminution de la densiteacute des sangliers
reacuteservoir secondaire (Zanella et al 2008) Drsquoautres deacutepartements comme la Cocircte drsquoOr la Corse les
Pyreacuteneacutees Atlantiques la Dordogne et la Charente notamment ont par la suite deacutetecteacute des animaux
sauvages infecteacutes essentiellement blaireaux sangliers et dans une moindre mesure cervideacutes avec
toutefois des situations eacutepideacutemiologiques tregraves diffeacuterentes selon les reacutegions toucheacutees Ces situations
deacutependent de la population animale impliqueacutee (espegraveces densiteacutes interactionshellip) et de
lrsquoenvironnement (climat paysagehellip) Dans tous les cas le geacutenotypage (spoligotype et VNTR) des
souches incrimineacutees dans la faune sauvage a montreacute qursquoelles eacutetaient identiques agrave celles retrouveacutees
dans les eacutelevages bovins voisins suggeacuterant un lien eacutepideacutemiologique entre cas domestiques et sauvages
(Zanella et al 2008 Payne et al 2012 Richomme et al 2010)
Depuis 2011 un reacuteseau de surveillance eacutepideacutemiologique de la tuberculose bovine dans la faune
sauvage appeleacute Sylvatub a eacuteteacute mis en place en France Les diffeacuterentes modaliteacutes de surveillance ont
pour but la deacutetection de lrsquoinfection chez les animaux sauvages dans les zones agrave risque et preacutesumeacutees
indemnes ainsi que le suivi de son eacutevolution afin de proposer des mesures de lutte aux autoriteacutes
sanitaires (Riviegravere et al 2011) La surveillance est baseacutee sur un classement du territoire national en 3
niveaux de risque Pour les zones situeacutees en risque 1 (risque faible correspondant agrave une absence de
bovins deacutetecteacutes) il est demandeacute aux chasseurs de signaler toute leacutesion suspecte lors de lrsquoexamen initial
de la venaison Concernant les zones ougrave il existe un risque aveacutereacute (niveau 2 ou 3) la surveillance peut
ecirctre agrave la fois eacuteveacutenementielle (ramassage et analyse des cadavres drsquoanimaux trouveacutes morts) etou
programmeacutee (analyse des cerfs et sangliers tueacutes agrave la chasse et les blaireaux pieacutegeacutes) Les reacutesultats des
analyses effectueacutees en 2013-2014 sont preacutesenteacutes dans la Figure 11
Chapitre 1
38
Figure 11 Reacutepartition et reacutesultats des analyses effectueacutees dans le cadre du dispositif Sylvatub chez le grand gibier drsquoaoucirct 2013 agrave aoucirct 2014 et chez les blaireaux en 2014 (Source Sylvatub)
Evolution de la situation sanitaire en Cocircte drsquoOr
Quelques geacuteneacuteraliteacutes sur lrsquoeacutelevage bovin en Cocircte drsquoOr
La Cocircte drsquoOr est un deacutepartement drsquoeacutelevage bovin principalement allaitant (231 270 tecirctes en 2013
dont 74 565 vaches allaitantes et 15 687 vaches laitiegraveres) Lrsquoeacutelevage allaitant est majoritairement
concentreacute dans les zones herbagegraveres de lrsquoouest du deacutepartement et lrsquoeacutelevage laitier sur le plateau de
Langres au nord de Dijon Lrsquoeacutelevage allaitant est extensif (les bovins sont au preacute drsquoavril agrave octobre) lieacute agrave
la production de broutards maigres destineacutes agrave ecirctre engraisseacutes principalement en Italie En Cocircte drsquoOr le
parcellaire des exploitations de bovins allaitant est souvent tregraves morceleacute
Situation eacutepideacutemiologique chez les bovins et mesures de lutte associeacutees
Historiquement la Cocircte drsquoOr est un deacutepartement en prise avec la tuberculose depuis de
nombreuses anneacutees (Figure 10) A la fin des anneacutees 50 le taux de preacutevalence annuel chez les bovins
atteignait 65 Les mesures de lutte collective mises en place ont permis drsquoabaisser ce taux en dessous
du seuil des 10 en 1969 puis de le faire diminuer lentement et reacuteguliegraverement jusqursquoen 1985 ougrave la
maladie semblait eacuteradiqueacutee A la fin des anneacutees 90 les mesures prophylactiques ont cesseacute dans le
Chapitre 1
39
deacutepartement et la vigilance vis-agrave-vis de la maladie a baisseacute (Communication personnelle DDPP 21)
Les premiers signes drsquoune nouvelle eacutepizootie sont apparus en 2002 sur le canton de Pouilly-en-
Auxois (zone Sud) puis en 2003 dans ceux de Venarey-Les-Laumes et Vitteaux (zone Nord) (Source
DDPP21) Localiseacutee agrave quelques eacutelevages agrave son deacutebut la zone de lrsquoinfection srsquoest peu agrave peu eacutelargie au
cours du temps Un plan de prophylaxie restreint aux communes infecteacutees a eacuteteacute mis en place en 2002
puis eacutelargi aux cantons en 2003 avec de plus une surveillance des cheptels en lien eacutepideacutemiologique
avec les troupeaux infecteacutes et ceux des communes dans lesquelles les bovins infecteacutes avaient patucircreacute
Mais ces mesures nrsquoont pas suffi agrave limiter la progression de lrsquoinfection puisque limiteacute agrave moins de 5 par
an jusqursquoen 2006 le nombre de foyers est passeacute de 11 en 2007 agrave 17 en 2009 (Figure 13) Le
renforcement de la lutte comprenant notamment le deacutepistage annuel de tous les bovins de plus de 12
mois dans tout le deacutepartement a permis la deacutecouverte de 45 foyers en 2010 signe que lrsquoinfection
circulait agrave bas bruit De 2011 agrave 2013 le nombre de foyers a eacuteteacute diviseacute par 2 et pour lrsquoanneacutee 2014 10
foyers seulement ont eacuteteacute mis en eacutevidence ce qui semble conforter une maitrise de lrsquoeacutepizootie (Figure
13)
En Cocircte drsquoOr les foyers bovins sont circonscrits agrave une zone geacuteographique bien deacutelimiteacutee dans le
deacutepartement et partagent le mecircme spoligotype M bovis spoligotype SB0134 dans la zone Sud et
M bovis spoligotype SB0120 dans la zone Nord (Figure 12) Depuis 3 ans toutefois les deux zones ont
tendance agrave se teacutelescoper agrave cause notamment de la deacutecouverte drsquoanimaux sauvages infecteacutes avec
M bovis SB0120 en zone Sud Les deux spoligotypes ont deacutejagrave eacuteteacute retrouveacutes simultaneacutement dans des
cheptels infecteacutes Ces observations associeacutees agrave lrsquoabsence drsquoinstallation peacuterenne de la maladie dans des
zones du deacutepartements ougrave les concentrations de bovins sont aussi eacuteleacuteveacutees que dans la zone infecteacutee
soulegravevent la question de lrsquoexistence de facteurs locaux assurant lrsquoentretien et la propagation de
lrsquoinfection entre les eacutelevages bovins Au vu du morcellement et de lrsquoimbrication des pacirctures la
contamination par voisinage est probablement le facteur primordial chez les bovins Une eacutetude reacutecente
(de type cas-teacutemoin) reacutealiseacutee dans des cheptels de Cocircte drsquoOr de Dordogne et des Ardennes a montreacute
que les contacts de voisinage entre troupeaux eacutetaient le facteur de risque principal drsquoinfection des
troupeaux (Marsot et al Article soumis) Toutefois la deacutetection drsquoanimaux sauvages agrave proximiteacute de
certains foyers bovins interroge sur le rocircle potentiel de la faune sauvage infecteacutee dans la contamination
des bovins et dans la recontamination rapide de plusieurs cheptels apregraves abattage total des bovins et
vide sanitaire (Figure 13) (ANSES 2011)
Chapitre 1
40
Figure 12 Localisation du parcellaire des cheptels infecteacutes par la tuberculose bovine entre 2010 et 2015 et spoligotypes incrimineacutes (Source CIREV)
Figure 13 Nombre de nouveaux foyers bovins et de foyers recontamineacutes entre 2002 et 2014 (Donneacutees DDPP21)
0
5
10
15
20
25
30
35
40
45
50
2002 2003 2004 2005 2006 2007 2008 2009 2010 2011 2012 2013 2014
No
mb
re d
e fo
yers
bo
vin
s
Foyers contamineacutes pour la 1egravere fois Foyers recontamineacutes
Chapitre 1
41
Situation eacutepideacutemiologique dans la faune sauvage et mesures de lutte associeacutees
Le premier animal sauvage infecteacute une biche a eacuteteacute deacutecouvert en 2003 agrave proximiteacute de 2 foyers
bovins de 2002 dans la zone Sud Entre 2003 et 2007 la surveillance active de la faune sauvage mise
en place et porteacutee par lrsquoONCFS (Office National de la Chasse et de la Faune Sauvage) et la DDPP et sur
le terrain par les chasseurs les pieacutegeurs et les veacuteteacuterinaires a permis de mettre en eacutevidence lrsquoinfection
chez 1 cervideacute (n=284) 2 sangliers (n= 160) mais aucun blaireau (n=63) Depuis 2007 des sangliers
infecteacutes sont retrouveacutes tous les ans notamment de jeunes animaux avec des leacutesions eacutevolutives
majoritairement dans la zone Sud ougrave les populations sont plus importantes Depuis 2009 lrsquoinfection
est retrouveacutee chez des blaireaux vivant agrave proximiteacute des cheptels infecteacutes majoritairement dans la zone
Nord (Hars Richomme and Boschiroli 2010 DDPP21)(Figure 14) La maladie circule donc dans un
systegraveme multi-hocirctes complexe et unique incluant les bovins les blaireaux les sangliers et les cervideacutes
(Payne 2014)
Figure 14 Nombre de cervideacutes sangliers et blaireaux infecteacutes deacutecouverts en Cocircte drsquoOr entre 2002 et 2014
Dans la crainte de lrsquoeacutetablissement drsquoun reacuteservoir sauvage chez le sanglier comme en Espagne
(Naranjo et al 2008) ou le blaireau au Royaume-Uni (Delahay et al 2001) des mesures de diminution
des densiteacutes ont eacuteteacute mises en place (augmentation des tableaux de chasse pour le sanglier et les
cervideacutes et intensification du pieacutegeage des blaireaux) le ramassage des viscegraveres drsquoanimaux tueacutes agrave la
chasse a eacuteteacute rendu obligatoire et lrsquoagrainage interdit
0
5
10
15
20
25
30
35
No
mb
re d
an
imau
x in
fect
eacutes
Cervideacutes Sangliers Blaireaux
Chapitre 1
42
C ECOLOGIE DE M BOVIS DANS LrsquoENVIRONNEMENT
1 Introduction
Avant les anneacutees 30 il eacutetait admis sine qua none qursquoen tant que bacteacuteries intracellulaires
obligatoires les bacteacuteries tuberculeuses ne survivaient pas en dehors de leur hocircte Toutefois degraves le
deacutebut du 20egraveme siegravecle la deacutetection de Bacillus tuberculosis virulent dans lrsquoeau et les biofilms des
canalisations drsquoeffluents de diffeacuterents sanatoria en Allemagne et au Royaume-Uni (Musehold en 1900
citeacute par Jensen 1954 et Conroy et al en 1921 et 1922 et Cummins et Ackland en 1929 citeacutes par (Williams
amp Hoy 1930) a souleveacute la question du rocircle potentiel de lrsquoenvironnement dans lrsquoeacutepideacutemiologie des
mycobacteacuteries tuberculeuses Concernant la tuberculose bovine les diffeacuterentes recherches meneacutees
pour deacutetecter et caracteacuteriser la persistance de M bovis dans lrsquoenvironnement ont pour objectif
principal de preacuteciser le rocircle de la transmission indirecte dans leacutepideacutemiologie de la maladie La
transmission indirecte semble en effet prendre toute son importance lorsque des reacuteservoirs sauvages
sont identifieacutes etou des systegravemes multi-hocirctes car ils ont la capaciteacute de transmettre lrsquoinfection aux
bovins (Corner 2006 Payne 2014)
Reacutepondre agrave cette question neacutecessite de connaitre les modaliteacutes de transfert des bacteacuteries de
lrsquoanimal ou lrsquohomme infecteacute vers lrsquoenvironnement la dureacutee de survie dans les diffeacuterentes matrices les
strateacutegies de survie de la bacteacuterie ainsi que les facteurs qui affectent sa survie
2 Origines de la contamination environnementale par M bovis
a Lrsquoexcreacutetion animale de Mycobacterium bovis
Lrsquohomme et les animaux infecteacutes ont la capaciteacute drsquoexcreacuteter le bacille via diffeacuterentes voies et
constituent donc une source de contamination potentielle pour lrsquoenvironnement Comme nous lrsquoavons
indiqueacute preacuteceacutedemment les voies drsquoexcreacutetion ainsi que les niveaux drsquoexcreacutetion deacutependent de la
localisation des leacutesions et de leur seacuteveacuteriteacute Les tableaux leacutesionnels deacutecrits preacuteceacutedemment indiquent
que M bovis est susceptible drsquoecirctre excreacuteteacutee principalement par voie orale ou nasale lors drsquoatteinte
pulmonaire etou des nœuds lymphatiques ceacutephaliques par voie digestive via les matiegraveres feacutecales et
par voie urinaire lors drsquoinfection geacuteneacuteraliseacutee (Phillips et al 2003 Corner et al 2012) Lrsquoexcreacutetion feacutecale
de bacteacuteries du MTBC en lrsquoabsence de leacutesions gastro-intestinales semble majoritairement lieacutee au
passage de mucus respiratoire chargeacute en bacilles dans le tube digestif La reacutesistance aux pH acides
(Vandal et al 2008) des bacteacuteries du MTBC leur permet de reacutesister au passage du tractus digestif et
drsquoecirctre excreacuteteacutees sous forme viable dans les matiegraveres feacutecales comme cela a eacuteteacute deacutemontreacute pour M bovis
dans certaines espegraveces animales (Corner Murphy and Gormley 2011 Gallagher and Clifton-Hadley
Chapitre 1
43
2000 Gavier-Widen et al 2001) pour M tuberculosis (El Kheacutechine et al 2009) et M africanum
(Cadmus et al 2009)
Chez les bovins lrsquoexcreacutetion du bacille a eacuteteacute deacutemontreacutee dans le mucus nasal et tracheacuteal et les
fegraveces drsquoanimaux infecteacutes expeacuterimentalement (Neill et al 1988 McCorry et al 2005 Phillips et al 2003
Maddock 1936) Le deacutelai drsquoexcreacutetion apregraves lrsquoinoculation la dureacutee de lrsquoexcreacutetion (plusieurs semaines) et
la quantiteacute de bacilles excreacuteteacutes sont proportionnels agrave la dose infectieuse utiliseacutee (Neill et al 1988)
Lrsquoexcreacutetion feacutecale apparait peu freacutequente et intermittente chez les bovins (Neill et al 1988 Goodchild
amp Clifton-Hadley 2001) Dans les pays industrialiseacutes ougrave le deacutepistage des animaux infecteacutes est en place
lrsquoexcreacutetion de M bovis dans le lait les fegraveces et les urines est consideacutereacutee comme insignifiante chez les
bovins (Morris et al 1994)
Chez le blaireau infecteacute naturellement ou expeacuterimentalement M bovis est excreacuteteacutee par 25 agrave
50 des animaux par voie respiratoire (mucus tracheacuteal et aeacuterosols) urinaire et feacutecale ainsi que par la
suppuration drsquoabcegraves reacutesultant de morsures (Corner et al 2011 Delahay et al 2000 Clifton-Hadley et
al 1993) Lrsquoisolement de M bovis dans des glandes salivaires de blaireaux suggegravere que la salive
constitue eacutegalement une voie possible drsquoexcreacutetion du bacille (Payne 2014) Les blaireaux preacutesentant un
stade avanceacute de la maladie et des leacutesions macroscopiques semblent ecirctre les plus excreacuteteurs en terme
de dureacutee et drsquointensiteacute drsquoexcreacutetion (Corner et al 2012 Gallagher amp Clifton-Hadley 2000 Jenkins et al
2008) toutefois lrsquoexcreacutetion du bacille a eacuteteacute deacutecrite chez des animaux sans leacutesions visibles (Jenkins et al
2008 Gavier-Wideacuten et al 2009) Dans les stades avanceacutes de la maladie les quantiteacutes excreacuteteacutees peuvent
atteindre 2105 UFCmL1 de mucus bronchique 25105 UFCmL1 drsquourine et 75104 UFCg1 de fegraveces
(Gallagher et al 1998) Le bacille est excreacuteteacute par voie feacutecale de 165 agrave 1305 jours avant la mort de
lrsquoanimal (Little et al 1982) Dans cette espegravece lrsquoexcreacutetion est geacuteneacuteralement intermittente mais des
individus laquo super excreacuteteurs raquo ont eacuteteacute deacutecrits et se caracteacuterisent par une excreacutetion persistante dans le
temps ou par lrsquoexcreacutetion du bacille par plusieurs voies simultaneacutement (Delahay et al 2000) Des
blaireaux laquo super excreacuteteurs raquo ont eacuteteacute mis en eacutevidence en Cocircte drsquoOr la voie feacutecale eacutetant toujours
positive chez ces animaux (Payne 2014) Lrsquointensiteacute de lrsquoexcreacutetion semble varier avec lrsquoacircge (les adultes
excregravetent plus que les jeunes) et la saison puisque lrsquoexcreacutetion semble supeacuterieure au printemps et en
eacuteteacute (Delahay et al 2013 King Murphy James Travis Porter Hung et al 2015)
Chez le sanglier lrsquoexcreacutetion de M bovis se produit par voie orale nasale feacutecale et urinaire
(Martiacuten-Hernando et al 2007 Santos Almeida et al 2015 Barasona et al 2015) Lrsquoexcreacutetion salivaire
est encore discuteacutee (Payne 2014) En Espagne lrsquoexcreacutetion par au moins une des voies a eacuteteacute mise en
eacutevidence par PCR sur 30 des sangliers analyseacutes (n = 39) et 25 des 39 animaux excreacutetaient par les trois
voies (Barasona et al 2015) Lrsquoexcreacutetion eacutetait majoritairement associeacutee agrave une maladie geacuteneacuteraliseacutee Au
Portugal sur 39 sangliers adultes ou subadultes infecteacutes analyseacutes 80 excreacutetaient le bacille par au
moins une voie et 13 eacutetaient des laquo super excreacuteteurs raquo (excreacutetant par trois voies diffeacuterentes) Environ
Chapitre 1
44
28 des sangliers excreacutetaient des quantiteacutes de bacilles supeacuterieures agrave 1 000 UFC par g ou mL drsquoexcreacuteta
par au moins une voie (Santos Almeida et al 2015) Chez le sanglier agrave notre connaissance lrsquoexcreacutetion
nrsquoa jamais eacuteteacute deacutecrite en lrsquoabsence de leacutesions
Chez les cervideacutes diffeacuterentes eacutetudes ont montreacute que le bacille eacutetait excreacuteteacute de faccedilon tregraves
sporadique dans les mucus nasal oro-pharyngeacute et tracheacuteal ainsi que dans les matiegraveres feacutecales lors
drsquoinfections naturelles ou expeacuterimentales (Lugton et al 1998 Santos Almeida et al 2015 Palmer et
al 2001) Dans cette espegravece une excreacutetion intermittente apparait degraves les premiers stades de la
maladie la seacuteveacuteriteacute des tableaux leacutesionnels rencontreacutes explique le niveau drsquoexcreacutetion eacuteleveacute (Palmer et
al 2001 Santos Almeida et al 2015)
Les carcasses drsquoanimaux infecteacutes peuvent constituer une source drsquoinfection directe pour les
animaux charognards qui srsquoen nourrissent mais eacutegalement une source de contamination indirecte pour
les bovins qui pacircturent sur le site de deacutecomposition de la carcasse (Morris et al 1994) Concernant les
animaux tueacutes agrave la chasse lrsquoabandon des viscegraveres ou leur ensevelissement repreacutesente eacutegalement un
risque pour les animaux charognards De ce fait en France la collecte et lrsquoeacutelimination des viscegraveres des
espegraveces sensibles sont obligatoires dans les zones toucheacutees par la tuberculose bovine
b Le transfert de Mycobacterium bovis par les effluents drsquoeacutelevage ou urbains
Les eacutetudes sur la contamination potentielle des effluents drsquoeacutelevage avec M bovis sont tregraves
rares probablement agrave cause de la difficulteacute agrave cultiver le bacille dans ces matrices complexes Les
multiples voies drsquoexcreacutetion du bacille et plus particuliegraverement lrsquoexcreacutetion par voie feacutecale mecircme si elle
est intermittente peuvent probablement contribuer agrave la contamination des effluents drsquoeacutelevage bovin
tels que les fumiers lisiers et eaux useacutees Lrsquoeacutepandage de ces effluents sur les pacirctures ou lrsquoirrigation des
cultures peut constituer un vecteur de contamination du sol des cultures et de lrsquoeau profonde ou de
surface et repreacutesenter un risque pour les animaux et lrsquohomme mais aucune donneacutee chiffreacutee nrsquoexiste
pour ces matrices en conditions naturelles Lrsquoabattage et la deacutecoupe de bovins tuberculeux peut
eacutegalement repreacutesenter une source de contamination des rejets drsquoabattoir (Jensen 1954) Au vu des
mesures actuelles de collecte stockage traitement et controcircle des effluents drsquoabattoir cette
contamination semble tregraves peu probable en France Toutefois elle reste complegravetement valable dans
les zones drsquoendeacutemie des pays nrsquoappliquant pas ces mesures et ne disposant pas de structures
drsquoabattage adeacutequates
Il est inteacuteressant de noter que quelques eacutetudes sont disponibles concernant la contamination
drsquoeffluents urbains avec M tuberculosis Jensen (1954) a montreacute que les effluents bruts de sanatoria
de 5 villes des Pays-Bas les effluents purifieacutes les boues digeacutereacutees et les boues seacutecheacutees (preacuteleveacutees 5
semaines apregraves lrsquoeacutepandage) contenaient des bacilles virulents de M tuberculosis Plus reacutecemment la
Chapitre 1
45
mise en culture de 800 eacutechantillons drsquoeau et 700 eacutechantillons de sol preacuteleveacutes dans 3 quartiers de
Teacuteheacuteran (Iran) entre 2012 et 2014 a deacutemontreacute la preacutesence de M tuberculosis dans 71 des eacutechantillons
drsquoeau (88) et 11 eacutechantillons de sol (15) Les eacutechantillons drsquoeau contamineacutes ont eacuteteacute retrouveacutes
principalement dans les rigoles de ciment acheminant les eaux useacutees et pluviales (5671) et dans les
eaux traiteacutees (1571) La bacteacuterie nrsquoa toutefois pas eacuteteacute isoleacutee du reacuteseau drsquoeau potable Environ 10 des
souches isoleacutees eacutetaient multi-reacutesistants aux antibiotiques (Velayati et al 2015)
3 Deacutetection et survie de Mycobacterium bovis dans lrsquoenvironnement
a Deacutetection de Mycobacterium bovis en conditions naturelles
A ce jour peu drsquoeacutetudes concernant la deacutetection de M bovis dans des substrats naturellement
contamineacutes ont eacuteteacute publieacutees La deacutetection de M bovis dans des eacutechantillons environnementaux
naturellement contamineacutes gracircce agrave des meacutethodes baseacutees sur la culture est exceptionnelle A
Woodchester Park dans le Gloucestershire (Sud-Ouest de lrsquoAngleterre) ougrave les densiteacutes de blaireaux
sont eacuteleveacutees et la preacutevalence importante (20 en 2000 et 35 en 2005) un suivi longitudinal du statut
des populations est reacutealiseacute depuis 1980 (Delahay et al 2000) et diffeacuterentes campagnes de
preacutelegravevements de sols de terriers et de latrines ont eacuteteacute reacutealiseacutees Le traitement drsquoeacutechantillons de sol et
latrines provenant de Woodchester Park traiteacute par immunocapture magneacutetique a permis pour la
premiegravere et seule fois lrsquoisolement de M bovis par culture dans des eacutechantillons environnementaux
(Sweeney et al 2006 Sweeney et al 2007) Cette technique est toutefois peu applicable agrave un grand
nombre drsquoeacutechantillons en termes de coucirct et de temps et nrsquoa jamais eacuteteacute reacuteutiliseacutee par la suite
Drsquoautres tentatives de deacutetection de M bovis par diffeacuterentes meacutethodes baseacutees sur la culture
dans des zones endeacutemiques de tuberculose bovine ont eacutechoueacute malgreacute le grand nombre drsquoeacutechantillons
environnementaux preacuteleveacutes (Pillai et al 2000 Fine Bolin et al 2011 Iivanainen et al 1999 Williams
amp Hoy 1930a Little et al 1982) La deacutetection de M bovis dans des eacutechantillons environnementaux par
la culture est compliqueacutee par la nature des eacutechantillons analyseacutes en effet ces derniers (sol seacutediments
eau stagnante matiegraveres feacutecales etc) contiennent des micro-organismes tregraves divers et abondants La
croissance de M bovis eacutetant tregraves lente les organismes agrave croissance rapide contaminent rapidement les
milieux de culture compliquant voire empecircchant lrsquoisolement de la bacteacuterie Des traitements de
deacutecontamination preacuteliminaires sont indispensables pour eacuteliminer cette microflore mais la
deacutecontamination reacuteduit en geacuteneacuteral fortement la viabiliteacute de M bovis reacuteduisant aussi la sensibiliteacute des
meacutethodes baseacutees sur la culture (Fine Bolin et al 2011) Des microorganismes voisins des
mycobacteacuteries et preacutesents dans les sols peuvent eacutegalement reacutesister aux proceacutedeacutes de deacutecontamination
et ecirctre retrouveacutes dans les isolements De plus lrsquointermittence et le faible niveau drsquoexcreacutetion chez les
Chapitre 1
46
animaux infecteacutes associeacutes au volume limiteacute des preacutelegravevements analyseacutes diminuent les chances de
deacutetection de M bovis dans lrsquoenvironnement (Fine OrsquoBrien et al 2011) Lrsquoameacutelioration des protocoles
de traitement des eacutechantillons avant la culture est fondamentale pour lrsquoisolement des souches
environnementales
Le deacuteveloppement des outils moleacuteculaires a permis de contourner les inconveacutenients lieacutes aux
techniques baseacutees sur la culture deacutecrits preacuteceacutedemment En Irlande lrsquoanalyse de preacutelegravevements reacutealiseacutes
en 2000 dans un eacutelevage bovin toucheacute plusieurs fois par la tuberculose a mis en eacutevidence par des
meacutethodes moleacuteculaires la preacutesence de 1 times 103 agrave 25 times 103 copies de gegravenes cibles de M bovis (mpb64 et
mpb70) dans des pacirctures et des sols drsquoentreacutee de terriers de blaireaux adjacents aux pacirctures (Young et
al 2005) La reacuteiteacuteration des analyses 30 mois plus tard en novembre 2002 a donneacute le mecircme reacutesultat
sur les terriers bien qursquoils soient resteacutes inhabiteacutes Le rocircle des blaireaux dans la contamination des bovins
est fortement suspecteacute dans cette ferme En Angleterre lrsquoanalyse moleacuteculaire de preacutelegravevements
environnementaux (terriers et latrines) reacutealiseacutes en 2003 autour ou dans les pacirctures drsquoeacutelevages bovins
infecteacutes dans six zones endeacutemiques de tuberculose bovine a mis en eacutevidence M bovis dans les sols
de terriers de blaireaux de 78 des fermes analyseacutees (n=60) Dans chaque ferme 47 des terriers et
29 des latrines analyseacutes eacutetaient positifs en PCR une forte proportion eacutetait accessible aux bovins
(Courtenay et al 2006)
Entre 2004 et 2013 lrsquoanalyse par deacutetection moleacuteculaire de latrines appartenantt agrave des groupes sociaux
de blaireaux infecteacutes de Woodchester Park a montreacute que selon les eacutetudes 13 agrave 100 des latrines eacutetaient
contamineacutees avec des quantiteacutes pouvant atteindre 11 times 108 copies de gegravenes par g de latrine
(Courtenay et al 2006 Travis et al 2011 Sweeney et al 2006) Lrsquoamplification de seacutequences ciblant
lrsquoARNr 16S dans certains eacutechantillons des eacutetudes preacutesenteacutees ci-dessus suggegravere que les bacilles deacutetecteacutes
eacutetaient viables (Young et al 2005 Courtenay et al 2006) toutefois la seacutequence de lrsquoARNr 16S cibleacutee
eacutetant speacutecifique de toutes les mycobacteacuteries agrave croissance lente les reacutesultats sont agrave consideacuterer avec
prudence Dans ces eacutetudes le nombre de blaireaux excreacuteteurs et leur niveau drsquoexcreacutetion sont
fortement correacuteleacutes agrave la contamination des terriers et des latrines
Dans le sud du Portugal une eacutetude reacutecente dans une zone endeacutemique a mis en eacutevidence par PCR ciblant
lrsquoIS6110 des bacteacuteries du MTBC dans 38 des 248 eacutechantillons preacuteleveacutes dans des zones drsquoagreacutegation
drsquoanimaux domestiques et sauvages zones drsquoabreuvement utiliseacutees par les bovins et la faune sauvage
(seacutediments et eau drsquoeacutetangs 1358 et 357 et de sources saisonniegraveres 1161) sol des zones de
nourrissage des animaux (528) du sol de boutis de sangliers (416) ainsi que dans le sol des zones de
nourrissage des vautours (119) et dans leurs fegraveces (19) (Santos Santos et al 2015) Lrsquoutilisation drsquoune
seconde cible PCR (RD12) a confirmeacute 4 eacutechantillons comme positifs pour M bovisM caprae (2
eacutechantillons de seacutediments drsquoeacutetang 1 de source et 1 de sol de nourrissage) sans pouvoir diffeacuterencier les
deux espegraveces Le plus grand nombre drsquoeacutechantillons positifs a eacuteteacute retrouveacute au printemps (tempeacuterature
Chapitre 1
47
moyenne de 16degC)
La PCR permet drsquoobtenir un reacutesultat rapide avec une speacutecificiteacute acceptable Lrsquoanalyse
drsquoeacutechantillons de sol drsquoeau de foin et de maiumls inoculeacutes avec M bovis par une meacutethode baseacutee sur la
culture et la PCR a montreacute que cette derniegravere permet de deacutetecter lrsquoADN de M bovis dans lrsquoeacutechantillon
sur une plus longue peacuteriode apregraves la contamination que la culture indeacutependamment du substrat
(Adams et al 2013) Toutefois les seuils de deacutetection eacuteleveacutes des systegravemes speacutecifiques de M bovis (de
4 times 104 agrave 5 times 105 par g ou mL drsquoeacutechantillon Tableau 3) limitent leur utilisation dans le screening
drsquoeacutechantillons environnementaux pour lesquels la charge bacteacuterienne est probablement infeacuterieure au
seuil de deacutetection Enfin la technique PCR ne donne aucune information sur la viabiliteacute et la virulence
des bacteacuteries deacutetecteacutees
b Dureacutees de survie de Mycobacterium bovis en conditions expeacuterimentales
Les eacutetudes expeacuterimentales sur la survie environnementale de M bovis ont deacutebuteacute dans les
anneacutees 1930 en Angleterre dans lrsquoobjectif de preacuteciser le rocircle de lrsquoenvironnement dans lrsquoeacutepideacutemiologie
de la tuberculose bovine et drsquoameacuteliorer les programmes drsquoeacuteradication de la maladie (Maddock 1933
Maddock 1934 Maddock 1936 Williams amp Hoy 1930) Mycobacterium bovis et M bovis BCG sont
capables de survivre dans les fegraveces les effluents le sol et les divers excreacutetas animaux de quelques jours
agrave quelques mois voire jusqursquoagrave deux ans () Les nombreuses conditions drsquoinoculation (concentration de
lrsquoinoculum souche utiliseacutee M bovis ou M bovis BCG origine de lrsquoinoculum culture ou broyat de
leacutesions) la varieacuteteacute des matrices inoculeacutees (matiegraveres feacutecales urine mucus organes sol eau etc) et
des conditions environnementales testeacutees (conditions controcircleacutees de laboratoire ou exposition aux
conditions climatiques) dans ces diffeacuterentes eacutetudes expliquent vraisemblablement de tels eacutecarts de
survie
La compilation de toutes ces donneacutees indique que la tempeacuterature lrsquoexposition aux UV et
lrsquohumiditeacute de la matrice semblent ecirctre les trois facteurs primordiaux intervenant dans la persistance
de M bovis quel que soit le substrat analyseacute Il est montreacute que dans lrsquoheacutemisphegravere Nord la dureacutee de
survie de M bovis est plus longue en automne hiver et printemps (Williams amp Hoy 1930 Tanner amp
Michel 1999 Fine Bolin et al 2011 Duffield amp Young 1985 Jackson et al 1995) saisons caracteacuteriseacutees
par des tempeacuteratures plus basses qursquoen eacuteteacute un ensoleillement minimal et une eacutevapotranspiration par
le couvert veacutegeacutetal limiteacutee Toutefois Fine et al (2011) ont montreacute que M bovis pouvait survivre jusqursquoagrave
48 jours dans lrsquoeau en eacuteteacute malgreacute les tempeacuteratures eacuteleveacutees et lrsquoensoleillement Les eacutetudes reacutealiseacutees en
Afrique du Sud et en Nouvelle-Zeacutelande ougrave les tempeacuteratures sont eacuteleveacutees sur de longues dureacutees avec
un ensoleillement important ont montreacute que la survie de M bovis est ameacutelioreacutee lorsque le sol contient
une humiditeacute suffisante sous une exposition ombrageacutee (Duffield amp Young 1985 Tanner amp Michel 1999)
Chapitre 1
48
Lrsquoexposition des bacilles aux UV diminue en effet fortement leur dureacutee de persistance dans
lrsquoenvironnement (Williams amp Hoy 1930a Genov 1965 Fine Bolin et al 2011) et la dureacutee de survie est
tregraves prolongeacutee lorsque les eacutechantillons sont enterreacutes agrave quelques centimegravetres sous la surface du sol
(Genov 1965) La survie de M bovis est optimale dans les matiegraveres feacutecales ougrave le bacille est proteacutegeacute
dans un environnement humide agrave lrsquoabri des radiations solaires et minimale lorsqursquoil est pulveacuteriseacute sur
les veacutegeacutetaux et soumis aux UV et agrave la dessiccation (Maddock 1933 Van Donsel amp Larkin 1977)
49
Reacutefeacuterences BCG M bovis Matrices inoculeacutees Concentration de
lrsquoinoculum Conditions
Meacutethodes de deacutetection
de M bovis Reacutesultats de survie (virulence) de M bovis
(Williams amp Hoy
1930)
X Fegraveces de bovin
Inconnue Expeacuterimentales
(tempeacuterature
fraicircche obscuriteacute)
Injection sous-
cutaneacutee agrave des cochons
drsquoInde (cuisse)
12 mois dans fegraveces naturellement infecteacutees
5104 agrave 5107 UFC 2 ans dans fegraveces artificiellement infecteacutees
X Lisier de bovin 5000 UFCcm3
Expeacuterimentales
(tempeacuterature
fraicircche obscuriteacute)
Injection sous-
cutaneacutee agrave des cochons
drsquoInde (cuisse)
Survie 4 mois avec diminution de la virulence du
bacille au cours du temps
X Fegraveces de bovin 5104 agrave 5107 UFC Environnementales
(pacircture)
Injection sous-cutaneacutee
agrave des cochons drsquoInde
(cuisse)
5 mois en hiver
4 mois en automne
2 mois au printemps eacuteteacute
4 mois si protection UV en eacuteteacute
6 mois si protection insectes-vers en automne
(Maddock 1933)
X
Sol
Sol + Fegraveces de bovin
Bouse
Inconnue Environnementales
(pacircture)
Injection sous-cutaneacutee
agrave des cochons drsquoInde
(cuisse)
178 j dans toutes les matrices
X Herbe
12105 01 m2 Environnementales
(pacircture)
Injection sous-
cutaneacutee agrave des cochons
drsquoInde (cuisse)
14 jours
12106 01 m2 28 jours
120106 01 m2 49 jours
(Maddock 1934) X Herbe
Inoculation
mensuelle de 5 agrave
25106 01 m2
reacutepeacuteteacutee
Environnementales
et expeacuterimentales
Ingestion drsquoherbe
contamineacutee
1422 cochons drsquoInde avec leacutesions
Tableau 1 Dureacutees de survie de M bovis dans diffeacuterentes matrices environnementales obtenues en conditions expeacuterimentales naturelles ou de laboratoire
50
(Maddock 1936)
X
Veaux infecteacutes
(excreacutetion prouveacutee
par injection agrave des
cochons drsquoInde)
Inconnue Environnementales
Pacircture seacutepareacutee en 3 2
veaux mis 1 mois apregraves
le retrait des veaux
infecteacutes 2 veaux 1 mois
plus tard 2 veaux 2
mois plus tard (laisseacutes 3
sem)
Aucun veau IDR +
Aucune leacutesion tuberculeuse agrave lrsquoautopsie
X
Vache infecteacutee
(excreacutetion prouveacutee
par injection agrave des
cochons drsquoInde)
Pacircture seacutepareacutee en 2 3
veaux mis 1 mois et 12
apregraves le retrait de la
vache infecteacutee (laisseacutes
53j) 3 veaux mis 1 mois
plus tard (laisseacutes 24j)
Abattage et autopsie 1 an plus tard aucune leacutesion
Bouses infectieuses 1 mois et frac12 apregraves retrait de la
vache infecteacutee puis neacutegatif 1 2 et 3 mois plus tard
Sol et herbe sous bouses neacutegatifs 3 4 et 5 mois apregraves
retrait de la vache infecteacutee
(Genov 1965) X
Fegraveces de bovin
Sang
Urine
Environnementales Culture
150 agrave 332 jours agrave 12-24degC sans drsquoUV
18 agrave 31 jours agrave 24-34degC avec UV
2 ans si enterreacute dans sol agrave 5 cm
11 mois si enterreacute agrave 1 cm
452 et 469 jours dans eaubouse et eauurine
(Van Donsel amp
Larkin 1977) X
Effluents animaux
Boues drsquoeacutepuration Environnementales
Abattage 90 en 11 jours pour sols arroseacutes avec
effluents 8 jours avec boues
Isolement apregraves 4 agrave 6 jours sur des radis et jusqursquoagrave
35 jours sur des laitues
Anon 1979 citeacute par
(Morris et al 1994)
Excreacutetas de
blaireaux conserveacutes
dans un terrier
Environnementales Culture
Urine 3 jours en eacuteteacute 28 jours en hiver
Mucus bronchique 7 et 70 jours
Fegraveces naturellement infecteacutes 14 et 28 jours
(Little et al 1982)
Sol foin litiegravere et
eau de blaireaux
inoculeacutes
Inconnue
Expeacuterimentales
Culture 1 culture positive (eau drsquoun abreuvoir)
51
(Duffield amp Young
1985)
Sol sec steacuterile ou non
Sol 30 humiditeacute
steacuterile ou non
Fegraveces
Culture
lt8 semaines dans sols secs ou humides non steacuteriles
avec 80 ombre (34degc) obscuriteacute (32degC) et dans
labo (23degC)
lt8 semaines dans sol humide steacuterile avec 80
ombre et obscuriteacute
Pas de survie plein soleil (43degC) dans sols et fegraveces
(Tanner amp Michel
1999)
Leacutesions pulmonaires
ou ganglionnaires de
buffle
Fegraveces de bovin
inoculeacutees
8107 UFC 50g
fegraveces Culture
Poumon 42 jours en hiver agrave lrsquoombre dans sol
humide ou sec (20degC)
Fegraveces 28 jours en hiver au soleil dans sol humide
ou sec
(Jackson et al
1995) X Rubans de coton
Environ 106 UFC
ruban
Environnementales
(rubans deacuteposeacutes
dans terriers de
possum et sols de
pacirctures ou de forecirct)
Culture
2 jours sur les sols de pacirctures
14 jours sur les sols forestiers et dans les terriers de
possum
(Scanlon amp Quinn
2000b) X Lisier bovin steacuteriliseacute 65 times 103 UFCmL Expeacuterimentales Culture 171 jours (concentration finale 2 UFCmL)
(Young et al 2005) X Sol steacuterile et non
steacuterile 108 UFC g sol
Expeacuterimentales
(4 15 25 37degC)
PCR pour les sols non
steacuteriles
Culture pour les sols
steacuteriles
Survie agrave 60 jours optimale agrave 37degC agrave -20kPa
drsquohumiditeacute dans le sol steacuterile
Deacutetection drsquoARNm pendant 15 mois (sol steacuterile)
(Palmer amp Whipple
2006) X
Betterave pommes
carottes pommes de
terre maiumls foin
11 times 106
UFC10g
Expeacuterimentales
(-20 8 et 23degC)
Culture
112 jours agrave -20degC pour tous les substrats
112 jours agrave 8degC pour pommes maiumls foin et pdeterre
112 jours agrave 23degC pour pommes maiumls et pdeterre
(Fine Bolin et al
2011) X Sol foin maiumls eau
50 times 104
UFCeacutechantillon Environnementales Culture
88 jours dans le sol en hiverprintemps
58 jours dans lrsquoeau 43 jours dans le foin et 37 jours
dans le maiumls en automnehiver
52
(Ghodbane et al
2014) X Sol steacuterile 108 UFC g sol
Expeacuterimentales
(18 agrave 24degC
obscuriteacute)
Culture
Survie 12 mois (concentration finale 150 UFCg
sol)
Virulence conserveacutee sur souris
Chapitre 1
53
Dans les fegraveces drsquoanimaux M bovis survit jusqursquo agrave 2 ans dans les bouses de bovins en conditions
de laboratoire (Williams amp Hoy 1930) et de 14 jours agrave 6 mois lorsque les fegraveces sont exposeacutees dans les
pacirctures (Williams amp Hoy 1930a Genov 1965 Morris et al 1994 Tanner amp Michel 1999) Dans les
effluents drsquoeacutelevages contamineacutes il a eacuteteacute montreacute que M bovis survivait de 4 mois dans du lisier en
conditions de laboratoire (Williams amp Hoy 1930a) et jusqursquoagrave 171 jours dans du lisier steacuteriliseacute conserveacute
agrave tempeacuterature ambiante et agrave lrsquoobscuriteacute (Scanlon amp Quinn 2000b) Lrsquoeacutepandage drsquoeffluents ou de boues
inoculeacutees avec M bovis BCG sur des cultures de radis et laitue a montreacute la reacuteduction de 90 des bacilles
en respectivement 11 et 8 jours M bovis a toutefois eacuteteacute isoleacutee de faccedilon sporadique 35 jours apregraves
arrosage (Van Donsel amp Larkin 1977)
Dans le sol les dureacutees de survie les plus longues pour M bovis et M bovis BCG ont eacuteteacute
observeacutees dans du sol steacuterile (63 de sable pH 65-68) et en conditions de laboratoire (Young et al
2005 Ghodbane et al 2014) Young et al (2005) ont amplifieacute lrsquoARNr 16S de M bovis BCG jusqursquoagrave 15
mois apregraves inoculation du sol alors que Ghodbane et al (2014) ont cultiveacute M bovis apregraves 12 mois de
seacutejour dans le sol (pH 83) Dans les conditions environnementales Fine et al (2011) ont confirmeacute
lrsquoeffet de la saison sur la survie de M bovis dans le sol sur du maiumls et du foin avec une survie maximale
en hiverprintemps (feacutevrier-mai tempeacuterature moyenne 29degC radiations solaires 800 Wm2) une
survie de 34 jours en automnehiver (novembre-janvier tempeacuterature moyenne 04degC radiations
solaires 243 Wm2) et une survie minimale de 12 jours pendant la peacuteriode printempseacuteteacute (mai-aoucirct
tempeacuterature moyenne 217degC radiations solaires 1500 Wm2) De faccedilon contradictoire lrsquoeacutetude de
Young et al (2005) a mis en eacutevidence une dureacutee de survie optimale (60 jours) de M bovis BCG agrave 37degC
et minimale (16 jours) agrave 4degC
La survie de M bovis a eacutegalement eacuteteacute mise en eacutevidence par meacutethode culturale sur diffeacuterents
aliments du beacutetail (Palmer amp Whipple 2006 Fine Bolin et al 2011) Apregraves 112 jours la bacteacuterie est
cultiveacutee sur tous les substrats inoculeacutes conserveacutes agrave -20degC (betteraves pommes carottes pommes de
terre maiumls et foin) sur les pommes le maiumls le foin et les pommes de terre conserveacutes agrave 8degC et
uniquement sur les pommes le maiumls et les pommes de terre conserveacutes agrave 23degC (Palmer amp Whipple
2006) Fine et al (2011) ont eacutegalement montreacute que M bovis survivait plus longuement en
automnehiver sur du maiumls (43 jours) et du foin (37 jours) qursquoagrave la peacuteriode printempseacuteteacute (survie
infeacuterieure agrave 3 jours)
La bacteacuterie peut eacutegalement survivre entre 2 et 6 semaines dans des leacutesions tuberculeuses
macroscopiques de poumons et nœuds lymphatiques de buffle (Tanner amp Michel 1999) la survie
optimale eacutetant observeacutee en hiver dans des situations ombrageacutees et humides Des reacutesultats identiques
sont observeacutes pour des eacutechantillons drsquourine (3 jours en eacuteteacute vs 28 jours en hiver) et des seacutecreacutetions
bronchiques (7 jours vs 70 jours) (Morris et al 1994)
Les diffeacuterentes eacutetudes preacutesenteacutees ci-dessus deacutemontrent que M bovis survit un temps variable dans
Chapitre 1
54
lrsquoenvironnement une fois excreacuteteacute par son hocircte Ce temps est fonction de lrsquoexcreacuteta et du substrat qui le
reccediloit de sa charge bacteacuterienne et des conditions climatiques Il existe donc une peacuteriode appeleacutee
fenecirctre temporelle critique pendant laquelle les animaux qui interagissent avec les matrices
contamineacutees par des bacilles viables preacutesentent un risque accru drsquoinfection Comme nous lrsquoavons vu
preacuteceacutedemment la fenecirctre temporelle critique est fortement deacutependante du pays ou de la reacutegion
eacutetudieacutes et des conditions climatiques
Tregraves peu de donneacutees existent sur la conservation de la virulence de M bovis apregraves un passage
en dehors de son hocircte La virulence du bacille a eacuteteacute confirmeacutee par injection agrave des cobayes dans des
fegraveces de bovins naturellement contamineacutees apregraves 178 jours drsquoexposition dans une pacircture et apregraves 12
mois drsquoincubation en conditions de laboratoire (Maddock 1933) Lrsquoeacutetude reacutecente de Ghodbane et al
(2014) a deacutemontreacute que M tuberculosis M canetti et M bovis conservaient leur virulence apregraves un
seacutejour de 12 mois dans le sol En effet lrsquoinjection intra-peacuteritoneacuteale de suspensions bacteacuteriennes agrave des
souris a entraineacute lrsquoapparition de leacutesions avec M canetti (nodules sur les poumons le foie et la rate) et
M tuberculosis (nodules sur les poumons et la rate) Malgreacute lrsquoabsence de leacutesions lors de lrsquoinjection de
M bovis la bacteacuterie a pu ecirctre isoleacutee en quantiteacute importante des 3 organes Toutefois les injections
eacutetaient reacutealiseacutees avec les colonies de mycobacteacuteries obtenues apregraves culture sur milieu geacuteloseacute induisant
un biais important De plus lrsquoingestion par des souris de sol inoculeacute par M tuberculosis apregraves 12 mois
drsquoincubation a provoqueacute des leacutesions pulmonaires (1 souris5) et des leacutesions de la rate (25) mais la
bacteacuterie a pu ecirctre cultiveacutee agrave partir des poumons de la rate de lrsquoestomac et de lrsquointestin chez toutes les
souris
4 Strateacutegies de survie des mycobacteacuteries dans lrsquoenvironnement
Lrsquoenveloppe particuliegravere des mycobacteacuteries deacutecrite preacuteceacutedemment est probablement un des
facteurs majeurs de la reacutesistance de M bovis en dehors de son hocircte dans des conditions
environnementales deacutefavorables Drsquoautres strateacutegies de survie sont probablement mises en jeu par
M bovis pour persister dans lrsquoenvironnement mais sont totalement inconnues agrave ce jour Les
mycobacteacuteries pathogegravenes et opportunistes ont deacuteveloppeacute diffeacuterents meacutecanismes pour survivre agrave
lrsquointeacuterieur des cellules de lrsquohocircte strateacutegies qui peuvent eacutegalement ecirctre utiliseacutees en dehors de lrsquohocircte
Les meacutecanismes moleacuteculaires de certains processus sont eacutegalement en cours de compreacutehension Les
principales strateacutegies de survie sont preacutesenteacutees dans la partie suivante
a Lrsquoeacutetat de dormance
La dormance est deacutefinie chez les bacteacuteries non sporuleacutees comme un eacutetat physiologique
Chapitre 1
55
reacuteversible permettant la survie de la bacteacuterie pendant une longue peacuteriode sans croissance ni reacuteplication
(Kaprelyants et al 1993) Le programme geacuteneacutetique de la bacteacuterie lui permet drsquoentrer en dormance (en
anglais Non-Replicating Persistence) lorsque les conditions exteacuterieures deviennent deacutefavorables
(notamment en conditions oligotrophiques) puis de retrouver sa capaciteacute agrave se diviser lorsque les
conditions redeviennent favorables La dormance est deacutecrite chez plusieurs espegraveces de mycobacteacuteries
Lrsquoentreacutee en dormance de M smegmatis (Dick et al 1998) M bovis (Lim et al 1999 Boon et al 2001)
et M tuberculosis (Wayne amp Hayes 1996) en culture est provoqueacutee par la rareacutefaction de lrsquooxygegravene
Lrsquoeacutetat de dormance est suggeacutereacute chez Map et pourrait expliquer sa longue survie dans le fumier le lisier
et le sol (Whittington et al 2004) Lrsquoeacutetat de dormance est eacutegalement deacutecrit chez M avium (Archuleta
et al 2005)
Chez les mycobacteacuteries lrsquoentreacutee en eacutetat de dormance est controcircleacutee par le facteur de
transcription DosR (Dormancy Survival Regulator) (Boon amp Dick 2002) Composeacute de 48 gegravenes
(Schnappinger et al 2003 Flores Valdez amp Schoolnik 2010) son expression est activeacutee in vitro par
lrsquooxyde nitrique (NO) (Voskuil et al 2003) le monoxyde de carbone (CO) (Shiloh et al 2008) le deacuteficit
en nutriments (Rodriguez et al 2008) et lrsquohypoxie (Rodriguez et al 2008) Lrsquoexpression de ce reacutegulon a
eacuteteacute deacutemontreacutee lors drsquoinfection expeacuterimentale de macrophages (Schnappinger et al 2003 Rodriguez et
al 2008) et drsquoanimaux modegraveles (Voskuil et al 2003) pour M tuberculosis et dans des nœuds
lymphatiques de bovins positifs en IDR pour M bovis (Jimeacutenez et al 2013) Ce reacutegulon permettrait non
seulement la survie de la bacteacuterie pendant la phase de dormance mais aussi la transition de la
respiration aeacuterobie agrave lrsquoanaeacuterobiose et inversement (Leistikow et al 2010) A lrsquoheure actuelle le rocircle de
certaines proteacuteines codantes et drsquoARN non codants de ce reacutegulon demeure toujours inconnu Une
eacutetude geacutenomique comparative a permis de constater que le reacutegulon de la dormance eacutetait conserveacute
chez les diffeacuterentes espegraveces de mycobacteacuteries y compris chez les mycobacteacuteries environnementales
(Gerasimova et al 2011) Cette donneacutee suggegravere que la dormance est une reacuteponse adaptative
probablement tregraves ancienne chez les mycobacteacuteries pour survivre dans des environnements ougrave les
concentrations en oxygegravene sont fluctuantes (Boon amp Dick 2012) et que cette adaptation a eacuteteacute mise agrave
profit par les mycobacteacuteries pour survivre dans les cellules immunitaires des hocirctes qursquoelles infectent
Aucune eacutetude nrsquoest agrave ce jour disponible concernant lrsquoentreacutee en dormance potentielle de
M bovis dans lrsquoenvironnement Le sol constitue toutefois un environnement hostile ougrave facteurs
biotiques et abiotiques pourraient deacuteclencher le passage de la bacteacuterie drsquoun eacutetat actif agrave un eacutetat
dormant (hypoxie carence en nutriments tempeacuterature hellip)
b La sporulation
Lrsquoeacutevocation de la sporulation chez les mycobacteacuteries est tregraves controverseacutee Ghosh et al (Ghosh
Chapitre 1
56
et al 2009) ont deacutecrit la sporulation expeacuterimentale de M marinum sous forme drsquoendospores dans des
cultures liquides acircgeacutees de 2 mois et dans des cultures solides de M bovis BCG de 8 mois Malgreacute des
veacuterifications pheacutenotypiques biochimiques et geacutenomiques deacutemontrant que ces spores eacutetaient bien
issues de M marinum ces reacutesultats ont eacuteteacute contesteacutes par Traag et al (Traag et al 2010) qui ont eacutechoueacute
agrave les reproduire Lamont et al (Lamont et al 2012) ont pour la premiegravere fois mis en eacutevidence
lrsquoapparition de morphotype spore-like dans des cultures liquides de Map drsquoorigine bovine (K-10) ovine
(7565) et humaine acircgeacutees drsquoun an en conditions oligotrophiques Le seacutequenccedilage de lrsquoARNr 16S et
lrsquoamplification de lrsquoIS900 speacutecifique de Map ont confirmeacute que les colonies bacteacuteriennes issues des
spores correspondaient bien agrave Map Le chauffage des spores obtenues agrave 70degC pendant 30 minutes ne
semble pas affecter sa virulence sur les macrophages Des eacutetudes compleacutementaires sont neacutecessaires
pour confirmer ces observations A notre connaissance la sporulation nrsquoa jamais eacuteteacute eacutevoqueacutee pour les
mycobacteacuteries du MTBC
c Interactions des mycobacteacuteries avec la faune du sol
La microfaune et plus particuliegraverement les amibes libres sont largement preacutesentes dans
lrsquoenvironnement et colonisent les eacutecosystegravemes aquatiques le sol (Khan 2006 Rodriacuteguez-Zaragoza
1994) et lrsquoair Elles ont eacuteteacute eacutegalement isoleacutees chez lrsquohomme Elles vivent preacutefeacuterentiellement dans les
biofilms et aux interfaces eau-sol eau-air et eau-plantes (Rodriacuteguez-Zaragoza 1994) ougrave elles
laquo broutent raquo les bacteacuteries les champignons et les algues dont elles reacutegulent les communauteacutes (Huws
et al 2005) Les mycobacteacuteries environnementales sont largement preacutesentes dans les milieux
aquatiques (Primm et al 2004) ougrave elles peuvent interagir avec les amibes De nombreuses eacutetudes
rapportent la capaciteacute des mycobacteacuteries environnementales agrave survivre et se multiplier dans les
trophozoiumltes etou les kystes drsquoamibes (Adekambi et al 2006 Drancourt et al 2007 Ben Salah amp
Drancourt 2010 Krishna-Prasad amp Gupta 1978 Mura amp Bull 2006) Les amibes pourraient mecircme
constituer un veacuteritable reacuteservoir pour M xenopi (Drancourt et al 2007 Thomas et al 2006)
La plupart des mycobacteacuteries pathogegravenes y compris M bovis sont capables de survivre dans
les amibes voire de srsquoy multiplier (Thomas amp McDonnell 2007 Mba Medie et al 2011 Greub amp Raoult
2004 Cirillo et al 1997 Adekambi et al 2006) La survie de M bovis a eacuteteacute confirmeacutee dans les
trophozoiumltes et les kystes drsquoAcanthamoeba castellanii jusqursquoagrave 5 mois apregraves infection en conditions
expeacuterimentales (Taylor et al 2003 Mardare et al 2013) La virulence vis-agrave-vis des macrophages
humains et des cellules intestinales de souris semble ecirctre exacerbeacutee par la reacuteplication intra-amibienne
(Cirillo et al 1997) En conditions naturelles lrsquoobservation de protozoaires isoleacutes de latrines de
blaireaux infecteacutes a mis en eacutevidence des BAAR sans pouvoir identifier preacuteciseacutement M bovis (Mardare
et al 2013) Des eacutetudes compleacutementaires sont neacutecessaires pour preacuteciser le rocircle de reacuteservoir eacuteventuel
Chapitre 1
57
des amibes pour M bovis Les interactions des mycobacteacuteries pathogegravenes avec les amibes pourraient
ecirctre du mecircme ordre que celles avec les macrophages de lrsquohocircte infecteacute notamment au niveau des
meacutecanismes de peacuteneacutetration (Danelishvili et al 2007) et de lrsquoinhibition de la fusion entre le phagosome
et le lysosome suggeacuterant qursquoune relation eacutevolutive des mycobacteacuteries avec les amibes auraient permis
aux mycobacteacuteries lrsquoacquisition de ces caracteacuteristiques de survie (Cirillo et al 1997)
La localisation intra-amibienne des mycobacteacuteries les protegravege des conditions
environnementales deacutefavorables des biocides tels que le chlore (Adekambi et al 2006 Whan et al
2006) des antibiotiques (Miltner amp Bermudez 2000) des diffeacuterents traitements comme lrsquoozonation de
lrsquoeau (Thomas et al 2008) et favorise leur disseacutemination et leur implication potentielle dans des
maladies humaines (Greub amp Raoult 2004) Lors de la peacuteneacutetration dans lrsquohocircte lrsquoamibe joue un rocircle de
laquo cheval de Troie raquo (Barker amp Brown 1994) proteacutegeant les bacteacuteries contre les premiegraveres deacutefenses de
lrsquohocircte
Lrsquoutilisation des amibes en co-cultures pour isoler des bacteacuteries reacutesistantes aux amibes dans
des eacutechantillons environnementaux et meacutedicaux a permis drsquoisoler des mycobacteacuteries drsquoeacutecosystegravemes
aquatiques (Thomas et al 2006 Thomas amp McDonnell 2007 Pagnier et al 2008) et de sols pollueacutes
(Wang et al 2006)
Les eacutetudes relatant les interactions entre M bovis et la meacuteso- et macrofaune sont rares Une
eacutetude reacutealiseacutee dans des eacutelevages bovins et caprins infecteacutes par Map a mis en eacutevidence la contamination
de 31 des vers de terre preacuteleveacutes sur les pacirctures freacutequenteacutees par les animaux (Fischer et al 2003)
Dans cette eacutetude Map a eacuteteacute retrouveacutee dans le corps et les fegraveces de vers de terre 1 et 2 jours apregraves
lrsquoingestion de bouses infecteacutees expeacuterimentalement suggeacuterant qursquoils peuvent probablement jouer un
rocircle de vecteurs dans la disseacutemination des mycobacteacuteries
Les eacutetudes disponibles bien que tregraves rares suggegraverent que la meacuteso- et macrofaune du sol
peuvent heacuteberger et probablement jouer un rocircle de vecteur des mycobacteacuteries Toutefois des eacutetudes
compleacutementaires sont indispensables concernant M bovis
d Interactions des mycobacteacuteries avec les diptegraveres
Le portage digestif et feacutecal de M bovis par des mouches communes (Musca domestica) en
contact avec des viscegraveres infecteacutes de bovins est deacutecrit La bacteacuterie survit de 12 jours sur leurs corps et
15 jours dans le tractus digestif de mouches mortes (Schlee 1957 citeacute par Kazda et al 2009)
e La formation de biofilms
Lrsquoaptitude agrave former des biofilms est connue pour de nombreuses espegraveces de mycobacteacuteries
Chapitre 1
58
environnementales agrave croissance rapide comme M smegmatis M fortuitum ou M abcessus ou agrave
croissance lente notamment M avium et M intracellulare (Falkinham et al 2001 Beumer et al 2010
Torvinen et al 2007) Lrsquoorganisation des mycobacteacuteries en biofilm leur confegravere une reacutesistance accrue
vis-agrave-vis des conditions deacutefavorables dans lrsquoenvironnement mais eacutegalement chez lrsquohocircte humain ougrave
elles peuvent coloniser les catheacuteters et provoquer des infections sanguines tregraves graves (El Helou et al
2013) Concernant les mycobacteacuteries du MTBC la formation de biofilms est deacutecrite pour
M tuberculosis dans des conditions expeacuterimentales tregraves particuliegraveres tregraves eacuteloigneacutees des conditions
naturelles (Kulka et al 2012) La formation drsquoagreacutegats (clumping ou cording) observeacutes lors de la culture
des bacteacuteries du MTBC en milieu liquide gracircce agrave une matrice extracellulaire riche en acides mycoliques
peut ecirctre assimileacutee agrave une forme organiseacutee de reacutesistance des bacteacuteries puisque sous cette forme les
bacteacuteries survivent agrave des concentrations tregraves eacuteleveacutees en antibiotique (Ojha et al 2008) Il a eacuteteacute montreacute
chez M tuberculosis que la formation de biofilms mettait en jeu des processus geacuteneacutetiques et
physiologiques diffeacuterents de la croissance planctonique (Ojha et al 2008) Toutefois comme il nrsquoa
jamais eacuteteacute deacutemontreacute que ces bacteacuteries se multiplient dans lrsquoenvironnement il semble peu probable
que les biofilms constituent une forme de reacutesistance pour M bovis
5 Facteurs de risque de contamination de lrsquoenvironnement et de transmission de Mycobacterium bovis aux animaux par lrsquoenvironnement contamineacute
Plusieurs eacutetudes portant sur les interactions entre bovins et faune sauvage ont montreacute que les
contacts directs eacutetaient tregraves peu freacutequents entre les bovins et les blaireaux (Benham amp Broom 1989
Drewe et al 2013 Boumlhm et al 2009) ainsi qursquoentre les cerfs les sangliers et les bovins (Cooper et al
2010 Kukielka et al 2013) La transmission indirecte de M bovis semble donc ecirctre le mode de
transmission preacutepondeacuterant entre la faune sauvage et les bovins voire entre les diffeacuterentes espegraveces
sauvages (Cowie et al 2015 Carrasco-Garcia et al 2015) Diffeacuterents facteurs de risque jouent un rocircle
sur ce mode de transmission qui neacutecessite un recouvrement des domaines vitaux des animaux
sauvages infecteacutes et excreacuteteurs avec les sites drsquoeacutelevage bovin (pacirctures et bacirctiments) Ces principaux
facteurs de risque sont deacutetailleacutes ci-dessous
a Risques lieacutes agrave lrsquoeacutecologie des espegraveces sauvages sensibles agrave Mycobacterium bovis
Concernant le blaireau et le sanglier les pacirctures peuvent constituer une grande partie de leur
domaine vital (Hutchings amp Harris 1999 Payne 2014) Elles sont utiliseacutees pour la recherche de
Chapitre 1
59
nourriture essentiellement des lombrics qui repreacutesentent une part importante de leur reacutegime
alimentaire (Tolhurst et al 2009 Cleary et al 2011 Baubet et al 2003) Une eacutetude reacutealiseacutee en Cocircte
drsquoOr a montreacute que la freacutequentation des pacirctures par les blaireaux est optimale au printemps peacuteriode
de mise au preacute des bovins par rapport agrave lrsquoautomne et agrave lrsquohiver Pour les sangliers la freacutequentation des
pacirctures est plus importante au printemps et en eacuteteacute (Payne 2014)
Concernant plus preacuteciseacutement les blaireaux ces derniers urinent et deacutefegravequent dans des latrines
pour deacutelimiter leurs territoires (Hutchings et al 2001) mais ils urinent eacutegalement sur les pacirctures
preacutefeacuterentiellement au niveau des lisiegraveres de forecirct des clocirctures et des haies qursquoils croisent au greacute de
leurs deacuteplacements dans un but de marquage (White et al 1993 Drewe et al 2013) Dans les pacirctures
il a eacuteteacute montreacute que le taux de contact entre bovins et latrines est eacuteleveacute (Hutchings amp Harris 1997 Drewe
et al 2013) Dans lrsquoeacutetude de Hutchings et Harris (1997) 48 des 50 vaches du troupeau ont cumuleacute 416
contacts avec 4 latrines preacutesentes dans la pacircture eacutetudieacutee et 33 animaux ont brouteacute sur les latrines (suivi
sur 1776 heures) Pour Drewe et al (2013) 1716 contacts avec 13 latrines ont eacuteteacute enregistreacutes pour les
33 bovins drsquoun troupeau sur les cinq mois de lrsquoeacutetude Toutefois le rang hieacuterarchique des bovins du
troupeau joue un rocircle important puisque les animaux domineacutes sont les plus enclins agrave brouter au
voisinage des latrines Enfin les bovins consomment lrsquoherbe au voisinage des latrines de faccedilon plus
importante lorsque lrsquoherbe de la pacircture est rase (en cas de surpacircturage ou de fauchage) (Hutchings amp
Harris 1997) Compte tenu du grand nombre de bacilles qui peuvent ecirctre excreacuteteacutes par la voie urinaire
du fort taux de contact bovinslatrines et de la consommation de lrsquoherbe sur les latrines par certains
bovins il semblerait que les latrines et plus particuliegraverement les zones de marquage urinaire dans les
pacirctures repreacutesentent un risque important de transmission indirecte de la maladie par inhalation ou
ingestion (Hutchings amp Harris 1999)
Bien qursquoil nrsquoexiste aucune eacutetude sur le sujet la contamination des pacirctures par des blaireaux
preacutesentant des eacutecoulements purulents de morsures a eacutegalement eacuteteacute envisageacutee (Gallagher amp Clifton-
Hadley 2000)
Les blaireaux vivent dans des terriers qursquoils creusent dans le sol en milieu forestier ou en
dehors notamment dans les pacirctures La contamination des sols drsquoentreacutee de terrier par M bovis
repreacutesente eacutegalement un risque pour les bovins qui peuvent se contaminer en explorant les entreacutees de
terrier localiseacutees dans les pacirctures par ingestion ou inhalation de la litiegravere expulseacutee des tunnels
(Benham 1985) De plus le sol excaveacute des tunnels par les blaireaux est attirant pour les bovins qui
peuvent en ingeacuterer pour combler des carences mineacuterales (Dewes 1996) ou srsquoy frotter la tecircte creacuteant
des aeacuterosols de poussiegravere potentiellement infectant (Phillips et al 2003) La contamination des sols de
terriers repreacutesente eacutegalement un risque important de transmission de lrsquoinfection dans le groupe social
de blaireaux qui partage le terrier Le sol du terrier constitue un biotope adeacutequat pour la survie de
M bovis agrave lrsquoabri des UV sans variation extrecircme de tempeacuterature et avec une humiditeacute relative
Chapitre 1
60
importante (Moore amp Roper 2003) Lrsquoentretien de la maladie dans un groupe social par le terrier
contamineacute a deacutejagrave eacuteteacute eacutevoqueacute (Rogers et al 2003)
La freacutequentation des pacirctures par les sangliers pour la recherche de nourriture constitue
eacutegalement un facteur de risque dans la transmission indirecte de la bacteacuterie aux bovins Lrsquoexploration
du sol avec leurs museaux agrave la recherche de lombrics larves et autre nourriture ainsi que les reliquats
de sol deacuteplaceacutes peuvent constituer un facteur de contamination potentiel de lrsquoenvironnement par la
deacuteposition drsquoexcreacutetas oro-nasaux infecteacutes
Les cervideacutes peuvent eacutegalement utiliser les preacutes pour se nourrir et ecirctre une source de
contamination indirecte pour les bovins (Brook amp McLachlan 2009)
b Risques lieacutes aux pratiques drsquoeacutelevage
Le seacutejour prolongeacute des bovins au preacute est susceptible drsquoaugmenter les contacts directs et
indirects avec la faune sauvage infecteacutee Lrsquoeacutelevage allaitant est caracteacuteriseacute par une longue peacuteriode de
pacircturage entre avril et octobre puis une hivernation en bacirctiments au moment des vecirclages Certaines
races rustiques notamment la race salers qui ont repeupleacute une partie des cheptels bovins infecteacutes
apregraves abattage total passent toute lrsquoanneacutee au pacircturage Ces longues peacuteriodes de pacircturage constituent
un facteur de risque non neacutegligeable pour les bovins qui partagent leur habitat avec des espegraveces
sauvages infecteacutees Le surpacircturage est eacutegalement un facteur de risque pour les bovins qui sont ameneacutes
agrave brouter les latrines de blaireaux et agrave ingeacuterer des quantiteacutes de sol potentiellement contamineacute plus
importantes (Hutchings amp Harris 1999)
Les zones drsquoabreuvement et de nourrissage des bovins dans la pacircture constituent des zones
attractives pour la faune sauvage notamment lrsquohiver lorsque les ressources alimentaires font deacutefaut
Le partage de fourrage entre bovins et cervideacutes est probablement une source de contamination des
bovins dans le Michigan (OrsquoBrien et al 2006 Brook amp McLachlan 2009 Brook et al 2012) drsquoautant plus
que la transmission indirecte de la maladie entre des cerfs et des veaux par le partage du foin a eacuteteacute
prouveacutee expeacuterimentalement (Palmer et al 2010 Palmer et al 2004) La freacutequentation des auges de
bovins et des zones de nourrissage au sol par les blaireaux qui viennent manger les reliquats
alimentaires est reacuteguliegraverement deacutecrite au Royaume-Uni ainsi que celle des reacuteserves de grains
drsquoensilage et de foin dans lesquels ils peuvent uriner et deacutefeacutequer (Garnett et al 2002 Tolhurst et al
2009 Ward et al 2010) Les pierres agrave sel souvent poseacutees au sol dans les preacutes attirent particuliegraverement
les cervideacutes (Payne et al 2015 Brook amp McLachlan 2009) Les abreuvoirs et les zones drsquoabreuvement
(trous drsquoeau ruisseau source) constituent eacutegalement un facteur de risque lorsqursquoils sont utiliseacutes par
les bovins et la faune sauvage la survie de la bacteacuterie excreacuteteacutee eacutetant optimale dans les zones humides
Le comportement de baignade des sangliers dans les trous drsquoeau et les souilles entraine une forte
Chapitre 1
61
freacutequentation de ces zones notamment en eacuteteacute (Payne 2014)
Finalement lrsquoeacutepandage de fumiers frais augmenterait le risque drsquoinfection drsquoun troupeau par
rapport agrave lrsquoeacutepandage drsquoengrais ou de fumiers non frais (stockeacutes au moins deux mois) les animaux se
contaminant par inhalation ou ingestion de substrats contamineacutes dans les deux mois suivant lrsquoeacutepandage
(Griffin et al 1996 Humblet et al 2009) Certaines preacuteconisations conseillent un temps de stockage
minimum des fumiers avant eacutepandage (au moins 6 mois suite aux reacutesultats de Scanlon et Quinn 2000a
et 2000b) ou leur traitement chimique (Scanlon amp Quinn 2000b Scanlon amp Quinn 2000a) un
compostage (McCallan et al 2014) voire lrsquointerdiction drsquoeacutepandage de fumiers pour les cheptels infecteacutes
(Phillips et al 2003)
c Risques lieacutes aux pratiques cyneacutegeacutetiques
Les zones drsquoagreacutegation des animaux sauvages telles que les zones drsquoagrainage ou de
nourrissage agrave points fixes et la reacutealisation de trous drsquoeau artificiels pour lrsquoabreuvement et la baignade
des animaux sauvages sont des zones agrave risque qui augmentent les interactions directes et indirectes
entre les animaux et donc les risques de transmission de M bovis entre eux (Corner 2006 Vicente et
al 2007 Delahay et al 2002 Coleman amp Cooke 2001) En Espagne lrsquoaugmentation des densiteacutes
drsquoonguleacutes sauvages (cervideacutes et sangliers) dans le Sud du pays a entraineacute le deacuteveloppement drsquoactiviteacutes
de chasse agrave but commercial amenant agrave la reacutealisation de parcs gigantesques pour contenir les animaux
sauvages Dans ces zones lrsquoagreacutegation des animaux sauvages de diffeacuterentes espegraveces (sangliers
cervideacutes cochons sauvages et bovins) autour des trous drsquoeau et des zones de nourrissage a favoriseacute la
transmission intra- et interspeacutecifique de la maladie (Vicente et al 2007 Cowie et al 2015) Chez le
sanglier la preacutevalence varie de 52 agrave 70 voire presque 100 dans certaines populations (Gortaacutezar et
al 2008 Naranjo et al 2008) Cowie et al (2015) ont montreacute que 40 agrave 66 des interactions indirectes
avaient lieu dans une fenecirctre temporelle de 3 jours en saison segraveche et 72 agrave 100 dans une fenecirctre
temporelle de 12 jours en saison humide Or dans ces zones la survie de M bovis a eacuteteacute estimeacutee agrave 3
jours pendant la saison segraveche et agrave 12 jours pendant la saison humide (Kukielka et al 2013) suggeacuterant
que si le pathogegravene est preacutesent dans une population lrsquoagreacutegation est un facteur de maintenance et de
disseacutemination de lrsquoinfection dans et entre les espegraveces De plus la preacutesence drsquoeacutelevages bovins agrave
proximiteacute de ces parcs de chasse entraine une freacutequentation importante des structures drsquoeacutelevage par
les animaux sauvages et un risque accru drsquoinfection pour les bovins des eacutelevages visiteacutes (Carrasco-
Garcia et al 2015)
Le nourrissage des cervideacutes dans le Michigan dans le but de maintenir les populations pour la
chasse est la cause du maintien de lrsquoinfection dans les populations sauvages qui se contaminent entre
eux via les aliments distribueacutes (OrsquoBrien et al 2006)
Chapitre 1
62
Comme nous lrsquoavons deacutejagrave dit preacuteceacutedemment les viscegraveres drsquoanimaux infecteacutes laisseacutes agrave la
disposition des animaux sauvages par les chasseurs peuvent repreacutesenter un risque important pour les
animaux charognards (Morris et al 1994)
d Risques lieacutes aux facteurs paysagers climatiques et peacutedologiques
La structure du paysage est un facteur qui influence le recouvrement des domaines vitaux des
animaux sauvages et des bovins La localisation des pacirctures ou des bacirctiments drsquoeacutelevage en lisiegravere de
forecirct augmente les risques drsquointeractions entre animaux sauvages et domestiques (Ward et al 2010)
lrsquointerface pacircture-forecirct repreacutesenteacutee par les lisiegraveres eacutetant un eacutecotone agrave risque pour lrsquoeacutemergence des
agents pathogegravenes multi-hocirctes (Boyard et al 2008) La fragmentation du paysage (preacutes champs forecircts
bosquets) augmente le deacutepocirct drsquourine par les blaireaux aux zones frontiegraveres entre deux milieux
augmentant le risque de contamination de ces sites (White et al 1993 Humblet et al 2009) Les lisiegraveres
sont tregraves freacutequenteacutees par les sangliers et les blaireaux (Payne 2014) et constituent une zone privileacutegieacutee
par les blaireaux pour eacutetablir leurs terriers et marquer leurs territoires par des mictions ou des latrines
(Hutchings et al 2002)
Comme nous lrsquoavons vu preacuteceacutedemment les conditions climatiques sont un facteur essentiel
dans la dureacutee de survie de M bovis dans les diffeacuterents substrats environnementaux contamineacutes et in
fine dans la transmission indirecte de la maladie aux animaux qui interagissent avec ces substrats Les
saisons influencent eacutegalement la freacutequentation des pacirctures par les animaux sauvages avec un risque
accru au printemps et en eacuteteacute vis-agrave-vis des blaireaux et des sangliers qui vermillent dans les pacirctures
notamment lors des nuits humides ougrave les vers de terre remontent en surface appeleacutees laquo nuits agrave
lombrics raquo (Kruuk 1978 Cleary et al 2011 Baubet et al 2003) Lors de temps plus sec voire de
seacutecheresse les points drsquoeau destineacutes aux bovins constituent des zones attractives pour la faune sauvage
(Vicente et al 2007 Santos Santos et al 2015 Barasona et al 2014) favorisant lrsquoagreacutegation des
animaux domestiques et sauvages
Le rocircle des facteurs peacutedologiques dans la survie de M bovis est peu connu agrave ce jour La texture
du sol qui conditionne la structure la porositeacute et le reacutegime hydrique du sol le pH et la matiegravere
organique disponible sont des facteurs importants dans la survie des bacteacuteries A notre connaissance
une seule eacutetude statistique reacutealiseacutee dans le Michigan a montreacute que la nature du sol et plus
preacuteciseacutement le pourcentage de sable eacutetait associeacute agrave un risque drsquoinfection plus eacuteleveacute chez les bovins
(Walter et al 2014) Une augmentation drsquo1 de la quantiteacute de sable dans le sol augmentait de 36
le risque pour des eacutelevages bovins drsquoecirctre infecteacutes Hormis cette eacutetude qui relie indirectement infection
animale et survie de la bacteacuterie dans le sol aucune eacutetude nrsquoest actuellement disponible sur le lien
direct entre nature du sol et survie de M bovis
Chapitre 1
63
A la lumiegravere des informations preacutesenteacutees dans ce chapitre il apparait qursquoune fois excreacuteteacutee en
dehors de son hocircte M bovis peut survivre longuement dans lrsquoenvironnement si les conditions sont
adeacutequates par la mise en jeu de strateacutegies de survie encore inconnues agrave lrsquoheure actuelle Les
connaissances sur les scheacutemas de circulation de la bacteacuterie entre les animaux infecteacutes et
lrsquoenvironnement (et vice et versa) srsquoameacuteliorent et permettent drsquoadapter les programmes de lutte contre
la maladie mais elles sont encore insuffisantes pour permettre la maitrise de tous les facteurs en jeu
Ces diffeacuterents facteurs sont scheacutematiseacutes dans la Figure 15
Figure 15 Repreacutesentation scheacutematique de la circulation de M bovis dans lrsquoenvironnement agricole (modifieacute drsquoapregraves Elliott et al 2014)
Chapitre 1
64
D PRINCIPAUX OUTILS DE DETECTION DE PHENOTYPAGE ET DE GENOTYPAGE APPLICABLES AU MTBC ET PLUS PARTICULIEREMENT A MYCOBACTERIUM BOVIS
Depuis la deacutecouverte du bacille de la tuberculose de nombreux outils ont eacuteteacute deacuteveloppeacutes pour
deacutetecter et discriminer les diffeacuterentes mycobacteacuteries du MTBC Ces meacutethodes microscopiques
pheacutenotypiques et biochimiques essentiellement deacuteveloppeacutees pour le diagnostic meacutedical ont eacuteteacute peu
agrave peu ameacutelioreacutees et associeacutees agrave de nouvelles techniques de deacutetection de caracteacuterisation et
drsquoidentification des bacilles telles que lrsquoamplification geacutenique et le seacutequenccedilage notamment ameacuteliorant
la rapiditeacute du diagnostic la discrimination des espegraveces la pertinence de lrsquoantibiotheacuterapie et permettant
le geacutenotypage des souches agrave des fins eacutepideacutemiologiques Adapteacutees agrave des eacutechantillons biologiques
divers tels que les crachats les liquides de lavage broncho-alveacuteolaire les tissus de biopsies chez
lrsquohomme ou les broyats de nœuds lymphatiques chez les bovins et les animaux sauvages ces outils ne
sont pas tous applicables aux eacutechantillons environnementaux heacutebergeant des communauteacutes
microbiennes tregraves complexes
1 La deacutetection des mycobacteacuteries par microscopie
La microscopie optique
Lrsquoexamen microscopique de frottis ou drsquoeacutetalement drsquoeacutechantillons biologiques est la premiegravere
eacutetape dans le diagnostic bacteacuteriologique de la tuberculose humaine Elle est parfois la seule meacutethode
de diagnostic dans les pays en voie de deacuteveloppement Les proprieacuteteacutes tinctoriales particuliegraveres de la
mycomembrane des mycobacteacuteries sont exploiteacutees dans les colorations utiliseacutees La coloration de
reacutefeacuterence est la coloration de Ziehl et Neelsen Cette coloration agrave chaud est baseacutee sur la reacutesistance de
la paroi des mycobacteacuteries agrave la deacutecoloration agrave lrsquoacide et lrsquoalcool apregraves traitement agrave la fuschine Les
mycobacteacuteries apparaissent comme des bacirctonnets rouges sur un fond bleu Des meacutethodes agrave froid
baseacutees sur les mecircmes reacuteactifs existent eacutegalement (meacutethode de Kinyoun) La coloration agrave lrsquoauramine
un fluorochrome permet drsquoobserver en microscopie agrave fluorescence les bacilles en vert-jaune brillant
sur un fond jaune-orangeacute Cette coloration est plus sensible que la coloration de ZN (Steingart et al
2006)
Ces meacutethodes de coloration ne permettent pas de discriminer les mycobacteacuteries entre elles et
ne sont pas speacutecifiques du genre Mycobacterium puisqursquoelles peuvent colorer drsquoautres actinobacteacuteries
comme les genres bacteacuteriens Nocardia Rhodococcus et Corynebacterium (Shinnick amp Good 1994)
Lrsquoutilisation de la microscopie optique nrsquoest pas deacutecrite pour lrsquoanalyse des eacutechantillons
environnementaux drsquoune part car le seuil de deacutetection est eacuteleveacute (de lrsquoordre de 104 BAARmL) et drsquoautre
Chapitre 1
65
part car la coloration est difficile agrave mettre en œuvre sur des matrices telles que du sol ou des matiegraveres
feacutecales
Lrsquohybridation avec des sondes nucleacuteotidiques FISH (Fluorescence in situ hybridization)
Cette meacutethode est baseacutee sur lrsquohybridation speacutecifique drsquoune sonde marqueacutee avec un
fluorochrome sur une reacutegion cible de lrsquoADN bacteacuterien Les sondes utiliseacutees ADN ARNm ou
oligonucleacuteotides syntheacutetiques (PNA peptide nucleic acid) drsquoenviron 20 bases peacutenegravetrent agrave lrsquointeacuterieur
des cellules preacutealablement permeacuteabiliseacutees ougrave elles srsquohybrident speacutecifiquement avec leur ADN cible
Lrsquoobservation se fait agrave lrsquoaide drsquoun microscope agrave eacutepifluorescence
La meacutethode FISH a surtout eacuteteacute deacuteveloppeacutee pour la deacutetection des mycobacteacuteries dans les
crachats dans des cultures liquides chez des animaux infecteacutes expeacuterimentalement (Hongmanee et al
2001 Stender Mollerup et al 1999 Stender Lund et al 1999) et dans des eacutechantillons biologiques
(Stender Mollerup et al 1999 Lefmann et al 2006 Rodriguez-Nuntildeez et al 2012) La meacutethode FISH
est peu utiliseacutee pour la recherche de mycobacteacuteries dans des eacutechantillons environnementaux mais elle
a permis de mettre en eacutevidence Maa et Map dans de lrsquoeau et des biofilms de canalisations (Lehtola et
al 2006 Torvinen et al 2007) Elle parait toutefois difficilement applicable agrave des matrices telles que le
sol agrave cause de lrsquoauto-fluorescence geacuteneacutereacutee par les particules de sol et du risque de non-speacutecificiteacute des
sondes
2 Les meacutethodes de culture des mycobacteacuteries
Le diagnostic de la tuberculose est indissociable de la culture car historiquement crsquoest la
meacutethode la plus utiliseacutee Elle est sensible et permet drsquoisoler les souches de les caracteacuteriser et de tester
leur sensibiliteacute aux antibiotiques La recherche de mycobacteacuteries par des meacutethodes baseacutees sur la
culture est tregraves standardiseacutee en diagnostic humain et veacuteteacuterinaire tout comme les traitements
preacuteliminaires indispensables A lrsquoopposeacute aucune meacutethode standardiseacutee nrsquoest disponible pour la
recherche de mycobacteacuteries dans lrsquoenvironnement De nombreux protocoles emprunteacutes agrave la
microbiologie clinique ont eacuteteacute adapteacutes pour la recherche de mycobacteacuteries environnementales
notamment dans lrsquoeau (Narang et al 2009 Radomski et al 2011 Norby et al 2007 Pickup et al 2006
Whiley et al 2012 Le Dantec et al 2002 Bland et al 2005) Les meacutethodes utiliseacutees pour cultiver les
mycobacteacuteries pathogegravenes sont moins nombreuses et sont deacutecrites ci-dessous
a Les diffeacuterents traitements preacutealables agrave la mise en culture des eacutechantillons
De nombreuses meacutethodes de deacutecontamination des eacutechantillons biologiques ont eacuteteacute deacutecrites
dans le diagnostic meacutedical de la tuberculose Le choix de la meacutethode agrave utiliser deacutepend essentiellement
Chapitre 1
66
du type drsquoeacutechantillon analyseacute (tissu crachat lait fegraveces eau solhellip) des milieux de culture employeacutes et
de sa compatibiliteacute avec les meacutethodes moleacuteculaires Les meacutethodes les plus utiliseacutees en microbiologie
humaine meacutethodes de Petroff Taquet-Tison et Kubica utilisent la soude comme agent deacutecontaminant
principal (Petroff 1915 Taquet amp Tison 1961 Kubica et al 1963) Lrsquoutilisation drsquoacides comme agent
deacutecontaminant tels que lrsquoacide oxalique (David et al 1989) ou lrsquoacide sulfurique (Hohn amp Lowenstein
1926) ou drsquoammoniums quaternaires comme le chlorure de ceacutetyl pyridinium (Smithwick et al 1975)
ou le chlorure de benzalkonium (Krasnow amp Wayne 1966) est reacuteserveacutee aux eacutechantillons tregraves contamineacutes
(urines fegraveces) ou aux biopsies (cutaneacutees coliques pulmonaires etc) Ces meacutethodes ne sont pas toutes
compatibles avec la culture en milieu liquide et lrsquoamplification geacutenique
Dans le domaine veacuteteacuterinaire les tissus animaux (broyats de ganglions ou de tissus) sont
deacutecontamineacutes selon les preacuteconisations de lrsquoANSES et suivent la norme AFNOR NF U47-104 baseacutee sur
lrsquoutilisation de lrsquoacide sulfurique et de la soude agrave 4 Malgreacute la reacutesistance des mycobacteacuteries aux agents
chimiques la deacutecontamination reacuteduit fortement la population initiale preacutesente dans lrsquoeacutechantillon et
abaisse la sensibiliteacute de la culture (Fine Bolin et al 2011)
Concernant lrsquoisolement de bacteacuteries du MTBC dans les matrices environnementales ou les
excreacutetas animaux les principales meacutethodes utiliseacutees deacutecoulent des meacutethodes citeacutees preacuteceacutedemment
La meacutethode drsquoimmunocapture magneacutetique utiliseacutee par Sweeney (2006 et 2007) a permis lrsquoisolement
de M bovis dans des sols de terriers et des latrines contamineacutes naturellement Cette meacutethode est
baseacutee sur lrsquoutilisation de microbilles magneacutetiques recouvertes avec un ligand speacutecifique drsquoun antigegravene
de lrsquoenveloppe de M bovis Son avantage est qursquoelle permet la capture et la concentration de la bacteacuterie
cible dans un eacutechantillon complexe sans alteacuterer sa viabiliteacute contrairement agrave la deacutecontamination
chimique Cette meacutethode a eacutegalement eacuteteacute testeacutee avec succegraves sur des broyats de nœuds lymphatiques
de bovins (Stewart et al 2012 Stewart et al 2013 Garbaccio amp Cataldi 2010)
b La culture sur milieu solide
Les milieux de culture solides agrave lrsquoœuf (milieux de Loumlwenstein-Jensen et milieu de Coletsos) sont
les plus utiliseacutes en France dans le diagnostic bacteacuteriologique chez lrsquohomme et lrsquoanimal Ils sont sensibles
et reacutepondent aux exigences culturales de la plupart des mycobacteacuteries tuberculeuses et non
tuberculeuses Le milieu de Coletsos est particuliegraverement indiqueacute pour la culture des mycobacteacuteries
exigeantes telles que M bovis et M africanum
Les milieux geacuteloseacutes Middlebrook 7H10 et 7H11 sont des milieux semi-syntheacutetiques non utiliseacutes
en diagnostic de routine en France mais tregraves utiliseacutes aux Etats-Unis Enrichis avec de lrsquoOADC meacutelange
drsquoacide oleacuteique drsquoalbumine bovine de dextrose de chlorure de sodium et de catalase ils permettent
une croissance plus rapide de M tuberculosis (Mejia et al 1999) et de M bovis (Idigoras et al 1995)
Chapitre 1
67
que les milieux agrave base drsquoœuf Lrsquoajout de seacuterum bovin et de sang de mouton lyseacute au milieu 7H11 enrichi
en OADC ameacuteliore fortement la croissance de M bovis agrave partir drsquoeacutechantillons animaux et raccourcit le
deacutelai drsquoobservation des colonies (Gallagher amp Horwill 1977) Ce milieu a eacuteteacute utiliseacute pour la mise en
culture drsquoeacutechantillons environnementaux tels que le sol ou les fegraveces apregraves un traitement preacutealable de
lrsquoeacutechantillon (Sweeney et al 2006 Fine OrsquoBrien et al 2011) mais son manque de seacutelectiviteacute neacutecessite
lrsquoajout drsquoun cocktail drsquoantibiotiques souvent composeacute de lrsquoassociation drsquoune carboxypeacutenicilline de
trimeacutethoprime et de polymyxine B et drsquoun antifongique (amphoteacutericine B) (Sweeney et al 2007 Fine
OrsquoBrien et al 2011) Lrsquoincubation des milieux solides ensemenceacutes est reacutealiseacutee agrave 37degC pendant au moins
8 semaines
c La culture en milieu liquide
Les milieux liquides ont eacuteteacute deacuteveloppeacutes pour pallier la croissance lente des mycobacteacuteries
drsquointeacuterecirct clinique sur les milieux solides afin de confirmer preacutecocement le diagnostic Diffeacuterents
systegravemes manuels ou automatiseacutes existent agrave lrsquoheure actuelle Ils sont essentiellement utiliseacutes dans le
diagnostic chez lrsquohomme plus rarement chez lrsquoanimal
Le systegraveme Bactectrade 460 (Becton Dickinson) baseacute sur la respiromeacutetrie radiomeacutetrique nrsquoest plus
utiliseacute en France mais il lrsquoa eacuteteacute pendant une vingtaine drsquoanneacutees Il eacutetait baseacute sur la mesure du dioxyde
de carbone marqueacute au 14C rejeteacute par les mycobacteacuteries en croissance dans un milieu contenant de
lrsquoacide palmitique marqueacute au 14C La libeacuteration de 14CO2 eacutetait mesureacutee reacuteguliegraverement de faccedilon manuelle
gracircce agrave un appareil adapteacute La contrainte drsquoutilisation de stockage et de recyclage de produits
radioactifs ainsi que lrsquoabsence drsquoautomatisation ont conduit agrave lrsquoarrecirct de sa commercialisation Son
utilisation a eacuteteacute deacutecrite pour la deacutetection de M bovis sur des tissus animaux et des fegraveces (Corner et al
2012 Thacker et al 2015)
Pour eacuteviter les inconveacutenients lieacutes agrave la radioactiviteacute des meacutethodes non radiomeacutetriques ont eacuteteacute
deacuteveloppeacutees La croissance bacteacuterienne en milieu liquide est mise en eacutevidence soit par la
consommation de lrsquooxygegravene soit par le deacutegagement de CO2 dans le tube La mise en eacutevidence de la
diminution de la pression partielle drsquooxygegravene dissous dans le milieu de culture lors de la croissance
bacteacuterienne est utiliseacutee dans les tubes BBLtrade MGITtrade de Becton Dickinson Lorsque lrsquoO2 diminue les sels
de rutheacutenium preacutesents au fond du tube eacutemettent une fluorescence Lrsquoutilisation de systegravemes
automatiseacutes tels que les systegravemes Bactectrade MGITtrade assurent une lecture en continue de la fluorescence
eacutemise Le systegraveme Bactectrade MGITtrade a eacuteteacute utiliseacutee pour la recherche de M bovis dans des broyats de
nœuds lymphatiques de bovins (Hines et al 2006 Stewart et al 2013) des tissus drsquoanimaux sauvages
(Murphy et al 2010 Witmer et al 2010) et dans du fromage frais (Harris et al 2007) Lrsquoaugmentation
du CO2 dans le milieu de culture lors de la croissance bacteacuterienne est agrave la base du systegraveme BacTALERT
Chapitre 1
68
3D de Biomeacuterieux Cette meacutethode colorimeacutetrique est baseacutee sur la diminution du pH suite agrave la libeacuteration
du CO2 Lrsquoautomate eacutevalue en continu le virage de couleur drsquoune pastille fixeacutee au fond du tube Les
systegravemes deacutecrits permettent eacutegalement la recherche de reacutesistance agrave diffeacuterents antibiotiques
(Piersimoni et al 2006) A notre connaissance cette technique nrsquoa jamais eacuteteacute utiliseacutee pour la culture
drsquoeacutechantillons environnementaux
Ces diffeacuterents systegravemes utilisent comme milieu liquide le Middlebrook 7H9 suppleacutementeacute en
OADC ou en ADC (Albumine bovine dextrose catalase) pour ameacuteliorer la croissance des mycobacteacuteries
et rendu plus seacutelectif par lrsquoajout drsquoun meacutelange drsquoantibiotiques et drsquoantifongiques pour eacuteviter la
croissance de bacteacuteries indeacutesirables Le plus utiliseacute est le PANTAtrade (polymyxine B amphoteacutericine acide
nalidixique trimeacutethoprime azlocilline) Il peut ecirctre compleacuteteacute avec de la vancomycine (effet contre les
bacteacuteries gram +) Lrsquoavantage majeur de ces systegravemes en milieu liquide est la diminution du deacutelai moyen
de deacutetection des mycobacteacuteries Avec le systegraveme Bactectrade M tuberculosis est deacutetecteacutee entre 8 et 30
jours en fonction de la charge bacteacuterienne initiale de lrsquoeacutechantillon La deacutetection de M bovis est reacutealiseacutee
en 15 jours contre environ 45 jours sur milieu solide Lorsque la culture est deacuteclareacutee positive la
preacutesence de BAAR doit ecirctre veacuterifieacutee par coloration sub-culture sur milieu de Loumlwenstein-Jensen
identification biochimique ou moleacuteculaire Lrsquoassociation de la culture en milieu liquide et solide donne
les meilleurs reacutesultats avec une limite de deacutetection de 10 agrave 102 bacillesmL drsquoeacutechantillon (Guillet-Caruba
et al 2014)
NB A la fin du 19egraveme siegravecle et au deacutebut du 20egraveme la discrimination des espegraveces M bovis M tuberculosis
et M avium se faisait principalement par inoculation de lrsquoisolat bacteacuterien au cobaye au lapin et au
poulet Des leacutesions pulmonaires heacutepatiques et spleacuteniques importantes chez le cobaye et le lapin
identifiaient M bovis des leacutesions retrouveacutees uniquement chez le cobaye ou le poulet identifiaient
respectivement M tuberculosis et M avium (Kazda et al 2009) Cette meacutethode eacutetait utiliseacutee pour
lrsquoidentification de M bovis dans les tissus animaux et les eacutechantillons environnementaux comme le sol
et lrsquoeau apregraves deacutecontamination avec un meacutelange de javel et de soude (Maddock 1933 Maddock 1934
Maddock 1936 Williams amp Hoy 1930)
3 Meacutethodes de deacutetection et drsquoidentification des bacteacuteries du MTBC
Pour des raisons eacutepideacutemiologiques sanitaires (isolement du malade pour eacuteviter la
transmission) et theacuterapeutiques (mise en place drsquoune antibiotheacuterapie adeacutequate) il est primordial de
diffeacuterencier les membres du MTBC des autres mycobacteacuteries et drsquoidentifier preacuteciseacutement lrsquoespegravece du
MTBC en cause
Chapitre 1
69
a Caracteacuterisation pheacutenotypique et biochimique des mycobacteacuteries du MTBC
Depuis la deacutecouverte du bacille de la tuberculose jusqursquoau deacuteveloppement des outils
modernes les diffeacuterentes espegraveces de mycobacteacuteries ont eacuteteacute identifieacutees gracircce agrave leurs caracteacuteristiques
pheacutenotypiques et biochimiques Certaines de ces caracteacuteristiques sont encore exploiteacutees agrave lrsquoheure
actuelle en compleacutement des meacutethodes de biologie moleacuteculaire plus rapides Lrsquoaspect des colonies sur
un milieu solide leur deacutelai drsquoapparition leur nombre leur aspect rugueux ou lisse et la preacutesence drsquoune
pigmentation ou non permet drsquoorienter lrsquoidentification vers une mycobacteacuterie du MTBC ou une
NTM Dans le MTBC par exemple M tuberculosis apparait en 20 jours sur milieux agrave lrsquoœuf sous forme
de colonies ivoire en chou-fleur drsquoaspect rugueux Les colonies de M bovis apparaissent tardivement
(6 agrave 10 semaines) elles sont non pigmenteacutees petites fines et lisses (Carbonnelle et al 2003) Les
principaux tests biochimiques utiliseacutes pour diffeacuterencier les diffeacuterentes espegraveces du MTBC sont le test agrave
la niacine la reacuteduction des nitrates lrsquohydrolyse du Tween la sensibiliteacute au TCH (acide thiophegravene-2-
carboxylique) agrave la cycloseacuterine et au pyrazinamide Les reacutesultats des observations pheacutenotypiques et
biochimiques permettant de diffeacuterencier les principales espegraveces du MTBC sont preacutesenteacutes dans le
Tableau 2 Les tests biochimiques neacutecessitent des cultures riches et ne sont donc pas reacutealisables avant
la croissance avanceacutee de la bacteacuterie Chronophages et fastidieux ces tests sont remplaceacutes par drsquoautres
techniques plus rapides
Tableau 2 Identification des principales mycobacteacuteries du MTBC par des observations pheacutenotypiques et des tests biochimiques
M tuberculosis M africanum M bovis M bovis BCG
Aspect des colonies Rugueuses Rugueuses Lisses Rugueuses
Pigmentation Non pigmenteacute Non pigmenteacute Non pigmenteacute Non pigmenteacute
Croissance Lente Lente Lente Lente
Niacin test + - - -
Reacuteduction nitrates + +- - -
TCH R S S S
Cycloseacuterine S S R R
Pyrazinamide S S R R
+ test positif - test neacutegatif R reacutesistant S sensible
b Lrsquoidentification des mycobacteacuteries par recherche de biomarqueurs
Diffeacuterentes techniques de recherche de biomarqueurs potentiels de M tuberculosis par
chromatographie ont eacuteteacute deacuteveloppeacutees reacutecemment La chromatographie en phase liquide (HPLC High-
Chapitre 1
70
Performance Liquid Chromatography) a eacuteteacute deacuteveloppeacutee pour lrsquoanalyse des acides mycoliques (Butler amp
Guthertz 2001) La chromatographie en phase gazeuse coupleacutee agrave de la spectromeacutetrie de masse (GC-
MS) est utiliseacutee pour identifier des acides gras speacutecifiques de la paroi de M tuberculosis comme lrsquoacide
tuberculosteacutearique ou lrsquoacide hexacosanoiumlque dans des crachats (Cha et al 2009 Kaal et al 2009)
Lrsquoidentification de biomarqueurs speacutecifiques permet de diffeacuterencier M tuberculosis des NTM en GC-
MS en diagnostic direct sur des crachats (Dang et al 2013)
Depuis quelques anneacutees la spectromeacutetrie de masse de type MALDI-TOF (matrix-assisted laser
desorption ionization-time of flight) est utiliseacutee pour lrsquoidentification rapide des mycobacteacuteries du
complexe tuberculosis et de la majoriteacute des autres mycobacteacuteries agrave partir de cultures en milieu solide
et liquide (El Kheacutechine et al 2011 Bille et al 2012 Lotz et al 2010 Biswas amp Rolain 2013) Cette
technique permet drsquoeacutetablir un spectre de masse des diffeacuterentes proteacuteines constituant la mycobacteacuterie
(profil peptidique obtenu apregraves digestion trypsique des proteacuteines totales) La comparaison du profil
proteacuteique obtenu avec des bases de donneacutees internationales de plus en plus eacutetoffeacutees permet
drsquoidentifier un grand nombre drsquoespegraveces de mycobacteacuteries Elle peut ecirctre eacutegalement utiliseacutee comme
meacutethode de spoligotypage (Shitikov et al 2012) Cette meacutethode est simple rapide reproductible et
neacutecessite des consommables peu coucircteux (hors achat et entretien de lrsquoappareil) La qualiteacute du profil
proteacuteique obtenu est toutefois fortement deacutependante de la qualiteacute du protocole drsquoextraction des
proteacuteines (Saleeb et al 2011) La technique MALDI-TOF est probablement une technique drsquoavenir pour
lrsquoidentification rapide des mycobacteacuteries agrave croissance lente (le deacutelai de croissance reste toutefois le
facteur limitant) et plus geacuteneacuteralement des microorganismes pathogegravenes drsquointeacuterecirct clinique dans les
eacutechantillons A lrsquoheure actuelle elle ne permet pas de diffeacuterencier les diffeacuterentes espegraveces du MTBC
Elle nrsquoest pour lrsquoinstant applicable qursquoagrave des isolats bacteacuteriens issus de culture et non agrave des eacutechantillons
complexes
c Lrsquoidentification des mycobacteacuteries par des meacutethodes drsquohybridation coupleacutees ou
non agrave la PCR
La technique baseacutee sur lrsquohybridation de lrsquoADN est une meacutethode sensible rapide et simple
drsquoemploi couramment utiliseacutee en bacteacuteriologie humaine Elle est reacutealiseacutee sur une culture liquide ou
solide La technique est baseacutee sur lrsquohybridation drsquoune sonde moleacuteculaire conjugueacutee agrave un marqueur
chimioluminescent compleacutementaire de lrsquoARN ribosomal de la bacteacuterie cible Le kit commercial
AccuProbe (Gen Probe) fournit diffeacuterentes sondes permettant drsquoidentifier les Complexes tuberculosis
avium gordonae kansaii (sans discrimination des espegraveces) ainsi que les espegraveces M avium et
M intracellulare
Le systegraveme le plus utiliseacute agrave lrsquoheure actuelle est le GenXpert MTBRIF (Cepheid) utilisant la PCR
Chapitre 1
71
en temps reacuteel multi-colorimeacutetrique Il utilise 5 sondes Beacon (sondes avec une structure en eacutepingle agrave
cheveux qui fluorescent lors de lrsquohybridation) speacutecifiques du locus rpoB (reacutegion de 81 pb deacuteterminant
la reacutesistance agrave la rifampicine) des membres du MTBC Lrsquointeacuterecirct de cette technique est agrave la fois pratique
puisque lrsquoopeacuterateur deacutepose lrsquoeacutechantillon deacutecontamineacute dans une cassette contenant tous les reacuteactifs
lyophiliseacutes lrsquoappareil gegravere toutes les autres eacutetapes et diagnostique car elle couple la deacutetection des
bacteacuteries du MTBC et la mise en eacutevidence drsquoune eacuteventuelle reacutesistance agrave la rifampicine antituberculeux
de premiegravere ligne chez lrsquohomme
Drsquoautres systegravemes tels que lrsquoINNO-Lipareg Mycobacteria v2 (Fujirebio) le GenoType MTBC et le
GenoType Mycobacterium CMAS (Hain Lifescience) sont baseacutes sur lrsquohybridation drsquoune sonde
speacutecifique drsquoun Complexe ou drsquoune espegravece sur la reacutegion intergeacutenique variable 16S-23S apregraves
amplification PCR Ils permettent drsquoidentifier les Complexes tuberculosis et avium ainsi que de 16 agrave 40
espegraveces de NTM
Ces techniques sont essentiellement utiliseacutees dans le diagnostic humain mais elles sont parfois
deacutecrites pour lrsquoidentification du Complexes tuberculosis en diagnostic animal (Oloya et al 2007 Pavlik
et al 2005 Bollo et al 1998 Lieacutebana et al 1995) Bien que deacutejagrave utiliseacutees sur des eacutechantillons
environnementaux comme lrsquoeau (Aronson et al 1999) elles sont agrave utiliser avec preacutecaution car des
erreurs drsquoidentification ont eacuteteacute rapporteacutees (croisement entre des mycobacteacuteries saprophytes et
pathogegravenes) (Turenne 2004) en outre elles ne permettent pas la discrimination des diffeacuterentes
espegraveces du MTBC
d Le seacutequenccedilage
Lrsquooutil geacutenotypique de reacutefeacuterence pour lrsquoidentification des diffeacuterentes espegraveces de bacteacuteries est
le seacutequenccedilage du gegravene codant lrsquoARN de la sous-uniteacute 16S du ribosome (gegravene rrs) De nombreux couples
drsquoamorces ont eacuteteacute proposeacutes dans la litteacuterature pour amplifier entiegraverement ou partiellement le gegravene rrs
avant seacutequenccedilage mais lrsquoidentification des mycobacteacuteries par cette meacutethode nrsquoest pas toujours
possible agrave cause de la forte conservation de cette seacutequence Afin drsquoameacuteliorer la discrimination de ces
espegraveces le seacutequenccedilage total ou partiel drsquoautres gegravenes cibles a eacuteteacute deacuteveloppeacute tels les gegravenes de meacutenage
ciblant rpoB codant la sous-uniteacute β de lrsquoARN polymeacuterase (Adeacutekambi amp Drancourt 2004 Lee et al 2000)
hsp65 codant une proteacuteine de reacuteponse au stress thermique de 65 kDa (McNabb et al 2004 Ringuet et
al 1999 Telenti et al 1993) gyrA et gyrB codant les sous-uniteacutes α et β de lrsquoADN gyrase (enzyme
intervenant dans lrsquoenroulement et le deacuteroulement de lrsquoheacutelice drsquoADN) recA intervenant dans la
recombinaison homologue de lrsquoADN et dans sa reacuteparation (Blackwood et al 2000) sodA codant la super
oxyde dismutase (Zolg amp Philippi-Schulz 1994) secA1 codant la proteacuteine SecA1 intervenant dans le
seacutecreacutetion des proteacuteines agrave travers la membrane plasmique (Zelazny et al 2005) ou la reacutegion inter-
Chapitre 1
72
geacutenique encadreacutee par les ARNr 16S et 23S (appeleacutee ITS Internal Transcribed Spacer) (Roth et al 1998)
Malgreacute une ameacutelioration de la discrimination par lrsquoanalyse combineacutee de ces diffeacuterentes cibles le
seacutequenccedilage ne permet pas drsquoidentifier toutes les diffeacuterentes bacteacuteries du MTBC A cause de la forte
conservation des seacutequences dans le MTBC le seacutequenccedilage drsquoun grand nombre de gegravenes est neacutecessaire
pour discriminer les espegraveces (Kirschner et al 1993 Boumlddinghaus et al 1990) ce qui nrsquoest pas reacutealisable
en routine
Depuis lrsquoidentification de la plupart des gegravenes et des mutations responsables de la reacutesistance
des mycobacteacuteries aux antituberculeux le seacutequenccedilage est un des moyens utiliseacute pour identifier les
souches reacutesistantes La reacutesistance agrave la rifampicine par exemple est lieacutee agrave plusieurs mutations sur le gegravene
rpoB modifiant la structure de la sous-uniteacute β de lrsquoARN polymeacuterase et empecircchant la fixation de la
rifampicine sur lrsquoARN polymeacuterase conduisant agrave son inefficaciteacute (Zhang amp Yew 2009) Ces mutations
peuvent ecirctre deacutetecteacutees par seacutequenccedilage du gegravene amplifieacute par PCR
e Deacutetection et geacutenotypage des mycobacteacuteries du MTBC les marqueurs utiliseacutes
Les seacutequences drsquoinsertion
Les seacutequences drsquoinsertion (IS) correspondent agrave de petits eacuteleacutements geacuteneacutetiques transposables
Elles contiennent uniquement les informations geacuteneacutetiques neacutecessaires agrave leur transposition dans le
geacutenome de la bacteacuterie (transposase) On deacutenombre 42 IS dans le premier geacutenome annoteacute de M bovis
AF212297 (Garnier et al 2003) La seacutequence drsquoinsertion IS6110 speacutecifique du MTBC est la cible
historique utiliseacutee pour la deacutetection moleacuteculaire du Complexe tuberculosis en PCR (Thierry Cave et al
1990 Thierry Brisson-Noeumll et al 1990 Dziadek et al 2001) La meacutethode de reacutefeacuterence pour le
geacutenotypage de M tuberculosis est la RFLP-IS6110 (Restriction Fragment Lenght Polymorphism sur
lrsquoIS6110) Crsquoest la seacutequence drsquoinsertion la plus abondante dans le geacutenome de M tuberculosis qui en
contient de 1 agrave plus de 25 copies (Thierry Cave et al 1990 Thierry Brisson-Noeumll et al 1990) Elle
jouerait un rocircle dans lrsquoadaptation des bacteacuteries du MTBC agrave leur hocircte et dans leur virulence (Otal et al
2008 Safi et al 2004)
De nombreux systegravemes drsquoamorces ont eacuteteacute deacuteveloppeacutes pour la deacutetection de lrsquoIS6110 par PCR
dans les eacutechantillons biologiques (Dziadek et al 2001 El Kheacutechine et al 2009 Lieacutebana et al 1996
Savelkoul et al 2006) Diffeacuterents kits commerciaux utiliseacutes agrave lrsquoheure actuelle sont baseacutes sur la deacutetection
de lrsquoIS6110 notamment dans le diagnostic animal (Courcoul et al 2014) Cette cible a deacutejagrave eacuteteacute utiliseacutee
pour la deacutetection de M bovis dans des eacutechantillons inoculeacutes en PCR nicheacutee (Adams et al 2013) ainsi
que pour la deacutetection moleacuteculaire du MTBC dans du sol de lrsquoeau des seacutediments et des matiegraveres feacutecales
drsquoanimaux (Santos Santos et al 2015) (Tableau 3)
Chapitre 1
73
LrsquoIS6110 est eacutegalement utiliseacutee pour le typage des bacteacuteries du MTBC La comparaison des
souches repose sur le nombre drsquoIS6110 et leurs positions sur le chromosome apregraves extraction de lrsquoADN
digestion par une endonucleacutease (PvuII qui coupe lrsquoIS6110 sur un seul site) eacutelectrophoregravese transfert
sur membrane et hybridation (Van Soolingen et al 1991) Chaque fragment contenant une copie de
lrsquoIS6110 produit une bande (Figure 16) Cette meacutethode standardiseacutee beacuteneacuteficie de banques de donneacutees
de profils drsquohybridation internationales qui permettent des investigations eacutepideacutemiologiques Cette
meacutethode est beaucoup moins discriminante sur les souches agrave faible nombre drsquoIS (Allix et al 2006)
comme M bovis qui nrsquoen contient qursquoune agrave cinq copies et M bovis BCG qui nrsquoen contient qursquoune seule
copie (Van Soolingen et al 1991 Skuce et al 1994) Le reacutesultat est obtenu en une dizaine de jours
neacutecessite un volume important de culture et des manipulations fastidieuses De ce fait il a eacuteteacute
remplaceacute par des meacutethodes plus rapides indeacutependantes de la culture le spoligotypage et le typage
MLVA
Figure 16 Profils de restriction disolats de M tuberculosis obtenus suite agrave une RFLP-IS6110 Les isolats 3 5 6 9 et 10 ont le mecircme profil et posseacutedaient un lien eacutepideacutemiologique S repreacutesente le marqueur de poids moleacuteculaire (Source web site CDC)
Les courtes seacutequences drsquoADN reacutepeacuteteacutees
Le locus DR
Les souches du Complexe tuberculosis contiennent une reacutegion chromosomique particuliegravere
unique et figeacutee caracteacuteriseacutee par des seacutequences reacutepeacuteteacutees de 36 pb appeleacutee locus DR (Direct Repeat)
qui appartient agrave la famille des ADN reacutepeacutetitifs CRISPR (Clustered Regularly Interspersed Short
Chapitre 1
74
Palindromic Repeats) Ces DR sont seacutepareacutes par des espaceurs de 35 agrave 41 pb dont les seacutequences sont
uniques et polymorphes Ce polymorphisme associeacute agrave la deacuteleacutetion de certains espaceurs par des
reacutearrangements geacuteneacutetiques est exploiteacute dans le spoligotypage (en anglais spoligotyping
SPacer OLIGOnucleacuteotide TYPING) (Kamerbeek et al 1997) Par exemple les espaceurs 3 9 16 et 39 agrave
43 sont absents chez M bovis M caprae M microti et M pinnipedii (Smith et al 2006) comme
preacutesenteacute dans la Figure 17 Le geacutenotypage par spoligotypage est baseacute sur lrsquoamplification par PCR du
locus DR le marquage des amplicons et leur hybridation par des oligonucleacuteotides fixeacutes agrave une
membrane (spoligotypage 2D) Le profil final est deacutetecteacute par chimioluminescence (Kamerbeek et al
1997) Dans la technologie Luminex les oligonucleacuteotides sont fixeacutes sur des microbilles agrave signature
spectrale diffeacuterente (spoligotypage 3D) (Zhang et al 2010) Le spolygotypage standard analyse 43
espaceurs et permet drsquoobtenir des donneacutees eacutepideacutemiologiques et phylogeacuteneacutetiques deacuteterminantes (Sola
et al 2001) Il est largement utiliseacute pour typer les souches de M bovis notamment en France (Haddad
et al 2001) Cette meacutethode est reacutealisable sur de faibles quantiteacutes drsquoADN mecircme impur et des bacteacuteries
non cultivables Crsquoest une technique robuste rapide simple de reacutealisation dont le reacutesultat est facile agrave
interpreacuteter car il fournit un code binaire (pour chaque espaceur preacutesent 1 ou absent 0) faisant lrsquoobjet
drsquoune convention internationale (Dale et al 2001) Toutefois le spoligotypage ne preacutesente pas un
pouvoir de reacutesolution suffisant pour la diffeacuterenciation de certains groupes clonaux car il ne cible qursquoun
seul locus sur lrsquoensemble du geacutenome
Chapitre 1
75
C
Figure 17 Principe du spoligotypage des bacteacuteries du MTBC dapregraves Kamerbeek et al (1997) A Repreacutesentation du locus DR de M tuberculosis H37Rv et M bovis BCG B Amplifications des espaceurs par un couple damorces (a et b) et amplicons obtenus C Profils drsquohybridation sur membrane (un carreacute noir signale la preacutesence de lrsquoespaceur les parties blanches leur absence)
76
Gegravene ou locus
cible Espegraveces cibles PCR
Taille du
produit PCR
(pb)
Limites de deacutetection
(bacteacuteries g-1 sol
ou mL-1)
Matrices analyseacutees
Quantification si
deacutetection positive (nb
gegravenes ou UFC g-1 sol
ou mL-1)
Reacutefeacuterence Utiliseacute par
mpb64 MTBC PCR classique 543 101-102 Sols (terriers et preacutes
adjacents) 10 - 25 times 103 (Gormley 1999) (Young et al 2005)
mpb70 MTBC PCR classique 471 101-102 Sols (terriers et preacutes
adjacents) 10 - 25 times 103 (Gormley 1999)
(Young et al 2005)
(Courtenay et al 2006)
Gegravene codant
lrsquoARNr 16S
Mycobacteacuteries
agrave croissance
lente
RT-PCR 605 102-103 Sols (terriers et preacutes
adjacents) -
(Young et al
2005)
(Young et al 2005)
(Courtenay et al 2006)
IS6110 MTBC PCR nicheacutee PCR1 252
PCR2 116 -
Sol Eau
Foin Maiumls -
(Adams et al
2013) (Adams et al 2013)
IS6110 MTBC PCR nicheacutee 110
Sol 40 times 104
Eau 50 times 105
Sol
Seacutediment
Eau et fegraveces
(Soo et al 2006) (Santos Santos et al 2015)
RD12 M bovis
M caprae PCR classique 306
Sol 10 times 106
Eau 50 times 105
Sol
Seacutediment
Eau et fegraveces
(Warren et al
2006) (Santos Santos et al 2015)
RD12
M microti
M tuberculosis
M africanum
M pinnipedii
PCR semi-
nicheacutee 369
Sol 40 times 104
Eau 50 times 105
Sol
Seacutediment
Eau et fegraveces
(Warren et al
2006) (Santos Santos et al 2015)
RD4 (zone
flanquante) M bovis qPCR 142
42 times 105
(Pontiroli et al 2011)
10 times 105
(Pontiroli et al 2011)
Sols de terriers
Latrines
68 times 104 agrave 54 times 106
(Sweeney et al 2007)
20 times 104 agrave 10 times 108
(Travis et al 2011)
(Sweeney et al
2007)
(Sweeney et al 2007)
(King Murphy James Travis
Porter Sawyer et al 2015)
(King Murphy James Travis
Porter Hung et al 2015)
(Pontiroli et al 2011)
(Roug et al 2014)
Rv1510 (RD4) MTBC sauf
M bovis
PCR classique 1 033 - Sols de terriers
Latrines
- (Huard et al
2003)
(Courtenay et al 2006)
(Sweeney et al 2006)
(Sweeney et al 2007)
Tableau 3 Cibles moleacuteculaires deacutecrites dans la litteacuterature pour la deacutetection du Complexe tuberculosis ou de M bovis dans les eacutechantillons environnementaux
Chapitre 1
77
Les mini-satellites
Les mini-satellites correspondent agrave de courtes seacutequences drsquoADN reacutepeacuteteacutees (de 10 agrave 1 000 pb) et
conseacutecutives Pour pallier le manque de discrimination du spoligotypage un typage sur diffeacuterents loci
a eacuteteacute deacuteveloppeacute le MLVA (Multi Locus VNTR Analysis) baseacute sur lrsquoanalyse drsquoeacuteleacutements situeacutes sur des
seacutequences intergeacuteniques (les MIRU Mycobacterial interspersed Repetitive Unit) composeacutes de
seacutequences reacutepeacuteteacutees en tandem dont le nombre et la seacutequence sont variables (notamment les VNTR
Variable Number Tandem Repeat et les ETR Exact Tandem Repeat) (Supply et al 2001 Supply et al
2006) Lrsquoeacutetude de loci disperseacutes dans le geacutenome apporte une discrimination robuste des souches Le
geacutenotypage des souches de M tuberculosis ou M bovis par la meacutethode MIRU-VNTR repose sur la
recherche du nombre drsquoexemplaires des seacutequences reacutepeacuteteacutees de chaque locus eacutetudieacute Plus le nombre
de loci analyseacutes est important plus la discrimination est fine ainsi le MIRU-VNTR peut porter sur 12
15 ou 24 loci pour le typage des souches de M tuberculosis selon lrsquoobjectif envisageacute (eacutepideacutemiologie ou
phylogeacutenie) (Supply et al 2001 Supply et al 2006) Concernant le typage des souches animales de
M bovis diffeacuterentes combinaisons de marqueurs VNTR ont eacuteteacute eacutevalueacutees (Roring et al 2002 Allix et al
2006 Skuce et al 2002 Boniotti et al 2009) et ont montreacute que lrsquoanalyse de 5 agrave 8 loci suffisait pour
obtenir un pouvoir de discrimination supeacuterieur agrave 90 En France 8 loci ont eacuteteacute seacutelectionneacutes sur leur
polymorphisme et leur pouvoir de discrimination et utiliseacutes pour typer plus de mille isolats cliniques
recueillis entre 1978 et 2011 (Hauer et al 2015) Dans cette eacutetude les reacutesultats de geacutenotypage se
preacutesentent sous forme drsquoun code agrave 8 chiffres (Tableau 4) Pour un mecircme spoligotype SB0120 le
geacutenotypage MIRU-VNTR permet de diffeacuterencier les souches retrouveacutees en Cocircte drsquoOr de celles de
Dordogne Ces reacutesultats ont eacutegalement permis de deacutemontrer lrsquohomogeacuteneacuteiteacute des profils geacuteneacutetiques
retrouveacutes chez les animaux domestiques et sauvages dans une zone geacuteographique confirmant
lrsquohypothegravese de la transmission interspeacutecifique de lrsquoinfection
Tableau 4 Spoligotypes et geacutenotypes MIRU-VNTR des souches de M bovis isoleacutees chez les bovins et la faune sauvage en Cocircte dOr et en Dordogne (drsquoapregraves Hauer et al 2015)
Spoligotype Geacutenotype MIRU-VNTR (8 loci cibleacutes)
Origine des isolats 2165 2461 0577 580 2163a 2163b 4052 3232
SB0120
Cocircte drsquoOr 5 5 4 3 11 4 5 6
Bovins blaireaux
sangliers
SB0120
Dordogne 5 3 5 3 9 4 5 6
Bovins blaireaux
sangliers
SB0134
Cocircte drsquoOr 6 4 5 3 6 4 3 6
Bovins blaireaux
sangliers cerfs
Chapitre 1
78
La combinaison du spoligotypage et de la MLVA optimise la discrimination des souches pour
les suivis eacutepideacutemiologiques et apporte des informations phylogeacuteneacutetiques inteacuteressantes Elle permet
eacutegalement de suivre lrsquoeacutemergence de variants clonaux chez un animal ou dans un troupeau infecteacute en
fonction du temps (Navarro et al 2015 Rodriguez-Campos et al 2014) Toutefois les loci agrave analyser
doivent ecirctre adapteacutes en fonction des complexes clonaux rencontreacutes dans la reacutegion ou le pays eacutetudieacutes
(Boniotti et al 2009 Hauer et al 2015) Diffeacuterentes bases de donneacutees internationales sont disponibles
pour la comparaison des reacutesultats de spoligotypage et de MIRU-VNTR comme SpolDB4 SITVITWEB
MIRU-VNTRplus et Mbovisorg (regroupe les spolygotypes des espegraveces deacuteleacuteteacutees pour RD9) (Couvin amp
Rastogi 2014 Smith amp Upton 2012)
Reacutegions de Diffeacuterence (RD) et Single Nucleotide
Polymorphism (SNP)
Les Reacutegions de Diffeacuterences (RD1 agrave RD14) identifieacutees chez les bacteacuteries du MTBC correspondent
agrave des deacuteleacutetions et des reacutearrangements geacuteneacutetiques irreacuteversibles et discriminants Ces RD sont
speacutecifiques drsquoune espegravece ou communes agrave plusieurs espegraveces Le plus grand nombre de RD est observeacute
dans le geacutenome de M bovis BCG qui preacutesente 14 RD par comparaison agrave M tuberculosis H37Rv (Gordon
et al 1999 Brosch et al 2002) En parallegravele six seacutequences deacuteleacuteteacutees chez M tuberculosis H37Rv (les
RvD) et une seacutequence absente de toutes les souches de M tuberculosis (TbD1) sont preacutesentes chez les
autres bacteacuteries du MTBC (Gordon et al 1999 Brosch et al 2002) Lrsquoabsence de RD4 est speacutecifique agrave
M bovis et M bovis BCG et permet de les diffeacuterencier des autres membres du MTBC (Huard et al 2003
Huard et al 2006) La RD4 a eacuteteacute utiliseacutee dans diffeacuterentes eacutetudes pour la deacutetection de M bovis dans les
eacutechantillons environnementaux (Courtenay et al 2006 Sweeney et al 2007 King Murphy James
Travis Porter Sawyer et al 2015 Travis et al 2011 Roug et al 2014 Pontiroli et al 2011) Dans une
eacutetude reacutecente Santos et al (2015) ont associeacute la deacutetection du MTBC par lrsquoIS6110 agrave une identification
de lrsquoespegravece par amplification de deux reacutegions de la RD12 pour diffeacuterencier M bovisM caprae des
autres espegraveces du Complexe (Tableau 3)
Lrsquointeacuterecirct des RD est bien plus phylogeacuteneacutetique que geacutenotypique puisque lrsquoeacutetude de leur
apparition successive au cours de lrsquoeacutevolution a permis drsquoeacutetablir preacuteciseacutement la phylogeacutenie des
diffeacuterentes espegraveces comme preacutesenteacute preacuteceacutedemment dans la Figure 3 (Huard et al 2006 Rodriguez-
Campos et al 2014 Dippenaar et al 2015)
Les Single Nucleotide Polymorphism (SNPs) correspondent agrave des mutations ponctuelles drsquoune
base dans un geacutenome Ils peuvent ecirctre synonymes (le changement de base nrsquoaffecte pas la seacutequence
proteacuteique correspondante) ou non synonymes (la seacutequence proteacuteique est affecteacutee) et repreacutesentent
Chapitre 1
79
une variabiliteacute du geacutenome utile pour la caracteacuterisation de la variabiliteacute drsquoun groupe clonal et lrsquoeacutetude
des relations phylogeacuteneacutetiques entre les souches (Mathema et al 2006)
f Identification par seacutequenccedilage complet des geacutenomes
Le seacutequenccedilage complet des geacutenomes (WGS Whole Genome Sequencing) beacuteneacuteficie depuis
quelques anneacutees de nouvelles technologies (NGS Next Generation Sequencing) plus rapides et moins
coucircteuses que la meacutethode de Sanger Diffeacuterentes technologies coexistent agrave lrsquoheure actuelle dont les
plus utiliseacutees sont le pyroseacutequenccedilage 454 (toutefois en fin drsquoutilisation) la technique Illumina et la
technique Ion PGM (Ion Personal Genome Machine) (Desikan amp Narayanan 2015) Ces techniques ont
reacutevolutionneacute le geacutenotypage en fournissant le plus haut niveau de discrimination geacuteneacutetique possible
Ces techniques parfois coupleacutees au typage des SNPs permettent de suivre la dynamique de
transmission drsquoune maladie de caracteacuteriser lrsquoantibioreacutesistance drsquoune souche et drsquoeacutevaluer la divergence
geacuteneacutetique de souches qui ont les mecircmes profils de restriction ou de geacutenotypage (Schuumlrch amp van
Soolingen 2012) Une eacutetude irlandaise sur des souches de M bovis isoleacutees de bovins et de blaireaux
vivant dans les mecircmes zones a permis gracircce au WGS et agrave lrsquoanalyse de 39 SNPs de deacutemontrer la
circulation et la persistance locales drsquoune souche entre animaux sauvages et domestiques (Biek et al
2012) Une eacutetude reacutecente portant sur le seacutequenccedilage complet de 82 souches de M bovis isoleacutees en
France entre 1978 et 2011 a permis de deacutemontrer la congruence entre le typage par
spoligotypageMIRU-VNTR et le typage des souches par analyse des SNP (Hauer 2015) suggeacuterant que
certains SNP pourraient ecirctre utiliseacutes comme cibles dans le geacutenotypage de M bovis Dans un futur
proche le WGS sera probablement lrsquooutil qui offrira lrsquoinformation la plus complegravete sur le geacutenotype des
souches les liens phylogeacuteneacutetiques entre souches et qui permettra de deacutecoder les scheacutemas de
transmission de lrsquoinfection entre animaux (Biek et al 2012 Pepperell et al 2013 Rodriguez-Campos et
al 2014)
Les puces (microarrays) agrave ADN ou ARN sont utiliseacutees pour la comparaison de geacutenomes complets
des espegraveces du MTBC et ont notamment apporteacute des informations sur lrsquoexpression de gegravenes de la
virulence chez M bovis (Voskuil et al 2004) ainsi que sur lrsquoidentification de seacutequences polymorphiques
distinguant les principaux complexes clonaux (Berg et al 2011 Muumlller et al 2009 Smith et al 2011)
Ces techniques appartiennent toutefois au domaine de la recherche et ne sont pas utiliseacutees en routine
g Meacutethode speacutecifique drsquoidentification des mycobacteacuteries animales utiliseacutee par le
Laboratoire National de reacutefeacuterence de la tuberculose bovine LNR TB
Lrsquoidentification des mycobacteacuteries isoleacutees sur des animaux est reacutealiseacutee au LNR TB de Maisons-
Chapitre 1
80
Alfort En premiegravere intention quatre systegravemes de deacutetection moleacuteculaires ciblant des seacutequences
drsquoinsertion (IS6110 IS1081 IS1245) et le gegravene atpE sont utiliseacutes en qPCR pour orienter lrsquoidentification
(Tableau 5)
Tableau 5 Systegravemes de deacutetection utiliseacutes en qPCR en premiegravere intention pour lidentification dune mycobacteacuterie dorigine animale et reacutesultats obtenus + amplification de la cible - absence drsquoamplification de la cible
ARNr 16S IS1245 IS901 IS6110 IS1081 IS1561rsquo RD4 RD7 atpE
Complexe tuberculosis
M bovis M tuberculosis
M microti M caprae
+ + + +
- - - -
- - - -
+ + + +
+ + + +
+ + - +
- + + +
- + + -
- - - -
Complexe avium + + +- - - - - + +
Autres mycobacteacuteries
+ - - - - - - + +
(Slana et
al 2010) (Slana et al
2010) (Courcoul et
al 2014) (cette thegravese)
(Radomski et al 2013)
Lrsquoidentification des espegraveces du complexe tuberculosis (IS6110 et IS1081 positives) est ensuite
reacutealiseacutee par geacutenotypage gracircce agrave la technique de spoligotypage (Zhang et al 2010) Lrsquoamplification ou
non des IS1245 et IS901 permet drsquoidentifier les espegraveces Mah (IS1245 et IS901 neacutegatives) Maa (IS1245
et IS901 positives) et Mah (IS1245 positive et IS901 neacutegative) (Slana et al 2010) Lrsquoidentification des
autres mycobacteacuteries repose sur le seacutequenccedilage de lrsquohsp65 puis de la comparaison par BLAST avec une
base de donneacutees (Telenti et al 1993)
De par le grand nombre drsquoespegraveces de mycobacteacuteries et la forte identiteacute des espegraveces dans le
Complexe MTBC la deacutetection et le geacutenotypage des souches du MTBC reposent obligatoirement sur
lrsquoutilisation conjointe de diffeacuterentes techniques En fonction des eacutechantillons disponibles et des
objectifs souhaiteacutes qursquoils soient diagnostiques eacutepideacutemiologiques ou phylogeacuteneacutetiques le choix des
techniques agrave utiliser doit ecirctre reacutefleacutechi et pertinent
82
Objectifs de la Thegravese
Depuis 2004 la situation de la Cocircte drsquoOr vis-agrave-vis de la tuberculose bovine est preacuteoccupante
Malgreacute la mise en place de moyens de surveillance et de lutte en premier lieu dans les cheptels bovins
puis dans la faune sauvage lrsquoeacuteradication de la maladie semble difficile Depuis 2011 on observe une
diminution du nombre des foyers bovins et une stabiliteacute du nombre de cas observeacutes dans la faune
sauvage avec une concentration spatiale des foyers dans des petites valleacutees Dans ces valleacutees ougrave
cohabitent faune sauvage et bovins la transmission indirecte du bacille entre animaux sauvages et
domestiques par le biais de lrsquoenvironnement est fortement suspecteacutee pour plusieurs cas bovins Bien
que plusieurs eacutetudes aient deacutemontreacute la preacutesence de M bovis dans diffeacuterentes matrices
environnementales naturelles sur les lieux de vie drsquoanimaux domestiques et sauvages infecteacutes
notamment en Angleterre et en Espagne aucune donneacutee nrsquoest disponible sur la contamination
environnementale potentielle par M bovis dans les zones endeacutemiques de tuberculose bovine en
France et plus particuliegraverement en Cocircte drsquoOr
Pour reacutepondre agrave cette question le premier objectif du travail de thegravese a eacuteteacute de deacutevelopper et de valider
sur des eacutechantillons inoculeacutes artificiellement des outils moleacuteculaires speacutecifiques et sensibles pour la
deacutetection des souches du MTBC et de M bovis dans les substrats environnementaux naturellement
contamineacutes tels que le sol lrsquoeau et les seacutediments et les matiegraveres feacutecales animales Nous avons
eacutegalement deacuteveloppeacute un protocole drsquoextraction-deacutecontamination des mycobacteacuteries du sol coupleacute agrave
une technique originale de suivi de croissance des mycobacteacuteries en milieu liquide dans lrsquoobjectif
drsquoisoler les souches environnementales de M bovis (Chapitre 2)
Les outils moleacuteculaires deacuteveloppeacutes ont eacuteteacute utiliseacutes pour la recherche de M bovis dans un grand nombre
drsquoeacutechantillons environnementaux (sol eau seacutediments fegraveces animales et membres de la micro- ou
meacuteso-faune des sols) preacuteleveacutes dans la zone endeacutemique de Cocircte drsquoOr et plus preacuteciseacutement dans des
zones partageacutees par la faune domestique et sauvage infecteacutee afin de preacuteciser le niveau de
contamination des lieux de vie des bovins et des animaux sauvages (Chapitre 3)
Face aux eacutetudes parfois controverseacutees sur le rocircle des facteurs abiotiques dans la persistance de
M bovis dans le sol nous avons rechercheacute lrsquoeffet de la nature du sol et de la tempeacuterature sur la dureacutee
de survie de M bovis SB0120 en conditions de laboratoire (Chapitre 4)
Nous avons enfin eacutetudieacute le rocircle de la faune du sol et plus particuliegraverement des lombrics tregraves abondants
dans les pacirctures et gros consommateurs de matiegravere organique dans la circulation et la disseacutemination
de M bovis dans les sols (Chapitre 5)
CHAPITRE 2
Deacuteveloppement et optimisation drsquooutils pour la deacutetection
de Mycobacterium bovis dans les eacutechantillons
environnementaux
PARTIE CONFIDENTIELLE agrave lrsquoexception des meacutethodes de deacutetection moleacuteculaire
86
CHAPITRE 2 - Deacuteveloppement et optimisation drsquooutils pour la
deacutetection de Mycobacterium bovis dans les eacutechantillons
environnementaux
A DEVELOPPEMENT DrsquoOUTILS MOLECULAIRES ADAPTES AUX MATRICES ENVIRONNEMENTALES
Les diffeacuterents systegravemes moleacuteculaires utiliseacutes pour la deacutetection de M bovis dans
lrsquoenvironnement citeacutes dans la litteacuterature sont syntheacutetiseacutes dans le Tableau 3 En consideacuterant la diversiteacute
et lrsquoabondance des mycobacteacuteries potentiellement preacutesentes dans lrsquoenvironnement nous avons
eacutelimineacute les systegravemes ciblant toutes les espegraveces du MTBC notamment lrsquoIS6110 car bien que tregraves
sensible et speacutecifique sur les eacutechantillons biologiques et tregraves utiliseacutee en diagnostic humain et
veacuteteacuterinaire nous avons montreacute qursquoelle nrsquoeacutetait pas utilisable en diagnostic environnemental En effet
nous avons effectueacute des essais preacuteliminaires avec le kit LSI VetMAXtrade Mycobacterium tuberculosis PCR
Kit (Life Technologies) utiliseacute en diagnostic veacuteteacuterinaire sur 25 eacutechantillons de sol provenant de zones
geacuteographiques non impacteacutees par la tuberculose bovine Pour tous ces sols lrsquoIS6110 a donneacute un signal
moleacuteculaire quantifiable teacutemoignant de lrsquoabsence de speacutecificiteacute de ce systegraveme pour la matrice sol
De plus M microti membre du MTBC agent de la tuberculose des petits rongeurs est susceptible
drsquoecirctre retrouveacute dans le sol (Courtenay et al 2006) et a deacutejagrave eacuteteacute isoleacute sur des feacutelins et des animaux
sauvages en Cocircte drsquoOr (Laprie et al 2013 Michelet et al 2015) Nous avons donc deacuteveloppeacute des
systegravemes moleacuteculaires qui nrsquoamplifient pas M microti
Enfin M bovis est une mycobacteacuterie pathogegravene de classe III qui neacutecessite une manipulation dans un
laboratoire de confinement L3 Pour des raisons techniques et pour srsquoaffranchir de la contrainte de
confinement dans la mise au point de la technique de PCR nous avons utiliseacute une des souches
atteacutenueacutees de M bovis et plus preacuteciseacutement M bovis BCG Pasteur 1173P2 manipulable dans un
laboratoire de type L2 disponible agrave lrsquoINRA de Dijon Nous avons donc choisi des cibles moleacuteculaires
eacutegalement preacutesentes chez M bovis BCG
Pour augmenter la speacutecificiteacute de la deacutetection moleacuteculaire dans les matrices environnementales
complexes nous avons choisi drsquoutiliser la PCR quantitative avec une sonde TaqManreg qui est formeacutee
par un nucleacuteotide lieacute de faccedilon covalente agrave un fluorophore (reporter) en 5rsquo et agrave un deacutesactivateur
(quencher) en 3rsquo Le clivage de la sonde par la Taq polymeacuterase apregraves lrsquohybridation agrave la seacutequence drsquoADN
compleacutementaire eacuteloigne le quencher du fluorophore qui eacutemet une fluorescence mesureacutee en temps
reacuteel dans un thermocycleur adapteacute
Chapitre 2
87
Le systegraveme moleacuteculaire deacutecrit preacuteceacutedemment par Sweeney et al (2007) ciblant les marges de
la reacutegion RD4 deacuteleacuteteacutee uniquement chez M bovis et M bovis BCG nous a particuliegraverement inteacuteresseacutes
Des amorces internes agrave la seacutequence RD4 ont eacuteteacute utiliseacutees en PCR pour la discrimination de M bovis
avec les autres membres du MTBC (pas de signal pour M bovis mais un signal pour les autres membres
du MTBC) (Huard et al 2003 Courtenay et al 2006) Des amorces de PCR classique ont eacutegalement eacuteteacute
dessineacutees sur les zones flanquantes de la reacutegion RD4 (Brosch et al 2002) Ces amorces ont eacuteteacute adapteacutees
agrave la technologie TaqManreg avec une amorce et une forward localiseacutees sur les marges de la seacutequence
deacuteleacuteteacutee et une sonde agrave cheval sur la zone de deacuteleacutetion (Sweeney et al 2007) La speacutecificiteacute et la
validation de ce systegraveme sur diffeacuterents eacutechantillons environnementaux tels que des sols de terriers
des fegraveces de blaireaux et de buffles africains ont eacuteteacute deacutemontreacutees dans plusieurs eacutetudes (Sweeney et al
2007 Roug et al 2014 Travis et al 2011 Pontiroli et al 2013 King et al 2015) Ce systegraveme de deacutetection
nous paraissant a priori speacutecifique pour la discrimination de M bovis dans le MTBC et adapteacute aux
matrices complexes nous lrsquoavons inclus dans lrsquoeacutetude Toutefois lrsquoamorce Reverse du systegraveme de
deacutetection a eacuteteacute modifieacutee par nos soins afin de reacuteduire lrsquoamplicon obtenu de 146 agrave 116 bases taille plus
adapteacutee agrave la technologie TaqManreg
Face au manque de systegravemes discriminants disponibles dans la litteacuterature nous avons dessineacute
deux nouveaux systegravemes Nous avons choisi de cibler la seacutequence drsquoinsertion IS1561rsquo (Rv3349c) car elle
est preacutesente chez tous les membres du MTBC sauf M microti (Gordon et al 1999 Huard et al 2003
Brodin et al 2002) Elle appartient agrave la famille des ISL3 qui regroupe des copies drsquoIS deacutefectueuses chez
M tuberculosis elles sont alors annoteacutees avec une apostrophe
La troisiegraveme cible a eacuteteacute choisie dans une reacutegion chromosomique de M tuberculosis appeleacutee
RD1 preacutesente chez les souches virulentes de M tuberculosis et M bovis mais partiellement deacuteleacuteteacutee
chez M bovis BCG (Mahairas et al 1996) et M microti (Brodin et al 2002) Cette reacutegion contient
notamment un cluster de 6 gegravenes (Rv3871 agrave Rv3876) qui fait partie des gegravenes codant les proteacuteines
antigeacuteniques ESAT-6 (Early Secreted Antigenic Target 6kD) et CFP-10 (Culture Filtrat Protein 10kD)
responsables de la reacuteponse immunitaire de lrsquohocircte M bovis BCG et M microti sont les deux seuls
membres du MTBC agrave preacutesenter une deacuteleacutetion de ce cluster deacuteleacutetion probablement en partie
responsable de leur absence de virulence et de leur utilisation en vaccination (Pym et al 2002) Chez
M bovis BCG la reacutegion aval de ce cluster est deacuteleacuteteacutee la reacutegion deacuteleacuteteacutee est appeleacutee RD1bcg (Figure 18)
Chez M microti crsquoest la reacutegion amont du cluster qui est absente la reacutegion deacuteleacuteteacutee est appeleacutee RD1mic
Nous avons plus particuliegraverement cibleacute le locus Rv3866 preacutesent chez M tuberculosis et dans toutes les
souches de M bovis et M bovis BCG mais absente chez M microti (Brodin et al 2002) M avium et
M smegmatis (Mahairas et al 1996)
Chapitre 2
88
Figure 18 Diffeacuterences geacutenomiques de la reacutegion RD1 entre M bovis BCG (A) M microti (C) par comparaison agrave M tuberculosis H37Rv (B) drsquoapregraves Brodin et al 2002 et localisation du locus Rv3866 RD1mic correspond agrave la seacutequence de M tuberculosis deacuteleacuteteacutee chez M microti RD1bcg correspond agrave la seacutequence de M tuberculosis deacuteleacuteteacutee chez M bovis BCG
Les seacutequences drsquointeacuterecirct ont eacuteteacute rechercheacutees pour les membres du MTBC et certaines MNT sur
la base de donneacutees GenBank Un alignement de ces seacutequences a eacuteteacute reacutealiseacute gracircce au portail drsquoanalyses
bioinformatiques MOBYLE PASTEUR pour rechercher les reacutegions conserveacutees et les reacutegions de
diffeacuterence Le dessin des oligonucleacuteotides a eacuteteacute reacutealiseacute grace au logiciel en ligne Primer3
(httpbioinfouteeprimer3-040) Les systegravemes de deacutetection ont eacuteteacute testeacutes in silico gracircce agrave lrsquooutil
BLAST (Basic Local Alignment Search Tool) afin drsquoeacutevaluer la speacutecificiteacute de nos systegravemes pour chaque
cible par rapport agrave une base de donneacutees de geacutenomes seacutequenceacutes
La validation (speacutecificiteacute et seuil de deacutetection) de ces trois systegravemes ainsi que leur utilisation
dans le screening drsquoun nombre important drsquoeacutechantillons environnementaux sont preacutesenteacutees dans le
Chapitre 3 dans lrsquoarticle intituleacute laquo First molecular detection of Mycobacterium bovis in environmental
samples from a French region with endemic bovine tuberculosis raquo Les reacutesultats des analyses reacutealiseacutees
en 2013 sont preacutesenteacutes dans un article publieacute en 2014 dans la revue de lrsquoAEEMA (association pour
leacutetude de leacutepideacutemiologie des maladies animales) en Annexe 1
A
B
C
Chapitre 2
89
CHAPITRE 3
Etude de la preacutevalence environnementale de
Mycobacterium bovis dans la zone endeacutemique de
tuberculose bovine de Cocircte drsquoOr et reacutesultats obtenus
en Charente
Elodie Barbier Maria Laura Boschiroli Eric Gueneau Murielle Rochelet Ariane Payne Krystel
de Cruz AnneLaure Blieux Ceacuteline Fossot Alain Hartmann
First molecular detection of Mycobacterium bovis in environmental samples from a French
region with endemic bovine tuberculosis
(Article accepteacute en cours de publication Journal of Applied Microbiology)
93
CHAPITRE 3 - Etude de la preacutevalence environnementale de
Mycobacterium bovis dans la zone endeacutemique de tuberculose
bovine de Cocircte drsquoOr et reacutesultats obtenus en Charente
A AVANT-PROPOS
Dans la synthegravese bibliographique preacutesenteacutee dans le chapitre 1 nous avons deacutecrit la situation
eacutepideacutemiologique complexe de la tuberculose bovine en Cocircte drsquoOr Dans ce deacutepartement
Mycobacterium bovis circule dans un systegraveme multi-hocirctes complexe incluant les bovins les blaireaux
les cervideacutes et les sangliers sur une zone geacuteographique limiteacutee (Payne 2014) La faune sauvage infecteacutee
et potentiellement excreacutetrice est suspecteacutee drsquoecirctre agrave lrsquoorigine de la contamination ou recontamination
de certains foyers bovins dans cette zone (ANSES 2011 Courcoul Moutou amp Vialard 2013) Le mode
de transmission indirecte est suspecteacute car les contacts directs entre bovins et animaux sauvages sont
rarement deacutecrits (Drewe OrsquoConnor Weber McDonald amp Delahay 2013 Kukielka et al 2013) Ce mode
de transmission repose sur la contamination de matrices environnementales par des mycobacteacuteries
viables et virulentes agrave la faveur de lrsquoexcreacutetion animale puis sur lrsquointeraction drsquoanimaux avec ces
matrices Deacutemontreacutee par des meacutethodes moleacuteculaires etou culturales en Angleterre et en Espagne sur
des sites partageacutes par les espegraveces sauvages et les bovins (King et al 2015 Santos et al 2015 Sweeney
et al 2006 2007) la contamination environnementale par M bovis nrsquoa pas encore fait lrsquoobjet drsquoeacutetudes
dans les zones endeacutemiques franccedilaises
Afin drsquoeacutevaluer la preacutevalence et lrsquoexistence de reacuteservoirs potentiels de M bovis dans
lrsquoenvironnement des bovins et des animaux sauvages infecteacutes en Cocircte drsquoOr nous avons deacuteveloppeacute et
valideacute des systegravemes de deacutetection moleacuteculaire ou modifieacute des systegravemes existants adapteacutes aux matrices
environnementales complexes pour deacutetecter les espegraveces du MTBC et plus speacutecifiquement M bovis
Ces systegravemes ont eacuteteacute utiliseacutes pour analyser plus de 400 eacutechantillons environnementaux (sol eau
seacutediments microfaune fegraveces) preacuteleveacutes en Cocircte drsquoOr Notre strateacutegie drsquoeacutechantillonnage a cibleacute des
pacirctures suspectes sur lesquelles des bovins infecteacutes ont transiteacute et autour desquelles des animaux
sauvages infecteacutes ont eacuteteacute deacutetecteacutes et plus particuliegraverement les zones pouvant ecirctre partageacutees par les
diffeacuterentes espegraveces notamment les zones drsquoabreuvement Nous avons eacutegalement analyseacute des
eacutechantillons provenant de sites freacutequenteacutes exclusivement par la faune sauvage notamment des sols
de terriers et des latrines de blaireaux localiseacutes agrave proximiteacute du site de pieacutegeage de blaireaux infecteacutes
ainsi que des seacutediments preacuteleveacutes dans les trous drsquoeau ou souilles freacutequenteacutes par les sangliers et
cervideacutes
Chapitre 3
95
B FIRST MOLECULAR DETECTION OF MYCOBACTERIUM BOVIS IN ENVIRONMENTAL SAMPLES FROM A FRENCH REGION WITH ENDEMIC BOVINE TUBERCULOSIS
E Barbier12 ML Boschiroli3 E Gueneau4 M Rochelet2 A Payne1 Krystel de Cruz3 AL Blieux5 C
Fossot6 A Hartmann1
1 INRA UMR 1347 Agroeacutecologie Dijon Cedex France
2 Universiteacute de Bourgogne Franche Comteacute UMR 1347 Agroeacutecologie Dijon Cedex France
3 Universiteacute Paris-Est Laboratoire National de Reacutefeacuterence de la Tuberculose Uniteacute de Zoonoses
Bacteacuteriennes Laboratoire de Santeacute Animale ANSES Maisons-Alfort Cedex France
4 Laboratoire Deacutepartemental de la Cocircte drsquoOr Dijon France
5 Welience SATT Grand-Est AgrOnov Breteniegravere France
6 Phytocontrol Paris MIN de Rungis Rungis Cedex France
Running head
Environmental detection of Mycobacterium bovis
Corresponding author
Alain Hartmann UMR 1347 Agroeacutecologie INRA 17 rue Sully BP 86510 21065 Dijon Cedex France E-
mail alainhartmanndijoninrafr
Chapitre 3
96
ABSTRACT
Aims
The aim of the study was to determine the prevalence of Mycobacterium bovis (the causative agent of
bovine tuberculosis bTB) in environmental matrices within a French region (Cocircte drsquoOr) affected by this
zoonotic disease
Methods and Results
We report here the development and the use of molecular detection assays based on qPCR (double
fluorescent dye labelled probe) to monitor the occurrence of Mycobacterium tuberculosis complex
(MTBC) or M bovis in environmental samples collected in pastures where infected cattle and wildlife
had been reported Three qPCR assays targeting members of the MTBC (IS1561rsquo and Rv3866 loci) or M
bovis (RD4 locus) were developed or refined from existing assays These tools were validated using
M bovis spiked soil water and feces samples Environmental samples were detected positive for the
presence of MTBC strains and M bovis in the environment of bTB-infected farms in the Cocircte drsquoOr
region
Conclusions
The development of molecular assays permitted testing of several types of environmental samples
including spring water sediment samples and soils from badger setts entrance located in the vicinity
of these farms which were repeatedly contaminated with M bovis (up to 87 times 103 gene copies per g
of badger sett soil) For the first time direct spoligotyping of soil DNA enabled identification of M bovis
genotypes from environmental matrices
Significance and Impact of the Study
All together these results suggest that M bovis occurs at low levels in environmental matrices in Cocircte
drsquoOr within the bTB-infected area Drinking contaminated water or inhaling contaminated bioaerosols
might explain cattle infection in some cases
Keywords
Mycobacterium bovis environment quantitative real-time PCR indirect transmission
Chapitre 3
97
INTRODUCTION
Bovine tuberculosis (bTB) is a worldwide spread zoonosis caused by Mycobacterium bovis a
pathogenic mycobacteria that belongs to the Mycobacterium tuberculosis complex (MTBC) This
disease affects cattle that are the natural host of M bovis but also a wide range of domestic or wild
mammals including humans (Morris et al 1994 Biet et al 2005) Infected wildlife can act as a bTB
reservoir maintaining and spreading M bovis infection in cattle Many studies including French ones
showed the circulation of the bacteria within multi-host systems including cattle and various wild
species as badgers (Meles meles) wild boar (Sus scrofa) or red deer (Cervus elaphus) (Little et al 1982
Aranaz et al 2004 Gortaacutezar et al 2008 Zanella et al 2008 Richomme et al 2010 Payne et al 2012)
The zoonotic nature of bTB may represent a public health threat Moreover animal disease leads to
important economic losses mainly due to cattle testing slaughtering campaigns and wildlife screening
trials
Infected animals can excrete M bovis via sputum feces urine and any type of aerosols (Little
et al 1982 Phillips et al 2003 Palmer et al 2004 Payne 2014) and thus contaminate their environment
(Phillips et al 2003) This may contribute to contamination of cattle pastures through deposition of
M bovis on cattle feedstuffs and into drinking water Indeed many studies confirmed the presence and
the persistence of M bovis in environmental substrates such as soil feces hay corn water (Young et
al 2005 Courtenay et al 2006 Sweeney et al 2006 Fine et al 2011 Ghodbane et al 2014) despite
many technical difficulties in isolating this organism from environmental substrates Altogether these
data strongly suggest that the environment may constitute a reservoir of M bovis from which cattle
might get infected However there is still very little direct evidence concerning these potential M bovis
environmental reservoirs and the way that animals might get infected through them
The European Commission declared that France was bTB-free in 2001 However the disease is
still a persistent problem in several regions including Cocircte drsquoOr (Middle East of France Figure 1)
Despite a very low national prevalence in cattle (0075 since 2012) complete eradication of the
disease seems difficult to achieve (Fediaevsky et al 2014) In Cocircte drsquoOr M bovis circulates in a multi-
host system including badgers wild boar and deer within a restricted area of the region (Payne 2014
Hauer et al 2015) As shown in Figure 1 two genotypes are found in this area M bovis spoligotype
SB0120 VNTR profile 5 5 4 3 11 4 5 6 in the northern bTB endemic area and M bovis spoligotype SB0134
VNTR profile 6 4 5 3 6 4 3 6 in the southern area Between 2002 and 2014 196 cattle breakdowns have
been registered in Cocircte drsquoOr (Cote drsquoOr Regional Veterinary Service personal communication) Wild
animals in the vicinity of infected cattle herds were found to be infected by M bovis with the same
genotypes (Hauer et al 2015)
Direct cross contamination between cattle has historically been considered as the main
Chapitre 3
98
transmission route of M bovis (Morris et al 1994 Phillips et al 2003) but environmental
contamination is suspected in many herd infections when cattle and wildlife share the same resources
(Garnett et al 2002 Palmer et al 2004) However this hypothesis has never been extensively explored
in France This is probably due to the lack of adequate detection tools to monitor M bovis under natural
environmental conditions Indeed most of the studies report the detection of M bovis from
experimentally inoculated environmental samples (Fine et al 2011 Adams et al 2013) Although the
culture method is the gold standard for direct diagnosis of M bovis in animals it is not well suited to
studying environmental samples due to the abundance of soil microbial communities and the slow
growth rate of M bovis To circumvent these drawbacks the use of highly selective media containing
antibiotics and antifungals in combination with sample decontamination is required in order to prevent
competition of fast growing organisms However such pretreatments strongly reduce M bovis viability
(Thornton et al 1998 Fine 2006) On the other hand molecular detection tools such as semi-
quantitative PCR have been used to detect M bovis in soils water and badger feces Following DNA
extraction detection of MTBC members or M bovis was done using primers targeting mpb64 mpb70
hsp65 16SrRNA genes or IS6110 insertion sequence (Young et al 2005 Courtenay et al 2006
Sweeney et al 2006 Adams et al 2013 Santos et al 2015) More recently a M bovis specific qPCR
(based on double fluorescent dye probe detection) was developed to target the RD4 deletion flanking
sequences (Sweeney et al 2007) This molecular system has been validated in several studies to test
environmental substrates such as soil badger feces and cow slurry (Pontiroli et al 2011 Travis et al
2011 King et al 2015a King et al 2015b) This RD4 based qPCR assay allowed to detect the occurrence
of M bovis within environmental matrices in UK (Sweeney et al 2007 King et al 2015a King et al
2015b) Finally Santos et al (Santos et al 2015) successfully used a hemi-nested PCR targeting the
RD12 locus which is common to M bovis and M caprae to monitor these species in environmental
substrates from bTB endemic regions located in the Iberian Peninsula
In this context the objectives of this study were firstly to develop new sensitive qPCR based
tools and to refine an existing one (based on RD4 deletion sequence) for the specific detection of MTBC
or M bovis in environmental matrices The sensitivity and specificity of qPCR based detection systems
were evaluated on a collection of mycobacterial strain DNAs but also on DNAs extracted from samples
(soil water and feces) spiked with known amounts of M bovis Secondly we aimed to screen large
numbers of environmental samples (soil sediments water feces) originating from both bTB endemic
or bTB free areas to assess the presence of MTBC and more specifically the presence of M bovis in
these areas
Chapitre 3
99
Figure 1 Distribution of bTB cumulative outbreaks in cattle in France (picture in the insert above) and per municipality in Cocircte drsquoOr from 2005 to 2013 Geographical location of the sampled areas are indicated by black stars and the location of the six positive areas are indicated by white stars and named A B C D E and F respectively
MATERIALS AND METHODS
Bacterial strains
A collection of bacterial strains belonging to the species M bovis M bovis BCG vaccine strain
and other mycobacterial species (Table 1) was used to test the specificity of the primers to detect
M bovis Crude thermo-lysates of strains corresponding to 20 bacterial spoligotypes of the M bovis
species 4 other species of the MTBC 2 sub-species of the Mycobacterium avium Complex (MAC) and
20 non tuberculous mycobacteria (NTM) Field strains belong to the National Reference Laboratory
collection and were previously identified by several genotyping methods using four qPCR assays done
in parallel and targeting IS6110 IS1081 IS1245 and atpE Namely MTBC members are positive for
Chapitre 3
100
IS6110 and IS1081 targets (Courcoul et al 2014) MAC members are positive for the IS1245 target
(Slana et al 2010) and finally NTM are positive for the atpE target (Radomski et al 2013) Furthermore
spoligotyping was used to discriminate the different MTBC species (Zhang et al 2010) IS901 was used
to discriminate between MAC members (Slana et al 2010) and hsp65 sequencing was used to
discriminate between other mycobacteria species (Telenti et al 1993) Attenuated M bovis BCG strain
Pasteur 1173P2 was used as target organism to validate the specificity and detection limits of the qPCR
assay in soil water and feces microcosms
Origin of environmental samples
Within the bTB endemic zone in Cocircte drsquoOr a total number of 356 environmental samples were
collected from 15 areas (an area corresponds to a municipality territory including at least one bTB
infected cattle farm) between April 2013 and December 2014 In those areas typical landscape is
observed ie limestone plateau with forest or farm fields overhang small hills The hillslopes are
covered with hydromorphic soils containing marl and claylimestone Alluvial valleys set at the bottom
of the slopes Cattle usually graze on those pastures during summer and spring Samples of soil
sediment and water were collected in areas frequented by both wildlife and cattle such as feeding
areas waterholes naturally occurring springs drinking troughs with open access to livestock and
wildlife Knowing that bTB infected badgers were discovered in the vicinity of the pastures grazed by
infected cattle (Cote drsquoOr Regional Veterinary Service personal communication) soils from badger
setts and latrines were also collected
Twenty-four agricultural soils were randomly chosen as control soils in French bTB-free areas
among a panel of soils from the French soil quality monitoring network (Reacuteseau de Mesures de la
Qualiteacute des Sols Infosol INRA Orleacuteans) Among them 11 french soils were collected from Brittany
(North West of France) Yvelines (Middle north of France) Haut Rhin (North east of France) while 13
soils were collected in the bTB-free area of Cocircte drsquoOr
In order to evaluate the detection limit of the qPCR assay we used two agricultural soils from two bTB-
free areas of Cocircte drsquoOr (Soil S1 and soil S2) one pond water and badger feces from animals tested as
bTB-free from non-infected areas Both soils were sampled in a bTB-free cattle pasture in November
2013 They were chosen for their contrasted structural and physico-chemical properties (Table 2)
DNA extraction and purification
DNAs from all soil sediment and latrine samples were extracted from 2 g-samples as following
Briefly 8 ml of lysis buffer (01 mol l-1 Tris pH 80 01 mol l-1 EDTA pH 80 01 mol l-1 NaCl and 2 (wv)
SDS) 4 g of silica beads (100 μm diameter) 5 g of ceramic beads (14 mm diameter) and 8 glass beads
(4 mm diameter) were added to each sample placed in a 15 ml conical centrifugation tube (Falcon
Chapitre 3
101
Dominique Dutscher France) Samples were disrupted for 3 times 30 s at 4 m s-1 in a FastPrep-24 Instrument
(MP Biomedicals Europe France) incubated for 30 min at 70degC and centrifuged for 5 min at 7000 g at
room temperature in a Sigma 3-18K centrifuge (Sigma-Aldrich Germany) Supernatants were
incubated on ice for 10 min with 110 volume of 3 mol l-1 potassium acetate pH 55 and then
centrifuged for 5 min at 14000 g One volume of ice-cold isopropanol was added to the supernatant
for DNA precipitation overnight at minus 20degC DNA was collected by centrifugation (30 min at 14000 g
4degC) and DNA pellets were washed with ice-cold ethanol (70 ) and dissolved in 100 μl of water Crude
DNA extracts (100 μl) were loaded on a Microbiospin column (Biorad France) filled with 100 mg of
polyvinyl polypyrrolidone (Sigma Chemical Co France) After centrifugation (4 min at 1000 g 4degC)
eluted DNA was further purified using the Geneclean Turbo Kit (Qiagen France) according to the
manufacturers instructions For water samples the microbial biomass was concentrated by filtering
from 100 ml to a maximum of 1 l of water sample (depending on water turbidity) through a 045 microm
Millipore HAWP filter (47 mm Merk Millipore Germany) Filters were then ground in liquid nitrogen
and DNA was extracted as described above Nematodes were extracted from soil using an Oostenbrink
elutriator (Oostenbrink 1960) DNA extraction was achieved as described above
Purified DNA concentrations were estimated using a NanoDropreg 2000 spectrophotometer
(Thermo Scientific France) at 260 nm wavelength
Choice of gene targets design of primers and probes for qPCR assays
A first qPCR detection system assay targeting the insertion sequence IS1561rsquo was designed
This insertion sequence is present in all the MTBC members except in M microti (Huard et al 2003
Huard et al 2006) The primers and double dye probe sequences are given in Table 3 IS1561rsquo contains
a defective transposase and belongs to the ISL3 family (Gordon et al 1999) In order to confirm the
specificity of the detection for MTBC a nested PCR protocol was additionally developed the first step
was a classical PCR using primers targeting a sequence of 222-bp sequence in the IS1561rsquo surrounding
the 76-bp qPCR product (Table 3) the second step was a qPCR assay targeting the 76-bp fragment To
confirm the results obtained with IS1561rsquo qPCR assay two supplementary qPCR targets were used The
first one was the Rv3866 locus located in the RD1 locus (Brodin et al 2002) The second one was the
RD4 flanking region (the RD4 locus is specifically deleted in M bovis) To improve the qPCR efficiency
the RD4 based assay was derived from a previously reported qPCR assay (Sweeney et al 2007) by
modifying the reverse primer to reduce the size of the qPCR product from 142 to 116 bp (Table 3) The
RD4 based qPCR assay designed by Sweeney et al (Sweeney et al 2007) has previously been validated
for environmental matrices in several studies (Pontiroli et al 2011 Travis et al 2011 King et al 2015a
King et al 2015b) Primers and probe were designed from alignments of mycobacterial genes from
GenBank (httpwwwncbinlmnihgovgenbank) such as M bovis AF212797 four strains of M bovis
Chapitre 3
102
BCG M tuberculosis H37Rv M africanum GM041182 and Mycobacterium canettii CIPT 140010059
(respective accession numbers NC_0029453 NC_0087691 NC_0168041 NC_0122071
GCA_0004632151 NC_0009623 NC_0157581 NC_0158481) using Primer3web
(httpprimer3utee) The specificity of the assay was tested in silico by BLAST analysis
(httpblastncbinlmnihgovBlastcgi) Primers and probes were synthesized by Eurogentec (Seraing
Belgium)
Quantitative real time PCR (qPCR) and nested PCR conditions
For the three qPCR assays reactions were carried out in a 25 microl reaction mix containing 125 microl
of Absolute QPCR ROX Mix (Thermo Scientific France) at a final concentration of 1X 1 microl of T4
bacteriophage gene 32 product (final concentration 25 ng microl-1 MP Biomedicals Europe France) 1 microl of
each primer (final concentration 400 nmol l-1) 05 microl of each probe (final concentration 200 nmol l-1)
4 microl of ultrapure water and 5 microl of diluted DNAs for the environmental samples screening Absolute
quantification was achieved using standard DNA dilutions A recombinant plasmid containing one copy
of the IS1561rsquo fragment of M bovis BCG str Pasteur 1173P2 cloned in the pCRregII-TOPOreg vector
(Invitrogen France) was used as a standard A calibration curve was obtained by amplification of serial
dilutions of the plasmid ranging from 1 to 106 copies per PCR reaction All the environmental
metagenomic DNA samples were screened with the IS1561rsquo based qPCR assay and final results were
expressed as the gene copy number per g of soil feces or water qPCR assays were performed in a VIIA
7 Real-Time PCR System (Thermo Fischer Scientific France) Initial denaturation was made at 95degC for
15 min followed by 45 cycles with denaturation at 95degC for 15 s and annealing and elongation at 60degC
for 1 min
Positive DNA extracts were then tested by the nested IS1561rsquo PCR protocol as follows the first PCR was
achieved in a 25 microl reaction mixture containing 1725 microl of ultrapure water for molecular biology 25
microl of PCR Buffer 10X with MgCl2 02 microl of dNTP mix at 25 mmol l-1 each 005 microl of Taq DNA polymerase
15 U microl-1 (Taq Core Kit 25 MP Biomedicals Europe France) 125 microl of each primers (final concentration
500 nmol l-1) and 25 microl of environmental DNA The nested PCR was achieved in a Veriti Thermal Cycler
(Applied Biosystems France) with a first step of denaturation at 94degC for 5 min then 30 cycles for 30
s at 94degC for denaturation 30 s at 60degC for annealing and 30 s at 72degC for elongation and 5 min at 72degC
for final elongation The qPCR was then achieved in the same conditions as described above using as a
target a 5 microl aliquot of a 100-fold dilution of the first PCR product
Preliminary screening of all environmental samples was done using the IS1561rsquo based qPCR
assay targeting MTBC species The IS1561rsquo positive samples were then further analysed using the
Rv3866 based qPCR to confirm the presence of MTBC and the RD4 based qPCR assay to confirm the
presence of M bovis All qPCR assays were done in triplicate
Chapitre 3
103
Determination of qPCR assays analytical sensitivity in spiked soil feces or water microcosms
Microcosms consisting of 2 g of fresh soil S1 and S2 sieved to 4 mm 2 g of fresh badger feces
or 100 mL of pond water were spiked in triplicate with 100 microl of 10-fold serially diluted suspensions of
M bovis BCG strain Pasteur to reach M bovis concentrations ranging from 25 to 25 times 106 CFU per
gram of soil or feces and 50 to 50 times 106 CFU per 100 mL of water Briefly inoculum suspensions were
realized as following a thirty days M bovis BCG liquid culture was vortexed for 15 s with glass beads (4
mm diameter) in order to disrupt aggregates Optical bacterial density was then measured at 600 nm
(Biophotometre Eppendorf France) to roughly estimate mycobacterial concentration Appropriate 10-
fold dilutions (10-1 to 10-6) were prepared in sterile water M bovis concentration was estimated by
spreading in triplicate 100 microL-aliquot of 10-4 10-5 and 10-6 diluted suspension onto Middlebrook 7H11
agar enriched with OADC 10 (Sigma-Aldrich France) Plates were incubated at 37degC for two months
and then M bovis colonies were counted Results were reported as CFU per ml of suspension
The spiked microcosms were incubated at room temperature for 3-4 hours prior to DNA
extraction as described in the section 3 To assess the reproducibility all qPCR assays were done in
triplicate and performed as previously using the three detection systems targeting IS1561rsquo Rv3866 and
RD4 Non-inoculated soil feces or water used to prepare the microcosms proved to be negative with
the three qPCR systems in preliminary experiments
Estimation of DNA quality for PCR amplification
Efficiency of qPCR might be impaired by the presence of PCR inhibitors in DNA extracts The
quality of DNA extracted from soil water and feces was assessed by measuring the capacity to amplify
a control DNA target in the presence of various dilutions of soil DNA extracts Briefly qPCR reactions
were done in a final volume of 25 microl containing 25 μl of 106 copies of the circularized pGEM-T Easy
plasmid DNA and 5 μl of water (control) or diluted soil DNA (5- 10- 20-fold dilutions) 1X ABsolute
qPCR SYBR Green ROX Mix (final concentration 500 nmol l-1 Thermo Scientific France) 25 ng microl-1 of T4
bacteriophage gene 32 product qPCR primers were SP6 (5rsquo - ATTTAGGTGACACTATAG - 3rsquo) and T7 (5rsquo -
TAATACGACTCACTATAGGG - 3rsquo) universal primers (final concentration 500 nmol l-1) targeting the
polylinker of the plasmid qPCR and cycling conditions were as following initial denaturation at 95degC
for 15 min followed by 40 cycles with denaturation at 95degC for 15 s annealing at 55degC and elongation
at 72degC for 30 s each The PCR product specificity was assessed by the melting curve analysis A 10-fold
reduction of the copy number amplified by qPCR in the presence of diluted soil DNA with respect to
the copy number obtained in the control reaction (water) was considered as 10 amplification
efficiency or conversely as 90 inhibition Although inhibition tests and detection of specific targets
were conducted with different mixes and primers the DNA polymerases from the ABsolute qPCR SYBR
Chapitre 3
104
Green ROX Mix (used for inhibition testing) and the ABsolute QPCR ROX Mix (used for mycobacterial
detection in environmental samples) are the same (Thermo-Start DNA polymerase) Thus the quality
of DNAs extracted from environmental samples was estimated by this method that is compatible with
acceptable experimental costs associated with the high number of analysed samples
Direct M bovis spoligotyping from environmental DNA
Environmental DNA samples that proved to be positive for the presence of MTBC were
submitted for spoligotyping using the Luminex method described previously by Zhang et al (Zhang et
al 2010) Spoligotypes were named according to an agreed international convention
(wwwmbovisorg)
RESULTS
Validation of the qPCR assays on pure bacterial strains
The results of the specificity test for the three qPCR assays are shown in Table 1 Using IS1561rsquo-
and Rv3866-based qPCR assays all spoligotypes of M bovis were detected with cycle threshold (Ct)
values below 25 cycles as well as all strains from the other species belonging to the MTBC (M caprae
M tuberculosis M pinnipedi) except the M microti species Using the RD4-based qPCR assay only
M bovis strains were detected with Ct values below 25 cycles For the three qPCR assays the 22 strains
of the NTM and MAC members were not detected or gave Ct values above 35 cycles Strains with Ct
values above 35 cycles when using high DNA concentration from pure culture are unlikely to give any
signal when testing soil metagenomic DNA where targets are highly diluted These results indicated
that IS1561rsquo- and Rv3866-based qPCR assays are highly specific for MTBC members and exclude
M microti whereas the RD4-based qPCR assay is specific for M bovis species
The calibration curve used for absolute quantification is presented in Figure 2 for the IS1561rsquo-
based qPCR assay A linear regression was established between the log (gene copy number) and the Ct
value over a wide range of target concentrations from 10 to 106 gene copies per PCR (r2= 09989) Below
10 target gene copies per reaction the detection was still possible but results were not reproducible
The quantification limit of the qPCR detection system was estimated to be 10 gene copies per PCR
reaction thus indicating a high sensitivity Similar calibration curves and detection thresholds were
obtained for the two other qPCR assays targeting Rv3866 and RD4 (data not shown)
Chapitre 3
105
Table 1 Strains used for testing oligonucleotides specificity Results are given with Ct (Cycle Threshold) ND (not detected) means that no signal was observed after 40 cycles
Strains Ct
IS1561rsquo
Ct
Rv3866 Ct RD4
Mycobacterium bovis BCG
BCG str Pasteur 1173P2
BCG str Danish 1331
lt25 lt25 lt25
Mycobacterium bovis
SB0089 SB0120 SB0121 SB0130
SB0134 SB0162 SB0166 SB0821
SB0823 SB0825 SB0826 SB0827
SB0878 SB0885 SB0912 SB0948
SB0999 SB1081
lt25 lt25 lt25
Others MTBC members
Mycobacterium microti ND ND ND
Mycobacterium tuberculosis lt25 lt25 ND
Mycobacterium caprae lt25 lt25 ND
Mycobacterium pinnipedi lt25 lt25 ND
Non tuberculous mycobacteria
Mycobacterium avium ND ND ND
Mycobacterium intracellulare ND ND ND
Mycobacterium aichiense ND ND ND
Mycobacterium arupense ND gt35 gt35
Mycobacterium celatum gt35 ND ND
Mycobacterium chelonae ND ND ND
Mycobacterium chitae ND ND ND
Mycobacterium confluentis gt35 ND gt35
Mycobacterium elephantis ND ND ND
Mycobacterium fortuitum ND ND ND
Mycobacterium intermedium ND ND ND
Mycobacterium kansai ND gt35 ND
Mycobacterium non chromogenicum ND ND ND
Mycobacterium parafinicum gt35 ND ND
Mycobacterium porcinum ND ND ND
Mycobacterium rufum ND ND ND
Mycobacterium rutilum ND ND ND
Mycobacterium terrae ND ND ND
Mycobacterium smegmatis ND gt35 ND
Mycobacterium szulgai ND gt35 ND
Mycobacterium thermoresitibile ND ND ND
Mycobacterium vaccae ND ND ND
Mycobacterium xenopi gt35 gt35 gt35
Chapitre 3
106
Detection threshold of qPCR assays in M bovis BCG spiked environmental matrices
The detection thresholds of the three qPCR assays were evaluated in complex environmental
matrices (soil feces and water) spiked with 25 to 25 times 106 CFU per g of soil or feces and 50 to 50 times
106 CFU per 100 ml of water The quality of DNAs extracted from microcosms was assessed as described
in the MampM section ldquoEstimation of DNA quality for PCR amplificationrdquo The results showed that 20-fold
dilutions of the DNA extracts did not inhibit the qPCR assay and were thus used for all qPCR assays
performed on spiked microcosms Results of M bovis detection in spiked microcosms are given in Table
4 for IS1561rsquo-based qPCR assay No signal was detected in control samples (not inoculated)
For M bovis cell densities above 250 CFU per g of soil 2500 CFU per g of feces and 5 CFU per
g of water (or 500 CFU per 100 ml of water) the detection by the qPCR assay yielded eight to nine
positive reactions out of nine repeats which allowed determination of the numbers of M bovis gene
copy number In contrast although M bovis was sometimes detected in spiked material with lower
densities than those described above quantification was unreliable A strong discrepancy between the
results obtained by the culture-based method and those obtained with the qPCR assay was observed
(10- to 100-fold higher for the qPCR assay) All of the microcosms were analysed with the two
complementary detection systems (targeting the Rv3866 locus and the flanking region of the RD4
deletion) and similar results were obtained (data not shown) Despite soils S1 and S2 being different
based on their physico-chemical characteristics detection limits were equivalent with the three qPCR
systems on these two soils
Extrapolating these results for natural environmental samples suggest that the three qPCR
assays allow a reliable quantification of MTBC (IS1561rsquo and Rv3866) or M bovis (RD4) targets in
environmental samples for Ct values below 39 Above this threshold detection may occur but lacks
reproducibility The word ldquotracesrdquo was then chosen in all further experiments in order to indicate that
MTBC or M bovis targets are present in very low quantities
Chapitre 3
107
Figure 2 Standard curve used for absolute quantification of M bovis in samples with IS1561rsquo qPCR assay (Mean values of ten standard curves) Error bars represent the 95 confidence interval associated to the values
Table 2 Properties of the two soils used for the qPCR sensitivity detection of Mycobacterium bovis
Table 3 Sets of primers tested in this study F forward primer R reverse primer P Double dye probe
Target locus Nucleotide sequence Location Size (bp)
IS1561rsquo F 5rsquo GATCCAGGCCGAGAGAATCTG 3rsquo
R 5rsquo GGACAAAAGCTTCGCCAAAA 3rsquo
P 5rsquo FAM ACGGCGTTGATCCGATTCCGC TAMRA 3rsquo
3742298-3742319
3742354-3782374
3742332-3742353
76
Nested IS1561rsquo F 5rsquo AGGAAGCGGTGTAGGTGATG 3rsquo
R 5rsquo AAGATCGACCCGCAGTG 3rsquo
3742212-3742232
3742416-3742434 222
RD4 F 5rsquo GCAGAAGCGCAACACTCTTG 3rsquo
R 5rsquo ATCTTAGCTGGTCAATAGCCATTTTT 3rsquo
P 5rsquo FAM AGTGGCCTACAACGGCGCTCTCC 3rsquo
1692712-1692736
1692802-1692827
1692743-1692768
116
Rv3866 F 5rsquo ACCCGCCAACGTCGAA 3rsquo
R 5rsquo CAATTGACGCAATACGGTATCG 3rsquo
P 5rsquo FAM CGAGCTAGCCGAATGCACAACCG 3rsquo
4278658-4278673
4278721-4278743
4278693-4278716
86
Location is given by reference to the M bovis AF212297 genome sequence accession ndeg NC_0029453
y = -33818x + 4394Rsup2 = 09989
15
20
25
30
35
40
45
1 2 3 4 5 6
Cyc
le T
hre
sho
ld C
t
DNA concentrations [log(IS1561 copies5microL-1)]
Site Clay Loam Sand Total
Carbon
Total Nitrogen
pH H2O
Organic matter
Land use
S1 397 485 118 361 038 775 0625 Pasture S2 192 463 345 292 0269 546 0506 Pasture
Chapitre 3
108
Molecular detection of M bovis in environmental samples
Our molecular detection systems were first evaluated on twenty-four soils from bTB-free zones
in France Whatever the detection system no signal was observed showing a good specificity of the
IS1561rsquo Rv3866 and RD4 qPCR systems
Between April 2013 and December 2014 63 samples of pasture soil 150 samples of setts soil 21
latrines samples 67 water samples from naturally occurring springs (and subsequent flowing water on
soil surface) within cattle pastures and 55 samples of spring sediments within 15 areas located in the
bTB-endemic zone in Cocircte drsquoOr were analysed Some spots were sampled several times (monthly from
January to June and in November) to assess the persistence of the MTBC signals Altogether 356 DNA
samples were primarily screened using the qPCR assay targeting IS1561rsquo locus Molecular signals
corresponding to MTBC were found in six areas out of 15 analysed and only positive results are
presented in Table 5 Apparent gene copy numbers were the average value from n = 2 to n = 3 repeats
These results indicate a MTBC contamination of the environment in 40 of the studied areas
Among the 36 positive samples 19 were found to be positive for MTBC detection with low
signals (indicated as ldquoTracesrdquo in Table 5) These samples include 7 badger sett soils 3 badger latrines
1 sediment sample and 8 water samples from naturally occurring springs in pastures More
interestingly high positive signals for MTBC were observed for the other 17 positive samples with 58
to 8 700 gene copies per g of soil It is worth noting that these samples only included badger sett soils
in area D and one nematodes sample in area C while no signal was recorded for the pasture soils
In order to confirm these positive results some of them were subjected to the nested IS1561rsquo
qPCR assay Eleven samples including eight badger sett soils two samples of water from springs and
one sample of nematodes isolated from spring sediments were confirmed to be positive for MTBC
detection with this nested qPCR assay Badger sett soils from area D were sampled five times over an
eight month-period The occurrence of MTBC signal was discontinuous in the seven soils from badger
setts that were sampled However MTBC signals were detected in several badger sett soils (badger sett
soils 1 4 and 5 in Table 5) following a seven to eight months period after M bovis was detected for the
first time The IS1561rsquo gene copy numbers for the positive badger sett soils ranged levels from
254 times 102 to 505 times 104 per g of soil The MTBC signal remained at high levels throughout the eight
month-period although the badgers moved out one month after the first sampling of the sett (Table
5) Furthermore confirmation of M bovis detection using the RD4 qPCR detection system was achieved
for a few IS1561rsquo positive samples Only three out of eight tested samples proved to be positive with
RD4-based detection system (see Table 5) This result definitely demonstrates the occurrence of M
bovis DNA in these samples
Spoligotyping was performed on positive DNA samples and the results are shown in Table 6 A
badger sett soil DNA was fully spoligotyped and found to belong to the SB0120 M bovis spoligotype
Chapitre 3
109
which is responsible for cattle and wildlife infection in this area of Cocircte drsquoOr Partial M bovis
spoligotypes were obtained for three badger sett soils DNAs The two other partial spoligotypes did
not allow identification of the species within the MTBC
Table 4 Evaluation of the detection of the IS1561rsquo qPCR assay with DNA extracted from control and spiked test matrices
Traces means that IS1561rsquo signal was detected but that quantification was unreliable (Ct value above 39)
Number of M bovis spiked per g of dry soil or per 100 mL of water
Number of positive PCR 9
reactions
Mean Ct Standard deviation Ct
Mean IS1561rsquo genome-equivalent per g of soil or feces or 100 mL of water
Soil S1
Control soil 0 ND ND ND
25 1 396 ND Traces 25101 1 394 ND Traces 25102 8 389 1 (654 plusmn 366) times103
25103 9 345 05 (217 plusmn 086) times105
25104 9 302 02 (218 plusmn 026) times106
25105 9 276 05 (132 plusmn 039) times107
Soil S2
Control soil 0 ND ND ND 25 0 ND ND ND 25101 5 404 06 Traces 25102 9 381 07 (800 plusmn 319) times103
25103 9 342 04 (141 plusmn 049) times105
25104 9 296 05 (374 plusmn 161) times106
25105 9 267 07 (265 plusmn 185) times107
Water Control water 0 ND ND ND 5101 1 401 ND Traces 5102 9 381 11 (220 plusmn 145) times104
5103 9 351 05 (148 plusmn 058) times105
5104 9 31 16 (320 plusmn 134) times106
5105 9 259 03 (819 plusmn 209) times107
5106 9 228 03 (698 plusmn 131) times108
Badger feces
Control feces 0 ND ND ND 25101 0 ND ND ND 25102 1 399 ND Traces 25103 9 379 105 (232 plusmn 077) times104
25104 9 347 159 (212 plusmn 187) times105
25105 9 312 203 (244 plusmn 286) times106
25106 9 261 059 (336 plusmn 107) times107
Chapitre 3
110
Table 5 Molecular detection of MTBC or M bovis targets in positive environmental samples from 6 areas in Cocircte drsquoOr Only positive samples are presented MTBC gene copy numbers were computed from calibration curve and amounts of DNA extracted from the various samples (n=2 ou n=3)
Area
Type of sample Sampling date Mean MTBC gene copy numbers g-1 of soil or feces (IS1561rsquo qPCR assay)
Confirmation after IS1561rsquo-nested PCR
Confirmation of presence of M bovis after RD4 qPCR assay
A Badger sett soil June 2013 Traces Negative NT dagger
B Spring water 1 January 2014 February 2014 March 2014
Traces
Traces
Traces
Negative Negative Negative
Negative Negative Negative
C Spring water 1 Spring water 2 Sediments Soil nematodes
February 2014 February 2014 February 2014 February 2014
Traces Traces Traces 582101
Positive Negative Negative Positive
NT
NT
NT
NT
D Badger sett soil 1 March 2014 November 2014
193103 577103
Positive NT
Positive NT
Badger sett soil 2 March 2014 May 2014 June 2014
870103 502102 143103
Positive Positive NT
Positive NT
NT Badger sett soil 3 March 2014
May 2014 June 2014
441102
741103 282103
Positive Positive NT
Negative NT
NT Badger sett soil 4 April 2014
May 2014 June 2014 November 2014
446103
236103 502102 607103
Positive NT
NT
NT
Negative NT
NT
NT Badger sett soil 5 April 2014
May 2014 June 2014 November 2014
248103
254102
202103
360103
Positive NT NT
NT
Positive NT
NT
NT Badger sett soil 6 November 2014 Traces NT NT Badger sett soil 7 November 2014 Traces NT NT Latrine 1 March 2014 Traces Negative NT Latrine 2 March 2014 Traces Negative NT
E Spring water 1 March 2014 March 2014 March 2014
Traces Negative NT Spring water 2 Traces Positive NT Spring water 3 Traces Negative NT Badger sett soil 1 March 2014
March 2014 Traces Negative NT
Badger sett soil 2 Traces Negative NT Latrine 1 March 2014 Traces Negative NT
F Badger sett soil 1 June 2014 June 2014
Traces Negative NT Badger sett soil 2 Traces Positive NT
Traces MTBC targets were present but MTBC genome equivalent were not calculated since Ct were close to the detection limit of the method dagger NT not tested
Chapitre 3
111
Table 6 Spoligotyping results of DNA positive samples extracted from environmental matrices Black squares represent positive hybridization signals and dots represent lack of hybridization
Figure 3 Spatial location of MTBC positive water and sediment samples in a pasture (site C) along the run off of one naturally occurring springs Star 1 indicates the sampling zone of positive spring sediment and nematodes sample Stars 2 and 3 indicate the 2 sampling sites of positive source water Contour lines are superimposed on aerial photography (450 m and 425 m contour lines are visible 5 m elevation interval between 2 consecutive lines)
Area Type of sample
Sampling date Hybridization pattern Spoligotype
D Badger sett soil 2
March 2014 M bovis SB0120
D Badger sett soil 1
March 2014 Partial M bovis spoligotype
D Badger sett soil 3
March 2014 Partial M bovis spoligotype
D Badger sett soil 5
November 2014
Partial M bovis spoligotype
C Water February 2014
Partial Mycobacterium spoligotype
Chapitre 3
112
DISCUSSION Mycobacterium bovis is described as an obligate intracellular microorganism transmitted by
direct contact between mammalian hosts However direct contacts between species seem to be scarce
and indirect transmission involving environmental matrices is likely to occur (Boumlhm et al 2009
Kukielka et al 2013) Many studies have clearly demonstrated the presence and the persistence of
M bovis in naturally infected environmental substrates in bTB endemic regions of United Kingdom
(Young et al 2005 Courtenay et al 2006 Sweeney et al 2006 Sweeney et al 2007 King et al 2015a
King et al 2015b) Evaluation of the environment as a reservoir of M bovis could help to understand
its epidemiological significance and its role in the indirect transmission of the disease among livestock
and wildlife To our knowledge the results from our study demonstrate for the first time the
contamination of environmental matrices with MTBC and more specifically M bovis in Cocircte drsquoOr which
is a region where bTB is endemic in cattle and wildlife
qPCR detection is a method of choice for M bovis environmental monitoring since it is a faster
alternative to culture based methods that are hampered by the slow-growing rate of M bovis and the
high number and diversity of microbial communities Moreover PCR is able to detect M bovis DNA in
the environment over a long period following initial contamination with a relatively high sensitivity
(Adams et al 2013)
The detection of M bovis in environmental samples (soil water and feces) that contain highly
diverse microbial communities including numerous described and potentially non-described NTM
(Falkinham 2009) was still a challenge The two qPCR assays developed in this study (IS1561rsquo and
Rv3866 targets) proved to be specific for the MTBC excluding M microti These results are in agreement
with the fact that IS1561rsquo is present in the genomes of all the members of the MTBC except in M microti
(Gordon et al 1999 Brodin et al 2002 Huard et al 2003) Itrsquos worth noting that M microti is quite
widespread in animals in France (Michelet et al 2015) and thus may also occur in environmental
matrices (Courtenay et al 2006) The refined RD4 qPCR assay was shown to be highly specific for
M bovis species in agreement with previous results (King et al 2015a) Although some strains gave Ct
gt 35 when concentrated in pure culture with these molecular assays these targets would not be
detected in natural samples due to extreme dilution in highly complex environmental DNAs
The DNA extraction method used in this study proved to be highly efficient in soils harboring
different physico-chemical characteristics (especially in clayey soils) (Dequiedt et al 2011 Locatelli et
al 2013) The large mass of samples (2 g) allowed a better representativeness of sampling to overcome
the potential heterogeneity of substrates ie a physical heterogeneity as well as a heterogeneous
distribution of mycobacteria in environmental samples The use of an amplification facilitator (T4
bacteriophage gene 32 product) allowed minimizing PCR inhibition by co-extracted contaminants
(Matheson et al 2010 Opel et al 2010) and significantly improving M bovis detection In previous
Chapitre 3
113
published studies (Travis et al 2011 Pontiroli et al 2011 Sweeney et al 2007) DNA extraction was
usually achieved from smaller masses (01 to 05 g) of soil or feces using commercial DNA extraction kit
in order to limit PCR inhibition
The three qPCR assays developed in this study using 20-fold dilutions of DNA extracts of spiked
samples allowed a reliable detection of spiked M bovis (BCG strain) at levels as low as 25 times 102 CFU to
25 times 103 CFU g-1 of sample providing an improved detection limit compared to a recent study where
detection limit was 40 times 104 CFU g-1 soil and 50 times 105 CFU ml-1 water (Santos et al 2015) In other
previous studies detection limits were estimated to 85 times 103 cells g1 for soil and 425 times 105 cells g1 for
fecal samples using the original RD4 qPCR assay (Pontiroli et al 2011 Travis et al 2011) As cell numbers
were estimated by fluorescent dye assisted flow cytometry comparison with our results is difficult
Finally King et al (King et al 2015a King et al 2015b) were able to detect 103 M bovis genome
equivalents per g-1 for badger feces
In our study the detection limits are equivalent for the three qPCR systems developed or refined
whatever the physico-chemical characteristics of the soils
The 10- to 100-fold discrepancy between bacterial numbers obtained using culture based methods and
qPCR based methods that we observed could be explained by the underestimation of cultivable
bacteria due to cell aggregation (linked to a strong hydrophobicity of mycobacteria cell wall) in
inoculum as well as the occurrence of viable but not cultivable cells (VBNC) and cell mortality (with
persistence of DNA) Similar discrepancies have already been observed in previous studies (eg Betelli
et al 2013)
Though similar detection thresholds were observed for the three systems in spiked samples
RD4 and RV3866 based qPCR assays seemed less sensitive than IS1561rsquo-based detection system in
environmental samples (range of 2 Ct between IS1561rsquo and the 2 other systems) However the
occurrence of RD4 based positive signals in several samples gave an additional proof of the specific
detection of M bovis Furthermore the direct spolygotyping of DNA extracted from environmental
matrices confirmed for the first time the presence of M bovis in several environmental samples in
France
All 63-pasture soil samples from the bTB endemic area in Cocircte drsquoOr were found to be negative
for the presence of MTBC (below the detection threshold) Failure to detect MTBC in pasture soil
samples can be related to the low prevalence of bTB in cattle in the sampling areas (herd prevalence
reached 08 in 2014 Cote drsquoOr Regional Veterinary Service) and the low prevalence of bTB infection
in badgers which territories overlap the sampled pastures (Payne et al 2012) This result might reflect
the high dilution rate of M bovis targets in soil DNA extracts which contains high numbers of other
microorganisms (abundant soil microbial communities containing up to 1010 genomes per g of soil)
The discontinuous (spatial and temporal) distribution of M bovis in soil pasture because of
Chapitre 3
114
intermittent andor low shedding (Neill et al 1988 Clifton-Hadley et al 1993) might also limit the
detection of M bovis in soil samples Despite the high number of soil sample analyzed financial and
time constraints have limited more intensive sampling efforts that could have overcome this limit
MTBC signal was detected in 101 (7 positive out of 67) of the water samples taken from
naturally occurring springs with the IS1561rsquo qPCR assay As the level of detection was close to the
detection threshold we were unable to quantify MTBC signals Interestingly one sample of nematodes
originated from sediments located in close vicinity of one of the positive sources was also found
positive for the presence of MTBC Although nematodes are known to host bacterial pathogen no data
is available concerning MTBC This intriguing result would require further investigations These water
springs (and accompanying flowing water) are located in pastures where both cattle and wildlife have
access for drinking In several pastures (four sites) water was sampled along the slope following the
gravitational water flow In two of these pastures we repeatedly found (two to three) positive samples
indicating multiple point contamination of spring and flowing water along these slopes (see Figure 3
for localization of positive samples in one of these pastures) This result is an important new finding of
our study that can support several hypotheses on the role of water in the dissemination of MTBC in
the environment and in animal contamination We can hypothesize from these results that bioaerosols
formation from contaminated water and subsequent inhalation by animals (cattle and wildlife) might
contribute to animal infection with MTBC members especially as the infective dose through this
transmission route is very low (lt 10 CFU) (Dean et al 2005 Corner et al 2007) Infection of animals
might also occur through oral route by repeatedly drinking contaminated water
A special effort was made to monitor the contamination of badger sett soils since bTB infected
badgers were detected in our study area Between 2009 and 2011 878 badgers had been trapped and
analyzed in the infected area of Cocircte drsquoOr (Payne et al 2012) Macroscopic lesions were present in 19
of them despite culture-based method detecting 56 of positive badgers Only one badger showed
widespread tuberculosis Moreover in this infected area numerous badger setts were located either
within pastures or in close vicinity (forest-pasture interface) Our study demonstrated an average
prevalence of MTBC of 73 (11 out of 150) in soils from badger setts with the highest gene copy
number up to 37 times 104 gene copies g-1 of soil observed among environmental samples analysed in our
study The prevalence of MTBC molecular signature in latrines was estimated to 7 (3 positive out of
21) Our results demonstrate that infected badgers might contaminate their close environment for the
first time in France Indeed infected badgers have been shown to excrete M bovis from the respiratory
digestive and urinary tracts and in bite wounds exudates (Gavier-Widen et al 2001 Corner et al 2012)
Our results are in agreement with previous findings from several studies in UK (Young et al 2005
Courtenay et al 2006 Sweeney et al 2007 King et al 2015a) that recovered M bovis from badger
setts latrines and adjacent fields with gene copy numbers ranging from 1 times 103 to 54 times 106 per g of
Chapitre 3
115
soil In Woodchester Park in UK where badgers populations have been extensively studied for more
than 20 years bTB prevalence was estimated around 30 in 2005 which might explain high numbers
of positive setts and latrines observed in this area (Courtenay et al 2006 King et al 2015a) As
suggested by Courtenay et al (Courtenay et al 2006) the low bTB prevalence of infected sett soils
observed in our study may be linked to the low prevalence and the low density of badgers excreting
the bacteria within their social group
Finally in soils of 7 badger setts we were able to detect M bovis signatures (RD4 positive qPCR)
either continuously (4 setts) or discontinuously (3 setts) The presence of M bovis was confirmed by
direct soil DNA spolygotyping (see Table 6) in soils of the setts The presence of M bovis molecular
signatures was proved over an eight month-period (despite the badgers moved out) These results are
in agreement with a previous study where positive molecular detection of M bovis was demonstrated
over a period of 15 months despite the badgers having moved out (Young et al 2005) To our
knowledge this is the first time that spoligotype SB0120 was observed by direct spoligotyping of soil
DNA this spoligotype being identical to the spoligotype of strains isolated from cattle and wildlife in
these sites (Payne et al 2012)
Although the molecular detection of MTBC or M bovis by specific qPCR assay doesnrsquot give any
information about mycobacterial viability Young et al (Young et al 2005) demonstrated that MTBC DNA
did not persist in soil for more than ten days suggesting that DNA detected in our environmental
samples could belong to viable cells
A remaining challenge is to develop adequate methods to cultivate M bovis from
environmental matrices Indeed culture based methods might also be impaired by the occurrence of
VBNC state or dormancy in M bovis present in environmental matrices as previously suggested by
Whittington et al (Whittington et al 2004) for Mycobacterium avium subsp paratuberculosis Further
work is needed to assess if VBNC or dormancy occur in M bovis under environmental conditions
Although we concentrated our sampling efforts on badger environment it is however
acknowledged that in Cocircte drsquoOr M bovis circulates in a multi-host system that includes also wild boar
and deer We can thus hypothesize that these wild animals can also contribute to the shedding of
M bovis in the environment
Further research is needed to understand the ecology of M bovis in the environment and its
epidemiological role in indirect transmission This knowledge would help to improve herd
management in reducing indirect contacts between livestock and wildlife in controlling water access
in areas considered as ldquoat riskrdquo and in preventing contacts with badger setts for example Biosecurity
should thus be an integral part of the long-term strategy for the management of bovine tuberculosis in
Cocircte drsquoOr
Chapitre 3
116
Acknowledgements
The authors thank F Chevalier and M Beacuteral from the CIREV (Cellule Inter-Reacutegionale
drsquoEpideacutemiologie Veacuteteacuterinaire) AE Taconet F Bartheacuteleacutemy and T Grosjean from the Cote drsquoOr Regional
Veterinary Service (DDPP21) We are grateful to the cattle breeders trappers and hunters who helped
us during this study We wish also to thank C Ferrier from ONCFS (National Game Hunting and Wildlife
Agency) Dominique Meunier and Luc Biju-Duval from INRA Dijon Ceacuteline Richomme from ANSES The
work was supported by the Conseil Reacutegional de Bourgogne and Conseil Geacuteneacuteral de Cocircte drsquoOr Interbev
GDS Cocircte drsquoOr
Conflict of interest
No conflict of interest declared
References
Adams AP Bolin SR Fine AE Bolin CA and Kaneene JB (2013) Comparison of PCR versus culture for detection of Mycobacterium bovis after experimental inoculation of various matrices held under environmental conditions for extended periods Appl Environ Microbiol 79 6501-6506
Aranaz A De Juan L Montero N Saacutenchez C Galka M Delso C Alvarez J Romero B Bezos J Vela AI Briones V Mateos A and Domiacutenguez L (2004) Bovine tuberculosis (Mycobacterium bovis) in wildlife in Spain J Clin Microbiol 42 2602-2608
Betelli L Duquenne P Grenouillet F Simon X Scherer E Gehin E and Hartmann A (2013) Development and evaluation of a method for the quantification of airborne Thermoactinomyces vulgaris by real-time PCR J Microbiol Methods 92 25-32
Biet F Boschiroli ML Thorel MF and Guilloteau LA (2005) Zoonotic aspects of Mycobacterium bovis and Mycobacterium avium-intracellulare complex (MAC) Vet Res 36 411-436
Boumlhm M Hutchings MR and White PCL (2009) Contact networks in a wildlife-livestock host community identifying high-risk individuals in the transmission of bovine TB among badgers and cattle PLoS One 4 e5016
Brodin P Eiglmeier K Marmiesse M Billault A Garnier T Niemann S Cole ST and Brosch R (2002) Bacterial artificial chromosome-based comparative genomic analysis identifies Mycobacterium microti as a natural ESAT-6 deletion mutant Infect Immun 70 5568-5578
Clifton-Hadley RS Wilesmith JW and Stuart Fa (1993) Mycobacterium bovis in the European badger (Meles meles) epidemiological findings in tuberculous badgers from a naturally infected population Epidemiol Infect 111 9-19
Corner LA OMeara D Costello E Lesellier S and Gormley E (2012) The distribution of Mycobacterium bovis infection in naturally infected badgers Vet J 194 166-172
Corner LA Costello E Lesellier S OMeara D Sleeman DP and Gormley E (2007) Experimental tuberculosis in the European badger (Meles meles) after endobronchial
Chapitre 3
117
inoculation of Mycobacterium bovis I Pathology and bacteriology Res Vet Sci 83 53-62
Courcoul A Moyen J-L Brugegravere L Faye S Heacutenault S Gares H and Boschiroli M-L (2014) Estimation of sensitivity and specificity of bacteriology histopathology and PCR for the confirmatory diagnosis of bovine tuberculosis using latent class analysis PLoS One 9 e90334
Courtenay O Reilly LA Sweeney FP Hibberd V Bryan S Ul-Hassan A Newman C Macdonald DW Delahay RJ Wilson GJ and Wellington EMH (2006) Is Mycobacterium bovis in the environment important for the persistence of bovine tuberculosis Biol Lett 71 1946-1952
Dean GS Rhodes SG Coad M Adam O Cockle PJ Clifford DJ Hewinson RG and Vordermeier HM (2005) Minimum infective dose of Mycobacterium bovis in cattle Infect Immun 73 6467-6471
Dequiedt S Saby NPa Lelievre M Jolivet C Thioulouse J Toutain B Arrouays D Bispo a Lemanceau P and Ranjard L (2011) Biogeographical patterns of soil molecular microbial biomass as influenced by soil characteristics and management Global Ecol Biogeogr 20 641-652
Falkinham JO (2009) Surrounded by mycobacteria nontuberculous mycobacteria in the human environment J Appl Microbiol 107 356-367
Fediaevsky A Courcoul A Boschiroli M-L and Reveillaud E (2014) Tuberculose bovine en France reacutesultats dune strateacutegie plus offensive Bull Epid Santeacute Anim Alim 64 4-11
Fine AE (2006) The role of indirect transmission in the epidemiology of bovine tuberculosis in cattle and white-tailed deer in Michigan Michigan Bovine Tuberculosis Bibliography and DatabasePaper httpdigitalcommonsunledumichbovinetb33
Fine AE Bolin Ca Gardiner JC and Kaneene JB (2011) A study of the persistence of Mycobacterium bovis in the environment under natural weather conditions in Michigan USA Vet Med Int 2011
Garnett BT Delahay RJ and Roper TJ (2002) Use of cattle farm resources by badgers (Meles meles) and risk of bovine tuberculosis (Mycobacterium bovis) transmission to cattle Proc R Soc Lond B Biol Sci 269 1487-1491
Gavier-Widen D Chambers MA Palmer N Newell DG and Hewinson RG (2001) Pathology of natural Mycobacterium bovis infection in European badgers (Meles meles) and its relationship with bacterial excretion Vet Rec 148 299-304
Ghodbane R Mba Medie F Lepidi H Nappez C and Drancourt M (2014) Long-term survival of tuberculosis complex mycobacteria in soil Microbiology 160 496-501
Gordon SV Heym B Parkhill J Barrell B and Cole ST (1999) New insertion sequences and a novel repeated sequence in the genome of Mycobacterium tuberculosis H37Rv Microbiology 145 881-892
Gortaacutezar C Torres MJ Vicente J Acevedo P Reglero M de la Fuente J Negro JJ and Aznar-Martiacuten J (2008) Bovine tuberculosis in Dontildeana Biosphere Reserve The role of wild ungulates as disease reservoirs in the last Iberian lynx strongholds PLoS One 3 e2776
Chapitre 3
118
Hauer A Cruz KD Cochard T Godreuil S Karaoui C Henault S Bulach T Banuls AL Biet F and Boschiroli M-L (2015) Genetic evolution of Mycobacterium bovis causing tuberculosis in livestock and wildlife in France since 1978 PLoS One 10 e0117103
Huard RC de Oliveira Lazzarini LC Butler WR van Soolingen D and Ho JL (2003) PCR-based method to differentiate the subspecies of the Mycobacterium tuberculosis complex on the basis of genomic deletions J Clin Microbiol 41 1637-1650
Huard RC Fabre M de Haas P Lazzarini LCO van Soolingen D Cousins D and Ho JL (2006) Novel genetic polymorphisms that further delineate the phylogeny of the Mycobacterium tuberculosis complex J Bacteriol 188 4271-4287
King HC Murphy A James P Travis E Porter D Hung Y-J Sawyer J Cork J Delahay RJ Gaze W Courtenay O and Wellington EM (2015a) The variability and seasonality of the environmental reservoir of Mycobacterium bovis shed by wild European badgers Sci Rep 5 1-7
King HC Murphy A James P Travis E Porter D Sawyer J Cork J Delahay RJ Gaze W Courtenay O and Wellington EM (2015b) Performance of a noninvasive test for detecting Mycobacterium bovis shedding in European badger (Meles meles) populations J Clin Microbiol 53 2316-2323
Kukielka E Barasona JA Cowie CE Drewe JA Gortazar C Cotarelo I and Vicente J (2013) Spatial and temporal interactions between livestock and wildlife in South Central Spain assessed by camera traps Prev Vet Med 112 213-221
Little TW Naylor PF and Wilesmith JW (1982) Laboratory study of Mycobacterium bovis infection in badgers and calves Vet Rec 111 550-557
Locatelli A Depret G Jolivet C Henry S Dequiedt S Piveteau P and Hartmann A (2013) Nation-wide study of the occurrence of Listeria monocytogenes in French soils using culture-based and molecular detection methods J Microbiol Methods 93 242-250
Matheson CD Gurney C Esau N and Lehto R (2010) Assessing PCR inhibition from humic substances Open Enzym Inhib J 3 38-45
Michelet L de Cruz K Zanella G Aaziz R Bulach T Karoui C Heacutenault S Joncour G and Boschiroli ML (2015) Infection with Mycobacterium microti in animals in France J Clin Microbiol 53 981-985
Morris RS Pfeiffer DU and Jackson R (1994) The epidemiology of Mycobacterium bovis infections Vet Microbiol 40 153-177
Neill S Hanna J OBrien J and McCracken R (1988) Excretion of Mycobacterium bovis by experimentally infected cattle Vet Rec 123 340-343
Oostenbrink M (1960) Estimating nematode populations by some selected methods Nematology 6(8) 165-2015
Opel KL Chung D and McCord BR (2010) A study of PCR inhibition mechanisms using real time PCR J Forensic Sci 55 25-33
Palmer MV Waters WR and Whipple DL (2004) Investigation of the transmission of
Chapitre 3
119
Mycobacterium bovis from deer to cattle through indirect contact Am J Vet Res 65 1483-1489
Payne A (2014) Rocircle de la faune sauvage dans le systegraveme multi-hocirctes de Mycobacterium bovis et risque de transmission entre faune sauvage et bovins eacutetude expeacuterimentale en Cocircte drsquoOr In Epidemiologie Universiteacute Claude Bernard Lyon 1 httpwwwthesesfr2014LYO10040
Payne A Boschiroli ML Gueneau E Moyen JL Rambaud T Dufour B Gilot-Fromont E and Hars J (2012) Bovine tuberculosis in ldquoEurasianrdquo badgers (Meles meles) in France Eur J Wildlife Res 59 331-339
Phillips CJC Foster CRW Morris PA and Teverson R (2003) The transmission of Mycobacterium bovis infection to cattle Res Vet Sci 74 1-15
Pontiroli A Travis ER Sweeney FP Porter D Gaze WH Mason S Hibberd V Holden J Courtenay O and Wellington EMH (2011) Pathogen quantitation in complex matrices a multi-operator comparison of DNA extraction methods with a novel assessment of PCR inhibition PLoS One 6 e17916
Radomski N Roguet A Lucas FS Veyrier FJ Cambau E Accrombessi H Moilleron R Behr MA and Moulin L (2013) atpE gene as a new useful specific molecular target to quantify Mycobacterium in environmental samples BMC Microbiol 13 277
Richomme C Boschiroli ML Hars J Casabianca F and Ducrot C (2010) Bovine tuberculosis in livestock and wild boar on the Mediterranean island Corsica J Wildl Dis 46 627-631
Santos N Santos C Valente T Gortazar C Almeida V and Correia-Neves M (2015) Widespread environmental contamination with Mycobacterium tuberculosis Complex revealed by a molecular detection protocol PLoS One 10 e0142079
Slana I Kaevska M Kralik P Horvathova A and Pavlik I (2010) Distribution of Mycobacterium avium subsp avium and M a hominissuis in artificially infected pigs studied by culture and IS901 and IS1245 quantitative real time PCR Vet Microbiol 144 437-443
Sweeney FP Courtenay O Hibberd V Hewinson RG Reilly LA Gaze WH and Wellington EMH (2007) Environmental monitoring of Mycobacterium bovis in badger feces and badger sett soil by Real-Time PCR as confirmed by immunofluorescence immunocapture and cultivation Appl Environ Microbiol 73 7471-7473
Sweeney FP Courtenay O Ul-Hassan A Hibberd V Reilly LA and Wellington EMH (2006) Immunomagnetic recovery of Mycobacterium bovis from naturally infected environmental samples Lett Appl Microbiol 43 364-369
Telenti A Marchesi F Balz M Bally F Bottger EC and Bodmer T (1993) Rapid identification of mycobacteria to the species level by polymerase chain reaction and restriction enzyme analysis J Clin Microbiol 31 175-178
Thornton CG MacLellan KM Brink TL Lockwood DE Romagnoli M Turner J Merz WG Schwalbe RS Moody M Lue Y and Passen S (1998) Novel method for processing respiratory specimens for detection of mycobacteria by using C-18-carboxypropylbetaine Blinded study J Clin Microbiol 36 1996-2003
Travis ER Gaze WH Pontiroli A Sweeney FP Porter D Mason S Keeling MJC Jones RM
Chapitre 3
120
Sawyer J Aranaz A Rizaldos EC Cork J Delahay RJ Wilson GJ Hewinson RG Courtenay O and Wellington EMH (2011) An inter-laboratory validation of a real time PCR assay to measure host excretion of bacterial pathogens particularly of Mycobacterium bovis PLoS One 6 e27369
Whittington R Marshall D Nicholls PJ Marsh IB and Reddacliff LA (2004) Survival and dormancy of Mycobacterium avium subsp paratuberculosis in the environment Appl Environ Microbiol 70 2989-3004
Young JS Gormley E and Wellington EM (2005) Molecular detection of Mycobacterium bovis and Mycobacterium bovis BCG (Pasteur) in soil Appl Environ Microbiol 71 1946-1952
Zanella G Durand B Hars J Moutou F Garin-Bastuji B Duvauchelle A Ferme M Karoul C and Boschiroli ML (2008) Mycobacterium bovis in wildlife in France J Wildl Dis 44 99-108
Zhang J Abadia E Refregier G Tafaj S Boschiroli ML Guillard B Andremont A Ruimy R and Sola C (2010) Mycobacterium tuberculosis complex CRISPR genotyping improving efficiency throughput and discriminative power of spoligotyping with new spacers and a microbead-based hybridization assay J Med Microbiol 59 285-294
Chapitre 3
121
C CAMPAGNES DE PRELEVEMENTS SUPPLEMENTAIRES
Entre deacutecembre 2014 et mars 2015 deux campagnes de preacutelegravevement drsquoeacutechantillons
environnementaux suppleacutementaires ont eacuteteacute reacutealiseacutees dans la zone endeacutemique de Cocircte drsquoOr ainsi
qursquoune campagne en Charente ougrave des foyers bovins et sauvages sont recenseacutes respectivement depuis
2006 et 2009
1 Fin des preacutelegravevements en Cocircte drsquoOr
Les preacutelegravevements ont concerneacute drsquoune part une forecirct domaniale agrave cheval sur deux communes
de la zone Sud (eacutetoiles blanches sur la Figure 19) dans laquelle nous nous sommes focaliseacutes sur les
trous drsquoeau freacutequenteacutes par les sangliers et les cervideacutes Cinq trous drsquoeau importants et freacutequenteacutes ont
eacuteteacute cibleacutes et sur chacun trois preacutelegravevements de solsseacutediments et deux preacutelegravevements drsquoeau ont eacuteteacute
reacutealiseacutes en deacutecembre 2014 Drsquoautre part des preacutelegravevements ont eacuteteacute effectueacutes dans les terriers de
blaireaux infecteacutes de la commune D citeacutee dans lrsquoarticle preacuteceacutedent que nous suivons depuis mars 2014
et dans laquelle 1 agrave 3 blaireaux infecteacutes sont pieacutegeacutes tous les ans ainsi que dans des terriers freacutequenteacutes
drsquoune commune limitrophe (commune G) sur laquelle un blaireau infecteacute a eacuteteacute pieacutegeacute en 2014 Quatorze
sols de terriers (5 sur la commune D et 9 sur la commune G) et cinq latrines proches des terriers (3 sur
D et 2 sur G) ont eacuteteacute preacuteleveacutes en mars 2015 Ces deux sites sont repreacutesenteacutes par les eacutetoiles noires sur
la Figure 19 Tous ces eacutechantillons ont eacuteteacute analyseacutes selon les techniques deacutecrites dans lrsquoarticle
preacuteceacutedent (section Mateacuteriel et meacutethodes)
Les vingt-cinq eacutechantillons preacuteleveacutes dans la forecirct domaniale de la zone Sud en deacutecembre 2014
nrsquoont donneacute aucun signal moleacuteculaire avec le systegraveme de deacutetection ciblant lrsquoIS1561rsquo Dans cette zone
forestiegravere aucun animal sauvage infecteacute nrsquoa eacuteteacute deacutetecteacute depuis 2013
Concernant les terriers et latrines de la commune D les cinq eacutechantillons de sols de terriers
analyseacutes et une des deux latrines ont donneacute un signal moleacuteculaire positif pour le MTBC Pour lrsquoun des
sols la quantification nrsquoa pas eacuteteacute possible avec la qPCR (Ct gt 39 traces) Pour les quatre autres sols
des valeurs de 10 times 104 agrave 46 times 104 copies de gegravenes ont eacuteteacute obtenues et de 12 times 104 copies de gegravenes
pour la latrine Trois des eacutechantillons de sol ont donneacute un reacutesultat positif avec la PCR nicheacutee ciblant
lrsquoIS1561rsquo Dans ces terriers les preacutelegravevements reacutealiseacutes en mars avril mai juin et novembre 2014 puis
mars 2015 ont montreacute une persistance du signal moleacuteculaire au cours du temps Pour quatre des
terriers le signal a eacuteteacute deacutetecteacute agrave chaque campagne de preacutelegravevements alors que pour trois autres la
deacutetection a eacuteteacute intermittente Il est inteacuteressant de noter que les valeurs obtenues en mars 2015 sont
supeacuterieures agrave celles obtenues en 2014 (voir article preacuteceacutedent) Plusieurs hypothegraveses sont eacutemises pour
Chapitre 3
122
expliquer ce reacutesultat La premiegravere est lieacutee au fait que les terriers eacutetaient habiteacutes lors de nos
preacutelegravevements en mars 2015 alors qursquoils ne lrsquoeacutetaient plus apregraves les preacutelegravevements de mars 2014
La surveillance videacuteo reacutealiseacutee sur ces terriers a montreacute que les terriers ont agrave nouveau eacuteteacute
occupeacutes agrave partir de septembre 2014 Il est possible que cette occupation ait geacuteneacutereacute lrsquoexteacuteriorisation de
sol profond et de litiegravere potentiellement plus contamineacutes et que nous ayons preacuteleveacute ce sol drsquoorigine
profonde La seconde hypothegravese est lieacutee au fait que nous avons preacuteleveacute plus profondeacutement dans le
terrier que dans les campagnes de preacutelegravevements preacuteceacutedentes (utilisation drsquoun aspirateur) Enfin il est
eacutegalement possible que le groupe de blaireaux peuplant ce terrier possegravede un ou plusieurs animaux
infecteacutes et excreacuteteurs agrave lrsquoorigine drsquoune contamination plus importante du sol du terrier Ces terriers sont
situeacutes dans le bosquet drsquoun preacute qui fournit aux bovins de lrsquoombre et de la fraicheur en eacuteteacute et un abri en
cas de pluie ou de vent La surveillance videacuteo de cette zone nous a permis drsquoobserver que plusieurs
veaux ont reacuteguliegraverement exploreacute les entreacutees de gueules et consommeacute les brins drsquoherbe autour de ces
entreacutees (surveillance videacuteo)
Sur les neuf eacutechantillons de sol preacuteleveacutes dans la commune G deux ont donneacute un signal positif
pour le MTBC Pour lrsquoun drsquoeux seules des traces de M bovis ont eacuteteacute deacutetecteacutees pour lrsquoautre le nombre
de copies de gegravenes a eacuteteacute estimeacute agrave 12 times 104 La PCR nicheacutee srsquoest reacuteveacuteleacutee neacutegative pour les deux
eacutechantillons
Chapitre 3
123
Figure 19 Localisation des communes de Cocircte drsquoOr sur lesquelles de foyers bovins et des animaux sauvages infecteacutes ont eacuteteacute deacutetecteacutes et localisation des preacutelegravevements environnementaux effectueacutes en 2015 (les preacutelegravevements positifs sont repreacutesenteacutes par des eacutetoiles noires) (Reacutealisation Marina Beacuteral CIREV Bourgogne)
2 Preacutelegravevements en Charente
Depuis 2006 25 foyers bovins ont eacuteteacute deacutetecteacutes en Charente avec 12 foyers pour la seule anneacutee
2014 et la deacutecouverte de foyers anciens qui eacutevoluaient agrave bas bruit (communication personnelle
DDPP16) Depuis 2009 lrsquoinfection est mise en eacutevidence chez les blaireaux (6 positifs en 2013 et 11 en
2014) et les sangliers (8 positifs en 2014 donneacutees Sylvatub) et un chevreuil en 2015 (Communication
personnelle Sylvatub) autour des foyers bovins avec les mecircmes spoligotypes Le systegraveme multi-hocirctes
observeacute diffegravere donc de celui de la Cocircte drsquoOr puisqursquoagrave lrsquoheure actuelle lrsquoinfection nrsquoa pas eacuteteacute observeacutee
chez les cerfs dont les densiteacutes sont beaucoup moins importantes qursquoen Cocircte drsquoOr Les foyers bovins
Chapitre 3
124
et sauvages sont majoritairement localiseacutes dans le sud du deacutepartement dans une zone limitrophe de
la Dordogne deacutepartement fortement infecteacute depuis 2005 (Figure 20)
A la demande de la DDPP16 nous sommes alleacutes preacutelever des eacutechantillons environnementaux
dans le Sud du deacutepartement dans sept communes fortement toucheacutees par la maladie (Figure 20) Entre
avril et mai 2015 55 eacutechantillons environnementaux (50 eacutechantillons de sol de terrier de blaireau 3
sols de souille 1 latrine de blaireau et 1 fegraveces de sanglier) ont eacuteteacute preacuteleveacutes sur 7 communes de
Charente Ces communes font lrsquoobjet de la deacuteclaration drsquoun ou plusieurs foyers bovins de tuberculose
bovine etou drsquoanimaux sauvages infecteacutes (Figure 20) De plus 3 eacutechantillons de matiegraveres feacutecales
preacuteleveacutes pendant lrsquoautopsie de 3 blaireaux (BLAIR 7057 BLAIR 7058 et BLAIR 7061) ont eacuteteacute analyseacutes
Six eacutechantillons positifs ont eacuteteacute mis en eacutevidence sur les 55 analyseacutes La qPCR ciblant lrsquoIS1561rsquo a permis
de deacutetecter et de quantifier M bovis dans quatre ADN de sol de terrier avec un nombre de copies de
gegravenes variant de 11 times 103 agrave 54 times 103 par g de sol Deux eacutechantillons suppleacutementaires (1 sol de souille
et 1 sol de terrier) ont donneacute un signal non quantifiable avec la qPCR (traces) Tous les eacutechantillons
positifs ont eacuteteacute preacuteleveacutes sur une mecircme commune sur 2 lieux-dits diffeacuterents Maine-Joli (4 sols de
terrier) et la Tuilerie (1 sol de terrier et 1 sol de souille) A notre connaissance 2 blaireaux et 3 sangliers
infecteacutes ont eacuteteacute deacutecouverts agrave proximiteacute du lieu-dit la Tuilerie deacutebut 2015 (Source DDPP16) Lors de
notre campagne de preacutelegravevement tous les terriers deacutetecteacutes positifs eacutetaient freacutequenteacutes
Les ADN des eacutechantillons positifs ont eacuteteacute envoyeacutes agrave lrsquoANSES pour geacutenotypage Aucun ADN nrsquoa toutefois
pu ecirctre spoligotypeacute
Les fegraveces de blaireaux (BLAIR 7057 BLAIR 7058 et BLAIR 7061) preacuteleveacutes lors des autopsies nrsquoont donneacute
aucun signal avec la qPCR
Chapitre 3
125
Figure 20 Localisation des communes de Charente sur lesquelles de foyers bovins et des animaux sauvages infecteacutes ont eacuteteacute deacutetecteacutes et localisation des preacutelegravevements environnementaux effectueacutes en 2015 (les preacutelegravevements positifs sont repreacutesenteacutes par des eacutetoiles noires) (Reacutealisation Marina Beacuteral CIREV Bourgogne)
3 Compleacutements paysage geacuteomorphologie et peacutedologie
La zone endeacutemique Nord de tuberculose bovine en Cocircte drsquoOr que nous avons principalement
eacutetudieacutee est situeacutee sur un territoire appeleacute lrsquoAuxois caracteacuteriseacute par un paysage bocager ponctueacute de
buttes et de plateaux calcaires ougrave affleure un calcaire dur et karstique Les pacirctures suspectes
preacutesentent un contexte paysager hydrologique et geacuteomorphologique commun Le parcellaire suspect
est localiseacute dans trois petites valleacutees parallegraveles au fond desquelles circule un cours drsquoeau en direction
du Nord-Ouest (Figure 21) Chaque cours drsquoeau est alimenteacute par des sources naissant entre les couches
calcaires et marneuses du sous-sol dans les pentes Ces reacutesurgences drsquoeau proviennent de lrsquoinfiltration
des preacutecipitations dans les plateaux calcaires agrave lrsquoorigine du deacuteveloppement drsquoun reacuteseau karstique
drainant drsquoimportants eacutecoulements souterrains et la reacutesurgence drsquoeau en surface agrave cause des marnes
impermeacuteables du sous-sol Les pacirctures sont situeacutees sur les pentes douces marneuses des buttes ainsi
que dans les valleacutees alluviales humides Au-dessus des pacirctures un bourrelet boiseacute marque la rupture
Chapitre 3
126
de pente il preacutesente en geacuteneacuteral des affleurements plus ou moins importants de calcaire dur (Figure
22) Les pacirctures sont majoritairement situeacutees sur la mecircme formation geacuteologique des marnes du Lias
ou du Jurassique infeacuterieur correspondant agrave des formations peacutedologiques particuliegraveres caracteacuteriseacutees
par des sols de pentes argilo-limoneux localement hydromorphes (sources et laquo mouilles raquo)
Il est inteacuteressant drsquoobserver que les cantons toucheacutes par la tuberculose bovine en Cocircte drsquoOr
possegravedent une densiteacute de bovins allaitant quasi-identique agrave celle des cantons indemnes localiseacutes dans
le sud-ouest du deacutepartement et limitrophe de la zone drsquoendeacutemie de bTB (Figure 23) Ces deux groupes
de cantons sont localiseacutes sur des formations peacutedologiques diffeacuterentes suggeacuterant que le type de sol des
pacirctures pourrait ecirctre un facteur de risque dans le maintien de la maladie et que la densiteacute des bovins
ne semble pas ecirctre un facteur majeur dans la distribution des foyers de bTB Lrsquoobservation de la
distribution du parcellaire des exploitations foyers de tuberculose bovine sur la carte peacutedologique du
deacutepartement montre que 74 des parcelles de ces exploitations sont localiseacutees sur cette uniteacute
paysagegravere (sols argilo-limoneux et hydromorphes des pentes 48 des parcelles sols calcaires humides
sur alluvions des valleacutees 14 et sols argileux des plateaux calcaires 12 ) (Figure 24Figure 24)
Une enquecircte (Etude PatRisk) reacutealiseacutee dans le cadre drsquoun Master drsquoEpideacutemiologie dans neuf
eacutelevages bovins deacuteclareacutes infecteacutes entre 2010 et 2014 a chercheacute agrave identifier les pacirctures laquo agrave risque raquo de
tuberculose et agrave les caracteacuteriser par des variables concernant les pratiques drsquoeacutelevage le voisinage la
faune sauvage et lrsquoenvironnement (pente type de sol relief exposition humiditeacute) Lrsquoanalyse
multivarieacutee a mis en eacutevidence deux expositions indicatrices de risque la pratique du fauchage avant
la mise agrave lrsquoherbe et la preacutesence de zones humides localiseacutees sur la pacircture (Bouchez-Zacria 2015) Ces
reacutesultats sont en accord avec le fait que la survie de M bovis est favoriseacutee par la preacutesence drsquohumiditeacute
et qursquoune faible hauteur drsquoherbe augmente la possibiliteacute de contact avec les mycobacteacuteries du sol Bien
que des eacutetudes plus preacutecises soient recommandeacutees il semble toutefois que les sols argilo-limoneux
hydromorphes des pacirctures suspectes constituent un facteur de risque agrave prendre en compte dans
lrsquoeacutepideacutemiologie de la maladie
Chapitre 3
127
Figure 21 Orographie et hydrographie de lrsquoAuxois Les valleacutees supportant les parcelles suspectes sont indiqueacutees par une flegraveche noire (Source BD Carthage et BD alti - IGN)
Figure 22 Localisation de lrsquoAuxois en Cocircte drsquoOr structure paysagegravere et geacuteomorphologie de ce territoire
Chapitre 3
128
Figure 23 Nombre de vaches allaitantes par canton dans le deacutepartement de la Cocircte drsquoOr (agrave gauche) et parcellaire des foyers bovins de 2010 agrave 2015 (agrave droite) (Source Recensement agricole 2010 ndash Agreste)
Chapitre 3
129
Figure 24 Reacutepartition spatiale des parcelles des eacutelevages bovins infecteacutes de 2010 agrave 2014 en Cocircte drsquoOr et repreacutesentation des couches peacutedologiques (Reacutealisation Marina Beacuteral CIREV Bourgogne)
Chapitre 3
130
D BILAN GLOBAL
Nous avons deacuteveloppeacute deux systegravemes de deacutetection moleacuteculaire adapteacutes aux matrices
environnementales ciblant les espegraveces du MTBC (IS1561rsquo et Rv3866) et modifieacute un systegraveme existant
speacutecifique de M bovis (RD4) Nous avons valideacute ces systegravemes sur trois matrices diffeacuterentes (sol eau
fegraveces) inoculeacutees avec M bovis BCG en deacutemontrant leur speacutecificiteacute soit pour le MTBC (excluant
M microti potentiellement preacutesent dans lrsquoenvironnement) soit pour M bovis Leur sensibiliteacute est
bonne puisque les seuils de deacutetection obtenus sont infeacuterieurs aux seuils deacutecrits dans la litteacuterature (en
comparant avec une meacutethode de deacutenombrement identique agrave la nocirctre) Le systegraveme RD4 est agrave ce jour le
seul systegraveme de deacutetection speacutecifique de M bovis dans les eacutechantillons environnementaux toutefois
drsquoapregraves notre expeacuterience il preacutesente une sensibiliteacute moins importante que les deux autres systegravemes
malgreacute des seuils de deacutetection eacutequivalents sur les eacutechantillons inoculeacutes au laboratoire
La deacutetection du MTBC par lrsquoamplification de lrsquoIS1561rsquo a eacuteteacute effective dans 42 eacutechantillons (sur
plus de 400) la PCR nicheacutee ciblant la mecircme seacutequence a mis en eacutevidence 14 eacutechantillons positifs et le
systegraveme RD4 trois eacutechantillons positifs Les niveaux de deacutetection varient fortement en fonction des
eacutechantillons lrsquoeffet de dilution reste tregraves important dans des matrices comportant des communauteacutes
microbiennes abondantes et diversifieacutees (quantiteacute relative drsquoADN cible faible par rapport agrave des
quantiteacutes eacuteleveacutees drsquoADN non cible) Lrsquoutilisation de la PCR quantitative a permis dans certains cas de
quantifier les cibles preacutesentes dans les eacutechantillons environnementaux Les sols de terriers preacutesentent
les concentrations en mycobacteacuteries du MTBC les plus eacuteleveacutees pouvant atteindre 46 times 104 copies du
gegravene cibleacute ce qui correspond agrave des quantiteacutes de cellules eacuteleveacutees sachant que le nombre de copies du
gegravene cible par geacutenome est de 1
Parmi les diffeacuterents sites preacuteleveacutes la contamination a eacuteteacute retrouveacutee exclusivement dans deux
eacutecosystegravemes particuliers les zones humides de pacirctures (sources et laquo mouilles raquo) et les sols de terrier
et latrines de blaireaux La contamination apparait le plus souvent localiseacutee agrave une zone geacuteographique
et ponctuelle mais un signal moleacuteculaire positif a eacuteteacute retrouveacute pendant un an dans les sols de terriers
de blaireaux localiseacutes sur une pacircture
Ces reacutesultats correacuteleacutes au contexte paysager et peacutedologique des parcelles suspectes nous ont
ameneacute agrave penser que certaines formations peacutedologiques seraient plus susceptibles de favoriser la
persistance de M bovis Nous avons donc eacutevalueacute lrsquoimpact de la nature du sol retrouveacute dans ces
parcelles sur la survie de M bovis Nous avons eacutegalement eacutevalueacute lrsquoimpact de la tempeacuterature sur la
dureacutee de survie de la bacteacuterie dans le sol
CHAPITRE 4
Impact de la tempeacuterature et de la nature du sol sur la
survie de Mycobacterium bovis SB0120
Elodie Barbier Murielle Rochelet Laurent Gal Maria Laura Boschiroli Alain Hartmann
Experimental investigation of the impact of the temperature and the soil type on Mycobacterium
bovis survival in the environment
(Article en cours de soumission)
133
CHAPITRE 4 - Impact de la tempeacuterature et de la nature du sol sur la
survie de Mycobacterium bovis SB0120
A AVANT-PROPOS
La deacutetection moleacuteculaire de Mycobacterium bovis dans les eacutecosystegravemes agricoles de la zone
endeacutemique de Cocircte drsquoOr a confirmeacute que les animaux infecteacutes et excreacuteteurs ont la capaciteacute de
contaminer les sols et lrsquoeau accessibles agrave la faune domestique et sauvage (Chapitre 2) La bacteacuterie ne
se multipliant pas dans lrsquoenvironnement sa survie dans les substrats environnementaux conditionne la
transmission indirecte du bacille entre un animal infecteacute excreacuteteur et un animal sain par le biais de ces
substrats
Des eacutetudes expeacuterimentales sur divers substrats environnementaux notamment du sol
inoculeacutes avec M bovis ou M bovis BCG ont suggeacutereacute que les principaux facteurs influenccedilant la survie
des bacteacuteries sont les facteurs climatiques tels que tempeacuterature humiditeacute et exposition aux UV (Fine
et al 2011 Duffield amp Young 1985 Jackson et al 1995 Young et al 2005) Bien que quelques eacutetudes
aient rechercheacute par la culture ou par des approches statistiques une eacuteventuelle association de la
contamination environnementale ou animale par des mycobacteacuteries avec la nature du sol les reacutesultats
sont difficilement comparables et extrapolables (Walter et al 2012 Dhand et al 2009 Reviriego et al
2000 Johnson-Ifearulundu amp Kaneene 1999 Ward amp Perez 2004)
Face agrave ce manque de connaissances nous avons donc souhaiteacute eacutevaluer la dureacutee de la survie de
M bovis dans deux sols de Cocircte drsquoOr preacutesentant des caracteacuteristiques physico-chimiques contrasteacutees et
ce agrave deux tempeacuteratures diffeacuterentes afin de rechercher un eacuteventuel impact de la tempeacuterature et de la
nature du sol sur la persistance de la bacteacuterie Pour cette eacutetude nous avons utiliseacute une souche
pathogegravene de M bovis (geacutenotype SB0120) ce geacutenotype est retrouveacute majoritairement dans la zone
Nord chez les bovins et dans la faune sauvage Deux meacutethodes de suivi de la population bacteacuterienne
ont eacuteteacute utiliseacutees la culture sur milieu geacuteloseacute reacutealiseacutee agrave partir de sols irradieacutes et la qPCR pour le suivi
des sols irradieacutes et biotiques
135
B EXPERIMENTAL INVESTIGATION OF THE IMPACT OF THE TEMPERATURE AND THE SOIL TYPE ON MYCOBACTERIUM BOVIS SURVIVAL IN THE ENVIRONMENT
Elodie Barbierab Murielle Rocheletab Laurent Galac Maria Laura Boschirolid Alain Hartmanna
a INRA UMR 1347 Agroeacutecologie Dijon Cedex France
b Universiteacute de Bourgogne Franche Comteacute Dijon Cedex France
c AgroSup Dijon UMR1347 Agroeacutecologie Dijon France
d Universiteacute Paris-Est Laboratoire National de Reacutefeacuterence de la Tuberculose Uniteacute des Zoonoses Bacteacuteriennes Laboratoire de
Santeacute Animale ANSES Maisons-Alfort Cedex France
Key words Mycobacterium bovis soil temperature culture quantitative PCR
Corresponding Author Alain Hartmann UMR 1347 Agroeacutecologie INRA 17 rue Sully BP 86510 21065
Dijon Cedex France E-mail alainhartmanndijoninrafr
INRA UMR1347 Agroeacutecologie BP 86510 F-21000 Dijon France
Chapitre 4
136
Abstract
Mycobacterium bovis the causative agent of the bovine tuberculosis (bTB) mainly affect cattle its
natural reservoir but also a wide range of domestic and wild mammals Besides direct transmission via
contaminated aerosols indirect transmission of the disease between wildlife and livestock is thought
to occur by inhalation or ingestion of environmental substrates contaminated by M bovis through
infected animal shedding We monitored the survival of M bovis in two soils chosen for their contrasted
physico-chemical properties at two temperatures The population of M bovis spiked in disinfected soils
was enumerated by a culture-based method after 14 30 60 90 120 and 150 days of incubation at 4degC
and 22degC The fate of M bovis spiked in disinfected or biotic soils was monitored using a qPCR based
assay targeting the IS1561rsquo locus The analysis of survival profiles using culture-based method showed
that M bovis survived longer at cooler temperature but impact of soil characteristics on M bovis
persistence was not obvious Futhermore qPCR based assay detected M bovis longer than the culture
based method with higher gene copy numbers in sterile soils than in biotic ones Impact of soil type on
M bovis persistence need to be further studied in order to shed light on the risk and mechanisms of
indirect transmission of the disease
Chapitre 4
137
INTRODUCTION
Mycobacterium bovis is a pathogenic mycobacteria responsible for bovine tuberculosis (bTB)
Tuberculosis caused by M bovis is a primarily respiratory disease that may also affect various organs in
animals (Morris et al 1994) Even if bTB mainly concerns cattle a wide range of domestic and wild
mammals may be infected as well (Biet et al 2005) On the other hand bTB a zoonotic disease
impacting human health (Peacuterez-Lago et al 2013) Although cattle is considered as the principal host of
M bovis wild fauna such as badgers (Meles meles) wild boar (Sus scrofa) and red deer (Cervus elaphus)
has recently been identified as potential wildlife reservoirs (Palmer 2013) Though the transmission of
M bovis among cattle usually occurs via the inhalation of contaminated aerosols released by infected
animals andor the close contacts (Morris et al 1994 Phillips et al 2003) the indirect transmission
either by inhalation of environmental bioaerosols or ingestion of infected environmental matrices may
be another potential routes of animal infection (Vicente et al 2007 Palmer et al 2004 Phillips et al
2003) Indirect transmission through feed or water has already been demonstrated between deer deer
and cattle (Palmer et al 2004 Palmer et al 2001) and badgers and cattle (Little et al 1982) in laboratory
experiments
Investigating this type of indirect transmission is challenging because it results at least from the
combination of three essential factors iei) the environmental contamination by shedding from
infected animals ii) the persistence of the bacteria under a virulent state in environmental matrices
and finally iii) the interaction between a new susceptible host with the contaminated matrices The
shedding step by animals has already been demonstrated for many species via oro-nasal mucus
sputum urine feces and wound discharges depending on the animal species (Corner et al 2012
Palmer et al 2004 Barasona et al 2015) Secondly a few previous experimental studies using various
environmental substrates spiked with M bovis showed that the bacteria was able to survive for a long
time period outside of its host (Williams amp Hoy 1930 Duffield amp Young 1985 Young et al 2005 Fine et
al 2011 Ghodbane et al 2014) For example using a culture-based method Ghodbane et al (2014)
recovered M bovis bacteria from sterile soils incubated in controlled laboratory conditions 12 months
after their inoculation whereas an incubation under the natural weather conditions of Michigan USA
allowed the persistence of the bacteria in sterile soils for 88 days (Fine et al 2011) Moreover several
studies have identified several climatic factors such as a cool temperature an adequate moisture and
a protection against solar radiation (ultra violet) as positive factors for the persistence of M bovis in
environmental matrices like feces (Williams amp Hoy 1930 Tanner amp Michel 1999) food supply such as
corn hay apples (Palmer amp Whipple 2006 Fine et al 2011) water (Little et al 1982 Fine et al 2011)
and soil (Duffield amp Young 1985 Fine et al 2011) (Duffield amp Young 1985 Williams amp Hoy 1930 Tanner
amp Michel 1999 Jackson et al 1995) When Fine et al (2011) compared the persistence of M bovis in
Chapitre 4
138
four substrates (corn hay water and soil) the longest survival was found in cool and moist soil (88
days in winterspring) suggesting that soil probably ensured better conditions of persistence for
M bovis than the other matrices tested Moreover M bovis was often detected in soils especially in
badger sett soil or in pasture soil (Courtenay et al 2006 Santos et al 2015 Sweeney et al 2007
2006 Young et al 2005 Barbier et al manuscript submitted)
More generally low pH and high iron content are often described as major factors driving
mycobacterial survival in soils especially for Mycobacterium avium ssp paratuberculosis (Johnson-
Ifearulundu amp Kaneene 1999 Ward amp Perez 2004) Concerning M bovis one statistical study
established a relationship between the occurrence of bTB in animals and soil parameters by showing
that every a 1 increase in the sandy content of soil led to an increase in odds of bTB infection by 4
(Walter et al 2014) suggesting that soil type may promote or hinder M bovis environmental
persistence
To our knowledge no comprehensive study focuses on the influence of soil biotic and abiotic
parameters on the persistence M bovis With the aim to further assess the role of the soil
characteristics on the persistence of Mbovis and thus to better understand the impact of the
environment in the epidemiology of the bTB disease the influence of both the physicochemical
characteristics of the soil and the temperature was studied under controlled laboratory conditions For
this purpose two different soils selected for their contrasted properties were spiked with known
concentrations of M bovis and incubated at two different temperatures to mimic seasonal
temperatures The persistence of M bovis over time was monitored using both a culture-based
method for the detection of viable M bovis and a molecular detection method based on a qPCR assay
MATERIAL AND METHODS
All experiments were conducted in a biosafety level 3 laboratory unless otherwise stated
Mycobacterium bovis cultivation and inoculum preparation
The strain Mycobacterium bovis SB0120 VNTR 5 5 4 3 11 4 5 6 that infects cattle and wild
animals such as badgers and wild boar in France was provided by the National Reference Laboratory of
bovine tuberculosis (Maisons-Alfort France) and grown aerobically at 37degC in Middlebrook 7H9
supplemented with 10 Middlebrook ADC Growth Supplement (Sigma-Aldrich France) for thirty days
The M bovis inoculum was prepared as following a 20 mL stationary phase culture was pelleted and
once the supernatant removed the cell aggregates were disrupted by vortexing with glass beads for 15
seconds The pellet was suspended in 1 mL of sterile water constituting the stock solution and serial
10-fold dilutions were subsequently plated in triplicate onto Middlebrook 7H11 supplemented with
10 heat inactivated bovine serum (Dominique Dutscher France) and 10 Middlebrook OADC
Chapitre 4
139
Growth Supplement (Sigma-Aldrich France) for enumeration Colonies were counted following
incubation for two months at 37degC The inoculum for the culture-based assay and the qPCR-based assay
were respectively prepared by diluting 10-fold and 1000-fold the M bovis stock solution
Soil characterization and microcosm preparation
Two different soils were collected in two cattle-grazed pastures at 0-30 cm depth in Cocircte drsquoOr
(a department located in the Middle East of France) The soil A is a clay loamy soil (calcisol) and soil B
is a loamy sandy soil Soils were prepared as following soil samples were gently air dried and sieved to
recover 4 mm aggregates The moisture contents of soils were determined by drying for 24 h at 105degC
and their physico-chemical parameters were determined by the Soil Analysis Laboratory of Arras
France Fresh sieved soil was used to prepare the microcosms
Vials of 15 mL (Falcon Dominique Dutscher France) were filled with 2 g of the soils A and B Half of the
soil vials were sterilized by gamma-irradiation up to an internal minimum dose of 35 KGray (Ionisos
France) All the microcosm soils (biotic and sterile) were then adjusted to 90 of the water holding
capacity We verified that neither soil A nor B were positive for M bovis with the culture-based method
(sterile soils) and with the qPCR assay (sterile and biotic soils) described later All the soil microcosms
were spiked with 100 microL of the M bovis inoculum 10- or 1000-fold diluted (for sterile and biotic soil
respectively) and stored in the dark in a biosafety level 3 laboratory until their analysis Sterile spiked
soils were incubated at 4degC and 22degC while biotic spiked soils were only incubated at 22degC Vials were
opened in a microbiological safety cabinet every fifteen days to regenerate their atmosphere
Schedule of microcosm sampling
In order to monitor M bovis cell numbers over time both plate enumeration and qPCR assay
were performed on irradiated soils while qPCR assay only was carried out for biotic soils At each
sampling date ie day 0 14 30 60 90 120 and 150 three mesocosms of each biotic and sterile soils
were analysed with the qPCR assay after DNA extractionpurification and four mesocosms of each
sterile soil at each temperature were analysed using culture-based method
DNA extraction and purification
DNAs from the soil samples were extracted as follows 8 mL of lysis buffer (100 mM Tris pH 80
100 mM EDTA pH 80 100 mM NaCl and 2 (wv) SDS) 4 g of silica beads (100 μm diameter) 5 g of
ceramic beads (14 mm diameter) and 8 glass beads (4 mm diameter) were added to each vial of soil
However as DNA extraction could not be achieved within the biosafety level 3 laboratory for practical
reason soil was suspended in 8 mL of lysis buffer and heated 2 hours at 80degC to kill viable bacteria The
following steps of DNA extraction were then done in a biosafety level 2 laboratory Samples were
Chapitre 4
140
disrupted for 3 times 30 s at 4 ms in a FastPrepreg-24 Instrument (MP Biomedicals Europe France)
incubated for 30 min at 70degC and centrifuged for 5 min at 7000 timesg at room temperature Supernatants
were incubated on ice for 10 min with 110 volume of 3 M potassium acetate pH 55 and then
centrifuged for 5 min at 14000 times g One volume of ice-cold isopropanol was added to the supernatant
for DNA precipitation overnight at minus 20degC DNA was collected by centrifugation (30 min at 14000 times g)
and DNA pellets were washed with ice-cold ethanol (70 ) and dissolved in 100 μL of water Crude DNA
extracts (100 μL) were loaded on Microbiospintrade columns (Biorad France) filled with 100 mg of
polyvinyl polypyrrolidone (PVPP Sigma Chemical Co France) After centrifugation (4 min at 1000 times g
4 degC) eluted DNA was further purified using the Geneclean Turbo Kit (Qiagen France) according to the
manufacturers instructions Purified DNA concentrations were estimated by spectrophotometry (λ =
260 nm) using a NanoDropreg 2000 spectrophotometer (Thermo Scientific France)
Estimation of DNA quality for PCR amplification
The quality of the purified DNAs extracted from the two soils was assessed by measuring the
capacity to amplify a control DNA target in the presence of various dilutions of soil DNA extracts Briefly
qPCR reactions were done in a final volume of 25 microL containing 25 μl of 106 copies of the circularized
pGEM-T Easy plasmid DNA and 5 μl of water (control) or diluted soil DNA (10- and 20-fold dilutions)
1X Absolute qPCR SYBR Green ROX (500 nM) Mix (Thermo Scientific) 25ngmicroL of T4 bacteriophage
gene 32 product qPCR primers were SP6 (5rsquondashATTTAGGTGACACTATAG-3rsquo) and T7 (5rsquo-
TAATACGACTCACTATAGGG-3rsquo) universal primers (final concentration 500 nM) targeting the polylinker
of the plasmid qPCR and cycling conditions were as following initial denaturation at 95degC for 15 min
followed by 40 cycles with denaturation at 95degC for 15 s annealing at 55degC and elongation at 72degC for
30 s each The analysis of the melting curves allowed to assess PCR specificity A tenfold reduction of
the copy number amplified by qPCR in the presence of diluted soil DNA with respect to the copy
number obtained in the control reaction (water) was considered as 10 amplification efficiency or
conversely as 90 inhibition of the PCR
Cuture-based method
The sterile soil microcosms were suspended in 9 mL of Middlebrook 7H9 supplemented with
ADC by vortexing the vials for 2 min Tenfold dilutions of soil suspensions were achieved and 100 microL
were plated in triplicate onto Middlebrook 7H11 agar enriched as described above and supplemented
with antibiotics and antifungal (100 mgL-1Ticarcillin 10 mgL-1 Trimethoprim 200 000 UIL-1 Polymyxin
B and 100 mgL-1 Cycloheximid Sigma-Aldrich France) for enumeration Plates were then incubated in
gas-permeable bags at 37degC for 2 months and checked every 15 days
Chapitre 4
141
Mycobacterium tuberculosis complex quantitative PCR (qPCR)
Mycobacterium bovis was detected using the Mycobacterium tuberculosis complex (MTBC)
specific qPCR targeting the IS1561rsquo locus (Forward 5rsquo- GATCCAGGCCGAGAGAATCTG -3rsquo Reverse 5rsquo-
GGACAAAAGCTTCGCCAAAA - 3rsquo and probe 5rsquo - FAM ACGGCGTTGATCCGATTCCGC TAMRA - 3rsquo) as
previously described (Barbier et al manuscript submitted for publication)
qPCR reactions were carried out in triplicate with a 25 microL reaction mix containing 125 microL of ABsolutetrade
QPCR ROX Mix (Thermo Scientific France) 1 microL (final concentration 25 ngmicroL-1) of T4 bacteriophage
gene 32 product (MP Biomedicals Europe France) 1 microL of each primer (final concentration 400 nmolL-
1) 05 microL of each probe (final concentration 200 nmol L-1) 4 microL of ultrapure water and 5 microL of 20-fold
diluted DNA Absolute quantification was achieved using standard DNA dilutions A recombinant
plasmid containing one copy of the IS1561rsquo fragment of M bovis BCG strain Pasteur 1173P2 cloned in
the pCRregII-TOPOreg vector (Invitrogen France) was used as a standard A calibration curve was obtained
by amplification of serial dilutions of the plasmid ranging from 1 to 106 copies per PCR reaction All
DNAs extracted from the soil samples were screened with the IS1561rsquo-based qPCR assay and results
were expressed as the number of M bovis gene copy number per g of soil qPCR assay was performed
in a VIIA 7 Real-Time PCR System (Thermo Fischer Scientific France) Initial denaturation was done at
95degC for 15 min followed by fourty-five cycles with denaturation at 95degC for 15 s annealing and
elongation at 60degC for 1 min Detection limits of this molecular system was previously determined in
soil (Barbier et al manuscript submitted for publication)
RESULTS
Physico-chemical characteristics of soils A and B
The physico-chemical characteristics of soils A and B are summarized in Table 1 The soil A is a
clay loamy soil with a slightly basic pH (775) and a high content of clay (clay account for 397 in
particle-size distribution and sand for only 118 ) while the soil B is a loamy sandy soil with an acidic
pH (546) and a high content of sand (sand account for 345 in particle-size distribution and clay for
only 192 ) Silt content was roughly equivalent in the two soils (485 g kg-1 for soil A vs 463 g kg-1 for
soil B) We chose to adjust soil moisture to 90 of their water field holding capacity leading to a
moisture content of 31 for soil A and 21 for soil B Finally the C-to-N ratio was 95 for soil A and
109 for soil B
Chapitre 4
142
Table 1 Physico-chemical parameters of soils A and B
Physical parameters Soil A Soil B Unit
Clay (lt 2 microm) 397 192 g kg-1
Silt 485 463 g kg-1
Sand 118 345 g kg-1
Water field capacity 352 247 g kg-1
Chemical parameters -
pH 775 546
Total C 361 292 g kg-1
Total N 38 269 g kg-1
C- to N- ratio 95 109 -
Organic matter 625 506 g kg-1
P 0031 0025 g kg-1
K 0158 00877 g kg-1
Ca 717 11 g kg-1
Na 00115 00129 g kg-1
Mg 00541 00789 g kg-1
Fe 0526 0227 g kg-1
CECa 183 108 cmol+kg-1b
aCEC Cation Exchange Capacity bcmol+kg-1 centimoles of positive cations per kg of soil
Effect of temperature and soil type on M bovis survival time measured by culture-based method
Theoretical detection limit of our culture-based method was estimated at 15 times 103 CFU g-1 soil
Total cultivable M bovis (CFU g-1 soil) were enumerated by the culture-based method described above
for each sterile soil incubated at 4degC and 22degC at day 0 14 30 60 90 120 and 150 At day 0 number
of M bovis was estimated at 15 (plusmn 06) times 107 CFU g-1 for soil A and 17 (plusmn 07) times 107 CFU g-1 for soil B by
the culture-based method
Two different patterns of mycobacterial survival were observed depending on the temperature
(Figure 1a and 1b) At 22degC M bovis cells decreased over the course of the experiment in the two
sterile soils A and B (Figure 1a) No more cultivable bacteria growth was observed from day 90 in soil B
and day 120 in soil A to the end of the experiment at day 150 Although M bovis SB0120 was detected
by culture for one month longer in soil A than in soil B no statistically significant difference of CFU
numbers was observed between the two soils at 22degC (Student-Newman-Keuls Test p = 0559) thus
no significant difference in M bovis survival was observed due to soil type
At 4degC M bovis cultivable cells were recovered in high concentrations in the two soils at every
sampling date (over the five months of the experiment) The M bovis CFU numbers remained stable
during the first 30 days in the two soils as indicated in Figure 1b We observed a slight decrease of the
Chapitre 4
143
CFU numbers in the two soils starting from day 60 to the end of the study In both soils M bovis CFU
numbers decreased significantly from day 0 to day 150 (Student-Newman-Keuls Test plt0001)
reaching 87 (plusmn 25) times 10 5 CFU g- 1 and 22 (plusmn 11) times 10 5 CFU g- 1 in soil A and B respectively
Effect of soil type and endogenous microbiota on M bovis survival time measured by qPCR assay
Survival of M bovis when biotic or sterile soils A and B were incubated at 22degC was monitored
using the qPCR based assay The M bovis gene copy numbers were estimated with IS1561rsquoqPCR-based
method for each sterile and biotic soils incubated at 22degC at day 0 14 30 60 90 120 and 150 (see
Figure 2) At day 0 number of M bovis gene copies per g of soil yielded 36 (plusmn 12) times 105 and 60 (plusmn 19)
times 105 respectively for sterile soils A and B and 12 (plusmn 07) times 105 and 25 (plusmn 15) times 105 respectively for
biotic soils A and B by the qPCR-based method
Overall high amounts of M bovis gene copies were detected in the four soils at the eight sampling
dates
A statistically significant interaction between time and soil condition (biotic vs sterile) was
observed when considering the two soils independently and dependently (Student-Newman-Keuls
Test plt0001) with higher amounts of gene copies recovered in sterile soils than in biotic ones from
day 0 to day 150 No significant differences were observed when comparing the gene copy numbers in
the two sterile soils or in the two biotic soils at each sampling date
Even though the M bovis gene copy numbers slightly oscillated in the four soils over the course of the
experiment a slight but significant decrease of these numbers was observed between day 0 and day
150 for each soil The highest decreases of M bovis were observed in the biotic soils with a rough loss
of 1 log for soil B and 075 log for soil A between day 0 and day 150 (Figure 2)
Finally qPCR assay allowed a longer M bovis detection in sterile soils A and B incubated at 22degC
than culture-based assay Indeed M bovis DNA was detected by qPCR for up to 150 days in the two
sterile soil samples while bacterial culture detected viable M bovis cells up to 60 days in soil D and 90
days in soil C
Chapitre 4
144
Figure 1 Detection of M bovis SB0120 by culture-based method in sterile soils A and B after incubation at 22degC (figure above) and 4degC (figure below) Error bars represent the standard deviation values on 8 replicates
Figure 2 Detection of M bovis gene copy numbers by IS1561rsquo qPCR-based assay on sterile and biotic soils A and B after incubation at 22degC Error bars represent standard deviation values on 9 replicates
10E+00
10E+01
10E+02
10E+03
10E+04
10E+05
10E+06
10E+07
10E+08
0 14 30 60 90 120 150
M b
ovi
s C
FU n
um
ber
per
g o
f
ster
ile s
oil
Sampling day
22degC
Soil A
Soil B
10E+00
10E+01
10E+02
10E+03
10E+04
10E+05
10E+06
10E+07
10E+08
0 14 30 60 90 120 150
M b
ovi
s C
FU n
um
ber
per
g o
f st
erile
so
il
Sampling day
4degC
Soil A
Soil B
10E+00
10E+01
10E+02
10E+03
10E+04
10E+05
10E+06
0 14 30 60 90 120 150
M b
ovi
s IS
15
61
co
py
nu
mb
er p
er g
of
ster
ile s
oil
Sampling day
Sterile soil A Sterile soil B Soil A Soil B
Chapitre 4
145
DISCUSSION
Contamination of the environment with M bovis is considered as essential for the persistence
of bovine tuberculosis in animals and its interspecific indirect transmission The survival of this
mycobacteria in soil has been documented (Duffield amp Young 1985 Jackson et al 1995 Fine et al 2011
Young et al 2005) but limited information is available on the soils used and the edaphic factors that
may affect M bovis persistence are poorly understood These experiments have dealt with controlled
or environmental parameters that promote or hinder M bovis persistence in soil such as temperature
moisture or UV exposition but to our knowledge impact of the soil type has not been thoroughly
addressed This study was designed to evaluate temperature and soil type impact on M bovis survival
In this study we demonstrated that M bovis SB0120 may survive several months in soil under
different laboratory conditions in two contrasted soils and that cool temperature (4degC) strongly
promotes M bovis persistence whatever the soil type while warmer temperature (22degC) shortens its
survival time Our results are in contradiction with a previous laboratory study on M bovis persistence
(Young et al 2005) where mycobacterial survival was longer at 25degC than at 4degC in a sterile sandy soil
(pH 65-68 60 days vs 16 days) but in agreement with previous field studies where lower temperature
increased M bovis survival time up to 88 days (Fine et al 2011 Jackson et al 1995)
We also pointed out that soil type may impact on M bovis survival time Indeed despite that
no statistical difference was observed the soil A that exhibits high clay and loam content and slightly
basic pH allowed persistence of cultivable M bovis for up to three months (90 days) while this
persistence was only two months in the acidic sandy loamy soil B when soils were incubated at 22degC
On the contrary a difference of M bovis survival was observed at 4degC between the two soils with a
better survival in soil A than in soil B suggesting an impact of soil type on M bovis survival at cool
temperature However we were hampered by the detection limit of the culture-based method in the
experiment on sterile soil at 22degC where CFU numbers and detection limit ovelaped after day 60
Enrichment broth are very useful to recover low number of bacteria in complex matrices such as
Salmonella typhimurium in food or Listeria monocytogenes in soil (Locatelli et al 2013 Zheng et al
2013) but mycobacterial enrichment broths are not yet available for mycobacteria
In our study clay soil with slightly basic pH seemed to promote M bovis SB0120 survival
although mycobacterial culture detection or persistence are usually associated with sandy soils
harbouring acidic pH as previously suggested by positive culture or statistical studies (Walter et al
2014 Johnson-Ifearulundu amp Kaneene 1999 Reviriego et al 2000 Santos et al 2015) suggesting an
improved survival of the mycobacteria such as M avium ssp paratuberculosis and M bovis in these
soils However a one year persistence of cultivable M bovis was demonstrated in a sandy soil with a
basic pH (83) spiked with 108 CFU g- 1 (Ghodbane et al 2014) suggesting that M bovis may survive a
Chapitre 4
146
long time in soils harbouring a wide range of pH Moreover iron content of soils A and B seemed to be
significantly higher than soils described in previous studies
In this study qPCR was able to detect the presence of M bovis DNA over a longer period of
time than bacterial culture in soils samples incubated at 22degC whatever the soil type As one previous
study reported that DNA of dead M bovis cells was no more amplified 10 days after cell death (Young
et al 2005) we hypothesized that our qPCR assay detected live M bovis up to 150 days after
inoculation of soils Viable M bovis cells have already been detected in sterile and biotic soil over a 15
month period targeting the 16S rRNA encoding gene (Young et al 2005) Physiological state of
pathogenic mycobacteria such as M bovis in soil is unknown but dormancy or a viable but non
cultivable state (VBNC) have been suggested to explain their survival in environmental matrices
(Whittington et al 2004 Adams et al 2013) Further analysis of the expression of dormancy-related
genes (Aacutelvarez et al 2009) under soil conditions might allow to support this hypothesis Further
researches are critical to ascertain the long bacterial survival in soil and to understand the genetic
mechanisms involved in soil persistence
Our laboratory conditions were far from field conditions where temperature fluctuations
moisture variations and UV radiation are detrimental to the mycobacteria Mycobacterium bovis
survival time obtained in this study is probably longer than survival time in environmental conditions
As the gap of knowledge is important about how M bovis survive in soils laboratory studies have to
be set out in order to determine the main factors involved in M bovis survival However experiments
dealing with the exposition of M bovis naturally or artificially contaminated samples to environmental
conditions must be carried out in specific open air enclosure with high levels of protection to exclude
mycobacterial dissemination (Fine et al 2011) or require climate chambers with a L3 biosafety level
Significant differences in the recovery of M bovis DNA between biotic and sterile soils were
observed with the qPCR-based assay from the first day of the experiment indicating that qPCR
detection is hampered when soil metagenomic DNA is concentrated although no inhibition was
observed
In our experiment the use two soils gave an insight on the impact of soil type on M bovis
survival However extensive studies on larger number of well-characterised and contrasted soils would
be required to target the key edaphic factors that promote M bovis survival in soil Further work is
needed to identify those key factors and determine if the population detected with qPCR consists of
non-culturable or dormant cells capable of resuscitation under specific growth conditions or following
inhalation or ingestion by animals
Chapitre 4
147
Funding information
The work was supported by the Regional Council of Burgundy (Elodie Barbier PhD fellowship) the Cocircte
drsquoOr General Council Interbev and the Cocircte drsquoOr GDS (Groupement de Deacutefense Sanitaire Livestock
health protection group) The funders had no role in study design data collection and interpretation
or the decision to submit the work for publication
Conflict of interest
The authors have no conflict of interest to declare
Acknowledgements
The authors would like to thank Jean-Marie Duez Julien Bador and Catherine Neuwirth from the
Plateau Technique de Biologie of the Hospital of Dijon
REFERENCES
Adams AP et al 2013 Comparison of PCR versus culture for detection of Mycobacterium bovis after experimental inoculation of various matrices held under environmental conditions for extended periods Applied and Environmental Microbiology 79(20) pp6501ndash6506
Aacutelvarez AH Estrada-Chaacutevez C amp Flores-Valdez MA 2009 Molecular findings and approaches spotlighting Mycobacterium bovis persistence in cattle Veterinary Research 40(3) pp1ndash16
Barasona JA et al 2015 DNA detection reveals Mycobacterium tuberculosis complex shedding routes in its wildlife reservoir the Eurasian wild boar Transboundary and Emerging Diseases (DECEMBER)
Biet F et al 2005 Zoonotic aspects of Mycobacterium bovis and Mycobacterium avium-intracellulare complex (MAC) Veterinary Research 36 pp411ndash436
Corner LAL et al 2012 The distribution of Mycobacterium bovis infection in naturally infected badgers Veterinary journal (London England 1997) 194(2) pp166ndash72
Courtenay O et al 2006 Is Mycobacterium bovis in the environment important for the persistence of bovine tuberculosis Biology letters 2(3) pp460ndash2
Dhand NK et al 2009 Association of farm soil characteristics with ovine Johnersquos disease in Australia Preventive Veterinary Medicine 89 pp110ndash120
Duffield BJ amp Young DA 1985 Survival of Mycobacterium bovis in defined environmental conditions Veterinary Microbiology 10 pp193ndash197
Fine AE et al 2011 A study of the persistence of Mycobacterium bovis in the environment under natural weather conditions in Michigan USA Veterinary medicine international 2011 p765430
Chapitre 4
148
Ghodbane R et al 2014 Long-term survival of tuberculosis complex mycobacteria in soil Microbiology (Reading England) 160(Pt 3) pp496ndash501
Jackson R De Lisle GW amp Morris RS 1995 A study of the environmental survival of Mycobacterium bovis on a farm in New Zealand New Zealand Veterinary Journal 43(7) pp346ndash352
Johnson-Ifearulundu Y amp Kaneene JB 1999 Distribution and environmental risk factors for paratuberculosis in dairy cattle herds in Michigan American journal of veterinary research 60(5) pp589ndash96
Little TW Naylor PF amp Wilesmith JW 1982 Laboratory study of Mycobacterium bovis infection in badgers and calves The Veterinary record 111(24) pp550ndash7
Locatelli A et al 2013 Nation-wide study of the occurrence of Listeria monocytogenes in French soils using culture-based and molecular detection methods Journal of microbiological methods 93(3) pp242ndash50
Morris RS Pfeiffer DU amp Jackson R 1994 The epidemiology of Mycobacterium bovis infections Veterinary microbiology 40(1-2) pp153ndash77
Palmer M V 2013 Mycobacterium bovis Characteristics of wildlife reservoir hosts Transboundary and Emerging Diseases 60(SUPPL1) pp1ndash13
Palmer M V Waters WR amp Whipple DL 2004 Investigation of the transmission of Mycobacterium bovis from deer to cattle through indirect contact American Journal of Veterinary Research 65(11) pp1483ndash1489
Palmer M V amp Whipple DL 2006 Survival of Mycobacterium bovis on feedstuffs commonly used as supplemental feed for white-tailed deer (Odocoileus virginianus) Journal of wildlife diseases 42(4) pp853ndash8
Palmer M V Whipple DL amp Waters WR 2001 Experimental deer-to-deer transmission of Mycobacterium bovis American journal of veterinary research 62(5) pp692ndash6
Peacuterez-Lago L Navarro Y amp Garciacutea-de-Viedma D 2013 Current knowledge and pending challenges in zoonosis caused by Mycobacterium bovis A review Research in Veterinary Science 97 ppS94ndashS100
Phillips CJC et al 2003 The transmission of Mycobacterium bovis infection to cattle Research in veterinary science 74(1) pp1ndash15
Reviriego FJ Moreno M a amp Domiacutenguez L 2000 Soil type as a putative risk factor of ovine and caprine paratuberculosis seropositivity in Spain Preventive Veterinary Medicine 43 pp43ndash51
Santos N et al 2015 Widespread environmental contamination with Mycobacterium tuberculosis Complex revealed by a molecular detection protocol Plos One 10(11) pe0142079
Sweeney FP et al 2007 Environmental monitoring of Mycobacterium bovis in badger feces and badger sett soil by real-time PCR as confirmed by immunofluorescence immunocapture and cultivation Applied and environmental microbiology 73(22) pp7471ndash3
Chapitre 4
149
Sweeney FP et al 2006 Immunomagnetic recovery of Mycobacterium bovis from naturally infected environmental samples Letters in applied microbiology 43(4) pp364ndash9
Tanner M amp Michel AL 1999 Investigation of the viability of M bovis under different environmental conditions in the Kruger National Park Journal of Veterinary Research (66) pp185ndash190
Vicente J et al 2007 Risk factors associated with the prevalence of tuberculosis-like lesions in fenced wild boar and red deer in south central Spain Veterinary Research 38(3) pp451ndash464
Walter WD et al 2014 Linking bovine tuberculosis on cattle farms to white-tailed deer and environmental variables using Bayesian hierarchical analysis PloS one 9(3) pe90925
Walter WD et al 2012 On-farm mitigation of transmission of tuberculosis from white-tailed deer to cattle literature review and recommendations Veterinary medicine international 2012 p616318
Ward MP amp Perez AM 2004 Association between soil type and paratuberculosis in cattle herds American Journal of Veterinary Research 65(1) pp10ndash14
Whittington R et al 2004 Survival and dormancy of Mycobacterium avium subsp paratuberculosis in the environment Applied and environmental microbiology 70(5) pp2989ndash3004
Williams RS amp Hoy WA 1930 The viability of B tuberculosis (bovinus) on pasture land in stored faeces and in liquid manure The Journal of hygiene 30(4) pp413ndash419
Young JS Gormley E amp Wellington EMH 2005 Molecular detection of Mycobacterium bovis and Mycobacterium bovis BCG (Pasteur) in soil Applied and environmental microbiology 71(4) pp1946ndash1952
Zheng Q et al 2013 Comparison of enrichment broths for the recovery of healthy and heat-injured Salmonella typhimurium on raw duck wings Journal of food protection 76(11) pp1963ndash8
Chapitre 4
150
C BILAN
Les reacutesultats preacutesenteacutes dans ce chapitre deacutemontrent la capaciteacute de M bovis SB0120 agrave survivre
plusieurs mois dans les deux sols de Cocircte drsquoOr sous une forme viable cette dureacutee eacutetant impacteacutee par
un facteur abiotique particulier la tempeacuterature Une survie drsquoau moins cinq mois est observeacutee agrave basse
tempeacuterature (4degC) dans les deux sols avec une tregraves faible diminution de la population bacteacuterienne
initialement inoculeacutee alors qursquoelle nrsquoest que de deux agrave trois mois agrave 22degC Bien que M bovis soit cultiveacutee
un mois de plus dans le sol C par rapport au sol D nous nrsquoavons pas mis en eacutevidence drsquoeffet de la nature
du sol sur la survie de la bacteacuterie agrave 22degC Toutefois agrave cause de contraintes techniques notre analyse
nrsquoa porteacute que sur deux sols limitant fortement lrsquointerpreacutetation des reacutesultats Lrsquoanalyse des sols inoculeacutes
et incubeacutes agrave 22degC par qPCR montre un tout autre profil de survie puisque M bovis est deacutetecteacutee en
quantiteacute importante pendant les cinq mois de lrsquoeacutetude sur les deux sols et les deux conditions (biotique
vs steacuterile)
Les reacutesultats divergents obtenus par la culture et la qPCR sur les sols incubeacutes agrave 22degC amegravenent
agrave faire diffeacuterentes hypothegraveses pour expliquer lrsquoabsence de croissance des bacteacuteries puisque la qPCR
deacutetecte lrsquoADN des cellules vivantes et mortes ainsi que lrsquoADN libre Nous pouvons donc envisager que
lrsquoADN deacutetecteacute entre 60 et 150 jours appartienne agrave des cellules viables non cultivables ou agrave des cellules
mortes entiegraveres ou lyseacutees Des eacutetudes compleacutementaires sont indispensables pour preacuteciser lrsquoeacutetat
physiologique des bacteacuteries devenues incultivables apregraves une incubation dans le sol
En extrapolant les reacutesultats obtenus aux conditions de tempeacuterature rencontreacutees en Cocircte drsquoOr
nous pouvons suggeacuterer une fenecirctre temporelle pendant laquelle la survie de M bovis dans le sol est
maximale La Figure 25 repreacutesente les tempeacuteratures minimales et maximales releveacutees agrave Dijon entre
janvier 2012 et deacutecembre 2015 Nous estimons que les tempeacuteratures rencontreacutees dans lrsquoAuxois sont
probablement plus proches des tempeacuteratures minimales observeacutees dans la plaine de Dijon-Longvic ougrave
se trouve la station meacuteteacuteo (anthropisation minimale altitude humiditeacute veacutegeacutetation) que des
maximales Ainsi la peacuteriode la plus agrave risque va de novembre agrave avril (tempeacuteratures minimales de
lrsquoair lt 5degC) et correspond agrave la fin de lrsquoautomne lrsquohiver et le deacutebut du printemps Durant le reste de
lrsquoanneacutee et en particulier en eacuteteacute (tempeacuteratures maximales de lrsquoair gt 22degC) les risques de survie de M
bovis semblent moins importants Ces conclusions restent des hypothegraveses dans la mesure ougrave la
tempeacuterature reacuteelle des sols nrsquoa pas eacuteteacute mesureacutee et la variation des autres facteurs climatiques nrsquoa pas
eacuteteacute prise en compte Cependant on peut penser qursquoen eacuteteacute (sous forte insolation) la tempeacuterature des
sols peut augmenter rapidement dans les quelques premiers centimegravetres de sol limitant la dureacutee de
survie de la bacteacuterie
Chapitre 4
151
Figure 25 Tempeacuteratures minimales et maximales observeacutees agrave Dijon-Longvic de 2012 agrave 2015 (Source Meacuteteacuteo France)
-5
0
5
10
15
20
25
30
35
1 2 3 4 5 6 7 8 9 10 11 12
Tmin2012
Tmin2013
Tmin2014
Tmin2015
Tmax2012
Tmax2013
Tmax2014
Tmax2015
4degC
22degC
CHAPITRE 5
Rocircle de la faune du sol dans la circulation de
Mycobacterium bovis eacutetude de la disseacutemination de
M bovis par les lombrics
E Barbier B Chantemesse M Rochelet L Fayolle L Bollache M L Boschiroli A Hartmann
Rapid dissemination of Mycobacterium bovis from cattle dung to soil by the earthworm
Lumbricus terrestris
(Article accepteacute en cours de publication Veterinary Microbiology)
155
CHAPITRE 5 - Rocircle de la faune du sol dans la circulation de
Mycobacterium bovis eacutetude de sa disseacutemination par les lombrics
A AVANT-PROPOS
Le rocircle de la faune et de la microfaune du sol dans la survie et la disseacutemination de
Mycobacterium bovis dans lrsquoenvironnement est inconnu aujourdrsquohui Comme nous lrsquoavons eacutevoqueacute dans
la synthegravese bibliographique les protozoaires plus particuliegraverement les amibes jouent un rocircle
vraisemblable dans la survie des mycobacteacuteries environnementales (Adekambi et al 2006 Mba Medie
et al 2011 Mura amp Bull 2006) voire dans la virulence des mycobacteacuteries opportunistes (Cirillo et al
1997) In vitro la survie intra-amibienne de M bovis pendant plusieurs mois a eacuteteacute deacutemontreacutee mais
aucune eacutetude de terrain eacutevaluant le rocircle potentiel des amibes comme reacuteservoir de M bovis nrsquoest
disponible Concernant la meacutesofaune les lombrics ont eacuteteacute identifieacutes comme vecteur potentiel dans la
transmission de M avium ssp paratuberculosis dans des eacutelevages bovins et caprins infecteacutes (Fischer
et al 2003) Veacuteritables ingeacutenieurs du sol ces animaux ont un rocircle important dans la circulation des
microorganismes en les transfeacuterant et les seacutelectionnant depuis la matiegravere organique qursquoils ingegraverent et
dont ils sont de gros consommateurs vers le sol qui les entoure par le biais de leurs turricules
Afin drsquoeacutevaluer le rocircle potentiel des lombrics dans la disseacutemination de M bovis dans le sol suite
agrave lrsquoingestion de matiegraveres feacutecales animales contamineacutees nous avons eacutetudieacute le transfert de M bovis BCG
depuis les fegraveces inoculeacutes vers le sol environnant par une espegravece de lombric aneacutecique Lumbricus
terrestris La population bacteacuterienne de M bovis a eacuteteacute deacutetecteacutee et quantifieacutee par qPCR dans le sol
environnant les turricules et le tube digestif des lombrics
Chapitre 5
157
B RAPID DISSEMINATION OF M BOVIS FROM CATTLE DUNG TO SOIL BY THE EARTHWORM LUMBRICUS TERRESTRIS
Elodie Barbierab Benoit Chantemesseab Murielle Rocheletab Leacuteon Fayolle a Loiumlc Bollachebc
Maria Laura Boschiroli d Alain Hartmann a
a INRA UMR 1347 Agroeacutecologie Dijon Cedex France
b Universiteacute de Bourgogne Franche Comteacute Dijon Cedex France
c Laboratoire Chrono-environnement UMR CNRS 6249 Besanccedilon France
d Universiteacute Paris-Est Laboratoire National de Reacutefeacuterence de la Tuberculose Uniteacute de Zoonoses Bacteacuteriennes Laboratoire de
Santeacute Animale ANSES Maisons-Alfort Cedex France
Key words Mycobacterium bovis spreading earthworms Lumbricus terrestris cattle soil quantitative
PCR
Running title Environmental dissemination of Mycobacterium bovis by earthworms
Corresponding Author Alain Hartmann UMR 1347 Agroeacutecologie INRA 17 rue Sully BP 86510 21065
Dijon Cedex France E-mail alainhartmanndijoninrafr
Chapitre 5
158
Abstract
Indirect transmission of the bovine tuberculosis (bTB) between wildlife and livestock is thought to occur
by inhalation or ingestion of environmental substrates contaminated through animal shedding The
role of the soil fauna such as earthworms in the circulation of Mycobacterium bovis from
contaminated animal feces is of interest in the epidemiology of bTB The objective of this study was to
assess the impact of earthworm activity on M bovis transfer from animal dung to castings and the
surrounding soil For this purpose microcosms of soil containing the anecic eathworms Lumbricus
terrestris were prepared and covered with cattle feces spiked with the M bovis BCG strain Pasteur to
carry out two separate experiments The dissemination the gut carriage and the excretion of M bovis
were all monitored using a specific qPCR-based assay Our results showed that the earthworm L
terrestris was able to rapidly disseminate M bovis from the contaminated cattle feces to the
surrounding soil through casting egestion Moreover contaminated earthworms were shown to shed
the bacteria for 4 days when transferred in a bTB-free soil This study highlights for the first time the
possible role of earthworms in the dissemination and the persistence of M bovis in soils in bTB endemic
areas
Chapitre 5
159
INTRODUCTION
Mycobacterium bovis is a pathogenic mycobacteria that can infect and cause bovine
tuberculosis (bTB) in cattle and in a wide range of domestic or wild mammals as well as in humans
(Biet et al 2005) bTB is a primarily respiratory disease but it may affect various organs in animals
(Morris et al 1994) Although cattle are considered as the principal host of M bovis wild fauna such
as badgers (Meles meles) wild boar (Sus scrofa) and red deer (Cervus elaphus) has also been identified
as potential wildlife reservoirs (Palmer 2013) Transmission of bTB between cattle mainly occurs by
inhalation of contaminated aerosols during close contacts with infected animals (Morris et al 1994
Phillips et al 2003) An inoculum of less than ten bacilli is enough to cause respiratory disease in cattle
(Dean et al 2005) Moreover indirect transmission by inhalation of environmental bioaerosols or
ingestion of infected soil and water could also be a potential way of animal contamination (Palmer et
al 2004 Vicente et al 2007)
Infected animals were shown to shed M bovis via oro-nasal mucus sputum urine feces and
wound discharges (Corner et al 2012 Palmer et al 2004) that may contaminate environment
especially pastures shared by cattle and wild animals Thus investigation of soil as a potential reservoir
of M bovis is an important step towards understanding the epidemiology of the bacteria To reach this
goal deciphering the ecological factors that govern the survival and the spread of M bovis is first
required
Among soil fauna earthworms constitute the largest animal biomass in most terrestrial
ecosystems especially in grasslands (Lavelle and Spain 2001) Earthworms play a role of engineers in
soil processes impacting on soil functions and ecosystem services (Blouin et al 2013) affecting soil
structure water movement organic matter decomposition nutrients releasing plant growth and
pollution remediation Earthworms are usually classified in three ecological types according to their
feeding habits i) the epigeic ones are restricted to the surface organic layers where they feed
themselves ii) the endogeic ones live in the subsurface where they burrow lateral tunnels to ingest soil
and organic matter and iii) the anecic ones feed preferentially on surface litter but live in burrows in
the soil (Lee 1985) Earthworms ingest soil particles mixed with organic matter that are mechanically
and chemically digested by both gut fluid and microbiota (Curry and Schmidt 2007) and release
castings (undigested soil egested on or under the soil surface) Earthworm gut microbiota acts as a
selective filter for soil microorganisms shapes microbial communities of soil and either up- or down-
regulates microbial populations (Byzov et al 2007 Egert et al 2004 Goacutemez-Brandoacuten et al 2011) It
has been shown that earthworms may have microbial beneficial activities especially in the transport of
bacterial inocula through soil (Thorpe et al 1996) but earthworms may also act as pests by promoting
the dissemination of plant pathogens such as Phytophtora cactorum and Fusarium reticulatum
Chapitre 5
160
(Montecchio et al 2015) More interestingly a few studies have described that human and animal
pathogenic bacteria viruses and parasites especially Escherichia coli O157H7 foot-and-mouth disease
virus and Cestoda tapeworms may be carried and spread by earthworms (Edwards and Bohlen 1996
Prysor Williams et al 2006)
Although animal dung is a very attractive food for earthworms (Lowe and Butt 2005) few
studies have evaluated the role of earthworms in the survival and the dissemination of potential
pathogenic mycobacteria from animal dung or manure to surrounding soil (Fischer et al 2003) In
addition since earthworms represent an important part of the diet of various animal species including
badgers and wild boar (Granval and Muys 1995 Lee 1985) the consumption of contaminated
earthworms (gut carriage) might be a source of contamination for wild fauna
With the aim to investigate the above hypothesis this current study was first devoted to assess
the impact of earthworm activity on M bovis transfer from animal dung to their released castings and
the surrounding soil Then the M bovis gut carriage and the casting contamination following ingestion
of infected feces were investigated
MATERIAL AND METHODS
Mycobacterium bovis strain preparation
Mycobacterium bovis BCG strain Pasteur 1173P2 was grown aerobically at 37degC in Middlebrook
7H9 broth supplemented with 10 Middlebrook ADC Growth Supplement (Sigma-Aldrich France) The
M bovis inoculum was prepared as following a 10 mL stationary phase culture was pelleted and once
the supernatant removed the cell aggregates were disrupted by vortexing with glass beads for 15
seconds The pellet was suspended in 1 mL of sterile water and serial 10-fold dilutions of the suspension
were spiked on modified Middlebrook 7H11 plates (Middlebrook 7H11 supplemented with 10 heat
inactivated bovine serum (Dominique Dutscher France) and 10 Middlebrook OADC Growth
Supplement (Sigma-Aldrich France) for enumeration The plates were always incubated at 37degC for 2
months A 10-fold dilution of the prepared M bovis suspension was also directly quantified with the
RD4 qPCR assay described in the section ldquoMycobacterium bovis quantitative PCR (qPCR)rdquo
Soil cattle feces and earthworms characteristics
A bTB-free pasture (that has never been grazed by bTB infected cattle) located in Burgundy
(Middle East of France) was selected to sample the soil used in this study This latter is a clayey loamy
soil with a pH of 775 Organic carbon and nitrogen contents were 361 and 38 gkg-1 respectively The
soil was sieved to 4 mm and stored at 4degC until its use
Fresh cattle feces were collected from a bTB-free Charolais cattle in a bTB-free area of Burgundy and
Chapitre 5
161
immediately used
The adult anecic earthworms (Lumbricus terrestris) were provided by LombrirsquoCarraz (France)
and kept at 4degC in large buckets containing soil and organic matter until starting the study Soil
earthworms and cattle feces used in this work were previously proved to be negative for the presence
of M bovis using the qPCR-based assay
DNA extraction and purification
DNAs from all the samples collected were extracted as described previously (Dequiedt et al
2011) Briefly 8 mL of lysis buffer (100 mM Tris pH 80 100 mM EDTA pH 80 100 mM NaCl and 2
(wv) SDS) 4 g of silica beads (100 μm diameter) 5 g of ceramic beads (14 mm diameter) and 8 glass
beads (4 mm diameter) were added to 2 g of sample in a 15 ml sterile vial - whereas 1 mL of lysis
buffer 05 g of silica beads 0625 g of ceramic beads 1 glass beads were added to 250 mg-samples
Samples were disrupted for 3 times 30 s at 4 ms in a FastPrepreg-24 Instrument (MP Biomedicals Europe
Illkirch France) incubated for 30 min at 70 degC and centrifuged for 5 min at 7000 timesg at room
temperature Supernatants were incubated on ice for 10 min with 110 volume of 3 M potassium
acetate pH 55 and then centrifuged for 5 min at 14000 times g One volume of ice-cold isopropanol was
added to the supernatant for DNA precipitation overnight at minus 20 degC DNA was collected by
centrifugation (30 min at 14000 times g) and DNA pellets were washed with ice-cold ethanol (70) and
dissolved in 100 μl of water Crude DNA extracts (100 μl) were loaded on Microbiospintrade columns
(Biorad Marnes-La-Coquette France) filled with 100 mg of polyvinyl polypyrrolidone (PVPP Sigma
Chemical Co France) After centrifugation (4 min 1000 times g 4 degC) eluted DNA was further purified
using the Geneclean Turbo Kit (Qiagen Courtaboeuf France) according to the manufacturers
instructions Purified DNA concentrations were estimated by spectrophotometry (λ = 260 nm) using a
NanoDropreg 2000 spectrophotometer (Thermo Scientific Illkirch France)
Mycobacterium bovis quantitative PCR (qPCR)
Mycobacterium bovis-specific qPCR was set up targeting the RD4 flanking region using
previously described forward RD4 flanking primer (5rsquo-TGTGAATTCATACAAGCCGTAGTCG-3rsquo) and probe
(5rsquo-6-FAM-AGCGCAACACTCTTGGAGTGGCCTAC-3rsquo TAMRA) (Sweeney et al 2007) In order to improve
qPCR efficiency the reverse RD4 flanking primer based assay was modified (5rsquo-
ATCTTAGCTGGTCAATAGCCATTTTT-3rsquo) to reduce the size of the qPCR product to 116 bp
qPCR reactions were carried out in triplicate with a 25 microL reaction mix containing 125 microL of
ABsolutetrade QPCR ROX Mix (Thermo Scientific France) 1 microL (final concentration 25 ngmicroL-1) of T4
bacteriophage gene 32 product (MP Biomedicals Europe France) 1 microL of each primer (final
concentration 400 nmolL-1) 05 microL of each probe (final concentration 200 nmol L-1) 4 microL of ultrapure
Chapitre 5
162
water and 5 microL of pure or diluted (10- or 20-fold) DNA Absolute quantification was achieved using
standard DNA dilutions A recombinant plasmid containing one copy of the RD4 fragment of M bovis
BCG strain Pasteur 1173P2 cloned in the pCRregII-TOPOreg vector (Invitrogen France) was used as a
standard A calibration curve was obtained by amplification of serial dilutions of the plasmid ranging
from 1 to 106 copies per PCR reaction All DNA samples extracted from the soil castings and feces
samples were screened with the RD4-based qPCR assay and results were expressed as the number of
M bovis gene copies per g of soil castings or feces We previously assessed that when Cycle threshold
(Ct) was above 39 detection was possible but unreliable (unpublished results) The word ldquotracesrdquo was
then chosen in all further experiments in order to indicate that M bovis targets are present in very low
quantities (below 250 CFUg-1 of soil) The qPCR assay was performed in a VIIA 7 Real-Time PCR System
(Thermo Fischer Scientific France) Initial denaturation was done at 95degC for 15 min followed by 45
cycles with denaturation at 95degC for 15 s annealing and elongation at 60degC for 1 min
Mesocosms preparation
Plastic jars (15 L) were filled with 1 kg of the clay-loamy soil Sterile water was added to each
soil sample to reach final soil moisture of 80 of the water-holding capacity (WHC) Once the soil was
packed down to obtain a 1 gcm-3 soil density 6 earthworms Lumbricus terrestris were placed into each
jar A 4-days delay was necessary to acclimatize the earthworms Nine control mesocosms where no
earthworms were added were processed and analysed
Evaluation of earthworm dissemination of M bovis to soil mesocosms
Aliquots of 50 g of cattle feces previously spiked with 1 mL of M bovis BCG suspension were
layered on the soil surface of each mesocosm Jars were covered with pierced fitted lids and kept in a
dark room at 20degC in a L2 confinement facility (day 0)
In order to sample fresh earthworm castings all the castings were removed 48 h before each
sampling date A sampling schedule ie 2 4 7 10 14 21 30 45 and 60 days was followed For each
sampling date 2 mesocosms with earthworms and one control mesocosm were analysed as following
First earthworm castings were collected at the surface of each mesocosm containing earthworms
pooled homogenized and then aliquoted in triplicate in 2 g-samples Secondly the cattle feces and
earthworms were removed out of the mesocosms and the remaining soil was mixed and aliquoted in
triplicate in 2 g-samples Finally the six earthworms contained in each mesocosm were rinsed twice
with sterile water dried with a paper towel and individually transferred into a 60 mL empty vessel
After a 48h incubation at 20degC in a dark room to release the gut content one 250 mg-aliquot of fecal
samples was sampled for each earthworm In the control mesocosm the cattle feces were removed
from the top of mesocosm the soil was then mixed sampled and analysed as described above All
Chapitre 5
163
samples were frozen at -20degC until their DNA extraction and RD4 qPCR analysis Gene copy numbers
found in each sample were box-cox transformed before analysis to normalize the data Generalized
Linear Models (GLMs) were used to analyze the effects of sampling date (number of days after the
M bovis spiked cattle feces was layered onto the soil) and treatment (feces castings and soil) on the
number of CFU Tests were performed using JMP 70 Software (SAS Institute Inc) Results were
considered to be significantly different for P values lt 005
Gut carriage and excretion of M bovis by earthworms
Five mesocosms each containing 6 earthworms were prepared as described in the section
ldquomesocosm preparationrdquo and were then covered with 50 g of cattle feces spiked with 1 mL of M bovis
BCG suspension After 7 days of incubation each earthworm was rinsed twice with sterile water dried
with a paper towel To estimate M bovis carriage on earthworm tegument 6 earthworms out of the
30 were deposited in an individual Petri plate containing modified Middlebrook 7H11 media
supplemented with antibiotics and antifungal (100 mgL-1Ticarcillin 10 mgL-1 Trimethoprim 200 000
UIL-1 Polymyxin B and 100 mgL-1 Cycloheximid Sigma-Aldrich France) After gently moving it on the
plate surface for few minutes the earthworms were individually transferred in a 200 mL vessel
containing 100 g of M bovis-free fresh soil as well as the 24 earthworms This step corresponded to
the starting point of the experiment (day 0)
At each sampling date ie day 1 2 3 4 and 5 the castings from series of 6 randomly selected
remaining vessels were collected pooled homogenized and one aliquot of 2 g was sampled Then the
earthworms from 1 series of 6 vessels were removed rinsed twice with sterile water dried with a paper
towel and individually transferred in a 60 mL empty vessel After 48h at 20degC in a dark room the
animals have released their gut contents and a 250 mg-aliquot of fecal sample was recovered from
each vessel containing earthworms All of the samples were stored at -20degC until further analysis
RESULTS
Earthworm dissemination of M bovis from cattle feces spiked with the mycobacteria to soil
mesocosms
For this purpose the protocol sketched in Figure 1 was carried out over a 60 days-period and
the results are summarized in Figure 2 Both the culture-based method and the RD4 qPCR assay were
used to estimate the concentration of the Mycobacterium bovis BCG suspension used to spike the cattle
feces Different values of 125 times 107 CFU per mL and of (65 plusmn 05) times 108 gene copies per mL were
determined with the culture and the molecular methods respectively However given that only the
RD4 qPCR assay was carried out to quantify M bovis throughout this experiment the M bovis genome
Chapitre 5
164
equivalent number contained in the 50 g of cattle feces was estimated to (65 plusmn 05) times 108 at the
beginning of the experiment (day 0 dashed line in Figure 2)
Figure 1 Schematic representation of the procedure followed for the evaluation of M bovis dissemination in
soil by earthworms
Overall M bovis was repeatedly detected in all of the analysed samples ie earthworm castings
fecal samples and soils from 2 days after depositing the cattle feces spiked with M bovis on the surface
of the mesocosms Gene copy number was significantly influenced by the sampling date (df= 8 sup2 =
14508 Plt00001) treatment (df= 2 sup2 = 22430 Plt00001) and interaction between sampling date
and treatment (df= 16 sup2 = 14464 Plt00001) However at the end of the experiment (day 60)
M bovis gene copy numbers were not statistically different (Average and P value) whatever the kind of
samples More precisely the earthworm fecal samples (open circle Figure 2) showed the highest
contamination (up to (57 plusmn 03) times 106 M bovis gene copies per g) at day 2 while the M bovis gene copy
numbers in the fecal samples decreased (20-fold factor) after day 4 to reach a final value of (25 plusmn 22)
times 105 gene copy number per g On the other hand the M bovis gene copy numbers per g of earthworm
castings were up to around 10-fold lower than those per g of earthworm fecal samples until day 45
after which similar M bovis gene concentrations were measured (full circle Figure 2) Although soil
samples from mesocosms also led to a M bovis positive signal from day 2 (44 plusmn 42) times 104 gene copy
number per g) it is worth noting that only 4 out of the 6 analyzed samples were positive at day 2 (full
square Figure 2) Afterwards (ie from day 4) all of the soil samples gave a positive signal for M bovis
with a strong increase (10-fold) of the M bovis gene copy number per g of soil between day 2 and day
(D0 + 2 4 7 10 14 21 30 45)
Mesocosm Deposition of M bovis spiked feces
D0
CASTING POOL
FECAL SAMPLE
48h00
Earthworm
Fresh soil
M bovis spiked cattle feces
SOIL
Sampling day
Chapitre 5
165
7 followed by minor fluctuations until day 60 For the control mesocosms (ie without any
earthworms) no M bovis signal were measured in the soil samples throughout the experiment until
day 60 where a positive signal value of (35 plusmn 19) times 104 M bovis per g of soil was obtained (cross symbol
on Figure 2) thus indicating a very slow spread of M bovis in the bulk soil in the absence of earthworms
Finally it should be noticed that the cattle feces remained undisturbed in the control mesocosms over
the course of the experiment whereas the earthworms totally ingested the spiked cattle feces in all of
the mesocosms containing these animals during the first week of the experiment
Figure 2 Concentration of M bovis DNA determined by RD4 qPCR in soil earthworm castings and fecal samples recovered at each sampling date Concentrations of M bovis DNA in control mesocosms are represented with crosses (times) The dashed line corresponds to the concentration of M bovis spiked in 50 g of cattle feces Error bars represent the standard deviation values of 3 replicates
3
4
5
6
7
8
0 5 10 15 20 25 30 35 40 45 50 55 60
Lo
g1
0(M
b
ovi
sg
ene
cop
y n
um
ber
) in
1 g
of
sam
ple
Days
Chapitre 5
166
Table 1 RD4-based qPCR detection of M bovis gene copy number contained in the earthworm castings recovered according to the protocol of Figure 2
Sampling day Number of analysed
casting poolsa Number of positive
casting poolsb M bovis gene copy number per gc
1 5 5 (196 plusmn 136) times 105
2 4 4 (104 plusmn 060) times 105
3 3 2 Tracesd
4 2 1 Traces
5 1 0 Negative a
From 6 vessels b
Threshold cycle (Ct) between 20 and 39 c Mean value calculated with the signal obtained for the positive earthworm casting pools
d Detected but not quantifiable (below 250 CFUg-1 of soil)
Evaluation of the carriage and the excretion of M bovis by earthworms
In order to check whether earthworms carry M bovis in their gut and are able to excrete the
bacteria the following experiment was realized (see detailed protocol in Figure 3) Earthworms were
artificially contaminated by feeding for 7 days from cattle feces spiked with M bovis then their castings
and their fecal samples were analysed for a 5-days period First no growth of M bovis was observed
from earthworm tegument surfaces suggesting the absence or the low carriage of M bovis by
earthworm teguments The results shown in Table 1 indicated that all of the earthworms produced
castings highly contaminated with M bovis 1 and 2 days after their introduction in the fresh M bovis
free soil (up to around 105 M bovis gene copy number per g) Then the percentage of positive M bovis
casting pools gradually decreased (66 and 50 for sampling day 3 and 4 respectively) until the
sampling at day 5 for which no M bovis molecular signal was detected Although positive signals were
obtained for the sampling days 3 and 4 only traces of M bovis were detected (Ct above 39) With
values ranging from 1 to 5 105 M bovis gene copy number per g of fecal sample (Table 2) the RD4
qPCR assay yielded M bovis positive signals for the sampling days 1 to 4 Nevertheless for each of
these sampling days some earthworms (17-50 ) led to negative M bovis qPCR signal thus indicating
the absence or a very low amount of M bovis in the earthworm feces (below the detection limit of the
qPCR) Finally all of the six earthworm fecal samples analysed at day 5 were M bovis free
Chapitre 5
167
Figure 3 Schematic representation of the procedure followed for the estimation of M bovis gut carriage and excretion by earthworms
Table 2 RD4-based qPCR detection of M bovis gene copy number contained in the earthworm fecal samples recovered according to the protocol of Figure 3
Sampling day Number of positive
fecal samplesa (n = 6) M bovis gene copy number per gb
1 5 (123 plusmn 082) times 105
2 4 (228 plusmn 214) times 105
3 3 (542 plusmn 049) times 105
4 4 (213 plusmn 164) times 105
5 0 Negative
a Threshold cycle (Ct) between 20 and 39 b Mean value calculated with the signal obtained for the positive earthworm fecal samples
Sampling day (D0 + 1 2 3 4 5)Mesocosm
(n = 5)
Deposition of M bovisspiked feces (n = 5 )
Investigation of M bovis tegumentcarriage on 6 earthworms
D0
7 days
Transfer of each earthworm
5 series of 6 vessels(n = 30)
FECAL SAMPLE
CASTING POOL
n = 6
48h00
n = number of remainingseries of 6 vessels
Earthworm
Fresh soil
M bovis spiked cattle feces
Chapitre 5
168
DISCUSSION
The results shown in Figure 1 evidenced for the first time that earthworms were able to widely
transfer M bovis from cattle feces to their castings after ingestion of cattle feces spiked with the
bacteria Owing to the concomitant ingestion of M bovis-free soil and dung remains and thus the
dilution of M bovis cells in the gut content the number of M bovis cells in castings gradually decreased
from day 4 until the end of the experiment These results were in good agreement with earlier reported
laboratory trials with Mycobacterium avium subsp paratuberculosis that is responsible for the Johnersquos
disease in cattle and goat (Fischer et al 2003) Prysor Williams et al (Prysor Williams et al 2006) have
also reported the excretion of the human food-borne pathogen Escherichia coli O157H7 in castings
after ingestion of spiked manure by earthworms
On the other hand this study demonstrated that when earthworms were maintained in the
M bovis infected environment they were able to shed M bovis in their castings for at least 60 days
(Tableau 1) However when transferred in a M bovis-free soil earthworms only shed the bacteria up
to 4 days This data suggests that M bovis is unable to colonize the gut of the earthworm L terrestris
over extended periods Moreover L terrestris may ingest 490 to 3500 mg dry mass g-1 fresh mass of
arable soil per day (Curry and Schmidt 2007) and has a mean gut transit time of about 12 hours (Taylor
and Taylor 2014) leading to the rapid egestion of the bacteria from earthworm guts through castings
Furthermore earthworms were able to transfer M bovis from cattle feces to the bulk soil by
releasing castings in the tunnels they burrow Our results showed that M bovis was still detected in
mixed soil over the course of the experiment (60 days) highlighting a long persistence of M bovis in
soil Rapid dispersal of the bacteria in soil can be fully imputed to earthworm activity since soil from
control mesocosms without earthworms remained free from M bovis-except at day 60 Moreover
bacteria movement was mainly due to M bovis earthworm excretion since no M bovis tegument
carriage was observed The M bovis signal observed in control mesocosms at day 60 is probably due
to the moving of M bovis cells during moisture transfer from dung to soil surface and to the presence
of remaining dung particles in soil (incomplete removal of contaminated dung before soil mixing) As
M bovis is sensitive to solar radiation (UV) and soil dryness the burying of M bovis in deep soil by
earthworms may contribute to enhance its survival in soil environment Indeed survival of M bovis in
soil may be enhanced up to 2 years when the bacteria is buried to a depth of 5 cm (Genov 1965) Our
experimental design based on 1 kg-soil mesocosms did not allow estimating the real spatial distribution
of the bacilli in soil by earthworms It has been shown that earthworms are potentially able to move
several meters apart from the area where they graze organic matter (Mather and Christensen 1988)
thus disseminating soil-borne plan pathogen through large surfaces of soil (Thorpe et al 1996)
Concerning potential spread of animal or human pathogens earthworms were demonstrated in
Chapitre 5
169
experimental boxes to significantly aid vertical and lateral movement of ingested bacteria both up to
40 cm from the inoculation point (Prysor Williams et al 2006)
Finally we showed that earthworms may carry M bovis for many days in their guts even when
transferred in a M bovis-free environment Earthworms are known to host animal parasites and
pathogen such as protozoan tapeworms (Cestoda) and nematodes they can transmit to birds and
mammals when earthworms are ingested (Edwards and Bohlen 1996) However to our knowledge
the fact that earthworms may act as a source of contamination for animals feeding from earthworms
has never been studied Earthworms that constitute a nutritious food resource are an important part
of the diet of badgers and wild boar (Baubet et al 2003 Cleary et al 2011) that often forage in cattle
pastures hosting high earthworm populations (Muldowney et al 2003) Badgers and wild boar are
able to eat up to 200 and 120 earthworms respectively in a night especially during worm nights when
humid and mild weather conditions are optimal for earthworms to ascend to the surface (Baubet et
al 2003 Kruuk and Parish 1981) In endemic area of bTB where fecal M bovis shedding may occur in
infected animals it would be interesting to evaluate M bovis carriage in earthworm gut in the vicinity
of infected animal dung Further studies are needed to assess the role of earthworms harbouring
M bovis gut carriage in the contamination of bTB sensible earthworm feeders
One field study reports the isolation of non-tuberculous mycobacteria such as Mycobacterium
avium complex members (M avium ssp paratuberculosis and M avium ssp hominisuis) as well as M
abcessus M scrofulaceum and M gastri) from 9 out of 109 earthworm feces in sites where cattle
goats and swine were severely infected with the Johnersquos disease or mycobacteriosis (Fischer et al
2003) However this study failed to isolate M avium ssp avium from earthworms feces in aviaries
infected with avian tuberculosis
Cattle feces were spiked with M bovis at a final concentration of 125 times 107 CFU per 50 g of
feces ie 25 times 105 CFU per g (estimated by culture-based enumeration of the inoculum) M bovis
concentration in spiked cattle feces was also measured by the qPCR-based method We observed
numbers of M bovis 52-fold higher when using the molecular detection method than with the culture-
based method This discrepancy is probably due first to the strong aggregation of mycobacteria cells in
the inoculum despite physical disruption of cell aggregates and second to the presence of dead or
viable but non-cultivable cells of M bovis non detected by culture-based method The amount of
M bovis spiked in cattle feces is equivalent or lower than the number of M bovis cells recorded per g
of urine or feces shed by infected badgers (Gallagher and Clifton-Hadley 2000 Sweeney et al 2007)
leading to the presence of millions of bacteria in a latrine (burrowed hole in the soil where badgers
defecate and urinate) But it is probably larger than M bovis cells deposited in infected cattle feces
(Neill et al 1988) For practical reasons we used cattle feces in our study but it could be interesting to
compare if dung from other animals than cattle would be as attractive for earthworms
Chapitre 5
170
Our study was based on a qPCR assay method detecting M bovis DNA in soil castings and feces
samples This molecular tool does not give any information on the viability of the M bovis cells egested
by the earthworms However as mycobacteria harbour a resistant waxy cell envelope (Brennan and
Nikaido 1995) and as Mycobacterium avium ssp paratuberculosis and other environmental
mycobacteria have been showed to largely survive through the gut passage (Fischer et al 2003) these
data strongly suggest that M bovis could resist through the passage of earthworm guts and remain
alive in castings Further research is needed to clarify if tuberculous mycobacteria such as M bovis are
digested or survive during transit through the earthworm gut and if their virulence is conserved
Evidencing M bovis presence with culture-based method in environmental matrices such as soil
castings or animal feces is still a challenge As M bovis is a slow growing mycobacteria environmental
samples have to be strongly decontaminated prior to culturing to overcome contamination with fast
growing microorganisms leading to a decrease in the sensitivity of the method
Surface and subsurface casting shedding of M bovis is all the more relevant because this
pathogen bacteria may survive in soil and feces for long periods (Young et al 2005) Castings are then
within the reach of animals especially cattle that may be infected by inhaling soil aerosols or by
ingesting contaminated casting soil during their exploratory foraging or feeding activities While
Mycobacterium bovis infective dose by the respiratory route is very low since only ten viable bacilli are
able to cause respiratory bovine tuberculosis (Dean et al 2005) 5000 bacilli would be required to
initiate infection through the oral route in cattle (Palmer et al 2004) As cattle usually avoid grazing
close to the feces of other cattle or wild animals (Hutchings and Harris 1997) further dissemination of
M bovis by earthworms away from infected feces might induce contamination risk for cattle grazing in
this area
Further work is still needed to study the occurrence of M bovis in earthworms under natural
field conditions in bTB-contaminated area Cultivation of environmental M bovis is also required to
characterize properly these environmental strains Finally virulence of M bovis contained in
earthworm castings remains to be deciphered
Funding information
The work was supported by the Regional Council of Burgundy (Elodie Barbier PhD fellowship) the Cocircte
drsquoOr General Council Interbev and the Cocircte drsquoOr GDS (Groupement de Deacutefense Sanitaire Livestock
health protection group) The funders had no role in study design data collection and interpretation
or the decision to submit the work for publication
Conflict of interest
The authors have no conflict of interest to declare
Chapitre 5
171
Acknowledgements
The authors would like to thank AE Taconet F Bartheacuteleacutemy and T Grosjean from the Cocircte drsquoOr Regional
Veterinary Service (DDPP21) F Chevalier and M Beacuteral from the CIREV (Cellule Inter-Reacutegionale
drsquoEpideacutemiologie Veacuteteacuterinaire)
References
Baubet E Ropert-Coudert Y Brandt S 2003 Seasonal and annual variations in earthworm consumption by wild boar (Sus scrofa scrofa L) Wildl Res 30 179 doi101071WR00113
Biet F Boschiroli ML Thorel MF Guilloteau LA 2005 Zoonotic aspects of Mycobacterium bovis and Mycobacterium avium-intracellulare complex (MAC) Vet Res doi101051vetres
Blouin M Hodson ME Delgado E a Baker G Brussaard L Butt KR Dai J Dendooven L Peres G Tondoh JE Cluzeau D Brun JJ 2013 A review of earthworm impact on soil function and ecosystem services Eur J Soil Sci 64 161ndash182 doi101111ejss12025
Brennan PJ Nikaido H 1995 The envelope of mycobacteria Annu Rev Biochem 64 29ndash63 doi101146annurevbiochem64129
Byzov BA Khomyakov N V Kharin SA Kurakov A V 2007 Fate of soil bacteria and fungi in the gut of earthworms Eur J Soil Biol 43 149ndash156 doi101016jejsobi200708012
Cleary GP Corner LAL OrsquoKeeffe J Marples NM 2011 Diet of the European badger (Meles meles) in the Republic of Ireland A comparison of results from an analysis of stomach contents and rectal faeces Mamm Biol 76 470ndash475 doi101016jmambio201010012
Corner LAL OrsquoMeara D Costello E Lesellier S Gormley E 2012 The distribution of Mycobacterium bovis infection in naturally infected badgers Vet J 194 166ndash72 doi101016jtvjl201203013
Curry JP Schmidt O 2007 The feeding ecology of earthworms - A review Pedobiologia (Jena) 50 463ndash477 doi101016jpedobi200609001
Dean GS Rhodes SG Coad M Adam O Cockle PJ Clifford DJ Hewinson RG Vordermeier HM 2005 Minimum infective dose of Mycobacterium bovis in cattle Infect Immun 73 6467ndash6471 doi101128IAI73106467
Edwards CA Bohlen PJ 1996 Biology and ecology of earthworms Chapman and Hall London
Egert M Marhan S Wagner B Scheu S Friedrich MW 2004 Molecular profiling of 16S rRNA genes reveals diet-related differences of microbial communities in soil gut and casts of Lumbricus terrestris L (Oligochaeta Lumbricidae) FEMS Microbiol Ecol 48 187ndash197 doi101016jfemsec200401007
Fischer OA Matlova L Bartl J Dvorska L Svastova P Du Maine R Melicharek I Bartos M Pavlik I 2003 Earthworms (Oligochaeta Lumbricidae) and mycobacteria Vet Microbiol 91 325ndash338 doi101016S0378-1135(02)00302-4
Chapitre 5
172
Gallagher J Clifton-Hadley RS 2000 Tuberculosis in badgers a review of the disease and its significance for other animals Res Vet Sci 69 203ndash17 doi101053rvsc20000422
Genov I 1965 The effects of certain physical and chemical agents on Mycobacterium tuberculosis Veteriaacuternı Med Nauk Sofia 2 97ndash107
Goacutemez-Brandoacuten M Aira M Lores M Domiacutenguez J 2011 Epigeic Earthworms exert a bottleneck effect on microbial communities through gut associated processes PLoS One 6 doi101371journalpone0024786
Granval P Muys B 1995 Predation on earthworms by terrestrial vertebrates in XII Congress ldquoThe Game and the Manrdquo Sofia Bulgaria p 480
Hutchings MR Harris S 1997 Effects of farm management practices on cattle grazing behaviour and the potential for transmission of bovine tuberculosis from badgers to cattle Vet J 153 149ndash62
Kruuk H Parish T 1981 Feeding specialization of the European badger Meles meles in Scotland UK J Anim Ecol 50 773ndash788 doi1023074136
Lavelle P Spain A 2001 Soil ecology Kluwer Academic Publishers Dordrecht
Lee KE 1985 Earthworms their ecology and relationship with soils and land use Academic P ed
Lowe CN Butt KR 2005 Culture techniques for soil dwelling earthworms A review Pedobiologia (Jena) 49 401ndash413 doi101016jpedobi200504005
Mather JG Christensen O 1988 Surface movements of earthworms in agricultural land Pedobiologia (Jena) 32 399ndash405
Montecchio L Scattolin L Squartini A Butt K 2015 Potential spread of forest soil-borne fungi through earthworm consumption and casting iForest 8 295ndash301 doi103832ifor1217-008
Morris RS Pfeiffer DU Jackson R 1994 The epidemiology of Mycobacterium bovis infections Vet Microbiol 40 153ndash77
Muldowney J Curry J Okeeffe J Schmidt O 2003 Relationships between earthworm populations grassland management and badger densities in County Kilkenny Ireland Pedobiologia (Jena) 47 913ndash919 doi101016S0031-4056(04)70289-9
Neill SD Hanna J OrsquoBrien J McCracken R 1988 Excretion of Mycobacterium bovis by experimentally infected cattle Vet Rec 123 340ndash343 doi101136vr12313340
Palmer M V 2013 Mycobacterium bovis Characteristics of wildlife reservoir hosts Transbound Emerg Dis 60 1ndash13 doi101111tbed12115
Palmer M V Waters WR Whipple DL 2004 Investigation of the transmission of Mycobacterium bovis from deer to cattle through indirect contact Am J Vet Res 65 1483ndash1489 doi102460ajvr2004651483
Phillips CJC Foster CRW Morris PA Teverson R 2003 The transmission of Mycobacterium bovis infection to cattle Res Vet Sci 74 1ndash15
Chapitre 5
173
Prysor Williams A Roberts P Avery LM Killham K Jones DL 2006 Earthworms as vectors of Escherichia coli O157H7 in soil and vermicomposts FEMS Microbiol Ecol 58 54ndash64 doi101111j1574-6941200600142x
Sweeney FP Courtenay O Hibberd V Hewinson RG Reilly L a Gaze WH Wellington EMH 2007 Environmental monitoring of Mycobacterium bovis in badger feces and badger sett soil by real-time PCR as confirmed by immunofluorescence immunocapture and cultivation Appl Environ Microbiol 73 7471ndash3 doi101128AEM00978-07
Taylor AR Taylor AFS 2014 Assessing daily egestion rates in earthworms using fungal spores as a natural soil marker to estimate gut transit time Biol Fertil Soils 50 179ndash183 doi101007s00374-013-0823-5
Thorpe IS Prosser JI Glover L a Killham K 1996 The role of the earthworm Lumbricus terrestris in the transport of bacterial inocula through soil Biol Fertil Soils 23 132ndash139
Vicente J Houmlfle U Garrido JM Fernaacutedez-De-Mera IG Acevedo P Juste R Barral M Gortazar C 2007 Risk factors associated with the prevalence of tuberculosis-like lesions in fenced wild boar and red deer in south central Spain Vet Res 38 451ndash464 doi101051vetres2007002
Young JS Gormley E Wellington EMH 2005 Molecular detection of Mycobacterium bovis and Mycobacterium bovis BCG (Pasteur) in soil Appl Environ Microbiol 71 1946ndash1952 doi101128AEM7141946
Chapitre 5
174
C COMPLEMENT PRELEVEMENT ET ANALYSE DE LOMBRICS DANS UNE PATURE SUSPECTE
Afin drsquoeacutevaluer la contamination naturelle eacuteventuelle des lombrics par M bovis nous avons
analyseacute 90 lombrics preacuteleveacutes en feacutevrier 2014 agrave proximiteacute drsquoune source permanente drsquoune des pacirctures
suspectes de lrsquoAuxois ougrave des bovins infecteacutes ont pacirctureacute lrsquoeacuteteacute preacuteceacutedent (Site C de lrsquoeacutetude
environnementale Chapitre 2) Les lombrics ont eacuteteacute preacuteleveacutes le mecircme jour que les trois eacutechantillons
positifs de ce site (eau seacutediments neacutematodes) Sur sept points de la pacircture choisis au hasard une
pelleteacutee drsquoenviron 20 kg de sol a eacuteteacute retourneacutee Les vers ont eacuteteacute seacutepareacutes du sol rinceacutes avec de lrsquoeau et
transfeacutereacutes individuellement dans des pots en plastique de 60 mL Les pots ont eacuteteacute stockeacutes 48 h agrave 20degC
dans lrsquoobscuriteacute afin de laisser les vers se vider Pour chaque point de la pacircture preacuteleveacute les fegraveces des
vers ont eacuteteacute pooleacutees et homogeacuteneacuteiseacutees puis 2 g ont eacuteteacute preacuteleveacutes et congeleacutes jusqursquoagrave la reacutealisation de
lrsquoanalyse Apregraves extraction de lrsquoADN chaque eacutechantillon a eacuteteacute analyseacute en qPCR avec le systegraveme de
deacutetection RD4 Les protocoles utiliseacutes sont deacutecrits dans le Mateacuteriel et Meacutethodes de lrsquoarticle preacuteceacutedent
Lrsquoidentification pheacutenotypique des 90 vers collecteacutes dans la pacircture a permis de mettre en
eacutevidence cinq genres de lombric diffeacuterents Lumbricus terrestris Lumbricus castaneus Nicodrilus sp
Octolosium cyaneum Allolobophora sp Aporrectodea caliginosa
Aucun des sept pools de fegraveces analyseacutes nrsquoa donneacute de signal positif en qPCR avec le systegraveme RD4
Chapitre 5
175
D BILAN
Les lombrics sont de gros consommateurs de matiegravere organique naturellement chargeacutee drsquoune
flore abondante et varieacutee et potentiellement drsquoune flore bacteacuterienne pathogegravene Toutefois il existe
tregraves peu de donneacutees sur la disseacutemination des bacteacuteries pathogegravenes pour lrsquohomme et les animaux par
les lombrics dans le sol ainsi que sur les risques lieacutes au portage intestinal de bacteacuteries pathogegravenes par
les lombrics pour les animaux qui les consomment
Lrsquoeacutetude deacutecrite dans lrsquoarticle preacuteceacutedent montre qursquoapregraves avoir ingeacutereacute des matiegraveres feacutecales
fraiches inoculeacutees les lombrics ont la capaciteacute de disseacuteminer rapidement M bovis dans le sol
environnant par le biais des turricules qursquoils eacutemettent agrave la surface du sol et en profondeur dans les
tunnels qursquoils creusent Lrsquoenfouissement de la bacteacuterie dans le sol permet probablement drsquoameacuteliorer la
persistance de la bacteacuterie en la proteacutegeant des UV et de la deacuteshydratation Drsquoautre part le transfert
de la bacteacuterie agrave quelques cm des deacutejections animales contamineacutees par le biais des turricules de surface
la rend plus accessible aux animaux (qui en geacuteneacuteral eacutevitent les fegraveces des autres animaux) Ainsi en
fonction des comportements exploratoires ou alimentaires des diffeacuterentes espegraveces freacutequentant les
pacirctures il existe un risque potentiel de contamination par du sol ou des turricules contamineacutes
(inhalation de bioaeacuterosols ou ingestion)
Nous avons eacutegalement montreacute que le tube digestif des lombrics ne constituait pas un reacuteservoir
peacuterenne de M bovis la bacteacuterie nrsquoeacutetait deacutetecteacutee que transitoirement (4 jours) apregraves avoir remis les
lombrics dans un sol sain Le portage digestif pendant les 4 premiers jours constitue toutefois un risque
pour les animaux qui consomment les lombrics en grande quantiteacute notamment les blaireaux et les
sangliers
Cette eacutetude soulegraveve les mecircmes inconnues que les eacutetudes preacuteceacutedentes principalement sur la
viabiliteacute lrsquoeacutetat physiologique et la conservation de la virulence de M bovis apregraves un transfert dans le
sol ou suite au passage de la bacteacuterie dans le tube digestif des lombrics Ces facteurs sont
indispensables pour permettre lrsquoinfection drsquoanimaux via des matrices environnementales contamineacutees
Les moyens drsquoeacutevaluation de ces diffeacuterents facteurs seront discuteacutes dans la partie Perspectives du
manuscrit
Chapitre 5
176
Discussion geacuteneacuterale
Discussion geacuteneacuterale
179
Depuis 2004 face agrave la recrudescence de la tuberculose bovine en Cocircte drsquoOr la mise en place
drsquoun programme de lutte sanitaire reacuteguliegraverement ameacutelioreacute contre la maladie a permis de controcircler et
drsquoameacuteliorer la situation dans lrsquoeacutelevage bovin depuis 2010 Dans la zone endeacutemique du deacutepartement la
circulation de Mycobacterium bovis dans un systegraveme multi-hocircte speacutecifiques impliquant les bovins et
des espegraveces sauvages complique fortement la lutte contre la maladie notamment par la difficulteacute agrave
geacuterer lrsquoinfection dans la faune sauvage malgreacute les mesures de controcircle entreprises jusqursquoagrave maintenant
La transmission indirecte de la maladie entre animaux sauvages et bovins par le biais de substrats
environnementaux contamineacutes est fortement suspecteacutee dans les zones ougrave bovins et faune sauvage
infecteacutes partagent les mecircmes territoires et ressources La transmission environnementale a deacutejagrave eacuteteacute
deacutemontreacutee en conditions expeacuterimentales (Little et al 1982 Palmer et al 2004) mais elle est tregraves
difficile agrave deacutemontrer en conditions naturelles et passe notamment par lrsquoeacutevaluation du niveau
drsquoexcreacutetion par les animaux infecteacutes du rocircle de lrsquoenvironnement comme reacuteservoir de la bacteacuterie de sa
survie dans un eacutetat viable et du maintien de sa virulence
Dans ce contexte les objectifs de ce travail eacutetaient drsquoune part drsquoeacutevaluer la contamination
environnementale en Cocircte drsquoOr sur des agroeacutecosystegravemes freacutequenteacutes par des bovins etou des animaux
sauvages potentiellement infecteacutes dans le but de mettre en eacutevidence des reacuteservoirs
environnementaux eacuteventuels Drsquoautre part nous avons chercheacute agrave preacuteciser les facteurs impliqueacutes dans
la persistance de M bovis dans le sol et sa disseacutemination par la micro- et meacutesofaune du sol
1 Des meacutethodes deacuteveloppeacutees (ou adapteacutees) puis valideacutees dans des matrices environnementales
Notre objectif initial pour lrsquoanalyse des eacutechantillons environnementaux eacutetait drsquoutiliser en
parallegravele la PCR quantitative apregraves extractionpurification de lrsquoADN pour la deacutetection et la quantification
de M bovis et une meacutethode baseacutee sur la culture afin drsquoisoler les souches environnementales pour les
caracteacuteriser Cependant les protocoles de preacutetraitement et de culture tireacutes de la litteacuterature
(immunocapture et protocoles de deacutecontamination) que nous avons utiliseacutes en premiegravere intention sur
les eacutechantillons environnementaux nrsquoont pas donneacute les reacutesultats escompteacutes du fait de la croissance tregraves
lente de M bovis et de la preacutesence dans les eacutechantillons de tregraves nombreux microorganismes
interfeacuterents agrave croissance rapide qui ont gecircneacute la culture (envahissement rapide des boicirctes en une
semaine) La culture de M bovis agrave partir drsquoeacutechantillons environnementaux reste agrave lrsquoheure actuelle un
challenge (Fine OrsquoBrien et al 2011 Fine Bolin et al 2011 Young et al 2005 King et al 2015)
notamment agrave cause de lrsquoabsence de milieux suffisamment seacutelectifs et de bouillons drsquoenrichissement
Discussion geacuteneacuterale
180
adapteacutes comme il en existe notamment pour lrsquoisolement de bacteacuteries pathogegravenes telluriques telles
que Listeria monocytogenes ou Pseudomonas aeruginosa En lrsquoabsence de meacutethodes disponibles nous
avons donc mis au point deux protocoles tregraves innovants agrave la fin de nos travaux un protocole
drsquoextractiondeacutecontamination des mycobacteacuteries dans le sol et une meacutethode de deacutetection des
mycobacteacuteries en culture liquide Ces deux meacutethodes sont en cours drsquoeacutevaluation par la SATT (Socieacuteteacute
drsquoAcceacuteleacuteration de Transfert de Technologies) en vue drsquoune valorisation sous forme de savoir-faire ou de
brevet Des eacutetudes compleacutementaires seront neacutecessaires pour deacutevelopper ces outils innovants
La recherche de reacuteservoirs environnementaux potentiels de M bovis en Cocircte drsquoOr a donc eacuteteacute
baseacutee sur lrsquoutilisation de meacutethodes moleacuteculaires adapteacutees aux matrices environnementales
Contrairement aux meacutethodes culturales la qPCR permet une deacutetection et une quantification rapide
des ADN cibles sans ecirctre gecircneacutee par la preacutesence de contaminants Face au grand nombre de
mycobacteacuteries potentiellement preacutesentes dans les sols de Cocircte drsquoOr dont M microti (Michelet et al
2015) nous avons choisi drsquoutiliser trois systegravemes ciblant trois seacutequences diffeacuterentes dans le geacutenome
des espegraveces du MTBC ou de M bovis que nous avons deacuteveloppeacutes et valideacutes pour leur speacutecificiteacute et leur
sensibiliteacute Lrsquoutilisation conjointe de ces trois systegravemes nous a permis drsquoaugmenter la speacutecificiteacute de la
deacutetection Toutefois le faible niveau de deacutetection observeacute dans certains eacutechantillons ne nous a pas
permis de quantifier le MTBC ou M bovis dans tous les eacutechantillons
2 Contamination environnementale les zones humides les eaux de ruissellement et les sols de terriers sont potentiellement agrave risque
Les systegravemes moleacuteculaires deacuteveloppeacutes ont eacuteteacute utiliseacutes pour lrsquoanalyse de plus de 400
eacutechantillons environnementaux varieacutes preacuteleveacutes dans les agroeacutecosystegravemes de la zone endeacutemique de
Cocircte drsquoOr et de Charente La contamination environnementale a eacuteteacute deacutetecteacutee de faccedilon reacutepeacuteteacutee dans
deux types drsquoeacutecosystegravemes bien particuliers les zones humides preacutesentes sur les pacirctures (sources et
laquo mouilles raquo) les terriers et latrines de blaireaux
Concernant les zones humides nous avons mis en eacutevidence une contamination dans lrsquoeau
circulante et les seacutediments de deux sources naturelles et permanentes situeacutees sur des pacirctures utiliseacutees
par deux cheptels diffeacuterents dans deux communes diffeacuterentes Cette faible contamination de lrsquoeau
etou des seacutediments retrouveacutee agrave lrsquoendroit de la reacutesurgence ainsi que sur diffeacuterents points en aval est
une donneacutee nouvelle puisqursquoagrave ce jour M bovis nrsquoavait eacuteteacute isoleacutee que dans lrsquoeau stagnante et ses
seacutediments (Santos et al 2015) Lors de nos campagnes de preacutelegravevements nous avons constateacute que ces
reacutesurgences parfois capteacutees dans des abreuvoirs eacutetaient parfois le seul moyen drsquoabreuvement des
bovins dans les pacirctures Ces zones humides semblent tregraves attractives pour les bovins au vu des indices
de preacutesence constateacutes (matiegraveres feacutecales et nombreuses empreintes) La persistance de M bovis eacutetant
Discussion geacuteneacuterale
181
favoriseacutee dans les matrices humides riches en matiegravere organique et dans lrsquoeau (Young et al 2005 Fine
Bolin et al 2011 Duffield amp Young 1985 Wint et al 2002) ces zones constituent un milieu favorable
agrave la survie de la bacteacuterie Dans notre eacutetude la mise en eacutevidence de la contamination des zones humides
dans des pacirctures suspectes corrobore le reacutesultat de lrsquoeacutetude PatRisk dans laquelle une association
significative a eacuteteacute eacutetablie entre la preacutesence de mouilles et le statut laquo agrave risque drsquoune pacircture raquo (Bouchez-
Zacria 2015)
Lrsquoorigine de la contamination de ces sources et laquo mouilles raquo est inconnue toutefois elles sont
accessibles aux bovins et animaux sauvages infecteacutes qui peuvent potentiellement excreacuteter la bacteacuterie
lors de lrsquoabreuvement ou du fouissement Vu le contexte geacuteomorphologique il est eacutegalement
envisageable que les infiltrations drsquoeau dans les plateaux calcaires coiffant les valleacutees entrainent avec
elles des matrices environnementales fortement contamineacutees (latrines sols) qui ressortent au niveau
des pacirctures avec toutefois un effet de dilution tregraves important
Des mesures de bioseacutecuriteacute visant agrave limiter les risques lieacutes aux zones humides sont deacutejagrave
preacuteconiseacutees par la DDPP 21 et le GDS 21 agrave lrsquoeacutechelle de la pacircture Lrsquoameacutenagement drsquoabreuvoirs peu
accessibles aux animaux sauvages et la condamnation par des clocirctures de lrsquoaccegraves des bovins aux
laquo mouilles raquo propices agrave la survie de la bacteacuterie sont fortement conseilleacutes aux eacuteleveurs des zones agrave
risque pour limiter le risque de transmission indirecte de la maladie Le drainage des pacirctures est une
solution envisageacutee pour asseacutecher les zones humides et canaliser les sources dans les pacirctures agrave risque
mais il est strictement reacuteglementeacute et soumis agrave autorisation avec enquecircte publique sous le controcircle de
la Police de lrsquoeau (Direction Deacutepartementale des Territoires) La correacutelation de nos reacutesultats sur la
contamination des zones humides avec le bilan des foyers bovins et des cas sauvages ainsi que les
reacutesultats de lrsquoeacutetude PatRisk ont initieacute une reacuteflexion entre la DDPP21 la CIREV le GDS et le SIRTAVA
(Syndicat Intercommunal pour la Reacutealisation des Travaux drsquoAmeacutenagement de la Valleacutee de lrsquoArmanccedilon)
pour eacutevaluer la faisabiliteacute de travaux de drainage dans les pacirctures agrave risque dans le respect de la
regraveglementation Un problegraveme se pose toutefois pour les pacirctures preacutesentant de tregraves nombreuses
reacutesurgences temporaires sur lesquelles le drainage nrsquoest pas envisageable Lrsquoutilisation de ces pacirctures
pour drsquoautres productions animales (ovins chevaux) ou veacutegeacutetales (production de foin la mise en
culture nrsquoest pas toujours possible agrave cause des pentes) est une solution
Notre eacutetude a eacutegalement mis en eacutevidence la contamination des sols de terriers de blaireaux
par M bovis Le geacutenotype retrouveacute dans les sols de terriers est identique agrave celui retrouveacute chez les
blaireaux infecteacutes pieacutegeacutes agrave proximiteacute confirmant que cette espegravece a la capaciteacute de contaminer son
milieu de vie (Young et al 2005 Courtenay et al 2006 Sweeney et al 2006 Sweeney et al 2007) Dans
cette espegravece des animaux excreacuteteurs et super excreacuteteurs ont eacuteteacute mis en eacutevidence en Cocircte drsquoOr (Payne
2014) expliquant les quantiteacutes importantes de bacilles retrouveacutees dans les sols de terrier (Young et al
Discussion geacuteneacuterale
182
2005 Courtenay et al 2006) et les latrines dans notre eacutetude La deacutetection drsquoun signal moleacuteculaire
pendant 12 mois dans plusieurs entreacutees drsquoun mecircme terrier suggegravere fortement que les sols de terriers
peuvent constituer un reacuteservoir environnemental pour M bovis La survie des mycobacteacuteries est
favoriseacutee dans ces structures complexes et profondes ougrave regravegne un microclimat propice (Moore amp Roper
2003) tant au niveau de la stabiliteacute de la tempeacuterature de lrsquohumiditeacute optimale que de la protection
contre les UV Ce reacuteservoir potentiel repreacutesente un risque pour le maintien de la maladie dans le groupe
social de blaireaux qui partage le terrier notamment pour les blairotins qui y naissent pendant lrsquohiver
mais aussi entre les groupes sociaux qui peuvent reacuteguliegraverement changer de terriers Lorsqursquoils sont
localiseacutes dans les pacirctures et sont accessibles aux bovins le sol des terriers contamineacutes repreacutesenterait
un risque potentiel pour les bovins qui les explorent notamment un risque drsquoinhalation de bioaeacuterosols
contamineacutes geacuteneacutereacutes agrave partir de sol Enfin les sols et latrines contamineacutes souvent en limite des plateaux
calcaires seraient susceptibles drsquoecirctre agrave lrsquoorigine de la contamination des pacirctures par le biais des
infiltrations drsquoeau des pheacutenomegravenes de reacutesurgence drsquoeau et de ruissellement
Les mesures de bioseacutecuriteacute preacuteconisant la clocircture des terriers situeacutes dans les pacirctures semblent
essentielles agrave respecter par les eacuteleveurs si lrsquoon considegravere la longue persistance de la bacteacuterie dans ce
biotope particulier Le repeuplement des terriers contamineacutes ne fait lrsquoobjet drsquoaucun controcircle agrave lrsquoheure
actuelle Sous le controcircle du CSRPN (Conseil Scientifique Reacutegional du Patrimoine Naturel) une eacutetude
est actuellement en cours en Cocircte drsquoOr pour lrsquoeacutetude de faisabiliteacute drsquoun protocole de deacutepeuplement des
terriers (pieacutegeage de tous les individus du groupe) associeacute agrave un traitement des tunnels par des reacutepulsifs
permanents pour eacuteviter la recolonisation immeacutediate
La preacutevalence de M bovis dans lrsquoenvironnement des animaux infecteacutes en Cocircte drsquoOr est
probablement sous-estimeacutee dans notre eacutetude agrave cause de limites temporelles spatiales et
meacutethodologiques auxquelles nous avons fait face En premier lieu il existe un deacutelai incompressible de
plusieurs semaines agrave plusieurs mois entre les trois eacutevegravenements suivants i) la contamination
environnementale par un animal excreacuteteur (bovin ou faune sauvage) ii) le diagnostic de lrsquoinfection chez
cet animal (apregraves prophylaxie et abattage pour les bovins en hiver et pieacutegeage pour les blaireaux au
printemps et en eacuteteacute) et iii) le preacutelegravevement et lrsquoanalyse drsquoeacutechantillons environnementaux dans les zones
de vie de ces animaux Cette limite temporelle est eacutegalement eacutevoqueacutee par drsquoautres auteurs (Fine
OrsquoBrien et al 2011) Il serait ideacuteal drsquoeffectuer les preacutelegravevements environnementaux dans un deacutelai le plus
court possible apregraves lrsquoexcreacutetion du bacille par un animal infecteacute toutefois ce deacutelai deacutepend du rythme
des prophylaxies et de la freacutequence du pieacutegeage des blaireaux De plus sur le terrain malgreacute notre
strateacutegie drsquoeacutechantillonnage ciblant les zones partageacutees preacutefeacuterentiellement par la faune domestique et
sauvage les surfaces agrave preacutelever restent importantes Lrsquoanalyse de la totaliteacute des surfaces cibleacutees est
humainement techniquement et eacuteconomiquement irreacutealisable le nombre de preacutelegravevements a donc ducirc
Discussion geacuteneacuterale
183
ecirctre optimiseacute pour aboutir agrave une analyse pertinente Les reacutesultats obtenus ne donnent de ce fait qursquoune
image partielle de la contamination reacuteelle des zones preacuteleveacutees Enfin la charge bacteacuterienne de certains
preacutelegravevements environnementaux nrsquoa pu ecirctre quantifieacutee signifiant que les quantiteacutes de cibles preacutesentes
eacutetaient proches des limites de deacutetection de nos systegravemes moleacuteculaires Pour ces diffeacuterentes raisons
nous ne pouvons donc pas exclure une contamination environnementale plus large avec des quantiteacutes
tregraves faibles de M bovis
3 Survie et circulation de M bovis dans lrsquoenvironnement
La survie des bacteacuteries dans le sol deacutepend des facteurs biotiques et abiotiques qui caracteacuterisent
ces sols Les diffeacuterentes eacutetudes sur la survie de M bovis ont essentiellement consideacutereacute les facteurs
abiotiques climatiques telles que la tempeacuterature lrsquohumiditeacute et lrsquoexposition aux UV dans un seul type
de sol (Young et al 2005 Jackson et al 1995 Duffield amp Young 1985) Nous avons montreacute que M bovis
SB0120 eacutetait cultivable au moins cinq mois agrave une tempeacuterature basse (4degC) avec une mortaliteacute
bacteacuterienne minime et jusqursquoagrave trois mois agrave 22degC Ces dureacutees de survie sont supeacuterieures agrave celles
rapporteacutees dans de preacuteceacutedentes eacutetudes (Fine Bolin et al 2011 Jackson et al 1995 Duffield amp Young
1985) Les sols retrouveacutes dans lrsquoAuxois ont agrave la fois des caracteacuteristiques diffeacuterentes des sols
habituellement deacutecrits comme favorables agrave la survie des mycobacteacuteries (sols sableux agrave pH acide) et
des caracteacuteristiques communes leur richesse en matiegravere organique et en fer Bien que nous nrsquoayons
pas pu mettre en eacutevidence un effet de la nature du sol dans notre eacutetude sur la survie de M bovis il
nous parait probable que les sols argilo-limoneux peacuterennisent la maladie dans les valleacutees de lrsquoAuxois
Notre expeacuterience limiteacutee par des contraintes techniques lieacutees agrave lrsquoutilisation du laboratoire de
confinement L3 du CHU de Dijon aurait neacutecessiteacute lrsquoutilisation drsquoun plus grand nombre de sols de points
de preacutelegravevements et de reacutepeacutetitions biologiques La concentration des foyers bovins (notamment des
recontaminations de cheptels) dans une zone limiteacutee du deacutepartement en Auxois interroge eacutegalement
sur le rocircle du contexte paysager et geacuteomorphologique commun des pacirctures suspectes dans la survie
de M bovis
Ces dureacutees de survie de M bovis dans les sols doivent ecirctre consideacutereacutees avec preacutecaution car
obtenues dans des conditions expeacuterimentales controcircleacutees qui ne miment que partiellement les
conditions environnementales reacuteelles (sols deacutebarrasseacutees de leur flore endogegravene agrave une tempeacuterature
drsquoincubation constante une humiditeacute optimale et une protection contre les UV) Il est probable qursquoen
conditions naturelles la survie de M bovis soit moins longue agrave cause des variations nyctheacutemeacuterales de
tempeacuteratures des variations des preacutecipitations des UV ainsi que de lrsquoeffet barriegravere potentiel de la flore
endogegravene des matrices environnementales La correacutelation de ces donneacutees avec les courbes de
Discussion geacuteneacuterale
184
tempeacuteratures permettent de montrer que la peacuteriode de novembre agrave avril ougrave les tempeacuteratures sont les
plus basses est compatible avec une survie optimale de M bovis dans les sols
Les reacutesultats obtenus dans notre eacutetude de survie ont souleveacute la question de la persistance de
M bovis dans le sol dans un eacutetat viable mais non cultivable (VNC) puisque la bacteacuterie nrsquoest plus
cultivable mais toujours deacutetecteacutee par qPCR Lrsquoeacutetat VNC de M bovis a deacutejagrave eacuteteacute suggeacutereacute dans des eacutetudes
de survie dans du sol (Adams et al 2013 Young et al 2005) Les mycobacteacuteries possegravedent une capaciteacute
particuliegravere agrave srsquoadapter et agrave survivre dans des conditions deacutefavorables en lrsquoabsence de nutriments
drsquooxygegravene sous diffeacuterents stress exogegravenes notamment dans les phagosomes de lrsquohocircte pour les
mycobacteacuteries pathogegravenes (Gengenbacher amp Kaufmann 2012 Boon amp Dick 2012 Voskuil et al 2003)
Les mycobacteacuteries disposent de diffeacuterents systegravemes agrave deux composants (permettant une transduction
de signal) leur permettant la perception sensorielle des conditions environnementales et en particulier
de la composition du milieu extracellulaire (Cook et al 2009 Cole et al 1998) Dans les substrats
environnementaux la bacteacuterie conserve probablement tout son potentiel de perception du milieu qui
lrsquoentoure et ses capaciteacutes drsquoadaptation mais elle ne peut pas interagir avec lui comme avec les cellules
de lrsquohocircte A lrsquoheure actuelle les meacutecanismes drsquoadaptation mis en jeu par M bovis pour survivre lors de
son excreacutetion par lrsquohocircte dans lrsquoenvironnement sont inconnus
Nous avons montreacute pour la premiegravere fois que les lombrics veacuteritables ingeacutenieurs du sol ont la
capaciteacute de disseacuteminer la bacteacuterie dans leur environnement par le biais des tunnels qursquoils creusent et
des turricules qursquoils eacutemettent agrave et sous la surface du sol Lrsquoenfouissement de la bacteacuterie agrave quelques
centimegravetres sous le sol favorise probablement sa survie Les lombrics peuvent eacutegalement jouer un rocircle
de vecteur quand ils portent du sol contamineacute dans leur tube digestif et qursquoils sont ingeacutereacutes par des
blaireaux ou des sangliers Cette eacutetude soulegraveve la probleacutematique du rocircle de la faune du sol dans la
survie et la circulation de M bovis dans lrsquoenvironnement et le risque que des reacuteservoirs cacheacutes de la
bacteacuterie entretiennent agrave bas bruit une contamination de lrsquoenvironnement
Perspectives
Perspectives
187
Lrsquoeacutevaluation de la contamination environnementale reacutealiseacutee en Cocircte drsquoOr et dans une moindre
mesure en Charente (terriers de blaireaux uniquement) nrsquoest pas extrapolable aux autres reacutegions
franccedilaises toucheacutees par la tuberculose A ce jour aucune investigation environnementale aussi
complegravete nrsquoa eacuteteacute meneacutee dans des deacutepartements fortement infecteacutes comme la Dordogne et les
Pyreacuteneacutees Atlantiques ougrave bovins et faune sauvage cohabitent Il serait donc inteacuteressant drsquoeacutevaluer le
niveau de contamination environnementale des zones partageacutees par les bovins et la faune sauvage afin
de veacuterifier si les zones humides et les terriers sont eacutegalement agrave risque dans ces deacutepartements De plus
les reacutesultats obtenus dans la zone endeacutemique de Cocircte drsquoOr ont suggeacutereacute le rocircle possible du contexte
geacuteomorphologique hydrologique et peacutedologique de lrsquoAuxois dans la contamination environnementale
et la persistance de M bovis Pour veacuterifier cette hypothegravese il serait inteacuteressant de comparer le contexte
des pacirctures suspectes de lrsquoAuxois avec les pacirctures suspectes des deacutepartements les plus toucheacutes par la
tuberculose bovine La caracteacuterisation physico-chimique du sol de ces pacirctures permettrait peut-ecirctre de
mettre en relation drsquoeacuteventuelles proprieacuteteacutes communes entre les sols incrimineacutes et la persistance de la
bacteacuterie La mise en eacutevidence de facteurs de risque paysagers hydrographiques etou peacutedologiques
permettrait eacuteventuellement de mettre au point un outil de preacutevision des risques en fonction du
contexte rencontreacute (pacirctures agrave risque) drsquoadapter au mieux la gestion sanitaire et drsquoameacuteliorer la
bioseacutecuriteacute dans les eacutelevages Si le rocircle des zones humides (laquomouilles raquo) et des sources dans drsquoautres
reacutegions est confirmeacute il conviendra de limiter lrsquoaccegraves des bovins agrave ces zones pour limiter la
contamination En Cocircte drsquoOr des approches de ce type (drainage et clocircture des zones humides) pour
empecirccher lrsquoaccegraves des animaux sont en cours en collaboration avec les eacuteleveurs les agences de bassin
lrsquoONEMA ainsi que les gestionnaires des zones humides agrave lrsquoeacutechelle du bassin versant de la Brenne
La dureacutee de survie deacutemontreacutee de M bovis dans les sols remet partiellement en cause la dureacutee
du deacutelai de carence de 2 mois avant reacuteinstallation drsquoanimaux suite agrave la peacuteriode de vide sanitaire dans
les exploitations qui ont eacuteteacute foyer de tuberculose Nos observations devront ecirctre discuteacutees dans le
cadre de lrsquooptimisation de ces pratiques
Sur le plan expeacuterimental notre expeacuterience de survie sur deux sols eacutetant insuffisante une eacutetude
de la survie de M bovis sur un grand nombre de sols aux caracteacuteristiques physico-chimiques
contrasteacutees permettrait drsquoidentifier les facteurs physico-chimiques du sol associeacutes agrave une survie
optimale de la bacteacuterie comme cela a pu ecirctre fait au sein de notre eacutequipe pour Listeria monocytogenes
(Locatelli et al 2013) Lrsquoidentification des facteurs eacutedaphiques favorisant la persistance de M bovis
permettra de mettre en eacutevidence les sols agrave risque
La recherche de M bovis dans des matrices environnementales se heurte aux limites des
techniques utiliseacutees Concernant la qPCR bien qursquoelle reste une technique de choix des efforts sont
encore neacutecessaires pour abaisser les seuils de deacutetection dans les matrices complexes faiblement
Perspectives
188
chargeacutees en mycobacteacuteries Drsquoautres techniques PCR pourraient ecirctre envisageacutees notamment la PCR
digitale (ou PCR numeacuterique) qui permet de gagner en sensibiliteacute et de limiter lrsquoimpact des inhibiteurs
mais dont lrsquoutilisation est encore limiteacutee par le coucirct De plus la qPCR deacutetecte lrsquoADN bacteacuterien sans
donner aucune indication sur la viabiliteacute de la cellule Devant les difficulteacutes agrave isoler M bovis par la
culture agrave partir de matrices environnementales complexes agrave cause notamment de sa croissance tregraves
lente une approche transcriptomique permettrait de preacuteciser lrsquoeacutetat viable de M bovis Cette approche
pourrait eacutegalement apporter des connaissances encore totalement inconnues agrave lrsquoheure actuelle sur les
meacutecanismes de survie mis en jeu par la bacteacuterie une fois excreacuteteacutee par son hocircte dans lrsquoenvironnement
et mettre en eacutevidence les gegravenes impliqueacutes dans son adaptation aux conditions environnementales et
donc sa persistance Dans lrsquoobjectif drsquoeacuteclaircir ces meacutecanismes qui semblent fondamentaux pour la
compreacutehension de lrsquoadaptation et de la survie de M bovis dans lrsquoenvironnement lrsquoutilisation drsquoune
approche transcriptomique permettrait drsquoidentifier les modifications transcriptionnelles agrave lrsquoeacutechelle du
geacutenome complet par une approche de seacutequenccedilage des ARN (RNA seq) et de certains gegravenes par RT-PCR
lors de lrsquointroduction de M bovis dans le sol La comparaison de ces donneacutees avec les reacutesultats obtenus
in vitro et in vivo dans diffeacuterentes conditions de stress (Rodriguez et al 2008 Jang et al 2009)
permettraient de veacuterifier si les mecircmes meacutecanismes sont impliqueacutes Les gegravenes du reacutegulon de la
dormance preacutesents chez M bovis sont surexprimeacutes dans les macrophages in vitro et in vivo (Rodriguez
et al 2008) il serait particuliegraverement inteacuteressant drsquoeacutevaluer leur niveau drsquoexpression lors de
lrsquointroduction de la bacteacuterie dans le sol Comme il est deacutelicat drsquoextraire lrsquoARN de matrices complexes
comme le sol lrsquoutilisation drsquoextraits aqueux de sol pourrait faciliter lrsquoeacutetude (Piveteau et al 2011) Pour
aller plus loin il serait inteacuteressant de comparer les transcriptomes de M bovis lorsque les conditions
abiotiques (type de sol tempeacuterature humiditeacute etc) et biotiques (comparaison absencepreacutesence du
microbiote endogegravene du sol) varient pour cerner plus finement les meacutecanismes drsquoadaptation deacuteployeacutes
face aux diffeacuterents stress
Les meacutethodes de culture de M bovis agrave partir de matrices environnementales complexes
comme le sol sont encore agrave ameacuteliorer Lrsquoisolement de souches environnementales de M bovis
permettrait gracircce aux nouvelles techniques de seacutequenccedilage (NGS) de faire de la geacutenomique
comparative sur geacutenome complet avec les mecircmes souches isoleacutees chez les animaux et drsquoameacuteliorer les
connaissances sur les meacutecanismes geacuteneacutetiques mis en jeu dans lrsquoadaptation et la persistance de la
bacteacuterie dans lrsquoenvironnement
Concernant la faune la persistance et la circulation des mycobacteacuteries dans lrsquoeau et le sol
pourraient ecirctre favoriseacutees par la micro- et la meacutesofaune preacutesentes dans ces matrices Les protozoaires
et particuliegraverement les amibes ont fait lrsquoobjet drsquoeacutetudes montrant la survie dans leurs trophozoiumltes et
leurs cystes de nombreuses espegraveces dont M bovis (Taylor et al 2003 Mba Medie et al 2011) Dans
notre eacutetude un des eacutechantillons de neacutematodes preacuteleveacute dans les seacutediments drsquoune source a donneacute un
Perspectives
189
signal moleacuteculaire reacuteveacutelant un lien potentiel entre neacutematodes et persistance des mycobacteacuteries Pour
des raisons de temps et de compeacutetences nous nrsquoavons pas exploreacute minutieusement ces eacuteventuels
reacuteservoirs cacheacutes de M bovis Devant lrsquoabondance et la diversiteacute de ces animaux dans les matrices que
nous analysons des recherches compleacutementaires nous semblent neacutecessaires pour preacuteciser leur rocircle
potentiel dans la persistance de M bovis
Les diffeacuterents reacutesultats obtenus pendant ce travail et les implications qui en ressortent ont eacuteteacute
reacuteguliegraverement partageacutes avec les instances sanitaires et agricoles de Cocircte drsquoOr afin drsquoenrichir les
connaissances sur lrsquoeacutepideacutemiologie tregraves complexe de cette maladie et de participer agrave lrsquoameacutelioration des
mesures de bioseacutecuriteacute qui en deacutecoulent Drsquoautres deacutepartements toucheacutes par la tuberculose bovine
inteacuteresseacutes par nos travaux nous ont fait parvenir des demandes drsquoanalyse environnementale dans des
zones ougrave la transmission indirecte de la maladie est fortement suspecteacutee
Les deux protocoles innovants pour lrsquoextraction des mycobacteacuteries du sol et le suivi de leur
croissance en milieu liquide deacuteveloppeacutes dans ce travail de thegravese font lrsquoobjet drsquoune eacutetude de valorisation
(protection intellectuelle et eacutetude de marcheacute) en collaboration avec la SATT Grand Est
Enfin les diffeacuterentes ideacutees et perspectives deacuteveloppeacutees dans cette thegravese ont eacuteteacute reprises en
partie dans le cadre de lrsquoappel drsquooffre Horizon 2020 agrave travers le deacutepocirct drsquoun projet europeacuteen
multipartenaire (scientifiques et entreprises) qui est coordonneacute par lrsquoINRA de Tours Nouzilly et qui
implique de nombreux partenaires en Allemagne (Lionex PME speacutecialiseacutee dans les outils de deacutetection
seacuterologique speacutecifiques) en Hollande (WUR) en UK (APHA) en Irlande (UCD) en Italie (Universiteacute de
Naples) Ce projet intituleacute DEPICT vise entre autre agrave une meilleure compreacutehension des meacutecanismes
infectieux de la reacuteaction inflammatoire et immunologique des animaux mais eacutegalement agrave deacuteterminer
le rocircle de lrsquoenvironnement dans la survie des pathogegravenes comme M bovis et Map entre autres La
diversiteacute et la persistance de M bovis et Map seront eacutetudieacutees dans lrsquoenvironnement afin de deacuteterminer
les facteurs influenccedilant la survie de ces pathogegravenes en particulier dans les sols Au travers de ce
programme et drsquoautres collaborations agrave venir avec lrsquoINRA de Tours le travail deacutebuteacute au cours de cette
thegravese sur M bovis sera eacutelargi agrave Map mycobacteacuterie eacutegalement difficilement cultivable et revecirctant une
grande importance pour la santeacute animale et la santeacute humaine (elle serait impliqueacutee dans la maladie de
Crohn) Les nouveaux outils (culture et deacutetection) deacuteveloppeacutes au cours de cette thegravese seront testeacutes sur
Map pour permettre lrsquoisolement drsquoun plus grand nombre de souches environnementales
Une des questions en suspens reste la deacutemonstration de la contamination de bovins agrave partir
de M bovis preacutesente dans des matrices environnementales Des expeacuteriences de contamination
expeacuterimentale en animalerie P3 agrave partir de sol ou drsquoeau contamineacutes pourraient permettre cette
deacutemonstration Lrsquoutilisation du modegravele cobaye espegravece tregraves sensible agrave lrsquoinfection par M bovis pour
reacutealiser des expeacuteriences preacuteliminaires pourrait ecirctre envisageacutee avant de concevoir des expeacuteriences sur
bovins Bien que ce modegravele animal soit assez eacuteloigneacute du modegravele bovin en ce qui concerne lrsquoeacutetude de la
Perspectives
190
transmission indirecte de M bovis (les cobayes geacutenegraverent par exemple beaucoup moins drsquoaeacuterosols que
les bovins quand ils boivent et ils ne ruminent pas) il constitue un modegravele plus facilement manipulable
et moins coucircteux Il permettrait drsquoaffiner les donneacutees sur les voies de transmission environnementales
potentielles avant de passer sur un modegravele bovin beaucoup plus contraignant de par la taille des
animaux et plus coucircteux
Reacutefeacuterences bibliographiques
Reacutefeacuterences bibliographiques
192
Acevedo-Whitehouse K Vicente J Gortaacutezar C Houmlfle U Fernaacutendez-de-Mera IG Amos W 2005 Genetic resistance to bovine tuberculosis in the Iberian wild boar Mol Ecol 14 3209ndash17
Adams AP Bolin SR Fine AE Bolin CA Kaneene JB 2013 Comparison of PCR versus culture for detection of Mycobacterium bovis after experimental inoculation of various matrices held under environmental conditions for extended periods Appl Environ Microbiol 79 6501ndash6506
Adekambi T Ben Salah S Khlif M Raoult D Drancourt M 2006 Survival of environmental mycobacteria in Acanthamoeba polyphaga Appl Environ Microbiol 72 5974ndash5981
Adeacutekambi T Drancourt M 2004 Dissection of phylogenetic relationships among 19 rapidly growing Mycobacterium species by 16S rRNA hsp65 sodA recA and rpoB gene sequencing Int J Syst Evol Microbiol 54 2095ndash105
Aimeacute B Lequen L Balageas A Haddad N Maugein J 2012 M bovis and M caprae infections in Aquitaine A clinico-epidemiologic study of 15 patients Pathol Biol (Paris) 60 156ndash159
Alexander K A Laver PN Michel AL Williams M van Helden PD Warren RM van Pittius NCG 2010 Novel Mycobacterium tuberculosis complex pathogen M Mungi Emerg Infect Dis 16 1296ndash1299
Allix C Walravens K Saegerman C Godfroid J Supply P Fauville-Dufaux M 2006 Evaluation of the epidemiological relevance of variable-number tandem-repeat genotyping of Mycobacterium bovis and comparison of the method with IS6110 restriction fragment length polymorphism analysis and spoligotyping J Clin Microbiol 44 1951ndash1962
Aacutelvarez AH Estrada-Chaacutevez C Flores-Valdez MA 2009 Molecular findings and approaches spotlighting Mycobacterium bovis persistence in cattle Vet Res 40 1ndash16
Ameni G Vordermeier M Firdessa R Aseffa A Hewinson G Gordon S V Berg S 2011 Mycobacterium tuberculosis infection in grazing cattle in central Ethiopia Vet J 188 359ndash361
ANSES 2011 Tuberculose bovine et faune sauvageRapport Avril 2011
Antoine D Jarlier V 2010 La tuberculose humaine agrave Mycobacterium bovis en France Bull eacutepideacutemiologique santeacute Anim Aliment Septembre
Aranaz A Cousins D Mateos A Domınguez L 2003 Elevation of Mycobacterium tuberculosis subsp caprae (Aranaz et al 1999) to species rank as Mycobacterium caprae comb nov sp Int J Syst Evol Microbiol 53 1785ndash1789
Aranaz A Lieacutebana E Goacutemez-Mampaso Enrique Galaacuten JC Cousins D Ortega A Blaacutezquez J Baquero F Mateos A Suacutearez G Domiacutenguez L 1999 Mycobacterium tuberculosis subsp caprae subsp nov a taxonomic study of a new member of the Mycobacterium tuberculosis complex isolated from goats in Spain Int J Syst Bacteriol 49 1263ndash1273
Archuleta RJ Yvonne Hoppes P Primm TP 2005 Mycobacterium avium enters a state of metabolic dormancy in response to starvation Tuberculosis (Edinb) 85 147ndash58
Aronson T Holtzman A Glover N Boian M Froman S Berlin OGW Hill H Stelma G J 1999 Comparison of large restriction fragments of Mycobacterium avium isolates recovered from AIDS and non-AIDS patients with those of isolates from potable water J Clin Microbiol 37 1008ndash1012
Ayele WY Neill SD Zinsstag J Weiss MG Pavlik I 2004 Bovine tuberculosis an old disease but a new threat to Africa Int J Tuberc Lung Dis 8 924ndash937
Ballesteros C Garrido JM Vicente J Romero B Galindo RC Minguijoacuten E Villar M Martiacuten-Hernando
Reacutefeacuterences bibliographiques
193
MP Sevilla I Juste R Aranaz A de la Fuente J Gortaacutezar C 2009 First data on Eurasian wild boar response to oral immunization with BCG and challenge with a Mycobacterium bovis field strain Vaccine 27 6662ndash6668
Barasona JA Latham MC Acevedo P Armenteros JA Latham ADM Gortazar C Carro F Soriguer RC Vicente J 2014 Spatiotemporal interactions between wild boar and cattle Implications for cross-species disease transmission Vet Res 45 1ndash11
Barasona JA Torres MJ Aznar J Gortaacutezar C Vicente J 2015 DNA detection reveals Mycobacterium tuberculosis complex shedding routes in its wildlife reservoir the Eurasian wild boar Transbound Emerg Dis (December) doi101111tbed12458
Barker J Brown MRW 1994 Trojan Horses of the microbial world protozoa and the survival of bacterial pathogens in the environment Microbiology 140 1253ndash1259
Baubet E Ropert-Coudert Y Brandt S 2003 Seasonal and annual variations in earthworm consumption by wild boar (Sus scrofa scrofa L) Wildl Res 30 179
Ben Salah I Drancourt M 2010 Surviving within the amoebal exocyst the Mycobacterium avium complex paradigm BMC Microbiol 10 99
Beacutenet J Boschiroli ML Dufour B Garin-Bastuji B 2006 Lutte contre la tuberculose bovine en France de 1954 agrave 2005 analyse de la pertinence eacutepideacutemiologique de lrsquoeacutevolution de la reacuteglementation Epideacutemiologie santeacute Anim 50 127ndash143
Benham PF 1985 The behaviour of badgers and cattle and some factors that affect the chance of contact between the species Appl Anim Behav Sci 14 390ndash391
Benham PF Broom DM 1989 Interactions between cattle and badgers at pasture with reference to bovine tuberculosis transmission Br Vet J 145 226ndash241
Bercovier H Kafri O Sela S 1986 Mycobacteria possess a surprisingly small number of ribosomal RNA genes in relation to the size of their genome Biochem Biophys Res Commun 136 1136ndash1141
Berg S Garcia-Pelayo MC Muumlller B Hailu E Asiimwe B Kremer K Dale J Boniotti MB Rodriguez S Hilty M Rigouts L Firdessa R MacHado A Mucavele C Ngandolo BNR Bruchfeld J Boschiroli ML Muumlller A Sahraoui N Pacciarini M Cadmus SI Joloba M Van Soolingen D Michel AL Djoslashnne B Aranaz A Zinsstag J Van Helden P Portaels F Kazwala R Kaumlllenius G Hewinson RG Aseffa A Gordon S V Smith NH 2011 African 2 a clonal complex of Mycobacterium bovis epidemiologically important in East Africa J Bacteriol 193 670ndash678
Bermingham ML Bishop SC Woolliams JA Pong-Wong R Allen AR McBride SH Ryder JJ Wright DM Skuce RA McDowell SW Glass EJ 2014 Genome-wide association study identifies novel loci associated with resistance to bovine tuberculosis Heredity (Edinb) 112 543ndash551
Beširović H Alić A Špičić S Cvetnić Ž Prašović S Velić L 2012 Bovine tuberculosis in Bosnia and Herzegovina caused by Mycobacterium caprae Vet Arh 82 341ndash349
Beumer A King D Donohue M Mistry J Covert T Pfaller S 2010 Detection of Mycobacterium avium subsp paratuberculosis in drinking water and biofilms by quantitative PCR Appl Environ Microbiol 76 7367ndash7370
Biek R OrsquoHare A Wright D Mallon T McCormick C Orton RJ McDowell S Trewby H Skuce RA Kao RR 2012 Whole genome sequencing reveals local transmission patterns of Mycobacterium bovis in sympatric cattle and badger populations PLoS Pathog 8 e1003008
Biet F Boschiroli ML Thorel MF Guilloteau LA 2005 Zoonotic aspects of Mycobacterium bovis and
Reacutefeacuterences bibliographiques
194
Mycobacterium avium-intracellulare complex (MAC) Vet Res36 411-436
Bille E Dauphin B Leto J Bougnoux M-E Beretti J-L Lotz A Suarez S Meyer J Join-Lambert O Descamps P Grall N Mory F Dubreuil L Berche P Nassif X Ferroni A 2012 MALDI-TOF MS Andromas strategy for the routine identification of bacteria mycobacteria yeasts Aspergillus spp and positive blood cultures Clin Microbiol Infect 18 1117ndash1125
Biswas S Rolain JM 2013 Use of MALDI-TOF mass spectrometry for identification of bacteria that are difficult to culture J Microbiol Methods 92 14ndash24
Blackwood KS He C Gunton J Turenne CY Wolfe J Kabani A M 2000 Evaluation of recA sequences for identification of Mycobacterium species J Clin Microbiol 38 2846ndash2852
Bland CS Ireland JM Lozano E Alvarez ME Primm TP 2005 Mycobacterial ecology of the Rio Grande Appl Environ Microbiol 71 5719ndash5727
Boadella M Barasona JA Diaz-Sanchez S Lyashchenko KP Greenwald R Esfandiari J Gortaacutezar C 2012 Performance of immunochromatographic and ELISA tests for detecting fallow deer infected with Mycobacterium bovis Prev Vet Med 104 160ndash164
Boadella M Lyashchenko K Greenwald R Esfandiari J Jaroso R Carta T Garrido JM Vicente J de la Fuente J Gortaacutezar C 2011 Serologic tests for detecting antibodies against Mycobacterium bovis and Mycobacterium avium subspecies paratuberculosis in Eurasian wild boar (Sus scrofa scrofa) J Vet Diagnostic Investig 23 77ndash83
Boumlddinghaus B Rogall T Flohr T Bloumlcker H Boumlttger EC 1990 Detection and identification of mycobacteria by amplification of rRNA J Clin Microbiol 28 1751ndash1759
Boumlhm M Hutchings MR White PCL 2009 Contact networks in a wildlife-livestock host community identifying high-risk individuals in the transmission of bovine TB among badgers and cattle PLoS One 4 e5016
Bollo E Guarda F Capucchio MT Galietti F 1998 Direct detection of Mycobacterium tuberculosis Complex and M avium Complex in tissue specimens from cattle through identification of specific rRNA sequences J Vet Med 45 395ndash400
Boniotti MB Goria M Loda D Garrone A Benedetto A Mondo A Tisato E Zanoni M Zoppi S Dondo A Tagliabue S Bonora S Zanardi G Pacciarini ML 2009 Molecular typing of Mycobacterium bovis strains isolated in Italy from 2000 to 2006 and evaluation of variable-number tandem repeats for geographically optimized genotyping J Clin Microbiol 47 636ndash644
Boon C Dick T 2012 How Mycobacterium tuberculosis goes to sleep the dormancy survival regulator DosR a decade later Future Microbiol 7 513ndash518
Boon C Dick T 2002 Mycobacterium bovis BCG response regulator essential for hypoxic dormancy J Bacteriol 184 6760ndash6767
Boon C Li R Qi R Dick T 2001 Proteins of Mycobacterium bovis BCG induced in the Wayne dormancy model J Bacteriol 183 2672ndash2676
Boulahbal F Robert J Trystram D Benoist A C Vincent V Jarlier V 1998 La tuberculose humaine agrave Mycobacterium bovis en France durant lrsquoanneacutee 1995 Bull eacutepideacutemiologique Hebd 48
Boyard C Vourcrsquoh G Barnouin J 2008 The relationships between Ixodes ricinus and small mammal species at the woodland-pasture interface Exp Appl Acarol 44 61ndash76
Brennan PJ Nikaido H 1995 The envelope of mycobacteria Annu Rev Biochem 64 29ndash63
Reacutefeacuterences bibliographiques
195
Brodin P Eiglmeier K Marmiesse M Billault A Garnier T Niemann S Cole ST Brosch R 2002 Bacterial artificial chromosome-based comparative genomic analysis identifies Mycobacterium microti as a natural ESAT-6 deletion mutant Infect70 5568ndash5578
Brook RK McLachlan SM 2009 Transdisciplinary habitat models for elk and cattle as a proxy for bovine tuberculosis transmission risk Prev Vet Med 91 197ndash208
Brook RK Wal E Vander van Beest FM McLachlan SM 2012 Evaluating use of cattle winter feeding areas by elk and white-tailed deer Implications for managing bovine tuberculosis transmission risk from the ground up Prev Vet Med 1ndash11
Brosch R Gordon SV Marmiesse M Brodin P Buchrieser C Eiglmeier K Garnier T Gutierrez C Hewinson G Kremer K Parsons LM Pym A Samper S van Soolingen D Cole S 2002 A new evolutionary scenario for the Mycobacterium tuberculosis complex Proc Natl Acad Sci U S A 99 3684ndash3689
Brosch R Gordon S V Garnier T Eiglmeier K Frigui W Valenti P Dos Santos S Duthoy S Lacroix C Garcia-Pelayo C Inwald JK Golby P Garcia JN Hewinson RG Behr M A Quail M A Churcher C Barrell BG Parkhill J Cole ST 2007 Genome plasticity of BCG and impact on vaccine efficacy Proc Natl Acad Sci U S A 104 5596ndash5601
Brotherstone S White IMS Coffey M Downs SH Mitchell AP Clifton-Hadley RS More SJ Good M Woolliams JA 2010 Evidence of genetic resistance of cattle to infection with Mycobacterium bovis J Dairy Sci 93 1234ndash1242
Broughan JM Crawshaw TR Downs SH Brewer J Clifton-Hadley RS 2013a Mycobacterium bovis infections in domesticated non-bovine mammalian species Part 2 A review of diagnostic methods Vet J 198 346ndash351
Broughan JM Downs SH Crawshaw TR Upton PA Brewer J Clifton-Hadley RS 2013b Mycobacterium bovis infections in domesticated non-bovine mammalian species Part 1 Review of epidemiology and laboratory submissions in Great Britain 2004-2010 Vet J 198 339ndash345
Buddle BM Wedlock DN Denis M Vordermeier HM Hewinson RG 2011 Update on vaccination of cattle and wildlife populations against tuberculosis Vet Microbiol 151 14ndash22
Butler WR Guthertz LS 2001 Mycolic acid analysis by High-Performance Liquid Chromatography for identification of Mycobacterium species Clin Microbiol Rev 14 704ndash726
Cadmus SI Adesokan HK Jenkins AO van Soolingen D 2009a Mycobacterium bovis and M tuberculosis in goats Nigeria Emerg Infect Dis 15 2066ndash2067
Cadmus SI Jenkins AO Godfroid J Osinusi K Adewole IF Murphy RL Taiwo BO 2009b Mycobacterium tuberculosis and Mycobacterium africanum in stools from children attending an immunization clinic in Ibadan Nigeria Int J Infect Dis 13 740ndash744
Cadmus SI Palmer S Okker M Dale J Gover K Smith N Jahans K Hewinson RG Gordon S V 2006 Molecular analysis of human and bovine tubercle bacilli from a local setting in Nigeria J Clin Microbiol 44 29ndash34
Cadmus SI Yakubu MK Magaji AA Jenkins AO van Soolingen D 2010 Mycobacterium bovis but also M africanum present in raw milk of pastoral cattle in north-central Nigeria Trop Anim Health Prod 42 1047ndash1048
Cambau E Lamaitre N Sougakoff W Jarlier V 2003 Reacutesistance aux antituberculeux Antibiotiques 5 29ndash37
Reacutefeacuterences bibliographiques
196
Carbonnelle B Dailloux M Lebrun L Maugein J Pernot C 2003 Cahier de biologie meacutedicale Ndeg29Mycobacteacuteries mycobacteacuterioses
Carrasco-Garcia R Barasona JA Gort Montoro V Sanchez-Vizcaino JM Vicente J 2015 Wildlife and livestock use of extensive farm resources in South Central Spain implications for disease transmission Eur J Wildl Res62(1) 65-78
Cassidy JP 2006 The pathogenesis and pathology of bovine tuberculosis with insights from studies of tuberculosis in humans and laboratory animal models Vet Microbiol 112 151ndash161
Cassidy JP Bryson DG Pollock JM Evans RT Forster F Neill SD 1999 Lesions in cattle exposed to Mycobacterium bovis-inoculated calves J Comp Pathol 121 321ndash37
Castets M Sarrat H 1968 Bacteriologic aspects of mycobacteria isolated at Dakar in 1967 Bull Soc Med Afr Noire Lang Fr 13 463ndash469
Cavalerie L Courcoul A Boschiroli ML Reacuteveillaud E Gay P 2014 Tuberculose bovine en France en 2014 une situation stable Bull eacutepideacutemiologique santeacute Anim Aliment 4ndash11
Cha D Cheng D Liu M Zeng Z Hu X Guan W 2009 Analysis of fatty acids in sputum from patients with pulmonary tuberculosis using gas chromatography-mass spectrometry preceded by solid-phase microextraction and post-derivatization on the fiber J Chromatogr A 1216 1450ndash1457
Chapinal N Elkin BT Joly DO Schumaker BA Stephen C 2012 Agreement between the caudal fold test and serological tests for the detection of Mycobacterium bovis infection in bison Prev Vet Med 105 326ndash330
Ciliberti A Gavier-Wideacuten D Yon L Hutchings MR Artois M 2015 Prioritisation of wildlife pathogens to be targeted in European surveillance programmes Expert-based risk analysis focus on ruminants Prev Vet Med 118 271ndash284
Cirillo JD Falkow S Tompkins LS Bermudez LE 1997 Interaction of Mycobacterium avium with environmental amoebae enhances virulence Infect Immun 65 3759ndash3767
Cleary GP Corner LAL OrsquoKeeffe J Marples NM 2011 Diet of the European badger (Meles meles) in the Republic of Ireland A comparison of results from an analysis of stomach contents and rectal faeces Mamm Biol 76 470ndash475
Clifton-Hadley RS Wilesmith JW Stuart F A 1993 Mycobacterium bovis in the European badger (Meles meles) epidemiological findings in tuberculous badgers from a naturally infected population Epidemiol Infect 111 9ndash19
Cole ST Brosch R Parkhill J Garnier T Churcher C Harris D Gordon S V Eiglmeier K Gas S Barry CE Tekaia F Badcock K Basham D Brown D Chillingworth T Connor R Davies R Devlin K Feltwell T Gentles S Hamlin N Holroyd S Hornsby T Jagels K Krogh A McLean J Moule S Murphy L Oliver K Osborne J Quail MA Rajandream MA Rogers J Rutter S Seeger K Skelton J Squares R Squares S Sulston JE Taylor K Whitehead S Barrell BG 1998 Deciphering the biology of Mycobacterium tuberculosis from the complete genome sequence Nature 396 190
Coleman JD Cooke MM 2001 Mycobacterium bovis infection in wildlife in New Zealand Tuberculosis (Edinb) 81 191ndash202
Collins CH Grange JM 1983 The bovine tubercle bacillus J Appl Bacteriol 55 13ndash29
Cooper SM Scott HM de la Garza GR Deck AL Cathey JC 2010 Distribution and interspecies contact of feral Swine and cattle on rangeland in South Texas implications for disease transmission J Wildl Dis 46 152ndash164
Reacutefeacuterences bibliographiques
197
Corner LAL 2006 The role of wild animal populations in the epidemiology of tuberculosis in domestic animals How to assess the risk Vet Microbiol 112 303ndash312
Corner LAL Costello E Lesellier S OrsquoMeara D Sleeman DP Gormley E 2007 Experimental tuberculosis in the European badger (Meles meles) after endobronchial inoculation of Mycobacterium bovis I Pathology and bacteriology Res Vet Sci 83 53ndash62
Corner LAL Murphy D Gormley E 2011 Mycobacterium bovis infection in the Eurasian badger (Meles meles) the disease pathogenesis epidemiology and control J Comp Pathol 144 1ndash24
Corner LAL OrsquoMeara D Costello E Lesellier S Gormley E 2012 The distribution of Mycobacterium bovis infection in naturally infected badgers Vet J 194 166ndash172
Coscolla M Gagneux S 2014 Consequences of genomic diversity in Mycobacterium tuberculosis Semin Immunol 26 431ndash444
Coscolla M Lewin A Metzger S Maetz-Rennsing K Calvignac-Spencer S Nitsche A Dabrowski PW Radonic A Niemann S Parkhill J Couacy-Hymann E Feldman J Comas I Boesch C Gagneux S Leendertz FH 2013 Novel Mycobacterium tuberculosis complex isolate from a wild chimpanzee Emerg Infect Dis 19 969ndash976
Cosivi O Grange JM Daborn CJ Raviglione MC Fujikura T Cousins D Robinson R A Huchzermeyer HF de Kantor I Meslin FX 1999 Zoonotic tuberculosis due to Mycobacterium bovis in developing countries Emerg Infect Dis 4 59ndash70
Coulombe F Divangahi M Veyrier F de Leacuteseacuteleuc L Gleason JL Yang Y Kelliher MA Pandey AK Sassetti CM Reed MB Behr M 2009 Increased NOD2-mediated recognition of N-glycolyl muramyl dipeptide J Exp Med 206 1709ndash1716
Courcoul A Moyen J-L Brugegravere L Faye S Heacutenault S Gares H Boschiroli ML 2014 Estimation of sensitivity and specificity of bacteriology histopathology and PCR for the confirmatory diagnosis of bovine tuberculosis using latent class analysis PLoS One 9 e90334
Courtenay O Reilly L A Sweeney FP Hibberd V Bryan S Ul-Hassan A Newman C Macdonald DW Delahay RJ Wilson GJ Wellington EMH 2006 Is Mycobacterium bovis in the environment important for the persistence of bovine tuberculosis Biol Lett 2 460ndash462
Cousins D V Bastida R Cataldi A Quse V Redrobe S Dow S Duignan P Murray A Dupont C Ahmed N Collins DM Butler WR Dawson D Rodriacuteguez D Loureiro J Romano MI Alito A Zumarraga M Bernardelli A 2003 Tuberculosis in seals caused by a novel member of the Mycobacterium tuberculosis complex Mycobacterium pinnipedii sp nov Int J Syst Evol Microbiol 53 1305ndash1314
Couvin D Rastogi N 2014 La mise en place de bases de donneacutees des geacutenotypes circulants du complexe Mycobacterium tuberculosis et des outils web permettant de mieux surveiller comprendre et controcircler lrsquoeacutepideacutemie de la tuberculose dans le monde EuroReference Cahier num 12
Cowie CE Hutchings MR Barasona JA Gortaacutezar C Vicente J White PCL 2015 Interactions between four species in a complex wildlife livestock disease community implications for Mycobacterium bovis maintenance and transmission Eur J Wildl Res62(1) 51-64
Cvetnic Z Katalinic-Jankovic V Sostaric B Spicic S Obrovac M Marjanovic S Benic M Kirin BK Vickovic I 2007 Mycobacterium caprae in cattle and humans in Croatia Int J Tuberc Lung Dis 11 652ndash658
Dale JW Brittain D Cataldi AA Cousins D Crawford JT Driscoll J Heersma H Lillebaek T Quitugua T Rastogi N Skuce RA Sola C Van Soolingen D Vincent V 2001 Spacer oligonucleotide typing of bacteria of the Mycobacterium tuberculosis complex Recommendations for standardised nomenclature
Reacutefeacuterences bibliographiques
198
Int J Tuberc Lung Dis 5 216ndash219
Dalley D Daveacute D Lesellier S Palmer S Crawshaw T Hewinson RG Chambers M 2008 Development and evaluation of a gamma-interferon assay for tuberculosis in badgers (Meles meles) Tuberculosis 88 235ndash243
Danelishvili L Wu M Stang B Harriff M Cirillo SLG Cirillo JD Bildfell R Arbogast B Bermudez LE 2007 Identification of Mycobacterium avium pathogenicity island important for macrophage and amoeba infection Proc Natl Acad Sci U S A 104 11038ndash11043
Dang NA Kuijper S Walters E Claassens M van Soolingen D Vivo-Truyols G Janssen H-G Kolk AHJ 2013 Validation of biomarkers for distinguishing Mycobacterium tuberculosis from non-tuberculous mycobacteria using gas chromatography-mass spectrometry and chemometrics PLoS One 8 e76263
Daniel TM 2006 The history of tuberculosis Respir Med 100 1862ndash1870
David H Levy Frebaut U Thorel MF 1989 Meacutethodes de laboratoire pour mycobacteacuteriologie clinique Institut Pasteur p 35-36
Dawson KL Bell A Kawakami RP Coley K Yates G Collins DM 2012 Transmission of Mycobacterium orygis (M tuberculosis complex species) from a tuberculosis patient to a dairy cow in New Zealand J Clin Microbiol 50 3136ndash3138
de Jong BC Antonio M Gagneux S 2010 Mycobacterium africanum-review of an important cause of human tuberculosis in West Africa PLoS Negl Trop Dis 4(9) e744
de la Rua-Domenech R 2006 Human Mycobacterium bovis infection in the United Kingdom Incidence risks control measures and review of the zoonotic aspects of bovine tuberculosis Tuberculosis (Edinb) 86 77ndash109
de la Rua-Domenech R Goodchild AT Vordermeier HM Hewinson RG Christiansen KH Clifton-Hadley RS 2006 Ante mortem diagnosis of tuberculosis in cattle a review of the tuberculin tests gamma-interferon assay and other ancillary diagnostic techniques Res Vet Sci 81 190ndash210
Dean GS Rhodes SG Coad M Adam O Cockle PJ Clifford DJ Hewinson RG Vordermeier HM 2005 Minimum infective dose of Mycobacterium bovis in cattle Infect Immun 73 6467ndash6471
Delahay RJ Cheeseman CL Clifton-Hadley RS 2001 Wildlife disease reservoirs the epidemiology of Mycobacterium bovis infection in the European badger (Meles meles) and other British mammals Tuberculosis (Edinb) 81 43ndash49
Delahay RJ De Leeuw ANS Barlow AM Clifton-Hadley RS Cheeseman CL 2002 The status of Mycobacterium bovis Infection in UK Wild Mammals A Review Vet J 164 90ndash105
Delahay RJ Langton S Smith GC Clifton-Hadley RS Cheeseman CL 2000 The spatio-temporal distribution of Mycobacterium bovis (bovine tuberculosis) infection in a high-density badger population J Anim Ecol 69 428ndash441
Delahay RJ Walker N Smith G Wilkinson D Clifton-Hadley R Cheeseman C Tomlinson A Chambers M 2013 Long-term temporal trends and estimated transmission rates for Mycobacterium bovis infection in an undisturbed high-density badger (Meles meles) population Epidemiol Infect 141 1445ndash1456
Delaune D Janvier F Rapp C Geacuterocircme P Fabre M Soler C Meacuterens A 2012 Actualiteacutes de lrsquoinfection agrave Mycobacterium bovis en France agrave propos de 4 cas Ann Biol Clin (Paris) 70 231ndash236
Desikan S Narayanan S 2015 Genetic markers genotyping methods amp next generation sequencing in Mycobacterium tuberculosis Indian J Med Res 141 761ndash774
Reacutefeacuterences bibliographiques
199
Desmond E Ahmed AT Probert WS Ely J Jang Y Sanders C A Lin SY Flood J 2004 Mycobacterium africanum Cases California Emerg Infect Dis 10 921ndash923
Dewes HF 1996 The rate of soil ingestion by dairy cows and the effect on available copper calcium sodium and magnesium N Z Vet J 44 199ndash200
Dick T Lee BH Murugasu-Oei B 1998 Oxygen depletion induced dormancy in Mycobacterium smegmatis FEMS Microbiol Lett 163 159ndash164
Dippenaar A Parsons SDC Sampson SL van der Merwe RG Drewe JA Abdallah AM Siame KK Gey van Pittius NC van Helden PD Pain A Warren RM 2015 Whole genome sequence analysis of Mycobacterium suricattae Tuberculosis 95(6) 682-688
Djelouadji Z Raoult D Drancourt M 2011 Palaeogenomics of Mycobacterium tuberculosis Epidemic bursts with a degrading genome Lancet Infect Dis 11 641ndash650
Dondo A Zoppi S Rossi F Chiavacci L Barbaro A Garrone A Benedetto A Goria M 2007 Mycobacteriosis in Wild Boar Results of 2000-2006 Activity in North-Western Italy Epideacutemiologie santeacute Anim 51 35ndash42
Drancourt M Adeacutekambi T Raoult D 2007 Interactions between Mycobacterium xenopi amoeba and human cells J Hosp Infect 65 138ndash142
Drewe JA OrsquoConnor HM Weber N McDonald RA Delahay RJ 2013 Patterns of direct and indirect contact between cattle and badgers naturally infected with tuberculosis Epidemiol Infect 141 1467ndash1475
Drewe JA Tomlinson AJ Walker NJ Delahay RJ 2010 Diagnostic accuracy and optimal use of three tests for tuberculosis in live badgers PLoS One 5 e11196
Duffield BJ Young DA 1985 Survival of Mycobacterium bovis in defined environmental conditions Vet Microbiol 10 193ndash197
Dutau G 2005 Petite histoire illustreacutee de la tuberculose The history of tuberculosis Arch Pediatr 12 88ndash95
Dziadek J Sajduda A Boruń M 2001 Specificity of insertion sequence-based PCR assays for Mycobacterium tuberculosis complex Int J Tuberc Lung Dis 5 569ndash574
EFSA 2014 The European Union summary report on trends ans sources of zoonotic agents and food-borne outbreaks in 2012 EFSA J 12 169
El Helou G Viola GM Hachem R Han XY Raad II 2013 Rapidly growing mycobacterial bloodstream infections Lancet Infect Dis 13 166ndash174
El Kheacutechine A Couderc C Flaudrops C Raoult D Drancourt M 2011 Matrix-assisted laser desorptionionization time-of-flight mass spectrometry identification of mycobacteria in routine clinical practice PLoS One 6 e24720
El Kheacutechine A Henry M Raoult D Drancourt M 2009 Detection of Mycobacterium tuberculosis complex organisms in the stools of patients with pulmonary tuberculosis Microbiology 155 2384ndash2389
Elliott GN Hough RL Avery LM Maltin C A Campbell CD 2014 Environmental risk factors in the incidence of Johnersquos disease Crit Rev Microbiol 7828 1ndash20
Evans JT Smith EG Banerjee A Smith RMM Dale J Innes JA Hunt D Tweddell A Wood A Anderson C Hewinson RG Smith NH Hawkey PM Sonnenberg P 2007 Cluster of human tuberculosis caused by Mycobacterium bovis evidence for person-to-person transmission in the UK Lancet 369 1270ndash1276
Reacutefeacuterences bibliographiques
200
Fabre M Koeck JL Le Flegraveche P Simon F Herveacute V Vergnaud G Pourcel C 2004 High genetic diversity revealed by variable-number tandem repeat genotyping and analysis of hsp65 gene polymorphism in a large collection of ldquoMycobacterium canettiirdquo strains indicates that the M tuberculosis complex is a recently emerged clone of M can J Clin Microbiol 42 3248ndash3255
Falkinham JO 2016 Nontuberculous mycobacteria community and nosocomial waterborne opportunistic pathogens Clin Microbiol Newsl 38 1ndash7
Falkinham JO Norton CD Mark W 2001 Factors influencing numbers of Mycobacterium avium Mycobacterium intracellulare and other Mycobacteria in drinking water distribution systems Appl Environ Microbiol 67 1225ndash1231
Fanning A Edwards S 1991 Mycobacterium bovis infection in human beings in contact with elk (Cervus elaphus) in Alberta Canada Lancet 338 1253ndash1255
Fediaevsky A Beacutenet J-J Boschiroli ML Riviegravere J Hars J 2012 La tuberculose bovine en France en 2011 poursuite de la reacuteduction du nombre de foyers Bull eacutepideacutemiologique santeacute Anim Aliment Novembre 4ndash12
Fediaevsky A Courcoul A Boschiroli ML Reveillaud E 2014 Tuberculose bovine en France reacutesultats drsquoune strateacutegie plus offensive Bull eacutepideacutemiologique santeacute Anim Aliment 64 4ndash11
Fediaevsky A Courcoul A Boschiroli ML Reveillaud E 2013 Tuberculose bovine en France des signaux favorables mais une situation toujours complexe dans certaines zones Bull Epideacutemiologique - Santeacute Anim Aliment 59 4ndash10
Fetene T Kebede N Alem G 2011 Tuberculosis infection in animal and human populations in three districts of Western Gojam Ethiopia Zoonoses Public Health 58 47ndash53
Fifis T Corner LAL Rothel JS Wood PR 1994 Cellular and humoral immune responses of cattle to purified Mycobacterium bovis antigens Scand J Immunol 39 267ndash274
Fine AE Bolin CA Gardiner JC Kaneene JB 2011a A study of the persistence of Mycobacterium bovis in the environment under natural weather conditions in Michigan USA Vet Med Int 2011 ID 765430 1-12
Fine AE OrsquoBrien DJ Winterstein SR Kaneene JB 2011b An effort to isolate Mycobacterium bovis from environmental substrates during investigations of bovine tuberculosis transmission sites (cattle farms and wildlife areas) in Michigan USA ISRN Vet Sci 2011 ID 787181 1-11
Fischer OA Matlova L Bartl J Dvorska L Svastova P Du Maine R Melicharek I Bartos M Pavlik I 2003 Earthworms (Oligochaeta Lumbricidae) and mycobacteria Vet Microbiol 91 325ndash338
Flores Valdez MA Schoolnik GK 2010 DosR-regulon genes induction in Mycobacterium bovis BCG under aerobic conditions Tuberculosis (Edinb) 90 197ndash200
Forrellad M Klepp L Gioffreacute A 2013 Virulence factors of the Mycobacterium tuberculosis complex Virulence 4 3ndash66
Frank W Reisinger EC Brandt-Hamerla W Schwede I Handrick W 2009 Mycobacterium microti--pulmonary tuberculosis in an immunocompetent patient Wien Klin Wochenschr 121 282ndash6
Frottier J Eliaszewicz M Arlet V Gaudillat C 1990 Infections caused by Mycobacterium africanum Bull Acad Natl Med 174 29ndash35
Gallagher J Clifton-Hadley RS 2000 Tuberculosis in badgers a review of the disease and its significance for other animals Res Vet Sci 69 203ndash217
Reacutefeacuterences bibliographiques
201
Gallagher J Horwill DM 1977 A selective oleic acid albumin agar medium for the cultivation of Mycobacterium bovis J Hyg (Lond) 79 155ndash160
Gallagher J Monies R Gavier-Widen M Rule B 1998 Role of infected non-diseased badgers in the pathogenesis of tuberculosis in the badger Vet Rec 142 710ndash714
Garbaccio S Macias A Shimizu E Paolicchi F Pezzone N Magnano G Zapata L Abdala A Tarabla H Peyru M Caimi K Zumaacuterraga M Canal A Cataldi A 2014 Association between spoligotype-VNTR types and virulence of Mycobacterium bovis in cattle Virulence 5 297ndash302
Garbaccio SG Cataldi A 2010 Evaluation of an immunomagnetic capture method followed by PCR to detect Mycobacterium bovis in tissue samples from cattle Rev Argent Microbiol 42 247ndash53
Garnett BT Delahay RJ Roper TJ 2002 Use of cattle farm resources by badgers (Meles meles) and risk of bovine tuberculosis (Mycobacterium bovis) transmission to cattle Proc Biol Sci 269 1487ndash1491
Garnier T Eiglmeier K Camus J-C Medina N Mansoor H Pryor M Duthoy S Grondin S Lacroix C Monsempe C Simon S Harris B Atkin R Doggett J Mayes R Keating L Wheeler PR Parkhill J Barrell BG Cole ST Gordon S V Hewinson RG 2003 The complete genome sequence of Mycobacterium bovis Proc Natl Acad Sci U S A 100 7877ndash7882
Gavier-Widen D Chambers MA Palmer N Newell DG Hewinson RG 2001 Pathology of natural Mycobacterium bovis infection in European badgers (Meles meles) and its relationship with bacterial excretion Vet Rec 148 299ndash304
Gavier-Wideacuten D Cooke MM Gallagher J Chambers MA Gortaacutezar C 2009 A review of infection of wildlife hosts with Mycobacterium bovis and the diagnostic difficulties of the ldquono visible lesionrdquo presentation N Z Vet J 57 122ndash131
Genov I 1965 The effects of certain physical and chemical agents on Mycobacterium tuberculosis Veteriaacuternı Med Nauk Sofia 2 97ndash107
Gerasimova A Kazakov AE Arkin AP Dubchak I Gelfand MS 2011 Comparative genomics of the dormancy regulons in mycobacteria J Bacteriol 193 3446ndash3452
Ghodbane R Mba Medie F Lepidi H Nappez C Drancourt M 2014 Long-term survival of tuberculosis complex mycobacteria in soil Microbiology 160 496ndash501
Ghosh J Larsson P Singh B Pettersson BMF Islam NM Sarkar SN Dasgupta S Kirsebom LA 2009 Sporulation in mycobacteria Proc Natl Acad Sci U S A 106 10781ndash10786
Goodchild A V Clifton-Hadley RS 2001 Cattle-to-cattle transmission of Mycobacterium bovis Tuberculosis (Edinb) 81 23ndash41
Gordon S V Heym B Parkhill J Barrell B Cole ST 1999 New insertion sequences and a novel repeated sequence in the genome of Mycobacterium tuberculosis H37Rv Microbiology 145 Pt 4 881ndash892
Gormley E 1999 Detection of Mycobacterium bovis lymphocyte stimulating antigens in culture filtrates of a recombinant Mycobacterium smegmatis cosmid library Vaccine 17 2792ndash2801
Gormley E Doyle MB Fitzsimons T McGill K Collins JD 2006 Diagnosis of Mycobacterium bovis infection in cattle by use of the gamma-interferon (Bovigam) assay Vet Microbiol 112 171ndash179
Gortaacutezar C Delahay RJ McDonald RA Boadella M Wilson GJ Gavier-Wideacuten D Acevedo P 2012 The status of tuberculosis in European wild mammals Mamm Rev 42 193ndash206
Gortaacutezar C Torres MJ Vicente J Acevedo P Reglero M de la Fuente J Negro JJ Aznar-Martiacuten J
Reacutefeacuterences bibliographiques
202
2008 Bovine tuberculosis in Dontildeana Biosphere Reserve The role of wild ungulates as disease reservoirs in the last Iberian lynx strongholds PLoS One 3(7) e2776
Grange JM 2001 Mycobacterium bovis infection in human beings Tuberculosis (Edinb) 81 71ndash77
Grange JM Yates MD 1994 Zoonotic aspects of Mycobacterium bovis infection Vet Microbiol 40 137ndash151
Grange JM Yates MD 1989 Incidence and nature of human tuberculosis due to Mycobacterium africanum in South-East England 1977-1987 Epidemiol Infect 103 127ndash132
Greub G Raoult D 2004 Microorganisms resistant to free-living amoebae Clin Microbiol Rev 17 413ndash433
Griffin JFT Rodgers CR Liggett S Mackintosh CG 2006 Tuberculosis in ruminants Characteristics of intra-tonsilar Mycobacterium bovis infection models in cattle and deer Tuberculosis 86 404ndash418
Griffin JM Martin SW Thorburn MA Eves JA Hammond RF 1996 A case-control study on the association of selected risk factors with the occurrence of bovine tuberculosis in the Republic of Ireland Prev Vet Med 27 217ndash229
Guillet-Caruba C Martinez V Doucet-Populaire F 2014 The new tools of microbiological diagnosis of tuberculosis La Rev meacutedecine interne 35 794ndash800
Gunn-Moore DA McFarland SE Brewer JI Crawshaw TR Clifton-Hadley RS Kovalik M Shaw DJ 2011 Mycobacterial disease in cats in Great Britain I Culture results geographical distribution and clinical presentation of 339 cases J Feline Med Surg 13 934ndash944
Gunn-Moore DA Jenkins PA Lucke VM 1996 Feline tuberculosis a literature review and discussion of 19 cases caused by an unusual mycobacterial variant Vet Rec 138 53ndash58
Gutierrez MC Brisse S Brosch R Fabre M Omaiumls B Marmiesse M Supply P Vincent V 2005 Ancient origin and gene mosaicism of the progenitor of Mycobacterium tuberculosis PLoS Pathog 1(1) e5
Haddad N Ostyn A Karoui C Masselot M Thorel MF Hughes SL Inwald J Hewinson RG Durand B 2001 Spoligotype diversity of Mycobacterium bovis strains isolated in France from 1979 to 2000 J Clin Microbiol 39 3623ndash3632
Harris NB Payeur J Bravo D Osorio R Stuber T Farrell D Paulson D Treviso S Mikolon A Rodriguez-Lainz A Cernek-Hoskins S Rast R Ginsberg M Kinde H 2007 Recovery of Mycobacterium bovis from soft fresh cheese originating in Mexico Appl Environ Microbiol 73 1025ndash1028
Hars J Richomme C Boschiroli ML 2010 La tuberculose bovine dans la faune sauvage en France Bull Epideacutemiologique - Santeacute Anim Aliment 38 28ndash31
Hauer A Cruz K De Cochard T Godreuil S Karaoui C Henault S Bulach T Banuls A Biet F Boschiroli ML 2015 Genetic evolution of Mycobacterium bovis causing tuberculosis in livestock and wildlife in France since 1978 PLoS One 10(2) e 0117103
Haydon DT Cleaveland S Taylor LH Laurenson MK 2002 Identifying reservoirs of infection A conceptual and practical challenge Emerg Infect Dis 8 1468ndash1473
Hennessee CT Seo JS Alvarez AM Li QX 2009 Polycyclic aromatic hydrocarbon-degrading species isolated from Hawaiian soils Mycobacterium crocinum sp nov Mycobacterium pallens sp nov Mycobacterium rutilum sp nov Mycobacterium rufum sp nov and Mycobacterium aromaticivorans sp nov Int J Syst Evol Microbiol 59 378ndash387
Hines N Payeur JB Hoffman LJ 2006 Comparison of the recovery of Mycobacterium bovis isolates using
Reacutefeacuterences bibliographiques
203
the BACTEC MGIT 960 system BACTEC 460 system and Middlebrook 7H10 and 7H11 solid media J Vet Diagn Invest 18 243ndash250
Hmama Z Pena-Diaz S Sunil J Av-Gay Y 2015 Immunoevasion and immunosuppression of the macrophage by Mycobacterium tuberculosis Immunol Rev 264 220ndash232
Hoefsloot W Van Ingen J Andrejak C Aumlngeby K Bauriaud R Bemer P Beylis N Boeree MJ Cacho J Chihota V Chimara E Churchyard G Cias R Daza R Daley CL Dekhuijzen PNR Domingo D Drobniewski F Esteban J Fauville-Dufaux M Folkvardsen DB Gibbons N Goacutemez-Mampaso E Gonzalez R Hoffmann H Hsueh PR Indra A Jagielski T Jamieson F Jankovic M Jong E Keane J Koh WJ Lange B Leao S Macedo R Mannsaringker T Marras TK Maugein J Milburn HJ Mlinkoacute T Morcillo N Morimoto K Papaventsis D Palenque E Paez-Pentildea M Piersimoni C Polanovaacute M Rastogi N Richter E Ruiz-Serrano MJ Silva A Da Silva MP Simsek H Van Soolingen D Szaboacute N Thomson R Fernandez TT Tortoli E Totten SE Tyrrell G Vasankari T Villar M Walkiewicz R Winthrop KL Wagner D 2013 The geographic diversity of nontuberculous mycobacteria isolated from pulmonary samples An NTM-NET collaborative study Eur Respir J 42 1604ndash1613
Hohn A Lowenstein 1926 Die Kulture des tuberklebacillus zur diagnose der tuberculose Zeut fur BaktI Orig 98 460ndash477
Hongmanee P Stender H Rasmussen OF 2001 Evaluation of a fluorescence in situ hybridization assay for differentiation between tuberculous and nontuberculous Mycobacterium species in smears of Lowenstein-Jensen and Mycobacteria Growth Indicator Tube cultures using peptide nucleic acid prob J Clin Microbiol 39 1032ndash1035
Hormisch D Brost I Kohring G Giffhorn F Kroppenstedt RM Stackebrandt E Faumlrber P 2004 Mycobacterium fluoranthenivorans sp nov a Fluoranthene and Aflatoxin B 1 Degrading Bacterium from Contaminated Soil of a Former Coal Gas Plant Syst Appl Microbiol 27 653ndash660
Huard RC Claudio L Lazzarini DO Butler WR Soolingen D Van Ho JL 2003 PCR-based method to differentiate the subspecies of the Mycobacterium tuberculosis complex on the basis of genomic deletions J Clin Microbiol 41 1637ndash1650
Huard RC Fabre M de Haas P Lazzarini LCO van Soolingen D Cousins D Ho JL 2006 Novel genetic polymorphisms that further delineate the phylogeny of the Mycobacterium tuberculosis complex J Bacteriol 188 4271ndash4287
Humblet MF Boschiroli ML Saegerman C 2009 Classification of worldwide bovine tuberculosis risk factors in cattle A stratified approach Vet Res 40(5) 1-24
Hutchings MR Harris S 1999 Quantifying the risks of TB infection to cattle posed by badger excreta Epidemiol Infect 122 167ndash173
Hutchings MR Harris S 1997 Effects of farm management practices on cattle grazing behaviour and the potential for transmission of bovine tuberculosis from badgers to cattle Vet J 153 149ndash162
Hutchings MR Service KM Harris S 2002 Is population density correlated with faecal and urine scent marking in European badgers (Meles meles) in the UK Mamm Biol 67 286ndash293
Hutchings MR Service KM Harris S 2001 Defecation and urination patterns of badgers Meles meles at low density in south west England- Acta Theriol (Warsz) 46 87ndash96
Huws SA McBain AJ Gilbert P 2005 Protozoan grazing and its impact upon population dynamics in biofilm communities J Appl Microbiol 98 238ndash244
Idigoras P Peacuterez-Trallero E Alcorta M Gutieacuterrez C Muntildeoz-Baroja I 1995 Rapid detection of tuberculous
Reacutefeacuterences bibliographiques
204
and non-tuberculous mycobacteria by microscopic observation of growth on Middlebrook 7H11 agar Eur J Clin Microbiol Infect Dis 14 6ndash10
Iivanainen E Martikainen PJ Vaumlaumlnaumlnen PK Vaananen PK Katila M 1999 Environmental factors affecting the occurrence of mycobacteria in brook waters J Appl Microbiol 86 673ndash681
Jackson R De Lisle GW Morris RS 1995 A study of the environmental survival of Mycobacterium bovis on a farm in New Zealand N Z Vet J 43 346ndash352
Jahans K Palmer S Inwald J Brown J Abayakoon S 2004 Isolation of Mycobacterium microti from a male Charolais-Hereford cross Vet Rec 155 373ndash374
Jalava K Jones JA Goodchild T Clifton-Hadley R Mitchell A Story A Watson JM 2007 No increase in human cases of Mycobacterium bovis disease despite resurgence of infections in cattle in the United Kingdom Epidemiol Infect 135 40ndash45
Jenkins AO Cadmus SI Venter EH Pourcel C Hauk Y Vergnaud G Godfroid J 2011 Molecular epidemiology of human and animal tuberculosis in Ibadan Southwestern Nigeria Vet Microbiol 151 139ndash147
Jenkins HE Morrison WI Cox DR Donnelly CA Johnston WT Bourne FJ Clifton-Hadley RS Gettinby G McInerney JP Watkins GH Woodroffe R 2008 The prevalence distribution and severity of detectable pathological lesions in badgers naturally infected with Mycobacterium bovis Epidemiol Infect 136 1350ndash1361
Jensen KE 1954 Presence and destruction of tubercle bacilli in sewage Bull World Health Organ 10 171ndash179
Jimeacutenez BA Hinojoza-Loza E Flores-Valdez MA Prado-Montes de Oca E Allen K Estrada-Chaacutevez C Herrera-Rodriacuteguez S Flores-Fernaacutendez JM Martiacutenez-Velaacutezquez M Hernaacutendez-Gutieacuterrez R Alvarez AH 2013 Expression of non-replicating persistence associated genes of Mycobacterium bovis in lymph nodes from skin test-reactor cattle Microb Pathog 61-62 23ndash28
Johnson L Dean G Rhodes S Hewinson G Vordermeier M Wangoo A 2007 Low-dose Mycobacterium bovis infection in cattle results in pathology indistinguishable from that of high-dose infection Tuberculosis (Edinb) 87 71ndash76
Kaal E Kolk AHJ Kuijper S Janssen H-G 2009 A fast method for the identification of Mycobacterium tuberculosis in sputum and cultures based on thermally assisted hydrolysis and methylation followed by gas chromatography-mass spectrometry J Chromatogr A 1216 6319ndash6325
Kamerbeek J Schouls L Kolk A van Agterveld M van Soolingen D Kuijper S Bunschoten A Molhuizen H Shaw R Goyal M van Embden J 1997 Simultaneous detection and strain differentiation of Mycobacterium tuberculosis for diagnosis and epidemiology J Clin Microbiol 35 907ndash914
Kaprelyants AS Gottschal JC Kell DB 1993 Dormancy in non-sporulating bacteria FEMS Microbiol Rev 104 271ndash285
Kasai H Ezaki T Harayama S 2000 Differentiation of phylogenetically related slowly growing mycobacteria by their gyrB sequences J Clin Microbiol 38(1) 301
Kazda J Pavlik I Falkinham JO Hruska K 2009 The Ecology of Mycobacteria Impact on Animalrsquos and Humans Health Springer Netherlands Dordrecht
Khan NA 2006 Acanthamoeba biology and increasing importance in human health FEMS Microbiol Rev 30 564ndash595
Kiers A Klarenbeek A Mendelts B Van Soolingen D Koeumlter G 2008 Transmission of Mycobacterium
Reacutefeacuterences bibliographiques
205
pinnipedii to humans in a zoo with marine mammals Int J Tuberc Lung Dis 12 1469ndash1473
King HC Murphy A James P Travis E Porter D Hung Y-J Sawyer J Cork J Delahay RJ Gaze W Courtenay O Wellington EM 2015a The variability and seasonality of the environmental reservoir of Mycobacterium bovis shed by wild European badgers Sci Rep 5 12318
King HC Murphy A James P Travis E Porter D Sawyer J Cork J Delahay RJ Gaze W Courtenay O Wellington EM 2015b Performance of a non-invasive test for detecting Mycobacterium bovis shedding in European badger (Meles meles) populations J Clin Microbiol 53(7) 2316-2323
Kirchheimer WF Storrs EE 1971 Attempts to establish the armadillo (Dasypus novemcinctus Linn) as a model for the study of leprosy I Report of lepromatoid leprosy in an experimentally infected armadillo Int J Lepr Other Mycobact Dis 39 693ndash702
Kirschner P Springer B Vogel U Meier A Wrede A Kiekenbeck M Bange F Bottger EC 1993 Genotypic identification of Mycobacteria by nucleic acid sequence determination report of a 2-year experience in a clinical laboratory J Clin Microbiol 31 2882ndash2889
Kleeberg HH 1984 Tuberculose humaine drsquoorigine bovine et santeacute publique Rev Sci Tech 3 33ndash54
Koeck JL Fabre M Simon F Daffeacute M Garnotel Eacute Matan A B Geacuterocircme P Bernatas JJ Buisson Y Pourcel C 2011 Clinical characteristics of the smooth tubercle bacilli ldquoMycobacterium canettiirdquo infection suggest the existence of an environmental reservoir Clin Microbiol Infect 17 1013ndash1019
Krasnow I Wayne LG 1966 Sputum Digestion I The mortality rate of tubercle bacilli in various digestion systems Am J Clin Pathol 45 352ndash355
Krishna-Prasad BN Gupta SK 1978 Preliminary report on engulfment and retention of mycobacteria by trophozoites of axenically grown Acanthamoeba castellanii Douglas Curr Sci 47 245ndash247
Kruuk H 1978 Foraging and spatial organisation of the European badger Meles meles L Behav Ecol Sociobiol 4 75ndash89
Kubica GP Dye WE Cohn ML Middlebrook GSD and D with N-A-L-CH for C of M 1963 Sputum digestion and decontamination with N-acetyl-L-cysteinemdashsodium hydroxide for culture of mycobacteria Am Rev Respir Dis 87 775ndash779
Kubica T Rusch-Gerdes S Niemann S 2003 Mycobacterium bovis subsp caprae Caused One-Third of Human M bovis-Associated Tuberculosis Cases Reported in Germany between 1999 and 2001 J Clin Microbiol 41 3070ndash3077
Kukielka E Barasona JA Cowie CE Drewe J a Gortaacutezar C Cotarelo I Vicente J 2013 Spatial and temporal interactions between livestock and wildlife in South Central Spain assessed by camera traps Prev Vet Med 112 213ndash221
Kulka K Hatfull G Ojha AK 2012 Growth of Mycobacterium tuberculosis biofilms J Vis Exp (60) e3820
Lamont EA Bannantine JP Armieacuten A Ariyakumar DS Sreevatsan S 2012 Identification and characterization of a spore-like morphotype in chronically starved Mycobacterium avium subsp paratuberculosis cultures PLoS One 7 e30648
Laprie C Duboy J Malik R Fyfe J 2013 Feline cutaneous mycobacteriosis a review of clinical pathological and molecular characterization of one case of Mycobacterium microti skin infection and nine cases of feline leprosy syndrome from France and New Caledonia Vet Dermatol 24(6) 561ndash9 e133ndash4
Lavania M Katoch K Sachan P Dubey A Kapoor S Kashyap M Chauhan DS Singh HB Sharma VD Jadhav RS Katoch VM 2006 Detection of Mycobacterium leprae DNA from soil samples by PCR
Reacutefeacuterences bibliographiques
206
targeting RLEP sequences J Commun Dis 38 269ndash273
Le Dantec C Duguet J-P Montiel A Dumoutier N Dubrou S Vincent V 2002 Occurrence of Mycobacteria in Water Treatment Lines and in Water Distribution Systems Appl Environ Microbiol 68 5318ndash5325
Lee H Park H Cho S Bai G Kim S 2000 Species identification of Mycobacteria by PCR-Restriction Fragment Length Polymorphism of the rpoB gene J Clin Microbiol 38 2966ndash2971
Lefmann M Schweickert B Buchholz P Goumlbel UB Ulrichs T Seiler P Theegarten D Moter A 2006 Evaluation of peptide nucleic acid-fluorescence in situ hybridization for identification of clinically relevant mycobacteria in clinical specimens and tissue sections J Clin Microbiol 44 3760ndash3767
Lehtola MJ Torvinen E Miettinen IT Keevil CW 2006 Fluorescence in situ hybridization using peptide nucleic acid probes for rapid detection of Mycobacterium avium subsp avium and Mycobacterium avium subsp paratuberculosis in potable-water biofilms Appl Environ Microbiol 72 848ndash853
Leistikow RL Morton RA Bartek IL Frimpong I Wagner K Voskuil MI 2010 The Mycobacterium tuberculosis dosR regulon assists in metabolic homeostasis and enables rapid recovery from nonrespiring dormancy J Bacteriol 192 1662ndash1670
Leys NM Ryngaert A Bastiaens L Wattiau P Top EM Verstraete W Springael D 2005 Occurrence and community composition of fast-growing Mycobacterium in soils contaminated with polycyclic aromatic hydrocarbons FEMS Microbiol Ecol 51 375ndash388
Li L Bannantine JP Zhang Q Amonsin A May BJ Alt D Banerji N Kanjilal S Kapur V 2005 The complete genome sequence of Mycobacterium avium subspecies paratuberculosis Proc Natl Acad Sci U S A 102 12344ndash12349
Libero G De Mori L 2014 The T-cell response to lipid antigens of Mycobacterium tuberculosis Front Immunol 5 1ndash9
Lieacutebana E Aranaz A Francis B Cousins D 1996 Assessment of genetic markers for species differentiation within the Mycobacterium tuberculosis complex J Clin Microbiol 34 933ndash938
Lieacutebana E Aranaz A Mateos A Vilafranca M Gomez-Mampaso E Tercero JC Alemany J Suarez G Domingo M Dominguez L 1995 Simple and rapid detection of Mycobacterium tuberculosis complex organisms in bovine tissue samples by PCR J Clin Microbiol 33 33ndash36
Lim A Eleuterio M Hutter B Murugasu-Oei B Dick T 1999 Oxygen depletion-induced dormancy in Mycobacterium bovis BCG J Bacteriol 181 2252ndash2256
Liss GM Wong L Kittle DC Simor A Naus M Martiquet P Misener CR 1994 Occupational exposure to Mycobacterium bovis infection in deer and elk in Ontario Can J Public Heal 85 326ndash329
Little TW Naylor PF Wilesmith JW 1982 Laboratory study of Mycobacterium bovis infection in badgers and calves Vet Rec 111 550ndash557
Lomme JR Thoen CO Himes EM Vinson JW King RE 1976 Mycobacterium tuberculosis infection in two East African oryxes J Am Vet Med Assoc 169 912ndash914
Lotz A Ferroni A Beretti J-L Dauphin B Carbonnelle E Guet-Revillet H Veziris N Heym B Jarlier V Gaillard J-L Pierre-Audigier C Frapy E Berche P Nassif X Bille E 2010 Rapid identification of mycobacterial whole cells in solid and liquid culture media by matrix-assisted laser desorption ionization-time of flight mass spectrometry J Clin Microbiol 48 4481ndash4486
Ludwig W Euzeacuteby J Schumann P Busse H-J Trujillo ME Kaumlmpfer P Whitman WB 2008 Road map
Reacutefeacuterences bibliographiques
207
of the phylum Actinobacteria Road Map of the phylum Actinobacteria Bergeys Manual of Systematic Bacteriology second Edition Volume five Springer Editions
Lugton IW Wilson PR Morris RS Nugent G 1998 Epidemiology and pathogenesis of Mycobacterium bovis infection of red deer (Cervus elaphus) in New Zealand N Z Vet J 46 147ndash156
Lyashchenko KP Greenwald R Esfandiari J OrsquoBrien DJ Schmitt SM Palmer M V Waters WR 2013 Rapid detection of serum antibody by dual-path platform VetTB assay in white-tailed deer infected with Mycobacterium bovis Clin Vaccine Immunol 20 907ndash911
Mackintosh CG Qureshi T Waldrup K Labes RE Dodds KG Griffin JFT 2000 Genetic resistance to experimental infection with Mycobacterium bovis in red deer (Cervus elaphus) Infect Immun 68 1620ndash1625
Maddock EC 1936 Experiments on the infectivity for healthy calves of bovine tubercle bacilli discharged in dung upon pasture Part I From tubercular calves fed with emulsions of Tubercle bacilli 1934-5 Part II From tubercular cows passing Tubercle bacilli in their dun J Hyg (Lond) 36 594ndash601
Maddock EC 1934 Further studies on the survival time of the bovine tubercle bacillus in soil soil and dung in dung and on grass with experiments on feeding guinea-pigs and calves on grass artificially infected with bovine tubercle bacilli J Hyg (Lond) 34 372ndash379
Maddock EC 1933 Studies on the survival time of the bovine tubercle bacillus in soil soil and dung in dung and on grass with experiments on the preliminary treatment of infected organic matter and the cultivation of the organism J Hyg (Lond) 33 103ndash117
Magee JG Ward AC 2012 Mycobacteriaceae Bergeys Manual of Systematic Bacteriology second Edition Volume five Springer Editions
Mahairas GG Sabo PJ Hickey MJ Singh DC Stover CK Mahairas GG Sabo PJ Hickey MJ Singh DC Stover CK 1996 Molecular analysis of genetic differences between Mycobacterium bovis BCG and virulent M bovis J Bacteriol 178 1274ndash1282
Mardare C Delahay RJ Dale JW 2013 Environmental amoebae do not support the long-term survival of virulent mycobacteria J Appl Microbiol 114 1388ndash1394
Marrakchi H Laneacuteelle MA Daffeacute M 2014 Mycolic acids Structures biosynthesis and beyond Chem Biol 21 67ndash85
Marri PR Bannantine JP Golding GB 2006 Comparative genomics of metabolic pathways in Mycobacterium species gene duplication gene decay and lateral gene transfer FEMS Microbiol Rev 30 906ndash925
Martiacuten-Hernando MP Houmlfle U Vicente J Ruiz-Fons F Vidal D Barral M Garrido JM de la Fuente J Gort 2007 Lesions associated with Mycobacterium tuberculosis complex infection in the European wild boar Tuberculosis 87 360ndash367
Mathema B Kurepina NE Bifani PJ Kreiswirth BN 2006 Molecular epidemiology of tuberculosis Current insights Clin Microbiol Rev 19 658ndash685
Matsuoka M Izumi S Budiawan T Nakata N Saeki K 1999 Mycobacterium leprae DNA in daily using water as a possible source of leprosy infection Indian J Lepr 71 61ndash67
Mba Medie F Ben Salah I Henrissat B Raoult D Drancourt M 2011 Mycobacterium tuberculosis complex mycobacteria as amoeba-resistant organisms PLoS One 6 e20499
McCallan L Mcnair J Skuce R 2014 A review of the potential role of cattle slurry in the spread of bovine
Reacutefeacuterences bibliographiques
208
tuberculosis 1ndash48
McCorry T Whelan AO Welsh MD McNair J Walton E Bryson DG Hewinson RG Vordermeier HM Pollock JM 2005 Shedding of Mycobacterium bovis in the nasal mucus of cattle infected experimentally with tuberculosis by the intranasal and intratracheal routes Vet Rec 157 613ndash618
McLaughlin AM Gibbons N Fitzgibbon M Power JT Foley SC Hayes JP Rogers T Keane J 2012 Primary isoniazid resistance in Mycobacterium bovis disease A prospect of concern Am J Respir Crit Care Med 186 110ndash111
McNabb A Eisler D Adie K Amos M Rodrigues M Stephens G Black WA Isaac-Renton J 2004 Assessment of partial sequencing of the 65-kilodalton heat shock protein gene (hsp65) for routine identification of Mycobacterium species isolated from clinical sources J Clin Microbiol 42 3000ndash3011
Mejia GI Castrillon L Trujillo H Robledo J A 1999 Microcolony detection in 7H11 thin layer culture is an alternative for rapid diagnosis of Mycobacterium tuberculosis infection Int J Tuberc Lung Dis 3 138ndash142
Menzies FD Neill SD 2000 Cattle-to-Cattle Transmission of Bovine Tuberculosis Vet J 160 92ndash106
Meredith A Del Pozo J Smith S Milne E Stevenson K McLuckie J 2014 Leprosy in red squirrels in Scotland Vet Rec 175 285ndash286
Michel AL Muumlller B van Helden PD 2010 Mycobacterium bovis at the animal-human interface a problem or not Vet Microbiol 140 371ndash381
Michel AL Venter L Espie IW Coetzee ML 2003 Mycobacterium tuberculosis infections in eight species at the national zoological gardens of South Africa 1991-2001 J Zoo Wildl Med 34 364ndash370
Michelet L de Cruz K Zanella G Aaziz R Bulach T Karoui C Heacutenault S Joncour G Boschiroli ML 2015 Infection with Mycobacterium microti in animals in France J Clin Microbiol 53 981ndash985
Mignard S Pichat C Carret G 2006 Mycobacterium bovis infection Lyon France Emerg Infect Dis 12 1431ndash1433
Miltgen J Morillon M Koeck J-L Varnerot A Briant J-F Nguyen G Verrot D Bonnet D Vincent V 2002 Two cases of pulmonary tuberculosis caused by Mycobacterium tuberculosis subsp canetti Emerg Infect Dis 8 1350ndash1352
Miltner EC Bermudez LE 2000 Mycobacterium avium grown in Acanthamoeba castellanii is protected from the effects of antimicrobials Antimicrob Agents Chemother 44 1990ndash1994
Mitscherlich E Marth EH 2012 Microbial survival in the environment Bacteria and rickettsiae important in human and animal health Springer S ed
Momotani E Ozaki H Hori M Yamamoto S Kuribayashi T Eda S Ikegami M 2012 Mycobacterium avium subsp paratuberculosis lipophilic antigen causes Crohnrsquos disease-type necrotizing colitis in mice Springerplus 1 1ndash10
Moore J Roper T 2003 Temperature and humidity in badger Meles meles setts Mamm Rev 33 308ndash313
Morris RS Pfeiffer DU Jackson R 1994 The epidemiology of Mycobacterium bovis infections Vet Microbiol 40 153ndash177
Mostafa HM Kazda J Irgens LM Luesse HG 1995 Acid-fast bacilli from former leprosy regions in coastal Norway showing PCR positivity for Mycobacterium leprae Int J Lepr other Mycobact Dis 63 97ndash99
Mukhopadhyay S Nair S Ghosh S 2012 Pathogenesis in tuberculosis Transcriptomic approaches to
Reacutefeacuterences bibliographiques
209
unraveling virulence mechanisms and finding new drug targets FEMS Microbiol Rev 36 463ndash485
Muumlller B Duumlrr S Alonso S Hattendorf J Laisse CJM Parsons SDC van Helden PD Zinsstag J 2013 Zoonotic Mycobacterium bovis ndashinduced tuberculosis in humans Emerg Infect Dis 19 899ndash908
Muumlller B Hilty M Berg S Garcia-Pelayo MC Dale J Boschiroli ML Cadmus SI Ngandolo BNR Godreuil S Diguimbaye-Djaibeacute C Kazwala R Bonfoh B Njanpop-Lafourcade BM Sahraoui N Guetarni D Aseffa A Mekonnen MH Razanamparany VR Ramarokoto H Djoslashnne B Oloya J Machado A Mucavele C Skjerve E Portaels F Rigouts L Michel A Muumlller A Kaumlllenius G Van Helden PD Hewinson RG Zinsstag J Gordon S V Smith NH 2009 African 1 an epidemiologically important clonal complex of Mycobacterium bovis dominant in Mali Nigeria Cameroon and Chad J Bacteriol 191 1951ndash1960
Mura M Bull T 2006 Replication and long-term persistence of bovine and human strains of Mycobacterium avium subsp paratuberculosis within Acanthamoeba polyphaga Appl Environ Microbiol 72 854ndash859
Murphy D Gormley E Costello E OrsquoMeara D Corner LAL 2010 The prevalence and distribution of Mycobacterium bovis infection in European badgers (Meles meles) as determined by enhanced post mortem examination and bacteriological culture Res Vet Sci 88 1ndash5
Musser JM Amin A Ramaswamy S 2000 Negligible genetic diversity of Mycobacterium tuberculosis host immune system protein targets Evidence of limited selective pressure Genetics 155 7ndash16
Namouchi A Didelot X Schoumlck U Gicquel B Rocha EPC 2012 After the bottleneck Genome-wide diversification of the Mycobacterium tuberculosis complex by mutation recombination and natural selection Genome Res 22 721ndash734
Narang R Narang P Mendiratta DK 2009 Isolation and identification of nontuberculous mycobacteria from water and soil in central India Indian J Med Microbiol 27 247ndash250
Naranjo V Gortaacutezar C Vicente J de la Fuente J 2008 Evidence of the role of European wild boar as a reservoir of Mycobacterium tuberculosis complex Vet Microbiol 127 1ndash9
Navarro Y Romero B Bouza E Domiacutenguez L Juan L de Garciacutea-de-Viedma D 2015 Detailed chronological analysis of microevolution events in herds infected persistently by Mycobacterium bovis Vet Microbiol 183 97-102
Neill SD Hanna J OrsquoBrien J McCracken R 1988 Excretion of Mycobacterium bovis by experimentally infected cattle Vet Rec 123 340ndash343
Neill SD Pollock JM Bryson DB Hanna J 1994 Pathogenesis of Mycobacterium bovis infection in cattle Vet Microbiol 40 41ndash52
Niemann S Richter E Daluumlgge-Tamm H Schlesinger H Graupner D Koumlnigstein B Gurath G Greinert U Ruumlsch-Gerdes S 2000 Two cases of Mycobacterium microti-derived tuberculosis in HIV-negative immunocompetent patients Emerg Infect Dis 6 539ndash542
Nishi JS Shury T Elkin BT 2006 Wildlife reservoirs for bovine tuberculosis (Mycobacterium bovis) in Canada Strategies for management and research Vet Microbiol 112 325ndash338
Norby B Fosgate GT Manning EJB Collins MT Roussel AJ 2007 Environmental mycobacteria in soil and water on beef ranches Association between presence of cultivable mycobacteria and soil and water physicochemical characteristics Vet Microbiol 124 153ndash159
Nugent G 2011 Maintenance spillover and spillback transmission of bovine tuberculosis in multi-host wildlife complexes a New Zealand case study Vet Microbiol 151 34ndash42
Reacutefeacuterences bibliographiques
210
OrsquoBrien DJ Schmitt SM Fitzgerald SD Berry DE Hickling GJ 2006 Managing the wildlife reservoir of Mycobacterium bovis the Michigan USA experience Vet Microbiol 112 313ndash323
OrsquoReilly LM Daborn CJ 1995 The epidemiology of Mycobacterium bovis infections in animals and man a review Tuber Lung Dis 76 Suppl 1 1ndash46
OIE 2009 Bovine Tuberculosis Terrestrial Manual 2009Chap 247
Ojha AK Baughn AD Sambandan D Hsu T Trivelli X Guerardel Y Alahari A Kremer L Jacobs WR Hatfull GF 2008 Growth of Mycobacterium tuberculosis biofilms containing free mycolic acids and harbouring drug-tolerant bacteria Mol Microbiol 69 164ndash174
Oloya J Kazwala R Lund A Opuda-Asibo J Demelash B Skjerve E Johansen TB Djoslashnne B 2007 Characterisation of mycobacteria isolated from slaughter cattle in pastoral regions of Uganda BMC Microbiol 7 95
Otal I Isabel O Gomez AB Kremer K de Haas P Garciacutea MJ Martiacuten C van Soolingen D 2008 Mapping of IS6110 insertion sites in Mycobacterium bovis isolates in relation to adaptation from the animal to human host Vet Microbiol 129 333ndash341
Pagnier I Raoult D La Scola B 2008 Isolation and identification of amoeba-resisting bacteria from water in human environment by using an Acanthamoeba polyphaga co-culture procedure Environ Microbiol 10 1135ndash1144
Palgrave CJ Benato L Eatwell K Laurenson IF Smith NH 2012 Mycobacterium microti infection in two meerkats (Suricata suricatta) J Comp Pathol 146 278ndash282
Palmer M V 2013 Mycobacterium bovis Characteristics of wildlife reservoir hosts Transbound Emerg Dis 60 1ndash13
Palmer M V C Thacker T Waters WR Robbe-Austerman S Harris BN 2010 Investigations on deer to deer and deer to cattle transmission of the vaccine Mycobacterium bovis bacillus Calmette-Gueacuterin (BCG) J Vaccines Vaccin 01 1ndash5
Palmer M V Waters WR Whipple DL 2004 Investigation of the transmission of Mycobacterium bovis from deer to cattle through indirect contact Am J Vet Res 65 1483ndash1489
Palmer M V Waters WR Whipple DL 2002 Lesion development in white-tailed Deer (Odocoileus virginianus) experimentally infected with Mycobacterium bovis Vet Pathol 39 334ndash340
Palmer M V Whipple DL 2006 Survival of Mycobacterium bovis on feedstuffs commonly used as supplemental feed for white-tailed deer (Odocoileus virginianus) J Wildl Dis 42 853ndash8
Palmer M V Whipple DL Rhyan JC Bolin CA Saari DA 1999 Granuloma development in cattle after intratonsilar inoculation with Mycobacterium bovis Am J Vet Res 60 310ndash315
Palmer M V Whipple DL Waters WR 2001 Experimental deer-to-deer transmission of Mycobacterium bovis Am J Vet Res 62 692ndash696
Panteix G Gutierrez MC Boschiroli ML Rouviere M Plaidy A Pressac D Porcheret H Chyderiotis G Ponsada M Van Oortegem K Salloum S Cabuzel S Bantildeuls AL Van de Perre P Godreuil S 2010 Pulmonary tuberculosis due to Mycobacterium microti a study of six recent cases in France J Med Microbiol 59 984ndash989
Parsons SDC Drewe JA van Pittius NCG Warren RM van Helden PD 2013 Novel cause of tuberculosis in meerkats South Africa Emerg Infect Dis 19 2004ndash2007
Reacutefeacuterences bibliographiques
211
Pavlik I Svastova P Bartl J Dvorska L Rychlik I 2000 Relationship between IS901 in the Mycobacterium avium complex strains isolated from birds animals humans and the environment and virulence for poultry Clin Diagn Lab Immunol 7 212ndash217
Pavlik I Trcka I Parmova I Svobodova J Melicharek I Nagy G Cvetnic Z Ocepek M Pate M Lipiec M 2005 Detection of bovine and human tuberculosis in cattle and other animals in six Central European countries during the years 2000 ndash 2004 Vet Med-Czech 2005 291ndash299
Pawlowski A Jansson M Skoumlld M Rottenberg ME Kaumlllenius G 2012 Tuberculosis and HIV co-infection PLoS Pathog 8(2) e1002464
Payne A 2014 Rocircle de la faune sauvage dans le systegraveme multi-hocirctes de Mycobacterium bovis et risque de transmission entre faune sauvage et bovins Etude expeacuterimentale en Cocircte drsquoOr Thegravese Universiteacute Lyon
Payne A Boschiroli ML Gueneau E Moyen JL Rambaud T Dufour B Gilot-Fromont E Hars J 2012 Bovine tuberculosis in ldquoEurasianrdquo badgers (Meles meles) in France Eur J Wildl Res 59 331ndash339
Payne A Chappa S Hars J Dufour B Gilot-Fromont E 2015 Wildlife visits to farm facilities assessed by camera traps in a bovine tuberculosis-infected area in France Eur J Wildl Res 62(1) 33-42
Pepperell CS Casto AM Kitchen A Granka JM Cornejo OE Holmes EC Holmes EC Birren B Galagan J Feldman MW 2013 The role of selection in shaping diversity of natural M tuberculosis populations PLoS Pathog 9 e1003543
Petroff SA 1915 Some cultural studies on the tubercle bacillus Bull Johns Hopkins Hosp 26 276ndash279
Pfyffer GE Auckenthaler R Van Embden JD a Van Soolingen D 1998 Mycobacterium canettii the smooth variant of M tuberculosis isolated from a Swiss patient exposed in Africa Emerg Infect Dis 4 631ndash634
Phillips CJC Foster CRW Morris PA Teverson R 2003 The transmission of Mycobacterium bovis infection to cattle Res Vet Sci 74 1ndash15
Pickup RW Rhodes G Bull TJ Arnott S Sidi-Boumedine K Hurley M Hermon-Taylor J 2006 Mycobacterium avium subsp paratuberculosis in lake catchments in river water abstracted for domestic use and in effluent from domestic sewage treatment works diverse opportunities for environmental cycling and human exposure Appl Environ Microbiol 72 4067ndash4077
Piersimoni C Olivieri A Benacchio L Scarparo C 2006 Current perspectives on drug susceptibility testing of Mycobacterium tuberculosis complex the automated nonradiometric systems J Clin Microbiol 44 20ndash28
Pillai SD Widmer KW Ivey LJ Coker KC Newman E Lingsweiler S Baca D Kelley M Davis DS Silvy NJ Adams LG 2000 Failure to identify non-bovine reservoirs of Mycobacterium bovis in a region with a history of infected dairy-cattle herds Prev Vet Med 43 53ndash62
Pollock JM Neill SD 2002 Mycobacterium bovis infection and tuberculosis in cattle Vet J 163 115ndash127
Pontiroli A Travis ER Sweeney FP Porter D Gaze WH Mason S Hibberd V Holden J Courtenay O Wellington EMH 2011 Pathogen quantitation in complex matrices a multi-operator comparison of DNA extraction methods with a novel assessment of PCR inhibition PLoS One 6 e17916
Prasad HK Singhal A Mishra A Shah NP Katoch VM Thakral SS Singh D V Chumber S Bal S Aggarwal S Padma M V Kumar S Singh MK Acharya SK 2005 Bovine tuberculosis in India potential basis for zoonosis Tuberculosis (Edinb) 85 421ndash428
Primm TP Lucero CA Falkinham JO 2004 Health impacts of environmental mycobacteria Clin Microbiol Rev 17 98ndash106
Reacutefeacuterences bibliographiques
212
Pritchard DG 1988 A Century of Bovine Tuberculosis 1888-1988 Conquest and Controversy J Comp Pathol 99 357ndash399
Prodinger WM Brandsta A Naumann L Pacciarini M Kubica T Boschiroli ML Aranaz A Cvetnic Z Ocepek M Skrypnyk A Erler W Niemann S Pavlik I Moser I 2005 Characterization of Mycobacterium caprae isolates from Europe by Mycobacterial Interspersed Repetitive Unit Genotyping J Clin Microbiol 43 4984ndash4992
Prodinger WM Eigentler A Allerberger F Schonbauer M Glawischnig W 2002 Infection of Red Deer Cattle and Humans with Mycobacterium bovis subsp caprae in Western Austria J Clin Microbiol 40 2270ndash2272
Prodinger WM Indra A Koksalan OK Kilicaslan Z Richter E 2014 Mycobacterium caprae infection in humans Expert Rev Anti Infect Ther12(12) 1501-1513
Pym AS Brodin P Brosch R Huerre M Cole ST 2002 Loss of RD1 contributed to the attenuation of the live tuberculosis vaccines Mycobacterium bovis BCG and Mycobacterium microti Mol Microbiol 46 709ndash717
Radomski N Betelli L Moilleron R Haenn S Moulin L Cambau E Rocher V Gonccedilalves A Lucas FS 2011 Mycobacterium behavior in wastewater treatment plant a bacterial model distinct from Escherichia coli and Enterococci Environ Sci Technol 45 5380ndash5386
Radomski N Roguet A Lucas FS Veyrier FJ Cambau E Accrombessi H Moilleron R Behr M Moulin L 2013 atpE gene as a new useful specific molecular target to quantify Mycobacterium in environmental samples BMC Microbiol 13 277
Radomski N Thibault VC Karoui C de Cruz K Cochard T Gutierrez C Supply P Biet F Boschiroli ML 2010 Determination of genotypic diversity of Mycobacterium avium subspecies from human and animal origins by Mycobacterial Interspersed Repetitive-Unit-Variable-Number Tandem-Repeat and IS1311 Restriction Fragment Length Polymorphism typing methods J Clin Microbiol 48 1026ndash1034
Rahim Z Moumlllers M Te Koppele-Vije A De Beer J Zaman K Matin MA Kamal M Raquib R Van Soolingen D Baqi M a Heilmann FGC Van Der Zanden AGM 2007 Characterization of Mycobacterium africanum subtype i among cows in a dairy farm in Bangladesh using spoligotyping Southeast Asian J Trop Med Public Health 38 706ndash713
Rauzier J Moniz-Pereira J Gicquel-Sanzey B 1988 Complete nucleotide sequence of pAL5OOO a plasmid from Mycobacterium fortuitum Gene 71 315ndash321
Renwick AR White PCL Bengis RG 2007 Bovine tuberculosis in southern African wildlife a multi-species host-pathogen system Epidemiol Infect 135 529ndash540
Reviriego-Gordejo FJ Vermeersch JP 2006 Towards eradication of bovine tuberculosis in the European Union Vet Microbiol 112 101ndash109
Richomme C Boschiroli ML Hars J Casabianca F Ducrot C 2010 Bovine tuberculosis in livestock and wild boar on the Mediterranean island Corsica J Wildl Dis 46 627ndash631
Ringuet H Akoua-Koffi C Honore S Varnerot A Vincent V Berche P Gaillard JL Pierre-Audigier C 1999 hsp65 sequencing for identification of rapidly growing mycobacteria J Clin Microbiol 37 852ndash857
Ritacco V Loacutepez B De Kantor IN Barrera L Errico F Nader A 1991 Reciprocal cellular and humoral immune responses in bovine tuberculosis Res Vet Sci 50 365ndash367
Rivero A Maacuterquez M Santos J Pinedo A Saacutenchez MA Esteve A Samper S Martiacuten C 2001 High rate of tuberculosis reinfection during a nosocomial outbreak of multidrug-resistant tuberculosis caused by
Reacutefeacuterences bibliographiques
213
Mycobacterium bovis strain B Clin Infect Dis 32 159ndash161
Riviegravere J Fediaevsky A Hars J Richomme C Calavas D Hendrikx P 2011 SYLVATUB Dispositif national de surveillance de la tuberculose bovine dans la faune sauvage Organisation institutionnelle et reacuteseau dacteurs 2011ndash2012
Rodriguez JG Burbano CS Nuntildeez C Gonzaacutelez CE Zambrano MM Garciacutea MJ Del Portillo P 2008 Rv3134cdevRdevS operon of Mycobacterium bovis BCG is differentially transcribed under ldquoin vitrordquo stress conditions Tuberculosis 88 273ndash282
Rodriacuteguez S Bezos J Romero B de Juan L Aacutelvarez J Castellanos E Moya N Lozano F Javed MT Saacuteez-Llorente JL Lieacutebana E Mateos A Domiacutenguez L Aranaz A 2011 Mycobacterium caprae infection in livestock and wildlife Spain Emerg Infect Dis 17 532ndash535
Rodriguez-Campos S Schuumlrch AC Dale J Lohan AJ Cunha M V Botelho A Cruz K Boschiroli ML Boniotti MB Pacciarini M Garcia-Pelayo MC Romero B de Juan L Domiacutenguez L Gordon S V van Soolingen D Loftus B Berg S Hewinson RG Aranaz A Smith NH 2012 European 2 - A clonal complex of Mycobacterium bovis dominant in the Iberian Peninsula Infect Genet Evol 12 866ndash872
Rodriguez-Campos S Smith NH Boniotti MB Aranaz A 2014 Overview and phylogeny of Mycobacterium tuberculosis complex organisms Implications for diagnostics and legislation of bovine tuberculosis Res Vet Sci 97 Suppl S5ndashS19
Rodriguez-Nuntildeez J Avelar FJ Marquez F Rivas-Santiago B Quintildeones C Guerrero-Barrera AL 2012 Mycobacterium tuberculosis complex detected by modified fluorescent in situ hybridization in lymph nodes of clinical samples J Infect Dev Ctries 6(1) 58ndash66
Rodriacuteguez-Zaragoza S 1994 Ecology of free-living Amoebae Crit Rev Microbiol 20(3) 225-241
Rogers LM Forrester GJ Wilson GJ Yarnell RW Cheeseman CL 2003 The role of setts in badger (Meles meles) group size breeding success and status of TB (Mycobacterium bovis) J Zool 260 209ndash215
Romero B Rodriguez S Bezos J Diaz R Francisca Copano M Merediz I Minguez O Marques S Palacios JJ De Viedma DG Saez JL Mateos A Aranaz A Dominguez L De Juan L 2011 Humans as source of Mycobacterium tuberculosis infection in cattle Spain Emerg Infect Dis 17 2393ndash2395
Roring S Scott A Brittain D Walker I Hewinson G Neill SD Skuce R 2002 Development of variable-number tandem repeat typing of Mycobacterium bovis comparison of results with those obtained by using existing exact tandem repeats and spoligotyping J Clin Microbiol 40 2126ndash2133
Roth A Fischer M Hamid ME Michalke S Ludwig W Mauch H 1998 Differentiation of phylogenetically related slowly growing mycobacteria based on 16S-23S rRNA gene internal transcribed spacer sequences J Clin Microbiol 36 139ndash147
Rothschild BM Martin LD Lev G Bercovier H Bar-Gal GK Greenblatt C Donoghue H Spigelman M Brittain D 2001 Mycobacterium tuberculosis complex DNA from an extinct bison dated 17000 years before the present Clin Infect Dis 33 305ndash311
Roug A Geoghegan C Wellington E Miller W A Travis E Porter D Cooper D Clifford DL Mazet J AK Parsons S 2014 Utility of a fecal real-time PCR protocol for detection of Mycobacterium bovis infection in African buffalo (Syncerus caffer) J Wildl Dis 50 140ndash142
Rowe MT Grant IR 2006 Mycobacterium avium ssp paratuberculosis and its potential survival tactics Lett Appl Microbiol 42 305ndash11
Safi H Barnes PF Lakey DL Shams H Samten B Vankayalapati R Howard ST 2004 IS6110 functions as a mobile monocyte-activated promoter in Mycobacterium tuberculosis Mol Microbiol 52 999ndash1012
Reacutefeacuterences bibliographiques
214
Saleeb PG Drake SK Murray PR Zelazny AM 2011 Identification of mycobacteria in solid-culture media by matrix-assisted laser desorption ionization-time of flight mass spectrometry J Clin Microbiol 49 1790ndash1794
Santos N Almeida V Gortaacutezar C Correia-Neves M 2015a Patterns of Mycobacterium tuberculosis-complex excretion and characterization of super-shedders in naturally-infected wild boar and red deer Vet Res 46 129
Santos N Santos C Valente T Gortaacutezar C Almeida V Correia-Neves M 2015b Widespread environmental contamination with Mycobacterium tuberculosis Complex revealed by a molecular detection protocol PLoS One 10 e0142079
Savelkoul PHM Catsburg A Mulder S Oostendorp L Schirm J Wilke H van der Zanden AGM Noordhoek GT 2006 Detection of Mycobacterium tuberculosis complex with Real Time PCR Comparison of different primer-probe sets based on the IS6110 element J Microbiol Methods 66 177ndash180
Scanlon MP Quinn PJ 2000a The survival of Mycobacterium bovis in sterilized cattle slurry and its relevance to the persistence of this pathogen in the environment Ir Vet J 53 412ndash415
Scanlon MP Quinn PJ 2000b Inactivation of Mycobacterium bovis in cattle slurry by five volatile chemicals J Appl Microbiol 89 854ndash861
Schnappinger D Ehrt S Voskuil MI Liu Y Mangan JA Monahan IM Dolganov G Efron B Butcher PD Nathan C Schoolnik GK 2003 Transcriptional adaptation of Mycobacterium tuberculosis within macrophages insights into the phagosomal environment J Exp Med 198 693ndash704
Schoepf K Prodinger WM Glawischnig W Hofer E Revilla-Fernandez S Hofrichter J Fritz J Koumlfer J Schmoll F 2012 A two-yearsrsquo survey on the prevalence of tuberculosis caused by Mycobacterium caprae in red deer (Cervus elaphus) in the Tyrol Austria ISRN Vet Sci 2012 245138
Schroumlder KH 1982 Isolation of M africanum from German patients Bull Int Union Tuberc 57 249ndash53
Schuumlrch AC van Soolingen D 2012 DNA fingerprinting of Mycobacterium tuberculosis from phage typing to whole-genome sequencing Infection Genetics and Evolution 12 602-609
Shepard CC 1960 The experimental disease that follows the injection of human leprosy bacilli into foot-pads of mice J Exp Med 112 445ndash454
Shiloh MU Manzanillo P Cox JS 2008 Mycobacterium tuberculosis Senses Host-Derived Carbon Monoxide during Macrophage Infection Cell Host Microbe 3 323ndash330
Shinnick TM Good RC 1994 Mycobacterial taxonomy Eur J Clin Microbiol Infect Dis 13 884ndash901
Shitikov E Ilina E Chernousova L Borovskaya A Rukin I Afanasrsquoev M Smirnova T Vorobyeva A Larionova E Andreevskaya S Kostrzewa M Govorun V 2012 Mass spectrometry based methods for the discrimination and typing of mycobacteria Infect Genet Evol 12 838ndash845
Shrikrishna D de la Rua-Domenech R Smith NH Colloff A Coutts I 2009 Human and canine pulmonary Mycobacterium bovis infection in the same household re-emergence of an old zoonotic threat Thorax 64 89ndash91
Shuralev E Quinn P Doyle M Duignan A Kwok HF Bezos J Olwill SA Gormley E Aranaz A Good M Davis WC Clarke J Whelan C 2012 Application of the Enfer chemiluminescent multiplex ELISA system for the detection of Mycobacterium bovis infection in goats Vet Microbiol 154 292ndash297
Skuce RA McCorry TP McCarroll JF Roring SMM Scott AN Brittain D Hughes SL Hewinson RG Neill SD 2002 Discrimination of Mycobacterium tuberculosis complex bacteria using novel VNTR-PCR
Reacutefeacuterences bibliographiques
215
targets Microbiology 148 519ndash528
Skuce RA Brittain D Hughes MS Beck LA Neill SD 1994 Genomic fingerprinting of Mycobacterium bovis from cattle by restriction fragment length polymorphism analysis J Clin Microbiol 32 2387ndash2392
Slana I Kaevska M Kralik P Horvathova A Pavlik I 2010 Distribution of Mycobacterium avium subsp avium and M a hominissuis in artificially infected pigs studied by culture and IS901 and IS1245 quantitative real time PCR Vet Microbiol 144 437ndash443
Smith NH 2012 The global distribution and phylogeography of Mycobacterium bovis clonal complexes Infect Genet Evol 12 857ndash865
Smith NH Berg S Dale J Allen AR Rodriguez S Romero B Matos F Ghebremichael S Karoui C Donati C Machado A da C Mucavele C Kazwala RR Hilty M Cadmus SI Ngandolo BNR Habtamu M Oloya J Muller A Milian-Suazo F Andrievskaia O Projahn M Barandiaraacuten S Maciacuteas A Muumlller B Zanini MS Ikuta CY Rodriguez CAR Pinheiro SR Figueroa A Cho S-N Mosavari N Chuang P-C Jou R Zinsstag J van Soolingen D Costello E Aseffa A Proantildeo-Perez F Portaels F Rigouts L Cataldi AA Collins DM Boschiroli ML Hewinson RG Ferreira Neto JS Surujballi O Tadyon K Botelho A Zaacuterraga AM Buller N Skuce R Michel A Aranaz A Gordon S V Jeon B-Y Kaumlllenius G Niemann S Boniotti MB van Helden PD Harris B Zumaacuterraga MJ Kremer K 2011 European 1 a globally important clonal complex of Mycobacterium bovis Infect Genet Evol 11 1340ndash1351
Smith NH Crawshaw T Parry J Birtles RJ 2009 Mycobacterium microti More diverse than previously thought J Clin Microbiol 47 2551ndash2559
Smith NH Kremer K Inwald J Dale J Driscoll JR Gordon S V Van Soolingen D Glyn Hewinson R Maynard Smith J 2006 Ecotypes of the Mycobacterium tuberculosis complex J Theor Biol 239 220ndash225
Smith NH Upton P 2012 Naming spoligotype patterns for the RD9-deleted lineage of the Mycobacterium tuberculosis complex wwwMbovisorg Infect Genet Evol 12 873ndash876
Smith RMM Drobniewski F Gibson A Montague JDE Logan MN Hunt D Hewinson G Salmon RL OrsquoNeill B 2004 Mycobacterium bovis infection United Kingdom Emerg Infect Dis 10 539ndash541
Smithwick RW Stratigos CB David HL 1975 Use of cetylpyridinium chloride and sodium chloride for the decontamination of sputum specimens that are transported to the laboratory for the isolation of Mycobacterium tuberculosis J Clin Microbiol 1 411ndash413
Soini H Musser JM 2001 Molecular diagnosis of mycobacteria Clin Chem 47 809ndash814
Sola C Filliol I Gutierrez MC Mokrousov I Vincent V Rastogi N 2001 Spoligotype database of Mycobacterium tuberculosis biogeographic distribution of shared types and epidemiologic and phylogenetic perspectives Emerg Infect Dis 7 390ndash396
Soo P-C Horng Y-T Hsueh P-R Shen B-J Wang J-Y Tu H-H Wei J-R Hsieh S-C Huang C-C Lai H-C 2006 Direct and simultaneous identification of Mycobacterium tuberculosis complex (MTBC) and Mycobacterium tuberculosis (MTB) by rapid multiplex nested PCR-ICT assay J Microbiol Methods 66 440ndash448
Sreevatsan S Pan X 1997 Restricted structural gene polymorphism in the Mycobacterium tuberculosis complex indicates evolutionarily recent global dissemination ProcNatl Acad Sci USA 94 9869ndash9874
Stahl DA Urbance JW 1990 The division between fast- and slow-growing species corresponds to natural relationships among the mycobacteria J Bacteriol 172 116ndash124
Reacutefeacuterences bibliographiques
216
Steingart KR Ng V Henry M Hopewell PC Ramsay A Cunningham J Urbanczik R Perkins MD Aziz MA Pai M 2006 Sputum processing methods to improve the sensitivity of smear microscopy for tuberculosis a systematic review Lancet Infect Dis 6 664ndash674
Stender H Lund K Petersen KH Rasmussen OF Hongmanee P Miorner H Godtfredsen SE 1999a Fluorescence In Situ Hybridization assay using peptide nucleic acid probes for differentiation between tuberculous and nontuberculous Mycobacterium species in smears of Mycobacterium cultures J Clin Microbiol 37 2760ndash2765
Stender H Mollerup TA Lund K Petersen KH Hongmanee P Godtfredsen SE 1999b Direct detection and identification of Mycobacterium tuberculosis in smear-positive sputum samples by fluorescence in situ hybridization (FISH) using peptide nucleic acid (PNA) probes Int J Tuberc lung Dis 3 830ndash837
Stewart LD McNair J McCallan L Gordon A Grant IR 2013 Improved detection of Mycobacterium bovis infection in bovine lymph node tissue using immunomagnetic separation (IMS)-based methods PLoS One 8 e58374
Stewart LD McNair J McCallan L Thompson S Kulakov LA Grant IR 2012 Production and evaluation of antibodies and phage display-derived peptide ligands for immunomagnetic separation of Mycobacterium bovis J Clin Microbiol 50 1598ndash1605
Stinear TP Mve-Obiang A Small PLC Frigui W Pryor MJ Brosch R Jenkin GA Johnson PDR Davies JK Lee RE Adusumilli S Garnier T Haydock SF Leadlay PF Cole ST 2004 Giant plasmid-encoded polyketide synthases produce the macrolide toxin of Mycobacterium ulcerans Proc Natl Acad Sci U S A 101 1345ndash1349
Sunder S Lanotte P Godreuil S Martin C Boschiroli ML Besnier JM 2009 Human-to-human transmission of tuberculosis caused by Mycobacterium bovis in immunocompetent patients J Clin Microbiol 47 1249ndash1251
Supply P Allix C Lesjean S Cardoso-Oelemann M Rusch-Gerdes S Willery E Savine E de Haas P van Deutekom H Roring S Bifani P Kurepina N Kreiswirth B Sola C Rastogi N Vatin V Gutierrez MC Fauville M Niemann S Skuce R Kremer K Locht C van Soolingen D 2006 Proposal for standardization of optimized Mycobacterial Interspersed Repetitive Unit-Variable-Number Tandem Repeat typing of Mycobacterium tuberculosis J Clin Microbiol 44 4498ndash4510
Supply P Lesjean S Savine E Kremer K Locht C 2001 Automated high-throughput genotyping for study of global epidemiology of Mycobacterium tuberculosis based on Mycobacterial Interspersed Repetitive Units J Clin 39 3563ndash3571
Supply P Marceau M Mangenot S Roche D Rouanet C Khanna V Majlessi L Criscuolo A Tap J Pawlik A Fiette L Orgeur M Fabre M Parmentier C Frigui W Simeone R Boritsch EC Debrie A-S Willery E Walker D Quail M a Ma L Bouchier C Salvignol G Sayes F Cascioferro A Seemann T Barbe V Locht C Gutierrez M-C Leclerc C Bentley SD Stinear TP Brisse S Meacutedigue C Parkhill J Cruveiller S Brosch R 2013 Genomic analysis of smooth tubercle bacilli provides insights into ancestry and pathoadaptation of Mycobacterium tuberculosis Nat Genet 45 172ndash179
Sweeney FP Courtenay O Hibberd V Hewinson RG Reilly LA Gaze WH Wellington EMH 2007 Environmental monitoring of Mycobacterium bovis in badger feces and badger sett soil by real-time PCR as confirmed by immunofluorescence immunocapture and cultivation Appl Environ Microbiol 73 7471ndash7473
Sweeney FP Courtenay O Ul-Hassan A Hibberd V Reilly LA Wellington EMH 2006 Immunomagnetic recovery of Mycobacterium bovis from naturally infected environmental samples Lett Appl Microbiol 43 364ndash369
Reacutefeacuterences bibliographiques
217
Tanner M Michel AL 1999 Investigation of the viability of M bovis under different environmental conditions in the Kruger National Park J Vet Res 185ndash190
Taquet A Tison F 1961 Nouvelle technique drsquoisolement des mycobacteacuteries par le lauryl sulfate de sodium Ann Inst Pasteur 100 676ndash680
Taylor C Jahans K Palmer S Okker M Brown J Steer K 2006 Mycobacterium microti isolated from two pigs Vet Rec 159 59ndash60
Taylor SJ Ahonen LJ de Leij FAAM Dale JW 2003 Infection of Acanthamoeba castellanii with Mycobacterium bovis BCG and survival of M bovis within the amoeba Appl Environ Microbiol 69 4316ndash4319
Telenti A Marchesi F Balz M Bally F Botrger EC Bodmer T 1993 Rapid identification of mycobacteria to the species level by Polymerase Chain Reaction and Restriction Enzyme Analysis J Clin Microbiol 31 175ndash178
Thacker TC Palmer M V Robbe-Austerman S Stuber TP Waters WR 2015 Anatomical distribution of Mycobacterium bovis genotypes in experimentally infected white-tailed deer Vet Microbiol180(1-2) 75-81
Thierry D Brisson-Noeumll A Vincent-Leacutevy-Freacutebault V Nguyen S Guesdon JL Gicquel B 1990a Characterization of a Mycobacterium tuberculosis insertion sequence IS6110 and its application in diagnosis J Clin Microbiol 28 2668ndash2673
Thierry D Cave MD Eisenach KD Crawford JT Bates JH Gicquel B Guesdon JL 1990b IS6110 an IS-like element of Mycobacterium tuberculosis complex Nucleic Acids Res 18 188
Thoen C LoBue P De Kantor I 2006 The importance of Mycobacterium bovis as a zoonosis Vet Microbiol 112 339ndash345
Thomas V Blanc D Bille J Greub G 2006 Biodiversity of amoebae resistant micro-organisms in an hospital water network Appl Environ Microbiol 72 2428ndash2438
Thomas V Loret J-F Jousset M Greub G 2008 Biodiversity of amoebae and amoebae-resisting bacteria in a drinking water treatment plant Environ Microbiol 10 2728ndash2745
Thomas V McDonnell G 2007 Relationship between mycobacteria and amoebae ecological and epidemiological concerns Lett Appl Microbiol 45 349ndash357
Thorel MF 1980 Isolation of Mycobacterium africanum from monkeys Tubercle 61 101ndash104
Tolhurst BA Delahay RJ Walker NJ Ward AI Roper TJ 2009 Behaviour of badgers (Meles meles) in farm buildings Opportunities for the transmission of Mycobacterium bovis to cattle Appl Anim Behav Sci 117 103ndash113
Torgerson PR Torgerson DJ 2010 Public health and bovine tuberculosis whatrsquos all the fuss about Trends Microbiol 18 67ndash72
Tortoli E 2014 Microbiological features and clinical relevance of new species of the genus Mycobacterium Clin Microbiol Rev 27 727ndash752
Tortoli E 2009 Clinical manifestations of nontuberculous mycobacteria Clin Microbiol Infect 15 906ndash910
Tortoli E 2003 Impact of genotypic studies on mycobacterial taxonomy the new mycobacteria of the 1990s Clin Microbiol Rev 16 319ndash354
Reacutefeacuterences bibliographiques
218
Torvinen E Lehtola MJ Martikainen PJ Miettinen IT 2007 Survival of Mycobacterium avium in drinking water biofilms as affected by water flow velocity availability of phosphorus and temperature Appl Environ Microbiol 73 6201ndash6207
Traag BA Driks A Stragier P Bitter W Broussard G Hatfull G Chu F Adams KN Ramakrishnan L Losick R 2010 Do mycobacteria produce endospores Proc Natl Acad Sci 107 878ndash881
Travis ER Gaze WH Pontiroli A Sweeney FP Porter D Mason S Keeling MJC Jones RM Sawyer J Aranaz A Rizaldos EC Cork J Delahay RJ Wilson GJ Hewinson RG Courtenay O Wellington EMH 2011 An inter-laboratory validation of a real time PCR assay to measure host excretion of bacterial pathogens particularly of Mycobacterium bovis PLoS One 6 e27369
Trujillo ME Velaacutezquez E Kroppenstedt RM Schumann P Rivas R Mateos PF Martiacutenez-Molina E 2004 Mycobacterium psychrotolerans sp nov isolated from pond water near a uranium mine Int J Syst Evol Microbiol 54 1459ndash1463
Truman R 2005 Leprosy in wild armadillos Lepr Rev 76 198ndash208
Tsairidou S Woolliams JA Allen AR Skuce RA McBride SH Wright DM Bermingham ML Pong-Wong R Matika O McDowell SWJ Glass EJ Bishop SC 2014 Genomic prediction for tuberculosis resistance in dairy cattle PLoS One 9(5) e96728
Turenne CY 2004 Mycobacterium saskatchewanense sp nov a novel slowly growing scotochromogenic species from human clinical isolates related to Mycobacterium interjectum and Accuprobe-positive for Mycobacterium avium complex Int J Syst Evol Microbiol 54 659ndash667
Van Donsel DJ Larkin EP 1977 Persistence of Mycobacterium bovis BCG in soil and on vegetables spray irrigated with sewage effluent and sludge J Food Prot 40 160ndash163
Van Ingen J Bendien SA de Lange WCM Hoefsloot W Dekhuijzen PNR Boeree MJ van Soolingen D 2009 Clinical relevance of non-tuberculous mycobacteria isolated in the Nijmegen-Arnhem region The Netherlands Thorax 64 502ndash506
Van Ingen J Rahim Z Mulder A Boeree MJ Simeone R Brosch R van Soolingen D 2012 Characterization of Mycobacterium orygis as M tuberculosis complex subspecies Emerg Infect Dis 18 653ndash655
Van Ingen J Van 2013 Diagnosis of nontuberculous mycobacterial infections Semin Respir Crit Care Med 34 103ndash109
Van Pittius NCG Perrett KD Michel AL Keet DF Hlokwe T Streicher EM Warren RM van Helden PD 2012 Infection of African Buffalo (Syncerus Caffer) by Oryx Bacillus A rare member of the antelope clade of the Mycobacterium Tuberculosis Complex J Wildl Dis 48 849ndash857
Van Soolingen D De Haas PEW Haagsma J Eger T Hermans PWM Ritacco V Alito A Van Embden JDA 1994 Use of various genetic markers in differentiation of Mycobacterium bovis strains from animals and humans and for studying epidemiology of bovine tuberculosis J Clin Microbiol 32 2425ndash2433
Van Soolingen D Hermans PWM De Haas PEW Soll DR Van Embden JDA 1991 Occurrence and stability of insertion sequences in Mycobacterium tuberculosis complex strains Evaluation of an insertion sequence-dependent DNA polymorphism as a tool in the epidemiology of tuberculosis J Clin Microbiol 29 2578ndash2586
Vandal OH Pierini LM Schnappinger D Nathan CF Ehrt S 2008 A membrane protein preserves intrabacterial pH in intraphagosomal Mycobacterium tuberculosis Nat Med 14 849ndash854
Varello K Pezzolato M Mascarino D Ingravalle F Caramelli M Bozzetta E 2008 Comparison of
Reacutefeacuterences bibliographiques
219
histologic techniques for the diagnosis of bovine tuberculosis in the framework of eradication programs J Vet Diagnostic Investig 20 164ndash169
Velayati AA Farnia P Mozafari M Malekshahian D Farahbod AM Seif S Rahideh S Mirsaeidi M 2015 Identification and genotyping of Mycobacterium tuberculosis isolated from water and soil samples of a metropolitan city CHEST J 147 1094
Vicente J Houmlfle U Garrido JM Fernaacutedez-De-Mera IG Acevedo P Juste R Barral M Gortaacutezar C 2007 Risk factors associated with the prevalence of tuberculosis-like lesions in fenced wild boar and red deer in south central Spain Vet Res 38 451ndash464
Voskuil MI Schnappinger D Rutherford R Liu Y Schoolnik GK 2004 Regulation of the Mycobacterium tuberculosis PEPPE genes Tuberculosis (Edinb) 84 256ndash262
Voskuil MI Schnappinger D Visconti KC Harrell MI Dolganov GM Sherman DR Schoolnik GK 2003 Inhibition of respiration by nitric oxide induces a Mycobacterium tuberculosis dormancy program J Exp Med 198 705ndash713
Wagner JC Buchanan G Bokkenheuser V Leviseur S 1958 An acid-fast bacillus isolated from the lungs of the Cape Hyrax Procavia capensis (Pallas) Nature 181 284ndash285
Walter WD Smith R Vanderklok M VerCauteren KC 2014 Linking bovine tuberculosis on cattle farms to white-tailed deer and environmental variables using Bayesian hierarchical analysis PLoS One 9 e90925
Wang Y Ogawa M Fukuda K Miyamoto H Taniguchi H 2006 Isolation and identification of mycobacteria from soils at an illegal dumping site and landfills in Japan Microbiol Immunol 50 513ndash524
Ward AI Judge J Delahay RJ 2010 Farm husbandry and badger behaviour Opportunities to manage badger to cattle transmission of Mycobacterium bovis Prev Vet Med 93 2ndash10
Warren RM Gey Van Pittius NC Barnard M Hesseling A Engelke E De Kock M Gutierrez MC Chege GK Victor TC Hoal EG Van Helden PD 2006 Differentiation of Mycobacterium tuberculosis complex by PCR amplification of genomic regions of difference Int J Tuberc Lung Dis 10 818ndash822
Waters WR Buddle BM Vordermeier HM Gormley E Palmer M V Thacker TC Bannantine JP Stabel JR Linscott R Martel E Milian F Foshaug W Lawrence JC 2011 Development and evaluation of an enzyme-linked immunosorbent assay for use in the detection of bovine tuberculosis in cattle Clin Vaccine Immunol 18 1882ndash1888
Wayne LG Hayes LG 1996 An in vitro model for sequential study of shiftdown of Mycobacterium tuberculosis through two stages of nonreplicating persistence Infect Immun 64 2062ndash2069
Wells AQ 1937 Tuberculosis in wild voles Lancet 229 1221
Wells AQ Robb-Smith AHT 1946 Murine type of tubercle bacillus (the vole acid-fast bacillus) Spec Rep Ser Med Res Cou London 259
Whan L Grant IR Rowe MT 2006 Interaction between Mycobacterium avium subsp paratuberculosis and environmental protozoa BMC Microbiol 6 63
Whiley H Keegan A Giglio S Bentham R 2012 Mycobacterium avium complex - the role of potable water in disease transmission J Appl Microbiol 113 223ndash32
White PCL Brown JA Harris S 1993 Badgers (Meles meles) cattle and bovine tuberculosis (Mycobacterium bovis) a hypothesis to explain the influence of habitat on the risk of disease transmission in southwest England Proc R Soc London B 253 277ndash284
Reacutefeacuterences bibliographiques
220
Whittington R Marshall D Nicholls PJ Marsh IB Reddacliff LA 2004 Survival and dormancy of Mycobacterium avium subsp paratuberculosis in the environment Appl Environ Microbiol 70 2989ndash3004
WHO 2014 Global leprosy update 2013 Wkly Epidemiol Rec 89 389ndash400
WHO 2014 Global Tuberculosis Report 2014
Williams RS Hoy WA 1930 The viability of B tuberculosis (bovinus) on pasture land in stored faeces and in liquid manure J Hyg (Lond) 30 413ndash419
Wirth T Hildebrand F Allix-Beacuteguec C Woumllbeling F Kubica T Kremer K Van Soolingen D Ruumlsch-Gerdes S Locht C Brisse S Meyer A Supply P Niemann S 2008 Origin spread and demography of the Mycobacterium tuberculosis complex PLoS Pathog 4(9) e1000160
Witmer G Fine AE Gionfriddo J Pipas M Shively K Piccolo K Burke P 2010 Epizootiologic survey of Mycobacterium bovis in wildlife and farm environments in northern Michigan J Wildl Dis 46 368ndash378
Wobeser G 2009 Bovine tuberculosis in Canadian wildlife an updated history Can Vet J 50 1169ndash1176
Wright DM Allen AR Mallon TR McDowell SWJ Bishop SC Glass EJ Bermingham ML Woolliams JA Skuce RA 2013 Field-isolated genotypes of Mycobacterium bovis vary in virulence and influence case pathology but do not affect outbreak size PLoS One 8(9) e74503
Young JS Gormley E Wellington EMH 2005 Molecular detection of Mycobacterium bovis and Mycobacterium bovis BCG (Pasteur) in soil Appl Environ Microbiol 71 1946ndash1952
Zanella G Durand B Hars J Moutou F Garin-Bastuji B Duvauchelle A Fermeacute M Karoui C Boschiroli ML 2008 Mycobacterium bovis in wildlife in France J Wildl Dis 44 99ndash108
Zelazny AM Calhoun LB L Shea YR Fischer SH 2005 Identification of Mycobacterium species by secA1 sequences J Clin Microbiol 43 1051ndash1058
Zeng W Zhang Y Zhao X Huang G Jiang Y Dong H Li X Wan K He C 2013 Occurrence of non-tuberculous mycobacteria species in livestock from northern China and first isolation of Mycobacterium caprae Epidemiol Infect 141 1545ndash1551
Zhang J Abadia E Refregier G Tafaj S Boschiroli ML Guillard B Andremont A Ruimy R Sola C 2010 Mycobacterium tuberculosis complex CRISPR genotyping Improving efficiency throughput and discriminative power of ldquospoligotypingrdquo with new spacers and a microbead-based hybridization assay J Med Microbiol 59 285ndash294
Zhang Y Yew W 2009 Mechanisms of drug resistance in Mycobacterium tuberculosis Int J Tuberc Lung Dis 13 1320ndash1330
Zolg JW Philippi-Schulz S 1994 The superoxide dismutase gene a target for detection and identification of mycobacteria by PCR J Clin Microbiol 32 2801ndash2812
Zuber B Chami M Houssin C Dubochet J Griffiths G Daffeacute M 2008 Direct visualization of the outer membrane of mycobacteria and corynebacteria in their native state J Bacteriol 190 5672ndash5680
Annexes
223
Annexe 1 Deacutetection de Mycobacterium bovis dans lrsquoenvironnement Premiers
reacutesultats des recherches meneacutees en Cocircte-drsquoOr
Deacutetection de Mycobacterium bovis dans lrsquoenvironnement
Premiers reacutesultats des recherches meneacutees en Cocircte-drsquoOr
Elodie Barbier1 et Alain Hartmann1
RESUME
Depuis les anneacutees 2000 en Cocircte-drsquoOr on assiste agrave une recrudescence des cas de tuberculose agrave
Mycobacterium bovis dans les cheptels bovins associeacutee agrave une contamination de la faune sauvage agrave
proximiteacute des eacutelevages contamineacutes Dans certaines zones du deacutepartement la faune sauvage infecteacutee
serait impliqueacutee dans le retour de la maladie aux bovins La transmission indirecte de la maladie par
lrsquoenvironnement semblerait une voie importante de la contamination entre faune sauvage et
domestique
Afin drsquoeacutevaluer la preacutevalence de M bovis dans lrsquoenvironnement et drsquoeacutevaluer le rocircle de lrsquoenvironnement
dans la transmission indirecte de la maladie nous avons deacuteveloppeacute des outils moleacuteculaires de
deacutetection adapteacutes aux matrices environnementales complexes (sol eau matiegraveres feacutecales) Ces outils agrave
la fois speacutecifiques et sensibles nous ont permis drsquoanalyser de nombreux preacutelegravevements reacutealiseacutes en 2013
Seuls deux eacutechantillons de sol se sont reacuteveacuteleacutes faiblement positifs
Mots-cleacutes tuberculose bovine bovins faune sauvage environnement PCR en temps reacuteel
SUMMARY
Since 2000 an increasing number of cases of bovine tuberculosis were found in cattle in Cocircte-drsquoOr and
in wild animals infected by Mycobacterium bovis in the vicinity of contaminated farms In some areas
wildlife could be considered as the likely source of cattle contamination We assume that
contamination between wild and domestic animals may occur indirectly by environmental factors (soil
water feces)
To quantify the prevalence of M bovis in the environment and to evaluate its role in indirect
transmission of the disease between species we developed molecular detection tools These are
adapted to complex environmental matrices (soil water faces) and are both specific and sensitive
They allowed us to analyze numerous samples collected in 2013 Two samples gave slight positive
results
Keywords Bovine tuberculosis Cattle Wildlife Environment Real time PCR
Texte de la confeacuterence preacutesenteacutee au cours de la Journeacutee scientifique AEEMA-RFSA 18 mars 2014
1 INRA UMR Agroeacutecologie 16 rue de Sully BP 86510 21065 Dijon France
Eacutepideacutemiol et santeacute anim 2014 65 Deacutetection de Mycobacterium bovis dans lrsquoenvironnement Premiers reacutesultats des recherches meneacutees en Cocircte-drsquoOr
224
I - Introduction
Deacuteclareacutee indemne de tuberculose bovine en
2000 la France connait depuis 2002 une
reacuteeacutemergence de lrsquoinfection dans certains
deacutepartements notamment en Cocircte-drsquoOr Entre
2002 et aujourdrsquohui 178 foyers ont eacuteteacute
successivement deacuteclareacutes dans ce deacutepartement
Pour la saison 2012-2013 la moitieacute des
cheptels infecteacutes avaient deacutejagrave eacuteteacute deacuteclareacutes
foyer une deux voire trois fois auparavant La
reacutepartition des cas bovins sur le deacutepartement
nrsquoest pas aleacuteatoire mais concentreacutee sur une
zone assez reacuteduite
Deux spoligotypes principaux sont retrouveacutes
chez les bovins SB0120 (VNTR 5544) appeleacute
plus simplement BCG-like dans la reacutegion de
Veacutenarey-Les-Laumes et Vitteaux et SB0134
(VNTR 5355) appeleacute GB35 dans la reacutegion de
Pouilly-en-Auxois
Parallegravelement agrave lrsquoinfection bovine et agrave
proximiteacute drsquoun cheptel foyer lrsquoinfection a eacuteteacute
deacutetecteacutee pour la premiegravere fois chez un animal
sauvage en 2003 chez un cerf Entre 2003 et
2007 seulement deux sangliers en plus du
cervideacute ont eacuteteacute trouveacutes infecteacutes Depuis 2007
et chaque anneacutee la maladie est reacuteguliegraverement
retrouveacutee chez des blaireaux et des sangliers
moins reacuteguliegraverement chez les cerfs Les
animaux sauvages infecteacutes sont
majoritairement trouveacutes agrave proximiteacute de
cheptels bovins infecteacutes
Lrsquohypothegravese drsquoun cycle eacutepideacutemiologique de
transmission de la maladie entre bovins et
faune sauvage est maintenant reconnue
[Richomme et al 2010] mecircme si celui-ci est
encore mal connu En France les exemples de
la forecirct de Brotonne [Zanella et al 2008] de la
Cocircte-drsquoOr de la Dordogne [Payne et al 2012]
de la Corse [Richomme et al 2010] et ceux de
lrsquoEspagne [Aranaz et al 2004] ont montreacute que
les spoligotypes et les profils VNTR (Variable
Number Tandem Repeat) des souches isoleacutees
chez les animaux sauvages tueacutes ou pieacutegeacutes agrave
proximiteacute des cheptels bovins infecteacutes sont
exactement les mecircmes que ceux des souches
bovines Cette identiteacute des souches reacutesulterait
drsquoune transmission directe ou indirecte entre
faune domestique et sauvage et vice et versa
Bien que quelques contacts directs soient
possibles entre faune sauvage et bovins [Boumlhm
et al 2009] il semble probable que la
transmission indirecte de M bovis soit la voie
de transmission preacutepondeacuterante entre bovin et
faune sauvage et inversement Il semble eacutetabli
que la transmission indirecte de M bovis soit la
principale voie de transmission entre bovins et
faune sauvage et inversement La transmission
indirecte ferait intervenir une excreacutetion de M
bovis par les animaux infecteacutes au niveau
respiratoire salivaire urinaire etou feacutecal les
animaux excreacuteteurs deacuteposeraient donc le
bacille dans lrsquoenvironnement par le biais de
leurs excreacutetats lrsquoenvironnement deviendrait
alors un reacuteservoir potentiel pour le bacille La
contamination drsquoanimaux sains pourrait se
produire par ingestion ou inhalation drsquoaeacuterosols
produits agrave partir des matrices
environnementales souilleacutees Par exemple la
formation de bioaeacuterosols contamineacutes par des
souches bacteacuteriennes drsquoactinomycegravetes
thermotoleacuterants est maintenant bien
documenteacutee agrave partir de matrices solides ou
liquides [Betteli et al 2013] Dans
lrsquoenvironnement des bovins ces bioaeacuterosols
pourraient ecirctre formeacutes agrave partir drsquoeau ou de
meacutelanges eau-seacutediments dans les zones
drsquoabreuvement ou agrave partir de particules fines
de sol (argiles)
Lrsquoexcreacutetion respiratoire salivaire urinaire et
feacutecale de M bovis par des blaireaux
tuberculeux a eacuteteacute documenteacutee en Angleterre
[Gallagher et al 1998 Gavier-Widen et al
2001 Corner et al 2011] De plus
Mycobacterium bovis a la capaciteacute de survivre
en dehors de ses hocirctes pendant des dureacutees
Eacutepideacutemiol et santeacute anim 2014 65 Deacutetection de Mycobacterium bovis dans lrsquoenvironnement Premiers reacutesultats des recherches meneacutees en Cocircte-drsquoOr
225
assez longues Cette dureacutee varie en fonction de
la nature du support contamineacute (eau sol foin
maiumls etc) de la tempeacuterature de lrsquohumiditeacute et
de lrsquoexposition aux UV [Courtenay et al 2006
Fine et al 2011 Ghodbane et al 2014] La
survie de M bovis dans des microcosmes de
sols contamineacutes soumis aux facteurs
climatiques a mecircme eacuteteacute deacutemontreacutee aux Etats-
Unis pour des dureacutees allant jusqursquoagrave environ 88
jours [Fine et al 2011]
La recherche de M bovis par une meacutethode
culturale dans lrsquoenvironnement drsquoanimaux
infecteacutes est souvent infructueuse [Pillai et al
2000 Michel et al 2007 Fine et al 2011] car
elle se heurte agrave plusieurs difficulteacutes i) la vitesse
de croissance tregraves lente de M bovis de lrsquoordre
de 45 jours qui impose un traitement preacutealable
des eacutechantillons de faccedilon agrave deacutetruire les micro-
organismes agrave croissance plus rapide ii) la
richesse des matrices analyseacutees (sol
seacutediments eau matiegraveres feacutecales etc) en
bacteacuteries saprophytes et en champignons tregraves
diversifieacutes qui rendent la deacutecontamination des
eacutechantillons tregraves difficile La deacutecontamination a
toutefois un effet non neacutegligeable sur la
viabiliteacute de M bovis et interfegravere avec la
sensibiliteacute de deacutetection par la mise en culture
[Fine 2006] Lrsquoimmunocapture magneacutetique a
eacuteteacute testeacutee avec succegraves en Angleterre pour isoler
et concentrer le bacille avant la mise en culture
[Sweeney et al 2006 et 2007]
La technique de PCR (Polymerase Chain
Reaction) est une technique indeacutependante de
la croissance de la bacteacuterie Quelques eacutetudes
ont permis la deacutetection par PCR du bacille dans
des sols et des latrines de blaireaux au
Royaume-Uni et en Irlande agrave proximiteacute de
cheptels bovins infecteacutes [Young et al 2005
Courtenay et al 2006 Sweeney et al 2006 et
Les objectifs de ce travail visaient en premier
lieu agrave mettre au point un systegraveme de deacutetection
moleacuteculaire agrave la fois speacutecifique du bacille et
assez sensible pour deacutetecter de faibles
concentrations dans les diffeacuterentes matrices
environnementales suspecteacutees pour leur rocircle
de reacuteservoir potentiel Ces nouveaux outils
moleacuteculaires nous ont permis drsquoanalyser les
eacutechantillons preacuteleveacutes dans des parcelles
partageacutees par les bovins et la faune sauvage
Ces parcelles ont eacuteteacute choisies car une
transmission indirecte de lrsquoinfection entre
animaux sauvages et domestiques eacutetait
suspecteacutee (origine de lrsquoinfection bovine
indeacutetermineacutee suite agrave lrsquoenquecircte
eacutepideacutemiologique et preacutesence drsquoanimaux
sauvages infecteacutes agrave proximiteacute) Cette approche
permettra drsquoeacutevaluer lrsquoimportance de la
transmission indirecte du bacille entre les
espegraveces et drsquoappreacutecier le rocircle joueacute par
lrsquoenvironnement
Cet article ne traite que de la deacutetection
moleacuteculaire la meacutethode fondeacutee sur la culture
eacutetant toujours en cours de validation
II - Mateacuteriel et meacutethodes
Souches bacteacuteriennes utiliseacutees
La souche vaccinale Mycobacterium bovis BCG
strain 1331 (SSI Copenhague Danemark) a eacuteteacute
utiliseacutee pour lrsquoeacutevaluation de la sensibiliteacute de la
PCR en temps reacuteel (qPCR) sur souche pure et
sur microcosmes de sol inoculeacute ainsi que pour
la reacutealisation des gammes eacutetalon de qPCR (PCR
quantitative) Cette souche a eacuteteacute cultiveacutee sur
geacutelose Coletsos pendant 30 jours puis remise
en suspension dans du milieu Middlebrook 7H9
enrichi en ADC (Sigma Aldrich) Apregraves mesure
de la densiteacute optique agrave 600nm la suspension a
eacuteteacute dilueacutee en cascade au 110 pour inoculer les
microcosmes de sol et drsquoeau Un
Eacutepideacutemiol et santeacute anim 2014 65 Deacutetection de Mycobacterium bovis dans lrsquoenvironnement Premiers reacutesultats des recherches meneacutees en Cocircte-drsquoOr
226
deacutenombrement des dilutions est ensuite
reacutealiseacute sur milieu Middlebrook 7H11 enrichi en
OADC (Sigma Aldrich)
Les ADN de 20 spoligotypes de M bovis et de
30 souches de mycobacteacuteries issus drsquoisolats
cliniques drsquoanimaux infecteacutes ont eacuteteacute fournis par
lrsquoANSES Maisons-Alfort pour les tests de
speacutecificiteacute (inclusion - exclusion) de la PCR en
temps reacuteel (tableau 1)
Extraction de lrsquoADN et purification
Lrsquoextraction drsquoADN et la purification des
microcosmes de sol drsquoeau et des eacutechantillons
environnementaux a eacuteteacute reacutealiseacutee selon le
protocole modifieacute deacutecrit par Ranjard et al
[2003]
Les eacutechantillons drsquoeau ont eacuteteacute filtreacutes sur des
membranes drsquoester de cellulose de 045 microm de
porositeacute Les filtres ont eacuteteacute broyeacutes avec de
lrsquoazote liquide puis lrsquoADN extrait selon la
meacutethode preacuteceacutedemment citeacutee agrave lrsquoexception de
la purification sur colonnes PVPP
(polyvinylpolypyrrolidone insoluble)
Les ADN extraits ont eacuteteacute quantifieacutes avec un
spectrophotomegravetre (NanoDropreg 2000 Thermo
Scientific) et dilueacutes au 15 110 et 120 pour la
PCR en temps reacuteel
Deacuteveloppement de trois systegravemes de
deacutetection moleacuteculaire
La bibliographie nous a permis de seacutelectionner
trois cibles geacuteneacutetiques situeacutees sur des locus
diffeacuterents tregraves speacutecifiques de M bovis [Huard
et al 2002 Brodin et al 2002] Les geacutenomes
entiegraverement seacutequenceacutes des mycobacteacuteries du
MTBC (complexe M tuberculosis comprenant
M bovis) et des MNT (mycobacteacuteries non
tuberculeuses) sur le site NCBI nous ont permis
drsquoaligner les seacutequences de ces trois gegravenes agrave la
recherche de zones tregraves conserveacutees dans le
MTBC Chaque systegraveme de deacutetection
moleacuteculaire en PCR temps reacuteel creacuteeacute comprend
une amorce Forward une amorce Reverse et
une sonde oligonucleacuteotidique marqueacutee par
deux fluorochromes (sonde TaqMan) choisies
agrave lrsquoaide du logiciel Primer3 (Simgenecom) Pour
chaque systegraveme nous avons eacutegalement conccedilu
des amorces de PCR agrave lrsquoexteacuterieur des seacutequences
preacuteceacutedentes de faccedilon agrave reacutealiser des PCR
nicheacutees (utiliseacutees uniquement en confirmation
des PCR en temps reacuteel positives)
Speacutecificiteacute et sensibiliteacute des systegravemes de
deacutetection sur souches pures sol et eau
La speacutecificiteacute des trois systegravemes de deacutetection a
eacuteteacute testeacutee par PCR en temps reacuteel sur lrsquoADN de
19 spoligotypes diffeacuterents de M bovis quatre
souches de mycobacteacuteries appartenant au
MTBC deux au Complexe avium et 20 MNT
(tableau 1) Nous avons eacutegalement veacuterifieacute que
la PCR en temps reacuteel ne donnait pas de signal
sur des ADN extraits de 22 sols provenant du
Morbihan des Yvelines du Haut-Rhin et de
Cocircte-drsquoOr preacuteleveacutes en dehors des zones ougrave
seacutevit la tuberculose bovine Ces sols preacutesentent
des caracteacuteristiques physico-chimiques varieacutees
Pour deacuteterminer la sensibiliteacute de deacutetection de
la PCR en temps reacuteel de chacun des systegravemes
la quantification a eacuteteacute reacutealiseacutee sur des dilutions
du gegravene cible cloneacute dans un plasmide (gamme
allant de 1 copie agrave 106 copies du gegravene rechercheacute
dans 5 microl) La limite de deacutetection de la PCR en
temps reacuteel dans le sol a eacuteteacute eacutevalueacutee par
lrsquoinoculation de suspension bacteacuterienne de M
bovis BCG dans un sol dont les caracteacuteristiques
physico-chimiques sont identiques aux sols
incrimineacutes en Cocircte-drsquoOr Chaque microcosme
de sol de 2 g a eacuteteacute inoculeacute avec des suspensions
bacteacuteriennes pour obtenir une concentration
finale en bacteacuteries variant de 10 agrave 105 CFU par
g de sol Des aliquotes de 100 ml drsquoeau non
steacuterile issue drsquoun ruisseau hors zone de
tuberculose ont eacuteteacute inoculeacutes avec ces mecircmes
concentrations
Eacutepideacutemiol et santeacute anim 2014 65 Deacutetection de Mycobacterium bovis dans lrsquoenvironnement Premiers reacutesultats des recherches meneacutees en Cocircte-drsquoOr
227
Tableau 1
Souches bacteacuteriennes et spoligotypes utiliseacutes pour eacutevaluer la speacutecificiteacute des trois systegravemes
de deacutetection moleacuteculaire
M bovis (spoligotypes) Mycobacteacuteries du MTBC Mycobacteacuteries atypiques Mycobacteacuteries du
Complexe avium
GB35 M microti M chitae Mavium
BCG M tuberculosis M elephantis M intracellulare
F1 M caprae M confluentis
F2 M pinnipedi M fortuitum
F5 M rufum
F6 M non chromogenicum
F7 M aichiense
F23 M thermoresitibile
F41 M xenopi
F57 M kansai
F61 M vaccae
F105 M arupense
F151 M chelonae
GB21 M porcinum
GB54 M parafinicum
SB0166 M celatum
SB0885 M intermedium
SB0999 M rutilum
SB1081 M smegmatis
M szulgai
Choix des sites de preacutelegravevements
Les eacutechantillons environnementaux ont eacuteteacute
preacuteleveacutes sur huit communes de Cocircte-drsquoOr
situeacutees dans la zone Nord entre mai et juillet
2013 Nous avons cibleacute avec lrsquoaide de la DDPP21
des parcelles
Sur lesquelles un ou plusieurs bovins
deacutecouverts infecteacutes lors de la prophylaxie
2012-2013 ont pacirctureacute en 2012 et pour
lesquels lrsquoorigine de la contamination reste
inexpliqueacutee
Autour desquelles la surveillance de la
faune sauvage a mis en eacutevidence lrsquoinfection
agrave M bovis chez des blaireaux pieacutegeacutes ou des
sangliers tueacutes agrave la chasse
Sur les pacirctures nous avons privileacutegieacute les zones
susceptibles drsquoecirctre freacutequenteacutees par les bovins
et la faune sauvage zones drsquoabreuvement
(trous drsquoeau sources abreuvoirs) zones de
nourrissage au sol De lrsquoeau du sol et des
seacutediments ont eacuteteacute reacutecolteacutes Autour des
parcelles les terriers et matiegraveres feacutecales de
blaireaux (latrines) ainsi que les zones
drsquoabreuvement de la faune sauvage ont eacuteteacute
preacuteleveacutes
Eacutepideacutemiol et santeacute anim 2014 65 Deacutetection de Mycobacterium bovis dans lrsquoenvironnement Premiers reacutesultats des recherches meneacutees en Cocircte-drsquoOr
228
III - Reacutesultats
Les systegravemes de deacutetection conccedilus sont
speacutecifiques des espegraveces appartenant au MTBC
in silico
Les gegravenes ou locus cibles ont eacuteteacute choisis soit
pour leur speacutecificiteacute agrave M bovis soit pour leur
absence chez M microti seule mycobacteacuterie
du complexe MTBC agrave avoir eacuteteacute deacutecrite dans
lrsquoenvironnement avec M bovis Les systegravemes de
deacutetection ciblant lrsquoIS1561 et la reacutegion Rv3866
ont eacuteteacute choisis car ces locus sont preacutesents chez
tous les membres du MTBC mais absents chez
M microti chez les mycobacteacuteries du complexe
avium ainsi que chez les MNT [Huard et al
2002 Brodin et al 2002] La reacutegion RD4
preacutesente chez tous les membres du MTBC est
absente chez M bovis [Huard et al 2002]
Deux systegravemes de deacutetection moleacuteculaire
ciblant lrsquoIS1561 et la reacutegion Rv3866 ont eacuteteacute
reacutealiseacutes agrave lrsquoaide du logiciel Primer3 Le troisiegraveme
systegraveme ciblant la zone encadrant la RD4 utilise
une amorce Forward et une sonde deacutejagrave deacutecrite
[Sweeney et al 2007] seule lrsquoamorce Reverse
a eacuteteacute redessineacutee afin drsquoobtenir un produit PCR
plus petit La sonde utiliseacutee ne peut se fixer que
sur lrsquoADN des souches preacutesentant la deacuteleacutetion La
speacutecificiteacute des systegravemes de deacutetection a eacuteteacute
deacutemontreacutee in silico par alignement de
seacutequences agrave lrsquoaide du programme BLASTreg
Aucune deacutetection croiseacutee avec drsquoautres
espegraveces que celles appartenant au MTBC nrsquoa
eacuteteacute mise en eacutevidence in silico pour les systegravemes
de deacutetection utiliseacutes
Speacutecificiteacute et sensibiliteacute des systegravemes de
deacutetection sur souches pures et microcosmes
de sol inoculeacutes
Sur les 18 spoligotypes de M bovis testeacutes tous
ont donneacute un signal fort et conserveacute lors de la
PCR en temps reacuteel (Cycle threshold = 20)
Dans le complexe MTBC nous avons testeacute nos
systegravemes sur M microti M tuberculosis M
caprae et M pinnipedii Le systegraveme RD4
preacutesente la plus grande speacutecificiteacute puisqursquoil
nrsquoamplifie que M bovis parmi les membres du
complexe testeacutes LrsquoIS1561 et la reacutegion Rv3866
donnent un signal identique pour M bovis M
caprae M tuberculosis et M pinnipedii mais
ces systegravemes nrsquoamplifient pas M microti
Mecircme srsquoil a eacuteteacute montreacute que M tuberculosis
peut survivre expeacuterimentalement dans le sol
(Ghodbane et al 2014) seuls M microti et M
bovis y ont eacuteteacute deacutejagrave eacuteteacute deacutecrits [Courtenay et
al 2006 Sweeney et al 2006 et 2007]
Sur les 20 souches de MNT testeacutees cinq ont
donneacute un tregraves faible signal (Ct gt 38) avec les
trois systegravemes de deacutetection Pour le complexe
avium seul M avium a eacuteteacute deacutetecteacute avec un
Ctgt38
Aucun des 22 sols testeacutes nrsquoa donneacute de signal en
PCR en temps reacuteel quel que soit le systegraveme de
deacutetection utiliseacute ce qui indique une tregraves bonne
speacutecificiteacute de ces systegravemes de deacutetection sur
une large gamme de sols aux caracteacuteristiques
physico-chimiques diffeacuterentes et contenant des
communauteacutes microbiennes varieacutees
En ce qui concerne la sensibiliteacute des systegravemes
la limite de deacutetection des cibles se situe entre 1
et 10 copies lorsqursquoon utilise des solutions de
plasmide dilueacutees La reacutepeacutetabiliteacute de la
quantification des dilutions de plasmides est
tregraves bonne entre 10 et 106 copies de gegravenes avec
les trois systegravemes En revanche en dessous de
10 copies la deacutetection est moins reacutepeacutetable
(non deacutetection dans certaines reacutepeacutetitions)
Sur les microcosmes de sol et drsquoeau inoculeacutes la
limite de deacutetection des cibles se situe agrave 103
copies du gegravene par g de sol et agrave 10 copies du
gegravene par ml drsquoeau
Le systegraveme ciblant lrsquoIS1561 semble ecirctre tregraves
leacutegegraverement plus sensible que les deux autres
Nous lrsquoavons donc utiliseacute en 1egravere intention pour
Eacutepideacutemiol et santeacute anim 2014 65 Deacutetection de Mycobacterium bovis dans lrsquoenvironnement Premiers reacutesultats des recherches meneacutees en Cocircte-drsquoOr
229
cribler lrsquoensemble des eacutechantillons
environnementaux
Premiers reacutesultats sur les eacutechantillons
environnementaux
Sur les huit communes cibleacutees 150
eacutechantillons de sol 18 eacutechantillons de
seacutediments cinq eacutechantillons drsquoeau et huit
eacutechantillons de fegraveces de blaireaux ont eacuteteacute
preacuteleveacutes Parmi les 150 sols preacuteleveacutes 28 lrsquoont
eacuteteacute dans des terriers de blaireaux et de renards
Parmi les terriers de blaireaux preacuteleveacutes
certains eacutetaient potentiellement des terriers de
blaireaux infecteacutes par M bovis car des
animaux pieacutegeacutes agrave proximiteacute se sont reacuteveacuteleacutes ecirctre
porteurs de M bovis Cependant en 2013 nous
nrsquoavons pu obtenir aucune certitude sur les
terriers reacuteellement occupeacutes par les blaireaux
porteurs de M bovis (pieacutegeage des blaireaux en
couleacutee et non pas en gueule de terrier)
Sur les 181 eacutechantillons analyseacutes seuls deux
eacutechantillons de sol dans deux communes
diffeacuterentes ont donneacute un signal en PCR en
temps reacuteel avec lrsquoIS1561 agrave lrsquoune des dilutions
seulement Ce signal est toutefois faible et nrsquoa
pas pu ecirctre reproduit avec les autres systegravemes
de deacutetection Un des eacutechantillons positifs
provient drsquoun terrier agrave proximiteacute duquel un
blaireau infecteacute a eacuteteacute pieacutegeacute Le second
eacutechantillon de sol a eacuteteacute preacuteleveacute dans une latrine
de blaireau creuseacutee sous une clocircture de preacute
Lrsquoanalyse des fegraveces de cette latrine nrsquoa toutefois
pas donneacute de signal
La campagne de preacutelegravevement 2014 sera
reacutealiseacutee pour augmenter le nombre
drsquoeacutechantillons preacuteleveacutes sur une zone plus
restreinte en particulier ougrave des contaminations
reacutecurrentes des cheptels bovins ont eacuteteacute et sont
observeacutees
IV - Discussion
Mycobacterium bovis lrsquoagent de la tuberculose
bovine est une bacteacuterie pathogegravene qui infecte
les bovins et certains mammifegraveres sauvages en
France La maicirctrise de la maladie chez les
bovins dans certains deacutepartements franccedilais
semble ecirctre entraveacutee par la proximiteacute
drsquoanimaux sauvages infecteacutes La transmission
indirecte du bacille entre faune sauvage et
domestique par lrsquointermeacutediaire de
lrsquoenvironnement permettrait drsquoexpliquer
certains cas de re-contaminations chez les
bovins Lrsquoenvironnement pourrait servir de
reacuteservoir de la bacteacuterie par le biais de
lrsquoexcreacutetion animale il pourrait en retour
infecter des animaux sains soit par inhalation
drsquoaeacuterosols contamineacutes soit par ingestion
(abreuvement) Quelques eacutetudes en
Angleterre et en Irlande ont mis en eacutevidence la
preacutesence de M bovis dans des sols de preacutes des
sols de terriers et des latrines de blaireaux agrave
proximiteacute de cheptels bovins infecteacutes
[Courtenay et al 2006 Sweeney et al 2006
et 2007 Young et al 2005 Travis et al 2011]
Des eacutetudes expeacuterimentales ont montreacute que M
bovis pouvait survivre plusieurs mois dans le sol
[Maddock 1933 Young et al 2005 Fine et al
2011 Ghodbane et al 2014] tout en restant
virulent [Maddock 1933 Ghodbane et al
2014] La deacutetection du bacille dans
lrsquoenvironnement freacutequenteacute par les animaux
domestiques ou sauvages infecteacutes nrsquoa jamais
eacuteteacute deacutecrite en France Dans certaines zones de
Cocircte-drsquoOr la transmission indirecte du bacille
aux bovins par lrsquoenvironnement est fortement
suspecteacutee
Dans cette eacutetude nous avons deacuteveloppeacute trois
systegravemes de deacutetection moleacuteculaire afin
drsquooptimiser la mise en eacutevidence de M bovis
Eacutepideacutemiol et santeacute anim 2014 65 Deacutetection de Mycobacterium bovis dans lrsquoenvironnement Premiers reacutesultats des recherches meneacutees en Cocircte-drsquoOr
230
dans des eacutechantillons environnementaux
complexes afin de srsquoaffranchir de la culture En
effet lrsquoisolement de M bovis dans les matrices
environnementales par la culture est long car
drsquoune part un traitement preacutealable est
neacutecessaire pour srsquoaffranchir de la flore
microbienne de contamination et drsquoautre part
le bacille a une croissance tregraves lente
Contrairement agrave la culture la technique de PCR
en temps reacuteel permet une deacutetection rapide du
bacille sous reacuteserve que le ou les gegravenes cibleacutes
soient tregraves speacutecifiques de faccedilon agrave deacutetecter et
quantifier une espegravece bacteacuterienne particuliegravere
dans un meacutelange drsquoADN complexe Mais cette
technique rencontre neacuteanmoins quelques
limites techniques
Les eacutechantillons environnementaux
contiennent une flore microbienne et animale
abondante et varieacutee lrsquoADN extrait contient une
grande diversiteacute de seacutequences geacuteneacutetiques agrave
lrsquoorigine de faux positifs Ainsi des espegraveces
proches de M bovis notamment les
mycobacteacuteries environnementales ainsi que
des espegraveces encore inconnues peuvent ecirctre
preacutesentes dans les eacutechantillons Il est donc
neacutecessaire de srsquoassurer de la speacutecificiteacute des
systegravemes de deacutetection La deacutetection du bacille
par la recherche associeacutee de trois cibles
geacuteneacutetiques avec le systegraveme TaqMan nous a
permis de deacutevelopper une meacutethode de
deacutetection moleacuteculaire speacutecifique et sensible de
M bovis dans des matrices complexes
Lrsquoextraction drsquoADN de matrices environ-
nementales srsquoaccompagne drsquoune co-extraction
drsquoinhibiteurs potentiels de la PCR (acides
humiques polypheacutenols polysaccharides et
meacutetaux) Lrsquoacide humique par exemple inhibe
lrsquoactiviteacute de la Taq polymeacuterase [Kermekchiev et
al 2009] et se lie sur des sites speacutecifiques de
lrsquoADN empecircchant son amplification [Opel et al
2010] La dilution des eacutechantillons drsquoADN
utiliseacutes en PCR en temps reacuteel permet la dilution
des inhibiteurs et limite leur action Toutefois
lorsque la cible rechercheacutee est preacutesente en
petit nombre de copies la dilution de lrsquoADN
rend la deacutetection moins efficace Crsquoest
pourquoi nous analysons lrsquoADN extrait pur et
dilueacute au 15 au 110 et au 120 en PCR en
temps reacuteel de faccedilon agrave rechercher la quantiteacute
drsquoinhibiteurs minimale permettant drsquoamplifier
la cible
Les limites de deacutetection de la PCR se situent au
mieux autour de 1000 copies de gegravenes par g de
sol Les seuils de deacutetection des trois systegravemes
que nous avons creacuteeacutes se situent au-dessus de
1000 copies du gegravene par g de sol ce qui
correspond agrave des valeurs en adeacutequation avec
drsquoautres reacutesultats obtenus sur la deacutetection de
bacteacuteries dans les sols [Ibekwe et Grieve
2003]
En 2013 seulement deux des eacutechantillons
analyseacutes ont donneacute un signal faible avec un des
systegravemes de deacutetection moleacuteculaire Il srsquoagissait
dans les deux cas drsquoeacutechantillons de sol un sol
de terrier probablement habiteacute par un blaireau
infecteacute et un sol de latrine Nous nrsquoavons pu
reacutepeacuteter les reacutesultats probablement agrave cause de
la faible charge en M bovis des eacutechantillons
analyseacutes en dessous de la limite de deacutetection
de nos outils Il reste difficile drsquoisoler M bovis
de lrsquoenvironnement malgreacute la sensibiliteacute et la
speacutecificiteacute des techniques utiliseacutees Plusieurs
eacutetudes malgreacute lrsquoabondance de preacutelegravevements
reacutealiseacutes ont eacuteteacute infructueuses [Pillai et al
2000 Michel et al 2007 Fine et al 2011]
Cette difficulteacute reacutesulte des limites techniques
preacuteceacutedemment citeacutees mais eacutegalement du
caractegravere variable et intermittent de lrsquoexcreacutetion
de M bovis par les animaux infecteacutes
Lrsquoexcreacutetion de la bacteacuterie par la voie
respiratoire les urines les matiegraveres feacutecales ou
les plaies de morsure nrsquoest pas systeacutematique
chez les blaireaux infecteacutes Le niveau
drsquoexcreacutetion deacutepend de la localisation et de
lrsquointensiteacute des leacutesions Il semble que lrsquoexcreacutetion
la plus importante soit trouveacutee chez des
animaux agrave un stade avanceacute de la maladie
[Gallagher et al 1998 Corner et al 2012] En
Eacutepideacutemiol et santeacute anim 2014 65 Deacutetection de Mycobacterium bovis dans lrsquoenvironnement Premiers reacutesultats des recherches meneacutees en Cocircte-drsquoOr
231
Cocircte-drsquoOr une eacutetude meneacutee entre 2009 et
2011 a montreacute que seuls 17 des blaireaux
pieacutegeacutes et analyseacutes (19 sur 1 146) preacutesentaient
des leacutesions visibles alors que la bacteacuterie eacutetait
retrouveacutee sur 56 des blaireaux (49 sur 878)
dans la zone de tuberculose bovine [Payne et
al 2012] Lrsquoabsence de leacutesion visible sur la
majoriteacute des blaireaux analyseacutes pourrait
expliquer une absence drsquoexcreacutetion ou une
faible excreacutetion de M bovis dans
lrsquoenvironnement
Une autre difficulteacute reacuteside dans la reacutealisation
spatiale et temporelle de lrsquoeacutechantillonnage Les
bacilles sont excreacuteteacutes par les animaux sur de
petites zones bien localiseacutees Hormis les
latrines de blaireaux bien visibles les
preacutelegravevements agrave lrsquoeacutechelle drsquoune parcelle drsquoun
terrier drsquoune source sont reacutealiseacutes de maniegravere
ponctuelle et au hasard ce qui diminue la
probabiliteacute de deacutetection de cette bacteacuterie De
plus nous avons toujours un temps de
deacutecalage entre le deacutepistage des bovins ou des
animaux sauvages infecteacutes et la date des
preacutelegravevements car la certitude du diagnostic
animal neacutecessite plusieurs semaines
V - Conclusion et perspectives
Nous avons deacuteveloppeacute des outils de PCR en
temps reacuteel agrave la fois speacutecifiques et sensibles
pour la deacutetection de M bovis dans les matrices
environnementales Malgreacute le nombre
important drsquoeacutechantillons analyseacutes seuls deux
eacutechantillons peu chargeacutes en M bovis ont pu
ecirctre mis en eacutevidence Ces outils de PCR en
temps reacuteel vont encore ecirctre optimiseacutes en 2014
et permettre lrsquoanalyse des nombreux
preacutelegravevements preacutevus dans des zones plus
cibleacutees de Cocircte-drsquoOr dans lesquelles bovins et
faune sauvage infecteacutes ont eacuteteacute retrouveacutes
Plusieurs meacutethodes de culture sont en cours
drsquoessai et seront utiliseacutees sur les eacutechantillons
preacuteleveacutes afin de montrer la viabiliteacute des
bacteacuteries dans lrsquoenvironnement
Eacutepideacutemiol et santeacute anim 2014 65 Deacutetection de Mycobacterium bovis dans lrsquoenvironnement Premiers reacutesultats des recherches meneacutees en Cocircte-drsquoOr
232
Bibliographie
Aranaz A De Juan L Montero N Saacutenchez C Galka M Delso C Alvarez J Romero B Bezos J Vela A I Briones V Mateos A Domiacutenguez L - Bovine tuberculosis (Mycobacterium bovis) in wildlife in Spain Journal of clinical microbiology 2004 42 2602-2608
Betelli L Duquenne P Grenouillet F Simon X Scherer E Geacutehin E Hartmann A - Development and evaluation of a method for the quantification of airborne Thermoactinomyces vulgaris by real-time PCR Journal of microbiological methods 2013 92 25-32
Boumlhm M Hutchings MR White PCL - Contact networks in a wildlife-livestock host community identifying high-risk individuals in the transmission of bovine TB among badgers and cattle PloS One 2009 4(4) e5016
Brodin P Eiglmeier K Marmiesse M Garnier T Niemann S Cole S T Brosch R Billault A - Bacterial Artificial Chromosome-Based Comparative Genomic Analysis Identifies Mycobacterium microti as a Natural ESAT-6 Deletion Mutant Bacterial Artificial Chromosome-Based Comparative Genomic Analysis Identifies Mycobacterium microti as a Natural ESAT-6 Infection and immunity 2002 70 10 5568-5578
Colinon C Deredjian A Hien E Brothier E Bouziri L Cournoyer B Hartman A Henry S Jolivet C Ranjard L Nazaret S - Detection and enumeration of Pseudomonas aeruginosa in soil and manure assessed by an ecfX qPCR assay Journal of applied microbiology 2013 114 1734-1749
Corner L A L Murphy D Gormley E - Mycobacterium bovis infection in the Eurasian badger (Meles meles) the disease pathogenesis epidemiology and control Journal of comparative pathology 2011 144 1-24
Corner L A L OrsquoMeara D Costello E Lesellier S Gormley E - The distribution of Mycobacterium bovis infection in naturally infected badgers Veterinary journal (London England 1997) 2012 194 166-172
Courtenay O Reilly L A Sweeney F P Hibberd V Bryan S Ul-Hassan A Newman C Macdonald D W Delahay R J Wilson G J Wellington E M H - Is Mycobacterium bovis in the environment important for the persistence of bovine tuberculosis Biology letters 2006 2 460-462
Fine AE - The Role of Indirect Transmission in the Epidemiology of Bovine Tuberculosis in Cattle and White-Tailed Deer in Michigan 2006 Dissertation for the degree of Doctor of Philosophy Michigan State University
Fine A E Bolin C A Gardiner J C Kaneene J B - A Study of the Persistence of Mycobacterium bovis in the Environment under Natural Weather Conditions in Michigan USA Veterinary medicine international 2011 2011 765430
Gallagher J Clifton-Hadley R S - Tuberculosis in badgers a review of the disease and its significance for other animals Research in veterinary science 2000 69 203-217
Gallagher J Monies R Gavier-Widen M Rule B - Role of infected non-diseased badgers in the pathogenesis of tuberculosis in the badger Veterinary Record 1998 142 710-714
Gavier-Widen D Chambers M A Palmer N Newell DG Hewinson R G - Pathology of natural Mycobacterium bovis infection in European badgers (Meles meles) and its relationship with bacterial excretion Veterinary Record 2001 148(10) 299-304
Ghodbane R Mba Medie F Lepidi H Nappez C Drancourt M - Long-term survival of tuberculosis complex mycobacteria in soil Microbiology (Reading England) 2014 160(Pt 3) 496-501
Goodchild A V Clifton-Hadley R S - Cattle-to-cattle transmission of Mycobacterium bovis Tuberculosis (Edinburgh Scotland) 2001 81 23-41
Huard R C Claudio L Lazzarini D O Butler W R Soolingen D Van Ho J L - PCR-Based Method To Differentiate the Subspecies of the Mycobacterium tuberculosis Complex on the Basis of Genomic Deletions Journal of clinical microbiology 2003 41 1637-1650
Ibekwe AM Grieve CM - Detection and quantification of Escherichia coli O157H7 in environmental samples by real-time PCR Journal of Applied Microbiology 2003 94(3) 421-431
Kermekchiev M B Kirilova L I Vail E E Barnes W M - Mutants of Taq DNA polymerase resistant to PCR inhibitors allow DNA amplification from whole blood and crude soil samples Nucleic acids research 2009 37 e40
Maddock EC - Studies on the survival time of the bovine tubercle bacillus in soil soil and dung in dung and on grass with experiments on the
Eacutepideacutemiol et santeacute anim 2014 65 Deacutetection de Mycobacterium bovis dans lrsquoenvironnement Premiers reacutesultats des recherches meneacutees en Cocircte-drsquoOr
233
preliminary treatment of infected organic matter and the cultivation of the organism The Journal of Hygiene 1933 33(1) 103-117
Michel AL de Klerk LM Gey van Pittius NC Warren RM van Helden PD - Bovine tuberculosis in African buffaloes observations regarding Mycobacterium bovis shedding into water and exposure to environmental mycobacteria BMC Veterinary Research 2007 3 23
Neill S Hanna J OrsquoBrien J McCracken R - Excretion of Mycobacterium bovis by experimentally infected cattle Veterinary Record 1988 123 340-343
Opel K L Chung D McCord B R - A study of PCR inhibition mechanisms using real time PCR Journal of forensic sciences 2010 55 25-33
Palmer M V Whipple D L Waters W R Mitchell V Diana L Ray W Transmission E D - Experimental Deer-To-Deer Transmission of Mycobacterium bovis Experimental deer-to-deer transmission of Mycobacterium bovis American journal of veterinary research 2001 62 692-696
Payne A Boschiroli M L Gueneau E Moyen J L Rambaud T Dufour B Gilot-Fromont E Hars J - Bovine tuberculosis in ldquoEurasianrdquo badgers (Meles meles) in France European Journal of Wildlife Research 2012 59 331-339
Pillai S D Widmer K W Ivey L J Coker K C Newman E Lingsweiler S Baca D Kelley M Davis D S Silvy N J Adams L G - Failure to identify non-bovine reservoirs of Mycobacterium bovis in a region with a history of infected dairy-cattle herds Preventive veterinary medicine 2000 43 53-62
Ranjard L Lejon DPH Mougel C Schehrer L Merdinoglu D Chaussod R - Sampling strategy in molecular microbial ecology influence of soil sample size on DNA fingerprinting analysis of fungal and bacterial communities Environmental Microbiology 2003 5(11) 1111-1120
Richomme C Boschiroli M L Hars J Casabianca F Ducrot C - Bovine tuberculosis in livestock and wild boar on the Mediterranean island Corsica Journal of wildlife diseases 2010 46 627-631
Sweeney F P Courtenay O Ul-Hassan A Hibberd V Reilly L A Wellington E M H - Immunomagnetic recovery of Mycobacterium bovis from naturally infected environmental
samples Letters in applied microbiology 2006 43 364-369
Sweeney F P Courtenay O Hibberd V Hewinson R G Reilly L A Gaze W H Wellington E M H - Environmental monitoring of Mycobacterium bovis in badger feces and badger sett soil by real-time PCR as confirmed by immunofluorescence immunocapture and cultivation Applied and environmental microbiology 2007 73 7471-7473
Travis ER Gaze WH Pontiroli A Sweeney FP Porter D Mason S Keeling MJC Jones RM Sawyer J Aranaz A Rizaldos EC Cork J Delahay RJ Wilson GJ Hewinson RG Courtenay O Wellington EMH - An inter-laboratory validation of a real time PCR assay to measure host excretion of bacterial pathogens particularly of Mycobacterium bovis PloS One 2011 6(11) e27369
Walter W D Anderson C W Smith R Vanderklok M Averill J J Vercauteren K C - On-farm mitigation of transmission of tuberculosis from white-tailed deer to cattle literature review and recommendations Veterinary medicine international 2012 2012 616318
Young J S Gormley E amp Wellington E M H - Molecular Detection of Mycobacterium bovis and Mycobacterium bovis BCG ( Pasteur ) in Soil Applied and environmental microbiology 2005 71 1946-1952
Zanella G Durand B Hars J Moutou F Garin-Bastuji B Duvauchelle A Fermeacute M Karoui C Boschiroli M L - Mycobacterium bovis in wildlife in France Journal of wildlife diseases 2008 44 99-108
235
Annexe 2 Preacuteparation du Middlebrook 7H9
Middlebrook 7H9 suppleacutementeacute pour la culture liquide de Mycobacterium bovis
Preacuteparation de la base 7H9 (pour 500 mL de milieu)
1 Diluer 235 g de Middlebrook 7H9 (Fluka analytical) dans 450 mL drsquoeau osmoseacutee 2 Autoclaver 15 min agrave 120degC
Preacuteparation de lrsquoenrichissement ADC (pour 50 mL)
1 Peser 25 g de BSA 1 g de Dextrose et 0005 g de catalase bovine 2 Ajouter 50 mL drsquoeau osmoseacutee 3 Agiter environ 30 min (agitation magneacutetique modeacutereacutee pour eacuteviter la formation de mousse) 4 Filtrer lrsquoenrichissement sur une membrane filtrante de porositeacute 022 microM (Conservation 1 mois agrave 4degC)
Preacuteparation du Middlebrook 7H9 enrichi
1 Apregraves lrsquoautoclavage une fois le milieu refroidi ajouter 50 mL drsquoADC (conservation 1 mois agrave 4degC)
237
Annexe 3 Preacuteparation du Middlebrook 7H11
Middlebrook 7H11 suppleacutementeacute pour la culture de Mycobacterium bovis
Preacuteparation des solutions stock (tous les produits ont eacuteteacute fournis par le laboratoire Sigma-Aldrich)
Vert malachite 06 dissoudre 03 g dans 50 mL drsquoeau osmoseacutee
TicarcillineTrimeacutethoprime peser 1 g ticarcilline et 01 g de trimeacutethoprime lactate ajouter 10 mL drsquoeau osmoseacutee steacuterile Vortexer jusqursquoagrave dissolution reacutepartir en Eppendorf de 1 mL congeler (conservation 6 mois agrave -20degC)
Cycloheximide peser 500 mg et dissoudre dans 10 mL drsquoeacutethanol absolu (conservation 1 an agrave 4degC)
Polymyxine B dilution dans eau osmoseacutee steacuterile pour concentration finale de 100 000 UI par mL
Sang mouton lyseacute Meacutelanger 25 mL de sang avec 25 mL drsquoeau osmoseacutee steacuterile bain-marie 10 min agrave 60degC puis utilisation immeacutediate
Preacuteparation de la base 7H11 (pour 1 L de milieu)
1 Diluer 189 g de Middlebrook 7H11 (Fluka analytical) dans 781 mL drsquoeau 2 Ajouter 1 mL de Vert de malachite 06 3 Ajouter un barreau aimanteacute dans la bouteille 4 Autoclaver 15 min agrave 120degC
Preacuteparation de lrsquoenrichissement OADC (pour 100mL)
1 Peser 5 g de BSA 2 g de Dextrose 085 g de NaCl et 0005g de catalase bovine 2 Ajouter 100 mL drsquoeau osmoseacutee 3 Agiter environ 30 min (agitation magneacutetique modeacutereacutee pour eacuteviter la formation de mousse) 4 Ajouter 125 microL drsquoacide oleacuteique 5 Filtrer lrsquoenrichissement sur une membrane filtrante de porositeacute 022 microM (Conservation 1 mois agrave 4degC)
Preacuteparation du Middlebrook 7H11 enrichi
1 Apregraves lrsquoautoclavage porter le milieu agrave 45-50degC 2 Ajouter 100 mL de seacuterum bovin inactiveacute (agitation magneacutetique) 3 Ajouter 100 mL drsquoOADC (agitation magneacutetique) 4 Ajouter 5 mL de sang de mouton lyseacute 5 Ajouter 1 mL de TicarcillineTrimeacutethoprime 2 mL de Polymyxine B 2 mL de cycloheximide
(agitation magneacutetique) (facultatif si ensemencement de cultures pures)(Concentrations finales respectives 100 mgL 10 mgL 200 000 UIL et 100 mgL)
6 Reacutepartition de 25 mL de milieu par boite (Conservation 1 mois agrave 4degC)
Reacutesumeacute La tuberculose bovine est une maladie infectieuse contagieuse causeacutee par Mycobacterium bovis Cette maladie touche les bovins
et de nombreuses espegraveces de mammifegraveres domestiques et sauvages ainsi que lrsquohomme La circulation de la bacteacuterie dans des systegravemes
multi-hocirctes varieacutes favorise lrsquoentretien de la maladie et la contamination des bovins vivant agrave proximiteacute des animaux sauvages infecteacutes En
marge de la transmission directe de M bovis par voie respiratoire la transmission indirecte aux bovins lieacutee agrave lrsquoinhalation ou agrave lrsquoingestion de
matrices environnementales contamineacutees par un animal infecteacute excreacuteteur est suspecteacutee dans plusieurs reacutegions du monde Lrsquoexistence de
reacuteservoirs environnementaux ougrave le bacille M bovis est capable de persister pourrait donc ecirctre un facteur important de la reacuteeacutemergence puis
du maintien de la maladie dans les systegravemes multi-hocirctes
En Cocircte drsquoOr deacutepartement fortement toucheacute par la tuberculose bovine depuis 2004 la transmission indirecte de la bacteacuterie entre la faune
sauvage infecteacutee et les bovins est suspecteacutee dans plusieurs eacutelevages Pour eacutevaluer la preacutesence et la survie de cette bacteacuterie dans
lrsquoenvironnement nous avons analyseacute un grand nombre drsquoeacutechantillons preacuteleveacutes dans des zones partageacutees par les bovins etou la faune sauvage
infecteacutes dans le but de deacuteterminer la distribution environnementale de M bovis Pour ce faire nous avons deacuteveloppeacute ou modifieacute des systegravemes
de deacutetection moleacuteculaire adapteacutes aux matrices environnementales complexes Nous avons eacutegalement eacutevalueacute lrsquoimpact de la tempeacuterature et
des proprieacuteteacutes physico-chimiques de deux sols sur la survie de M bovis ainsi que le rocircle de la meacutesofaune du sol (lombrics en particulier)
dans la disseacutemination de la bacteacuterie agrave partir de matiegravere organique contamineacutee Lrsquoeacutetude environnementale a mis plus particuliegraverement en
eacutevidence la contamination de deux biotopes les zones humides des pacirctures et les sols de terriers de blaireaux De plus les eacutetudes
expeacuterimentales ont montreacute que M bovis pouvait survivre plusieurs mois dans le sol agrave 4degC et que les lombrics pouvaient disseacuteminer la bacteacuterie
dans le sol voire jouer un rocircle potentiel de vecteur pour les animaux qui les consomment Ces reacutesultats apportent de nouvelles connaissances
sur la persistance et la circulation de M bovis dans lrsquoenvironnement en Cocircte drsquoOr et permettront de proposer des ameacuteliorations aux mesures
de bioseacutecuriteacute deacutejagrave existantes dans les eacutelevages bovins
Mots Cleacutes Mycobacterium bovis environnement sol eau fegraveces bovins faune sauvage qPCR culture
Abstract Bovine tuberculosis is an infectious disease caused by Mycobacterium bovis This disease affects multiple species of domestic
livestock notably cattle wild mammals and humans The circulation of the bacteria within complex multi-host systems promotes the
maintenance of the disease and the reinfection of cattle Besides the classical direct transmission of the bacteria through the respiratory
route indirect transmission through inhalation or ingestion of environmental substrates contaminated by the excretions of infected animals
is suspect in several countries Environmental contamination with M bovis appears to be a crucial factor in the persistence of the infection in
multi-host systems
In Cocircte dOr a French department impacted by bovine tuberculosis since 2004 the indirect transmission of the bacteria from infected wildlife
to cattle is now suspect in several cases To assess this mode of transmission we evaluated environmental contamination with M bovis on a
large number of samples collected in areas shared by cattle and infected wildlife For this purpose we both developed and modified molecular
detection systems adapted for complex environmental substrates We also assessed the impact of temperature and soil physicochemical
properties on the survival of M bovis as well as the role of earthworms in the dissemination of the bacteria from contaminated organic
material The environmental study showed contamination of two habitats in particular wetland pastures and soil badger setts Moreover
results indicated that M bovis can survive in soil for several months at 4degC and that worms may disseminate the bacteria in the soil and even
serve as a vector for the mammals that consume them This study increases our understanding of the persistence and the circulation of
M bovis in the environment in Cocircte dOr In addition these results will be used to improve the existing biosecurity measures on cattle farms
in the region
Key words Mycobacterium bovis environment soil water feces cattle wildlife qPCR culture