examining rare and low-frequency genetic variants...
TRANSCRIPT

DOI: 10.1161/CIRCGENETICS.114.000718
1
Examining Rare and Low-Frequency Genetic Variants Previously Associated
with Lone or Familial Forms of Atrial Fibrillation in an Electronic Medical
Record System: A Cautionary Note
Running title: Weeke et al.; Rare and Low-Frequency Genetic Variants and AF
Peter Weeke, MD1,2; Joshua C. Denny, MD1,3; Lisa Basterache, MS3; Christian Shaffer, BS1;
Erica Bowton, PhD4; Christie Ingram,BS1; Dawood Darbar, MD1; Dan M. Roden, MD1
1Department of Internal Medicine, 3Department of Biomedical Informatics, Vanderbilt University Medical Center, Nashville, TN; 2Department of Cardiology, Copenhagen University
Hospital, Gentofte, Denmark; 4Institute for Clinical & Translational Research, Vanderbilt University School of Medicine, Nashville, TN
Correspondence:
Dan M. Roden, MD
Professor of Medicine and Pharmacology
Assistant Vice-Chancellor for Personalized Medicine
Vanderbilt University School of Medicine
1285 Medical Research Building IV
Nashville, TN 37232
Tel: 615-322-0067
Fax: 615-343-4522
E-mail: [email protected]
Journal Subject Codes: [146] Genomics, [89] Genetics of cardiovascular disease, [8] Epidemiology
3;;;; Chrirrr stiaaaan n nn Shafffffefefeferrrr,,, , B
; Dan n n n M. Roddddenenenen, ,,, MMMMD
a ly va b
artrtrtmmmem nt of Inteernnnal MMMedicicici ine, 333DDDeppartttmmennt of BiBiBiiomomomomedddiccal IInffforrrmmmam ticccsc , VaVandereerbily rMeMeMedidd cal CeCeCeCenttteeere , NaNaNaashviviviv lllllle,e,e,e, TTTNNN;;; 2222DeDeDeDepapapartttmemm nnntn ooof f ff CaCaCarddddioioiololll gygygyy, CoCoCoopepepp nhnhnhnhaggggeeen UUUUnnival, Gentofte, Deeeenmnnmark; 4InInInstitute fof r Clinnnnicii al & TrTrTrT ana slational l l l Research, Vanderb
UnUnUnUnivivivvererersisisiitytytyty SSSSchchchchoooooool l l l ofofoff MMMMededede icccinininine,ee NNNNasasasa hvhvhvhvililillelelee, ,,, TNTNTNN
by guest on July 3, 2018http://circgenetics.ahajournals.org/
Dow
nloaded from
by guest on July 3, 2018http://circgenetics.ahajournals.org/
Dow
nloaded from
by guest on July 3, 2018http://circgenetics.ahajournals.org/
Dow
nloaded from
by guest on July 3, 2018http://circgenetics.ahajournals.org/
Dow
nloaded from
by guest on July 3, 2018http://circgenetics.ahajournals.org/
Dow
nloaded from
by guest on July 3, 2018http://circgenetics.ahajournals.org/
Dow
nloaded from
by guest on July 3, 2018http://circgenetics.ahajournals.org/
Dow
nloaded from
by guest on July 3, 2018http://circgenetics.ahajournals.org/
Dow
nloaded from

DOI: 10.1161/CIRCGENETICS.114.000718
2
Abstract:
Background - Studies in individuals or small kindreds have implicated rare variants in 25
different genes in lone and familial atrial fibrillation (AF) using linkage and segregation analysis,
functional characterization, and rarity in public databases. Here we used a cohort of 20,204
patients of European or African ancestry with electronic medical records (EMRs) and exome
chip data to compare the frequency of AF among carriers and non-carriers of these rare variants.
Methods and Results - The exome chip included 19/115 rare variants, in 9 genes, previously
associated with lone or familial AF. Using validated algorithms querying a combination of
clinical notes, structured billing codes, ECG reports, and procedure codes, we identified 1,056
AF cases (>18 years) and 19,148 non-AF controls (>50 years) with available genotype data on
the Illumina HumanExome BeadChip v.1.0 in the Vanderbilt electronic medical record-linked
DNA repository, BioVU. Known correlations between AF and common variants at 4q25 were
replicated. None of the 19 variants previously associated with AF were overrepresented among
AF cases (P >0.1 for all), and the frequency of variant carriers among non-AF controls was
>0.1% for 14/19. Repeat analyses using non-AF controls aged >60 (n=14,904) >70 (n=9,670),
and >80 (n=4,729) years old did not influence these findings.
Conclusions - Rare variants previously implicated in lone or familial forms AF present on the
exome chip are detected at low frequencies in a general population but are not associated with
AF. These findings emphasize the need for caution when ascribing variants as pathogenic or
causative.
Key words: Genome Wide Association Study, single nucleotide polymorphism, genetics, association studies, atrial fibrillation, gene mutation, family study, penetrance, exome, genome, GWAS
, ,
aaaaililililabababablelelele gegegegenonononotytytytypepepepe ddddaaataaaa
ic meddddicicicicalalalal rrrrececececorororordddd-l-lllinii k
s w
N m
P s
1 7
=
n p y p present on
sitoryryry,, BiBiBiBioVoVVoVU.. KKKnown correlations betwwweeeeen AF and commomomon variants at 4q25 w
NNNoN nnne of the 191919 varariaiaiaantnntsss prprp evvvioioiousussly aaassss ociaiated d d wiwiwithththt AAAAF weeereere oveveerrrrepepprerereseseentntntedededd aaam
P >0>0>0>0.1... for aaalllll ),, aaand thhhe ffffrerer ququenencyyy oof vavvarianantt caaarrrieeerss amomm ngngg nnnonn-A-A-A-AF cococontttroools wwas
14/19. Repeat aaanannalyyyyseses usususing ggg nooon-AF cononontrt olss aggggedede >6060 ((((n=n=n=141414,9,9,90400 ) >70 (n=9,67
=4,7,, 29) ) yeyy ars olllldd dd didididid ddd not influence thhhhese fifififi ddnddingsg .
nnnss - RaRaRarere vvarariaiaiantnttss pprereviviviiououslslslyy imimimi plplplicicicattatedededd iiiinn lololol nene oorr fafafaf mimimilililiialalal fofofof rmrmss AFAFAFAF prpresesenentt onon
by guest on July 3, 2018http://circgenetics.ahajournals.org/
Dow
nloaded from

DOI: 10.1161/CIRCGENETICS.114.000718
3
Atrial fibrillation (AF) is the most prevalent arrhythmia encountered in clinical practice and is
significantly associated with increased risks of stroke, heart failure, and death.1 While there are a
many clinical correlates of AF (e.g. hypertension and diabetes), genetic variability is being
increasingly recognized as an important determinant for developing AF.2-11 12-16 To date, rare
genetic variants in the exonic regions of 25 genes encoding cardiac ion channels, gap junction
proteins, and signaling molecules have been linked with in lone or familial forms of AF.2-11 In
addition, genome-wide association studies (GWAS) have associated multiple common variants
with AF in a general population.13-16 Overall, the effects of common variants on AF
susceptibility are small, whereas the effects of rare variants on AF susceptibility have not been
determined, although they are presumed to be large.17 Establishing the prevalence of such
variants in large population studies with available phenotypic information can be useful i) to
estimate the effect sizes associated with genetic variants; ii) to avoid falsely predicting
individuals being at risk for disease; and iii) to evaluate penetrance. In the present study, we
studied a large cohort of patients with available genotyping data from the Illumina HumanExome
BeadChip v.1.0 genotyping array (“the exome chip”) that interrogates ~250,000 rare and
common exonic variants and linked to de-identified electronic medical records (EMR).18 We
tested the hypothesis that AF is more prevalent in carriers of rare variants previously designated
as pathogenic for AF.
Methods
BioVU and Exome Chip Study cohorts
BioVU links DNA samples extracted from discarded blood samples to de-identified EMRs, as
previously described.18 In 2012-2013, ~40,000 subjects in BioVU were genotyped on the exome
variants on AF
ceptiiibibibilililitytt hhah ve nonoottt bbb
17
l t
e
w
arge cohort of patients ith a ailable genot ping data from the Ill mina H manE
, alllthththouououghghgh ttthehehey y y are presumed to be largeee.171717 Establishingg thehehe prevalence of such
laara gggeg populatioonnn sttuddddiesss s wwiw thhhh avvvaiilabbblee phehenootytyyypipipic infofformaatiiionn n ccan bebebebe uuuseeefulll l iii)i t
e effefeffectctctc ssssizizizes asassosocicii tatatede wwwittitithhhh genenenetiititic vvaririiianantsttt ; ; iiiiiii)))) totototo avovovooidididid ffafalslsll ely y prprprp edicctititingng
being gg at risk for dididiisease;;; and dd iiiiii)i)i)i) to ev llaluate pppenetrance. In thhhhe prpp esent study,y,y, w
hhort ff iti ts iithh iaillablbl t ipi ddat ffr thhe IlIlll imina HH EnE by guest on July 3, 2018http://circgenetics.ahajournals.org/
Dow
nloaded from

DOI: 10.1161/CIRCGENETICS.114.000718
4
chip (http://genome.sph.umich.edu/wiki/Exome_Chip_Design). Subjects for exome chip
genotyping were drawn from the following BioVU cohorts:
1. An elderly cohor -up.
2. A rare diseases cohort: subjects with FDA defined rare diseases.
3. A longitudinal cohort:
a. Healthy longitudinal set = subjects with 1 or 2 notes/year for at least 5 years.
b. Sick longitudinal set = subjects with 3, 4, 5, or 6 notes/year for at least 5 years.
4. The Vanderbilt Electronic Systems for Pharmacogenomic Assessment (VESPA)
cohort: subjects with drug response phenotypes.19
5. A cancer cohort: subjects with cancer according to the Vanderbilt Tumor Registry.
6. A Pediatric cohort: subjects <18 years old with a minimum threshold of clinical
.
For the present study, we included all subjects of European American (EA) and African
American (AA) ancestry from groups 1-5 above.
AF phenotype
To identify AF cases, we used a validated algorithm20 that uses a combination of queries of
structured billing codes (i.e. International Classification of Diseases, version 9-CM [ICD-9],
procedure codes from Current Procedural Terminology [CPT]), and unstructured “natural
language” from clinical notes and electrocardiograms. In brief, AF cases were defined as
individuals who were >18 years old, had an ICD-9 diagnosis for AF or flutter (ICD-9: 427.3,
427.31, and 427.32) or a cardiologist diagnosis of AF as identified by a natural language
processing tool from the unstructured free text of the “ECG impression”. In all instances, patients
with a history of a heart transplant were excluded (CPT: 33935, 3394, and 580; ICD-9: V42.1,
c Assessment (VEEEESPSPSS
c
r
AA) ancestr from gro ps 1 5 abo e
A A A cacacancncncererer cohohohhooort: subjects with cancerr aaaccording to the VaVaVVanderbilt Tumor Reg
AAAA Pediatric cocoohoorttt: suuuubjbjbjb ectstts <188 yeaeears oolld wiwiwiwiththth aa mmminnimumumm ttthhhrh eshohohoholdlll oofff f cllllinnic
.
the prpp esent studdddy,y,y, we iiini cllluddddeddd allllll subbbjejej cts ffof Europepp an AAAAme iirican ((EA)) and Afr
AAAAA)) t ffr 11 55 bab by guest on July 3, 2018http://circgenetics.ahajournals.org/
Dow
nloaded from

DOI: 10.1161/CIRCGENETICS.114.000718
5
996.83).
For the present study, non-AF controls were defined as individuals who were >50 years
old and who were not identified as a case by the case algorithm. For sensitivity analyses, we
identified three additional control cohorts of individuals who were not identified as cases and
who were >60 years, >70 years, and >80 years old. In addition, we also defined a “definite non-
AF” control cohort as individuals -9 codes
for AF or atrial flutter (ICD-9: 427.3, 427.31, and 427.32), had no free text mentions AF in their
clinical notes, and no free-text references (including synonyms) to direct-current cardioversion,
atrial tachycardia or multifocal atrial tachycardia, atrioventricular nodal ablation, as previously
described.20 AF case and control numbers are derived from algorithm deployment into BioVU
on January 15, 2014. We stratified all analyses by AA and EA ancestry, as described further
below.
Genotyping and Quality Control
We excluded samples with genotype-phenotype sex discrepancies, samples with genotyping
efficiency <98%, related samples as determined by identity-by-descent, and unintended
duplicates. We removed SNPs that were monomorphic, had low call-rates (<98%), or were
common (minor allele frequency (MAF) >5%) and were out of Hardy-Weinberg equilibrium
(HWE) (HWE P<0.0001).
Ancestry
In order to reduce the influence of population stratification on our results, we identified and
analyzed separately individuals of EA and AA ancestry by calculating principal components
using EIGENSOFT21; the first and second principal components were retained for further
analysis.
ect-current cardioveveveversr
al abbbllllatititition, as pppreeeviviviviou
0 o
e
ed samples ith genot pe phenot pe se discrepancies samples ith genot pin
0 AFAFAF cccasasase e anaa d d d cccontrol numbers are deriveveved d from algorithmmm ddddeployment into Bio
1115,,,, 2014. We sttrattiffffied d aala l annnaala yyysses bybby AAAA andndnd EEEAAA annnccestryryy, asasas descrcrcrc ibii edededd fuuurtttht e
ggg and QQQualityyy CCControlll
edd lpl iithh ot hph ot didi iie lpl iithh t ipi by guest on July 3, 2018http://circgenetics.ahajournals.org/
Dow
nloaded from

DOI: 10.1161/CIRCGENETICS.114.000718
6
SNP identification
We performed a literature review to identify studies implicating rare genetic variants with lone or
familial forms of AF using the following search terms in PubMed: ((atrial fibrillation (mesh))
OR (atrial fibrillation)) AND ((genetics (mesh)) OR (genetic*)) AND ((mutation (mesh)) OR
(mutation*) OR (polymorphism, single nucleotide (mesh)) OR (polymorphism, single
nucleotide) OR (monogenic*) OR (gwas)). In total, we identified 565 articles meeting these
search criteria as of January 12, 2014. These articles implicated 115 rare genetic variants that had
evidence of i) co-segregation with AF; ii) supporting electrophysiological studies or; iii) were
associated with AF based on large studies of lone AF probands and/or SNP rarity. Supplemental
Table 1 lists additional rare variants not included in a recent review by Olesen et al.22 We cross-
referenced the 115 rare SNPs with the exome chip. Overlapping SNPs were retained for further
analysis.
As a positive control for our experiment, we also examined the relationship between
AF/non-AF phenotype status and previously-described common AF variants at the 4q25 locus
among EAs (rs6843082, rs2200733, and rs17042171).13, 16, 23
Statistical Analyses
Dichotomous and continuous variables were compared with Fisher’s exact test and the Kruskal-
Wallis test, respectively. Continuous values are presented as medians and interquartile ranges
(IQR). We determined SNP associations with AF using exact logistic regression. We performed
both unadjusted and adjusted analyses (age, sex, first and second principal components [PC]),
which yielded similar estimates unless otherwise noted.
We also performed a series of interaction analyses using logistic regression analysis with
2 main model effect terms (“presence or absence of rare AF variants” and “presence or absence
ical studies or; iii)i))) wwwwe
SNPPPP ra iiiritttyt . SSSuS ppppppplelelelem
22 c
t r
a n
F phenot pe stat s and pre io sl described common AF ariants at the 4q25 lo
s adddddididid titititionononalalalal raaarerere variants not included in nn a recent review bybybyy Olesen et al.22 We c
thheh 115 eeexxomemme chihhipprare SNPNN s wwwiw thh ttthheh . OvOverrlalalappppppppinng SSNPss ewewerrere rettttaiaiaiineeedd d d forrr fuff r
a popp sitive controllll ffffor our exppperimiii ent,t,t, we alllso exa imiiined ddd thhhe relall tiiiionshipp between
F hph ot stat dnd ii lsl dde iribbedd AAFF iri ts t hth 44q2525 ll by guest on July 3, 2018http://circgenetics.ahajournals.org/
Dow
nloaded from

DOI: 10.1161/CIRCGENETICS.114.000718
7
of common AF variants”) and a multiplicative interaction term (“presence or absence of rare AF
variants” X “presence or absence of common AF variants”) to test the hypothesis that the co-
occurrence of both common AF variants and rare AF variants modulates the risk of developing
AF (i.e. SNP X SNP interaction). We tested the co-occurrence of each of common vs. each rare
variant and we also collapsed rare and common variants into an allelic test based on the presence
and absence of the variant. We only evaluated SNP X SNP interactions for SNPs that were
identified from studies based on EA populations.
To reduce the risk of misclassification due to age-dependent development of AF, we
repeated all association analyses using additional non-AF control cohorts, with no AF as
described above at age cut-offs of >60, >70, and >80 years. Conservation for single base variants
and prediction of functional effects were assessed using PhastCons, Genome Evolution Rate
Profiler (GERP), Grantham scores, and PolyPhen2.
All analyses were performed using PLINK/SEQ (http://atgu.mgh.harvard.edu/plinkseq)
and SAS, version 9.2 (SAS institute Inc., Cary, NC, USA). A two-sided p-value <0.05 was
considered to be statistically significant.
Ethics
Approval for the present study was obtained from the Vanderbilt Institutional Review Board
(IRB).
Results
We identified 20,204 individuals (18,424 EAs and 1,780 AAs) with available genotype data and
also passed all QC parameters. Table 1 lists the clinical characteristics for AF cases and the non-
, stratified by EA and AA ancestry. Overall, the AF cases were older
(77.8 [IQR: 69.4-83.8] years) than the non- -79.4]
eveloppment of AFFF,,,, wwww
orts, witititithhhh no AAAAF FFF asasass
a a
i t
E
k
ersion 9 2 (SAS instit te Inc Car NC USA) A t o sided p al e <0 05 as
abovvve ee atatat aaagegege cutututt-o-- ffs of >60, >70, and >808080 yyears. Conservaatititiion for single base va
ioononn of functionnaaal efffffectssss wwwew reee asssseessedeed ussinng PPPhahahasttCooonns, GGGeeenomomome EvEvEvEvooolo uuutiiioi nn RRRRat
ERP)P)PP), GrGrGrGranthhhamam sscocoreres, aanddndnd PolyPyPyPhehehehe 2n2n2.
analyyses were ppperfffof rm dded usinggg PPPLIL NKNKNK/S/S/S/SEQEQEQQ ((httptp://a// tgtgu.mggh.hh hahh rvard.edu/plp ink
isi 99 22 ((SASASS iinstiit t IIn CCa NCNC USUSA)A) AA t idid ded lal <00 0505 by guest on July 3, 2018http://circgenetics.ahajournals.org/
Dow
nloaded from

DOI: 10.1161/CIRCGENETICS.114.000718
8
years, P<0.001). The median age was 74.2 (IQR: 66.6-81.5) years for non-
years (n=13,715), 79.4 (IQR: 74.8-84.6) years for the non- s (n=8,856),
and 84.6 (IQR: 81.9-88.5) years for the non- (n=4,384). For the
definite non-AF controls (n=4,514), the median age was 78.0 (IQR: 66.6-83.9).
Previously-described associations between common genetic variants at the 4q25 locus
and AF were replicated in the present study in both adjusted (rs6843082, odds ratio [OR]=1.28,
95% confidence interval [CI]: 1.15-1.42, p=5.3x10-6; rs2200733, OR=1.33, CI: 1.16-
1.50,p=3.0x10-5; rs17042171, OR=1.33, CI: 1.17-1.51, p=1.9x10-5) and unadjusted additive
models (data not shown).
Cross referencing the 115 rare SNPs previously implicated with lone or familial forms of
AF with the exome chip yielded a total of 19 overlapping SNPs, in 9 genes: NPPA (n=1),
KCNE4 (n=1), GREM2 (n=1), SCN5A (n=11), GATA4 (n=1), KCNJ8 (n=1), KCND3 (n=1),
KCNA5 (n=1), and KCNJ2 (n=1). In the parent studies suggesting a role for these SNPs in lone
or familial forms of AF, 14 SNPs were identified among individuals of EA ancestry, 4 SNPs in
individuals of AA ancestry, and 1 SNP in Asian subjects.
None of the 19 SNPs were significantly overrepresented among EA/AA AF cases
compared to the respective non-AF controls (P>0.1 for all; Table 2). Of the 19 SNPs, 14/19 (10
in EAs and 4 in AAs) were -AF controls (number of risk
allele carriers range among 17,416 EA non-AF controls, range: 19-179 [0.11-1.03%]; number of
risk allele carriers range among 1,732 AA non-AF controls, range: 24-112 [1.39-6.47%]).
Four of the 10 EA SNPs that were common among the non-AF controls were
very rare or absent in the 1,008 EA AF cases (range:0-1), and an additional 4 EA SNPs were rare
among both EA AF cases (range: 0-1 [0-0.1%]) and EA non-AF controls (range: 2-14 [0.01-
d unadjusted addititittivivivive
r
e
= )
= l
forms of AF 14 SNPs ere identified among indi id als of EA ancestr 4 SNP
ss rererefefeferererencncnciniii g g ththththe 115 rare SNPs previoooouususlyl implicated wiiithththh lone or familial for
e exxxxome chip yyiiieldddedddd a tototot tal ofofofo 19119 ovvverlappppingg SSSNPNPNPs, inn 9 egeeneeesss: NPPPPPPA (nn=1)1)1)1),
=1), GRGRGRGREMEMEME 2 (((nn=1)1)1), SCSCSCS N555AAA (((n=11)11)1), GAGAGAGATATATATA4444 (n=n=n==1)11)1), KCKCKCK NJNJNJNJ888 (((nn=1)))), KCKCKCK NDNDND3333 (((nn=1)11
=1)), ,, and KCNJ2222 (n(n(( 11=11).). InII thhhhe papp rent studididid es suggggegg stiniii g gg a rolllel fffor these SNPs in l
foff fof AAFF 1144 SNSNPPs idid itififi ded iindidi iidd lal fof EEAA st 44 SNSNPP by guest on July 3, 2018http://circgenetics.ahajournals.org/
Dow
nloaded from

DOI: 10.1161/CIRCGENETICS.114.000718
9
0.08%]). All four SNPs originally reported to be of AA ancestry were common among both AA
AF cases and AA non-AF controls (Table 2).
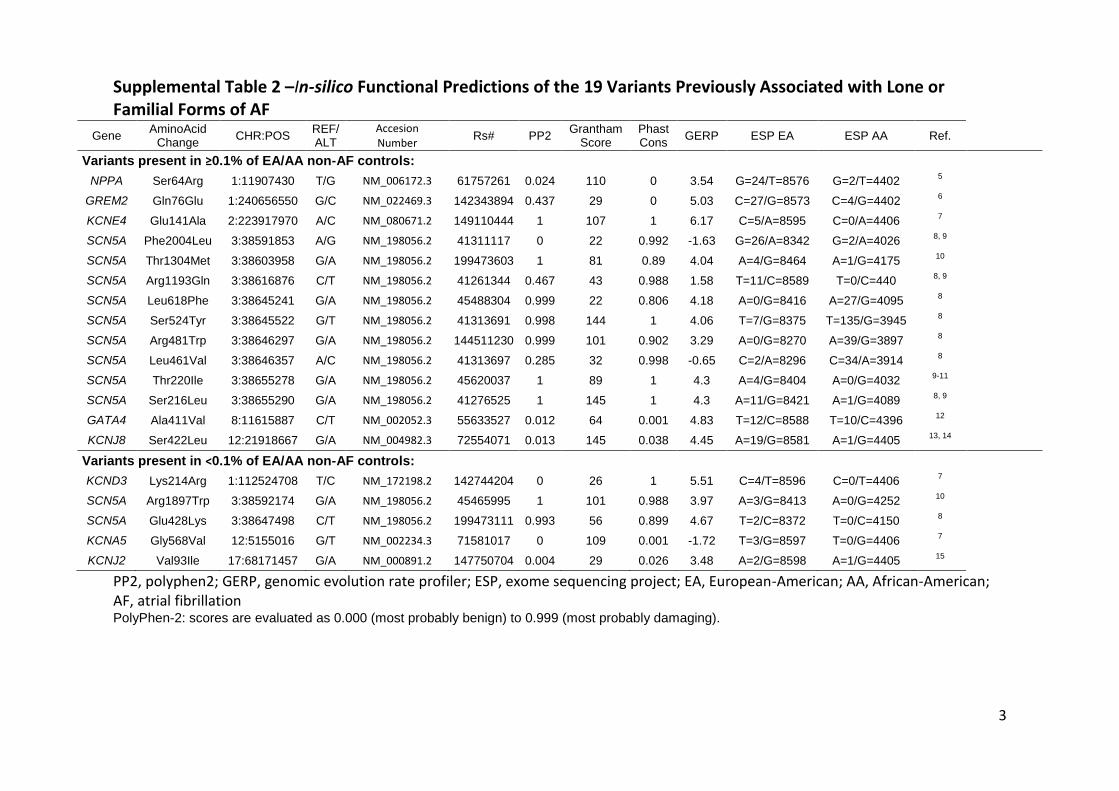
Supplemental Table 2 presents the in silico functional predictions of the 19 SNPs and the
derived EA and AA MAFs from the exome sequencing project of 6500 individuals.25 Of the 19
variants, 9 were predicted to disrupt protein function by PolyPhen2 (the PolyPhen2 score ranges
from 0 to 1, with the threshold for probably damaging at 0.85).
Repeat analyses using the definite EA and AA non-AF controls (n=4514 and n= 541,
respectively), the EA and AA non-AF controls aged >60 years (n=13,715 and n=1,189,
respectively), the EA and AA non-AF controls aged >70 years (n=8,856 and n=714,
respectively), and the EA and AA non-AF controls aged >80 years (n=4,388 and n=341,
respectively), did not influence the results.
SNP x SNP interaction
Table 3 presents the number of rare variant carriers as a function of common AF variant carrier
status and AF status. Here there was no significant interaction between co-occurrence of
common AF variants (rs6843082, rs2200733, and rs17042171) and rare genetic variants and AF
susceptibility (P=0.47). We also found no association between overall rare AF variant carrier
status (ignoring common variants) and AF susceptibility (P=0.81).
Discussion
In the present study we found that the rare or low-frequency variants interrogated by the exome
chip and previously implicated in lone or familial forms of AF were not associated with AF in a
general population; a substantial proportion of the non-AF controls carried rare or low frequency
variants previously labeled as being AF causative or pathogenic. Moreover, co-occurrence of
common variants and rare variants on the exome chip did not appear to modulate AF
715 and n=1,189,
56 and ddd n=71717171444,4,
y
y
P
e a
AF stat s Here there as no significant interaction bet een co occ rrence of
y), aaandndndnd ttttheheheh EEEEA AA aaand AA non-AF controlssss agagaged >80 years (nnnn=4== ,388 and n=341,
y)),) dddid not inflululueencce theehe rrresulululultts.
P intetetterarararactctctc iioi n
sents the number ffoff rare variant carriiei rs as a function offf f common AF variant ca
AAAFF stat HHe thhe isi ififiic t iinte ctiio bbet co ff by guest on July 3, 2018http://circgenetics.ahajournals.org/
Dow
nloaded from

DOI: 10.1161/CIRCGENETICS.114.000718
10
susceptibility risk. These data stress the need for caution when assigning pathogenicity of rare
variants.
We evaluated 19 rare variants previously implicated with lone or familial forms of AF by
linking genotypic information with phenotypic data from the Vanderbilt EMR. Notably, we did
not find any of the 19 rare variants previously implicated with lone or familial forms AF to be
enriched among patients with AF in either EA or AA populations. In fact, 14/19 variants were
relatively common among non-AF controls (>0.1%), which suggests that these variants may not
be causative (i.e. the monogenic cause of AF). It is possible that these variants are associated
with very low penetrance or contribute in some as yet undefined fashion to AF susceptibility. For
example, we can postulate that we did not identify a true pathogenic effect of the variants
because of a missing “second hit”; the “two-hit” hypothesis wherein the combination of a rare
genetic variant and another risk factor (e.g. a common AF variant or a clinical risk factor [e.g.
hypertension]) modifies the risk of AF.26 We recently reported an interaction between rare and
common AF risk variants modulating the penetrance of familial AF,27 but this signal was not
present in the general population studied here. While we determined that 14/19 variants are not
strongly correlated with AF, we are unable to determine the correlation with AF for 5/19 as they
were very rare or absent among both cases and controls.
For most genetic studies, replication is vital as it adds confidence and lessens the risk of a
false positive chance finding. However, for many studies implicating rare genetic variants,
replication is not possible given the rarity of the phenotype in question (unable to accrue enough
cases), the rarity of the variant in question (investigators would need to screen thousands of
subjects to gain adequate statistical power), or both. In the absence of readily available
replication cohorts for rare variants, variant function or causality can be inferred by providing
e variants are assocccciaiaiaiat
on to AAAAFFFF suscepptptibibibibililililiit
w
r
i e
o
F risk ariants mod lating the penetrance of familial AF 27 b t this signal as n
we cacacan nn popopostststuluuu attte e e that we did not identify aaa trttrue pathogenic efefefffect of the variants
aaa mmmmissing “secocoond hhhhiiiti ”;” ttthheh ““““twtwooo-hit””” hhypopothesesisss wwwheeereein tthhe cccooombiiiinananan tiononn offf f aaa r
iant anananandd dd aaanotttheheh rr ririi kksksk fffactooorrr (e(e(e(e.g. a aa cocococommmmmmm onon AAAFFF vavavariiririannttt orooror aa cclilll niiiicacacacalll riskkk fffacactotot rr [e[[
on])]) modifies thhe riiiiskkkk offf f AFAFAFF.2666 WeWW recentlylyl reppported d an iiiinteractiiion between rare
FFF iri ksk iiant dd lla iti thhe t fof ff ililiiall AFAF 2727 bb t thihi isi lal by guest on July 3, 2018http://circgenetics.ahajournals.org/
Dow
nloaded from

DOI: 10.1161/CIRCGENETICS.114.000718
11
convincing functional data. For rhythm disorders such as AF, function or causality is commonly
inferred through strong genotype-phenotype co-segregation patterns2, 3, 7, 9, 28, evidence of
electrophysiological effects3, 5-10, 29, and/or rarity of the variant in question in various control
populations under the assumption that truly pathogenic variants are under a strong negative
selection pressure.2, 4, 8, 9, 28 While such methods represent contemporary approaches for inferring
function or causality in studies involving arrhythmia susceptibility, they provide no information
regarding variant effects on disease susceptibility.
The idea that rare variants previously associated with disease may represent false positive
or at best weak associations, as has been suggested by others using public databases or isolated
cases of normal individuals harboring previously-reported pathogenic variants. For example,
Ploski et al. recently reported a patient with a normal phenotype and the Q247X deletion in
muscle ring finger protein 1 (TRIM63), a variant which has previously been reported by family
and functional studies as a cause of hypertrophic cardiomyopathy (HCM). 30, 31 In addition,
several studies have questioned the pathogenicity of genetic variants previously associated with
rare monogenic syndromes such as catecholaminergic polymorphic ventricular tachycardia,
Brugada syndrome, HCM, dilated cardiomyopathy, and arrhythmogenic right ventricular
cardiomyopathy,32-34 based on variant prevalence reported by the exome variant server
(http://evs.gs.washington.edu/EVS/).25 However, this resource does not include phenotypic
information, and did include some subjects with cardiovascular diseases such as DCM or
extreme lipid values.35, 36 This study suggests broadly-phenotyped EMR-linked genotype data
may be a useful resource for evaluating rare variants for pathogenicity.
Limitations
The rarity of the SNPs investigated limits the power of our study. In addition, while stratifying
may represent false e e e ppppo
blic dddd ttatt bbbabases or isisisisoooola
rmal individuals harboring previously-reported pathogenic variants. For examp
n
g m
,
dies ha e q estioned the pathogenicit of genetic ariants pre io sl associated
rmalalalal iiindndndndivivivididididuaaalslslsls harboring previously-reeepopported pathogeg nic c vvvariants. For examp
. rererer cecc ntly repoorttted aaa pattttieieiei nt wwwiiti hhh aa nooorrmaal phhhenenototototyype aand thththe Q2Q2Q2247XXX X deeeleeetionoon in
g fingggerererr pppprroteeinini 1111 (((TRTRTRRIMII 636363)))), a variririananananttt t hwhwhhiicichhhh hahahahassss ppprererereviiiouuououslslly bbbbeenn reerereportrtt ddeded bbby ffafam
nal studies as a cause offf hyhyh pepp rtrophphhiciii cardididiiomyoyy papp thhhhy yy (H(H(H(HCMCMCMCM))). 30303030, 31 In addition,
dddiie hha iti ded thhe athho ini icit ff iti iri ts ii lsl iciat ded by guest on July 3, 2018http://circgenetics.ahajournals.org/
Dow
nloaded from

DOI: 10.1161/CIRCGENETICS.114.000718
12
by race is important to reduce the likelihood of ancestral informative markers driving and
association, it does mean an additional loss of power in our analysis of individuals of both EA
and AA ancestry. The rarity of the SNPs investigated limits the power of our study. Hence, we
acknowledge the fact that we may be underpowered to truly detect an association between rare
genetic variants previously associated with monogenic or familial forms of AF. A power
calculation is depicted in the supplemental appendix (Supplemental Figure 1). While we tried to
eliminate the effects associated with population stratification by performing separate analysis for
EAs and AA, our results may have been influenced by differences in population substructure.
Although our results suggest that caution should be exercised before extrapolating study findings
on genetic correlates of lone or familial forms of AF to a general form of AF, it is important to
note that this conclusion is based on the findings examining 19 SNPs (with one originally
reported in Asian subjects and so not meaningfully evaluated here). Also, low numbers for the
AA AF cases may have influenced our results. The HumanExome BeadChip v.1.0 includes rare
variants (nonsynonymous variants 3 times and in 2 call sets; splice and stop variants seen 2
times and in 2 call sets) observed among ~9000 individuals of EA ancestry, ~2000 of AA
ancestry, and ~500 samples of Hispanic and Asian ancestry
(http://genome.sph.umich.edu/wiki/Exome_Chip_Design). Thus, singletons, which may be
associated with AF in families, are not interrogated. In the present study we did not perform
orthogonal validation of genotyped variants on the exome chip; genotype data from this platform
is known to yield reliable results.37, 38
While we in the present manuscript were able to replicate known AF associations which
do add confidence in the data, we acknowledge the fact that our study cohort represents patients
either hospitalized or seen in a hospital setting which may have affected our results. Hence,
population substrucucucuctututtu
xtrapolllal titititing stuttt dydydydy ffffiiiin
correlates of lone or familial forms of AF to a general form of AF, it is importan
Asian subjects and so not meaningfully evaluated here). Also, low numbers for
s
ons non mo s ariants 3 times and in 2 call sets; splice and stop ariants seen
corrererelalalaattetetess ofooo lononone or familial forms of AAAF FF tto a general form m of AF, it is importan
iss cccconclusion isisis baaseeede oooon nn the ee ffif ndnndingggs examaminnninining g gg 19 SSSNNPs (wiwwithththh oneeee oooriiigiiiinalllll yy
Asiaiannn susususubjbjbjecctstst aandndd sso o nottt mmmmeanininini gffgfgfulululu llyly eevavaluuuattatatededed hhhherre))e)e). AlAlAlsoso, lolololowwww numbmbbererss ffofor
es may yy have inffffllull enceddd d our resulltl s. TTThehh HHumanEEExome BBBeadCdCdCdChihihihippp v.1.0 includes
iiant 33 iti dd iin 22 llll et lpliic dd st iiant by guest on July 3, 2018http://circgenetics.ahajournals.org/
Dow
nloaded from

DOI: 10.1161/CIRCGENETICS.114.000718
13
extrapolation of our results to other study cohorts should be done with caution.
Conclusion
A set of rare variants previously implicated with lone or familial forms AF were detected at low
frequencies in a general population, where they were not associated with AF. Collectively, our
results suggest that evaluating the contribution of rare variants to human phenotypes continues to
require assessment of variant frequencies and associated phenotypes in the general population.
Moreover, our data also suggest that the variants present on the exome chip may be too common
to represent truly causative mutations in the setting of diseases of a presumed monogenic
etiology.
Funding Sources: The datasets used for the analyses described were obtained from Vanderbilt University Medical Center’s BioVU which is supported by institutional funding and by the Vanderbilt CTSA grant UL1 TR000445 from NCATS/NIH. This work was also in part supported by NIH grants U19 HL65962, and R01 HL092217). PW was funded by an unrestricted research grant from the Tryg Foundation (J.nr. 7343-09, TrygFonden, Denmark).
Conflict of Interest Disclosures: None
References:
1. Stewart S, Hart CL, Hole DJ, McMurray JJ. A population-based study of the long-term risks associated with atrial fibrillation: 20-year follow-up of the Renfrew/Paisley study. Am J Med.2002;113:359-364.
2. Darbar D, Kannankeril PJ, Donahue BS, Kucera G, Stubblefield T, Haines JL, et al. Cardiac sodium channel (SCN5A) variants associated with atrial fibrillation. Circulation.2008;117:1927-1935.
3. Xia M, Jin Q, Bendahhou S, He Y, Larroque MM, Chen Y, et al. A Kir2.1 gain-of-function mutation underlies familial atrial fibrillation. Biochem Biophys Res Commun. 2005;332:1012-1019.
4. Posch MG, Boldt LH, Polotzki M, Richter S, Rolf S, Perrot A, et al. Mutations in the cardiac transcription factor GATA4 in patients with lone atrial fibrillation. Eur J Med Genet.
p y
esumemememed d d d momomomononononogegegeg ninnn c ccc
o eMedical Center’s BioVU which is supported by institutional funding and by the
b sa
ouuuurcccces: Thhe ee ddatttaseetsss ussededed forr thhhe anaaalyyysees deeesccribebed wwere ooobtaiaiiinnned frfrfrommm VaaandeMeddddicicicalalalal CCCenntetet ’’r’r’s s BiBiBiioVo U UU whwhwhwhich isiisis supupupu pooo trtrt dededd bbbyyyy ininnsttststittutututtiiioionanallll fundndndndinining anandddd bybyb ttthhheCTSA grant ULULULUL11 TRTRTRTR000000004040445454545 fffrorororom m m m NCNCNCN ATATATSSSS/N/N/N/NIHIHIHH.... ThThThT isisisi wwwworororo k k k k wawawaass s s alaaa so in part
byy NIH gggrants UUU19191919 HHHL6L6L66595959626626 , ,, andd d R0R0R00111 HLHHL090909092222 17)7)7)7). PWPWPW was ffffunddedd d byy an unresaaant from thhhthe ee e TrTrTrygygygyg FFFFououooundndndndatatatioioioion n nn (JJJJ.n.nn.nr.rr 777734344343-33-3-09090909, TrTrTrTrygyygygFoFoFoFonddndndenenenen,, DeDeDeDenmnmmnmararark)k)k)k)..
by guest on July 3, 2018http://circgenetics.ahajournals.org/
Dow
nloaded from

DOI: 10.1161/CIRCGENETICS.114.000718
14
2010;53:201-203.
5. Delaney JT, Muhammad R, Blair MA, Kor K, Fish FA, Roden DM, et al. A KCNJ8 mutation associated with early repolarization and atrial fibrillation. Europace. 2012;14:1428-1432.
6. Medeiros-Domingo A, Tan BH, Crotti L, Tester DJ, Eckhardt L, Cuoretti A, et al. Gain-of-function mutation S422L in the KCNJ8-encoded cardiac K(ATP) channel Kir6.1 as a pathogenic substrate for J-wave syndromes. Heart Rhythm. 2010;7:1466-1471.
7. Abraham RL, Yang T, Blair M, Roden DM, Darbar D. Augmented potassium current is a shared phenotype for two genetic defects associated with familial atrial fibrillation. J Mol Cell Cardiol. 2010;48:181-190.
8. Olesen MS, Yuan L, Liang B, Holst AG, Nielsen N, Nielsen JB, et al. High prevalence of long QT syndrome-associated SCN5A variants in patients with early-onset lone atrial fibrillation. Circ Cardiovasc Genet. 2012;5:450-459.
9. Mann SA, Otway R, Guo G, Soka M, Karlsdotter L, Trivedi G, et al. Epistatic effects of potassium channel variation on cardiac repolarization and atrial fibrillation risk. J Am Coll Cardiol. 2012;59:1017-1025.
10. Muller, II, Melville DB, Tanwar V, Rybski WM, Mukherjee A, Shoemaker MB, et al. Functional modeling in zebrafish demonstrates that the atrial-fibrillation-associated gene GREM2 regulates cardiac laterality, cardiomyocyte differentiation and atrial rhythm. Dis Model Mech. 2013;6:332-341.
11. Olson TM, Michels VV, Ballew JD, Reyna SP, Karst ML, Herron KJ, et al. Sodium channel mutations and susceptibility to heart failure and atrial fibrillation. JAMA. 2005;293:447-454.
12. Weeke P, Muhammad R, Delaney JT, Shaffer C, Mosley JD, Blair M, et al. Whole-exome sequencing in familial atrial fibrillation. Eur Heart J. 2014;35:2477-2483.
13. Ellinor PT, Lunetta KL, Glazer NL, Pfeufer A, Alonso A, Chung MK, et al. Common variants in KCNN3 are associated with lone atrial fibrillation. Nat Genet. 2010;42:240-244.
14. Ellinor PT, Lunetta KL, Albert CM, Glazer NL, Ritchie MD, Smith AV, et al. Meta-analysis identifies six new susceptibility loci for atrial fibrillation. Nat Genet. 2012;44:670-675.
15. Pfeufer A, van Noord C, Marciante KD, Arking DE, Larson MG, Smith AV, et al. Genome-wide association study of PR interval. Nat Genet. 2010;42:153-159.
16. Benjamin EJ, Rice KM, Arking DE, Pfeufer A, van Noord C, Smith AV, et al. Variants in ZFHX3 are associated with atrial fibrillation in individuals of European ancestry. Nat Genet.2009;41:879-881.
lone atrial fibrillaaaatitititionooo
l Episisisistatatatatitititicccc efefefeffefefefectctctctssss ofooc0
g M3
TM Michels VV Balle JD Re na SP Karst ML Herron KJ et al Sodi m cha
chananannenenel lll vavavaririririattioioioon on cardiac repolarizatioioioion and atrial fibrillllllaaation risk. J Am Coll 012;2;2;;55595 :1017-777 101001025222 .
IIII,,, MMMMelvillelele DDBBB, TTaaanwaaar r V,V RRybbbsski WMWM, MuMuukkkheerjjeeee AA, Shohooemmmakaa errr MMMB,B,B, et all. moddddelelelininininggg in z bbebebrarafififi hshsh demmmoooonstratetetes thththt atttt ttthhehe aatrtrtrriaiaiaialll-fifififibrrrillilillllalatititionon-assosososociciciiatededd ggenenee gulates cardiaccc c lallateteteerarararalililitytytyty,, cacacaardrdrdr ioioioomymymymyococococytytyte e e dididiifffffffferererennnntititiatatatatioioioion n n anananand d d d atatatriririialalalal rhythm. Dis M3;6; :332-341.
TMTTM MiMi hch lel VVVV BBallll JJDD RR SSPP KK st MMLL HH KJKJ et ll SS dodii hch by guest on July 3, 2018http://circgenetics.ahajournals.org/
Dow
nloaded from

DOI: 10.1161/CIRCGENETICS.114.000718
15
17. Manolio TA, Collins FS, Cox NJ, Goldstein DB, Hindorff LA, Hunter DJ, et al. Finding the missing heritability of complex diseases. Nature. 2009;461:747-753.
18. Roden DM, Pulley JM, Basford MA, Bernard GR, Clayton EW, Balser JR, et al. Development of a large-scale de-identified DNA biobank to enable personalized medicine. Clin Pharmacol Ther. 2008;84:362-369.
19. Bowton E, Field JR, Wang S, Schildcrout JS, Van Driest SL, Delaney JT, et al. Biobanks and electronic medical records: enabling cost-effective research. Sci Transl Med. 2014;6:234cm233.
20. Ritchie MD, Denny JC, Crawford DC, Ramirez AH, Weiner JB, Pulley JM, et al. Robust replication of genotype-phenotype associations across multiple diseases in an electronic medical record. Am J Hum Genet. 2010;86:560-572.
21. Patterson N, Price AL, Reich D. Population structure and eigenanalysis. PLoS Genet.2006;2:e190.
22. Olesen MS, Nielsen MW, Haunso S, Svendsen JH. Atrial fibrillation: the role of common and rare genetic variants. Eur J Hum Genet. 2014;22:297-306.
23. Gudbjartsson DF, Holm H, Gretarsdottir S, Thorleifsson G, Walters GB, Thorgeirsson G, et al. A sequence variant in ZFHX3 on 16q22 associates with atrial fibrillation and ischemic stroke. Nat Genet. 2009;41:876-878.
24. Arnestad M, Crotti L, Rognum TO, Insolia R, Pedrazzini M, Ferrandi C, et al. Prevalence of long-QT syndrome gene variants in sudden infant death syndrome. Circulation. 2007;115:361-367.
25. Tennessen JA, Bigham AW, O'Connor TD, Fu W, Kenny EE, Gravel S, et al. Evolution and functional impact of rare coding variation from deep sequencing of human exomes. Science.2012;337:64-69.
26. Weeke P, Mosley JD, Hanna D, Delaney JT, Shaffer C, Wells QS, et al. Exome Sequencing Implicates an Increased Burden of Rare Potassium Channel Variants in the Risk of Drug-Induced Long QT Interval Syndrome. J Am Coll Cardiol. 2014;63:1430-1437.
27. Ritchie MD, Rowan S, Kucera G, Stubblefield T, Blair M, Carter S, et al. Chromosome 4q25 variants are genetic modifiers of rare ion channel mutations associated with familial atrial fibrillation. J Am Coll Cardiol. 2012;60:1173-1181.
28. Weeke P, Parvez B, Blair M, Short L, Ingram C, Kucera G, et al. Candidate gene approach to identifying rare genetic variants associated with lone atrial fibrillation. Heart Rhythm.2014;11:46-52.
29. Olson TM, Alekseev AE, Moreau C, Liu XK, Zingman LV, Miki T, et al. KATP channel mutation confers risk for vein of Marshall adrenergic atrial fibrillation. Nat Clin Pract
alysis. PLoS Genetttt...
on: thehehehe rrrrololololeeee ofofofof ccccomomomommmmn
rtsson DF, Holm H, Gretarsdottir S, Thorleifsson G, Walters GB, Thorgeirsson n s
a ny 3
neticicicic vvvararariaiaiantnn s.. EuEEE r J Hum Genet. 2014;2222:2:2:2922 7-306.
rttttssssson DF, Holmmm HH,H GGreeeetatat rsdododod ttiiir S, TThorrleeifssssonononon GGG, WaWW lteersss GBGBGBB, ThThhhooorgegegeiiri ssssoon ncecece vvvarianttt iin ZZZFHXHXX3 onoon 16q6qq22222 aassocoociateess wiwiwithhth aatrrialll ffibriiillllatioioonnn annnd d d ischchhemmmic s2009090909;4;4;441:1:1:1 8878 6-66 87878778.88
ad M, ,, Crotti L,,, RoRR gngg um TTTO,O,OO IIns lloliiai RRR,,, PePP drdd azziniii MMM, ,, FFerrandiddd CCCC,,, et al. Prevalenynndrome ggeneneneneee vavavavaririrrianananantststs iiiin nn n susuusudddddddenennen iiinfnffnfanaanant tt dedededeatattath hhh sysyysyndnndndrorooromemmeme. CiCiCiCircrcrcrcululuulatatatatioioiionnnn... 20202007;115:3
by guest on July 3, 2018http://circgenetics.ahajournals.org/
Dow
nloaded from

DOI: 10.1161/CIRCGENETICS.114.000718
16
Cardiovasc Med. 2007;4:110-116.
30. Ploski R, Pollak A, Muller S, Franaszczyk M, Michalak E, Kosinska J, et al. Does p.Q247X in TRIM63 Cause Human Hypertrophic Cardiomyopathy? Circ Res. 2014;114:e2-5.
31. Chen SN, Czernuszewicz G, Tan Y, Lombardi R, Jin J, Willerson JT, et al. Human molecular genetic and functional studies identify TRIM63, encoding Muscle RING Finger Protein 1, as a novel gene for human hypertrophic cardiomyopathy. Circ Res. 2012;111:907-919.
32. Jabbari J, Jabbari R, Nielsen MW, Holst AG, Nielsen JB, Haunso S, et al. New exome data question the pathogenicity of genetic variants previously associated with catecholaminergic polymorphic ventricular tachycardia. Circ Cardiovasc Genet. 2013;6:481-489.
33. Risgaard B, Jabbari R, Refsgaard L, Holst AG, Haunso S, Sadjadieh A, et al. High prevalence of genetic variants previously associated with Brugada syndrome in new exome data. Clin Genet. 2013;84:489-495.
34. Andreasen C, Nielsen JB, Refsgaard L, Holst AG, Christensen AH, Andreasen L, et al. New population-based exome data are questioning the pathogenicity of previously cardiomyopathy-associated genetic variants. Eur J Hum Genet. 2013;21:918-928.
35. Norton N, Li D, Rampersaud E, Morales A, Martin ER, Zuchner S, et al. Exome sequencing and genome-wide linkage analysis in 17 families illustrate the complex contribution of TTN truncating variants to dilated cardiomyopathy. Circ Cardiovasc Genet. 2013;6:144-153.
36. Lange LA, Hu Y, Zhang H, Xue C, Schmidt EM, Tang ZZ, et al. Whole-Exome Sequencing Identifies Rare and Low-Frequency Coding Variants Associated with LDL Cholesterol. Am J Hum Genet. 2014;94:233-245.
37. Peloso GM, Auer PL, Bis JC, Voorman A, Morrison AC, Stitziel NO, et al. Association of low-frequency and rare coding-sequence variants with blood lipids and coronary heart disease in 56,000 whites and blacks. Am J Hum Genet. 2014;94:223-232.
38. Auer PL, Teumer A, Schick U, O'Shaughnessy A, Lo KS, Chami N, et al. Rare and low-frequency coding variants in CXCR2 and other genes are associated with hematological traits. Nat Genet. 2014;46:629-634.
ndrome in new exomomomome
H Andrdrdrdreaeaeaeasesesesennnn LLLL eeeetttt aala-based exome data are questioning the pathogenicity of previously cardiomyopatg
N, Li D, Rampersaud E, Morales A, Martin ER, Zuchner S, et al. Exome sequene Nv
L nRare and Lo Freq enc Coding Variants Associated ith LDL Cholesterol A
-basssededede eeexoxoxomemm ddddataa a are questioning the papapaththtt ogenicity of preevivvv ously cardiomyopatgeneneneeete ic variiaiai ntnttnts.sss EuEuEuEur rrr JJJ HuHuHuH m m m m GeGeGeGenenenen t. 2020202 13;2;21:9191918-8-8-8-9299 8.8.8.8.
NNN,,, LLLLi D, RaRaRammpeeersaauuud EEE, MoM raraleees A,,, MMarrtiin ERERER, ZuZuchhhnner S,S,, et t aaal. Exxxomomomeee sequqquene-wiiiideedede llllininini kkkagege aananallylysis s ininn 11117777 famimimililililiesesese iiillllllllususttrtrt atatateee thtththeee coompmmpmplelel x ccontntntntrririribububub tiiionon oof fff TTTTTTNvariants to dilatatttededede ccccararara dididiiomomomo yoyoyoyopapapaaththththy.y.y.. CiCCiC rcrcrc CCCarararardidd ovvvvasassasccc c GeGeGeenenenenettt. 20202013131313;6;; :144-153.
LLLA, Hu YY,, ZhZZhZhanannang g gg H,HHH, XXXXueueueue CCCC,,, ScSSchmhmhmhmididididt tt t EMEMEMEM, , TaTaTT ngnggng ZZZZZ,Z,Z, eeet alalalal... WhWhWhWholollole-e-e-e-ExExExExomomoome SequenRRR dd LLo FF CC dodiin VVa iri ts AA iiatedd ii hth LLDLDL CChhollest lol AA by guest on July 3, 2018
http://circgenetics.ahajournals.org/D
ownloaded from

DOI: 10.1161/CIRCGENETICS.114.000718
17
Table 1: Characteristics EA/AA AF cases and non-AF controls
EA AF Cases EA non-AF Controls AA AF Cases AA non-AF
Controls
N 1,008 17,416 48 1,732
Age, (IQR) 78.1 (69.7-83.8) 69.9 (60.9-79.6) 71.3 (58.3-81.6) 65.5 (57.6-77.5)
Female gender, (%) 458 (46.0) 9088 (52.2) 26 (54.2) 1063 (61.4)
AF, atrial fibrillation; EA, European American; AA, African American, IQR, interquartile range R, interquartitititilllle rangegeee
by guest on July 3, 2018http://circgenetics.ahajournals.org/
Dow
nloaded from

DOI: 10.1161/CIRCGENETICS.114.000718
18
Table 2: Prevalence of 19 Variants Previously Associated with Lone or Familial Forms of AF
Gene AminoAcidChange
GenotypePhenotype
Segregation
EP function
Discovery”cohort”ancestry
EAMAF
Risk allelecarriers amongEA AF Cases,
(%)
Risk allele carriers among EA non-AF
Controls*, (%)
EAP†
AAMAF
Risk allele carriers
among AA AF Cases, (%)
Risk allele carriers among
AA non-AF Controls*, (%)
AAP† Ref
-AF controls:
NPPA Ser64Arg S LoF EA 0.003 5/1008 (0.50) 120/17415 (0.69) 0.69 0.0006 0/48 2/1732 (0.11) 0 7
GREM2 Gln76Glu
Increase-in-
inhibitory-effect
EA 0.004 10/1008 (1.00) 133/17415 (0.76) 0.46 0 0/48 0/1732 - 10
KCNE4 Glu141Ala Possible-change EA 0.0005 0/1008 19/17414 (0.11) 0.62 0 0/48 0/1732 - 9
SCN5A Phe2004Leu GoF EA 0.0052 13/1008 (1.30) 179/17414 (1.03) 0.42 0.0003 0/48 1/1731 (0.06) 0 2, 24
SCN5A Thr1304Met GoF EA 0.0009 0/1008 35/17416 (0.20) 0.26 0.0003 0/48 1/1732 (0.06) 0 8
SCN5A Arg1193Gln GoF EA 0.0010 4/1008 (0.40) 33/17416 (0.19) 0.14 0 0/48 0/1732 - 2, 24
SCN5A Leu618Phe - AA 0 0/1006 0/17381 - 0.0071 1/48 (2.1) 24/1725 (1.39) 0.50 2
SCN5A Ser524Tyr - AA 0.0005 0/1008 18/17416 (0.10) 0.62 0.0328 5/48 (10.4) 112/1732 (6.47) 0.25 2
SCN5A Arg481Trp - AA 8x10-5 0/1008 3/17416 (0.02) 1.00 0.0101 2/48 (4.2) 34/1732 (1.96) 0.25 2
SCN5A Leu461Val S - EA/AA 0.0001 1/1008 (0.10) 3/17382 (0.02) 0.20 0.0127 1/48 (2.01) 44/1727 (2.55) 1 2
SCN5A Thr220Ile LoF EA 0.0010 1/1008 (0.10) 35/17416 (0.20) 0.72 0.0006 0/48 2/1732 (0.12) 1 8, 11, 24
SCN5A Ser216Leu GoF EA 0.0015 3/1008 (0.30) 53/17416 (0.30) 1.00 0 0/48 0/1732 - 2, 24
GATA4 Ala411Val - EA 0.0015 1/1008 (0.10) 56/17410 (0.32) 0.37 0.0011 1/48 (2.1) 3/1731 (0.17) 0.10 4
KCNJ8 Ser422Leu GoF EA 0.0010 2/1008 (0.02) 36/17416 (0.21) 1.00 0 0/48 0/1732 - 5, 6
Variants present in <0.1% of EA/AA non-AF controls:
KCND3 Lys214Arg No Change EA 0.0004 0/1005 14/17380 (0.08) 1.00 0 0/48 0/1727 - 9
SCN5A Arg1897Trp LoF EA 8x10-5 1/1004 (0.10) 2/17353 (0.01) 0.16 0 0/48 0/1724 - 8
SCN5A Glu428Lys S - EA 0.0003 0/1008 12/17415 (0.07) 1.00 0 0/48 0/1732 - 2
KCNA5 Gly568Val S GoF EA 0.0002 0/1001 6/17329 (0.03) 1.00 0 0/48 0/1724 - 9
KCNJ2 Val93Ile S GoF Asian 2x10-5 0/1007 1/17420 (0.006) 1.00 0 0/48 0/1731 - 3
*Free from AF at 50 years old †Unadjusted p-values shown; the adjusted p-values (age, sex, PC1, and PC2) yielded similar estimatesS, Genotype-phenotype segregation; EP, electrophysiological; LoF, loss-of-function; GoF, gain-of-function; EA, European-American; AA, African-American; AF, atrial fibrillation
0.0.0.0.62626262 0 0 0 0
0.424222 0.0.0.0.0000000003030303
E
EAAAA 0....0000000009 0/1008 35353535/17416 (0.20) 0.26 0.0003
EAAAA 0.0.00.0000000010101010 4/4/4/4/1010101008080808 ((((0.0.00 4000)))) 333/117474747416161616 (00.0.0 1999))) 0....14141414 0 0 0
AAAAAAAA 000 0/10000006 0/0/0/0/17171 38381 -- 0.000071
AA 0.0000005000 0/1008 1811 /17416666 (0.10) 0.62 0.0328
AA 8x10101010-5555 0/0/0/0 101010100808080 3/3/3/3/1717174116666 (0(0(0(0.00002)2)2) 1.111 00000000 0.0101
EEEA/AA 0.000 0000000001010101 1/11 1010101008080808 ((((0.000 10101010)))) 3/3/3/3/17171717383838382222 (0(0(0(0.00002)2)2)2) 0.000 20202020 0.000 0127
by guest on July 3, 2018http://circgenetics.ahajournals.org/
Dow
nloaded from

DOI: 10.1161/CIRCGENETICS.114.000718
19
Table 3: No interaction between common AF risk variants at the 4q25 locus and 14 rare variants
previously implicated with lone or familial forms of AF among EAs
No AF, (%) AF, (%)
Absence of common 4q25 variants* 459 (60.5) 25 (3.3)
Presence of common 4q25 variants* 259 (34.2) 15 (2.0)
* rs6843082, rs2200733, and rs17042171 AF, atrial fibrillation; SNP, single nucleotide polymorphism, EAs, European Americans Chi-square statistic = 0.85
memememeriririricacacacansnsnsns
by guest on July 3, 2018http://circgenetics.ahajournals.org/
Dow
nloaded from

Dawood Darbar and Dan M. RodenPeter Weeke, Joshua C. Denny, Lisa Basterache, Christian Shaffer, Erica Bowton, Christie Ingram,
NoteFamilial Forms of Atrial Fibrillation in an Electronic Medical Record System: A Cautionary Examining Rare and Low-Frequency Genetic Variants Previously Associated with Lone or
Print ISSN: 1942-325X. Online ISSN: 1942-3268 Copyright © 2014 American Heart Association, Inc. All rights reserved.
TX 75231is published by the American Heart Association, 7272 Greenville Avenue, Dallas,Circulation: Cardiovascular Genetics
published online November 19, 2014;Circ Cardiovasc Genet.
http://circgenetics.ahajournals.org/content/early/2014/11/19/CIRCGENETICS.114.000718World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://circgenetics.ahajournals.org/content/suppl/2014/11/19/CIRCGENETICS.114.000718.DC1Data Supplement (unedited) at:
http://circgenetics.ahajournals.org//subscriptions/
is online at: Circulation: Cardiovascular Genetics Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer this process is available in the
located, click Request Permissions in the middle column of the Web page under Services. Further information aboutnot the Editorial Office. Once the online version of the published article for which permission is being requested is
can be obtained via RightsLink, a service of the Copyright Clearance Center,Circulation: Cardiovascular Genetics Requests for permissions to reproduce figures, tables, or portions of articles originally published inPermissions:
by guest on July 3, 2018http://circgenetics.ahajournals.org/
Dow
nloaded from

1
Supplemental Material

2
Supplemental Table 1 –Additional Rare and low frequency Variants Associated with Lone or Familial Forms of AF
Gene AminoAcid
Change CHR:POS REF/ALT
Accession Number
Rs# Ref
KCNQ1 p.Gly229Asp 11:2593245 G/A NM_000218.2 rs199472708 1 KCNQ1 p.Val241Phe 11:2593280 G/T NM_000218.2 NA 2 KCNJ2 p.Glu229Val 17:68172076 A/T NM_000891.2 NA 3
JPH2 p.Glu169Lys 20:42788922 C/T NM_020433.4 NA 4
3
Supplemental Table 2 –In-silico Functional Predictions of the 19 Variants Previously Associated with Lone or Familial Forms of AF
Gene AminoAcid
Change CHR:POS
REF/ALT
Accesion Number
Rs# PP2 Grantham
Score Phast Cons
GERP ESP EA ESP AA Ref.
Variants present in ≥0.1% of EA/AA non-AF controls: NPPA Ser64Arg 1:11907430 T/G NM_006172.3 61757261 0.024 110 0 3.54 G=24/T=8576 G=2/T=4402
5
GREM2 Gln76Glu 1:240656550 G/C NM_022469.3 142343894 0.437 29 0 5.03 C=27/G=8573 C=4/G=4402 6
KCNE4 Glu141Ala 2:223917970 A/C NM_080671.2 149110444 1 107 1 6.17 C=5/A=8595 C=0/A=4406 7
SCN5A Phe2004Leu 3:38591853 A/G NM_198056.2 41311117 0 22 0.992 -1.63 G=26/A=8342 G=2/A=4026 8, 9
SCN5A Thr1304Met 3:38603958 G/A NM_198056.2 199473603 1 81 0.89 4.04 A=4/G=8464 A=1/G=4175 10
SCN5A Arg1193Gln 3:38616876 C/T NM_198056.2 41261344 0.467 43 0.988 1.58 T=11/C=8589 T=0/C=440 8, 9
SCN5A Leu618Phe 3:38645241 G/A NM_198056.2 45488304 0.999 22 0.806 4.18 A=0/G=8416 A=27/G=4095 8
SCN5A Ser524Tyr 3:38645522 G/T NM_198056.2 41313691 0.998 144 1 4.06 T=7/G=8375 T=135/G=3945 8
SCN5A Arg481Trp 3:38646297 G/A NM_198056.2 144511230 0.999 101 0.902 3.29 A=0/G=8270 A=39/G=3897 8
SCN5A Leu461Val 3:38646357 A/C NM_198056.2 41313697 0.285 32 0.998 -0.65 C=2/A=8296 C=34/A=3914 8
SCN5A Thr220Ile 3:38655278 G/A NM_198056.2 45620037 1 89 1 4.3 A=4/G=8404 A=0/G=4032 9-11
SCN5A Ser216Leu 3:38655290 G/A NM_198056.2 41276525 1 145 1 4.3 A=11/G=8421 A=1/G=4089 8, 9
GATA4 Ala411Val 8:11615887 C/T NM_002052.3 55633527 0.012 64 0.001 4.83 T=12/C=8588 T=10/C=4396 12
KCNJ8 Ser422Leu 12:21918667 G/A NM_004982.3 72554071 0.013 145 0.038 4.45 A=19/G=8581 A=1/G=4405 13, 14
Variants present in <0.1% of EA/AA non-AF controls: KCND3 Lys214Arg 1:112524708 T/C NM_172198.2 142744204 0 26 1 5.51 C=4/T=8596 C=0/T=4406
7
SCN5A Arg1897Trp 3:38592174 G/A NM_198056.2 45465995 1 101 0.988 3.97 A=3/G=8413 A=0/G=4252 10
SCN5A Glu428Lys 3:38647498 C/T NM_198056.2 199473111 0.993 56 0.899 4.67 T=2/C=8372 T=0/C=4150 8
KCNA5 Gly568Val 12:5155016 G/T NM_002234.3 71581017 0 109 0.001 -1.72 T=3/G=8597 T=0/G=4406 7
KCNJ2 Val93Ile 17:68171457 G/A NM_000891.2 147750704 0.004 29 0.026 3.48 A=2/G=8598 A=1/G=4405 15
PP2, polyphen2; GERP, genomic evolution rate profiler; ESP, exome sequencing project; EA, European-American; AA, African-American; AF, atrial fibrillation PolyPhen-2: scores are evaluated as 0.000 (most probably benign) to 0.999 (most probably damaging).

4
Supplemental Figure 1 - Power calculation
Figure Legend: We employed a 1:18 case control ratio based on our study cohort (1,056 AF cases and 19,148 non-AF controls) assuming an additive gene-only model with a MAF=0.01 using a nominal two-sided p-value of <0.05 as our alpha. The effect sizes are reported as beta coefficient. Power calculations were performed using Quanto, (http://hydra.usc.edu/gxe/).

5
1. Hasegawa K, Ohno S, Ashihara T, Itoh H, Ding WG, Toyoda F, et al. A novel KCNQ1 missense mutation identified in a patient with juvenile-onset atrial fibrillation causes constitutively open IKs channels. Heart Rhythm. 2014;11:67-75
2. Ki CS, Jung CL, Kim HJ, Baek KH, Park SJ, On YK, et al. A KCNQ1 mutation causes age-dependant bradycardia and persistent atrial fibrillation. Pflugers Arch. 2014;466:529-540
3. Deo M, Ruan Y, Pandit SV, Shah K, Berenfeld O, Blaufox A, et al. KCNJ2 mutation in short QT syndrome 3 results in atrial fibrillation and ventricular proarrhythmia. Proc Natl Acad Sci U S A. 2013;110:4291-4296
4. Beavers DL, Wang W, Ather S, Voigt N, Garbino A, Dixit SS, et al. Mutation E169K in junctophilin-2 causes atrial fibrillation due to impaired RyR2 stabilization. Journal of the American College of Cardiology. 2013;62:2010-2019
5. Abraham RL, Yang T, Blair M, Roden DM, Darbar D. Augmented potassium current is a shared phenotype for two genetic defects associated with familial atrial fibrillation. J Mol Cell Cardiol. 2010;48:181-190
6. Muller, II, Melville DB, Tanwar V, Rybski WM, Mukherjee A, Shoemaker MB, et al. Functional modeling in zebrafish demonstrates that the atrial-fibrillation-associated gene GREM2 regulates cardiac laterality, cardiomyocyte differentiation and atrial rhythm. Dis Model Mech. 2013;6:332-341
7. Mann SA, Otway R, Guo G, Soka M, Karlsdotter L, Trivedi G, et al. Epistatic effects of potassium channel variation on cardiac repolarization and atrial fibrillation risk. Journal of the American College of Cardiology. 2012;59:1017-1025
8. Darbar D, Kannankeril PJ, Donahue BS, Kucera G, Stubblefield T, Haines JL, et al. Cardiac sodium channel (SCN5A) variants associated with atrial fibrillation. Circulation. 2008;117:1927-1935
9. Arnestad M, Crotti L, Rognum TO, Insolia R, Pedrazzini M, Ferrandi C, et al. Prevalence of long-QT syndrome gene variants in sudden infant death syndrome. Circulation. 2007;115:361-367
10. Olesen MS, Yuan L, Liang B, Holst AG, Nielsen N, Nielsen JB, et al. High prevalence of long QT syndrome-associated SCN5A variants in patients with early-onset lone atrial fibrillation. Circ Cardiovasc Genet. 2012;5:450-459
11. Olson TM, Michels VV, Ballew JD, Reyna SP, Karst ML, Herron KJ, et al. Sodium channel mutations and susceptibility to heart failure and atrial fibrillation. JAMA : the journal of the American Medical Association. 2005;293:447-454
12. Posch MG, Boldt LH, Polotzki M, Richter S, Rolf S, Perrot A, et al. Mutations in the cardiac transcription factor GATA4 in patients with lone atrial fibrillation. Eur J Med Genet. 2010;53:201-203
13. Medeiros-Domingo A, Tan BH, Crotti L, Tester DJ, Eckhardt L, Cuoretti A, et al. Gain-of-function mutation S422L in the KCNJ8-encoded cardiac K(ATP) channel Kir6.1 as a pathogenic substrate for J-wave syndromes. Heart Rhythm. 2010;7:1466-1471
14. Delaney JT, Muhammad R, Blair MA, Kor K, Fish FA, Roden DM, et al. A KCNJ8 mutation associated with early repolarization and atrial fibrillation. Europace. 2012;14:1428-1432

6
15. Xia M, Jin Q, Bendahhou S, He Y, Larroque MM, Chen Y, et al. A Kir2.1 gain-of-function mutation underlies familial atrial fibrillation. Biochem Biophys Res Commun. 2005;332:1012-1019