epitope mapping and characterization of a neutralizing monoclonal antibody against human adenovirus...
TRANSCRIPT

Accepted Manuscript
Title: Epitope mapping and characterization of a neutralizingmonoclonal antibody against human adenovirus type 3
Author: Xingui Tian Chenyang Li Chunyan Xue Xiao LiZhichao Zhou Rong Zhou
PII: S0168-1702(13)00292-XDOI: http://dx.doi.org/doi:10.1016/j.virusres.2013.08.013Reference: VIRUS 96056
To appear in: Virus Research
Received date: 4-7-2013Revised date: 28-8-2013Accepted date: 29-8-2013
Please cite this article as: Tian, X., Li, C., Xue, C., Li, X., Zhou,Z., Zhou, R., Epitope mapping and characterization of a neutralizingmonoclonal antibody against human adenovirus type 3, Virus Research (2013),http://dx.doi.org/10.1016/j.virusres.2013.08.013
This is a PDF file of an unedited manuscript that has been accepted for publication.As a service to our customers we are providing this early version of the manuscript.The manuscript will undergo copyediting, typesetting, and review of the resulting proofbefore it is published in its final form. Please note that during the production processerrors may be discovered which could affect the content, and all legal disclaimers thatapply to the journal pertain.

Page 1 of 20
Accep
ted
Man
uscr
ipt
1
Epitope mapping and characterization of a neutralizing monoclonal 1
antibody against human adenovirus type 32
3
Xingui Tian, Chenyang Li, Chunyan Xue, Xiao Li, Zhichao Zhou, Rong Zhou*4
State Key Lab of Respiratory Disease, Guangzhou Institute of Respiratory Disease, The5
First Affiliated Hospital of Guangzhou Medical University, Guangzhou 510120, China6
7
*Corresponding author: Dr Zhou, State Key Lab of Respiratory Disease, The First 8
Affiliated Hospital of Guangzhou Medical University, 151 Yan Jiang Road, 9
Guangzhou 510120, China. Phone: +86 20 34281614. Fax: +86 20 34281614. Email: 10
E-mail addresses: [email protected] (X. Tian), [email protected] (C. Li), 12
[email protected] (C. Xue), [email protected] (X. Li), [email protected] (Z. Zhou), 13
[email protected] (R. Zhou)14

Page 2 of 20
Accep
ted
Man
uscr
ipt
2
Abstract15
Human adenovirus serotype 3 (HAdV-3) has occurred as a global epidemic in recent 16
years causing serious diseases such as pneumonia in pediatric and adult patients.17
Development of reliable diagnostic reagents and identification of neutralizing epitopes18
is important for the surveillance and control of infection. In this study, a neutralizing 19
monoclonal antibody (MAb) MAb 1B6 was generated using the HAdV-3 virion. MAb 20
1B6 specially recognized the HAdV-3 virus particles and the HAdV-3 hexon protein, 21
but not the virus particles or the hexon protein of HAdV-7 and HAdV-4 by western-blot 22
analysis and indirect enzyme-linked immunosorbent assay (ELISA). Analysis using a23
series of peptides from the hexon protein and chimeric adenovirus (Ad) particles of 24
epitope mutants revealed that MAb 1B6 bound to the exposed region (amino acid 25
positions 414-424 of hexon) in hypervariable region 7 (HVR7). ELISA demonstrated 26
that MAb 1B6 could recognize the corresponding regions of other HAdV-3 genotypes 27
that have some residues substituted. The identification of the neutralizing epitope and 28
the generation of MAb 1B6 may be useful for clinical serotype-specific diagnosis, 29
subunit vaccine construction for HAdV-3 infection, and virion structural analysis.30
31
Key words: human adenovirus type 3, neutralizing monoclonal antibody, epitope 32
identification33

Page 3 of 20
Accep
ted
Man
uscr
ipt
3
1. Introduction 34
Human adenoviruses (HAdV) belong to the Mastadenovirus genus and cause a broad 35
spectrum of diseases in pediatric and adult patients, including epidemic 36
keratoconjunctivitis, acute respiratory illnesses, and acute gastroenteritis (1, 2). To 37
date, 51 human adenovirus serotypes have been identified and classified into six groups 38
(A to F). HAdV species C (HAdV-1, -2, -5, and -6), species B (HAdV-3, -7, and -14) 39
and species E (HAdV-4) are most commonly found in patients with respiratory40
infection (3, 4). Among these, the major epidemic strains are HAdV-3 of subspecies B141
that have caused severe respiratory diseases, such as acute respiratory disease and lethal42
pneumonia epidemics and outbreaks in Asia, Europe, and America (5-10). 43
Currently, there is no effective medicine or vaccine against HAdV-3. Neutralizing 44
monoclonal antibodies (MAb) may be a promising prophylactic or therapeutic 45
medicine against virus disease. Neutralizing MAbs also could be useful for identifying 46
neutralizing epitopes, which is of great importance in the molecular design of vaccines. 47
For diagnosis and therapy, the typing of HAdV infection is valuable and essential 48
because of the numerous serotypes present. Thus, type-specific MAbs against HAdV-3 49
may be used to develop rapid and reliable immunological tests for HAdV-3 infection. 50
The adenovirus capsid is an icosahedron composed of three structural proteins: hexon, 51
penton base and fiber. The hexon protein is the major antigenic determinant (11). 52
Type-specific neutralizing epitopes of hexons have been proposed to reside within 53
seven high variable regions (HVRs) of the hexon, in which HVR7 could be further 54
subdivided into three highly variable regions (12). Our previous study demonstrated 55
that HVR1, 2, 5 and 7 regions of HAdV-3 containing neutralizing epitopes (13). 56
However, the HVR7 of HAdV-3 is long (30 amino acids) and requires further detailed 57
study. 58
To understand the neutralization epitopes of HAdV-3 better, we produced a 59
neutralizing MAb termed MAb 1B6 against the HAdV-3-gz01 viruses. Epitope 60
mapping revealed that MAb 1B6 recognized the HVR7 of HAdV-3 hexon. The data 61
show that MAb 1B6 cross-detected the HVR7s of other HAdV-3 strains but not 62

Page 4 of 20
Accep
ted
Man
uscr
ipt
4
HAdV-7 and HAdV-4. The findings of this study will be useful for the reliable and 63
stable serotype-specific diagnosis of adenovirus type 3 infection, and adenovirus 64
structural analysis.65
66
2. Methods and materials67
2.1 Virus strains and cells68
The following strains of adenoviruses used in this study were obtained as previously 69
described or maintained in our laboratory (11, 13): HAdV-3 GZ01 strain (GenBank 70
accession no. DQ099432), HAdV-4 GZ01 strain (GenBank accession no. KF006344), 71
HAdV-7 GZ08 strain (GenBank accession no. GQ478341), recombinant HAdV-5, 72
recombinant adenovirus Ad3EGFP encoding a HAdV-3 GZ-01 genome and enhanced 73
green fluorescent protein (eGFP) with an E3 region deletion, hexon chimeric 74
adenovirus rAd3egf/H7 with the hexon gene from HAdV-7 replacing the HAdV-3 75
hexon gene of the Ad3EGFP, and epitope mutants (rAdH7R1, rAdH7R2, rAdH7R3, 76
rAdH7R4) from rAd3egf/H7 replaced with corresponding HAdV-3 epitopes. All 77
adenoviruses were cultured in HEp-2 cells or AD293 cells. The cells were maintained 78
in Dulbecco’s modified Eagle’s medium (DMEM)(Gibco, USA) supplemented with 79
100 IU/ml penicillin, 100 mg/ml streptomycin, and 10% fetal bovine serum. 80
Adenovirus particles were purified by standard CsCl gradient centrifugation, as 81
previously described (14). The virus particle (VP) titers were determined by 82
spectrophotometry using a conversion factor of 1.1 × 1012 VPs per absorbance unit at 83
260 nm.84
85
2.2 Western blot analysis86
HAdV-3 virions were used to immunize BALB/c mice and screen MAbs. Production 87
and selection of mouse monoclonal antibodies against HAdV-3 were performed as 88
described previously (24). To analyze the reactivity of MAbs with the adenovirus 89
capsid proteins, total protein from purified adenoviruses was separated by 10% SDS 90
polyacrylamide gels. The proteins were transferred onto PVDF membranes using a wet 91

Page 5 of 20
Accep
ted
Man
uscr
ipt
5
blotter. The membranes were blocked with 5% skim-milk in phosphate buffered saline92
(PBS) and then incubated with the MAb at final dilutions of 1:10,000, washed and93
exposed to a 1:10,000 dilution of goat anti-mouse IgG (H+L)-horseradish peroxidase94
(HRP) conjugate affinity-purified secondary antibody (Bio-Rad, China). The blot was 95
developed using Immobilon Western chemiluminescence HRP substrate (Millipore, 96
USA).97
98
2.3 Virus neutralization test99
For in vitro adenovirus neutralization experiments, purified MAbs were serially diluted 100
2-fold in DMEM, and 50 µl aliquots of each dilution were mixed with 50 µl 101
recombinant adenoviruses (Ad3EGFP or rAd3egf/H7) with 2 × 105 VPs. The 102
antibody-virus mixtures were incubated for 1 h at 37°C and then transferred to 96-well 103
plates containing 85–95% confluent monolayers of HEp-2 cells. Monolayers were104
cultured in RPMI Medium 1640 without phenol red (Gibco, USA) and serum for 72 h. 105
Infected cells were analyzed using a Varioskan Flash Multimode Reader (Thermo 106
Scientific) to measure the EGFP expression. Neutralizing titers were defined as the 107
minimum concentration of MAb that inhibited 90% of EGFP expression.108
109
2.4 Expression of recombinant protein fragments110
A pQE30 expression vector was used to express different serotype hexon peptides with 111
a hexahistidine (His) tag, and a pGEX-4T-3 vector was used to produce short peptides 112
(HVRs) with an N-terminal glutathione of S-transferase (GST) tag. The primers used 113
for cloning these fragments are shown in Table 1. Each primer contained an engineered 114
restriction enzyme site to facilitate the cloning. The three HIS fusion proteins were 115
purified by Ni-NTA beads affinity chromatography (Novagen, USA) according to the 116
manufacturer’s instructions, and the GST fusion peptides were purified by affinity 117
chromatography using GST-Bind Resin (Novagen, USA) under native conditions. The 118
control protein GST was purified under the same conditions.119
120
2.5 Indirect Enzyme-linked immunosorbent assay (ELISA) analysis121

Page 6 of 20
Accep
ted
Man
uscr
ipt
6
For ELISA, 96-well plates (Nunc Maxisorp) were coated overnight at 4°C with fusion 122
peptides (about 2 µg/ml) or virus particles (about 1010 VPs/ml) in PBS (pH 7.4) and 123
were washed one time with 0.05% Tween-20 in phosphate-buffered saline (PBST) and124
blocked for 2 h with 2% bovine serum albumin (BSA) in PBST. Then, 100 µl/well 125
MAb ascites was added to each well and incubated at a dilution of 1:5000 for 1 h at 126
37°C. The plates were washed three times with PBST and incubated for 1 h with a 127
1:10,000 dilution of goat anti-mouse IgG (H+L)-HRP conjugate affinity-purified 128
secondary antibody (Bio-Rad, China). After washing four times with PBST, the plates129
were developed with tetramethylbenzidine (TMB) substrate, stopped with 2M H2SO4, 130
and analyzed at 450 nm using an ELISA plate reader (Thermo Scientific Multiskan131
MK3).132
133
2.6 Peptide competition ELISA134
The epitope detected by MAb was confirmed by competitive inhibition ELISA. 135
Optimized concentrations of the MAbs were determined by serial dilution. Briefly, the 136
HAdV-3 GZ01 virions in PBS (pH 7.4) were used to coat 96-well plates overnight at137
4°C. In separate tubes, constant concentrations of MAb at a final dilution of 1:1000 138
were added to increasing concentrations of competitor peptide (0.1, 1, 10, or 100 139
µg/ml) in blocking buffer and incubated for 30 min at 37°C. The virion-coated plates 140
were washed one time with washing buffer and incubated with blocking buffer for 2 h 141
at 37°C. Then each of the MAb-peptide mixtures was added to duplicate wells, and the 142
plates were incubated for 1 h at 37°C. A control MAb in blocking buffer without 143
peptide was included in each plate. The subsequent processes were performed as 144
described for indirect ELISA (section 2.5). 145
146
2.7 Immunocytochemistry147
To test whether 1B6 could be used to detect adenovirus-infected cells, 148
immunocytochemistry was performed. Briefly, the AD293 cells were infected with 2 × 149
105 VP Ads in 24-well plates and were cultured for 2 days at 37°C. After removal of the 150
media, the plates were rinsed once with PBS, and immediately fixed with methanol 151

Page 7 of 20
Accep
ted
Man
uscr
ipt
7
containing 0.3% H2O2 for 5 min in a −20°C freezer. They were then blocked with 152
blocking buffer, goat sera (5%) in PBST for 1 h, with gentle rocking, and then cells 153
were incubated with MAb at a final dilution of 1:1000 in blocking buffer for 1 h at 154
37°C, washed and exposed to a 1:2000 dilution of goat anti-mouse IgG (H+L)-HRP 155
conjugate affinity-purified secondary antibody (Bio-Rad, China). The cells were 156
developed using DAB reagent and observed by microscopy.157
158
2.8 Statistical Analysis159
The data are presented as the mean ± standard error. Statistical significance was 160
determined using Prism 5.0 software. Comparisons among groups were performed by 161
ANOVA with Bonferroni’s test to account for multiple comparisons. P values of less 162
than 0.05 were considered statistically significant.163
3. Results164
3.1 Generation and identification of neutralizing MAb 1B6 directed against 165
HAdV-3 hexon166
Purified HAdV-3 GZ01 was used to immunize mice and a monoclonal IgG1 isotype 167
antibody, MAb 1B6, was obtained. The ascites fluid titer by ELISA was 160,000.168
ELISA analysis indicated that MAb 1B6 reacted with its parental antigen, whole 169
HAdV-3 GZ01 virus particles, which suggested the epitope recognized by MAb 1B6170
was presented on the surface of the virion. 171
Western blotting demonstrated that MAb 1B6 specifically detected the hexon protein 172
of the HAdV-3 GZ01 capsid (Fig. 1A), which was also confirmed by indirect-ELISA 173
using the truncated hexon peptide expressed in Escherichia coli (Fig. 1B). MAb 1B6 174
did not react with the HAdV-4 GZ01 or HAdV-7 GZ08 hexon (Fig. 1A and B), and 175
showed a lack of reactivity with the negative control (data not shown). 176
MAb 1B6 neutralized Ad3EGFP but not rAd3egf/H7, and the neutralizing titer of 177
MAb 1B6 was 10μg/ml (Fig. 1C). Immunocytochemistry analysis using MAb 1B6 178
specifically detected cells infected with HAdV-3 GZ01 virus, but not those infected 179
with HAdV-4 GZ01 or HAdV-7 GZ08 viruses (data not shown). Thus, the epitope 180

Page 8 of 20
Accep
ted
Man
uscr
ipt
8
recognized by MAb 1B6 was serotype 3-specific and located within the region of the 181
truncated hexon protein containing all 7 HVRs, which are thought to be continuous 182
based on their reactivity with the denatured protein and native virion.183
184
3.2 Mapping of the epitope recognized by MAb 1B6185
The hexon crystallographic and phylogenetic analysis revealed that seven 186
hypervariable regions (HVRs) might contain serotype-specific residues (21, 25). 187
Sequence alignment between the Ad3 and Ad7 hexon protein clearly demonstrated 188
variation of one or more amino acids in HVR-1, -2, -4, -5, -6 and -7, in which HVR7189
was further subdivided into three highly variable regions named HVR7s, HVR8 and 190
HVR9.191
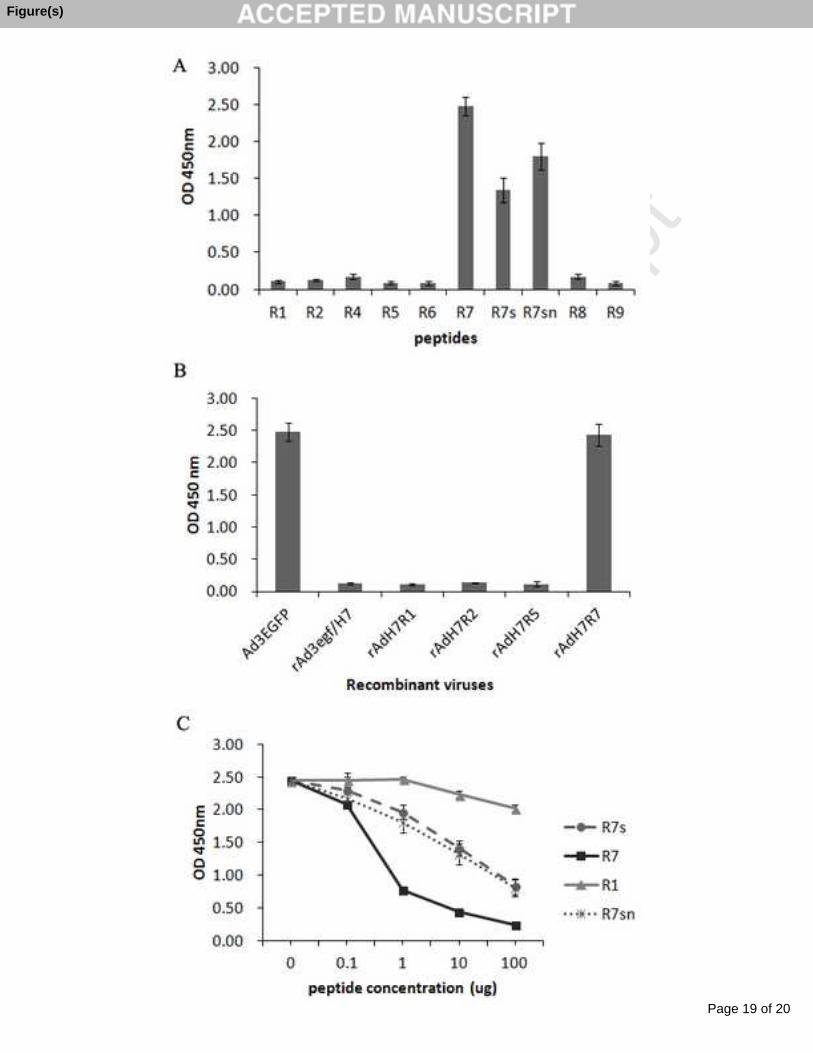
To identify the region of the HAdV-3 hexon bound with MAb 1B6, we expressed 192
recombinant HVR peptides with GST tag. Indirect-ELISA revealed that MAb 1B6193
reacted with R7-GST and R7s-GST but not others (Fig. 2A), suggesting MAb 1B6194
recognized HVR7s of the hexon. To access the epitope on a native virion, we used a 195
previously constructed series of recombinant adenoviruses with chimeric hexon to react 196
with MAb 1B6 by indirect-ELISA. This analysis also confirmed HVR7 as the region 197
bound with MAb 1B6 (Fig. 2B). 198
Further peptide competitive inhibition ELISA was performed to demonstrate the 199
accuracy of the identified epitope for MAb 1B6. Increasing concentrations of peptides 200
were incubated with MAb 1B6. The R1-GST fragment was used as a negative control. 201
The mixtures were then assayed by ELISA with HAdV-3 virus particle-coated plates. 202
The fragments R7 and R7sn competed with virions for MAb 1B6 binding in a 203
dose-dependent manner, while the control peptide R1 did not. Higher concentrations of204
peptide R7s could partly compete with virions for MAb 1B6 binding compared with R7 205
(Fig. 2C). Thus, R7sn was the epitope recognized by MAb 1B6.206
207
3.3 Characterization of the identified epitope for MAb 1B6208
MAb 1B6 showed unique reactivity to the hexon of adenovirus serotype 3, therefore, it 209
is necessary to determine the distribution of the epitope among different groups/types 210

Page 9 of 20
Accep
ted
Man
uscr
ipt
9
of human adenovirus. Alignment of the hexon protein sequences available from 211
GenBank revealed that the epitope was unique for serotype 3 adenovirus (13), but212
global HAdV-3 strains showed amino acid substitutions in the region (Fig. 3A). Three 213
variants (M1 [GPGNRYQGIK], M2 [GPGHTYQGIK] and M3 [GPGHTFQGVK]) 214
that represented all variants from HAdV-3 strains were also constructed to detect the215
reactive range of MAb 1B6. ELISA demonstrated that MAb 1B6 bound to all the 216
variants, but not the control peptide of adenovirus serotype 7 (Fig. 3B). Therefore, 217
MAb 1B6 could be a useful tool for distinguishing human adenovirus serotype 3 from218
other adenoviruses. 219
220
4. Discussion221
In this study, we generated MAb 1B6, which neutralized HAdV-3 (Ad3EGFP) but not 222
HAdV-7 (rAd3egf/H7) and specifically detected the HAdV-3 hexon protein (Fig. 1). 223
Epitope mapping on HAdV-3 hexon revealed that MAb 1B6 recognized the exposed 224
region (amino acid positions 414-424 of the hexon) corresponding to the HVR7 (Fig. 225
2). 226
Hexon crystallographic and phylogenetic analysis revealed seven HVRs might contain 227
serotype-specific residues that are exposed to the capsid surface (15). However, to our 228
knowledge, among the 52 adenoviruses, few type-specific neutralization antibody 229
recognition sites have been identified (13, 16-19). In our previous study, we identified 230
HVR3 as the only epitope shared by HAdV-3 and HAdV-7 using MAb 5D4 (20).231
Although HVR7 of HAdV-3 is a neutralizing region, the precise neutralizing epitope 232
was not clear (13). In this paper, HVR7 was further subdivided into three fragments by 233
sequence alignment, of which HVR7sn (GPGHRYQGIK) was detected by the 234
neutralizing MAb 1B6 (Fig. 2B). The reactivity of MAb 1B6 with the peptide 235
“GPGHRYQG” was reduced, demonstrating that the amino acids “IK” affected MAb 236
1B6 binding (Fig. 2B). However, the epitope “GPGHRYQGIK” is not included in the 237
predicted neutralizing regions for HAdV-3 by homology modeling (18, 21). This 238
indicates the structural complexity of HAdV-3 HVR7, which may contain several 239

Page 10 of 20
Accep
ted
Man
uscr
ipt
10
neutralization epitopes. The HVR7sn also could be a candidate position for displaying 240
exogenous epitope (22). 241
MAb 1B6 detected and neutralized HAdV-3 but not HAdV-7 and HAdV-4 (Fig. 1), 242
therefore, it may be used to distinguish HAdV-3 from other serotype HAdVs. The 243
epitope was unique for HAdV-3. However, it is necessary to identify the distribution of 244
the epitope among HAdV-3 strains. Although the HAdV-3 genome is relatively stable 245
across time and geographic space, it still yields small-scale nucleotide variations (23). 246
Several studies found that the HAdV-3 hexon sequences showed a high heterogeneity 247
that affected HVRs and formed multiple genetic lineages (24-26). In these studies, the 248
amino acid changes were conserved among the genotypes, and the epitope 249
“GPGHRYQGIK” had three genotypes, “GPGHRYQGIK”, “GPGHTYQGIK” and 250
“GPGNRYQGIK” with only one substitution at position 147 H to N or at the position 251
148 R to T. This was in accordance with our result using globally known HAdV-3 252
hexon sequences from GenBank alignment. In recent years, a rare mutant with a 253
“GPGHTFQGVK” genotype was detected. Indirect-ELISA analysis demonstrated 254
reactivity of MAb 1B6 with the three variants was reduced but not abolished (Fig. 3B), 255
which suggested these substitutions may be tolerated.256
257
Acknowledgements258
This work was supported by the National Natural Science Foundation of China (NSFC, 259
31070150) and Guangdong and Macau Joint Center of Innovative Drugs for 260
Respiratory Pathogens (2010B091000018).261
262
Authors’ contributions263
TXG performed expression of recombinant protein fragments, ELISA analysis and 264
western blot assays, analyzed the data, and drafted the manuscript. LCY produced the 265
monoclonal antibody. XQY collected serum samples and performed some ELISA 266
analysis. LX and ZZC prepared the viruses particles. ZR designed and supervised the 267

Page 11 of 20
Accep
ted
Man
uscr
ipt
11
work, and edited the final version of this manuscript. All authors read and approved the 268
final version of the manuscript.269
270
Competing interests271
The authors declare that they have no competing interests.272
273
References274
1. Lenaerts, L., De Clercq, E., Naesens, L., 2008. Clinical features and treatment of 275adenovirus infections. Rev Med Virol. 18(6), 357-374.2762. Echavarria, M., 2009. Adenovirus, p. 463–488. In A. J. Zuckerman, J. E. Banatvala, 277B. D. Schoub, P. D. Griffiths, and P. Mortimer (ed.), Principles and practice of clinical 278virology, 6th ed. John Wiley and Sons, San Diego, CA.2793. Pavia, A.T., 2011. Viral infections of the lower respiratory tract: old viruses, new 280viruses, and the role of diagnosis. Clin Infect Dis. 52 Suppl 4, S284-9.2814. Alharbi, S., Van Caeseele, P., Consunji-Araneta, R., Zoubeidi, T., Fanella, S., Souid, 282A.K., Alsuwaidi, A.R., 2012. Epidemiology of severe pediatric adenovirus lower 283respiratory tract infections in Manitoba, Canada, 1991-2005. BMC Infect Dis. 12:55.2845. Tabain, I., Ljubin-Sternak, S., Cepin-Bogovic, J., Markovinovic, L., Knezovic, I., 285Mlinaric-Galinovic, G., 2012. Adenovirus respiratory infections in hospitalized 286children: clinical findings in relation to species and serotypes. Pediatr Infect Dis J. 28731(7), 680-684.2886. Tsou, T.P., Tan, B.F., Chang, H.Y., Chen, W.C., Huang, Y.P., Lai, C.Y., Chao, Y.N., 289Wei, S.H., Hung, M.N., Hsu, L.C., Lu, C.Y., Shao, P.L., Mu, J.J., Chang, L.Y., Liu, 290M.T., 2012. Community Outbreak of Adenovirus, Taiwan, 2011. Emerg Infect Dis. 18, 2911825–1832.2927. Rojas, L.J., Jaramillo, C.A., Mojica, M.F., Escalante, M.P., Delgado, P., 2012. 293Molecular typing of adenovirus circulating in a Colombian paediatric population with 294acute respiratory infection. Epidemiol Infect. 140(5), 818-822.2958. Gao, X.Q., Jin, Y., Xie, Z.P., Gao, H.C., Xie, L.Y., Zhang, J., Duan, Z.J., 2012. [The 296epidemiological study of adenovirus in children with respiratory tract infections in 297Nanjing area from 2010 to 2011]. Bing Du Xue Bao. 28(5), 531-535. 2989. Barrero, P.R., Valinotto, L.E., Tittarelli, E., Mistchenko, A.S., 2012. Molecular 299typing of adenoviruses in pediatric respiratory infections in Buenos Aires, Argentina 300(1999-2010). J Clin Virol. 53(2), 145-150. 30110. Guo, L., Gonzalez, R., Zhou, H., Wu, C., Vernet, G., Wang, Z., Wang, J., 2012. 302Detection of three human adenovirus species in adults with acute respiratory infection 303in China. Eur J Clin Microbiol Infect Dis. 31(6), 1051-1058.30411. Tian, X., Su, X., Li, H., Li, X., Zhou, Z., Liu, W., Zhou, R., 2011. Construction and 305characterization of human adenovirus serotype 3 packaged by serotype 7 hexon. Virus 306Res. 160(1-2), 214-220.307

Page 12 of 20
Accep
ted
Man
uscr
ipt
12
12. Rux, J.J., Kuser, P.R., Burnett, R.M., 2003. Structural and phylogenetic analysis of 308adenovirus hexons by use of high-resolution x-ray crystallographic, molecular 309modeling, and sequence-based methods. J Virol. 77(17), 9553-9566.31013. Qiu, H., Li, X., Tian, X., Zhou, Z., Xing, K., Li, H., Tang, N., Liu, W., Bai, P., 311Zhou, R., 2012. Serotype-specific neutralizing antibody epitopes of human adenovirus 312type 3 (HAdV-3) and HAdV-7 reside in multiple hexon hypervariable regions. J Virol. 31386(15), 7964-7975.31414. Wu, H., Dmitriev, I., Kashentseva, E., Seki, T., Wang, M., Curiel, D.T., 2002. 315Construction and characterization of adenovirus serotype 5 packaged by serotype 3 316hexon. J Virol. 76(24), 12775-12782.31715. Rux, J.J., Burnett, R.M., 2000. Type-specific epitope locations revealed by X-ray 318crystallographic study of adenovirus type 5 hexon. Mol Ther. 1(1), 18-30.31916. Bradley, R.R., Maxfield, L.F., Lynch, D.M., Iampietro, M.J., Borducchi, E.N., 320Barouch, D.H., 2012. Adenovirus serotype 5-specific neutralizing antibodies target 321multiple hexon hypervariable regions. J Virol. 86(2), 1267-1272.32217. Pichla-Gollon, S.L., Drinker, M., Zhou, X., Xue, F., Rux, J.J., Gao, G.P., Wilson, 323J.M., Ertl, H.C., Burnett, R.M., Bergelson, J.M., 2007. Structure-based identification of 324a major neutralizing site in an adenovirus hexon. J Virol. 81(4), 1680-1689.32518. Yuan, X., Qu, Z., Wu, X., Wang, Y., Liu, L., Wei, F., Gao, H., Shang, L., Zhang, 326H., Cui, H., Zhao, Y., Wu, N., Tang, Y., Qin, L., 2009. Molecular modeling and 327epitopes mapping of human adenovirus type 3 hexon protein. Vaccine. 27(37), 3285103-5110.32919. Toogood, C.I., Crompton, J., Hay, R.T., 1992. Antipeptide antisera define 330neutralizing epitopes on the adenovirus hexon. J Gen Virol. 73 (6), 1429-1435.33120. Qiu, H., Zhou, Z., Feng, K., Tian, X., Li, X., Li, H., Xing, K., Chen, J., Li, C., Zhou, 332R., 2009. Epitope mapping and cross-reactivity analysis of the monoclonal antibodies 333against hexon protein of human adenovirus type 3. Virus Res. 146(1-2), 58-65.33421. Yuan, X.H., Wang, Y.C., Jin, W.J., Zhao, B.B., Chen, C.F., Yang, J., Wang, J.F., 335Guo, Y.Y., Liu, J.J., Zhang, D., Gong, L.L., He, Y.W., 2012. Structure-based 336high-throughput epitope analysis of hexon proteins in B and C species human 337adenoviruses (HAdVs). PLoS One. 7(3), e32938.33822. Tian X, Su X, Li X, Li H, Li T, Zhou Z, et al. Protection against Enterovirus 71 with 339Neutralizing Epitope Incorporation within Adenovirus Type 3 Hexon. PLoS One. 3402012;7(7):e41381.34123. Mahadevan, P., Seto J., Tibbetts, C., Seto, D., 2010. Natural variants of human 342adenovirus type 3 provide evidence for relative genome stability across time and 343geographic space. Virology. 397(1), 113-118.34424. Choi, E.H., Kim, H.S., Park, K.H., Lee, H.J., 2006. Genetic heterogeneity of the 345hexon gene of adenovirus type 3 over a 9-year period in Korea. J Med Virol. 78(3), 346379-383.34725. Mizuta, K., Matsuzaki, Y., Hongo, S., Ohmi, A., Okamoto, M., Nishimura, H., 348Itagaki, T., Katsushima, N., Oshitani, H., Suzuki, A., Furuse, Y., Noda, M., Kimura, H., 349Ahiko, T., 2009. Stability of the seven hexon hypervariable region sequences of 350

Page 13 of 20
Accep
ted
Man
uscr
ipt
13
adenovirus types 1-6 isolated in Yamagata, Japan between 1988 and 2007. Virus Res. 351140(1-2), 32-39.35226. Biere, B., Schweiger, B., 2010. Human adenoviruses in respiratory infections: 353sequencing of the hexon hypervariable region reveals high sequence variability. J Clin 354Virol. 47(4), 366-371.355
356

Page 14 of 20
Accep
ted
Man
uscr
ipt
14
Table 1. Primers used for the production of recombinant truncated hexon proteins.356
aNumbers show the location of the His tag or GST-fused peptides in HAdV-3 hexon 357
protein.358
Restriction sites are underlined in the primer pairs used for PCR and cloning.359
Primer pairs without underlines were annealed to generate the cloned fragments.360
361
Clone Positiona Primer Oligonucleotide sequence (5' to 3')
pGEX-R1 136-146 R1-r ggccgcagttactgcattgtctcgattcgttgtaactatgg
R1-u aattccatagttacaacgaatcgagacaatgcagtaactgc
pGEX-R2 172-183 R2-r ggccgcaatgggcttttcttctccttcagtagtggtaatgtcgg
R2-u aattccgacattaccactactgaaggagaagaaaagcccattgc
pGEX-R4 244-254 R4-r ggccgcagtttcaacccctccttcggttgttggttttactttgg
R4-u aattccaaagtaaaaccaacaaccgaaggaggggttgaaactgc
pGEX-R5 265-278 R5-r ggccgcaatttcaggcgctaaagctcctgcaacagcatccctaccatcgg
R5-u aattccgatggtagggatgctgttgcaggagctttagcgcctgaaattgc
pGEX-R6 299-307 R6-r ggccgcatttgcatgagagttatcagacgttccgg
R6-u aattccggaacgtctgataactctcatgcaaatgc
pGEX-R7 414-443 R7-r cgatgcggccgcttcattagctgtatcaacattagc
R7-u cgtgaattccggaccagggcacaggtatcaag
pGEX-R7s 414-421 R7S-r ggccgcgccttgatacctgtgccctggtccgg
R7S-u aattccggaccagggcacaggtatcaaggcgc
pGEX-R7sn 414-423 R7Sn-r ggccgctttaatgccttgatacctgtgccctggtccgg
R7Sn-u aattccggaccagggcacaggtatcaaggcattaaagc
pGEX-R8 422-433 R8-r ggccgcttcccatccattagcgtcatcggttttaactttaatgg
R8-u aattccattaaagttaaaaccgatgacgctaatggatgggaagc
pGEX-R9 433-443 R9-r ggccgcttcattagctgtatcaacattagcatctttgg
R9-u aattccaaagatgctaatgttgatacagctaatgaagc
pQE-H 112-500 H-uSphI aaagcatgcagctttaagccctattccggcac
H-rSalI aaagtcgacttactcggccgttcatgtactcgtagg

Page 15 of 20
Accep
ted
Man
uscr
ipt
15
Figure legends361
Fig.1. Analysis of MAb 1B6 specific binding to HAdV-3 hexon. (A) Western-blot 362
indicated that MAb 1B6 reacted with the naive hexon protein of HAdV-3 but not 363
HAdV-4 and HAdV-7. Purified virions HAdV-3 (lane 1), HAdV-7 (lane 2) and 364
HAdV-4 (lane 3) were separated by SDS-PAGE and then transferred to nitro-cellulose 365
membranes and incubated with MAb 1B6. (B) ELISAs indicated that MAb 1B6 reacted 366
with the virus particles and recombinant truncated hexon fragments of HAdV-3 but not 367
HAdV-4 and HAdV-7. 96-well plates were coated with the purified naive virus 368
particles of HAdV-3, HAdV-4 and HAdV-7, and recombinant truncated hexon 369
fragments of HAdV-3, HAdV-4 and HAdV-7, and then reacted with MAb 1B6. Each 370
experiment was repeated independently at least three times, and the mean values and 371
standard deviations (indicated by the error bars) are shown. (C) In vitro neutralization 372
test indicated MAb 1B6 efficiently neutralized Ad3EGFP but not rAd3egf/H7. 373
374
Fig. 2. Identification of the epitope for MAb 1B6 by ELISA. (A) Epitope mapping was 375
performed by indirect-ELISA of MAb 1B6 with 7 HVRs fusion fragments R1 to R7. 376
The identified R7 peptide was further truncated and used for additional peptide 377
scanning. The location of the respective peptide in the hexon protein is shown in Table 378
1. (B) A series of recombinant adenovirus particles with chimeric hexons were reacted379
with MAb 1B6 by indirect-ELISA. (C) Peptide competition ELISA. Increasing 380
concentrations of the identified peptides R7s, R7sn, R7 or peptide R1 (as a negative 381
control) were pre-incubated with a constant concentration of MAb 1B6, which was then 382
used to detect HAdV-3 particles bound to microtiter plates. Each experiment was 383
repeated independently at least three times, and the mean values and standard 384
deviations (indicated by the error bars) are shown.385
386
Fig. 3. Reactivity of MAb 1B6 with GST-fused peptides from different HAdV-3 strains 387
by ELISA. (A) Sequence alignment of the epitope for MAb 1B6 in the hexon protein 388
from HAdV-3 strains available from GenBank. The following 4 peptides (R7sn, V2, 389
V3, V4) represented all known variants occurring in the HAdV-3 hexon from390

Page 16 of 20
Accep
ted
Man
uscr
ipt
16
GenBank. (B) Analysis of MAb 1B6 reactivity with GST-fused proteins R7sn, V2, V3, 391
V4 by indirect-ELISA. Recombinant HAdV-3 hexon fragment was used as a positive 392
control (H), and the GST-fused fragment Ad7R7 from the corresponding region of 393
HAdV-7 was used as a negative control (Ad7R7). Each experiment was repeated 394
independently at least three times, and the mean values and standard deviations 395
(indicated by the error bars) are shown.396
397

Page 17 of 20
Accep
ted
Man
uscr
ipt
17
Highlights397
We generated a neutralizing monoclonal antibody MAb 1B6398
MAb 1B6 specifically recognized HAdV3 but not other adenovirus serotypes399
MAb 1B6 recognized amino acid position 414-424 of the HAdV3 hexon400
The neutralization epitope is the first reported for HAdV3401
402
403

Page 18 of 20
Accep
ted
Man
uscr
ipt
Figure(s)
Page 19 of 20
Accep
ted
Man
uscr
ipt
Figure(s)

Page 20 of 20
Accep
ted
Man
uscr
ipt
Figure(s)