estudo quÍmico e avaliaÇÃo da atividade …livros01.livrosgratis.com.br/cp100725.pdf · carinho,...
TRANSCRIPT

UNIVERSIDADE FEDERAL DA PARAÍBA
LABORATÓRIO DE TECNOLOGIA FARMACÊUTICA
PROGRAMA DE PÓS-GRADUAÇÃO EM PRODUTOS NATURAIS
E SINTÉTICOS BIOATIVOS
ESTUDO QUÍMICO E AVALIAÇÃO DA ATIVIDADE
HIPOGLICEMIANTE E ANTIOXIDANTE DE Bauhinia
cheilantha (Bong.) Steudel.
ADRIANA MARIA FERNANDES DE OLIVEIRA
JOÃO PESSOA 2008

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

ADRIANA MARIA FERNANDES DE OLIVEIRA
ESTUDO QUÍMICO E AVALIAÇÃO DA ATIVIDADE
HIPOGLICEMIANTE E ANTIOXIDANTE DE Bauhinia
cheilantha (Bong.) Steudel.
Dissertação apresentada ao Programa de Pós-
graduação em Produtos Naturais e Sintéticos
Bioativos da Universidade Federal da Paraíba,
em cumprimento às exigências para a
obtenção do título de Mestre em Produtos
Naturais e Sintéticos Bioativos. Área de
Concentração: Farmacologia
ORIENTADOR (a): Profa. Dra. Temilce Simões de Assis
CO- ORIENTADOR (a): Profa. Dra. Tania Maria Sarmento da Silva
JOÃO PESSOA
2008

ADRIANA MARIA FERNANDES DE OLIVEIRA
ESTUDO QUÍMICO E AVALIAÇÃO DA ATIVIDADE
HIPOGLICEMIANTE E ANTIOXIDANTE DE Bauhinia
cheilantha (Bong.) Steudel.
Aprovada em: 07/03/2008
Banca examinadora
Profa. Dra. Temilce Simões de Assis (Orientadora)
Profa. Dra. Tania Maria Sarmento da Silva (Co-orientadora)
Prof. Dr. Luciano Augusto de Araújo Ribeiro (Examinador externo)
Profª. Dra. Liana Clébia Soares Lima de Moraes (Examinadora
interna)

Aos meus Pais, Rijalma e Neuman,
pelo incentivo, apoio e confiança.

AGRADECIMENTOS À Deus, que me dá forças nas horas de desânimos, por iluminar-me nas horas
das escolhas, por conduzir-me confiante na concretização deste trabalho. Obrigada,
Senhor...
Aos meus pais, Rijalma e Neuman, por respeitarem as minhas escolhas, pelo
carinho, pela compreensão, apoio e presença na conquista de meus ideais;
Aos meus irmãos, Andrea e Rijalma Júnior, por serem meus eternos
companheiros e amigos, e que sempre estiveram ao meu lado, ajudando-me, mesmo que
silenciosamente, na conclusão deste trabalho;
A todos os meus familiares, especialmente meu tios Sarmento e Norma e minha
primas pela acolhida e incentivo;
A Sócrates Golzio pelo amor, dedicação e carinho durante todo o período de
realização deste trabalho;
À minha orientadora, Profa. Dra. Temilce Simões de Assis, pela acolhida,
confiança, paciência e por todo o conhecimento transmitido;
À co-orientadora, Profa. Dra. Tânia Maria Sarmento, por todo o auxílio e
dedicação;
Aos meus professores da Pós-Graduação, especialmente os professores Eduardo
de Jesus Oliveira, Emídio Vasconcelos Leitão da Cunha, Bagnólia Araújo, Fátima
Wanderlei, Reinaldo Nóbrega pelo conhecimento transferido, por auxiliar-me na busca
da realização de meus ideais profissionais;
À equipe do Laboratório de Farmacologia Funcional Prof. George Thomas pela
acolhida e conhecimentos compartilhados;

À equipe do Prof. Marcelo Sobral da Silva, pela receptividade e amizade que
sempre me dedicaram;
À equipe do Laboratório de Psicofarmacologia Prof. Elizaldo Carlini pela
acolhida;
À banca examinadora, Prof. Luciano Ribeiro e Liana Clébia pela disponibilidade
com que aceitaram realizar esse trabalho;
À turma de mestrado pelas conquistas alcançadas e pelos obstáculos superados;
Aos amigos de laboratório Ana Paloma, Antonio Cláudio, Stanley Chavez,
Kristerson Reinaldo, Maria, Kiriak, Wilson, Fernando Oliveira, Cibele pelo clima de
amizade e alegria que sempre mantiveram no ambiente de trabalho. Um agradecimento
especial a Roberto Jefferson pela orientação e amizade, durante esse tempo de trabalho;
Aos amigos Aline, Camila, Gabriela, Marcela, Steno,Vivianne pela amizade
construída;
Às amigas Anna Cláudia, Danielle e Vanine pelo companheirismo e amizade
durante todo esse tempo de convivência;
Aos amigos Fernanda, Nádja, Tayse, Aldo que, mesmo distante, sempre
torceram pela minha vitória;
Aos funcionários e técnicos do LTF, especialmente Tânia Maria, Raimundo
Nonato, Vicente Carlos, Ataide e Alexsandro Marinho pela ajuda na realização deste
trabalho;
Ao funcionário e amigo José Crispim Duarte cuja ajuda e confiança foi
imprescindível para a realização deste;
Ao LTF, CAPES e FAPESP pelo apoio financeiro;

A todos que contribuíram de alguma forma para que este trabalho pudesse ser
realizado.

SUMÁRIO
Resumo
Abstract
Lista de Tabelas……………………………………………..…………………….. I
Lista de Figuras........................................................................................................ II
Lista de Gráficos...................................................................................................... III
Lista de Fluxograma................................................................................................ IV
Lista de abreviaturas............................................................................................... V
INTRODUÇÃO…………………………………………………………………… 2
FUNDAMENTAÇÃO TEÓRICA
1. Considerações sobre o gênero Bauhinia..................................................... 6
2. Pâncreas.............................................................................................................. 8
2.1. Anatomia....................................................................................................... 8
2.2. Fisiologia....................................................................................................... 9
3. Insulina................................................................................................................ 10
3.1. Estrutura......................................................................................................... 11
3.2. Síntese............................................................................................................ 12
3.3. Secreção......................................................................................................... 14
3.4. Padrão bifásico de secreção da insulina.......................................................... 16
3.5. Mecanismo de ação da insulina...................................................................... 17
3.6. Distribuição e degradação da insulina......................................................... 18
3.7. Ações da insulina............................................................................................ 19
3.8. Glicose no sangue........................................................................................... 20
4. Diabetes Mellitus.............................................................................................. 23
4.1. Conceito.......................................................................................................... 23
4.2. Breve histórico................................................................................................ 23
4.3. Prevalência e Epidemiologia........................................................................... 24
4.4. Classificação................................................................................................... 25
4.4.1. DM Tipo 1................................................................................................ 25
4.4.2. DM Tipo 2................................................................................................ 26
4.4.3. MODY (Diabetes de maturidade com início na juventude ou tipo
1,5)............................................................................................................................. 27

4.4.4. Diabetes mellitus Gestacional.................................................................. 28
4.5. Diagnóstico de DM........................................................................................ 29
4.6. Sintomas......................................................................................................... 30
4.7. Complicações ................................................................................................ 31
4.8. Tratamento..................................................................................................... 32
4.9. Diagnóstico, prevenção e possível cura......................................................... 36
4.10. Diabetes experimental.................................................................................. 38
4.10.1. Aloxana.................................................................................................. 39
4.10.2. Estreptozotocina (STZ).......................................................................... 41
4.11. Diabetes mellitus e as plantas...................................................................... 44
5. Antioxidante.......................................................................................................
5.1. Introdução...................................................................................................... 48
5.5.1. Atividade seqüestradora do radical DPPH•.............................................. 55
5.5.2. . Teor de fenólicos totais........................................................................... 57
5.5.3. Capacidade antioxidante equivalente ao trolox (CAET).......................... 58
OBJETIVOS
1. OBJETIVO GERAL.......................................................................................... 60
2. OBJETIVOS ESPECÍFICOS............................................................................. 60
METODOLOGIA
1. Estudo químico da planta................................................................................... 62
2. Estudo farmacológico........................................................................................ 65
3. Estudo antioxidante............................................................................................ 67
3.1. Atividade seqüestradora do radical
DPPH•.................................................
67
3.2. Teor de fenólicos
totais..............................................................................
68
3.3. Capacidade antioxidante equivalente ao trolox (CAET)........................... 69
4. Análise Estatística.............................................................................................. 70
RESULTADOS
1. Identificação estrutural ...................................................................................... 72
2. Toxicidade aguda...............................................................................................
3. Atividade hipoglicemiante.................................................................................
83
83
4. TESTES ANTIOXIDANTES

4.1. Atividade seqüestradora do radical DPPH ..................................................... 90
4.2. •Total de fenólicos Totais (FT)....................................................................... 94
4.3. Capacidade equivalente ao Trolox.................................................................. 96
DISCUSSÃO............................................................................................................. 99
CONCLUSÕES........................................................................................................ 106
PERSPECTIVAS..................................................................................................... 108
REFERÊNCIAS....................................................................................................... 110
ANEXO

RESUMO
ESTUDO QUÍMICO E AVALIAÇÃO DA ATIVIDADE HIPOGLICEMIANTE E ANTIOXIDANTE DE Bauhinia cheilantha (Bong.) Steudel.
Adriana Maria Fernandes de Oliveira Mestrado em Produtos Naturais e Sintéticos Bioativos (Farmacologia)
Laboratório de Tecnologia Farmacêutica, UFPB, Caixa Postal 5009, Cep 58051-970 João Pessoa, Paraíba, Brasil
Bauhinia cheilantha (Bong) Steudel. (MSE), é uma planta pertencente à família das Leguminosas, vulgarmente conhecida como “Pata-de-vaca”, “Mororó” ou “Unha-de-boi” devido ao formato de suas folhas. As plantas do gênero Bauhinia são conhecidas para o tratamento de algumas enfermidades, onde a mais conhecida é o diabetes mellitus (DM). O objetivo deste trabalho foi estudar a atividade hipoglicemiante induzindo DM tipo 1 com aloxana, em camundongos, e correlacionar com a atividade antioxidante de MSE através da determinanção do teor de fenólicos totais (FT), da atividade sequestradora do radical DPPH·+ e da capacidade antioxidante equivalente ao Trolox (CAET - ABTS). Foram realizadas 3 coletas nos meses de janeiro de 2005, agosto 2006 e fevereiro de 2007, onde a partir do extrato bruto etanólico, foram obtidos três frações: fração aquosa, fração hexânica e fração acetato de etila. As amostras foram testadas nas doses de 300, 600 e 900 mg/kg e administrada pelas vias i.p. e oral (gavage). O extrato bruto da primeira coleta reduziu a glicemia de camundongos diabéticos em média 46 % da glicemia, as frações obtidas do extrato não foram testadas. O extrato da 2a coleta não apresentou atividade significativa. O extrato bruto da 3a coleta apresentou uma redução dos níveis glicêmicos de forma expressiva em todas as doses testadas, sendo que a dose de 300 mg/Kg houve uma redução em todos os tempos. A atividade antioxidante determinada pelo teste do DPPH da 1a coleta (CE50 = 11,67 ± 1,43 µg/mL), da 2a coleta (CE50 = 26,68 ± 0,23 µg/mL) e 3a coleta (CE50 = 21,77 ± 0,126 µg/mL). As frações hexânicas das 3 coletas não apresentaram efeito hipoglicemiante. A fração aquosa da 3a coleta reduziu a glicemia de animais diabético na dose de 300 mg/Kg em 12,3 ± 3,9 %; representados no gráfico de barras. A fração acetato de etila não reduziu a glicemia. Todas as frações ou extrato bruto que apresentaram efeito biológico foram administrados por gavage, exceto a o extrato bruto da 2 a coleta (i.p.). Na 2a coleta, a fração acetato de etila apresentou a melhor atividade antioxidante (DPPH – [CE50 = 9,32 ± 0,081 µg/mL] e ABTS – [CE50 = 2,9 ± 0,05 µg/mL]) e FT (341,55 ± 1,41 EAG/g). O precipitado MeOH apesar de não apresentar atividade hipoglicemiante, apresentou a melhor atividade antioxidante (DPPH – [CE50 = 9,35 ± 0,09µg/mL] e ABTS – [CE50 = 5,24 ± 0,13µg/mL]) e FT (540,27 ± 22,42 EAG/g) na 3a coleta, seguido pela fração acetato de etila. Palavras-chave: Bauhinia cheilantha, diabetes, antioxidante.

ABSTRACT
Bauhinia cheilantha (Bong) Steudel. (MSE), is a plant which belongs to the Leguminosae family, popularly known as Pata-de-vaca”, “Mororó” or “Unha-de-boi” due to the shape of its leaves. Plants of the Bauhinia genus are used in the treatment of several illnesses, which the most recognized is diabetes mellitus (DM). The aim of this work was study the hypoglycemic activity of the plant, inducing type 1 DM with alloxan, in mice and compare the antioxidative activity of MSE through: Determination of total phenolic content (TF), DPPH• radical scavenging assay or ABTSH• radical cation scavenging. Leaves of MSE were collected on the following months: January, 2005; august, 2006 and February 2007. From the ethanol crude extract, three fractions were obtained: aqueous fraction, hexane fraction and acetate fraction. The samples were tested on the following doses: 300, 600 or 900 mg/kg, administered via i.p. or orally (gavage). The first plant collecting crude extract reduced glycemia levels by 46 %. Fractions derived from this extract were not tested. Crude extracts from the second plant collecting did not present significant hypoglycemic activity. The crude extract from the 3rd collecting presented a reduction of glycemia in all doses tested on the 180 min. The dose of 300 mg/Kg showed a hipoglicemiant effect on the following times: 60, 120 and 180 min. The antioxidant activity determined by the DPPH• test of the 1st collecting CE50 = 11.67 ± 1.43 µg/mL), 2nd collecting (CE50 = 26.68 ± 0.23 µg/mL) and 3rd collecting (CE50 = 21.77 ± 0.126 µg/mL). The hexane fractions from all three collecting did not presented hypoglycemic effect. The aqueous fraction from the 3rd collecting reduced glycemia of diabetic animals on the dose of 300 mg/Kg by 12.3 ± 3.9 %. The ethyl acetate fraction did not present any hypoglicemiant effect.. All crude extract or fractions which show biological effect were administered by gavage, except the crude extract (i.p.), 2nd collecting The ethyl acetate fraction obtained from the 2nd collecting, presented the best antioxidant activity (DPPH – [CE50 = 9.32 ± 0.081 µg/mL] or ABTS – [CE50 = 2.9 ± 0.05 µg/mL]) and FT (341.55 ± 1.41 EAG/g). Although the methanol precipitate did not present a hypoglycemic activity, it has the best antioxidant activity (DPPH – [CE50 = 9.35 ± 0.09µg/mL] or ABTS – [CE50 = 5.24 ± 0.13µg/mL]) and FT (540.27 ± 22.42 EAG/g) from the 3rd collecting followed by the ethyl acetate fraction. Key words: Bauhinia cheilantha, diabetes, antioxidant.

LISTA DE TABELAS
Tabela 1: Resumo dos efeitos da insulina sobre o metabolismo de carboidratos,
lipídios e proteínas no fígado, músculo e tecido adiposo......................................... 20
Tabela 2: Transportadores teciduais de glicose....................................................... 22
Tabela 3: Total de pessoas com diabetes em alguns países da América do Sul,
Central e do Norte.................................................................................................... 24
Tabela 4: Características clínicas de pacientes com Diabetes mellitus tipo 1 e 2 27
Tabela 5: Critérios para o diagnóstico de diabetes.................................................. 29
Tabela 6: Diagnóstico do diabetes mellitus e alterações da tolerância á glicose
de acordo com valores de glicose plasmática (mg/dl).............................................. 30
Tabela 7: Fatores de risco para o diabetes mellitus................................................. 30
Tabela 8: Tipos de insulina..................................................................................... 33
Tabela 9: Perfil farmacológico e terapêutico dos fármacos mais comumente
utilizados no tratamento do diabetes........................................................................ 34
Tabela 10: Famílias das plantas que possuem atividade hipoglicemiante.............. 44
Tabela 11: Plantas brasileiras antidiabéticas........................................................... 46
Tabela 12: Dados de RMN de 1H (400 MHz) e RMN de 13C (100 MHz) do
flavonóide 1 (afzelina) em DMSO........................................................................... 74
Tabela 13: Dados de RMN de 1H (400 MHz) e RMN de 13C (100 MHz) do
flavonóide 2 (quercitrina) em DMSO...................................................................... 80

LISTA DE FIGURAS Figura 1: Foto de uma espécie do gênero Bauhinia.................................................. 6
Figura 2: Formato das folhas de plantas do gênero Bauhinia................................... 7
Figura 3: Exsicata de B. cheilantha........................................................................... 7
Figura 4: Flavonóide Caempferol-3,7-O-(r)-diraminosideo..................................... 8
Figura 5: Anatomia do Pâncreas............................................................................... 9
Figura 6: Distribuição das células na ilhota de Langerhans...................................... 10
Figura 7: Estrutura da insulina: A. Molécula da pró insulina humana; B. Molécula
da insulina dobrada..................................................................................................... 12
Figura 8: Síntese de insulina..................................................................................... 14
Figura 9: Modelo para acoplamento do metabolismo da glicose e secreção de
insulina nas células β.................................................................................................. 16
Figura 10: Receptor da insulina................................................................................ 18
Figura 11: Modelo de um transportador de glicose.................................................. 21
Figura 12: Estrutura química da aloxana............................................................ 39
Figura 13: Estrutura da Estreptozotocina (STZ)....................................................... 41
Figura 14: Mecanismos propostos da toxicidade induzida pela STZ. Dois
mecanismos diretos: (I) metilação do DNA, induzida por CH3+ ou CH3• e (II)
modificação do DNA, induzida pela radicais reativos de oxigênio ou NO pela
STZ. O2-...................................................................................................................... 43
Figura 15: Geração de EROs induzida pela hiperglicemia e conseqüente ativação
das vias patológicas.................................................................................................... 50
Figura 16: Processo de glicação não enzimática. Adição de glicose a proteínas
com subseqüente degradação do produto de glicação. Formação de
deoxiglicosanas que reagem com proteínas para formar AGEs................................. 52
Figura 17: Ativação de PKC e formação de EROs na hiperglicemia....................... 53
Figura 18: Estrutura de um flavonóide...................................................................... 54
Figura 19: Estrutura do 1,1-difenil-2-picril-hidrazil (DPPH)................................... 55
Figura 20: Estrutura do DPPH e sua redução pelo antioxidante............................... 56
Figura 21: Formação do cátion radical ABTS·+........................................................ 58
Figura 22: Estrutura do ácido ascórbico.................................................................... 67

Figura 23: Cromatograma do PPT solúvel em MeOH.............................................. 72
LISTA DE GRÁFICOS
Gráfico 1: Efeito do extrato etanólico bruto das folhas de MSE (1ª coleta) em
camundongos diabéticos induzidos com aloxana. Valores são média ± E.P.M.
(n=6). ANOVA Two-Way seguido do Pós-Testes de Bonferroni, *P<0,01............. 83
Gráfico 2: Efeito do extrato etanólico bruto das folhas de MSE (1ª coleta), nas
concentrações de 300, 600 e 900 mg/Kg, comparados com o grupo controle com
relação à porcentagem de redução da glicemia em camundongos. Valores são
média ± E.P.M. (n=6). * P> 0,01. ANOVA one-way............................................... 84
Gráfico 3: Efeito do extrato etanólico bruto das folhas de MSE (2ª coleta) em
camundongos diabéticos induzidos com aloxana. Valores são média ± E.P.M.
(n=6). ANOVA Two-Way seguido do Pós-Testes de Bonferroni............................ 85
Gráfico 4: Efeito da fração AcOEt das folhas de MSE (2ª coleta) em
camundongos diabéticos induzidos com aloxana. Valores são média ± E.P.M.
(n=6). ANOVA Two-Way seguido do Pós-Testes de Bonferroni............................ 85
Gráfico 5: Efeito do extrato etanólico bruto das folhas de MSE (3ª coleta), em
camundongos diabéticos induzidos com aloxana. Valores são Média ± E.P.M.
(n=6). ANOVA Two-Way seguido do Pós-Testes de Bonferroni............................ 86
Gráfico 6: Efeito do extrato etanólico bruto das folhas de MSE (3ª coleta), nas
concentrações de 300, 600 e 900 mg/Kg, comparados com o grupo controle com
relação à porcentagem de redução da glicemia em camundongos. Valores são
média ± E.P.M. (n=6). ANOVA one-way................................................................. 87
Gráfico 7: Efeito da fração aquosa das folhas de B. cheilantha (MSE) (3ª coleta),
em camundongos diabéticos induzidos com aloxana. Valores são média ± E.P.M.
(n=6). *P<0,05. ANOVA Two-Way seguido do Pós-Testes de Bonferroni............. 88
Gráfico 8: Efeito da fração aquosa das folhas de MSE (3ª coleta), nas
concentrações de 300, 600 e 900 mg/Kg, comparados com o grupo controle, com
relação à porcentagem de redução da glicemia em camundongos. Valores são
média ± E.P.M. (n=6). *P<0,05. ANOVA one way.................................................. 88
Gráfico 9: Efeito do PPT MeOH das folhas de MSE (3ª coleta), em camundongos
diabéticos induzidos com aloxana. Valores são média ± E.P.M.
89

(n=6)...........................................................................................................................
Gráfico 10: Efeito do PPT diclorometano das folhas de MSE (3ª coleta), em
camundongos diabéticos induzidos com aloxana. Valores são média ± E.P.M.
(n=6)..........................................................................................................................................................................................
89
Gráfico 11: Atividade seqüestradora de radical livre dos extratos/amostras de
MSE, usando o radical DPPH•, da 1a coleta................................................................ 90
Gráfico 12: Atividade seqüestradora de radical livre dos extratos/amostras de
MSE, usando o radical DPPH•, da 2a coleta................................................................ 91
Gráfico 13: Atividade seqüestradora de radical livre dos extratos/amostras de
MSE, usando o radical DPPH•, referente a 3a coleta.................................................. 92
Gráfico 14: Correlação linear do teor de Fenólicos Totais (FT) em função de
1/CE50 dos extratos/frações de MSE da 3a coleta....................................................... 93
Gráfico 15: Teor de fenólicos totais dos extratos/frações de MSE referente à 2a
coleta............................................................................................................................ 94
Gráfico 16: Teor de fenólicos totais dos extratos/frações de MSE referente à 3a
coleta............................................................................................................................ 95
Gráfico 17: Atividade antioxidante equivalente ao Trolox (CAET) dos
extratos/amostras de MSE referentes à 2a coleta......................................................... 96
Gráfico 18: Atividade antioxidante equivalente ao Trolox (CAET) dos
extratos/amostras de MSE referentes à 3a coleta......................................................... 97
Gráfico 19: Correlação linear do teor de Fenólicos Totais (FT) em função de
1/CE50 dos extratos/frações de MSE da 3º coleta........................................................ 97
LISTA DE FLUXOGRAMA
Fluxograma 1: Obtenção do extrato e frações de B. cheilantha................................. 63

LISTA DE ABREVIATURAS ABTS - Ácido 2, 2´-azinobis-[3-etilbenzotiazolina-6-sulfônico]
ACTH - Adenocorticotrófico
ADA - Associação Americana de Diabetes
AGEs - Produtos finais de glicação avançada
AL - Aloxana
ANVISA - Agência Nacional de Vigilância Sanitária
ATP - Adenosina trifosfato
CAET - Capacidade antioxidante equivalente ao trolox
CLAE - Cromatografia Líquida de Alta Eficiência
COSY - Espectrocospia de correlação homonuclear
CV - Cardiovascular
CZI - Insulina zíncica cristalina
DAG - Diacilglicerol
DM - Diabetes mellitus
DPPH - 1,1-Difenil-2-picril-hidrazil
E.P.M - Erro Padrão da Média
EROs - Espécies reativas de oxigênio
FADH2 - Flavina adenina dinucleotídeo
FPG - Glicose plasmática de jejum
FT - Fenólicos Totais
GDM - Diabetes Mellitus Gestational
GIP - Peptídeo inibitório gástrico
GLP1 - Peptídeo semelhante ao glucagon (GLP1)
GLUT - Transportador de glicose
GSH - Glutationa
Hz - Hertz
HDL - Lipoproteína de alta densidade
HLA - Antígeno Leucocitário Humano
HMBC - Hetero Multi Bond Correlation

HMQC - Hetero Multi Quantum Correlation
i.p. - intraperitoneal
IR - Receptor de insulina
J - Constante de acoplamento
KATP - Canais de Potássio sensíveis à ATP
LDL - Lipoproteína de baixa densidade
MODY - Diabetes da maturidade com início na juventude
MSE - Mororó Sem Espinho
NADH - Nicotinamida adenina dinucleotídeo-P
NPH- Neutral Protamina Hagedorn
OGTT - Teste de tolerância oral à glicose
OMS - Organização Mundial de Saúde
PI - Percentagem de inibição
PKC - Proteína cinase C
PPT - Precipitado
Reagente NP ácido difenilbórico etanolamina
RMN - Ressonância Magnética Nuclear
ROS - ver EROs
S.N.C - Sistema Nervoso Central
Sl - Singleto largo
STZ - Estreptozotocina
TCA - Ciclo do ácido tricarboxílico
TGF-β - Fator de crescimento transformador β
v.o. - via oral
VDCC - Canais de Cálcio sensíveis a voltagem
VEGF Fator de crescimento endotelial vascular

1. INTRODUÇÃO
A Organização Mundial de Saúde (OMS) define Planta Medicinal como sendo
“todo e qualquer vegetal que possui, em um ou mais órgãos, substâncias que podem ser
utilizadas com fins terapêuticos ou que sejam precursores de fármacos semi-sintéticos”
(WORLD HEALTH ORGANIZATION, 1998). A utilização de plantas com fins
medicinais para tratamento, cura e prevenção de doenças é uma das mais antigas formas
de prática medicinal da humanidade. No início da década de 90, a OMS divulgou que
65-80% da população dos países em desenvolvimento dependiam das plantas
medicinais como única forma de acesso aos cuidados básicos de saúde (AKERELE,
1993).
Vários países desenvolvidos estão voltados para o uso da medicina tradicional,
segundo recente levantamento realizado nos Estados Membros da União Européia,
cerca de 1400 preparações são amplamente utilizadas para tratamento de doenças e com
fins cosméticos na França, Holanda, Bélgica e Alemanha (HOAREAU & DASILVA,
1999). Segundo SILVA (2006), a Organização Mundial de Saúde estima que 80% da
população mundial dependem da medicina tradicional para suas necessidades básicas de
saúde, e que, quase 85% da medicina tradicional envolvem o uso de plantas medicinais,
extratos vegetais e seus princípios ativos.
O Brasil é o país com a maior biodiversidade do mundo, estima-se que mais de
20% do número de espécies do planeta seja encontrada no Brasil. O País possui a mais
diversa flora, número superior a 55 mil espécies descritas, o que corresponde a 22% do
total mundial (BRASIL, 2006). Esta rica biodiversidade é acompanhada por uma grande
aceitação de uso de plantas medicinais e conhecimento tradicional associado
(RODRIGUES, 2006). Apenas 8% das espécies vegetais da flora brasileira foram
estudadas em busca de compostos bioativos e 1.100 espécies vegetais foram avaliadas
em suas propriedades medicinais (GARCIA et al., 2003).
Atualmente, aproximadamente 48% dos medicamentos empregados na
terapêutica advêm, direta ou indiretamente, de produtos naturais, especialmente de
plantas medicinais (BALUNAS e KINGHORN, 2005) que permanecem uma
importante fonte para obtenção de medicamentos. Segundo a Agência Nacional de

vigilância sanitária (ANVISA) há cerca de 400 espécies de fitoterápicos registrados,
com 60 plantas medicinais, sendo destas 10 nativas do Brasil. As plantas que mais
possuem registro na ANVISA na forma de seus derivados para obtenção de
fitoterápicos são: Ginkgo biloba (Ginkgo), Aesculus hippocastanum (Castanha da
índia), Panax ginseng (Ginseng), Senna alexandrina (Sene), Peumus boldus (Boldo),
Cynara scolymus (alcachofra), Passiflora incarnata (Maracujá), Valeriana officinalis
(Valeriana) e Arnica Montana (Arnica).
Dentre os diversos reinos da natureza, o reino vegetal é o que tem contribuído de
forma mais significativa para o fornecimento de metabólitos secundários. O isolamento
das primeiras substâncias puras do reino vegetal começou a acontecer no século XVII.
Este século, juntamente com o XIX, caracteriza-se pelos trabalhos de extração,
principalmente de ácidos orgânicos e de alcalóides. É desta época o isolamento de
morfina (1806), quinina e estricnina (1820) (PINTO et al., 2002), muitos destes de
grande valor agregado devido às suas aplicações como medicamento, cosméticos,
alimentos e agroquímicos (PHILLIPSON et al., 1998). A avaliação do potencial
terapêutico de plantas medicinais e de alguns de seus constituintes, tais como
flavonóides, alcalóides, triterpenos, sesquiterpenos, taninos, lignanas, etc, tem sido
objeto de incessantes estudos, onde já foram comprovadas as ações farmacológicas
através de testes pré-clínicos com animais (HAVSTEEN, 1983; CALIXTO et al., 1990;
SAMUELSSON, 1992; CECHINEL FILHO, 1995). Muitas destas substâncias têm
grandes possibilidades de futuramente virem a ser aproveitadas como agentes
medicinais. Entre os diversos exemplos de substâncias oriundas de plantas e de importância
atualmente, podemos mencionar a forscolina, obtida de Coleus forskohlii, que apresenta
promissores efeitos contra hipertensão, glaucoma, asma e certos tumores (DE SOUZA,
1993), a artemisinina, presente em Artemisia annua, que exerce potente atividade
antimalárica (KAMCHONWONGPAISON, 1996), e o diterpeno anticancerígeno taxol,
isolado de plantas do gênero Taxus, que após sua síntese em escala industrial, já se
encontra disponível no mercado farmacêutico, constituindo-se numa grande esperança
para pessoas portadoras de câncer nos ovários e pulmões (KINGSTON, 1991;
HORWITZ, 1994; Fitoterapia, 1995; CORRÊA, 1995).
As plantas medicinais da flora nativa, no Brasil, são consumidas com pouca ou
nenhuma comprovação de suas propriedades farmacológicas, propagadas por usuários
ou comerciantes (VEIGA-JUNIOR, 2005). A garantia da qualidade dos produtos

fitoterápicos certamente não tem acompanhado o crescimento na popularidade destes
produtos. A falta de testes clínicos que demonstrem de forma inequívoca a sua eficácia,
e a ausência de reprodutibilidade dos produtos fitoterápicos são as maiores críticas que
esta forma de terapia tem sofrido ao longo dos anos. Existem poucos estudos na
literatura realizados com o objetivo específico de se comparar a qualidade dos
fitoterápicos comercializados, mas os poucos estudos realizados reforçam a idéia
bastante generalizada de que a qualidade dos fitoterápicos é extremamente variável.
O estudo farmacológico das drogas vegetais (planta inteira ou partes), além de
constituir um campo inesgotável de novos conhecimentos científicos e geradores de
riquezas, pode contribuir notavelmente para o aprimoramento da medicina tradicional
(SIXEL, 1998). Relatos etno-botânicos indicam que há aproximadamente 1200 plantas
no mundo com potencial atividade anti-diabética (ALARCON-AGUILAR et al., 2002).
Estudos atuais sobre novas drogas hipoglicemiantes vêm sendo realizados, com
enfoque especial para as plantas medicinais usadas na medicina popular (CECHINEL-
FILHO, YUNES, 1998; BARBOSA-FILHO et al., 2005). Entre as inúmeras espécies de
interesse medicinal, encontram-se as plantas do gênero Bauhinia, amplamente utilizada
na medicina popular como hipoglicemiante.
Um dos fatores de extrema importância para a descoberta de princípios ativos
naturais consiste, principalmente, na interação entre a química e a farmacologia. Quanto
mais estreita for esta colaboração, mais rápida e consistentemente serão alcançados os
objetivos almejados (CECHINEL-FILHO, YUNES, 1998). Assim, é extremamente
necessária uma integração entre as duas áreas de concentração, visando uma agregação
dos conhecimentos e ampliando a visão do aluno que participa das duas áreas
mencionadas. É interessante mencionar que estudos in vivo e in vitro (JENNINGS et al., 1991)
entre medicamentos com atividade hipoglicemiante e antioxidante reduziram de forma
significativa o aparecimento de algumas complicações diabéticas em pacientes com
DM2 como retinopatia quando comparados com medicamentos com atividade
hipoglicemiante apenas. Nesse estudo, o controle glicêmico dos pacientes de ambos os
grupos não diferiu, o que apóia a hipótese de que um paciente que faz uso de um
medicamento com ambas as propriedades (hipoglicemiante e antioxidante) pode ter
mais benefícios no controle da glicemia e prevenção de complicações oriundas do
diabetes do que um paciente que faz uso apenas de um medicamento com propriedades
hipoglicemiantes.

1. Considerações sobre o gênero Bauhinia
Entre as inúmeras espécies vegetais de interesse medicinal, encontram-se as
plantas do gênero Bauhinia (Figura 1), pertencentes à família Leguminosae, as quais
são encontradas principalmente nas áreas tropicais do planeta, compreendendo
aproximadamente 300 espécies (SILVA; CECHINEL-FILHO, 2002), distribuídos,
principalmente, na Ásia, África e América Latina (ILDIS and CHCD, 1992). Este
gênero inclui árvores (perenes e decíduas), arbustos e lianas, com folhas simples
bilobuladas e flores de várias cores e tamanhos, únicas ou em rácemos, corimbos ou
panículas. Os frutos são deiscentes ou indeiscentes, com aproximadamente 15-20 cm de
comprimento. As sementes são largas (PONOMARENKO; PAVLOVA, 2000).
Fonte: www.rain-tree.com/Plant-Images/Bauhinia_p1.jpg
Figura 1: Foto de uma espécie do gênero Bauhinia
No Brasil, as plantas do gênero Bauhinia são conhecidas com “Pata-de-vaca”,
Mororó, “Unha-de-boi”, devido ao formato da folha (Figura 2). As folhas, caules e
raízes das espécies de Bauhinia, especialmente B. manca, B. rufescens, B. forficata, B.
cheilantha (Figura 3) e B. splendens, são amplamente utilizadas no Brasil e em outros
países em forma de chás e outras preparações fitoterápicas para o tratamento de várias
enfermidades, principalmente infecções, processos dolorosos e diabetes
(ACHENBACH, 1988; TESKE, 1995; GUPTA, 1995).

Fonte: http://herbaria.plants.ox.ac.uk
Figura 2: Formato das folhas de plantas do gênero Bauhinia
Figura 3: Exsicata de B. cheilantha
Pouco se conhece a respeito da atividade farmacológica das plantas do gênero
Bauhinia, porém, este gênero é mais freqüentemente estudado quanto à sua possível
ação hipoglicemiante, uma vez que na medicina popular estas plantas são usadas para o
tratamento de diabetes (TESKE, 1995; GUPTA, 1995). Carmela Juliani (1929) foi a
primeira autora a descrever a atividade hipoglicêmica da B. forficata, realizando

experimentos em cães e coelhos submetidos à hiperglicemia adrenalítica e
pancreactomizados onde, ela observou que esta espécie vegetal continha compostos
capazes de diminuir a glicemia. A partir daí outras espécies têm sido estudadas quanto a
essa atividade, como B. divaricata, B. monandra e B. variegata. Outros estudos têm
sido realizados para avaliar outras atividades farmacológicas deste gênero, como
atividade antimicrobiana (CECHINEL FILHO et al., 1995), efeitos sobre a musculatura
lisa (WAZLAWIK et al, 1994) e sistema nervoso central (SCHMELING et al., 2000).
Algumas espécies de plantas do gênero Bauhinia foram e estão sendo estudadas
fitoquimica e farmacologicamente e, com isto, muitos compostos foram isolados e
identificados. Assim, diferentes classes de compostos orgânicos de interesse medicinal
existentes nessas espécies foram relatados, incluindo lactonas, flavonóides, terpenóides,
esteróides, taninos e quinonas (SILVA, CECHINEL-FILHO, 2002). DE SOUSA et al.
(2004) isolaram das folhas de Bauhinia forficata o flavonóide Caempferol-3,7-O-(r)-
diraminosideo (Figura 4), conferindo a esta substância a capacidade de diminuir os
níveis de glicemia nos modelos animais estudados.
Figura 4: Flavonóide Caempferol-3,7-O-(r)-diraminosideo
2. Pâncreas 2.1. Anatomia
O pâncreas (Figura 5) é conhecido como uma glândula mista, porque ele possui
funções exócrinas e endócrinas. A função endócrina é formada por aglomerados de
células chamadas de ilhotas pancreáticas (ilhotas de Langerhans). As células das ilhotas

secretam os hormônios insulina e glucagon no sangue. Como uma glândula exócrina, o
pâncreas secreta suco pancreático, através do duto pancreático, que esvazia dentro do
duodeno (GRAAFF, 2002).
Fonte: www.esadi.com.br/images/pancreas2.jpg
Figura 5: Anatomia do Pâncreas 2.2. Fisiologia
O pâncreas é uma glândula mista, composta por uma porção exócrina, os ácinos,
que produzem enzimas digestivas, e outra endócrina, as ilhotas de Langerhans, que
sintetizam e liberam vários hormônios (FLATT, 1992) (Figura 6).
As ilhotas de Langerhans são pequenas massas de células, muito vascularizadas,
distribuídas por toda a porção exócrina do pâncreas (FLATT, 1992). Existem cerca de 1
milhão de ilhotas no pâncreas, que constituem 1 a 1,5% da massa pancreática humana.
Cada ilhota contém, em média, 2.500 células de quatro tipos diferentes (KAHN, 1996).
Células B ou β: no ser humano, constituem a única fonte de insulina, perfazem de 60 a
70% e ocupam preferencialmente a parte central da ilhota;
• Células A ou α: secretam glucagon. Representam 25% da população celular da
ilhota e se distribuem na sua periferia ou rodeando os capilares que penetram na
ilhota;
• Células D ou δ: secretam somatostatina e representam 10% das células
insulares. Distribuem-se na periferia da ilhota e também junto aos capilares, de
modo que guardam uma estreita relação com as células A;

• Células PP: secretam polipeptídeo pancreático. Ocupam 5% da massa celular e
se localizam em torno dos capilares e na periferia da ilhota, um pouco mais
profundamente que as células A e D (KAHN, 1994; DRAZNIN et al., 1989).
Células exócrinas
SecretamCélulasCélulas endócrinas GlucagonCel. α
Cel. D
Insulina
SomatostatIlhotas de Langerhans Cel. β
Células Alfa
Células Beta
Células D
Fonte: SILVERTHORN, 2003
Figura 6: Distribuição das células na ilhota de Langerhans.
As ilhotas recebem aproximadamente 10% do fluxo sanguíneo total do pâncreas.
O suprimento sanguíneo do pâncreas endócrino é disposto de tal modo que o sangue
venoso de um tipo de célula banha os outros tipos celulares. Pequenas artérias penetram
o cerne da ilhota, distribuindo sangue por uma rede de capilares fenestrados e daí
convergindo em vênulas que carreiam o sangue para a borda da ilhota. Assim, o sangue
venoso das células β carreia insulina para as células α e δ (BELL et al., 1980).
3. Insulina
O hormônio Insulina foi purificado e cristalizado por Abel em 1921, depois de
muitos anos após a sua descoberta. Sanger determinou a seqüência de aminoácidos da
insulina em 1958, a proteína foi sintetizada em 1963, e Hodgkin e colaboradores
elucidaram a estrutura tridimensional da insulina em 1972 (KAHN, ROTH, 2004).
A Insulina é o hormônio anabólico mais conhecido e é essencial para a
manutenção da homeostase de glicose e do crescimento e diferenciação celular
(CARVALHEIRA, et al., 2002).

A insulina:
• Foi o primeiro hormônio a ser isolado de fontes animais em uma forma
que podia ser administrada terapeuticamente aos seres humanos;
• Foi o primeiro hormônio a ter suas estruturas primária e terciária
determinadas;
• Foi o primeiro hormônio a ter o seu mecanismo de ação elucidado;
• Foi o primeiro hormônio a ser dosado por radioimunoensaio, por Yalow
and Berson;
• Foi o primeiro hormônio conhecido a ser sintetizado a partir de um
precursor maior (pró-hormônio);
• Foi o primeiro hormônio a ser sintetizado pela tecnologia do DNA
recombinante (BELL et al., 1980; DE GROOT, 1994; UNGER et al.,
1978).
3.1. Estrutura
A insulina é formada por duas cadeia peptídicas retas (denominadas cadeias A e
B) que são mantidas juntas (Figura 7A). Seu peso molecular é de 6.000 dáltons. A
cadeia A, contendo 21 aminoácidos, e a cadeia B, contendo 30 aminoácidos, são
conectadas por duas pontes dissulfeto (JEFFERSON, 1980). A cadeia A é ácida e
apresenta uma ponte intracatenária 6-11, enquanto a B é básica. A insulina porcina
difere da humana apenas no aminoácido da posição B30 (DRAZNIN et al., 1989).
A estrutura terciária da insulina é determinada pelos aminoácidos N-terminal e
C-terminal da cadeia A e pela natureza hidrofóbica dos aminoácidos no C-terminal da
cadeia B. Essa estrutura terciária é importante para a atividade biológica da insulina,
que reside na cadeia B (GRANNER, ANDREONE, 1985).
A atividade biológica do hormônio depende não só da seqüência de
aminoácidos, mas também de sua estrutura espacial e do dobramento tridimensional de
sua molécula (Figura 7B). Esse último permite agrupar na superfície as porções
carboxila e amino da cadeia A e a porção distal da cadeia B (DRAZNIN et al., 1989).

Peptídio C
Cadeia
Peptídeo
Cadeia
A. Fonte: IZE-LUDLOW, SPERLING, 2005
B. Fonte: lqes.iqm.unicamp.br/images/lqes_empauta_novid
Figura 7: Estrutura da insulina: A. Molécula da pró insulina humana; B. Molécula da
insulina dobrada.
3.2. Síntese
As células β (ou B) das ilhotas pancreáticas sintetizam insulina, de um precursor
de cadeia simples com 110 aminoácidos, denominado pré-pró-insulina. Depois da
translocação através da membrana do retículo endoplasmático rugoso, o peptídeo

sinalizador, com 24 aminoácidos, da pré-pró-insulina é clivado rapidamente para formar
pró-insulina (DAVIS, 2006) (Figura 8).
A pró-insulina humana, que tem peso molecular de 9 kDa, é composta por 86
aminoácidos , onde as cadeias A e B da insulina encontram-se unidas entre si por um
peptídeo intermediário (DRAZNIN et al., 1989). Enquanto a pró-insulina está sendo
orientada para o aparelho de Golgi, são estabelecidas ligações dissulfeto que produzem
a molécula “dobrada” de pró-insulina com peso molecular de 9.000 kDa. As cadeias A e
B da insulina, conectadas por pontes dissulfeto, estão ligadas por um peptídeo de
conexão (peptídeo C) através de dois resíduos básicos, cada um dos quais fica
localizado ao nível do C-terminal da cadeia B e do N-terminal da cadeia A
(JEFFERSON, 1980).
Durante a conversão da pró-insulina humana à insulina, quatro aminoácidos
básicos e o peptídeo conector ou peptídeo C são removidos por proteólise. Isto ocasiona
as cadeias peptídicas A e B da molécula de insulina, que contém uma ligação dissulfito
intrasubunidade e duas ligações intersubunidade. (DE MEYTS, 2004) (Figura 8).
A conversão da pró-insulina para insulina começa no Complexo de Golgi,
continua nos grânulos secretórios, e é quase completa no tempo de secreção. Assim,
quantidade equimolar do peptídeo C e insulina é liberada na circulação. O peptídeo C
não tem função biológica conhecida. Pequenas quantidades de insulina e pró-insulina
também são liberadas das células β. Isto reflete se a exocitose de grânulos em que a
conversão de pró-insulina para insulina não é completa ou a secreção ocorre por outros
caminhos. Já que meia-vida da pró-insulina na circulação é mais longa do que a da
insulina, mais de 20% da insulina imunoreativa plasmática é, na realidade, pró-insulina
e intermediários (DE MEYTS, 1994).
Duas endopeptidases distintas dependentes de Ca2+, encontradas nos grânulos
das ilhotas e em outras células neuroendócrinas, são responsáveis pela conversão da
pró-insulina em insulina. Estas endoproteases, PC2 e PC3, tem domínios catalíticos
relatados como subtilisina e cliva as sequências Lis-Arg ou Arg-Arg (STEINER, 1996).
PC2 seletivamente cliva a junção da cadeia A-peptídeo C. PC3 preferencialmente cliva
a junção da cadeia B-peptídeo C, mas tem algumas ações na junção da cadeia A.
Embora existam outros membros da família de endoproteases como a PC1, a furina, a
PC2 e a PC3 parecem ser as enzimas responsáveis por processar a pró-insulina em
insulina (STEINER, 1996).

Fonte: CINGOLANI, 2004
Figura 8: Síntese de insulina
3.3. Secreção
O controle da secreção de insulina nas células β e a ação biológica deste
hormônio peptídeo nos seus tecidos alvos assegura a função vital da homeostase da
glicose (LANG, 1999).
Depois de sua síntese no retículo endoplasmático, a insulina é processada para
sua forma biologicamente ativa e estocada em grânulos secretórios à espera de sua
liberação (RORSMAN; RENSTROM, 2003). Estudos ultra-estruturais tem mostrado
que uma única célula β contém mais do que 10.000 grânulos secretórios (DEAN, 1973;
OLOFSSON, 2002).
A secreção de insulina (Figura 9) é um processo rigorosamente regulado,
projetado para fornecer concentrações estáveis de glicose no sangue, durante o jejum e
período de alimentação (DAVIS, 2006). Algumas substâncias podem estimular a

secreção de insulina, os secretagogos, que podem ser divididos em dois grupos: os
iniciadores e os potenciadores.
Os iniciadores são capazes de estimular a secreção de insulina por si próprios e
incluem nutrientes, semelhantes à glicose e drogas, semelhantes a sulfoniluréias. Todas
essas substâncias agem pela inibição da atividade dos Canais para Potássio Sensíveis à
ATP (KATP), mas, enquanto os nutrientes podem ser metabolizados para afetar o
fechamento dos canais, as drogas ligam-se diretamente ao canal e bloqueiam sua
atividade. Os Potenciadores da secreção de insulina incluem um número de hormônios
[por exemplo, glucagon e peptídeo semelhante ao glucagon (GLP1)], transmissores,
(por exemplo a acetilcolina) e aminoácidos. Estes agentes amplificam a secreção de
insulina induzida pelos iniciadores, mas não podem aumentar a secreção de insulina por
eles mesmos, porque eles não fecham os canais KATP e são apenas capazes de exercer
seus efeitos depois que os iniciadores têm exercido seu efeito de inibição dos KATP
(ASHCROFT, 1999).
As ilhotas de Langerhans são ricamente inervadas pelos nervos adrenérgicos e
colinérgicos. A estimulação dos receptores α2 adrenérgicos inibe a secreção de insulina,
enquanto agonistas dos receptores β2 adrenérgicos e estimulação do nervo vagal
aumenta a liberação de insulina (DAVIS, 2006).
Os níveis de glicose sanguíneos são fortemente controlados pela secreção de
insulina das células β pancreáticas e pela ação da insulina no fígado, músculo e outros
tecidos alvos. As células β é capaz de adaptar a secreção de insulina de acordo com as
flutuações na concentração de glicose no sangue. A glicose entra na célula β por
transporte facilitado, mediado pelos GLUT2, um subtipo específico de transportador de
glicose (DAVIS, 2006). Sua fosforilação pela glicocinase para glicose-6-fosfato
determina a taxa de glicólise e a taxa de geração de piruvato (NEWGARD,
MCGARRY, 1995; MATSCHINSKY, 1996). Nas células β, o piruvato é o principal
produto da glicólise (ISHIHARA, et al. 1999). Comparado com outros tipos celulares,
uma alta proporção de carbonos derivados de glicose entra na mitocôndria na forma de
piruvato e entra no ciclo do Ácido Tricarboxílico (TCA). A transferência de elétrons do
ciclo do TCA para a cadeia respiratória pelo NADH e FADH2 promove a geração de
ATP, que é exportado para o citosol (MAECHLER, WOLLHEIM, 2001).
A mudança da razão adenosina trifosfato (ATP)/adenosina bifosfato (ADP)
promove o fechamento dos KATP. Isto ocasiona a despolarização da membrana e
abertura dos Canais de Cálcio abertos por voltagem (CaV) (MAECHLER,

WOLLHEIM, 1994) (Figura 9). Isto é a etapa chave pelo qual a glicose estimula a
secreção de insulina, como o aumento do Ca+2 citosólico é o principal estimulador da
exocitose, processo pelo qual os grânulos secretórios contendo insulina se fundem com
a membrana plasmática (LANG, 1999; RORSMAN, 1997).
Canal KATP
Captação de glicose
Glicólise, respiração
glicocinase
F
F
F F
Canal KCa
2+Liberação de insulina
Grânulos de estoque Fonte: www.betacell.org/.../insulin-secretion-w500.jpg
Figura 9: Modelo para acoplamento do metabolismo da glicose e secreção de
insulina nas células β
3.4. Padrão bifásico de secreção da insulina
A secreção de insulina, em resposta à estimulação pela glicose, exibe um
característico padrão bifásico e consiste de uma primeira fase rápida e transiente,
acompanhada por uma segunda fase sustentada durante a qual a secreção de insulina
continua a uma taxa reduzida, mas ainda aumentada com relação ao controle pré-
estimulatório. Apenas uma fração das vesículas das células β é mobilizada durante a
estimulação (RORSMAN; RENSTROM, 2003). O componente rápido (secreção da
primeira fase) corresponde à liberação dos grânulos dependentes de Ca+2, pertencentes

aos grânulos prontamente liberáveis, enquanto o componente sustentado (segunda fase)
reflete fusão mais lenta dos grânulos e é resultante de sua translocação para o sítio de
liberação (RORSMAN et al, 2000; BARG et al., 2002a).
A primeira e a segunda fase da secreção de insulina em ilhotas de rato têm, de
acordo com relatos, a quantidade de 0,14% dos grânulos por min e 0,05% dos grânulos
por min, respectivamente (ANELLO et al., 1999). Sabendo-se que muitas células β
contêm aproximadamente 10.000 grânulos, esta taxa de liberação corresponde a
aproximadamente 15 e 5 grânulos por minutos (BRATANOVA-TOCHKOVA et al.,
2002).
A exocitose finalmente resulta na fusão da membrana dos grânulos secretórios
com a membrana plasmática, acompanhada pela liberação do conteúdo dos grânulos,
insulina, dentro do espaço extracelular, comunicando com o fluxo sanguíneo capilar.
Apenas uma pequena quantidade de insulina é liberada sob condições estimulatória
máxima (LANG, 1999). Como em outras células endócrinas e em neurônios, a exocitose
em células β é regulada e a quantidade de insulina circulante no sangue depende mais da
taxa de exocitose do que da taxa de biossíntese de insulina (BARG, 2003).
3.5. Mecanismo de ação da insulina O receptor de insulina (IR) é uma proteína de membrana tetramérica, consistindo
de duas subunidades α idênticas e duas β idênticas (Figura 10). Os IR estão presentes
não apenas nos alvos clássicos como fígado, músculo, tecido adiposo, mas também em
praticamente todas as células mamárias, células sanguíneas circulantes, neurônios e
células gonadais. O número de receptores varia de poucos, como 40 por célula nos
eritrócitos, a 300.000 por célula nos adipócitos e hepatócitos (KHAN, PESSIN, 2002).
As subunidades α estão localizadas inteiramente na face extracelular da membrana
plasmática e contêm o sítio de ligação à insulina. As subunidades β são um peptídeo
transmembrana, possuem atividade tirosina-cinase específica no domínio intracelular.
(KASUGA et al., 1982; KASUGA et al., 1983) A ligação da insulina ao receptor
tirosina cinase na superfície externa da célula induz uma autofosforilação nos muitos
resíduos tirosina localizados no lado interno das células (LIZCANO, ALESSI, 2002).

Zona de ligação da insulina
Pontes dissulfeto entre as duas cadeias α
Pontes dissulfeto entre as cadeias α e β
Domínio catalítico tirosina-cinase
Fonte: www.scielo.org.co/.../rfmun/v53n4/v53n4a05f1.jpg Figura 10: Receptor da insulina
3.6. Distribuição e degradação da insulina
Insulina circula no sangue como um monômero livre e seu volume de
distribuição se aproxima do volume de distribuição do fluido extracelular. Sob
condições de jejum, o pâncreas secreta aproximadamente 40µg (1 unidade) de
insulina por hora dentro da veia porta, atingindo uma concentração no sangue porta
de 2 a 4 ng/ml (50 a 100 µunidades/ml) e na circulação periférica de 0,5 ng/ml (12
µunidades/ml) ou aproximadamente 0,1 nM (DAVIS, 2006).
A meia vida plasmática da insulina é de 5 a 8 minutos e sua velocidade de
depuração metabólica é de 800 ml/min (KAHN, 1996). Essa fugaz meia-vida
plasmática deve-se à retenção do hormônio por certos tecidos que a retiram da
circulação; em apenas uma passagem, o fígado retém 50% da insulina plasmática e o
rim retém 40%. A pró-insulina tem uma meia-vida pelo menos duas vezes maior e
não é convertida em insulina fora do pâncreas (DRAZNIN, 1989).
A glicose é o principal estímulo para a secreção de insulina em seres humanos
e é um fator permissivo essencial para a ação de muitos outros secretagogos. O açúcar
(glicose) é mais efetivo em provocar a secreção de insulina quando dado oralmente
do que quando administrado intravenosamente, porque a ingestão de glicose (ou
carboidrato) induz a liberação de hormônios gastrintestinais e estimula a atividade
vagal. Muitos hormônios promovem a secreção de insulina. Os mais potentes são o
Peptídeo Inibitório Gástrico (GIP) e Peptídeo-1 Semelhante ao Glucagon (GLP-1).
Liberação de insulina também é estimulada pela gastrina, secretina, colecistocinina,

peptídeo intestinal vasoativo, peptídeo liberador de gastrina e enteroglucagon
(MATCHINSKY; BANTING, 1995).
A insulina é metabolizada sobretudo no rim e no fígado por enzimas
específicas que abrem as ligações dissulfeto e separam a cadeias A das cadeias B.
Pouquíssima insulina é excretada de forma inalterada na urina (JEFFERSON, 1980).
A degradação proteolítica da insulina no fígado ocorre primariamente depois da
internalização do hormônio e seu receptor. O caminho primário para internalização é
endocitose mediada pelo receptor, GLUT-4. O complexo da insulina e seu receptor é
internalizado em pequenas vesículas chamadas endossomas, onde a degradação é
iniciada. Alguma insulina é liberada para lisossomas para degradação
(DUCKWORTH, 1988).
3.7. Ações da insulina
A insulina é um hormônio caracterizado por promover todos os processos que
facilitam o depósito de substratos em forma de macromoléculas nos tecidos e inibir
aqueles que produzem o efeito oposto (DRAZNIN et al., 1989) (Tabela 1). Os tecidos
alvos importantes para a regulação da homeostase da glicose são o fígado, músculo e
tecido adiposo, mas a insulina exerce potente efeito regulatório em outros tipos
celulares. Insulina é o primeiro hormônio responsável por controlar a captação, uso e
armazenamento de nutrientes celulares. Ação anabólica da insulina inclui a
estimulação do uso e armazenamento de glicose, aminoácido e lipídios, entretanto ele
inibe processos catabólicos semelhante à quebra de glicogênio, gordura e proteína
(DAVIS, 2006).

Tabela 1: Resumo dos efeitos da insulina sobre o metabolismo de carboidratos, lipídios e proteínas no fígado, músculo e tecido adiposo.
Tipo de metabolismo
Células hepáticas Tecido adiposo Músculo
Metabolismo de carboidratos
↓gliconeogênese
↓glicogenólise
↑glicólise
↑glicogênese
↑captação de glicose
↑síntese de glicerol
↑glicólise
↑glicogênese
Metabolismo de lipídios
↑lipogênese
↓lipólise
↑síntese de triglicerídeos ↑síntese de ácidos graxos
_____
Metabolismo de proteínas
↓ degradação das proteínas
______ ↑ captação de Aminoácidos ↑síntese de proteína
Fonte: Rang et al, 2004 3.8. Glicose no sangue
Os níveis normais de glicose sanguínea em um indivíduo normal variam de 60
a 110 mg/dl. A concentração normal de glicose sanguínea em um indivíduo que não
se alimentou nas últimas 3 a 4 horas é aproximadamente 90 mg/dl. Após uma refeição
contendo grande quantidade de carboidratos, este nível raramente sobe acima de 140
mg/dl (BELFIORE, MOGENSEN, 2000).
Transporte de glicose e os GLUTs
O transporte de glicose é mediado por carreadores, referidos como uma
família de transportadores de glicose, com diferentes distribuições nos tecidos
(Tabela 2), propriedades cinéticas e especificidade de açúcar. Correntemente há 13
membros desta família (GLUT 1-12). No presente, os membros melhor
caracterizados desta família são os transportadores de glicose classe I (GLUT 1-4)
(KHAN, PESSIN, 2002). Cada membro dessa família consiste de uma cadeia simples
de polipeptídio, com aproximadamente 500 resíduos (Figura 11). A estrutura comum
está presente em 12 segmentos transmembrana (STRYER, 1996).

Fonte: STRYER, 1996
Figura 11: Modelo de um transportador de glicose
O transportador GLUT-1 é onipresente e responsável pela captação de glicose
basal. A isoforma GLUT-2 é primariamente expressa nas células β e no fígado, e tem
uma relativa baixa afinidade pela glicose. GLUT-3 tem alta afinidade e é expresso
durante o desenvolvimento fetal e nos neurônios de adultos. A proteína GLUT-4 é
predominantemente restrita ao tecido adiposo e músculo e responsável pela captação de
glicose dependente de insulina. No estado basal, GLUT-4 está localizado
morfologicamente em um “sistema tubulovesicular”, presente no compartimento
intracelular, enquanto que na presença de insulina, GLUT-4 está localizado na
membrana plasmática do tecido adiposo, músculo cardíaco e esquelético (SLOT, et. al.
1991; SLOT, et al. 1991; SMITH, et al. 1991).

Tabela 2: Transportadores teciduais de glicose
Nome Local de expressão
GLUT1 Vários tecidos; níveis elevados nos eritrócitos, no cérebro, na
placenta, no rim, no cólon e nos tecidos fetais.
GLUT2 Fígado, células β insulares, rim e intestino delgado.
GLUT3 Vários tecidos; níveis altos no cérebro, na placenta e no rim.
GLUT4 Músculo esquelético, coração, tecido adiposo e gordura
marron.
GLUT5 Intestino delgado.
GLUT6
(GLUT3P1)
?
SGLT1 Borda em escova do intestino delgado Fonte: WOOD, TRAYHURN, 2003
4. DIABETES MELLITUS 4.1. CONCEITO
Os mecanismos homeostáticos mantêm os níveis de glicose sanguínea dentro de
uma faixa estreita de 81-99 mg/dl. Este controle é realizado por modulação
hormonal,sendo, basicamente, dois os hormônios reguladores: glucagon e a insulina.
Esses dois hormônios são secretados e liberados pelo pâncreas: glucagon pelas células α
das ilhotas de Langerhans e a insulina pelas células β. Estados em que a homeostasia do
metabolismo de carboidrato, proteínas e lipídeos não é regulada de maneira apropriada
pela insulina, resultam, primariamente, em um aumento dos níveis de glicose sanguínea
em jejum e pós-prandial. Se esse desequilíbrio homeostático não for restabelecido, mas
sim continuar por um período de tempo longo, expondo o sistema endócrino a uma
sobrecarga, ocorre uma exacerbação dos distúrbios metabólicos que resulta em
hiperglicemia, a qual pode evoluir para a síndrome chamada Diabetes mellitus
(TIWARI, RAO, 2002).
O Diabetes Mellitus (DM) é uma desordem, caracterizada por hiperglicemia,
metabolismo alterado dos lipídios, carboidratos e proteínas, e um aumento do risco de
complicações das doenças vasculares (DAVIS, 2006). DM inclui doenças clinicamente

e etiologicamente diferentes, que tem a hiperglicemia em comum, representando mais
uma síndrome do que uma simples doença (BELFIORE, MOGENSEN, 2000).
Suas principais manifestações incluem distúrbios metabólicos, que acarretam um
estado crônico de hiperglicemia (RAMALHO, 1998), sendo a principal causa de
cegueira, retinopatia, nefropatia, neuropatia, amputação dos membros inferiores,
distúrbios cardiovasculares, hipertensão e infarto (MARLES, FARNSWORTH, 1995).
4.2. BREVE HISTÓRICO
DM é uma doença tão antiga como a própria humanidade, sendo hoje a mais
importante patologia que envolve o pâncreas endócrino (RAMALHO, 1998). Seus
sintomas foram descritos há cerca de 3500 anos, no Papiro de Ebers no antigo Egito
(MARLES, FARNSWORTH, 1995). Seu nome, “diabetes mellitus”, origina-se das
palavras Gregas “siphon” e “sugar” e descrevem os sintomas mais óbvios dos diabetes
não controlado: a passagem de uma grande quantidade de urina, que é doce devido ao
seu conteúdo de açúcar. Em 1921, dois cientistas canadenses, Frederick Banting e
Charles Best, descobriram que uma substância misteriosa era produzida por um
pequeno grupo de células, conhecidas como Ilhotas de Langerhans, no pâncreas. Esta
substância, denominada insulin (do Latim islet, que é insula, ilha) provavelmente foi a
descoberta mais importante da história do diabetes. Quando a insulina tornou-se
disponível para tratamento do diabetes, em 1922, foi visto como um milagre médico,
transformando a vida de muitas pessoas (CLARK, 2004)
4.3. PREVALÊNCIA E EPIDEMIOLOGIA
DM é a doença endócrina mundial mais comum. Estima-se que
aproximadamente 173 milhões de pessoas sofram da doença. O número de pessoas com
DM dobrará nos próximos 25 anos, chegando a aproximadamente 370 milhões de
pessoas em 2030 (WHO, 1999, 2003). A maior parte deste aumento ocorrerá nos países
em desenvolvimento (Tabela 3) (BARBOSA-FILHO et al., 2005).

Tabela 3: Total de pessoas com diabetes em alguns países da América do Sul, Central e do Norte País 2000 2030 América do Sul Argentina 1.426.152 2.457.044 Bolívia 206.824 554.527 Brasil 4.553.003 11.305.516 Chile 494.932 1.047.405 Colômbia 883.401 2.410.362 Paraguai 102.237 324.326 Peru 754.087 1.960.957 América Central Cuba 479.612 875.643 Jamaica 80.631 197.573 México 2.178.507 6.130.209 Panamá 59.220 153.308 Trinidad 60.259 124.780 América do Norte Canadá 2.006.107 3.542.974 EUA 17.701.942 30.312.264 MUNDIAL 176.525.312 370.023.002 Fonte: BARBOSA-FILHO et al., 2005
Diabetes é uma situação clínica freqüente, acometendo cerca de 7,6% da
população adulta entre 30 e 69 anos (MALERBI, FRANCO, 1992) e 0,3% das gestantes
(ADA, 2001).
4.4. CLASSIFICAÇÃO
Em 1997, a ADA (American Diabetes Association) emitiu um novo diagnóstico
e critério de classificação (THE EXPERT COMMITTEE ON THE DIAGNOSIS AND
CLASSIFICATION OF DIABETES MELLITUS, 1997). Em 2003, modificações foram
feitas respeitando o diagnóstico da glicose em jejum prejudicada (IFG) (THE EXPERT
COMMITTEE ON THE DIAGNOSIS AND CLASSIFICATION OF DIABETES
MELLITUS: FOLLOW-UP REPORT ON THE DIAGNOSIS OF DIABETES
MELLITUS, 2003). A classificação de DM abaixo inclui as 4 classes clínicas mais
frequentes:
4.4.1. DM Tipo 1: É uma doença auto-imune, resultando da destruição específica das
células β (produtoras de insulina), nas ilhotas de Langerhans (TISCH, MCDEVITT,
1996). DM Tipo 1 compreende duas fases: Insulitis, quando uma população mista de
leucócitos invade as ilhotas; e o diabetes, quando a maioria das células β morrem não

havendo produção suficiente de insulina para regular os níveis de glicose sanguínea,
resultando em hiperglicemia (MATHIS et al., 2001). Uma combinação de fatores
ambientais e genéticos acionam um ataque imune nas células β, ocorrendo em
indivíduos geneticamente susceptíveis (WATKINS, 2003).
Este tipo de DM ocorre principalmente em crianças não-obesas ou adultos
jovens, mas também afeta indivíduos de outras idades. Pacientes adultos podem reter
algum tipo de função celular residual, enquanto crianças e adolescentes freqüentemente
mostram efeito precoce da falta de insulina, com o diabetes aparecendo abruptamente
em dias ou semanas e rapidamente progredindo para complicações agudas, que podem
ser as primeiras manifestações da doença, particularmente na presença de fatores como
a infecções ou estresse (BELFIORE, MOGENSEN, 2001).
Há algumas formas de diabetes Tipo 1 que não possuem etiologia conhecida
(idiopática). Alguns desses pacientes têm insulinopenia permanente e são propensas a
cetoacidose, mas não há evidências de autoimunidade (McLARTY et al., 1990).
4.4.2. DM Tipo 2: Resulta de uma resistência a insulina e secreção de insulina
compensatória insuficiente. A doença tem um início insidioso e pode permanecer
assintomático e não diagnosticado por um longo período (BELFIORE, MOGENSEN,
2000).
DM Tipo 2 é a forma mais comum de diabetes nos adultos, mas sua prevalência
nas crianças está aumentando. Pacientes pediátricos com DM tipo 2 são susceptíveis a
ter sobrepeso ou obesidade, e apresenta glicosúria sem cetonúria, poliúria ausente ou
passageira e polidipsia, e pouca ou nenhuma perda de peso (IZE-LUDLOW,
SPERLING, 2005).
A patologia da DM tipo 2 é caracterizada por resistência periférica à insulina,
regulação anormal da estocagem de glicogênio e declínio da função das células β,
eventualmente levando a falha dessas células (MAHLER, ADLER, 1999). Os eventos
primários são julgados serem um déficit inicial na secreção de insulina, em muito
pacientes, e relativa deficiência à insulina em associação com resistência periférica à
insulina (OLEFSKY, 1989).
Diferenças étnicas e geográficas na incidência de DM Tipo 2 indica que a
doença é uma desordem metabólica complexa, de etiologia diferente, com fatores de
risco social, comportamental e ambiental. É claramente forte o componente hereditário
da doença, que é susceptível ser de natureza multigênica (KIESS, BRANSKI, 2005).

Tabela 4: Características clínicas de pacientes com Diabetes mellitus tipo 1 e 2
Tipo 1 Tipo 2
Idade do início Geralmente <20 anos Geralmente > 30 anos
Massa corporal Baixa a normal Obeso
Insulina Plasmática Baixa a ausente Normal a inicialmente alta
Glucagon Plasmático Alta, pode ser
suprimida
Alta, resistente à supressão
Glicose Plasmática Aumentada Aumentada
Sensibilidade à insulina Normal Reduzida
Terapia Insulina Agentes Orais, insulina Adaptado: GUYTON, 2007 4.4.3. MODY (Diabetes de maturidade com início na juventude ou Tipo 1,5)
Na medida em que têm sido elucidados os processos de patogênese do diabetes,
tanto em relação a marcadores genéticos como aos mecanismos de doença, tem crescido
o número de tipos distintos de diabetes, permitindo uma classificação mais específica e
definitiva. Portanto, novas categorias têm sido acrescidas à lista de tipos específicos de
diabetes, incluindo defeitos genéticos da célula β e da ação da insulina, processos de
doenças que danificam o pâncreas, diabetes relacionado a outras endocrinopatias e os
casos decorrentes do uso de medicamentos (GROSS et al., 2002).
Diabetes da maturidade com início na juventude (MODY-maturity-onset
diabetes of the young) é uma forma geneticamente monogênica heterogênea de diabetes
não insulino dependente, caracterizada pelo início precoce, em relação ao DM tipo 2,
geralmente antes dos 25 anos de idade, freqüentemente em adolescentes e crianças e
pela herança autossômica dominante. Não há associação com o sistema HLA (Human
Leukocyte Antigens), nem evidências de autoimunidade mediada por células. É
estimado que 2-5% dos pacientes com DM podem ter esse tipo de diabetes. Estudos
clínicos têm mostrado que indivíduos MODY pré-diabéticos têm sensibilidade à
insulina normal, mas sofrem de um defeito na secreção de insulina estimulada pela
glicose, sugerindo que uma disfunção nas células β pancreáticas é um defeito primário
desta desordem (BELFIORE, MOGENSEN, 2000) 4.4.4. GDM (Diabetes mellitus Gestacional)
O GDM é definido como um grau de intolerância à glicose, com o início durante
a gestação. Deve ser distinguido pela ligeira deterioração da tolerância à glicose que

pode ocorrer durante a gestação normal (particularmente no 3º trimestre de gravidez). A
prevalência de GDM compreende o intervalo de 2-3% das gestantes, dependendo das
diferenças raciais/étnicas da subpopulação estudada (BELFIORE, MOGENSEN, 2000).
Os fatores de risco associados ao diabetes gestacional são semelhantes aos
descritos para o diabetes tipo 2, incluindo, ainda, idade superior a 25 anos, ganho
excessivo de peso na gravidez, deposição excessiva de gordura corporal central, baixa
estatura, crescimento fetal excessivo, hipertensão ou pré-eclâmpsia na gravidez,
antecedentes obstétricos de morte fetal ou neonatal (GROSS et al., 2002).
GDM é um problema sério e seu reconhecimento é importante para prevenir a
associação de morbidade e mortalidade perinatal e complicações maternais (parto
cesareano e hipertensão crônica). No GDM geralmente a paciente retorna o estado de
tolerância normal à glicose depois do parto, mas 60% das mulheres com GDM podem
desenvolver diabetes dentro de 15 anos após o parto (BELFIORE, MOGENSEN, 2000).
4.5. DIAGNÓSTICO DE DM
Os critérios diagnósticos baseiam-se na glicose plasmática de jejum (FPG -
fasting plasma glucose) (8 horas), nos pontos de jejum, entre as refeições, e de 2h após
uma sobrecarga oral de 75g de glicose - teste oral de tolerância à glicose (OGTT - oral
glucose tolerance test) e na medida da glicose plasmática casual (Tabela 5) (GROSS et
al., 2002)
Tabela 5: Critérios para o diagnóstico de diabetes
• Sintomas: poliúria, polidipsia, perda de peso inexplicável;
• Glicose plasmática casual*: > 200 mg/dl (11.1 mmol/l)
• FPG (glicose plasmática de jejum**): > 126 mg/dl (7 mmol/l)
• OGTT (teste de tolerância oral à glicose)*** Adaptado: American Diabetes Association (ADA), 2004.
* Casual é a medida dos níveis de glicose em qualquer período do dia, onde o paciente não está em jejum; ** Jejum é definido como ingestão não calórica por pelo menos 8h, antes do teste; ***O teste deve ser executado Segundo a Organização Mundial de Saúde, usando uma sobrecarga de glicose contendo o equivalente a 75g de glicose dissolvida em água.
A medida apenas da glicose plasmática de jejum é considerada pela ADA o
método de escolha para o diagnóstico do diabetes e o teste oral de tolerância à glicose

não deveria ser utilizado rotineiramente, mas apenas em algumas situações clínicas ou
para fins de pesquisa (THE EXPERT COMMITTEE ON THE DIAGNOSIS AND
CLASSIFICATION OF DIABETES MELLITUS, 1997). A glicose plasmática de jejum
é mais econômica, de fácil execução, favorecendo a realização em um maior número de
pessoas e apresenta um menor coeficiente de variação inter-individual do que o OGTT
(GROSS et al., 2002). Tabela 6: Diagnóstico do diabetes mellitus e alterações da tolerância á glicose de acordo com valores de glicose plasmática (mg/dl)
CATEGORIA Jejum OGTT (75g-2H) Casual Normal <110 <140 Glicose Plasmática
de jejum (FPG)
alterada
≥110 e <126
____
Tolerância à glicose
diminuída <126 ≥140 e <200 ____
Diabetes mellitus ≥126 ≥200 ≥200 com sintomas
Diabetes gestacional ≥110 ≥140 ____ Fonte: GROSS et al., 2002
O rastreamento de diabetes deve ser realizado em todo indivíduo com mais de 45
anos de idade a cada 3 anos, ou mais precocemente e mais freqüentemente em
indivíduos assintomáticos quando apresentarem fatores de risco para o desenvolvimento
de diabetes (Tabela 7). Tabela 7: Fatores de risco para o diabetes mellitus
• Idade acima de 45 anos; • Obesidade (>120% peso ideal); • Histórico familiar de diabetes em parentes de 1º grau; • Diabetes gestacional; • Hipertensão arterial sistêmica; • HDL-colesterol abaixo de 35mg/dl e/ou triglicerídeos
acima de 250 mg/dl; • Alterações prévias da regulação da glicose; • Indivíduos membros de populações de risco (afro-
americanos, hispano-americanos e outras). Fonte: GUYTON, 2006

4.6. SINTOMAS Os sintomas do diabetes clássico são causados pela prolongada hiperglicemia.
Uma vez a concentração de glicose excede o limiar para reabsorção (aproximadamente
180 mg/dL), a glicosúria resulta em diurese osmótica (poliúria), desidratação e sede
(polidipsia). Ao longo do tempo, a crescente diminuição da captação de glicose pelos
tecidos, resulta em glicosúria crônica, quebra dos aminoácidos para gliconeogênese, e
quebra dos triglicerídeos para suprir os ácidos graxos para a cetogênese, contribuindo
assim com a perda de peso (HALLER et al., 2005).
4.7. COMPLICAÇÕES As complicações semelhantes podem ser devastadoras nos efeitos a longo prazo,
incluindo cegueira, causada pela retinopatia; falha renal, causada pela nefropatia
diabética e diminuição da sensibilidade à dor, causada pela neuropatia diabética.
Embora, os sintomas raramente afetem crianças e adolescentes com diabetes, mudanças
microvasculares subclínicas, podem ser detectadas durante um período precoce. A
infância e a adolescência são períodos durantes os quais a educação intensiva e
tratamento podem prevenir ou atrasar o início das complicações (GLASTRAS et. al.,
2005).
Doenças microvasculares específicas causadas pelo diabetes na retina, glomérulo
e nervos têm patofisiologia similar. A hiperglicemia causa anormalidades no fluxo
sanguíneo e aumento da permeabilidade vascular, isso reflete diminuição da atividade
das substâncias vasodilatadores, semelhante ao óxido nítrico, aumento da atividade de
substâncias vasoconstritores, semelhante a angiotensina II, endotelina I, e a elaboração
de fatores de permeabilidade semelhante a fatores de crescimento endotelial vascular
(VEGF- vascular endothelial growth factor). Anormalidades quantitativas e qualitativas
da matrix extracelular contribuem para um aumento irreversível na permeabilidade
vascular. Com o tempo, ocorre perda celular microvascular, em parte como um
resultado da morte celular programada, e há progressiva oclusão capilar devido ambos a
superprodução de matriz extracelular induzida por fatores de crescimento, tais como
fator de crescimento e transformação β (TGF-β - transforming growth factor) e a
deposição de proteínas plasmáticas extravazadas ácido Schiff positivas. A hiperglicemia
pode também diminuir a produção de fatores tróficos para as células endoteliais e
neuronais. Juntas essa mudanças levam ao edema, isquemia e neovascularização
induzida por hipóxia na retina, proteinúria, expansão da matriz mesangial e

glomerulosclerose nos rins e degeneração axonal multifocal em nervos periféricos
(BROWNLEE, 2001).
A Hiperglicemia é um fator patogênico primário no desenvolvimento das
complicações (NISHIKAWA et al., 2000). Muitas vias bioquímicas podem ser ativadas
na presença de hiperglicemia, incluindo acúmulo de poliol, formação de produtos finais
de glicação avançada, estresse oxidativo e ativação de proteína cinase C (GABBAY,
1975; GIUGLIANO et al., 1996). O efeito combinado deste mecanismo resulta em
mudanças celulares, funcionais e estruturais (GLASTRAS et. al., 2005).
A elevação dos níveis de glicose no diabetes causa dislipidemia (elevação dos
níveis plasmáticos de colesterol), aumento da lipoproteína de baixa densidade (LDL),
aumento dos triglicerídeos e diminuição da concentração do lipoproteína de alta
densidade (HDL) (EL-HILALY et al., 2006). A dislipidemia, nos dois tipos de diabetes,
exerce um papel significante na manifestação e desenvolvimento da aterosclerose
prematura, levando a doença cardiovascular (CV) e, juntos, eles são as maiores causas
de morbidade cardiovascular e mortalidade em pacientes diabéticos (NATHAN et al.,
1997; FEHER, 2004; WILSON TANG et al., 2004; ADA, 2005).
Em relação ao controle glicêmico, outros fatores são relevantes para o
desenvolvimento das complicações. Durante o diabetes, a idade, histórico familiar de
complicações, fumo, dislipidemia e hipertensão são conhecidas por influenciar no
desenvolvimento das complicações. A presença de uma complicação vascular está
associada com o desenvolvimento de outras (ROSSING et al., 2002) 8. TRATAMENTO
Pessoas com DM Tipo 1 são dependentes de insulina para sobreviver. Deve ser
salientado que não há outra alternativa para o tratamento, a não ser insulina, que deve
ter o regime de tratamento individualizado (KIESS, BRANSKI, 2005). O tratamento
com insulina deve ser realizado durante toda a vida, sem interrupção (MOGENSEN,
2007) (Tabela 8).

Tabela 8: Tipos de insulina Tipo de insulina Início da ação Pico Duração
Insulina de ação ultra rápida
Lispro 15-30 min 30-90 min 3-5 horas
Aspart 10-20 min 40-50 min 3-5 horas
Insulina de Ação Rápida
Regular (R) 30 min -1 horas 2-5 horas 5-8 horas
Insulina de Ação Intermediária e Lenta
NPH (N) 1-2 horas 4-12 horas 18-24 horas
Lantus 1-1½ horas
Sem pico; a insulina é
liberada em um nível
constante
20-24 horas
Levemir ou detemir 1-2 horas 6-8 horas Até 24 horas
Insulinas Pré-mixadas*
Humulin 70/30 30 min 2-4 horas 14-24 horas
Novolin 70/30 30 min 2-12 horas Até 24 horas
Novolog 70/30 10-20 min 1-4 horas Até 24 horas
Humalog mix 75/25 15 min 30 min.-2½ horas 16-20 horas
*As insulinas pré-mixadas são uma combinação de proporções específicas de insulinas de ação intermediária e de ação rápida em um frasco ou caneta (os números indicam a porcentagem de cada tipo de insulina).
O principal efeito indesejável da insulina é a hipoglicemia. A hipoglicemia é
comum e, quando muito grave, pode causar lesão cerebral. A insulinoterapia intensiva
resulta em aumento de três vezes na ocorrência de hipoglicemia grave. A hipoglicemia
induzida pela insulina pode ser seguida de hiperglicemia de rebote (“efeito Somogyi”),
devido à liberação de hormônios contra-reguladores (OWENS, 2001).
Algumas classes de agentes orais (Tabela 9) teoricamente podem prevenir ou
atrasar a diminuição gradual da função das células β pela redução da resistência à
insulina ou necessitar da secreção máxima de insulina. A vantagem geral de agentes
orais inclui potencial adesão e conveniência para o paciente e familiares. A
disponibilidade da terapia oral para o tratamento de DM tipo 2 tem expandido,
fornecendo uma grande flexibilidade para médicos e pacientes. Alguns agentes
oferecem uma maior vantagem terapêutica pela redução da resistência à insulina e suas
complicações, incluindo hipertensão, doenças cardiovasculares, hiperlipidemia. A meta

do tratamento é a normalização do valor da glicose sanguínea e controle das suas
complicações. A última meta do tratamento é a diminuição dos riscos das complicações
agudas e crônicas associadas com diabetes (ADA, 1999)
Tabela 9: Perfil farmacológico e terapêutico dos fármacos mais comumente utilizados no tratamento do diabetes
Classe terapêutica Mecanismo primário de
ação
Possíveis eventos adversos Monitorização Comentários
Sulfoniluréias Secretagogo de insulina
• Hipoglicemia • Ganho de peso
• Glicemia de jejum em 2 semanas
• A1C (3º mês)
• Glimepirida, gliclazida e glipizida são opções preferenciais
Metformina
Inibição da liberação
hepática de glicose
• Diarréia relacionada à dose (auto-limitada a 7 a 10 dias)
• Acidose láctica em pacientes com problema renal
• Creatinina sérica no início do tratamento
• Glicemia de jejum em 2 semanas
• A1C em 3 meses
• Menor ganho de peso em monoterapia ou em tratamento combinado
• Dose máxima efetiva: 2 a 2,5g/dia (titulada)
• Contra-indicações: creatinina > 1,4 em mulheres e >1,6 em homens tratamento de ICC; doença hepática; abuso de álcool
Inibidores da alfa-glicosidase (acarbose)
Retardo na absorção de carboidratos pelo intestino
• Diarréia, dor abdominal e flatulência (intensidade relacionada à dose)
• Glicemia pós-prandial no início
• A1C após 3 meses
• Deve ser tomado no início da refeição
• Dose deve ser lentamente titulada para reduzir eventos adversos
Tiazolidinedionas (rosiglitazona e pioglitazona)
Aumento da sensibilidade à
insulina
• Edema, ganho de peso, ICC, aumento do risco de fraturas.
• Segurança cardiovascular da rosiglitazona sendo reavaliada em função de
• AST e ALT no início do tratamento
• Monitorizar para sinais de sobrecarga líquida e anemia
• Redução da glicemia pode não ser aparente nas primeiras 4 semanas
• A máxima eficácia pode não ser atingida nos primeiros 4-6 meses
• Contra-indiciada se ALT > 2,5

possível aumento do risco de infarto do miocárdio
vezes o limite do normal, em doença hepática, abuso de álcool.
• Cautela no uso de rosiglitazona em ICC ou DCV significativa ou em diabéticos em tratamento com insulina ou nitratos(*)
Glinidas Secretagogo de
insulina de curta duração
• Hipoglicemia
• Glicemia de jejum em 2 semanas
• Glicemia pós-prandial no início
• A1C após 3 meses
• Estimula a secreção pancreática de insulina
• Duração de ação: 1-2 horas
Inibidores da DPP-4 (sitagliptina e vildagliptina)
Restauração dos níveis de GLP-1 e de
GIP
• Efeitos adversos não clinicamente significantes
• Glicemia pós-prandial no início
• Glicemia de jejum após 2 semanas
• A1C após 3 meses
• Ausência de ganho de peso
• Incidência bastante reduzida de hipoglicemias
Análogos do GLP-1 (exenatida)
Estímulo à produção de
insulina, inibição da secreção de glucagon e aumento da saciedade
• Sintomas gastrintestinais
• Glicemia pós-prandial no início
• Glicemia de jejum após 2 semanas
• A1C após 3 meses
• Redução de peso • Indicada para
tratamento combinado com sulfoniluréia, metformina ou glitazona
(*) Recomendações do FDA Advisory Panel em 30 de julho de 2007 Adaptado de: Medical Guidelines for Clinical Practice for the Management of Diabetes Mellitus. American Association of Clinical Endocrinologists (AACE)
4.9. DIAGNÓSTICO, PREVENÇÃO E POSSÍVEL CURA
4.9.1. DIAGNÓSTICO DO DIABETES TIPO 1
Anticorpos das ilhotas
DM Tipo 1 é causada pela destruição autoimune ou disfunção das células
secretoras de insulina, nas ilhotas de Langerhans e representa o ponto final de um
declínio progressivo da função das células β (ATKINSON, MACLAREN, 1994;
EISENBARTH, 1986). Este longo período antes da manifestação do desenvolvimento

da DM Tipo 1 oferece uma larga janela de oportunidade não apenas para predizer o
início da doença, mas também para intervir com agentes terapêuticos seguros
(EISENBARTH, 1986).
Semelhante a muitas doenças auto-imune, DM Tipo 1 é caracterizada pelas
respostas autoimunes celular e humoral, dirigida contra múltiplos antígenos alvos das
células pancreáticas. Múltiplos anticorpos estão presentes nos pacientes mais
recentemente diagnosticados com DM Tipo 1 e sua presença é altamente preditiva da
progressão da doença em outros parentes de primeiro grau. O diagnóstico imunológico
das doenças autoimunes invoca principalmente a detecção de autoanticorpos no soro
dos pacientes. O corrente uso de marcadores para predizer o estudo são anticorpos das
células das ilhotas (ICA), autoanticorpo ácido glutâmico descarboxilase (GAD65 AA),
autoanticorpo IA-2 proteína semelhantes à tirosina fosfatase (IA-2/ICA512 AA) e
autoanticorpo insulina (IAA).
4.9.2. PREVENÇÃO
O processo da doença pode ser prevenido usando diferentes abordagens
terapêuticas. Estratégias de prevenção podem ser classificadas baseadas no tempo de
intervenção. (1) Prevenção Primária: objetiva retardar a doença de alto risco na
população, antes de alguma evidência sorológica de imunidade das ilhotas; (2)
Prevenção secundária: objetiva retardar e, possivelmente, suprimir os danos às células β
em indivíduos euglicêmicos, com evidências de imunidade às ilhotas e (3) Prevenção
terciária: Começa após o início do diabetes, objetiva induzir uma remissão prolongada
ou regeneração das células β. Isto pode ser útil para prevenir a função das células β
remanescentes, que tem sido relacionado com a reversão do quadro de destruição destas
células (GROUP, 1998; GROUP, 1987; ROSENBLOOM et al., 2000; BOUGNERES et
al., 1990). Voluntários sadios com parentes de primeiro grau com DM tipo 1
representam uma população altamente motivada e acessível para ser investigada e
incluída na triagem preventiva primária e secundária. Se estes voluntários forem
diagnosticados posteriormente como pacientes diabéticos serão incluídos na estratégia
de prevenção terciária (CASU et al., 2005).

4.9.3. CURA
• Transplante do pancreas ou ilhotas
A maior meta das investigações clínicas em pacientes com DM tipo 1 é restabelecer
a secreção de insulina fisiológica depois da disfunção pancreática ou das ilhotas nos
pacientes diabéticos. Embora o transplante do pâncreas tem provado ser efetivo em
normalizar os níveis de glicose com parcial ou completa restauração da produção de
insulina em grupos seletivos de pacientes, o transplante de ilhotas deu resultados menos
promissores (BOTTINO et al., 2002). Isto pode assinalar que o transplante do pâncreas
ou das ilhotas não pode ser considerado como um procedimento para salvar vidas, e os
riscos ou benefícios deste procedimento devem ser minuciosamente pesados,
particularmente se lidam com pacientes pediátricos que necessitam de uma longa terapia
imunossupressora crônica para prevenir a rejeição do enxerto. A melhoria dos novos
regimes imunossupressivos é um importante objetivo para conseguir o transplantes do
pâncreas e ilhotas como um padrão de cura para DM tipo 1 (BOTTINO et al., 2002).
• Terapia com células tronco, terapia gênica e outras terapias
Com o aumento do interesse pelo transplante das ilhotas, numerosos impedimentos
tem surgido, incluindo o número insuficiente de doadores para render um número
suficiente de ilhotas transplantadas e a necessidade de uma longa terapia
imunossupressora para prevenir a rejeição e a recorrência da autoimunidade. Estes
maiores obstáculos têm deslocado o interesse para abordagens alternativas, criando
menos imunoreatividade e potencialmente intermináveis fontes alternativas das ilhotas
ou para regenerar as células β (TRUCCO, 2005). A inuficiência de ilhotas
transplantadas pode ser superada pelo número de novas abordagens, incluindo linhagens
de células produtoras de insulina transformadas, transferência de tipos celulares
diferentes, capazes de produzir insulina, transdiferenciação in vivo de células do fígado
e isolamento de ilhotas porcinas xenogenéticas (BOTTINO et al., 2002; TRUCCO,
2004, TRUCCO, 2005;). Células tronco do embrião podem aumentar as fontes
potenciais ilimitadas de células produtoras de insulina (CASU et al., 2005).
Uma estratégia semelhante é baseada na conversão de células não-ilhotas dos
pacientes de diferentes linhagens em células produtoras de insulina. Isto tem sido
realizado usando engenharia genética dos hepatócitos que são capazes de produzir
insulina depois que são transferidos com pdx1 sob o controle do promotor insulina 1 de
ratos (FERBER et al., 2000). Embora isso não tenha sido já confirmado, esta

observação sugere que a engenharia para substituir células β ou células produtoras de
insulina obtidas de células de diferentes linhagens pode ser explorada para restabelecer
a secreção de insulina (CASU et al., 2005). 4.10. DIABETES EXPERIMENTAL
O diabetes experimental pode ser induzido em animais, por vários mecanismos.
Infelizmente, porém, em grande número de métodos, o agente utilizado não é capaz de
desenvolver a maior parte da fisiopatologia do diabetes humano. Os principais
mecanismos de produção de diabetes são: estresse, infecções, toxinas, ou manipulações,
incluindo a pancreatectomia, lesões do sistema nervoso central, uso de hormônios anti-
insulínicos, exposição à hidrocortisona ou hormônio adenocorticotrófico (ACTH),
indução por vírus e o uso de agentes químicos beta-citotóxicos (LERCO et al., 2003).
A indução de diabetes experimental nos ratos, usando substâncias químicas que
destroem células β pancreáticas é muito conveniente e simples. As substâncias indutoras
de diabetes mais usuais são aloxana e estreptozotocina (STZ) (SZKUDELSKI, 2001).
4.10.1. ALOXANA Aloxana (2, 4, 5, 6-tetraoxipirimidina) (Figura 12) foi primeiro descrita por
Brugnatelli em 1818. Wöhler and Liebig usaram o nome “aloxana” e descreveram sua
síntese a partir da oxidação do ácido úrico (LENZEN, PANTEN, 1988). As
propriedades diabetogênicas desta droga foram relatadas muitos anos mais tarde por
DUNN, SHEEHAN, MCLETHIE (1943), que estudaram o efeito de sua administração
em coelhos e relataram a necrose específica nas ilhotas pancreáticas. Desde então,
aloxana tem sido comumente utilizada como modelo animal para DM tipo 1.
HN
NH
O O
O
O
Figura 12: Estrutura química da aloxana

A aloxana exerce sua ação diabetogênica quando ela é administrada
parenteralmente: intravenosa, intraperitoneal ou subcutânea. A dose de aloxana
requerida para induzir diabetes depende da espécie animal, da via de administração e
estado nutricional. As ilhotas humanas são consideravelmente mais resistentes a aloxana
do que ilhotas de ratos e camundongos (EIZIRIK et al., 1994). A dose intravenosa desta
droga mais freqüentemente usada para induzir diabetes em ratos é 65 mg/kg de peso do
animal (GRUPPUSO et al., 1990; BOYLAN et al., 1992). Animais em jejum são mais
susceptíveis a aloxana (KATSUMATA et al., 1992; SZKUDELSKI et al., 1998),
considerando que o aumento da glicose sanguínea, proveniente da alimentação, fornece
proteção parcial ao agente (BANSAL et al., 1980; SZKUDELSKI et al., 1998).
Lukens, 1948, revisando os trabalhos de vários autores, cita que animais que não
respondem a uma primeira injeção de aloxana podem ser refratários a injeções
posteriores de doses similares. Nesse aspecto, os autores têm observado que os animais
que apresentam este comportamento geralmente são do grupo em que o jejum não foi
convenientemente observado. Esses mesmos animais, refratários às doses subseqüentes,
tornam-se diabéticos após 60 horas de jejum, com uma dose padrão de aloxana.
Outro aspecto a considerar são as vias de administração da droga. A via
endovenosa tem sido a de escolha na maioria das espécies animais, onde os efeitos da
aloxana são mais evidentes. As vias subcutânea e intraperitoneal também são
satisfatórias em ratos, enquanto que, em coelhos, doses muito elevadas devem ser
ministradas por essas vias para que se atinja o mesmo efeito diabetogênico. Finalmente,
a via oral é usualmente insatisfatória para a ação da aloxana, muito embora o diabetes
possa ser produzido em ratos, coelhos e gatos, se a droga for rapidamente ingerida com
comida, após um jejum prévio prolongado (LUKENS, 1948). Quando a via endovenosa
é usada, a velocidade de infusão da droga é importante. Doses efetivas de aloxana não
produzem diabetes se as mesmas são injetadas muito lentamente (LERCO et al., 2003).
LUKENS (1948) afirma a importância do peso, em relação à susceptibilidade do
animal aos efeitos tóxicos da aloxana. Em um estudo detalhado em ratos, esse
pesquisador revelaram que a toxicidade da aloxana altera-se numa relação linear com o
peso do animal,
A aloxana é uma substância hidrofílica e instável. Sua meia-vida, no pH neutro e
à 37ºC, é aproximadamente de 1,5 min e é mais longa em temperaturas mais baixas
(LENZEN, MUNDAY, 1991). Por outro lado, quando a dose diabetogênica é usada, o
tempo de decomposição da aloxana é sufucientemente reduzido para permitir que ela

chegue ao pâncreas em quantidades que são deletérias as ilhotas pancreáticas
(SZKUDELSKI, 2001).
A ação da aloxana no pâncreas é precedida por sua rápida captação pelas células
β (WEAVER et al., 1978a; BOQUIST et al., 1983). A rápida captação pelas células
secretoras de insulina tem sido proposta ser uma importante característica determinante
da diabetogenicidade da aloxana. Outro aspecto diz respeito à formação de espécies
reativas de oxigênio (HEIKKILA et al., 1976). Uma captação similar da aloxana
acontece no fígado. Contudo, o fígado e outros tecidos são mais resistentes a espécies
reativas de oxigênio, em comparação a células β, e essa resistência os protegem contra a
toxicidade da aloxana (MALAISSE et al., 1982; TIEDGE et al., 1997). Nas células β do
pâncreas a aloxana exibe alta afinidade por compostos celulares contendo tiol (SH), ela
reduz a glutationa (GSH), cisteína e grupos sulfidril ligantes de proteína (incluindo
enzimas contendo SH) são muitos susceptíveis a sua ação (LENZEN, MUNDAY,
1991).
A aloxana, por si só, não é tóxica. Ela é reduzida extracelularmente para ácido
dialúrico na presença de um agente redutor (por exemplo: cisteína). Oxidação do ácido
dialúrico na presença de oxigênio resulta na produção de radicais, ânions superóxidos e
peróxido de hidrogênio, o último pode se difundir através da membrana plasmática e
entra no interior da célula (ZHANG et al., 1995; SZKUDELSKI, 2001).
Estudos indicam que a glicose diminui a citoxicidade da aloxana, in vitro e in
vivo. O efeito protetor da glicose contra a morte por necrose das células β pode ser
devido a interação do açúcar com o transportador de glicose (GLUT2), resultando em
captação limitada de aloxana (JÖRNS et al., 1997).
Usando aloxana para induzir diabetes, os animais devem ser examinados depois
de um certo período, para minimizar os efeitos adversos da ação da aloxana. Deve ser
enfatizado que o intervalo da dose diabetogênica da aloxana é muito restrita (LENZEN
et al., 1996).
4.10.2. ESTREPTOZOTOCINA (STZ)
Estreptozotocina (STZ-2-deoxi-(3-(metil-3-nitrosoureido)-D-glicopiranose),
estrutura representada na figura 13, é sintetizada pelo Streptomycetes achromogenes e é
usado para induzir diabetes em modelos experimentais (IARC, 1978).

O
NH
OH
CH2OH
H
OH
C
N
CH3
NO
O
HO
Figura13: Estrutura da Estreptozotocina (STZ)
O intervalo da dose da STZ não é tão restrita como da aloxana. A dose
freqüentemente usada, intravenosamente, para induzir DM tipo 1 é entre 40 e 60 mg/kg
de peso do animal (GANDA et al., 1976), mas doses mais altas são também usadas. DM
tipo 2 é facilmente induzida em ratos, pela administração intravenosa ou intraperitoneal,
com 100 mg/kg de peso do animal de STZ, no dia do nascimento (SZKUDELSKI,
2001).
A ação da STZ nas células β é acompanhada pelas características da insulina
sanguínea e concentração da glicose. Duas horas depois da injeção, a hiperglicemia é
observada com a concomitante diminuição da insulina sanguínea. Aproximadamente
seis horas mais tarde, hipoglicemia ocorre com um aumento dos níveis de insulina.
Finalmente a hiperglicemia se desenvolve e há uma diminuição dos níveis de insulina
sanguínea (WEST et al., 1996). Estas mudanças nas concentrações de glicose e insulina
refletem as anormalidades nas funções das células β. STZ prejudica a oxidação da
glicose (BEDOYA et al., 1996) e diminui a biosíntese e secreção da insulina (BOLAFFI
et al., 1987; NUKATSUKA et al., 1990b).
STZ é captada pelas células β pancreáticas via transportador de glicose
(GLUT2). A expressão reduzida de GLUT2 previne a ação diabetogênica da STZ
(SCHNEDL et al., 1994; THULESEN et al., 1996).
Com relação ao mecanismo de ação, STZ pode danificar o DNA, que ativa a
sintetase de ADP ribose, levando a depleção da síntese de pró insulina (YAMAMOTO
et al., 1981a; YAMAMOTO et al., 1981b; UCHIGATA et al., 1982). Muitos
mecanismos pelos quais STZ danifica as células β têm sido propostos (Figura 14): (1)
STZ é um forte agente alquilante, causando alquilação direta do DNA pelo •CH3 ou

CH3+ , via a decomposição da STZ (BENNET, PEGG, 1981; JOHANSSON, TJA¨LVA,
1978), (2) A geração de espécies reativas de oxigênio ou óxido nítrico (NO) pela STZ
participa da toxicidade da STZ na diabetogênese (ROBBINS et al., 1980; PAPACCIO
et al., 1986). Baixas doses de STZ podem causar inflamação não-específica das ilhotas,
com infiltração pelas células mononucleares e produção e liberação de NO derivado de
macrófago bem como de superóxido, isto é, pode produzir efeitos citotóxicos
indiretamente através da inflamação (LIKE, ROSSINI, 1976; LIKE et al., 1978;
ANDRADE et al., 1993).
O
NH
OH
CH2OH
H
OH
C
N
CH3
NO
O
HO
Adaptado: MURATA et al., 1999
ESTREPTOZOTOCINA (STZ)
Decomposição Decomposição ou
metabolização
CH3+ ou •CH3 •NO e/ou O2
-
(I) Metilação do DNA (II) Modificação do DNA
Figura 14: Mecanismos propostos da toxicidade induzida pela STZ. Dois mecanismos
diretos: (I) metilação do DNA, induzida por CH3+ ou CH3• e (II) modificação do DNA,
induzida pela radicais reativos de oxigênio ou NO pela STZ. O2-.
Alguns autores preferem utilizar a STZ ao invés da aloxana, pois esta primeira
produz um diabético permanente e apresenta menor taxa de mortalidade quando
comparada com a AL (ISLAS-ANDRADE, 2000). Porém ambas as drogas produzem
hiperglicemia sendo que a AL tem um custo financeiro menor do que a STZ, por isso a
Aloxana foi utilizada neste estudo.

4.11. DIABETES MELLITUS E AS PLANTAS MEDICINAIS
Muitas espécies de plantas têm sido usadas etnofarmacologicamente ou
experimentalmente para tratar dos sintomas do diabetes mellitus (IVORRA et al., 1989;
LAMBA et al., 2000; SAXENA, VIKRAM, 2004). Estas plantas estão distribuídas em
mais de 725 gêneros em 183 famílias (Tabela 10), estendendo-se desde as algas
marinhas e fungos até plantas (NEGRI, 2005). Tabela 10: Famílias das plantas que possuem atividade hipoglicemiante
Família Espécies citadas Total de espécies
Fabaceae 127 18.000
Asteraceae 98 21.300
Lamiaceae 36 3.500
Liliaceae 35 6.460
Poaceae 30 10.000
Euphorbiaceae 30 7.000
Fonte: Marles, Farnsworth, 1995
A maioria das plantas que são utilizadas como antidiabéticas ao serem avaliadas
farmacologicamente demonstraram ter atividade hipoglicemiante e possuir constituintes
químicos que podem ser utilizados como modelos para novos agentes hipoglicemiantes.
Entretanto, as análises posteriores revelaram grande variedade de mecanismos de ação
que podem levar ao efeito hipoglicemiante, nem todos terapeuticamente úteis
(MARLES, FARNSWORTH, 1995; HUO et al., 2003; SAID et al., 2002).
O mecanismo de ação pelos quais as plantas reduzem a taxa de glicose do
sangue pode ser atribuído aos seguintes fatores: aumento da liberação de insulina
através da estimulação das células β-pancreáticas; resistência aos hormônios que
aumentam a taxa de glicose; aumento do número e da sensibilidade do sítio receptor de
insulina; diminuição da quebra de glicogênio; inibição da α-glicosidase; aumento do
consumo de glicose nos tecidos e órgãos; resistência à peroxidação de lipídeos; correção
da desordem metabólica causada em lipídeos e proteínas (MARLES, FARNSWORTH,
1995; HUO et al., 2003; SAID et al., 2002, LI et al., 2004; VOLPATO et al., 2002).
Várias espécies são estudadas quanto à atividade farmacológica hipoglicemiante.
Extratos dos frutos, folhas e raízes de Momordica charantia Linn (Cucurbitaceae) são
eficientes no tratamento do diabetes tipo II (MIURA et al., 2004; SENANAYAKE et
al., 2004). O extrato de Mormodica charantia continuou a exercer um efeito

hipoglicêmico, mesmo quando a maior parte das células β foi destruída, indicando um
efeito insulinomimético direto (GROVER et al., 2001). Plantas comestíveis, tais como
Allium cepa L. (Liliaceae) (cebola) e Allium sativum L. (Liliaceae) (alho) usadas no
tratamento do diabetes são caracterizadas por possuir concentração baixa de carboidrato
e gordura, além de impedir as complicações cardiovasculares diabéticas
(BALUCHNEJADMOJARAD et al., 2003).
Aloe vera (L.) Burm. Fil. (Liliaceae) (babosa) tem sido muito usada em todo o
mundo, devido às suas propriedades medicinais. O extrato aquoso da folha de Aloe vera
mostrou atividade hipoglicêmica em ratos com diabetes tipo I e II. No diabetes tipo II
apresentou maior atividade do que a droga padrão, a glibenclamida (OKYAR et al.,
2001).
Extrato aquoso das folhas secas de Eucalyptus globulus Labill (Myrtaceae)
exerceu atividade antidiabética sobre os camundongos que tiveram o diabetes mellitus
induzido por STZ. Entretanto, não apresentou efeito nos camundongos com diabetes
induzido por aloxano (WORLD HEALTH ORGANIZATION, 2002)
No Brasil, aproximadamente 200 plantas são usadas empiricamente no controle
do diabetes mellitus. Destas, 52 têm sido experimentalmente estudadas e a atividade
hipoglicemiante é detectada na maioria delas. B. forficata, popularmente conhecida
como “Pata-de-vaca” é a espécie mais estudada. Alguns estudos confirmam a atividade
outros não (BARBOSA-FILHO et al., 2005). Outras espécies do gênero Bauhinia são
amplamente utilizadas. O extrato de Bauhinia divaricata Linn (Leguminosae) é usado
para tratar variedade de enfermidades, tais como problemas gastrintestinais e
respiratórios, sendo também usado freqüentemente no tratamento do diabetes. A
atividade hipoglicemiante desta planta é atribuída à sua capacidade de inibir a α-amilase
(ANKLI et al., 2002). A fração n-butanólica do extrato das folhas de Bauhinia forficata
Link (Leguminosae) foi efetiva no decréscimo do nível de glicose, provavelmente
agindo através da redução da sua absorção intestinal (SILVA et al., 2002). No entanto,
esta melhora não parece estar relacionada à inibição da glicogênese, nem mesmo o
extrato parece agir de maneira similar à insulina ou sulfoniluréias, embora possa agir
através da inibição da neoglicogênese de maneira similar à biguanida (PEPATO et al.,
2002). O extrato aquoso das folhas de Bauhinia megalandra G. (Leguminosae) inibiu a
absorção da glicose pelo intestino, efeito este dependente da concentração do extrato
(GONZALEZ-MUJICA et al., 2003).

Tabela 11: Plantas brasileiras antidiabéticas NOME POPULAR SINONÍMIA NOME CIENTÍFICO PARTE USADA
1. Abagiru Bagiru Chrysobalamus icaco Folhas
2. Bardana Carrapicho-grande, pega
massa
Arctium lappa Raiz, sementes,
folhas
3. Cajueiro Acaju Anacardium occidentale Cascas, folhas,
frutos
4. Carambola Camerunga Averrhoa carambola Raízes, folhas
5. Carqueja Carqueja amargosa Baccharis genistelloides Toda
6 Dente-de-leão Alface-de-cão, taraxaco Taraxacum officinale Folhas
7. Estévia Kaahette Stevia rebaudiana Folhas
8. Eucalipto Árvore da febre Eucaliptus globulus Folhas
9. Graviola Jaca-de-pobre, graviola
do norte
Anona muricata Raiz, fruto, folhas
10. Jambo Jambo vermelho, jambo
rosa
Eugenia jambolana Polpa de fruto,
caroço
11. Jambolão Jamelão Szygium jambolanum Fruto
12. Jucá Pau-ferro, jacaína Caesalpinea férrea Cascas, sementes,
raízes, favas
13. Melão-de-são-caetano Erva-de-são-caetano Mormodica charantia Fruto
14. Pata-de-vaca Unha-de-vaca. Mororó Bauhinia forficata Folhas
15. Quixaba Quixabeira, sacutiaba Bumelia sartorum Folhas
16. Romã Punica de muitos grãos,
romeira
Punica granatum Caule, raiz, flores e
frutos
17. Sálvia Selima fina, salva dos
jardins
Salvia officinallis Toda a planta
Fonte: ALMEIDA, AGRA, 1986
Há muitas substâncias extraídas de plantas que reduzem o nível de glicose no
sangue e a grande variedade de classes químicas indica que vários de mecanismos de
ação devem estar envolvidos na redução do nível de glicose no sangue. Algumas destas
substâncias podem ter potencial terapêutico, enquanto outras podem produzir
hipoglicemia como efeito colateral devido à sua toxicidade, especialmente
hepatotoxicidade (IVORRA et al., 1989; PÉREZ GUTIÉRREZ, 2002a; WANG, NG,
1999; BAE et al., 1999; PÉREZ GUTIÉRREZ et al., 1998; LAMBA et al., 2000).

5. ANTIOXIDANTE 5.1. INTRODUÇÃO
As Plantas constituem uma importante fonte de produtos naturais ativos, que
diferem amplamente em termos de propriedades biológicas e estruturais. Nos anos
recentes, a prevenção do câncer e doenças cardiovasculares tem sido associada com a
ingestão de frutos frescos, vegetais e chás ricos em antioxidantes naturais (VIRGILI et
al., 2001; JOHNSON, 2001). Há fortes sugestões que a grande ingestão destes
compostos está associada com o menor risco de mortalidade destas doenças, bem como
de diabetes (MCCUNE, JOHNS, 2002).
Existem no nosso organismo processos normais que geram radicais livres. A
oxidação é a transferência de elétrons de um átomo a outro e representa uma parte
essencial da vida aeróbica de nosso organismo, uma vez que o oxigênio é o último
aceptor de elétrons na cadeia de transporte de elétrons levando a produção de energia na
forma de ATP. No entanto, problemas podem aparecer quando a cascata de elétrons
torna-se desemparelhada, gerando radicais livres. Exemplos de radicais livres do
oxigênio, conhecido como espécies reativas de oxigênio, incluem o superóxido (O2•-),
peroxila (ROO•), alcoxila (RO•), hidroxila (HO•), e óxido nítrico (NO•). Os radicais
livres hidroxila e alcoxila são muito reativos e rapidamente atacam moléculas em
células próximas, e provavelmente o dano causado por eles é inevitável e é tratado por
processos de reparo (AMES et al., 1993; PIETTA, 2000). Por outro lado, o ânion radical
superóxido, hidroperóxidos lipídicos e o óxido nítrico são menos reativos. Em
acréscimo a estas espécies reativas de oxigênio, existem no organismo espécies não
radicalares, tal como o peróxido de hidrogênio (H2O2) e o ácido hipocloroso (HOCl)
(PIETTA, 2000).
As espécies reativas de oxigênio desempenham importantes papéis no
organismo. Alguns são positivos e relacionados aos seus envolvimentos na produção de
energia, fagocitose, regulação do crescimento e sinalização celular e síntese de
compostos biologicamente importantes (HALLIWELL, 1997). Entretanto, as espécies
reativas de oxigênio podem ser muito danosas para o organismo, a partir do momento
em que elas podem atacar lipídios em membranas celulares, proteínas em tecidos ou
enzimas, carboidratos e DNA, induzir oxidação de tecidos, podendo causar danos à
membrana, modificação de proteínas (incluindo enzimas), e danos ao DNA. Este dano
oxidativo é considerado a principal causa do processo de envelhecimento e de várias

doenças degenerativas associadas a isto, tal como doenças do coração, cataratas,
disfunção cognitiva e câncer (BLAKE e WINYARD, 1995; HALLIWELL e
GUTTERIDGE, 1998).
Radicais livres estão sendo associados com a patogênese de várias desordens,
semelhante ao câncer, diabetes, doenças cardiovasculares, doenças autoimunes e
desordens neurodegenerativas e também estão implicadas com a idade. Antioxidantes
estão emergindo como agentes profiláticos e terapêuticos, que seqüestram radicais livres
e previnem os danos causados por eles (RATNAM, et al., 2006).
Diabetes mellitus é uma desordem metabólica caracterizada por hiperglicemia e
insuficiência da secreção ou insensibilidade do receptor a insulina endógena (ROLO,
PALMEIRA, 2006). O quadro de hiperglicemia em diabetes está quase sempre
acompanhado de disfunções lipídicas (dislipidemias e aumento de ácidos graxos livres)
e protéicas (inativação de enzimas e fragmentação de proteínas,entre outras) (KUROKI
et al., 2003). Essas alterações metabólicas podem levar ao desenvolvimento de
patologias associadas a diabetes como catarata, microangiopatias, nefropatias,
aterosclerose, infecções e alterações neurológicas (KING, BROWNLEE, 1996,
UCHIDA, 2000, LEE et al., 2003). Hiperglicemia causa estresse oxidativo. Há muitas
evidências que o estresse oxidativo está envolvido na etiologia de várias complicações
diabéticas (GIUGLIANO, CERIELLO, 1996; MCDONAGH, HOKAMA, 2000).
Estresse oxidativo resulta quando a taxa de produção oxidante excede a taxa de
sequestro oxidante. No diabetes ou na resistência à insulina, a falha na captação de
glicose estimulada pela insulina, no tecido adiposo e músculos, leva a um aumento na
concentração alta de glicose no sangue. O aumento do fluxo de glicose aumenta a
produção oxidante e prejudica as defesas antioxidantes por múltiplos caminhos (KING,
LOEKEN, 2004). Estresse oxidativo crônico ou excessivo pode interferir com a função
normal dos tecidos pela hiperglicemia diabética, pelo aumento do fluxo sanguíneo e
distúrbios hemodinâmicos na retina (KUNISAKI, et al 1995; KOWLURU, KENNEDY,
2001), contratibilidade das células do músculo liso vascular (SHARPE, et al. 1998), e
diminuição da condutibilidade dos nervos periféricos (HOUNSOM, et al. 2001).
Quando as células β são expostas a altas concentrações de glicose por um
período prolongado de tempo, a glicose satura a rota normal de glicólise e desvia para
um caminho alternativo, nos quais as espécies reativas de oxigênio (EROs) são geradas
por distintos processos metabólicos, dentro e fora da mitocôndria (ROBERTSON,
2006). Uma grande quantidade de dados enfatizam quatro caminhos metabólicos como

os maiores contribuintes para os danos celulares induzidos por hiperglicemia (Figura
15)(NISHIKAWA, et al 2000; Brownlee, 2001; Robertson, 2004): (1) aumento do fluxo
através da via dos polióis; (2) aumento da formação dos produtos finais de glicação
avançada (AGEs); (3) ativação da isoforma da proteína cinase C (PKC); e (4) aumento
do fluxo da via da hexosamina (Figura 16) (KING, LOEKEN, 2004).
DANO MACROMOLECULAR
↑ ROS MITOCONDRIAL
Via da Proteína cinase C
↑ DAG
Via dos AGEs
Via da hexosamina
↑ AGEs
Via dos Polióis
↑PKC
↑ GLICOSE
↑ UDP-GlcNac
↑ Sorbitol
Adaptação: ROLO, A. P., PALMEIRA, 2006 Figura 15: Geração de EROs induzida pela hiperglicemia e conseqüente ativação das
vias patológicas
1. Aumento do fluxo através da via dos polióis: Hiperglicemia resulta no aumento da
conversão enzimática da glicose para o poliálcool sorbitol, com diminuição
concomitante da glutationa e NADPH (BROWNLEE, 2001), resultando na perda de
equivalentes redutores antioxidantes ocasionando um aumento do estresse oxidativo
associado com ROS intracelular A grande produção de superóxidos induzida pela
hiperglicemia inibe a glicose dehidrogenase – 6- fosfato (NISHIKAWA, et al. 2000), a
enzima limitante da via das pentoses, que é requerida para fornecer equivalentes
redutores para o sistema de defesa antioxidante. O produto NADPH é o principal
redutor da célula (ZHANG, et al. 2000; WU, et al. 2001) e a depleção do estoque de
NADPH das células pode inibir outras enzimas que sejam dependentes dessa coenzima
como o sistema glutationa redox (reciclagem de GSH) e NO-sintase (formação de óxido
nítrico) (ROCHA, et al. 2006). O sorbitol é metabolizado em frutose pela sorbitol

dehidrogenase, aumentando a razão NADH/NAD+ (BROWNLEE, 2001). O aumento
nos níveis de frutose leva a um aumento na formação de seus derivados frutose-3-
fosfato e 3- deoxiglicosonas que são agentes da glicação não enzimática mais potentes
do que a glicose e, portanto, o fluxo de glicose na via dos polióis pode aumentar a
formação de AGEs (ROCHA, et al. 2006).
2. Aumento da formação dos produtos finais de glicação avançada (AGEs): AGEs
têm sido implicados na patogênese da maioria das complicações microvasculares da
diabetes mellitus: nefropatia, neuropatia e retinopatia (BASTA, et al. 2004; COOPER,
2004; STITT, et al. 2004).
Os açúcares redutores, como a glicose, podem sofrer uma reação de adição
nucleofílica com os grupos amino livres das proteínas (Figura 16), formando iminas
(base de Schiff). (ROCHA, et al. 2006). A formação da base de Schiff é relativamente
rápida e altamente reversível. Subseqüentemente, o rearranjo da base de Schiff origina o
produto de Amadori. Esta reação é mais rápida do que a reação reversa (ULRICH,
CERAMI, 2001). Conseqüentemente, o produto de glicação Amadori leva ao acúmulo
de proteínas, iniciando o processo de glicação avançada. AGEs surge da autooxidação
da glicose para glioxal, decomposição do produto Amadori para 3-deoxiglicosona, e
fragmentação do gliceraldeído-3-fosfato e dihidroacetona fosfato para metilglioxal
(BROWNLEE, , 2004).
Esses derivados são mais reativos do que os açúcares que lhe deram origem e formam
ligações irreversíveis com grupos amino livres de proteína para formar ligações
cruzadas estáveis (SAKURAI E TSUCHIYA, 1988, WOLFF et al., 1991).
O acúmulo desses produtos finais de glicação avançada é comum no processo de
envelhecimento, na aterosclerose e no Diabetes mellitus e está associado,
especialmente, a proteínas de vida longa como o colágeno, o cristalino e proteínas dos
nervos (YIM et al., 1995).

Fonte: ROCHA, et al. 2006
Figura 16: Processo de glicação não enzimática. Adição de glicose a proteínas
com subseqüente degradação do produto de glicação. Formação de deoxiglicosanas que
reagem com proteínas para formar AGEs.
3. Ativação de isoformas da proteína cinase C (PKC): o aumento nos níveis de
glicose ativa a via de síntese de novo de diacilglicerol (DAG) (Figura 17). O aumento
do conteúdo de DAG ativa várias isoformas de PKC, que são sinalizadores
intracelulares importantes e estão envolvidos na regulação de muitas funções
vasculares. Essa ativa de PKC é responsável por muitas patologias das complicações do
Diabetes (BROWNLEE, 2001).
A ativação da PKC, induzida pela glicose leva a um aumento na produção de
matriz celular e citocinas, assim como uma exacerbação da contratilidade,
permeabilidade e proliferação celular vascular; ativação de fosfolipase A2 citosólica e
inibição de ATPase dependente de Na+/K+ . Esses processos estão associados a várias
anormalidades nos tecidos renal, cardiovascular e na retina (KOYA, KING, 1998).
Além disso, PKC ativa NAD(P)H oxidases na superfície das células vasculares,
levando a um aumento na formação de EROs que contribuem para a fisiopatologia de
várias doenças vasculares como hipercolesterolemia, aterosclerose e hipertensão
(INOGUCHI et al., 2003a e b).

Células do endotélio vascularCélulas do músculo liso
↑ Níveis de glicose
Ativação de PKC
Ativação de NAD(P)H-oxidase
↑ Formação de EROs
↑ Síntese de novo de DAG
Adaptado de: ROCHA, et al. 2006
Figura 17: Ativação de PKC e formação de EROs na hiperglicemia.
4. Aumento do fluxo da via da hexosamina: Sob condições metabólicas usuais, 2-5%
da glicose que entra na célula é diretamente dentro da via da hexosamina, começando
com a conversão da frutose-6-fosfato para glicosamina-6-fosfato, pela enzima
glutamina:frutose-6-fosfato amidotransferase (JAMES, et al. 2002). Durante a
hiperglicemia, devido ao aumento dos nutrientes disponíveis, boa parte do excesso de
glicose é desviado para a via da hexosamina. O produto final deste caminho, UDP-N-
acetilglucosamina, é o substrato para a glicosilação de importantes fatores intracelulares
(MCCLAIN, CROOK, 1996), incluindo fatores de transcrição, assim afetando a
expressão de muitos genes e levando ao desenvolvimento de complicações
microvasculares do DM (GABRIELY et al., 2002; GOLDBERG et al., 2002).
Alguns autores postularam que esses mecanismos principais responsáveis pelos
danos oxidativos hiperglicêmicos, na verdade, estariam interligados e associados a um
aumento na formação de espécies reativas de oxigênio (estresse oxidativo), como
mostrou WAJCHENBERG (2002).
Os flavonóides constituem uma classe de compostos polifenólicos de ampla
distribuição no reino vegetal. Encontrados em plantas principalmente na forma de
glicosídeos, são os pigmentos amarelos, laranjas, azuis e vermelhos das flores,
responsáveis também pela cor amarela das folhas no outono. São importantes para o

crescimento normal, desenvolvimento e defesa das plantas. Atuam como atrativos
visuais favorecendo a polinização, como um mecanismo de defesa diante do ataque de
insetos e microorganismos e como protetores da radiação ultravioleta por suas
propriedades antioxidantes (MUSCHIETTI; MARTINO, 2007). Apresentam um núcleo
fundamental com 15 átomos de carbono, formado por dois anéis aromáticos ligados
entre si por uma cadeia de três átomos de carbono (C6-C3-C6) (Figura 18) (SIMÕES, et
al. 2001).
Figura 18: Estrutura de um flavonóide
Os antioxidantes vegetais são de natureza muito variada, mas, indubitavelmente,
os flavonóides constituem o grupo mais representativo encontrado com elevada
diversidade de formas (RICE-EVANS et al., 1995) com descrição de mais de mil
compostos distintos até a presente data (HARBONE, 1988). Existem outros encontrados
em plantas com propriedades antioxidantes já descritas na literatura pertinente, como
terpenóides, cumarinas, saponinas, curcuminas, ácido caféico, ácidos hidro cinânimicos,
entre outros (RICE-EVANS et al., 1995, 1996).
Há muitos métodos diferentes para determinar a capacidade antioxidante de uma
amostra. Estes métodos diferem quanto ao princípio do teste e condições experimentais.
A maioria deles é baseada na reação em que o radical livre é gerado e como esta reação
é inibida pela adição do composto ou amostra, que é objeto da medida da capacidade
antioxidante (ALONSO et al., 2002). Entre os métodos utilizados para determinar a
capacidade antioxidante de uma amostra estão: (1) Teor de fenólicos totais, (2)
Atividade seqüestradora do radical DPPH•; (3) Capacidade antioxidante equivalente ao
trolox (CAET).

Métodos utilizados:
5.5.1. Atividade seqüestradora do radical DPPH•:
É um método simples e rápido para estimar a capacidade antioxidante de
amostras utilizando o radical livre estável 1,1-difenil-2-picril-hidrazil (DPPH•) (Figura
19), pela a adição de compostos seqüestradores (HUANG et al., 2005).
Figura 19: Estrutura do 1,1-difenil-2-picril-hidrazil (DPPH)
A medida do consumo do radical DPPH• permite determinar exclusivamente a
habilidade intrínseca da substância doar átomos de hidrogênios ou elétrons destas
espécies reativas em um sistema homogêneo. O método é baseado na redução de uma
solução alcoólica de DPPH•, na presença de antioxidantes, devido à formação da forma
não-radicalar DPPH-H (Figura 20). A adição de um antioxidante resulta em uma
diminuição da absorbância proporcional à concentração e a atividade antioxidante do
composto. A coloração azul-violeta muda gradualmente para verde, podendo tornar-se
amarelo e a diminuição da absorbância, à 515nm, é monitorada durante a reação
(PAIXÃO et al., 2007).
Difenil-picril-hidrazina (não-radical) Difenil-picril-hidrazil (radical livre)
Fonte: (PAIXÃO et al., 2007). Figura 20: Estrutura do DPPH e sua redução pelo antioxidante

5.5.2.Teor de fenólicos totais
Compostos fenólicos ou constituintes polifenólicos constituem um dos grupos
mais numerosos e amplamente distribuídos de substâncias, com mais de 8000 estruturas
fenólicas conhecidas (HARBONE, 1980). Polifenóis são classificados em compostos
não-flavonóides e flavonóides. Flavonóides representam um grupo único de
fitoquímicos fenólicos, de ocorrência mais ampla (HARBORNE, 1964; HASLAM,
1998). Trabalhos recentes realçam a potencial propriedade benéfica dos flavonóides e
componentes polifenólicos para a saúde, conhecidos por serem poderosos doadores de
hidrogênio e sequestradores de espécies reativas de oxigênio e nitrogênio in vitro
(RICE-EVANS et al., 1996; KERRY, RICE-EVANS, 1999).
O ensaio de Fenólicos Totais tem sido usado durante muitos anos como uma
media de fenólicos totais nos produtos naturais (PRIOR et al., 2005). É um ensaio
comumente aceito e rotineiramente praticado nos laboratórios de pesquisas de
antioxidantes em todo o país, por ser um método conveniente, simples e reprodutível.
(HUANG et al., 2005).
O reagente de Folin-Ciocalteu, que contém ânions heteropoli-fosfotungstatos-
molibdatos, reage com compostos fenólicos em meio básico, e com a retirada de um
próton fenólico, há a formação de um ânion fenolato, que é capaz de reduzir o reagente,
em um complexo molibdênio-tugstênio de coloração azul, por um provável mecanismo
de transferência de elétrons (HUANG et al., 2005; PRIOR et al., 2005; ROGINSKY,
LISSI, 2005).
5.5.3. Capacidade antioxidante equivalente ao trolox (CAET).
O ensaio da Capacidade antioxidante equivalente ao trolox (CAET) ou teste do
ABTS (ácido 2, 2´-azinobis-[3-etilbenzotiazolina-6-sulfônico]) foi primeiro relatado por
Miller, Rice (MILLER et al., 1993), que é baseada na habilidade sequestradora de
antioxidantes para gerar o radical monocátion (ABTS•+) (PRIOR et al., 2005) (Figura
21). Este cátion é gerado pela oxidação do ABTS com persulfato de potássio e é
reduzido na presença de antioxidantes doadores de hidrogênio. A reação produz uma
coloração azul-esverdeada ou verde-azulada, de modo que a extensão da descoloração
indica a percentagem de inibição do cátion ABTS•+. Essa percentagem de inibição é
determinada como uma função da concentração e do tempo tomando a reatividade do
Trolox como padrão, sendo que ambos são determinados sob as mesmas condições.
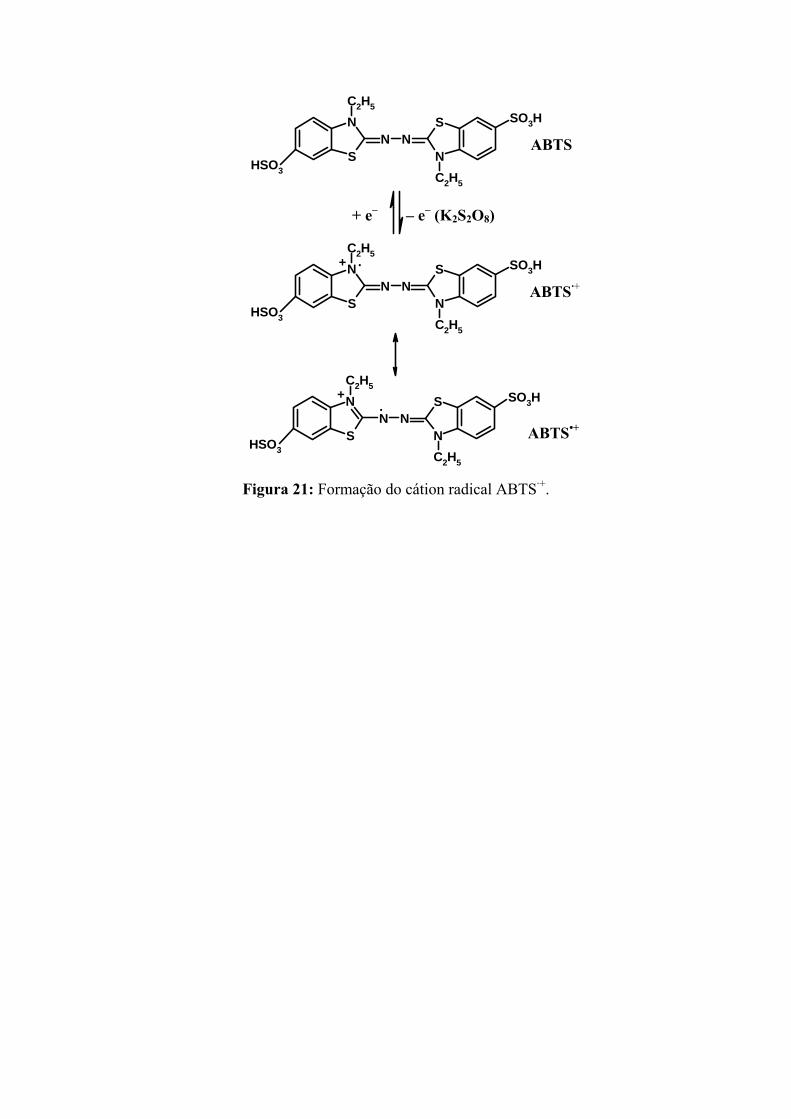
S
NS
NN N
SO3H
C2H5
C2H5
HSO3
S
NS
NN N
SO3H
C2H5
C2H5
HSO3
.
S
NN N
SO3H
C2H5
S
NC2H5
HSO3
.
+
+
ABTS
+ e– – e– (K2S2O8)
ABTS·+
ABTS•+
Figura 21: Formação do cátion radical ABTS·+.

OBJETIVO GERAL
Avaliar a atividade hipoglicemiante e antioxidante de Bauhinia cheilantha,
utilizando extratos e frações de diferentes épocas de coleta, contribuindo para o
estudo fitoquímico e farmacológico desta planta.
OBJETIVOS ESPECÍFICOS
Isolar e identificar os principais constituintes químicos com atividade
hipoglicemiante e antioxidante;
Avaliar e comparar a atividade hipoglicemiante e antioxidante do extrato e das
frações de Bauhinia cheilantha;
Realizar um estudo comparativo da atividade hipoglicemiante e antioxidante do
extrato e das frações de B. cheilantha.

METODOLOGIA
1. FITOQUÍMICA DA PLANTA Três (3) coletas de Bauhinia cheilantha (“Mororó sem espinho”-MSE) foram
feitas em épocas diferentes. A primeira coleta (1a) foi realizada em janeiro de 2005. A
2a coleta foi feita em agosto de 2006 e a 3a coleta foi feita em fevereiro de 2007, todas
as coletas foram feitas na região semi-árida, no município de Vieirópolis, Paraíba. A
espécie foi identificada pela botânica Profa. Dra. Maria de Fátima Agra e uma exsicata
encontra-se no Herbário do Laboratório de Tecnologia Farmacêutica, UFPB, sob o
número 6887.
As plantas obtidas em períodos diferentes foram submetidas ao mesmo processo
de obtenção do extrato etanólico bruto e particionamento, com solventes em ordem
crescente de polaridade (Fluxograma 1).
Inicialmente, as partes aéreas de Bauhinia cheilantha foram coletadas, secas em
estufa e pulverizadas. O pó seco foi extraído com etanol em temperatura ambiente. O
líquido extrator foi evaporado sob pressão reduzida a 40 0C, fornecendo o extrato
etanólico. Deste extrato uma alíquota foi separada para os ensaios biológicos e o
restante foi submetido a partição com solventes orgânicos.
O extrato etanólico foi suspenso com metanol:água (1:1) e extraído primeiro
com hexano (300,0 mL x 5 vezes) e em seguida com acetato de etila (300,0 mL x 5
vezes). Os solventes foram evaporados e obtendo-se o extrato hexânico e o extrato
acetato de etila. Após o processo de partição houve a formação de um precipitado
(PPT), onde uma parte mostrou-se solúvel em diclorometano (CH2Cl2) e a outra parte
em metanol (MeOH). Após evaporação do metanol foi obtido um extrato amarelo com
teste positivo para flavonóides. Este extrato amarelo foi cromatografado em coluna de
Sephadex LH-20 utilizando metanol como eluente. Desta coluna foram coletadas 11
frações e a fração de número 7 apresentou-se como um precipitado amarelo amorfo de
coloração amarelo laranja intenso com reagente NP (ácido difenilbórico etanolamina),
um reagente específico para flavonóides. Esta coloração amarelo laranja indica a
presença de hidroxilas em posição orto para os flavonóides.
A fração 7 (0,237 g), obtida do PPT solúvel em MeOH, foi solubilizada em
solução metanólica (90%) e injetada no sistema de Cromatografia Líquida de Alta
Eficiência (HPLC – High Perfomance Liquid Chromatography).

Fluxograma 1: Obtenção do extrato e frações de B. cheilantha
A caracterização estrutural dos constituintes químicos isolados de Bauhinia
cheilantha foi realizada através da análise dos espectros obtidos através dos métodos
espectroscópicos Infravermelho e Ressonância Magnética Nuclear (RMN) de
Hidrogênio e Carbono 13, utilizando técnicas uni e bidimensionais (COSY, HMQC,
HMBC) além de comparações com modelos da literatura.
Extrato Bruto
Planta seca
• Maceração (etanol 95%);
• Rotaevaporador rotativo.
• Estudo biológico • MeOH:H2O (1:1)
• AcOEt
• Hexano Ext. Bruto
Fr. Hexânica Fr. AcOEt Fr. Hidroalcóolica
PPT MeOH PPT CHCl2

Ressonância Magnética Nuclear (RMN)
Os espectros de Ressonância Magnética Nuclear, 1H e 13C e as técnicas
bidimensionais, foram registrados em espectrômetro da JOEL que opera a 400 MHz . O
solvente empregado foi DMSO.
Cromatografia Líquida de Alta Eficiência (HPLC)
Para a análise em Cromatografia Líquida de Alta Eficiência (HPLC) o sistemaa
consistiu de uma bomba de solvente modelo LC-10Avp, equipado com um detector
UV-Vis SPD 10AVvp (Shimadzu). As amostras foram injetadas em um injetor
Rheodyne 7125i com um “loop” de 200 µL. A separação comatográfica foi feita em
uma coluna C18 (25cm x 4.6mm, Shim-pack®), pré-coluna C18 (1 cm x 4 mm, Shim-
pack®). O solvente utilizado foi Metanol (MeOH), Tedia Company, grau HPLC. Para
filtração das amostras foram usados cartuchos com membrana millipore com poros de
0,45 µm de diâmetro.
2. ESTUDO FARMACOLÓGICO
2.1. Animais experimentais
Foram utilizados camundongos Swiss (Mus musculus) de ambos os sexos, com
peso variando entre 35-45 g, com aproximadamente 3 meses de idade. Os animais
foram provenientes do biotério Prof. Dr. Thomas George, LTF/UFPB, e mantidos em
gaiolas de polietileno, alimentados com ração comercial (tipo pellets), com livre acesso
à água. Sob condições de temperatura entre 21 ± 2 °C, com um ciclo de claro e escuro
de 12 horas, sendo a fase clara de 6:00 às 18:00 horas e a fase escura das 18:00 às 6:00
horas.
Este trabalho foi aprovado pelo Comitê de Ética em Pesquisa animal do
Laboratório de Tecnologia Farmacêutica (CEPA: 1201/05).
2.2 Teste de Toxicidade Aguda
Dois grupos de 10 camundongos de ambos os sexos, receberam doses de 1000 e
2000 g, via intra peritoneal (i.p.). Após a administração dos extratos os animais foram
observados durante 48 horas em intervalos 12 horas onde o registro das mortes, se

houvesse, era efetuado. Nas primeiras 4 horas do experimento, os animais foram
observados e o seu comportamento avaliado quanto a possíveis efeitos tóxicos das
substâncias.
2.3. Indução do diabetes em camundongos
O diabetes tipo 1 foi induzido através de injeção de 70 mg/Kg de peso de
aloxana na veia caudal dos camundongos. Os animais foram mantidos em jejum por,
pelo menos, 12 horas antes da indução do diabetes; e tiveram água disponível ad
libitum. A avaliação dos níveis glicêmicos foi verificada 48h após a indução com
aloxana. A divisão dos grupos aconteceu da seguinte maneira: (1) um grupo controle,
(2) um grupo aloxana, (3) um grupo padrão, que recebeu insulina regular e (4) grupos
experimentais. Durante o experimento para a verificação da atividade hipoglicemiante,
os grupos controles receberam solução diluente e grupos teste, receberam extratos e
frações por via oral (v.o.) (gavage) ou via intraperitoneal (i.p.), nas doses de 300, 600 e
900 mg/Kg. Apenas animais com glicemia acima de 200 mg/dL foram utilizados nos
experimentos.
As concentrações de glicose para todos os experimentos foram verificadas
através de um glicosímetro comercial (Accucheck Active - Roche). A glicemia foi
medida antes da administração do extrato/fração e após a administração da substância
teste, a glicemia foi verificada nos seguintes intervalos: 60, 120 e 180 minutos. A
porcentagem da variação da glicemia para cada grupo foi calculada em relação ao nível
glicêmico no tempo 0 (zero) e obtida de acordo com a seguinte equação: Variação da
glicemia (%) =Gx − G0/G0 x100 onde G0: a concentração de glicose sanguínea antes da
administração da amostra teste e Gx: a concentração de glicose sanguínea após a
administração da substância teste, nos seguintes valores glicêmicos: 0, 60, 120 e 180
minutos, respectivamente (SEPICI, 2004).

3. ESTUDO ANTIOXIDANTE
3.1. Atividade seqüestradora do radical DPPH•
A atividade seqüestradora de radical livre foi medida usando radical DPPH•, em
que a sua taxa de clareamento é monitorada por um comprimento de onda característico
(517 nm), na presença da amostra. Na forma de radical, o DPPH• absorve a 517nm, mas
sobre redução pelo antioxidante ou espécies radicalares sua absorção diminui.
Em um tubo de ensaio com 5 mL de uma solução de DPPH• (23,6 µg/mL em
EtOH) foi misturado quantidades apropriadas (provenientes de uma solução estoque de
1 mg/mL e obtidas através de uma triagem preliminar), acompanhadas pela
homogeneização. Após 30 min de agitação em aparelho de ultra-som, a quantificação do
radical DPPH• foi realizada em espectofotômetro UV-visível a 517nm. As análises
foram realizadas em triplicata (n = 3). A percentagem da atividade seqüestradora (%
AS) foi calculada pela equação:
% AS = 100x(Acontrole - Aamostra)/Acontrole
onde Acontrole é a absorbância do controle, uma solução que contém apenas o radical
DPPH· e EtOH, e Aamostra é a absorbância do radical na presença dos extratos ou do
padrão ácido ascórbico (Figura 22).
As medidas foram calculadas e os resultados expressos como média ± E.P.M. A
eficiência antiradicalar foi estabelecida usando análise de regressão linear no intervalo
de confiança de 95% (p<0,05) obtido pelo programa de estatística GraphPad Prism 4.0.
Os resultados estão representados pelo valor da CE50, que representa a concentração da
amostra necessária para seqüestrar 50% dos radicais DPPH•. Os extratos e substâncias
são considerados ativos quando apresentam CE50 <500 µg/mL (CAMPOS et al.,
2003).Como padrão de referência foi utilizado o ácido ascórbico.
O
OH OH
OHOH
Figura 22: Estrutura do ácido ascórbico

3.2. Teor de fenólicos totais A determinação do teor de fenólicos totais presentes nos extratos EtOH bruto e
as frações da B. cheilantha foi realizada utilizando o Reagente de Folin Ciocalteau, com
o ácido gálico como o composto fenólico padrão.
Uma alíquota de 500 µL das frações hexânica, aquosa e do precipitado solúvel
em diclorometano (PPT dicloro) e 100 µL da fração AcOET, precipitado solúvel em
MeOH e do extrato etanólico bruto, todas provenientes de uma solução estoque de
1mg/mL, solubilizados em EtOH foi transferida para um balão volumétrico de 5 mL,
adicionando-se 100 µL do reagente de Folin-Ciocalteu e 3 mL de água destilada,
agitando-se por 1 min. Em seguida, 300 µL de Na2CO3 (15%) foram acrescentados à
mistura e agitados por 30 segundos. Finalmente a solução teve seu volume aferido para
5 mL com água destilada. Após duas horas, a absorbância das amostras foi medida a
760 nm utilizando-se cubetas de vidro. A concentração dos compostos fenólicos totais
foi determinada como micrograma de equivalente de ácido gálico por grama de amostra
(mg EAG/g), a partir da curva de calibração construída com padrões de ácido gálico
(0,5 a 25 µg/mL) considerando-se o Erro Padrão da Média (E.P.M.). A equação da
curva de calibração do ácido gálico foi: A=0, 0951C + 0, 1319, com o coeficiente de
correlação de r2 = 0, 997, onde C é a concentração do ácido gálico e A é a absorbância a
760 nm.
4.3. Capacidade antioxidante equivalente ao trolox (CAET).
A determinação da CAET foi determinada de acordo com RE et al. (1999),
usando o Trolox (ácido 6-hidróxi-2,5,7,8-tetrametilcroman-2-carboxílico) como
composto padrão.
O cátion ABTS•+ foi preparado pela reação de 7mM de ABTS com 2,45mM de
persulfato de potássio. A mistura foi deixada no escuro, a temperatura ambiente, por 12-
16h, antes do uso. Para a avaliação da atividade a solução ABTS•+ foi diluída em etanol
(1:90 v/v, aproximadamente) até obter um absorbância de 0,7 ± 0,02, no comprimento
de onda de 734nm. Quantidades apropriadas das amostras (provenientes de uma solução
estoque de 1 mg/mL e obtidas através de uma triagem preliminar) ou do padrão Trolox
(concentrações finais de 6 a 18 µM) foram misturadas com 5 mL da solução de ABTS•+.
As soluções foram agitadas e, após 10 minutos, a absorbância das amostras e do padrão,
utilizando-se cubetas de vidro. As análises foram realizadas em triplicata e a
percentagem de inibição (% I) em relação ao branco foi calculada pela equação:

% I = 100x(Abranco - Aamostra)/Abranco
A eficiência da inibição do ABTS·+ foi estabelecida utilizando a análise de
regressão linear no intervalo de confiança de 95 % (p<0,05) obtido pelo programa de
estatística GraphPad Prism 4.0. Os resultados foram expressos através da CE50 ±
E.P.M, que representa a concentração da amostra necessária para obter metade da
inibição máxima do cátion radical ABTS·+.
3. ANÁLISE ESTATÍSTICA
Os resultados foram expressos com Média ± Erro Padrão da Média (E.P.M.) e
analisados estatisticamente empregando-se a Análise de Variância (ANOVA) one-way e
two-way, seguido do teste de Bonferroni, onde os valores de p < 0,05 foram
considerados significantes.
A relação entre os testes antioxidantes foi estabelecida utilizando o programa
Excel 2003.
Todos os dados foram analisados pelo Programa GraphPad Prism© versão 4.0
(GraphPad Software Inc., San Diego CA, EUA).

RESULTADOS E DISCUSSÃO
1. Identificação estrutural
A fração 7 foi injetada no sistema de Cromatografia Líquida de Alta Eficiência
(HPLC), obtendo-se três picos (Figura 23): o primeiro, obtido em quantidade muito
pequena, não foi utilizado para análises posteriores, o segundo e o terceiro pico foram
coletados, evaporados e submetidos a análise em aparelho de Ressonância Magnética
Nuclear de Hidrogênio e Carbono 13, sendo identificado dois flavonóides.
Cromatograma
-1,00E-03
4,90E-02
9,90E-02
1,49E-01
1,99E-01
2,49E-01
2,99E-01
0 1 2 3 4 5 6 7 8 9 1
Tempo (min)
Abs
orbâ
ncia
(AU
)
Substância 3
Substância 2
Cromatograma (260nm) Cromatograma (320nm)
Substância 1
0
Figura 23: Cromatograma do PPT solúvel em MeOH
O espectro de RMN de 1H (Figura 23) da substância 2 indicou a presença de um
flavonóide glicosilado. Dois sinais em δ 6,17 (sl) e 6,37 (sl) foram atribuídos ao anel A
do flavonóide para os hidrogênios H-6 e H-8, respectivamente. Dois dubletos em δ 7,73
(J=8,0 Hz) e 6,90 (J=8,0 Hz) correspondem ao anel B p-hidroxilado. Dois singletos
largos em δ 0,77 para um grupo metila e outro em δ 5,28, em adição aos sinais entre δ
3-4 sugeriu a presença de uma ramnose ligada na aglicona. O espectro de RMN de 13C
(Figura 25) mostrou a presença dos sinais em δ 70,17; 70,39; 70,70 e 71,18 para
carbonos hidroximetínicos, um sinal em δ 17,5 para um carbono metílico e em 101,6
para um carbono anomérico confirmou a presença da ramnose ligada na aglicona. Este
espectro também mostrou os sinais em δ 177,61; 156,70 e 134,16 sugerindo a presença

de uma cetona α-β-insaturada de um flavonol. Correlações observadas nos espectros de
COSY (Figura 26), HMQC (Figura 27) e HMBC (Figura 28) permitiu o completo
assinalamento dos sinais de 1H e 13C do flavonóide 2. Os valores dos deslocamentos
químicos do C-3 (3 ppm) para campo mais alto e do C-2 (10 ppm) para campo mais
baixo (MARKHAM et al., 1978) sugeriu que a unidade do açúcar estivesse ligada no C-
3. Os sinais restantes confirmaram os padrões de oxigenação nos anéis A e B. Os
valores dos deslocamentos químicos de cada açúcar foram atribuídos pelo COSY
(Figura 26). Por exemplo, começando pela metila 3H-6” (δ 0,77) foi possível verificar o
acoplamento com H-5” (m; δ 3,10) e assim por diante. O espectro de HMBC (Figura
28) mostrou a correlação do hidrogênio anomérico H-1” (δ 5,28) com o C-3 (134,16)
que confirmou a posição da ramnose em C-3. Com base nestas evidências foi possível
identificar o flavonóide glicosilado como sendo 3-O-α-L-ramnopiranosil kanferol
(afzelina). Estes dados estão de acordo com Ribeiro et al. (2002). Os dados de RMN
estão na tabela 12. A afzelina foi isolada anteriormente de outras espécies de Bauhinia,
como: Bauhinia microstachya (Raddi) Macbr (MEYRE-SILVA, 2001), Bauhinia
racemosa (EL-HOSSARY et al., 2000) e Bauhinia uruguayensis (IRIBARREN,
POMILIO, 1989).

Tabela 12: Dados de RMN de 1H (400 MHz) e RMN de 13C (100 MHz) do flavonóide 1 (afzelina) em DMSO
O
O
O
OH
OH
OH
O
OHOH
OH
CH3
2
34
5
6
78 1'
2'
3'
4'
5'
6'
1"
2"3"
4"
5" 6"
1
1H-13C-HMQC-1JCH
1H-13C-HMBC-nJCHC δC δH (Mult, 2JCH
3JCH2 156,70 3 134,16 H-1” 4 177,61 5 161,32 H-6 7 165,67 H-6, H-8 9 157,11 H-8 10 103,73 H-6 1’ 120,59 H-3’, H-5’ 4’ 160,15 H-2’, H-6’ CH 6 99,16 6,17 (sl) H-8 8 94,02 6,37 (sl) H-6 2’,6’ 130,66 7,73 (d; 8,0) H-6’/H-2’ 3’,5’ 115,51 6,90 (d; 8,0) Ramnose 1” 101,82 5,28 (sl) 2” 70,17 3,97 (sl) 3” 70,39 3,46 (d; 8,0) H-4’ 4” 71,18 3,08 (m) H-3’, H-5’ 5” 70,70 3,10 (m) H-3” 6” 17,57 0,77 (sl) H-5”

Figu
ra 2
3: E
spec
tro d
e R
MN
1 H (4
00 M
Hz)
em
DM
SO d
a su
bstâ
ncia
2.

Figu
ra 2
4: E
spec
tro d
e R
MN
13C
(100
MH
z) e
m D
MSO
, da
subs
tânc
ia 2
.

Figura 25: Espectro de RMN 1H x1H- COSY (400 MHz) em DMSO da substância 2
Figura 26: Espectro de RMN de HMQC em DMSO, da substância 2

Figura 27: Espectro de RMN de HMBC em DMSOda substância 2

O espectro de RMN de 1H (Figura 28) da substância 3 mostrou sinais em δ 6,86
(d; 5,2 Hz), δ 7,29 (sl) e 7,24 (d; 8,0 Hz) que foram atribuídos aos hidrogênios H-5’, H-
2’ e H-6’, respectivamente, sugerindo a presença de oxigenação nas posições 3 e 4 do
anel B na estrutura do flavonóide. O espectro de RMN de 13C (Figura 29) mostrou os
sinais em δ 148,83 e 145,47 referentes aos carbonos quaternários C-3’ e C-4’,
respectivamente e os sinais em δ 115,72; 115,61 e 121,11 assinalados para os carbonos
metínicos aromáticos em C-2’, C-5’ e C-6’, respectivamente, confirmando o padrão de
oxigenação no anel B. Os valores dos sinais restantes da molécula tanto de 1H como de 13C são similares aos deslocamentos químicos do flavonóide 2, incluindo a correlação
no HMBC do C-3 (δ 134,14) e o hidrogênio anomérico H-1” (δ 5,25) que confirmou a
posição da ramnose em C-3 e permitiu identificar o flavonóide como 3-O-α-L-
ramnopiranosil quercetina (quercitrina). Estes dados estão de acordo com a literatura
(RIBEIRO et al., 2002).
Flavonóides são característicos da família Leguminosae e possuem grande
ocorrência no gênero Bauhinia. Quercitrina tem sido isolada de muitas espécies,
incluindo Bauhinia purpurea (RAMACHANDRAN AND JOSHI, 1967), Bauhinia
reticulata (RABATE, 1938), Bauhinia splendens (CECHINEL FILHO ET AL., 1995),
Bauhinia tomentosa (SUBRAMANIAN AND NAIR, 1963), Bauhinia uruguayensis
(IRIBARREN AND POMILIO, 1989) and Bauhinia vahlii (SULTANA ET AL., 1985).

Tabela 13: Dados de RMN de 1H (400 MHz) e RMN de 13C (100 MHz) do flavonóide
2 (quercitrina) em DMSO
O
O
O
OH
OH
OH
O
OHOH
OH
CH3
OH
2
34
5
6
78 1'
2'
3'
4'
5'
6'
1"
2"3"
4"
5" 6"
2
1H-13C-HMQC-1JCH1H-13C-HMBC-nJCH
C δC δH (Mult, MHz) 2JCH3JCH
2 156,93 3 134,14 H-1” 4 177,64 5 161,29 H-6 7 165,83 H-6, H-8 9 157,11 H-8 10 103,60 H-6, H-8 1’ 120,69 H-5’ 3’ 148,83 H-2’ 4’ 145,47 H-5’ H-2’ CH 6 99,16 6,17 (sl) H-8 8 93,91 6,36 (sl) H-6 2’ 115,72 7,29 (sl) H-6’ 5’ 115,61 6,86 (d; 5,2) 6’ 121,11 7,24 (d; 8,0) H2’ Ramnose 1” 101,87 5,27 (sl) 2” 70,16 3,93 (sl) 3” 70,41 3,51 (sl) 4” 71,28 3,14 (sl) H-3’, H-5’ 5” 70,68 3,22 (d; 3,1) 3H-6 6” 17,61 0,81 (sl)

Figu
ra 2
8: E
spec
tro d
e R
MN
1H
(400
MH
z) e
m D
MSO
, da
subs
tânc
ia 3

Figu
ra 2
9: E
spec
tro d
e R
MN
13C
(100
MH
z) e
m D
MSO
, da
subs
tânc
ia 3

2. Toxicidade aguda Os resultados da toxicidade aguda dos extratos de Bauhinia cheilantha (BONG)
STEUD (designada MSE de acordo com seu nome popular mororó sem espinho)
obtidos foram realizados nas concentrações de 1000 e 2000 mg/Kg i.p. e 5000 mg/Kg
v.o. em camundongos de ambos o sexos, não havendo mortes após 48 horas.Uma vez
que as espécies não apresentaram toxicidade nas concentrações citadas, não foi possível
estabelecer uma concentração inicial para testar os extratos nos animais. Baseado em
artigos sobre a atividade hipoglicemiante de outras espécies de Bauhinia, as
concentrações de 300, 600 e 900 mg/Kg de peso foram escolhidas e estabelecidas como
as concentrações efetivas para a realização dos testes (ANEXO).
3. Atividade hipoglicemiante
O extrato etanólico bruto de Bauhinia cheilantha (MSE), da 1a coleta, reduziu a
glicemia dos animais na dose de 900 mg/Kg, em média 46% (gráficos 1 e 2). A redução
da glicemia foi acentuada desde a primeira medição alcançando os menores valores
glicêmicos aos 180 minutos após a administração de MSE.
0 60 120 1800
100
200
300
400
500
600
Aloxana 300 mg/kg 600 mg/kg 900 mg/kg
ControleInsulina + Aloxana
* * *
***
Tempo (min)
Glic
emia
mg/
dL
Gráfico 1: Efeito do extrato etanólico bruto das folhas de MSE (1ª coleta) em
camundongos diabéticos induzidos com aloxana. Valores são média ± E.P.M. (n=6).
ANOVA Two-Way seguido do Pós-Testes de Bonferroni, *P<0,01.

0
25
50
75ControleMSE 300MSE 600MSE 900
*%
de
redu
ção
da g
licem
ia
Gráfico 2: Efeito do extrato etanólico bruto das folhas de MSE (1ª coleta), nas
concentrações de 300, 600 e 900 mg/Kg, comparados com o grupo controle com relação
à porcentagem de redução da glicemia em camundongos. Valores são média ± E.P.M.
(n=6). * P> 0,01. ANOVA one-way.
A quantidade obtida na 1a coleta foi insuficiente para testar a atividade
hipoglicemiante das frações resultantes do extrato bruto e repetir os experimentos,
portanto uma segunda coleta foi necessária. Dessa forma não foi possível comparar as
frações das coletas subsequentes com a 1a coleta.
O extrato bruto testado da 2ª coleta, não apresentou resultados expressivos,
quanto a sua atividade hipoglicemiante (Gráfico 3). Entretanto, as frações resultantes do
particionamento do extrato de MSE foram testadas, porque apresentaram resultados
promissores com relação à sua atividade antioxidante, que será demonstrada a seguir.
No entanto, nenhuma delas apresentou atividade hipoglicemiante significativa; onde
está representado no Gráfico 4, o resultado obtido com a Fração Acetato de Etila
(AcOEt).

0 60 120 1800
100
200
300
400
500
600
Aloxana300 mg/kg600 mg/kg900 mg/kg
ControleInsulina + Aloxana
* * *
Tempo (min)
glic
emia
(mg/
dl)
Gráfico 3: Efeito do extrato etanólico bruto das folhas de MSE (2ª coleta) em
camundongos diabéticos induzidos com aloxana. Valores são média ± E.P.M. (n=6).
ANOVA Two-Way seguido do Pós-Testes de Bonferroni.
0 60 120 1800
100
200
300
400
500
600
Aloxana300 mg/kg600 mg/kg900 mg/kg
ControleInsulina +Aloxana
* * *
Tempo (min)
glic
emia
(mg/
dl)
Gráfico 4: Efeito da fração AcOEt das folhas de MSE (2ª coleta) em camundongos
diabéticos induzidos com aloxana. Valores são média ± E.P.M. (n=6). ANOVA Two-
Way seguido do Pós-Testes de Bonferroni.

Todos os gráficos a seguir são referentes às amostras obtidas da 3a coleta. Nos
gráficos 5 e 6 são mostrados o efeito nas doses de 300, 600 e 900 mg/Kg nos diferentes
grupos, tratados com extrato etanólico bruto de MSE. A dose de 300 mg/Kg foi capaz
de diminuir os níveis glicêmicos dos animais em todos os tempos considerados e as
doses de 600 e 900 mg/Kg diminuíram a glicemia de forma expressiva apenas no tempo
de 180’. Estes resultados estão de acordo com aqueles encontrados por ALMEIDA et
al., (2006). Após um tratamento agudo com o extrato metanólico de Bauhinia
cheilantha em ratos, ALMEIDA et al. verificaram uma significativa diminuição dos
níveis de glicose plasmática nos animais.
0 60 120 1800
100
200
300
400
500
600
Aloxana300 mg/kg600 mg/kg900 mg/kg
ControleInsulina + Aloxana
*
* * *
* **
Tempo (min)
glic
emia
(mg/
dl)
Gráfico 5: Efeito do extrato etanólico bruto das folhas de MSE (3ª coleta), em
camundongos diabéticos induzidos com aloxana. Valores são Média ± E.P.M. (n=6).
ANOVA Two-Way seguido do Pós-Testes de Bonferroni.

-6
-4
-2
0
2
4
6Controle300 mg/kg600 mg/kg900 mg/kg
% re
duçã
o da
glic
emia
Gráfico 6: Efeito do extrato etanólico bruto das folhas de MSE (3ª coleta), nas
concentrações de 300, 600 e 900 mg/Kg, comparados com o grupo controle com relação
à porcentagem de redução da glicemia em camundongos. Valores são média ± E.P.M.
(n=6). ANOVA one-way.
A fração hexânica, não mostrou atividade hipoglicemiante. Essas diferenças
quanto à atividade hipoglicemiante pode ser ocasionada por diversos fatores, como:
época da coleta e condições ambientais dos animais.
As doses de 600mg/kg e 900 mg/kg da fração Acetato de etila (AcOEt) não
apresentaram um efeito hipoglicemiante significativo, quando comparado com o
controle.
A fração aquosa, administrada oralmente, das folhas de MSE apresentou uma
discreta redução dos níveis glicêmicos, quando comparados com o controle, na dose de
300 mg/Kg (Gráfico 7). A dose de 300 mg/kg demonstrou uma maior atividade
hipoglicemiante média (Gráfico 8).

0 60 120 1800
100
200
300
400
500
600
Aloxana300 mg/kg600 mg/kg900 mg/kg
ControleInsulina + Aloxana
*
** *
Tempo (min)
glic
emia
(mg/
dl)
Gráfico 7: Efeito da fração aquosa das folhas de B. cheilantha (MSE) (3ª coleta), em
camundongos diabéticos induzidos com aloxana. Valores são média ± E.P.M. (n=6).
*P<0,05. ANOVA Two-Way seguido do Pós-Testes de Bonferroni.
-10
-5
0
5
10
15 *Controle300 mg/kg600 mg/kg900 mg/kg
% re
duçã
o da
glic
emia
Gráfico 8: Efeito da fração aquosa das folhas de MSE (3ª coleta), nas concentrações de
300, 600 e 900 mg/Kg, comparados com o grupo controle, com relação à porcentagem
de redução da glicemia em camundongos. Valores são média ± E.P.M. (n=6). *P<0,05.
ANOVA one way.

O efeito hipoglicêmico causado pelos extratos/frações pode estar relacionado;
pela ação da amostra em tecidos alvos da insulina, como, por exemplo, músculo e tecido
adiposo (estimulando a captação de glicose), pela inibição da absorção de glicose pelo
intestino ou ainda pela inibição da reabsorção da glicose pelos rins.
Além dos extratos/frações, foram testados o precipitado solúvel em MeOH e
solúvel em Diclorometano (Gráfico 9 e 10, respectivamente), ambos os precipitados não
apresentaram efeito hipoglicemiante.
0 60 120 1800
100
200
300
400
500
600
Aloxana300 mg/kg600 mg/kg900 mg/kg
ControleInsulina + Aloxana
* * *
Tempo (min)
glic
emia
(mg/
dl)
Gráfico 9: Efeito do PPT MeOH das folhas de MSE (3ª coleta), em camundongos
diabéticos induzidos com aloxana. Valores são média ± E.P.M. (n=6).
0 60 120 1800
100
200
300
400
500
600
Aloxana300 mg/kg600 mg/kg900 mg/kg
ControleInsulina + Aloxana
* * *
Tempo (min)
glic
emia
(mg/
dl)
Gráfico 10: Efeito do PPT diclorometano das folhas de MSE (3ª coleta), em
camundongos diabéticos induzidos com aloxana. Valores são média ± E.P.M. (n=6).

4. Testes antioxidantes
O teste de DPPH foi realizado em todas as coletas.O conteúdo de fenólicos totais, e o
teste do ABTS foram feitos apenas nas duas últimas coletas (2ª e 3a coletas).
4.1. Atividade seqüestradora do radical DPPH•
A capacidade de seqüestrar radicais livres foi avaliada no extrato bruto e nas
frações hexânica, acetato de etila e aquosa, além dos precipitados solúveis em MeOH e
em diclorometano das duas últimas coletas de MSE sendo os resultados expressos como
valores de CE50. Valores de CE50 e PI (percentagem de inibição) caracterizam a
capacidade antioxidante de compostos puros, mas para extratos estes parâmetros podem
também serem usados para indicar que extratos são mais adequados como fontes de
compostos antioxidantes puros e podem ser usados como guia para futura purificação e
isolamento (ARGOLO et al., 2004).
De acordo com os resultados obtidos, o extrato e frações desta espécie
apresentam uma boa atividade anti-radicalar, considerando que os extratos e substâncias
são considerados ativos quando apresentam CE50 <500 µg/mL (CAMPOS et al., 2003).
Na 1ª coleta, foi feito apenas o teste de DPPH do extrato bruto. Comparando os
valores dos extratos observa-se que o extrato bruto da 1ª coleta apresentou o menor
valor da CE50, portanto melhor atividade antioxidante (Gráfico 11).
0 10 20 30
Ext. EtOH 1.coleta
Ext. EtOH 2.coleta
Ext. EtOH 3.coleta
26,683 ± 0,228
11,672 ± 1,43
23,457 ± 0,034
CE50 (µg/mL)
Gráfico 11: Atividade seqüestradora de radical livre dos extratos/amostras de MSE,
usando o radical DPPH•, da 1a coleta.

Na 2a coleta da planta (agosto de 2006), os valores variaram de 9,6 ± 0,468 a
38,986 ± 0,139 µg/mL. A fração AcOEt apresentou uma grande atividade seqüestradora
de radical DPPH• (CE50 = 9,32 ± 0,081 µg/mL), acompanhada pela fração aquosa
(CE50= 9,6 ± 0,468 µg/mL), ext. EtOH (CE50 = 26,683 ± 0,228 µg/mL) e Fr. hexânica
(CE50 = 38,986 ± 0,139 µg/mL), como mostrado na Gráfico 12.
0 10 20 30 40
Ext. EtOH
Fr. Hex.
Fr. AcOEt
Fr. Aquosa
PPT dicloro
Ác. asc. 2,48 ± 0,003
28,41 ± 0,413
9,6 ± 0,468
9,32 ± 0,081
38,98 ± 0,139
26,683 ± 0,228
CE50 (µg/ml)
ras
t
Amos
Gráfico 12: Atividade seqüestradora de radical livre dos extratos/amostras de MSE,
usando o radical DPPH•, da 2a coleta.
Em relação a 3a coleta (fevereiro de 2007), os valores da CE50 apresentaram um
intervalo de variação de 18,079 ± 0,255 a 30,753 ± 0,153 µg/mL. Da mesma forma
como ocorreu na 2a coleta, a fração AcOEt apresentou a maior atividade anti-radicalar
(CE50 = 18,079 ± 0,255 µg/mL). No entanto, diferente da coleta anterior, a fração
AcOEt foi seguida pela fração hexânica (CE50 = 20,405 ± 0,15 µg/mL), extrato bruto
(CE50= 21,77 ± 0,126 µg/mL) e fração aquosa (CE50 = 30,753 ± 0,153 µg/mL), como
demonstrado no Gráfico 13.

0 10 20 30 40
Ext. EtOH
Fr. Hex.
Fr. AcOEt
Fr. Aquoso
PPT MeOH
Ácido ascórbico 2,48 ± 0,003
9,35 ± 0,09
30,753 ± 0,153
14,78 ± 0,391
20,405 ± 0,15
23,457 ± 0,034
CE50 (µg/mL)
Am
ostr
as
Gráfico 13: Atividade seqüestradora de radical livre dos extratos/amostras de MSE,
usando o radical DPPH•, referente a 3a coleta.
Estes resultados estão de acordo com aqueles publicados por ADEROGBA et
al., 2007, onde comprovam que o extrato bruto e as frações, obtidos com solventes mais
polares (AcOEt e butanol), mostram considerável capacidade antioxidante. No entanto,
discordam de ARGOLO et al. (2004) que afirmam que a CE50 da fração hexano é muito
alta (17 mg antioxidante/g DPPH) comparado com os outros extratos/frações, que
possuem o valor da CE50 na taxa de 2,06-2,32 mg antioxidante/g DPPH. Afirmam ainda
que o valor da CE50 obtida para a fração hexânica foi significativamente diferente dos
valores obtidos para os extratos etanólicos, clorofórmio, acetato de etila e aquoso que
mostraram resultado similar. Portanto, a fração hexânica é uma fonte pobre de
antioxidante.
Comparando o resultado da CE50 do extrato etanólico bruto das três coletas
realizadas, observa-se que o extrato da 1a coleta apresentou o maior capacidade
antioxidante (CE50 = 11,672 ± 1,43 µg/mL), quando comparado com os extratos das
outras duas coletas, que apresentaram valores de CE50 semelhantes (CE50 = 26,683 ±
0,228 µg/mL e CE50 = 21,77 ± 0,126 µg/mL).
A diferença observada entre os valores da CE50 das amostras, pode ser atribuída
ao fato de que plantas possuem uma variabilidade sazonal com relação aos seus
constituintes, sendo natural que apresentem quantidades de metabólicos diferentes,
considerando que foram coletadas em épocas diferentes do ano.

Os precipitados solúvel em MeOH (PPT MeOH) e solúvel em diclorometano
(PPT CH2Cl2) foram obtidos em pouca quantidade nas duas coletas realizadas, não
sendo possível realizar os testes antioxidantes com amostras das duas coletas,
impossobilitando a comparação entre as duas amostras. O PPT MeOH mostrou forte
atividade seqüestradora de radical livre (CE50= 9,35 ± 0,09 µg/mL).
O gráfico 14 mostra a curva de regressão linear entre o teor de fenólicos totais
em função dos valores de 1/CE50 da atividade seqüestradora do radical DPPH das
amostras obtidas na 3a coleta. O coeficiente de correlação linear (R2) obtido de todas as
amostras analisadas foi 0,609. As amostras obtidas da 2a coleta não apresentaram
coeficiente de correlação linear (R2) significante (dados não mostrados).
DASGUPTA (2007) não encontraram correlação entre atividade antioxidante e
conteúdo de fenólicos totais/flavonóides, como determinado pelo coeficiente de
regressão linear (R2). Algumas plantas têm alto conteúdo de fenóis/flavonóides, mas
baixa atividade antioxidante.
Gráfico 14: Correlação linear do teor de Fenólicos Totais (FT) em função de 1/CE50 da
atividade seqüestradora do radical DPPH , dos extratos/frações de MSE da 3a coleta.
4.2. Total de fenólicos Totais (FT)
Os valores de FT dos diferentes extratos/frações das duas coletas de Bauhinia
cheilantha estão expressos nos gráficos 15 e 16. Os valores referentes à 2a coleta estão
compreendidos no intervalo de 78,67 a 341,55 mg EAG/g da amostra, sendo o AcOEt
aquele que possui a maior quantidade de compostos fenólicos. Quanto a análise feita

nos extratos/frações da 3a coleta, os valores de fenólicos totais estão entre 37,5 a 180,79
mg EAG/g da amostra, dentre os extratos/frações analisados, o AcOEt foi o que
apresentou a maior quantidade de compostos fenólicos, semelhante ao que ocorreu com
a 2a coleta, o que pode estar relacionado a uma grande concentração de flavonóides e
compostos polifenóis, o que, segundo LUO, BASILE; KENNELLY, 2002, contribuem
significativamente para a atividade antioxidante total de muitas frutas e vegetais.
0 50 100 150 200 250 300 350
Ext. EtOH
Fr. Hex.
Fr. AcOEt
Fr. Aquosa
PPT dicloro 68,56 ± 0,67
78,67 ± 5,17
341,55 ± 1,41
132,39 ± 0,98
87,74 ± 4,96
Fenólicos Totais (mg EAG/g da amostra)
Amos
tras
Gráfico 15: Teor de fenólicos totais dos extratos/frações de MSE referente à 2a coleta
0 100 200 300 400 500 600
Ext. EtOH
Ext. Hex
Ext. AcOEt
Ext. Aquoso
PPT MeOH 540,27 ± 22,42
63,9 ± 13,59
180,79 ± 4,8
37,5 ± 1,15
84,96 ± 3,03
Fenólicos Totais (mg EAG/g da amostra)
Am
ostr
as
Gráfico 16: Teor de fenólicos totais dos extratos/frações de MSE referente à 3a coleta

No entanto, apesar da fração AcOEt apresentar a maior quantidade de FT nas
duas coletas da planta, a ordem dos valores de fenólicos totais dos outros
extratos/frações estudados é diferente para as referidas coletas. Na 2º coleta, a ordem
foi: Fr. AcOEt (341,55 mg EAG/g) > Fr. Hex (132,39 mg EAG/g) > Fr. EtOH (87,74
mg EAG/g) > Fr. Aquoso (78,67 mg EAG/g). Na 3º coleta, as amostras apresentaram-se
na seguinte ordem: Fr. AcOEt (180,79 mg EAG/g) > ext. EtOH (84,96 mg EAG/g) > Fr.
aquoso (63,9 mg EAG/g) > Fr. hexano (37,5 mg EAG/g).
A diferença observada entre os valores dos fenólicos totais das amostras,
oriundas de épocas diferentes do ano, pode ser atribuída à complexidade química dos
produtos vegetais, bem como a variações diversas como a circadiana, dos locais de
origem, solo, clima e época do ano, que uma planta está sujeita (MODESTO FILHO,
1989).
O PPT MeOH apresentou grande quantidade de FT (540,27 mg EAG/g),
podendo esse resultado ser atribuído a presença de grande quantidade de compostos
fenólicos. O PPT dicloro apresentou uma quantidade bem menor de FT (68,56 mg
EAG/g).
4.3. Capacidade equivalente ao Trolox
A análise de CAET foi realizada nos extratos/frações de MSE das 2 (duas)
coletas realizadas (2a e 3a coletas). Na 2a coleta, como representado no gráfico 17, a
fração AcOEt (2,9 ± 0,03 µg/mL ) apresentou uma CE50 semelhante ao composto
padrão Trolox (2,03 ± 0,05 µg/mL). Os outros extratos/frações apresentaram CE50 na
seguinte ordem: Fr. hexânica (7,16 ± 0,04 µg/mL), ext EtOH (15,18 ± 0,14 µg/mL), Fr.
aquosa (16,02 ± 0,05 µg/mL).

0 10 20 30 40
Ext. EtOH
Fr. Hex.
Fr. AcOEt
Fr. Aquosa
PPT dicloro
Trolox 2,03 ± 0,03
21,12 ± 0,132
16,02 ± 0,05
2,9 ± 0,05
15,183 ± 0,14
7,16 ± 0,04
CE50 (µg/mL)
Am
ostr
as
Gráfico 17: Atividade antioxidante equivalente ao Trolox (CAET) dos
extratos/amostras de MSE referentes à 2a coleta
Na 3a coleta, CE50 expressas no gráfico 18, a fração AcOEt assim como ocorreu
na 2a coleta, também apresentou o menor valor da CE50 (10,99 ± 0,09 µg/mL),
indicando que esse extrato possui uma forte capacidade em inibir a formação do cátion
radical ABTS•+, acompanhada pelo ext. EtOH (18,87 ± 0,32 µg/mL), fração aquosa
(31,89 ± 0,62 µg/mL), fração hexânica (36,33 ± 0,38 µg/mL). Esses dados são coerentes
com o fato de que os compostos mais polares possuem maior capacidade seqüestradora
de radicais livres.
0 10 20 30 40
Ext. EtOH
Fr. Hex
Fr. AcOEt
Fr. Aquosa
PPT MeOH
Trolox 2,03 ± 0,03
5,24 ± 0,13
31,89 ± 0,622
10,99 ± 0,09
36,33 ± 0,38
18,87 ± 0,32
CE50 (µg/mL)
Am
ostr
as
Gráfico 18: Atividade antioxidante equivalente ao Trolox (CAET) dos
extratos/amostras de MSE referentes à 3a coleta.

O gráfico 19 mostra a curva de regressão linear entre o teor de fenólicos totais
em função dos valores de 1/CE50 da atividade inibidora do cátion radical ABTS•+ das
amostras obtidas na 3a coleta. O coeficiente de correlação linear (R2) obtido de todas as
amostras analisadas foi 0,9702, este resultado sugere que 97% da capacidade inibidora
cátion radical radical ABTS•+ das amostras estudadas deve-se a contribuição de teor de
fenólicos totais. As amostras obtidas da 2a coleta não apresentaram coeficiente de
correlação linear (R2) significante (dados não mostrados).
y = 0,0005x + 0,0086R2 = 0,9702
00,010,020,030,040,050,060,070,080,090,1
0 50 100 150 200
Teor de Fenólicos Totais (mg de EAG/g)
ABTS
(1/C
E 50)
Gráfico 19: Correlação linear do teor de Fenólicos Totais (FT) em função de 1/CE50 da
atividade inibidora do cátion radical ABTS•+, dos extratos/frações de MSE da 3º coleta.
Há alguns estudos mostrando que o conteúdo de fenólicos totais de extratos
podem ser positivamente correlacionados com o potencial antioxidante (HUKKANEN
et al., 2006; KUTI, KONURU, 2004; VALENTÃO et al., 2002; GOVINDARAJAN et
al., 2003), por outro lado, recentes estudos não encontraram significativa correlação
entre o conteúdo de fenólicos totais e atividade antioxidante (KÄHKÖNEN et al.,
1999).

DISCUSSÃO Bauhinia cheilantha, popularmente conhecida como mororó-sem-espinho
(MSE), foi o objeto do estudo dessa dissertação, onde a atividade hipoglicemiante e
antioxidante desta planta foi avaliada. A escolha dessa planta é resultante de uma
pesquisa (NAPRALERT) que mostra a escassez de estudos relacionados com as
possíveis atividades farmacológicas do gênero Bauhinia. B. forficata, vulgarmente
conhecida como pata-de-vaca, é a espécie mais conhecida e estudada do gênero,
principalmente quando a atividade em questão é a hipoglicemiante. Esta planta é a
primeira de uma série de espécies do gênero com um possível uso como agente
coadjuvante no tratamento do DM. O estudo desta espécie visa, também, preservar a
espécie, uma vez que, a sua ocorrência é restrita a algumas áreas e os moradores da
região, frequentemente devastam a área para a criação de gado e ocultivo de plantações
diversas. O estudo fitoquímico e farmacológico desta espécie e outras relacionadas ao
gênero, ajudaria na preservação da mesma, ao aliar um possível interesse farmacológico
com o interesse econômico da população local.
Durante a execução deste projeto até o presente, poucos estudos foram
publicados atribuindo uma atividade hipoglicemiante a esta espécie, como o trabalho
feito por ALMEIDA et al., 2006. Neste estudo, foi realizado um total de 3 coletas para a
obtenção de material para a execução dos experimentos. Inicialmente, foi coletado uma
quantidade suficiente da planta para os estudos químicos e farmacológicos. Os testes
farmacológicos para determinar a atividade hipoglicemiante do extrato bruto etanólico,
na dose de 900 mg/kg, após repetidos testes para confirmar os resultados, reduziram a
glicemia em 46 %. As doses de 300 e 600 mg/kg não reduziram a glicemia
significativamente. Infelizmente, uma grande parte do material obtido das frações foi
utilizado e os resutados compilados não puderam ser aproveitados por ajustes feitos na
metodologia, troca do modelo animal (ratos por camundongos) e animais que seriam
aproveitados de acordo com a sua glicemia, na tentativa de reduzir o erro padrão da
média. A seguir, algumas razões são descritas pelas quais houve um gasto maior doque
o planejado, inicialmente:
Após um período de validação da metodologia que durou cerca de 3 meses e o
estabelecimento da concentração de aloxana de 150mg/Kg de peso para ratos machos e
120 mg/Kg de peso para fêmeas para a indução de diabetes nestes animais, alguns
parâmetros previamente descritos, foram alterados no projeto a fim de otimizar as

medições e os resultados apresentados. Os intervalos de tempo para a aferição da
glicemia nos animais foram alterados de 0, 15, 30, 60, 120, 180 minutos, para 0, 60,
120, 180 minutos. Essa alteração ocorreu em virtude de os valores glicêmicos nos
tempos de 15 e 30 minutos variarem pouco em relação ao tempo 0, além de reduzir o
estresse da coleta das amostras de sangue, o que ocasionaria valores alterados nas
medições. Outra espécie foi utilizada trocando a espécie animal, ratos por
camundongos. A troca do modelo animal deve-se às seguintes razões:
1. Com o progresso do fracionamento dos extratos das espécies de
Bauhinia, ocorreu um problema relacionado com a quantidade de
substância disponível para os experimentos farmacológicos. À medida
em que uma fração era particionada, quantidades menores eram
obtidas e frequentemente a quantidade era insuficiente para a
realização de experimentos com o número mínimo de animais,
requerido para um experimento.
2. Os camundongos são animais mais resistentes à indução de diabetes
por aloxana, ou seja, a mortalidade pós indução é menor, resultando
em um número de animais diabéticos após a administração de aloxana
maior, os valores glicêmicos são mais estáveis e menos variáveis em
seu estado basal. Frequentemente era necessário solicitar um número
maior de ratos (comparados aos camundongos) por serem animais
mais sensíveis a indução com aloxana. Frequentemente, os animais
não resistiam aos experimentos, levando a uma repetição dos mesmos
com um gasto maior de substância por dia testado.
3. O ciclo reprodutivo do camundongo é menor, consequentemente é
mais fácil obter camundongos com o peso adequado comparado com
ratos.
Ao mudar de espécie animal, a concentração de aloxana teve que ser
padronizada para os camundongos e determinada a via de indução (i.p. ou endovenosa)
por ter sido verificado que os valores glicêmicos mais constantes foram obtidos por via
endovenosa. A padronização do experimento durou cerca de 3 meses. Uma vez
consolidado, os experimentos tiveram que ser refeitos e concentração dos extratos foram
testados em níveis crescentes até os resultados já descritos terem sido obtidos. Dessa
forma, uma segunda coleta foi realizada para a verificaçãoda atividade, não apenas do
extrato bruto, mas também das frações resultantes da partição. Após repetidos testes

com o extrato bruto da 2a coleta, e com a constatação de que o efeito farmacológico
praticamente desaparecera, foi feito a atividade antioxidante pelo método do DPPH dos
extratos da 1a (CE50 = 11,67 ± 1,43 µg/mL) e da 2a coleta (CE50 = 26,68 ± 0,23 µg/mL),
para comparar esses resultados com aqueles referentes a atividade hipoglicemiante,
obtendo uma redução significativa da atividade anti-radicalar, justificando assim, o
decréscimo da atividade hipoglicemiante. A redução significativa da atividade
hipoglicemiante com as coletas sucessivas, sugere uma possível variação nos
constituintes da planta. A sazonalidade é um fator conhecido entre plantas. O teor de
compostos voláteis pode variar de acordo com a hora do dia em que as folhas ou
amostras de partes da plantas foram coletadas (SIXEL, PECINALLI, 2005). Apesar da
espécie em estudo não possuir em sua composição uma quantidade significativa de
óleos voláteis, a literatura reporta que a exposição da planta ao sol e à chuva altera a
proporção e a quantidade de compostos resultantes do metabolismo secundário da
planta podendo interferir na atividade biológica da mesma. As coletas, realizadas em
meses diferentes (janeiro, agosto e fevereiro de anos diferentes) provavelmente, resultou
em diferentes proporções de constituíntes ativos. A diferença de 6- 7 meses entre uma
coleta e outra poderia explicar a diferença de atividade entra as coletas, além da
precipitação pluvial ser maior nos meses do inverno. A quantidade de material vegetal
da 2a coleta foi bastante inferior à quantidade da primeira e uma 3a coleta foi necessária
para ser possível testar as frações e comparar os resultados não só com relação à
atividade hipoglicemiante mas para realizarmos os testes de atividade antioxidante
(DPPH, Fenólicos totais e ABTS) da planta. Todos os experimentos duvidosos foram
repetidos para assegurar que o resultado obtido poderia ser utilizado. Das três partições
obtidas (fração hexânica, fração aquosa e fração acetato de etila), apenas a fração
aquosa originada do material da última coleta realizada apresentou atividade
hipoglicemiante. Não foi possível testar a atividade antioxidante da fração hexânica da
1a coleta, pela indisponibilidade de material. A fração hexânica oriunda da 2a coleta, ao
ser testada, mostrou uma redução significativa na quantidade de fenólicos totais (132,39
± 0,98 EAG/g) e no teste do ABTS (CE50 = 7,16 ± 0,04) quando comparada com os
valores obtidos nos mesmo experimentos na 3a coleta: fenólico totais (FT) (37,5 ± 1,15
EAG/g) e no teste do ABTS (CE50 = 36,33 ± 0,38). No teste do DPPH, entretanto, a
fração hexânica apresentou um aumento em sua atividade, como mostrado pelos valores
seguintes: 2a coleta (CE50 = 38,98 ± 0,14) e 3a coleta (CE50 = 20,41 ± 0,15). O

coeficiente de correlação linear entre os testes revelou que apenas o teste FT e o teste do
ABTS apresentam um valor significativo: R2 = 0,97, mostrando que 97 % da capacidade
redutora dos constituíntes da amostra (fração) é devido aos componentes fenólicos
presentes na mesma. O método é aplicado para o estudo de antioxidantes hidrossolúveis
e lipofílicos, compostos puros e alimentos (RE et al., 1999).
De fato, as frações, extrato bruto e precipitados, em termos de atividade
antioxidante no teste do ABTS e FT obtidos na 2a coleta, apresentam a mesma
sequência em atividade antioxidante: Fr. AcOEt > Fr. Hex > Ext. EtOH > Fr. Aquosa >
PPT CH2Cl2 (diclorometano). Na 3ª coleta, em ambos os testes ABTS e FT as amostras
apresentaram-se na seguinte ordem de atividade: Fr. AcOEt > ext. EtOH > Fr. aquosa >
Fr. Hex.. Apesar do coeficiente de correlação linear entre os testes do DPPH e o de FT
apresentar um R2 = 0,690 (baixa correlação), o extrato bruto obtido da 2a coleta que
apresentou uma equivalência em termos de atividade e na 3a coleta, o PPT MeOH e a
fração AcOEt mantiveram o seu teor proporcional de fenólicos totais e a sua ordem de
atividade antioxidante (DPPH), este ensaio é considerado um ensaio válido e fácil para
avaliar a atividade seqüestradora de antioxidantes, já que o radical é estável e não
precisa ser gerado por reações como em outros ensaios de atividade seqüestradora.
Além disso, este ensaio vem sendo aplicado a uma grande variedade de produtos, como
alimentos e extratos (SANCHEZ-MORENO, 2002; ROGINSKY, LISSI, 2005).
Apesar de haver alguma correlação entre as frações e extratos, alguns estudos
mostram que a correlação entre a atividade antioxidante pode ser correlacionada
(GOVINDARAJAN et al., 1999; VALENTÃO et al., 2002; KUTI, KONURO, 2004;
HUKKANEN et al, 2006) enquanto outros mostram que não existe uma correlação
significativa entre o conteúdo de fenólicos totais e a atividade antioxidante
(KÄHKÖNEN et al., 1999). Aparentemente não existe uma correlação entre a atividade
hipoglicemiante e a atividade antioxidante, já que a amostra que apresentou maior
atividade antioxidante nos testes do DPPH e ABTS bem como maior teor de FT, a
fração AcOEt, não foi a fração com maior atividade hipoglicemiante. O extrato bruto
etanólico da 1a coleta foi a amostra com maior atividade, seguido da fração aquosa da 3ª
coleta, seguindo pela fração AcOEt da 3ª coleta. A fração aquosa da 3ª coleta por sua
vez, apresentou uma das menores atividades antioxidantes e das 5 amostras de frações,
PPT e extrato bruto, ficando na quarta colocação em termos de atividade antioxidante,
corroborando para uma correlação inversa. O extrato bruto da 1a coleta foi a única
amostra qua apresentou uma correlação entre as atividades em questão: o teste do DPPH

do extrato bruto da 1acoleta (CE50 = 11,67 ± 1,43 µg/mL), da 2a coleta (CE50 = 26,68 ±
0,23 µg/mL) e 3ª coleta (CE50 = 21,77 ± 0,126 µg/mL). Um argumento contra a
correlação entre a atividade hipoglicemiante e a antioxidante é a ausência de atividade
redutora da glicemia do PPT MeOH, sendo que o mesmo PPT apresentou os melhores
valores em termos de atividade antioxidante e teor de fenólicos totais em comparação
com as demais amostras. Com resultados igualmente opostos em termos de atividade, o
PPT CH2Cl2, apresentou uma das menores atividades antioxidantes e em teor de FT e
atividade hipoglicemiante nula. Substâncias que simultaneamente reduzem a glicemia,
restauram a secreçãode insulina e inibem o estresse oxidativo causado pela
hiperglicemia, parecem ser opções terapêuticas interessantes para a prevenção de
complicações diabéticas vasculares. O medicamento Diamicron MR (gliclazida) é um
hipoglicemiante oral pertencente à classe das sulfoniluréias. Vários estudos in vivo e in
vitro, demonstraram que Diamicron mostra uma atividade antioxidante independente da
redutora da glicemia em pacientes com DM tipo 2 (JENNINGS et al., 1991).
Comparada com a glibenclamida, em um estudo com pacientes com retinopatia,
Diamicron produziu uma diminuição significativa e sustentada dos níveis de lipídios
peroxidados e um aumento na atividade da enzima eritrocitária superóxido dismutase. É
interessante notar que o controle glicêmico dos pacientes de ambos os grupos não
diferiu, apoiando a hipótese de que o efeito de Diamicron devido às suas propriedades
antioxidantes, tem um impacto significativo na manutenção da integridade do sistema
vascular do diabético.
A atividade hipoglicemiante parece ser dose-dependente apenas no experimento
realizado com o extrato bruto da 1a coleta que apresentou atividade na maior dose
testada (900 mg/kg), o que não foi observado em experimentos com outras frações.
Duas vias foram utilizadas para a administração das amostras nos animais: a
intraperitoneal e a via oral (gavage). Os experimentos realizados com a administração
por via oral apresentaram resultados expressivos na atividade redutora da glicemia
quando comparados com a via intraperitoneal. Estes resultados são positivos, uma vez
que a administração de preparações de plantas consideradas medicinais é usualmente
feita através de chás e decoctos. Se o efeito da planta predomina através da via oral,
então provavelmente, a(s) substância(s) não é (são) destruída(s) pelo suco gástrico e
deve(m) ser absorvida(s) pela mucosa intestinal.
Os resultados apresentados neste trabalho originaram a vontade de saber um
pouco mais sobre algumas das questões apresentadas:

• Porque a atividade redutora da glicemia diminuiu nas coletas subsequentes?
• Poderia haver uma correlação entre a atividade hipoglicemiante e antioxidante
em alguns meses específicos do ano ou não é possível correlacionar?
• Que substância(s) possui(em) atividade redutora da glicemia? Qual(is) é(são)
a(s) classe(s) da(s) sustância(s)?
• Algum efeito hipoglicemiante poderia ser visualizado se o estudo fosse crônico
nas frações e doses que não apresentaram efeito durante o estudo agudo?
CONCLUSÕES
1. O extrato bruto etanólico da 1a coleta apresentou a melhor atividade
hipoglicemiante;
2. A fração hexânica de Bauhinia cheilantha das 3 coletas realizadas não possui
atividade hipoglicemiante; a fração aquosa possui discreta atividade
hipoglicemiante;
3. Os extratos/frações da planta proveniente da 3ª coleta possuem melhor atividade
antioxidante comparado com os extratos/frações da 2ª coleta;
4. Os flavonóides isolados do PPT MeOH, quercitrina e afzelina não são os
responsáveis pela atividade hipoglicemiante da planta;
5. Os resultados obtidos indicam que provavelmente existe mais de uma substância
ativa, uma vez que duas (frações acetato de etila e aquosa) das 3 frações
resultantes do particionamento do extrato bruto etanólico apresentaram um
efeito hipoglicemiante;
6. Não há correlação direta entre a atividade hipoglicemiante e a atividade
antioxidante.

PERSPECTIVAS
De acordo com os resultados obtidos, onde a espécie em estudo apresentou uma
variação sazonal de seus constituintes e conseqüentemente, na sua atividade biológica
sugere-se a necessidade de encontrar marcadores químicos que possam validar os
resultados obtidos de plantas e de outros produtos naturais bem como conceber a
possibilidade de, ao comercializar um produto natural, de alguma forma, notificar a
potência do produto de acordo com um ou mais marcadores químicos.
Além disso, pretende-se relacionar as diferenças entre a atividade
hipoglicemiante, a atividade antioxidante com a época da coleta. Bem como, comparar
o efeito farmacológico de B. cheilantha com outras espécies já caracterizadas (ex: B.
forficata, B. monandra) na literatura pela sua ação redutora da glicemia, uma vez que
alguns dados sobre a atividade hipoglicemiante das espécies citadas anteriormente
variam de acordo com o grupo de pesquisa.
Em adição aos experimentos a serem realizados neste projeto já citados
anteriormente, pretende-se realizar um estudo crônico da atividade de B. cheilantha para
determinarmos se há diferença na atividade hipoglicemiante quando comparado com o
estudo agudo.
Por fim, pretende-se realizar um estudo comparativo de extratos e frações das
plantas que apresentarem o melhor efeito hipoglicemiante durante o estudo comparativo
sazonal, em um modelo de intestino invertido para acessar a absorção dos constituintes
que são absorvidos pela mucosa ileal de ratos. O objetivo da continuidade deste estudo é
tentar elucidar o(s) constituinte(s) do material em estudo com possível atividade
biológica que poderia(m) estar atuando como componente(s) ativo(s), uma vez que não
foi possível designar a(s) substância(s) ativa(s), mas apenas excluir que isoladamente,
os flavonóides isolados, não apresentaram efeito.

ACHENBACH, H., STOCKER, M., CONSTENLA, M. A. F. Phytochemistry, 27, 1835, 1988. ADEROGBA, M. A.; MCGAW, L. J.; OGUNDAINI, A. O.; ELOFF, J. N. Antioxidant activity and cytotoxicity study of the flavonol glycosides from Bauhinia galpinii. Bioactive Natural Products, 21(7), 591-599, 2007. AKERELE, O. Summary of WHO guidelines for the assessment of herbal medicines. Herbal Gram, 28, 13, 1993. AKHTAR, F.M., ALI, M.R. Journal Pakistan Medical Association, 34, 239, 1984. ALARCON-AGUILAR, F.J., ROMAN-RAMOS, R., FLORES-SAENZ, J.L., AGUIRRE-GARCIA, F. Investigation on the hypoglycaemic effects of extracts of four Mexican medicinal plants in normal and alloxan-diabetic mice. Phytother Res, 16, 383-386, 2002. ALMEIDA, E.R., GUEDES, M.C., ALBUQUERQUE, J.F.C., XAVIER, H. Hypoglycemic effect of Bauhinia cheilandra in rats. Fitoterapia 77: 276–278, 2006. ALMEIDA, R.N., AGRA, M.F. Plantas medicinais. Revista Brasileira de Farmacognosia, 1986. ALONSO, A. M., DOMÍNGUEZ, C., GUILLÉN, D. A., BARROSO, C. G. Determination of antioxidant power of red and white wines by a new electrochemical method and its correlation with polyphenolic content. Journal of Agricultural and Food Chemistry, 50(11), 3112–3115, 2002. American Diabetes Association (ADA). Gestational diabetes mellitus. Clinical Practice Recommendations 2001. Diabetes Care; 24(Suppl 1):S77-9, 2001. American Diabetes Association (ADA). Standards of Medical Care in Diabetes. Diabetes Care, 27 ( SUPPL 1), 2004. American Diabetes Association. Diagnosis and classification of diabetes mellitus. Diabetes Care, 28, S37–S42, 2005. American Diabetes Association. Standards of medical care for patients with diabetes mellitus: position statement. Diabetes Care, 22(1S), 32S–41S, 1999. AMES, B.N.; SHIGENAGA, M.K.; HAGEN, T.M. Oxidants, Antioxidants, and the Degenerative Diseases of Aging. Proceedings of the National Academy of Sciences of the United States of America, 90 (17),. 7915-7922, 1993. ANDRADE, J., CONDE, M., SOBRINO, F. BEDOYA, F.J. Activation of peritoneal macrophages during the prediabetic phase in lowdose streptozotocin-treated mice. FEBS Lett, 327, 32–34,1993.

ANELLO, M., GILON, P., HENQUIN, J. C. Alterations of insulin secretion from mouse islets treated with sulphonylureas: perturbations of Ca2+ regulation prevail over changes in insulin content. British Journal of Pharmacology, 127,1883–1891, 1999. ANKLI, A.; HEINRICH, M.; BORK, P.; WOLFRAM, L.; BAUERFEIND, P.; BRUN, R.; SCHMID, C.; WEISS, C.; BRUGGISSER, R.; GERTSCH, J.; WASESCHA, M.; STICHER, O. Yucatec Mayan medicinal plants: evaluation based on indigenous uses. Journal of Ethnopharmacology, 79, 43-52, 2002. ARGOLO, A.C.C., SANT’ANA, A.E.G., PLETSCH, M., COELHO, L.C.B.B. Antioxidant activity of leaf extracts from Bauhinia monandra. Bioresource Technology 95, 229–233, 2004. ASHCROFT, F. M, RORSMAN, P. Electrophysiology of the pancreatic b-cell. Prog Biophysics Molecular Biology , 54, 87±143, 1989. ATKINSON, M.A., MACLAREN, N.K. The pathogenesis of insulin-dependent diabetes mellitus. The New England Journal of Medicine, 331, 1428 –36, 1994. BALUCHNEJADMOJARAD, T.; ROGHANI, M.; HOMAYOUNFAR, H.; HOSSEINI, M. Beneficial effect of aqueous garlic extract on the vascular reactivity of streptozotocin diabetic rats. Journal of Ethnopharmacology, 85, 139-144, 2003. BALUNAS, M. J., KINGHORN, D. Drug Discovery from medicinal plants. Life Sciences. 78. p. 431-41. 2005. BANSAL, R., AHMAD, N., KIDWAI, J.R: Alloxan-glucose interaction: effect of incorporation of 14C-leucine into pancreatic islets of rats. Acta Diabetologica Latina, 17, 135-143, 1980. BARBOSA-FILHO, J. M., VASCONCELOS, T. H.C., ALENCAR, A. A., BATISTA, L. M., OLIVEIRA, R.A.G., GUEDES, D. N., FALCÃO, H. S., MOURA, M. D., DINIZ, M. F.F.M., MODESTO-FILHO, J. Plants and their active constituents from South, Central, and North America with hypoglycemic activity. Revista Brasileira de Farmacognosia,15(4), 392-413, 2005. BARBOSA-FILHO, J.M., VASCONCELOS, T.H.C., ALENCAR, A.A., BATISTA, L.M., OLIVEIRA, R.A.G., GUEDES, D.N., FALCÃO, H.S., MOURA, M.D., DINIZ, M.F.F.M., MODESTO-FILHO, J. Plants and their active constituents from South, Central, and North America with hypoglycemic activity. Revista Brasileira de Farmacognosia, 15: 392-413, 2005. BARG, S. Mechanisms of Exocytosis in Insulin-Secreting B-Cells and Glucagon Secreting A-Cells. Pharmacology and Toxicology, 92, 3–13, 2003. BASTA, G., SCHMIDT, A.M., DE CATERINA, R. Advanced glycation end products and vascular inflammation: implications for accelerated atherosclerosis in diabetes. Cardiovascular Research, 63, 582–592, 2004.

BECKER, D.J., CAKAN, N., LAPORTE, R.E., et al. High risk DQ alleles improve IDDM prediction in ICA + ve first-degree relatives with decreased insulin secretion [abstract]. Diabetologia; 38(Suppl 1):A42, 1995. BEDOYA, F.J., SOLANO, F., LUCAS, M. N-monomethyl-arginine and nicotinamide prevent streptozotocin-induced double strand DNA break formation in pancreatic rat islets. Experientia, 52, 344-347, 1996. BELFIORE, F., MOGENSEN, C.E. (eds). New Concepts in Diabetes and Its Treatment – Switzerland: Basel, Karger, 2000. BELL, G.L, PICTET, R.L., RUTTER, W.J., CORDELL, B., TISCHER, E., GOODMAN, H.M. Sequence of human insulin gene, Nature, 284: 26, 1980. BENNET, R.A., PEGG, A.E. Alkylation of DNA in rat tissues following administration of streptozotocin. Cancer Research, 41, 2786–2790, 1981. BERNE, R. M., LEVY, M. N., KOEPPEN, B. M. Fisiologia, 4. ed. - Rio de Janeiro: Guanabara Koogan, 1998. BLAKE, D.; WINYARD, P. G. Immunopharmacology of Free Radical species. San Diego: Academic Press, 1995.
BOLAFFI, J.L., NAGAMATSU, S., HARRIS, J., GRODSKY, G.M. Protection by thymidine, an inhibitor of polyadenosine diphosphate ribosylation, of streptozotocin inhibition of insulin secretion. Endocrinology, 120, 2117-2122, 1987. BOQUIST, L., NELSON, L., LORENTZON, R: Uptake of labelled alloxan in mouse organs and mitochondria in vivo and in vitro. Endocrinology, 113, 943-948, 1983. BOTTINO, R., BALAMURUGAN, A.N., GIANNOUKAKIS, N, et al. Islet/pancreas transplantation: challenges for pediatrics. Pediatric Diabetes, 3, 210–23, 2002. BOTTINO, R., TRUCCO, M., BALAMURUGAN, A.N. et al. Pancreas and islet cell transplantation. Best Practice and Research Clinical Gastroenterology, 16, 457– 74, 2002. BOUGNERES, P.F, LANDAIS, P., BOISSON, C., et al. Limited duration of remission of insulin dependency in children with recent overt type I diabetes treated with low-dose cyclosporin. Diabetes, 39,1264–72, 1990. BOYLAN, J.M., BRAUTIGAN, D.L., MADDEN, J., RAVEN, T., ELLIS, L., GRUPPUSO, P.A. Differential regulation of multiple hepatic protein tyrosine phosphatases in alloxan diabetic rats. Journal of Clinical Investigation, 90, 174-179, 1992. BRASIL. Ministério da Saúde. Portaria nº 971, de 03 de maio de 2006. Aprova a Política Nacional de Práticas Integrativas e Complementares (PN- BRASIL PIC) no Sistema Único de Saúde. D.O.U. Poder Executivo, Brasília, 04 mai. 2006.

BRATANOVA-TOCHKOVA, T. K., CHENG, H., DANIEL, S., GUNAWARDANA, S., LIU, Y-J., MULVANEY-MUSA, J., SCHERMERHORN, T., STRAUB, S. G., YAJIMA, H., SHARP, G. W. G. Triggering and augmentation mechanisms, granule pools, and biphasic insulin secretion. Diabetes, 51 [Suppl 1]:S83–S90, 2002. BROWNLEE, M. Biochemistry and molecular cell biology of diabetic complications. Nature, 414, 813–820, 2001. BROWNLEE, M. Glycation and diabetic complications. Diabetes, 43, 836–841, 1994. CALIXTO, J. B.; YUNES, R.A.; RAE, G. A.; MEDEIROS, Y.S. In Bradykinin Antagonists: Basic and Clinical Research; R.M. Burch, Marcel Dekker Inc. New York, 1990, p. 88. CAMPOS, M.G.; WEBBY, R.F.; MARKHAM, K.R.; MITCHELL, K.A.; DA CUNHA, A.P. Age-induced diminution of free radical scavenging capacity in bee pollens and the contribution of constituent flavonoids. Journal of Agricultural and Food Chemistry, 51 (3), 742-745, 2003. CARVALHEIRA, J. B. C., ZECCHIN, H. G., SAAD, M. J. A. Vias de Sinalização da Insulina, Arquivo Brasileiro de Endocrinologia e Metabologia, 46/4, 419-425, 2002. CASU, A., TRUCCO, M., PIETROPAOLO, M. A Look to the Future: Prediction, Prevention, and Cure Including Islet Transplantation and Stem Cell Therapy. Pediatric Clinics of North América, 52, 1779– 1804, 2005. CECHINEL FILHO, V. Tese de Doutorado em Química, UFSC, Florianópolis, SC, 1995. CECHINEL FILHO, V.; BREVIGLIERI, E.; WILLAIN FILHO, A.; SANTOS A.R.S. Revista Brasileira de Farmacognosia, 76, 115. 1995. CECHINEL-FILHO, V., YUNES, R.A. Estratégias para a obtenção de compostos farmacologicamente ativos a partir de plantas medicinais. Conceitos sobre modificação estrutural para otimização da atividade. Química nova, 21, 99-105, 1998. CINGOLANI, H. E. Fisiologia Humana de Houssay/ Horacio E. Cingolani e Alberto B. Houssay; trad. Adriane Belló Klein...[et. al.],- 7. ed. atual. ampl. – Porto Alegre: Artmed, 2004. CLARK, M. Understanding diabetes. England: John Wiley & Sons Ltd, 2004. COLMAN, P.G., MCNAIR, P., MARGETTS, H., et al. The Melbourne Pre-Diabetes Study: prediction of type 1 diabetes mellitus using antibody and metabolic testing. Med J Aust,169, 81– 4, 1998. COOPER, M.E. Importance of advanced glycation end products in diabetes-associated cardiovascular and renal disease. American Journal of Hypertension, 17, 31S–38S, 2004. CORRÊA, A. G.; Taxol: da descoberta ao uso terapêutico. Quím. Nova, 18, 460, 1995.

Da SILVA, E. G., BEHR, G. A., ZANOTTO-FILHO, A., LORENZI, R., PASQUALI, M. A. De B., RAVAZOLO, L. G., BORDIGNON-JR, C. L., Da SILVA, F. A., ABOY, A. L., BASSANI, V. L., HENRIQUES, A. T., REGINATTO, F. H., DAL-PIZZOL, F., MOREIRA, J. C. F. Antioxidant Activities and Free Radical Scavenging Potential of Bauhinia microstachya (RADDI) MACBR. (Caesalpinaceae) Extracts Linked to Their Polyphenol Content. Biological and Pharmaceutical Bulletin, 30(8): 1488—1496, 2007. DAI, Y., VAUGHT, T.D., BOONE, J., et al. Targeted disruption of the alpha1,3-galactosyltransferase gene in cloned pigs. Nature Biotechnology, 20, 251–5, 2002. DASGUPTA, N., DE, B. Antioxidant activity of some leafy vegetables of India: A comparative study. Food Chemistry, 101, 471–474, 2007. DAVIS, S. N. Insulin, Oral Hypoglycemic Agents, and The Pharmacology Of The Endocrine Pancreas. In: GOODMAN, A.; GILMAN, H. L. The Pharmacological Basis of Therapeutics. 11. ed. Rio de Janeiro: Mc Graw-Hill, 2006. DE GROOT, L.J.: Endocrinology, 3 ed, Philadelphia, 1994, WB Saunders. DE MEYTS, P. The structural basis of insulin and insulin-like growth factor-I receptor binding and negative co-operativity, and its relevance to mitogenic versus metabolic signalling. Diabetologia, 37, S135-148, 1994. DE SOUSA, E., ZANATTA, L., SEIFRIZ, I., CRECZYNSKI-PASA, T. B., PIZZOLATTI, M. G., SZPOGANICZ, B., SILVA, F. R. M. B. Hypoglycemic Effect and Antioxidant Potential of Kaempferol-3,7-O-(r)-dirhamnoside from Bauhinia forficata Leaves J. Nat. Prod., 67, 829-832, 2004. DE SOUSA, E., ZANATTA, L., SEIFRIZ, I., CRECZYNSKI-PASA, T. B., PIZZOLATTI, M. G., SZPOGANICZ, B., SILVA, F. R. M. B. Hypoglycemic Effect and Antioxidant Potential of Kaempferol-3,7-O-(r)-dirhamnoside from Bauhinia forficata Leaves. Journal of Natural Products, 67, 829-832, 2004. DE SOUZA, N. J.; Journal of Ethnopharmacology, 38,177, 1993. DEAN, P.M. Ultrastrutural morphometry of the pancreatic β-cell. Diabetologia, 9, 115-119, 1973. Diabetes 2004. Disponível em: <www.diabetes.org.br. Acessada em: novembro. 2004. DRAZNIN, B., MELMED, S., LEROITH, D. Insulin secretion. In: Molecular and Cellular Biology of Diabetes Mellitus, vol. I. Alan R. Liss Inc., New York, 1989. DUCKWORTH, W.C. Insulin degradation: Mechanisms, products, and significance. Endocrine Reviews., 9, 319-345, 1988. DUNN, J.S., SHEEHAN, H.L., MCLETHIE, N.G.B. Necrosis of islets of Langerhans produced experimentally. Lancet, 1: 484-487, 1943.

EISENBARTH, G.S. Type I diabetes mellitus: a chronic autoimmune disease. The New England Journal Medicine, 314, 1360– 8, 1986. EISENBARTH, G.S., GIANANI, R., YU, L., et al. Dual-parameter model for prediction of type I diabetes mellitus. Proc Assoc Am Physicians,110, 126– 35, 1998. EIZIRIK, D.L., PIPELEERS, D.G., LING, Z., WELSH, N., HELLERSTROM, C., ANDERSSON, A. Major species differences between humans and rodents in the susceptibility to pancreatic beta-cell injury. Proc Natl Acad Science USA, 91, 9253-9256, 1994. EL-HILALY, J., TAHRAOUI, A., ISRAILI, Z. H., LYOUSSI, B. Hypolipidemic effects of acute and sub-chronic administration of an aqueous extract of Ajuga iva L. whole plant in normal and diabetic rats. Journal of Ethnopharmacology, 105, 441–448, 2006. FEHER, M.D. Diabetes: preventing coronary heart disease in a high risk group. Heart (British Cardiac Society), 90, iv18–iv21 (Review), 2004. FERBER, S., HALKIN, A., COHEN, H, et al. Pancreatic and duodenal homeobox gene 1 induces expression of insulin genes in liver and ameliorates streptozotocin-induced hyperglycemia. Nature Medicine, 6,568 – 72, 2000. Fitoterapia - Suplemento dedicado inteiramente à química e farmacologia do taxol e derivados, 62, 1995. FLATT, P.R.: Nutrient Regulation of Insulin Secretion. Portland Press, 1992. GABBAY, K.H. Hyperglycemia, polyol metabolism, and complications of diabetes mellitus. Annual Review Medicine, 26, 521– 36, 1975. GABRIELY, I., YANG, X.M., CASES, J.A., MA, X.H., ROSSETTI, L., BARZILAI, N. Hyperglycemia induces PAI-1 gene expression in adipose tissue by activation of the hexosamine biosynthetic pathway. Atherosclerosis, 160, 115–122, 2002. GANDA, O.P., ROSSI, A.A., LIKE, A.A. Studies on streptozotocin diabetes. Diabetes, 25: 595-603, 1976. GARCIA, E.S.; SILVA, A.C.P.; GILBERT, B.; CORRÊA, C.B.V.; CAVALHEIRO, M.V.S.; SANTOS, R.R.; TOMASINI, T. Fitoterápicos. Campinas: André Tosello, 1996. 17p. GIUGLIANO D, CERIELLO A Oxidative stress and diabetic vascular complications. Diabetes Care, 19, 257–267, 1996. GIUGLIANO, D., CERIELLO, A., PAOLISSO, G. Oxidative stress and diabetic vascular complications. Diabetes Care,19, 257– 67, 1996.

GLASTRAS, S. J., MOHSIN, F., DONAGHUE, K. C., Complications of Diabetes Mellitus in Childhood. Pediatric Clinics of North America, 52, 1735– 1753, 2005. GOLDBERG, H.J., WHITESIDE, C.I., FANTUS, I.G. The hexosamine pathway regulates the plasminogen activator inhibitor-1 gene promoter and Sp1 transcriptional activation through protein kinase C-′βI and -′δ. Journal of Biological Chemistry, 277, 33833–33841, 2002. GOVINDARAJAN R., RASTOGI S., VIJAYAKUMAR M., SHIRWAIKAR A., RAWAT A. K. S., MEHROTRA S., PUSHPANGADAN P. Studies on the Antioxidant Activities of Desmodium gangeticum. Biological and Pharmaceutical Bulletin 26,1424—1427, 2003. GRAAFF, Van De. Human Anatomy, 6. ed. – The McGraw−Hill Companies, 2001. GRANNER, D.K, ANDREONE, T.L.: Insulin modulation of gene expression, Diabetes Metabolism Review, 1,139, 1982. GREENBAUM, C.J, SEARS, K.L, KAHN, S.E, et al. Relationship of beta-cell function and autoantibodies to progression and nonprogression of subclinical type 1 diabetes: follow-up of the Seattle Family Study. Diabetes; 48, 170–5, 1999. GROSS, J. L., SILVEIRO, S.P., CAMARGO, J. L., REICHELT, A. J., AZEVEDO, M. J. Diabetes Melito: Diagnóstico, Classificação e Avaliação do Controle Glicêmico. Arquivo Brasileiro Endocrinologia Metabolismo, 46, 16-26, 2002. GROUP, T.D.C. Effect of intensive therapy on residual beta-cell function in patients with type 1 diabetes in the diabetes control and complications trial: a randomized, controlled trial. Annals of Internal Medicine, 128, 517– 23, 1998. GROUP, T.D.R. Effects of age, duration and treatment of insulin-dependent diabetes mellitus on residual beta-cell function: observations during eligibility testing for the Diabetes Control and Complications Trial (DCCT). J Clin Endocrinol Metab, 65, 30–6, 1987. GROVER, J. K.; VATS, V.; RATHI, S.S.; DAWAR, R. Traditional Indian anti-diabetic plants Attenuate progression of renal damage in streptozotocin induced diabetic mice. Journal of Ethnopharmacology, 76, 233-236, 2001. GRUPPUSO, P.A., BOYLAN, J.M., POSNER, B.I., FAURE, R., BRAUTIGAN, D.L: Hepatic protein phosphotyrosine phosphatase. Dephosphorylation of insulin receptor and epidermal growth factor receptors in normal and alloxan diabetic rats. Journal of Clinical Investigation, 85, 1754-1760, 1990. GUERRA, M. P.; NODARI, R. O. Farmacognosia: da planta ao medicamento. Capítulo I, p. 14, 5° ed. Ver. Ampl. Ed. Da UFRGS/ Ed. Da UFSC, Porto Alegre/ Florianópolis, 2003.

GUILLAUSSEAU, P.J., MASSIN, P., DUBOIS-LAFORGUE, D., TIMSIT, J.,VIRALLY, M., GIN, H. et al. Maternally inherited diabetes and deafness: a multicenter study. Annals of Internal Medicine, 8,134-721, 2001. GUPTA, M. P., ed., 270 Plantas Medicinales Iberoamericanas; alleres de Editorial Presencia, Santafé de Bogotá: Colômbia, p. 347, 1995. HALLER, M. J., ATKINSON, M. A., SCHATZ, D., Type 1 Diabetes Mellitus: Etiology, Presentation, and Management. Pediatric Clinics of North America, 52, 1553– 1578, 2005. HALLIWELL, B. Antioxidants and human disease: A general introduction. Nutrition Reviews, 55 (1), 44-49, 1997.
HALLIWELL, B.; GUTTERIDGE, J. M. C. The chemistry of free radicals and related “reactive species”. Free Radicals in Biology and Medicine. Oxford: Oxford University Press, 1998.
HARBONE, J. B. Plant phenolics. In E. A. Bell & B. V. Charlwood (Eds.). Encyclopedia of plant physiology, secondary plant products. New York: Springer-Verlag, Berlin Heidelberg, 8: 329–395, 1980. HARBORNE J. B. THE FLAVONOIDS: ADVANCES IN RESEARCH SINCE 1988. Chapman & Hall, New York, 1998 HARBORNE J.B. (Ed) Biochemistry of Phenolic Compounds, Academic Press, London, 1964; HARRISON, L.C. Risk assessment, prediction and prevention of type 1 diabetes. Pediatr Diabetes, 2, 71–82, 2001. HASLAM, E. Practical Polyphenolics. Cambridge University Press, Cambridge, UK, 1998; HAVSTEEN, B. Flavonoids, a class of natural products of high pharmacological potency Biochemistry Pharmacology, 32, 1141. 1983. HEIKKILA, R.E., WINSTON, B., COHEN, G., BARDEN, H: Alloxan induced diabetes, evidence for hydroxyl radicals as a cytotoxic intermediate. Biochemical Pharmacology, 25, 1085-1092, 1976. HOAREAU, L., DA SILVA, E. J. Medicinal plants: a re-emerging health aid. Electronic Journal of Biotechnology, 2 (2), 1999. HORWITZ, S. B. How to make taxol from scratch. Nature, 367, 593, 1994. HOU, C. C.; LIN, S. J.; CHENG, J. T.; HSU, F. L. Antidiabetic dimeric guianolides and a lignan glycoside from Lactuca indica. Journal of Naturals Products, 66, 625-629, 2003.

HOUNSOM L., CORDER R., PATEL J., TOMLINSON D.R.. Oxidative stress participates in the breakdown of neuronal phenotype in experimental diabetic neuropathy. Diabetologia 44, 424–428, 2001. HUANG, D., OU, B., PRIOR, R. L. The chemistry behind antioxidant capacity assays. Journal of Agricultural and Food Chemistry, 53, 1841–1856, 2005. HUANG, D., OU, B., PRIOR, R. L. The chemistry behind antioxidant capacity assays. Journal of Agricultural and Food Chemistry, 53: 1841–1856, 2005. HUKKANEN A. T., PÖLÖNEN S. S., KÄRENLAMPI S. O., KOKKO H. I., Journal of Agricutural and Food Chemistry, 54, 112—119, 2006. IARC Working Group, Some N-nitroso compounds. In: IARC Monographs on the Evaluation of the Carcinogenic Risk of Chemicals to Humans, pp. 337–349. IARC Scientific Publications No. 17, IARC, Lyon, 1978. ILDIS and CHCD, Phytochemical Dictionary of the Leguminosae; Chapman & Hall: London, Vol. 1, p 118, 1992. INOGUCHI, T.; TSUBOUCHI, H.; ETOH, T.; KAKIMOTO, M.; SONTA, T.; UTSUMI, H.; SUMIMOTO, H.; YU, H.Y.; SONODA, N.; INUO, M.; SATO, N.; SEKIGUCHI, N.; KOBAYASHI, K. E NAWATTA, H. A Possible Target of Antioxidative therapy for diabetic vascular complications-vascular NAD(P)H oxidase. Current Medicinal Chemistry. (10): 1759-1764. 2003a. INOGUCHI, T.; SONTA, T.;TSUBOUCHI, H.; ETOH, T.; KAKIMOTO, M.; SONODA, N.; SATO, N.; SEKIGUCHI, N.; KOBAYASHI, K; SUMIMOTO, H.; UTSUMI, H. E NAWATTA, H. Protein kinase C-dependent increase in reactive oxygen species (ROS) production in vascular tissues of diabetes: role of vascular NAD(P)H oxidase. Journal of the American Society of Nephrology, (14), S227-S232. 2003b. ISHIHARA, H., WANG, H., DREWES, L. R., WOLLHEIM, C. B. Overexpression of monocarboxylate transporter and lactate dehydrogenase alters insulin secretory responses to pyruvate and lactate in b cells. Journal of Clinical Investigation, 104, 1621–1629, 1999. ISLAS-ANDRADE, S.; MONSALVE, M.C.R.; DE LA PEÑA, J.E.; POLANCO, A.C.; PALOMINO, M.A.; VELASCO, A.F. Streptozotocin and alloxan in experimental diabetes: comparison of the two models in rats. Acta Histochemica et Cytochemica, 33 (3), 201-208, 2000. IVORRA, M. D.; PAYÁ, M.; VILLAR, A. A review of natural products and plants as potential antidiabetic drugs. Journal of Ethnopharmacology, 27, 243-275, 1989. IZE-LUDLOW, D., SPERLING, M. A. The Classification of Diabetes Mellitus: A Conceptual Framework. Pediatric Clinics of North America, 52, 1533– 1552, 2005.

JAMES, L.R., TANG, D., INGRAM, A., LY, H., THAI, K., CAI, L., SCHOLEY, J.W. Flux through the hexosamine pathway is a determinant of nuclear factor Κb dependent promoter activation. Diabetes 51, 1146–1156, 2002. JEFFERSON, L.S. Role of insulin in the regulation of protein synthesis. Diabetes 29:487, 1980. JENNINGS, P.E.; SCOTT, N.A.; SANIABADI, A.R., BELC; H.J. F., Effects of gliclazide on platelet reactivity and free radicals in type II diabetic patients: clinical assessment. Metabolism; 41(suppl 1), 36-39, 1992. JOHANSSON, E.B., TJA¨LVA, H. Studies on the tissue-disposition and fate of [14C] streptozotocin with special reference to the pancreatic islets. Acta Endocrinologica, 89, 339–351, 1978. JOHNSON, I.T. Antioxidants and antitumour properties. In: POKORNY, J., YANISHLIEVA, N., GORDON, M. (Eds.), Antioxidants in Food. Woodhead Publishing Ltda., Cambridge, 100–123, 2001. JÖRNS, A., MUNDAY, R., TIEDGE, M., LENZEN, S: Comparative toxicity of alloxan, N-alkylalloxans and ninhydrin to isolated pancreatic islets in vitro. Journal of Endocrinology, 155, 283-293, 1997. JULIANI, C. Ação hipoglicemiante da unha de vaca. Revista Medica de Pharmacia, Chimica e Physica 2, 165-169, 1929. KÄHKÖNEN M. P., HOPIA A. I., VUORELA H. J., RAUHA J., PIHLAJA K., KUJALA T. S., HEINONEN M. Antioxidant Activity of Anthocyanins and Their Aglycons. Journal of Agricutural and Food Chemistry, 47, 3954—3962, 1999. KAHN, S.E. Regulation of β-cell function in vivo. Diabetes Metabolism Review, 4, 372, 1996. KAMCHONWONGPAISON, S.; MESHNICK, S. R.; Gen. Pharmac., 27, 587, 1996. KASUGA, M., FUJITA-YAMAGUCHI, Y., BLITHE, D., KAHN, C.R. Tyrosine-specific protein kinase activity is associated with purified insulin receptor. Proc Natl Acad Sci USA 80:2137–2141, 1983. KASUGA, M., KARLSSON, F.A., KAHN, C.R. Insulin stimulates the phosphorylation of the 95–000 dalton subunit of its own receptor. Science 215: 185–187, 1982. KATSUMATA, K., KATSUMATA, K., KATSUMATA, Y. Protective effect of diltiazem hydrochloride on the occurrence of alloxan- or streptozotocin-induced diabetes in rats. Horm Metabolism Research, 24, 508-510, 1992. KERRY, N.; RICE-EVANS, C. Inhibition of Peroxynitrite-Mediated Oxidation of Dopamine by Flavonoid and Phenolic Antioxidants and Their Structural Relationships J. Neurochem., 73, 247-253, 1999.

KHAN A. H., PESSIN J. E. Insulin regulation of glucose uptake: a complex interplay of intracellular signalling pathways. Diabetologia 45:1475–1483, 2002. KIESS, W., BRANSKI, D. Diabetes in Childhood and Adolescence. Switzerland: Karger, 2005 KING, G. L. E BROWNLEE, M. The cellular and molecular mechanisms of diabetic complications. Endocrinology and Metabolism Clinics of North America. 25 (2): 255-270. 1996. KING, G. L., LOEKEN, M. R. Hyperglycemia-induced oxidative stress in diabetic complications. Histochemistry and Cell Biology, 122, 333–338, 2004. KING, H.; AUBERT, R.E. E GERMAN, W. H. Global Burden of Diabetes, 1995-2025. Diabetes Care (21): 1414-1431, 1998. KINGSTON, D. G. I.; The chemistry of Taxol. Pharmac. Ther., 52, 1, 1991. KOIKE, C., FRIDAY, R.P, NAKASHIMA I, et al. Isolation of the regulatory regions and genomic organization of the porcine alpha1,3-galactosyltransferase gene. Transplantation, 70, 1275– 83, 2000. KOWLURU R.A., KENNEDY A. Therapeutic potential of anti-oxidants and diabetic retinopathy. Expert Opinion Investigation Drugs, 10,1665–1676, 2001. KUNISAKI M., BURSELL S-E., CLERMONT A.C., ISHII H., BALLAS L.M., JIROUSEK M.R., UMEDA F., NAATA H., KING G.L. Vitamin E prevents diabetes-induced abnormal retinal blood flow via the diacylglycerol-protein kinase C pathway. Am Journal of Physiol Endocrinol Metabolism, 269, 239–246, 1995. KUROKI, T.; ISSHIKI, K. E KING, G. L. Oxidative stress: the lead or supporting actor in the pathogenesis of diabetic complication. Journal of the American Society of Nephrology. 14, 216-220, 2003. KUTI J. O., KONURU H. B. Journal of Agricutural and Food Chemistry, 52, 117—121, 2004. LAMBA, S. S.; BUCH, K.Y.; LEWIS, H.; LAMBA, H. J. Phytochemicals as potential hypoglycemic agents. Studies in Natural Products Chemistry, 21, 457- 495, 2000. LANG, J. Molecular mechanism and regulation of insulin exocytosis as a paradigm of endocrine secretion. European Journal of Biochemistry, 259, 3-17, 1999. LARNER J. The pharmacological basis of therapeutics. 7th ed. New York: Mac Millan; 1985. LEAHY, J. L., CEFOLU, W. T. Insulin Therapy, 1. ed. - New York: Marcel Dekker, 2002. LEE, H. B.; HA, H. E KING, G. L. Reactive oxygen species and diabetic nephropathy. Journal of the American Society of Nephrology, 14, 209-210. 2003.

LENZEN, S., MUNDAY, R. Thiol-group reactivity, hydrophilicity and stability of alloxan, its reduction products and its N-methyl derivatives and a comparison with ninhydrin. Biochemical Pharmacology, 42, 1385-1391, 1991. LENZEN, S., PANTEN, U. Alloxan: history and mechanism of action. Diabetologia, 31, 337-342, 1988. LENZEN, S., TIEDGE, M., JORNS, A., MUNDAY, R. Alloxan derivatives as a tool for the elucidation of the mechanism of the diabetogenic action of alloxan. In: Lessons from Animal Diabetes, E SHAFRIR (ed), Birkhauser, Boston, 1996, pp 113-122. LERCO, M. M., SPADELLA, C. T., MACHADO, J. L. M., SCHELLINI, S. A., PADOVANI, C. R. Caracterização de um modelo experimental de Diabetes mellitus, induzido pela aloxana em ratos. Estudo clínico e çaboratorial. Acta Cirúrgica Brasileira, 18: 132-142, 2003. LI, W. L.; ZHENG, H. C.; BUKURU, J.; De KIMPE, N. Natural medicines used in the traditional chinese medical system for therapy of diabetes mellitus. Journal of Ethnopharmacology, 92, 1-21, 2004. LIKE, A.A., APPLE, M.C., WILLIAMS, R.M., ROSSINI, A.A. Streptozotocin-induced pancreatic insulitis in mice: Morphologic and physiologic studies. Laboratory Investigation, 38, 470–486, 1978. LIKE, A.A., ROSSINI, A.A. Streptozotocin-induced pancreatic insulitis: New model in mice. Science, 193: 415–417, 1976. LINO, C. D.; DIOGENES, J. P. L.; PEREIRA, B. A.; FARIA, R. A. P. G.; NETO, M. A.; ALVES, R. S.; De QUEIROZ, M. G. R.; De SOUSA, F. C. F.; VIANA, G. S. B. Antidiabetic activity of Bauhinia forticata extracts in alloxan-diabetic rats. Biological and Pharmaceutical Bulletin., 27, 125- 127, 2004. LIZCANO, J. M.; ALESSI, D. R. The insulin signalling pathway. Current Biology, 12, 236-238, 2002. LUKENS, F.D.W. Alloxan diabetes. Physiological Review; 28, 304-30, 1948. LUO, X. D., BASILE, M. J., KENNELLY, E. J. Polyphenolic antioxidants from the fruits of Chrysophyllum cainito L. (star apple). Journal of Agricultural and Food Chemistry, 50, 1379–1382, 2002. MAECHLER, P., KENNEDY, E.D., POZZAN, T., WOLLHEIM, C.B. Mitochondrial activation directly triggers the exocytosis of insulin in permeabilized pancreatic b-cells. EMBO J. 16, 3833–3841, 1997. MAHLER, R. J., ADLER, M. L. Type 2 Diabetes Mellitus: Update on Diagnosis, Pathophysiology, and Treatment. The Journal of Clinical Endocrinology & Metabolismo, 84, 1168-1171, 1999.

MALAISSE, W.J., MALAISSE-LAGAE, F., SENER, A., PIPELEERS, D.G. Determinants of the selective toxicity of alloxan to the pancreatic B cell. Proc Natl Acad Sci USA, 79: 927-930, 1982. MALERBI D, FRANCO L. Multicenter study of the prevalence of diabetes mellitus and impaired glucose tolerance in the urban Brazilian population aged 30-69 yr. Diabetes Care;15, 1509-16, 1992. MARKHAM, K. R.; TERNAI, B.; STANLEY, R.; GEIGER, H.; MABRY, T. J. Carbon-13 NMR studies of flavonoids II. Naturally occurring glycosides and their acylated derivatives. Tetrahedron, 34, 1389-1397, 1978. MARLES, R. J, FARNSWORTH N. R. Antidiabetic plants and their active constituents. Phytomedicine, 2, 137-89, 1995. MATHIS, D., VENCE, L., BENOIST, C. β-Cell death during progression to diabetes. Nature, 414, 792-798, 2001. MATSCHINSKY, F. M. A lesson in metabolic regulation inspired by the glucokinase glucose sensor paradigm. Diabetes, 45, 223–241, 1996. MCCLAIN, D.A., CROOK, E.D., 1996. Hexosamines and insulin resistance. Diabetes 45, 1003–1009, 1995. MCCUNE, L.M., JOHNS, T. Antioxidant activity in medicinal plants associated with the symptoms of diabetes mellitus used by the Indigenous Peoples of North American boreal forest. Journal of Ethnopharmacology, 82, 197–205, 2002. MCDONAGH PF, HOKAMA JY. Microvascular perfusion and transport in the diabetic heart. Microcirculation, 7, 163–181, 2000. McLARTY, D. G., ATHAIDE, I., BOTTAZZO, G. I., SWAI, A.B.M. ALBERTI, K.G.M.M. Islet cell antibodies are not specifically associated with insulin dependent diabetes rural Tanzanian Africans. Diabetes Research and Clinical Practice, 9, 219-224, 1990. MILLER, N. J., DIPLOCK, A. T., RICE-EVANS, C., DAVIES, M. J., GOPINATHAN, V., MILNER, A. A novel method for measuring antioxidant capacity and its application to monitoring the antioxidant status in premature neonates. Clinical Science, 84: 407-412, 1993. MIURA, T.; ITOH, Y.; IWAMOTO, N.; KATO, M.; ISHIDA, T. Suppressive activity of the fruit of Momordica charantia with exercise on blood glucose in type 2 diabetic mice. Biological and Pharmaceutical Bulletin, 27, 248-250, 2004. MODESTO FILHO, J. Estudos a curto e a médio prazo da atividade Hipoglicemiante da Bumelia sartorium Mart. (quixaba) em diabéticos não insulino dependentes. Relatório Depto. Méd. Interna, Centro de Cienc. Saúde, Univ. Fed, Paraíba, 19, 1989.

MOGENSEN, C. E. Pharmacotherapy of Diabetes: New Developments. New York: Springer, 2007. MURATA, M., TAKAHASHI, A., SAITO, I., KAWANISHI, S. Site-specific DNA Methylation and Apoptosis: Induction by Diabetogenic Streptozotocin. Biochemical Pharmacology, 57: 881–887, 1999. MUSCHETTI, L. V.; MARTINO, V. S. Atividades biológicas dos flavonóides naturais. In: YUNES, R. A.; CECHINEL-FILHO, V. Química de produtos naturais, novos fármacos e moderna farmacognosia. 1 ed. Itajaí: Univali editora, 2007.
NATHAN, D.M., MEIGS, J., SINGER, D.E. The epidemiology of cardiovascular disease in type 2 diabetes mellitus: how sweet it is . . . or is it? Lancet, 350, SI4–SI9, 1997.
NISHIKAWA, T., EDELSTEIN, D., BROWNLEE, M. The missing link: a single unifying mechanism for diabetic complications. Kidney Intertional, 77, S26– 30, 2000. NISHIKAWA, T., EDELSTEIN, D., DU, X.L., YAMAGISHI, S.-I., MATSUMURA, T., KANEDA, Y., YOREK, M.A., BEEBE, D., OATES, P.J., HAMMES, H.-P., GIARDINO, I., BROWNLEE, M.,. Normalizing mitochondrial superoxide production blocks three pathways of hyperglycaemic damage. Nature, 404, 787–790, 2000a. NUKATSUKA, M., YOSHIMURA, Y., NISHIDA, M., KAWADA, J. Importance of the concentration of ATP in rat pancreatic beta cells in the mechanism of streptozotocin-induced cytotoxicity. Journal of Endocrinology, 127, 161-165, 1990. OKYAR, A.; CAN, A.; AKEV, N.; BAKTIR, G.; SUTLÜPINAR, N. Effect of Aloe vera leaves on blood glucose level in type I and type II diabetic rat models. Phytother. Res., v. 15, 157-161, 2001. OLEFSKY, J.M. Pathogenesis of non-insulin dependent diabetes (type 2). In: DEGROOT,L.J., BESSER, G.M., CAHILL, J.C., eds. Endocrinology, 2nd ed.Philadelphia: Saunders; 1369–1388, 1989. OLOFSSON, C.S., GOPEL, S.O. BARG, S., GALVANOVSKIS, J., MA, X., SALEHI, A., 25. RORSMAN P., ELIASSON, L. Fast insulin secretion reflects exocytosis of docked granules in mouse pancreatic B-cells. Pflugers Arch, 444, 43-51, 2002. OSOWSKI, S., ROSTOCK, M., BARTSCH, H.H., MASSING, U. Pharmaceutical comparability of different therapeutic Echinacea preparations. Forsch Komplementarmed Klass Naturheilkd, 7, 294-300, 2000. OWENS, D.R., ZINMAN, B., BOLLI, G.B. Insulins today and beyond. Lancet 358, 739-746, 2001. PAIXÃO, N., PERESTRELO, R., MARQUES, J. C., CÂMARA, J. S. Relationship between antioxidant capacity and total phenolic content of red, rosé and white wines. Food Chemistry, 105, 204–214, 2007.

PAPACCIO, G., PISANTI, F.A., FRASCATORE, S. Acetyl-homocysteine- thiolactone-induced increase of superoxide dismutase counteracts the effect of subdiabetogenic doses of streptozocin. Diabetes, 35, 470–474, 1986. PAYET, B.; SING, A.S.C.; SMADJA, J. Comparison of the concentrations of phenolic constituents in cane sugar manufacturing products with their antioxidant activities. Journal of Agricultural and Food Chemistry, 54 (19), 7270-7276, 2006. PEPATO, M. T.; KELLER, E. H.; BAVIERA, A. M.; KETTELHUT, I. C.;VENDRAMINI, R. C.; BRUNETTI, I. L. Antidiabetic activity of Bauhinia forticata decoction in streptozotocin-diabetic rats. Journal of Ethnopharmacology, 81, 191-197, 2002. PEREZ GUTIÉRREZ, R. M. Compuestos aislados de plantas com actividad antiinflamatoria, antiviral e hipoglicemiante. México: Instituto Politécnico Nacional, p. 139-185, 2002a.b. PEREZ GUTIÉRREZ, R. M.; ZAVALA, S. M. A.; PEREZ, G. S.; PEREZ, G. C. Antidiabetic effect of compounds isolated from plants. Phytomedicine, 5, 55-75, 1998. PHILLIPSON, G.W., ANDERSON, A.C. Journal of Ethnopharmacology, 25, 61, 1998. PIETROPAOLO, M., TRUCCO, M., CAKAN, N, et al. Autoantibodies to GAD65 and ICA512/IA-2 improve IDDM prediction in first-degree relatives with decreased first phase insulin response and ICA positivity [abstract]. Autoimmunity, 24(Suppl 1):51, 1996. PIETTA, P.G. Flavonoids as antioxidants. Journal of Natural Products, 63 (7), 1035-1042, 2000. PINTO, A.C., SILVA, D.H.S., BOLZANI, V.D., LOPES, N.P., EPIFANIO, R.D. Current status, challenges and trends on natural products in Brazil. Química Nova, 25, 45-61, 2002. PONOMARENKO, S. F.; PAVLOVA, M. E. Seed Structure in the Genus Bauhinia L. (fam. Fabaceae Lindl.) in the Context of Taxonomy. Biology Bulletin, 30 (4), 361–369, 2000. PRIOR, R. L., WU, X., SCHAICH, K. Standardized Methods for the Determination of Antioxidant Capacity and Phenolics in Foods and Dietary Supplements. Journal of Agricutural and Food Chemistry, 53, 4290-4302, 2005. RAMALHO, A. C. R. Insulina e hipoglicemiantes orais. In: Silva P. Farmacologia. 9 ed. Rio de Janeiro: GUANABARA:746-8, 1998. RANG, H. P., DALE, M. M., RITTER, J. M., MOORE, P. K. Farmacologia. 5 ed. Rio de Janeiro: Elsevier, 2004.

RATNAM, D. V.; ANKOLA, D. D.; BHARDWAJ, V.; SAHANA, D. K.; RAVI KUMAR, M. N. V. Review. Role of antioxidants in prophylaxis and therapy: a pharmaceutical perspective. Journal of Controlled Release, 113, 189-207, 2006. RE, R., PELLEGRINI, N., PROTEGGENTE, A., PANNALA, A., YANG, M., RICE-EVANS, C. Antioxidant activity applying an improved abts radical cation decolorization assay. Free Radical Biology and Medicine, 26, 1231–1237, 1999. REAVEN, G.M. The role of insulin resistance in human disease. Diabetes, 37, 1595–1607, 1998. RIBEIRO, A. B.; SILVA, D. H. S.; BOLZANI, V. S. Flavonóis glicosilados antioxidantes de Nectandra grandiflora (Lauraceae), Eclet. Quimica, 27, 34-44, 2002. RICE-EVANS C. A.; MILLER N.J.; BOLWELL P. G.; BRAMLEY P. M.; PRIDHAM J. B. The relative antioxidant activities of plant-derived polyphenolic flavonoids. Free Radical Research, 22, 375, 1995. RICE-EVANS, C.A.; MILLER, N.J.; PAGANGA, G. Free Radical Biology and Medicine, 20, 933-956, 1996. ROBBINS, M.J., SHARP, R.A., SLONIM, S.A., BURR, I.M. Protection against streptozotocin–induced diabetes by superoxide dismutase. Diabetologia, 18, 55–58, 1980. ROBERTSON, R P. Oxidative stress and impaired insulin secretion in type 2 diabetes Current Opinion in Pharmacology, 6, 615–619, 2006. ROBERTSON, R.P.Chronic oxidative stress as a central mechanism for glucose toxicity in pancreatic islet beta cells in diabetes. Journal of Biological Chemistry, 279, 42351–42354, 2004. ROCHA F. D., TEIXEIRA V. L., PEREIRA R. C., KAPLAN M. A. C. Diabetes mellitus e estresse oxidativo: produtos naturais como alvo de novos modelos terapêuticos. Revista Brasileira de Farmacognosia, 87(2), 49-54, 2006. RODRIGUES, A. G. Fitoterapia no Sistema Único de Saúde. Anais da V Jornada Catarinense e I Jornada Internacional de Plantas Medicinais. Joinville, 2006. p. 68-69. ROGINSKY, V., LISSI, E.A. Review of methods to determine chain-breaking antioxidant activity in food. Food Chemistry, 92 (2), 235-254, 2005. ROLO, A. P., PALMEIRA, C. M. Diabetes and mitochondrial function: Role of hyperglycemia and oxidative stress. Toxicology and Applied Pharmacology, 212, 167–178, 2006. RORSMAN, P. The pancreatic beta-cell as a fuel sensor: an electrophysiologist’s viewpoint. Diabetologia, 40, 487–495, 1997. RORSMAN, P., ELIASSON, L., RENSTRÖM, E., GROMADA, J., BARG, S., GÖPEL, S.: The cell physiology of biphasic insulin secretion. News Physiology Science, 15, 72–77, 2000.

RORSMAN, P.; RENSTROM, E. Insulin granule dynamics in pancreatic beta cells. Diabetologia, 46: 1029-1045, 2003. ROSENBLOOM, A.L., SCHATZ, D.A., KRISCHER, J.P, et al. Therapeutic controversy: prevention and treatment of diabetes in children. Journal of Clinical Endocrinology and Metabolism, 85, 494– 522, 2000. ROSSING, P., HOUGAARD, P., PARVING, H.H. Risk factors for development of incipient and overt diabetic nephropathy in type 1 diabetic patients: a 10-year prospective observational study. Diabetes Care, 25, 859– 64, 2002. SAID, O.; KHALIL, K.; FULDER, S.; AZAIZEH, H. Ethnopharmacological survey of medicinal herbs in Israel, the Golan Heights and the West Bank Region. Journal of Ethnopharmacology, 83, 251-265, 2002. SAKURAI, T., TSUCHIYA, S. Superoxide production from nonenzymatically glycated protein. FEBS letters, 236 (2), 406-410, 1988. SALTIEL, A.R., KAHN, C.R. Insulin signalling and the regulation of glucose and lipid metabolism. Nature, 414,799–806, 2001 . SAMUELSSON, G.; Acta Pharm. Fenn. 101, 33, 1992. SANCHEZ-MORENO, C. Review: Methods used to evaluate the free radical scavenging activity in foods and biological systems. Food Science and Technology International, 8, 121-137, 2002. SAXENA, A.; VIKRAM, N. K. Role of selected Indian plants in management of type 2 diabetes: a review. J. Altern. Complem. Med., 10, 369-378, 2004. SCHMELING, L. O.; GADOTTI, V. M.; LIZ, F. H.; MACHADO, C.; MEYRE-SILVA, C.; SANTOS, A. R. S.; CECHINEL FILHO, V. VI Seminário Integrado de Iniciação Científica, Camboriú, Brasil, 2000. SCHNEDL, W.J., FERBER, S., JOHNSON, J.H., NEWGARD, C.B. STZ transport and cytotoxicity. Specific enhancement in GLUT2-expressing cells. Diabetes, 43, 1326-1333, 1994. SENANAYAKE, G. V. K.; MARUYAMA, M.; SHIBUYA, K.; SAKONO, M.; FUKUDA, N.; MORISHITA, T.; YUKIZAKI, C.; KAWANO, M.; OHTA, H. The effects of bitter melon (Momordica Charantia) on serum and liver triglyceride levels in rats. Journal of Ethnopharmacology, 91, 257-262, 2004. SHARPE P.C., LIU W-H., YUE K.K.M., MCMASGTER D., CATHERWOOD M.A., MCGINTY A.M., TRIMBLE E.R.. Glucose-induced oxidative stress in vascular contractile cells: comparison of aortic smooth muscle cells and retinal pericytes. Diabetes, 47, 801–809, 1998. SILVA, F. R. M. B.; SZPOGANICZ, B.; PIZZOLATTI, M. G.; VIEIRA WILLRICH, M. A.; de SOUSA, E. Acute effect of Bauhinia forticata on serum glucose levels in

normal and alloxan-induced diabetic rats. Journal of Ethnopharmacology, 83, 33-37, 2002. SILVA, K.L.; CECHINEL-FILHO, V. Plantas do gênero Bauhinia: composição química e potencial farmacológico. Química Nova, 25 (3), 449-454, 2002. SILVA, S. R.; BUITRÓN, X.; OLIVEIRA, L. H. O.; MARTINS, M. V. Plantas Medicinais no Brasil: aspectos Gerais sobre legislação e comércio. Disponível em: http://www.traffic.org/publications/trafficc_portu.pdf. Acesso em 17 dez de 2006. SILVERTHORN, D. U. Human Physiology an Integrated Approach, 2. ed. – New Jersey: Prentice-Hall, Inc. A Pearson Education Company, 2003. SIMÕES, C.M.O.; SCHENKEL, E.P.; GOSMANN, G.; MELLO, J.C.P.; MENTZ, L.A.; PETROVICK, P.R. Farmacognosia: da planta ao medicamento. Editora da Universidade Federal de Santa Catarina, Florianópolis, 2001. SIXEL, P. J., PECINALLI, N. R. Características Farmacológicas Gerais Das Plantas Medicinais. Infarma, 74 (16), 13-14, 2005. STEINER, D.F., ROUILLE, Y., GONG, Q., MARTIN, S., CARROLL, R., CHAN, S. J.. The role of prohormone convertases in insulin biosynthesis: Evidence for inherited defects in their action in man and experimental animals. Diabetes Metabolism, 22, 94-104, 1996. STITT, A.W., FRIZZELL, N., THORPE, S.R. Advanced glycation and advanced lipoxidation: possible role in initiation and progression of diabetic retinopathy. Current Pharmaceutical Design, 10, 3349–3360, 2004. STRYER, L. Bioquímica. 4ª Ed.- Rio de Janeiro: Editora Guanabara Koogan, 1996. SZKUDELSKI, T. The Mechanism of Alloxan and Streptozotocin Action in β Cells of the Rat Pancreas. Physiol. Res. 50: 536-546, 2001. SZKUDELSKI, T., KANDULSKA, K., OKULICZ, M. Alloxan in vivo does not only exert deleterious effects on pancreatic B cells. Physiology Research, 47, 343-346, 1998. TESKE, M; TRENTINI, A. M. M. Compêndio de Fitoterapia; Herbarium Lab. Botânico Curitiba: Paraná, 1995. The Expert Committee on the Diagnosis and Classification of Diabetes Mellitus: Report of the Expert Committee on the Diagnosis and Classification of Diabetes Mellitus. Diabetes Care 20, 1183–1197,1997. The Expert Committee on the Diagnosis and Classification of Diabetes Mellitus: Follow-up report on the diagnosis of diabetes mellitus. Diabetes Care, 26, 160–3167, 2003.

THULESEN, J., ORSKOV, C., HOLST, J.J., POULSEN, S.S. Short-term insulin treatment prevents the diabetogenic action of streptozotocin in rats. Endocrinology, 138: 62-68, 1997. TIEDGE, M., LORTZ, S., DRINKGERN, J., LENZEN, S. Relation between antioxidant enzyme gene expression and antioxidative defense status of insulin-producing cells. Diabetes, 46: 1733-1742, 1997. TISCH, R., MCDEVITT, H. Insulin-dependent diabetes mellitus. Cell. 85, 291–297, 1996. TIWARI, A. K., RAO, J. M. Diabetes mellitus and multiple therapeutic approaches of phytochemicals: Present status and future prospects. Current Science, 83 (1), 30-38, 2002. TRUCCO, M. Regeneration of the pancreatic beta cell. The Journal of Clinical Investigation, 115, 5–12, 2005. TRUCCO, M. Stem cells and diabetes. Pediatric Diabetes, 5(Suppl 2):2–4, 2004. UCHIDA, K. Role of reactive aldehyde in cardiovascular diseases. Free Radical Biology & Medicine, 28 (12), 1685-1696. 2000. UCHIGATA, Y., YAMAMOTO, H., KAWAMURA, A., OKAMOTO, H. Protection by superoxide dismutase, catalase, and poly (ADPribose) synthetase inhibitors against alloxan–streptozotocininduced islet DNA strand breaks and against the inhibition of proinsulin synthesis. Journal of Biological Chemistry, 257, 6084–6088, 1982. ULRICH, P., CERAMI, A. Protein glycation, diabetes, and aging. Recent Progress and Hormone Research, 56, 1–21, 2001. UNGER, R.H., DOBBS, R.E., ORCI, L.: Insulin, glucagon and somatostatin secretion in the regulation of metabolism, Annual Review Physiology, 40, 307, 1978. VALENTÃO P., FERNANDES E., CARVALHO F., ANDRADE P. B., SEABRA R. M., BASTOS M. L., Biological and Pharmaceutical Bulletin 25, 1324—1327, 2002. VARDI, P., CRISA, L., JACKSON, R.A. Predictive value of intravenous glucose tolerance test insulin secretion less than or greater than the first percentile in islet cell antibody positive relatives of type 1 (insulin-dependent) diabetic patients. Diabetologia, 34, 93–102, 1991. VERGE, C.F., GIANANI, R., KAWASAKI, E, et al. Prediction of type I diabetes in first-degree relatives using a combination of insulin, GAD, and ICA512bdc/IA-2 autoantibodies. Diabetes, 45, 926–33, 1996. VIRGILI, F., SCACCINI, C., PACKER, L., RIMBACH, G. CARDIOVASCULAR Disease And Nutritional Phenolics. In: POKORNY, J., YANISHLIEVA, N., GORDON, M. (Eds.), Antioxidants in Food. Woodhead Publishing Ltda, Cambridge, pp. 87–99, 2001.

VOLPATO, G. T.; DAMASCENO, D. C.; CALDERON, I. M. P.; RUDGE, M. V. C. Revisão de plantas brasileiras com comprovado efeito hipoglicemiante no controle do Diabetes mellitus. Revista Brasileira Pl. Med., 4, 35-45, 2002. WAJCHENBERG, B. L. Disfunção endotelial na diabetes Tipo 2. Arquivos Brasileiros de Endecrinoligia e Metabologia, 46 (5), 514-519, 2002. WANG, H. X.; NG, T. B. Natural products with hypoglycemic, hypotensive, hypocholesterolemic, antistherosclerotic and antithrombotic activities. Life Science, 65, 2663-2677, 1999. WATKINS, P. J. ABC Of Diabetes,5. ed.- London: BMJ books, 2003. WAZLAWIK, E.; SILVA, M. A.; PETERS, R.R.; SIMÕES, C.M.O.; RIBEIRO-DO VALE, R.M. IX Reunião Anual da FSBE, Caxambú, Brasil, 1994. WEAVER, D.C., MCDANIEL, M., LACY, P.E: Alloxan uptake by isolated rat islets of Langerhans. Endocrinology, 102: 1847-1855, 1978a. WEST, E., SIMON, O.R., MORRISON, E.Y. Streptozotocin alters pancreatic beta-cell responsiveness to glucose within six hours of injection into rats. West Indian Medical Journal, 45, 60-62, 1996. WILLIAMS, A. D., BRAUMAN, J. I., NELSON, N. J., FLORY, P. J. Journal of the Americun Clinical Society, 30, 89, 1967. WILSON TANG, W.H., MAROO, A., YOUNG, J.B. Ischemic heart disease and congestive heart failure in diabetic patients. Medical Clinics of North America, 88, 1037–1061, xi–xii (Review), 2004. WOLFF, S. P.; JIANG, Z. Y. E HUNT, J. V. Protein glycation and oxidative stress in diabetes mellitus and ageing. Free Radical Biology & Medicine, (10), 339-352, 1991. WOOD, I.S., TRAYHURN, P. Glucose transporters (GLUT and SGLT): expanded families of sugar transport proteins. British Journal of Nutrition, 89, 3-9, 2003. WORLD HEALTH ORGANIZATION. Definition, Diagnosis e Classificacion of Diabetes Mellitus and its complications: Report of a WHO Consultation. Part 1: Diagnosis and Clasification of Diabetes mellitus. Geneva: World Health Organization, 1999. WORLD HEALTH ORGANIZATION. Regulatory situation of herbal medicines. A worldwide review, Geneva, 1998. WORLD HEALTH ORGANIZATION. Screening for Type 2 Diabetes: Report of a World Health Organization and International Diabetes Federation meeting. Geneva: World Health Organization, 2003. WORLD HEALTH ORGANIZATION. WHO. Monographs on Selected Medicinal Plants. Report of a WHO study group. WHO Technical Report Series, WHO, Geneva, 2, 211, 2002.

WU, G., HAYNES, T.E., LI, H., YAN, W., MEININGER, C.J. Glutamine metabolism to glucosamine is necessary for glutamine inhibition of endothelial nitric oxide synthesis. Biochemical Journal, 353, 245–252, 2001. YAMAMOTO, H., UCHIGATA, Y., OKAMOTO, H. DNA strand breaks in pancreatic islets by in vivo administration of alloxan or streptozotocin. Biochemical Biophysical Research Communications, 103, 1014–1020, 1981b. YAMAMOTO, H., UCHIGATA, Y., OKAMOTO, H. Streptozotocin and alloxan induce DNA strand breaks and poly (ADPribose) synthetase in pancreatic islets. Nature, 294, 284–286, 1981a. YIM, H. S.; KANG, S-O.; HAH, Y-C.; CHOCK, P.B. E YIM, M.B. Free radicals generated during the glycation reaction of amino acids by methylglyoxal. A model study of protein cross-linked free radicals. The Journal of Biological Chemsitry, 270 (24), 28228-28233. ZHANG, H., OLLINGER, K., BRUNK, U. Insulinoma cells in culture show pronounced sensitivity to alloxan-induced oxidadive stress. Diabetologia, 38, 635−641, 1995. ZHANG, H., ZDOLSEK, J.M., BRUNK, U.T: Alloxan cytotoxicity involves lysosomal damage. APMIS, 100, 309-316, 1992. ZHANG, Z., APSE, K., PANG, J., STANTON, R.C. High glucose inhibits glucose-6-phosphate dehydrogenase via cAMP in aortic endothelial cells. Journal of Biological Chemistry, 275, 40042–40047, 2000.

TRIAGEM FARMACOLÓGICA COMPORTAMENTAL NOME PLANTA:_______________________ DOSE:_____________PARTE USADA____________ VEÍCULO:________________ DATA:____/_____/_____ VIA DE ADMINISTRAÇÃO:____________ ESPÉCIE ANIMAL:_________________ SEXO: _________________ RESPONSÁVEL TÉCN.:______________________________________________
ATIVIDADE FARMACOLÓGICA
Quantificação dos efeitos (0) sem efeito, (-) efeito diminuído, (+) efeito aumentado,
(++) efeito intenso até 30` 1h 2h 3h 4h
1 – SNC a – Estimulante Hiperatividade Irritabilidade Agressividade Tremores Convulsões Piloereção Movimento intenso das vibrissas Outras_____________________ b – Depressora Hipnose Ptose palpebral Sedação Anestesia Ataxia Reflexo do endireitamento Catatonia Analgesia Resposta ao toque diminuído Perda do reflexo corneal Perda do reflexo auricular c – Outros comportamentos Ambulação Bocejo excessivo Limpeza Levantar Escalar Vocalizar Sacudir a cabeça Contorções abdominais Abdução das patas do trem posterior Pedalar Estereotipia 2 - SN AUTÔNOMO Diarréia Constipação Defecação Respiração forçada Lacrimejamento Micção Salivação Cianose Tono muscular Força para agarrar 3 – MORTE
Observações:__________________________________________________________________________________________________________________________________________________________________________________


Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas

Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo