endomorphin-2 in the medial nts attenuates the responses to baroreflex activation
TRANSCRIPT

B R A I N R E S E A R C H 1 0 7 3 – 1 0 7 4 ( 2 0 0 6 ) 3 6 5 – 3 7 3
ava i l ab l e a t www.sc i enced i rec t . com
www.e l sev i e r. com/ l oca te /b ra in res
Research Report
Endomorphin-2 in the medial NTS attenuatesthe responses to baroreflex activation
Eddy Viard, Hreday N. Sapru⁎
Department of Neurological Surgery, MSB H-586, University of Medicine and Dentistry of New Jersey-New Jersey Medical School, 185 SouthOrange Ave., Newark, NJ 07103, USA
A R T I C L E I N F O
⁎ Corresponding author. Fax: +1 973 972 5986.E-mail address: [email protected] du (H.N. S
0006-8993/$ – see front matter © 2005 Elsevidoi:10.1016/ j.brainres.2005.12.10 2
Available online 3 February 2006
A B S T R A C T
Article history:Accepted 27 December 2005
We have previously reported that microinjections of endomorphin-2 (E-2; an endogenousmu-receptor agonist) into the medial subnucleus of the NTS (mNTS) elicit depressor andbradycardic responses via activation of ionotropic glutamate receptors located on secondarymNTS-neurons. Based on this report, it was hypothesized that activation of secondarymNTS neurons by E-2 may result in an exaggeration of baroreflex responses. In order to testthis hypothesis, baroreflex responses were studied in adult, urethane-anesthetized,artificially ventilated, male Wistar rats before and after the microinjections of E-2 into themNTS. Baroreceptors were stimulated by applying pressure increments (80–100 mm Hg) inthe carotid sinus and by electrical stimulation (stimulus intensity: 0.5 V, frequencies 5, 10,and 25 pulses/s, pulse duration: 1 ms) of the aortic nerve for 30-s periods. Baroreceptorstimulation elicited depressor and bradycardic responses. Microinjections (100 nl) of E-2 (0.4mmol/l) into the mNTS attenuated the baroreflex responses. Microinjections of naloxone(an opioid receptor antagonist) into the mNTS (0.5 mmol/l) did not alter baroreflexresponses. Based on these results, it was concluded that activation ofmu-opioid receptors inthe mNTS attenuates baroreflex responses. Possible mechanisms for excitatory effects of E-2 in the mNTS resulting in depressor and bradycardic responses, on one hand, andinhibitory effects resulting in attenuation of baroreflex responses, on the other, arediscussed.
© 2005 Elsevier B.V. All rights reserved.
Keywords:BaroreflexBlood pressureEndomorphinHeart rateMu-opioid receptorNaloxone
1. Introduction
Baroreceptor afferents are known to make their primarysynapse in the medial subnucleus of the nucleus tractussolitarius (mNTS) (Ciriello et al., 1994). Although there is ageneral consensus that an excitatory amino acid (probablyglutamate) is the neurotransmitter released at the barorecep-tor terminals in the NTS (Talman et al., 1984), otherendogenous substances present in this nucleus may play amodulatory role in baroreflex. Endomorphins (two tetrapep-tides, endomorphin-1 and -2), isolated from human and
apru).
er B.V. All rights reserved
bovine brains, have been reported to possess a high affinityand selectivity for themu-opioid receptors and are consideredto be endogenous ligands for these receptors (Hackler et al.,1997; Zadina et al., 1997). The presence of endomorphin-likeimmunoreactivity (Martin-Schild et al., 1999; Pierce andWessendorf, 2000) and opiate receptors (Atweh and Kuhar,1977; Lord et al., 1977) in the mNTS has been demonstrated.We recently reported that microinjections of endomorphin-2(E-2) into the mNTS elicited depressor and bradycardicresponses (Kasamatsu et al., 2004). This result was unexpectedconsidering that endomorphins are known to possess inhib-
.
366 B R A I N R E S E A R C H 1 0 7 3 – 1 0 7 4 ( 2 0 0 6 ) 3 6 5 – 3 7 3
itory actions on neurons (Baraban et al., 1995; Dun et al., 2000;Guyenet et al., 2002; Hayar and Guyenet, 1998; Wu et al., 1999),and this effect is expected to elicit an increase in bloodpressure (BP) and heart rate (HR) based on known circuitry ofcardiovascular regulatory areas in the medulla (Dampney,1994; Sapru, 2002). The depressor and bradycardic responsesobserved in our earlier study were mediated via activation ofionotropic glutamate receptors located on the secondarymNTS neurons (Kasamatsu et al., 2004). Based on this report(Kasamatsu et al., 2004), it was hypothesized that activation ofsecondarymNTSneurons by E-2may result in an exaggerationof baroreflex responses. The present study was designed totest this hypothesis. However, the results showed that E-2attenuated baroreflex responses. Possible mechanisms forexcitatory effects of E-2 in the mNTS resulting in depressorand bradycardic responses, on one hand, and inhibitoryeffects resulting in attenuation of baroreflex responses, onthe other, are discussed.
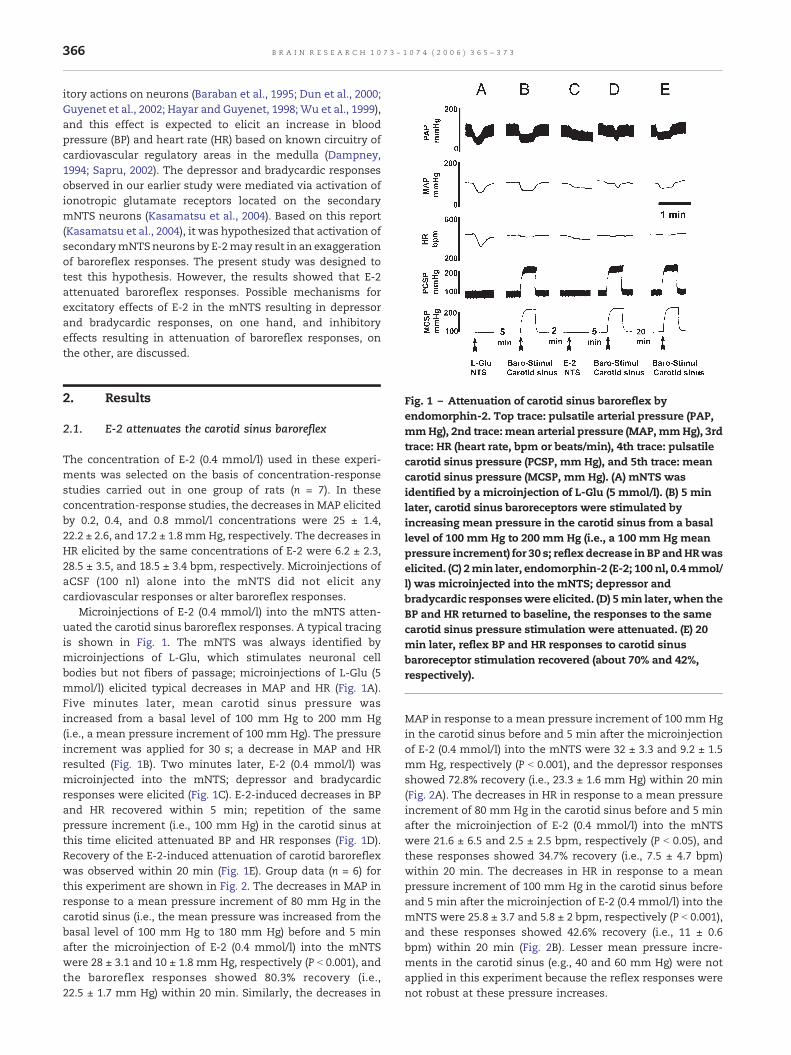
Fig. 1 – Attenuation of carotid sinus baroreflex byendomorphin-2. Top trace: pulsatile arterial pressure (PAP,mmHg), 2nd trace: mean arterial pressure (MAP,mmHg), 3rdtrace: HR (heart rate, bpm or beats/min), 4th trace: pulsatilecarotid sinus pressure (PCSP, mm Hg), and 5th trace: meancarotid sinus pressure (MCSP, mm Hg). (A) mNTS wasidentified by a microinjection of L-Glu (5 mmol/l). (B) 5 minlater, carotid sinus baroreceptors were stimulated byincreasing mean pressure in the carotid sinus from a basallevel of 100 mm Hg to 200 mm Hg (i.e., a 100 mm Hg meanpressure increment) for 30 s; reflex decrease in BP andHRwaselicited. (C) 2min later, endomorphin-2 (E-2; 100 nl, 0.4mmol/l) was microinjected into the mNTS; depressor andbradycardic responseswere elicited. (D) 5min later, when theBP and HR returned to baseline, the responses to the samecarotid sinus pressure stimulation were attenuated. (E) 20min later, reflex BP and HR responses to carotid sinusbaroreceptor stimulation recovered (about 70% and 42%,respectively).
2. Results
2.1. E-2 attenuates the carotid sinus baroreflex
The concentration of E-2 (0.4 mmol/l) used in these experi-ments was selected on the basis of concentration-responsestudies carried out in one group of rats (n = 7). In theseconcentration-response studies, the decreases in MAP elicitedby 0.2, 0.4, and 0.8 mmol/l concentrations were 25 ± 1.4,22.2 ± 2.6, and 17.2 ± 1.8 mmHg, respectively. The decreases inHR elicited by the same concentrations of E-2 were 6.2 ± 2.3,28.5 ± 3.5, and 18.5 ± 3.4 bpm, respectively. Microinjections ofaCSF (100 nl) alone into the mNTS did not elicit anycardiovascular responses or alter baroreflex responses.
Microinjections of E-2 (0.4 mmol/l) into the mNTS atten-uated the carotid sinus baroreflex responses. A typical tracingis shown in Fig. 1. The mNTS was always identified bymicroinjections of L-Glu, which stimulates neuronal cellbodies but not fibers of passage; microinjections of L-Glu (5mmol/l) elicited typical decreases in MAP and HR (Fig. 1A).Five minutes later, mean carotid sinus pressure wasincreased from a basal level of 100 mm Hg to 200 mm Hg(i.e., a mean pressure increment of 100 mm Hg). The pressureincrement was applied for 30 s; a decrease in MAP and HRresulted (Fig. 1B). Two minutes later, E-2 (0.4 mmol/l) wasmicroinjected into the mNTS; depressor and bradycardicresponses were elicited (Fig. 1C). E-2-induced decreases in BPand HR recovered within 5 min; repetition of the samepressure increment (i.e., 100 mm Hg) in the carotid sinus atthis time elicited attenuated BP and HR responses (Fig. 1D).Recovery of the E-2-induced attenuation of carotid baroreflexwas observed within 20 min (Fig. 1E). Group data (n = 6) forthis experiment are shown in Fig. 2. The decreases in MAP inresponse to a mean pressure increment of 80 mm Hg in thecarotid sinus (i.e., the mean pressure was increased from thebasal level of 100 mm Hg to 180 mm Hg) before and 5 minafter the microinjection of E-2 (0.4 mmol/l) into the mNTSwere 28 ± 3.1 and 10 ± 1.8 mm Hg, respectively (P b 0.001), andthe baroreflex responses showed 80.3% recovery (i.e.,22.5 ± 1.7 mm Hg) within 20 min. Similarly, the decreases in
MAP in response to a mean pressure increment of 100 mm Hgin the carotid sinus before and 5 min after the microinjectionof E-2 (0.4 mmol/l) into the mNTS were 32 ± 3.3 and 9.2 ± 1.5mm Hg, respectively (P b 0.001), and the depressor responsesshowed 72.8% recovery (i.e., 23.3 ± 1.6 mm Hg) within 20 min(Fig. 2A). The decreases in HR in response to a mean pressureincrement of 80 mm Hg in the carotid sinus before and 5 minafter the microinjection of E-2 (0.4 mmol/l) into the mNTSwere 21.6 ± 6.5 and 2.5 ± 2.5 bpm, respectively (P b 0.05), andthese responses showed 34.7% recovery (i.e., 7.5 ± 4.7 bpm)within 20 min. The decreases in HR in response to a meanpressure increment of 100 mm Hg in the carotid sinus beforeand 5 min after the microinjection of E-2 (0.4 mmol/l) into themNTS were 25.8 ± 3.7 and 5.8 ± 2 bpm, respectively (P b 0.001),and these responses showed 42.6% recovery (i.e., 11 ± 0.6bpm) within 20 min (Fig. 2B). Lesser mean pressure incre-ments in the carotid sinus (e.g., 40 and 60 mm Hg) were notapplied in this experiment because the reflex responses werenot robust at these pressure increases.

Fig. 2 – Bar graph showing the effects of endomorphin-2 on carotid baroreflex responses. (A) The decrease in MAP in responseto a mean pressure increase in the carotid sinus from the basal level of 100 mm Hg to 180 mm Hg (i.e., a mean pressureincrement of 80 mm Hg) 5 min after the microinjection of E-2 (100 nl, 0.4 mmol/l) into the mNTS was significantly attenuated(P b 0.001) when compared to the reflex decrease inMAP before themicroinjection of E-2. The decreases inMAP in response to amean pressure increment of 100 mm Hg in the carotid sinus 5 min after the same concentration of E-2 was also significantlyattenuated (P b 0.001) when compared to the reflex decrease in MAP before the microinjection of E-2. (B) The decrease in HR inresponse to a mean pressure increment of 80 mm Hg in the carotid sinus 5 min after the microinjection of the sameconcentration of E-2 into the mNTS was significantly attenuated (P b 0.05) when compared to the reflex decrease in HR beforethe microinjection of E-2. The decreases in HR in response to a mean pressure increment of 100 mm Hg in the carotid sinus 5min after the same concentration of E-2was also significantly attenuated (P b 0.001) when compared to the reflex decrease in HRbefore the microinjection of E-2. There was a recovery from attenuation of the reflex depressor and bradycardic responses 20min after the microinjection of E-2.
Fig. 3 – Attenuation of aortic baroreflex by endomorphin-2.Top trace: pulsatile arterial pressure (PAP,mmHg), 2nd trace:mean arterial pressure (MAP, mm Hg), 3rd trace: HR (heartrate, bpm or beats/min). (A) mNTS was identified by amicroinjection of L-Glu (5 mmol/l). (B) 5 min later, aorticdepressor nerve was stimulated electrically at 5, 10, and 25pulses/s (the interval between each stimulus frequency was30–35 s); decrease in BP and HR was elicited by aortic nervestimulation at each frequency. Five minutes after the lastaortic nerve stimulation, E-2 (100 nl, 0.4 mmol/l) was injectedinto the mNTS (large arrow between panels B and C); usualdepressor and bradycardic responses were elicited (notshown). (C) 5 min later when the BP and HR returned tobaseline, aortic nerve was again stimulated at 5, 10, and 25pps; the depressor and bradycardic responses wereattenuated. (D) 20 min later, reflex BP and HR responses toaortic nerve stimulation showed almost complete recovery.
367B R A I N R E S E A R C H 1 0 7 3 – 1 0 7 4 ( 2 0 0 6 ) 3 6 5 – 3 7 3
2.2. E-2 attenuates the aortic baroreflex
The aortic nerve was electrically stimulated at three differentfrequencies: 5, 10, and 25 pulses/s (pps). We have previouslyreported that repeated stimulation of the aortic nerve did notlead to desensitization of the baroreflex responses (Kasa-matsu and Sapru, 2005). Microinjections of E-2 (0.4 mmol/l)into the mNTS attenuated the responses to the aortic nervestimulation. A typical tracing is shown in Fig. 3. ThemNTSwasidentified by microinjection of L-Glu (5 mmol/l) (Fig. 3A). Fiveminutes later, the aortic nerve was stimulated at 5, 10, and 25pps which resulted in decreases in MAP and HR (Fig. 3B). Theonset of responses for each stimulus was immediate, whereasthe peak effects were elicited within 4–5 s after the stimula-tion. The MAP and HR recovered to basal levels within 4–5 safter the cessation of the stimulation. Two minutes after thelast aortic nerve stimulation, E-2 (0.4 mmol/l) was micro-injected in the NTS; a decrease in MAP and HR was elicitedwhich recovered within 5 min. Five minutes after themicroinjection of E-2, the aortic nerve was again stimulatedat the same frequencies; the decreases in MAP and HR weresignificantly attenuated (Fig. 3C). Recovery from E-2-inducedattenuation of aortic baroreflex responses was observedwithin 20 min (Fig. 3D). Group data (n = 7) for this experimentare shown graphically in Fig. 4. The decreases in MAP (Fig. 4A)in response to electrical stimulations of 5 pps before and 5minafter the microinjection of E-2 (0.4 mmol/l) were 16 ± 1.9 and6.4 ± 0.9mmHg, respectively (P b 0.001). Recovery from the E-2-induced attenuation of BP response was observed within 20min (BP response to aortic nerve stimulation at 5 pps was17.1 ± 3.2 mmHg at this time). Similarly, the decreases in MAPin response to electrical stimulation at 10 pps before, 5 min

Fig. 4 – Bar graph showing the effects of endomorphin-2 on aortic baroreflex responses. (A) The decreases in MAP in responseto 5 pps electrical stimulation of the aortic nerve 5 min after the microinjection of E-2 (100 nl, 0.4 mmol/l) into the mNTS weresignificantly attenuated (P b 0.001) when compared to the decreases in MAP before the microinjection of E-2. The decreases inMAP in response to 10 pps nerve stimulation 5 min after the microinjection of E-2 into the mNTS were significantly attenuated(P b 0.001) compared the similar decreases before the microinjection of E-2. The decreases in MAP in response to 25 pps nervestimulation 5min after themicroinjection of E-2 into themNTSwere also significantly attenuated (P b 0.001) compared to thoseelicited before the microinjection of E-2. (B) The decreases in HR in response to 5 pps electrical stimulation of the aortic nerve 5min after themicroinjection of E-2 (100 nl, 0.4mmol/l) into themNTSwere significantly attenuated (P b 0.001)when compared tothe decreases in HR before themicroinjection of E-2. The decreases in HR in response to 10 pps nerve stimulation 5min after themicroinjection of E-2 into the mNTS were significantly attenuated (P b 0.001) compared the similar decreases before themicroinjection of E-2. The decreases in HR in response to 25 pps nerve stimulation 5min after the microinjection of E-2 into themNTSwere also significantly attenuated (P b 0.001) compared to those elicited before themicroinjection of E-2. The attenuationof reflex BP and HR responses by E-2 showed recovery within 20 min of the microinjection of E-2.
368 B R A I N R E S E A R C H 1 0 7 3 – 1 0 7 4 ( 2 0 0 6 ) 3 6 5 – 3 7 3
and 20 min after E-2 were 30 ± 2.6, 12.4 ± 2.5 (P b 0.001), and27.8 ± 2.6 mmHg (92.6% recovery), respectively. The decreasesin MAP in response to electrical stimulation at 25 pps before, 5min and 20 min after E-2 were 39.2 ± 2.2, 18.5 ± 4.8 (P b 0.001),and 34.2 ± 1.7 mm Hg (87.2% recovery), respectively. Thedecreases in HR (Fig. 4B) in response to electrical stimulationsof 5 pps before and 5 and 20min after themicroinjection of E-2were 27.2 ± 9.4, 3.5 ± 1.8 (P b 0.001) and 30 ± 10.5 bpm (completerecovery), respectively. The decreases in HR at 10 pps before, 5and 20 min after the microinjection of E-2 were 61.4 ± 12.6,11.4 ± 1.7 (P b 0.001), and 57.1 ± 16.2 bpm (92.9% recovery),respectively. The decreases in HR at 25 pps before, 5 and 20min after the microinjection of E-2 were 103.5 ± 18.7, 28.5 ± 6.3(P b 0.001), and 82.8 ± 13.7 bpm (80% recovery), respectively.
2.3. E-2 does not alter response to L-Glu or NMDA
In another group of rats (n = 5), E-2 did not alter responses toNMDA. In this experiment, the decreases in MAP in responseto microinjections of NMDA (10 μmol/l) into the mNTS beforeand after the microinjection of E-2 (0.4 mmol/l) were 65 ± 9.4and 67 ± 5.1 mm Hg, respectively (P N 0.05), and the decreasesin HR in response to NMDA before and after themicroinjectionof E-2 were 146 ± 33.2 and 142 ± 23.3 bpm, respectively(P N 0.05).
2.4. Naloxone does not attenuate the baroreflex responses
The role of endogenous opioid receptors in baroreflexresponses was tested using aortic nerve stimulation. First the
concentration of naloxone that blocked the responses tomaximally effective concentration of E-2 (0.4 mmol/l) wasdetermined as follows. Microinjection of E-2 (0.4 mmol/l) intothe mNTS, previously identified by microinjections of L-Glu,elicited decreases in MAP (25 ± 2.2 mm Hg) and HR (56 ± 6.7bpm). E-2-induced depressor and bradycardic responses re-coveredwithin 20min. At this time, naloxone (0.5mmol/l) wasmicroinjected at the samemNTS-site. Subsequent microinjec-tion of E-2 (0.4 mmol/l), within 5 min, failed to elicit anyresponse indicating that naloxone completely blocked theeffects of the maximally effective concentration of E-2.Naloxone-induced blockade of E-2 responses lasted for 20min; at this time, another microinjection of E-2 (0.4 mmol/l)showed almost complete recovery of depressor (15 ± 2.7 mmHg) and bradycardic (40 ± 3.1 bpm) responses. The specificity ofnaloxone (0.5 mmol/l) was indicated by the observation thatthis antagonist did not alter the depressor and bradycardicresponses to L-Glu (5mmol/l); the depressor responses to L-Glubefore and after the microinjection of naloxone (0.5 mmol/l)were 47.5 ± 5.9 and 50 ± 5.4 mm Hg, respectively (P N 0.05), andthe bradycardic responses to L-Glu before and after themicroinjection of this antagonist were 82.5 ± 14.3 and82.5 ± 13.7 bpm, respectively (P N 0.05). Smaller concentrationsof naloxone (e.g., 0.25 mmol/l) did not attenuate the cardio-vascular responses to E-2 (0.4 mmol/l).
The effect of naloxone (0.5mmol/l) on aortic baroreflex wastested in another group of rats (n = 4) as follows. The depressorresponses to 10 pps electrical stimulation before and aftermicroinjection of naloxonewere 45 ± 4.5 and 42.5 ± 3.2mmHg,respectively (P N 0.05), and the bradycardic responses to the

369B R A I N R E S E A R C H 1 0 7 3 – 1 0 7 4 ( 2 0 0 6 ) 3 6 5 – 3 7 3
same stimulation before and after naloxone were 87.5 ± 24.2and 70 ± 17.7 bpm, respectively (P N 0.05). Thus, blockade ofopioid receptors in themNTS did not alter the aortic baroreflexresponses. A typical tracing of these results is shown in Fig. 5.
2.5. Histology
The mNTS sites, where different agents were microinjected,were marked in 12 rats. Composite diagrams of the markedsites are shown in Figs. 6A and B inwhich each spot representsa site of microinjection. The sites marked with ink werelocated in the mNTS, 0.5–0.6 mm rostral to the calamusscriptorius, 0.5–0.6 mm lateral to the midline, and 0.4–0.6 mmdeep from the dorsal medullary surface.
Fig. 6 – Drawings of the coronal sections of the medullashowing marked mNTS sites. (A) 0.5 mm rostral to the
3. Discussion
The main finding of this study is that microinjections of E-2,an endogenous agonist of μ-opioid receptors (Zadina et al.,1997), into themNTS of the rat attenuated the reflex responses(i.e., decrease in BP and HR) to the stimulation of carotid sinusand aortic baroreceptors. Isolated carotid sinus preparationwas used for pressure-induced stimulation of the barorecep-tors in this region (Viard and Sapru, 2004). Electrical stimula-tion of the aortic nerve was used because, in the rat, this nervecontains predominantly baroreceptor afferents (Numao et al.,1985; Sapru and Krieger, 1977; Sapru et al., 1981). Although thepresence of a sparse population of chemoreceptor afferents in
Fig. 5 – Effect of microinjections of naloxone into the mNTSon aortic baroreflex. (A) The mNTS was identified bymicroinjections of L-Glu (100 nl; 5 mmol/l); usual depressorand bradycardic responses were elicited. (B) 5 min later,aortic nerve (AN) was stimulated electrically at 10 pps; reflexdepressor and bradycardic responses were elicited.Naloxone (0.5 mmol/l) (large arrow between panels B and C)was microinjected into the mNTS; no cardiovascularresponses were elicited by naloxone (not shown). (C) 5 minlater, the aortic nerve was stimulated at the same frequency;naloxone did not alter the responses to the AN stimulation.
calamus scriptorius (CS). (B) 0.6 mm rostral to the CS. Eachdark spot represents microinjection site in one animal. Thesites marked with ink were located in mNTS, 0.5–0.6 mmrostral to the calamus scriptorius, 0.5–0.6 mm lateral to themidline, and 0.4–0.6 mm deep from the dorsal medullarysurface. Abbreviations: AP: area postrema, cc: central canal,nAmb: nucleus ambiguus, NTS: nucleus tractus solitarius,py: pyramids; 10: dorsal motor nucleus of vagus, 12:hypoglossal nucleus.
this nerve has been reported in the rat (Brophy et al., 1999;Cheng et al., 1997), subsequent studies have shown that theseafferents may not be of functional importance in the reflexregulation of cardiovascular function (Kobayashi et al., 1999).
Our finding that E-2 attenuated baroreflex responses is inagreement with earlier reports in which other opioid receptoragonists (e.g., enkephalins, β-endorphin, and morphine) havebeen shown to attenuate or abolish the responses tobaroreceptor stimulation (Gordon, 1990; Miura and Takayama,1983; Wang and Li, 1988). Local distortion of brain tissuecaused by microinjections was not responsible for theattenuation of baroreflex responses because microinjectionsof aCSF alone into the mNTS did not attenuate theseresponses. The selection of 100 nl volume for microinjectionsinto the mNTS was based on our earlier studies in whichresponses to L-Glu or E-2 using this volume were greater thanthose elicited by 50 nl volumemicroinjections (Kasamatsu andSapru, 2005; Kasamatsu et al., 2004).
The effects of E-2 were mediated via opioid receptorsbecause prior microinjections of naloxone (an opioid receptor

Fig. 7 – A hypothetical model showing possible mechanismsof E-2 effects in the mNTS. Ionotropic glutamate receptors(NMDA, AMPA, and Kainate receptors), located on thepostsynaptic neurons (a) within the mNTS, are accessible toendogenously released glutamate (Glu). GABA released fromthe GABAergic neurons (b) located in the mNTS acts onpresynaptic glutamate terminals (c) and inhibits glutamaterelease from them. GABA also acts on postsynaptic mNTSneurons (a) and inhibits them. Exogenously microinjectedE-2 acts on mu-opioid receptors located on the GABAergicneurons and inhibits them. Thus, inhibition of Glu releasefrom the presynaptic glutamatergic terminals by GABAergicneurons is removed, and there is an increase in Glu releasefrom these terminals. Endogenously released Glu excitespostsynaptic neurons via ionotropic glutamate receptors.E-2-induced inhibition of GABAergic neurons also results indisinhibition of the postsynaptic mNTS neurons. Net result isexcitation of postsynaptic mNTS neurons which results indepressor and bradycardic responses. Mu-opioid receptorsare located either exclusively or preponderantly on theglutamatergic baroreceptor afferent terminals (d); E-2 acts onopioid receptors and decreases Glu release from theseterminals in response to baroreceptor stimulation andbaroreflex is attenuated.
370 B R A I N R E S E A R C H 1 0 7 3 – 1 0 7 4 ( 2 0 0 6 ) 3 6 5 – 3 7 3
antagonist) prevented these responses. However, microinjec-tions of naloxone alone did not alter the baroreflex responsesindicating that opioid receptors in the mNTS are not normallyinvolved in regulating the baroreflex function. This observa-tion regarding naloxone is also in agreementwith prior reports(Gordon, 1990; Gordon et al., 1994; Wang and Li, 1988).
We have previously reported that microinjections of E-2into the mNTS elicit depressor and bradycardic responses(Kasamatsu et al., 2004). Based on our current knowledge ofthe organization of medullary neural structures regulatingcardiovascular function (Dampney, 1994; Sapru, 2002), E-2-induced depressor, and bradycardic responses must be theresult of excitation of mNTS neurons. Increased excitability ofmNTS neurons is expected to cause a potentiation rather thanattenuation of baroreflex responses observed in this study.Attenuation of baroreflex responses can be explained by eitherE-2-induced reduction in responses of postsynaptic mNTSneurons involved in baroreflex or inhibition of neurotrans-mitter release from the baroreceptor terminals. Reduction inthe postsynaptic responses of the neurons involved inbaroreflex does not seem to be the mechanism by whichattenuation of baroreflex is elicited by E-2 because theresponses to microinjections of NMDA, which acts predomi-nantly postsynaptically (Malenka and Nicoll, 1999), remainedunaltered after microinjections of E-2 into the mNTS. There-fore, attenuation of baroreflex responses by E-2 may beascribed to a reduction in the release of neurotransmitterfrom the baroreceptor terminals. Since glutamate has beenreported to be the neurotransmitter released from barorecep-tor terminals in the mNTS (Talman et al., 1984), E-2 mayinhibit its release by a presynaptic action. In this context, itmay be pointed out that opioid receptors have been reportedto be present on presynaptic terminals of vagal afferents in theNTS (Aicher et al., 2000), and opioid peptides have been shownto inhibit glutamate release from nerve terminals (Guyenet etal., 2002; Hayar and Guyenet, 1998; Nicol et al., 1996). Theinhibitory effect of E-2 on baroreflex, on one hand, andexcitatory effect on mNTS neurons resulting in depressorand bradycardic responses, on the other hand, can bereconciled as follows. It is hypothesized (Fig. 7) that theactivity of mNTS neurons may be under dual control: (1)inhibitory influence from projections of GABAergic neuronspresent in themNTS (Izzo et al., 1992; Maqbool et al., 1991) andexcitatory influence of glutamatergic projections from otherareas of the brain, e.g., insular cortex (Owens et al., 1999;Torrealba and Müller, 1996) to the mNTS. The inhibitory effectof GABA neurons in the mNTS may be presynaptic (i.e.,inhibition of the release of glutamate from the terminals) aswell as postsynaptic (i.e., hyperpolarization of the postsynap-tic neuron). Inhibition of GABAergic neurons by E-2 via mu-opioid receptors would result in increased release of gluta-mate from presynaptic terminals and reduction of inhibitoryinfluence on the postsynaptic neurons (disinhibition). Thus, E-2-induced excitation of postsynaptic mNTS neurons can beexplained by the preponderance of excitatory influence ofglutamatergic terminals on them and the disinhibition ofthese neurons. This effect would explain the mechanism ofthe depressor and bradycardic responses (Kasamatsu et al.,2004). As explained above, attenuation of baroreflex may bethe result of inhibition of neurotransmitter (glutamate) release
from the baroreceptor terminals via a presynaptic opioidreceptor action; for this explanation to be true, opioidreceptors must be present either exclusively or preponder-antly on the baroreceptor terminals. Supporting this hypoth-esis is the report in which only 33% of the vagal afferentscontained labeling for opioid receptors suggesting that manyglutamatergic terminals in the mNTS do not contain opioidreceptors (Aicher et al., 2000; Silverman et al., 2005).
In summary, microinjections of E-2 into the mNTS elicitdepressor and bradycardic responses and attenuation ofbaroreflex responses. The physiological significance of ourresults is not clear at the present time. Since microinjections

371B R A I N R E S E A R C H 1 0 7 3 – 1 0 7 4 ( 2 0 0 6 ) 3 6 5 – 3 7 3
of naloxone alone into the mNTS did not exert any cardio-vascular effects and did not alter baroreflex responses, it isunlikely that endomorphins play a role in the regulation ofcardiovascular function under normal circumstances. How-ever, opioidergic mechanisms may come into play in yetunidentified situations which affect cardiovascular function.For example, it can be speculated that opioidergic mechan-isms may be activated during nociception. Noxious stimula-tion is known to elicit pressor and tachycardic responses(Boscan et al., 2002; Ness and Gebhart, 1990). Under theseconditions, depressor and bradycardic responses elicited byendomorphin in the mNTS may mitigate the pressor andtachycardic responses elicited by painful stimuli. Inhibition ofbaroreflex in such situations would also be useful so thatcompensatory cardiovascular adjustments to painful stimuliare not mitigated by an intact baroreflex.
4. Experimental procedures
4.1. General procedures
Experiments were done in adult male Wistar rats (Charles RiverLaboratories, Wilmington, MA, USA), weighing 300–350 g (n = 60).All animals were housed under controlled conditions with a 12-h light/dark cycle. Food andwater were available to the animals adlibitum. The experimental procedures were performed in accor-dance with the guidelines for animal experimentation describedin the “Guiding Principles for Research Involving Animals andHuman Beings” (Am J Physiol Regul Integr Comp Physiol 283: R281–R283, 2002), and the Institutional Animal Care and Use Committeeof this university approved the experimental protocols. Thenumber of animals used was the minimum required for statisticalanalyses of the data, and every effort was made to minimize anysuffering to the animals.
Anesthesia was induced by administration of isoflurane (3% in100% oxygen) via a nose-mask. The trachea was cannulated withpolyethylene tubing (PE 240), the rats were artificially ventilatedusing a rodent ventilator (Harvard Instruments, Holliston, MA,model 683) with the respiratory volume and rate set at 2.5 ml and50 strokes/min, respectively. Anesthesia was maintained bytracheal administration of isoflurane (2–3% in 100% oxygen)contained in a bag connected to the inlet of the ventilator. Thefemoral vein and artery on one side were cannulated withpolyethylene tubing (PE 50) for intravenous injections of variousagents and for monitoring BP, respectively. A pressure transducer(Grass Instruments, West Warwick, RI, USA, model P23 Db) wasused to measure the BP. HR wasmonitored by a tachograph (GrassInstruments, model 7P4) that was triggered by the BP waves.Urethane (1.2–1.4 gm/kg) was injected intravenously in 6–7aliquots at 2-min intervals. The tracheal inhalation of isofluranewas discontinued after the administration of the third aliquot ofurethane. The depth of anesthesiawas established by pinching thehind paw of the rat; absence of a BP response and/or withdrawal ofthe limb indicated that the rat was properly anesthetized. Theanimals were immobilized by intravenous administration ofdecamethonium (3 mg/kg) and artificially ventilated throughoutthe experiment. Before the administration of the neuromuscularblocker, the depth of anesthesia was always ascertained asmentioned earlier in order to ensure that the animal wasadequately anesthetized. The respiratory volume and rate on theventilator were adjusted so that the end-tidal CO2, monitored bymeasurement of expired gas with an infrared CO2 analyzer (Micro-Capnometer, Columbus Instruments, OH, USA) was maintainedbetween 3.5 and 4.5%. Rectal temperature was monitored contin-uously and maintained at 37 ± 0.5 °C using an infrared lamp
connected to a temperature controller. All of the tracings wererecorded on a polygraph (Grass Instruments, model 7D).
4.2. Microinjection technique
The rats were placed in a prone position in a stereotaxicinstrument (David Kopf Instruments, Tujunga, CA, USA, model1430) with the bite bar 18 mm below the interaural line. Themedulla was exposed by removing the dorsal neck muscles, partsof the occipital bone and dura. The atlanto-occipital membranewas incised. Three or four barreled glassmicropipettes (tip size 20–40 μm) were mounted on a micromanipulator (David KopfInstruments, model 1460 with an AP slide 1262), and each barrelwas connected via PE tubing to one of the channels on apicospritzer (General Valve Corp, Fairfield, NJ, USA). One of thebarrels contained L-glutamate (L-Glu), and the contents of theother barrels varied according to the requirements of theexperiment being conducted. The coordinates for the mNTSwere 0.5–0.6 mm rostral and 0.5–0.6 mm lateral to the calamusscriptorius and 0.5–0.6 mm deep from the dorsal medullarysurface. For making microinjections, the micropipette wasinserted into the brain perpendicularly. The site eliciting depres-sor and bradycardic responses was identified by microinjectionsof L-Glu (5 mmol/l) into the mNTS. The volume of microinjections(100 nl) was visually confirmed by the displacement of fluidmeniscus in the barrel containing the injectate under a modifiedbinocular horizontal microscope with a graduated reticule in oneeye-piece (World Precision Instruments, Sarasota, FL, USA, modelPZMH). The duration of microinjection was 10 s. Controls formicroinjections consisted of artificial cerebrospinal fluid (aCSF;see Drugs and chemicals).
4.3. Stimulation of carotid sinus baroreceptors
The procedure for the stimulation of carotid sinus baroreceptorshas been described elsewhere (Viard and Sapru, 2004). Briefly, thecarotid sinus on one side was isolated from the general circulationby ligating the internal carotid, pterygopalatine, superior thyroid,ascending pharyngeal, lingual, and external carotid arteries(Sapru and Krieger, 1977). A cannula was placed in the commoncarotid artery pointing rostrally, and another cannula was placedin the external carotid artery pointing caudally. The isolatedcarotid sinus was immediately perfused with a warm (37 °C)physiological solution (pH 7.4) of the following composition (mM):NaCl 125, KCl 3, MgSO4 1.3, CaCl2 2.4, dextrose 10, and HEPES 5.Cannulae (PE50) in the common carotid and external carotidarteries served as an inlet and outlet for the perfusion fluid,respectively. The physiological saline was saturated with 100%oxygen (O2) and perfused through the carotid sinus (with intactinnervation) at a rate of 1.5–2 ml/min. The perfusion solutionflowed into the carotid sinus in a pulsatile manner; the crest andthe trough of the pulses were 114 ± 0.8 and 86 ± 0.8 mm Hg,respectively. The amplitude of pressure pulses in the isolatedcarotid sinus was adjusted so that the mean basal perfusionpressure, monitored by a pressure transducer (Statham P23 Db),was 100 mmHg. Different levels of mean pressure were generatedby the height of the water column in the perfusion assembly. Inorder to stimulate the carotid baroreceptors, the mean pressure inthe carotid sinus was changed to the desired level using selectorswitches connected to the water column included in the perfusionassembly. This procedure allowed an abrupt increase in the meancarotid sinus pressure from the basal perfusion pressure. Themean pressure increment was maintained for 30 s at the end ofwhich the selector switches were returned to their originalpositions and the basal perfusion pressure resumed. For example,during the application of an 80 mm Hg mean pressure increment,the pressure increased from the mean basal perfusion pressure of100mmHg to amaximummean pressure of 180mmHg. Similarly,

372 B R A I N R E S E A R C H 1 0 7 3 – 1 0 7 4 ( 2 0 0 6 ) 3 6 5 – 3 7 3
during the application of a 100 mm Hg pressure increment, thepressure in the carotid sinus was increased from the mean basalperfusion pressure of 100 mmHg to amaximummean pressure of200 mm Hg, and the crest and trough of the pressure pulses were213.3 ± 1.2 and 186.6 ± 1.2 mm Hg, respectively. It should be notedthat throughout the paper, the pressure increments to stimulatethe carotid baroreceptors refer to mean pressure increments fromthe mean basal perfusion pressure within the carotid sinus. Themagnitude and sequence of mean pressure increments wereselected at random.
4.4. Aortic nerve stimulation
The procedure for aortic nerve stimulation has been describedpreviously (Gonzalez et al., 1983; Kasamatsu and Sapru, 2005).Briefly, either the left or right aortic nerve was identified under anoperation microscope as it joined the superior laryngeal nerve.The nerve was then exposed in the neck as far caudally as possibleand sectioned at a level close to the chest cavity. The caudal end ofthe aortic nerve was placed on a bipolar electrode made fromsilver wire (A-M Systems; outer diameter 0.003 mm), and theelectrode tip, along with the nerve-end placed on it, wasembedded in a silicone elastomer (Kwik-Sil, WPI, FL). Theelectrode was connected to a stimulator (model S88, GrassInstruments) via an isolation unit (model SIU5A, Grass Instru-ments) and stimulated electrically. The intensity of the stimuluswas 0.5 V, the duration of pulses was 1 ms, and the frequency ofpulses was variable. Each cycle of stimulations consisted of threefrequencies of electrical stimulus (5, 10, and 25 pulses/s). Aninterval of at least 5–10 min was allowed between different cyclesof stimulations. The stimuli at each frequency were applied for 30s, and the interval between the stimulations at different frequen-cies was 30 s. The responses to aortic nerve stimulation wereobserved before and 5 min after the microinjection of E-2 into theNTS. When E-2 microinjections into the NTS attenuated thebaroreflex responses, a period of at least 20 min was allowed forthe recovery of the reflex responses.
4.5. Histology
The accuracy of our placement of microinjections into the mNTS,where L-Glu elicited baroreceptor reflex-like responses, has beenrepeatedly demonstrated by histological examination in ourprevious publications (Viard and Sapru, 2004). In this study,typical sites of microinjections were marked by a unilateralmicroinjection (10–50 nl) of diluted India ink (n = 12). The animalswere perfused with arterial administration of heparinized normalsaline followed by 10% formalin, the brains were then removedand fixed in 10% formalin for 24 h. After the fixation procedurewas completed, serial sections were cut (30 μm) in a cryostat andmounted on slides. The sections were then stained with cresylviolet, and the microinjection or recording site (marked with Indiaink) was identified under a microscope (Olympus Provis, Middle-bush, NJ, USA, model AX70). The sections were photographed andcompared with a standard atlas (Paxinos et al., 1986).
4.6. Statistical analyses
Themeans and standard error of themeans (SEM) were calculatedformaximumdecreases inmean arterial pressure (MAP) and HR inresponse to microinjections of E-2 or L-Glu into the mNTS, carotidsinus pressure increments, and aortic nerve stimulations. Com-parisons of decreases in MAP elicited by different concentrationsof E-2 weremade by using a one-way analysis of variance followedby Tukey–Kramer multiple comparison test. Paired t test was usedfor comparisons of the maximum changes in MAP and HR elicitedby microinjections of E-2 into the mNTS before and after themicroinjections of naloxone at the same site. Comparisons of the
maximum changes in MAP and HR elicited by carotid sinuspressure increments and aortic nerve stimulations before andafter the microinjections of E-2 or naloxone into the mNTS werealso made by using paired t test. Baroreflex responses (i.e., reflexdecrease in BP and HR) were compared before and 5 min after themicroinjection of E-2 into the mNTS. The comparisons were madein the same animal and paired t test was applied. Although E-2microinjection induced a decrease in MAP and HR, this effectlasted for less than 5 min. Therefore, carotid sinus pressurestimulus was applied 5 min after the microinjection of E-2 whenthe BP and HR had returned to basal levels. In all cases, thedifferences were considered significant at P b 0.05.
4.7. Drugs and chemicals
The following drugs and chemicals were used. E-2 (a μ-receptoragonist) (Kasamatsu et al., 2004), naloxone hydrochloride (com-petitive antagonist at μ, κ, and δ receptors) (Miura and Takayama,1983), L-glutamate monosodium, N-methyl-D-aspartic acid(NMDA; an agonist at the NMDA receptors), isoflurane, andurethane. All solutions for the microinjections were freshlyprepared in aCSF. The composition of aCSF (pH 7.4) was as follows(inmmol/l): 128 NaCl, 3 KCl, 1.2 CaCl2, 0.8 MgCl2, 3.4 dextrose, and 5HEPES (Kasamatsu and Sapru, 2005; Kasamatsu et al., 2004). Thecontrol solutions for microinjections consisted of 100 nl of aCSF.Where applicable, the concentration of drugs injected into themNTS refers to their salts. All drugs were obtained from SigmaChemicals (St. Louis, MO). Isoflurane was purchased from BaxterPharmaceutical Products (Deerfield, IL).
Acknowledgments
This work was supported in part by N.I.H. grants HL24347 andHL076248 awarded to Dr. H. N. Sapru.
R E F E R E N C E S
Aicher, S.A., Goldberg, A., Sharma, S., Pickel, V.M., 2000. Mu-opioidreceptors are present in vagal afferents and their dendritictargets in the medial nucleus tractus solitarius. J. Comp.Neurol. 422, 181–190.
Atweh, S.F., Kuhar, M.J., 1977. Autoradiographic localization ofopiate receptors in rat brain: I. Spinal cord and lower medulla.Brain Res. 124, 53–67.
Baraban, S.C., Stornetta, R.L., 1995. Effects of morphine andmorphine withdrawal on adrenergic neurons of the rat rostralventrolateral medulla. Brain Res. 676, 245–257.
Boscan, P., Kasparov, S.K., 2002. Somatic nociception activates NK1receptors in the nucleus tractus solitarii to attenuate thebaroreceptor cardiac reflex. Eur. J. Neurosci. 16, 907–920.
Brophy, S., Ford, T.W., Carey, M., 1999. Activity of aorticchemoreceptors in the anaesthetized rat. J. Physiol. 514,821–828.
Cheng, Z., Powley, T.L., Schwaber, J.S., Doyle, F.J., 1997. A laserconfocal microscopic study of vagal afferent innervation of rataortic arch: chemoreceptors as well as baroreceptors. J. Auton.Nerv. Sys. 67, 1–14.
Ciriello, J., Hochstenbach, S.L., Roder, S., 1994. Central projectionsof baroreceptor and chemoreceptor afferent fibers in the rat. In:Barraco, I.R.A. (Ed.), Nucleus of the Solitary Tract. CRC Press,Boca Raton, pp. 35–50.
Dampney, R.A., 1994. Functional organization of centralpathways regulating the cardiovascular system. Physiol. Rev.74, 323–364.

373B R A I N R E S E A R C H 1 0 7 3 – 1 0 7 4 ( 2 0 0 6 ) 3 6 5 – 3 7 3
Dun, N.J., Dun, S.L., Wu, S.Y., Williams, C.A., 2000. Endomorphins:localization, release and action on rat dorsal horn neurons.J. Biomed. Sci. 7, 213–220.
Gonzalez, E.R., Krieger, A.J., 1983. Central resetting of baroreflex inthe spontaneously hypertensive rat. Hypertension 5, 346–352.
Gordon, F.J., 1990. Opioids and central baroreflex control: a site ofaction in the nucleus tractus solitarius. Peptides 11, 305–309.
Gordon, F.J., 1994. Opioids and the nucleus of the tractus solitarius;effects on cardiovascular and baroreflex function. In: Barraco, I.R.A. (Ed.), Nucleus of the Solitary Tract. CRC Press, Boca Raton,FL, pp. 283–287.
Guyenet, P.G., Stornetta, R.L., Schreihofer, A.M., Pelaez, N.M.,Hayar, A., Aicher, S., 2002. Opioid signalling in the rat rostralventrolateral medulla. Clin. Exp. Pharmacol. Physiol. 29,238–242.
Hackler, L., Zadina, J.E., Ge, L.J., 1997. Isolation of relatively largeamounts of endomorphin-1 and endomorphin-2 from humanbrain cortex. Peptides 18, 1635–1639.
Hayar, A., Guyenet, P.G., 1998. Pre- and postsynaptic inhibitoryactions of methionine-enkephalin on identified bulbospinalneurons of the rat RVL. J. Neurophysiol. 80, 2003–2014.
Izzo, P.N., Sykes, R.M., 1992. Gamma-Aminobutyric acidimmunoreactive structures in the nucleus tractus solitarius: alight and electron microscopic study. Brain Res. 591, 69–78.
Kasamatsu, K., Sapru, H.N., 2005. Attenuation of aortic baroreflexresponses by microinjections of endomorphin-2 into therostral ventrolateral medullary pressor area of the rat. Am. J.Physiol. 289, R59–R67.
Kasamatsu, K., Chitravanshi, V.C., 2004. Depressor andbradycardic responses to microinjections of endomorphin-2into the NTS are mediated via ionotropic glutamate receptors.Am. J. Physiol. 287, R715–R728.
Kobayashi, M., Cheng, Z.B., Tanaka, K., 1999. Is the aortic depressornerve involved in arterial chemoreflexes in rats? J. Auton. Nerv.Sys. 78, 38–48.
Lord, J.A.H., Waterfield, A.A., Hughes, J., Kosterlitz, H.W., 1977.Endogenous opioid peptides: multiple agonists and receptors.Nature 267, 495–499.
Malenka, R.C., Nicoll, R.A., 1999. Long-term potentiation—Adecade of progress? Science 285, 1870–1874.
Maqbool, A., T.Batten, F.C., McWilliam, P.N., 1991. Ultrastructuralrelationships between GABAergic terminals and cardiac vagalpreganglionic motoneurons and vagal afferents in the cat: acombined HRP tracing and immunogold labelling study. Eur. J.Neurosci. 3, 501–513.
Martin-Schild, S., Gerall, A.A., Kastin, A.J., Zadina, J.E., 1999.Differential distribution of endomorphin 1- and endomorphin2-like immunoreactivities in the CNS of the rodent. J. Comp.Neurol. 405, 450–471.
Miura, M., Takayama, K., 1983. Naloxone inhibition of the carotid
sinus nerve reflex in the nucleus of the solitary tract of the cat.Brain Res. 288, 389–392.
Ness, T.J., Gebhart, G.F., 1990. Visceral pain: a review ofexperimental studies. Pain 41, 167–234.
Nicol, B., Rowbotham, D.J., Lambert, D.G., 1996. Mu- andkappa-opioids inhibit K+ evoked glutamate release from ratcerebrocortical slices. Neurosci. Lett. 218, 79–82.
Numao, Y., Saito, M., Terui, N., Kumada, M., 1985. The aorticnerve-sympathetic reflex in the rat. J. Auton. Nerv. Sys. 13,65–79.
Owens, N.C., Sartor, D.M., Verberne, A.J., 1999. Medial prefrontalcortex depressor response: role of the solitary tract nucleus inthe rat. Neuroscience 89, 1331–1346.
Paxinos, G., Watson, C., 1986. The Rat Brain in StereotaxicCoordinates. Academic Press.
Pierce, T.L., Wessendorf, M.W., 2000. Immunocytochemicalmapping of endomorphin-2-immunoreactivity in rat brain.J. Chem. Neuroanat. 18, 181–207.
Sapru, H.N., 2002. Glutamate circuits in selected medullo-spinalareas regulating cardiovascular function. Clin. Exp. Pharmacol.Physiol. 29, 491–496.
Sapru, H.N., Krieger, A.J., 1977. Carotid and aortic chemoreceptorfunction in the rat. J. Appl. Physiol. 42, 344–348.
Sapru, H.N., Gonzalez, E.R., 1981. Aortic nerve stimulation in therat: cardiovascular and respiratory responses. Brain Res. Bull. 6,393–398.
Silverman, M.B., Hermes, S.M., Zadina, J.E., Aicher, S.A., 2005.Mu-opioid receptor is present in dendritic targets ofendomorphin-2 axon terminals in the nuclei of the solitarytract. Neuroscience 135, 887–896.
Talman, W.T., Granata, A.R., 1984. Glutamatergic mechanisms inthe nucleus tractus solitarius in blood pressure control. Fed.Proc. 43, 39–44.
Torrealba, F., Müller, C., 1996. Glutamate immunoreactivity ofinsular cortex afferents to the nucleus tractus solitarius in therat: a quantitative electron microscopic study. Neuroscience71, 77–87.
Viard, E., Sapru, H.N., 2004. Carotid baroreflex in the rat: role ofglutamate receptors in the medial subnucleus of the solitarytract. Neuroscience 126, 785–794.
Wang, Q., Li, P., 1988. Inhibition of baroreflex followingmicroinjection of GABA or morphine into the nucleus tractussolitarii in rabbits. J. Auton. Nerv. Sys. 25, 165–172.
Wu, S.Y., Dun, S.L., Wright, M.T., Chang, J.K., Dun, N.J., 1999.Endomorphin-like immunoreactivity in the rat dorsal horn andinhibition of substantia gelatinosa neurons in vitro.Neuroscience 89, 317–321.
Zadina, J.E., Hackler, L., L.-Ge, J., 1997. A potent and selectiveendogenous agonist for the mu-opiate receptor. Nature 386,499–502.