estimation of likely cancer cure using first- and …drops/ramkipapers/ramki's papers... ·...
TRANSCRIPT

Estimation of Likely Cancer Cure Using First- and Second-Order Product
Densities of Population Balance Models
ERIC SHERER,1 ROBERT E. HANNEMANN,1,2 ANN RUNDELL,2 and DORAISWAMI RAMKRISHNA1
1School of Chemical Engineering, Forney Hall of Chemical Engineering, Purdue University, 480 Stadium Mall Way, WestLafayette, IN 47907, USA; and 2Wheldon School of Biomedical Engineering, Purdue University, West Lafayette, IN 47907,
USA
(Received 26 September 2006; accepted 30 March 2007; published online 14 April 2007)
Abstract—The objective of chemotherapy is to eradicate allcancerous cells. However, due to the stochastic behavior ofcells, the elimination of all cancerous cells must be discussedprobabilistically. We hypothesize, and demonstrate in theresults, that the mean and standard deviation of a cancer cellpopulation, derived through the probabilistic interpretationof population balance equations, are sufficient to estimate thelikelihood of cancer eradication. Our analysis of a binary celldivision model reveals that an expected cancer populationthat is six standard deviations less than one cell provides agood estimate for the treatment durations that nearly ensurestreatment successes. This approximation is evaluated andtested on two other physiologically likely scenarios: variablepatient response to chemotherapy and the presence of adormant population. We find that early identification ofindividual patient susceptibility to the chemotherapeuticagent is extremely important to all patients as treatmentadjustments for non-responders greatly enhances their like-lihood of cure while responders need not be subjected toneedlessly harsh treatments. Presence of a dormant popula-tion increases both the required treatment duration andpopulation variability, but the same estimation methodholds. This work is a step toward using stochastic modelsfor a quantitative evaluation of chemotherapy.
Keywords—Stochastic equations, Chemotherapy, Modeling.
INTRODUCTION
Oncologists are very careful to distinguish betweencancer cure and cancer remission. They prefer to utilizethe less reassuring terminology of cancer remissionpartly because the detection threshold of currentscreening techniques can be as high as 1010 cancerouscells.16 Using these technologies, it is impossible todistinguish between a cancer in remission and one thathas been cured.Hence, it is not uncommon, tomaximize
the probability of patient survival, for chemotherapytreatments to last for months or even years beyond theonset of remission with no detectable cancerous cells.
The design of cancer treatments must balance theeradication of the cancer with the patient’s generalwell-being. The cancer is likely to return if the treat-ment intensity is insufficient; however, harsher treat-ments which increase the likelihood of cure may causeintolerable collateral damage and side-effects for thepatient. The optimal treatment intensity and durationmaximizes the survival likelihood while minimizingcollateral damage and patient discomfort. As anexample, an extended period (up to 2 years) of main-tenance chemotherapy has been shown to be necessaryfor prolonged remission of childhood acute lympho-blastic leukemia.13 High drug metabolite levels areassociated with a decreased risk of relapse22 and in-creased cytotoxicity in vitro.3
To assist with the design of the optimal treatmentstrategies, mathematical models have been used toestimate cancer cell populations and the response tochemotherapy. To name a few, analytical approacheshave been used to examine the effect on specific cellularresponses such as drug resistance,11 quiescent popula-tions,2,7 spatial and diffusion effects,1,4 and cell cycle-specific drugs5,9 for use in optimal treatmentdesigns.10,12,17,24 To date, most mathematical modelsemployed to evaluate treatments are first-orderapproximations of the state of the cancerous cellpopulation; cell behavior is averaged rather than con-sidering all possible behaviors of individual cells. Be-cause each cell is governed by events whose timing isuncertain, the state of the population becomes uncer-tain. This uncertainty underlies the refusal of anoncologist to guarantee cancer cure.
Unless continuous and comprehensive measure-ments of the state are taken, the actual state can onlybe spoken of in terms of probabilities and likelihoods.This is not to say that population-averaged models are
Address correspondence to Doraiswami Ramkrishna, School of
Chemical Engineering, Forney Hall of Chemical Engineering, Pur-
due University, 480 Stadium Mall Way, West Lafayette, IN 47907,
USA. Electronic mail: [email protected]
Annals of Biomedical Engineering, Vol. 35, No. 6, June 2007 (� 2007) pp. 903–915
DOI: 10.1007/s10439-007-9310-1
0090-6964/07/0600-0903/0 � 2007 Biomedical Engineering Society
903

inept, quite the contrary. When there are a largenumber of cells, the random fluctuations are insignifi-cant once they are averaged over the entire population.However, as the cancer population approachesextinction, the behavior of individual cancerous cellsbecomes an important factor in determining thepatient’s likelihood of cure.
Fredrickson,8 in his studies of sterilization, showedthat complete eradication of a cell population cannever be guaranteed, but the likelihood of eliminationcould be computed as a function of the sterilizationparameters. He derived analytical solutions for the cellnumber probability distribution, which includes thelikelihood of complete sterility or having zero residualcells, for a homogenous population undergoing pure,independent death processes. That is, an inoculation ofN0 cells is exposed to sterilization conditions where celldivision is inhibited and death is induced. A time tlater, the probability of having N cells is given by PN(t)where the probability of complete sterility during theprocess is P0(t). Complete sterility can never be guar-anteed, but its likelihood can be computed as a func-tion of the time spent in a sterilizing environment andthe sterilizing intensity. Analyzing chemotherapytreatments are extensions of this principle. Eventhough cancer cure cannot be guaranteed, estimatingthe likelihood of curing cancer provides a quantitativebasis on which potential treatment protocols can beevaluated.
The modeling of random processes is accomplishedeither directly by random equations (e.g., the Langevinequation, Ito equation, etc.) or by deterministic dif-ferential equations in the probabilities for variousrealizations of the random process. A purely compu-tational approach based on Monte Carlo simulations(which consist of artificial realizations of the process) isalso popularly used in modeling random processes.
In what follows, we illustrate how stochastic equa-tions of population balance models19–21 can be utilizedto estimate the likelihood of complete eradication ofcancerous cells. The probabilistic interpretation ofpopulation balance equations yields the cell numberprobability distribution (which describes the likelihoodof having a given discrete number of cells) in which theprobability of having zero cancer cells is the likelihoodof cancer eradication. While, this distribution is diffi-cult to calculate directly, the continuous properties ofthe expected population mean and standard deviationobtained from the stochastic formulation give insightson the behavior of the pure probabilistic model. Thusanalysis of an age-structured binary cell division modelreveals that an expected population that is six standarddeviations less than one cancer cell provides a goodestimate for a treatment duration that will result in a
nearly 100% chance of having zero cancerous cells fora given patient.
MODEL DEVELOPMENT AND METHODS
Cure of a clinical cancer is assured only when nocancerous cells remain. Since the timing of cell divisionand death are stochastic events, the number of cells ata given time is described by a cell number probabilitydistribution. Thus, the likelihood of complete eradi-cation of cancerous cells, or the probability of cure, ata given time is the probability of having zero cells atthat time. The number of cancerous cells can be esti-mated by the expected population size (which is themean number of the cell number probability distribu-tion and thus a continuous rather than discrete quan-tity) predicted by a population balance model. Whilethe estimated population size describes whether thenumber of cancerous cells is near zero, the certaintywith which one is in this estimate is related to thestandard deviation of the population size. Wehypothesize, and demonstrate in the results, that thefirst and second moments describing the mean andstandard deviation of the cancer cell population to-gether provide an estimate of the likelihood of cancereradication.
Introduction to Number and Product Densities
Variations in the population arise due to uncer-tainty in the timing of cellular events. For example, inan unstructured model, each cell undergoes a numberof transitions at pre-specified rates. Multiplying thetransition rate by an infinitesimal time interval dt givesthe probability that an individual cell makes thattransition during that time interval. This randomnessimplies that the actual number of cells in a populationat time t, N(t), can only be known in a probabilisticsense.
In an age-structured model, the same uncertainty inthe timing of transitions exists, but the probability oftransition also depends on the time passed since a cell’slast transition, s. Neither the number of cells nor theages of those cells are known exactly. Hence, it isimpossible to know the actual number density, nðs; tÞ;or the actual number of cells exactly. This uncertaintyis contained in a master probability density function Jv,where the probability that there are v cells having agiven actual number density is
Pr nðs; tÞ ¼Xv
i¼1dðs� siÞ
( )
¼ Jvðs1; s2; . . . ; sv; tÞds1ds2 . . . dsv:
SHERER et al.904

For a detailed description of the theory, seeRamkrishna.18,19 It is possible to derive a differentialequation for this master probability density but ingeneral because of its combinatorial complexity itssolution is not often desired. However, this function isthe basis for other probabilistic functions which canbe used to describe the system in somewhat lesserdetail.
The expected number density, n1ðs; tÞ; or first-orderproduct density, is a commonly used property which isderived from the master probability density function.The first-order product density is the averaging over allage combinations of the probability density function:
E½nðs; tÞ� ¼ n1ðs; tÞ
¼X1
v¼1
1
v� 1ð Þ!Yv�1
r¼1
Z1
0
Jvðs1; s2; . . . ; sv�1; s; tÞdsr
It is of interest to note that multiplying the expectednumber density by ds represents all circumstances inwhich a cell can exist in the infinitesimal age range sand s + ds. So, n1ðs; tÞds; besides being the averagenumber of cells in the range ðs; sþ dsÞ; is also theprobability that one cell exists in that age range.
When the total number density is large, the cellbehavior can be adequately described by the deter-ministic expected number density since fluctuations incell behavior average out. Most population balancemodels are of sufficiently large systems and use for-mulations of the expected number density dynamics.However, if the number of cells is small, such as whenthe eradication of all cancerous cells is of concern, thevariation in individual cell behavior becomes impor-tant, and a more intimate knowledge of the proba-bility density function is required. Additionalinformation is contained in each of the subsequentmoments of the total population density. For exam-ple, while the expected number density also providesthe first moment, or mean, of the total populationdensity
E½NðtÞ� ¼ N1ðtÞ ¼Z1
0
n1ðs; tÞds;
deviations in the total number density are capturedthrough the variance, V[N(t)], where
V½NðtÞ� ¼ E½NðtÞ2� � E2½NðtÞ�:
Just as calculation of E[N(t)] required knowledge of thefirst-order product density, n1ðs; tÞ; calculation ofE[N(t)2] depends upon the second-order product den-sity, n2ðs; s0; tÞ: The kth-order product density is givenby:
nkðsð1Þ; sð2Þ; . . . ; sðkÞ; tÞ ¼X1
v¼k
1
v� kð Þ!
Yv�k
r¼1
Z1
0
Jvðs1; s2; . . . ; sv�k; sð1Þ; sð2Þ . . . ; sðkÞ; tÞdsr: 19
While additional information regarding the cellnumber probability distribution is contained in sub-sequent moments, these calculations for age-structuredmodels can become computationally expensive. Toestimate the likelihood of cancer eradication, the needfor the full probability distribution can be circum-vented. As shown in this article, the first and secondmoments, which yield the mean and standard deviationof the cancer cell population, are sufficient.
Age-Structured Stochastic Equations of Binary CellDivision
The mathematical models developed and utilized tostudy the cancerous cell population response to che-motherapeutic agents are based on a number ofassumptions. Foremost, the population is homoge-neous. That is, extrinsic cellular variations due tomutation or other sources are neglected. Intrinsic ra-ther than extrinsic stochastic behavior is studiedherein. Also, no external effects are considered otherthan death induced by a hypothetical chemotherapytreatment. The target of this model at present is for cellcycle non-specific drugs where there is no loss orchange in drug efficacy during treatment. The effect ofthe immune system on removing cancerous cells isneglected along with cell signaling, nutritionalrequirement, potentially evolving drug resistance, orother environmental and spatial effects.
The cellular replication of mammalian cells is binarydivision: the division of one mother cell produces twodaughter cells. In a population balance model of bin-ary division, the mother cell is removed from thenumber density while two daughter cells are simulta-neously created. Using an age-structured model wherethe age of a cell, s, is the time since its last transition,cell division produces two cells of age zero. Each celldivides at a rate CðsÞ; which is assumed to be inde-pendent of external conditions, and dies at an age-independent rate, k, which is assumed to be a fixedconstant whose magnitude is determined by intransientdrug concentrations.
The model parameter values utilized herein arebased either from in vitro cultures of established leu-kemia cell lines24 or from those observed clinically.26
For example, clinical proliferation fractions rangebetween 10 and 100%,26 hence, quiescent rates eval-uated yield proliferation fractions of 12, 54, 72, and
Estimation of Likelihood of Cancer Cure 905

93%. Since external effects on cells are neglected, thein vitro doubling times of roughly 24 h24 are used as abasis for division rates rather than those of clinicalcancers (e.g. 26.42 days for lymphomas26). Theremaining text in this section determines the first twoproduct densities which will be used to compute themean and standard deviation of the cell population.Previously, Ramkrishna and Borwanker21 have shownthat the mean and variance are functions of the firsttwo product densities of the population balanceequations.
First-Order Product Density
The population balance equations for the first-orderproduct density, n1ðs; tÞ; under binary division haveassumed that a dividing mother cell forms two new-born daughter cells of age zero. But the formation oftwo cells at the boundary contradicts the definition of anumber density where a single entity only can exist at agiven age at one time. The expected number density,n1ðs; tÞ; is actually a combination of the number den-sity for doublets formed via binary division, n21ðs; tÞ(where the superscript is not to be misconstrued as anexponent), and the number density of singlet cells thatremains after one of the doublets has transitioned, viadivision or death, n11ðs; tÞ; where n1ðs; tÞ ¼ 2n21ðs; tÞþn11ðs; tÞ (See Fig. 1).
The dynamics of the first-order product densityvector composed of the double and single numberdensities:
n1ðs; tÞ ¼n21ðs; tÞn11ðs; tÞ
� �;
is described by the population balance equation:
@n1ðs; tÞ@t
þ @n1ðs; tÞ@s
¼ Dout1 ðsÞn1ðs; tÞ ð1Þ
subject to the boundary and initial conditions:
n1ð0; tÞ ¼R1
0
Din1 ðsÞn1ðs; tÞds
n1ðs; 0Þ ¼n21ðs; 0Þn11ðs; 0Þ
� �¼ N2
1ð0Þg21ðs; 0ÞN1
1ð0Þg11ðs; 0Þ
� �
where
Dout1 ðsÞ ¼
�2 CðsÞ þ 2kð Þ 0
2 CðsÞ þ 2kð Þ � CðsÞ þ kð Þ
� �and
Din1 ðsÞ ¼
2CðsÞ CðsÞ0 0
� �:
While the literal initiation of a cancer will be a seriesof cellular events that will lead to the development of acancer, herein it is assumed that no information isavailable about the cancer until its initial clinicalobservation. So, the initial condition for the model isbased upon these macroscopic measurements. The totalnumber of cancerous cells can be determined fromdirectobservation of the cancer and the initial age distribu-tions and singlet fraction can be found using methodsdeveloped by Sherer et al.25 This method relies on BrdUpulse-labeling of cells in the S cell cycle phase to createtwo subpopulations in unbalanced growth, from whichtransition rates can be determined, amidst the balancedgrowth of the total population. The initial age distri-butions can be calculated from these transition rates.
Note that the initial number density for the i-tuple isthe product of the total number of i-tuple cells, Ni
1ðtÞ;and the age distribution, gi1ðs; tÞ; at time t=0 where:
Ni1ðtÞ ¼
Z1
0
ni1ðs; tÞds:
These total number densities define the total first-orderproduct density as demonstrated by Ramkrishna andBorwanker,21
E½NðtÞ� ¼ N1ðtÞ ¼ 2N21ðtÞ þN1
1ðtÞ:
Second-Order Product Density
The second-order product density correlates likelyage combinations of cells where n2ðs; s0; tÞdsds0 is theprobability that one cell exists in both infinitesimal ageintervals ðs; sþ dsÞ and ðs0; s0 þ dsÞ: Since a double orsingle cell is possible in each interval, all combinationsof cell multiples need be considered in the second-orderproduct density. So, the second-order product densityvector is given by
FIGURE 1. When a mother cell undergoes binary division, itis replaced by two identical daughter cells: Such doubletshave total population density n2
1ðs; tÞ. Loss of one of thedoublet cells through division or death leaves a residual sin-gle cell which is part of the single cell total population density,n1
1ðs; tÞ: All cells, regardless of affiliation, are assumed to di-vide at a rate CðsÞ and die at a rate k with the arrows denotingpossible transitions and associated rates.
SHERER et al.906

n2ðs; s0; tÞ ¼
n2;22 ðs; s0; tÞn2;12 ðs; s0; tÞn1;22 ðs; s0; tÞn1;12 ðs; s0; tÞ
266664
377775;
N2ðtÞ ¼
N2;22 ðtÞ
N2;12 ðtÞ
N1;22 ðtÞ
N1;12 ðtÞ
2
66664
3
77775¼Z1
0
Z1
0
n2ðs; s0; tÞdsds0
;
where ni;j2 ðs; s0; tÞ denotes an i-tuple of age s and aj-tuple of age s¢ at time t. Similar to the first-orderproduct density, when one of the doublet cells dividesor dies, it either forms a doublet or is removed whilethe residual cell remains that continues its pairing. Thesecond-order population balance equation andboundary conditions are
@n‘2ðs; s0; tÞ@t
þ @n2ðs; s0; tÞ
@sþ @n2ðs; s
0; tÞ@s0
¼ Dout2 ðs; s0Þn2ðs; s0; tÞ
n2ðs; 0; tÞ ¼Z1
0
Din2 ðs0Þn2ðs; s0; tÞds0 þD1ðs; tÞ
ð2Þ
respectively, where for binary division
Din2 ðs0Þ ¼
2Cðs0Þ Cðs0Þ 0 00 0 0 00 0 2Cðs0Þ Cðs0Þ0 0 0 0
2664
3775; and
D1ðs; tÞ ¼
00
2CðsÞn21ðs; tÞ0
2
664
3
775:
The inhomogeneous vector in the boundary conditionis due to the formation of a double and single pairwhenever a double divides. The dividing cell creates adouble at the boundary that is paired with the residualcell. So, while each product density equation is closedwith respect to higher order product densities, the ith-order product density is a function of the ith and(i) 1)st product densities.
If cellular independence is assumed initially, theinitial second-order product densities can be written interms of its first-order counterparts. In making a pair,one item is selected from the first-order product densityand the second item can be any of the remainder.
n2ðs; s0; 0Þ ¼N2
1ð0Þg21ðsÞðN21ð0Þ � 1Þg21ðsÞ
N21ð0Þg21ðsÞN1
1ð0Þg11ðsÞN1
1ð0Þg11ðsÞN21ð0Þg21ðsÞ
N11ð0Þg11ðsÞðN1
1ð0Þ � 1Þg11ðsÞ
2
664
3
775
In addition, cellular independence leads to initial var-iance of zero. Solution of the above system yields thesecond-order product density vector.
The general principles of product densities derivedby Ramkrishna and Borwanker21 can be applied to theabove system to yield a total second-order productdensity:
N2ðtÞ ¼ 4N2;22 ðtÞ þ 2N2;1
2 ðtÞ þ 2N1;22 ðtÞ þN1;1
2 ðtÞ;
where the expected population size and its standarddeviation are given by:
E NðtÞ½ � ¼ N1ðtÞ
Var NðtÞ½ �ð Þ1=2 ¼ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiN1
1ðtÞ þ 4N21ðtÞ þN2ðtÞ � N1ðtÞð Þ2
q :
ð3Þ
Simulation Techniques
To correlate the likelihoods of cure with the ex-pected population and the standard deviation bothMonte Carlo simulation techniques and analyticalsolutions to the first- and second-order product densityequations (Eqs. 1 and 2) are employed. The first- andsecond-order product densities yield the expectedpopulation and standard deviation but not the tran-sient likelihood of cure. The aggregate of multipleMonte Carlo simulations can be used to estimate thelikelihood of cure through time as well as the expectedpopulation and standard deviation.
The population balance equations were solved usingthe method of characteristics in order of increasingproduct density number. This method produces a ser-ies of ordinary differential equations describing eachcharacteristic curve whose origins can be traced back
Dout2 ðs; s0Þ ¼
�2 CðsÞ � Cðs0Þ � 2kð Þ 0 0 02 Cðs0Þ þ kð Þ �2CðsÞ � Cðs0Þ � 3k 0 02 CðsÞ þ kð Þ 0 �CðsÞ � 2Cðs0Þ � 3k 0
0 2 CðsÞ þ kð Þ 2 Cðs0Þ þ kð Þ �CðsÞ � Cðs0Þ � 2k
2664
3775;
Estimation of Likelihood of Cancer Cure 907

to either the initial or boundary condition. The first-order product density utilized the method of successivegenerations; the first generation is known and eachsubsequent generation can be found directly from itsparent generation.14 The second-order product densityrequired the use of integrating factors to describe theinhomogeneous differential equations of the charac-teristic curves. The resulting differential equations aresolved iteratively until the reciprocal dependence of thefinal solution and boundary condition converges.
To estimate the likelihood of cure during a treat-ment, 100 Monte Carlo simulations were performedusing the constant number simulation method ofMantzaris,15 with a maximum sample size of 106 cells,which is an expansion of the work of Shah et al.23
Briefly, the initial sample size is known, but the ages ofthe initial sample are selected randomly with weightsbased on the known initial age distributions. Thesample then undergoes a series of quiescent periodspreceding a single cellular event. The duration of eachquiescent interval is a random period based on unal-tered state paths of the cells in the system. Once theperiod is identified, the state of each cell in the systemis updated and a single cellular event is performed;randomly selected based on the state of the each cellbefore a new quiescent interval is selected.
A number of approximations were made to reducethe computations time. First, the age-dependent tran-sition rates CðsÞ correspond to normalized Gaussiandistribution division age distributions with a mean land standard deviation r6:
CðsÞ ¼ 2ffiffiffiffiffiffi2pp e
� s�lffiffi2p
r
� �2
erfc s�lffiffi2p
r
� � :
These transition rates are approximated by 15th-orderpolynomials with unnoticeable differences between theanalytical and polynomial curves. In particular, thisgreatly reduces the computational burden of thenumerous integrations over various intervals. Lastly,the age range is terminated at least four standarddeviations from the mean. Since normalized Gaussiantransition age distribution are used, this age intervalencompasses >99.8% of cell divisions while limitingthe age range under consideration to a reasonablemaximum.
RESULTS
Herein, the model of binary cell division and twovariants are analyzed for patterns of cure by employingMonte Carlo methods and solutions of the first- andsecond-order product density models. The models are
used to demonstrate that the expected population sizeand its standard deviation, which are given by first andsecond product densities according to Eq. (3), canprovide an estimate of the likelihood of cure. The firstvariant of this process considers a variable patient re-sponse to chemotherapy where the death rate is arandom variable. If patient responses to a treatmentare non-uniform; the time at which individual patientsare cured will vary significantly. Non-responding out-liers will reach cure at a much slower rate. As the non-responders have significant cancerous cell populationsfor extended durations, this dramatically influences thelikelihood of cure estimates. The second variant con-siders the possibility of cells entering and leaving aprolonged dormant state in which they are insuscep-tible to treatment yet cannot divide. This can take avariety of forms such as physical location, resistancedevelopment, or physiological state and will lead togreater population size variation.
Binary Division
The objective of this study is to identify the condi-tions on the first and second product densities whennearly all patients are cured as predicted by the MonteCarlo simulations. For a given patient, smaller expectedcancer cell populations imply that cure is more likely,but how small is small enough? In addition, the stan-dard deviation provides a sense of confidence in theexpected population, but how certain is certain enoughand does this give additional insights on cures? In theforthcoming section we examine the trends of theexpected population size and its standard deviation forqualities that can be used to approximate cure scenarios.
Expected Population
Can one simply claim cancer is cured when the pre-dicted cancer cell numbers are less than one (or somepre-defined value less than one?). Although this seemsreasonable, this study demonstrates that the expectedpopulation alone is not a reliable indicator of cure. Toillustrate the difficulties in relying upon the expectednumber, the expected cell population was found when99% of Monte Carlo simulated patients were likelycured for a small parameter space of division and deathrates (see Table 1). The expected cancerous cell popu-lations vary by nearly an order of magnitude (0.0210–0.1542 cells).If a sufficient cure is assumed to occur atan arbitrary fixed value of 0.1 cells, then some of thepatients will be under-treated with detrimental conse-quences and others over-treated which is sub-optimal(see Table 2). Using this cure threshold, the range ofcure percentages begins at 95.6% for the slow divisionand low death while the treatment for the fast division
SHERER et al.908

and slow death case is 99.7%. If a sufficient cure isassumed to occur at a lower fixed value of 0.01 cells forthe expected value, then all patients with high divisionand high death rates will be over-treated spending anextraneous 44.2% of time on medication beyond whatis considered necessary for likely cure. These discrep-ancies would likely be even greater if a broaderparameter space is sampled. Thus, an expected popu-lation appropriate for gauging likely cure for a givensituation is inestimable a priori.
Standard Deviation
If providing a mean cell population threshold aloneis insufficient to claiming cancer cure, what aboutclaiming a cancer cure when the expected cell popula-tion number is less than one and the standard deviationof that cell population is also less than one? This seemslike a more precise indicator of likely cure as this sce-nario requires both a low cancerous cell population
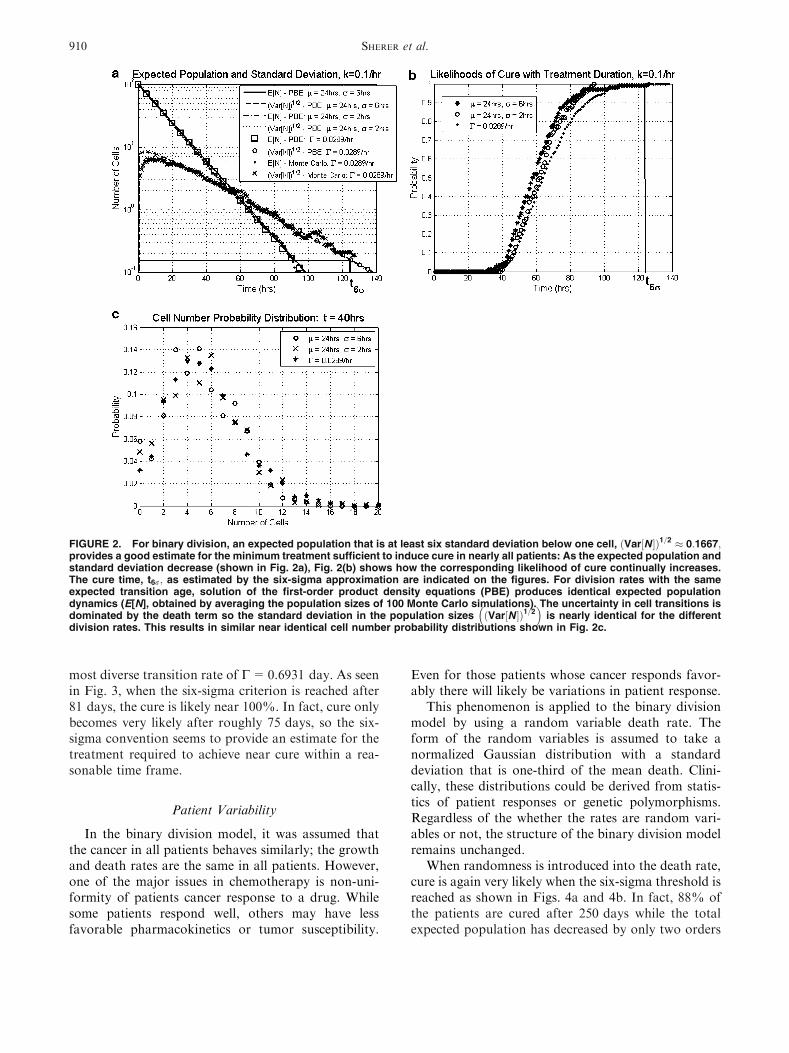
and a certainty in this estimate. However, while astandard deviation less than one indicates that patientsare entering remission; the variation in population isstill significant enough where not all patients will becured. As seen in Fig. 2, when the standard deviationand expected population are both below one, the curepercentage is roughly 80%. To find treatments neces-sary to nearly ensure cure ðP0ðtÞ � 1Þ; the range ofpopulation sizes must be restricted further. The meanand standard deviation of the population size shouldbe reduced to an appropriate level.
To identify this appropriate level, an approximationis made of the total cell number probability distribu-tion using the mean and standard deviations. It is de-sired to have a cancerous population with less than onecell. If a normalized Gaussian distribution is assumedwith a mean that is one standard deviation less thanone, then less than 67% of the distribution will be lessthan one. (The actual rates of cure are roughly 90% inFig. 2.) While the actual distribution will most likelybe of a different shape, this estimate provides a firstapproximation for the expected level of cure. Requir-ing that the mean is multiple standard deviations lessthan one only enhances the likelihood of cure.
Three transition rates, whose transition age distri-butions have identical means (24 h) but differentstandard deviations (2, 6 h, or C=0.0289 h), wereevaluated to test the impact of transition variability onthe rate of cure. The identical expected populationdynamics and minimal differences in the populationstandard deviations in Fig. 2a, lead to marginal dif-ferences in the likelihood of cure as a function oftreatment duration as seen in Fig. 2b. Though, asanticipated, the most diverse transition rate,C=0.6931 day, does asymptotically approach 100%cure most reluctantly. The similarity of total cellnumber standard deviations between division rates isdue to magnitude of the death rate, which is the samefor the division rates, relative to the division rate. Theuncertainty in the total number of cells is due mostly tothe uncertainty in the timing of the death process. Thisleads to similar cell number probability distributions(Fig. 2c) and their corresponding standard deviations.
For the cases tested in Fig. 2b, we found that curewas induced in all 100 Monte Carlo simulations if themean is six standard deviations less than one. Inter-estingly, although not surprisingly, this matches theindustrial convention of six-sigma: with high standardsof quality control, the number of defects will be min-imal. In this case, a stricter standard could be imposed,but since near cure is reached within the six-sigmaconvention; the benefits of extra treatment will beminimal. The six-sigma criterion is then tested for alarger initial cell population, 1010 cells, which is ob-served in acute myeloid leukemia16 remission, using the
TABLE 1. Comparison of treatment durations, t99, and ex-pected populations to reach at least 99% cure, E[N(t99)], for
10,000 simulated patients.
Death rates
(per day)
Division rates (per day)
C = 0.25 C = 0.75
k = 0.8 t99 = 15.4 days t99 = 129.5 days
E[N(t99)] = 0.0210 cells E[N(t99)] = 0.1542 cells
k = 1.2 t99 = 8.7 days t99 = 17.4 days
E[N(t99)] = 0.0257 cells E[N(t99)] = 0.0398 cells
Based on 10,000 Monte Carlo simulations of a binary division
model with N1(0) = 100 cells and an age-independent transition
rate, the expected cell population at which 99% of the 10,000
simulated patients is achieved, E[N(t99)], is unknown a priori. This
was found to vary by a factor of seven (0.0210–0.1542 cells) for the
division and death rate combinations tested.
TABLE 2. Using an expected number of cells to determinetreatment duration leads to potential over and under
exposures.
Death rates
(per day)
Division rates (per day)
C = 0.25 C = 0.75
k = 0.8 P0 (E[N] = 0.1) = 95.6% P0 (E[N] = 0.1) = 99.7%
%Dt = )18.8 %Dt = 7.0
P0(E[N] = 0.01) = 99.5% P0(E[N] = 0.01) = 100%
%Dt = 9.1 %Dt = 44.2
k = 1.2 P0(E[N] = 0.1) = 96.0% P0(E[N] = 0.1) = 96.7%
%Dt = )17.2 %Dt = )12.6
P0(E[N] = 0.01) = 99.7% P0(E[N] = 0.01) = 99.9%
%Dt = 10.3 %Dt = 20.5
Based on 10,000 Monte Carlo simulations of a binary division
model with N1(0) = 100 cells and an age-independent transition
rate, %Dt is the percentage difference from the optimal treatment,
the value is positive if the treatment was too long and negative if the
treatment was insufficient. P0(E[N]) is the probability of having no
cancerous cells for a given expected number of cells, E[N].
Estimation of Likelihood of Cancer Cure 909
most diverse transition rate of C=0.6931 day. As seenin Fig. 3, when the six-sigma criterion is reached after81 days, the cure is likely near 100%. In fact, cure onlybecomes very likely after roughly 75 days, so the six-sigma convention seems to provide an estimate for thetreatment required to achieve near cure within a rea-sonable time frame.
Patient Variability
In the binary division model, it was assumed thatthe cancer in all patients behaves similarly; the growthand death rates are the same in all patients. However,one of the major issues in chemotherapy is non-uni-formity of patients cancer response to a drug. Whilesome patients respond well, others may have lessfavorable pharmacokinetics or tumor susceptibility.
Even for those patients whose cancer responds favor-ably there will likely be variations in patient response.
This phenomenon is applied to the binary divisionmodel by using a random variable death rate. Theform of the random variables is assumed to take anormalized Gaussian distribution with a standarddeviation that is one-third of the mean death. Clini-cally, these distributions could be derived from statis-tics of patient responses or genetic polymorphisms.Regardless of the whether the rates are random vari-ables or not, the structure of the binary division modelremains unchanged.
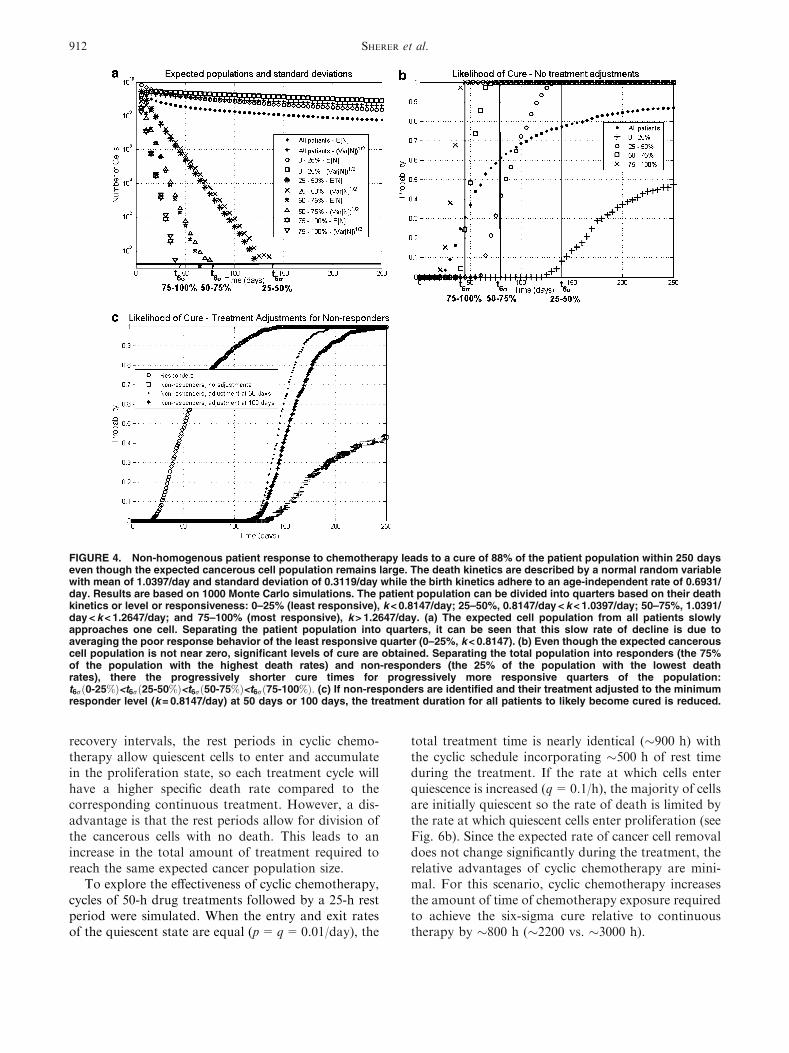
When randomness is introduced into the death rate,cure is again very likely when the six-sigma threshold isreached as shown in Figs. 4a and 4b. In fact, 88% ofthe patients are cured after 250 days while the totalexpected population has decreased by only two orders
FIGURE 2. For binary division, an expected population that is at least six standard deviation below one cell, ðVar½N �Þ1=2 � 0:1667;provides a good estimate for the minimum treatment sufficient to induce cure in nearly all patients: As the expected population andstandard deviation decrease (shown in Fig. 2a), Fig. 2(b) shows how the corresponding likelihood of cure continually increases.The cure time, t6r; as estimated by the six-sigma approximation are indicated on the figures. For division rates with the sameexpected transition age, solution of the first-order product density equations (PBE) produces identical expected populationdynamics (E[N], obtained by averaging the population sizes of 100 Monte Carlo simulations). The uncertainty in cell transitions isdominated by the death term so the standard deviation in the population sizes ðVar½N �Þ1=2
� �is nearly identical for the different
division rates. This results in similar near identical cell number probability distributions shown in Fig. 2c.
SHERER et al.910

of magnitude. This difference is due to a handful ofnon-responders to the medication with large cell pop-ulations while the majority of patients are at or nearcure. The expected cancer population for the quarter ofthe population with the lowest death rates (leastresponsive) only decreases by an order of magnitudeafter 100 days. When a distinction is made betweenresponders and non-responders, where the quarter ofthe population with the least drug activity (lowestdeath rates) is considered non-responders, there is asignificant difference in the cure levels associated withtreatments durations to reach near cure as estimatedby the six-sigma threshold (see Fig. 4b). So, while al-most all patients will likely enter clinical remissionwithin 250 days, the amount of treatment required forlikely cure is vastly different between the patients basedon their drug responsiveness: 45 days for mostresponsive quarter of the population (highest deathrates), 80 days for the second most responsive quarter,and 140 days for the third most responsive quarter (seeFig. 4a).
If the non-responders could be identified (eitherthrough measuring genetic polymorphism, relevantenzyme levels, drug concentration, or a surrogatemarker) their treatments could be adjusted accord-ingly. If this patient adjustment occurs after 100 days,230 days of treatment are needed to reach the six-sig-ma criterion while if the treatment modification can bemade after 50 days, only 195 days are needed for apatient to reach near cure based on six-sigma (seeFig. 4c). However, if the non-responders cannot bedifferentiated from the responders, then it is impossible
to tell which patients are most likely in remission andall patients must be treated identically. Treatmentoutcomes for this ‘‘average’’ patient are inefficient forboth responders and non-responders. The respondersare exposed to treatments even after they are likelycured while many non-responders are never cured dueto the finite duration of treatments. Thus, identifica-tion of the non-responders is critical as it not onlyincreases their survival, but the financial burden andpotential collateral damage of responders can be dra-matically reduced relative to treatment suggested bythe aggregate model.
Quiescence
For many cancers, there is a possibility that cellscould ‘‘escape’’ and be impervious to the drug due tospatial location, quiescent periods, or somatic muta-tions. Ideally, chemotherapy should be applied until allof the cells in hiding are removed, so though the ex-pected number of cells may be low, if some are in aquiescent state then the cure percentage may not behigh. In modeling these effects, we make the assump-tion that cells impervious to chemotherapy are com-pletely dormant in that they neither die nor divide.Also, all proliferation cells, single and paired cells,enter the dormant state as a single cell of age zero at afixed rate q and re-enter proliferation as a single cell ofage zero at a fixed rate p. These fixed transition ratesimply that dormant transitions are completely random:the likelihood of entering dormancy does not dependon the age of a cell and the likelihood of leaving dor-mancy does not depend on the time duration spent indormancy. These rates give the dynamics of the first-order product density of dormant cells, nQ1 ðs; tÞ; subjectto the initial condition n
Q1 ðs; 0Þ ¼ N
Q1 ð0Þg
Q1 ðs; 0Þ (see
Fig. 5).The effect of the dormant cancerous cell population
is revealed in the changing slope of the expected pop-ulation dynamics as shown by the lines labeled withcontinuous treatment, which represent averagedMonte Carlo simulation results, in Fig. 6a. When theentry and exit rates of the quiescent state are equal(p= q=0.01/day), the majority of cells are initially ina proliferating state, so there is a large, nearly constantrate of death. After the majority of the proliferatingcells have been exterminated, the dormant cell popu-lation dwarfs the number of proliferating cells, and therate at which the expected total population approacheszero cells depends on the rate at which the dormantcells become active. This shift in death rate corre-sponding with the emergence of the quiescent popu-lation implies a potential relative advantage for cyclicover continuous chemotherapy may arise. In additionto the likely decrease in toxic side effects due to the
FIGURE 3. The six-sigma estimate of cure time, t6r, is eval-uated for a model of acute myeloid leukemia (based on 100Monte Carlo simulations). The binary division model is ap-plied to an initial population of 1010 cells with C = 0.6391/dayand k = 1.0397/day.
Estimation of Likelihood of Cancer Cure 911
recovery intervals, the rest periods in cyclic chemo-therapy allow quiescent cells to enter and accumulatein the proliferation state, so each treatment cycle willhave a higher specific death rate compared to thecorresponding continuous treatment. However, a dis-advantage is that the rest periods allow for division ofthe cancerous cells with no death. This leads to anincrease in the total amount of treatment required toreach the same expected cancer population size.
To explore the effectiveness of cyclic chemotherapy,cycles of 50-h drug treatments followed by a 25-h restperiod were simulated. When the entry and exit ratesof the quiescent state are equal (p= q=0.01/day), the
total treatment time is nearly identical (�900 h) withthe cyclic schedule incorporating �500 h of rest timeduring the treatment. If the rate at which cells enterquiescence is increased (q=0.1/h), the majority of cellsare initially quiescent so the rate of death is limited bythe rate at which quiescent cells enter proliferation (seeFig. 6b). Since the expected rate of cancer cell removaldoes not change significantly during the treatment, therelative advantages of cyclic chemotherapy are mini-mal. For this scenario, cyclic chemotherapy increasesthe amount of time of chemotherapy exposure requiredto achieve the six-sigma cure relative to continuoustherapy by �800 h (�2200 vs. �3000 h).
FIGURE 4. Non-homogenous patient response to chemotherapy leads to a cure of 88% of the patient population within 250 dayseven though the expected cancerous cell population remains large. The death kinetics are described by a normal random variablewith mean of 1.0397/day and standard deviation of 0.3119/day while the birth kinetics adhere to an age-independent rate of 0.6931/day. Results are based on 1000 Monte Carlo simulations. The patient population can be divided into quarters based on their deathkinetics or level or responsiveness: 0–25% (least responsive), k < 0.8147/day; 25–50%, 0.8147/day < k < 1.0397/day; 50–75%, 1.0391/day < k < 1.2647/day; and 75–100% (most responsive), k > 1.2647/day. (a) The expected cell population from all patients slowlyapproaches one cell. Separating the patient population into quarters, it can be seen that this slow rate of decline is due toaveraging the poor response behavior of the least responsive quarter (0–25%, k < 0.8147). (b) Even though the expected cancerouscell population is not near zero, significant levels of cure are obtained. Separating the total population into responders (the 75%of the population with the highest death rates) and non-responders (the 25% of the population with the lowest deathrates), there the progressively shorter cure times for progressively more responsive quarters of the population:t6r 0-25%ð Þ<t6r 25-50%ð Þ<t6r 50-75%ð Þ<t6r 75-100%ð Þ: (c) If non-responders are identified and their treatment adjusted to the minimumresponder level (k = 0.8147/day) at 50 days or 100 days, the treatment duration for all patients to likely become cured is reduced.
SHERER et al.912

With the presence of multiple stages, such as qui-escent and proliferating cells, the age distributionsbecome transient even when an age-independent deathrate is applied. While the specific division rate isindependent of changes in the age distribution for theage-independent division rate studied above, for age-dependent division rates, the transient age distribu-tions do become significant. This leads to fluctuationsin the specific growth rate which produces periodicoscillations in the expected number density (seeFigs. 6c and 6d). However, the validity of the six-sigma approximation for minimal sufficient treatmentis not affected by the increase in population variation(see Figs. 6e and 6f). Regardless of the age depen-dence or treatment type, when comparing the times tocure, as estimated using the six-sigma threshold, thepresence of a dormant population increases thetreatment duration (Figs. 6e and 6f) necessary toreach a desired level of cure relative to non-dormantmodels (Figs. 2–4).
DISCUSSION
The idealistic objective of chemotherapy is to cureall patients, but this would require chemotherapytreatments that are unreasonable in duration or dos-age. A practical alternative is to maximize survivalrates subject to patient limits. These limits could in-clude factors that affect patient discomfort such as:drug dosage and duration, monetary costs, and phys-ical well-being where the marginal benefit of over-treatment must be weighed against all costs involved.To this end both over-treatment and under-treatmentare non-optimal situations and the question becomes
one of identification of the appropriate treatmentstrategy that maximizes the cancer survival rates; orrather, one that minimizes treatment burden while stillnearly ensuring cure. However, calculation of the cellnumber probability distribution required for such ananalysis is difficult. To this end, we investigated whe-ther an approximation of the system state, as describedby the expected population size and its standarddeviation, is sufficient to identify treatments that sat-isfy a more qualitative objective: almost ensure a givenpatient is cured without undue burden assuming thatwhether or not the necessary treatments are feasible isanother matter altogether.
Using a binary cell division model system, andoperating under the assumptions that all cells in apatient are governed by the same set of kinetics andexternal influences other than death due to chemo-therapy can be neglected, we found that the expectedpopulation size and its standard deviation can identifytreatments that cure nearly all patients. A small can-cerous cell population is desired, but the expectedpopulation alone is a poor prognosticator; confidencethat the population is small is required. Based on ourmodel predictions, we found that a mean that is onestandard deviation below one cancerous cell indicatesthat some patients have been cured, and a mean that issix standard deviations below one cell correlates wellwith extremely high cure percentages. Notably, thissix-sigma criterion does not necessitate prolongedchemotherapy treatments.
When the binary division model was applied to ascenario where patient response was non-uniform, wefound that identification of non-responders is ex-tremely beneficial to both the responders and non-responders. If patients are treated equivalently, allpatients are subjected to lengthy treatments whichlikely may not end in cure. If a patient’s response to atreatment can be quantified, treatment adjustments canbe made for non-responders so they will be more likelyto enter remission and for responders so they will nothave to endure unnecessary periods of treatment.Lastly, the presence of a temporarily dormant andnon-responsive tumor population lengthens the nec-essary period of chemotherapy as treatment must begiven until no cells are dormant and all proliferatingcells are removed. In such a case, cyclic chemotherapymay have potential relative advantage (i.e., similarresults for less cumulative drug administration or elsesimilar cumulative drug administration with the benefitof recovery periods) over continuous infusions. Theperiod of time spent in dormancy is a very uncertainevent and leads to greater population variation. This isespecially prevalent for low expected population sizes.However, the six-sigma criterion still identifies treat-ments that are very likely to result in cure.
FIGURE 5. Binary division model modified with the additionof a quiescent state. If cells can become temporarily dormant,cell behavior will depend on the whether a cell is dormant,nQ
1 ðs; tÞ; or actively proliferating, n21ðs; tÞ or n1
1ðs; tÞ: Arrowsdenote possible transitions and associated rates: entry rate tothe dormant state, q, re-entry rate to proliferation, p, deathrate, k, proliferation rate, CðsÞ.
Estimation of Likelihood of Cancer Cure 913

While this present work is limited to cell cycle non-specific treatment with intransient rate functions toensure the clarity of the illustrative examples, themodel framework can be adjusted for time-dependentfunctions. (Furthermore, the solution methodologywould not require any modifications.) There are manyfactors that would lead to transient birth or death ratesincluding drug schedules and evolving drug resistance.For example, drug schedules could produce transientdrug levels that dynamically alter the birth and deathrates. Sherer et al.24 use constitutive pharmacokineticmodel to predict a changing exposure to toxic agentswhile a pharmacodynamic model translates these druglevels into a dynamic, and cell cycle-specific, cell death
for an age-structured model. A similar constitutivepharmacodynamic model could be written describingthe changes in the birth rate due to drug exposures.Alternatively, cells may develop drug resistance or up/down regulate pathways in response to a drug whichwould affect the efficacy of subsequent treatments. Thedevelopment of drug resistance could be accounted forin the model by including discrete resistance levels10 orby introducing a second continuous variable whichaccounts for the observed or anticipated changes inpharmacodynamics.
The model situations presented above are grosssimplifications of the real problem but they enablethe exploration of utilizing stochastic mathematical
FIGURE 6. The six-sigma criterion holds for a binary division model with a quiescent cell population: based on the aggregate of1000 Monte Carlo simulations. The figures in the left column (a, c, and e) contain simulations with a low rate of transition in bothdirections between quiescence and proliferation (p = q = 0.01/day). The figures in the right column (b, d, and f) contain simulationresults for a higher rate of transition to quiescence (p = 0.01/day, q = 0.1/day). (a, b) The division rate is a fixed constant, C = 0.0289/h, while the death rate is k = 0.1/h when the drug is applied. For a continuous application of chemotherapy, the initially steepdecline in the expected population becomes tempered as the fraction of dormant cells becomes prominent. This occurs morerapidly in figure (b) due to the shorter residence time in the proliferation state. Eventually, the lifetime in dormancy limits the rate ofcure. In cyclic chemotherapy, 50 h boluses are separated by a 25 h rest period. (c, d) The division rate corresponds to a normalizedGaussian division age-distribution of mean of 24 h and standard deviation of 4 h while the death rate is k = 0.1/h. (e and f) There is adifference in the likelihood of cure as defined by the six-sigma threshold between age-structured and unstructured models. Acyclic chemotherapy schedule where 50-h treatments are separated by a 25-h rest period may have marginal benefits in certaincases (e) but not in others (f).
SHERER et al.914

models to estimate cure. The question remains,though: how will this approach be applied to a realsystem? What is the appropriate model framework andwhat are the model parameters? In addition, potentialclinical treatments will be constrained by a patient’shealth, so the treatments required to achieve likely cureare not feasible. These are non-trivial problems as themodel framework will likely vary between diseases,patient response are difficult to predict, and key rateparameters are difficult to measure.
The application of this approach would come in theform of an inverse model. That is, the model itselfwould need to be matched to accumulated patient data.In this case, the most basic data required would includetreatment protocols and the distribution of durations inwhich patients remained in remission. Potential modelframeworks and rate parameters could be identifiedthat matched the patient data. However, any patientdiscrimination data such as drug concentration or tu-mor proliferation indices or genetic predispositionscould also be utilized (and may be necessary) in modeland parameter refinement. Once the model is identifiedfor a cancer, predictions could be made (as exemplifiedin this paper) on an individual basis for incoming pa-tients. As mentioned early, the clinically related studiesby Lillyman and Lennard13 and Schmiegelow et al.22
and the in vitro work of Bokkerink et al.3 hint at thepositive relationship between metabolite levels, thecorresponding death rates, and the resulting probabilityof cure. By modeling this system, the appropriatedurations of treatment or treatment adjustments can bemade quantitatively for individual patients. While themethodology presented in this work is not immediatelyready for clinical application, it does provide a startingpoint for which more quantitative analyses of cancertreatments can be based.
REFERENCES
1Adam, J. A. A simplified mathematical model of tumorgrowth. Math. Biosci. 81:229–244, 1986.2Andersen, L. K., and M. C. Mackey. Resonance in peri-odic chemotherapy: A case study of acute myelogenousleukemia. J. Theor. Biol. 209:113–130, 2001.3Bokkerink, J. P. M., E. H. Stet, R. A. De Abreu, F. J. M.Damen, T. W. Hulscher, M. A. H. Bakker, and J. A. VanBaal. 6-Mercaptopurine: Cytotoxicity and biochemicalpharmacology in human malignant T-lymphoblasts. Bio-chem. Pharmacol. 45:1455–1463, 1993.4Chaplain, M. A. J. Avascular growth, angiogenesis andvascular growth in solid tumours: The mathematicalmodeling of the stages of tumour development. Math.Comput. Model. 23:47–87, 1996.5Cojocaru, L., and Z. Agur. A theoretical analysis ofinterval drug dosing for cell-cycle-phase-specific drugs.Math. Biosci. 109:85–97, 1992.
6Eakman, J. M., A. G. Fredrickson, and H. M. Tsuchiya.Statistics and dynamics of microbial cell populations.Chem. Eng. Prog. Symp. Ser. 62:37–49, 1966.7Fister, K. R., and J. C. Panetta. Optimal control appliedto cell-cycle-specific cancer chemotherapy. SIAM J. Appl.Math. 60:1059–1072, 2000.8Fredrickson, A. G. Stochastic models for sterilization.Biotechnol. Bioeng. 8:167–182, 1966.9Gaffney, E. A. The application of mathematical modelingto aspects of adjuvant chemotherapy scheduling. J. Math.Biol. 48:375–422, 2004.
10Gardner, S. N. Modeling multi-drug chemotherapy: Tai-loring treatments to individuals. J. Theor. Biol. 214:181–207, 2002.
11Goldie, J. H., and A. J. Coldman. Drug Resistance inCancer. Cambridge: Cambridge University Press, 1998.
12Iliadis, A., and D. Barbolosi. Optimizing drug regimens incancer chemotherapy by an efficacy-toxicity mathematicalmodel. Comput. Biomed. Res. 33:211–226, 2000.
13Lillyman, J. S., and L. Lennard. Mercaptopurine metab-olism and risk of relapse in childhood lymphoblastic leu-kaemia. The Lancet 343:1188–1190, 1994.
14Liou, J. J., F. Srienc, and A. G. Fredrickson. Solutions ofpopulation balance models based on a successive genera-tions approach. Chem. Eng. Sci. 52:1529–1540, 1997.
15Mantzaris, N. V. Stochastic and deterministic simulationsof heterogeneous cell population dynamics. J. Theor. Biol.241:690–706, 2006.
16Michalek, J., and J. Smarda. Detection of minimal residualdisease in acute myeloid leukemia. Scripta Med. 73:223–228, 2000.
17Murray, J. M. Some optimal control problems in cancerchemotherapy with a toxicity limit. Math. Biosci. 100:49–67, 1990.
18Ramakrishna, D.Statistical Models of Cell Populations. In:Advances in Biochemical Engineering, Vol 11, edited byA. Fiechter and T. K. Ghose. Berlin: Springer Verlag, 1979,pp. 1–47.
19Ramkrishna, D. Population Balances: Theory and Appli-cation to Particulate Systems in Engineering. San Diego:Academic Press, 2000, 355 pp.
20Ramkrishna,D., and J.D. Borwanker. Apuristic analysis ofpopulation balance – I.Chem. Eng. Sci. 28:1423–1435, 1973.
21Ramkrishna,D., and J.D. Borwanker. Apuristic analysis ofpopulation balance – II.Chem.Eng. Sci. 29:1711–1721, 1974.
22Schmiegelow, K., H. Schrøder, G. Gustafsson, J. Kris-tinsson, A. Glomstein, T. Salmi, and L. Wranne. Risk ofrelapse in childhood acute lymphoblastic leukemia is re-lated to RBC methotrexate and mercaptopurine metabo-lites during maintenance chemotherapy. J. Clin. Oncol.13:345–351, 1995.
23Shah, B. H., J. D. Borwanker, and D. Ramkrishna. MonteCarlo simulation of microbial population growth. Math.Biosci. 31:1–23, 1976.
24Sherer, E., R. E. Hannemann, A. Rundell, and D. Ram-krishna. Analysis of resonance chemotherapy in leukemiatreatment via multi-staged population balance models. J.Theor. Biol. 240:648–661, 2006.
25Sherer, E., R. E. Hannemann, A. Rundell, E. Tocce, andD. Ramkrishna. Determination of age-dependent cell cycletransition rates. Submitted.
26Steel, G. G. Growth Kinetics of Tumours: Cell PopulationKinetics in Relation to the Growth and Treatment ofCancer. Oxford: Clarendon Press, 1977, 351 pp.
Estimation of Likelihood of Cancer Cure 915