employment of tuberculostasis in serum-agar medium for the study
TRANSCRIPT

INFECTION AND IMMUNITY, Aug. 1971, p. 130-137Copyright ( 1971 American Society for Microbiology
Vol. 4, No. 2Printed in U.S.A.
Employment of Tuberculostasis in Serum-AgarMedium for the Study of Productioni and
Activity of MycobactinIVAN KOCHAN, DAVID L. CAHALL, AND CAROLE A. GOLDEN
Departinetit of Microbiology, Miami University, Oxford, Ohio 45056
Received for publication 15 April 1971
Mycobactin (M), an iron-chelating product of tubercle bacilli, neutralized serum
tuberculostasis by removing growth-essential iron from transferrin (Tr) and supply-ing the metal to the bacteria. The competition for iron between Tr and M has beendemonstrated by the agar-plate diffusion test. This test is suitable not only for thestudy of Tr-iron-M interplay but also for the evaluation of serum tuberculostasis.Extremely poor solubility of M in water and consequently its association with li-poidal cell wall of tubercle bacillus was overcome by the use of water-dispersible andsurface-active Tween 80. The addition of Tween 80 to culture media insured thepresence of M in spent media; otherwise M was extracted from bacillary cells witha solution of Tween 80 or a mixture of ethanol and Tween 80. Although M was pro-duced irrespective of the amount of iron present in culture medium, its productionin iron-poor medium was more prolific than in iron-rich medium. M extracted fromBCG or H37Rv cells neutralized serum tuberculostasis as effectively for the homolo-gous as for heterologous strains. However, the extract of virulent bacilli was muchmore active in the neutralization than similar extract prepared from attenuatedcells; whether this difference is of quantitative or qualitative nature remains to bedetermined.
When tubercle bacilli are exposed to mamma-lian serum, their multiplication stops because theserum fails to provide the parasite with growth-essential iron (6, 7). Serum iron complexed withiron-chelating protein transferrin (Tr) is unavail-able for the utilization by tubercle bacilli unlessthis Tr-iron union is disturbed by a bacillary prod-uct known as mycobactin (M). Possession of avery strong iron-chelating property enables M toremove iron from Tr and make the metal availablein utilizable form to the bacilli (9).The production ofM has been considered as an
emergency mechanism triggered in the growingbacilli by subjecting them to a stress of iron defi-ciency (11). It is possible, however, that M isneeded for the transport of iron into bacillary cellat all times, and, therefore, it may be producedirrespective of the amount of iron present in theculture medium. There may exist only a quantita-tive difference: in an iron-deficient medium moreM is produced than in an iron-rich medium. Thispossibility we have studied by the use of a tech-nique which we found to be most suitable notonly for the detection of M or the study of itsgrowth-promoting role in tuberculostatic serum
but also for the demonstration and titration ofserum tuberculostasis.
MATERIALS AND MIETHODSMethods used in our previous studies of the Tr-
iron-M interplay in serum tuberculostasis consumedmuch of our time and effort (6-9). Usually, results ofan experiment were obtained after 6 weeks of work andwaiting. Tubercle bacilli were exposed to experimentalserum mixtures for 2 weeks, and then samples in var-ious dilutions were plated on bacillary growth-support-ing medium; after 3 to 4 weeks of incubation, tlhenumbers of bacilli on plates were determined by count-ing colonies. Recently we developed a technique whichsignificantly reduced time and work in the study ofserum tuberculostasis and associated Tr-iron-M inter-play. Since the adoption of the technique constitutes amajor change in our work, the first part of this reportdeals with the reliability and sensitivity of the tech-nique in the demonstration and titration of serumtuberculostasis; the second part presents results ob-tained in the study of the production and activityof M.
Agar-plate diffusion test. The demonstration, titra-tion, and neutralization of serum tuberculostasis wereperformed in 60- by 15-mm plastic petri plates (no.3002, Falcon Plastic) which were filled with serum-
130
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 27
Jan
uary
202
2 by
139
.47.
78.2
34.

MYCOBACTIN IN SERUM-AGAR MEDIUM
agar medium. The medium consisted of tuberculostaticserum diluted 1:4 in iron-void Dubos agar medium(3).
Hemolysis-free serum samples were collected afterthe clotting of blood obtained from cows in a localslaughter house. Sera were sterilized by filtrationthrough bacteria-retaining Seitz filters. Chemicalanalysis was performed to determine the serum ironlevel (SI) and the total iron-binding capacity (TIBC)for each serum sample (10). The unbound iron-bind-ing capacity (UIBC), a value which in this study indi-cates the amount of added iron required to saturate Trin 100 ml of undiluted serum, was calculated by sub-tracting SI from TIBC. Since bacillary multiplicationdepends upon availability of iron (6), the value forUIBC indicated the approximate amount of iron whichwas required for the neutralization of serum tuber-culostasis.
The iron-void Dubos agar medium was prepared indouble-distilled water to which all ingredients of themedium were added except ferric ammonium citrate.Although efforts were made to prevent iron contam-ination of the medium, iron determinations showedthat 0.09 jig of the metal was present in 1 ml of themedium. In the absence of tuberculostatic serum, thisminute amount of iron is sufficient to promote a fairlygood bacillary growth on the surface of the medium.The mixture of 3 ml of serum and 9 ml of iron-void
Dubos agar medium was prepared separately for eachplate in a 1 5-ml tube. To prevent solidification of agar,the liquefied medium and serum before and after themixing were kept in a water bath at 56 C. The smallamount of contaminating iron in the iron-void Dubosmedium was bound to Tr of the serum during incuba-tion of the mixture for 30 min at 56 C. After the incu-bation period, a horizontally leveled plate received 2ml of the serum-medium which was allowed to solidifyat room temperature. A well (1 cm in diameter) wasmade in the center of the plate by putting an acid-cleaned, siliconized glass cylinder on the preparedlayer of gelled medium and then adding to the platethe remaining 10 ml of the medium. After solidifica-tion of agar, the surface of the serum-medium wasinoculated with a 0.25-ml suspension of washed BCGstrain of Mycobacterium bovis adjusted to 1 Klett unitin saline. The bacillary inoculum was spread uniformlyover the entire surface of the medium. Plates wereincubated in a drying chamber at 37 C for 24 hr toevaporate the water of the inoculum. After the incuba-tion, the glass cylinders were removed with sterileforceps and the wells were filled with 0.4 ml of salinecontaining materials tested for the antagonistic effecton serum tuberculostasis. The inoculated plates withcharged wells were incubated at 37 C in a high humid-ity chamber for a 2- to 3-week period.The degree of neutralization of serum tuberculo-
stasis around wells was determined by measuring thearea covered by bacillary growth. Results were re-corded photographically by taking pictures of platesplaced on a template which had a scale indicating0.5-cm intervals.
Mycobactin. Our previous results (9) showed that M
promotes bacillary multiplication in tuberculostaticserum by removing iron from Tr and supplying themetal to tubercle bacilli. Usually the presence of Mwas most readily detected on bacillary cells grown iniron-poor medium (11). A possibility exists, however,that M is needed for the iron transport at all times,and, therefore, it may be present on bacillary cell sur-faces irrespective of the amount of iron present in thegrowth medium. This possibility, as well as questionspertaining to the production, excretion, and the spec-ificity ofM were investigated in this study.
Attenuated bovine strain BCG of M. bovis andvirulent human strain H37Rv of M. tuberculosis weregrown for 6 weeks at 37 C in various iron-rich andiron-poor media in the presence or absence of Tween80. The cultures were killed during a 1-hr exposure to100 C, and the heat-killed bacilli were separated fromspent media by filtration through a Whatman no. 3filter paper in a Buchner funnel. Bacilli were washedwith saline free of medium. The extraction of M fromthe washed bacillary cells was done by suspending 1 gof moist-weight bacilli in 10 ml of 70% ethanol andmixing the suspension with a Teflon-enclosed magneticstirrer during a 24-hr extraction period. After the ex-traction, bacilli were separated from ethanol extractby centrifugation and filtration through a bacteria-retaining Seitz filter. Alcohol was evaporated at 100 C,and the remaining 3 ml of water contained poorlywater-soluble materials. After we found that M iseasily solubilized in the presence of Tween 80, extrac-tion of soluble material was performed by the use ofalcohol containing 0.05%,' of Tween 80. It will beshown later that the presence of Tween 80 not onlyhelps in the solubilization of M (and probably otherethanol-extractable bacillary materials) but also in theremoval of M from the bacillary cells. The extract,after the evaporation of alcohol, contained 0.3 ,g ofiron per 1 ml of the solution. The serum tuberculo-stasis neutralizing quality of extracts obtained fromBCG and H37Rv cells and of spent media was eval-uated by the agar-plate diffusion test.
In some experiments a solution of purified M wasused to assess its tuberculostasis neutralizing activityand to compare this activity to that of BCG andH37Rv cell extracts. This pure, iron-free M was ex-tracted from cells of M. phlei by the method describedby Francis et al. (5) and was generously supplied forthis study by G. A. Snow.
RESULTS
Reliability and sensitivity of the agar-plate dif-fusion test. Previous experiments performed intubes containing tuberculostatic sera showed thattuberculostasis can be neutralized with iron or M(9). In the present study, this antagonistic activityof iron and M against serum tuberculostasis wastested by the use of the agar-plate diffusion test.Tuberculostatic bovine serum diluted 1:4 in iron-void Dubos agar medium served as a medium onwhich BCG cells were spread uniformly. The wells
VOL. 3, 1971 131
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 27
Jan
uary
202
2 by
139
.47.
78.2
34.

KOCHAN, CAHALL, AND GOLDEN
were filled with various concentrations of iron orpurified M.
Figure 1 shows that with each increasing con-centration of iron or M, an increased surface areacovered by bacillary growth was observed aroundthe wells. The correlation between the amounts ofthe bacillary growth promoters and the areascovered by bacillary growth was determined by amethod devised for the study of antibiotic activityin plate diffusion assays (1). When logarithmicconcentrations of iron and M were plotted againstrespective diameters of bacillary growth zones,two parallel straight lines were obtained. Thegood correlation between the quantities of growthpromoters in the wells and corresponding areascovered by bacillary growth indicates that theagar-plate diffusion test is a reliable assay whichcan be used for the evaluation of serum tuber-culostasis.The bacillary growth promoted by iron differed
from that induced by M (Fig. 1). Around ironwells the growth was heavy and elevated withrather sharply defined edges, whereas the growtharound M wells was more spread and poorlydelineated. It seems that the presence of M in themedium stimulates the formation of BCG colonies
which are very similar to colonies of cord-pro-ducing virulent strains.
It was of considerable interest to determine ifthe agar-plate diffusion test was sensitive enoughto evaluate the degree of tuberculostasis in varioussera, and, if so, if the strength of tuberculostasisin a serum could be predicted on the basis of itsUIBC value. Two bovine sera, one with 410 andthe other with 224 value for UIBC, were tested bythe agar-plate diffusion test for the tuberculo-static activity in the presence of various concentra-tions of iron (Fig. 2). Plates whose wells werefilled with iron-void saline solution showed nobacillary growth; wells which received ironshowed bacillary multiplication around them, andthe area covered by the growth increased with theincreased concentration of iron. At each iron con-centration, plates containing medium with thelower UIBC (serum B) showed more growth thanthose with high UIBC (serum A). In a similarstudy 10 normal human sera were tested for thecorrelation between the strength of their tuber-culostasis and UIBC values. Most sera had UIBCvalues around 250 and showed similar growth-supporting ability in the presence of 5 ug of iron.Some sera had different values for UIBC and thei
IRON
MYCOBACTIN
0.6 2.5 10.0 40.0pg/WELL
FIG. 1. Bacillary growth oii tufberculostatic bovine serum-agar medium around wells wvhich received 0.6, 2..,10.0, and 40.0 ,ug of iron or mycobactin. The unbound iron-binding capacity of serum was 380.
132 INFEC. IMMUN.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 27
Jan
uary
202
2 by
139
.47.
78.2
34.
MYCOBACTIN IN SERUM-AGAR MEDIUM
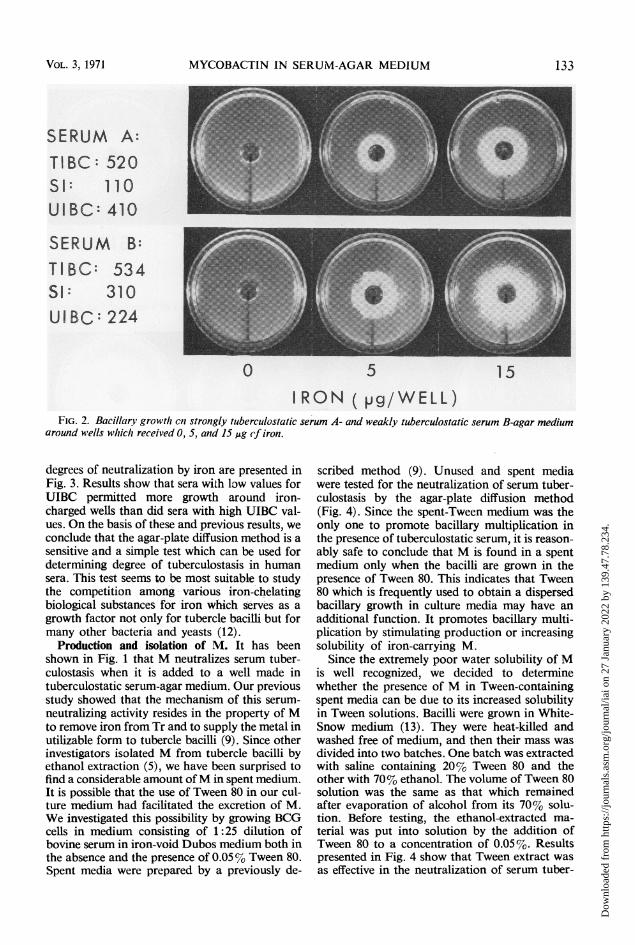
SERUM A:TIBC: 5205 1: 110UIBC: 410
SERUM B:TI BC: 534 E\UIBC: 224
0 5 15IRON (pg/WELL)
FIG. 2. Bacillary growtlh Ce strongly tuberculostatic serum A- and weakly tuberculostatic serum B-agar mediumaround wells which received 0, 5, and 15 Aig of iron.
degrees of neutralization by iron are presented inFig. 3. Results show that sera with low values forUIBC permitted more growth around iron-charged wells than did sera with high UIBC val-ues. On the basis of these and previous results, weconclude that the agar-plate diffusion method is asensitive and a simple test which can be used fordetermining degree of tuberculostasis in humansera. This test seems to be most suitable to studythe competition among various iron-chelatingbiological substances for iron which serves as agrowth factor not only for tubercle bacilli but formany other bacteria and yeasts (12).
Production and isolation of M. It has beenshown in Fig. 1 that M neutralizes serum tuber-culostasis when it is added to a well made intuberculostatic serum-agar medium. Our previousstudy showed that the mechanism of this serum-neutralizing activity resides in the property of Mto remove iron from Tr and to supply the metal inutilizable form to tubercle bacilli (9). Since otherinvestigators isolated M from tubercle bacilli byethanol extraction (5), we have been surprised tofind a considerable amount ofM in spent medium.It is possible that the use of Tween 80 in our cul-ture medium had facilitated the excretion of M.We investigated this possibility by growing BCGcells in medium consisting of 1:25 dilution ofbovine serum in iron-void Dubos medium both inthe absence and the presence of 0.05% Tween 80.Spent media were prepared by a previously de-
scribed method (9). Unused and spent mediawere tested for the neutralization of serum tuber-culostasis by the agar-plate diffusion method(Fig. 4). Since the spent-Tween medium was theonly one to promote bacillary multiplication inthe presence of tuberculostatic serum, it is reason-ably safe to conclude that M is found in a spentmedium only when the bacilli are grown in thepresence of Tween 80. This indicates that Tween80 which is frequently used to obtain a dispersedbacillary growth in culture media may have anadditional function. It promotes bacillary multi-plication by stimulating production or increasingsolubility of iron-carrying M.
Since the extremely poor water solubility ofMis well recognized, we decided to determinewhether the presence of M in Tween-containingspent media can be due to its increased solubilityin Tween solutions. Bacilli were grown in White-Snow medium (13). They were heat-killed andwashed free of medium, and then their mass wasdivided into two batches. One batch was extractedwith saline containing 20% Tween 80 and theother with 70% ethanol. The volume of Tween 80solution was the same as that which remainedafter evaporation of alcohol from its 70% solu-tion. Before testing, the ethanol-extracted ma-terial was put into solution by the addition ofTween 80 to a concentration of 0.05%. Resultspresented in Fig. 4 show that Tween extract wasas effective in the neutralization of serum tuber-
VOL. 3, 1971 133
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 27
Jan
uary
202
2 by
139
.47.
78.2
34.
KOCHAN, CAHALL, AND GOLDEN
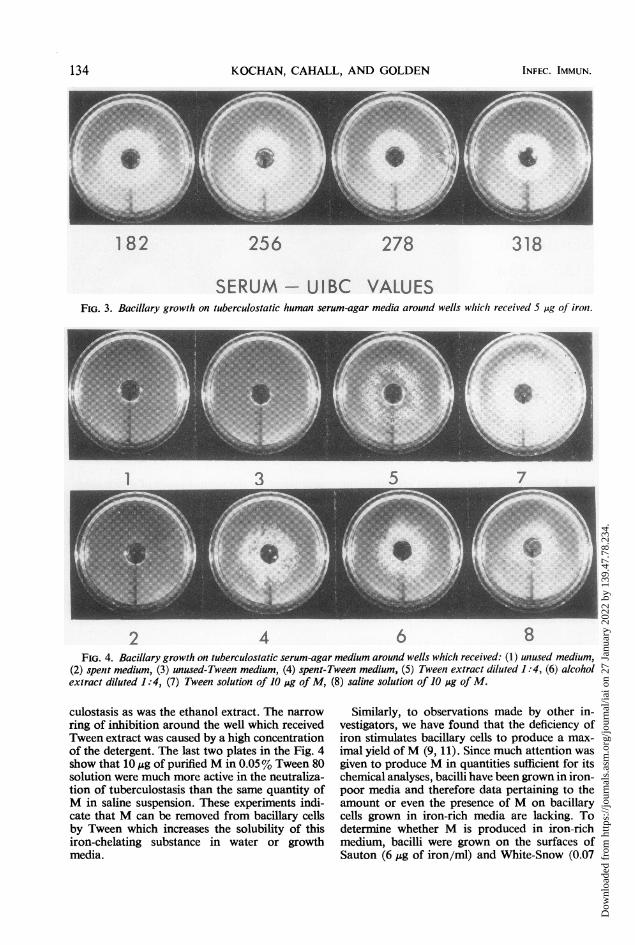
182 256 278 318
SERUM - UIBC VALUESFIG. 3. Bacillary growlh on tuberculostatic human serum-agar media arountd wells which received 5 ,ug of iron.
5 71
2 4 6 8FIG. 4. Bacillary growth on tuberculostatic serum-agar medium around wells which received: (1) unused medium,
(2) spent medium, (3) unused-Tween medium, (4) spent-Tween medium, (5) Tween extract diluted 1:4, (6) alcoholextract diluted 1:4, (7) Tween solution of 10 jig of M, (8) saline solution of 10 pig of M.
culostasis as was the ethanol extract. The narrowring of inhibition around the well which receivedTween extract was caused by a high concentrationof the detergent. The last two plates in the Fig. 4show that IO,g of purified M in 0.05% Tween 80solution were much more active in the neutraliza-tion of tuberculostasis than the same quantity ofM in saline suspension. These experiments indi-cate that M can be removed from bacillary cellsby Tween which increases the solubility of thisiron-chelating substance in water or growthmedia.
Similarly, to observations made by other in-vestigators, we have found that the deficiency ofiron stimulates bacillary cells to produce a max-imal yield ofM (9, 11). Since much attention wasgiven to produce M in quantities sufficient for itschemical analyses, bacilli have been grown in iron-poor media and therefore data pertaining to theamount or even the presence of M on bacillarycells grown in iron-rich media are lacking. Todetermine whether M is produced in iron-richmedium, bacilli were grown on the surfaces ofSauton (6 Mig of iron/ml) and White-Snow (0.07
134 INFEC. IMMUN.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 27
Jan
uary
202
2 by
139
.47.
78.2
34.
MYCOBACTIN IN SERUM-AGAR MEDIUM
SAUTON -
CE LS
EXTRACT
CELLSEXTRACT
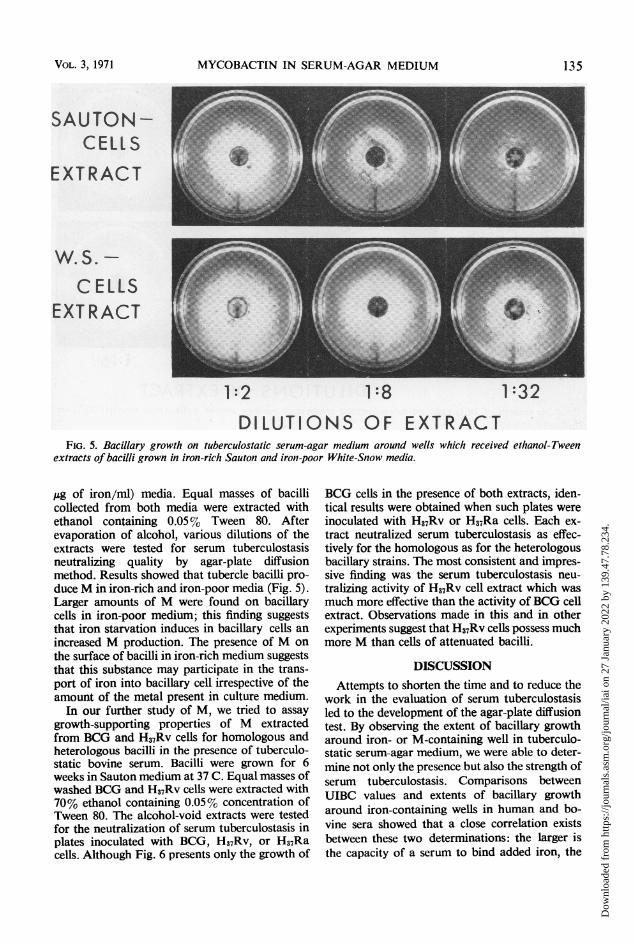
1:2 1:8 1:32
DILUTIONS OF EXTRACTFIG. 5. Bacillary growth on tuberculostalic serum-agar medium around wells which received ethanol-Tween
extracts of bacilli grown in iron-rich Sauton and iron-poor White-Snow media.
Ag of iron/ml) media. Equal masses of bacillicollected from both media were extracted withethanol containing 0.05% Tween 80. Afterevaporation of alcohol, various dilutions of theextracts were tested for serum tuberculostasisneutralizing quality by agar-plate diffusionmethod. Results showed that tubercle bacilli pro-duce M in iron-rich and iron-poor media (Fig. 5).Larger amounts of M were found on bacillarycells in iron-poor medium; this finding suggeststhat iron starvation induces in bacillary cells anincreased M production. The presence of M onthe surface of bacilli in iron-rich medium suggeststhat this substance may participate in the trans-port of iron into bacillary cell irrespective of theamount of the metal present in culture medium.
In our further study of M, we tried to assaygrowth-supporting properties of M extractedfrom BCG and H37Rv cells for homologous andheterologous bacilli in the presence of tuberculo-static bovine serum. Bacilli were grown for 6weeks in Sauton medium at 37 C. Equal masses ofwashed BCG and H37Rv cells were extracted with70% ethanol containing 0.05% concentration ofTween 80. The alcohol-void extracts were testedfor the neutralization of serum tuberculostasis inplates inoculated with BCG, H37Rv, or H37Racells. Although Fig. 6 presents only the growth of
BCG cells in the presence of both extracts, iden-tical results were obtained when such plates wereinoculated with H37Rv or H37Ra cells. Each ex-tract neutralized serum tuberculostasis as effec-tively for the homologous as for the heterologousbacillary strains. The most consistent and impres-sive finding was the serum tuberculostasis neu-tralizing activity of H37Rv cell extract which wasmuch more effective than the activity of BCG cellextract. Observations made in this and in otherexperiments suggest that H37Rv cells possess muchmore M than cells of attenuated bacilli.
DISCUSSIONAttempts to shorten the time and to reduce the
work in the evaluation of serum tuberculostasisled to the development of the agar-plate diffusiontest. By observing the extent of bacillary growtharound iron- or M-containing well in tuberculo-static serum-agar medium, we were able to deter-mine not only the presence but also the strength ofserum tuberculostasis. Comparisons betweenUIBC values and extents of bacillary growtharound iron-containing wells in human and bo-vine sera showed that a close correlation existsbetween these two determinations: the larger isthe capacity of a serum to bind added iron, the
VOL. 3, 1971 135
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 27
Jan
uary
202
2 by
139
.47.
78.2
34.

KOCHAN, CAHALL, AND GOLDEN
BCG-CE LLSEXTRACT
H37 Rv-CELLSEXTRACT
UNDIL. 1:4 1:16
DILUTIONS OF EXTRACTtuberculostatic serum-agar medium arounzd wells which received BCG or
smaller is the growth-supporting area around theiron-containing well.Although the close correlation between UIBC
values and the strength of tuberculostasis in sera
of normal animals has been well established, it isnot definite that the UIBC value alone could givea predictable degree of serum tuberculostasis insera of tuberculous patients. IfM were present intuberculous sera, it would remove iron from Trand consequently these sera would show a highUIBC value but at the same time be weakly or
nontuberculostatic. The comparison betweenUIBC values and tuberculostatic activities in sera
of tuberculous patients and the same values insera of normal individuals should enable one todetermine whether or not M is present in theblood of patients.The beneficial role of serum tuberculostasis in
native resistance of animals to infection withtubercle bacilli is suggested by circumstantial evi-dence. For instance, guinea pigs (whose blood haslittle or no tuberculostatic activity) are most sus-
ceptible to infection with tubercle bacilli, whereashumans (with strong tuberculostatic serum) are
quite resistant. The question is why men, in spiteof the tuberculostasis in their sera, succumb to theinfection with tubercle bacilli. Ten years ago,
while investigating tuberculostasis in various"racial sera," we found that about 10% of humansera demonstrated a low degree of tuberculostaticactivity (8). This finding as well as the results
presented in Fig. 3 show that normal individualspossess varying degrees of tuberculostasis in theirsera which could be a determining factor in ex-
plaining their varying susceptibilities to infectionwith tubercle bacilli.
Bacterial products which enable the survivaland the spread of parasites in a host are consid-ered as virulence factors. Factors which enablemultiplication of tubercle bacilli in an infectedhost are not known. Since M overcomes serum
tuberculostasis and ensures bacillary survival inserum, it may be considered as a good candidatefor recognition as a bacillary virulence factor. Be-cause of this consideration, much effort was andis devoted in our laboratory to study the produc-tion and the mode of operation of M.
Results presented in this report show that M ispresent on bacillary cells irrespective of theamount of iron present in the culture medium.However, more M was produced in iron-poor thanin iron-rich medium. This compensatory response
to iron starvation, the permanent presence of Mon bacillary cells, and the inability of other iron-carrying biological substances to supply bacilliwith iron (unpublished results) suggest that M notonly makes iron available for bacillary utilizationbut also participates in the transport of iron intobacillary cells.
Probably because of its poor water solubility,M remains associated with bacillary cells. Resultspresented in this report show that M was not
FIG. 6. Growth of BCG cells onH37Rv cell extracts.
INFEC. IMMUN.136
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 27
Jan
uary
202
2 by
139
.47.
78.2
34.

MYCOBACTIN IN SERUM-AGAR MEDIUM
found in spent medium unless bacilli were cul-tured in the presence of Tween 80; 0.05% of thisdetergent in a medium was sufficient to causesome release of M. Thus, the presence of Tween80 in medium devised for the growth of tuberclebacilli insures better bacillary multiplication notonly by producing a dispersed growth but also byincreasing solubility of M. In Tween-containingmedium, M is soluble in spite of its hydrophobicnature and fulfills its iron-transporting functionin an efficient way.
This study showed that M extracted either fromBCG or H37Rv cells is as effective in the neutrali-zation of serum tuberculostasis for homologousas for heterologous strains of tubercle bacilli.However, considerably more M was extractedfrom the same quantity of virulent than attenu-ated bacilli. Preliminary results showed that thesecond ethanol extraction of virulent cells yieldsas much M as the first extraction of the attenuatedcells. This persistence ofM on virulent cells couldeither be due to a large amount or a firm associa-tion with these cells.
It has been observed by Dubos and Middle-brook that in medium containing Tween 80, viru-lent and attenuated bacilli grow without the pro-duction of cords, and therefore, their growthcannot be differentiated from that produced bytotally avirulent cells (4). Triton A20, anotherwater-dispersible, nonionic, surface-active agent,inhibited the growth of avirulent bacilli while per-mitting virulent strains to multiply. It has beensuggested that virulent cells differ from avirulentcells by possessing on their surfaces a largeramount of certain hydrophobic substance and,therefore, are more resistant to the activity ofTriton. Since the surface-active substances had aneffect not only on the colonial morphology butalso on the survival of tubercle bacilli, it washoped that the study of the hydrophobic surfacefactors would yield a better understanding of themechanism of infection and immunity in tuber-culosis (2).
Results presented in this study are pertinent tothe above observations and may shed a new lighton factors associated with bacillary surface whichenable tubercle bacillus to behave as a parasite.We found that, depending upon the concentrationof Tween 80, M can either be released or removedfrom the surface of bacillary cells. The addition of
purified M to the medium of agar-plate diffusiontest induced the formation of BCG colonies whichwere thinly spread on the surface of agar and uni-formly exhibited a definite serpentine pattern ofgrowth. In the absence of M, the BCG colonieswere elevated, smooth, and only with an occa-sional indication of some cord formation. ThisM-induced difference in the colonial morphology,the extraction of M with Tween 80, and the pres-ence of large amount of M on virulent cells indi-cate that M could be one of the lipoidal factorsdescribed previously by Dubos and Middlebrook(4).
ACKNOWLEDGMENTS
We thank G. A. Snow for providing mycobactin P for thisstudy and E. D. Weinberg for helpful critiques of the manuscriptduring its preparation.
This investigation was supported by grant GB-241.14 from theNational Science Foundation and by funds provided by the Com-mittee on Faculty Research, Miami University.
LITERATURE CITED
l. Cooper, K. E. 1963. The theory of antibiotic inhibition zones,p. 1-86. In F. Kavanagh (ed.), Analytical microbiology,Academic Press Inc., New York.
2. Dubos, R. J. 1949. The tubercle bacillus and tubercuiosis.Amer. Scientist 3i:353-370.
3. Dubos, R. J., and G. Middlebrook. 1947. Media for tuberclebacilli. Amer. Rev. Tuberc. 56:334-345.
4. Dubos, R. J., and G. Middlebrook. 1948. The effect of wettingagents on the growth of tubercle bacilli. J. Exp. Med.88:81-88.
5. Francis, J., H. M. Macturk, J. Madinaveitia, and G. A. Snow.1953. Mycobactin, a growth factor for Mycobacteriumjohliei. I. Isolation from Mvcobacterium phlei. Biochem. J.55:596-607.
6. Kochan, I. 1969. Mechanism of tuberculostasis in mam-malian serum. I. Role of transferrin in human serumii tuber-culostasis. J. Infec. Dis. 119:11-18.
7. Kochan, I., C. A. Golden, and J. A. Bukovic. 1969. Mecha-nism of tuberculostasis in mammalian serumn. II. Inductionof serum tuberculostasis in guinea pigs. J. Bacteriol. 100:64-70.
8. Kochan, I., C. Patton, and K. Ishak. 1963. Tuberculostaticactivity of normal human sera. J. Immunol. 90:711-719.
9. Kochan, I., N. R. Pellis, and C. A. Golden. 1971. Mechanismof tuberculostasis in mammalian serum. III. Neutralizationof serum tuberculostasis by mycobactin. Infec. Immun.3:553-558.
10. Landers, J. W., and B. Zak. 1958. Determination of serumcopper and iron in a single small sample. Amer. J. Clin.Pathol. 29:590-594.
11. Snow, G. A. 1970. Mycobactins: iron-chelating growth factorsfrom mycobacteria. Bacteriol. Rev. 34:99-125.
12. Weinberg, E. D. 1966. Roles of metallic ions in host-parasiteinteractions. Bacterio!. Rev. 30:136-151.
13. White, A. J., and G. A. Snow. 1968. Methods for the separa-tion and identification of mycobactins from various speciesof mycobacteria. Biochem. J. 108:593-597.
VOL. 3, 1971 137
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 27
Jan
uary
202
2 by
139
.47.
78.2
34.