enhancementof neutrophil function result of prior...
TRANSCRIPT

Enhancement of Neutrophil Function
as a Result of Prior Exposure to Chemotactic Factor
DENNIS E. VANEPPS and MARYLYNNGARCIA, Departments of Medicine andMicrobiology, University of New Mexico, School of Medicine, Albuquerque, NewMexico 87131
A B S T R AC T Exposure of human polymorphonuclearleukocytes (PMN) to chemotactic factor, as well as themigration of PMNthrough a 5-gm pore-size membrane,results in a PMNpopulation with enhanced chemi-luminescence, enhanced capacity for superoxideanion production, and increased Escherichia coli bac-tericidal activity. The enhanced PMN response re-sulting from exposure to chemotactic factor was ob-served with several chemotactic stimuli, including amixture of casein and autologous serum, chemotacticC5 fragment, and formyl-L-methionyl-L-leucine-L-phenylalanine (f-Met-Leu-Phe). Enhanced levels ofchemiluminescence were observed with both solublestimuli (concanavalin A and phorbol myristate ace-tate) as well as particulate stimuli (opsonized zymosan).
Once activated by chemotactic factor, PMNretainedtheir enhanced stimulated chemiluminescence in theabsence of chemotactic factor for at least 2.5 h. En-hanced activity could not be correlated with a shift inthe number of immunoglobulin (Ig)G Fc receptorpositive or complement receptor positive PMN. Invivo studies with guinea pigs indicated that PMNattracted to an intraperitoneal injection of casein, likethose attracted through a chemotaxis memnbrane in vitroin response to casein, showed markedly enhancedstimulated chemiluminescence when compared withperipheral blood PMNfrom the same animal. Such amechanism to stimulate PMNfunction may enhancethe effectiveness of PMNin host defense at inflam-matory foci.
INTRODUCTIONChemotaxis is believed to be an important mechanismby which polymorphonuclear leukocytes (PMN)' as
D. E. Van Epps was the recipient of an Arthritis Fouinda-tion Senior Investigatorship Award.
Received for publication 5 July 1979 and in revised form26 March 1980.
1 Abbreviations used in this paper: Con A, concanavalin A;FCS, fetal calf serum; HBSS, Hanks' balanced salt solutioin;PMA, phorbol myristate acetate; PMN, polymorphonuclearleukocytes.
well as other cells are recruited to inflammatory sites.The attraction of these cells has been attributed tothe activation of a variety of chemotactic factors in vivo,including: kallikrein; chemotactic C5 fragment fromthe complement system; chemotactic factors derivedfrom cells involved in host defense such as lympho-cytes, monocytes, and neutrophils; and factors liber-ated from infecting microorganisms (1). It has alsobeen shown that chemotactic factors can stimulateneutrophil chemiluminescence (2), cause cell aggrega-tion (3), increase intracellular levels of cyclic guanosine5'-monophosphate (cGMP) (4); increase microtubuleand microfilament assembly (5, 6), stimulate degranu-lation (7), increase adherence (6, 8), and stimulatesuperoxide anion production (6, 9). A recent study byGallin et al. (10) has shown that under appropriateconditions, factors such as phorbol myristate acetate,concanavalin A (Con A), and the calcium ionophoreA23187, which stimulate neutrophil secretion of enzymes,increase adherence and alter the binding of thechemotactic peptide, f-Met-Leu-Phe (10). All of thesecited studies indicate that the neutrophils exposedto chemotactic stimuli or agents capable of stimulatingdegranulation undergo metabolic, cytoskeletal, andsurface receptor alterations. The present study demon-strates that neutrophils that either have been exposedto a chemotactic stimulus or have migrated in responseto this stimulus are functionally more active in systemsmeasuring stimulated chemiluminescence, superoxideanion generation, and bactericidal activity than thosethat have not been exposed to chemotactic factor.
METHODSCell preparation. Human PMIN were prepared from
heparinized peripheral blood by Ficoll-Hypaque centrifuga-tion (11). The cell pellet containing PMN and erythro-cytes was resuspended in Hanks' balanced salt solution(HBSS) and mixed with Plasmagel (HTI, Buffalo, N. Y.) at aconcentration of 1 ml of Plasmagel per 5 cm3 of blood. Theleukocyte-rich suipernate obtained after 30 min of 37°C in-cubation was then centrifuged and the cell pellet mixedwith 40C distilled water for 30 s to lyse residual erythro-cytes. Cells were then washed twice and resuispend(led in
167J. Clin. Invest. © The American Society for Clinical Investigation, Inc. 0021-9738/80/08/0167/09 $1.00
Volume 66 August 1980 167-175

HBSS supplemented with 5% fetal calf serum (FCS) to aconcentration of 2 x 107 PMN/ml. This preparation contained> 90% neutrophils. The cell suspension was then dividedinto three parts and each was treated in one of the followingways: (a) one part was incubated at 37°C for 3 h in HBSSsupplemented with 5% FCS; (b) one part was incubated at37°C for 3 h in 5%FCSplus chemotactic factor; (c) the remain-ing cells in media supplemented with 5% FCS were placedin the upper compartment of an enlarged Boyden type chemo-tactic chamber (12) (1 ml/chamber) and allowed to migratefor 3 h at 37°C toward one of several chemotactic factors. Inthe third case, cells were obtained after they had respondedto chemotactic factor and appeared in the lower compartmentof the chemotactic chamber. In each case, cells were washedtwice to remove media and chemotactic factor and testedin the appropriate assay. The three cell preparations repre-sent (a) control cells, (b) cells exposed to chemotactic factor,and (c) cells that had actually migrated through a membranein response to the same concentration of chemotactic factor.
Guinea pig peripheral blood and peritoneal exudate PMNwere obtained from male, outbred, Hartley guinea pigs 4 hafter an intraperitoneal injection of 10 ml of 50% casein-saturated Eagle's Minimum Essential Media (Casein suppliedby Fischer Scientific Co., Pittsburgh, Pa.). Casein prepara-tions were centrifuged at 20,000 g for 20 min and filteredthrough a 0.22-gm pore-size filter to remove any particulatebefore injection. Peritoneal exudate cells were obtained bylavage using 100 ml of HBSSwith 5 U of heparin/ml to washthe peritoneal cavity. The fluids were centrifuged and thecell pellet washed and resuspended to 1 x 107 cells/ml. Dif-ferential counts on these cells indicated that they contained>90% neutrophils. Peripheral blood was obtained by cardiacpuncture from the same animal after peritoneal lavage. Bloodwas heparinized and PMNwere prepared by Ficoll-Hypa(luecentrifugation followed by Plasmagel sedimentation oferythrocytes. Residual erythrocytes were lysed by a 30-s treat-ment with 4°C distilled water, and cells were resuspended to1 x 107 cells/ml. Differential counts showed these prepara-tions to contain >90% PMN. In each case, peritoneal exu-date cells were treated for 30 s with distilled water forcomparison with peripheral blood PMN where residualerythrocytes were similarly lysed.
Chemotactic factors. Three types of chemotactic factorswere used in this study. The first was a mixture containing50% casein-saturated minimum essential media and 10%fresh autologous normal serum. This preparation proved tobe a potent chemotactic mixture and was the primary chemo-tactic factor used to attract large numbers of cells through thechemotaxis membrane. This mixture was found to be bothchemotactic and chemokinetic as determined by titration ex-periments usinlg the leading front assay for chemotaxis (13)and by determining that maximal migration in the presence ofa positive chemotactic gradient far exceeded the maximalchemokinetic response observed with no gradient present.The second factor used was a partially purified preparationof chemotactic C5 fragment obtained from fresh serum after itsactivation with zymosan (20 mng/ml zymosani for 60 mmin at37°C). The chemotactic C5 fragment was obtained by pas-sage of activated human serum over a Sephadex G-75 column.The low molecular weight fractions with chemotacticactivity were concentrated by ultrafiltration, using a mem-brane permeable to molecules of <2,000 mol wt. Thispreparation was chemotactic for human PMNand its activitywas 95% inhibitable with anti-human C5. C5a was titratedfor optimal chemotaxis in a quantitative chemotaxis assay(11) and used at this concentration. Finally, in some studiesa preparation of the synthetic chemotactic peptide f-Met-Leu-Phe (kindly provided by Dr. Richard Freer of the Medical
College of Virginia) was used at a concentration of 10 nM,which proved to be optimal for migration. In all cases, pre-exposure of PMN for 1 h to the concentrations of eachchemotactic factor used in the study, followed by removal ofthe factor, deactivated these cells to the same factor later, asindicated by >50% inhibition in a quantitative chemotaxisassay (11).
Chemiluminescence assay. PMNchemiluminescence wasassayed as previously described in phosphate-buffered saline,pH 7.4, using zymosan that had been opsonized with freshnormal human serum as a stimulant (14). When guinea pigPMNwere used, zymosan was opsonized in the same mannerwith fresh guinea pig serum. PMN(5 x 106) were added in thedark to 2 mg of zymosan in a final volume of 1 ml. In someinstances soluble factors were used to stimulate chemi-luminescence, including 100 ,ug/ml Con A and 1.0 ,ug/mlphorbol myristate acetate (PMA) (Sigma Chemical Co., St.Louis, Mo.). When soluble stimuli were used, 10 nM luminol(Sigma Chemical Co.) was added to the mixture (15). 5 mg/mlPMAwas prepared in dimethyl sulfoxide and diluted to theappropriate concentration in media just prior to testing. As-says were performed in 18-h dark-adapted Beckman poly Qvials (Beckman Instruments, Inc., Fullerton, Calif.), andanalyzed in a nonrefrigeratedl Nuclear-Chicago B-scintillationcounter (Nuclear-Chicago Corp., Des Plaines, Ill.) used out ofcoincidence with a window setting of 0, 10. Chemilumines-cence, unless otherwise stated, was measured using 1-mincounts at 6-min intervals for a minimum of 49 min. In eachcase, data are presented either as the chemiluminescencecurve obtained or as the integrated area under the curve.
Superoxide anion assay. Superoxide anion generation wasmeasured by the reduction of cytochrome c (16). Cells to betested were washed and resuspended in HBSSwithouit phenolred to 1 x 107 PMN/ml. In some cases PMNwere stimulatedwith normal serum-opsonized zymosan (2 mg) by mixingzymosan with 5 x 106 PMN, centrifuging for 45 s at 1,000 g,and resuspending in 1.5 ml of an 80-,uM cytochrome c solu-tion prepared in HBSSwithout phenol red. Samples were thenincubated for 5 min at 37°C. Unstimulated samples consistedof 5 x 106 PMNincubated in 1.5 ml of 80 ,tM cytochrome c,for 30 min at 37°C. The longer incubation period was essentialfor unstimulated cells to achieve a measurable release ofsuperoxide anion. Superoxide anion release was assessed bydetermining the reduction of cytochrome c using spectro-photometric analysis at an optical density of 550 nM. Theamount of cytochrome c reduced in each case was deter-mined using a molar extinction coefficient of 2.1 x 104 andexpressing the result as nanomoles of cytochrome c redIucedper 106 PMN.
Bactericidal assay. Bactericidal activity was assayed aspreviously described (17), using Escherichia coli K 12 as atarget microorganismii. Mixtures containing -2.5 x 108 bac-teria, 0.3 ml of opsonin (fresh normal serum diluted 1:3), and5 x 106 PMNwere adjusted to 1 ml with HBSSand used toassay PMN bactericidal activity. Mixtures were sampledafter 0-, 30-, 60-, and 90-min incubations at 37°C by remov-ing 0.001 ml of the mixture with a calibrated loop, diltutingthis in 1 ml distilled water and subsequently mixing 0.1 mlof this suspension with Bacto antibiotic medium 2 agar (DifcoLaboratories, Detroit, Mich.) in a pour plate. Pour plateswere inctubated overnight and the number of viable bac-teria determined by colony counts.
Determination of IgG and complement receptor positivePMN. PMNbearing IgG receptors were quantitated by boththe rosette technique (17) and fluorescence microscopy, usingthe method of Winfield et al. (18). PMNbearing complementreceptors were quantitated using the rosette techniquie.Rosette assays for IgG receptors were performed with bovine
168 D. E. Van Epps and M. L. Garcia
erythrocytes coated with rabbit IgG antibovine erythrocyteantibodies (17). Complement receptor-bearing cells were de-termined using sheep erythrocytes coated with rabbit IgManti-sheep erythrocyte antibody and human complementthat had been absorbed with sheep erythrocytes (17).PMN(2 x 106 in 0.2 ml) were mixed with 0.2 ml of a 1%solu-tion of each of the erythrocyte preparations, centrifuged at800 g for 4 min, and incubated at room temperature for 15 min.Cells were gently resuspended and the number of rosettes(three or more erythrocytes bound to a PMN) were quanti-tated in a hemocytometer and expressed as a percent of thetotal PMNpresent. Fewer than 3% rosettes were observedwhen uncoated bovine erythrocytes and uncoated or IgM-coated sheep erythrocytes were used.
IgG receptor-bearing PMNwere also quantitated using amodification of the method of Winfield et al. (18). Prepara-tions of rabbit b4 allotype IgG and rabbit anti-b4 IgG wereprepared by DEAEion exchange chromatography (18). PMN(2 x 106) were incuibated with 0.08 mg of rabbit b4 IgG at4°C for 30 min. Cells were then washed twice and incubatedfor 30 min at 4°C with 40 ,ul of anti-b4 IgG which had beentitrated to give optimal binding. Cells were again washedtwice and subsequently incubated with fluorescein isothio-cyanate-conjugated goat anti-rabbit IgG for 30 min at 4°C.
After an additional two washes, cells were observed byfluorescence microscopy and the number of positive cellsquantitated. Fluorescence was inhibited by aggregatedhuman IgG and no reduction in fluorescence was observedfollowing absorption of the rabbit b4 or anti-b4 with humanerythrocytes.
RESULTS
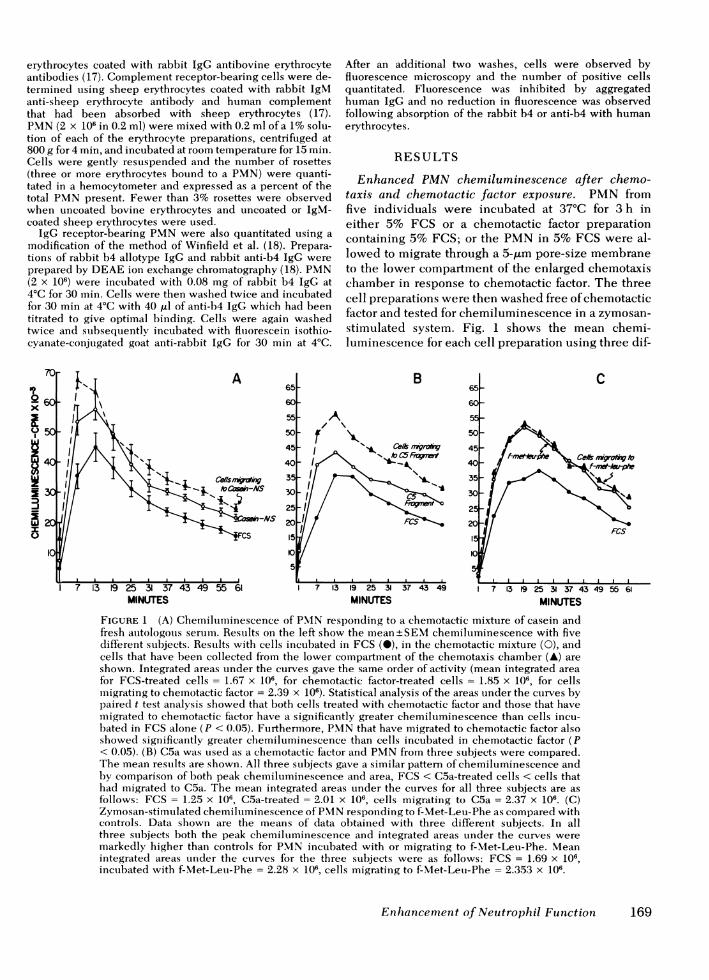
Enhanced PMNchemiluminescence after chemo-taxis and chemotactic factor exposure. PMN fromfive individuals were incubated at 37°C for 3 h ineither 5% FCS or a chemotactic factor preparationcontaining 5% FCS; or the PMNin 5% FCS were al-lowed to migrate through a 5-,tm pore-size membraneto the lower compartment of the enlarged chemotaxischamber in response to chemotactic factor. The threecell preparations were then washed free of chemotacticfactor and tested for chemiluminescence in a zymosan-stimulated system. Fig. 1 shows the mean chemi-luminescence for each cell preparation using three dif-
MINUTES MINUTES3 9 25 31 37 43 49 55 61
MINUTES
FIGURE 1 (A) Chemilumiiinescence of PMNresponding to a chemotactic mixture of casein andfresh autologous serum. Results on the left show the mean+SEMchemiluminescence with fivedifferent subjects. Results with cells incubated in FCS (M), in the chemotactic mixture (0), andcells that have been collected from the lower compartment of the chemotaxis chamber (A) areshown. Integrated areas under the curves gave the same order of activity (mean integrated areafor FCS-treated cells = 1.67 x 106, for chemotactic factor-treated cells = 1.85 x 106, for cellsmigrating to chemotactic factor = 2.39 x 106). Statistical analysis of the areas under the cturves bypaired t test analysis showed that both cells treated with chemotactic factor and those that havemigrated to chemotactic factor have a significantly greater chemiluminescence than cells incu-bated in FCS alone (P < 0.05). Furthermore, PMNthat have migrated to chemotactic factor alsoshowed significantly greater chemiluminescence than cells incubated in chemotactic factor (P< 0.05). (B) C5a was used as a chemotactic factor and PMNfrom three subjects were compared.The mean results are shown. All three subjects gave a similar pattern of chemiluminescence andby comparison of both peak chemiluminescence and area, FCS < C5a-treated cells < cells thathad migrated to C5a. The mean integrated areas under the curves for all three subjects are asfollows: FCS = 1.25 x 106, C5a-treated = 2.01 x 106, cells migrating to C5a = 2.37 x 106. (C)Zymosan-stimulated chemiluminescence of PMN responding to f-Met-Leu-Phe as compared withcontrols. Data shown are the means of data obtained with three different subjects. In allthree subjects both the peak chemiluminescence and integrated areas under the curves weremarkedly higher than controls for PMNincubated with or migrating to f-Met-Leu-Phe. Meanintegrated areas under the curves for the three subjects were as follows: FCS = 1.69 x 106,incubated with f-Met-Leu-Phe = 2.28 x 106, cells migrating to f-Met-Leu-Phe = 2.353 x 106.
Enhancement of Neutrophil Function 169

ferent chemotactic stimuli, including a casein-auto-logous serum mixture, C5a, and f-Met-Leu-Phe. Asshown, cells treated with any of the chemotactic fac-tors showed markedly enhanced chemiluminescencewhen compared with the control cells incubated withFCS. Furthermore, the preparation of PMNthat hadactually migrated through the 5-gm pore-size mem-
brane in response to chemotactic factor showed in mostcases even greater chemiluminescence than cellssimply incubated in chemotactic factor. This differencewas consistent in all cell preparations and was apparentin both the peak chemiluminescence as well as theintegrated areas under the chemiluminescence curves.
The mean integrated areas for each set of experimentsare indicated in the legend for Fig. 1. In the absenceof zymosan, no difference in resting (unstimulated)chemiluminescence was observed. Furthermore, intests on six different cell preparations, no significantdifference in the stimulated chemiluminescence was
observed between PMNtested immediately after cellisolation (51,865+5,731) and those that had been incu-bated in FCS for 3 h at 37'C (50,289+6,081).
Similar chemiluminescence studies were performedsubstituting soluble Con A and PMAfor the insolublezymosan stimulant in a system containing 10 nMluminol. As shown in Table I, a simnilar enhancedchemiluminescence was observed in PMNpreincu-bated in chemotactic factor (casein-autologous serum
mixture) and subsequently washed. Likewise, an even
greater chemiluminescence was observed in PMNallowed to migrate to chemotactic factor. Data inTable I were obtained using 100 ,ug/ml Con A and 1 ,ug/mlPMA. In all cases, the chemiluminescence of PMNmigrating to chemotactic factor was greater than PMNincubated in chemotactic factor, which was greater thanPMN incubated in media alone. This order was
independent of the concentration of Con A or PMN,because this same order was maintained at 10-or 100-fold lower concentrations, although the peakresponse at lower concentrations was proportionallydecreased.
Duration of enhanced chemiluminescence.Whether the enhanced chemiluminescence of PMNresulting from exposure to chemotactic factor repre-
TABLE IChemiluminescence Activity of Control PMN, Chemotactic Factor-treated PMN, and
PMNPrepared by Chemotaxis after Stimulation with Con A and PMA
Peak chemiluminescence response with each cell preparation
Incubated withchemotactic Prepared by
Experiments Control factor* chemotaxis
cpm
Stimulated with Con At1 419,994 431,382 627,8012 198,414 348,403 768,0813 49,212 76,096 189,4284 70,588 102,499 131,1515 58,954 63,820 72,5846 246,717 764,440 768,9957 54,811 59,027 65,5668 51,406 105,862 249,575
Mean+±1 SD5 143,762+ 135,129 243,941 +253,784 359,147+309,025Stimulated with PMAt
1 75,257 118,228 156,1072 223,898 306,409 372,3023 98,213 277,268 562,0604 135,055 247,104 352,6095 223,693 457,570 762,5806 89,254 199,926 219,6537 463,145 768,045 781,1368 289,955 696,917 749,417
Mean±tl SD§ 199,809+131,406 383,933+236,489 494,483+253,177
* Chemotactic factor used was a mixture of casein and autologous serum.t Con A and PMAwere used at 100 and 1.0 ,ug/ml concentrations, respectively.§ The difference between control cells and both chemotactic factor-treated cells and those pre-pared by chemotaxis was significant, P < 0.05, with both PMAand Con A. Statistical analysiswas performed by the nonparametric equivalent to two-way analysis of variance followed by thenonparametric equivalents of Newman-Keuls test for multiple comparisons.
170 D. E. Van Epps and M. L. Garcia

sents a permanent elevation in PMNresponsivenessor a transient condition was investigated. A portion ofPMNthat had migrated to the casein-autologouis serumilinixture or chemotactic C5 fragment and PMNthat hadbeen incubated in 5% FCS for 3 h were washed,counted, resuspended in phosphate-buffered saline,and tested in the chemilumiiinescence assay. Remainingcells were washed and resuspended in 5% FCS for2.5 h at 37°C. Subse(luently, cells were again washed,counted, and resuspended to 1 x 107 PIN/mll in phos-phate-buiffered saline. Each preparation- was thentested in the zymosan-stimulated chemnilumiinescenceassay a second time. Those cells that hald respondedto chemotactic factor still yielded a significantly greaterclhemilumiinlesceince than did conitrol cells, as indicatedin the representative results of three experimenits withtwo chemotactic preparations shown in Fig. 2. Further-more, both cell preparations gave a response com-parable to that obtained prior to the additional 2.5-hincubation in media alone. This prolonged acti-vation was also observed in PMIN simiply incubated inthe caseini-autologous serumii miiixture or chemotacticC5 fragmenit, anid indicates that this enhancemiientwas mainitainied for at least 2.5 h.
Superoxide atnion release fromti chleiniotactic factor-stuintlate(dPAIN. Stimulation of PMNwith opsonizedzymnosain not only results in chemiiiluminiescence, butalso in aIn increased prodluction acnd secretioni of super-oxide ainioni. Studies were designed to determiniewhether PMNstimultlated witlh chemnotactic factor and
extensively washed showed enhanced production ofsuperoxide anion. The same three groups of cells wereprepared, washed, and tested for superoxide aniongeneration in both zymosan-stimiiulated and unstimu-lated assays. The chemotactic mixture used was thecasein-atutologous serum mixtuire. Table II shows theresults of coontrol and chemotactic factor-stimulatedPMNfrom several different cell donors. It is evidentthat the secretion of superoxide anion as measuredby cytochrome c reduction is significantly elevatedboth in PMNincubiated with this chemiiotactic mixtureand in those that have migrated througlh the 5-am pore-size membrane in response to chemotactic factor. Thisincrease in superoxide anion release was apparent 1)othin the presence and absence of opsonized zymosan.The observed reduction of cytochrome c in theseexperiments was attributed to the production of super-oxide anion since cytochrome c reduction was totallyabrogated by the addition of 50 ,ug of superoxidedismiuitase.
Enihantced bactericidal activity b1 chenmotactic fac-tor-stiin ulalted neutrophils. Since both chemi-lumtiniescence aind superoxide anion production havebeen linked with bacterial killing (19, 20), experimentswere performed to determiine wlhether chemiotacticf actor enhancied PMN bactericidal activity. PMNfrom five dlonors were preparecl aind treated for 3 h at37°C xwith either 5% FCS or the casein-auitologoussertumii mnixtture; or the PMIN were allowed to migratein respoinse to the chemotactic preparation. These cell
in 55
x 50
45
~J40 I
135 3 13Frogment 2
w /0I
15 /I0-
5
7 13 1925 31 37 43 491 7 13 19 25 31 37 4349MINUTES
FIGURE 2 M\1aintenance of PMNactivation in the absence of chiemilotactic factor. The left sidle ofthe figtire shows the chemilumtiiinleseenice of PMIN that have imiigratedl to chenimotactic C5 fragimienit(0), andl the right sicle, that of PMNthtt have migrated to the caseini-auitologouis serumiil mixttire(0) for 3 h. Both are comilpared with PMNinculbated in FCS (0). Solidl lines shiow the restilts ofPXIN tested imilmedliately after the imiigration to chemotactic facitor or incuhation with FCS andldashed lines indicate the responise after cells wvere washe(d ainid incuhated ani additional 2.5 h in10% FCS in the ahsence of chemilotactic factor.
Enihlancem2enit (f Neutrophil Fntctiont 171

TABLE IISuperoxide Anion Generation by PMNActivated
with Chemotactic Factor
Release of superoxide anion from PMNpreparations in nanomoles of cytochrome c-
reduced/106 PMN
Incubatedwith
chemotactic Prepared byExperiments Control factor* chemotaxis
Unstimulated1 2.3 5.2 6.62 2.8 3.8 6.63 7.6 7.6 8.54 3.8 10.4 8.15 3.3 9.0 8.66 10.0 11.9 15.2
Mean+SDt 5.0+3.1 8.0+3.1 8.9±3.2
Zymosan-stimulated1 22.8 38.1 25.72 39.0 44.7 48.53 30.4 43.8 47.64 45.7 47.6 46.65 32.3 43.8 40.46 43.8 40.9 41.97 16.6 40.0 35.2
Mean±SDI 32.9±10.8 42.7±3.2 40.8±8.2
* The chemotactic factor used was a casein-autologous serummixture.t Statistical analysis gave a P < 0.05 for comparison of controlcells to chemotoactic factor-treated cells and those migratingto chemotactic factor in both zymosan-stimulated and unstim-ulated experiments. The difference between chemotactic fac-tor-treated cells and those migrating to chemotactic factor wasnot significant with either unstimulated or stimulated PMNby the paired t test.
preparations were then tested for bactericidal activityusing E. coli as a target microorganism. As shown inFig. 3, the bactericidal activity of PMNexposed tochemotactic factor and of those that migrated in re-sponse to cheinotactic factor is significantly greater(P < 0.05 by a paired t test analysis) than that of controlPMN. Although cells that migrated through the 5-,umpore-size membrane show a slightly greater bacteri-cidal activity than cells simply incubated in chemo-tactic factor, the difference was not statistically signifi-cant by a paired t test with this sample size.
Surface receptor analysis. It is feasible that modu-lation of surface opsonic receptors could account for thedifferences in the PMN responses described here.PMNpreparations incubated for 3 h at 37°C in either5% FCS or the casein-autologous serum mixture, aswell as PMNthat had migrated to this chemotacticpreparation, were tested for Fc and complement re-
ceptor positive cells. Table III shows the comparativeresults of tests for Fc receptor positive cells using boththe erythrocyte antibody-rosetting technique and thefluorescence technique. The table also shows results oftests for complement receptor positive cells assayed byerythrocyte-IgM antibody-complement rosette forma-tion. No significant difference in the percentage ofcells bearing Fc or complement receptors wasobserved among any of the three preparations.
Enhanced chemiluminescence in vivo by stimulatedguinea pig peritoneal exudate neutrophils. To equateour in vitro findings with an in vivo model, guineapigs were injected intraperitoneally with a 50%-saturated solution of casein in minimum essentialmedia. 4 h later, PMNwere removed from the peri-toneal cavity, washed, and compared by chemilumines-cence to peripheral blood PMNisolated from the sameguinea pig. Results (Fig. 4) indicate that PMNrespond-ing in vivo to an inflammatory stimulus show enhancedchemiluminescence compared with PMN obtainedfrom the peripheral blood of the same guinea pig. Thesedata support the in vitro evidence presented previ-ously and indicate that, in vivo, PMNmigrating to an
a,0
U,
0Z.B
Ez
Minutes
FIGURE 3 Enhancement of PMNbactericidal activity as a re-sult of chemotaxis and chemotactic factor exposure. Resultsshow the mean + 1 SDat each point for PMNfrom five differentsubjects. Killing curves are shown for FCS-treated PMN(0),chemotactic factor- (casein-autologous serum) treated cells(A), and cells that had migrated to the casein-autologousserum mixture (0). Both cells incubated with chemotacticfactor and those that had migrated to chemotactic factorshowed significantly greater bacterial killing (P < 0.05) at 30,60, and 90 min by paired t test.
172 D. E. Van Epps and M. L. Garcia

inflammatory foci show an enhanced stimulated chemi-lutminescence similar to those responding to a chemio-tactic stimulus in vitro.
DISCUSSION
Many previous studies have shown that the directaddition of chemotactic factors to PMNpreparationscan stimulate neutrophil function and metabolism.These changes include the following: (a) chemotacticformylated peptides stimulate neuitrophil chemi-luminescence (2); (b) exposure to C5a causes neuitro-phil aggregation (3); (c) bacterial derived chemotacticfactors, formyl-methionine-alanine, and trypsin-acti-vated complemiient increase intracellular levels ofcGMP (4); (d) f-Met-Leu-Phe and C5a increase as-sembly of microtubules (5, 6); (e) C5a, C3a, C567 (7),and formylated peptides (21) stimulate neutrophildegranulation (7); (f) f-Met-Leu-Phe, C5a, and bac-terial derived chemotactic factors increase celladherence (6, 8); and (g) f-Met-Leu-Phe stimutlatesneutrophil production of stuperoxide anion (6, 9). Thepresent study demonstrates that P\IN that have beenexposed to or have migrated to the chemotactic mixttureof caseiin and auitologous serumil are "activated" as indi-cated by enhanced chemiltuminescencee, superoxicleanIioIn productioni, and bactericidal activity. It is im-portant to note that in this study cells were washedfree of chemotactic factor in the media before testingsince many of the previous studies mentioned demon-strate the effect of direct addition of chemotactic factorto PMNpreparations. Althouigh washing the cells be-fore testing removes chemotactic factor in the media, it
TABLE IIIComparison of Sturface Fc and Conplenment Receptors
with Variouis Preparations of PMN
Per cent positive P\MN in each cell preparation*
Incubated with Cells miiigratinigReceptor cheiniotactic to chemiiotactic
assay Control tfactor ftactort
EA rosette (21)§ 79±+14 - 73+24EAC rosette (15)§ 67±+13 - 70±21Fluorescent Fe (5)§
technique 97±2 95+2 98±2
* Results are expressed as the meian percent positive cells-1 SD.t Chemotactic factor used was a mixtture of casein and autol-ogouIs sermn. Statistical analysis by paired t test showedl nosignificant difference between control and chemotactic factor-treated preparationis or conitrol P.MN compared with cells mi-gratinig to chemotactic factor.§ Numbers in parentheses equal the number of different donorcell preparations tested. EA, erythrocyte anitibody; EAC, eryth-rocyte-IgM antibody-complement.
00.x
z
zJ
-i
L&IU
A16p0
140
bx 12w
> 1.0M
i 0.8z40.6
'4
0
0
0
A
0.4
02-
P P
PB PEC
MINUTES
FIGURE 4 Chemiluminiescenice of guinlea pig peripheralblood (PB) PMN compared with caseini-stimuitilated peritonealexudate (PEC) PMN from the samile animilal. MIean+SEMIchemiltumiiinescence oni three different guiniea pig prepara-tions are shown on the left and integrated areas for each pairedpreparatiot) are showvn on the right.
does not mneani that all chemiotactic factor has beenremoved fromi the systemii since somie may be bound tocells or internalized. Although the effect of boundchemotactic factor on the assays performed is unknown,it 1hias been shown that, at least in the chemiluminies-cenc e assay systemii, the response of PMNto the chemo-tactic factor f-Met-Leu-Phe peaks at 2 min andl dropsoff exponentially thereafter (2). Becatuse PMNin thisstudy were incubbated for 3 h with chemotactic facetor,the chemiltumiiinescenice of PMNstimuitilated by chemno-tactic f:actor would have been far beyond its peak. Inthis study, enhanced stimulated chemiluminescenceoccurred with all tested chemotactic factors, includingchemotactic C5 fragment, f-Met-Leu-Phe, and ac mixtureof caseini aind fresh acutologouis serumi. Enhancemiienitwas generally greater in cells that hald actutallly migratedthrough al 5-pmi pore-size clhemiiotaxis membrane, thainin those simply incubated for the samiie timne in chemo-tactic f'actor. Additional sttudies (Fig. 2) show that PMNrespondling to chemnotactic factor (either the casein-autologous serum mixtture or chemiiotactic C5 fragment)maiintained this enhalnced chelmiiluminiescenece for atleast 2.5 h in the absence of chemotactic factor. It ispossible that the differences observed here betweencontrol PMNand those migratiing in response to achemotactic f:actor arose fromn the isolationi of a miioreactive subpopulation of P.MN moving through themiembrane. Such ta subpopulation of Fc receptor posi-
Enhanicemenit of Neutrophil Function 173
1.8 r
I

tive granulocytes has been described in a previousreport by Klempner and Gallin (22). In our study, how-ever, comparisons have not shown a significant dif-ference among the number of Fc or complement re-ceptor-bearing PMNin control preparations, chemo-tactic factor-treated preparationis, or cells prepared bychemotaxis (Table III).
Because enhanced chemiluminescence of PMNexposed to chemotactic factor persisted for at least 2.5 hin the absence of chemotactic factor, it may be thatactivation itself involves some more permanent bio-logic changes, possibly in cell metabolism, cytoskele-ton, or membrane structure. This would seem to pre-clude simply a short-term shift in intracellular regula-tors such as a cAMPor cGMPsince elevated levels ofcyclic nucleotides resulting from exposure of PMNtochemotactic factors and chemotactic modulators gen-erally decrease within 1 h (4).
Previous studies (6, 8) have shown that chemotacticfactors increase cell adherence, which could cause in-creased chemiluminescence and superoxide anion gen-eration in systems stimulated with zymosan. Similarly,adherence is also essential to the bactericidal assaysystem. Although increased cell adherence in chemo-tactic factor-activated PMNmay affect assays involvingzymosan and bacteria, it should not affect assays medi-ated by soluble stimuli. Since chemnotactic factor-activated PMNshow enhanced chemiluminescence inresponse to both Con A and PMN(Table I), it is ap-parent that adherence is not the only mechanism in-volved in this enhancement. Likewise, experimentsmeasuring superoxide anion generation by chemotacticfactor-activated PMNin the absence of zymosan alsoindicate that these cells do not need a particulatestimulus to show enhanced superoxide anion pro-duction.
Studies by Gallin et al. (10) have shown that PMN-degranulating agents such as PMA, Con A, and thecalcium ionophore A23187 increase PMNadherenceand affect chemotactic factor binding under the properconditions. These agents may act by exposing newmembrane at the cell surface as a result of the exocytosisof specific granules, a process also known to occurwhen neutrophils are exposed to chemotactic factor(7, 21). Whether or not such a phenomenon exposesnew opsonic receptors or granule-associated oxidaseactivity (23) is unknown. Such alterations in the cellmembrane could explain both increased chemi-luminescence and superoxide anion production in thechemotactic factor-activated neutrophil. Although exo-cytosis of granules may expose new membrane, thedata in Table III indicate that the percentage of Fcand complement receptor positive cells in control andchemotactic factor-treated cells does not differ. This isof interest, but does not discouint a change in the num-ber of receptors per cell.
It is apparent from Fig. 4 that the migration of PMNto the casein-stimulated peritoneal cavity of the guineapig results in a population of PMNwith enhancedchemiluminescence much like that observed in our invitro studies with human PMN. This enhancement invivo supports the contention that chemotactic factoractivation of PMNrepresents an important mechanismof increasing host defense at specific locations wherethese factors are generated.
It is of interest to note the similarity between studieswith activated or elicited macrophages and the studiespresented here with PMN. Recent studies by Johnstonet al. (16) demonstrate that both elicited and activatedperitoneal macrophages show enhanced stimulatedsuperoxide anion release as compared with residentmacrophages. Other investigators (24) have foundthat activated macrophages give an enhanced chemi-luminescence when stimulated with opsonized par-ticles. It appears from our study that peripheral bloodPMNmay be stimulated by chemotactic factor in vitroto show both enhanced chemiluminescence andsuperoxide anion generation. Furthermore, casein-elicited PMNobtained from the peritoneal cavity ofguinea pigs show enhanced chemiluminescence and inthis way resemble activated or elicited macrophages.
A recent study by Cross and Lowell (25) showedthat stimulated lymphocyte supernates also in-crease the bactericidal activity of PMN. Such an activa-tion could be due to the presence of PMNchemotacticfactors-known to be present in such preparations(26)-which could activate PMNin a manner similarto that observed here with other chemotactic factors. Inaddition, a report by Issekutz et al. (27), publishedduring the course of our study, indicates that PMNbactericidal activity for Staphylococcus aureus, E.coli, and Streptococcusfaecalis is increased by chemo-tactic factor exposure. Our study furthers their work andshows that (a) zymosan-, PMA-, and Con A-stimulatedchemiluminescence, as well as zymosan-stimulatedand unstimulated superoxide anion production, areincreased in PMNthat have migrated to or are exposedto chemotactic factor in a system where cells arewashed free of exogenious chemotactic factor; (b) thisenhanced activity persists for at least 2.5 h in the ab-sence of chemotactic factor; (c) enhanced PMNchemi-luminescence is also observed with casein-elicitedguinea pig PMNin vivo; and (d) enhanced PMNfunc-tion due to chemotactic factor exposure is not the resultof an increased number of Fc or comnplement receptorpositive PMN, as determined by the methods em-ployed here.
ACKNOWLEDGMENTS
Wewish to thank Drs. Kenneth Tung and Aneiko Meng forproviding guinea pig polymorphonuclear leukocytes essentialto this study, Dr. Richard Freer of the Medical College of
174 D. E. Van Epps and M. L. Garcia

Virginia for providing us with f-Met-Leu-Phe, Dr. Kay Knightof the University of Illinois for providing us with anti-b4allotype antisera, and Carol Arnold for her excellent secre-tarial assistance.
This study was supported by a grant from the NationalCancer Institute, CA20819; the Department of Health, Edu-cation, and Welfare; and the Arthritis Foundation.
REFERENCES1. Wilkinson, P. C. 1974. Chemotaxis and Inflammation.
Churchill Livingstone, Edinburgh, UK.2. Hatch, G. E., D. E. Gardner, and D. E. Menzel. 1978.
Chemiluminescence of phagocytic cells caused by N-formylmethionyl peptides. J. Exp. Med. 147: 182-195.
3. O'Flaherty, J. T., D. L. Kreutzer, and P. A. Ward. 1977.Neutrophil aggregation and swelling induced by chemo-tactic agents. J. Immunol. 119: 232-239.
4. Hatch, G. E., W. K. Nichols, and H. R. Hill. 1977. Cyclicnucleotide changes in human neutrophils induced bychemoattractants and chemotactic modulators. J. Im-munol. 119: 450-456.
5. Gallin, J. I., E. K. Gallin, H. L. Malech, and E. B. Cramer.1978. Structural and ionic events during leukocyte chemo-taxis. In Leukocyte Chemotaxis: Methods, Physiology,and Clinical Implications. J. I. Gallin and P. G. Quie,editors. Raven Press, New York. 123-140.
6. Boxer, L. A., M. Yoder, S. Bonsib, M. Schmidt, P. Ho, R.Jersild, and R. L. Baehner. 1979. Effects of a chemotacticfactor, N-formylmethionyl peptide, on adherence, super-oxide anion generation, phagocytosis and microtubuleassembly of human polymorphonuclear leukocytes. J.Lab. Clin. Med. 93: 506-514.
7. Becker, E. L., H. J. Showell, P. M. Henson, and L. S.Hsu. 1974. The ability of chemotactic factors to inducelysosomal enzyme release. I. The characteristics of re-lease, the importance of surfaces, and the relation ofenzyme release to chemotactic responsiveness. J.Immunol. 112: 2047-2054.
8. Smith, C. W., J. C. Hollers, R. A. Patrick, and C. Hassett.1979. Motility and adhesiveness in human neutrophils,effects of chemotactic factors. J. Clin. Invest. 63: 221-229.
9. Simchowitz, L., and I. Spilberg. 1979. Generation ofsuperoxide radicals by human peripheral neutrophils acti-vated by chemotactic factor. Evidence for the role ofcalcium. J. Lab. Clin. Med. 93: 583-593.
10. Gallin, J. I., D. G. Wright, and E. Schiffmann. 1978. Roleof secretory events in modulating human neutrophilchemotaxis.J. Clin. Invest. 62: 1364-1374.
11. Van Epps, D. E., and R. C. Williams, Jr. 1976. Suppressionof leukocyte chemotaxis by human IgA myeloma com-ponents. J. Exp. Med. 144: 1227-1242.
12. Van Epps, D. E., and L. L. Paxton. 1975. Chemotaxis as apreparative technique for human polymorphonuclearleukocytes. J. Lab. Clin. Med. 86: 309-314.
13. Zigmond, S. H., and J. G. Hirsch. 1973. Leukocyte loco-
motion and chemotaxis: new methods for evaluation anddemonstration of cell-derived chemotactic factor. J.Exp. Med. 137: 387-410.
14. Van Epps, D. E., J. S. Goodwin, and S. Murphy. 1978.Age-dependent variations in polymorphonuclear leuko-cyte chemiluminescence. Infect. Immun. 22: 57-61.
15. Harvath, L., and B. R. Andersen. 1979. Defective initia-tion of oxidative metabolism in polymorphonuclear leuko-cytes. N. Engl. J. Med. 300: 1130-1135.
16. Johnston, R. B., Jr., C. A. Godzik, and Z. A. Cohn. 1978.Increased superoxide anion production by immunologi-cally activated and chemically elicited macrophages. J.Exp. Med. 148: 115-127.
17. Van Epps, D. E., K. Reed, and R. C. Williams, Jr. 1978.Suppression of human PMN bactericidal activity byhuman IgA paraproteins. Cell. Immunol. 36: 363-376.
18. Winfield, J. B., P. I. Lobo, and M. E. Hamilton. 1977. Fcreceptor heterogeneity: immunofluorescent studies of B,T, and "third population" lymphocytes in human bloodwith rabbit IgG b4/anti b4 complexes. J. Immunol. 119:1778-1784.
19. Allen, R. C., S. J. Yevich, R. A. Orth, and R. W. Steele.1974. The superoxide anion and single molecular oxygen:their role in the microbicidal activity of the polymorpho-nuclear leukocyte. Biochem. Biophys. Res. Commun.60: 909-917.
20. Klebanoff, S. J. 1975. Antimicrobial systems of the poly-morphonuclear leukocyte. In The Phagocytic Cell in HostResistance. J. A. Bellanti and D. H. Dayton, editors.Raven Press, New York. 45-59.
21. Showell, H. J., R. J. Freer, S. H. Zigmond, E. Schiffmann,S. Aswanikumar, C. Corcoran, and E. L. Becker. 1976.The structure activity relations of synthetic peptides aschemotactic factors and inducers of lysosomal enzymesecretion for neutrophils. J. Exp. Med. 143: 1154-1169.
22. Klempner, M. S., and J. I. Gallin. 1978. Separation andfunctional characterization of human neutrophil subpopu-lations. Blood. 51: 659-669.
23. Iverson, D., L. DeChatetet, J. Spitznagel, and P. Wang.1977. Comparison of NADHand NADPHoxidase ac-tivities in granules isolated from human polymorpho-nuclear leukocytes with a fluorometric assay. J. Clin.Invest. 59: 282-290.
24. Schleupner, C. J., and L. A. Glasgow. 1978. Peritonealmacrophage activation indicated by enhanced chemi-luminescence. Infect. Immun. 21: 886-895.
25. Cross, A. S., and G. H. Lowell. 1978. Stimulation of poly-morphonuclear leukocyte bactericidal activity by super-natants of activated human mononuclear cells. Infect.Immun. 22: 502-507.
26. Altman, L. C. 1978. Chemotactic lymphokines: a review.In Leukocyte Chemotaxis: Methods, Physiology, andClinical Implications. J. I. Gallin and P. G. Quie, editors.Raven Press, New York. 267-286.
27. Issekutz, A. C., K. Lee, and W. D. Biggar. 1979. En-hancement of human neutrophil bactericidal activity bychemotactic factors. Infect. Immun. 24: 295-301.
Enhancement of Neutrophil Function 175