evaluaciÓn de la resistencia genÉtica de … · evaluaciÓn de la resistencia genÉtica de...
TRANSCRIPT

EVALUACIÓN DE LA RESISTENCIA GENÉTICA DE ESPECIES DE Passiflora spp A Fusarium spp, AGENTE CAUSAL DE LA “SECADERA”
JENNIFER LONDOÑO ARAMBURO
UNIVERSIDAD NACIONAL DE COLOMBIA
FACULTAD DE CIENCIAS AGROPECUARIAS
COORDINACION GENERAL DE POSTGRADOS
PALMIRA
2012

EVALUACIÓN DE LA RESISTENCIA GENÉTICA DE ESPECIES DE Passiflora spp A Fusarium spp, AGENTE CAUSAL DE LA “SECADERA”
JENNIFER LONDOÑO ARAMBURO
Trabajo de grado para optar al título de:
MAGÍSTER EN CIENCIAS AGRARIAS ÉNFASIS PROTECCIÓN DE CULTIVOS
DIRIGIDO POR:
EYDER DANIEL GÓMEZ LÓPEZ
Ingeniero Agrónomo, Ph. D.
UNIVERSIDAD NACIONAL DE COLOMBIA
FACULTAD DE CIENCIAS AGROPECUARIAS
COORDINACION GENERAL DE POSTGRADOS
PALMIRA
2012


“La facultad y los jurados de la tesis de grado
no se harán responsables de las
ideas emitidas”
Articulo 24, resolución 04 de 1974

DEDICATORIA
Este trabajo está dedicado primeramente a Dios por permitirme alcanzar este
importante logro, al haberse constituido en mi permanente acompañante y guía,
fortaleciéndome siempre en los momentos cruciales en cada paso de mi vida. A
mis padres JORGE ENRIQUE y MARÍA EUGENIA, y mi hermano JULIO
ENRIQUE, quienes con su continua ayuda y apoyo fijaron en mi sendero de la
superación y consagración.
JENNIFER

AGRADECIMIENTOS
Quiero expresar mis más sinceros agradecimientos a una persona que admiro mucho, al Dr Eyder Daniel Gómez, por su dirección, paciencia, comprensión, apoyo permanente y formación profesional.
Al Dr. Edgar Iván Estrada, por su estimulo a continuar estudiando, por su apoyo incondicional y amistad.
Al Dr. Carlos Alberto Huertas, por su ayuda, generosidad y amistad. Un ejemplo a seguir.
A la I.A., M.Sc. Martha Liliana Bonilla, por la confianza que depositó en mí, y al acompañarme en cada una de las etapas de este trabajo.
Al I.A., M.Sc. Andrés Mauricio Posso, por abrirme las puertas en el laboratorio de Biología Molecular, por su enseñanza y generosa ayuda.
A la I.A., M.Sc. Carolina Cardozo, por su apoyo en el proceso de este trabajo y por brindarme buenos momentos durante este trayecto.
Al Ministerio de Agricultura y Desarrollo Rural, por contribuir con el apoyo económico de este trabajo.
A la Corporación Colombiana de Investigación Agropecuaria “CORPOICA” Palmira, por abrir las puertas para el desarrollo de este trabajo, y contribuir con el apoyo científico y logístico.
Al Dr. Jaime Eduardo Muñoz, por su asesoría en los análisis estadísticos.
A CENICAFE, por donar varias de las especies de Passifloras, utilizadas en la evaluación de la resistencia.
A la Universidad Nacional de Colombia sede Palmira, por la formación que me ha brindado.
A mis amigos Carlos Alberto Hoyos y Neidy Yalile Quiñonez, por colaborarme, apoyarme en gran parte de este trabajo y por su bonita amistad.
A todas las personas que de una u otra forma colaboraron, aportando su granito de arena para culminar este trabajo. A todos, Dios los bendiga.

I
CONTENIDO
Pág.
INTRODUCCIÓN 22
1. OBJETIVOS 25
1.1 OBJETIVO GENERAL 25
1.2 Objetivos específicos 25
2. MARCO TEÓRICO 26
2.1 La familia Passifloraceae 26
2.2 El género Passiflora 26
2.3 DESCRIPCIÓN TAXONÓMICA DEL MARACUYÁ 27
2.4 Passiflora edulis sims var flavicarpa Degner (Maracuyá) 28
2.4.1 Condiciones y requerimientos agroecológicos del Maracuyá 30
2.5 Importancia Económica 30
2.6 Enfermedades del Maracuyá 33
2.6.1 Muerte descendente en ramas 33
2.6.2 Enfermedades foliares: Mancha parda 33
2.6.3. Enfermedades en frutos 34
2.6.4. LA SECADERA DEL MARACUYÁ 34
2.7 Género Fusarium sp 36
2.7.1 Clasificación taxonómica del Género Fusarium sp 36
2.8 Alternativa de control de la secadera en maracuyá 37

II
3. MATERIALES Y MÉTODOS 39
3.1 LOCALIZACIÓN GEOGRÁFICA 39
3.2 Estandarización de una metodología eficiente de inoculación para hongos del género Fusarium spp agente causal de la secadera
39
3.2.1 Método de inmersión de raíces 39
3.2.2 Método de incisión de tallo 40
3.3 Evaluación de la respuesta de la inoculación del hongo Fusarium sp. a las diferentes especies de Passifloras
41
3.3.1Diseño experimental 41
3.4 Identificación morfológica y molecular de las especies del género Fusarium aislados de Passifloras con síntomas de secadera en el Valle del Cauca, Colombia.
44
3.4.1 Caracterización morfológica. 45
3.4.2 Caracterización molecular 46
3.4.3 EXTRACCIÓN DE ADN DE Fusarium 46
3.4.3.1 Preparación de los aislados para la extracción de
ADN 46
3.4.3.2 Procedimiento de la extracción de ADN 46
3.4.3.3 PCR para amplificación de la región ITS1 – ITS4 y
posterior secuenciación de los aislados del género
Fusarium que manifiestan síntomas de secadera en
plantas de maracuyá
48
3.4.3.4 Purificación y secuenciación del fragmento
amplificado de la región ITS1-ITS4 49

III
3.5 Cepas de referencias 49
3.6. Caracterización molecular de las especies de Passifloras seleccionadas para la evaluación de resistencia a Fusarium sp
50
3.6.1 Colecta del material vegetal 50
3.6.2 Material vegetal 52
3.6.3 Extracción de ADN 52
3.6.4 Evaluación de cantidad y calidad de ADN 52
3.6.5 Marcadores moleculares RAMs 53
3.6.6 Análisis de la información 54
4. RESULTADOS Y DISCUSIÓN 55
4.1 Estandarización de inoculación para hongos del género Fusarium
spp agente causal de la secadera 55
4.2 Evaluación de la respuesta de la inoculación del hongo Fusarium
sp. a las diferentes especies de Passifloras 56
4.3 Identificación morfológica y molecular de las especies del género
Fusarium aislados de Passifloras con síntomas de secadera en el
Valle del Cauca, Colombia
59
4.3.1 Caracterización morfológica de las especies encontradas
en este estudio. 59
4.3.2 Fusarium solani 60
4.3.3 Fusarium oxysporum 66
4.3.4 Fusarium verticillioides 71
4.3.5 Fusarium brachygibbosum 73

IV
4.4 Identificación de los aislados por secuenciación 75
4.5 Caracterización molecular de las especies de Passifloras
seleccionadas. 81
4.5.1 Extracción de ADN 81
4.5.2 Características de los cebadores 82
4.5.3 Análisis descriptivo 85
4.5.4 Heterocigosidad esperada (He) y porcentaje de loci
polimórficos 88
5. CONCLUSIONES 90
6. RECOMENDACIONES 92
BIBLIOGRAFÍA 93
ANEXOS 102

V
LISTA DE FIGURAS
Pág.
Figura 1. Proceso de inoculación del hongo Fusarium sp. a las especies de Passifloras
40
Figura 2. Proceso de inoculación del hongo Fusarium sp. a las especies de Passifloras, a través del método incisión de tallo
41
Figura 3. Evaluación del método de inmersión de raíces e incisión de tallo
55
Figura 4. Prueba de promedio Duncan P (0.005) 57
Figura 5. Evaluación de la resistencia de Passifloras resistentes al hongo del género Fusarium sp. agente causal de la secadera del maracuyá
58
Figura 6. Evaluación de la resistencia de especies de Passifloras susceptibles al hongo del género Fusarium sp. agente causal de la secadera del maracuyá
58
Figura 7. MUC1. A. Abundantes clamidosporas individuales y en parejas y B. microconidos
60
Figura 8. MUC1. Colonia blanca con micelio aéreo. 61
Figura 9. MUC2. A – C. microconidos B - D. clamidosporas 61
Figura 10. MUC 2. Colonia blanca con escaso micelio aéreo. 62
Figura 11. MUC 4. A, B - C. micro, meso y macroconidios 62
Figura 12. MUC 4. Colonia crema con escaso micelio aéreo. 63
Figura 13. MUC7. A - B. micro y macroconidios. C – D. Clamidosporas en cadena.
63
Figura 14. MUC7. Colonia blanca con micelio aéreo de estructura algodonosa.
64
Figura 15. MUC12. A. microconidios. C – D. Clamidosporas en cadenas e 64

VI
individuales
Figura 16. MUC12. Colonia blanca con escaso micelio aéreo. 65
Figura 17. MUC13. A. Clamidosporas individuales B. microconidios 65
Figura 18. MUC13. Colonia blanca con escaso micelio aéreo. 66
Figura 19. MUC3. A - B. clamidosporas en pareja e individuales, C. macro y microconidio
67
Figura 20. MUC3. Colonia violeta, escaso micelio aéreo. 67
Figura 21. MUC5. A - B. meso y microconidio 68
Figura 22. Colonia violeta, escaso micelio aéreo. 68
Figura 23. A - B. micro, meso y macroconidio C – D. Clamidosporas individuales y en cadena
69
Figura 24. MUC9. Colonia blanca-violeta, escaso micelio aéreo. 69
Figura 25. MUC10. A - B. abundantes clamidosporas en cadena, pareja e individuales. C - D micro, meso y macroconidio
70
Figura 26. MUC10. Colonia violeta, escaso micelio aéreo. 70
Figura 27. MUC6. A. Células agrandadas. B. microconidios 71
Figura 28. MUC6. Colonia blanca - violeta, con micelio aéreo. 72
Figura 29. MUC8. A. micro, meso y macroconidios. B. Células agrandadas
72
Figura 30. MUC8. Colonia blanca – rosado, micelio aéreo de estructura algodonosa.
73
Figura 31. MUC11. A – B. Estructuras desconocidas. C. microconidios 74
Figura 32. MUC11. Colonia blanca – violeta con escaso micelio aéreo. 75
Figura 33. Amplificación en gel agarosa de los hongos del género Fusarium
76
Figura 34. Árbol filogenético, la raíz es Acremonium, donde muestra la agrupación de las 13 especies de Fusarium sp. aislados del
78

VII
cultivo de maracuyá con síntomas de secadera, colectadas en seis municipios del Valle del Cauca y amplificado por PCR-ITS, con los iniciadores ITS 1 e ITS 4. Y cuatro cepas de referencias. 0.1 indica la distancia genética. El árbol filogenético se construyó con el progama Treecom. Los porcentajes se calcularon con 1000 réplicas
Figura 35. Árbol filogenético relacionando las secuencias ITS de los diferentes aislados de F. solani, mediante el programa Treecom versión 1.3b., se indica valores de “bootstrap” > 98%.
80
Figura 36. Árbol filogenético relacionando las secuencias ITS de los diferentes aislados de F. oxysporum, mediante el programa Treecom versión 1.3b., se indica valores de “bootstrap” > 96%.
80
Figura 37. ADN de las accesiones de las especies de Passiflloras, extraído mediante la metodología de Dellaporta et al (1983) con modificaciones
82
Figura 38. Heterocigocidad y porcentaje de loci polimórficos (95%) para cada uno de los cebadores RAMs utilizados
85
Figura 39.
Dendrograma de distancia genética de 28 introducciones de Passifloras, basado en el coeficiente de similitud de Dice Nei-Li (1978) y calculado con los datos combinados de los cuatro cebadores RAMs
87

VIII
LISTA DE TABLAS
Pág.
Tabla 1. Departamentos con mayor producción de maracuyá en Colombia.
32
Tabla 2. Escala de evaluación de daño en tallos y raíces en maracuyá propuesta por Rodríguez et al., 1991
43
Tabla 3. Aislamientos obtenidos de cultivo de maracuyá en el Valle del Cauca, presentando síntomas de marchitamiento (Secadera).
44
Tabla 4. Cepas de referencia. 50
Tabla 5. Introducciones y especies de Passifloras seleccionada para la evaluación de resistencia.
51
Tabla 6. Cebadores RAMs utilizados en la caracterización molecular. 53
Tabla 7. Análisis de Varianza 56
Tabla 8. Análisis del programa BLAST para comparar la máxima identidad de las 13 especie de Fusarium sp.
76
Tabla 9. Bandas totales y polimórficas obtenida con los cebadores RAMs utilizados para la evaluación de la diversidad genética.
83
Tabla 10. Valores de Heterocigosidad y porcentaje de lous polimórficos (95%) para los cebadores RAMs utilizados.
84

IX
LISTA DE ANEXOS
Pág.
ANEXO A. Medios de cultivos 102
ANEXO B. Tabla 1. Aislamiento de ADN genómico 103
ANEXO C. Tabla 2. Evaluación de la resistencia de 28 introducciones de 17 especies de Passifloras.
104
ANEXO D. Tabla 3. Secuencia de los 13 aislamientos de los hongos del género Fusarium
114

X
Abreviaturas empleadas ADN Ácido desoxirribonucleico
ANDr Ácido desoxirribonucleico ribosómico
ARN Ácido ribonucleico
Blast Basic Local Alignement Sequence Tool
CECT Colección Española de Cultivos Tipo
CLA Carnation Leaf Agar
Cm Centímetro
CTAB Bromuro de cetil-trimetil-amonio
dNTPs Desoxinucleotidos trifosfato
EDTA Ácido etilendiaminotetracético
F. Fusarium
g Gramo
gmol-1 Gramos mol
He Heterocigocidad esperada
ITS Región espaciadora intergénica
ITS Internal Transcribed Spacer Regions
Kb kilobases (1000 pb)
Kg Kilogramos
M Molar
mg Miligramo
mL Mililitro
mm Milímetros

XI
mM Milimolar
msnm Metro sobre nivel del mar
NCBI National Center For Biotecnology Information
ng nanogramo
°C Grados centígrados
PCR Polymerase Chain Reaction (reacción en cadena de la polimerasa)
PDA Agar Patata Dextrosa
P. Passiflora
RAMs Random Amplified Microsatellites
Rpm Revoluciones por minuto
Seg Segundos
sp. Especie
spp. Especies
TE Tris-EDTA
µg Microgramos
µL Microlitro
µm Micrómetro
U Unidades
UPGMA Unweighted pair-group method with arithmetic mean
V Voltios

XII
RESUMEN
El maracuyá amarillo Passiflora edulis Sims var flavicarpa Degener, representa un
renglón importante en la fruticultura colombiana, por su aceptación en el mercado
nacional y por su potencial de exportación.
El departamento del Valle del Cauca es considerado como el de mayor
participación en la producción nacional de esta fruta, a pesar que los cultivos
presentan alta incidencia de una enfermedad vascular conocida como secadera,
causado por el hongo Fusarium oxysporum f. sp. passiflorae.
Una solución a esta problematica es el uso de portainjertos resistentes para
controlar los hongos fitopatógenos del suelo y prevenir pérdidas en el maracuyá,
por esta razón se debe buscar especies del género Passiflora que sean
resistentes o tolerantes. Es así como Corpoica mantiene el banco de
germoplasma de Passifloras en el Centro de Investigación La Selva, algunas
especies con potencial comercial, que expresan una notoria variabilidad genética,
interespecífica e intraespecífica. De este banco se seleccionaron 28
introducciones pertenecientes a 17 especies de Passifloras para evaluar la
resistencia a Fusarium oxysporum f. sp. passiflorae causante de la secadera del
maracuyá. A estas introducciones se le realizó un estudio de diversidad genética
para determinar la distancia filogenética entre las especies seleccionadas.
Para la evaluación de las Passifloras seleccionadas, se probaron dos
metodologías para inocular: el método de incisión de tallo y el método de
inmersión de raíces. Se utilizó el aislamiento más patogénico (MUC5), obtenido en
estudios anteriores.
Se aislaron 13 hongos del género Fusarium, obtenidos a partir de tallo y raíz de
plantas afectadas con síntomas de secadera del maracuyá detectados en cinco
municipios del departamento del Valle del Cauca. Una vez obtenidos los aislados

XIII
se identificaron morfológica y molecularmente. En este estudio se determinó que
los 13 aislamientos pertenecen al género Fusarium; estos aislamientos se
agruparon en cuatro especies: F. verticillioides, Fusarium oxysporum, Fusarium
brachygibbosum, y Fusarium solan;, además se encontró que hay diferencias inter
e intraespecies.
Palabras clave: Fusarium, región ITS, variabilidad genética, Passifloras,
resistencia.

XIV
SUMMARY
The yellow passion fruit Passiflora edulis Sims var flavicarpa Degener, it’s an
important asset for Colombian fruit, for its acceptance in the domestic market and
export potential.
The department of Valle del Cauca is considered the largest share in domestic
production of this fruit, though the crops have high incidence of vascular disease,
known as dry wilt caused by the fungus Fusarium oxysporum f. sp. passiflorae.
A solution to this problem is the use of resistant rootstocks to control fungal
pathogens and prevent soil loss in the passion fruit, therefore species of Passiflora
that are resistant or tolerant should be sought. Thus Corpoica keeps Passifloras
genebank Research Center La Selva, including some species with commercial
potential, which express a remarkable genetic variability, interspecific and
intraspecific. From this bank 28 introductions species were selected belonging to
17 Passifloras to assess resistance to Fusarium oxysporum f. sp. passiflorae
causing passion fruit dry wilt. These introductions will be a study of genetic
diversity, to determine the phylogenetic distance between selected species.
To evaluate the selected Passifloras, two methods were tested to inoculate, cut the
stem and the roots immersion. Insulation was used most pathogenic (MUC5)
obtained in previous studies.
13 Fusarium fungi were isolated, they were obtained from the stem and the root of
plants affected with symptoms of dry wilt of passion fruit in five municipalities in the
department of Valle del Cauca. After obtaining the isolated they were identified
morphologically and molecularly. This study found that 13 isolated belong to the
genus Fusarium, these isolated were grouped in four species: F. verticillioides,
Fusarium oxysporum, Fusarium brachygibbosum, and Fusarium solani, it was also
found that inter and intraspecies differences.

XV
Keywords: Fusarium, ITS region, genetic variability, Passifloras, resistance.

‐ 22 ‐
INTRODUCCIÓN
El género Passiflora de la familia Passifloraceae está conformado por 450
especies, ubicadas en su mayoría en el trópico. Se destacan por su valor
económico, medicinal y ornamental. Solo algunas especies son cultivadas, la
mayor parte son silvestres cuyos frutos son consumidos localmente. Esta planta
se adapta a un sin número de hábitats. Colombia es el país con la mayor
diversidad de Passifloras en el mundo. Se registran 165 especies en la región
Andina por encima de los 1.800 msnm; se concentran las especies del subgénero
Tacsonia (curubas), y las especies del subgénero Passiflora (granadillas, badea,
chulupa) con una distribución desde el nivel del mar hasta los 2.400 msnm
aproximadamente (Escobar, 1991).
En Colombia, Brasil y Ecuador se cultiva dos especies de maracuyá: la de color
púrpura Passiflora edulis Sims y la amarilla P. edulis Sims var flavicarpa Degener,
siendo el segundo el más cultivado comercialmente. En Colombia el área cultivada
a nivel comercial está dominada por una sola especie: el maracuyá amarillo
Passiflora edulis Sims var flavicarpa Degener (García, 2002).
En el Valle del Cauca, el maracuyá Passiflora edulis Sims var flavicarpa Degener,
es uno de los frutales con un gran potencial agrícola desde el punto de vista
económico por su aceptación en los mercados nacionales e internacionales, los
altos rendimientos en el área sembrada y por la generación de empleo.
En el cultivo de maracuyá se han venido incrementando diversos problemas
fitosanitarios que limitan su producción, razón por la cual su siembra se ha
desestimulado (González et al., 2002). Una de las más importantes, es la
secadera del maracuyá según algunos autores es causada por hongos del género
Fusarium de difícil control por los sistemas convencionales, disminuyendo los

‐ 23 ‐
rendimientos en forma considerable e incrementando los costos de producción,
motivo por el cual, se considera necesario incrementar los conocimientos con
relación al cultivo con miras a buscar alternativas de manejo que mejoren su
sostenibilidad. El Valle del Cauca es considerado por la encuesta nacional
agropecuaria 2006 (CCI, 2007), uno de los departamentos con mayor participación
en la producción nacional de esta fruta (31%); no obstante debido a su alta
susceptibilidad los cultivos presentan alta incidencia de secadera. (Proyecto
Corpoica Palmira, 2011).
En Colombia existen registros de que esta enfermedad ocasionó en el norte del
Valle del Cauca, entre el 30 y el 100% de muerte de las plantas en el periodo de
1997-1999 (Torres et al., 2000). Actualmente su incidencia está entre un 10 y
40%; es de resaltar que los cultivos comerciales en este momento alcanzan una
vida útil de 15 meses aproximadamente.
Una alternativa para el control de esta enfermedad es la utilización de variedades
o genotipos que ofrezcan resistencia genética a este patógeno del suelo, los
cuales deben conservar las características agronómicas y de calidad exigida por el
mercado nacional e internacional.
Según lo reportado por Meletti et al., (2005) en estudios de evaluacion de la
resistencia de especies de Passifloras, describen que han sido identificada
algunas especies con resistencia a Fusarium oxysporum como Passiflora giberti,
P. alata y P. caerulea.
Teniendo en cuenta que las especies silvestres pueden presentar incompatibilidad
con los brotes a injertar dado que sus tallos son muy finos, se ha estandarizado
otra alternativa de manejo, el método de injertación. El método de injerto
hipocotiledonal, que consiste en germinar las semillas del patrón y la semilla del
portainjerto; cuando estas tienen dos hojas verdaderas se corta el patrón y se
hace una hendidura en el centro del tallo y sobre este se injerta la copa cortando

‐ 24 ‐
por debajo del los cotiledones (Nogueira et al., 2005), el método de injerto de cuña
o hendidura y el método del injerto de empalme (Gardner 1987).
Con el presente trabajo, se evaluó la resistencia genética de 28 introducciones,
pertenecientes a 17 especies de Passiflora spp a Fusarium spp, agente causal de
la “secadera”.

‐ 25 ‐
1. OBJETIVOS
1.1 OBJETIVO GENERAL
Evaluar bajo condiciones controladas la resistencia de 28 introducciones de
Passifloras spp a Fusarium spp agente causal de la secadera.
1.2 Objetivos específicos
Estandarizar una metodología eficiente de inoculación para hongos del
género Fusarium spp agente causal de la secadera.
Evaluación de la respuesta de la inoculación del hongo Fusarium sp. a las
diferentes especies de Passifloras.
Identificar morfológica y molecularmente las especies del género Fusarium
aislados de Passifloras con síntomas de secadera en el Valle del Cauca
COLOMBIA.
Caracterizar molecularmente las 28 introducciones de Passifloras
seleccionadas para la evaluación de resistencia a Fusarium sp.

‐ 26 ‐
2. MARCO TEÓRICO
El nombre de Passiflora fue dado por Pluckenet en 1696, el cual se deriva del latín
“Flor Passionis” flor de la pasión (Escobar, 1988; Vecchia, 2000). Inicialmente, fue
la flor que atrajo la atención tanto de expertos botánicos como de coleccionistas
por su gran belleza, sin embargo los frutos poco a poco se fueron consolidando
como alimentos muy apetecidos por su valor nutricional y medicinal tanto en
Europa como en América del Norte (Ijjász, 1999).
2.1 La familia Passifloraceae
Es una familia de plantas de tamaño medio, las cuales pueden encontrarse en
forma de lianas o enredaderas herbáceas con zarcillos axilares, hasta arboles
bajos y arbustos. Su descripción taxonómica corresponde a que presentan hojas
siempre alternas, pecioladas con estípulas pequeñas o a veces caedizas, en
general ovadas o bilobuladas, muy a menudo con nectarios en el haz, cáliz floral
en forma de copa; fruto en forma de baya globosa, con semillas, de testa dura y
endospermo carnoso. (Killip, 1938; Uribe, 1972; Cronquist, 1978; Heywood, 1985;
Escobar, 1988).
2.2 El género Passiflora.
El género Passiflora, pertenece a la familia Passifloraceae, se distribuye en
regiones tropicales y subtropicales desde el nivel del mar hasta altitudes
superiores a 3000 m.s.n.m., pero la mayor riqueza en especies se encuentra en
las regiones moderadamente cálidas y templadas, entre 400 y 2000 m.s.n.m. Los
siguientes autores hacen una revisión taxonómica de este género donde Feuillet y
McDougal, 2003; Ulmer y McDougal, 2004, redujo de 22 (Killip, 1938) a cuatro los

‐ 27 ‐
subgéneros: Astrophea (57 especies), Deidamioides (13 especies), Decaloba (214
especies) y Passiflora (236 especies).
Cerca del 90% de las 520 especies de Passiflora son originarias en América
(Escobar, 1988). En Colombia se han registrado 165 especies (Ocampo et al.,
2007), 58 de ellas se han considerado como endémicas. Sin embargo, varias
especies andinas están amenazadas y algunas ya se consideran extintas. La
elevada diversidad de la zona cafetera colombiana, hogar del 59% de las especies
de Passifloraceae, ha sido afectada por la intensa fragmentación de las selvas
andinas (Kattan, 1997).
En Colombia las Passifloras comerciales como el maracuyá (Passiflora edulis
Sims var flavicarpa Degener), la granadilla (Passiflora ligularis) representan un
renglón relevante en la fruticultura. En el mercado de las frutas, Colombia se
encuentra dentro de los tres primeros productores y exportadores de Maracuyá de
Suramérica, teniendo en cuenta además que esta provee el 90% de la producción
a nivel mundial (CCI, 2007).
2.3 DESCRIPCIÓN TAXONÓMICA DEL MARACUYÁ
• División: Espermatofita
• Clase: Dicotiledonea
• Orden: Parietales
• Familia: Passifloraceae
• Género: Passiflora
• Subgénero: Passiflora
• Especie: Passiflora edulis Sims var flavicarpa Degener
• Nombre común Maracuyá, parchita,
Fuente: Kilip, 1938 y Cronquist, 1978; Escobar y Forero, 1993.

‐ 28 ‐
2.4 Passiflora edulis sims var flavicarpa Degner (Maracuyá)
El maracuyá, es una planta de la región del Amazonas. Este es considerado el
centro de origen de unas 150-200 especies del género Passiflora (García, 2002).
Esta planta cuenta con 550 especies que en estado silvestre se desarrollan en Sur
América, Asia y en el Pacífico Sur (Agostini, et al. 2002). De las especies del
género Passiflora se destacan entre 50 a 60 especies que producen frutos
comestibles, de interés comercial (Araújo, et al., 2002) de las cuales en Colombia,
prácticamente el área cultivada está dominada por una sola especie, el maracuyá
amarillo Passiflora edulis Sims var flavicarpa Degener, la cual según García,
(2002) es una mutación del maracuyá morado Passiflora edulis. El rango óptimo
de altitud para siembra, oscila entre los 900 y 1500 msnm, con temperaturas
promedio de 24ºC y mínimo 5 horas/luz/día, requerimientos de agua entre 800-
1500 mm/año, además de riego en la época seca. Tiene fácil adaptación a
diferentes tipos de suelos si estos son profundos, fértiles, con pH cercano a la
neutralidad y buen drenaje interno y externo.
Se puede propagar por semilla, estaca, acodo e injerto; comercialmente se hace
por semilla, la cual, previa elección se siembra en germinadores en donde dura 30
días, luego se lleva a almácigos por 30 días, transcurridos los cuales se traslada a
los sitios definitivos en el campo. Desde que se traslada a campo hasta la floración
transcurren 180 días. Presentan tres cosechas grandes, cada una de dos meses,
intercaladas con dos “mitacas o traviesas”, de cuatro meses cada una. Las
cosechas coinciden con los periodos de verano en la zona; los periodos lluviosos
inducen las floraciones (Paredes, 1996).
La fruta puede alcanzar un peso promedio mínimo de 130 g, el rendimiento de
jugo puede alcanzar el 36% y el contenido de ácidos solubles entre 13 - 18 º Brix,
y se utiliza directamente en refrescos, o ser industrializada para la elaboración de
cremas alimenticias, dulces cristalizados, sorbetes, licores, confites, néctares,
jaleas, refrescos y concentrados. La cáscara es utilizada en Brasil para preparar

‐ 29 ‐
raciones alimenticias de ganado bovino, pues es rica en aminoácidos, proteínas,
carbohidratos y pectina. Este último elemento hace que se emplee en la industria
de la confitería para darle consistencia a jaleas y gelatinas (García, 2002).
Se considera la maracuyá, una fruta de especial importancia a nivel industrial no
solo por su sabor intenso, alta acidez, agradable aroma y elevado contenido de
vitamina C (Knigth & Sauls, 2005), sino también por las propiedades medicinales
de su follaje y frutos, atribuidas a la presencia de alcaloides (Gupta, 1995) por lo
que son utilizadas en América y algunos países Europeos en la medicina
tradicional. En el follaje del maracuyá se encuentran alcaloides indólicos como
harmana, harmina y harmol y compuestos como la Passiflorina, que le confieren
efecto tranquilizante, antiespasmódico, anticonvulsivo y regulador de la presión
arterial, además, es usado en la elaboración de perfumes y esencias (Gupta,
1995). La semilla contiene un 20-25 % de aceite, que según el Instituto de
Tecnología y Alimentos de Brasil se puede usar en la fabricación de aceites, tintas
y barnices. Este aceite puede ser refinado para otros fines como el alimenticio, ya
que su calidad se asemeja al de la semilla de algodón en cuanto a valor
alimenticio y a la digestibilidad; además contiene un 10% de proteína. Otro
subproducto que se extrae es la maracuyina, un tranquilizante muy apreciado en
Brasil (García, 2002).

‐ 30 ‐
2.4.1 Condiciones y requerimientos agroecológicos del Maracuyá
CONDICIONES AGROECOLÓGICAS PLAGAS Y ENFERMEDADES
ALTITUD m.s.n.m <1200 PLAGAS Gusano cosechero
(Agroulis sp). Arañita roja (Tetraichus sp). Mosca sonsa (Dasiops sp). Encrespador cogollo (Trips
sp). Lorito verde (Diabrótica sp). Tortuguilla (Ceroplastes
sp).
RADIACIÓN H/día >4 TEMPERATURA ºC 20-30 PRECIPITACIÓN mm 1000-2000
HUMEDAD % PENDIENTE %
ZONA DE VIDA
Bosque espinoso subtropical Bosque muy seco tropical.
Bosque seco tropical.
NIVEL DE NUTRIENTES DEL
SUELO.
N Kg/ha 150 ENFERMEDADES Mancha parda
(Alternaria sp) Pudrición del fruto
(Phytophthora nicotianae var. Parasíticas)
Mancha por septoria (Septoria passiflorae.
Roña (Cladosporium herbarum).
Secadera (Fusarium sp.) Antracnosis
(Colletrotrichum gleosporoides penzingi)
P2O5 Kg/ha 45 K2O Kg/ha 160 PH Kg/ha 5.5-6.5
PROFUNDIDAD cm 50
TEXTURA Clase Franca, Franco-arenoso, franco-
arcilloso
DISTANCIA DE SIEMBRA 3*3, 3*4, 2.5*6
DENSIDAD DE SIEMBRA (Plantas/Ha) 1111, 833, 666
VIDA UTIL ±17 meses
COSECHA: inicia de los 8 después de la siembra y es continua durante mínimo 14 meses.
LABORES CULTURALES DESYERBAS: 4 por año
incluida 1 aplicación de herbicidas.
PODAS: de formación. TUTORADO: espaldera o
emparrado. Control sanitario: insectos
y hongos.
Fuente: fundación sonrisas de colores (2004). Secretaria agricultura del Huila. Acuerdo de productividad codena frutícola. 2006
2.5 Importancia Económica
En Colombia el cultivo comercial se inició en los años 60, por medio de la
importación de plantas procedentes de Hawaii, Brasil y Venezuela (Chacon, 1988).
Solo hasta los 80 se lanzó al mercado internacional (como jugo y concentrado),

‐ 31 ‐
contribuyendo al desarrollo del cultivo en zonas del Valle del Cauca y la zona
marginal baja cafetera de los departamentos de Caldas, Quindío, Risaralda y
Tolima, así como Huila. La superficie dedicada varía entre 2.500 y 7.000 hectáreas
y el 70% de la producción se exporta, dejando el 30% para el mercado interno. El
rendimiento medio alcanza las 20 t/ha, y su costo medio de producción, es de US$
180/t. Colombia participa en el mercado mundial de manera variable; en 1993
aportó del 60 al 70%, aunque en el 1994 contribuyó sólo el 7.3%. En Colombia, el
ICA promovió ampliamente el cultivo al tiempo que se dio un fuerte soporte técnico
y créditos accesibles a los productores por parte de la Caja de Crédito Agrario y de
la Federación Nacional de Cafeteros. En la actualidad, el maracuyá, representa un
reglón importante en la fruticultura, por su aceptación en el mercado nacional y por
su potencial de exportación. (UDEA, 2008). De acuerdo con las estadísticas del
2005, la producción mundial de maracuyá fue de 640.000 toneladas, la cual el
70% de la producción fue Brasil seguido por Ecuador y Colombia. Para el 2007,
Colombia ocupo el segundo lugar en producción a nivel mundial, los frutos
presentan un sabor particular intenso y una alta acidez, es buena fuente de
proteínas, minerales, carbohidratos, grasas, vitamina A y niacina, constituyéndose
en una base importante para la elaboración de bebidas industrializadas (Carvajal,
2007 y Gómez, et al., 1995), el cual es exportado de Colombia hacia Puerto rico y
Estados Unidos principalmente, con participaciones del 35% y 40 %
respectivamente, con un valor total de US$1'862.956. También se exportan
mezclas de jugos que incluyen el maracuyá, cuyo principal destino es Puerto Rico,
país al cual se destina el 84% de la producción nacional con un valor aproximado
de US$ 678.244 (MADR, 2006 & CCI, 2007). La producción de Maracuyá en
Colombia es de cerca de 80.000 Tn/año, en un área de 4800 hectáreas. El Huila y
Valle del Cauca, son los departamentos con mayor área y volumen en producción,
este valor corresponde al 22.2 Y 24.3% de la producción de frutas a nivel nacional.
Los municipios con mayor producción en el país se muestran en la tabla 1.

‐ 32 ‐
Tabla 1. Departamentos con mayor producción de maracuyá en Colombia.
Departamento Participación (%) Departamento Participación (%)
Valle del Cauca 24.3 Córdoba 10.3
Huila 22.2 Santander 6.4
Meta 14.0 Otros 22.8
Fuente: MADR. 2008
Fuente: ministerio de agricultura y desarrollo rural, 2008
De acuerdo con el Anuario estadístico (MADR, 2009), en Colombia se sembraron
en el 2008, 6372 ha, con una producción de 109.486 ton y un rendimiento
promedio de 17.182 ton/ha. (CCI, 2007).

‐ 33 ‐
2.6 Enfermedades del Maracuyá
En 1982, Bedoya y Medina encontraron que el cultivo difícilmente alcanzaba los 3
años como consecuencia del ataque de fitoparasitos, especialmente por la
presencia en las zonas donde su siembra era intensiva. En 1997, el periodo de
producción en la Unión (Valle) se había reducido a 17 meses, a pesar del intenso
programa de manejo del problema. Esto es debido al uso indiscriminado de
agroquímicos, los cuales han originado una disminución de la población de
insectos polinizadores y por ende afectando el ciclo normal del maracuyá. Esta
especie de uso comercial como muchas otras, se encuentra expuesta al ataque de
diversas enfermedades, las cuales son causadas por bacterias, hongos,
nematodos y virus; algunos de ellos causan pérdidas considerables en la
producción.
2.6.1 Muerte descendente en ramas, enfermedad de la parte aérea de la planta,
causado por el hongo del género Colletotrichum sp., su sintomatología se
caracteriza por necrosis total o parcial de brotes y zarcillos el cual se extiende
rápidamente por las ramas en forma descendente; puede formar chancros de color
castaño.
2.6.2 Enfermedades foliares: Mancha parda: causada por un hongo del género
Alternaria sp, su sintomatología se da en las hojas, las cuales presentan manchas
redondeadas de color castaño uniforme y halo clorótico. Bacteriosis: causada por
la bacteria Xanthomonas campestris pv passiflorae, su sintomatología se observan
en las hojas, presentando manchas irregulares, acuosas, que se inician
generalmente en el borde de ellas.

‐ 34 ‐
2.6.3. Enfermedades en frutos: Mancha aceitosa: causada por el Oomiceto
Phytophthora su sintomatología comienza, cuando en el fruto aparecen manchas
muy pequeñas (< a 1mm), se extienden y se une a otras hasta formar una gran
mancha castaño-rojizo con bordes de apariencia aceitosa. Roña: causado por el
hongo del género Colletotrichum sp, su sintomatología se caracteriza por
presentar lesiones circulares, pequeñas y de color café, las cuales al avanzar, se
tornan corchosas, que se distribuyen por todo el fruto. Chancro del fruto o
antracnosis: causado por el hongo del género Colletotrichum sp, su
sintomatología se da en frutos que han iniciado la maduración, se observan
pequeñas manchas de color café claro, las cuales crecen lentamente, forman
chancros, reblandecen el tejido y en estados avanzados, deterioran
completamente el endocarpio. (Torres et al., 1999).
El cultivo del Maracuyá, no solo se ve afectado en frutos y aéreas foliares,
también ocurre afecciones en la raíz, los cuales dan origen a pudriciones en los
tallos y ramas, causan muerte de tejidos y defoliaciones. La enfermedad de raíz
más importante es la secadera del maracuyá.
2.6.4 LA SECADERA DEL MARACUYÁ
Esta enfermedad es conocida como pudrición seca, marchitez, fusariosis o
secadera (James, 2000; Espinoza y Mendoza, 2001). Como principal agente
causal de la secadera se ha registrado a Fusarium oxysporum f. sp. passiflorae
(Díaz, 1998; Rico et al., 2001; Gómez, 2008), se caracteriza por penetrar las
diferentes capas de la corteza de la raíz hasta alcanzar el sistema vascular. La
muerte de la planta se debe a la impermeabilización y taponamiento de los haces
vasculares inducido por Fusarium (Díaz et al., 1998). Una vez establecido, la
colonización, el hongo es transportado rápidamente a través del xilema,
desarrollando la sintomatología característica de marchitez, esta sintomatología se

‐ 35 ‐
presentan desde semillero, en plántulas estrangula el hipocotilo ocasionando
volcamiento y muerte (Torres et al., 2000). Cuando se presenta su ataque en
campo, la sintomatología se caracteriza por marchitez de toda la planta, clorosis
en las hojas, seguidas de necrosis y fuerte defoliación. (Roncero et al., 2000; Rico
et al., 2001; Agrios, 2005 La enfermedad presenta una distribución en focos y en
casos extremos puede llegar a producir la muerte del 90 a 100% de una plantación
en estados tempranos en campo (Torres et. al, 2000).
Rodríguez, 1997, citando a diversos autores, los cuales sugieren en sus estudios
presencia de un complejo fungoso, compuesto por Fusarium, se encontrarían
Phytophthora, Rhizoctonia y posiblemente Phytium.
En Colombia existen registros de que esta enfermedad ocasionó, en el norte del
Valle del Cauca, entre el 90 y el 100% de muerte a plantas en el periodo de 1997-
1999 (Torres et al., 2000).
La diseminación de este patógeno se relaciona con el material vegetal de
propagación porque la selección de semillas no es la adecuada y no cumple con
los estandares mínimos de calidad, lo que lleva a una alta infección de
microorganismos fungosos, uno de ellos Fusarium. También inciden factores como
el agua de riego, las lluvias fuertes o el viento, que son considerados agentes de
diseminación de conidios y estructuras reproductivas del hongo. (Alexopoulos y
Mins, 1985 y Agrios, 2005). Esta enfermedad se ve favorecida por el mal drenaje,
bajo contenido de materia orgánica de los suelos y periodos lluviosos continuos
(Bedoya y Medina, 1982; Torres et al., 2000 y Sánchez de P., 2003).
Fusarium es un hongo cosmopolita que afecta muchas especies vegetales y es
difícil de controlar. Existe una gran diversidad de razas o formas especiales de
Fusarium spp., de las cuales algunas son patogénicas y su gran mayoría son
saprofitas, pueden ser utilizadas como controladores biológicos. Según Cook y
Baker (1989) existen más de 76 cepas de Fusarium oxysporum reportadas que

‐ 36 ‐
causan enfermedad. La mayoría de los hongos del género Fusarium están
relacionados con enfermedades del sistema vascular en plantas.
2.7 Género Fusarium sp.
Especies pertenecientes a este género presentan macroconidios fusoides,
ligeramente curvados, septados y con una célula basal pedicelada. Con
microconidios y clamidosporas terminales o intercalares, que pueden estar
ausentes o presentes (Nelson et al., 1994).
El género Fusarium comprenden 70 especies descritas, que a su vez están
agrupadas en 12 secciones. Cada sección es en realidad un conjunto de especies
relacionadas entre sí. Más de la mitad de las especies son parásitos de plantas y
entre ellos se encuentran algunos de los más serios patógenos del mundo agrícola
(Leslie and Summerell, 2006).
2.7.1 Clasificación taxonómica del Género Fusarium sp.
División: Ascomycota
Subdivisión: Pezizomycotina
Clase: Sordariomycetes
Subclase: Hypocreomycetidae
Orden: Hypocreales
Familia: Hypocreaceae
Género: Fusarium
Especies: F. oxysporum, F. solani, F. verticillioides, F. brachygibbosum, etc.

‐ 37 ‐
Algunas especies tienen un estado sexual reconocido dentro de la familia
Nectriaceae y pertenecen al género Gibberella o Nectria.
Fuente: De Hoog et al., 2000 y Leslies and Summerell, 2006.
2.8 Alternativa de control de la secadera en maracuyá
Está comprobado que el uso indiscriminado e intensivo de agroquímicos en los
cultivos, crean resistencia de los patógenos a ellos, además ocasionan deterioro
ambiental, incrementos en los costos de producción y efectos negativos en la
calidad de los productos los cuales afectan la salud humana y el ambiente.
(Sánchez, 2003).
En este sentido, en el tercer milenio, la competitividad se dará en términos de
precios sociales y ambientales, además de los propios a la actividad productiva.
La mejor alternativa para el control seria la utilización de variedades o genotipos
que ofrezcan resistencia a estos patógenos del suelo y conserven las
características agronómicas y de calidad exigida por el mercado nacional e
internacional.
El uso de cultivares resistentes, asociado al manejo integrado de enfermedades,
es la medida más eficaz, económica y ecológica de control (Vilela et al., 2002). Sin
embargo, aún no existen cultivares de maracuyá amarillo que cumplan con esta
característica, lo que es uno de los grandes desafíos para el mejoramiento
genético. Una solución, a corto plazo identificada por varios investigadores como
Sharma et al., (2002), sería el uso de portainjertos resistentes para controlar los
hongos del suelo y prevenir pérdidas en el maracuyá, para lo cual se debe buscar
en especies del género Passiflora que sean resistentes o tolerantes.
En Venezuela y Brasil, se han realizado estudios sobre patrones para el manejo
de la secadera del maracuya, no obstante en Colombia los avances en este tema

‐ 38 ‐
no han sido significativos, afortunadamente se cuenta con una colección nacional
de germoplasma de Passifloras en Corpoica C.I La Selva, las cuales se puede
utiizar como nueva alternativa de manejo de la secadera, enfermedad limitante en
el cultivo de maracuyá amarillo, mediante la utilización de portainjertos resistentes,
a los principales patógenos de origen fúngico, contribuyendo al incremento en la
rentabilidad del cultivo. (Proyecto Corpoica Palmira, 2011).
Varios autores, entre los cuales se destaca Ferreira & Oliveira, 1991, describen
una amplia variabilidad genética existente en el género Passiflora. Tamayo, 1999,
destaca que, gran parte de esta variabilidad genética se encuentra en Colombia la
cual puede ser utilizada en la búsqueda de especies tolerantes a enfermedades.
Meletti et al., (2005) describe que ya han sido identificada algunas especies con
resistencia a Fusarium oxysporum como Passiflora giberti, P. alata y P. caerulea,
además esta última, mostró resistencia a Phytophthora sp. Así mismo Tadeu, et
al., 2002, reportó que se ha encontrado resistencia a la muerte precoz por
Fusarium en P. nitida, P. Laurifolia y algunas accesiones de P. suberosa, P.
coccinea y P. setacea. Estas resistencias no solo son importantes para el uso de
estos materiales como portainjertos, sino también para su uso posterior dentro de
los programas de mejoramiento genético, dado que se ha encontrado en estudios
realizados en Brasil de compatibilidad genética, entre especies silvestres de
Passiflora y las comerciales (Lage, et al. 2004).

‐ 39 ‐
3. MATERIALES Y MÉTODOS
3.1 LOCALIZACIÓN GEOGRÁFICA
La presente investigación se realizó en el laboratorio de fitopatología y en el
invernadero de CORPOICA Palmira; y el laboratorio de biología molecular de la
Universidad Nacional de Colombia, municipio de Palmira; Valle del Cauca (1050
msnm; 23º C y 70% de Humedad Relativa; 3º 30’ 45.6’’N y 76º 18’ 29.91’’W).
3.2 Estandarización de una metodología eficiente de inoculación para hongos del género Fusarium spp agente causal de la secadera.
Se establecieron dos pruebas de patogenicidad. Se inocularon plantas de
maracuyá (P. flavicarpa) y badea (P. quadrangularis) obtenidas a partir de semilla
y sembradas en sustrato estéril y conservadas en condiciones controladas. Se
seleccionaron 5 plántulas por método y se evaluó el aislado con mayor virulencia
encontrados en las pruebas previas a este estudio de resistencia.
3.2.1 Método de inmersión de raíces: A partir de un cultivo monospórico del
aislado del hongo mas patogénico (Fusarium sp), se le adiciona agua destilada
estéril, se recoge cuidadosamente con una espátula, después la solución se filtra a
través de un embudo, el cual se le coloca una gasa estéril, con el fin de recolectar
la solución conidial sin residuos de PDA. De esta solución, se toma 1µl, mezclado
con azul de algodón y se coloca en la cámara de Neubauer para realizar el conteo
de esporas. Después de obtener el número de esporas, se procede a preparar la
solución inoculo a una concentración de 1* 106 UFC. La concentración de esporas
fue calculada mediante la siguiente fórmula:

‐ 40 ‐
Concentración de esporas = N/5* 1 /0.004
Donde, N = Número de esporas contadas en un cuadrante.
Luego, para realizar la poda de las raíces se sacan las plántulas cuidadosamente
del vaso donde están sembradas, se hace un lavado de las raíces con el fin de
limpiarles el sustrato (turba) y luego con unas tijeras estériles se cortan las puntas
de las raíces. Posteriormente, las plántulas se sumergen en los recipientes que
contienen el inoculo durante 24 horas. Finalizado este proceso, las plántulas se se
sembraran en bolsas con suelo estéril. (Figura 1).
Figura 1. Proceso de inoculación del hongo Fusarium sp. a las especies de Passifloras.
3.2.2 Método de incisión de tallo: Se toma un cultivo puro del mismo aislado del
hongo patógenico de origen monosporico (Fusarium sp), con un sacabocado se
toma una porción del aislado de diámetro 0.5 mm, y se coloca en la base del tallo
de la planta, el cual se le hace una incisión con un bisturí de cuchilla fina y estéril.
Después de colocar el hongo se envuelve con parafim, para fijar el hongo con la
herida del tallo. (Figura 2).

‐ 41 ‐
Figura 2. Proceso de inoculación del hongo Fusarium sp. a las especies de Passifloras, a través del método incisión de tallo.
3.3 Evaluación de la respuesta de la inoculación del hongo Fusarium sp. a las diferentes especies de Passifloras.
3.3.1 Diseño experimental
El diseño que se utilizó fue completamente al azar donde se evaluaron 28
introducciones de 17 especies de Passifloras como tratamientos. La unidad
experimental la conformarón 520 plantas sembradas en bolsas plásticas. Se

‐ 42 ‐
incluyó un testigo de maracuyá comercial. La variable de respuesta fue porcentaje
(%) de plantas enfermas.
De los 13 aislamientos de Fusarium que se utilizaron en este estudio, se usó para
la prueba de resistencia el hongo que presentó más patogénicidad, MUC5 (sus
siglas provienen de Maracuyá - Universidad Nacional - Corpoica Palmira), este
hongo se seleccionó en pruebas de patogenicidad exploratorias.
Se realizó un análisis de varianza y prueba de promedio de Duncan p (0.05)
utilizando el paquete estadístico SAS versión 9.1.
Las introducciones y especies de Passifloras se evaluaron semanalmente en la
casa de malla, a partir de la semana siguiente de la inoculación, mediante la
escala de evaluación utilizada por Rodriguez et al., 1999 (Tabla 2). La escala se
modificó de acuerdo a los síntomas que se observaron. Se llevo registro
fotográfico.

‐ 43 ‐
Tabla 2. Escala de evaluación de daño en tallos y raíces en maracuyá propuesta por Rodríguez et al., 1999.
Grado de daño
% de daño
síntoma
1
0 - 10
Plantas sanas o base del tallo sano.
3
11-20
Base del tallo ligeramente necrosado, Clorosis leve de las hojas
5
21-50
Base del tallo necrosadas puede presentar un color violeta,
marchitamiento de las hojas
7
50-75
Tercio inferior del tallo necrosado. Marchitamiento y defoliación severa
de las hojas
9
76-100
Tallos necrosados o muertos
‐ 44 ‐
3.4 Identificación morfológica y molecular de las especies del género Fusarium aislados de Passifloras con síntomas de secadera en el Valle del Cauca, Colombia.
Un total de 13 hongos (MUC1, MUC2, MUC3, MUC4, MUC5, MUC6, MUC7,
MUC8, MUC9, MUC10, MUC11, MUC12 y MUC13) pertenecientes al género
Fusarium y provenientes del departamento del Valle del Cauca, localizados en los
municipios Roldanillo, Palmira, La Victoria, Bolívar y Limites La Unión-Toro, fueron
aislados en CORPOICA Palmira, del hospedero Passiflora edulis Sims var
flavicarpa Degener (maracuyá comercial), de tejidos del tallo y de raíces, para ello
se desinfestaron previamente, siguiendo la metodología descrita por Seel et al.,
(1973). Estos hongos fueron aislados por Bonilla et al., (2008),
Para su identificación morfológica se utilizó el manual de Nelson et al., (1983);
Leslie et al., (2006). Además se registraba el sitio geográfico donde se tomó la
muestra (raíz-tallo), edad de cultivo aislado y tejido de donde fue aislado (tabla 3).
Tabla 3. Aislamientos obtenidos de cultivo de maracuyá en el Valle del Cauca, presentando síntomas de marchitamiento (Secadera).
CODIGO FECHA LATITUD LONGITUD ALTITUD EN M
MUNICIPIO FINCA EDAD DEL
CULTIVO
TEJIDO COLECTADO
MUC1 01.14.10 4° 32.82” 76° 38,06” 938 Roldanillo La esperanza
4 Meses Raíz
MUC2 01.14.09 4° 56’ 56” 76° 3’ 29,48” 927 Limites la Unión-Toro
Veragua 10 Meses Raíz
MUC3 10.28.10 4° 30’ 33,7”
76° 3’ 07,6” 933 La victoria Primavera 10 Meses Tallo-Raíz
MUC4 01.03.11 3°31´48” 76°81´13” 1000 Palmira CORPOICA 10 Meses Tallo-Raíz
MUC5 10.27.09 4° 30’ 33,7”
76° 3’ 07,6” 933 La victoria Primavera 4 Meses Tallo-Raíz
MUC6 01.14.10 4° 34.01” 76° 11’ 20,54”
965 Bolivar La Argentina 4 Meses Raíz
MUC7 01.14.10 4° 32,82” 76° 38,06” 938 Roldanillo La esperanza
4 Meses Raíz
MUC8 01.14.10 4° 34,01” 76° 11’ 20,54”
965 Bolivar La Argentina 4 Meses Raíz
MUC9 01.14.10 4° 32.82” 76° 38,06” 938 Roldanillo La esperanza
4 Meses Raíz
MUC10 01.14.10 4° 34.01” 76° 11’ 20,54”
965 Bolivar Veragua 4 Meses Raíz

‐ 45 ‐
MUC11 01.14.10 4° 56’ 56” 76° 3’ 29,48” 927 Limites la Unión-Toro
La Argentina 4 Meses Raíz
MUC12 01.14.10 4° 32.82” 76° 38,06” 938 Roldanillo La esperanza
4 Meses Raíz
MUC13 10.27.09 4° 30’ 33,7”
76° 3’ 07,6” 933 La victoria Primavera 10 Meses Tallo-Raíz
MUC (Maracuyá Universidad Nacional – Corpoica)
3.4.1 Caracterización morfológica.
Teniendo en cuenta los criterios de muchos autores (Gerlach y Nirenberg (1982);
Singh et al. (1991); Nelson et al. (1993); López (2003), López y López, (2004) y
Leslie et al., (2006)) relacionados con las características de las especies del
género Fusarium sobre diferentes medios de cultivos, se seleccionó el medio PDA
(papa, dextrosa, agar) y medio CLA (Carnation Leaf Agar; (Singleton, Mihail y
Rush, 1992)) para realizar la caracterización morfológica de las especies aisladas.
La mayoría de estas clasificaciones se basan en características macro y
microscópicas del aislado y se considera un género anamorfico dentro de los
ascomicetes. (de Hoog et al., 1998; 2000).
Cuatro pequeñas fracciones de la estructura vegetativa de los aislados, se
dispusieron en placas de petri de 9cm de diámetro y fueron incubadas a una
temperatura de 28°C.
La caracterización de los aislados fue establecida teniendo en cuenta el tipo de
pigmento de la colonia, así como su hábito de crecimiento. Para la caracterización
de los parámetros morfológicos (esporodoquios, clamidospora, forma y tamaño de
los conidios) se realizó a través del microscopio, en un aumento de 10, 40 y 100x.,
el color que desarrollan depende de la especie y el medio de cultivo donde se
siembra, el cual puede ser blanquecino, crema, purpura etc. El micelio aéreo suele
ser abundante y de aspecto algodonoso (de Hoog, 1998; Samson et al., Leslie and
Summerell, 2006).

‐ 46 ‐
3.4.2 Caracterización molecular.
Las características morfológicas que a nivel de género y especies distinguen a un
hongo se limitan con frecuencia a permitir su identificación; a ello también se
aplican las técnicas fisiológicas y bioquímicas. Avances técnicos importantes han
estimulado el uso de técnicas moleculares que son universalmente aplicables,
como la reacción de cadena de la polimerasa (PCR) ha permitido el análisis
molecular de las células fúngicas o aún esporas, material seco, u organismos
extintos, como también a la selección de oligonucleótidos específicos para
hongos, almacenado en bancos de genes (Olive and Bean, 1999; Wassenaar and
Newell, 2000; Leslie and Summerell, 2006).
3.4.3 EXTRACCIÓN DE ADN DE Fusarium
3.4.3.1 Preparación de los aislados para la extracción de ADN.
Todos los aislados seleccionados provienen de cultivos monosporicos y
sembrados en medio de cultivo PDA en una cámara de flujo laminar vertical e
incubados a una temperatura de 28ºC durante 7 a 10 días, tiempo suficiente para
el desarrollo del hongo.
3.4.3.2 Procedimiento de la extracción de ADN.
El procedimiento utilizado para el ADN cromosómico fue descrito por Wilson
(1987), con modificaciones realizadas por Gómez (2008), como se explica a
continuación.
Se recoge el micelio del aislado de Fusarium sp., con una espátula metálica
estéril, depositándolo en un microtubo Eppendorf de 2 mL esterilizado,

‐ 47 ‐
inmediatamente se muele con un punzón hasta deshacer gran parte del micelio.
Posteriormente se añade 750 L de la solución tampón (250 mL Tris- HCl 100mM
pH 7.2; 250 mL EDTA 100mM y 250 mL dodecil sulfato sódico (SDS) al 10%, se
agita y se mantiene todo en frío. Después, se añadió 3 L de una solución de
proteinasa K (20 mg/mL), se agitó y se incubó a 37 ºC durante 1 h.
A continuación se añade 100 L de NaCl 5M con el fin de proporcionar la
concentración salina necesaria para que el Cetyl Trimethyl Ammonium Bromide
(CTAB) no se acompleje con el ADN. Se adiciona 80 L de solución CTAB/NaCl
(CTAB al 10% en NaCl 0.7 M), de nuevo se agitó en homogenizador vortex y se
incubó a 65 ºC durante 10 min.
Para la purificación del ADN se añade 700 L de cloroformo/alcohol isoamílico
(24:1), se agita y se centrifuga a 12000 rpm durante 10 min., para eliminar los
complejos formados por el CTAB. El sobrenadante se transfiere a un nuevo
microtubo, al que se le añadió un volumen equivalente (entre 600-700 L) de
fenol/cloroformo/alcohol isoamílico (25:24:1), agitándolo después. El fenol
desnaturaliza las proteínas y las precipita, separándose del ADN que se mantiene
disuelto. Posteriormente la mezcla se centrifuga a 12000 rpm durante 5 min., para
eliminar los restos del complejo CTAB; se repite el proceso si es necesario. Las
proteínas quedan en la interfase y los ácidos nucleícos en la fase superior acuosa.
El sobrenadante (500-700 L), que contiene los ácidos nucleídos, se transfieren a
otro microtubo al que se le añade entre 500-600 L de isopropanol para precipitar
el ADN; se agita suavemente y se centrífuga a 12000 rpm durante 10 min. Se
elimina el isopropanol y se añadió 500 L de etanol frío al 70%; se agita y se
centrifuga durante 5 min. Transcurrido este tiempo, se elimina el etanol y se seca
la muestra al vacío.

‐ 48 ‐
Finalmente, se resuspende el ADN en 50 L de tampón TE (Tris 10 mM, EDTA
1mM, pH 8) y se añaden 3 L de RNAsa (25 mg/mL) para eliminar los restos de
RNA, dejando el ADN extraído a 5 ºC durante 24 h.
Para comprobar si la extracción fue correcta se visualizó en gel de agarosa, al
1.2% (p/v). Para la conservación del DNA extraído se conserva en microtubos a -
20ºC.
3.4.3.3 PCR para amplificación de la región ITS1 – ITS4 y posterior secuenciación de los aislados del género Fusarium que manifiestan síntomas de secadera en plantas de maracuyá
La región ITS del ADNr fue amplificado utilizando los cebadores universales ITS1
(5´-TCCGTAGGTGAACCTGCGG-3´) y ITS4 (5´TCCTCCGCTTATTGATATGC-3´)
White et al., (1990). La mezcla para la amplificación se realizó en un volumen 50
µL que contenía 5µL de 10x PCR tampón (500 mM KCl, 100mM Tris HCl (pH 9),
1% Tritón X-100), 3µL de 50 mM MgCl2, 3µL de ADN a una concentración entre
100 y 180 ng., 0.4 µL de cada dNTP´s a una concentración de 100 mM en total,
2,5 unidades de Taq ADN polimerasa (Eppendorf) y 1 µL de 20 µM de cada
cebador ITS1 / ITS4. La amplificación fue realizada en un termociclador PTC-
100TM programmable thermal controller siguiendo los ciclos de temperatura que
se describen a continuación: Iniciadores ITS1/ITS4, desnaturalización inicial 95ºC
por 60seg, ciclos 35, segunda desnaturalización 95ºC a 30seg, primer
alineamiento 55ºC a 60seg, extracción ADN 72ºc a 60seg, extensión final ADN
72ºC a 6 min.
El producto fue visualizado por electroforesis en gel de agarosa, donde se obtuvo
fragmentos entre 500 a 700 pares de bases (pb). Luego fueron enviados a
secuenciar al instituto de Biotecnología de Corea. Una vez obtenida estas
secuencias se ensamblaron manualmente con los programas informáticos

‐ 49 ‐
CHROMAS y PHYDIT. Una vez producidas las secuencias completas de cada una
de los aislados, se determinó la identidad a nivel de especies de cada uno de ellos
mediante el programa BLAST (Basic Local Alignment Search Tool) obtenida a
través de NCBI (National Center for Biotecnology information, 2008).
3.4.3.4 Purificación y secuenciación del fragmento amplificado de la región ITS1-ITS4
Los productos amplificados de la región ITS1-ITS4 de los aislados de los hongos
proveniente del cultivo de maracuyá se purificaron utilizando el kit Gen Elude TM
PCR clean-Up (SIGMA); se conservó el ADN purificado a -20ºC hasta su uso.
Aproximadamente 2µl del producto de la PCR purificada fueron requeridos en
cada secuenciación. La secuenciación fue llevada a cabo en Korea por Macrogen.
3.5 Cepas de referencias.
En este trabajo se utilizaron tres (3) cepas de referencias del Genbank, y dos (2)
cepas suministradas por la Colección Española de Cultivos (CECT) y utilizadas en
el trabajo doctoral de E, D. Gómez y Tabla 4.

‐ 50 ‐
Tabla 4. Cepas de referencia.
Referencia Especie
CECT2715 F. oxysporum
CECT20232 F. solani
JN664959 F. verticillioides
JN232128 F. verticillioides
GQ505450 F. brachygibbosum
3.6 Caracterización molecular de las especies de Passifloras seleccionadas para la evaluación de resistencia a Fusarium sp.
3.6.1 Colecta del material vegetal
Teniendo en cuenta la información de la distribución de las especies, se
identificaron sitios de muestreo en 5 departamentos; Antioquia, Quindío, Risaralda,
Huila y Valle del Cauca. En Corpoica Palmira se conserva un banco de trabajo con
139 introducciones de 32 especies de Passifloras de las cuales se evaluaron 28
introducciones de 17 especies. El 52% fue material donado por las instituciones
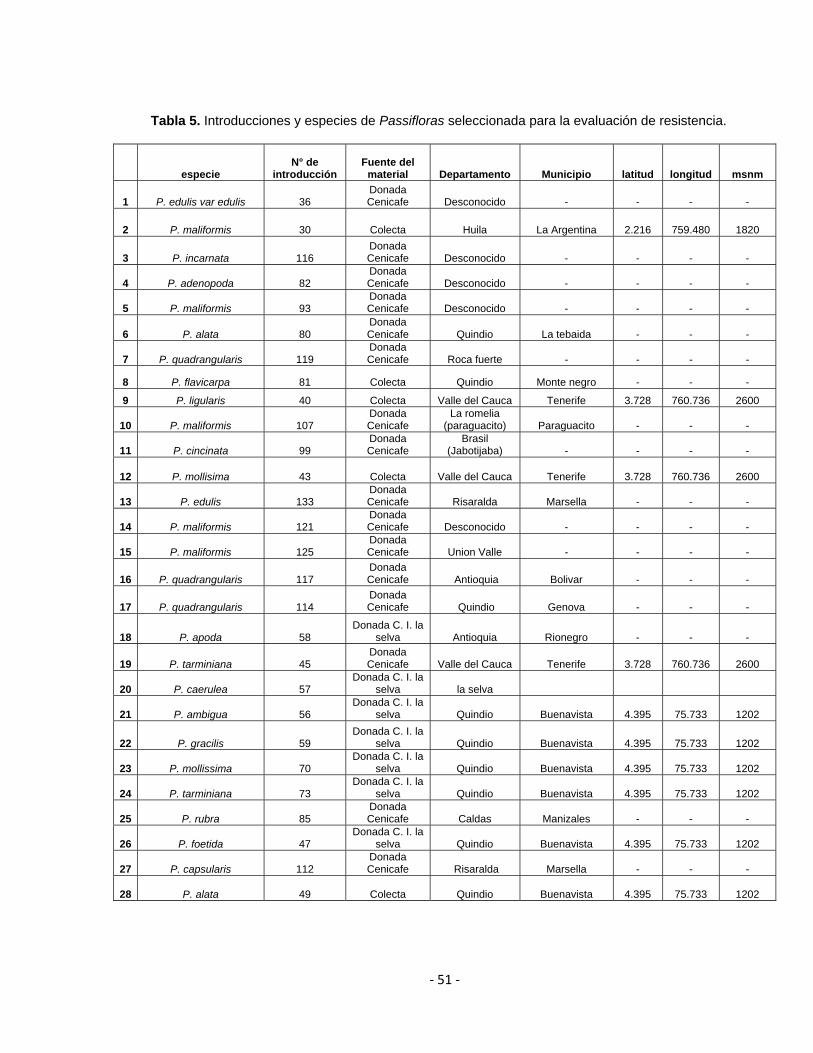
Corpoica La Selva y Cenicafé. Tabla 5.
‐ 51 ‐
Tabla 5. Introducciones y especies de Passifloras seleccionada para la evaluación de resistencia.
especie
N° de introducción
Fuente del material Departamento Municipio latitud longitud msnm
1 P. edulis var edulis 36 Donada Cenicafe Desconocido - - - -
2 P. maliformis 30 Colecta Huila La Argentina 2.216 759.480 1820
3 P. incarnata 116 Donada Cenicafe Desconocido - - - -
4 P. adenopoda 82 Donada Cenicafe Desconocido - - - -
5 P. maliformis 93 Donada Cenicafe Desconocido - - - -
6 P. alata 80 Donada Cenicafe Quindio La tebaida - - -
7 P. quadrangularis 119 Donada Cenicafe Roca fuerte - - - -
8 P. flavicarpa 81 Colecta Quindio Monte negro - - -
9 P. ligularis 40 Colecta Valle del Cauca Tenerife 3.728 760.736 2600
10 P. maliformis 107 Donada Cenicafe
La romelia (paraguacito) Paraguacito - - -
11 P. cincinata 99 Donada Cenicafe
Brasil (Jabotijaba) - - - -
12 P. mollisima 43 Colecta Valle del Cauca Tenerife 3.728 760.736 2600
13 P. edulis 133 Donada Cenicafe Risaralda Marsella - - -
14 P. maliformis 121 Donada Cenicafe Desconocido - - - -
15 P. maliformis 125 Donada Cenicafe Union Valle - - - -
16 P. quadrangularis 117 Donada Cenicafe Antioquia Bolivar - - -
17 P. quadrangularis 114 Donada Cenicafe Quindio Genova - - -
18 P. apoda 58 Donada C. I. la
selva Antioquia Rionegro - - -
19 P. tarminiana 45 Donada Cenicafe Valle del Cauca Tenerife 3.728 760.736 2600
20 P. caerulea 57 Donada C. I. la
selva la selva
21 P. ambigua 56 Donada C. I. la
selva Quindio Buenavista 4.395 75.733 1202
22 P. gracilis 59 Donada C. I. la
selva Quindio Buenavista 4.395 75.733 1202
23 P. mollissima 70 Donada C. I. la
selva Quindio Buenavista 4.395 75.733 1202
24 P. tarminiana 73 Donada C. I. la
selva Quindio Buenavista 4.395 75.733 1202
25 P. rubra 85 Donada Cenicafe Caldas Manizales - - -
26 P. foetida 47 Donada C. I. la
selva Quindio Buenavista 4.395 75.733 1202
27 P. capsularis 112 Donada Cenicafe Risaralda Marsella - - -
28 P. alata 49 Colecta Quindio Buenavista 4.395 75.733 1202

‐ 52 ‐
3.6.2 Material vegetal
Se colectaron hojas jóvenes de 28 accesiones de 17 especies de Passifloras. Las
colectas se realizaron utilizando cortes del tejido vegetal, envuelto en bolsa de
plástico y conservado en cajas de icopor con hielo para su transporte hasta el
momento de su procesamiento en el Laboratorio de Biología Molecular de la
Universidad Nacional de Colombia sede Palmira.
3.6.3 Extracción de ADN
Las muestras fueron maceradas utilizando la misma bolsa donde se colecto la
muestra. Para la extracción de ADN se utilizó la metodología descrita por
Dellaporta et al (1983) con algunas modificaciones.
3.6.4 Evaluación de cantidad y calidad de ADN
Para la evaluación de la calidad y cantidad del ADN obtenido se realizaron geles
de agarosa al 0,8% corridos en tampón TBE 0.5X (Tris-borato 0045M; EDTA
0.001M) a 100 voltios durante una hora. Estos fueron teñidos con bromuro de
etidio a una concentración final de 0.5 ug/ml. Las concentraciones de ADN se
determinaron mediante comparación con concentraciones conocidas de ADN del
bacteriófago lambda. Se realizaron diluciones de todas las muestras extraídas a
una concentración final de 5 ng/ul en tubos nuevos, las cuales fueron utilizadas
como ADN de trabajo para las reacciones de PCR.

‐ 53 ‐
3.6.5 Marcadores moleculares RAMs
Se evaluaron 5 cebadores para RAMs (tabla 6) con las condiciones de
amplificación por PCR descritas por Bonilla et al (2008) para uchuva Physallis
peruviana.
Tabla 6. Cebadores RAMs utilizados en la caracterización molecular.
Cebador Secuencia CGA DHBCGACGACGACGACGA CT DBDCTCTCTCTCTCTCTC TG HVHTGTGTGTGTGTGTGT
CCA DDBCCACCACCACCA GT VHV GTG TGT GTG TG
La amplificación fue realizada en un termociclador PTC-100TM programmable
thermal controller siguiendo los ciclos de temperatura que se describe a
continuación. Desnaturalización inicial 95°C por 5min, desnaturalización 95°C por
30seg, hibridación para el cebador CCA 50°C por 45seg, para los cebadores TG y
CT 55°C por 45seg y el cebador 58°C por 45seg, extensión 72°C por 2min, ciclos
de 37 desde el paso 2, extensión final 72°C por 7min.
Las condiciones de amplificación para RAMs utilizados en la investigación fue:
Buffer TAQ, dNTPs, Cebador, MgCl2, ADN, Taq polimerasa, con una
concentración inicial de 10X, 1.25mM, 50μm, 25mM, 5 ng /μl, 5 U/μl, y una
concentración final de 1X, 0.2mM, 2μM, 1.5mM, 10 ng, 0.625 U, para un volumen
total de 25 μl por muestra.
Los productos amplificados se visualizaron en geles de poliacrilamida al 8% (37:1
acrilamida – bisacrilamida) corridos a 140 voltios por una hora y teñidos bromuro
de etidio.

‐ 54 ‐
3.6.6 Análisis de la información
La información de los patrones de bandas obtenidos se registró en una matriz
binaria de presencia (1) o ausencia (0). Para la selección de bandas polimórficas
se consideró como locus polimórfico aquel en el cual la frecuencia del alelo más
común fue menor al 95%. A partir de esta matriz y usando los programas
SIMQUAL del paquete NTSYS- pc (Numerical Taxonomy System for Personal
Computer) y el programa TFPGA (Tools for Population Genetic Analisys) se
realizaron los análisis estadísticos.
La similitud genética se calculó mediante el coeficiente de Dice Nei- Li (1978)
)2(
2
cba
aS
donde a = bandas compartidas por ambos individuos, b = bandas
presentes en el individuo (1) pero no en (2), y c = bandas presentes en el individuo
(2) pero no en (1). El análisis de agrupamiento se realizó con el programa SAHN
de NTSYS –pc (versión 2.02g, 1998) utilizando el método UPGMA, método
gráfico de agrupamiento por parejas, que usa el promedio aritmético no
ponderado. El dendrograma se construyó con el programa TREE de NTSYS –pc
(versión 2.02g, 1998). Se realizó un análisis de correspondencia múltiple (ACM)
para asociar columnas y filas de la matriz binaria determinando el nivel de
asociación o determinar proximidad (Joseph et al. 1992). Para estimar la
diversidad genética se utilizaron los parámetros de heterocigosidad promedio
esperada (He) y el porcentaje de loci polimórficos (P), los cuales se estimarán
sobre todos los loci y el promedio de los mismos de acuerdo con la fórmula no
sesgada de Nei (1973) así:
H= 1-f (i)2
f(i): Frecuencia del alelo i en la población.
‐ 55 ‐
4. RESULTADOS Y DISCUSIÓN
4.1 Estandarización de inoculación para hongos del género Fusarium spp agente causal de la secadera.
La evaluación de los dos métodos para la inoculación del hongo del género
Fusarium, se realizó durante un mes con dos especies, P. flavicarpa 81 y P.
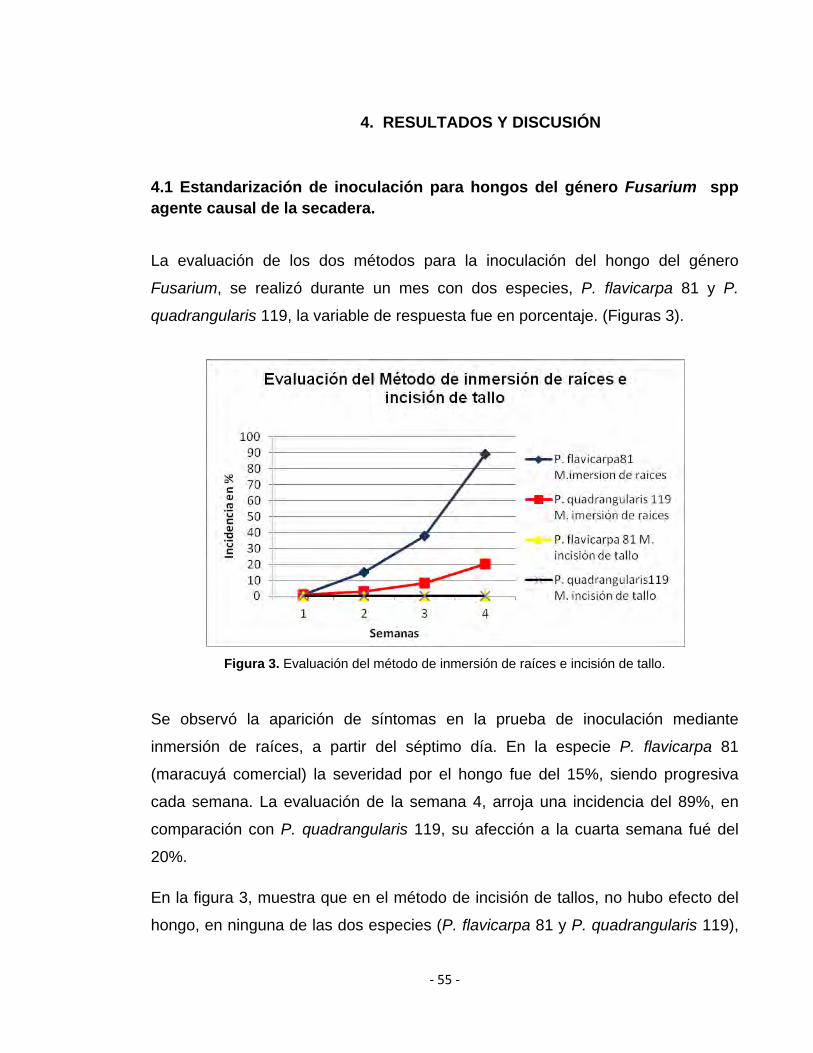
quadrangularis 119, la variable de respuesta fue en porcentaje. (Figuras 3).
Figura 3. Evaluación del método de inmersión de raíces e incisión de tallo.
Se observó la aparición de síntomas en la prueba de inoculación mediante
inmersión de raíces, a partir del séptimo día. En la especie P. flavicarpa 81
(maracuyá comercial) la severidad por el hongo fue del 15%, siendo progresiva
cada semana. La evaluación de la semana 4, arroja una incidencia del 89%, en
comparación con P. quadrangularis 119, su afección a la cuarta semana fué del
20%.
En la figura 3, muestra que en el método de incisión de tallos, no hubo efecto del
hongo, en ninguna de las dos especies (P. flavicarpa 81 y P. quadrangularis 119),

‐ 56 ‐
durante las cuatro semanas no se observó síntomas del hongo, y la incisión en la
base del tallo se cicatrizó.
4.2 Evaluación de la respuesta de la inoculación del hongo Fusarium sp. a las diferentes especies de Passifloras.
La variable de respuesta fue por porcentaje (%) de plantas enfermas (incidencia).
Se realizó un análisis de varianza y prueba de promedio Duncan (Tabla 7 y Figura
4), el cual muestra que hay diferencias significativas a nivel de especies (0.05%).
Especies o introducciones que presenten un porcentaje de plantas enfermas
superior al 10% se considerarán susceptibles al patógeno inoculado.
Tabla 7. Análisis de varianza
Variable dependiente: ABCURVA
Fuente DF Suma de
cuadrados
Cuadrado de la
media F-Valor Pr > F
Modelo 28 12308683.33 455877.16 34.32 0.0001
Error 308 4090836.05 13281.94
Total correcto
336 16399519.38
R-
cuadradoCoef Var Raíz MSE
ABCURVA Media
0.750551 89.96989 115.2473 128.0954
Fuente DF Tipo III SS Cuadrado
de la media
F-Valor Pr > F
Tratamiento 28 12308683.33 455877.16 34.32 0.0001
En la Figura 4 se observa que a través de la aplicación del método de Duncan se
forman cinco grupos entre los tratamientos, los cuales el promedio de las
poblaciones dentro de los grupos no difieren. Se observa que la especie más
susceptible al hongo del genero Fusarium, fue P. tarminiana 45, con una media de
656.33.

‐ 57 ‐
Figura 4. Prueba de promedio Duncan P (0.005)
Las figuras 5 y 6 muestran el progreso de la enfermedad de las 28 introducciones
evaluadas, de las cuales 15 tienen un área bajo la curva menor al 10%, indicando
que existen niveles de resistencia en estas introducciones (Figura 5). Trece (13)
de las quince (15) especies (P. cincinata 99, P. maliformis 121, P. capsularis 112,
P. foetida 47, p. edulis purpura 133, P. rubra 85, P. maliformis 125, P. maliformis
30, P. alata 49, P. quadrangularis 117) no presentaron ningún grado de afección.
En la Figura 6, se observa que 10 de las introducciones de Passifloras evaluadas
(P. tarminiana 45, P. flavicarpa 81, P. gracilis 59, P. quadrangularis 119, P.
tarminiana 73, P. maliformis 93, P. apoda 58, P. ambigua 56, P. mollissima 70, P.
alata 80), son susceptibles al hongo del género Fusarium sp., agente causal de la
secadera, los síntomas de la infección a estas introducciones, superan el 50%.
Tres de las trece llegan hasta el 30% de la afección.
EE E E E E E E E E E E E E E E E E
D
A
CC C C
C D
B B A B
Introducciones
Media

‐ 58 ‐
Figura 5. Evaluación de la resistencia de Passifloras resistentes al hongo del género Fusarium sp. agente causal de la secadera del maracuyá.
Figura 6. Evaluación de la resistencia de especies de Passifloras susceptibles al hongo del género
Fusarium sp. agente causal de la secadera del maracuyá.

‐ 59 ‐
También se observó que dentro de las mismas especies, hubo introducciones
resistentes y susceptibles, como es el caso de la especie P. maliformis, las
introducciones 121, 125 y 30 muestran resistencia al hongo fitopatógeno, en
comparación con P. maliformis 93 que se muestra susceptible, llegando a un 90%
de incidencia. Lo mismo sucedió con las especie P. quadrangularis y P. alata.
Las especies P. edulis var edulis, P. alata y P. maliformis y, han sido reportadas
por Meletti et al., (2005), Holguin y Posada (1990) como resistentes, lo cual se
corrobora con los resultados obtenidos en el presente estudio. Sin embargo, se
observó que en las introducciones pertenecientes a las especies P. maliformis y
P. alata, se presentaron diferencias en la respuesta a la resistencia, posiblemente
a la diversidad genética existente por la naturaleza alógama de la especie.
4.3 Identificación morfológica y molecular de las especies del género Fusarium aislados de Passifloras con síntomas de secadera en el Valle del Cauca Colombia.
Morfológica y molecularmente se identificaron 13 aislados de Fusarium, los cuales
6 pertenecen a Fusarium solani (MUC1, MUC2, MUC4, MUC7, MUC12 y MUC13),
4 a Fusarium oxysporum (MUC3, MUC5, MUC9 y MUC10), dos a Fusarium
verticillioides (MUC6 y MUC8) y uno a Fusarium brachygibbosum (MUC11).
4.3.1 Caracterización morfológica de las especies encontradas en este estudio
Esta caracterización se basó en la descripción de la colonia (macroscópica) en
medio PDA, así como de las estructuras del hongo (microscópica) en medio CLA.
En las figuras 7 a la 32 Se puede observar las características morfológicas de
cada especie.

‐ 60 ‐
4.3.2 Fusarium solani
Su estado sexual corresponde a Haemanectria haematococca (Berkeley &
Broome) Samuels &Nirenberg. Con un sinónimo común de Nectria haematococca.
Las características encontradas en el medio de Cultivo PDA, fueron de color
blanco y crema, abundante micelio. Las estructuras observadas en el medio CLA a
través del microscopio fue: los Macroconidios son relativamente amplios, rectos o
ligeramente curvos, con dos o más septas, con los extremos redondeados.
Microconidios: su forma es oval, fusiforme y reniforme, con 0 a 2 septas.
Clamidosporas: son abundantes, se forman a la tercera semana de su siembra, y
se encuentran en pares o individuales.
Características en medio de cultivo CLA.
Figura 7. MUC1. A. Abundantes clamidosporas individuales y en parejas y B. micro y mesoconidos.
100X 100X

‐ 61 ‐
Características en PDA
Figura 8. MUC1. Colonia blanca con micelio aéreo.
Características en medio de cultivo CLA.
Figura 9. MUC2. A – C. microconidos B - D. clamidosporas.
100X
100X
40X
40X

‐ 62 ‐
Características en PDA
Figura 10. MUC 2. Colonia blanca con escaso micelio aéreo.
Características en medio de cultivo CLA.
Figura 11. MUC 4. A, B - C. micro, meso y macroconidios.
40x
40x
100x
‐ 63 ‐
Características en PDA
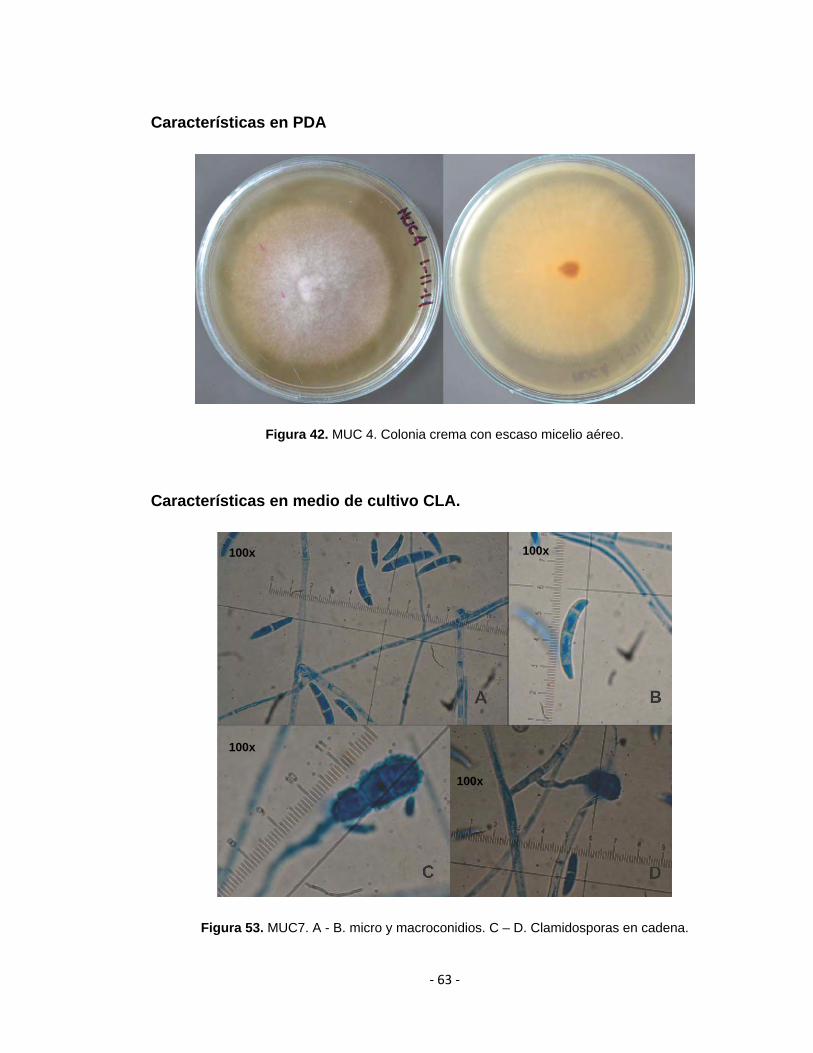
Figura 42. MUC 4. Colonia crema con escaso micelio aéreo.
Características en medio de cultivo CLA.
Figura 53. MUC7. A - B. micro y macroconidios. C – D. Clamidosporas en cadena.
100x
100x
100x
100x
‐ 64 ‐
Características en PDA
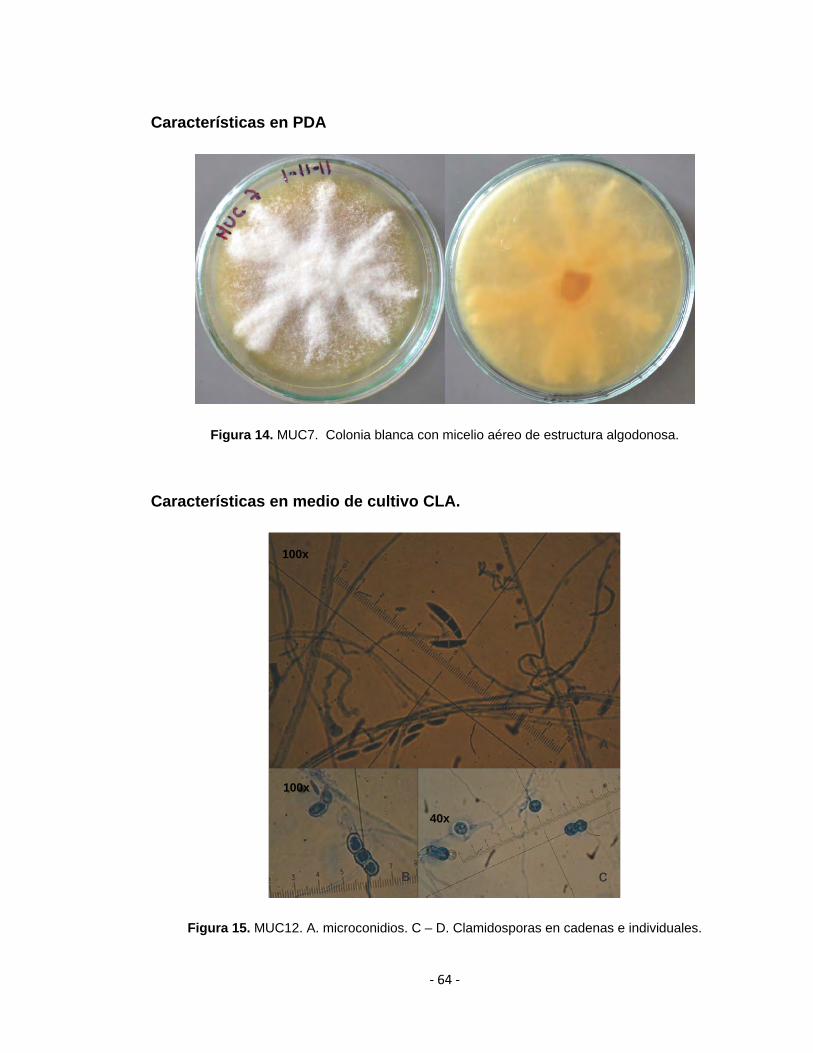
Figura 14. MUC7. Colonia blanca con micelio aéreo de estructura algodonosa.
Características en medio de cultivo CLA.
Figura 15. MUC12. A. microconidios. C – D. Clamidosporas en cadenas e individuales.
100x
100x
40x

‐ 65 ‐
Características en PDA
Figura 66. MUC12. Colonia blanca con escaso micelio aéreo.
Características en medio de cultivo CLA.
Figura 77. MUC13. A. Clamidosporas individuales B. micro y macroconidios.
100x100x

‐ 66 ‐
Características en PDA
Figura 18. MUC13. Colonia blanca con escaso micelio aéreo.
4.3.3 Fusarium oxysporum
Su estado sexual es desconocido. Las características encontradas en el medio de
Cultivo PDA, fueron: colonias de color blanco, purpura y rosado pálido, algunas
con abundante micelio. Las estructuras observadas a través del medio CLA
fueron: Macroconidio: relativamente amplio, recto y generalmente con tres septas.
Microconidio: su forma es oval, fusiforme y no posee septas. Clamidosporas: se
forman en la tercera semana de su siembra. Se encuentra en parejas.
‐ 67 ‐
Características en medio de cultivo CLA.
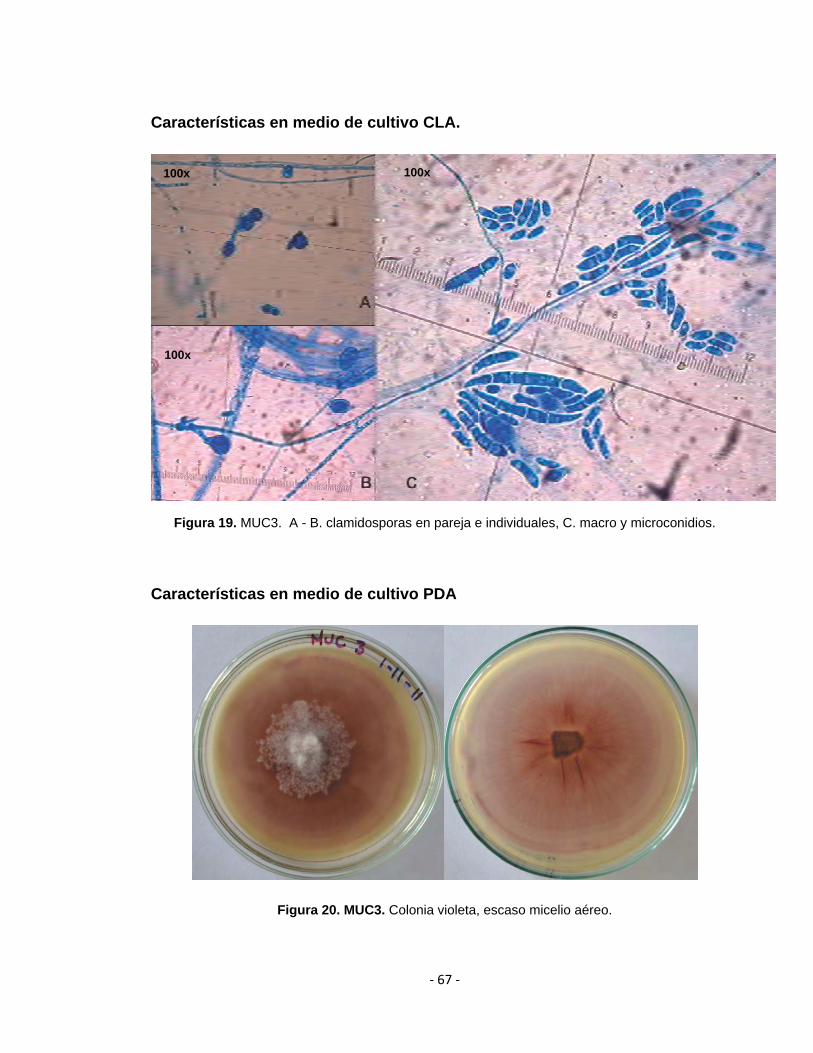
Figura 19. MUC3. A - B. clamidosporas en pareja e individuales, C. macro y microconidios.
Características en medio de cultivo PDA
Figura 20. MUC3. Colonia violeta, escaso micelio aéreo.
100x100x
100x

‐ 68 ‐
Características en medio de cultivo CLA.
Figura 21. MUC5. A - B. meso y microconidios.
Características en medio de cultivo PDA
Figura 22. MUC5. Colonia violeta, escaso micelio aéreo.
40x 40x

‐ 69 ‐
Características en medio de cultivo CLA.
Figura 23. MUC9. A - B. micro, meso y macroconidio C – D. Clamidosporas individuales y en cadena.
Características en medio de cultivo PDA
Figura 24. MUC9. Colonia blanca-violeta, escaso micelio aéreo.
100x
100x 100x
100x

‐ 70 ‐
Características en medio de cultivo CLA.
Figura 25. MUC10. A - B. abundantes clamidosporas en cadena, pareja e individuales. C - D micro, meso y macroconidio.
Características en medio de cultivo PDA
Figura 26. MUC10. Colonia violeta, escaso micelio aéreo.
100x
100x 100x
100x

‐ 71 ‐
4.3.4 Fusarium verticillioides
Su estado sexual corresponde a Gibberella moniliformis. Las características
encontradas en el medio de Cultivo PDA, fueron: colonias de color blanco y rosado
pálido, con abundante micelio. Las estructuras observadas a través del medio CLA
fueron: Macroconidio: relativamente amplio, recto y generalmente con tres septas.
Microconidio: su forma es oval, fusiforme y contiene de cero a una septa; no forma
clamidosporas.
Características en medio de cultivo CLA.
Figura 27. MUC6. A. Células agrandadas. B. microconidios.
100x100x
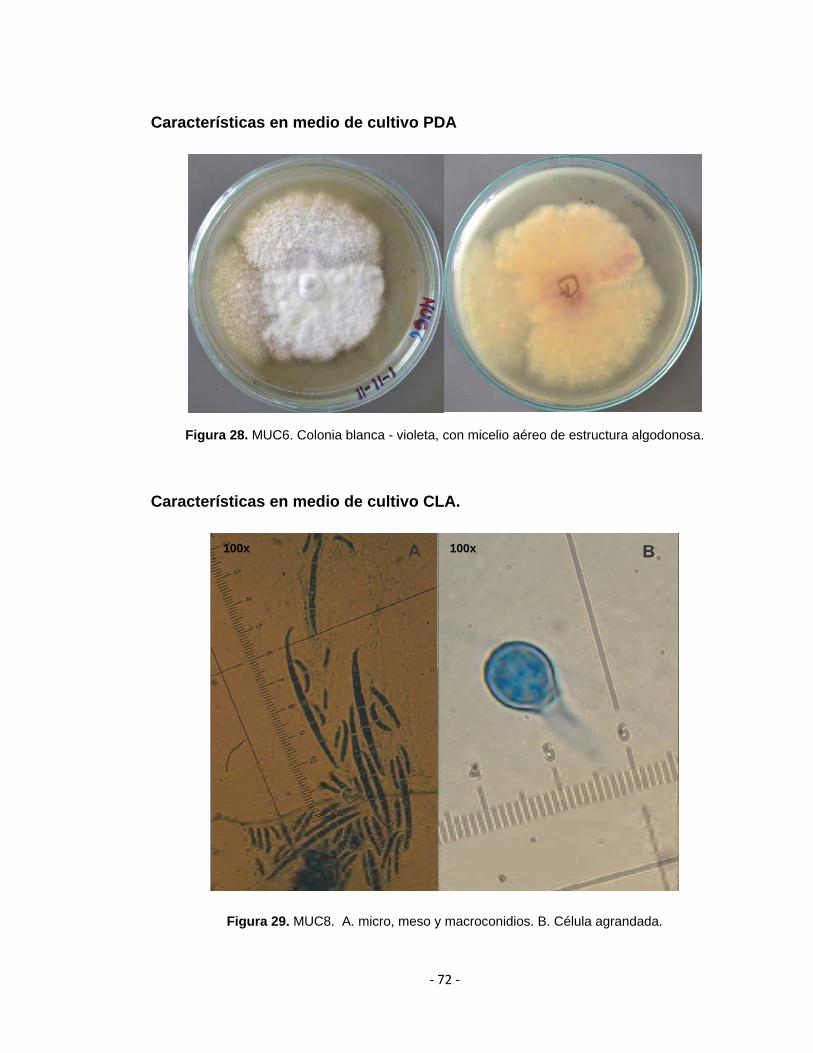
‐ 72 ‐
Características en medio de cultivo PDA
Figura 28. MUC6. Colonia blanca - violeta, con micelio aéreo de estructura algodonosa.
Características en medio de cultivo CLA.
Figura 29. MUC8. A. micro, meso y macroconidios. B. Célula agrandada.
100x100x
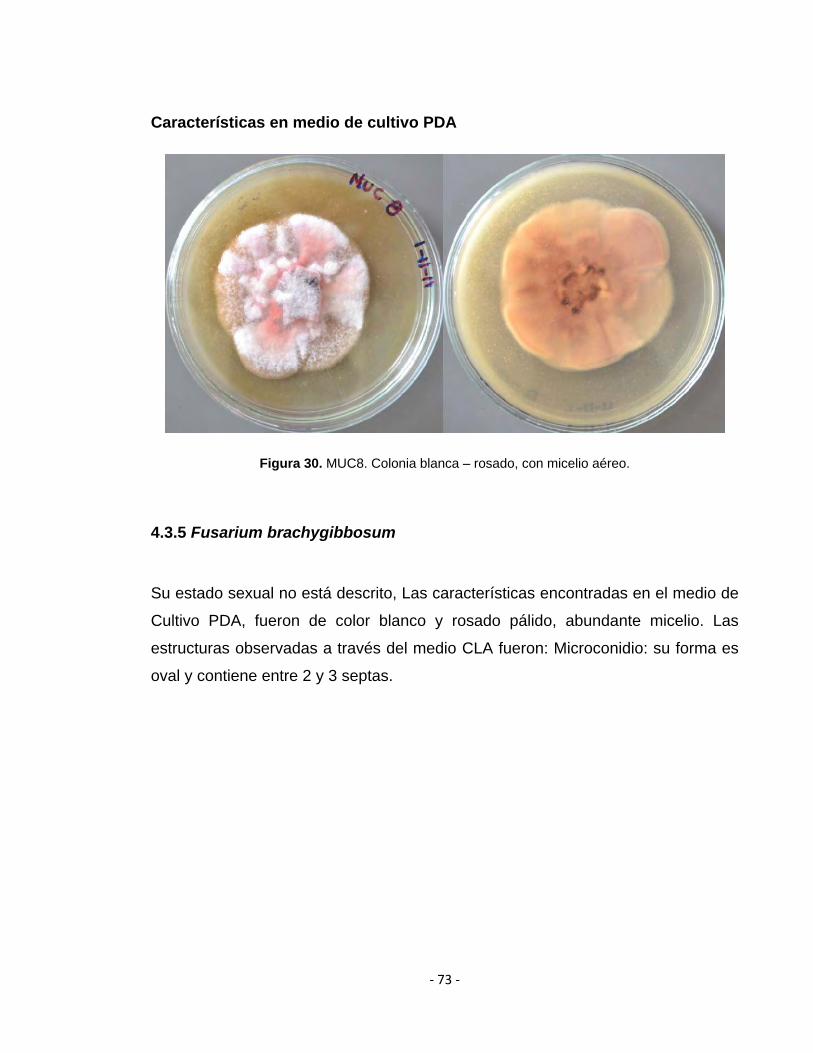
‐ 73 ‐
Características en medio de cultivo PDA
Figura 30. MUC8. Colonia blanca – rosado, con micelio aéreo.
4.3.5 Fusarium brachygibbosum
Su estado sexual no está descrito, Las características encontradas en el medio de
Cultivo PDA, fueron de color blanco y rosado pálido, abundante micelio. Las
estructuras observadas a través del medio CLA fueron: Microconidio: su forma es
oval y contiene entre 2 y 3 septas.

‐ 74 ‐
Características en medio de cultivo CLA.
Figura 81. MUC11. A – B. Estructuras desconocidas. C. microconidios.
100x 100x
100x

‐ 75 ‐
Características en medio de cultivo PDA
Figura 32. MUC11. Colonia blanca – violeta con escaso micelio aéreo.
4.4 Identificación de los aislados por secuenciación
Los productos se analizaron mediante la secuenciación de cadena simple. La
reacción de cada secuenciación consistió en aproximadamente 500 a 600
nucleótidos (Figura 33). La secuencia de datos fue proporcionado como formato
alfabetico como archivo ABI, mostrando la secuencia completa y como
electroferograma que es legible mediante el programa informático CHROMAS
VERSION 1.5 (C. McCarthy School o Health Science, Griffith Universiyty.
Queensland. Australia), para obtener la secuencia del fragmento completo se
ensamblaron manualmente los fragmentos continuos generados por los cebadores
“forward” y “reverse”, usando el programa PHYDIT. La identificación más próxima
fue determinada en la base de datos (National Center for Biotecnology information,
utilizando el programa BLAST (Basic Local Alignment Search Tool;
http://www.ncbi.nlm.nih.gov/). Tabla 8. Las secuencias fueron alineados para
generar arboles filogenéticos y matrices de similitud de nucleótidos usando el
programa PHYDIT (http://plaza.snu.ac.kr/~jchun//phydit/).

‐ 76 ‐
Figura 93. Amplificación en gel agarosa de los hongos del género Fusarium.
Tabla 8. Análisis del programa BLAST para comparar la máxima identidad de las 13 especie de Fusarium sp.
AISLADO DEL
HONGO
IDENTIFICACIÓN MACRO Y
MICROSCOPICA
ACCESIÓN GENBANK
ESPECIE MAS CERCANA PORCENTAJE
DE SIMILARIDAD
MUC1 Fusarium sp. JN127379.1 Fusarium solani 99%
MUC 2 Fusarium sp. GU066713.1 Fusarium solani 100%
MUC 3 Fusarium sp. EU727455.1 Fusarium oxysporum 100%
MUC 4 Fusarium sp. AB258993.1 Fusarium solani 100%
MUC 5 Fusarium sp. GU205817.1 Fusarium oxysporum 100%
MUC 6 Fusarium sp. HM210092.1 Fusarium verticillioides 99%
MUC 7 Fusarium sp. EU625405.1 Fusarium solani 100%
MUC 8 Fusarium sp. HM210092.1 Fusarium verticillioides 99%
MUC 9 Fusarium sp. HQ451888.1 Fusarium oxysporum 99%
MUC 10 Fusarium sp. JF429684.1 Fusarium oxysporum 99% MUC 11 Fusarium sp. JF300441.1 Fusarium brachygibbosum 100% MUC 12 Fusarium sp. GU066713.1 Fusarium solani 99%
MUC 13 Fusarium sp. GU066713.1 Fusarium solani 100%
Los resultados de la identificación por secuenciación se muestra en la tabla 9. Se
confirmó por esta técnica que los aislados MUC1, MUC2, MUC4, MUC7, MUC12
1 2 3 4 5 6 7 M
500PB

‐ 77 ‐
y MUC13 pertenecen a la especie F. solani, para los aislados MUC3, MUC5,
MUC9 y MUC10, pertenece a la especie F. oxysporum, los aislamientos MUC6 y
MUC8 a la especie Fusarium verticillioides. Uno de los 13 aislados, resultó ser
identificado como F. brachygibbosum, Estas especies obtuvieron una homología
del 99 al 100%, Mediante BLAST.
Con las secuencias de nucleótidos generadas, se construyó un árbol filogenético
(Figura 34) cuya raíz es Acremonium del Genbank accesión N° DQ914675 donde
se agrupan los 13 aislados de Fusarium sp. asociado con la secadera en
maracuyá, en cuatro grupos: El grupo I incluye dos aislamientos MUC6 y MUC8,
estos dos aislamientos se agruparon con las cepas de referencia JN664959 y
JN232128 (Fusarium verticillioides); el Grupo II, con el aislamiento MUC11, y la
cepa de referencia GQ505450 (Fusarium brachygibbosum), siendo el primer
reporte que se da en esta especie en Colombia.; el Grupo III, incluye 4
aislamientos MUC3, MUC5, MUC9 y MUC10, junto con la cepa de referencia
CECT2715 (Fusarium oxysporum); y el Grupo IV con 6 aislamientos, que
corresponden a MUC1, MUC2, MUC4, MUC7, MUC12 y MUC13, junto con la cepa
de referencia CECT20232 (Fusarium solani). Se demuestra que no solo F.
oxysporum y F. solani afectan a este cultivo, como indica las referencia
bibliográficas, si no también especies de Fusarium como verticillioides y
brachygibbosum; que no habían sido reportadas como patógenos en especies de
Passifloras. Además se observa una alta variabilidad de especies de Fusarium en
el departamento del Valle del Cauca.

‐ 78 ‐
0.1
Acremonium
MUC4
CECT20232
MUC3
MUC12
MUC5
MUC13
MUC7
MUC10
JN664959
MUC8
JN232128
MUC6
gq5054-ITS
MUC11
MUC9
CECT2715
MUC2
MUC1
100
100
97
99
93
100
93
Figura 104. Árbol filogenético, la raíz es Acremonium, donde muestra la agrupación de las 13 especies de Fusarium sp. aislados del cultivo de maracuyá con síntomas de secadera, colectadas en seis municipios del Valle del Cauca y amplificado por PCR-ITS, con los iniciadores ITS 1 e ITS 4. Y cuatro cepas de referencias. 0.1 indica la distancia genética. El árbol filogenético se construyó con el progama Treecom. Los porcentajes se calcularon con 1000 réplicas.
I
II
III
IV

‐ 79 ‐
También se observó, que dentro del grupo de F. oxysporum, se forman otros
grupos, lo cual demuestra que hay una variabilidad intraespecie. En el grupo II (F.
brachygibbosum) se puede observar que tiene una homología del 100% con el
aislamiento MUC11; esta especie se ha reportado en los Estados Unidos como
patógeno micotico, pero no atacando plantas (O’Donnell, 2009). Con las cepas de
referencia de F. verticilliodes, se agrupa MUC6 Y MUC8, el cual MUC6 se agrupa
con JN232128 y MUC8 con JN664959; ambos con una homología del 100%. En la
Figura 35 se observa que dentro de la especie solani, se conforman 2 grupos; de
los 6 aislamientos, cinco tienen una homología del 100%, que corresponden a
MUC1, MUC2, MUC7, MUC12 y MUC13. Dentro del grupo de F. oxysporum
(Figura 36) se puede analizar que a pesar de que hay una alta homología de que
son oxysporum, se forman tres grupos, lo que indica que hay una variabilidad
intraespecie. Los aislamientos MUC3 y MUC5 tiene una homología del 100%, esto
puede ser, porque provienen del mismo municipio (La Victoria), pero atacando al
cultivo en edades diferentes. El análisis filogenético de las secuencias de ADN ha
sido utilizado para distinguir y evaluar la relación genética entre las especies de
Fusarium estrechamente relacionados (Yli-Matilla, 2002; Harrow, 2010).

‐ 80 ‐
0.1
Acremonium
MUC4
CECT20232
MUC12
MUC7
MUC13
MUC1
MUC2
100
100
Figura 115. Árbol filogenético relacionando las secuencias ITS de los diferentes aislados de F. solani, mediante el programa Treecom versión 1.3b., se indica valores de “bootstrap” > 98%.
Acremonium
MUC9
CECT2715
MUC10
MUC3
MUC5
100
97
97
100
Figura 126. Árbol filogenético relacionando las secuencias ITS de los diferentes aislados de F. oxysporum, mediante el programa Treecom versión 1.3b., se indica valores de “bootstrap” > 96%.

‐ 81 ‐
4.5 Caracterización molecular de las especies de Passifloras seleccionadas.
A las especies utilizadas en la evaluación de la resistencia, se le realizó un estudio
de diversidad, ya que en algunas accesiones de la misma especie, hubo respuesta
diferente frente al patógeno Fusarium, causante de la secadera.
4.5.1 Extracción de ADN
La maceración del material vegetal como proceso mecánico inicial utilizado para
romper la pared celular y exponer los constituyentes celulares permitió la
liberación del ADN en el tampón de extracción, el cual contiene detergentes y
soluciones que aseguran que una cantidad significativa de ADN no sea atrapada
en los desechos o restos celulares, y/o que el ADN esté completamente disociado
de proteínas y otros contaminantes que pueda interferir en procesos posteriores
(Howell, 1973).
Mediante la metodología de Dellaporta et al (1983) se obtuvieron concentraciones
de ADN entre 50 y 100 ng/ul), de buena calidad para la amplificación por PCR de
los RAMs (Figura 37). Se realizaron algunas modificaciones adicionales a la
metodología consistentes en la realización de una precipitación con fenol:
cloroformo: alcohol isoamílico en proporción 24:24:1 y la dilución final del ADN en
100 ul de tampón TE 1X para asegurar una buena concentración y la eliminación
de otras sustancias que pudieran interferir en las amplificaciones.

‐ 82 ‐
Figura 137. ADN de las accesiones de las especies de Passiflloras, extraído mediante la metodología de Dellaporta et al (1983) con modificaciones.
La cuantificación del ADN mediante comparación con concentraciones conocidas
del bacteriófago Lambda permitió determinar de manera adecuada las
concentraciones de ADN obtenidas mediante las dos metodologías. Esta técnica
permite realizar la cuantificación de manera rápida y menos costosa ya que no
requiere de la utilización de equipos especializados como espectrofotómetros o
fluorómetros ni la utilización de reactivos tóxicos y peligrosos, lo que la propone
como una metodología de fácil y rápida implementación para la cuantificación de
ADN total.
4.5.2 Características de los cebadores
Para la evaluación de la diversidad genética de la colección se seleccionaron
cuatro de los cinco cebadores RAMs utilizados por presentar algún grado de
polimorfismo. El cebador GT no amplificó para ninguna de las muestras
posiblemente por condiciones no apropiadas de PCR, como la temperatura. Los
cuatro cebadores utilizados generaron un patrón de 71 bandas y el número de
bandas por cebador estuvo entre 15 para CCA y 20 para CT (Tabla 9).
M 5 22 23 25 27 30 31 32 33 34 35 36 37 39 40 41 43 44 45 46 48 54

‐ 83 ‐
Tabla 9. Bandas totales y polimórficas obtenida con los cebadores RAMs utilizados para la evaluación de la diversidad genética.
En uchuva utilizando la técnica RAMs se obtuvo un patrón de 50 bandas con siete
cebadores para la evaluación de la diversidad genética de 43 accesiones del
banco de germoplasma de la Universidad Nacional de Colombia sede Palmira
(Bonilla et al. 2008). En mora, se obtuvo un patrón de 58 bandas al utilizar seis
cebadores RAMs en la evaluación de la diversidad genética de 38 accesiones
(Morillo et al. 2005). En guayaba Psidium sp se obtuvieron 72 bandas con 5
cebadores RAMs para la evaluación de la diversidad genética de árboles nativos
de guayaba (Sanabria et al. 2006), En un estudio de la diversidad genética de la
colección nacional de nacedero de la Fundación Centro para la Investigación en
Sistemas Sostenibles de Producción Agropecuaria (CIPAV) se obtuvo un patrón
de 71 bandas con pesos moleculares entre 1500 y 120 pb con cinco cebadores
RAMs (Posso 2010). Al comparar el número de locus RAMs generados por los
cuatro cebadores en este estudio (71 en total) se propone que fue un número
adecuado para estimar parámetros genéticos para evaluar la diversidad de las 28
introducciones de 17 especies de Passifloras y es confiable para el análisis
estadístico (Nybom y Bartish 2000).
Se ha considerando que la máxima diversidad genética se conseguirá en una
población que tienda hacia el equilibrio génico, con valores de He cercanos a 0.5.;
los valores de heterocigosidad esperada (He) calculados mediante el programa
TFPGA considerando todas las accesiones del banco como un solo grupo y para
Cebador Nº bandas totales Nº bandas polimórficas
CT 20 18
CGA 19 15
TG 17 15
CCA 15 9
TOTAL 71 57

‐ 84 ‐
cada uno de los cebadores, mostraron que el valor más alto fue para el cebador
CGA (0,32) y el más bajo para el cebador CCA (0,24). El valor de porcentaje de
locus polimórfico más alto se encontró con el cebador CGA (79%) y el más bajo
con el cebador CCA (60%) (Tabla 10).
Tabla 10. Valores de Heterocigosidad y porcentaje de lous polimórficos (95%) para los cebadores RAMs utilizados.
Posiblemente el alto polimorfismo encontrado con el cebador CGA (79%) y He
(0,32) y el cebador CT con (76%) y He (0,26) se deba a una mayor frecuencia
entre los sitios de hibridación y las cadenas de ADN complementarias de esa
secuencia, lo que evidencia la presencia de dos regiones microsatélites CGA y
CT.
El valor alto de polimorfismo y de Heterocigosidad encontrados con los
cebadores CGA y TG permiten concluir que estos cebadores fueron los que
hicieron el mayor numero de cortes a la diversidad y resultan apropiados para su
utilización en futuras investigaciones sobre la evaluación de la diversidad genética
y estructura poblacional de accesiones de Passifloras. Sin embargo, los otros
cebadores también mostraron un número elevado de cortes. De manera general
hubo correlación entre Heterocigosidad y polimorfismo en los cebadores
evaluados (Figura 38).
Cebador Heterocigosidad
esperada (He)
% locus polimórficos (95%)
CT 0,30 70
CGA 0,32 79
TG 0,26 76
CCA 0,24 60
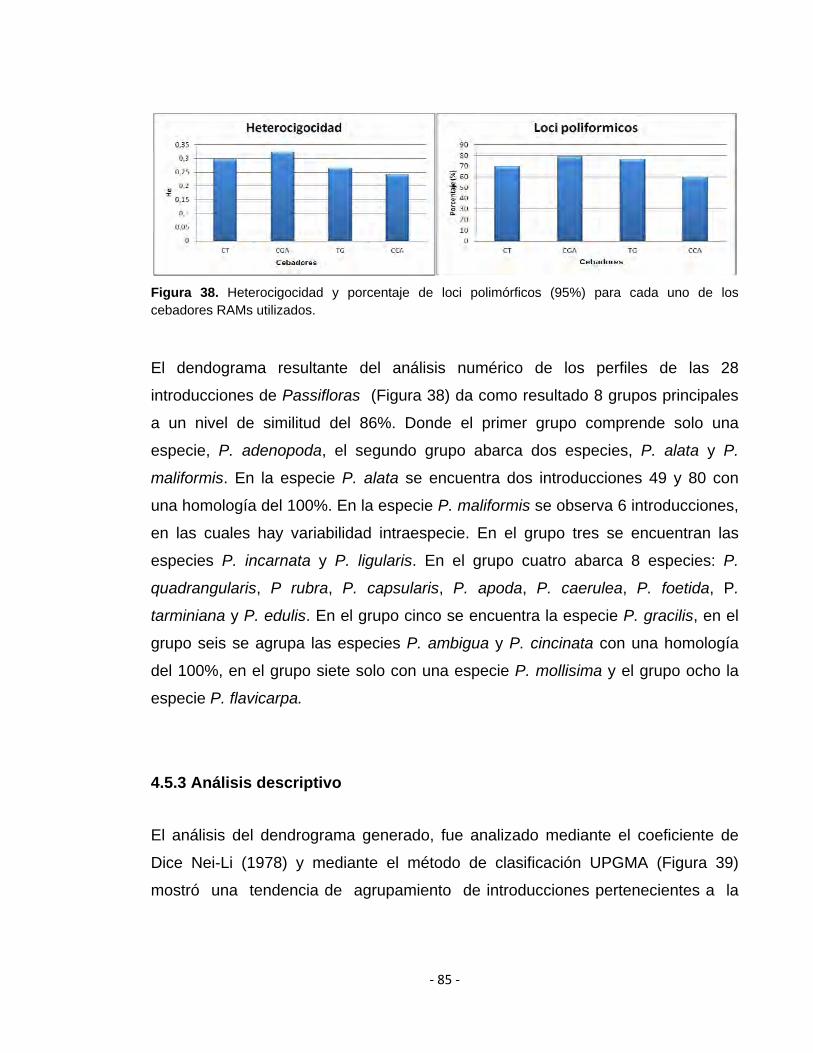
‐ 85 ‐
Figura 38. Heterocigocidad y porcentaje de loci polimórficos (95%) para cada uno de los cebadores RAMs utilizados.
El dendograma resultante del análisis numérico de los perfiles de las 28
introducciones de Passifloras (Figura 38) da como resultado 8 grupos principales
a un nivel de similitud del 86%. Donde el primer grupo comprende solo una
especie, P. adenopoda, el segundo grupo abarca dos especies, P. alata y P.
maliformis. En la especie P. alata se encuentra dos introducciones 49 y 80 con
una homología del 100%. En la especie P. maliformis se observa 6 introducciones,
en las cuales hay variabilidad intraespecie. En el grupo tres se encuentran las
especies P. incarnata y P. ligularis. En el grupo cuatro abarca 8 especies: P.
quadrangularis, P rubra, P. capsularis, P. apoda, P. caerulea, P. foetida, P.
tarminiana y P. edulis. En el grupo cinco se encuentra la especie P. gracilis, en el
grupo seis se agrupa las especies P. ambigua y P. cincinata con una homología
del 100%, en el grupo siete solo con una especie P. mollisima y el grupo ocho la
especie P. flavicarpa.
4.5.3 Análisis descriptivo
El análisis del dendrograma generado, fue analizado mediante el coeficiente de
Dice Nei-Li (1978) y mediante el método de clasificación UPGMA (Figura 39)
mostró una tendencia de agrupamiento de introducciones pertenecientes a la

‐ 86 ‐
misma especie, revelando un nivel aproximado de similitud de 0,96, la formación
de 16 grupos donde se observa la separación de las diferentes especies.
La introducción perteneciente a la especie P. flavicarpa 81 fue las más distanciada
genéticamente del resto de las introducciones a un nivel de similitud del 0,67. Esta
introducción fue una especie colectada en estado silvestre en el municipio de
Montenegro Quindío.
El grupo VII con las introducciones de las especies P. caerulea 57 y P. foétida 47
pertenecen al subgénero Passiflora supersección Passiflora, y el grupo IX con las
especies P. rubra 85 y P. capsularis 112 pertenecen al subgénero Decaloba,
supersección Decaloba, sección Xerogona (Figura 39).
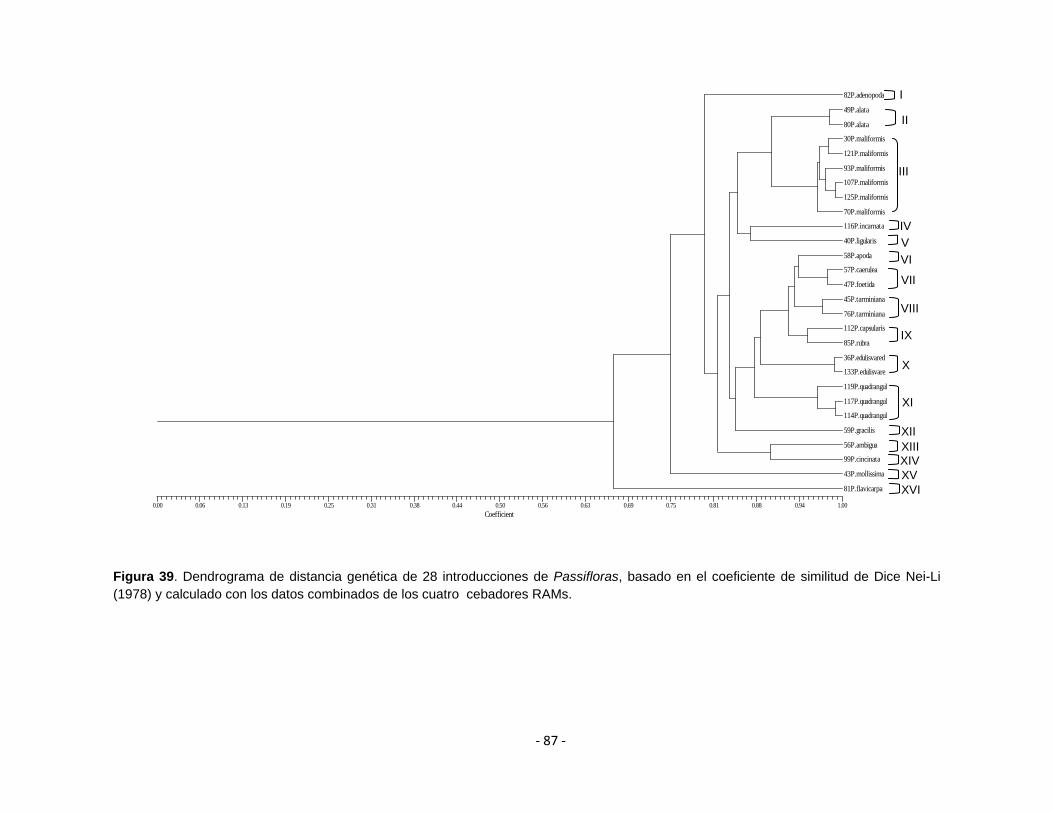
‐ 87 ‐
Figura 39. Dendrograma de distancia genética de 28 introducciones de Passifloras, basado en el coeficiente de similitud de Dice Nei-Li (1978) y calculado con los datos combinados de los cuatro cebadores RAMs.
Coefficient0.00 0.06 0.13 0.19 0.25 0.31 0.38 0.44 0.50 0.56 0.63 0.69 0.75 0.81 0.88 0.94 1.00
82P.adenopoda
49P.alata
80P.alata
30P.maliformis
121P.maliformis
93P.maliformis
107P.maliformis
125P.maliformis
70P.maliformis
116P.incarnata
40P.ligularis
58P.apoda
57P.caerulea
47P.foetida
45P.tarminiana
76P.tarminiana
112P.capsularis
85P.rubra
36P.edulisvared
133P.edulisvare
119P.quadrangul
117P.quadrangul
114P.quadrangul
59P.gracilis
56P.ambigua
99P.cincinata
43P.mollissima
81P.flavicarpa
XIV
I
II
III
IVV
VI
VII
VIII
IX
X
XI
XII
XVXVI
XIII

‐ 88 ‐
4.5.4 Heterocigosidad esperada (He) y porcentaje de loci polimórficos
La He para la colección total fue de 0,2853 lo que revela un alto grado de
diversidad en las introducciones evaluadas. Se escogió la heterocigosidad
insesgada de Nei (1978), pues esta presenta una corrección por el número de
muestras el cual en este estudio es bajo, (28 introducciones de Passifloras),
teniendo en cuenta que en Colombia cuenta con 165 especies (Morillo et al. 2005).
La He se interpreta como la probabilidad de colectar dos genes diferentes en un
muestreo con reemplazamiento del total de genes en una población. También se
le denomina como diversidad genética, aunque esta denominación no es
aconsejable ya que puede confundirse con variación genética “en general” y no
solo al valor de He (Caujapé 2006). El valor encontrado en este análisis es alto ya
que se ha estimado que 0 ≤ He ≤ 0,3 (Nei 1975; Selander 1979; citado por
Caujapé 2006). En un estudio de caracterización molecular de materiales
cultivados de gulupa (Passiflora edulis var. edulis) también se reportó alta
diversidad genética (0,291), probablemente producto del método de propagación,
de su procedencia diversa y el poco tiempo de establecimiento de los cultivos en
el país.
En accesiones de mora Rubus sp (Morillo et al. 2005) y uchuva Physallis
peruviana (Bonilla et al. 2008) del banco de germoplasma de la Universidad
Nacional de Colombia sede Palmira y utilizando RAMs se reportaron valores de
He de 0,31 y 0,25 respectivamente; valores altos comparados con resultados
obtenidos en cultivares comerciales de tomate Solanum lycopersicum en donde la
He fue de 0,18 (Restrepo 2003).
El porcentaje de locus polimórficos encontrado para la colección de Passifloras fue
de 71,83%, lo que nuevamente confirma que las introducciones incluyen una
buena diversidad y heterogeneidad genéticas.

‐ 89 ‐
Estas introducciones analizadas en este estudio, también fueron evaluadas en su
nivel de resistencia a la enfermedad secadera del maracuyá encontrándose
diferentes niveles de resistencia incluso entre introducciones de la misma especie,
lo cual puede ser explicado por la alta diversidad genética que se confirmó en este
estudio, y posiblemente a que en algunos casos corresponden a especies
colectadas en estado silvestre que como es conocido, pueden representar una
fuente de genes valiosos de resistencia.
Los resultados obtenidos sugieren la necesidad de continuar la caracterización del
todo el banco de trabajo de Passifloras de Corpoica Palmira e incluso trabajos
encaminados a recolectar nuevas introducciones que representen de una manera
más amplia la diversidad que puede existir en este género, y que fortalecerán
futuros trabajos de mejoramiento genético.

‐ 90 ‐
5 CONCLUSIONES
La metodología de inoculación de inmersión de raíces fue práctica y
eficiente para inocular al hongo del género Fusarium a 420 plantas
inoculadas, pertenecientes a especies de Passifloras.
De las 28 introducciones evaluadas, 15 son resistentes o tolerantes al
hongo patógeno Fusarium sp. agente causal de la secadera del maracuyá.
Se identificaron 13 aislados del género Fusarium que afectan el cultivo de
maracuyá en 5 municipios del Valle del Cauca, de los cuales cuatro
especies diferentes atacan al cultivo de maracuyá: Fusarium oxysporum,
Fusarium brachygibbosum, Fusarium verticilliodes y Fusarium solani.
Se encontró que hay diferencias inter e intraespecies dentro de las especies
perteneciente al género Fusarium identificas en este estudio.
Nuevamente se confirma que la técnica de la PCR unida a la amplificación
de la región ITS1_ITS4, es una herramienta adecuada para la identificación
de especies pertenecientes al género Fusarium. Se comprueba que la
región ITS es apta en la identificación de especies pertenecientes al
género Fusarium.

‐ 91 ‐
EL valor de He obtenido en el análisis molecular indica que existe una alta
diversidad genética entre las 28 introducciones de Passifloras analizadas
en este estudio.
Los marcadores moleculares RAMs utilizados en este estudio demostraron
ser adecuados para la evaluación de la diversidad genética de Passifloras.
La técnica molecular utilizada en este estudio permite evidenciar un alto
grado de polimorfismo y sensibilidad para la discriminación de las
introducciones y de las especies de Passifloras.
La diversidad y variabilidad encontradas en el género Passifloras puede
deberse al hecho de que las especies son alógamas, auto-incompatibles y
se cruzan con facilidad.
Las diferencias observadas a nivel molecular sirven de apoyo al estudio de
la variabilidad en Passiflora, monitorear hibridaciones interespecíficas, y el
ingreso de genes de las especies silvestres para las cultivadas a través de
programas de mejoramiento genético.

‐ 92 ‐
6 RECOMENDACIONES
Evaluar en campo las especies que salieron resistentes al hongo Fusarium
oxysporum (MUC5).
Evaluar la patogenicidad de las otras especies de Fusarium en las especies de Passifloras resistentes.
Realizar los portainjertos con las especies resistentes y evaluar la compatibilidad de los tallos.
Probar los portainjertos en campo.

‐ 93 ‐
BIBLIOGRAFÍA
AGRIOS G. 2005. Department of Plant Pathology. University of Florida. Fifth Edition
AGOSTINI J., FARALDO H., HABERLE R., RYBAK, Y ANDERSON G. 2002. Causa Y Control De La Antracnosis Del Maracuyá En Plantaciones Comerciales De Misiones, Argentina. Citrusmisiones (28): 27-35. ALEXOPOULOS, C y MIMS, C. 1985. Introducción a la micología. Editorial Omega. Barcelona, España. 630 p. BEDOYA L. J. y MEDINA L. O. 1982. Etiología de la pudrición radicular del cultivo de Maracuyá Amarillo Passiflora edulis f. flavicarpa, Degener.. Universidad Nacional de Colombia. Facultad de Ciencias Agropecuarias Palmira. 76 p. BONILLA M. L., ESPINOSA K., POSSO A. M., VASQUEZ H., MUÑOZ J. E. 2008. Caracterización molecular de 43 accesiones de uchuva de seis departamentos de Colombia. Acta Agronómica 57(2) 109-115. BORGES A., JUNQUEIRA V., COSTA A., ARAÚJO A., SILVA M., PEIXOTO R. y FREITAS F. 2006. Índices de Cruzabilidade do Maracujazeiro-azedo comercial (Passiflora edulis Sims var flavicarpa Degener) com espécies de Passifloras silvestres e progenies de retrocruzamentos, visando à obtenção de resistencia a doenças e autocompatibilidade, Planaltina: Embrapa Cerrados. CARDOZO B., BONILLA M. Y MURCIA N. 2011. Proyecto: “Portainjertos tolerantes como una estrategia para prevenir la secadera del maracuyá en Colombia.” Corporación Colombiana de Investigación Agropecuaria CORPOICA.
CARVAJAL L. 2007. Pulpa de frutas tropicales. En: Universidad de Antioquia, Facultad de Química Farmacéutica, http://huitoto.udea.edu.co/FrutasTropi cales/maracuya.html; consulta: julio 2007.
CAUJAPE, J. 2006. Brújula para botánicos desorientados en la genética de poblaciones. EXEGEN Ediciones. Las Palmas de Gran Canaria. España. 132 p.
CCI. 2007. Costos de comercialización de la maracuyá en la web:

‐ 94 ‐
http://www.cci.org.co/publicaciones/1_Mayo1107%20Comercializacion%20Maracuyá.pdf. CHACON, A. C. 1988. El cultivo de Maracuyá en el Valle del Cauca. Passiflora edulis var. Flavicarpa Degener. Memorias. Primer concurso Nacional de Frutales de Clima Frio. Centro Nacional Investigación Palmira. Colombia. pp 76-89. CRONQUIST, A. 1978. The Evolution and Classification of Flowering Plants. Allen Press. New York Botanical Garden, USA. 396 p. COOK, J. R; BAKER, K. F. 1989. The nature and practice of biological control of plant pathogens. St. Paul, Minnesota. The American Phytopathological Society. 539 p. DE HOOG, G.S., AND GERRITS VAN DEN ENDE A.H. 1998. Molecular diagnostics of clinical strains of filamentous Basidiomycetes. Mycoses, Vol., 41, p 183-189.
DE HOOG, G.S.; GUAARRO J.; GENÉ, J. AND FIGUERAS, M.J. 2000. Atlas of clinical fungi., centraalbureau voor Schimmelcultures, Utrecht, The Netherlands. Universitat Rovira I virgili, Reus, spain. P 1126.
DELLAPORTA S. L., WOOD, J., HICKS, J. B. 1983. A plant DNA inipreparation: Versión II. Plan. Molec Biol Rep 14:19-21. DÍAZ, M.; TORRES, C.; y BRACON N. 1998. Microorganismos fungosos asociados a la semilla de maracuyá Passiflora edulis var. flavicarpa en el norte del Valle del Cauca Boletín de ASCOLFI. VOL. 24 N°. 6 PP 30-31.
ESCOBAR, L. K. 1988. Flora de Colombia. Passifloraceae. Bogotá. Universidad Nacional de Colombia. vol.10. 135p.
ESCOBAR, L. K. 1991. La sistemática y evolución de las Passifloras. pp. 51–54. En: Memorias. 1er Simposium Internacional de Passifloras. Palmira, Colombia.
ESCOBAR, A. & FORERO, R. 1993. Análisis técnico y económico del uso de la cáscara fresca de Maracuyá Passiflora edulis en la ceba comercial de Bóvinos. Tesis de grado. Universidad Nacional de Colombia- Palmira. 80 p.
FERREIRA, F. Y J. OLIVEIRA. 1991. Germoplasma de Passiflora. In: A. São José (ed.). A Cultura do Maracujá. FUNEP. Jaboticabal. pp. 187-200.

‐ 95 ‐
FEUILLET, C. Y MCDOUGAL, J. 2003. A new infrageneric classification of Passiflora. Passiflora 14(1):1-4.
.GARCÍA T. 2002. Cultivo De Maracuyá Amarillo (Guía Técnica). Centro Nacional De Tecnología Agropecuaria Y Forestal. El Salvador. 33p.
GARDNER MAH, O’BRIEN MF, STAFFORD EG, POHLNER PG, MCGIFFIN DC, KIRKLIN JW. 1987. A comparison of aortic valve replacement with a viable cryopreserved and fresh allograft valves with a note on chromosomal studies. J Thorac Cardiovasc Surg; 94: 812-823.
GERLACH, W.; H.; NIRENBER G. 1982 “The genus Fusarium Pictorial Atlas”Mitt. Biol. Bundesanst. Ld. Forstw. Berlin-Dahlem 209:1- 406, 1982.
GÓMEZ C., SCHWENTESIUS R. Y GOMEZ T. 1995. La Producción y el Mercado Mundial del Maracuyá, Centro de Investigaciones Económicas, Sociales y Tecnológicas de la Agroindustria y la Agricultura Mundial (CIESTAAM).
GÓMEZ E.D. 2008. caracterización de cepas toxigenicas del género Fusarium mediante técnicas de biología molecular. Tesis doctoral p 62, 115-120.
GONZÁLEZ, O.; SÁNCHEZ DE P., M.; MIRANDA, J.; MUÑOZ, J. 2002. Actividad de esterasas en rizósfera del maracuyá (Passiflora edulis var, flavicarpa) en diferentes estados de desarrollo y condiciones sanitarias. Memorias del XXIII congreso ASCOLFI. Nuevas tendencias en Fitopatología. Universidad Nacional de Colombia, Bogotá. Julio 3-6/2002. pp 38.
GRAHAM, J., McNICOL, R. J. 1995. An examination of the ability of RAPD markers to determine the relationships within and between Rubus species. Theor Appl genet 90 (7-8): 1128 – 1132.
GUPTA M. 1995. 270 Plantas Medicinales Iberoamericanas. Convenio Andrés Bello - CYTED. Editorial Presencia, Bogotá. Citado en: Rodríguez R., Perea M. 2004. Aislamiento y cultivo de protoplastos en maracuyá. Acta Biológica Colombiana, Vol. 9 Nº. 2.
HARROW, S.A.; FERROKHI-NEJAD, R.; PITMAN, A.R.; SCOTT, I.A.W.; BENTLEY, A.; HIDE, C.; CROMEY, M.G. 2010. Characterization of New Zealand Fusarium populations using a polyphasic approach differentiates the F. avenaceum/F. acuminatum/F. tricinctum species complex in cereal and grassland systems. Fungal Biol. 114, 293–311.
HEYWOOD, V.H. 1985. Las plantas con flores. Edición en Español. Editorial Reverté S.A. España. Pp 98-99.

‐ 96 ‐
HOLGUIN JM., POSADA, J. 1990. Resistenica de passifloráceas a Nectria haematococca Berk y Br. Tesis. Facultad de Ciencias Agropecuarias, Universidad Nacional de Colombia. Medellín. 41p.
HOWELL, S.H. 1973. The Isolation and Analysis of DNA from Eukaryotic Cells. In: Molecular Techniques and Approaches in Developmental Biology, Chrispeels, M.J., Ed., Wiley - Interscience Publication, New York. 127-130p.
IJJÁSZ, I., y EMOKE, S. 1999. Maracuyá (Fruto de la pasión). Editorial Panamericana. Santa Fé de Bogotá. Colombia. 48 p.
JAMES, R. L., PEREZ, R., DUMROESE, R. K., AND WENNY, D. L. 2000. Virulence of Fusarium oxysporum on Douglas-fir germinants: Comparison of isolates from nursery soil and roots of healthy and diseased seedlings. Pages 49-64 in: Proc. Fourth Meeting of IUFRO Working Party S7.03-04 (Disease and Insects in Forest Nurseries). Research Paper 781. A. Lilja and J. R. Sutherland, eds. Finnish Forest Research Institute, Helsinki,Finland. JOSEPH, H., ANDERSON, R., TATHAM, R., BLACK, W. 1992. Multivariate data Analysys with Readings. 3rd Ed. Riverside. 554 p.
KATTAN, G. H. 1997. Transformación de paisajes y fragmentación de hábitats. En: Chaves, M. E. y Arango, N. (eds.). Informe Nacional sobre el Estado de la Biodiversidad – Colombia. Instituto Alexander von Humboldt, PNUMA, Ministerio del Medio Ambiente, Bogotá. Tomo 2:76-82.
KILLIP, E.P. 1938. The American Species of Passifloraceae Field Museum of Natural History Publication, Botanical Series 19 (1, 2): 1-613.
KNIGHT R. Y SAULS J. 2005. La maracuyá o parchita. Universidad de Florida. IFAS Extensión. Pág. 7.
LAGE D., JUNQUEIRA N., BORGER T., BRAGA M., FALEIRO F, BELLON G. y ALENCAR C. 2004. Compatibilidade genética entre o maracujazeiro-azedo comercial (Passiflora edulis Sims var flavicarpa Degener) e espécies de Passifloras silvestres. Embrapa Cerradosl.
LARKIN, R Y FRAVEL D. Efficacy of Various Fungal and Bacterial Biocontrol Organisms for Control of Fusarium Wilt of Tomato. Plant Disease. Vol. 82 No. 9: 1022-1028. LESLIE J.F. AND SUMMERELL, B.A. 2006. The Fusarium Laboratory Manual. Blackwell publishing. P 369.

‐ 97 ‐
LÓPEZ, D. 2003. “Contribución al diagnóstico del género Fusarium Link en Cuba”. Tesis presentada en opción al título académico de Master en Ciencia Agrícolas. Instituto de Investigaciones de Sanidad Vegetal. Ministerio de la Agricultura, 2003.
LÓPEZ, D; Y LÓPEZ, M. 2004 “Influencia del pH y de los medios de cultivo en la Expresión de los caracteres de valor diagnóstico de las especies del Género Fusarium en Cuba” Rev. Fitosanidad 8(3):7-11. 2004.
MADR. 2006. Estadísticas cultivo de maracuyá. Anuario estadístico de frutas y hortalizas. Ministerio de Agricultura y Desarrollo Rural.
MADR. 2009. Estadísticas cultivo de maracuyá. Anuario estadístico de frutas y hortalizas. Ministerio de Agricultura y Desarrollo Rural.
MAG (Ministerio de agricultura y ganadería). 1991. Aspectos Técnicos sobre Cuarenta y Cinco Cultivos Agrícolas de Costa Rica. Dirección General de Investigación y Extensión Agrícola. Ministerio de Agricultura y Ganadería. San José, Costa Rica. En la página web: http://www.mag.go.cr/bibioteca_virtual_ciencia/tec_maracuyá.pdf consultado 1-2-07.
MARULANDA, M. L., MÁRQUEZ, M. 2001. Caracterización de la diversidad Genética de Rubus glaucus Benth con Marcadores Moleculares ( RAPD ). En: Actual Biol 23 (74): 57 – 63.
MELETTI L., SOARES M., BERNACCI L. y PASSOS I. 2005. Melhoramento genético do maracujá: passado e futuro. En: Maracujá: germoplasma e melhoramento genético. Planaltina: Embrapa Cerrados. cap. 3, p. 55-78.
MORILLO, Y., MORILLO, A. C., MUÑOZ, J. E., VASQUEZ, H. D., ZAMORANO, A. 2005. Caracterización molecular con microsatélites aleatorios RAMs de la colección de mora, Rubus spp, de la Universidad Nacional de Colombia sede Palmira. Acta Agronómica 54 (2)
NEI, M. 1973. Analysis of genetic diversity in subdivided populations. Proc Nat Acad Sci USA 70: 3321-3326.
NEI, M. 1975. Molecular population genetics and evolution. North Holland, Amsterdam. NEI, M. 1978. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics 89: 583-590.

‐ 98 ‐
NELSON, P.E.; TOUSSOUN, T.A., and MARASAS, W.F.O. 1983. Fusarium species an illustrated manual for identification. Pennsylvania State University Press, University Park. P 193.
NELSON P.E., DESJARDINS A. E., PLATTNER R. D. 1993. Fumonisins, mycotoxins producen by Fusarium species: Biology chemistry, and significance. Ann. Rev. Phytopathol., 31:233-252.
NELSON, P.E.; DIGNANI, M.C., AND ANAISSIE, E.J. 1994. Taxonomy, Biology, and Clinical Aspects of Fusarium species. Clin. Microbial. Rev., Vol., 18 (2), p 157.
NOGUEIRA G., RONCATTO G., RUGGIERO C., OLIVEIRA J. y BRAGA E. 2005. Porpagacao vegetativa do maracuzaeiro- conquista de novas adesoes. Faleiro, F.G, Junqueira, NTV, Braga, MF. (eds). Maracujá, Germoplasma e Melhoramiento genético. Capitulo 14. 345-358 pp.
NYBOM, M., BARTISH, I. 2000. Effects of life history traits and sampling strategies on genetic diversity estimates obtained with RAPD markers in plants. In: perspectives in plant ecology, evolution and systematic. Washington, D.C, USA, Vol. 3. p 93-114.
OCAMPO P., J.; COPPENS D'EECKENBRUGGE, G.; RESTREPO, M. T.; SALAZAR, M. H.; CAETANO, C. M.; VILLEGAS, A. M.; Y JARVIS, A. 2007. A new Passsifloraceae species checklist for Colombia: diversity and biogeographic distribution. Biota Col. (en imprenta).
O'DONNELL K., SUTTON DA., RINALDI MG., GUEIDAN C., CROUS PW., GEISER DM. 2009. Novel multilocus sequence typing scheme reveals high genetic diversity of human pathogenic members of the Fusarium incarnatum-F. equiseti and F. chlamydosporum species complexes within the United States. J Clin Microbiol. Dec;47(12):3851-61.
OLIVE, D.M., AND BEAN P. 1999. Principles and Applications of methods for DNA based typing of microbial organisms. J. Clin. Microbiol., Vol., 37, p1661-1669.
PAREDES C., J.C. 1996. El papel de las comercializadoras frutícolas y su impacto en la normalización de la producción de maracuyá en la zona del distrito RUT. Universidad del Valle. Facultad de ciencias sociales y Economía. Cali. pp 1-44
POSSO, A. 2010, Tesis: DIVERSIDAD GENÉTICA DE ACCESIONES DE NACEDERO Trichanthera gigantea (Humb. & Bonpl.) Nees MEDIANTE RAM´S (Random Amplified Microsatellites) Universidad del Valle Facultad de Ciencias Naturales y Exactas programa Académico de Biología Santiago de Cali.

‐ 99 ‐
RESTREPO, E. 2003. Estudio de la diversidad del tomate cultivado tipo chonto Lycopersicum esculentum en las zonas productoras de Colombia. Trabajo de Maestría en Recursos Fitogenéticos Neotropicales. Universidad Nacional de Colombia, Sede Palmira.
RICO, L., B. GUERRERO, A. LÓPEZ, C. MUÑOZ, L. GUEVARA, R.G. GUEVARA, I. TORRES Y M. GONZÁLEZ. 2001. Búsqueda de resistencia natural en plantas de chile (Capsicum spp.) contra aislados del complejo fúngico que causa pudrición de raíz. En: Memorias XXVIII Congreso Nacional de Fitopatología. Querétaro, México
RODRIGUEZ, H. GOMEZ, E. Y MARMOLEJO DE LA T. 1999. Tesis de ingeniería agronómica. Alternativas de manejo de la enfermedad pudrición seca o secadera del maracuyá Passiflora var. flavicarpa en el zona norte del Valle del Cauca. Universidad Nacional de Colombia sede Palmira. 76 p.
RODRIGUEZ R., H. 1997. Prediagnostico y evaluación de la enfermedad secadera del maracuyá Passiflora var. flavicarpa en zonas del norte del Valle y Cauca. Universidad Nacional de Colombia. Sede Palmira. Problema especial 31 p.
SAMSON, R. A.; HOEKSTRA, E.S. AND FRISVAD, J.C. 2004. Introduction to food borne fungi, 7th edition. Centraalbureau voor Schimmecultures, Ultrecht.
SANCHEZ M. 2003. Tesis Doctoral: Actividad Biológica en la Rizosfera del Maracuyá Passiflora var. flavicarpa en diferentes sistemas de manejo, estados de desarrollo y condiciones fitosanitarias. Universidad Politécnica de Madrid. Escuela Técnica superior de Ingenieros Agrónomos. P. 76-70.
SANABRIA, H., GARCÍA, M., DÍAZ, H., MUÑOZ, J.E. 2006. Caracterización molecular con marcadores RAMs de árboles nativos de Psidium guajava (guayaba) en el Valle del Cauca. Acta Agronómica 55 (1): 23-30.
SHARMA R., VILELA N. y GOMES A. 2002. Comportamento de Espécies e Híbrido de Passiflora em Relação ao Nematóide-dasgalhas, Meloidogyne arenaria. Boletim de Pesquisa e Desenvolvimento 29. Embrapa Cerrados. ISSN 1676-918X. 11p.
SINGH, K.; FRISVAD, J; TRAEN, J Y MATHUR, S. 1991 “An Illustrated Manual en Identification of some Seed- borne Aspergilli; Fusarium; Penicillia and their Mycotoxins”. Danish Goverment Institute of Seed Pathology for Developing Countries. Demark 1ra Ed, 72-80pp, 1991.

‐ 100 ‐
SINGLETON LL, MIHAIL JD, RUSH CM. 1992. Methods for research on soilborne phytopathogenic fungi. St Paul: APS Press.
TADEU V., COSTA L., BORGES A., CHAVES C. Y FIALHO F. 2002. Produção de Mudas de Maracujazeiro-azedo por Enxertia em Estacas Herbáceas Enraizadas de Espécies de Passifloras Silvestres, Ministerio da Agricultura, Pecuária e abastecimento, ISSN 1517-1469 (Comunicado 70)
TAMAYO PJ. 1999. Informe Técnico: Estudio para el control de la secadera (Nectria haematococca berk. & Br.) de la granadilla (Passiflora ligularis Juss.): Evaluación de patrones existentes y prácticas de manejo integrado. Corporación Colombiana de Investigación Agropecuaria CORPOICA.
TORRES C., SÁNCHEZ M DE P., BRAVO N., DE LA TORRE F., GÓMEZ E. D. 1999. Enfermedades fungosas y bacterianas en el cultivo del maracuyá Passiflora edulis var. flavicarpa. Editorial Nurani. 4-9 p.
TORRES C., SANCHEZ., GÓMEZ E. Y BRAVO N., MARMOLEJO F., RODRÍGUEZ H., DÍAZ M., MENESES S., PANESSO F. Y ÄGREDO M. 2000. Enfermedades fungosas y bacterianas en el cultivo de maracuyá Passiflora edulis Sims var flavicarpa en dos agroecosistemas. Fitopatología Colombiana 24(2): 47-54.
UDEA. 2008. Pulpas de frutas tropicales. Producción, transformación y Comercialización. Maracuyá. En la web: http://huitoto.udea.edu.co/FrutasTropicales/maracuyácomer.html. Consultado 1-2-08.
ULMER, T. Y MACDOUGAL, J. 2004. Passiflora. Passions flowers of the world. Portland, Oregon. Timber Press. 430 p.
URIBE, L. 1972. Catálogo ilustrado de las plantas de Cundinamarca. Vol. 15. Universidad Nacional, Instituto de Ciencias Naturales. Bogotá Colombia. 165 p.
VECCHIA, M. 2000. IL genere Passiflora. http://www.Passiflora.it/. 7 p.
VILELA N., DE RIBAMAR J., DE OLIVEIRA A., DA COSTA R., GOMES A. 2002. Comportamento em Relação às Doenças e Desempenho Agronômico de Cultivares de Maracujazeiro-azedo Cultivadas sem Agrotóxicos no Distrito Federal. Embrapa Cerrados. ISSN 1676-918X. 20p.

‐ 101 ‐
WASSENAAR, T.M, AND NEWELL, D.G. 2000. Genotyping of Campylobacter spp. Appl. Environ. Microbiol. Vol., 66, p 1-9.
WHITE, T.J; BRUNS, T.; LEE, S., AND TAYLOR, J. 1990. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics, p 315-322 in: Inns, M. A; Gelfand, D. H.; Sninsky, J. J. and White, T. J. (Eds). PCR protocols: a guide to methods and applications. Academic Press, San Diego.
YLI-MATTILA, T.; PAAVANEN-HUHTALA, S.; PARIKKA, P.; KONSTANTINOVA, P., and GAGKAEVA, T. Y. 2004. Molecular ande Morphological Diversity of Fusarium Species in Finland and North-Western Russia. Eur. J. Plant Pathol., Vol., 110 (5-6), p 573 – 585.

‐ 102 ‐
ANEXOS
ANEXO A. MEDIOS DE CULTIVO
Composición de los medios de cultivo referida a un litro de agua destilada
AGAR PATATA DEXTROSA (PDA)
39 g de bacto -.patata dextrosa agar
1000 ml de agua destilada
Se esteriliza en el autoclave durante 15 minutos a 121°C y se reparte en caja de
petri.
CARNATION LEAF AGAR (CLA)
20 g de agar
1000 ml de agua destilada
Hojas de Clavel
Las hojas de clavel se deben desinfectar con hipoclorito de sodio 5% (solución
comercial) durante 5 minutos, lavar posteriormente con agua destilada estéril y
luego con alcohol al 70% por 1 minuto, se repite el lavado con agua estéril y
posteriormente se deja secar bajo la cámara de flujo antes de agregarlas al medio
o secarlas en a 70 grados ºC.
Agregar de 3 a 5 hojas de clavel, cuando el medio esté tibio.
Medidas de las hojas: 5-8mm (1cm de largo aprox.) por placa de 9mm de
diámetro.

‐ 103 ‐
ANEXO B. Tabla 1. Aislamiento de ADN genómico
Con este método se obtienen de 50 a 100 ug de ADN por cada 100 mg de tejido liofilizado y molido.
Pasos
1 Coloque 50 mg de tejido liofilizado y molido en un tubo de 1,5 o 2 ml.
2 Agregue 800 ul de solución amortiguadora CTAB. Mezcle por inversión, para homogenizar el tejido con la solución amortiguadora.
3 Incube y mueva los tubos con suavidad en un agitador de balance en un baño a 65º C durante 90 min.
4 Retire los tubos los tubos del horno y deje enfriar durante 5 a 10 min.
5 Agregue 500 ul de FCI (Fenol cloroformo isoamilico 25:24:1) agite los tubos por inversión durante 10 min a temperatura ambiente.
6 Centrifugue a 13500 rpm a temperatura ambiente durante 5 min. para formar la parte acuosa (liquido claro de color amarillo) y la fase orgánica (de color verde oscuro)
7 Recupere aproximadamente 750 ul de la fase superior acuosa y vacíela en un tubo nuevo de 1.5 o 2.0 ml.
8 Agregar 500 ul CI (Cloroformo isoamilico 24:1) agitar 5 min. Centrifugar 5 min a 13500 rpm. Recuperar la fase acuosa en un tubo nuevo.
9 Agregue ½ volumen de isopropanol (2-propanol) al 100% previamente enfriado en un refrigerados a -20 ºC. Mezcle por inversiòn para favorecer la precipitación de ADN. Incube las muestras toda la noche a -20ºC.
10 Centrifugue a 13500 rpm a temperatura ambiente durante 10 min. para precipitar y formar la pastilla de ADN en el fondo del tubo. Deseche el isopropanol por decantación.
11
Agregue 1 ml de alcohol al 75% lave suavemente la pastilla de ADN. Centrifugue a 13500 por 1 min. Deseche el alcohol por decantación. Deje q se evapore a temperatura ambiente hasta que la pastilla seque. Si todavía siente olor a alcohol, esto indica que la pastilla no esta completamente seca.
12 Vuelva a suspender la pastilla en 100 ul de TE o agua doble destilada. Guarde las muestras a 4ºC hasta utilizarlas; si no va utilizarlas mucho tiempo, almacénelas a -20ºC.

‐ 104 ‐
ANEXO C. Tabla 2. Evaluación de la resistencia de 28 introducciones de 17 especies de Passifloras.
Especies e introducciones
evaluadas Evaluaciones Semanales
P. edulis var edulis (36)
1 2 3 4 5 6 7 8 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0,2 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0,2 0,2 0 0 0 0 0 0 0 0 0 0 0 0 0,4 0,4 0,4 0,4 0 0 0 0 0,2 0,2 0,2 0,2 0 0 0 0 0,5 0,5 0,5 0,5 0 0 0 0 0,1 0,1 0,1 0,1 0 0 0 0 0,1 0,1 0,1 0,1 0 0 0 0 0,1 0,1 0,1 0,1 0 0 0 0 0 0 0 0 0 0 0 0 0,1 0,1 0,1 0,1 0 0 0 0 0 0 0 0
P. maliformis (30)
0 0 0 0 0,2 0,2 0,5 2 0 0 0 0 0 0,2 0,5 0,5 0 0 0 0 0 0 0 1,5 0 0 0 0 0 0 0 1 0 0 0 0 0,2 0,2 0,5 1,5 0 0 0 0,2 0,8 1 2 2 0 0 0 0 0,2 0,2 0,5 1,5 0 0 0 0,3 0,4 0,5 0,7 0,7 0 0 0 0 0 0,2 0,5 1 0 0 0 0 0 0,2 0,3 0,3 0 0 0 0 0 0 0 0 0 0 0 0,1 0,1 0,1 0,1 0,1 0 0 0 0 0 0 0 0,7 0 0 0 0 0 0 0,2 0,5 0 0 0 0 0 0 0,3 0,3 0 0 0 0 0 0,1 0,1 0,2 0 0 0 0 0 0 0,3 0,3 0 0 0 0 0,3 0,3 0,5 0,5 0 0 0 0 0 0 0,2 0,2 0 0 0 0 0 0 0,2 1
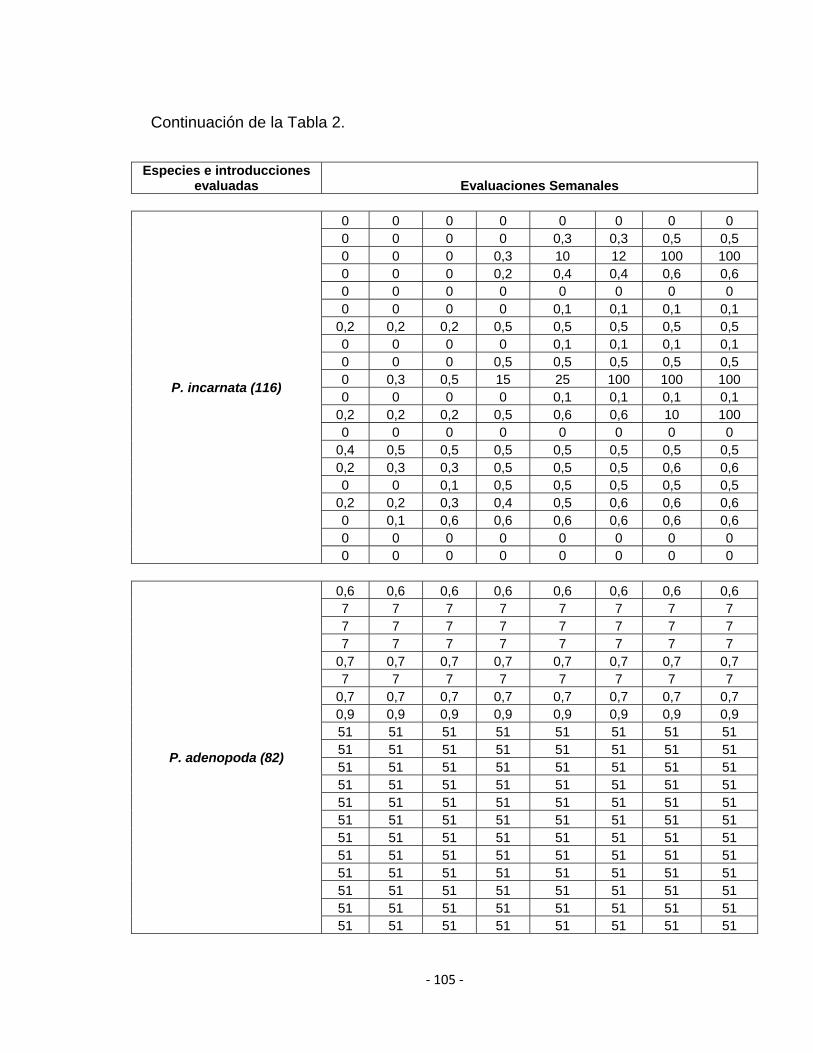
‐ 105 ‐
Continuación de la Tabla 2.
Especies e introducciones evaluadas Evaluaciones Semanales
P. incarnata (116)
0 0 0 0 0 0 0 0 0 0 0 0 0,3 0,3 0,5 0,5 0 0 0 0,3 10 12 100 100 0 0 0 0,2 0,4 0,4 0,6 0,6 0 0 0 0 0 0 0 0 0 0 0 0 0,1 0,1 0,1 0,1
0,2 0,2 0,2 0,5 0,5 0,5 0,5 0,5 0 0 0 0 0,1 0,1 0,1 0,1 0 0 0 0,5 0,5 0,5 0,5 0,5 0 0,3 0,5 15 25 100 100 100 0 0 0 0 0,1 0,1 0,1 0,1
0,2 0,2 0,2 0,5 0,6 0,6 10 100 0 0 0 0 0 0 0 0
0,4 0,5 0,5 0,5 0,5 0,5 0,5 0,5 0,2 0,3 0,3 0,5 0,5 0,5 0,6 0,6 0 0 0,1 0,5 0,5 0,5 0,5 0,5
0,2 0,2 0,3 0,4 0,5 0,6 0,6 0,6 0 0,1 0,6 0,6 0,6 0,6 0,6 0,6 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
P. adenopoda (82)
0,6 0,6 0,6 0,6 0,6 0,6 0,6 0,6 7 7 7 7 7 7 7 7 7 7 7 7 7 7 7 7 7 7 7 7 7 7 7 7
0,7 0,7 0,7 0,7 0,7 0,7 0,7 0,7 7 7 7 7 7 7 7 7
0,7 0,7 0,7 0,7 0,7 0,7 0,7 0,7 0,9 0,9 0,9 0,9 0,9 0,9 0,9 0,9 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51 51

‐ 106 ‐
Continuación Tabla 2.
Especies e introducciones
evaluadas Evaluaciones Semanales
P. maliformis pura (93)
15 15 100 100 100 100 100 100
0 1 1 1 1 15 100 100
0 1 12 20 100 100 100 100
15 15 100 100 100 100 100 100
0,5 0,5 10 15 100 100 100 100
90 90 100 100 100 100 100 100
0 0 0 100 100 100 100 100
15 15 100 100 100 100 100 100
0 0 1 1 15 50 100 100
1 12 12 100 100 100 100 100
0 0 0 0 0 0,2 0,2 0,2
12 12 100 100 100 100 100 100
51 51 100 100 100 100 100 100
0 0,3 1 20 50 50 100 100
1 1 1 15 50 75 100 100
15 15 50 90 100 100 100 100
0 0 1 1 15 15 100 100
0 0 0 100 100 100 100 100
0,5 0,5 10 25 25 50 100 100
0,7 0,7 2 2 5 5 5 5
P. alata (80)
30 50 100 100 100 100 100 100
40 100 100 100 100 100 100 100
60 100 100 100 100 100 100 100
0 15 100 100 100 100 100 100
0,2 0,3 100 100 100 100 100 100
0 0 0 15 100 100 100 100
70 100 100 100 100 100 100 100
0 100 100 100 100 100 100 100

‐ 107 ‐
Continuación Tabla 2.
Especies e introducciones
evaluadas Evaluaciones Semanales
P. quadrangularis (119)
0 10 10 12 12 100 100 100
0 0 0 0,5 0,5 1 1,5 1,5
0 0 0 0 0 0 0 0
0 0,5 0,5 0,5 10 12 12 12
10 10 15 15 15 15 15 15
0,5 0,6 19 100 100 100 100 100
0,5 0,5 1 2 3 3 5 5
0 0 0,9 100 100 100 100 100
12 12 12 12 12 12 12 12
0 0 0 10 100 100 100 100
0 0 0 0 0 0,5 0,5 1
0 0 0 0 0 0 0 0
0,7 0,7 0,7 1 100 100 100 100
0 0 15 15 100 100 100 100
0 10 100 100 100 100 100 100
10 10 10 20 100 100 100 100
0 0 0 0 0 0 0 0
0 0 0 0 0 0,5 0,5 1
0 0 0 0 0 15 100 100
0 0 0 0,5 1 1 1 1
P. ligularis (40)
0,6 0,9 0,9 0,9 1,5 1,5 1,5 1,5
0,5 0,5 0,5 0,6 0,6 0,6 0,6 0,6
0,5 0,5 0,5 0,5 1 1 1 1
0,2 0,5 0,5 0,6 0,6 0,6 1 1
0,2 7 7 7 7 7 7 7
0,5 0,6 0,6 1 1 1 1 1
0,9 1 1 1 1 1 1 1
0,5 0,5 0,6 0,6 1 1 1 1
P. ambigua (56)
1,5 2 90 100 100 100 100 100
2 2 15 100 100 100 100 100
2 100 100 100 100 100 100 100
1,5 2 70 100 100 100 100 100
2,5 2,5 50 100 100 100 100 100
2 2 5 100 100 100 100 100

‐ 108 ‐
Continuación Tabla 2.
Especies e introducciones
evaluadas Evaluaciones Semanales
P. gracilis(59)
2 2 100 100 100 100 100 100
2 3 3 3 3 3 3 3
10 100 100 100 100 100 100 100
2 2 2 2 2 100 100 100
50 100 100 100 100 100 100 100
2 2 2 2 2 2 2 2
P. falvicarpa 81
0,5 15 50 100 100 100 100 100
2 10 100 100 100 100 100 100
2 20 25 100 100 100 100 100
4 25 35 100 100 100 100 100
2 25 25 100 100 100 100 100
3 15 100 100 100 100 100 100
P. liguralis (21)
0 0 100 100 100 100 100 100
0,9 1 1 100 100 100 100 100
0,1 0,5 0,3 0,3 0,4 1 1 1
0,9 1,5 1,5 1,5 1,5 2 2 2
0 0 0 0 0 0 0 0
P. cincinata Jabotijaba (99)
0 0 0 0 0 0 0 0
0 0 0 0 0 0 0 0
0 0 0 0 0 0 0 0
0 0 0 0 0 0 0 0
P. mollissima (43)
0,5 0,5 0,5 1 1 1 1 1
0 0 0 0 0 0 0 0
1,5 1,5 2 2 2 2 2 2
0,7 0,9 0,9 1,5 1,5 2 2 2
1,7 1,7 1,7 2 2 2 2 2
0,2 0,2 0,2 0,9 0,9 1 1 1
0,2 0,2 0,9 1,5 1,5 1,5 1,5 1,5
0 0 0,5 0,5 0,5 0,5 0,5 0,5
0 10 25 100 100 100 100 100
1 1 1 1 1,5 1,5 2 2

‐ 109 ‐
Continuación Tabla 2.
Especies e introducciones
evaluadas Evaluaciones Semanales
P. edulis purpura (133)
0,5 0,5 2 2,5 2,5 2,5 2,5 2,5
0 0 0 0,7 1 1 1 1
0 0 0 0 0,2 0,2 0,2 0,2
0 0 0 0 0 0 0 0
0,6 0,6 1 1 1 1 1 1
0,5 0,5 0,9 0,9 1,5 1,5 1,5 1,5
0 0 0 0,5 0,5 0,5 0,5 0,5
0,7 0,7 1 1 1,5 1,5 1,5 1,5
0 0 0 0,7 1 1 1 1
0 0,5 0,5 1 1 1,5 1,5 1,5
0,5 0,6 0,9 1 1 1 1 1
0 0 0,5 0,9 1,5 1,5 1,5 1,5
0,5 0,5 0,5 1 1 1 1 1
0,5 0,5 0,5 1 1 1 1 1
0,5 0,5 0,5 1 1 1 1 1
P. tarminiana (73)
7 7 100 100 100 100 100 100
15 15 100 100 100 100 100 100
9 9 100 100 100 100 100 100
8 8 8 50 70 100 100 100
15 15 100 100 100 100 100 100
7 7 100 100 100 100 100 100
P. tarminiana (70)
15 15 20 20 50 60 100 100
10 100 100 100 100 100 100 100
7 8 8 100 100 100 100 100
7 8 8 8 15 25 50 100
8 8 10 10 15 20 50 100
15 15 20 100 100 100 100 100
P. rubra (85)
0 0 0 0 0 0 0 0
0 0 0 0 0 0 0 0
0 0 0 0 0 0 0 0
1 1 1 1 1 1 1 1
0 0 0 0 0 0 0 0

‐ 110 ‐
Continuación Tabla 2.
Especies e introducciones
evaluadas Evaluaciones Semanales
P. foetida (47)
0 0 0 0 0 0 0 0
0,1 0,1 0,1 1 1 1 1 1
0,5 0,5 0,5 1 1 1 1 1
0 0 0 0 0 0 0 0
P. vitifolia (87) 1 1 1 1 1 1 1 1
0 0 100 100 100 100 100 100
P. alata (20) 0 0 0,5 0,7 0,7 0,7 0,7 0,7
0,9 0,9 1 1 1,5 1,5 1,5 1,5
P. alata (79) 0 0 0 0 0 0 0 0
0 0,5 0,5 1 1 1 1 1
P. capsularis (112) 0 0 0 0,5 0,5 1 1 1
0 0 0,5 0,5 0,5 0,5 0,5 0,5
P. caerulea (57) 0 0 0 0 0 0 0 0
0 0 0 0 0 0 0 0
P. tarminiana (45)
7 100 100 100 100 100 100 100
6 100 100 100 100 100 100 100
5 100 100 100 100 100 100 100
5 100 100 100 100 100 100 100
10 100 100 100 100 100 100 100
5 100 100 100 100 100 100 100
P. quadrangularis (117)
0,2 0,2 0,2 0,2 0,2 0,2 0,2 0,2
0 0 0 0 0 0 0 0
0,2 0,2 0,2 0,2 0,2 0,2 0,2 0,2
0 0 0 0 0 0 0 0
0,2 0,2 0,2 0,2 0,2 0,2 0,2 0,2
0 0 0 0 0 0 0 0

‐ 111 ‐
Continuación Tabla 2.
Especies e introducciones
evaluadas Evaluaciones Semanales
P. alata (49)
0 0 0 0 0 0 0 0
0 0 0 0 0 0 0 0
0 0 0 0 0 0 0 0
0,3 0,3 0,3 0,3 0,3 0,3 0,5 0,5
0 0 0 0 0 0 0 0
0 0 0 0 0 0 0 0
0 0 0 0 0 0 0 0
0 0 0 0 0 0 0 0
0,3 0,3 0,3 0,3 0,3 0,3 0,3 0,3
P. quadrangularis (114)
0 0 0 0 0,2 0,2 0,2 0,2
0 0 0 0 0 0 0 0
0 0 0 0 0 0 0 0
0 0 0 0 0 0 0 0
0,5 0,5 0,5 0,5 0,5 0,5 0,5 0,5
10 100 100 100 100 100 100 100
0,2 0,6 0,6 0,6 0,6 0,6 0,6 0,6
0 0 0 0 0 0 0 0
0 0,1 0,1 0,1 0,1 0,1 0,1 0,1
0 0,5 0,5 0,5 0,5 0,5 0,5 0,5
0 0 0,7 0,7 0,7 0,7 0,7 0,7
0 0 0 0 0 0 0 0
0 0 0 0 0 0 0 0
0 0 0 0 0 0 0 0
0 0 0 0 0 0 0 0
0,3 0,5 0,5 0,5 0,5 0,5 0,5 0,5
0 0 0 0 0 0 0 0
0 0 0,2 0,2 0,2 0,2 0,2 0,2
0 0,6 0,6 0,6 0,6 0,6 0,6 0,6
0 0 0 0 0 0 0 0

‐ 112 ‐
Continuación Tabla 2.
Especies e introducciones evaluadas Evaluaciones Semanales
P. malimorfis blanda (121)
0,5 0,5 0,5 0,5 0,5 0,5 0,5 0,5 0 0 0,2 0,2 0,2 0,2 0,2 0,2 0 0 0 0 0 0 0 0 0 0 0 0 0 0,7 1 1
0,7 0,7 1 1 1 1 1 1 0 0 0 0 0 0 0 0
0,6 0,6 0,6 0,6 1 1 1 1 0,7 0,7 0,7 0,7 1 1 1 1 0,5 0,5 0,5 0,5 0,5 0,5 0,5 0,5 0,2 0,2 0,3 0,3 0,3 0,3 0,3 0,3 0,5 0,5 2 2,5 2,5 2,5 2,5 2,5 0,6 0,6 0,7 0,7 0,7 0,7 0,7 0,7 0,5 0,5 0,5 0,5 0,5 0,5 0,5 0,5 0 0 0 0 0 0 0 0 0 0 0 0 0,2 0,2 0,2 0,2 0 0 0 0 0,2 0,2 0,2 0,2 0 0 0,3 0,4 0,4 0,6 0,6 0,6
0,2 0,2 0,2 0,2 0,5 0,5 0,5 0,5 0,5 0,5 0,5 0,5 0,5 0,5 0,5 0,5 0,5 0,5 0,7 0,9 0,9 1 1 1
P. maliformis (125)
0 0 0,2 0,2 0,2 0,2 0,2 0,2 0 0 0,7 0,7 0,7 0,7 0,7 0,7
0,5 0,5 0,5 0,5 0,5 0,5 0,5 0,5 0,5 0,5 0,5 0,5 0,5 0,5 0,5 0,5 0 0 0 0 0 0 0 0 0 0,5 0,6 0,6 0,6 0,6 0,6 0,6 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0,1 0,3 0,3 0,3 0,3 0,3 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0,3 0,3 0,3 0,3 0,3 0,3 1 0 0,2 0,2 0,2 0,2 0,2 0,2
0,2 0,2 0,2 0,2 0,2 0,2 0,2 0,2 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0,2 0,2 0,2 0,2 0,2 0,2

‐ 113 ‐
Continuación Tabla 2.
Especies e introducciones
evaluadas Evaluaciones Semanales
P. edulis purpura (133)
0,5 0,5 0,5 0,5 0,5 0,5 0,5 0,5 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
0,2 0,2 0,2 0,2 0,2 0,2 0,2 0,2 0,5 0,5 0,5 0,5 0,5 0,5 0,5 0,5 0 0 0,3 0,3 0,3 0,3 0,3 0,3
0,5 0,5 0,5 0,5 0,5 0,5 0,5 0,5 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
0,5 0,5 0,5 0,5 0,5 0,5 0,5 0,5 0,2 0,5 0,5 0,5 0,5 0,5 0,5 0,5 0 0,6 0,7 0,7 0,7 0,7 0,7 0,7
0,2 0,3 0,3 0,3 0,3 0,3 0,3 0,3 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
0,2 0,2 0,2 0,2 0,2 0,2 0,2 0,2 0,6 0,6 0,7 0,7 0,7 0,7 0,7 0,7
P. apoda (58)
0,5 0,5 2 2 2,5 2,5 2,5 2,5 0 2 2 2 2 2,5 2,5 2,5 2 3 100 100 100 100 100 100 2 2 5 100 100 100 100 100
P. maliformis (107)
0 0 0 0 0 0 0 0 2,5 2,5 2,5 2,5 2,5 2,5 2,5 2,5 0 0,2 0,2 0,2 0,2 0,2 0,2 0,2
0,5 0,5 0,5 0,5 0,5 0,5 0,5 0,5 0,5 0,5 0,5 0,5 0,5 0,5 0,5 0,5 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
0,5 0,5 15 15 15 15 100 100 0 0 0 0 0 0 0 0 0 0,3 0,7 0,7 0,7 0,7 0,7 0,7 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
0,5 0,5 0,5 0,5 0,5 0,5 0,5 0,5 0,5 0,5 0,5 0,5 0,5 0,5 0,5 0,5 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0

‐ 114 ‐
ANEXO D. Tabla 3. Secuencia de los 13 Aislamientos.
AISLADO DEL
HONGO ESPECIE SECUENCIA
MUC1 Fusarium
solani
GGAGGGATCATTACCGAGTTATACAACTCATCAACCCTGTGAACATACCTATAACGTTGCCTCGGCGGGAACAGACGGCCCCGTAACACGGGCCGCCCCCCGCCAGAGGACCCCCTAACTCTGTTTCTATAATGTTTCTTCTGAGTAAACAAGCAAATAAATTAAAACTTTCAACAACGGATCTCTTGGCTCTGGCATCGATGAAGAACGCAGCGAAATGCGATAAGTAATGTGAATTGCAGAATTCAGTGAATCATCGAATCTTTGAACGCACATTGCGCCCGCCAGTATTCTGGCGGGCATGCCTGTTCGAGCGTCATTACAACCCTCAGGCCCCCGGGCCTGGCGTTGGGGATCGGCGGAAGCCCCCTGCGGGCACAACGCCGTCCCCCAAATACAGTGGCGGTCCCGCCGCAGCTTCCATTGCGTAGTAGCTAACACCTCGCAACTGGAGAGCGGCGCGGCCACGCCGTAAAACACCCAAC
MUC2 Fusarium
solani
GTACCTGCGGAGGGATCATTACCGAGTTATACAACTCATCAACCCTGTGAACATACCTATAACGTTGCCTCGGCGGGAACAGACGGCCCCGTAACACGGGCCGCCCCCGCCAGAGGACCCCCTAACTCTGTTTCTATAATGTTTCTTCTGAGTAAACAAGCAAATAAATTAAAACTTTCAACAACGGATCTCTTGGCTCTGGCATCGATGAAGAACGCAGCGAAATGCGATAAGTAATGTGAATTGCAGAATTCAGTGAATCATCGAATCTTTGAACGCACATTGCGCCCGCCAGTATTCTGGCGGGCATGCCTGTTCGAGCGTCATTACAACCCTCAGGCCCCCGGGCCTGGCGTTGGGGATCGGCGGAAGCCCCCTGCGGGCACAACGCCGTCCCCCAAATACAGTGGCGGTCCCGCCGCAGCTTCCATTGCGTAGTAGCTAACACCTCGCAACTGGAGAGCGGCGCGGCCACGCCGTAAAACACCCAACTTCTGAATGTTGACCTCGAATCAGGTAGGAATACCCGCTGAACTTAA

‐ 115 ‐
Continuación Tabla 3
AISLADO DEL
HONGO ESPECIE SECUENCIA
MUC3 Fusarium
oxysporum
GCGGAGGGATCATTACCGAGTTTACAACTCCCAAACCCCTGTGAACATACCACTTGTTGCCTCGGCGGATCAGCCCGCTCCCGGTAAAACGGGACGGCCCGCCAGAGGACCCCTAAACTCTGTTTCTATATGTAACTTCTGAGTAAAACCATAAATAAATCAAAACTTTCAACAACGGATCTCTTGGTTCTGGCATCGATGAAGAACGCAGCAAAATGCGATAAGTAATGTGAATTGCAGAATTCAGTGAATCATCGAATCTTTGAACGCACATTGCGCCCGCCAGTATTCTGGCGGGCATGCCTGTTCGAGCGTCATTTCAACCCTCAAGCACAGCTTGGTGTTGGGACTCGCGTTAATTCGCGTTCCTCAAATTGATTGGCGGTCACGTCGAGCTTCCATAGCGTAGTAGTAAAACCCTCGTTACTGGTAATCGTCGCGGCCACGCCGTTAAACCCCAACTTCTGAATGTTGACCTCGGATCAGGTAGGAATACCCGCTGAACTTAAGCA
MUC4 Fusarium
solani
GAGGGATCATTACCGAGTCTAAACAACTCATCAACCCTGTGAACATACCTAAAACGTTGCTTCGGCGGGAACAGACGGCCCCGTAAAACGGGCCGCCCCCGCCAGAGGACCCCTAACTCTGTTGCTATATGTATCTTCTGAGTAAACAAGCAAATAAATTAAAACTTTCAACAACGGATCTCTTGGCTCTGGCATCGATGAAGAACGCAGCGAAATGCGATAAGTAATGTGAATTGCAGAATTCAGTGAATCATCGAATCTTTGAACGCACATTGCGCCCGCCAGTATTCTGGCGGGCATGCCTGTTCGAGCGTCATTACAACCCTCAGGCCCCCGGGCCTGGCGTTGGGGATCGGCGAGGCGCCCCCTGTGGGCACGCGCCGTACCCCCAAATACAGTGGCGGTCCCGCCGCAGCTTCCATTGCGTAGTAGCTAACACCT
MUC5 Fusarium
oxysporum
TTTGCGGAGGGATCATTACCGAGTTTACAACTCCCAAACCCCTGTGAACATACCACTTGTTGCCTCGGCGGATCAGCCCGCTCCCGGTAAAACGGGACGGCCCGCCAGAGGACCCCTAAACTCTGTTTCTATATGTAACTTCTGAGTAAAACCATAAATAAATCAAAACTTTCAACAACGGATCTCTTGGTTCTGGCATCGATGAAGAACGCAGCAAAATGCGATAAGTAATGTGAATTGCAGAATTCAGTGAATCATCGAATCTTTGAACGCACATTGCGCCCGCCAGTATTCTGGCGGGCATGCCTGTTCGAGCGTCATTTCAACCCTCAAGCACAGCTTGGTGTTGGGACTCGCGTTAATTCGCGTTCCTCAAATTGATTGGCGGTCACGTCGAGCTTCCATAGCGTAGTAGTAAAACCCTCGTTACTGGTAATCGTCGCGGCCACGCCGTTAAACCCCCAACTTCTGAATGTTGACCTCGGATCAGGTAGGAATACCCGCTGAACTTAAGCA
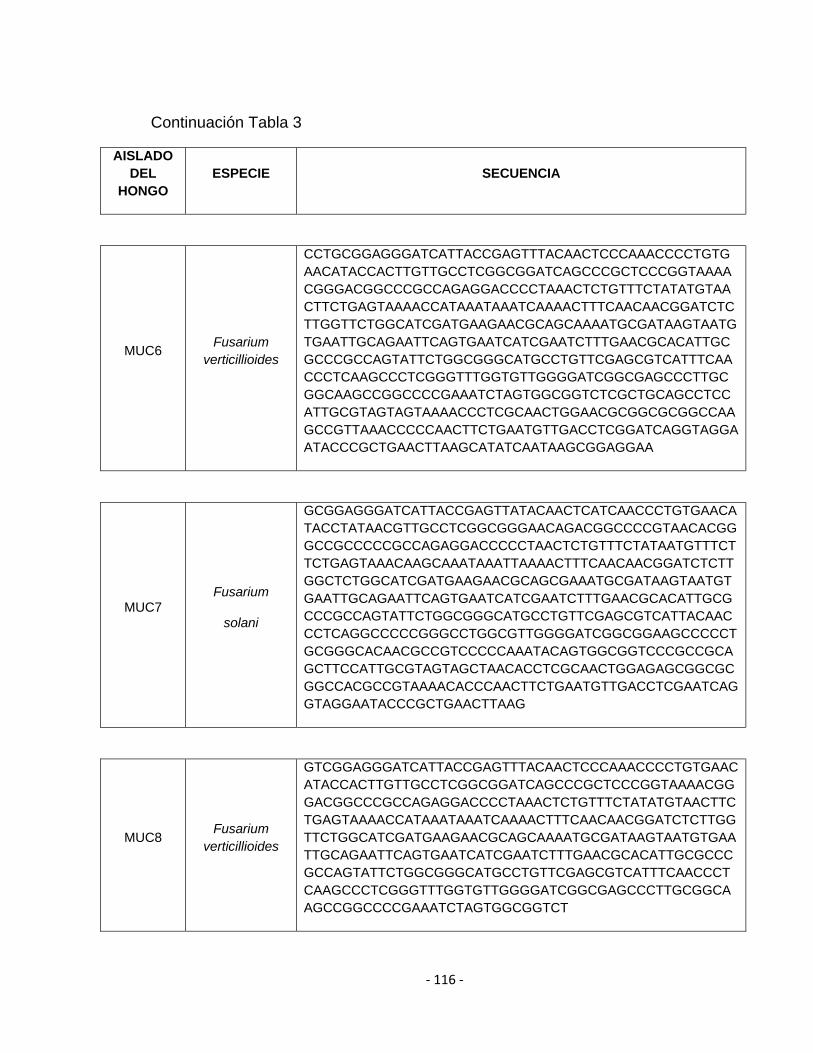
‐ 116 ‐
Continuación Tabla 3
AISLADO DEL
HONGO ESPECIE SECUENCIA
MUC6 Fusarium
verticillioides
CCTGCGGAGGGATCATTACCGAGTTTACAACTCCCAAACCCCTGTGAACATACCACTTGTTGCCTCGGCGGATCAGCCCGCTCCCGGTAAAACGGGACGGCCCGCCAGAGGACCCCTAAACTCTGTTTCTATATGTAACTTCTGAGTAAAACCATAAATAAATCAAAACTTTCAACAACGGATCTCTTGGTTCTGGCATCGATGAAGAACGCAGCAAAATGCGATAAGTAATGTGAATTGCAGAATTCAGTGAATCATCGAATCTTTGAACGCACATTGCGCCCGCCAGTATTCTGGCGGGCATGCCTGTTCGAGCGTCATTTCAACCCTCAAGCCCTCGGGTTTGGTGTTGGGGATCGGCGAGCCCTTGCGGCAAGCCGGCCCCGAAATCTAGTGGCGGTCTCGCTGCAGCCTCCATTGCGTAGTAGTAAAACCCTCGCAACTGGAACGCGGCGCGGCCAAGCCGTTAAACCCCCAACTTCTGAATGTTGACCTCGGATCAGGTAGGAATACCCGCTGAACTTAAGCATATCAATAAGCGGAGGAA
MUC7 Fusarium
solani
GCGGAGGGATCATTACCGAGTTATACAACTCATCAACCCTGTGAACATACCTATAACGTTGCCTCGGCGGGAACAGACGGCCCCGTAACACGGGCCGCCCCCGCCAGAGGACCCCCTAACTCTGTTTCTATAATGTTTCTTCTGAGTAAACAAGCAAATAAATTAAAACTTTCAACAACGGATCTCTTGGCTCTGGCATCGATGAAGAACGCAGCGAAATGCGATAAGTAATGTGAATTGCAGAATTCAGTGAATCATCGAATCTTTGAACGCACATTGCGCCCGCCAGTATTCTGGCGGGCATGCCTGTTCGAGCGTCATTACAACCCTCAGGCCCCCGGGCCTGGCGTTGGGGATCGGCGGAAGCCCCCTGCGGGCACAACGCCGTCCCCCAAATACAGTGGCGGTCCCGCCGCAGCTTCCATTGCGTAGTAGCTAACACCTCGCAACTGGAGAGCGGCGCGGCCACGCCGTAAAACACCCAACTTCTGAATGTTGACCTCGAATCAGGTAGGAATACCCGCTGAACTTAAG
MUC8 Fusarium
verticillioides
GTCGGAGGGATCATTACCGAGTTTACAACTCCCAAACCCCTGTGAACATACCACTTGTTGCCTCGGCGGATCAGCCCGCTCCCGGTAAAACGGGACGGCCCGCCAGAGGACCCCTAAACTCTGTTTCTATATGTAACTTCTGAGTAAAACCATAAATAAATCAAAACTTTCAACAACGGATCTCTTGGTTCTGGCATCGATGAAGAACGCAGCAAAATGCGATAAGTAATGTGAATTGCAGAATTCAGTGAATCATCGAATCTTTGAACGCACATTGCGCCCGCCAGTATTCTGGCGGGCATGCCTGTTCGAGCGTCATTTCAACCCTCAAGCCCTCGGGTTTGGTGTTGGGGATCGGCGAGCCCTTGCGGCAAGCCGGCCCCGAAATCTAGTGGCGGTCT

‐ 117 ‐
Continuación Tabla 3
AISLADO DEL
HONGO ESPECIE SECUENCIA
MUC9 Fusarium
oxysporum
TTGTTGCCTCGGCGGATCAGCCCGCTCCCGGTAAAACGGGACGGCCCGCCAGAGGACCCCTAAACTCTGTTTCTATATGTAACTTCTGAGTAAAACCATAAATAAATCAAAACTTTCAACAACGGATCTCTTGGTTCTGGCATCGATGAAGAACGCAGCAAAATGCGATAAGTAATGTGAATTGCAGAATTCAGTGAATCATCGAATCTTTGAACGCACATTGCGCCCGCCAGTATTCTGGCGGGCATGCCTGTTCGAGCGTCATTTCAACCCTCAAGCACAGCTTGGTGTTGGGACTCGCGTTAATTCGCGTTCCTCAAATTGATTGGCGGTCACGTCGAGCTTCCATAGCGTAGTAGTAAAACCCTCGTTACTGGTAATCGTCGCGGCCACGCCGTTAAACCCCAACTTCTGAATGTTGACCTCGGATCAGGTAGGAATACCCGCTGAAC
MUC10 Fusarium
oxysporum
TGACATACCACTTGTTGCCTCGGCGGATCAGCCCGCTCCCGGTAAAACGGGACGGCCCGCCAGAGGACCCCTAAACTCTGTTTCTATATGTAACTTCTGAGTAAAACCATAAATAAATCAAAACTTTCAACAACGGATCTCTTGGTTCTGGCATCGATGAAGAACGCAGCAAAATGCGATAAGTAATGTGAATTGCAGAATTCAGTGAATCATCGAATCTTTGAACGCACATTGCGCCCGCCAGTATTCTGGCGGGCATGCCTGTTCGAGCGTCATTTCAACCCTCAAGCACAGCTTGGTGTTGGGACTCGCGTTAATTCGCGTTCCTCAAATTGATTGGCGGTCACGTCGAGCTTCCATAGCGTAGTAGTAAAACCCTCGTTACTGGTAATCGTCGCGGCCACGCCGTTAAACCCCAACTTCTGAATGTTGACCTCGGATCAGGTAGGAATACCCGCTGAACTTAAGCATATCAA
MUC11 Fusarium
brachygibbosum
CTGCGGAGGGATCATTACCGAGTTTACAACTCCCAAACCCCTGTGAACATACCTTTATGTTGCCTCGGCGGATCAGCCCGCGCCCCGTAAAACGGGACGGCCCGCCGCAGGAACCACAAAACTCTGATTTTAGTGTAACTTCTGAGTCTAAAAAACAAATAAATCAAAACTTTCAACAACGGATCTCTTGGTTCTGGCATCGATGAAGAACGCAGCAAAATGCGATAAGTAATGTGAATTGCAGAATTCAGTGAATCATCGAATCTTTGAACGCACATTGCGCCCGCCAGTATTCTGGCGGGCATGCCTGTTCGAGCGTCATTTCAACCCTCAAGCCCCCGGGTTTGGTGTTGGGGATCGGGCTGTACTCCAGCCCGGCCCCGAAATCTAGTGGCGGTCTCGCTGCAGCCTCCATTGCGTAGTAGCTAACACCTCGCAACTGGAACGCGGCGCGGCCAAGCCGTTAAA

‐ 118 ‐
Continuación Tabla 3
AISLADO DEL
HONGO ESPECIE SECUENCIA
MUC12 Fusarium
solani
GCGGAGGGATCATTACCGAGTTATACAACTCATCAACCCTGTGAACATACCTATAACGTTGCCTCGGCGGGAACAGACGGCCCCGTAACACGGGCCGCCCCCGCCAGAGGACCCCCTAACTCTGTTTCTATAATGTTTCTTCTGAGTAAACAAGCAAATAAATTAAAACTTTCAACAACGGATCTCTTGGCTCTGGCATCGATGAAGAACGCAGCGAAATGCGATAAGTAATGTGAATTGCAGAATTCAGTGAATCATCGAATCTTTGAACGCACATTGCGCCCGCCAGTATTCTGGCGGGCATGCCTGTTCGAGCGTCATTACAACCCTCAGGCCCCCGGGCCTGGCGTTGGGGATCGGCGGAAGCCCCCTGCGGGCACAACGCCGTCCCCCAAATACAGTGGCGGTCCCGCCGCAGCTTCCATTGCGTAGTAGCTAACACCTCGCAACTGGAGAGCGGCGCGGCCACGCCGTAAAACACCCAACTTCTGAATGTTGACCTCGAATCAGGTAGGAATACCCGCTGAACTTAAGC
MUC13 Fusarium
solani
TGCGGAGGGATCATTACCGAGTTATACAACTCATCAACCCTGGTGACATACCTATAACGTTGCCTCGGCGGGAACAGACGGCCCCGTAACACGGGCCGCCCCCGCCAGAGGACCCCCTAACTCTGTTTCTATAATGTTTCTTCTGAGTAAACAAGCAAATAAATTAAAACTTTCAACAACGGATCTCTTGGCTCTGGCATCGATGAAGAACGCAGCGAAATGCGATAAGTAATGTGAATTGCAGAATTCAGTGAATCATCGAATCTTTGAACGCACATTGCGCCCGCCAGTATTCTGGCGGGCATGCCTGTTCGAGCGTCATTACAACCCTCAGGCCCCCGGGCCTGGCGTTGGGGATCGGCGGAAGCCCCCTGCGGGCACAACGCCGTCCCCCAAATACAGTGGCGGTCCCGCCGCAGCTTCCATTGCGTAGTAGCTAACACCTCGCAACTGGAGAGCGGCGCGGCCACGCCGTAAAACACCCAACTTCTGAATGTTGACCTCGAATCAGGTAGGAATACCCGCTGAACTTAAGC

‐ 119 ‐
Hay una fuerza motriz más poderosa que el vapor,
la electricidad y la energía atómica: la voluntad.
Albert Einstein