enraizamento de estacas e teor de glicosÍdeos … · (dados sem transformação) para mudas de...
TRANSCRIPT

UNIVERSIDADE ESTADUAL PAULISTA
FACULDADE DE CIÊNCIAS AGRÁRIAS E VETERINÁRIAS
CAMPUS DE JABOTICABAL
ENRAIZAMENTO DE ESTACAS E TEOR DE
GLICOSÍDEOS CARDIOATIVOS EM MUDAS DE Nerium
oleander L.
Denise Renata Pedrinho
Engenheira Agrônoma
JABOTICABAL - SÃO PAULO - BRASIL
Julho de 2006

2
UNIVERSIDADE ESTADUAL PAULISTA
FACULDADE DE CIÊNCIAS AGRÁRIAS E
VETERINÁRIAS
CAMPUS DE JABOTICABAL
ENRAIZAMENTO DE ESTACAS E TEOR DE
GLICOSÍDEOS CARDIOATIVOS EM MUDAS DE Nerium
oleander L.
Denise Renata Pedrinho
Orientadora: Profa. Dra. Kathia Fernades Lopes Pivetta
Co-Orientador: Prof. Dr. Silvio Favero
Tese apresentada à Faculdade de Ciências Agrárias e Veterinárias - Unesp, Campus de Jaboticabal, como parte das exigências para a obtenção do título de Doutor em Agronomia (Produção Vegetal).
JABOTICABAL – SP
Julho de 2006

3
DADOS CURRICULARES DA AUTORA
Denise Renata Pedrinho - nascida em 22 de junho de 1968, em Jaú, SP, filha
de Dilson Pedrinho e Apparecida Eugenia do Carmo Pedrinho, formada em
dezembro de 1992, pela Faculdade de Agronomia Dr. Francisco Maeda -
FAFRAM - Ituverava, SP. Ingressou em março de 2001, no Curso de Mestrado
em Agronomia na FCAV/UNESP, Área de Concentração em Produção Vegetal.
Desenvolveu o trabalho intitulado: ‘Efeito de armazenamento de hastes e
estacas no enraizamento de roseiras para corte nas quatro estações do ano’,
concluindo em julho de 2002. Em agosto de 2002, ingressou no curso de
Doutorado na mesma Universidade, sendo bolsista da CAPES. Atualmente é
professora no Curso de Agronomia e Gestão Ambiental da Universidade para o
Desenvolvimento do Estado e da Região do Pantanal – UNIDERP – MS.

4
Dedico e Ofereço
Aos meus Pais – “Dilson Pedrinho (in memorian) e
Eugenia Pedrinho”
Ao meu esposo Rogério Possionatto Giroldo
A toda minha família: Dilma, Lú, Tata, Jú, Junior, Cris e
Davi.

5
HOMENAGEM ESPECIAL
A MINHA QUERIDA ORIENTADORA – Profa. Dra. Kathia Fernandes Lopes
Pivetta, por todos os momentos especiais que passamos. Pela dedicação,
confiança, amizade e paciência. Pela força com que me conduziu até o
presente momento. Pelo grande incentivo em minha vida profissional e pelo
amor em orientar.
AO MEU CO-ORIENTADOR – Prof. Dr. Silvio Favero, pela grande contribuição
no trabalho desenvolvido. Pela orientação, estímulo e principalmente paciência.

6
AGRADECIMENTOS ESPECIAIS
A DEUS, que sempre e em todos os momentos me acolhe e ilumina.
Obrigada.
Ao Prof. Dr. Silvio Fávero
A Profa. M.Sc.Soraya Sólon
A Profa . M.Sc. Rosemary Mattias
Ao técnico da Horta do Campus III da UNIDERP, Marlos.
A acadêmica do Curso de Farmácia Danielle, pela dedicação e carinho.
A minha querida amiga Patrícia Unger Cesar Pizetta

7
AGRADECIMENTOS
À CAPES, pela concessão da bolsa de estudo.
À Coordenação do curso de pós-graduação em Agronomia: Produção
Vegetal, pela oportunidade.
A Universidade para o Desenvolvimento do Estado e da Região do
Pantanal - UNIDERP – MS
MUITO OBRIGADA À TODOS

8
SUMÁRIO
Página
LISTA DE TABELAS........................................................................................... viii
LISTA DE FIGURAS............................................................................................ x
RESUMO.............................................................................................................. 01
ABSTRACT.......................................................................................................... 03
1. INTRODUÇÃO................................................................................................. 05
2. REVISÃO DE LITERATURA........................................................................... 06
2.1. Aspectos gerais da espécie Nerium oleander L....................................... 06
2.2. Enraizamento de estacas........................................................................... 07
2.3. Aspectos gerais das plantas tóxicas........................................................ 08
2.4. Aspectos gerais dos metabólitos secundários.......................................... 12
2.5. Relação entre fertilidade de solo, estresse hídrico e saturação de
bases na produção de principio ativos..................................................... 15
2.6. Cromatografia por adsorção em CCD....................................................... 19
3. MATERIAL E MÉTODOS................................................................................ 20
3.1. Experimento I - Efeito de diferentes doses de nitrogênio no teor de
glicosídeos cardioativos...........................................................................
23
3.2. Experimento II - Efeito de diferentes doses de calcário (saturação de
base) no teor de glicosídeos cardioativos..............................................
23
3.3. Experimento III - Efeito do estresse hídrico, verificado por meio da
capacidade de campo na concentração de glicosídeos
cardioativos...............................................................................................
24
3.4. Análise quantitativa de glicosídeos cardioativos..................................... 25
4. RESULTADOS E DISCUSSÃO....................................................................... 27
4.1. Enraizamento de estacas herbáceas de duas variedades de Nerium
oleander L. em diferentes concentrações de IBA, no verão e no
inverno.......................................................................................................
27
4.2. Efeito de diferentes doses de nitrogênio na concentração de
glicosídeos cardioativos...........................................................................
37
4.3. Efeito de diferentes doses de calcário (Saturação de Base) no teor de
glicosídeos cardioativos.............................................................................
40

9
4.4. Efeito do estresse hídrico, verificado por meio da capacidade de campo
no teor de glicosídeos cardioativos...........................................................
44
4.5. Considerações gerais................................................................................. 45
5. CONCLUSÕES................................................................................................ 49
6. REFERÊNCIAS BIBLIOGRÁFICAS................................................................ 50

10
LISTA DE TABELAS
TABELAS Página
TABELA 1. Quadrados médios e médias obtidos nas análises de
variância para porcentagem de enraizamento em
estacas de espirradeira (Nerium oleander L.), de flores
branca e rosa, retiradas no verão e no inverno e tratadas
com diferentes concentrações de IBA (mg.kg-1).
Jaboticabal, SP, 2006........................................................
28
TABELA 2. Quadrados médios e médias obtidos nas análises de
variância para número de raízes em estacas de
espirradeira (Nerium oleander L.), de flores branca e
rosa, retiradas no verão e no inverno e tratadas com
diferentes concentrações de IBA (mg.kg-1). Jaboticabal,
SP, 2006............................................................................
31
TABELA 3. Quadrados médios e médias obtidos nas análises de
variância para comprimento médio de raízes (cm) em
estacas de espirradeira (Nerium oleander L.), de flores
branca e rosa, retiradas no verão e no inverno e tratadas
com diferentes concentrações de IBA (mg.kg-1).
Jaboticabal, SP, 2006........................................................
33
TABELA 4. Quadrados médios e médias obtidos nas análises de
variância para massa seca de raízes (g) em estacas de
espirradeira (Nerium oleander L.), de flores branca e
rosa, retiradas no verão e no inverno e tratadas com
diferentes concentrações de IBA (mg.kg-1). Jaboticabal,
SP, 2006............................................................................
35
TABELA 5. Quadrados médios obtidos nas análises de variância
para massa fresca (g), massa seca (g) e teor de
glicosídeos cardioativos (Mg) em mudas de Nerium
oleander L. tratadas com diferentes doses de
adubaçãonitrogenada.........................................................
37

11
TABELA 6. Quadrados médios obtidos nas análises de variância
para massa fresca (g), massa seca (g) e teor de
glicosídeos cardioativos (Mg) em mudas de Nerium
oleander L. tratadas com diferentes doses de calcário.....
41
TABELA 7. Quadrados médios obtidos nas análises de variância
para massa fresca (g), massa seca (g) e teor de
glicosídeos cardioativos (Mg) em mudas de Nerium
oleander L. cultivadas com diferentes quantidades de
água...................................................................................
45

12
LISTA DE FIGURAS
FIGURAS Página
FIGURA 1. Curva de regressão entre as concentrações de IBA
(mg.kg-1) e a porcentagem de enraizamento (dados
transformados em arc sen 100/x ), para estacas de
espirradeira de flores rosa e branca no verão e no
inverno. Jaboticabal, SP, 2006............................................
29
FIGURA 2. Curva de regressão entre as concentrações de IBA
(mg.kg-1) e o número de raiz (dados transformados em arc
sen 100/x ), para estacas de espirradeira de flores rosa
e branca no verão e no inverno. Jaboticabal, SP, 2006......
32
FIGURA 3. Curva de regressão entre as concentrações de IBA
(mg.kg-1) e o comprimento médio de raiz (dados
transformados em arc sen 100/x ), para estacas de
espirradeira de flores rosa e branca no verão e no
inverno. Jaboticabal, SP, 2006..........................................
34
FIGURA 4. Curva de regressão entre as concentrações de IBA
(mg.kg-1) e a massa seca de raiz (dados transformados
em arc sen 100/x ), para estacas de espirradeira de
flores rosa e branca no verão e no inverno. Jaboticabal,
SP, 2006..............................................................................
36
FIGURA 5. Curva de regressão entre doses de nitrogênio (sem a
presença de adubação nitrogenada; 2,5 g. N-1; 5 g. N-1;
7,5 g. N-1) e a massa fresca (dados sem transformação)
para mudas de espirradeira. Campo Grande, MS,
2006....................................................................................
38
FIGURA 6. Curva de regressão entre doses de nitrogênio (sem a
presença de adubação nitrogenada; 2,5 g. N-1; 5 g. N-1;
7,5 g. N-1) e a massa seca (dados sem transformação)
para mudas de espirradeira. Campo Grande, MS,
2006.....................................................................................
38

13
FIGURA 7. Curva de regressão entre doses de nitrogênio (sem a
presença de adubação nitrogenada; 2,5 g. N-1; 5 g. N-1;
7,5 g. N-1) e o teor de glicosídeos cardioativos (dados sem
transformação) para mudas de espirradeira. Campo
Grande, MS, 2006................................................................
39
FIGURA 8. Perfil cromatográfico obtido em cromatografia de camada
delgada de sílica - gel das amostras de glicosídeos
cardioativos de plantas de Nerium oleander L. de quatro
tratamentos (sem a presença de adubação nitrogenada;
2,5 g. N-1; 5 g. N-1; 7,5 g. N-1). Campo Grande, MS,
2006.....................................................................................
39
FIGURA 9. Curva de regressão entre doses de calcário (zero, 25%,
50%, 75% e 100%) e massa fresca (dados sem
transformação) para mudas de espirradeira. Campo
Grande, MS, 2006................................................................
41
FIGURA 10. Curva de regressão entre doses de calcário (zero, 25%,
50%, 75% e 100%) e massa seca (dados sem
transformação) para mudas de espirradeira. Campo
Grande, MS, 2006................................................................
42
FIGURA 11. Curva de regressão entre doses de calcário (zero, 25%,
50%, 75% e 100%) e teor de glicosídeos cardioativos
(dados sem transformação) para mudas de espirradeira.
Campo Grande, MS, 2006..................................................
43
FIGURA 12. Perfil cromatográfico obtido em cromatografia em camada
delgada de Sílica-gel das amostras de glicosídeos
cardioativos de plantas de Nerium oleander L. entre doses
de calcário (zero, 25%, 50%, 75% e 100%). Campo
Grande, MS, 2006................................................................
44
FIGURA 13. Curva de regressão entre capacidade de campo (25%,
50%, 75% e 100%) e massa fresca (dados sem
transformação) para mudas de espirradeira. Campo
Grande, MS, 2006................................................................
45
FIGURA 14. Curva de regressão entre capacidade de campo (25%,

14
50%, 75% e 100%) e massa seca (dados sem
transformação) para mudas de espirradeira. Campo
Grande, MS, 2006................................................................
46
FIGURA 15. Curva de regressão entre capacidade de campo (25%,
50%, 75% e 100%) e teor de glicosídeos cardioativos
(dados sem transformação) para mudas de espirradeira.
Campo Grande, MS, 2006...................................................
47
FIGURA 16. Perfil cromatográfico obtido em cromatografia em camada
delgada de sílica gel das amostras de glicosídeos
cardioativos de plantas de Nerium oleander L capacidade
de campo (25%, 50%, 75% e 100%). Campo Grande, MS,
2006.....................................................................................
47

15
ENRAIZAMENTO DE ESTACAS E TEOR DE GLICOSÍDEOS CARDIOATIVOS EM MUDAS DE NERIUM OLEANDER L.
RESUMO - A espirradeira, Nerium oleander L., apresenta grande importância
pela utilização no meio urbano e pela alta toxicidade da substância glicosídeos
cardioativos. Esta espécie é propagada por estacas, porém, não há estudos
sobre fatores que influenciam este processo. Muitas plantas utilizadas no meio
urbano apresentam substâncias consideradas tóxicas cuja produção pode ser
influenciada por vários fatores como fertilidade do solo e estresse hídrico. Este
trabalho objetivou, portanto, estudar o efeito das estações do ano e do ácido
indolbutírico (IBA) no enraizamento de estacas de 2 variedades de espirradeira,
determinadas pela coloração das flores (rosa e branca), bem como, estudar o
teor de glicosídeos cardioativos em mudas de Nerium oleander L. Os
experimentos sobre enraizamento de estacas foram instalados na UNESP,
Campus de Jaboticabal, no verão e no inverno. O delineamento experimental
foi em blocos casualizados, seguindo esquema fatorial 2 x 2 x 4 (2 variedades,
combinadas com 2 estações do ano e com 4 concentrações de IBA – 0, 1000,
2000 e 4000 mg.kg-1). As avaliações foram realizadas 60 dias após a estaquia,
observando-se: porcentagem de enraizamento; número médio, comprimento e
massa seca de raízes. Concluiu-se que o enraizamento das variedades de
Nerium oleander L. definidas pela coloração da flor, ou seja, rosa e branca foi
superior no verão, ou seja, 97% para ambas as variedades quando comparado
com o inverno (77% e 56%, respectivamente para as variedades de flores rosa
e branca); a variedade de flores rosa apresentou maior porcentagem de
enraizamento e massa seca de raiz no inverno e maior número e comprimento
de raiz no verão; o acido indolbutírico foi efetivo para aumentar a porcentagem
de enraizamento nas concentrações de 1000 e 2000 mg.L-1 e número,
comprimento e massa seca de raiz na concentração de 2000 mg.L-1. Para
estudar o efeito do nitrogênio, calagem (saturação de bases) e quantidade de
água no solo (capacidade de campo) na concentração de glicosídeos
cardioativos em mudas de Nerium oleander L., o trabalho foi conduzido na
Universidade para o Desenvolvimento do Estado e da Região do Pantanal –
UNIDERP, em Campo Grande, MS, utilizando o delineamento experimental foi

16
em blocos casualizados. Para o estudo do efeito do nitrogênio, foram 4
tratamentos, (sem a presença de adubação nitrogenada; 2,5 g planta-1 de N;
5,0 g planta-1 de N; 7,5/g planta-1 de N), 5 repetições e 4 plantas por parcela,
totalizando 80 plantas. Para o estudo do efeito da saturação de bases foram 5
tratamentos (testemunha, 25% da saturação de bases recomendada; 50% da
saturação de bases recomendada; 75% da saturação de bases recomendada;
100% da saturação de bases recomendada) 4 repetições e 4 plantas por
parcela, totalizando 80 plantas e para o estudo do efeito da capacidade de
campo foram 4 tratamentos (25% da capacidade de campo - CC; 50% CC;
75% CC; 100% CC), 5 repetições e 4 plantas por parcela, totalizando 80
plantas. As avaliações foram realizadas 60 dias após o plantio das mudas. A
análise quantitativa dos glicosídeos cardioativos foi realizada por meio de
ensaio gravimétrico após extração seletiva dos glicosídeos. Concluiu-se a
adubação nitrogenada e a calagem aumentaram a produção de biomassa e
diminuíram no teor de glicosídeos cardioativos e o aumento da quantidade de
água no solo aumentou a produção de biomassa até 75% da capacidade de
campo e aumentou também o teor de glicosídeos cardioativos.
Palavras-chave: estaquia, IBA, espirradeira, oleandrina

17
ROOTING OF CUTTINGS AND GRADE OF CARDIOATIVE GLUCOSIDES IN
SEEDLINGS OF NERIUM OLEANDER L.
ABSTRACT – The oleandrine, Nerium oleander L., shows a great importance
by the use in urban center and by the rise of toxicity of the substance
cardioative glucoside. This specie is propagated by cutting, but there are not
studies about the factors that have influence in this process. Many plants
utilized in the urban center shows substances considered toxic whose
production could be influenced by some factors, like fertility of the soil and
hydric stress. This research had the objective, therefore, study the effect of the
seasons of the year and the indolbutiric acid (IBA) in rooting of cuttings of two
varieties of oleandrine, determined by the color of the flowers (rose and white),
so study the grade of cardioative glucosides in seedlings of Nerium oleander L.
The experiments about rooting of cuttings were carried in the UNESP, Campus
of Jaboticabal, São Paulo State, Brazil, in summer and winter. The experimental
delineation was in randomized blocks, in factorial 2 x 2 x 4 (2 varieties,
combined with 2 seasons of the year and 4 concentration of IBA – 0, 1000,
2000 and 4000 mg.Kg-1. The evaluations were realized 60 days after the
cutting, observing: percentage of rooting; number medium, length and dray
mass to varieties of Nerium oleander L. defined by the color of the flowers,
being rose and white higher in the summer, with 97% when compared with
winter (77% and 56% respectily for the varieties with flowers rose and white);
the variety with rose flower showed better percentage of rooting and dry mass
in the winter and better number and length of root in summer; the indolbutiric
acid increased the percentage of rooting in the concentrations of 1000 and 2000
mg. kg-1and number, length and dry mass of the root in concentration of 2000
mg. kg-1. To study the effect of nitrogen fertilization, saturation for bases, and
quantity of water in the soil (field capacity) in concentration of cardioative
glucosides in seedlings of Nerium oleander L., the research were carried in the
UNIDERP, in Campo Grande city, MS state, Brazil, using the experimental
delineation in randomized blocks. To the study of the effect of the nitrogen,
were 4 treatments (without nitrogen; 2,5g of N per plant; 5,0 g of N per plant;
and 7,5 g of N per plant), 5 replications and 4 plants by parcel, totalizing 80
plants. To the study of saturation for bases were 5 treatments (witness, 25%,

18
50%, 75% and 100% of saturation for bases recommended), 4 replications and
4 plants by parcel, totalizing 80 plants, and to the study of the field capacity
were 4 treatments (25%; 50%; 75% and 100% of the field capacity), 5
replications and 4 plants by parcel, totalizing 80 plants. The evaluations were
realized 60 days after the seedlings were planted. The quantitative analysis of
the cardioative glucosides was realized by gravimetric test, after selective
extraction of the glucoside. Were conclude that the nitrogen fertilization and the
saturation for bases increase the biomass production and reduce the grade of
cardioative glucoside, and the increase of the quantity of water in the soil raised
the biomass production until 75% of the field capacity and increased the grade
of cardioative glucosides too.
Key-words: cutting; IBA; oleandrine; oleander.

19
1. INTRODUÇÃO
A espécie (Nerium oleander L.), pertencente à família Apocynaceae,
apresenta grande importância pela sua utilização generalizada no paisagismo
de vários países, tanto em praças públicas, residências, como na arborização
urbana. É uma planta arbustiva muito ramificada e florífera que atinge de 3 a 4
m de altura. A propagação comercial é feita por estaquia herbácea, entretanto,
a produção de mudas por este processo é influenciada por diversos fatores,
que podem ser inerentes à própria planta ou às condições do ambiente, como,
a variedade, a época de coleta das estacas e a utilização de auxinas.
A espirradeira é uma planta ornamental considerada altamente tóxica
devido à presença de metabólitos secundários (glicosídeos cardioativos), em
todas as suas partes, podendo causar danos em potencial quando ingerida
pelo homem ou animal.
Embora existam relatos de fatores fitotécnicos influenciando na
concentração de princípios ativos tóxicos (metabólitos secundários) de diversas
espécies vegetais, utilizadas para as mais variadas finalidades, entre elas
medicinais, não foi encontrado na literatura trabalhos com o objetivo de diminuir
a concentração destes metabólitos, em plantas consideradas tóxicas e que são
amplamente utilizadas no paisagismo e na arborização urbana, como é o caso
da ornamental Nerium oleander L., visando uma convivência com menores
riscos à população.
Diante do exposto, estudos que possam interferir na qualidade das
mudas e na quantidade do princípio ativo de espécies ornamentais
consideradas tóxicas, influenciando a diminuição dos mesmos, são muito
importantes, visto que, nada foi encontrado sobre o assunto.
Neste contexto, este trabalho objetivou estudar o efeito das estações do
ano e do ácido indolbutírico (IBA) no enraizamento de estacas de 2 variedades
de espirradeira, determinadas pela coloração das flores (rosa e branca), bem
como, estudar o efeito da adubação nitrogenada, da saturação de bases e da
capacidade de campo na biomassa e no teor de glicosídeos cardioativos em
mudas de Nerium oleander L.

20
2. REVISÃO DE LITERATURA
2.1. Aspectos gerais da espécie Nerium oleander L.
A espécie Nerium oleander L., comumente chamada de espirradeira ou
oleandro, é originaria do Mediterrâneo e pertence à Família Apocynaceae. É
um arbusto grande ou arvoreta, apresentando 3 a 5m de altura, lactescente,
muito ramificado e florífero, de folhas persistentes e coriáceas. As flores são
brancas, róseas ou vermelhas e formam-se na primavera e verão. Há diversas
variedades que se diferenciam pelo colorido das flores, as quais podem se
simples ou dobradas, bem como as folhas variegadas. É muito cultivada em
parques, jardins e arborização de ruas, inclusive em regiões de clima frio e
mesmo áridas (LORENZI e SOUZA, 2001).
De acordo com OLIVEIRA et al. (2003), a espécie Nerium oleander é
muito apreciada como ornamento, porém, em alguns paises da África do Sul,
são comumente utilizadas em suicídio. Segundo os autores a ingestão dessa
planta causa distúrbios gastrintestinais intensos, caracterizados por náuseas,
vômitos, cólicas abdominais, diarréia e arritmia cardíaca.
Seu emprego como abortivo tem provocado inúmeros acidentes tóxicos,
alguns deles fatais, mãe e feto morrem juntos. A análise fotoquímica desta
planta mostrou que as folhas, cascas, raízes, sementes e flores, contêm vários
glicosídeos cardioativos (metabólitos secundários), sendo o principal deles a
oleandrina, bem como, flavonóides e outros compostos comuns a diversas
plantas, sem atividade apreciável, como o ácido betulínico e outros de estrutura
triterpenóide livre ou combinado com ácido cumárico, dos quais o cis e o trans-
carenino tem atividade citotóxica (LORENZI e MATOS, 2002).
SIMÕES et al. (2000) consideram a espirradeira uma planta de grande
importância toxicológica devido à presença de metabólitos secundários
(glicosídeos cardioativos), sendo que, por esse motivo, todas as partes da
planta são consideradas tóxicas e quando ingeridas causam náuseas, vômitos,
diarréia, sintomas neurológicos como desorientação e dor de cabeça e ainda
arritmia cardíaca.

21
De acordo com STEYN (1934), há também uma série de plantas
ornamentais, cujo néctar é tóxico e que, conseqüentemente, quando visitadas
por abelhas melíferas produz mel tóxico. Destaca ainda que pode existir muitos
casos de intoxicação por mel e que até as abelhas podem ser intoxicadas pelo
mel produzido a partir dessas plantas tóxicas. O autor comenta que Nerium
oleander L. pode ter produzido mel tóxico na África do Sul e que o mel de
Rhododendron luteum, é tido como causa de intoxicação em massa de
soldados gregos numa campanha na Ásia Menor, na antiguidade; ressalta
ainda que o conhecimento da toxidez das plantas se remota aos nossos
antepassados.
2.2. Enraizamento de estacas
A estaquia é um processo de propagação assexuada altamente
desejável, principalmente pelo fato das plantas originadas serem geralmente
idênticas, além de simples, rápido e não requer técnicas especiais
(HARTMANN et al., 1997).
Estaca é qualquer parte destacada da planta-mãe, capaz de regenerar
uma nova planta completa. A formação de raízes adventícias ocorre em duas
fases que, geralmente, são seqüenciais: a iniciação, caracterizada pela divisão
e diferenciação de certas células em um primórdio radicular, e o crescimento,
no qual a raiz primordial se expande por meio da divisão e alongamento das
células (JANICK, 1966).
As estacas são classificadas quanto à posição no ramo, em apicais,
medianas e basais e, quanto ao estádio de desenvolvimento, em lenhosas,
herbáceas e semilenhosas ou semi-herbáceas (SOUZA, 1977).
Estacas lenhosas seriam aquelas que possuem tecidos fortes,
endurecidos e resistentes, enquanto herbáceas as têm com aspecto suculento,
pouco consistente. As semilenhosas ou semi-herbáceas apresentam um
intermediário entre os dois extremos (SOUZA e INFORZATO, 1959).
A diferenciação e o desenvolvimento de raízes na propagação de
plantas por estaquia dependem de diversos fatores, tais como características
intrínsecas da espécie, tipo de estaca, ambiente de enraizamento e presença

22
de indutores (HARTMANN et al., 1997), além de outros, como substrato,
irrigação, fotoperíodo, pragas e doenças.
As auxinas desempenham um papel importante, sendo que, o efeito
principal sobre o enraizamento de estacas está ligado à sua ação sobre a
indução de primórdios radiculares (KRAMER e KOZLOWSKI, 1960; HAISSIG,
1970; HARTMANN et al., 1997).
A aplicação exógena de auxina também proporciona maior
porcentagem, velocidade, qualidade e uniformidade de enraizamento de
estacas (HARTMANN et al., 1997).
Segundo KRAMER e KOZLOWSKI (1960), os fitormônios artificiais
(reguladores de crescimento), como os ácidos naftalenoacético (ANA),
indolbutírico (IBA) e indolacético (AIA) e seus sais de ésteres de potássio, são
habitualmente utilizados no tratamento de estacas com vista a um melhor
enraizamento.
Muitos estudos têm sido realizados para traçar a curva de resposta de
diferentes espécies, cultivares e tipos de estaca às diferentes concentrações de
auxina, entre eles LORETI e HARTMANN (1965), GRAZIANO (1981),
HOWARD (1985), THOMPSON (1986), PETRECHEN (1988), STOLTZ e
ANDERSON (1988), SHARMA e CLAVERIE (1989), PIVETTA (1990),
PIVETTA (1994), MARTINS (1998), BASTOS (2002), PEDRINHO (2002),
SARZI (2002), OLIVEIRA et al. (2003), RAMOS et al. (2003), CARVALHO
(2005) e FARIA e SACRAMENTO (2005).
A época de coleta das estacas influencia o enraizamento das espécies,
sendo a primavera e o verão as estações que proporcionam maior capacidade
às plantas de enraizar; porém, a coleta pode ser realizada em qualquer época
do ano, dependendo apenas da disponibilidade e necessidade de material
(MARTINS, 1998).
O melhor enrazaimento que ocorre, para muitas espécies, nas estações
de primavera e verão é devido ao fato de serem períodos de intenso
crescimento vegetativo, em que as estacas estão mais herbáceas, facilitando,
de modo geral, o desenvolvimento de raízes em espécies com dificuldade de
enraizamento (FACHINELLO et al., 1995).

23
HARTMANN e LORETI (1965) verificaram que estacas de oliveira (Olea
europea L.) enraizaram bem na primavera ou verão e muito pouco no inverno.
ONO e RODRIGUES (1996) explicam este fato pela baixa taxa fotossintética
no inverno, em conseqüência da baixa luminosidade e temperatura ambiente.
Assim, os metabólicos da fotossíntese são reduzidos, sendo insuficientes para
a iniciação e desenvolvimento das raízes.
FERNANDES et al. (1979), ao realizarem o estaqueamento de azaléia
(Rhododendron simsii, Planch) em épocas diferentes, correspondentes a
quatro estações do ano, observaram que o estaqueamento feito na primavera
foi o que apresentou o maior número de plantas enraizadas.
BASTOS (2002), também verificou em seu trabalho de enraizamento de
estacas de carambola (Averrhoa carambola L.) que no verão o enraizamento
foi superior.
CARVALHO et al. (2005), afirmam que o verão é a melhor época para a
o enraizamento de estacas de lichieira (Litchi chinensis).
2.3. Aspectos gerais das plantas tóxicas
Definem-se como plantas tóxicas todo vegetal que, ingerido pelo
homem ou animais, em condições naturais, é capaz de causar danos que
refletem na saúde e vitalidade (HOEHNE, 1939; BARCELLOS, 1998;
TOKARNIA et al., 2000; HARAGUCHI, 2003).
Existem em todas as partes do mundo plantas que encerram venenos ou
princípios tóxicos, capazes de perturbar ou aniquilar as funções essenciais à
vida dos organismos animais e do próprio homem (HOEHNE, 1939).
Segundo BARCELLOS (1998), muitas plantas tóxicas também são
plantas ornamentais e utilizadas no paisagismo, sendo facilmente encontradas
nos jardins causando inúmeros acidentes. O autor afirma que vive-se em um
mundo privilegiado em que tem-se amplo conhecimento sobre os usos das
plantas, inclusive sobre os efeitos tóxicos de inúmeras espécies, principalmente
quando comparados aos nossos antepassados. Entretanto, destaca que,
apesar deste amplo conhecimento, ocorrem hoje inúmeros acidentes com
plantas tóxicas, e geralmente acontece com espécies já caracterizadas como
tóxicas, o que leva a crer que este conhecimento não é bem difundido.

24
Muitos fatores influenciam na condição de uma planta ser tóxica ou não,
tais como a concentração do princípio considerado tóxico, o qual pode ser
afetado pela fertilidade do solo, luminosidade e umidade (HOHENE, 1939).
BARCELLOS (1998) cita que o poder de elaborar substâncias tóxicas,
cuja composição química é extremamente variável, (alcalóides, glicosídeos,
toxalbuminas, saponinas, compostos cianogenéticos, enzimas, etc) se distribui
de modo absolutamente irregular nos diferentes grupos que integram o reino
vegetal, havendo casos em que numa mesma espécie existam, lado a lado,
variedades inócuas e venenosas. Estas substâncias podem ser encontradas
em todas as partes de um vegetal, contudo em um ou mais órgãos haverá um
acúmulo preferencial dessas substâncias, tais como: tecidos com crescimento
ativo, células epidérmicas, bainhas vasculares e ainda vasos lactíferos.
Em diversas ocasiões e em diferentes grupos sociais, o sumo de plantas
tóxicas foi empregado como meio de execução da pena capital como no caso
de Sócrates, condenado a beber uma taça de sumo de cicuta. Já os grupos
primitivos, por outro lado, utilizavam plantas tóxicas para pescas, (plantas
ictiotóxicas), para preparar iscas envenenadas e assim abater feras e animais
predadores. Usavam-nas no fabrico de venenos sagitários, para caça e para
guerra e ainda como medicamentos. Este último uso foi incorporado pelos
civilizados e muitas plantas consideradas venenosas são hoje cultivadas e
constituem valiosos recursos no tratamento de várias enfermidades, sendo
que, de plantas tóxicas para animais de sangue frio, fizeram-se inseticidas que
ajudam o homem a combater os insetos nocivos e vetores de doenças, com
isso, tem-se observado que muitas plantas consideradas tóxicas são evitadas
por animais ou insetos em sua dieta (HOEHNE, 1939).
BARCELLOS (1998) destaca que o modo de ação destes princípios
tóxicos dependem da forma de intoxicação, podendo esta ser via ingestão,
contato e até inalação de odores. Além disso, a reação do ser humano,
também é bastante variável, em função, principalmente, da faixa etária,
nutrição e a própria constituição genética do individuo.
O mesmo autor explica que o estudo sobre os efeitos da ingestão de
plantas ornamentais tóxicas em seres humanos tem apenas como base
indicações de plantas com potencial tóxico, já que não há a possibilidade da

25
realização de testes experimentais, ao contrário de estudos feitos em animais
experimentalmente intoxicados.
Em algumas obras clássicas como HOEHNE (1939) destacam-se
centenas de plantas como sendo tóxicas. No entanto, a margem de certeza
sobre a toxicidade de uma planta é limitada por uma série de fatores. Para
ocorrer à intoxicação, seja por ingestão ou pelo contato com a pele, devem ser
vencidos, no processo, os mecanismos próprios de defesa de cada organismo.
Assim, uma planta pode ser potencialmente tóxica e, apesar disso, não
provocar a intoxicação; nesse caso, há uma convicção equivocada de ausência
de toxidade. Apesar dessa margem de incerteza, existe um número elevado de
plantas tóxicas documentadas.
HOEHNE (1939) explica que a intoxicação pode ser aguda ou crônica.
Aguda ocorre quase sempre por ingestão, geralmente crianças ou pessoas não
esclarecidas, que desconhecem as plantas e as ingerem. Existe também a
exposição crônica, evidenciada por manifestações cutâneas em decorrência do
contato sistemático com vegetais, além da utilização continuada de certas
espécies vegetais sob a forma de pó para inalação, fumos ou infusões, a fim de
obter efeitos alucinógenos ou entorpecentes. Sendo assim, é extremamente
importante, listar as plantas ornamentais que apresentam substâncias que
comprovadamente causam problemas ao ser humano e aqueles onde já foram
observados efeitos nocivos após a ingestão, contato ou inalação.
Dados do Sistema Nacional de Informações Tóxico-Farmacológicas da
Fundação Oswaldo Cruz (FIOCRUZ, 2006) mostram que no ser humano, os
sinais causados pela ingestão de plantas tóxicas no organismo vão desde
cólicas até a morte, passando por irritações na pele, convulsões e vômitos.
Para BARCELLOS (1998), as plantas ornamentais são as que se
encontram mais próximas ao homem sendo de fácil acesso, plantas essas que
embelezam trazendo sensação de bem-estar. Entretanto, podem também
trazer danos irreparáveis, pois por trás de tanta beleza pode existir na sua
constituição química, elemento não apropriado à espécie humana e animais.
O termo tóxico quando aplicado a um vegetal é adjetivo bastante vago,
porque cada um o interpreta de modo diferente. Os químicos dão-lhe um
sentido, os leigos outro e os fisiologistas ainda outro. Dizer o que é tóxico é tão
difícil como explicar o que faz bem e o que faz mal, porque o que para um é

26
veneno, para outro pode ser completamente inócuo, e mesmo as substâncias
mais tóxicas, quando usadas em doses moderadas podem ser terapêuticas;
entretanto, existem algumas substâncias tóxicas que sempre e em qualquer
via, são nocivas (HOEHNE, 1939).
Segundo dados da FIOCRUZ (2006), cerca de 60% dos casos de
intoxicação por plantas tóxicas no Brasil ocorrem com crianças menores de
nove anos, sendo a maioria acidental, por desconhecimento do potencial tóxico
da espécie.
OLIVEIRA et al. (2003) afirma que as intoxicações por plantas
correspondem a uma parcela significativa do número de atendimentos em
hospitais, ocupando no Brasil o nono lugar, sendo responsável por 2% dos
casos de envenenamentos.
2.4. Aspectos gerais dos metabólitos secundários
SCHENKEL et al. (2003) afirmam que, o número de vegetais superiores
é estimado em mais de 250 mil espécies, sendo que, muitas dessas espécies
produzem substâncias capazes de exercer ação tóxica sobre organismos vivos.
Essas substâncias são denominadas metabólitos secundários e tem função de
defesa contra predadores.
Os vegetais produzem uma grande quantidade de compostos orgânicos
que parecem não ter função direta no seu crescimento e desenvolvimento. Tais
substâncias são conhecidas como metabólitos secundários, produtos
secundários ou produtos naturais. Os metabólitos secundários não apresentam
ação direta sobre os processos como fotossíntese, respiração, transporte de
solutos, translocação e síntese de proteínas, assimilação de nutrientes,
diferenciação ou síntese de carboidratos e lipídeos (TAIZ e ZEIGER, 2004).
Segundo os mesmos autores, os metabólitos secundários também
diferem dos metabólitos primários (aminoácidos, nucleotídeos, açucares e acil
lipídeos) por apresentarem distribuição restrita no reino vegetal, ou seja,
metabólitos secundários específicos são restritos a uma espécie vegetal ou um
grupo de espécies relacionadas, enquanto que metabólitos primários são
encontrados em todo reino vegetal.

27
De acordo com CARVALHO et al. (2003) as plantas são ricas em
substâncias químicas que aparentemente não estão relacionadas com os
processos metabólicos normais como a fotossíntese a respiração e o
crescimento. Estas, chamadas de substâncias químicas secundárias, devem
estar relacionadas, pelo menos parcialmente, com a imobilidade das plantas,
uma vez que elas não podem escapar das pressões ambientais, sendo suas
únicas defesas as estruturas físicas e a composição química.
GOTTILIEB (1996) sugere que as plantas sintetizam apenas metabólitos
primários e genéricos, sendo todos igualmente necessários à vida vegetal.
Esses metabólitos genéricos chamados pelo autor de metabólitos especiais,
também poderiam adaptar o organismo vegetal a pressão de herbivoria, mas
sua função de proteção seria apenas acidental, e não predestinada a essa
atividade.
CASTRO et al. (2004) explica que a biossíntese de metabólitos
secundários é realizada por rotas metabólicas específicas do organismo
vegetal, ocorrendo estreita relação entre essas rotas e aquelas responsáveis
pela síntese de metabólitos primários. Essas rotas metabólicas são
interconectadas, e as rotas que sintetizam metabólitos primários fornecem
moléculas que são utilizadas como precursoras nas principais rotas de síntese
de metabólitos secundários. Para os autores, embora se faça a divisão em
metabolismos primário e secundário, o metabolismo deve ser considerado
como um todo, na produção de metabólitos secundários e primários.
Embora o nível de metabólitos secundários seja controlado
geneticamente, a quantidade e a concentração desses compostos variam
acentuadamente em função das condições ambientais. Apresentam-se como
importantes fatores ambientais a luz, a latitude, a temperatura, as propriedades
químicas e físicas do solo, os ventos, os macro e micro nutrientes e a
disponibilidade hídrica (PALEVITCH, 1987). Segundo o autor, a produção de
biomassa e de fotoquímicos também está estreitamente relacionada com
aspectos agronômicos como o preparo do solo, fertilização e irrigação.
GROS et al. (1985) comentam que a característica mais importante da
maioria dos metabólitos secundários é a sua distribuição restrita na natureza,
que se limita a uma espécie ou a espécies relacionadas, permitindo a ela se
adequar às condições impostas pelo ambiente. É possível que alguns desses

28
compostos não sejam essenciais para o organismo que os produzem, mas em
geral devem ter algum significado biológico, presumindo-se que possuam
alguma função, provavelmente especifica.
GOTTLIEB et al, (1996) citam que outra característica dos metabólitos
secundários é a de proteger o metabolismo vegetal contra a ação de agentes
destruidores, como em situações de estresse.
Segundo TAIZ e ZEIGER (2004), os compostos que agem como defesa
nas plantas, contra fungos, bactérias e herbívoros, podem também torná-los
indesejáveis para o homem.
Os metabólitos secundários são divididos em três grupos principais,
sendo estes: terpenos, compostos fenólicos e compostos nitrogenados. Os
terpenos são tóxicos e agem na defesa dos vegetais, é o maior grupo de
produtos secundários e nessa classe estão inclusos os glicosídeos cardioativos
– triterpenos e óleos essenciais – monoterpenos (VICKERY e VICKERY, 1981;
HARBONE e BAXTER 1995; TAIZ e ZEIGER, 2004).
Os glicosídeos cardioativos são definidos por TAIZ e ZEIGER (2004),
especificamente, como triterpenos ativos, contra herbívoros vertebrados. São
compostos que contêm um ou mais resíduos de açúcar ligados, e apresentam
um gosto amargo, sendo extremamente tóxicos. Em humanos, eles
apresentam efeitos drásticos na musculatura cardíaca devido a sua influencia
nas ATPases ativadas por Na+/K+. Em doses cuidadosamente reguladas, eles
diminuem e fortalecem os batimentos cardíacos.
Os glicosídeos cardioativos são divididos em dois grupos, um com
compostos de cadeia de vinte e três carbonos, os cardenolídeos, e outro
composto de cadeia de vinte e quatro carbonos, os bufadienolídeos. Sua
atividade cardíaca esta associada a uma cadeia insaturada de lactona e a
estereoquímica da molécula. Os cardenolídeos são encontrados em varias
famílias vegetais, especialmente em Apocynaceae e nas espécies Digitalis,
enquanto bufadienolídeos são encontrados nas famílias Ranunculaceae e
Liliaceae (OLIVEIRA et al., 2003).
Segundo o mesmo autor esses glicosídeos são usados na medicina para
o tratamento da insuficiência cardíaca, e intoxicações podem ocorrer depois do
consumo de flores, folhas ou sementes de plantas que contem glicosídeos
cardioativos. Sintomas gastrintestinais são normalmente os primeiros

29
envolvidos. Eles incluem náuseas, vômitos, dores abdominais, diarréia, e
anorexia. Sintomas neurológicos são tardios e incluem vertigem, dor de
cabeça, tontura, fadiga, debilidade e alucinações, sendo que, overdoses levam
a parada cardíaca e a morte.
Para VICKERY e VICKERY (1981), embora os glicosídeos cardioativos
serem usados na medicina para o tratamento de insuficiência cardíaca,
intoxicações podem ocorrer depois do consumo de chás preparados por partes
da planta ou depois do consumo de flores, folhas ou sementes.
2.5. Relação entre fertilidade de solo, estresse hídrico e saturação de
bases na produção de principio ativos
De acordo com SALOMON (1994), variações nas condições edáficas
(fertilidade, teor de matéria orgânica e pH) e climáticas (temperatura e
precipitação) no cultivo influenciam a produção de óleo essencial, ou seja,
princípios ativos em flores de camomila.
MING (1992), estudando a adubação orgânica em Lippia alba, verificou
maiores produções de biomassa em níveis crescentes de incorporação de
matéria orgânica e uma relação inversa quanto aos teores de óleo essencial. O
autor discute um papel de defesa dos óleos essenciais em ambientes
desfavoráveis, enquanto em ambientes mais favoráveis, a energia da planta
estaria voltada para atividades de crescimento e desenvolvimento, do
metabolismo primário.
HORNOK (1983) ao utilizar quatro níveis de N (0, 80, 160 e 240
kg/ha), quatro de P (0, 50, 100 e 150 kg/ha) e quatro de K (0, 60, 120 e 180
kg/ha) em Ocimum basilicum, observou que o teor de óleo essencial aumentou
proporcionalmente com os níveis de N, P e K.
Para DRAGAR e MENARY (1995) soluções nutritivas contendo baixas
concentrações de N resultou em baixas taxas de crescimento, mas favoreceu
alto rendimento de óleo essencial para Olearia phlogopappa Labill. D. C.
Altos níveis de N no solo causam aumento na massa de folhas de
Mentha arvensis, embora tenha havido redução de óleo essencial com baixos
níveis de mentol (MAIA et al., 2001).

30
TAIZ e ZEIGER (2004) comentam que a deficiência ou excesso de
nutrientes podem interferir na biomassa e quantidade do principio ativo das
plantas.
Segundo TOKARNIA et al. (2000) a fertilidade do solo em pastagens,
influencia a toxidez de Manihot spp. e outras plantas cianogênicas.
Trabalhando com Lathyrus tingitanus e Mimosa sp., NOWACKI et al.
(1975) verificaram que com o aumento da adubação nitrogenada houve síntese
desproporcional do aminoácido lisina, o qual, uma vez acumulado torna-se
tóxico.
O mesmo ocorreu para Tabernaemontana pachysiphon Stapf sob três
concentrações de fertilizantes nitrogenados de liberação lenta (0, 3 e 6 kg m-3
de Osmocote) em casa de vegetação, quando HOFT et al. (1996) encontraram
maior acúmulo de principio ativo com aumento da fertilização nitrogenada.
BECKER et al. (2000), estudando o efeito da adubação nitrogenada e
calagem na produção de alcalóides totais em quebra-pedra Phyllanthus niruri
L., observaram um aumento gradual na concentração de alcalóides na matéria
seca da parte aérea à medida que aumentaram as doses de nitrogênio e
calcário aplicadas. Destacam ainda que, apesar do pouco conhecimento das
exigências e requerimentos nutricionais, a correção do solo e a fertilização
nitrogenada mostraram-se importantes na produção de biomassa e no aumento
do teor de alcalóides de quebra-pedra.
Por outro lado, SCHERMEISTER et al. (1960) observou que, em Atropa
belladonna L. um aumento no suprimento de nitrogênio promoveu maior
acúmulo de proteínas e compostos nitrogenados nos tecidos, enquanto os
níveis de alcalóides encontrados foram relativamente baixos sugerindo,
portanto, não haver relação direta entre alcalóides e proteínas.
Resultados semelhantes foram obtidos por MAPELI et al. (2005), onde
estudando o efeito do nitrogênio e fósforo na produção de biomassa e óleo
essencial em Camomilla recutita L., observaram que a adubação não
influenciou na produção de óleo essencial, porém, aumentou a produção de
biomassa.
OLIVEIRA JUNIOR et al. (2005) estudando o teor e rendimento de óleo
essencial na massa fresca de arnica em função da adubação e da calagem,
concluiu que, para massa fresca, há efeito positivo da calagem quando

31
aplicada juntamente com adubo orgânico. Entretanto, para o teor de óleo
essencial, o autor afirma que a calagem influenciou negativamente.
UNANDER e BLUMBERG (1990) afirmam que os princípios ativos das
plantas medicinais podem ser influenciados pelas condições ambientais, como
fertilidade do solo, pH, umidade, temperatura qualidade da luz.
Com relação ao estresse hídrico, STEPONKUS (1990) afirma que, a
deficiência de água altera vários processos bioquímicos e fisiológicos nas
plantas, induzindo respostas metabólicas e fisiológicas como o fechamento
estomático, declínio na taxa de crescimento, acúmulo de solutos e substâncias
antioxidantes, podendo causar injurias as plantas, no entanto a planta ajusta-se
fisiologicamente ao estado de menor disponibilidade de água.
TAIZ e ZEIGER (2004) definem déficit hídrico como todo conteúdo de
água de um tecido ou célula que está abaixo do conteúdo de água mais alto
exibido no estado de maior hidratação. Os autores afirmam que o estresse
hídrico tem vários efeitos sobre o crescimento, um dos quais é a limitação da
expansão foliar. A área foliar é importante, porque em geral a fotossíntese é
proporcional a ela. No entanto, a expansão foliar rápida pode afetar
desfavoravelmente a disponibilidade de água.
CARVALHO et al. (2003) estudando a disponibilidade de água no solo
com relação ao crescimento de artemísia, concluiu que quanto maior a
disponibilidade de água no solo, maior a massa seca e fresca da planta.
DHRU et al. (1991), estudando duas espécies Nerium oleander L. e
Urginea indica, demonstraram que o déficit hídrico reduziu a produção de
biomassa de ambas as plantas.
Segundo KUDREV (1994), a deficiência hídrica interrompe o processo
de crescimento, não apenas diminuindo o acúmulo de massa fresca e seca,
mas também alterando o processo de crescimento e acelerando o processo de
catabólicos.
EGERT e TEVINI (2002), não verificaram alterações significativas nos
teores de clorofila, proteínas e compostos antioxidantes quando expuseram a
planta condimentar Allium schoenoprosum à seca. Entretanto, TAIZ e ZEIGER
(2004) explicam que o estresse hídrico reduz, tanto a fotossíntese quanto o
consumo de assimilados nas folhas em expansão. Como conseqüência o
estresse hídrico diminui indiretamente a quantidade de fotossintatos exportados

32
das folhas. Entretanto, SANTOS et al. (2004) estudando o efeito do estresse
hídrico no teor de óleo essencial em Hyptis pectinata L. afirma que para as
plantas submetida à falta de água, o teor de óleo foi aproximadamente 55%
maior em relação aquelas irrigadas.
PALEVITCH (1987) comenta que, embora o nível de metabólitos
secundários seja controlado geneticamente, a quantidade e a concentração
desses compostos variam acentuadamente em função das condições
ambientais. Apresentam-se como importantes fatores ambientais a luz, a
latitude, a temperatura, as propriedades químicas e físicas do solo, os ventos,
os macro e micro nutrientes e a disponibilidade hídrica. Segundo o autor, a
produção de biomassa e de fotoquímicos também está estreitamente
relacionada com aspectos agronômicos como o preparo do solo, fertilização e
irrigação.
2.6. Cromatografia por adsorção em CCD
De acordo com SHARAPIN et al. (2000), a cromatografia baseia-se em
diferentes mecanismos de separação que empregam uma fase estacionária e
uma fase móvel, as quais são capazes de realizar a migração diferencial de
cada componente da amostra a ser analisada. Os diferentes mecanismos de
separação fundamentam-se em competição eletrostática, tamanho molecular,
capacidade de partição e ionização das substâncias a serem separadas com
relação às diferentes afinidades pela fase estacionária e fase móvel do
sistema.
Segundo o mesmo autor, na cromatografia por adsorção, o processo
cromatográfico ocorre pela capacidade adsortiva da fase fixa (sólido
adsorvente). A interação entre uma molécula e o adsorvente existirá por meio
de forças eletrostáticas. Como a maioria das fases fixas adsortivas é polar, as
substâncias de maior polaridade tendem a permanecerem retidas na fase fixa
ocorrendo, nesse caso, deslocamento de substâncias apolares pela afinidade
com o eluente (fase móvel menos polar do que a fase fixa).

33
Quando se utiliza um adsorvente póla, o eluente deve apresentar menor
polaridade, sendo que, o contrário também é verdadeiro. A migração diferencial
para o uso de adsorvente polar ocorrerá no sentido de deslocar as substâncias
mais apolares, por terem mais afinidade pela fase móvel e, reter as substâncias
mais polares, por terem mais afinidade pela fase fixa (WAGNER e
BLADT,1996).
Segundo os mesmos autores, no desenvolvimento tecnológico do
cromatográfo, emprega-se a fase fixa aderida a um suporte plano. Esse tipo de
sistema é conhecido como cromatografia em camada delgada e é
caracterizado por ser um método rústico (simples), de baixo custo e rápido,
entretanto, sua resolução oferece resultados qualitativos e em experimentos
comparativos com amostras padrão de concentração conhecida, pode fornecer
resultados semi-quantitativos. Os resultados são obtidos por manchas que se
revelam em diferentes locais da cromatoplaca (Rf: retention factor).

34
3. MATERIAL E MÉTODOS
O trabalho foi conduzido em duas etapas:
Etapa I: Enraizamento de estacas herbáceas de duas variedades de
Nerium oleander L. (rosa e branca) em diferentes concentrações
de IBA, no verão e no inverno
As estacas foram coletadas de duas variedades de Nerium oleander L,
definidas pela coloração das flores (rosa e branca), no município de
Jaboticabal, SP.
Foram utilizadas estacas herbáceas, com aproximadamente 15 cm de
comprimento, com um par de folhas e duas gemas.
As estacas cortadas foram tratadas com ácido indolbutírico via pó,
sendo cada concentração misturada com talco industrial. Para o tratamento,
passou-se a extremidade basal da estaca em um recipiente contendo a
mistura, sendo logo depois estaqueadas em bandejas de poliestireno com 72
células, preenchidas com vermiculita de granulometria média. As bandejas
foram mantidas em câmara de nebulizaçao intermitente do Viveiro
Experimental de Plantas Ornamentais e Florestais da FCAV/UNESP, município
de Jaboticabal, SP. O município localiza-se a 21o 15’ 22” de latitude Sul e 48o
18’ 58” de longitude Oeste, a uma altitude média de 595 metros acima do nível
do mar. O clima, de acordo com classificação de Köppen, é do tipo Cwa, ou
seja, subtropical úmido com estiagem no inverno. No período de 1971 a 2005 a
média da temperatura máxima foi de 28,9ºC, a média da temperatura mínima
de 16,8ºC, a média da temperatura média de 22,2ºC e a média da umidade
relativa do ar de 70,8%, segundo dados coletados na Estação
Agrometeorológica da FCAV/UNESP, Campus de Jaboticabal.
Os tratamentos utilizados foram 2 épocas de estaquia (verão – 12/02/04,
e inverno – 21/07/2004) e 4 concentrações de ácido indolbutírico - IBA (0,
1000, 2000 e 4000 mg.kg-1) para 2 variedades de espirradeira determinadas
pela coloração das flores (rosa e branca).
O delineamento experimental utilizado foi o inteiramente casualizado,
seguindo o fatorial 2 x 2 x 4 (duas estações, duas variedades e quatro

35
concentrações de IBA), apresentando assim 16 tratamentos e quatro
repetições perfazendo um total de 64 parcelas. Cada parcela constituída por
três estacas, num total de192.
As avaliações foram feitas 60 dias após a estaquia, anotando-se o
número de estacas enraizadas (calculando posteriormente a porcentagem de
enraizamento), número de raízes, comprimento médio de raízes e massa seca
de raízes.
Como critério adotado no experimento para a contagem de raízes, toda
estaca que apresentava pelo menos uma raiz adventícia com comprimento
igual ou superior a 0,2cm foi considerada enraizada e estas raízes foram
anotadas e medidas. A medição do comprimento destas raízes foi feita com
régua, calculando a média por estaca.
Para a avaliação da massa seca das raízes, o material foi submetido à
secagem em estufa de aeração forçada, a 70°C, até que permanecesse em
massa constante, utilizando-se para a pesagem uma balança digital de
precisão (quatro dígitos decimais).
Para as análises estatísticas os dados de porcentagem de enraizamento
foram transformados em arc sen 100/x ; os de número de raízes em 50,0+x
e os de comprimento médio de raízes e peso da matéria seca de raízes não
foram transformados.
Os dados coletados, transformados quando necessários, foram
analisados estatisticamente, sendo realizada a análise de variância e, quando
cabível, o desdobramento dos graus de liberdade de concentrações de IBA,
para avaliar a regressão polinomial, a fim de se verificar o comportamento das
variáveis em função do aumento da concentração de IBA (BANZATTO e
KRONKA, 1992).
Etapa II: Determinação de glicosídeos cardioativos em mudas de Nerium
oleander L.
O trabalho foi conduzido a campo, instalado na Horta Experimental do
Campus III da Universidade para o Desenvolvimento do Estado e da Região do
Pantanal – UNIDERP, Campo Grande (MS), em Neossolo Quartzarênico. O

36
município de Campo Grande está localizado a latitude de 20º 26’ 34’’ e
longitude de 54º 38’ 47’’ a 532,1 m do nível do mar. O clima na região é tropical
úmido, com uma estação de chuvas no verão e outra de seca no inverno. A
temperatura média anual é de 26º C e o índice de precipitação chega a 1.500
mm por ano.
As análises cromatográficas foram realizadas no Laboratório de
Farmacologia também da UNIDERP.
O trabalho foi composto por três experimentos: (1) Efeito de diferentes
doses de nitrogênio na concentração de glicosídeos cardioativos; (2) Efeito de
diferentes doses de calcário na concentração de glicosídeos cardioativos; (3)
Efeito do estresse hídrico, verificado por meio da capacidade de campo, na
concentração de glicosídeos cardioativos.
As mudas de Nerium oleander L. variedade rosa, foram produzidas por
estaquia, cujas estacas, utilizadas nos três experimentos, foram obtidas de
matrizes localizadas no Campus III da UNIDERP.
As estacas foram cortadas, medindo aproximadamente 15 cm de
comprimento com um par de folhas e duas gemas e foram tratadas com ácido
indolbutírico, via pó, na concentração de 2000 mg.Kg-1 e estaqueadas, em
bandejas de isopor com 72 células, contendo vermiculita como substrato. As
estacas foram postas para enraizar no dia 04/02/2005. Imediatamente após o
estaqueamento, as bandejas foram colocadas em estufa com nebulizaçao
intermitente, até o enraizamento total das plantas, ou seja, aproximadamente
60 dias.
Após o enraizamento no dia 12/04/2005, as mudas foram transplantadas
para o campo, de acordo com cada experimento.
Para todos os experimentos o delineamento experimental utilizado foi o
de blocos casualizados.
As avaliações dos três experimentos foram realizadas aos 60 dias após
o transplante das mudas, cortando-se o caule rente ao solo com tesoura de
poda, sempre entre as 7:00 e 9:30 horas. Separou-se 4 folhas/planta da altura
mediana da planta para extração e determinação do teor de glicosídeos
cardioativos. Anotou-se a massa fresca da parte aérea pesando-se a cada
planta em balança analítica, em seguida, foram colocadas separadamente em
sacos de papel tipo Kraft, previamente identificados, para determinação da

37
massa seca da parte aérea, realizada após secagem em estufa de aeração
forçada.
3.1. Experimento I - Efeito de diferentes doses de nitrogênio no teor de
glicosídeos cardioativos
O experimento foi composto por quatro tratamentos (sem a presença de
adubação nitrogenada; 2,5 g. N-1; 5,0 g. N-1; 7,5 g. N-1), cinco repetições e
quatro plantas por parcela, totalizando 80 plantas.
O experimento foi conduzido em estufa de cultivo, coberta com filme
plástico de 150 micras e com as laterais protegidas por tela de sombreamento
30%. As mudas foram transplantas em sulcos, espaçadas com 2 m entre linhas
e 0,5 m entre plantas. Para cada tratamento realizou-se a adubação de base
com P e K, de acordo com análise de solo e recomendações de RIBEIRO et al.
(1999). Entre cada tratamento, respeitou-se uma distância de 0,5 m, como
margem de segurança para que não houvesse interferência de um tratamento
para o outro.
A aplicação da adubação nitrogenada (sulfato de amônia – 20% de N) foi
feita na cova de plantio no transplante das mudas. A quantidade de sulfato de
amônia indicada para cada tratamento, foi previamente misturada com
aproximadamente 1 L de solo e colocado ao fundo da cova, evitando-se desta
forma o contato das raízes com o adubo. O sistema de irrigação foi por
gotejamento.
3.2. Experimento II - Efeito de diferentes doses de calcário (saturação de
base) no teor de glicosídeos cardioativos
As mudas foram transplantadas para canteiros de 1,20 x 2,0 m a campo,
sendo a distância entre os canteiros de 0,5 m. A adubação química de base (N-
P-K) e a orgânica foram realizadas de acordo com análise de solo e
recomendações de RIBEIRO et al. (1999). A saturação de bases (V%) também
foi determinada após a análise de solo, sendo a recomendação de 2,8
toneladas por hectare. As diferentes quantidades de calcário aplicadas em
cada tratamento foram determinadas por meio da fórmula:

38
NC= (V2 – V1). CTC / 100, onde:
V1= 100. SB / T, onde:
SB = soma de bases (Ca + Mg + K + Na);
T ( CTC pH) = SB + (H + Al).
A saturação de bases foi atingida utilizando-se calcário filler (de reação
rápida).
O experimento constou de cinco tratamentos (testemunha; 25% da
saturação de bases recomendada - 168 g de calcário por canteiro; 50% da
saturação de bases recomendada - 336 g de calcário por canteiro; 75% da
saturação de bases recomendada - 504 g de calcário por canteiro; 100% da
saturação de bases recomendada - 672 g de calcário por canteiros) 4
repetições e 4 plantas por parcela, totalizando 80 plantas.
3.3. Experimento III - Efeito do estresse hídrico, verificado por meio da
capacidade de campo na concentração de glicosídeos cardioativos
O experimento foi composto por quatro tratamentos (25% da
capacidade de campo CC; 50% CC; 75% CC; 100% CC), cinco repetições e
quatro plantas por parcela, totalizando 80 plantas.
Conduziu-se o experimento em casa de vegetação, coberta com filme
plástico de 150 micras e com as laterais protegidas por tela de sombreamento
30%. As mudas foram transplantadas para vasos plásticos com capacidade
para 1 L, preenchidos 50% com composto orgânico (esterco de galinha) e 50%
com solo comum.
A capacidade de campo foi determinada previamente (CARVALHO et
al., 2003), por meio de curva de retenção de umidade, a partir do substrato
seco, peneirado e homogeneizado em laboratório.
O estabelecimento e controle dos níveis hídricos foram feitos por meio
de monitoramento diário da massa de uma amostra, constituída de dez vasos
por nível hídrico, incluindo o substrato e a planta. Previamente foi determinada
a massa referência, ou seja, a massa que cada vaso precisa ter, em cada nível
hídrico. Isso foi feito a partir dos dados da massa do vaso vazio, da massa do
vaso com substrato seco e na capacidade de campo (CARVALHO et al., 2003).
A cada dia após a pesagem dos vasos foi comparada essa massa com a

39
massa referência, foi calculada a diferença, completando-se a massa do vaso
com o volume de água correspondente.
3.4. Análise quantitativa de glicosídeos cardioativos
A análise quantitativa da glicosídeos cardioativos foi realizada por meio
da extração seletiva da glicosídeos cardioativos que, por sua vez, foi baseada
em proposta metodológica utilizada por SHARAPIN et al. (2000) para extração
de digoxina e digitoxina de Digitalis lanata considerando, também, as
características físico-químicas da glicosídeos cardioativos descritas pelo Index
Merck (MERCK, 2001).
Inicialmente, foi preparado o extrato metanólico por meio da extração
exaustiva do esgotamento da matéria fresca (3g da planta fragmentada com
faca em partículas menores do que 2 mm) com 60 mL de metanol PA, por
maceração sob agitação, seguido de eliminação do solvente extrator até
obtenção de 30 mL, sob exaustão em capela.
Buscando garantir o esgotamento, a matéria fresca foi extraída três
vezes consecutivas com metanol na proporção de 1:20, da seguinte maneira:
1ª maceração – agitação 15 minutos (agitador magnético), descanso 6 horas,
agitação 15 minutos (agitador magnético), descanso 24 horas, filtração; 2ª
maceração – agitação 15 minutos (agitador magnético), descanso 2 horas,
filtração; 3ª maceração – agitação 15 minutos (agitador magnético), descanso 2
horas, filtração.
Ao extrato metanólico proveniente da maceração foi adicionado 20 mL
de água destilada, originando um extrato hidrometanólico (50 mL), que foi
inicialmente, desengordurado com hexano por meio de partição líquido-líquido,
visando eliminar as substâncias lipofílicas indesejáveis (interferentes). Esta
extração foi realizada três vezes consecutivas com 40 mL de hexano PA.
Para extração da glicosídeos cardioativos, a fase hidrometanólica
desengordurada foi particionada com clorofórmio PA, também, por três vezes
consecutivas com 40 mL. Nesta etapa, adicionou-se cerca de 50 mL de água
destilada para favorecer a afinidade da glicosídeos cardioativos pelo
clorofórmio.

40
O teor de glicosídeos cardioativos foi obtido por meio da determinação
do peso da fase clorofórmica após eliminação total do solvente (sob exaustão
em capela) e da água residual (dessecador sob vácuo).

41
4. RESULTADOS E DISCUSSÃO
4.1. Enraizamento de estacas herbáceas de duas variedades de
espirradeira, Nerium oleander L., em diferentes concentrações de
IBA, no verão e no inverno
Na Tabela 1, são apresentados os resultados referentes à porcentagem
de enraizamento de estacas de espirradeira de flores rosas e brancas, no
verão e no inverno.
Observa-se que a interação entre os fatores foi significativa somente
entre as variedades e as estações do ano. A porcentagem de enraizamento foi
significativamente superior no verão onde ambas às variedades apresentaram
97% de enraizamento. No inverno, houve diferença significativa entre as duas
variedades sendo que a de flores rosas apresentou maior porcentagem de
enraizamento (77%) quando comparada com a de flores brancas (56%). Já no
verão, não houve diferença significativa entre as duas variedades.
FACHINELLO et al. (1995) explicam que esta superioridade do verão,
juntamente com a primavera, é em virtude de serem períodos de intenso
crescimento vegetativo, em que as estacas estão mais herbáceas, facilitando,
de modo geral, o desenvolvimento de raízes em espécies com dificuldade de
enraizamento. ONO e RODRIGUES (1996) explicam ainda, que no inverno, a
taxa fotossintética é baixa, em conseqüência da baixa luminosidade e
temperatura ambiente; assim, os metabólicos da fotossíntese são reduzidos,
sendo insuficientes para a iniciação e desenvolvimento das raízes. MARTINS
(1998) reforça que o verão e a primavera proporcionam maior capacidade às
plantas de enraizar. Maiores porcentagens de enraizamento obtidas no verão
e/ou na primavera em detrimento do inverno, também foram observadas para
outras espécies como oliveira, Olea europea L. (HARTMANN e LORETI, 1965),
azaléia, Rhododendron simsii Planch (FERNANDES et al., 1979), carambola,
Averrhoa carambola L. (BASTOS, 2002) e lichieira, Litchi chinensis
(CARVALHO et al., 2005).

42
Tabela 1. Quadrados médios e médias obtidos nas análises de variância para porcentagem de enraizamento em estacas de espirradeira (Nerium oleander L.), de flores branca e rosa, retiradas no verão e no inverno e tratadas com diferentes concentrações de IBA (mg.kg-1). Jaboticabal, SP, 2006.
CV GL Porcentagem de enraizamento1
Variedade (V) 1 684,70 **
Estação (E) 1 10510,55 **
IBA (I) 3 211,96 **
V x E 1 758,74 **
V x I 3 106, 64 NS
E x I 3 30,45 NS
V x E x I 3 106,41 NS
Blocos 3 84,61 NS
Resíduo 45 45,22
CV(%) 9,93
Regressões R2 (%)
Regressão linear 302,58 * 47,6
Regressão quadrática 198,68 * 78,8
Regressão cúbica 134,62 NS 100,0
Médias
Branca Rosa Média
Verão 80, 731(97,41) 2/aA 80,39 (97,21) aA 80,56 (97,31) a
Inverno 48,22 (55,61) bB 61,65 (77,45) bA 54,93 (66,99) b
Média 64,48 (81,44) B 71,02 (89,42) A
1/ Dados transformados em arc sen 100/x . 2/ Dados não transformados. NS - não significativo (P>0,05); * significativo (P< 0,05); ** significativo (P<0,01) Médias seguidas da mesma letra maiúscula na linha e minúscula na coluna, não diferem entre si pelo teste de Tukey (5%)
43
Alguns estudos (CORRÊA, 2001, CHALFUN, 1989; HESS, 1963,
OVERBEEK et al., 1946) têm mostrado que as variedades com flores coloridas
têm apresentado maior porcentagem de enraizamento ou enraizamento mais
rápido quando comparadas com as de flores brancas, como mostrou este
estudo na época de inverno.
Relacionada à aplicação de IBA, houve diferença entre as
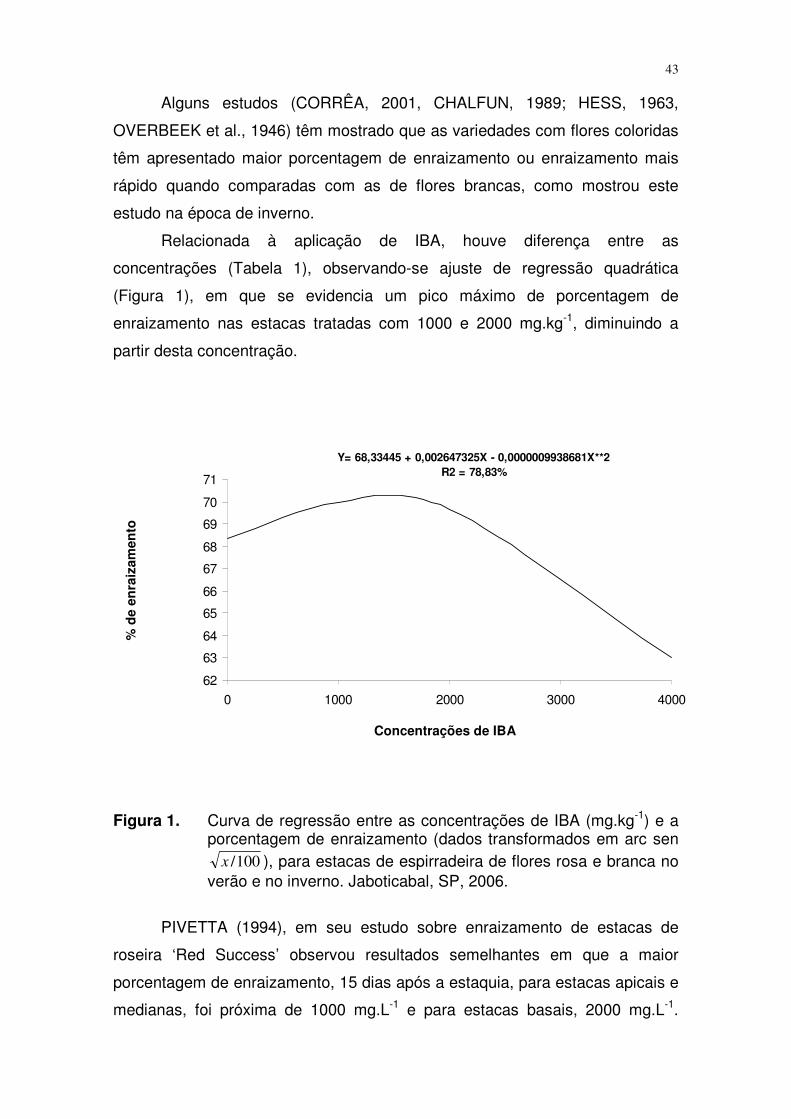
concentrações (Tabela 1), observando-se ajuste de regressão quadrática
(Figura 1), em que se evidencia um pico máximo de porcentagem de
enraizamento nas estacas tratadas com 1000 e 2000 mg.kg-1, diminuindo a
partir desta concentração.
Y= 68,33445 + 0,002647325X - 0,0000009938681X**2R2 = 78,83%
62
63
64
65
66
67
68
69
70
71
0 1000 2000 3000 4000
Figura 1. Curva de regressão entre as concentrações de IBA (mg.kg-1) e a porcentagem de enraizamento (dados transformados em arc sen
100/x ), para estacas de espirradeira de flores rosa e branca no verão e no inverno. Jaboticabal, SP, 2006.
PIVETTA (1994), em seu estudo sobre enraizamento de estacas de
roseira ‘Red Success’ observou resultados semelhantes em que a maior
porcentagem de enraizamento, 15 dias após a estaquia, para estacas apicais e
medianas, foi próxima de 1000 mg.L-1 e para estacas basais, 2000 mg.L-1.
% d
e en
raiz
amen
to
Concentrações de IBA

44
MARTINS (1998) também obteve curva semelhante quando tratou estacas de
lichieira ‘Bengal’ com IBA.
Igualmente, BASTOS (2002) verificou ajuste de regressão quadrática
para porcentagem de enraizamento de estacas de caramboleira, com o
aumento da concentração de IBA.
Na Tabela 2, são apresentados os resultados referentes ao número de
raízes por estaca. Semelhantemente ao que ocorreu para porcentagem de
enraizamento, a interação entre os fatores foi significativa somente entre as
variedades e as estações do ano. O número de raízes por estaca também foi
significativamente superior no verão, para ambas as variedades e, também, as
estacas de espirradeira de flores rosas tiveram número de raízes
significativamente superior quando comparadas com as de flores brancas,
porém, esta diferença foi verificada no verão.
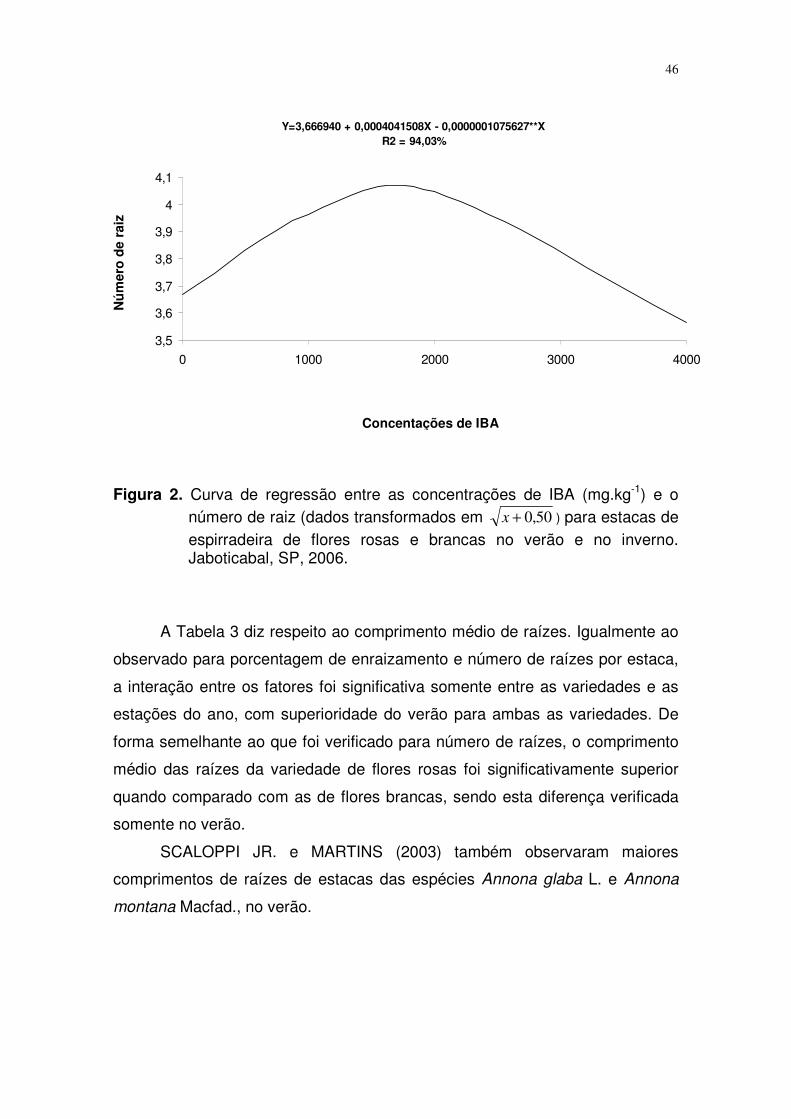
No que diz respeito à aplicação de IBA, observa-se (Tabela 2 e Figura 2)
ajuste de regressão quadrática, onde o número de raízes é crescente até a
concentração de 2000 mg.K-1, decrescendo a partir deste ponto.
Vários autores também observaram aumento do número de raízes em
função da utilização de IBA, para diferentes espécies, entre eles PIVETTA
(1994), QUINTANILHA (1995), MARTINS (1998), BASTOS (2002) e
CARVALHO et al. (2005).

45
Tabela 2. Quadrados médios e médias obtidos nas análises de variância para número de raízes em estacas de espirradeira (Nerium oleander L.), de flores branca e rosa, retiradas no verão e no inverno e tratadas com diferentes concentrações de IBA (mg.kg-1). Jaboticabal, SP, 2006.
CV GL Número médio de raízes1
Variedade (V) 1 0, 20 NS
Estação (E) 1 9,56 **
IBA (I) 3 0,68 **
V x E 1 0,40 *
V x I 3 0,10 NS
E x I 3 0,07 NS
V x E x I 3 0,15 NS
Blocos 3 0,12 NS
Resíduo 45 0,06
CV(%) 6,21
Regressões R2 (%)
Regressão linear 0,24 NS 8,8
Regressão quadrática 2,33 ** 94,0
Regressão cúbica 0,16 NS 100,0
Médias
Branca Rosa Média
Verão 4,081(16,15) 2/ aB 4,35 (18,42) aA 4,22 (17,31) a
Inverno 3,47 (11,54) bA 3,42 (11,20) bA 3,45 (11,40) b
Média 3,78 (13,79) A 3,89 (14,63) A
1/ Dados não transformados em 50,0+x . 2/ Dados não transformados. NS - não significativo (P>0,05); * significativo (P< 0,05); ** significativo (P<0,01) Médias seguidas da mesma letra maiúscula na linha e minúscula na coluna, não diferem entre si pelo teste de Tukey (5%)
46
Y=3,666940 + 0,0004041508X - 0,0000001075627**XR2 = 94,03%
3,5
3,6
3,7
3,8
3,9
4
4,1
0 1000 2000 3000 4000
Figura 2. Curva de regressão entre as concentrações de IBA (mg.kg-1) e o
número de raiz (dados transformados em 50,0+x ) para estacas de espirradeira de flores rosas e brancas no verão e no inverno. Jaboticabal, SP, 2006.
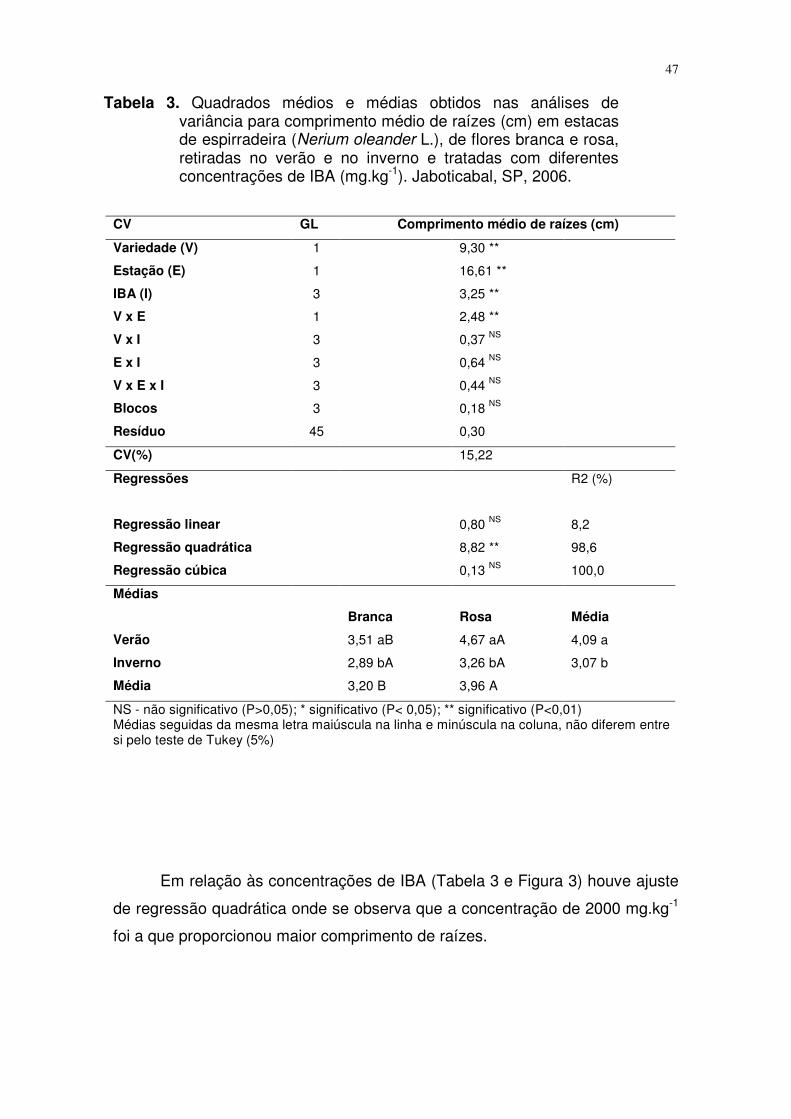
A Tabela 3 diz respeito ao comprimento médio de raízes. Igualmente ao
observado para porcentagem de enraizamento e número de raízes por estaca,
a interação entre os fatores foi significativa somente entre as variedades e as
estações do ano, com superioridade do verão para ambas as variedades. De
forma semelhante ao que foi verificado para número de raízes, o comprimento
médio das raízes da variedade de flores rosas foi significativamente superior
quando comparado com as de flores brancas, sendo esta diferença verificada
somente no verão.
SCALOPPI JR. e MARTINS (2003) também observaram maiores
comprimentos de raízes de estacas das espécies Annona glaba L. e Annona
montana Macfad., no verão.
Núm
ero
de r
aiz
Concentações de IBA
47
Tabela 3. Quadrados médios e médias obtidos nas análises de variância para comprimento médio de raízes (cm) em estacas de espirradeira (Nerium oleander L.), de flores branca e rosa, retiradas no verão e no inverno e tratadas com diferentes concentrações de IBA (mg.kg-1). Jaboticabal, SP, 2006.
CV GL Comprimento médio de raízes (cm)
Variedade (V) 1 9,30 **
Estação (E) 1 16,61 **
IBA (I) 3 3,25 **
V x E 1 2,48 **
V x I 3 0,37 NS
E x I 3 0,64 NS
V x E x I 3 0,44 NS
Blocos 3 0,18 NS
Resíduo 45 0,30
CV(%) 15,22
Regressões R2 (%)
Regressão linear 0,80 NS 8,2
Regressão quadrática 8,82 ** 98,6
Regressão cúbica 0,13 NS 100,0
Médias
Branca Rosa Média
Verão 3,51 aB 4,67 aA 4,09 a
Inverno 2,89 bA 3,26 bA 3,07 b
Média 3,20 B 3,96 A
NS - não significativo (P>0,05); * significativo (P< 0,05); ** significativo (P<0,01) Médias seguidas da mesma letra maiúscula na linha e minúscula na coluna, não diferem entre si pelo teste de Tukey (5%)
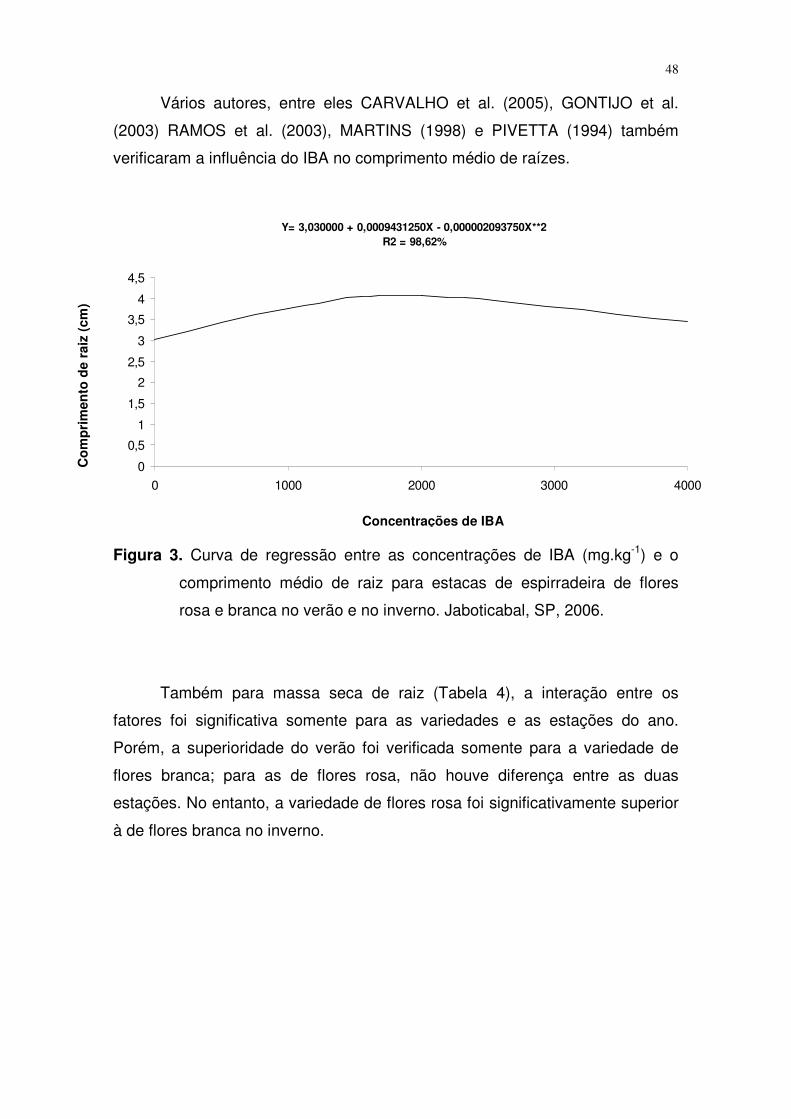
Em relação às concentrações de IBA (Tabela 3 e Figura 3) houve ajuste
de regressão quadrática onde se observa que a concentração de 2000 mg.kg-1
foi a que proporcionou maior comprimento de raízes.
48
Vários autores, entre eles CARVALHO et al. (2005), GONTIJO et al.
(2003) RAMOS et al. (2003), MARTINS (1998) e PIVETTA (1994) também
verificaram a influência do IBA no comprimento médio de raízes.
Y= 3,030000 + 0,0009431250X - 0,000002093750X**2R2 = 98,62%
0
0,5
1
1,5
2
2,5
3
3,5
4
4,5
0 1000 2000 3000 4000
Figura 3. Curva de regressão entre as concentrações de IBA (mg.kg-1) e o
comprimento médio de raiz para estacas de espirradeira de flores
rosa e branca no verão e no inverno. Jaboticabal, SP, 2006.
Também para massa seca de raiz (Tabela 4), a interação entre os
fatores foi significativa somente para as variedades e as estações do ano.
Porém, a superioridade do verão foi verificada somente para a variedade de
flores branca; para as de flores rosa, não houve diferença entre as duas
estações. No entanto, a variedade de flores rosa foi significativamente superior
à de flores branca no inverno.
Com
prim
ento
de
raiz
(cm
)
Concentrações de IBA
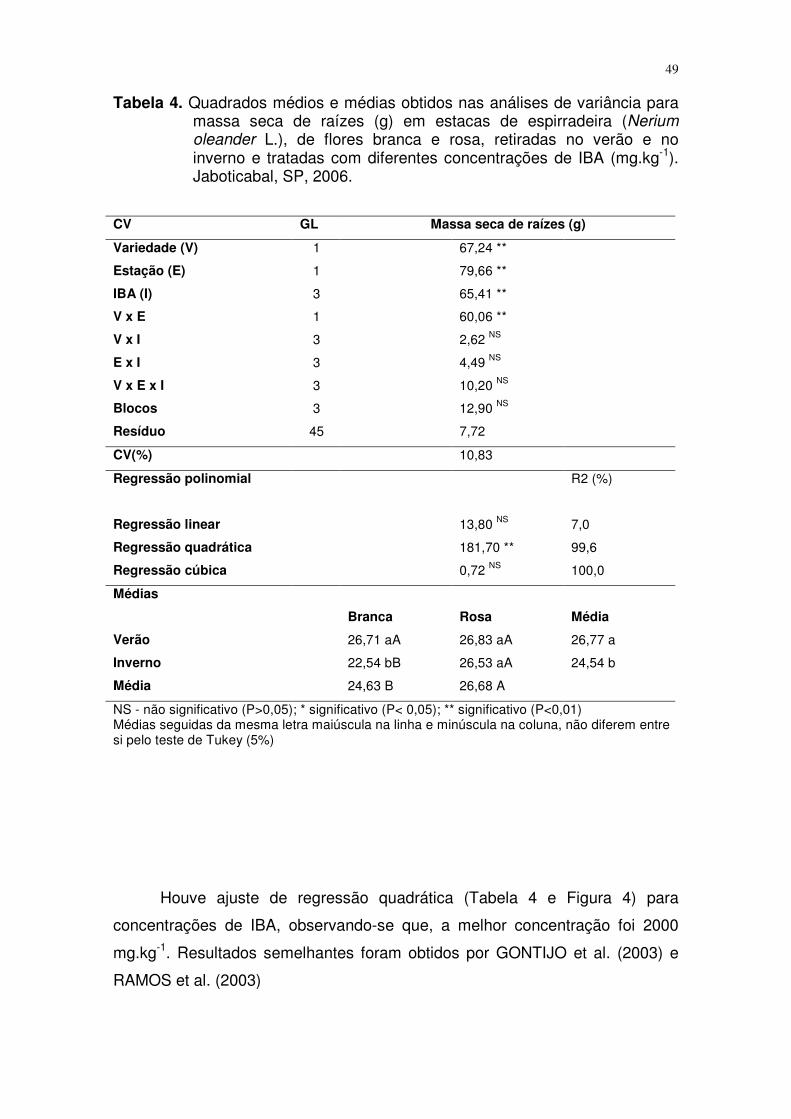
49
Tabela 4. Quadrados médios e médias obtidos nas análises de variância para massa seca de raízes (g) em estacas de espirradeira (Nerium oleander L.), de flores branca e rosa, retiradas no verão e no inverno e tratadas com diferentes concentrações de IBA (mg.kg-1). Jaboticabal, SP, 2006.
CV GL Massa seca de raízes (g)
Variedade (V) 1 67,24 **
Estação (E) 1 79,66 **
IBA (I) 3 65,41 **
V x E 1 60,06 **
V x I 3 2,62 NS
E x I 3 4,49 NS
V x E x I 3 10,20 NS
Blocos 3 12,90 NS
Resíduo 45 7,72
CV(%) 10,83
Regressão polinomial R2 (%)
Regressão linear 13,80 NS 7,0
Regressão quadrática 181,70 ** 99,6
Regressão cúbica 0,72 NS 100,0
Médias
Branca Rosa Média
Verão 26,71 aA 26,83 aA 26,77 a
Inverno 22,54 bB 26,53 aA 24,54 b
Média 24,63 B 26,68 A
NS - não significativo (P>0,05); * significativo (P< 0,05); ** significativo (P<0,01) Médias seguidas da mesma letra maiúscula na linha e minúscula na coluna, não diferem entre si pelo teste de Tukey (5%)
Houve ajuste de regressão quadrática (Tabela 4 e Figura 4) para
concentrações de IBA, observando-se que, a melhor concentração foi 2000
mg.kg-1. Resultados semelhantes foram obtidos por GONTIJO et al. (2003) e
RAMOS et al. (2003)
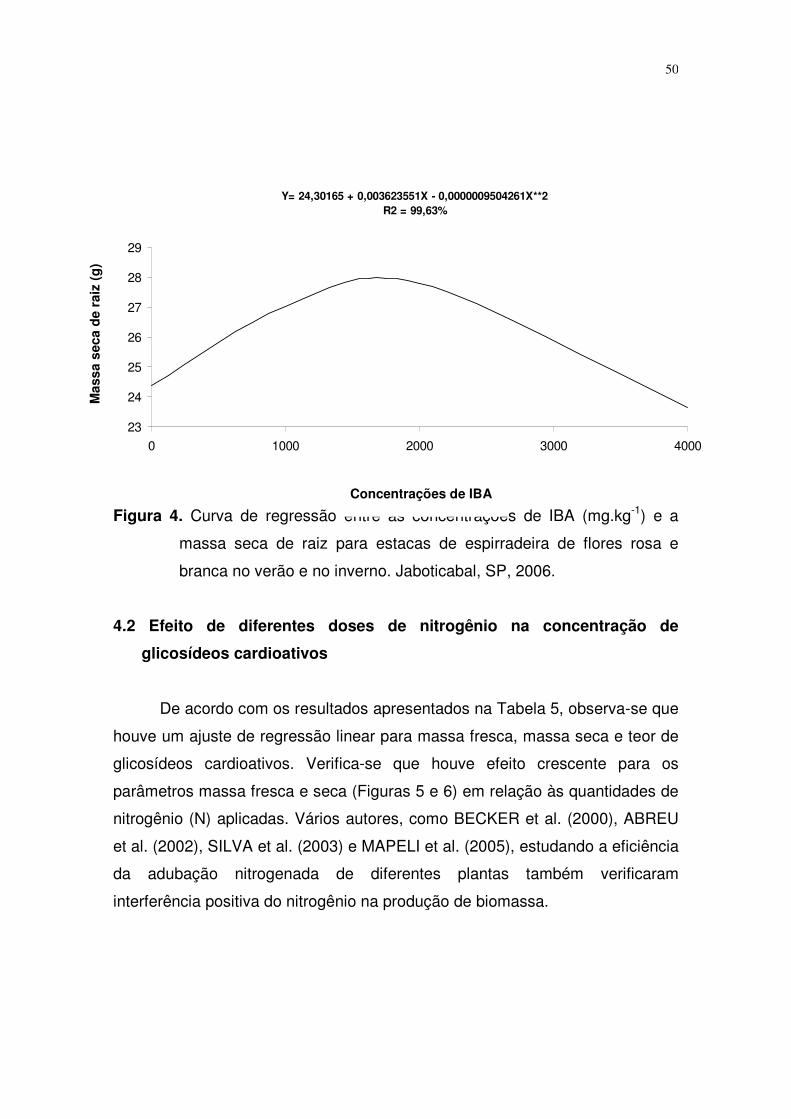
50
Y= 24,30165 + 0,003623551X - 0,0000009504261X**2R2 = 99,63%
23
24
25
26
27
28
29
0 1000 2000 3000 4000
Figura 4. Curva de regressão entre as concentrações de IBA (mg.kg-1) e a
massa seca de raiz para estacas de espirradeira de flores rosa e
branca no verão e no inverno. Jaboticabal, SP, 2006.
4.2 Efeito de diferentes doses de nitrogênio na concentração de
glicosídeos cardioativos
De acordo com os resultados apresentados na Tabela 5, observa-se que
houve um ajuste de regressão linear para massa fresca, massa seca e teor de
glicosídeos cardioativos. Verifica-se que houve efeito crescente para os
parâmetros massa fresca e seca (Figuras 5 e 6) em relação às quantidades de
nitrogênio (N) aplicadas. Vários autores, como BECKER et al. (2000), ABREU
et al. (2002), SILVA et al. (2003) e MAPELI et al. (2005), estudando a eficiência
da adubação nitrogenada de diferentes plantas também verificaram
interferência positiva do nitrogênio na produção de biomassa.
Mas
sa s
eca
de r
aiz
(g)
Concentrações de IBA
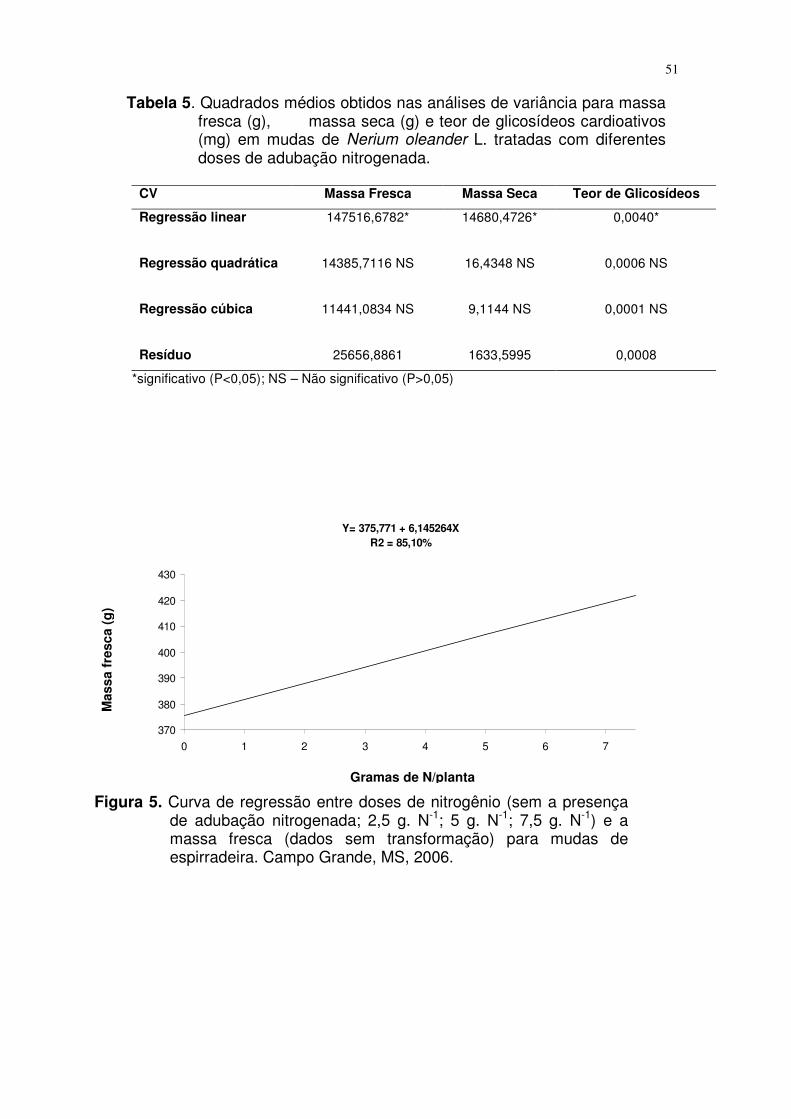
51
Tabela 5. Quadrados médios obtidos nas análises de variância para massa fresca (g), massa seca (g) e teor de glicosídeos cardioativos (mg) em mudas de Nerium oleander L. tratadas com diferentes doses de adubação nitrogenada.
CV Massa Fresca Massa Seca Teor de Glicosídeos
Regressão linear 147516,6782* 14680,4726* 0,0040*
Regressão quadrática 14385,7116 NS 16,4348 NS 0,0006 NS
Regressão cúbica 11441,0834 NS 9,1144 NS 0,0001 NS
Resíduo 25656,8861 1633,5995 0,0008
*significativo (P<0,05); NS – Não significativo (P>0,05)
Y= 375,771 + 6,145264XR2 = 85,10%
370
380
390
400
410
420
430
0 1 2 3 4 5 6 7
Figura 5. Curva de regressão entre doses de nitrogênio (sem a presença
de adubação nitrogenada; 2,5 g. N-1; 5 g. N-1; 7,5 g. N-1) e a massa fresca (dados sem transformação) para mudas de espirradeira. Campo Grande, MS, 2006.
Mas
sa fr
esca
(g)
Gramas de N/planta
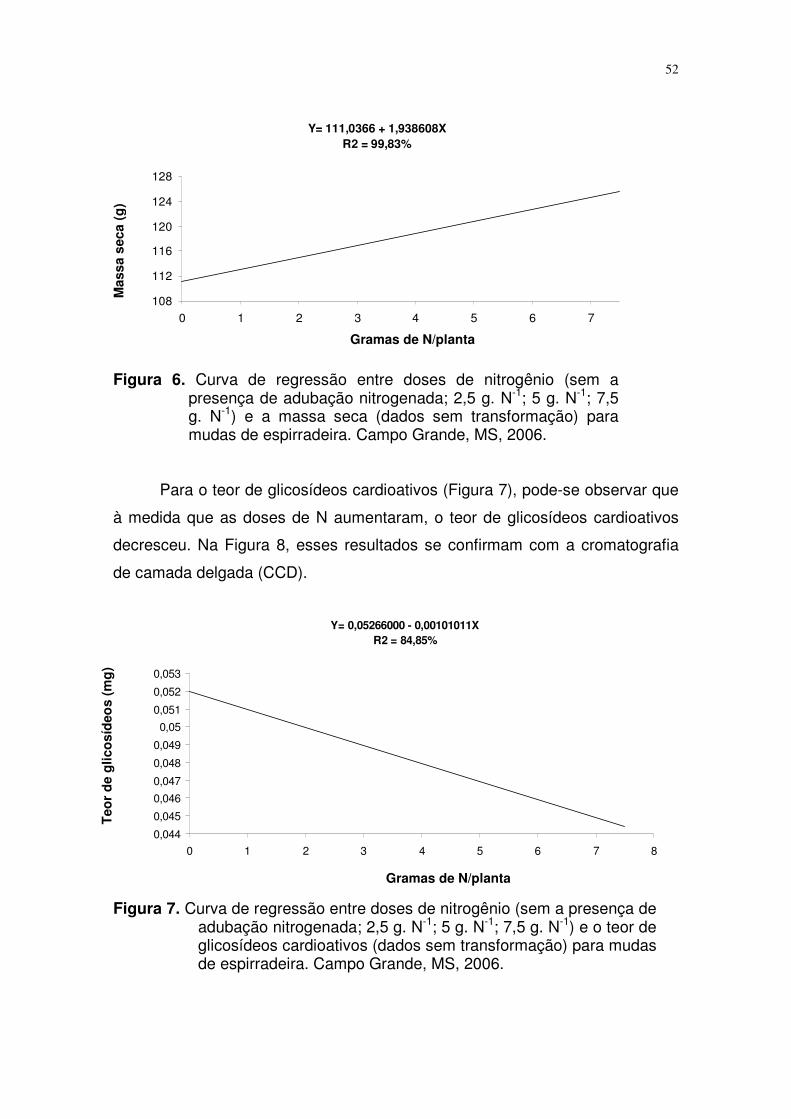
52
Y= 111,0366 + 1,938608XR2 = 99,83%
108
112
116
120
124
128
0 1 2 3 4 5 6 7
Figura 6. Curva de regressão entre doses de nitrogênio (sem a
presença de adubação nitrogenada; 2,5 g. N-1; 5 g. N-1; 7,5 g. N-1) e a massa seca (dados sem transformação) para mudas de espirradeira. Campo Grande, MS, 2006.
Para o teor de glicosídeos cardioativos (Figura 7), pode-se observar que
à medida que as doses de N aumentaram, o teor de glicosídeos cardioativos
decresceu. Na Figura 8, esses resultados se confirmam com a cromatografia
de camada delgada (CCD).
Y= 0,05266000 - 0,00101011XR2 = 84,85%
0,044
0,045
0,0460,047
0,048
0,049
0,050,051
0,052
0,053
0 1 2 3 4 5 6 7 8
Figura 7. Curva de regressão entre doses de nitrogênio (sem a presença de
adubação nitrogenada; 2,5 g. N-1; 5 g. N-1; 7,5 g. N-1) e o teor de glicosídeos cardioativos (dados sem transformação) para mudas de espirradeira. Campo Grande, MS, 2006.
Gramas de N/planta
Mas
sa s
eca
(g)
Teor
de
glic
osíd
eos
(mg)
Gramas de N/planta

53
Figura 8. Perfil cromatográfico obtido em cromatografia em camada delgada de sílica - gel das amostras de glicosídeos cardioativos de plantas de Nerium oleander L obtidas de quatro tratamentos (sem a presença de adubação nitrogenada; 2,5 g. N-1; 5 g. N-1; 7,5 g. N-1).
A adubação nitrogenada, portanto, mostrou ser uma prática muito
importante para a Nerium oleander L., sendo benéfica no desenvolvimento da
planta e reduzindo o teor da substância tóxica.
Também MING (1992), estudando a adubação orgânica em erva-cidreira
(Lippia alba) verificou maiores produções de biomassa em níveis crescentes de
matéria orgânica, entretanto observou uma relação inversa, quanto aos teores
de óleo essencial. O autor comenta o papel de defesa dos óleos essenciais em
ambientes desfavoráveis, enquanto em ambientes mais favoráveis, a energia
da planta estaria voltada para atividades de crescimento e desenvolvimento, do
metabolismo primário.
Contrariamente ao resultado obtido neste trabalho, o aumento da
concentração de nitrogênio aumentou o teor de metabólitos secundários em
Mimosa sp. (NOWACKI et al., 1975), em Tabernaemontana pachysiphon
(HOFT et al. 1996) e em quebra-pedra, Phyllanthus niruri (BECKER et al.,
2000).
Já MAPELI et al. (2005), estudando o efeito do nitrogênio e do fósforo no
teor de óleo essencial, em camomila (Chamomilla recutita L.), verificaram que
as adubações não influenciaram na produção de óleo essencial.
Zero
2,5 g. N-1
5 g. N-1
7,5 g. N-1
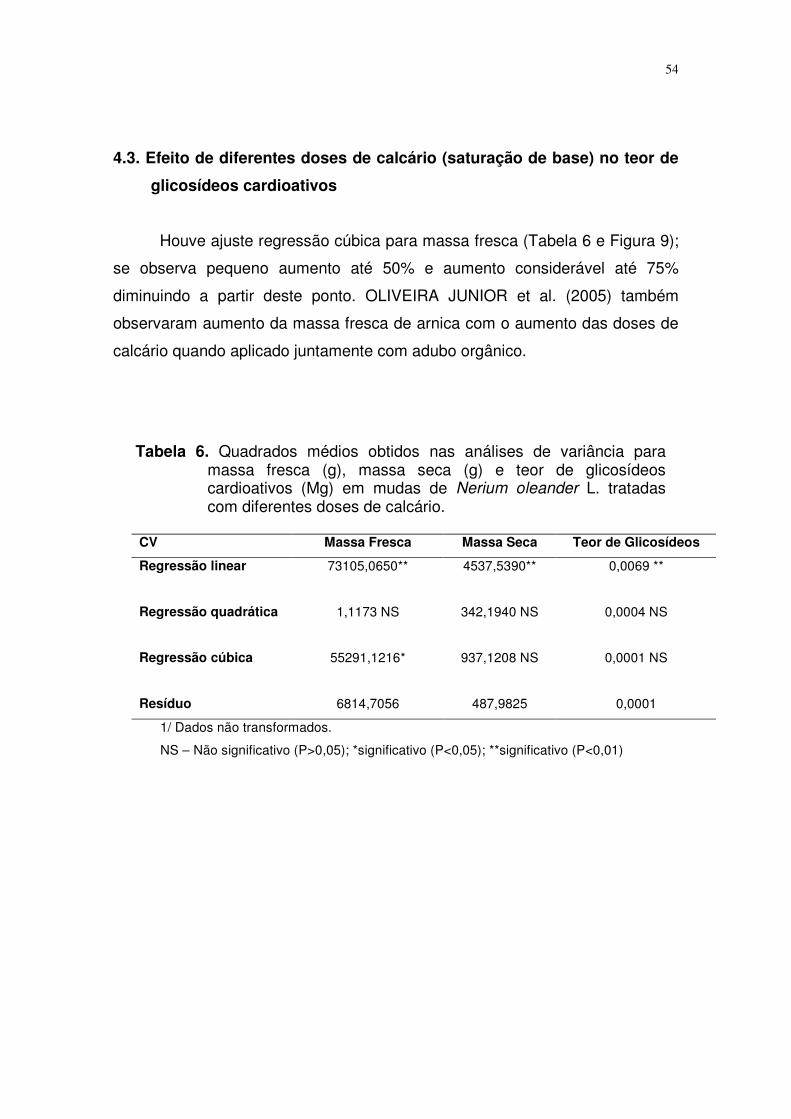
54
4.3. Efeito de diferentes doses de calcário (saturação de base) no teor de
glicosídeos cardioativos
Houve ajuste regressão cúbica para massa fresca (Tabela 6 e Figura 9);
se observa pequeno aumento até 50% e aumento considerável até 75%
diminuindo a partir deste ponto. OLIVEIRA JUNIOR et al. (2005) também
observaram aumento da massa fresca de arnica com o aumento das doses de
calcário quando aplicado juntamente com adubo orgânico.
Tabela 6. Quadrados médios obtidos nas análises de variância para
massa fresca (g), massa seca (g) e teor de glicosídeos cardioativos (Mg) em mudas de Nerium oleander L. tratadas com diferentes doses de calcário.
CV Massa Fresca Massa Seca Teor de Glicosídeos
Regressão linear 73105,0650** 4537,5390** 0,0069 **
Regressão quadrática 1,1173 NS 342,1940 NS 0,0004 NS
Regressão cúbica 55291,1216* 937,1208 NS 0,0001 NS
Resíduo 6814,7056 487,9825 0,0001
1/ Dados não transformados.
NS – Não significativo (P>0,05); *significativo (P<0,05); **significativo (P<0,01)

55
Y= 390,8600 + 8,970550X + 0,2976580X**2 - 0,001982880X**3 R2 =96,58
0
100
200
300
400
500
600
0 25 50 75 100
Figura 9. Curva de regressão entre doses de calcário (zero,
25%, 50%, 75% e 100%) e massa fresca (dados sem transformação) para mudas de espirradeira. Campo Grande, MS, 2006.
Para massa seca, houve um ajuste de regressão linear positiva (Tabela
6 e Figura 10).
Também BECKER et al. (2000), estudando o efeito da calagem na
produção de alcalóides totais em quebra-pedra Phyllanthus niruri L.,
observaram aumento de massa seca da parte aérea à medida que
aumentaram as doses de calcário aplicadas.
Mas
sa fr
esca
(g)
Saturação de bases (V%)
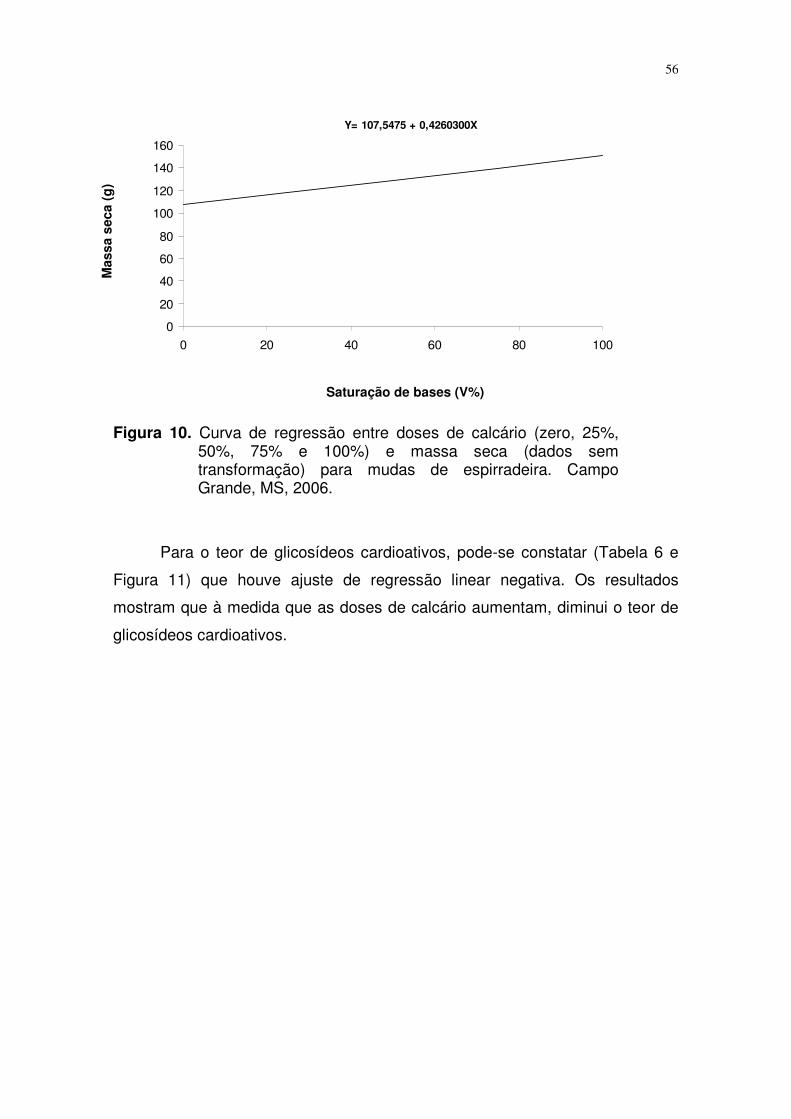
56
Y= 107,5475 + 0,4260300X
0
20
40
60
80
100
120
140
160
0 20 40 60 80 100
Figura 10. Curva de regressão entre doses de calcário (zero, 25%, 50%, 75% e 100%) e massa seca (dados sem transformação) para mudas de espirradeira. Campo Grande, MS, 2006.
Para o teor de glicosídeos cardioativos, pode-se constatar (Tabela 6 e
Figura 11) que houve ajuste de regressão linear negativa. Os resultados
mostram que à medida que as doses de calcário aumentam, diminui o teor de
glicosídeos cardioativos.
Mas
sa s
eca
(g)
Saturação de bases (V%)
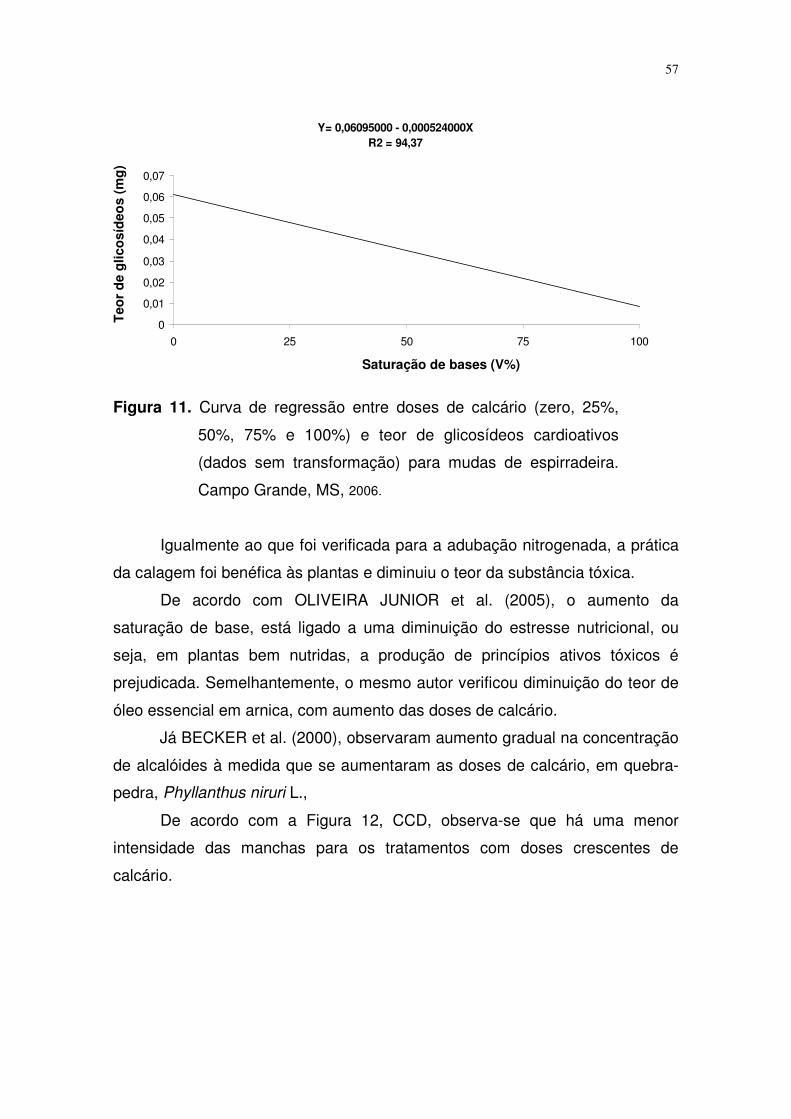
57
Y= 0,06095000 - 0,000524000XR2 = 94,37
0
0,01
0,02
0,03
0,04
0,05
0,06
0,07
0 25 50 75 100
Figura 11. Curva de regressão entre doses de calcário (zero, 25%,
50%, 75% e 100%) e teor de glicosídeos cardioativos
(dados sem transformação) para mudas de espirradeira.
Campo Grande, MS, 2006.
Igualmente ao que foi verificada para a adubação nitrogenada, a prática
da calagem foi benéfica às plantas e diminuiu o teor da substância tóxica.
De acordo com OLIVEIRA JUNIOR et al. (2005), o aumento da
saturação de base, está ligado a uma diminuição do estresse nutricional, ou
seja, em plantas bem nutridas, a produção de princípios ativos tóxicos é
prejudicada. Semelhantemente, o mesmo autor verificou diminuição do teor de
óleo essencial em arnica, com aumento das doses de calcário.
Já BECKER et al. (2000), observaram aumento gradual na concentração
de alcalóides à medida que se aumentaram as doses de calcário, em quebra-
pedra, Phyllanthus niruri L.,
De acordo com a Figura 12, CCD, observa-se que há uma menor
intensidade das manchas para os tratamentos com doses crescentes de
calcário.
Teor
de
glic
osíd
eos
(mg)
Saturação de bases (V%)

58
Figura 12. Perfil cromatográfico obtido em cromatografia em camada delgada de Sílica-gel das amostras de glicosídeos cardioativos de plantas de Nerium oleander L. entre doses de calcário (zero, 25%, 50%, 75% e 100%). Campo Grande, MS, 2006.
4.4. Efeito do estresse hídrico, verificado por meio da capacidade de
campo no teor de glicosídeos cardioativos.
Pelos resultados obtidos na Tabela 7, observa-se que houve ajuste de
regressão cúbica tanto para massa fresca, como massa seca em plantas de
Nerium oleander L. quando submetidas a diferentes quantidades de água. Nas
Figuras 13 e 14, observa-se que o aumento da quantidade de água no solo
promove aumento da massa fresca e seca das plantas até 75% da capacidade
de campo, decrescendo a partir deste ponto.
DHRU et al. (1991), também verificaram que o déficit hídrico reduziu a
produção de biomassa de Nerium oleander L. e Urginea indica.
Resultados semelhantes também foram descritos por CARVALHO et al.
(2003), que concluíram que quanto maior a disponibilidade de água no solo,
maior a massa fresca e seca das plantas de artemísia.
Zero
25%
50%
75%
100%

59
Tabela 7. Quadrados médios obtidos nas análises de variância para massa fresca (g), massa seca (g) e teor de glicosídeos cardioativos (Mg) em mudas de Nerium oleander L. cultivadas com diferentes quantidades de água.
CV Massa Fresca Massa Seca Teor de Glicosídeos
Regressão linear 171,7803** 24,6661** 3,2070**
Regressão quadrática 34,9140* 5,5704* 0,2192
Regressão cúbica 75,3337** 13,4286** 0,1759
Resíduo 4,7336 0,7995 0,0740
NS – Não significativo (P>0,05); *significativo (P<0,05); **significativo (P<0,01)
Y= 26,22800 - 1,563533X + 0,03260400X**2 - 0,0001851627X**3R2 = 100%
0
2
4
6
8
10
12
14
16
25 40 55 70 85 100
Figura 13. Curva de regressão entre capacidade de campo (25%,
50%, 75% e 100%) e massa fresca (dados sem transformação) para mudas de espirradeira. Campo Grande, MS, 2006.
Mas
sa fr
esca
(g)
% Capacidade de Campo

60
Y= 11,40600 - 0,6706800X + 0,01381360X**2 - 0,00007817600X**3R2 = 100%
0
1
2
3
4
5
6
7
25 40 55 70 85 100
Figura 14 Curva de regressão entre capacidade de campo (25%, 50%,
75% e 100%) e massa seca (dados sem transformação) para mudas de espirradeira. Campo Grande, MS, 2006
TAIZ e ZEIGER (2004) relatam que a área foliar é consideravelmente
aumentada em plantas, cuja capacidade de campo do solo é próxima a 100%.
STEPONKUS (1990) afirma que a deficiência de água altera vários
processos bioquímicos e fisiológicos, e induz respostas metabólicas e
fisiológicas como o fechamento estomático, declínio na taxa de crescimento,
acúmulo de solutos e ajusta-se fisiologicamente ao estado de menor
disponibilidade de água.
SANTOS et al. (2004) estudando o efeito do estresse hídrico na
produção de massa foliar da espécie Hyptis pectinata L., observaram que
quanto menor o estresse hídrico, maior a massa foliar.
Para o teor de glicosídeos cardioativos constata-se que houve ajuste de
regressão linear positiva (Tabela 7 e Figura 15), ou seja, quanto maior
quantidade de água fornecida para a planta, maior o teor de glicosídeos. Esses
resultados são confirmados pela Figura 16, onde por meio da CCD pode-se
verificar claramente que os teores de glicosídeos cardioativos aumentam
gradativamente com o aumento da capacidade de campo, apresentando
manchas mais intensas.
Mas
sa s
eca
(g)
% Capacidade de Campo
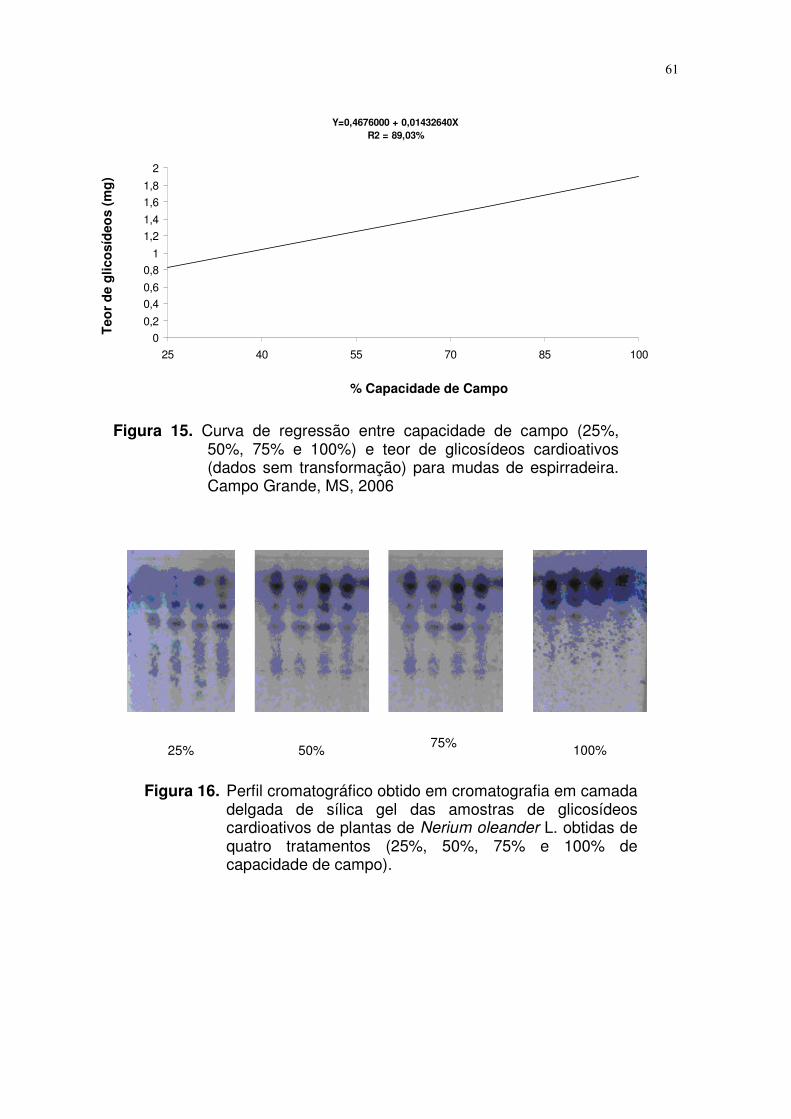
61
Y=0,4676000 + 0,01432640XR2 = 89,03%
00,20,40,60,8
11,21,41,61,8
2
25 40 55 70 85 100
Figura 15. Curva de regressão entre capacidade de campo (25%,
50%, 75% e 100%) e teor de glicosídeos cardioativos (dados sem transformação) para mudas de espirradeira. Campo Grande, MS, 2006
25%
50%
75%
100%
Figura 16. Perfil cromatográfico obtido em cromatografia em camada delgada de sílica gel das amostras de glicosídeos cardioativos de plantas de Nerium oleander L. obtidas de quatro tratamentos (25%, 50%, 75% e 100% de capacidade de campo).
Teor
de
glic
osíd
eos
(mg)
% Capacidade de Campo

62
TAIZ e ZEIGER (2004) explicam que o estresse hídrico reduz, tanto a
fotossíntese quanto o consumo de assimilados nas folhas em expansão. Como
conseqüência o estresse hídrico diminui indiretamente a quantidade de
fotossintatos exportados.
EGERT e TEVINI (2002), não verificaram alterações significativas nos
teores de clorofila, proteínas e compostos antioxidantes quando expuseram a
condimentar Allium schoenoprosum à seca.
4.5. Considerações gerais
É fato que a espirradeira, Nerium oleander L. e várias outras plantas
tóxicas são amplamente utilizadas no meio urbano, seja em jardins, na
arborização urbana ou em ambientes internos. Desta forma, além do trabalho
de conscientização da população, o uso de práticas fitotécnicas que possam
diminuir o teor das substâncias tóxicas possibilitarão um convívio com menos
riscos; no entanto, as pesquisas ainda são muito incipientes.
Este trabalho possibilitou verificar que a adubação nitrogenada e a
calagem, além de contribuírem para o melhor desenvolvimento das mudas,
reduziram o teor de glicosídeos cardioativos e, também, que o manejo da água
é muito importante, devendo ser fornecido somente o necessário para o
desenvolvimento da planta.

63
5. CONCLUSÕES
O enraizamento das variedades de Nerium oleander L. definidas pela
coloração da flor, ou seja, rosa e branca foi superior no verão, ou seja, 97%
para ambas as variedades quando comparado com o inverno (77% e 56%,
respectivamente para as variedades de flores rosa e branca)
A variedade de flores rosa apresentou maior porcentagem de
enraizamento e massa seca de raiz no inverno e maior número e comprimento
de raiz no verão.
O acido indolbutírico foi efetivo para aumentar a porcentagem de
enraizamento nas concentrações de 1000 e 2000 mg.L-1 e número,
comprimento e massa seca de raiz na concentração de 2000 mg.L-1.
A adubação nitrogenada e a calagem aumentaram a produção de
biomassa em plantas de Nerium oleander L. e diminuíram no teor de
glicosídeos cardioativos produzido pelas plantas.
O aumento da quantidade de água no solo até 75% da capacidade de
campo aumentou a produção de biomassa em plantas de Nerium oleander L. e
também o teor de glicosídeos cardioativos produzido pelas plantas.

64
6. REFERÊNCIAS BIBLIOGRÁFICAS
ABREU, I.N.; PINTO, J.E.B.P.; FURTINI NETO, A.E.; BERTOLLUCCI, S.K.V.;
LADEIRA, A.; GEROMEL, C. Nitrogênio e fósforo na produção vegetal e na
indução de mucilagem em plantas de insulina. Horticultura Brasileira,
Brasília, v.20, n.4, p.536-540, 2002.
BANZATTO, D. A.; KRONKA, S. N. Experimentação Agrícola. 2.ed.
Jaboticabal: FUNEP, 1992. 247p.
BARCELLOS, D.C. Plantas ornamentais tóxicas: estado e arte. 1998.
Disponível em: < http//www.plantastoxicas.hpg.ig.com.br/estado_arte.htm>.
Acesso em: 12 fev. 2006.
BASTOS, D.C. Efeito da época de coleta, estádio do ramo e do tratamento com
IBA no enraizamento de estacas de caramboleira (Averrhoa carambola L.).
Dissertação (Mestrado em Agronomia) – Faculdade de Ciências Agrárias e
Veterinárias, Universidade Estadual Paulista, Jaboticabal, 2002.
BECKER, L.; FURTINI NETO, A.E.; PINTO, J.E.B.P.; CARDOSO, M.G.;
SANTOS, C.D.;
BARBOSA, J.M.; LAMEIRA, O.A.; SANTIAGO, E.J.A. Crescimento e produção
de alcalóides totais de quebra-pedra em função da calagem e da adubação
nitrogenada. Horticultura Brasileira, Brasília, v.18, n.2, p.100-104, 2000.
CARVALHO, C.M.; CUNHA, R.J.P.; RODRIGUES, J.D. Enraizamento de
estacas semilenhosas de lichieira utilizando ácido indolbutírico. Revista
Brasileira de Fruticultura, Jaboticabal, SP. v.27, n.1, p. 95-97, 2005.
CARVALHO, L.M.; CASALI, V.W.D.; SOUZA, M.A.; CECON, P.R.
Disponibilidade de água no solo e crescimento de artemísia. Horticultura
brasileira. Brasília. v.21, n.4, p.726-730, 2003.

65
CASTRO, G.H.; FERREIRA, A. F.; SELVA, H.J.D.; MOSQUIM,R.P.
Contribuição ao estudo de plantas medicinais: metabólitos secundários.
2. ed. Viçosa: Gráfica Suprema. 2004. 113p.
CHALFUN, N.N.J. Fatores bioquímicos e fisiológicos no enraizamento de
estacas de Hibiscus rosa-sinensis L. 1989. 85f. Tese (Doutorado em
Fitotecnia) - Universidade Federal de Viçosa, Viçosa, 1989.
CORRÊA, C.D.S. Efeito do tipo de estaca, do ácido indolbutírico e do boro
no enraizamento de estacas de azaléia (Rhododendron simsii Planch.).
2001. 61f. Momografia (Trabalho de Graduação em Agronomia) – Faculdade
de Ciências Agronômicas, Universidade Estadual Paulista, Jaboticabal, 2001.
DHRU, P.; GUPTA, S.K.; PAL, D. Effect of moisture, temperature and radiation
stress on therapeutic yield and biomass in Nerium oleander and Urginea
indica. Advances in Plant Sciences, v.4, n.1, p. 54-60, 1991.
DRADAR, V.A.; MENARY, R.C. Mineral nutrition of Olearia phologopappa:
Effect of growth, essential oil yield and composition. Soil Sci. Plant Anal. v. 26,
p. 1299-313. 1995.
EGERT, M.; TEVINI, M. Influence of drought on some physiological parameters
symptomatic for oxidative stress in leaves of chives (Allium shoenoprasum).
Environmental and Experimental Botany, v. 48, p. 43-49, 2002.
FACHINELLO, J.C., HOFFMANN, A., NACHTIGL, J.C., KERSTEN, E.,
FORTES, G.R.L. Propagação de plantas frutíferas de clima temperado.
2.ed. Pelotas: Editora e Gráfica UFPEL, 1995. 179p.
FARIA, J.C.; SACRAMENTO, C.K. Enraizamento e crescimento de estacas
herbáceas do cacaueiro (clones CEPEC 42, TSH 516 e TSH 1188) em função
da aplicação do acido indolbutírico. Revista Brasileira de Fruticultura,
Jaboticabal, SP. v.25, n.1, p. 192-194, 2005.

66
FERNANDES F.M.; VIEIRA, R.D.; FERNANDES, P.D.; DE SÁ, M.E.
Multiplicação vegetativa de azaléia (Rhododendron simsii, Planch); efeito de
fitormônios e épocas de estaqueamento. Cientifica, v.7. n.2, p. 179-182, 1979.
FIOCRUZ – Sistema nacional de informações tóxico-farmacológicas da
Fundação Oswaldo Cruz (SINITOX). Disponível em: <http://www.fiocruz.br.>
Acesso em: 10 abr. 2006
GONTIJO, T.C.A.; RAMOS, J.D.; MENDONÇA, V.; PIO, R.; ARAUJO NETO,
S.E.; CORRÊA, F.L.O. Enraizamento de diferentes tipos de estacas de
aceroleira utilizando ácido indolbutírico. Revista Brasileira de Fruticultura. v.
25, n. 2, p. 290-292. 2003.
GOTTLIBEB, O.R.; KAPLAN, M.A.C.; BORIN, M.R.M.B. Biodiversidade: um
enfoque quimico-biológico. Rio de Janeiro: UFRJ. 1996. 268p.
GRAZIANO, T.T. Estudos sobre o enraizamento de estacas e o
desenvolvimento inicial de mudas de primavera (Bougainvillea spectabilis
Willd.) e alamanda (Allamanda cathartica L.). 1981. 64f. Dissertação
(Mestrado em Agronomia) – Faculdade de Ciências Agronômicas, Universidade
Estadual Paulista, Jaboticabal, 1981.
GROS, E.G.; POMILIO. A.B.; SELDES, A.M.;BURTON, G. Introducction al
studio de los productos naturales. Washington: The General Secretariat of
the Organization of american Status. 1985. 196p.
HAISSIG, B.E. Influence of indole-3-acetic acid on adventitious root primordia of
Brittle Willow. Planta, v.95, p.27-35. 1970.
HARAGUCHI, M. Plantas tóxicas de interesse na pecuária. Instituto
Biológico. São Paulo. v.65, n.1, p.37-39. 2003
HARBONE, J.B.; BAXTER, H. (1995). Phytochemical dictionary: a handbook
of bioactive e compounds from plants. Taylor and Francis: London. 1985.

67
HARTMANN, H. T.; KESTER, D. E.; DAVIES JR., F. T.; GENEVE, R. L. Plant
propagation: principles and practices. 6.ed. New Jersey: Prentice-Hall, 1997.
770p.
HARTMANN, H. T.; LORETI, F. Seasonal variation in rooting leafy olive cuttings
under mist. American Society for Horticultural Science, Alexandria, v.87,
p.194-198, 1965.
HESS, C.E. A physiological comparison of rooting en easy and difficult-t-root
cuttings. Proceedings Plant Propagation Society, v.1962, p.265-268, 1963.
HOEHNE, F.C. Plantas e substâncias vegetais tóxicas e medicinais. São
Paulo: Graphicars, 1939. 355p.
HOFT, M.; VERPOORTE, R.; BECK, E. Growth and alkaloid contents in leaves
of Tabernaemantana pachysiphon Sapf (Apocinaceae) as influenced by light
intensity, water and nutrient supply. Oecologia, Heidelberg, v.17, p. 160-169,
1996.
HORNOK, L. Influence of nutrition on the yield and content of active compounds
in some essential oil plants. Acta horticulturae, n. 132, p. 239-247, 1983.
HOWARD, B.H. Factors affecting the response of leafless winter cut-tings of
apple and plum to IBA applied in powder formulation. Journal of Horticultural
Science, London, v.60, n.2, p.161-168, 1985.
JANICK, J. A ciência da horticultura. Rio de Janeiro: Freitas Bastos, 1966.
485 p.
KRAMER, P. J.; KOZLOWSKI, T. T. Fisiologia das árvores. Lisboa: Fundação
Calouste Gulbenkian, 1960. p. 745.
KUDREV, T.G. Água: vida das plantas. São Paulo: Ícone, 178p. 1994.

68
LORENZI, H.; SOUZA, H.M.. Plantas ornamentais no Brasil: arbustivas,
herbáceas e trepadeiras. 3.ed. Nova Odessa, SP: Instituto Plantarum, 2001.
1088p.
LORENZI, H.; MATOS, A. J. F. Plantas medicinais no Brasil: nativas e
exóticas. Nova Odessa, SP: Instituto Plantarum, 2002. 512p.
LORETI, F.; HARTMANN, H.T. Propagation of olive trees by rooting leafy
cuttings under mist. American Society for Horticultural Science, v. 85, p.
257-265, 1965.
MAIA, N.B.; BOVI, O.A.; PERECIN, M.B.; GRANJA, N.P. Crescimento e
qualidade do óleo essencial de alfavaca-cravo em hidroponia. Horticultura
Brasileira V.19, suplemento, 2001.
MAPELI, N.C.; VIEIRA, M.C.; HEREDIAZ, N.A.; SIQUEIRA, J.M. Produção de
biomassa e de óleo essencial dos capítulos florais da camomila em função do
nitrogênio e fósforo. Horticultura Brasileira, Brasília, v. 23, n.1, p. 32-37.
2005.
MARTINS, A. B. G. Enraizamento de estacas enfolhadas de três variedades
de lichia (Litchi chinensis Sonn.). 1998. 100f. Tese (Doutorado em
Agronomia) – Faculdade de Ciências Agronômicas, Universidade Estadual
Paulista, Jaboticabal, 1998.
MERCK. Manual Merck de medicina. 17. ed. Rio de Janeiro: Rocca, 2001.
2736p.
MING, L.C. Influência de diferentes níveis de adubação orgânica na
produção de biomassa e teor de óleos essenciais de Lippia alba (Mill.)
N.E. Br. Verbenaceae. Curitiba. 1992. 206p. Dissertação (Mestrado em
ciências biológicas) – Universidade Federal do Paraná.

69
NOWACKI, E.; JURZYSTA, M.; GORSKI, P. Effect of availability of nitrogen on
alkaloid synthesis in Solanaceae. Bulletin de Academie Polonaise des
Sciences, Warsaw, v. 23, n. 3, p. 219-225, 1975.
OLIVEIRA JUNIOR, A.C.; FAQUIM,V.; PINTO,J.E.B.P.; LIMA SOBRINHO,
R.R.;
BERTOLUCCI, S.K.V. Teor e rendimento de óleo essencial no peso fresco de
arnica, em função de calagem e adubação. Horticultura Brasileira. Brasília.
V.23, n.3, p.735-739. 2005
OLIVEIRA, R.B.; GODOY, S.A.P.; COSTA, F.B. Plantas tóxicas:
conhecimento e prevenção de acidentes. Ribeirão Preto: Holos, 2003. 64p.
ONO, E.; RODRIGUES, J.D. Aspectos da fisiologia do entraizamento de
estacas caulinares. Jaboticabal: FUNEP, 1996. 83p.
OVERBEEK, J. van, GORDON, S.A., GREGORY, L.E. An analysis of the
function of the leaf in the process of root formation in cuttings. American
Journal Botany, v.33, p.100-107, 1946.
PALEVITCH, D. Recent advances in cultivation of medicinal plants. Acta
horticulturae. V.208, p.29-34. 1987.
PEDRINHO, D. R. Efeito do armazenamento de estacas e de hastes no
enraizamento de roseiras para corte nas quatro estações. 2002. 37f.
Dissertação (Mestrado em Agronomia) – Faculdade de Ciências Agrárias e
Veterinárias, Universidade Estadual Paulista, Jaboticabal, 2002.
PETRECHEN, E. de P. Efeito do ácido indolbutírico no enraizamento de
estacas herbáceas de goiabeira (Psidium guajava L.) das cultivares ‘Rica’
e ‘Paluma’ em câmara de nebulização intermitente. 1988. 50 f. Dissertação
(Mestrado em Agronomia) - Faculdade de Ciências Agrárias e Veterinárias,
Universidade Estadual Paulista, Jaboticabal, 1988.

70
PIVETTA, K. F. L. Efeito do ácido indolbutírico no enraizamento de estacas
herbáceas de nogueira-macadâmia (Macadamia integrifolia Smith) e
desenvolvimento inicial das mudas. 1990. 91f. Dissertação (Mestrado em
Agronomia) - Faculdade de Ciências Agrárias e Veterinárias, Universidade
Estadual Paulista, Jaboticabal, 1990.
PIVETTA, K. F. L. Estudos sobre enraizamento de estacas enfolhadas de
roseira (Rosa sp) "Red Success". 1994. 151f. Tese (Doutorado em
Agronomia) - Faculdade de Ciências Agrárias e Veterinárias, Universidade
Estadual Paulista, Jaboticabal, 1994.
QUINTANILHA, L.F. Enraizamento de porta-enxertos de roseira (Rosa
canina ‘Inermis’) com uso de AIB e dois substratos. 1995. 44f. Dissertação
(Mestrado em Ciências) - Faculdade de Agronomia Eliseu Maciel, Escola
Federal de Pelotas, Pelotas, 1995.
RAMOS, J.D.; MATOS, L.E.S.; GONTIJO, T.C.A.; PIO, R.; JUNQUEIRA, K.P.;
SANTOS, F.C. Enraizamento de estacas herbáceas de ‘Mirabolano’ (Prunus
cerasifera Ehrn.) em diferentes substratos e concentrações de ácido
indolbutírico. Revista Brasileira de Fruticultura, Jaboticabal, SP. v.25, n.1, p.
189-191, 2003.
RIBEIRO, A.C.; GUIMARÃES, P.T.G.; ALVARES, V.C. Recomendação para o
uso de corretivos e fertilizantes em Minas Gerais: 5a aproximação.Viçosa:
UFV, 1999. 359p.
SALOMON, I. Ecobiology of the chamomille (Chamomille recutita
L.).Horticultura brasileira. Brasilia. v. 12, n. 2, p. 226-229, 1994.
SANTOS, T.S.; SANTOS, M.F.; MENDONÇA, M.C.; SILVA JUNIOR, C.D.;
SILVA-MANN, R.; ARRIGONI-BLANK, M.F.; BLANK, A.F. Efeito do estresse
hídrico na produção de massa foliar e teor de óleo essencial em sambacaitá
(Hyptis pectinata L.). Horticultura brasileira. v. 22, n. 2, 2004. Suplemento
CD Room.

71
SARZI, I. Efeito das estações do ano e do acido indolbutírico no
enraizamento de estacas de variedades de mini-roseira (Rosa spp.).
Dissertação (Mestrado em Agronomia) – Faculdade de Ciências Agrárias e
Veterinárias, Universidade Estadual Paulista, Jaboticabal, 2002.
SCALOPPI JUNIOR, E.J.; MARTINS, A.B.G. Clonagem de quatro espécies de
Annonaceae potenciais como porta-enxertos. Revista Brasileira de
Fruticultura, Jaboticabal, SP. v.25, n.2, p. 95-97, 2003.
SCHENKEL, E.P.; GOSMANN, G. MELLO, J.C.P.; MENTZ, L.A.; PETROVICK,
P.R. Farmacognosia: da planta ao medicamento. 5.ed. Porto Alegre:
UFRGS; Florianópolis: Ed. UFSC, 2003. p.403-434.
SCHERMEISTER, L.J.; CRANE, F,A.; VOIGT, R.F. Nitrogenous constituents of
Atropa belladonna L. grown sources of externally supplied nitrogen. Journal of
the American Pharmaceutical Association, Washington, v.49, n.11, p.698-
705, 1960.
SHARAPIN, N.; ROCHA, L.M.; CARVALHO, E.S.; LÚCIO, E.M.R. DE A.;
SANTOS, E.V.M. DOS; ALMEIDA, J.M.L. de. Fundamentos de tecnologia de
produtos fitoterápicos. 12. ed. Rio de Janeiro: Merck e Co, 2000. p.87-89.
SHARMA, S.D.; CLAVERIE, J. Seasonal rooting behavior of cuttings of plum
cultivar as influenced by IBA treatments. Scientia Horticulturae, Amsterdan,
v.40, p. 297-303, 1989.
SILVA, T.R.B.; ART, O.; SORALTO, R.P. Adubação nitrogenada e resíduos
vegetais no desenvolvimento de feijoeiro em sistema de plantio direto. Acta
Scientiarum, Maringá. v.25, n.1. p.81-87, 2003.
SIMÕES, C.M.O.; SCHENKEL, E.P.; GOSMAN, G.; MELLO, J.C.P.; MENTZ,
L.A.; PETROVICK, P.R. Farmacognosia: da planta ao medicamento. 3: ed.
Porto Alegre: Editora Universidade/UFRGS, 2000. 329p.

72
SOUZA, H.M. Os tipos de estacas. Suplemento Agrícola, n. 1141, p.7, 1977.
SOUZA, H.M., INFORZATO, R. Enraizamento de estacas de Camellia japonica
L. var Alba Plam, por meio de hormônios vegetais. Bragantia, v.18, p. 5-8.
1959.
STEPONKUS, P.L. Cold acclimation and freezing injury from a perspective of
plasma membrane. In: Katterman, F. (ed.). Environmental injury to plants.
London: Academic Press, 1990. cap.1, p. 1-15.
STEYN, D.G. The poisoning of human beings by wild plants, ornamental
plants and domestic poisons. Farming in south Africa, Reprint n.6, p.8, 1934.
STOLTZ, L.P.; ANDERSON, R.G. Rooting of single node cotting of roses. Acta
Horticulturae, v. 227, p.230-235, 1988.
TAIZ, L.; ZEIGER, E. Fisiologia vegetal. 3.ed. Porto Alegre: Artmed, 2004.
719p.
THOMPSON, W.K. Effects of origin, time of collection, auxins and planting
media on rooting of cuttings of Epacris impressa. Labill. Scientia Horticulturae,
v. 30, p. 127-134, 1986.
TOKARNIA, C.H.; DOBEREINER, J.; PEIXOTO, P.V. Plantas tóxicas do
Brasil. Rio de Janeiro: Helianthus, 2000. 320p.
UNANDER, D.W.; BLUMBERG, B.S. In vitro activity of Phyllanthus
(Euphorbiaceae) species against the DNA polymerase of hepatitis vírus: effects
of growing environment and inter- and intra-specific differences. Economic
Botany, Bronx, v. 45, n. 2, p. 225-242, 1990.
VICKERY, M.L.; VICKERY, B. Secondary plant metabolism. Hong Kong: The
Macmillan Press Ltd., 1981. 184p.

73
WAGNER, H.; BLADT, S. Plant drug analysis a thin layer chromatography atlas.
2.ed. Berlin: Springer, 1996. 348p.