engineering triterpene and methylated triterpene ... · engineering triterpene and methylated...
TRANSCRIPT

Engineering Triterpene and Methylated TriterpeneProduction in Plants Provides Biochemical andPhysiological Insights into Terpene Metabolism1[OPEN]
Zuodong Jiang, Chase Kempinski, Caroline J. Bush, S. Eric Nybo2, and Joe Chappell*
Plant Biology Program (Z.J., C.K., J.C.) and Department of Pharmaceutical Sciences (C.J.B., S.E.N., J.C.),University of Kentucky, Lexington, Kentucky 40536–0596
Linear, branch-chained triterpenes, including squalene (C30), botryococcene (C30), and their methylated derivatives (C31–C37),generated by the green alga Botryococcus braunii race B have received significant attention because of their utility as chemical andbiofuel feedstocks. However, the slow growth habit of B. braunii makes it impractical as a production system. In this study, weevaluated the potential of generating high levels of botryococcene in tobacco (Nicotiana tabacum) plants by diverting carbon flux fromthe cytosolic mevalonate pathway or the plastidic methylerythritol phosphate pathway by the targeted overexpression of an avianfarnesyl diphosphate synthase along with two versions of botryococcene synthases. Up to 544 mg g21 fresh weight of botryococcenewas achieved when this metabolism was directed to the chloroplasts, which is approximately 90 times greater than thataccumulating in plants engineered for cytosolic production. To test if methylated triterpenes could be produced in tobacco, wealso engineered triterpene methyltransferases (TMTs) from B. braunii into wild-type plants and transgenic lines selected for high-level triterpene accumulation. Up to 91% of the total triterpene contents could be converted to methylated forms (C31 and C32) bycotargeting the TMTs and triterpene biosynthesis to the chloroplasts, whereas only 4% to 14% of total triterpenes were methylatedwhen this metabolism was directed to the cytoplasm. When the TMTs were overexpressed in the cytoplasm of wild-type plants, upto 72% of the total squalene was methylated, and total triterpene (C30+C31+C32) content was elevated 7-fold. Altogether, theseresults point to innate mechanisms controlling metabolite fluxes, including a homeostatic role for squalene.
Terpenes and terpenoids represent a distinct class ofnatural products (Buckingham, 2003) that are derivedfrom two universal five-carbon precursors: isopentenyldiphosphate (IPP) and dimethylallyl diphosphate(DMAPP). In eukaryotic fungi and animals, IPP andDMAPP are synthesized via the mevalonate (MVA)pathway, whereas in prokaryotes, they are synthesizedvia the methylerythritol phosphate (MEP) pathway. Inhigher plants, the pathways are present in separatecompartments and are believed to operate indepen-dently. The MVA pathway in the cytoplasm is pre-dominantly responsible for sesquiterpene (C15),triterpene (C30), and polyprenol (greater than C45) bi-osynthesis and associated with the endoplasmic retic-ulum (ER) system. The MEP pathway resides inplastids and is dedicated to monoterpenes (C10),
diterpenes (C20), carotenoids (C40), and long-chainphytol biosynthesis. All these compounds are usuallyproduced by plants for a variety of physiological (i.e.hormones, aliphatic membrane anchors, and main-taining membrane structure) and ecological (i.e. de-fense compounds and insect/animal attractants) roles(Kempinski et al., 2015). Terpenes are also important forvarious industrial applications, ranging from flavorsand fragrances (Schwab et al., 2008) to medicines(Dewick, 2009; Niehaus et al., 2011; Shelar, 2011).
The utility of terpenes as chemical and biofuel feed-stocks has also received considerable attention recently.Isoprenoid-derived biofuels include farnesane(Renninger and McPhee, 2008; Rude and Schirmer,2009), bisabolene (Peralta-Yahya et al., 2011), pinene di-mers (Harvey et al., 2010), isopentenal (Withers et al.,2007), and botryococcene (Moldowan and Seifert, 1980;Hillen et al., 1982; Glikson et al., 1989; Mastalerz andHower, 1996). The richness of branches within these hy-drocarbon scaffolds correlate with their high-energycontent, which enables them to serve as suitable alter-natives to crude petroleum (Peralta-Yahya and Keasling,2010). Indeed, some of them are already major contrib-utors to current-day petroleum-based fuels. One of thebest examples of this is the triterpene oil accumulating inthe green alga Botryococcus braunii race B, which is con-sidered a major progenitor to oil and coal shale deposits(Moldowan and Seifert, 1980). This alga has been wellstudied, and the major constituents of its prodigioushydrocarbon oil are a group of triterpenes including
1 This work was supported by the National Institute of Food andAgriculture, U.S. Department of Agriculture (grant no. 201006141622).
2 Present address: Department of Pharmaceutical Sciences, Collegeof Pharmacy, Ferris State University, Big Rapids, MI, 49307.
* Address correspondence to [email protected] author responsible for distribution of materials integral to the
findings presented in this article in accordance with the policy de-scribed in the Instructions for Authors (www.plantphysiol.org) is: JoeChappell ([email protected]).
Z.J. and J.C. conceived the research plan; Z.J., C.K., C.J.B., andS.E.N. performed the experiments; Z.J., C.K., and J.C. wrote the article.
[OPEN] Articles can be viewed without a subscription.www.plantphysiol.org/cgi/doi/10.1104/pp.15.01548
702 Plant Physiology�, February 2016, Vol. 170, pp. 702–716, www.plantphysiol.org � 2016 American Society of Plant Biologists. All Rights Reserved. www.plantphysiol.orgon January 5, 2020 - Published by Downloaded from
Copyright © 2016 American Society of Plant Biologists. All rights reserved.

squalene (C30), organism-specific botryococcene (C30),methylated squalene (C31–C34), and methylatedbotryococcene (C31–C37; Metzger et al., 1988; Huangand Poulter, 1989; Okada et al., 1995), which can bereadily converted into all classes of combustible fuelsunder hydrocracking conditions (Hillen et al., 1982).The unique biosynthetic mechanism for the triter-
penes in B. braunii was recently described by Niehauset al. (2011), and a series of novel squalene synthase-like genes were identified (Fig. 1). In short, squalenesynthase-like enzyme, SSL-1, performs a head-to-headcondensation of two farnesyl diphosphate (FPP) mole-cules into presqualene diphosphate, followed by a re-ductive rearrangement to yield squalene (C30) by theenzyme SSL-2, or is converted by SSL-3 to formbotryococcene through a different reductive rear-rangement (Niehaus et al., 2011). Methylated deriva-tives are the dominant triterpene species generated byB. braunii race B (Metzger, 1985; Metzger et al., 1988),and these derivatives are known to yield higher qualityfuels due to their high energy content and the hydro-cracking products derived by virtue of having morehydrocarbon branches. Triterpene methyltransferases(TMTs) that can methylate squalene and botryococcenehave been successfully characterized by Niehaus et al.(2012). TRITERPENE METHYLTRANSFERASE1(TMT-1) and TMT-2 prefer squalene C30 as their sub-strate for the production of monomethylated (C31) ordimethylated (C32) squalene, while TMT-3 prefersbotryococcene as its substrate for the biosynthesis ofmonomethylated (C31) or dimethylated (C32) botryo-coccene (Fig. 1). These TMTs are believed to be insolu-ble enzymes; they exhibit large hydrophobic areas, andtheir activities were only observed in vitro using yeastmicrosomal preparations (no activity was observedwhen expressed in bacteria; Niehaus et al., 2012).
Like the majority of identified methyltransferases,these TMTs utilize the methyl donor S-adenosyl me-thionine (SAM), which is ubiquitous in prokaryotes andeukaryotes (Scheer et al., 2011; Liscombe et al., 2012). Inplants, SAM is one of the most abundant cofactors(Fontecave et al., 2004; Sauter et al., 2013) and is syn-thesized exclusively in the cytosol (Wallsgrove et al.,1983; Ravanel et al., 1998, 2004; Bouvier et al., 2006).While it is used predominantly as a methyl donor in themethylation reaction (Ravanel et al., 2004), it also servesas the primary precursor for the biosynthesis of ethyl-ene (Wang et al., 2002b), polyamines (Kusano et al.,2008), and nicotianamine (Takahashi et al., 2003), whichplay a variety of important roles for plant growth anddevelopment (Huang et al., 2012; Sauter et al., 2013).The SAM present in organelles, like the chloroplast,appears to be imported from the cytosol by specificSAM/S-adenosylhomocysteine exchange transportersthat reside on the envelope membranes of plastids(Ravanel et al., 2004; Bouvier et al., 2006). The importedSAM is involved in the biogenesis of Asp-derivedamino acids (Curien et al., 1998; Jander and Joshi,2009; Sauter et al., 2013) and serves as the methyl donorfor the methylation of macromolecules, such as plastidDNA (Nishiyama et al., 2002; Ahlert et al., 2009) andproteins (Houtz et al., 1989; Niemi et al., 1990; Ying et al.,1999; Trievel et al., 2003; Alban et al., 2014), and smallmolecule metabolites, such as prenylipids (e.g. plasto-quinone, tocopherol, chlorophylls, and phylloquinone;Bouvier et al., 2005, 2006; DellaPenna, 2005).
Although plants andmicrobes are the natural sourcesfor useful terpenes, most of them are produced in verysmall amounts and often as complex mixtures. In con-trast, B. braunii produces large quantities of triterpenes,but its slow growth makes it undesirable as a via-ble production platform (Niehaus et al., 2011).
Figure 1. Depiction of the catalytic roles of novel SSLand TMT enzymes in B. braunii race B and their pu-tative contributions to the triterpene constituents(Niehaus et al., 2011; Niehaus et al., 2012). SSL-1 cat-alyzes the condensation of two farnesyl diphos-phate (FPP) molecules to presqualene diphosphate(PSPP), which is converted to either squalene orbotryococcene by SSL-2 or SSL-3, respectively.Squalene can also be synthesized directly from thecondensation of two FPP molecules catalyzed bysqualene synthase (SQS). TMT-1 and TMT-2 transferthe methyl donor group from S-adenosylmethionine(SAM) to squalene to form monomethylated anddimethylated squalene, whereas TMT-3 acts onbotryococcene to form monomethylated and dime-thylated botryococcene (Niehaus et al., 2012).
Plant Physiol. Vol. 170, 2016 703
Triterpene Metabolism in Plants
www.plantphysiol.orgon January 5, 2020 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

Nevertheless, metabolic engineering and syntheticbiology offer many strategies to manipulate terpenemetabolism in various biological systems to achievehigh-value terpene production with high yield andhigh fidelity for particular practical applications(Nielsen and Keasling, 2011). Many successeshave been achieved in engineering valuable terpenesin heterotrophic microbes, such as Escherichia coli(Nishiyama et al., 2002; Martin et al., 2003; Ajikumaret al., 2010) and Saccharomyces cerevisiae (Ro et al., 2006;Takahashi et al., 2007; Westfall et al., 2012; Zhuangand Chappell, 2015). The strategies developed inthese efforts usually take advantage of specific mi-crobe strains whose innate biosynthetic machineryis genetically modified to accumulate certain pre-nyldiphosphate precursors (e.g. IPP or FPP), whichcan be utilized by other introduced terpene synthase(s) for the production of the desired terpene(s). Forexample, greater than 900 mg L21 bisabolene wasproducedwhen bisabolene synthase genes from plantswere introduced into FPP-overproducing E. coli or S.cerevisiae strains (Peralta-Yahya et al., 2011). Highlevels of farnesane production for diesel fuels werealso achieved by reductive hydrogenation of its pre-cursor farnesene, which was generated from a genet-ically engineered yeast (e.g. Saccharomyces cerevisiae)strain using plant farnesene synthases (Renninger andMcPhee, 2008; Ubersax and Platt, 2010). However,terpene production using microbial platforms is stilldependent on exogenous feedstocks (i.e. sugars) andelaborate production facilities, both of which addsignificantly to their production costs.
Compared with microbial systems, engineering ter-pene production in plant systems seems like an attrac-tive target as well. This is because plants can takeadvantage of photosynthesis by using atmospheric CO2as their carbon resource instead of relying on exoge-nous carbon feedstocks. Moreover, crop plants such astobacco (Nicotiana tabacum) can generate a large amountof green tissues efficiently when grown for biomassproduction (Schillberg et al., 2003; Andrianov et al.,2010), making them a robust, sustainable, and scalableplatform for large-scale terpene production. Nonethe-less, compared with microbial platforms, there areonly a few examples of elevating terpene production inbioengineered plants. This is due partly to higher plantsbeing complex multicellular organisms, in which ter-pene metabolism generally utilizes more complexinnate machinery that can be compartmentalized in-tracellularly and to cell/tissue specificities (Lange andAhkami, 2013; Kempinski et al., 2015). Significant ef-forts have been made to overcome these obstacles toimprove the production of valuable terpenes inplants, including monoterpenes (Lücker et al., 2004;Ohara et al., 2010; Lange et al., 2011), sesquiterpenes(Aharoni et al., 2003; Kappers et al., 2005; Wu et al.,2006; Davidovich-Rikanati et al., 2008), diterpenes(Besumbes et al., 2004; Anterola et al., 2009), andtriterpenes (Inagaki et al., 2011; Wu et al., 2012).Among these, engineering terpene metabolism into a
subcellular organelle, where the engineered enzymes/pathways can utilize unlimited/unregulated precur-sors as substrates, appears most successful. For ex-ample, Wu et al. (2006, 2012) expressed an avianfarnesyl diphosphate synthase (FPS) with foreignsesquiterpene/triterpene synthases targeted to theplastid to divert the IPP/DMAPP pool from the plas-tidic MEP pathway to synthesize high levels of thenovel sesquiterpenes patchoulol and amorpha-4,11-diene up to 30 mg g21 fresh weight and the triterpenesqualene up to 1,000 mg g21 fresh weight. This strategyappears to be particularly robust because it avoidspossible endogenous regulation of sesquiterpene andtriterpene biosynthesis, which occurs normally in thecytoplasm, and relies upon more plastic precursorpools of IPP/DMAPP inherent in the plastid, whichare primarily derived from the local CO2 fixation(Wright et al., 2014).
The goal of this study was to evaluate the prospectsfor engineering advanced features of triterpene me-tabolism from B. braunii into tobacco and, thus, to probethe innate intricacies of isoprenoid metabolism inplants. In order to achieve this, we first introduced thekey steps of botryococcene biosynthesis into specificsubcellular compartments of tobacco cells under thedirection of constitutive or trichome-specific promoters.The transgenic lines expressing the enzymes in thechloroplast were found to accumulate the highest levelsof botryococcene. Triterpene methyltransferases werenext introduced into the same intracellular compart-ments of selected high-triterpene-accumulating lines. Ahigh yield of methylated triterpenes was also achievedin transgenic lines when the TMTs were targeted to thechloroplast. Through careful comparison of the levels oftriterpenes and the methylated triterpene products inthe various transgenic lines, we have also gained adeeper insight into the subcellular distribution of thetriterpene products in these transgenic lines as well as abetter understanding of methylation metabolism forspecialized metabolites in particular compartments.These findings all contribute to our understanding ofthe regulatory elements that control carbon fluxthrough the innate terpene biosynthetic pathways op-erating in plants.
RESULTS
Targeting Botryococcene Biosynthesis to the CytoplasmVersus Chloroplasts
An earlier study demonstrated that plastid-targetedengineering of a foreign squalene synthase (SQS) andan FPS can successfully divert carbon flux from theMEP pathway to accumulate a high level of squalenein transgenic tobacco (Wu et al., 2012). That studyrevealed that the availability of IPP/DMAPP precur-sors was adequate but that strong regulatory mecha-nisms were absent in the chloroplast for novel squalene(C30) production to occur. This, in turn, led us to utilize
704 Plant Physiol. Vol. 170, 2016
Jiang et al.
www.plantphysiol.orgon January 5, 2020 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
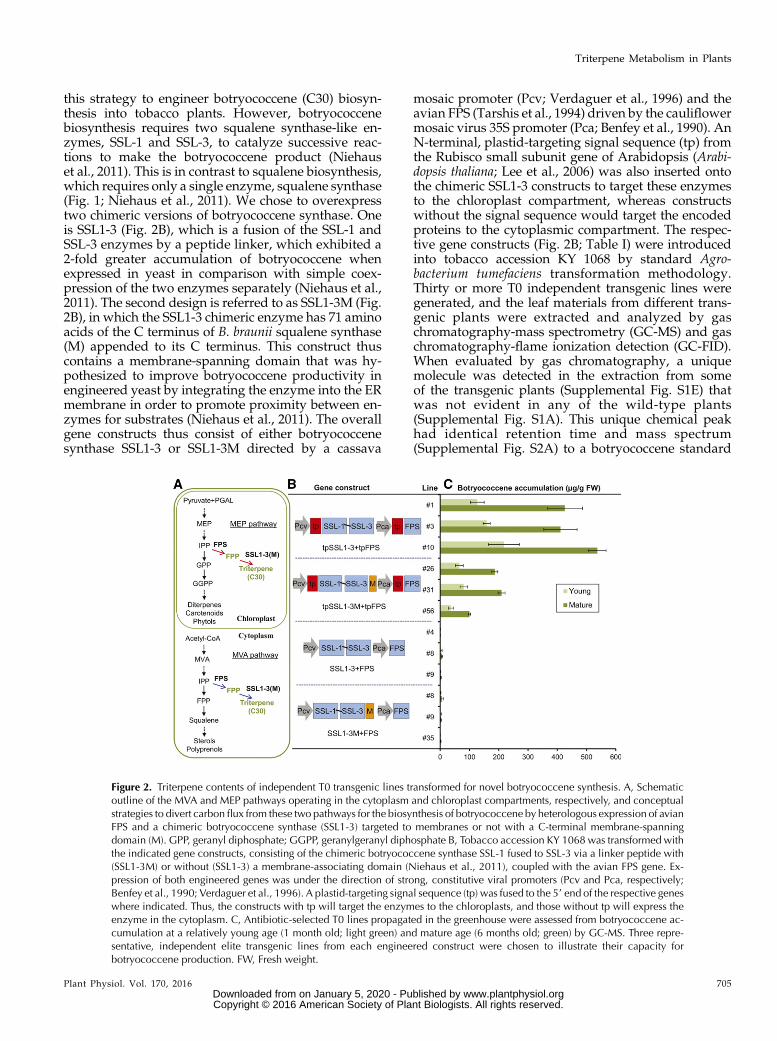
this strategy to engineer botryococcene (C30) biosyn-thesis into tobacco plants. However, botryococcenebiosynthesis requires two squalene synthase-like en-zymes, SSL-1 and SSL-3, to catalyze successive reac-tions to make the botryococcene product (Niehauset al., 2011). This is in contrast to squalene biosynthesis,which requires only a single enzyme, squalene synthase(Fig. 1; Niehaus et al., 2011). We chose to overexpresstwo chimeric versions of botryococcene synthase. Oneis SSL1-3 (Fig. 2B), which is a fusion of the SSL-1 andSSL-3 enzymes by a peptide linker, which exhibited a2-fold greater accumulation of botryococcene whenexpressed in yeast in comparison with simple coex-pression of the two enzymes separately (Niehaus et al.,2011). The second design is referred to as SSL1-3M (Fig.2B), in which the SSL1-3 chimeric enzyme has 71 aminoacids of the C terminus of B. braunii squalene synthase(M) appended to its C terminus. This construct thuscontains a membrane-spanning domain that was hy-pothesized to improve botryococcene productivity inengineered yeast by integrating the enzyme into the ERmembrane in order to promote proximity between en-zymes for substrates (Niehaus et al., 2011). The overallgene constructs thus consist of either botryococcenesynthase SSL1-3 or SSL1-3M directed by a cassava
mosaic promoter (Pcv; Verdaguer et al., 1996) and theavian FPS (Tarshis et al., 1994) driven by the cauliflowermosaic virus 35S promoter (Pca; Benfey et al., 1990). AnN-terminal, plastid-targeting signal sequence (tp) fromthe Rubisco small subunit gene of Arabidopsis (Arabi-dopsis thaliana; Lee et al., 2006) was also inserted ontothe chimeric SSL1-3 constructs to target these enzymesto the chloroplast compartment, whereas constructswithout the signal sequence would target the encodedproteins to the cytoplasmic compartment. The respec-tive gene constructs (Fig. 2B; Table I) were introducedinto tobacco accession KY 1068 by standard Agro-bacterium tumefaciens transformation methodology.Thirty or more T0 independent transgenic lines weregenerated, and the leaf materials from different trans-genic plants were extracted and analyzed by gaschromatography-mass spectrometry (GC-MS) and gaschromatography-flame ionization detection (GC-FID).When evaluated by gas chromatography, a uniquemolecule was detected in the extraction from someof the transgenic plants (Supplemental Fig. S1E) thatwas not evident in any of the wild-type plants(Supplemental Fig. S1A). This unique chemical peakhad identical retention time and mass spectrum(Supplemental Fig. S2A) to a botryococcene standard
Figure 2. Triterpene contents of independent T0 transgenic lines transformed for novel botryococcene synthesis. A, Schematicoutline of the MVA and MEP pathways operating in the cytoplasm and chloroplast compartments, respectively, and conceptualstrategies to divert carbon flux from these two pathways for the biosynthesis of botryococcene by heterologous expression of avianFPS and a chimeric botryococcene synthase (SSL1-3) targeted to membranes or not with a C-terminal membrane-spanningdomain (M). GPP, geranyl diphosphate; GGPP, geranylgeranyl diphosphate B, Tobacco accession KY 1068 was transformed withthe indicated gene constructs, consisting of the chimeric botryococcene synthase SSL-1 fused to SSL-3 via a linker peptide with(SSL1-3M) or without (SSL1-3) a membrane-associating domain (Niehaus et al., 2011), coupled with the avian FPS gene. Ex-pression of both engineered genes was under the direction of strong, constitutive viral promoters (Pcv and Pca, respectively;Benfey et al., 1990; Verdaguer et al., 1996). A plastid-targeting signal sequence (tp) was fused to the 59 end of the respective geneswhere indicated. Thus, the constructs with tp will target the enzymes to the chloroplasts, and those without tp will express theenzyme in the cytoplasm. C, Antibiotic-selected T0 lines propagated in the greenhouse were assessed from botryococcene ac-cumulation at a relatively young age (1 month old; light green) and mature age (6 months old; green) by GC-MS. Three repre-sentative, independent elite transgenic lines from each engineered construct were chosen to illustrate their capacity forbotryococcene production. FW, Fresh weight.
Plant Physiol. Vol. 170, 2016 705
Triterpene Metabolism in Plants
www.plantphysiol.orgon January 5, 2020 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
(Niehaus et al., 2011) andwas confirmed as botryococceneby 1H-NMR and 13C-NMR analyses (Supplemental Figs.S3 and S4).
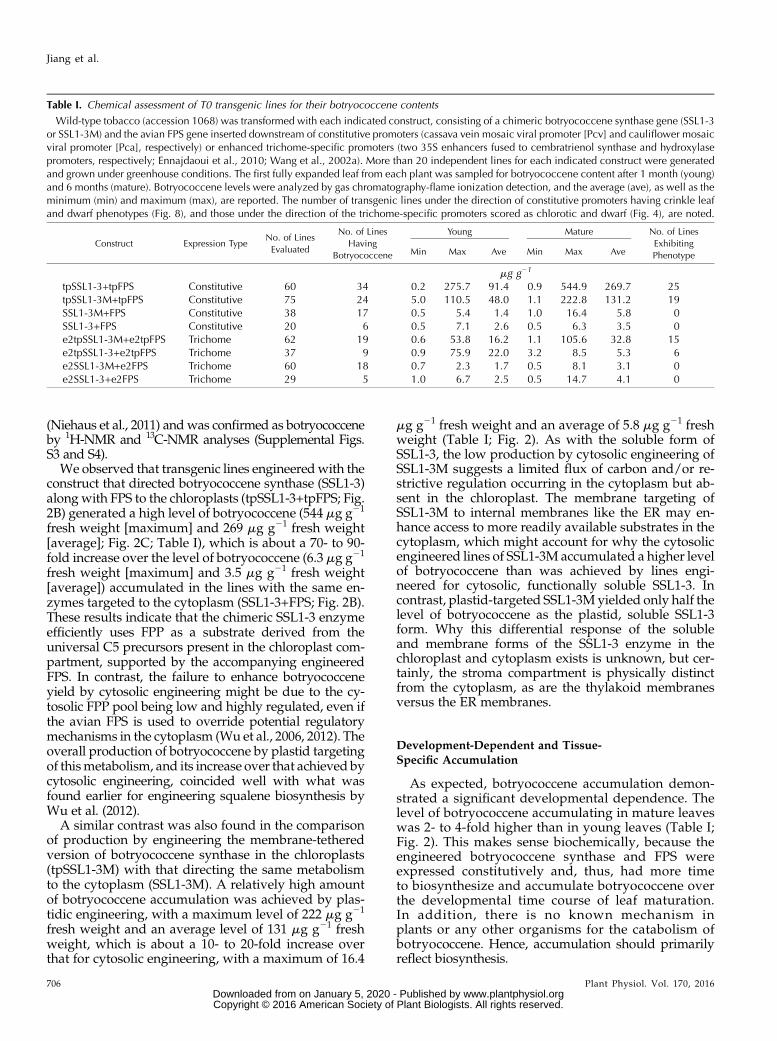
We observed that transgenic lines engineeredwith theconstruct that directed botryococcene synthase (SSL1-3)along with FPS to the chloroplasts (tpSSL1-3+tpFPS; Fig.2B) generated a high level of botryococcene (544 mg g21
fresh weight [maximum] and 269 mg g21 fresh weight[average]; Fig. 2C; Table I), which is about a 70- to 90-fold increase over the level of botryococcene (6.3 mg g21
fresh weight [maximum] and 3.5 mg g21 fresh weight[average]) accumulated in the lines with the same en-zymes targeted to the cytoplasm (SSL1-3+FPS; Fig. 2B).These results indicate that the chimeric SSL1-3 enzymeefficiently uses FPP as a substrate derived from theuniversal C5 precursors present in the chloroplast com-partment, supported by the accompanying engineeredFPS. In contrast, the failure to enhance botryococceneyield by cytosolic engineering might be due to the cy-tosolic FPP pool being low and highly regulated, even ifthe avian FPS is used to override potential regulatorymechanisms in the cytoplasm (Wu et al., 2006, 2012). Theoverall production of botryococcene by plastid targetingof thismetabolism, and its increase over that achieved bycytosolic engineering, coincided well with what wasfound earlier for engineering squalene biosynthesis byWu et al. (2012).
A similar contrast was also found in the comparisonof production by engineering the membrane-tetheredversion of botryococcene synthase in the chloroplasts(tpSSL1-3M) with that directing the same metabolismto the cytoplasm (SSL1-3M). A relatively high amountof botryococcene accumulation was achieved by plas-tidic engineering, with a maximum level of 222 mg g21
fresh weight and an average level of 131 mg g21 freshweight, which is about a 10- to 20-fold increase overthat for cytosolic engineering, with a maximum of 16.4
mg g21 fresh weight and an average of 5.8 mg g21 freshweight (Table I; Fig. 2). As with the soluble form ofSSL1-3, the low production by cytosolic engineering ofSSL1-3M suggests a limited flux of carbon and/or re-strictive regulation occurring in the cytoplasm but ab-sent in the chloroplast. The membrane targeting ofSSL1-3M to internal membranes like the ER may en-hance access to more readily available substrates in thecytoplasm, which might account for why the cytosolicengineered lines of SSL1-3M accumulated a higher levelof botryococcene than was achieved by lines engi-neered for cytosolic, functionally soluble SSL1-3. Incontrast, plastid-targeted SSL1-3M yielded only half thelevel of botryococcene as the plastid, soluble SSL1-3form. Why this differential response of the solubleand membrane forms of the SSL1-3 enzyme in thechloroplast and cytoplasm exists is unknown, but cer-tainly, the stroma compartment is physically distinctfrom the cytoplasm, as are the thylakoid membranesversus the ER membranes.
Development-Dependent and Tissue-Specific Accumulation
As expected, botryococcene accumulation demon-strated a significant developmental dependence. Thelevel of botryococcene accumulating in mature leaveswas 2- to 4-fold higher than in young leaves (Table I;Fig. 2). This makes sense biochemically, because theengineered botryococcene synthase and FPS wereexpressed constitutively and, thus, had more timeto biosynthesize and accumulate botryococcene overthe developmental time course of leaf maturation.In addition, there is no known mechanism inplants or any other organisms for the catabolism ofbotryococcene. Hence, accumulation should primarilyreflect biosynthesis.
Table I. Chemical assessment of T0 transgenic lines for their botryococcene contents
Wild-type tobacco (accession 1068) was transformed with each indicated construct, consisting of a chimeric botryococcene synthase gene (SSL1-3or SSL1-3M) and the avian FPS gene inserted downstream of constitutive promoters (cassava vein mosaic viral promoter [Pcv] and cauliflower mosaicviral promoter [Pca], respectively) or enhanced trichome-specific promoters (two 35S enhancers fused to cembratrienol synthase and hydroxylasepromoters, respectively; Ennajdaoui et al., 2010; Wang et al., 2002a). More than 20 independent lines for each indicated construct were generatedand grown under greenhouse conditions. The first fully expanded leaf from each plant was sampled for botryococcene content after 1 month (young)and 6 months (mature). Botryococcene levels were analyzed by gas chromatography-flame ionization detection, and the average (ave), as well as theminimum (min) and maximum (max), are reported. The number of transgenic lines under the direction of constitutive promoters having crinkle leafand dwarf phenotypes (Fig. 8), and those under the direction of the trichome-specific promoters scored as chlorotic and dwarf (Fig. 4), are noted.
Construct Expression TypeNo. of Lines
Evaluated
No. of Lines
Having
Botryococcene
Young Mature No. of Lines
Exhibiting
PhenotypeMin Max Ave Min Max Ave
mg g21
tpSSL1-3+tpFPS Constitutive 60 34 0.2 275.7 91.4 0.9 544.9 269.7 25tpSSL1-3M+tpFPS Constitutive 75 24 5.0 110.5 48.0 1.1 222.8 131.2 19SSL1-3M+FPS Constitutive 38 17 0.5 5.4 1.4 1.0 16.4 5.8 0SSL1-3+FPS Constitutive 20 6 0.5 7.1 2.6 0.5 6.3 3.5 0e2tpSSL1-3M+e2tpFPS Trichome 62 19 0.6 53.8 16.2 1.1 105.6 32.8 15e2tpSSL1-3+e2tpFPS Trichome 37 9 0.9 75.9 22.0 3.2 8.5 5.3 6e2SSL1-3M+e2FPS Trichome 60 18 0.7 2.3 1.7 0.5 8.1 3.1 0e2SSL1-3+e2FPS Trichome 29 5 1.0 6.7 2.5 0.5 14.7 4.1 0
706 Plant Physiol. Vol. 170, 2016
Jiang et al.
www.plantphysiol.orgon January 5, 2020 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

To determine if the various transgenic plants accu-mulated triterpenes in other tissues besides leaves,the triterpene chemical profiles across various tissueswere determined for select transgenic lines (Fig. 3). Thesqualene accumulating lines selected for comparisonwere homozygous T2 generation plants, while the linesselected for botryococcene accumulation were T1 het-erozygous lines that had been selected for transgenepresence under antibiotic pressure. The squalene-accumulating lines were previously described by Wuet al. (2012) and designated as tpSS+tpFPS #5. Thebotryococcene lines were tpSSL1-3M+tpFPS #31and tpSSL1-3+tpFPS #10. Triterpene content (eithersqualene or botryococcene) was found in all tissuesexamined, but the levels varied dramatically. Leafaccumulation was greatest and up to 64-fold morethan that found in roots. Low amounts of triterpenes,never exceeding 25 mg g21, were also observed inother tissues, like veins and stems.
Trichome-Specific Expression ofBotryococcene Metabolism
Trichomes are specialized organs located on the sur-face of the aerial parts of plant species, which can bethe site of abundant specialized metabolite biosyn-thesis, accumulation, and secretion. In tobacco, forinstance, up to 15% of the leaf dry weight has beenattributed to the secretion of leaf exudate from tri-chomes (Wagner et al., 2004). Such a large contribu-tion to leaf biomass relative to the actual volume ofthe glandular trichomes makes trichome engineeringan attractive target for metabolite bioengineering
(Ennajdaoui et al., 2010). Hence, in an effort to directbotryococcene biosynthesis to secretory trichomes, thetrichome-specific promoters of the cembratrienol syn-thase and cembratrienol hydroxylase genes (Wanget al., 2002a; Ennajdaoui et al., 2010) were used to directbotryococcene synthase and FPS expression, respec-tively, to the secretory trichomes of tobacco. Tostrengthen the overall trichome-specific expression, the35S double enhancer element was also appended to the59 end of each of the trichome promoters (Wu et al.,2012). Four constructs harboring SSL1-3 or SSL1-3Mwith FPS, plus or minus chloroplast-targeting N-terminalsequences (tp), were thus introduced into the Nicoti-ana tabacum accession 1068, an accession documentedto have high trichome density (Nielsen, 1982). Almost30 independent transgenic lines were generated foreach construct, the resulting transgenic lines werepropagated in greenhouse facilities, and differentlysized leaves were analyzed for their botryococcenecontent (Table I).
Targeting botryococcene metabolism to the chloro-plasts of trichomes resulted in only a modest accumu-lation of botryococcene in young and maturing leaftissue (less than 30 mg g21 fresh weight on average),which was 10- to 20-fold greater than that accumulatedin the lines with the enzymes directed to the cytoplasmof trichome cells (Table I). However, more unexpect-edly, plants with putative enhanced, trichome-specificexpression of SSL1-3 and FPS targeted to the chloro-plast showed a strong chlorotic, white, mottling, anddwarf phenotype (Fig. 4), which certainly contributedto difficulties in propagating these materials. Thisphenotype was also much more severe than anythingobserved with the trichome-specific expression of
Figure 3. Triterpene accumulation in dif-ferent tissues of transgenic lines. Selecttransgenic lines and wild-type plants weregrown in tissue culture for 3 months. Plantmaterials from different tissues (root, stem,vein, and leaf) were then sampled for theirtriterpene contents. The gene constructand the accumulating triterpene moni-tored are indicated. FW, Fresh weight.
Figure 4. A, Phenotype of a transgenic plant express-ing trichome-specific, plastid-targeted botryococcenebiosynthesis (tpSSL1-3+tpFPS; right) in comparisonwith a wild-type control plant (left). The control plantwas initiated from seed,while the transgenic line is a T0generation plant transferred from the tissue culture re-generation process. The plants were grown undergreenhouse conditions for approximately 2 months. B,Closeup of the transgenic line in A.
Plant Physiol. Vol. 170, 2016 707
Triterpene Metabolism in Plants
www.plantphysiol.orgon January 5, 2020 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.
squalene biosynthesis (Wu et al., 2012). These adversephenotypes were surprising, because we had hoped toengineer botryococcene metabolism away from tissuesimportant for normal growth and development. In-stead, we speculate that the trichome-specific pro-moters in combination with the double 35S enhancersmay not behave in the anticipated manner (i.e. are nottrichome specific) and exhibit ectopic expression(higher than that of the other constitutive promotersused here) in tissues that are more sensitive to this typeof metabolism (e.g. meristems), resulting in the delete-rious phenotypes described.
Engineering Triterpene Methyltransferases into ParticularSubcellular Compartments of Tobacco Plants
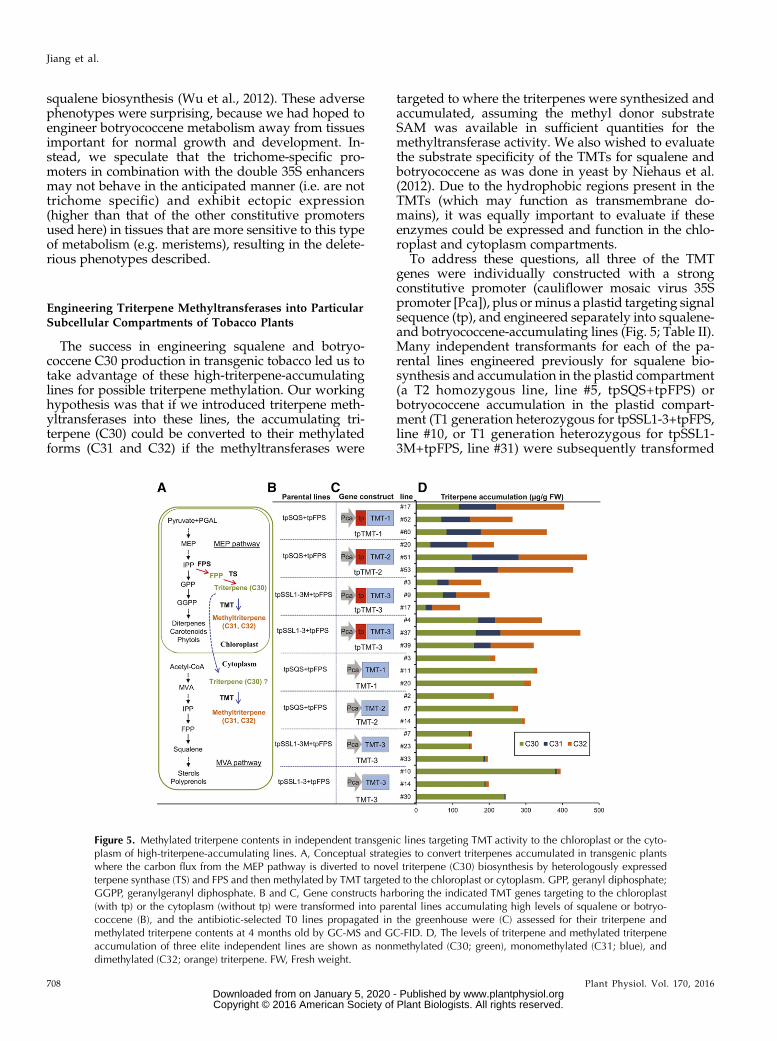
The success in engineering squalene and botryo-coccene C30 production in transgenic tobacco led us totake advantage of these high-triterpene-accumulatinglines for possible triterpene methylation. Our workinghypothesis was that if we introduced triterpene meth-yltransferases into these lines, the accumulating tri-terpene (C30) could be converted to their methylatedforms (C31 and C32) if the methyltransferases were
targeted to where the triterpenes were synthesized andaccumulated, assuming the methyl donor substrateSAM was available in sufficient quantities for themethyltransferase activity. We also wished to evaluatethe substrate specificity of the TMTs for squalene andbotryococcene as was done in yeast by Niehaus et al.(2012). Due to the hydrophobic regions present in theTMTs (which may function as transmembrane do-mains), it was equally important to evaluate if theseenzymes could be expressed and function in the chlo-roplast and cytoplasm compartments.
To address these questions, all three of the TMTgenes were individually constructed with a strongconstitutive promoter (cauliflower mosaic virus 35Spromoter [Pca]), plus orminus a plastid targeting signalsequence (tp), and engineered separately into squalene-and botryococcene-accumulating lines (Fig. 5; Table II).Many independent transformants for each of the pa-rental lines engineered previously for squalene bio-synthesis and accumulation in the plastid compartment(a T2 homozygous line, line #5, tpSQS+tpFPS) orbotryococcene accumulation in the plastid compart-ment (T1 generation heterozygous for tpSSL1-3+tpFPS,line #10, or T1 generation heterozygous for tpSSL1-3M+tpFPS, line #31) were subsequently transformed
Figure 5. Methylated triterpene contents in independent transgenic lines targeting TMT activity to the chloroplast or the cyto-plasm of high-triterpene-accumulating lines. A, Conceptual strategies to convert triterpenes accumulated in transgenic plantswhere the carbon flux from the MEP pathway is diverted to novel triterpene (C30) biosynthesis by heterologously expressedterpene synthase (TS) and FPS and then methylated by TMT targeted to the chloroplast or cytoplasm. GPP, geranyl diphosphate;GGPP, geranylgeranyl diphosphate. B and C, Gene constructs harboring the indicated TMT genes targeting to the chloroplast(with tp) or the cytoplasm (without tp) were transformed into parental lines accumulating high levels of squalene or botryo-coccene (B), and the antibiotic-selected T0 lines propagated in the greenhouse were (C) assessed for their triterpene andmethylated triterpene contents at 4 months old by GC-MS and GC-FID. D, The levels of triterpene and methylated triterpeneaccumulation of three elite independent lines are shown as nonmethylated (C30; green), monomethylated (C31; blue), anddimethylated (C32; orange) triterpene. FW, Fresh weight.
708 Plant Physiol. Vol. 170, 2016
Jiang et al.
www.plantphysiol.orgon January 5, 2020 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

with each of the methyltransferases targeted to the cy-toplasm (TMT-1, TMT-2, and TMT-3) or plastid com-partment (tpTMT-1, tpTMT-2, and tpTMT-3), and theresulting transformants were screened for their tri-terpene chemical content and composition. More spe-cifically, each of the transgenic lines was evaluatedfor triterpene (C30), monomethylated (C31), anddimethylated (C32) triterpene contents by GC-MS(Supplemental Figs. S1 and S2). Because of the largenumber of transgenic lines generated and evaluated,Table II summarizes the data for all the various lines,and Figure 5 illustrates the design strategies and pro-vides analysis for three example lines for each TMTconstruct.When the transgenic TMT-1 enzyme was targeted to
the chloroplasts of the squalene-accumulating line, themaximum level of squalene methylation was 91%, withthe average across all the lines being 65% (Table II). Ofthe methylated forms, approximately two-thirds wasin the dimethylated form (Table II; Fig. 5). When TMT-2was plastid targeted in the same parental backgroundline, the maximum level of squalene methylation was82% of the total, with an average of 51% for the 36 linesaccumulating methylated squalenes (Table II). In con-trast to the TMT-1 lines, only about one-third of themethylated squalene was in the dimethylated form inthe TMT-2 plastid-targeted lines (Table II; Fig. 5).When TMT-1 or TMT-2 was targeted to the cyto-
plasm rather than the chloroplasts, only 4% of the totalsqualene, on average, was methylated by either of themethyltransferases. However, 100% of the methylatedsqualenewas in the dimethylated form (Table II; Fig. 5).The small amount of squalene available to the
methyltransferases under these conditions could arisefrom cytosolic squalene synthesized by the native ma-chinery or that synthesized by mistargeted engineeredsqualene synthase not properly transported into thechloroplast.
In contrast, only 5% (average) of the total squalenewas methylated when TMT-3 was targeted to thechloroplast of the high-squalene-producing line, sug-gesting that TMT-3 exhibited weak catalytic activitytoward squalene (Supplemental Table S1), which wasalso observedwhen substrate specificity for TMT-3 wasinvestigated in yeast (Niehaus et al., 2012). Targetedexpression of TMT-3 to the cytoplasm in the same linedid not result in any methylation products, also cor-roborating the inability of TMT-3 to utilize the limitedamounts of squalene found in this compartment(Supplemental Table S1).
When the TMT-3 enzyme was introduced into thebotryococcene-accumulating lines, we observed a largeproportion of methylated botryococcene only whenTMT-3was targeted to the chloroplasts (Table II; Fig. 5).Using parental line tpSSL1-3M+tpFPS, 87% of thebotryococcene was maximally methylated, with 54% ofthe total botryococcene being methylated on averageacross all 18 lines evaluated. In comparison, maximally,66% of the botryococcene was methylated in parentalline tpSSL1-3+tpFPS, but more typically, 35% (average)was methylated (Table II). The apparent improved ef-ficiency of botryococcenemethylation in the tpSSL1-3Mline versus the tpSSL1-3 line, however, might be moreof a reflection on the total botryococcene levels ratherthan the efficiency of themethylation reaction itself. Theline engineered with tpSSL1-3M accumulates about
Table II. Chemical assessment of T0 transgenic lines targeting TMT activity to the chloroplast or the cytoplasm of high-squalene-accumulating(tpSQS) or botryococcene-accumulating (tpSSL1-3 and tpSSL1-3M) lines for their methylated triterpene contents
More than 30 independent lines were generated for each transformation construct, consisting of one of the three TMT genes targeting triterpenemethyltransferase activity to the chloroplast (with tp) or the cytoplasm (without tp) of the indicated parental lines. The squalene-accumulating line(tpSQS+tpFPS #5) or the botryococcene-accumulating lines (tpSSL1-3+tpFPS #10 and tpSSL1-3M+tpFPS #31) were transformed with the indicatedTMT construct and evaluated after 4 months by GC-FID/GC-MS for methylated triterpenes. The number of transgenic lines accumulating methylatedsqualene or botryococcene was scored, along with the average (ave) percentage of nonmethylated (C30), monomethylated (C31), or dimethylated(C32) triterpenes relative to total triterpene (C30+C31+C32) content. The percentage of methylated triterpene (C31+C32) to total triterpene isdenoted as conversion, and the number of plants with less than 10%, between 10% and 50%, and more than 50% conversion for each construct wascounted accordingly. The highest amount of conversion as well as the average conversions are noted. The average total triterpene content for eachline is also noted, as is the average triterpene content for three or four plants for each of the parental (control) lines.
Construct Parental Line
No. of
Lines
Evaluated
No. of Lines
Having Methylated
Triterpene
Triterpene/Total (Ave) Conversion: (C31+C32)/Total Total
Triterpene
(Ave)
mg g21C30 C31 C32
No. ,10%
No. 10%–
50%
No. .50%
Highest Ave
tpTMT-1 tpSQS+tpFPS #5 75 47 34% 25% 41% 2 26 19 91% 65% 222tpTMT-2 tpSQS+tpFPS #5 69 36 50% 31% 18% 2 17 17 82% 51% 301TMT-1 tpSQS+tpFPS #5 36 11 96% 0 4% 11 0 0 7% 4% 251TMT-2 tpSQS+tpFPS #5 33 12 96% 0 4% 12 0 0 6% 4% 241tpTMT-3 tpSSL1-3M+tpFPS #31 76 18 46% 17% 37% 3 3 12 87% 54% 131TMT-3 tpSSL1-3M+tpFPS #31 40 19 94% 3% 3% 15 4 0 14% 6% 126tpTMT-3 tpSSL1-3+tpFPS #10 69 29 65% 10% 25% 0 23 6 66% 35% 283TMT-3 tpSSL1-3+tpFPS #10 40 17 97% 1% 2% 16 1 0 10% 3% 294
tpSQS+tpFPS #5 4 0 100% 0 0 4 0 0 0 0 234tpSSl1-3+tpFPS #10 3 0 100% 0 0 3 0 0 0 0 237tpSSL1-3M+tpFPS #31 3 0 100% 0 0 3 0 0 0 0 125
Plant Physiol. Vol. 170, 2016 709
Triterpene Metabolism in Plants
www.plantphysiol.orgon January 5, 2020 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

half as much botryococcene as tpSSL1-3, and assumingsimilar expression levels of TMT-3 in both parental linesand comparable amounts of SAM availability, thegreater percentage of methylated botryococcene intpSSL1-3M may simply reflect the smaller pool ofbotryococcene available for secondary modifications.Little to none of the chloroplast-synthesized botryo-coccene was methylated when either TMT-1 or TMT-2was targeted to the chloroplast (Supplemental TableS1), further demonstrating the striking substrate pref-erence of TMT-3 for botryococcenes and of TMT-1 andTMT-2 for squalene.
By comparison, only a small proportion of meth-ylated botryococcene was formed when TMT-3 wastargeted to the cytoplasm. For parental line tpSSL1-3M+tpFPS, 14% maximal and on average only 6% oftotal botryococcene was methylated, whereas 10%maximal and on average only 3% of the botryococceneproduced in parental line tpSSL1-3+tpFPS was meth-ylated (Table II). As suggested above for squalene, thelow level of methylated botryococcene produced bycytosolic TMT-3 could arise from either botryococcene(C30) produced by mistargeted SSL1-3(M), generat-ing triterpene substrate in the cytoplasm, or leakage ofbotryococcene from the chloroplast-synthesized pool.
The observation of methylated squalene in the lineswith squalene biosynthesis targeted to the chloroplast,yet the methyltransferases expressed in the cytoplasm,raised a question about what pool of squalene was be-ing methylated. Was it the squalene synthesized in thechloroplast by the engineered squalene synthase, orcould it be a reflection of the squalene synthesizedby the native biosynthetic machinery operating in as-sociation with the ER? Because differentiating be-tween pools of native versus engineered squalene in thetpSQS+tpFPS line has proven to be technically difficult,an alternative approach was sought. Gene constructsdirecting expression of the TMT-1 and TMT-2 en-zymes targeted to the chloroplast and cytoplasm were
introduced into control, wild-type tobacco, and theresulting transgenic lines were screened for methylatedsqualene. Interestingly, a significant proportion ofmethylated squalene (average 41% of total squalene)was observed when TMT-1 was targeted to the chlo-roplasts (Table III), where there is no evidence forsqualene biosynthesis or accumulation (Aharoni et al.,2003). In this case, one plausible explanation is thatsome mistargeting of the engineered TMT-1 results inthe methylation of cytoplasmic biosynthesized squa-lene. In contrast, when TMT-1 and TMT-2 expressionwere targeted to the cytoplasm of wild-type plants, ahigh proportion of methylated squalene (average 72%and 67% of total squalene, respectively) was found(Table III; Fig. 6). Evenmore surprising, the level of totalsqualene (C30+C31+C32) in transgenic lines expressingTMT-1 was elevated to a maximum of 55 mg g21 freshweight and an average of 36 mg g21 fresh weight (TableIII). This was about 4- to 7-fold greater than the level ofendogenous squalene (C30) accumulating in wild-typeplants (Table III; Fig. 6). Equally important, the level ofnonmethylated squalene remained relatively constantin all these lines at 6 to 10 mg g21 fresh weight, withall the additional triterpene accumulating as mono-methylated and dimethylated squalene.
Developmental Triterpene Accumulation and Methylation
To explore the possible influence of developmentprocesses on the methylation status of squalene andbotryococcene, three independent lines for each ex-pression vector combination (squalene biosynthesistargeted to the plastid compartment with plastid-targeted TMT-1 or TMT-2, and botryococcene biosyn-thesis plus TMT-3 directed to the plastid compartment)were grown in the greenhouse for approximately 3months, then leaves at four developmental positions onthe plants were profiled for their triterpene levels and
Table III. Chemical assessment of T0 transgenic lines targeting TMT activity to the chloroplast or the cytoplasm of wild-type plants for theirmethylated squalene contents
Independent lines were generated for each of the indicated constructs consisting of either TMT-1 or TMT-2 targeted to the chloroplast (with tp) orthe cytoplasm (without tp) of wild-type plants. T0 antibiotic-selected transgenic plants were propagated in the greenhouse for up to 5 months beforetheir triterpene and methylated triterpene contents was determined by GC-MS. The number of transgenic plants accumulating methylated squalenewere scored, and their average (ave) percentage of nonmethylated (C30), monomethylated (C31), or dimethylated (C32) squalene was determinedrelative to the total squalene (C30+C31+C32). The percentage of methylated squalene (C31+C32) to total triterpene is denoted as conversion, andthe number of plants with less than 10%, between 10% and 50%, and more than 50% conversion for each construct was counted accordingly. Thehighest amount of conversion as well as the average conversions are noted. The average total squalene content for each line is also noted, as is theaverage squalene content for three of the wild-type control plants.
Construct Parental Line
No. of
Lines
Evaluated
No. of Lines
Having
Methylsqualene
Triterpene/Total (Ave) Conversion: (C31+C32)/Total Total
Triterpene
(Ave) mg g21C30 C31 C32No. ,10%
No. 10%–
50%
No. .50%
Highest Ave
tpTMT-1 Wild type 9 2 65% 0 35% 0 1 1 61% 41% 8.9tpTMT-2 Wild type 7 0 100% 0 0 0 0 0 0 0 6.3TMT-1 Wild type 14 3 24% 10% 66% 0 0 3 82% 72% 36.0TMT-2 Wild type 5 2 38% 11% 50% 0 1 1 85% 67% 16.8
Wild type 3 0 100% 0 0 0 0 0 0 0 8.2
710 Plant Physiol. Vol. 170, 2016
Jiang et al.
www.plantphysiol.orgon January 5, 2020 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

methylation status. The data in Figure 7 represent theaverage determinations for leaves from three indepen-dent transformants for each engineered combination.As expected, the total triterpene levels showed a suc-cessive increase with leaf maturation. The more matureleaves generally had more total triterpene accumulation(Fig. 7, left axis). However, the average ratio of methyl-ated (C31+C32) to total triterpene (C30+C31+C32) at thevarious leaf positions remained essentially the same,
from 55% to 75%, for all the transgenic lines (Fig. 7B,right axis).
Phenotypes of Triterpene-Accumulating Plants
Over 75% of the botryococcene-accumulating linesdirecting this metabolism to the chloroplast exhibitedmoderate to distinguishing phenotypes, includingdwarfing, chlorosis, and mottling (Table I; Fig. 8, A–C).
Figure 6. Methylated triterpene contents in independent transgenic lines targeting TMTactivity to the chloroplast or cytoplasm of wild-type plants. A, Conceptual strategies for how endogenous squalene biosynthesized by wild-type plants in the cytoplasm might bemethylated bymethyltransferases (TMT-1 and TMT-2) directed to the chloroplast or cytoplasm. B andC,Gene constructs harboring TMTgenes targeting the triterpenemethyltransferase to the chloroplast (with tp) or the cytoplasm (without tp) were transformed intowild typeplants (B), and the antibiotic-selected T0 lines propagated in the greenhouse (C)were assessed for their triterpene contents (4months old)by GC-MS. D, The levels of squalene (C30; green) and methylated squalenes (C31, blue; C32, orange) accumulating in three inde-pendent lines for each construct were determined in the first fully mature leaf of each line by GC-MS. FW, Fresh weight.
Figure 7. Accumulation of total triterpenecontent and the fraction of triterpene con-verted to methylated triterpenes over a timecourse of leaf development. Transgenic linesdirecting squalene (tpSQS+tpFPS) biosyn-thesis and squalenemethylation by TMT-1 orTMT-2 to the chloroplast compartment, ordirecting botryococcene (tpSSL1-3M+tpFPS)biosynthesis and botryococcene methyla-tion by TMT-3 to the chloroplast compart-ment, were grown under greenhouseconditions for 4 months. Leaves at thenoted positions relative to the apex of theplant were collected, and their triterpeneand methylated triterpene contents weredetermined by GC-MS. Absolute levels oftotal triterpenes (C30, C31, and C32) arereported in the histogram (blue), whilethe fraction of triterpene converted tomethylated forms are denoted by thescatterplot (red). FW, Fresh weight.
Plant Physiol. Vol. 170, 2016 711
Triterpene Metabolism in Plants
www.plantphysiol.orgon January 5, 2020 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

These phenotypes were not observed in any transgeniclines in which the engineered metabolism was targetedto the cytoplasm (Table I) and was obviously differ-ent from what was observed in any of squalene-accumulating plants (Fig. 8E, left; Wu et al., 2012). Thevisual observations thus indicated that botryococceneaccumulation had some undefined effects on chloroplastdevelopment, plant morphology, and growth, whilesqualene accumulation did not. Moreover, there werenot any noticeable differences in phenotypes betweentriterpene-accumulating plants and their respectivemethylated triterpene counterpart plants (Fig. 8, Dand E); hence, the methylation of botryococcenes wasnot able to restore the wild-type phenotype.
DISCUSSION
This work successfully transplanted several steps oftriterpene metabolism occurring in the alga B. brauniirace B into tobacco plants, leading to a high-level ac-cumulation of botryococcene and methylated triter-penes. The most robust triterpene accumulation reliedupon the strategy of diverting the five-carbon precur-sors IPP and DMAPP from theMEP pathway operatingin the chloroplast to triterpene (C30) biosynthesis by thecoexpression of an FPS plus a triterpene synthase. Theaccumulating triterpenes (C30) in the transgenic plantscould be further modified by targeting yet another en-zyme activity, TMT, to the chloroplasts of these trans-genic plants. Hence, engineering novel expression ofthe enzymes FPS, triterpene synthases, and TMTs cre-ated a new metabolic channel redirecting carbon flux
from the MEP pathway to the biosynthesis and accu-mulation of unique and unusual triterpenes (Figs. 1, 2A,and 5A).
The strategy was successful in taking advantageof engineering terpene metabolism in the plantchloroplasts. First, chloroplasts offer an abundanceof carbon passing through the MEP pathway, anddiverting an intermediate and carbon flux from thispathway does not adversely impact the biosyntheticneeds in the chloroplasts for large amounts of carotenoidsand chlorophylls. The second, equally important, ob-servation is that the chloroplast provides an idealenvironment for heterologous terpene production,perhaps due to the endogenous regulation of the MEPpathway in plastids, allowing for a new carbon sink inthe form of the introduced triterpenes, as comparedwith the MVA pathway operating in the cytosol pre-venting the high IPP/DMAPP flux into these com-pounds (Kempinski et al., 2015).
This approach has now been demonstrated to beapplicable for the metabolic engineering of varioustypes of terpene compounds, including monoterpenes,sesquiterpenes, and triterpenes in tobacco plants.However, we also note that the accumulation level ofeach type of terpene differed substantially between therespective terpene targets. Plants engineered for tri-terpene production accumulated 200 to 1,000 mg g21
fresh weight of triterpene, whereas sesquiterpene pro-duction has not exceeded 30 mg g21 fresh weight andmonoterpene accumulation ismaximally around 1mg g21
fresh weight (Wu et al., 2006; Kempinski et al., 2015).Such stark differences strongly suggest that the limita-tion in specific terpene class accumulation lies with
Figure 8. Phenotypes of transgenic lines expressingbotryococcene and squalene biosynthetic genes. Sometransgenic lines targeting botryococcene biosynthesisto the chloroplast (tpSSL1-3+tpFPS), and hence accu-mulating high levels of botryococcene, are dwarfed(A, left plant; right, wild-type plant) and exhibit chlo-rotic, mottled, and wrinkled leaf morphologies (B andC). High methylated botryococene accumulating lines(D, right plant) generated by engineering tpTMT-3engineered into a line (tpSSL1-3+tpFPS) accumulatingonly non-methylated botryococcene (D, left plant) didnot result in a wild type phenotype (E, right plant). Incontrast, plants targeting squalene biosynthesis to thechloroplast (tpSQS+tpFPS) and accumulating highlevels of squalene (E, left plant) did not exhibit suchphenotypes, nor when triterpene methyltransferases(TMT-1 or TMT-2) were also targeted to the chloroplastof squalene accumulating line (E, middle plant) com-pared to the wild type (E, right plant).
712 Plant Physiol. Vol. 170, 2016
Jiang et al.
www.plantphysiol.orgon January 5, 2020 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

the engineered terpene synthase or that certain intro-duced terpene compounds may have differing effectson physiological homeostasis and growth. Consistentwith this notion, we found that overexpression andchloroplast targeting of the soluble form of SSL1-3 withFPS yielded similar levels of botryococcene accumu-lation to that for squalene, which was achieved byplastid-targeted engineering of a yeast, soluble squa-lene synthase along with the avian FPS. However, twotimes more botryococcene was observed with SSL1-3than the SSL1-3M enzyme form with FPS targeted tothe chloroplast. This suggests that the chimeric enzymeSSL1-3 functions as well as the single yeast squalenesynthase enzyme targeted to the chloroplast and ex-hibits a higher catalytic capacity than SSL1-3M could.TMTs are functionally insoluble enzymes that exhibit
an unexpectedly high catalytic activity for the methyl-ation reaction when engineered into both the chloro-plast and cytoplasm compartments of the appropriatetransgenic plant lines. Up to 91% of the C30 triterpenesaccumulating in the high-yielding lines was subse-quently transformed to monomethylated or dimethy-lated triterpene when one of the three TMT genestargeted methyltransferase activity to the chloroplast.The methylation ratio of 51% to 91% by TMTs directedto the plastid compartment versus 3% to 14% by TMTstargeted to the cytosol provides additional evidencethat the distribution of triterpene C30 in the high-triterpene-accumulating transgenic lines remained inthe chloroplast. This was not unexpected, because theC30 triterpenes are supposedly synthesized in thechloroplast and methylation in the cytosol would re-quire some mechanism, either active or passive, to ex-port the novel triterpene out of the chloroplasts to thecytoplasm.Therefore, in order to account for the small but sig-
nificant methylation of triterpenes occurring in the cy-toplasm, at least four possible routes remain plausible.First, the methylated squalene produced by targetingTMT-1 and TMT-2 to the cytoplasm in the wild-typeplants proves that natively synthesized squalene can bemethylated by TMTs. Second, the small amount ofmethylated botryococcene generated in plants in whichTMT-3 was directed to the cytoplasm while highbotryococcene biosynthesis was directed to the chlo-roplasts [tpSSL1-3(M)+tpFPS] could arise from a lowlevel of botryococcene (C30) biosynthesized by mis-targeted SSL1-3(M) (Table II). This notion implies thatTMTs can methylate cytosolic triterpene (C30) pro-duced by mistargeted triterpene synthase as well as bythe native triterpene machinery. Third, expressing thetpTMT-1 in wild-type plants also resulted in methyl-ated products, which must be derived from cytosolicendogenous squalene catalyzed by mistargeted TMT-1.This evidence, not surprisingly, suggests that ourchloroplast-targeting strategy is not 100% effective andsupports our contention that mistargeted TMTs are alsoable to methylate cytosol-localized triterpenes. Fourth,the cytosolic engineered TMTs may have a way toaccess the plastid-localized squalene. The recent
discovery that plastid envelope-localized substrates canbe accessed by enzymes targeted to the ER membranethrough a continuity of ER and chloroplast (Mehrshahiet al., 2013) offers one possible explanation. Of course,the methylation status of triterpenes could come aboutby some combination of routes, which might also bevariable upon plant development and growth habit.
An issue raised during the initial phases of this workwas whether there would be sufficient SAM to supportthe formation of the methylated triterpenes. This con-cern arose because of an appreciation for how impor-tant SAM is to the methylation of macromolecules aswell as very diverse small molecules (Bouvier et al.,2006; Sauter et al., 2013) and its known biosynthesis inthe cytoplasm (Ravanel et al., 1998, 2004). Fortunately,concern for SAM availability seemed unfounded re-gardless of whether the methylation reactions weretargeted to the chloroplasts or the cytoplasm. Althoughthere was reduced TMT efficiency in the cytoplasm, thisis most likely due to the reduced amount of triterpeneavailable for methylation and, thus, reflects TMT effi-ciency and not SAM availability.
Equally interesting was the observation that plantsengineered for botryococcene accumulation tended toexhibit distinct phenotypic outcomes like dwarfism,chlorosis, andmottling, while plants accumulating highlevels of squalene did not show any of these adverseeffects. Why this might be so is currently unknown.However, if one could discern how the plants were ableto accumulate high levels of squalene without anynegative impact on growth performance, then onemight be able to use this information in the engineeringof advanced accumulation mechanisms for terpeneslike botryococcene. One suggestion worth examining ishow botryococcenes versus squalene might differentiallyinterdigitate into membranes and disrupt normal bio-chemical functions.Hence, engineering alternativemeansfor sequestering these molecules could alleviate physio-logical consequences and improve overall accumulation(e.g. engineering in lipid droplet-forming proteins).
Finally, squalene biosynthesis is known to be a keycommitted step in sterol biosynthesis, and squalene hasbeen suspected of serving a regulatory role (Wu et al.,2012). The results presented here, where introducingcytosolic forms of the TMT enzymes elevated overallsqualene and methylated squalene levels 4- to 7-foldhigher than normal, directly address this issue. Bydiverting squalene to its methylated forms, some innatemechanism had to be evoked to allow for additionalsqualene production to occur. This, we suggest, is pro-viding an important glimpse into the regulatory com-plexity of squalene biosynthesis, which is crucial for thehomoeostatic control of sterol biosynthesis in the plants.
MATERIALS AND METHODS
Expression Vector Construction and Plant Transformation
The design of gene constructs and assembly for engineering botryococcenebiosynthesis were based on the work described previously by Wu et al. (2006,
Plant Physiol. Vol. 170, 2016 713
Triterpene Metabolism in Plants
www.plantphysiol.orgon January 5, 2020 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

2012) using standard molecular methodologies. Gene constructs consisted ofa peptide fusion of SSL-1 (GenBank accession no. HQ585058.1) and SSL-3(GenBank accession no. HQ585060.1) connected by a triplet repeat peptidelinker of Gly-Gly-Ser-Gly, with or without appending the C-terminal end (71amino acids) of the Botryococcus braunii squalene synthase (GenBank accessionno. AF205791.1) onto the C terminus of SSL-3 and the FPS gene (P08836; Tarshiset al., 1994). The chimeric SSL1-3 genes and FPS gene were inserted down-stream of the strong constitutive promoters Pcv (Verdaguer et al., 1996) and Pca(Benfey et al., 1990), respectively. For trichome-specific expression of triterpenebiosynthesis, the trichome-specific promoters Pcbt (Ennajdaoui et al., 2010) andPcyp16 (Wang et al., 2002a) were fused to 59 end of botryococcene synthasegenes and the FPS gene, respectively. The duplicated cauliflower mosaic virus35S enhancer elements (Benfey et al., 1990) were fused to the 59 end of eachtrichome promoter. A chloroplast-targeting signal sequence encoding for thefirst 58 amino acids of the Arabidopsis (Arabidopsis thaliana) Rubisco smallsubunit gene (NM23202; Lee et al., 2006) was fused in frame with the 59 end ofthe respective terpene synthase genes. The gene cassettes were assembled to-gether in a helper vector described by Wu et al. (2012) by standard molecularbiology methods, and the various DNA segments were verified by DNA se-quencing. The gene cassettes were then introduced into pBDON, a modified Tiplasmid vector harboring a hygromycin resistance gene byDNA recombination(Wu et al., 2006).
The triterpene methyltransferase genes TMT-1 (JN828962.1), TMT-2(JN828963.1), and TMT-3 (JN828964.1) were inserted directly into the planttransformation vector pKYLx71 (Schardl et al., 1987) harboring a 35S viralpromoter and a kanamycin resistance gene. In order to target TMT genes to thechloroplast, the chloroplast-targeting signal sequence noted above was theninserted in frame with the 59 termini of the respective TMT genes.
The engineered Ti plasmid vectors were introduced into Agrobacteriumtumefaciens GV3850 by electroporation, and the resulting A. tumefaciens lineswere used to genetically engineer tobacco (Nicotiana tabacum) T1 accession 1068(Nielsen, 1982), or transgenic line tpSQS+tpFPS #5 (T2 homozygous generation)with a high level of squalene, as described previously by Wu et al. (2012), orthe high-botryococcene-accumulating transgenic lines (tpSSL1-3+tpFPS-10 ortpSSL1-3M+tpFPS-31; T1 heterozygous generation) generated in this study.Leaf explants were transformed with the respective gene constructs, and theresulting calli were selected on tissue culture medium with hygromycin (50 mgL21) for engineering botryococcene biosynthesis and with both hygromycin(50 mg L21) and kanamycin (250 mg L21) for engineering methylated triterpenebiosynthesis. The culture medium (1 L) contained 4.2 g ofMurashige and Skoogsalts (Phytotechnology Laboratories), 0.112 g of B5 vitamins (PhytotechnologyLaboratories), 30 g of sucrose, 9 g of agar, 1 mg of indole-3-acetic acid, and2.5 mg of benzylaminopurine (Sigma). The selected calli were grown understerile tissue culture conditions to regenerate plantlets. The selected T0 plantletswere then propagated in the greenhouse and assessed for triterpene content byGC-MS or GC-FID analysis.
Plant Propagation and Segregation Selection
All the T0 plantlets after hygromycin or kanamycin selection were grown incommoncommercialvermiculite/soilblends inagreenhouseandfertilizedweeklywith water-soluble fertilizer (20-20-20 for nitrogen, phosphorus, and potassium).Insect controlwasperformedasneeded.TheT0plantswereallowedtoflowerin thegreenhouse, and the T1 seed was collected for subsequent cycles of propagation.Segregation of the hygromycin and kanamycin resistance trait in the T1 seed lineswas also evaluated by germinating sterilized seeds on 50mgL21 hygromycin and250mgL21 kanamycin in T-tissue culture medium (4.2 g ofMurashige and Skoogsalts, 0.112 g of B5 vitamins, 30 g of Suc, and 9 g of agar in 1 L of medium).
Triterpene (Squalene, Botryococcene, MethylatedSqualene, and Methylated Botryococcene) Determinations
Fifty to 150 mg of transgenic leaf material was collected from the up-permost, fully expanded leaves of tobacco plants grown in a greenhouse. Theother plant tissues (roots, stems, and veins) were collected from plants grownin tissue culture for chemical analysis. The terpene content for each samplewas determined by the methods described previously by Wu et al. (2012).Each plant sample was ground in liquid nitrogen, then extracted with 2 to4 mL of a hexane:ethyl acetate (85:15, v/v) mixture containing 200 ng ofa-cedrene as an external standard for quantification and calculations of re-covery. The extracts were concentrated to 500 mL under a nitrogen streamwithout drying the sample. The concentrated extracts were then partially
purified by passing through a silica column (500 mg, prepared in a glasswool-plugged glass pipette) and further eluted with 1 mL of the hexanesolvent. After concentration of the combined eluate under a stream ofnitrogen, aliquots were injected onto a GC-MS device equipped with an HP5-MS capillary column (30 m 3 0.32 mm, 0.25-mm phase thickness) with atemperature program of 70°C for 1 min followed by a 4°C min21 gradient to250°C. Mass spectra were recorded at 70 eV, scanning from 35 to 500 atomicmass units, and experimental samples were compared with standards thatwere used previously in earlier studies (Niehaus et al., 2011, 2012; Wu et al.,2012) for verification.
The structure of purified botryococcene from tobacco was determined by1H-NMR and 13C-NMR spectral analyses, which were also described in anearlier study (Wu et al., 2012). Botryococcenewas extracted from leaf materialof a transgenic line (tpSSL1-3+tpFPS #10) targeting the chimeric botryo-coccene synthase SSL1-3 and FPS to the plastid compartment under the di-rection of the constitutive promoters.
One hundred grams of leaf material was ground in liquid nitrogen and thenextracted with 1.2 L of hexane:ethyl acetate (85:15, v/v); the extract was con-centrated to 5 mL and fractionated on a silica column with 5-mL aliquots ofhexane as the eluting solvent. Fractions were monitored by GC-MS for thedesired triterpene compound. Enriched fractions were pooled and concentratedunder nitrogen, and the entire sample was processed by silica HPLC usinghexane as the eluting solvent (Niehaus et al., 2012; Wu et al., 2012). Alterna-tively, the crude extract was resuspended in hexane and fractionated via silicagel chromatography, with a final purification step provided by HPLC. Recov-ery of a 6-mg purified botryococcene sample with a 50% yield was obtained.1H-NMR and 13C-NMR spectra were recorded on a 400-MHz Varian J-NMRspectrometer at 300 K, and chemical shifts were referenced relative to solventpeaks, namely dH 7.24 and dC 77 for CDCl3.
Supplemental Data
The following supplemental materials are available.
Supplemental Figure S1. Triterpene and methylated triterpene contentsdetermined in leaf extracts from elite transgenic plants by GC-MS.
Supplemental Figure S2. Mass spectra of C30 botryococcene produced intransgenic lines, C31 monomethylated botryococcene and C32 dimethy-lated botryococcene produced in lines expressing TMT-3 in a botryococceneaccumulating line, C31 monomethylated squalene and C32 dimethylatedsqualene produced in lines expressing TMT-1, TMT-2 or TMT-3 in a squa-lene accumulating line.
Supplemental Figure S3. (6E,10R,11E,13R,16E)-botryococcene (C30botryococcene) isolated from plants 1H-NMR (400 MHz).
Supplemental Figure S4. 13C-NMR (100 MHz) of botryococcenes isolatedfrom plants.
Supplemental Table S1. Screen of T0 transgenic lines targeting select TMTactivities to the chloroplast or the cytoplasm of high squalene (tpSQS+tpFPS)and botryococcene (tpSSL1-3+tpFPS) accumulating lines for their methylatedtriterpene content.
ACKNOWLEDGMENTS
We thank Scott Kinison for outstanding support for this project and all theChappell laboratory personnel for constructive comments throughout thecourse of this work.
Received October 1, 2015; accepted November 23, 2015; published November24, 2015.
LITERATURE CITED
Aharoni A, Giri AP, Deuerlein S, Griepink F, de Kogel WJ, VerstappenFW, Verhoeven HA, Jongsma MA, Schwab W, Bouwmeester HJ (2003)Terpenoid metabolism in wild-type and transgenic Arabidopsis plants.Plant Cell 15: 2866–2884
Ahlert D, Stegemann S, Kahlau S, Ruf S, Bock R (2009) Insensitivity ofchloroplast gene expression to DNA methylation. Mol Genet Genomics282: 17–24
714 Plant Physiol. Vol. 170, 2016
Jiang et al.
www.plantphysiol.orgon January 5, 2020 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

Ajikumar PK, Xiao WH, Tyo KEJ, Wang Y, Simeon F, Leonard E, MuchaO, Phon TH, Pfeifer B, Stephanopoulos G (2010) Isoprenoid pathwayoptimization for taxol precursor overproduction in Escherichia coli.Science 330: 70–74
Alban C, Tardif M, Mininno M, Brugière S, Gilgen A, Ma S, MazzoleniM, Gigarel O, Martin-Laffon J, Ferro M, et al (2014) Uncovering theprotein lysine and arginine methylation network in Arabidopsis chlo-roplasts. PLoS ONE 9: e95512
Andrianov V, Borisjuk N, Pogrebnyak N, Brinker A, Dixon J, Spitsin S,Flynn J, Matyszczuk P, Andryszak K, Laurelli M, et al (2010) Tobaccoas a production platform for biofuel: overexpression of ArabidopsisDGAT and LEC2 genes increases accumulation and shifts the compo-sition of lipids in green biomass. Plant Biotechnol J 8: 277–287
Anterola A, Shanle E, Perroud PF, Quatrano R (2009) Production of taxa-4(5),11(12)-diene by transgenic Physcomitrella patens. Transgenic Res 18:655–660
Benfey PN, Ren L, Chua NH (1990) Combinatorial and synergistic prop-erties of CaMV 35S enhancer subdomains. EMBO J 9: 1685–1696
Besumbes O, Sauret-Güeto S, Phillips MA, Imperial S, Rodríguez-Concepción M, Boronat A (2004) Metabolic engineering of isoprenoidbiosynthesis in Arabidopsis for the production of taxadiene, the firstcommitted precursor of taxol. Biotechnol Bioeng 88: 168–175
Bouvier F, Linka N, Isner JC, Mutterer J, Weber APM, Camara B (2006)Arabidopsis SAMT1 defines a plastid transporter regulating plastid bio-genesis and plant development. Plant Cell 18: 3088–3105
Bouvier F, Rahier A, Camara B (2005) Biogenesis, molecular regulation andfunction of plant isoprenoids. Prog Lipid Res 44: 357–429
Buckingham J (2003) Dictionary of Natural Products (CD ROM). Chapman& Hall, London
Curien G, Job D, Douce R, Dumas R (1998) Allosteric activation of Ara-bidopsis threonine synthase by S-adenosylmethionine. Biochemistry 37:13212–13221
Davidovich-Rikanati R, Lewinsohn E, Bar E, Iijima Y, Pichersky E, SitritY (2008) Overexpression of the lemon basil alpha-zingiberene synthasegene increases both mono- and sesquiterpene contents in tomato fruit.Plant J 56: 228–238
DellaPenna D (2005) A decade of progress in understanding vitamin Esynthesis in plants. J Plant Physiol 162: 729–737
Dewick PM (2009) Medicinal Natural Products: A Biosynthetic Approach,Ed 3. John Wiley & Sons, Chichester, UK.
Ennajdaoui H, Vachon G, Giacalone C, Besse I, Sallaud C, Herzog M,Tissier A (2010) Trichome specific expression of the tobacco (Nicotianasylvestris) cembratrien-ol synthase genes is controlled by both activat-ing and repressing cis-regions. Plant Mol Biol 73: 673–685
Fontecave M, Atta M, Mulliez E (2004) S-Adenosylmethionine: nothinggoes to waste. Trends Biochem Sci 29: 243–249
Glikson M, Lindsay K, Saxby J (1989) Botryococcus: a planktonic greenalga, the source of petroleum through the ages. Transmission electronmicroscopical studies of oil shales and petroleum source rocks. OrgGeochem 14: 595–608
Harvey BG, Wright ME, Quintana RL (2010) High-density renewablefuels based on the selective dimerization of pinenes. Energy Fuels 24:267–273
Hillen LW, Pollard G, Wake LV, White N (1982) Hydrocracking of theoils of Botryococcus braunii to transport fuels. Biotechnol Bioeng 24:193–205
Houtz RL, Stults JT, Mulligan RM, Tolbert NE (1989) Post-translationalmodifications in the large subunit of ribulose bisphosphate carboxylase/oxygenase. Proc Natl Acad Sci USA 86: 1855–1859
Huang LC, Hsiao LJ, Pu SY, Kuo CI, Huang BL, Tseng TC, Huang HJ,Chen YT (2012) DNA methylation and genome rearrangement charac-teristics of phase change in cultured shoots of Sequoia sempervirens.Physiol Plant 145: 360–368
Huang Z, Poulter CD (1989) Tetramethylsqualene, a triterpene fromBotryococcus braunii var. showa. Phytochemistry 28: 1467–1470
Inagaki YS, Etherington G, Geisler K, Field B, Dokarry M, Ikeda K,Mutsukado Y, Dicks J, Osbourn A (2011) Investigation of the potentialfor triterpene synthesis in rice through genome mining and metabolicengineering. New Phytol 191: 432–448
Jander G, Joshi V (2009) Aspartate-derived amino acid biosynthesis inArabidopsis thaliana. The Arabidopsis Book 7: e0121 doi: 10.1199/tab.0123.
Kappers IF, Aharoni A, van Herpen TWJM, Luckerhoff LLP, Dicke M,Bouwmeester HJ (2005) Genetic engineering of terpenoid metabolismattracts bodyguards to Arabidopsis. Science 309: 2070–2072
Kempinski C, Jiang Z, Bell S, Chappell J (2015) Metabolic engineering ofhigher plants and algae for isoprenoid production. Adv Biochem EngBiotechnol 148: 161–199
Kusano T, Berberich T, Tateda C, Takahashi Y (2008) Polyamines: es-sential factors for growth and survival. Planta 228: 367–381
Lange BM, Ahkami A (2013) Metabolic engineering of plant monoterpenes,sesquiterpenes and diterpenes: current status and future opportunities.Plant Biotechnol J 11: 169–196
Lange BM, Mahmoud SS, Wildung MR, Turner GW, Davis EM, Lange I,Baker RC, Boydston RA, Croteau RB (2011) Improving peppermintessential oil yield and composition by metabolic engineering. Proc NatlAcad Sci USA 108: 16944–16949
Lee DW, Lee S, Lee GJ, Lee KH, Kim S, Cheong GW, Hwang I (2006)Functional characterization of sequence motifs in the transit peptide ofArabidopsis small subunit of Rubisco. Plant Physiol 140: 466–483
Liscombe DK, Louie GV, Noel JP (2012) Architectures, mechanisms andmolecular evolution of natural product methyltransferases. Nat ProdRep 29: 1238–1250
Lücker J, Schwab W, Franssen MCR, Van Der Plas LHW, BouwmeesterHJ, Verhoeven HA (2004) Metabolic engineering of monoterpene bio-synthesis: two-step production of (+)-trans-isopiperitenol by tobacco.Plant J 39: 135–145
Martin VJJ, Pitera DJ, Withers ST, Newman JD, Keasling JD (2003) En-gineering a mevalonate pathway in Escherichia coli for production ofterpenoids. Nat Biotechnol 21: 796–802
Mastalerz M, Hower JC (1996) Elemental composition and molecularstructure of Botryococcus alginite in Westphalian cannel coals fromKentucky. Org Geochem 24: 301–308
Mehrshahi P, Stefano G, Andaloro JM, Brandizzi F, Froehlich JE,DellaPenna D (2013) Transorganellar complementation redefines thebiochemical continuity of endoplasmic reticulum and chloroplasts.Proc Natl Acad Sci USA 110: 12126–12131
Metzger P (1985) Structures of some botryococcenes: branched hydrocar-bons from the b-race of the green alga Botryococcus braunii. Phyto-chemistry 23: 2995–3002
Metzger P, Casadevall E, Coute A (1988) Botryococcene distribution instrains of the green alga Botryococcus braunii. Phytochemistry 27: 1383–1388
Moldowan JM, Seifert WK (1980) First discovery of botryococcane in pe-troleum. J Chem Soc Chem Commun 912
Niehaus TD, Kinison S, Okada S, Yeo YS, Bell SA, Cui P, DevarenneTP, Chappell J (2012) Functional identification of triterpene methyl-transferases from Botryococcus braunii race B. J Biol Chem 287: 8163–8173
Niehaus TD, Okada S, Devarenne TP, Watt DS, Sviripa V, Chappell J(2011) Identification of unique mechanisms for triterpene biosynthesis inBotryococcus braunii. Proc Natl Acad Sci USA 108: 12260–12265
Nielsen J, Keasling JD (2011) Synergies between synthetic biology andmetabolic engineering. Nat Biotechnol 29: 693–695
Nielsen MTJG (1982) Inheritance pattern for secreting and nonsecretingglandular trichomes in tobacco. Crop Sci 22: 1051–1053
Niemi KJ, Adler J, Selman BR (1990) Protein methylation in pea chloro-plasts. Plant Physiol 93: 1235–1240
Nishiyama R, Ito M, Yamaguchi Y, Koizumi N, Sano H (2002) Achloroplast-resident DNA methyltransferase is responsible for hyper-methylation of chloroplast genes in Chlamydomonas maternal gametes.Proc Natl Acad Sci USA 99: 5925–5930
Ohara K, Matsunaga E, Nanto K, Yamamoto K, Sasaki K, Ebinuma H,Yazaki K (2010) Monoterpene engineering in a woody plant Eucalyptuscamaldulensis using a limonene synthase cDNA. Plant Biotechnol J 8:28–37
Okada S, Murakami M, Yamaguchi K (1995) Hydrocarbon composition ofnewly isolated strains of the green microalga Botryococcus braunii. JAppl Phycol 7: 555–559
Peralta-Yahya PP, Keasling JD (2010) Advanced biofuel production inmicrobes. Biotechnol J 5: 147–162
Peralta-Yahya PP, Ouellet M, Chan R, Mukhopadhyay A, Keasling JD,Lee TS (2011) Identification and microbial production of a terpene-based advanced biofuel. Nat Commun 2: 483
Plant Physiol. Vol. 170, 2016 715
Triterpene Metabolism in Plants
www.plantphysiol.orgon January 5, 2020 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.

Ravanel S, Block MA, Rippert P, Jabrin S, Curien G, Rébeillé F, Douce R(2004) Methionine metabolism in plants: chloroplasts are autonomousfor de novo methionine synthesis and can import S-adenosylmethioninefrom the cytosol. J Biol Chem 279: 22548–22557
Ravanel S, Gakière B, Job D, Douce R (1998) The specific features ofmethionine biosynthesis and metabolism in plants. Proc Natl Acad SciUSA 95: 7805–7812
Renninger NS, McPhee DJ April 10, 2008. Fuel compositions comprisingfarnesane and farnesane derivatives and method of making and usingsame. US Patent Application No. 20080083158A1
Ro DK, Paradise EM, Ouellet M, Fisher KJ, Newman KL, Ndungu JM, HoKA, Eachus RA, Ham TS, Kirby J, et al (2006) Production of the anti-malarial drug precursor artemisinic acid in engineered yeast. Nature440: 940–943
Rude MA, Schirmer A (2009) New microbial fuels: a biotech perspective.Curr Opin Microbiol 12: 274–281
Sauter M, Moffatt B, Saechao MC, Hell R, Wirtz M (2013) Methioninesalvage and S-adenosylmethionine: essential links between sulfur, eth-ylene and polyamine biosynthesis. Biochem J 451: 145–154
Schardl CL, Byrd AD, Benzion G, Altschuler MA, Hildebrand DF, HuntAG (1987) Design and construction of a versatile system for the ex-pression of foreign genes in plants. Gene 61: 1–11
Scheer M, Grote A, Chang A, Schomburg I, Munaretto C, Rother M,Söhngen C, Stelzer M, Thiele J, Schomburg D (2011) BRENDA, theenzyme information system in 2011. Nucleic Acids Res 39: D670–D676
Schillberg S, Fischer R, Emans N (2003) Molecular farming of recombinantantibodies in plants. Cell Mol Life Sci 60: 433–445
Schwab W, Davidovich-Rikanati R, Lewinsohn E (2008) Biosynthesis ofplant-derived flavor compounds. Plant J 54: 712–732
Shelar DB, Shirote PJ (2010) Natural product in drug discovery: Back tofuture. Journal of Pharmacy Research 3: 2007
Takahashi M, Terada Y, Nakai I, Nakanishi H, Yoshimura E, Mori S(2003) Role of nicotianamine in the intracellular delivery of metals andplant reproductive development. Plant Cell 15: 1263–1280
Takahashi S, Yeo Y, Greenhagen BT, McMullin T, Song L, Maurina-Brunker J, Rosson R, Noel JP, Chappell J (2007) Metabolic engineeringof sesquiterpene metabolism in yeast. Biotechnol Bioeng 97: 170–181
Tarshis LC, Yan M, Poulter CD, Sacchettini JC (1994) Crystal structure ofrecombinant farnesyl diphosphate synthase at 2.6-A resolution. Bio-chemistry 33: 10871–10877
Trievel RC, Flynn EM, Houtz RL, Hurley JH (2003) Mechanism of multiplelysine methylation by the SET domain enzyme Rubisco LSMT. NatStruct Biol 10: 545–552
Ubersax JA, Platt DM December 9, 2010. Genetically modified microbesproducing isoprenoids. US Patent Application No. 20100311065 A1
Verdaguer B, de Kochko A, Beachy RN, Fauquet C (1996) Isolation andexpression in transgenic tobacco and rice plants, of the cassava veinmosaic virus (CVMV) promoter. Plant Mol Biol 31: 1129–1139
Wagner GJ, Wang E, Shepherd RW (2004) New approaches for studyingand exploiting an old protuberance, the plant trichome. Ann Bot (Lond)93: 3–11
Wallsgrove RM, Lea PJ, Miflin BJ (1983) Intracellular localization of as-partate kinase and the enzymes of threonine and methionine biosyn-thesis in green leaves. Plant Physiol 71: 780–784
Wang E, Gan S, Wagner GJ (2002a) Isolation and characterization of theCYP71D16 trichome-specific promoter from Nicotiana tabacum L. J ExpBot 53: 1891–1897
Wang KL, Li H, Ecker JR (2002b) Ethylene biosynthesis and signalingnetworks. Plant Cell (Suppl) 14: S131–S151
Westfall PJ, Pitera DJ, Lenihan JR, Eng D, Woolard FX, Regentin R,Horning T, Tsuruta H, Melis DJ, Owens A, et al (2012) Production ofamorphadiene in yeast, and its conversion to dihydroartemisinic acid,precursor to the antimalarial agent artemisinin. Proc Natl Acad Sci USA109: E111–E118
Withers ST, Gottlieb SS, Lieu B, Newman JD, Keasling JD (2007) Iden-tification of isopentenol biosynthetic genes from Bacillus subtilis by ascreening method based on isoprenoid precursor toxicity. Appl EnvironMicrobiol 73: 6277–6283
Wright LP, Rohwer JM, Ghirardo A, Hammerbacher A, Ortiz-Alcaide M,Raguschke B, Schnitzler JP, Gershenzon J, Phillips MA (2014) Deox-yxylulose 5-phosphate synthase controls flux through the methylery-thritol 4-phosphate pathway in Arabidopsis. Plant Physiol 165: 1488–1504
Wu S, Jiang Z, Kempinski C, Nybo SE, Husodo S, Williams R, Chappell J(2012) Engineering triterpene metabolism in tobacco. Planta 236: 867–877
Wu S, Schalk M, Clark A, Miles RB, Coates R, Chappell J (2006) Redi-rection of cytosolic or plastidic isoprenoid precursors elevates terpeneproduction in plants. Nat Biotechnol 24: 1441–1447
Ying Z, Mulligan RM, Janney N, Houtz RL (1999) Rubisco small and largesubunit N-methyltransferases: bi- and mono-functional methyltransfer-ases that methylate the small and large subunits of Rubisco. J Biol Chem274: 36750–36756
Zhuang X, Chappell J (2015) Building terpene production platforms inyeast. Biotechnol Bioeng 112: 1854–1864
716 Plant Physiol. Vol. 170, 2016
Jiang et al.
www.plantphysiol.orgon January 5, 2020 - Published by Downloaded from Copyright © 2016 American Society of Plant Biologists. All rights reserved.