evolutionary and transcriptional analysis of karyopherin ... · pdf fileate nucleocytoplasmic...
TRANSCRIPT

Evolutionary and Transcriptional Analysis ofKaryopherin � Superfamily Proteins*□S
Yu Quan‡, Zhi-Liang Ji‡§, Xiao Wang‡, Alan M. Tartakoff¶, and Tao Tao‡§
In eukaryotes, karyopherin � superfamily proteins medi-ate nucleocytoplasmic transport of macromolecules. Weinvestigated the evolutionary and transcriptional patternsof these proteins using bioinformatics approaches. Noobvious homologs were found in prokaryotes, but an ex-tensive set of �-karyopherin proteins was found in yeast.Among 14 �-karyopherins of Saccharomyces cerevisiae,eight corresponded to their human orthologs directlywithout diversification, two were lost, and the remainingfour proteins exhibited gene duplications by differentmechanisms. We also identified �-karyopherin orthologsin Caenorhabditis elegans, Drosophila melanogaster,Danio rerio, Xenopus tropicalis, Gallus gallus, and Musmusculus. �-Karyopherins were ubiquitously but nonuni-formly expressed in distinct cells and tissues. In yeastand mice, the titer of some �-karyopherin transcriptsappeared to be regulated both during the cell cycle andduring development. Further virtual analysis of promoterbinding elements suggested that the transcription fac-tors SP1, NRF-2, HEN-1, RREB-1, and nuclear factor Yregulate expression of most �-karyopherin genes.These findings emphasize new mechanisms in func-tional diversification of �-karyopherins and regulation ofnucleocytoplasmic transport. Molecular & CellularProteomics 7:1254–1269, 2008.
Nucleocytoplasmic transport of proteins, RNAs, and ribo-somes is essential in eukaryotic cells. In this process, the�-karyopherins (or importin-� superfamily members) play acentral role as they transport cargoes across the nuclearpore complex (1–9). Members of this family of 95–145-kDaproteins have a so-called importin-� N-terminal (IBN_N)1
domain (or RanGTP binding domain) at their N terminus andseveral “HEAT repeat” motifs that mostly occupy the C-terminal portion of the structure. The HEAT repeat motifsare able to assume different conformations in different func-tional states. The flexibility of these HEAT repeat motifsfacilitates the accommodation of their binding partners byan induced fit type of mechanism (10). These features dis-tinguish �-karyopherins from transporters that deliver car-goes to other subcellular organelles. �-Karyopherins havebeen found from yeast to humans. In Saccharomyces cer-evisiae, 14 �-karyopherins have been shown to transportcorresponding cargoes in and out of the nucleus (11, 12),whereas in mammalian cells, about 20 �-karyopherins par-ticipate in these events (7). The family includes both importand export receptors.
Nuclear proteins larger than the “diffusion limit” of thenuclear pore complex (�50 kDa) generally have a nuclearlocalization signal for nuclear import, whereas smaller pro-teins that are actively excluded from the nucleus have anuclear export signal (3). The transport of these cargoes ismediated by karyopherins in conjunction with the smallGTPase Ran, which cycles between its GDP-bound (in thecytoplasm) and GTP-bound (in the nucleus) forms. RanGTPconfers directionality upon transport by controlling the con-formation of �-karyopherins and their ability to bind cargo(13, 14). Although most karyopherins are constitutively ex-pressed in cells (6), emerging evidence shows that expres-sion of at least some karyopherin genes is regulated (15, 16)presumably due to varying requirements for transport, e.g.during development (17). For example, expression of Kpnb1and Ranbp5 is regulated during rodent spermatogenesis(18). Moreover the expression of IPO13 in rats and humansis regulated both hormonally and developmentally (16, 19).Although �-karyopherins have common features, the bio-logical importance of their functions in nuclear transport isimpressively varied. A little explored aspect of �-karyo-pherin biology is their evolution. In particular, it is unclearhow the 14 yeast �-karyopherins gave rise to 20 members inhuman. Moreover it is unknown whether the cell type ex-pression of �-karyopherin genes reflects correspondingfunctional requirements.
To address the above questions, we used bioinformaticsapproaches. We also investigated the expression of each�-karyopherin member through the systematic analysis oftranscriptional profiles and identification of putative transcrip-tion factor binding sites.
From the ‡School of Life Sciences and Key Laboratory for CellBiology and Tumor Cell Engineering, the Ministry of Education ofChina, Xiamen University, Xiamen, Fujian 361005, China and ¶De-partment of Pathology and Cell Biology Program, Case WesternReserve University School of Medicine, Cleveland, Ohio 44106
Received, October 22, 2007, and in revised form, February 25, 2008Published, MCP Papers in Press, March 18, 2008, DOI 10.1074/
mcp.M700511-MCP2001 The abbreviations used are: IBN_N, importin-� N-terminal;
BLAST, Basic Local Alignment Search Tool; GNF, Genomics Instituteof the Novartis Research Foundation; SPM, specificity measure;TFBS, transcription factor binding site; IDs, identities; NF-Y, nuclearfactor Y; NRF-2, nuclear respiratory factor 2; RREB-1, Ras-respon-sive element-binding protein 1; HEN-1, helix-loop-helix protein 1;dpc, days postcoitus; HEAT, huntingtin, elongation factor 3, proteinphosphatase 2A, Tor1.
Research
© 2008 by The American Society for Biochemistry and Molecular Biology, Inc.1254 Molecular & Cellular Proteomics 7.7This paper is available on line at http://www.mcponline.org

EXPERIMENTAL PROCEDURES
Identification of Orthologous Proteins—The assignment of ortholo-gous proteins is critical for evolutionary analyses of �-karyopherins.For this purpose, the orthologous proteins were searched and com-pared in eight species: Homo sapiens, Mus musculus, Gallus gallus,Danio rerio, Xenopus tropicalis, Drosophila melanogaster, Caenorh-abditis elegans, and S. cerevisiae. Each assignment of orthologs wasmade on the basis of multiple lines of evidences. 1) As the funda-mental method, the putative orthologs were identified or verified byBLAST searching against assembled genomes, adopting the defaultparameters of the National Center for Biotechnology Information(NCBI) blastp for sequence alignment. 2) Most �-karyopherin or-thologs can be identified by searching the NCBI Clustering of Ortholo-gous Groups (COG) database (20). 3) The “Orthologue Prediction”function of the Ensembl database also helps to identify putativeorthologs of �-karyopherins in whole genomes (21).
Construction of Phylogenetic Trees—The protein sequences andexon information of karyopherin � superfamily members of eightorganisms were derived from either the Ensembl genome databaserelease 46 (21) or GenBankTM. Corresponding protein sequenceswere extracted from GenBank and Ensembl (supplemental Table 1).To illustrate the evolution of �-karyopherins from yeast to human,phylogenetic trees were generated. Multiple sequence alignment onprotein sequences was first demonstrated using MUSCLE3.6 (22),then refined manually with consideration of the secondary structure,and finally visualized by ClustalX 1.83 (23). 14 individual phylogenetictrees (Fig. 1) were then constructed based on the multiple alignmentsusing the neighbor-joining method provided by the software MEGA4(24) under the Poisson correction amino acid substitution model withuniform rates among sites (25, 26). Bootstrap analysis was conductedusing 1,000 replicates to test the robustness of these phylogenetictrees.
Investigation of Transcription Patterns—The gene expression mi-croarray data sets GNF1H_GCRMA and GNF1M_GCRMA used in thisstudy were retrieved from GNF SymAtlas (27), which covers 24,277human genes in 79 tissues and 32,905 mouse genes in 61 tissues.The gene expression patterns of �-karyopherins were analyzed quan-titatively as described in previous work (28). Geometric comparison(similarity measure) was used to indicate how similar two gene ex-pression profiles are. A value of similarity measure close to 1 indicateshigh similarity of gene expression patterns regardless of their expres-sion levels. Such genes may have related biological roles. Specificitymeasure (SPM) was calculated to compare expression in distincttissues (28). Such information is helpful for understanding the phys-iological significance of genes. Three different microarray data sets ofS. cerevisiae (based on treatment with � factor and analysis of cdc15and cdc28 mutants) (29, 30) were adopted to investigate possible cellcycle-dependent expression of �-karyopherins.
Investigation of Regulatory Elements—400 bp of nucleotide se-quences from �300 to �100 bp of the transcript start of �-karyo-pherin genes in eight species were derived from the Ensembl data-base (21) or GenBank for the transcription factor binding sites (TFBSs)analysis. The MAPPER search engine is a program developed to aidthe molecular biologist in determining what eukaryotic transcriptionfactor binding elements may exist in a given DNA sequence (31). Inthis study, the putative TFBSs were identified by screening the up-stream regulatory sequences of �-karyopherin genes. Default scoreand E-value thresholds (0 and 10, respectively) of the MAPPERsearch engine were specified. To reduce the false positives, onlyspecies-specific TFBS models were adopted for predictions, e.g.TFBS models built from human, mouse, or fly data were only appliedto human, mouse, or fly promoter sequences, respectively. Thesepredictions were further combined with information on gene regula-tion to identify possible regulatory patterns.
RESULTS AND DISCUSSION
Evolution of the Karyopherin � Superfamily
The Sequence Alignments—Previous studies have identi-fied 14 �-karyopherins in bakers’ yeast (S. cerevisiae) andabout 20 in human (32). A few studies that have investigated�-karyopherin divergence and multiplication during evolutionwere based on limited data (33, 34). Through ortholog analy-ses, 138 �-karyopherin genes (132 annotated and six novel)were identified in human (20 genes), S. cerevisiae (14 genes),and six other organisms: M. musculus (20 genes), G. gallus(17 genes), X. tropicalis (18 genes), D. rerio (23 genes), D.melanogaster (16 genes), and C. elegans (10 genes) (Table I).The IDs of these sequences in Ensembl or GenBank can befound in the supplemental Table 1.
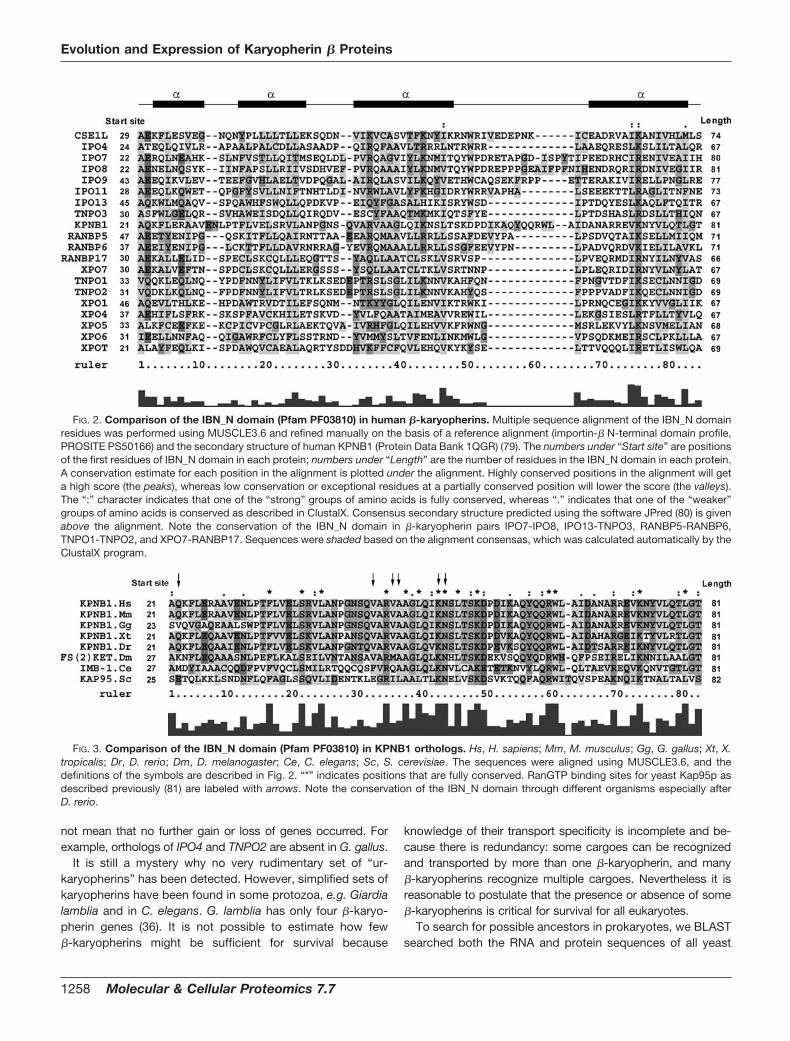
Multiple sequence alignment of full-length protein se-quences indicated the extensive diversity of �-karyopherinswithin a species (data not shown). Because the full-lengthsequence alignment within a species cannot provide an inclu-sive description of �-karyopherin evolution, local structuralclues were searched to provide complementary information.The IBN_N domain (Pfam PF03810) plays a pivotal role incargo release and thus carries distinctive “structural signa-tures” of the karyopherin � family (33, 35). We then comparedthe IBN_N domains of �-karyopherins within a species. Theconservation of IBN_N domains within a species is compar-atively poor (Fig. 2). This diversity suggests that �-karyo-pherins may not all be derived from one or a small number ofancestors. In other words, it is inappropriate to probe thekinship of �-karyopherins on the basis of multiple sequencealignment of the IBN_N domain or full-length sequence withina species. Exceptions were found in some pairs of �-karyo-pherins that share domain composition, length, and start sites(e.g. IPO7-IPO8, IPO13-TNPO3, RANBP5-RANBP6, TNPO1-TNPO2, and XPO7-RANBP17), suggesting that they areparalogous pairs. Although it is difficult to identify an alto-gether constant IBN_N domain among human �-karyo-pherins, several amino acid residues in this region are rela-tively conserved (Val/Ile/Leu46, Val/Ile/Leu74, and Lys/Arg75)(Fig. 2). These amino acids may be critical to maintain domainstructure and bind RanGTP. The comparison of IBN_N do-mains among species was also undertaken using KPNB1proteins as an example (Fig. 3). By contrast, the IBN_N do-main shows strong conservation among species, especiallyafter D. melanogaster, which agrees well with the results ofmultiple sequence alignment of full-length proteins (supple-mental materials).
The Ancestors of �-Karyopherins—On the basis of the mul-tiple sequence alignments of full-length protein sequences,the �-karyopherins were allocated to 14 separate trees withhigh confidence (with bootstrap values �50%). They areCSE1L, IPO4, IPO7-IPO8, IPO9, IPO11, IPO13-TNPO3,KPNB1, RANBP5-RANBP6, TNPO1-TNPO2, XPO1, XPO4-XPO7-RANBP17, XPO5, XPO6, and XPOT. It is interesting
Evolution and Expression of Karyopherin � Proteins
Molecular & Cellular Proteomics 7.7 1255
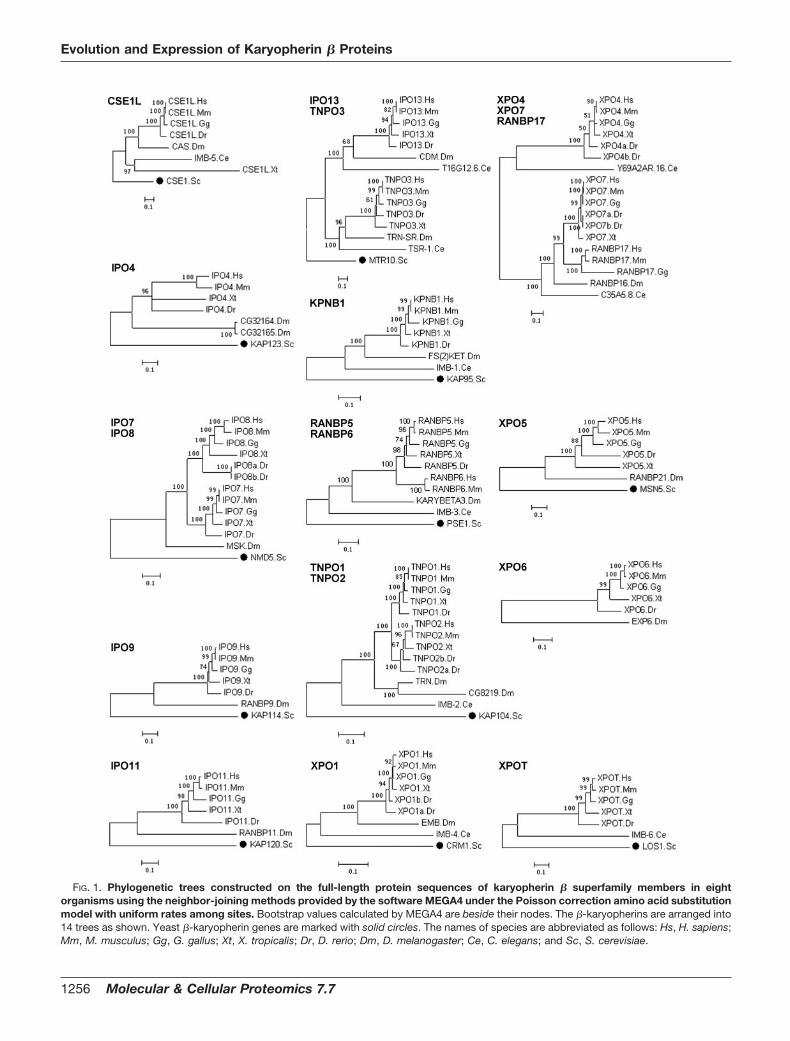
FIG. 1. Phylogenetic trees constructed on the full-length protein sequences of karyopherin � superfamily members in eightorganisms using the neighbor-joining methods provided by the software MEGA4 under the Poisson correction amino acid substitutionmodel with uniform rates among sites. Bootstrap values calculated by MEGA4 are beside their nodes. The �-karyopherins are arranged into14 trees as shown. Yeast �-karyopherin genes are marked with solid circles. The names of species are abbreviated as follows: Hs, H. sapiens;Mm, M. musculus; Gg, G. gallus; Xt, X. tropicalis; Dr, D. rerio; Dm, D. melanogaster; Ce, C. elegans; and Sc, S. cerevisiae.
Evolution and Expression of Karyopherin � Proteins
1256 Molecular & Cellular Proteomics 7.7

that these 14 phylogenetic trees do not all correspond toancestors in S. cerevisiae. As summarized by phylogeneticanalyses (Fig. 1) and ortholog analyses (Table I), only 12 yeast�-karyopherin genes have orthologs in multicellular organ-isms judging from multiple sequence alignments and phylo-genetic analysis. The other two genes, KAP122 and SXM1, donot have orthologs. Actually before the diversification of�-karyopherins stabilized, gain and loss of genes was com-mon. Thus, seven �-karyopherin genes of S. cerevisiae(KAP114, KAP120, KAP122, KAP123, MSN5, NMD5, andSXM1) are absent in C. elegans, although two new genes weregained: C35A5.8 (the possible ancestor of human RANBP17and XPO7) and Y69A2AR.16 (ancestor of human XPO4). Gainand loss were also observed in D. melanogaster whereY69A2AR.16 (ancestor of human XPO4) and LOS1 (ancestorof human XPOT) are absent, and a new �-karyopherin gene,
Exp6 (ancestor of human XPO6), emerges. These changespresumably equip distinct species with an ample but minimalrepertoire of carriers to transport critical cargoes.
Sequence analysis of seven other yeasts, Candida glabrata(14 genes), Debaryomyces hansenii (13 genes), Eremotheciumgossypii (14 genes), Kluyveromyces lactis (13 genes), Yarrowialipolytica (12 genes), Schizosaccharomyces pombe (13 genes),and Cryptococcus neoformans (nine genes), and two plants,Arabidopsis thaliana (17 genes) and Oryza sativa (14 genes),showed that the absence of selected �-karyopherin genes is notrare during evolution. The causes of gene loss may be related tounique aspects of physiology, behavior, and development. Asno new types of �-karyopherin genes were detected in D. rerio(although there are duplicates of some known �-karyopheringenes), the diversification and amplification of �-karyopheringenes became stable no later than D. rerio. However, this does
TABLE ISummary of �-karyopherin genes of eight organisms, including H. sapiens, M. musculus, G. gallus, X. tropicalis, D. rerio, D. melanogaster,
C. elegans, and S. cerevisiae
Orthologous gene groups are separated by shading. Note that IPO7 and IPO8 may come from the same common ancestor gene, NMD5,in S. cerevisiae; IPO13 and TNPO3 may come from the same common ancestor gene, MTR10, in S. cerevisiae; RANBP6 and RANBP5 maycome from the same common ancestor gene, PSE1, in S. cerevisiae; TNPO1 and TNPO2 may come from the same common ancestor gene,KAP104, in S. cerevisiae; and XPO7 and RANBP17 may come from the same common ancestor gene, C35A5.8, in C. elegans. Slashesrepresent no homologous genes found.
Evolution and Expression of Karyopherin � Proteins
Molecular & Cellular Proteomics 7.7 1257
not mean that no further gain or loss of genes occurred. Forexample, orthologs of IPO4 and TNPO2 are absent in G. gallus.
It is still a mystery why no very rudimentary set of “ur-karyopherins” has been detected. However, simplified sets ofkaryopherins have been found in some protozoa, e.g. Giardialamblia and in C. elegans. G. lamblia has only four �-karyo-pherin genes (36). It is not possible to estimate how few�-karyopherins might be sufficient for survival because
knowledge of their transport specificity is incomplete and be-cause there is redundancy: some cargoes can be recognizedand transported by more than one �-karyopherin, and many�-karyopherins recognize multiple cargoes. Nevertheless it isreasonable to postulate that the presence or absence of some�-karyopherins is critical for survival for all eukaryotes.
To search for possible ancestors in prokaryotes, we BLASTsearched both the RNA and protein sequences of all yeast
FIG. 2. Comparison of the IBN_N domain (Pfam PF03810) in human �-karyopherins. Multiple sequence alignment of the IBN_N domainresidues was performed using MUSCLE3.6 and refined manually on the basis of a reference alignment (importin-� N-terminal domain profile,PROSITE PS50166) and the secondary structure of human KPNB1 (Protein Data Bank 1QGR) (79). The numbers under “Start site” are positionsof the first residues of IBN_N domain in each protein; numbers under “Length” are the number of residues in the IBN_N domain in each protein.A conservation estimate for each position in the alignment is plotted under the alignment. Highly conserved positions in the alignment will geta high score (the peaks), whereas low conservation or exceptional residues at a partially conserved position will lower the score (the valleys).The “:” character indicates that one of the “strong” groups of amino acids is fully conserved, whereas “.” indicates that one of the “weaker”groups of amino acids is conserved as described in ClustalX. Consensus secondary structure predicted using the software JPred (80) is givenabove the alignment. Note the conservation of the IBN_N domain in �-karyopherin pairs IPO7-IPO8, IPO13-TNPO3, RANBP5-RANBP6,TNPO1-TNPO2, and XPO7-RANBP17. Sequences were shaded based on the alignment consensas, which was calculated automatically by theClustalX program.
FIG. 3. Comparison of the IBN_N domain (Pfam PF03810) in KPNB1 orthologs. Hs, H. sapiens; Mm, M. musculus; Gg, G. gallus; Xt, X.tropicalis; Dr, D. rerio; Dm, D. melanogaster; Ce, C. elegans; Sc, S. cerevisiae. The sequences were aligned using MUSCLE3.6, and thedefinitions of the symbols are described in Fig. 2. “*” indicates positions that are fully conserved. RanGTP binding sites for yeast Kap95p asdescribed previously (81) are labeled with arrows. Note the conservation of the IBN_N domain through different organisms especially afterD. rerio.
Evolution and Expression of Karyopherin � Proteins
1258 Molecular & Cellular Proteomics 7.7

FIG. 4. The exon arrangements of 20 human �-karyopherin genes. The transcript IDs in Ensembl are ENST00000261412,ENST00000356861, ENST00000379719, ENST00000256079, ENST00000252512, ENST00000377354, ENST00000265388,ENST00000372343, ENST00000290158, ENST00000354464, ENST00000361565, ENST00000389352, ENST00000389538,ENST00000255305, ENST00000265351, ENST00000304658, ENST00000389161, ENST00000262982, ENST00000357602, andENST00000259569, respectively. The shaded blocks are translated regions, whereas the transparent blocks are non-translated regions.The numbers under the blocks are the length of the exon (in bp), and the numbers in parenthesis indicate the length of translated regionswhen an exon contains a non-translated region. Note the similarity between TNPO1-TPNP2, IPO7-IPO8, and XPO7-RANBP17. RANBP6has only one exon and is thought to originate from RANBP5 through retroposition.
Evolution and Expression of Karyopherin � Proteins
Molecular & Cellular Proteomics 7.7 1259

�-karyopherins against 327 bacterial and 27 archaebacterialgenomes. Mild similarity (sequence identity, 26%) was foundfor Sxm1p to an �143-amino acid fragment (fragment ofsensory transduction histidine kinase in Clostridium acetobu-tylicum). Other �-karyopherins also show mild similarity (nomore than 30% sequence identity) to some short prokaryoticsequences, many of which are fragments of uncharacterizedhypothetical proteins. These sequences are remotely homol-ogous to HEAT repeats.
Gene Duplications—The expansion of the karyopherin �
family is accompanied by repeated gene amplification. Se-quence alignment shows that there are five pairs of human�-karyopherins (10 proteins) that closely resemble each other:IPO7-IPO8, IPO13-TNPO3, RANBP5-RANBP6, TNPO1-TNPO2, and XPO7-RANBP17. Because each pair of �-karyo-pherins may be derived from a common ancestor, exon anal-ysis within species and among species was undertaken (Figs.4 and 5 and the supplemental materials). We found that these�-karyopherin genes are well conserved over species judgingfrom both the length and nucleotide acid composition ofeach exon. Such conservation can be traced back to theearliest common ancestor, the PSE1 gene of S. cerevisiae.Comparison of exon arrangements among �-karyopheringenes indicates that for IPO7-IPO8, TNPO1-TNPO2, andXPO7-RANBP17 both members of each pair have almostthe same exon number, length, and order (Fig. 4). Thisobservation strongly supports the hypothesis that these�-karyopherin pairs are derived from the same ancestors bygene duplication.
Pairs of duplicated �-karyopherin genes were also found inD. melanogaster (CG32164-CG32165 and Trn-CG8219) andin D. rerio (ipo8a-ipo8B, tnpo2a-tnpo2b, xpo1a-xpo1b,xpo4a-xpo4b, and xpo7a-xpo7b) (Table I). These gene pairsexhibit high identity of protein sequence: 99.9% for ipo8a-ipo8b, 97.7% for CG32164-CG32165, 94.8% for xpo1a-
xpo1b, 91.9% for tnpo2a-tnpo2b, 84.1% for xpo7a-xpo7b,68.6% for Trn-CG8219, and 63.9% for xpo4a-xpo4b. Therenevertheless is no evidence that the duplications persist inhigher organisms, and some duplicated �-karyopherin genepairs may diverge to the point that they acquire specializedneofunctions (37). Thus gene duplication in D. melanogasterand D. rerio could in fact contribute to speciation (37).
Retroposition—As discussed previously, gene duplicationis one of several mechanisms that could have made it possi-ble for the karyopherin � family to expand. RANBP5 andRANBP6 are very closely related judging from sequencealignment and ortholog analysis. To draw the evolutionarypath of RANBP5 and RANBP6, their exon arrangement wasstudied. Fig. 5 shows, remarkably, that the exon numbergrows from one exon for PSE1, the putative ancestor ofhuman RANBP5 in S. cerevisiae, to 26 exons in D. rerio andfinally to 29 exons in H. sapiens. The multiplication of exonspresumably reflects the high phenotypic complexity of mam-mals (38). The enormous increase of the untranslated regions(0 bp in S. cerevisiae to about 2,709 bp in human) is likely tocontribute to regulation of gene transcription (39, 40). How-ever, unlike RANBP5 and other �-karyopherins, the RANBP6gene has only one exon in both human and mouse (Fig. 5).Further BLAST searches against the Ensembl genome data-base found that RANBP6 orthologs exist only in mammals.We therefore suggest that the emergence of RANBP6 mayresult from the retroposition of RANBP5 gene, which leads toproduction of a new functional pseudogene with only a singleexon (37). Judging from known RANBP6 orthologs, the ret-roposition of RANBP6 from ancient RANBP5 may have hap-pened before the divergence of mammals and birds. Thisinference is supported by the previous evidence that RANBP6is a functional retrocopy of RANBP5 (41). Additional orthologanalysis found that some RANBP6 orthologs contain severalexons, e.g. Canis familiaris (three exons), Oryctolagus cunic-
FIG. 5. The exon arrangements of RANBP5 and RANBP6 in different organisms, including H. sapiens (Hs), M. musculus (Mm), G.gallus (Gg), X. tropicalis (Xt), D. rerio (Dr), D. melanogaster (Dm), C. elegans (Ce), and S. cerevisiae (Sc). The transcript IDs of these�-karyopherin genes in Ensembl or GenBank are ENST00000357602, ENSMUST00000032898, XM_416978.2, ENSXETT00000050713,XM_692846.2, CG1059-RA, and C53D5.6 YMR308C for RANBP5 and ENST00000361966 and ENSMUST00000046742 for RANBP6. Theshaded blocks are translated regions, whereas the transparent blocks are non-translated regions. The numbers under the blocks are the lengthof exons, and numbers in parentheses are the length of translated regions when an exon contains a non-translated region. Note theconservation of exon arrangements of RANBP5 after D. rerio and the single exon RANBP6 in human and mouse after retroposition.
Evolution and Expression of Karyopherin � Proteins
1260 Molecular & Cellular Proteomics 7.7

ulus (six exons), and Sorex araneus (12 exons). Because noortholog was found other than in mammals, it is reasonable toconclude that the retroduplication event happened in thespeciation of mammals. After the retroposition, the RANBP6is further diversified in mammals: introns are species-specif-ically inserted into the only exon. This may explain the multipleexons of RANBP6 orthologs. Similar phenomena have beenreported previously (42, 43).
In conclusion, karyopherin � superfamily proteins are notfound in prokaryotes. �-Karyopherin evolution is suggested toproceed along 14 lines from yeast, although two genes(KAP122 and SXM1) are lost as summarized in Table I. Someyeast �-karyopherins (KAP95, KAP114, KAP120, KAP123,CSE1, MSN5, CRM1, and LOS1) can be directly linked to theirorthologs in man (KPNB1, IPO9, IPO11, IPO4, CSE1L, XPO5,XPO1, and XPOT), respectively. Other �-karyopherins (PSE1,NMD5, MTR10, and KAP104) underwent diversification duringtheir evolution.
The selective pressure for efficient communication betweenthe nucleus and the cytoplasm likely prompts the multiplica-tion and diversification of karyopherins to such an extent thatonly weak sequence homology is often observed between�-karyopherins in a species. Considering the evolutionary his-tory and functional importance of �-karyopherins, reclassifica-tion of the karyopherin � superfamily may become necessary.Classification should consider not only their overall structuralcharacteristics but their entire sequence, their cargo specificity,and their interactions with both Ran and nucleoporins.
Expressions of the Karyopherin � Superfamily in Cellsand Tissues
The functional importance of �-karyopherins determinestheir expression. Motivated by this premise, we thereforeexamined the gene expression patterns of �-karyopherinsby statistically analyzing gene expression microarray data,classifying them as comparatively high expression (SPM �
4), above average (SPM � 2), and below average (SPM � 2).�-Karyopherin genes are thought to be “housekeeping”
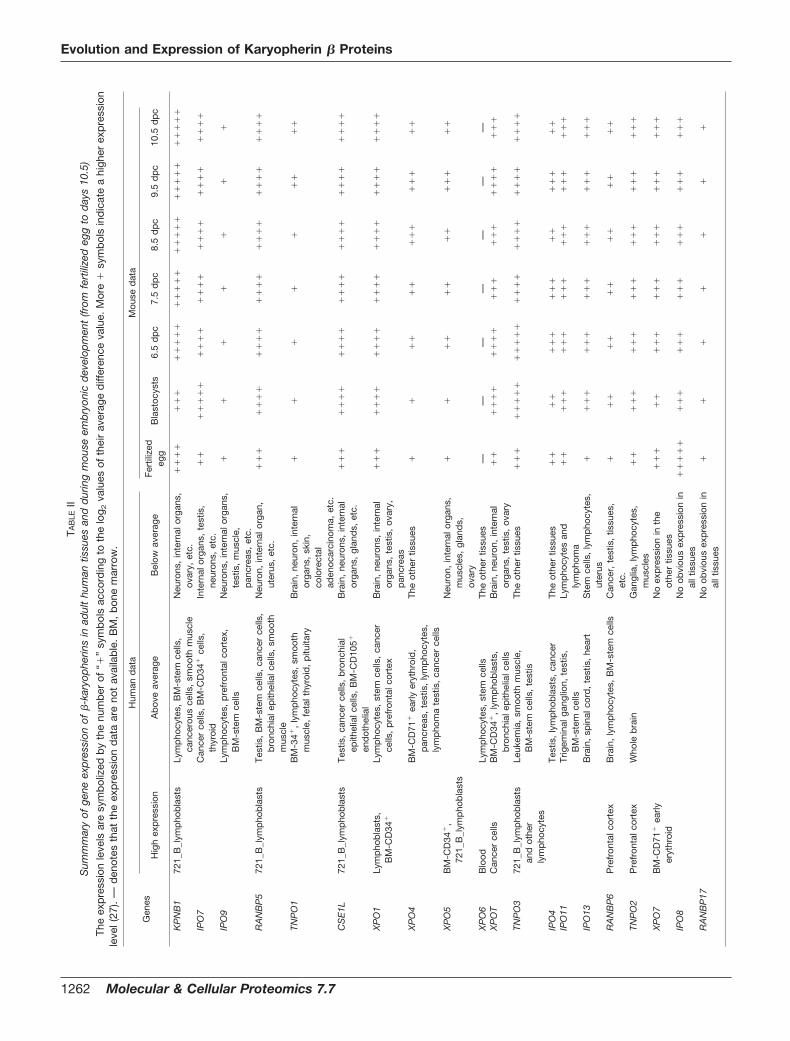
genes, and Table II shows that they are ubiquitously ex-pressed. Nevertheless they are not evenly expressed in allhuman tissues. 12 of 20 human �-karyopherin genes (CSE1L,IPO4, IPO7, IPO9, KPNB1, RANBP5, TNPO1, TNPO3, XPO1,XPO4, XPO5, and XPOT) show comparatively high expressionin tissues that proliferate actively, e.g. lymphocytes, tumors,testis, and stem cells. The high expression of these genesmay be due to their ability to carry cargoes needed for cellproliferation such as histones, glucocorticoid receptors, andRNAs (7). For example, CSE1L, which transports importin-�
(44), is overexpressed in colon cancer cell lines, breast can-cers, and liver neoplasms (45), and excess CSE1L may reduceimportin-�/�-dependent import. Higher expression of CSE1Lin tumors may reflect deregulation of nuclear transport.Among the overexpressed genes, KPNB1, RANBP5, CSE1L,
XPO1, XPO5, and TNPO3 are more highly expressed in lym-phoblasts than in other proliferating tissues. A second groupof karyopherin genes (IPO11, IPO13, RANBP6, TNPO2, andXPO6) is expressed more strongly in brain and spinal cordthan in other tissues. Different expression patterns of �-karyo-pherin genes among tissues were also documented in mousemicroarray data; however, the patterns are different fromthose in man. Tissue expression screening of �-karyopheringenes in the mouse revealed the restricted expression of Ipo8in both oocytes and fertilized eggs, whereas, curiously, suchrestriction was not observed in man. Mouse Ranbp17 mRNAis abundantly expressed in testis and pancreas (46). Thisfinding is supported by our ongoing analysis of microarraydata. We also observed that Ranbp17 has high expression inbrown fat and skeletal muscle.
Developmental Regulation of Karyopherin � SuperfamilyGene Expression—During the cell cycle and during develop-ment, there is a continuous need for precise transport of largemolecules in and out of the nucleus. Regulation of expressionof �-karyopherins could be important for these processes (16,19). We therefore examined the gene expression patterns of�-karyopherins during mouse development (27) by analyzinggene microarray data (Fig. 6). Most �-karyopherin genes in-crease their expression gradually during development, reachtheir peak expression by day 6.5–9.5, and then drop by day10.5. Through the similarity measure of gene expression pro-files in the early embryonic period (from fertilized egg to day10.5), we grouped 19 mouse �-karyopherin genes (Xpo6 dataare not available) into six expression patterns. The pattern thatwas found most frequently is characteristic of 10 genes(Cse1l, Ipo4, Ipo7, Ipo11, Ipo13, Ranbp5, Tnpo2, Tnpo3,Xpo1, and Xpot) (Fig. 6a) whose expression increases signif-icantly from fertilized egg to blastocysts and then fluctuatesslightly until day 10.5. Both Ranbp6-Xpo5 and Tnpo1-Xpo4pairs show expression peaks; however, the former pair is atday 6.5 and day 9.5 (Fig. 6b), and the latter pair is primarily atday 9.5 (Fig. 6c). Ipo9 and Ranbp17 are nearly constant (Fig.6d), whereas Xpo7, Kpnb1, and Ipo8 decrease in blastocysts,and all but Ipo8 then rebound (Fig. 6e). Expression of Xpo7and Kpnb1 rebounds on day 6.5 and remains high until day10.5 (Fig. 6f). In conclusion, �-karyopherin transcript levels dovary during development. Differences in their cargo specificitymay explain these changes.
Cell Cycle-dependent Transcriptional Regulation of Karyo-pherin � Superfamily Genes—As nuclear import of macromol-ecules can be regulated during the cell cycle due to alter-ations in the nuclear pore complex (47), it is of interest to askwhether expression of �-karyopherin genes is regulated in acell cycle-dependent manner. Different approaches havebeen adopted to identify, correct, and complete microarraydata sets of S. cerevisiae synchronized with mating factor orin cell cycle mutants. KAP95 shows the most obviouschanges during cell cycle progression (29, 30). In addition, aperiodic least square regression suggests cell cycle regulation
Evolution and Expression of Karyopherin � Proteins
Molecular & Cellular Proteomics 7.7 1261
TAB
LEII
Sum
mm
ary
ofge
neex
pre
ssio
nof
�-k
aryo
phe
rins
inad
ult
hum
antis
sues
and
dur
ing
mou
seem
bry
onic
dev
elop
men
t(fr
omfe
rtili
zed
egg
tod
ays
10.5
)
The
exp
ress
ion
leve
lsar
esy
mb
oliz
edb
yth
enu
mb
erof
“�”
sym
bol
sac
cord
ing
toth
elo
g 2va
lues
ofth
eir
aver
age
diff
eren
ceva
lue.
Mor
e�
sym
bol
sin
dic
ate
ahi
gher
exp
ress
ion
leve
l(27
).—
den
otes
that
the
exp
ress
ion
dat
aar
eno
tav
aila
ble
.B
M,
bon
em
arro
w.
Gen
es
Hum
and
ata
Mou
sed
ata
Hig
hex
pre
ssio
nA
bov
eav
erag
eB
elow
aver
age
Fert
ilize
deg
gB
last
ocys
ts6.
5d
pc
7.5
dp
c8.
5d
pc
9.5
dp
c10
.5d
pc
KP
NB
172
1_B
_lym
pho
bla
sts
Lym
pho
cyte
s,B
M-s
tem
cells
,ca
ncer
ous
cells
,sm
ooth
mus
cle
Neu
rons
,in
tern
alor
gans
,ov
ary,
etc.
��
��
��
��
��
��
��
��
��
��
��
��
��
��
��
��
IPO
7C
ance
rce
lls,
BM
-CD
34�
cells
,th
yroi
dIn
tern
alor
gans
,te
stis
,ne
uron
s,et
c.�
��
��
��
��
��
��
��
��
��
��
��
��
��
IPO
9Ly
mp
hocy
tes,
pre
fron
talc
orte
x,B
M-s
tem
cells
Neu
rons
,in
tern
alor
gans
,te
stis
,m
uscl
e,p
ancr
eas,
etc.
��
��
��
�
RA
NB
P5
721_
B_l
ymp
hob
last
sTe
stis
,B
M-s
tem
cells
,ca
ncer
cells
,b
ronc
hial
epith
elia
lcel
ls,
smoo
thm
uscl
e
Neu
ron,
inte
rnal
orga
n,ut
erus
,et
c.�
��
��
��
��
��
��
��
��
��
��
��
��
��
TNP
O1
BM
-34�
,ly
mp
hocy
tes,
smoo
thm
uscl
e,fe
talt
hyro
id,
pitu
itary
Bra
in,
neur
on,
inte
rnal
orga
ns,
skin
,co
lore
ctal
aden
ocar
cino
ma,
etc.
��
��
��
��
�
CS
E1L
721_
B_l
ymp
hob
last
sTe
stis
,ca
ncer
cells
,b
ronc
hial
epith
elia
lcel
ls,
BM
-CD
105�
end
othe
lial
Bra
in,
neur
ons,
inte
rnal
orga
ns,
glan
ds,
etc.
��
��
��
��
��
��
��
��
��
��
��
��
��
�
XP
O1
Lym
pho
bla
sts,
BM
-CD
34�
Lym
pho
cyte
s,st
emce
lls,
canc
erce
lls,
pre
fron
talc
orte
xB
rain
,ne
uron
s,in
tern
alor
gans
,te
stis
,ov
ary,
pan
crea
s
��
��
��
��
��
��
��
��
��
��
��
��
��
�
XP
O4
BM
-CD
71�
early
eryt
hroi
d,
pan
crea
s,te
stis
,ly
mp
hocy
tes,
lym
pho
ma
test
is,
canc
erce
lls
The
othe
rtis
sues
��
��
��
��
��
��
��
XP
O5
BM
-CD
34�
,72
1_B
_lym
pho
bla
sts
Neu
ron,
inte
rnal
orga
ns,
mus
cles
,gl
and
s,ov
ary
��
��
��
��
��
��
�
XP
O6
Blo
odLy
mp
hocy
tes,
stem
cells
The
othe
rtis
sues
——
——
——
—X
PO
TC
ance
rce
llsB
M-C
D34
�,
lym
pho
bla
sts,
bro
nchi
alep
ithel
ialc
ells
Bra
in,
neur
on,
inte
rnal
orga
ns,
test
is,
ovar
y�
��
��
��
��
��
��
��
��
��
��
��
TNP
O3
721_
B_l
ymp
hob
last
san
dot
her
lym
pho
cyte
s
Leuk
emia
,sm
ooth
mus
cle,
BM
-ste
mce
lls,
test
isTh
eot
her
tissu
es�
��
��
��
��
��
��
��
��
��
��
��
��
��
��
IPO
4Te
stis
,ly
mp
hob
last
s,ca
ncer
The
othe
rtis
sues
��
��
��
��
��
��
��
��
�IP
O11
Trig
emin
alga
nglio
n,te
stis
,B
M-s
tem
cells
Lym
pho
cyte
san
dly
mp
hom
a�
��
��
��
��
��
��
��
��
��
�
IPO
13B
rain
,sp
inal
cord
,te
stis
,he
art
Ste
mce
lls,
lym
pho
cyte
s,ut
erus
��
��
��
��
��
��
��
��
��
�
RA
NB
P6
Pre
fron
talc
orte
xB
rain
,ly
mp
hocy
tes,
BM
-ste
mce
llsC
ance
r,te
stis
,tis
sues
,et
c.�
��
��
��
��
��
��
TNP
O2
Pre
fron
talc
orte
xW
hole
bra
inG
angl
ia,
lym
pho
cyte
s,m
uscl
es�
��
��
��
��
��
��
��
��
��
�
XP
O7
BM
-CD
71�
early
eryt
hroi
dN
oex
pre
ssio
nin
the
othe
rtis
sues
��
��
��
��
��
��
��
��
��
��
IPO
8N
oob
viou
sex
pre
ssio
nin
allt
issu
es�
��
��
��
��
��
��
��
��
��
��
��
RA
NB
P17
No
obvi
ous
exp
ress
ion
inal
ltis
sues
��
��
��
�
Evolution and Expression of Karyopherin � Proteins
1262 Molecular & Cellular Proteomics 7.7

of KAP123 and NMD5 (48), and an approach based on cubicsplines suggests modulation of MTR10 (49). Bayesian mod-eling techniques also indicate that KAP114 and KAP95 aremodulated (50). Thus, although it would be useful to haveadditional data sets, cell cycle-dependent regulation of�-karyopherin expression could impact nuclear import andexport during the cell cycle.
Mechanism of Regulation of Karyopherin � SuperfamilyGenes—DNA regulatory elements are crucial for understand-ing gene expression because the binding of correspondingfactors determines the timing, location, and level of geneexpression. Computational methods have been developed toidentify and localize regulatory elements in a high throughputmanner (51, 52). Moreover similar clusters of TFBSs can befound in the promoter regions of orthologous genes (53). Acomputational tool, MAPPER (31), was therefore adopted toidentify putative TFBSs of human �-karyopherin genes (Fig. 7)and their orthologs in eight model species (supplemental dataand supplemental Table 2).
We observed that the predicted promoter binding sites of�-karyopherin genes lack TATA boxes but are rich in GCcontent (Fig. 7 and supplemental materials). Although theTATA box is often required for transcription by nuclear RNApolymerases, several in vivo studies show that the TATA boxis more specifically important for cellular proliferation, trans-
formation, and control of the cell cycle (54, 55) than for reg-ulation of housekeeping genes (56, 57). Mammalian promot-ers lacking a TATA box often contain SP1 (GGGCGG)/NF-1(TGGNNNNNNGCCAA) binding sites and rely on them toinitiate gene expression (58–60). SP1 is ubiquitously ex-pressed and plays a key role in maintaining basal transcriptionof housekeeping genes (61). SP1 is also implicated in medi-ating development-specific gene expression (62). It is pre-dicted that the SP1 binding site is one of the most commonTFBSs for �-karyopherin genes: 13 of 20 human �-karyo-pherin genes (CSE1L, IPO8, IPO9, IPO13, KPNB1, RANBP5,RANBP6, RANBP17, TNPO1, TNPO2, TNPO3, XPO1, andXPOT) are predicted to have one or multiple SP1 binding sitesin their proximal regulatory regions (Table III and Fig. 7). TheCAAT box is another frequent candidate binding site for�-karyopherin genes: seven of 20 human �-karyopheringenes (CSE1L, IPO9, KPNB1, RANBP5, RANBP6, TNPO2,and XPO1) and 10 of 20 mouse �-karyopherin genes (Cse1l,Ipo9, Ipo11, Kpnb1, Ranbp5, Tnpo1, Tnpo2, Xpo1, Xpo4, andXpo6) were predicted to posses this CAAT box in their up-stream regulatory regions. The CAAT box is also ubiquitouslydistributed, being present in about 30% of eukaryotic promot-ers (63, 64). In higher eukaryotes, it is involved in many typesof promoters: developmentally controlled (65), cell-cycle reg-ulated (66), and housekeeping (67). It is interesting that some-
FIG. 6. Gene expression patterns of �-karyopherins in mouse early development. The vertical axis is the log2 ratio of gene expressionlevels; the horizontal axis is the developmental period abbreviated as follows (days postcoitus (dpc)): T1, fertilized egg; T2, blastocysts; T3, dpc6.5; T4, dpc 7.5; T5, dpc 8.5; T6, dpc 9.5; and T7, dpc 10.5. The gene expression of mouse �-karyopherin genes was grouped into six patterns:a, Cse1l, Ipo4, Ipo7, Ipo11, Ipo13, Ranbp5, Tnpo2, Tnpo3, Xpo1, and Xpot; b, Ranbp6 and Xpo5; c, Tnpo1 and Xpo4; d, Ipo9 and Ranbp17;e, Kpnb1 and Xpo7; and f, Ipo8.
Evolution and Expression of Karyopherin � Proteins
Molecular & Cellular Proteomics 7.7 1263

FIG. 7. Transcription factor bindingsites of human �-karyopherin genespredicted by the MAPPER search en-gine. The regulatory analyses of the 20�-karyopherin genes are illustrated lineby line. The numbers under the lines arethe positions (in bp) from the start ofthe transcript (position �1). The TFBSsare indicated above or beneath thelines. Five mostly potential TFBSs (SP1,NRF-2, Hen-1, RREB-1, and CAAT box)are highlighted by symbols. USF, up-stream stimulatory factor; PPAR, per-oxisome proliferator-activated recep-tor; COUP-TF, chicken ovalbuminupstream promoter-transcription fac-tor; CREB, cAMP-response element-binding protein; RXR, retinoid X recep-tor; VDR, vitamin D receptor; ER,estrogen receptor; NF, nuclear factor;SRY, sex-determining region Y-chro-mosome protein; FTF, �-1-fetoproteintranscription factor; HLF, hepatic leu-kemia factor; ATF, activating transcrip-tion factor; CDP, CCAAT displacementprotein; NF-GMa, nuclear factor forgranulocyte/macrophage colony-stim-ulating factor; HLTF, helicase-like tran-scription factor.
Evolution and Expression of Karyopherin � Proteins
1264 Molecular & Cellular Proteomics 7.7

FIG. 7—continued
Evolution and Expression of Karyopherin � Proteins
Molecular & Cellular Proteomics 7.7 1265

times SP1 can interact with nuclear factor Y (NF-Y), alsoknown as CAAT box DNA-binding protein, to regulate down-stream genes cooperatively (68, 69). Nuclear respiratory fac-tor 2 (NRF-2), known as GA-binding protein, was also re-ported to cooperate with SP1 in the activation of severalwidely expressed housekeeping genes and genes that controlcell cycle, differentiation, development, and other key cellularfunctions (70). The TFBS for NRF-2 is also found frequently(12 of 20 genes) in promoters of human �-karyopherin genes(IPO4, IPO7, IPO8, IPO9, IPO11, IPO13, RANBP17, TNPO1,TNPO3, XPO5, XPO6, and XPOT). There are some TFBSs,e.g. the binding sites for Ras-responsive element-binding pro-tein 1 (RREB-1), helix-loop-helix protein 1 (HEN-1), aryl hy-drocarbon receptor nuclear transporter, and B-cell-specificactivator protein, that are also predicted to be potentialTFBSs of human or mouse �-karyopherin genes. These tran-scription factors are known to participate in the regulation ofdevelopment, differentiation, cell cycle, and other karyo-pherin-related cellular processes (38, 71, 72). Moreover sev-eral further TFBSs are predicted to be frequently involved inthe regulation of �-karyopherin genes of different experimen-tal models (Table III), e.g. five of 18 X. tropicalis �-karyopheringenes have binding sites for Staf, 12 of 16 D. melanogaster�-karyopherin genes have binding sites for Broad-complexisoform 4, three of 10 C. elegans �-karyopherin genes havebinding sites for UNC-86, and three of 14 S. cerevisiae�-karyopherin genes have binding sites for Ste12p.
Despite our conclusion that selected TFBSs are sharedamong �-karyopherin genes within a species (Fig. 8 and sup-plemental materials), many of the putative TFBSs seem to be
species-specific. For example, putative binding sites of SP1,NRF-2, HEN-1, RREB-1, and NF-Y emerge frequently in up-stream regions of �-karyopherin genes in human and mouse,whereas Broad-complex serial binding sites are preferred inthe fly (Table III and supplemental materials). These resultssuggest that functional regulatory factors change quickly dur-ing evolution and that the gain and loss of functional TFBSsare frequent during evolution (73). These and related conceptswill be refined as the �-karyopherins and �-karyopherin genesof additional species are characterized.
Conclusions
We analyzed karyopherin � superfamily members with re-gard to their evolution and expression. Although bacteria doencode HEAT repeat proteins, such as CpcE and CpcF (36),the karyopherin � family itself appears to be absent fromprokaryotes. Interestingly yeast Ran proteins are extremelysimilar to mammalian Ran or “Ran proteins of other species.”(74). Moreover most nucleoporins have homologs in all extanteukaryotic lineages, and the existence of distant prokaryotichomologs of several nucleoporins has been proposed (75). Bycontrast, there is more variability among the �-karyopherins.
Given the incomplete evolutionary record, we found theseemingly sudden appearance of 14 �-karyopherin membersin S. cerevisiae. Yet these 14 yeast �-karyopherins are notuniformly represented in higher organisms. Moreover they arenot all required for eukaryotic cells considering that C. elegansand some protozoa (e.g. G. lamblia) lack many of them. Fromyeast to man, eight �-karyopherin genes appear to haveevolved directly without diversification, but KAP104, MTR10,NMD5, and PSE1 diverged by gene duplication or retroposi-tion. Although the number of �-karyopherins increases from14 in yeast to 20 in human, only 10 �-karyopherins have beenidentified so far in C. elegans although the size of the genomeof C. elegans (97 Mbp) is much larger than that of S. cerevisiae(12 Mbp). It will be of interest to know how higher organismsthat have lost �-karyopherins still maintain their normal phys-iological functions.
Expression analysis of �-karyopherins showed that theirtranscripts vary with cell differentiation and proliferation.Moreover in yeast, the titer of some �-karyopherin transcriptsappears to be regulated during the cell cycle, and in mice,titers vary during development, suggesting changing roles ofnucleocytoplasmic transport during cell differentiation. Virtualanalysis of promoter binding elements showed that the com-binations of transcription factors SP1, NRF-2, HEN-1,RREB-1, and NF-Y regulate expression of most �-karyopheringenes.
We thus used bioinformatics techniques to give a system-atic overview of �-karyopherins: their sequence and structuralfeatures, their evolution, their transcriptional patterns, andtheir gene regulatory motifs. Future studies will need to ad-dress how a full set of �-karyopherins first appeared and
TABLE IIIThe top putative TFBSs of �-karyopherin genes in eight organisms: H.sapiens (Hs), M. musculus (Mm), G. gallus (Gg), X. tropicalis (Xt), D.rerio (Dr), D. melanogaster (Dm), C. elegans (Ce), and S. cerevisiae
(Sc)
The number following the abbreviated taxa indicates the number of�-karyopherin genes in the designated organism, whereas the num-ber after a TFBS indicates the number of �-karyopherin genes thatwere predicted by MAPPER search engine to be regulated by theTFBS. ARNT, aryl hydrocarbon receptor nuclear transporter; Bsap,B-cell-specific activator protein.
Species TFBSs
Hs (20) SP1 (13), NRF-2 (12), HEN-1 (10),RREB-1 (9), CAAT box (7)
Mm (20) CAAT box (10), ARNT (9), Bsap (9)Gg (17) NF-�B (2)Xt (18) Staf (5)Dr (23) HNF-1 (11)Dm (16) Broad-complex_1 (12),
Broad-complex_2 (7),Broad-complex_3 (7),Broad-complex_4 (13),Hunchback (6)
Ce (10) UNC-86 (3)Sc (14) STE12 (3)
Evolution and Expression of Karyopherin � Proteins
1266 Molecular & Cellular Proteomics 7.7

FIG. 8. Transcription factor binding sites of XPO1 orthologous genes predicted by the MAPPER search engine. The regulatoryanalyses of XPO1 over species are illustrated line by line: Hs, H. sapiens; Mm, M. musculus; Gg, G. gallus; Xt, X. tropicalis; Dr, D. rerio; Dm,D. melanogaster; Ce, C. elegans; and Sc, S. cerevisiae. The numbers under the lines are the positions (in bp) from the start of the transcript(position �1). The TFBSs are indicated above or beneath the lines. TF, transcription factor; PPAR, peroxisome proliferator-activated receptor;NF, nuclear factor; Bsap, B-cell-specific activator protein. NF-GMa, nuclear factor for granulocytes/macrophage colony-stimulating factor;DREF, DNA replication-related element binding factor; Su(H), suppressor of hairless protien; SU_h, suppressor of hairless protein.
Evolution and Expression of Karyopherin � Proteins
Molecular & Cellular Proteomics 7.7 1267

diversified (from nine to 14 members) in yeast, whether thereare additional functional �-karyopherins that have not beenfound, and how the �-karyopherins have co-evolved alongwith nucleoporins, components of the Ran GTPase cycle, andcargo diversity (70, 76–78).
* This work was supported by Grant 2006AA02A310 from the Min-istry of Science and Technology, China; Grants 3047085, 20423002,and 90608007 from the National Natural Science Foundation of Chi-na; Grant C0510003 from the Natural Science Foundation of FujianProvince; Grant 2005-383 from the Ministry of Education of China;and Intramural Fund XK0014 from Xiamen University (to T. T.) and byGrant 30400573 from the National Natural Science Foundation ofChina and a grant from the Program for New Century ExcellentTalents of Ministry of Education of China (to Z.-L. J.). The costs ofpublication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked “advertisement”in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
□S The on-line version of this article (available at http://www.mcponline.org) contains supplemental material.
§ To whom correspondence may be addressed: School of LifeSciences, Xiamen University, Xiamen City, Fujian 361005, China.Tel./Fax: 86-592-2182880; E-mail: [email protected] (Tao Tao) [email protected] (Zhiliang Ji).
REFERENCES
1. Bednenko, J., Cingolani, G., and Gerace, L. (2003) Importin � contains aCOOH-terminal nucleoporin binding region important for nuclear trans-port. J. Cell Biol. 162, 391–401
2. Chook, Y. M., and Blobel, G. (2001) Karyopherins and nuclear import. Curr.Opin. Struct. Biol. 11, 703–715
3. Gorlich, D., and Kutay, U. (1999) Transport between the cell nucleus andthe cytoplasm. Annu. Rev. Cell Dev. Biol. 15, 607–660
4. Macara, I. G. (2001) Transport into and out of the nucleus. Microbiol. Mol.Biol. Rev. 65, 570–594
5. Mattaj, I. W., and Englmeier, L. (1998) Nucleocytoplasmic transport: thesoluble phase. Annu. Rev. Biochem. 67, 265–306
6. Nakielny, S., and Dreyfuss, G. (1999) Transport of proteins and RNAs in andout of the nucleus. Cell 99, 677–690
7. Pemberton, L. F., and Paschal, B. M. (2005) Mechanisms of receptor-mediated nuclear import and nuclear export. Traffic 6, 187–198
8. Pemberton, L. F., and Paschal, B. M. (2006) Scientists share nuclearsecrets at Jekyll Island. Traffic 7, 751–760
9. Poon, I. K., and Jans, D. A. (2005) Regulation of nuclear transport: centralrole in development and transformation? Traffic 6, 173–186
10. Conti, E., Muller, C. W., and Stewart, M. (2006) Karyopherin flexibility innucleocytoplasmic transport. Curr. Opin. Struct. Biol. 16, 237–244
11. Hodges, J. L., Leslie, J. H., Mosammaparast, N., Guo, Y., Shabanowitz, J.,Hunt, D. F., and Pemberton, L. F. (2005) Nuclear import of TFIIB ismediated by Kap114p, a karyopherin with multiple cargo-binding do-mains. Mol. Biol. Cell 16, 3200–3210
12. Mosammaparast, N., and Pemberton, L. F. (2004) Karyopherins: from nu-clear-transport mediators to nuclear-function regulators. Trends CellBiol. 14, 547–556
13. Kuersten, S., Ohno, M., and Mattaj, I. W. (2001) Nucleocytoplasmic trans-port: Ran, � and beyond. Trends Cell Biol. 11, 497–503
14. Macara, I. G. (1999) Nuclear transport: randy couples. Curr. Biol. 9,R436–R439
15. Hogarth, C. A., Calanni, S., Jans, D. A., and Loveland, K. L. (2006) Importin� mRNAs have distinct expression profiles during spermatogenesis. Dev.Dyn. 235, 253–262
16. Zhang, C., Sweezey, N. B., Gagnon, S., Muskat, B., Koehler, D., Post, M.,and Kaplan, F. (2000) A novel karyopherin-� homolog is developmentallyand hormonally regulated in fetal lung. Am. J. Respir. Cell Mol. Biol. 22,451–459
17. Vrailas, A. D., Marenda, D. R., Cook, S. E., Powers, M. A., Lorenzen, J. A.,Perkins, L. A., and Moses, K. (2006) smoothened and thickveins regulate
Moleskin/Importin 7-mediated MAP kinase signaling in the developingDrosophila eye. Development 133, 1485–1494
18. Loveland, K. L., Hogarth, C., Szczepny, A., Prabhu, S. M., and Jans, D. A.(2006) Expression of nuclear transport importins � 1 and � 3 is regulatedduring rodent spermatogenesis. Biol. Reprod. 74, 67–74
19. Tao, T., Lan, J., Presley, J. F., Sweezey, N. B., and Kaplan, F. (2004)Nucleocytoplasmic shuttling of lgl2 is developmentally regulated in fetallung. Am. J. Respir. Cell Mol. Biol. 30, 350–359
20. Tatusov, R. L., Fedorova, N. D., Jackson, J. D., Jacobs, A. R., Kiryutin, B.,Koonin, E. V., Krylov, D. M., Mazumder, R., Mekhedov, S. L., Nikolskaya,A. N., Rao, B. S., Smirnov, S., Sverdlov, A. V., Vasudevan, S., Wolf, Y. I.,Yin, J. J., and Natale, D. A. (2003) The COG database: an updatedversion includes eukaryotes. BMC Bioinformatics 4, 41
21. Birney, E., Andrews, D., Caccamo, M., Chen, Y., Clarke, L., Coates, G.,Cox, T., Cunningham, F., Curwen, V., Cutts, T., Down, T., Durbin, R.,Fernandez-Suarez, X. M., Flicek, P., Graf, S., Hammond, M., Herrero, J.,Howe, K., Iyer, V., Jekosch, K., Kahari, A., Kasprzyk, A., Keefe, D.,Kokocinski, F., Kulesha, E., London, D., Longden, I., Melsopp, C., Meidl,P., Overduin, B., Parker, A., Proctor, G., Prlic, A., Rae, M., Rios, D.,Redmond, S., Schuster, M., Sealy, I., Searle, S., Severin, J., Slater, G.,Smedley, D., Smith, J., Stabenau, A., Stalker, J., Trevanion, S., Ureta-Vidal, A., Vogel, J., White, S., Woodwark, C., and Hubbard, T. J. (2006)Ensembl 2006. Nucleic Acids Res. 34, D556–D561
22. Edgar, R. C. (2004) MUSCLE: multiple sequence alignment with high ac-curacy and high throughput. Nucleic Acids Res. 32, 1792–1797
23. Thompson, J. D., Gibson, T. J., Plewniak, F., Jeanmougin, F., and Higgins,D. G. (1997) The CLUSTAL_X windows interface: flexible strategies formultiple sequence alignment aided by quality analysis tools. NucleicAcids Res. 25, 4876–4882
24. Kumar, S., Tamura, K., and Nei, M. (2004) MEGA3: integrated software forMolecular Evolutionary Genetics Analysis and sequence alignment. Brief.Bioinform. 5, 150–163
25. Dohm, J. C., Vingron, M., and Staub, E. (2006) Horizontal gene transfer inaminoacyl-tRNA synthetases including leucine-specific subtypes. J. Mol.Evol. 63, 437–447
26. Montanini, B., Blaudez, D., Jeandroz, S., Sanders, D., and Chalot, M. (2007)Phylogenetic and functional analysis of the Cation Diffusion Facilitator(CDF) family: improved signature and prediction of substrate specificity.BMC Genomics 8, 107
27. Su, A. I., Cooke, M. P., Ching, K. A., Hakak, Y., Walker, J. R., Wiltshire, T.,Orth, A. P., Vega, R. G., Sapinoso, L. M., Moqrich, A., Patapoutian, A.,Hampton, G. M., Schultz, P. G., and Hogenesch, J. B. (2002) Large-scaleanalysis of the human and mouse transcriptomes. Proc. Natl. Acad. Sci.U. S. A. 99, 4465–4470
28. Wang, Y. P., Liang, L., Han, B. C., Quan, Y., Wang, X., Tao, T., and Ji, Z. L.(2006) GEPS: the Gene Expression Pattern Scanner. Nucleic Acids Res.34, W492–W497
29. Spellman, P. T., Sherlock, G., Zhang, M. Q., Iyer, V. R., Anders, K., Eisen,M. B., Brown, P. O., Botstein, D., and Futcher, B. (1998) Comprehensiveidentification of cell cycle-regulated genes of the yeast Saccharomycescerevisiae by microarray hybridization. Mol. Biol. Cell 9, 3273–3297
30. de Lichtenberg, U., Jensen, L. J., Fausboll, A., Jensen, T. S., Bork, P., andBrunak, S. (2005) Comparison of computational methods for the identi-fication of cell cycle-regulated genes. Bioinformatics 21, 1164–1171
31. Marinescu, V. D., Kohane, I. S., and Riva, A. (2005) MAPPER: a searchengine for the computational identification of putative transcription factorbinding sites in multiple genomes. BMC Bioinformatics 6, 79
32. Harel, A., and Forbes, D. J. (2004) Importin �: conducting a much largercellular symphony. Mol Cell 16, 319–330
33. Gorlich, D., Dabrowski, M., Bischoff, F. R., Kutay, U., Bork, P., Hartmann,E., Prehn, S., and Izaurralde, E. (1997) A novel class of RanGTP bindingproteins. J. Cell Biol. 138, 65–80
34. Malik, H. S., Eickbush, T. H., and Goldfarb, D. S. (1997) Evolutionaryspecialization of the nuclear targeting apparatus. Proc. Natl. Acad. Sci.U. S. A. 94, 13738–13742
35. Bayliss, R., Littlewood, T., Strawn, L. A., Wente, S. R., and Stewart, M.(2002) GLFG and FxFG nucleoporins bind to overlapping sites on impor-tin-�. J. Biol. Chem. 277, 50597–50606
36. Mans, B. J., Anantharaman, V., Aravind, L., and Koonin, E. V. (2004)Comparative genomics, evolution and origins of the nuclear envelopeand nuclear pore complex. Cell Cycle 3, 1612–1637
Evolution and Expression of Karyopherin � Proteins
1268 Molecular & Cellular Proteomics 7.7

37. Zhang, J. Z. (2003) Evolution by gene duplication: an update. Trends Ecol.Evol. 18, 292–298
38. Mukhopadhyay, N. K., Cinar, B., Mukhopadhyay, L., Lutchman, M., Ferdi-nand, A. S., Kim, J., Chung, L. W., Adam, R. M., Ray, S. K., Leiter, A. B.,Richie, J. P., Liu, B. C., and Freeman, M. R. (2007) The zinc finger proteinras-responsive element binding protein-1 is a coregulator of the andro-gen receptor: implications for the role of the Ras pathway in enhancingandrogenic signaling in prostate cancer. Mol Endocrinol. 21, 2056–2070
39. Kenyon, J. R., and Craig, I. W. (1999) Analysis of the 5� regulatory region ofthe human Norrie’s disease gene: evidence that a non-translated CTdinucleotide repeat in exon one has a role in controlling expression.Gene (Amst.) 227, 181–188
40. Seroussi, E., Shani, N., Ben-Meir, D., Chajut, A., Divinski, I., Faier, S., Gery,S., Karby, S., Kariv-Inbal, Z., Sella, O., Smorodinsky, N. I., and Lavi, S.(2001) Uniquely conserved non-translated regions are involved in gen-eration of the two major transcripts of protein phosphatase 2C�. J. Mol.Biol. 312, 439–451
41. Vinckenbosch, N., Dupanloup, I., and Kaessmann, H. (2006) Evolutionaryfate of retroposed gene copies in the human genome. Proc. Natl. Acad.Sci. U. S. A. 103, 3220–3225
42. Wang, P. J. (2004) X chromosomes, retrogenes and their role in malereproduction. Trends Endocrinol. Metab. 15, 79–83
43. Bai, Y., Casola, C., Feschotte, C., and Betran, E. (2007) Comparativegenomics reveals a constant rate of origination and convergent acquisi-tion of functional retrogenes in Drosophila. Genome Biol. 8, R11
44. Goldfarb, D. S., Corbett, A. H., Mason, D. A., Harreman, M. T., and Adam,S. A. (2004) Importin �: a multipurpose nuclear-transport receptor.Trends Cell Biol. 14, 505–514
45. Behrens, P., Brinkmann, U., and Wellmann, A. (2003) CSE1L/CAS: its rolein proliferation and apoptosis. Apoptosis 8, 39–44
46. Koch, P., Bohlmann, I., Schafer, M., Hansen-Hagge, T. E., Kiyoi, H., Wilda,M., Hameister, H., Bartram, C. R., and Janssen, J. W. (2000) Identifica-tion of a novel putative Ran-binding protein and its close homologue.Biochem. Biophys. Res. Commun. 278, 241–249
47. Makhnevych, T., Lusk, C. P., Anderson, A. M., Aitchison, J. D., andWozniak, R. W. (2003) Cell cycle regulated transport controlled by alter-ations in the nuclear pore complex. Cell 115, 813–823
48. Johansson, D., Lindgren, P., and Berglund, A. (2003) A multivariate ap-proach applied to microarray data for identification of genes with cellcycle-coupled transcription. Bioinformatics 19, 467–473
49. Luan, Y., and Li, H. (2004) Model-based methods for identifying periodicallyexpressed genes based on time course microarray gene expressiondata. Bioinformatics 20, 332–339
50. Lu, X., Zhang, W., Qin, Z. S., Kwast, K. E., and Liu, J. S. (2004) Statisticalresynchronization and Bayesian detection of periodically expressedgenes. Nucleic Acids Res. 32, 447–455
51. Bussemaker, H. J., Li, H., and Siggia, E. D. (2001) Regulatory elementdetection using correlation with expression. Nat. Genet. 27, 167–171
52. Liu, Y., Liu, X. S., Wei, L., Altman, R. B., and Batzoglou, S. (2004) Eukaryoticregulatory element conservation analysis and identification using com-parative genomics. Genome Res. 14, 451–458
53. Wray, G. A., Hahn, M. W., Abouheif, E., Balhoff, J. P., Pizer, M., Rockman,M. V., and Romano, L. A. (2003) The evolution of transcriptional regula-tion in eukaryotes. Mol. Biol. Evol. 20, 1377–1419
54. Davidson, I., Martianov, I., and Viville, S. (2004) TBP, a universal transcrip-tion factor? Med. Sci. (Paris) 20, 575–579
55. Johnson, S. A., Dubeau, L., White, R. J., and Johnson, D. L. (2003) TheTATA-binding protein as a regulator of cellular transformation. Cell Cycle2, 442–444
56. Wang, L. H., and Chen, L. (1996) Organization of the gene encoding humanprostacyclin synthase. Biochem. Biophys. Res. Commun. 226, 631–637
57. Azizkhan, J. C., Jensen, D. E., Pierce, A. J., and Wade, M. (1993) Tran-scription from TATA-less promoters: dihydrofolate reductase as a model.Crit. Rev. Eukaryot. Gene Expr. 3, 229–254
58. Zhao, J., and Ennion, S. J. (2006) Sp1/3 and NF-1 mediate basal transcrip-tion of the human P2X1 gene in megakaryoblastic MEG-01 cells. BMCMol. Biol. 7, 10
59. Emami, K. H., Burke, T. W., and Smale, S. T. (1998) Sp1 activation of aTATA-less promoter requires a species-specific interaction involvingtranscription factor IID. Nucleic Acids Res. 26, 839–846
60. Nichols, A. F., Itoh, T., Zolezzi, F., Hutsell, S., and Linn, S. (2003) Basaltranscriptional regulation of human damage-specific DNA-binding pro-tein genes DDB1 and DDB2 by Sp1, E2F, N-myc and NF1 elements.Nucleic Acids Res. 31, 562–569
61. Samson, S. L., and Wong, N. C. (2002) Role of Sp1 in insulin regulation ofgene expression. J. Mol. Endocrinol. 29, 265–279
62. Thomas, K., Wu, J., Sung, D.Y., Thompson, W., Powell, M., McCarrey, J.,Gibbs, R., and Walker, W. (2007) SP1 transcription factors in male germcell development and differentiation. Mol. Cell. Endocrinol. 270, 1–7
63. Bucher, P. (1990) Weight matrix descriptions of four eukaryotic RNA poly-merase II promoter elements derived from 502 unrelated promoter se-quences. J. Mol. Biol. 212, 563–578
64. Mantovani, R. (1999) The molecular biology of the CCAAT-binding factorNF-Y. Gene (Amst.) 239, 15–27
65. Berry, M., Grosveld, F., and Dillon, N. (1992) A single point mutation is thecause of the Greek form of hereditary persistence of fetal haemoglobin.Nature 358, 499–502
66. Mantovani, R. (1998) A survey of 178 NF-Y binding CCAAT boxes. NucleicAcids Res. 26, 1135–1143
67. Roy, B., and Lee, A. S. (1995) Transduction of calcium stress throughinteraction of the human transcription factor CBF with the proximalCCAAT regulatory element of the grp78/BiP promoter. Mol. Cell. Biol. 15,2263–2274
68. Roder, K., Wolf, S. S., Beck, K. F., and Schweizer, M. (1997) Cooperativebinding of NF-Y and Sp1 at the DNase I-hypersensitive site, fatty acidsynthase insulin-responsive element 1, located at �500 in the rat fattyacid synthase promoter. J. Biol. Chem. 272, 21616–21624
69. Wright, K. L., Moore, T. L., Vilen, B. J., Brown, A. M., and Ting, J. P. (1995)Major histocompatibility complex class II-associated invariant chaingene expression is up-regulated by cooperative interactions of Sp1 andNF-Y. J. Biol. Chem. 270, 20978–20986
70. Rosmarin, A. G., Resendes, K. K., Yang, Z., McMillan, J. N., and Fleming,S. L. (2004) GA-binding protein transcription factor: a review of GABP asan integrator of intracellular signaling and protein-protein interactions.Blood Cells Mol. Dis. 32, 143–154
71. Zhang, S., Qian, X., Redman, C., Bliskovski, V., Ramsay, E. S., Lowy, D. R.,and Mock, B. A. (2003) p16 INK4a gene promoter variation and differ-ential binding of a repressor, the ras-responsive zinc-finger transcriptionfactor, RREB. Oncogene 22, 2285–2295
72. Brown, L., and Baer, R. (1994) HEN1 encodes a 20-kilodalton phospho-protein that binds an extended E-box motif as a homodimer. Mol. Cell.Biol. 14, 1245–1255
73. Doniger, S. W., and Fay, J. C. (2007) Frequent gain and loss of functionaltranscription factor binding sites. PLoS Comput. Biol. 3, e99
74. Kadowaki, T., Goldfarb, D., Spitz, L. M., Tartakoff, A. M., and Ohno, M.(1993) Regulation of RNA processing and transport by a nuclear guaninenucleotide release protein and members of the Ras superfamily. EMBOJ. 12, 2929–2937
75. Bapteste, E., Charlebois, R. L., MacLeod, D., and Brochier, C. (2005) Thetwo tempos of nuclear pore complex evolution: highly adapting proteinsin an ancient frozen structure. Genome Biol. 6, R85
76. Denning, D. P., and Rexach, M. F. (2007) Rapid evolution exposes theboundaries of domain structure and function in natively unfolded FGnucleoporins. Mol. Cell. Proteomics 6, 272–282
77. Devos, D., Dokudovskaya, S., Williams, R., Alber, F., Eswar, N., Chait, B. T.,Rout, M. P., and Sali, A. (2006) Simple fold composition and modulararchitecture of the nuclear pore complex. Proc. Natl. Acad. Sci. U. S. A.103, 2172–2177
78. Stavru, F., Nautrup-Pedersen, G., Cordes, V. C., and Gorlich, D. (2006)Nuclear pore complex assembly and maintenance in POM121- andgp210-deficient cells. J. Cell Biol. 173, 477–483
79. Cingolani, G., Petosa, C., Weis, K., and Muller, C. W. (1999) Structure ofimportin-� bound to the IBB domain of importin-�. Nature 399, 221–229
80. Cuff, J. A., Clamp, M. E., Siddiqui, A. S., Finlay, M., and Barton, G. J. (1998)JPred: a consensus secondary structure prediction server. Bioinformat-ics 14, 892–893
81. Lee, S. J., Matsuura, Y., Liu, S. M., and Stewart, M. (2005) Structural basisfor nuclear import complex dissociation by RanGTP. Nature 435,693–696
Evolution and Expression of Karyopherin � Proteins
Molecular & Cellular Proteomics 7.7 1269