experimental evidence of a linear relationship between inorganic mercury loading and methylmercury...
TRANSCRIPT

Experimental Evidence of a LinearRelationship between InorganicMercury Loading and MethylmercuryAccumulation by Aquatic BiotaD I A N E M . O R I H E L , * , † , ‡
M I C H A E L J . P A T E R S O N , ‡
P A U L J . B L A N C H F I E L D , ‡
R . A ( D R E W ) B O D A L Y , ‡ , | A N DH O L G E R H I N T E L M A N N §
Clayton H. Riddell Faculty of Environment, Earth, andResources, University of Manitoba, Winnipeg, Manitoba,Canada R3T 2N2, Freshwater Institute, Fisheries and OceansCanada, 501 University Crescent, Winnipeg, Manitoba,Canada R3T 2N6, and Department of Chemistry, TrentUniversity, 1600 West Bank Drive, Peterborough, Ontario,Canada K9J 7B8
Developing effective regulations on mercury (Hg) emissionsrequires a better understanding of how atmospheric Hgdeposition affects methylmercury (MeHg) levels in aquaticbiota. This study tested the hypothesis that MeHgaccumulation in aquatic food webs is related to atmosphericHg deposition. We simulated a range of inorganic Hgdeposition rates by adding isotopically enriched Hg(II) (90.9%202Hg) to 10-m diameter mesocosms in a boreal lake.Concentrations of experimentally added (“spike”) Hg weremonitored in zooplankton, benthic invertebrates, andfish. Some Hg(II) added to the mesocosms was methylatedand incorporated into the food web within weeks,demonstrating that Hg(II) deposited directly to aquaticecosystems can become quickly available to biota.Relationships between Hg(II) loading rates and spikeMeHg concentrations in zooplankton, benthic invertebrates,and fish were linear and significant. Furthermore, spikeMeHg concentrations in the food web were directlyproportional to Hg(II) loading rates (i.e., a percent changein Hg(II) loading rate resulted in, statistically, the samepercent change in MeHg concentration). This is the firstexperimental determination of the relationship between Hg-(II) loading and MeHg bioaccumulation in aquatic biota.We conclude that changes in atmospheric Hg depositioncaused by increases or decreases in Hg emissions willultimately affect MeHg levels in aquatic food webs.
IntroductionDuring the last century, more than 200 million kilograms ofmercury (Hg) were emitted to the atmosphere from anthro-pogenic activities (1). Lake sediments and peat bogs reveal
that Hg levels in the environment have increased on a globalscale since the onset of industrialization (2). Currently, thelargest anthropogenic source of Hg emissions to the atmo-sphere is the combustion of coal in power plants and inresidential and commercial boilers (3). Mercury emitted fromthese sources can be dispersed long distances in theatmosphere and subsequently returned to the Earth’s surfacevia wet deposition of oxidized species [Hg(II)], although drydeposition may also be important in some areas (4). Naturalprocesses in lakes and their watersheds convert atmospheri-cally derived Hg(II) to the potent toxin methylmercury(MeHg) (5). Methylmercury is transferred through the dietand strongly biomagnifies in aquatic food webs, which resultsin high concentrations in fish and other top predators (5).
Proposals to reduce anthropogenic Hg emissions to theatmosphere are intended to lower MeHg levels in fish, butit is unclear whether these policies will be effective inachieving their goal. The amount by which Hg emissionsneed to be reduced to achieve an acceptable level of MeHgin fish remains a complex, unresolved issue. Developingeffective policies on Hg emissions requires models that canpredict how changes in atmospheric Hg deposition affectlevels of MeHg in fish. This relationship is difficult to quantifybecause many factors affect Hg cycling in aquatic ecosystems.Fish MeHg levels are correlated with several environmentalfactors, including watershed characteristics, lake morphom-etry, water chemistry, and food web structure (6-8). Becausethis suite of factors inevitably varies from one ecosystem tothe next, lakes receiving similar rates of Hg deposition mayhave quite different fish MeHg levels (9).
We tested the hypothesis that MeHg concentrations inaquatic biota are related to atmospheric Hg deposition bysimulating different rates of Hg deposition in a boreal lake.We isolated sections of the lake in large mesocosms andadded a different amount of isotopically enriched Hg(II) toeach mesocosm. Adding Hg(II) enriched in a stable isotopeallowed us to monitor the methylation and subsequentbioaccumulation of the experimentally added (“spike”) Hg.In previous papers on this experiment (known as theMESOSIM project), we concluded that major biogeochemicalprocesses, including methylation, were Hg(II)-limited andresponded linearly to increases in Hg(II) loading (10, 11). Inthis paper, we examine the bioaccumulation of spike MeHgin the food webs of the mesocosms. First, we describe howquickly spike MeHg was incorporated into the food web.Second, we analyze the dose-response relationship betweenrates of Hg(II) loading and concentrations of spike MeHg inzooplankton, benthic invertebrates, and fish. Specifically,we test the hypothesis that the bioaccumulation of spikeMeHg in the food web was directly proportional to the rateof Hg(II) loading. By this, we mean that a percent change inHg(II) loading rate resulted in the same percent change inthe spike MeHg concentrations in biota.
Experimental SectionStudy Design. Mesocosms were installed in a 2-m deep littoralzone of Lake 240 at the Experimental Lakes Area (ELA;Ontario, Canada). The construction and physicochemicalcharacteristics of the mesocosms were described previously(10, 12). Briefly, mesocosms were 10 m in diameter, open tothe atmosphere and sediments, and contained oligotrophic,circumneutral water and sandy sediments. As described inref 10, isotopically enriched Hg(II) (90.9% 202Hg) was addedto the mesocosms once a week for 8 weeks, beginning onJune 26, 2002. Each mesocosm was randomly assigned aloading rate of 1×, 2×, 3×, 4×, 5×, 6×, 7×, 8×, 12×, or 15×
* Corresponding author phone: 204-984-8751; fax: 204-984-2404;e-mail: [email protected].
† University of Manitoba.‡ Fisheries and Oceans Canada.§ Trent University.| Current affiliation: Penobscot River Mercury Study (Maine), 115
Oystercatcher Place, Salt Spring Island, British Columbia, CanadaV8K 2W5.
Environ. Sci. Technol. 2007, 41, 4952-4958
4952 9 ENVIRONMENTAL SCIENCE & TECHNOLOGY / VOL. 41, NO. 14, 2007 10.1021/es063061r CCC: $37.00 2007 American Chemical SocietyPublished on Web 06/19/2007

the wet deposition rate of total Hg at the ELA in the 1999water year (7.1 µg Hg m-2 year-1) (13). All references to “Hg-(II) loading rate” refer to the rate of experimental Hg(II)addition.
Zooplankton and Benthic Invertebrate Sampling. Be-tween June 26 and September 4, 2002, zooplankton werecollected every 2 weeks between 13:00 and 16:00 with a sweepnet (150-µm mesh). Because some non-zooplankton materialwas inadvertently collected, the term “zooplankton” refersto all particles in the water column greater than 150 µm.
Between September 6 and 11, 2002, benthic invertebrateswere collected using a petit ponar dredge and light traps.Benthic invertebrates were sampled only in September toavoid disturbing the sediments during the experiment.Dredge samples were collected from six different locationsin each mesocosm and then sieved as described in ref 12.Light traps (12) were set on the sediment surface after duskand collected at dawn. In the laboratory, benthic invertebratesfrom dredges and light traps were sorted and combined intomajor taxonomic groups.
Eight benthic invertebrate taxa were common in allmesocosms: Amphipoda (scud; generalist), Gomphidae(dragon fly larva; predator), Hexagenia (mayfly nymph;collector-gatherer), Hydracarina (water mite; predator),Oligochaeta (aquatic worm; detritivore), Planorbidae (ram-horn snail; herbivore/detritivore), Tanypodinae (midge flylarva; predator), and other Chironomidae (midge fly larva;herbivore/detritivore). Three taxa (Amphipoda, Gomphidae,and Hydracarina) were analyzed in all mesocosms. The otherfive taxa were only analyzed in mesocosms 2×, 5×, and 12×.
Zooplankton and benthic invertebrate samples werestored in sealed plastic bags and frozen within 3-6 h ofcollection. Samples were freeze-dried, homogenized, weighedto the nearest 0.001 mg in aluminum weighing vessels, andtransferred to Teflon vials for MeHg analysis.
Fish Stocking and Sampling. On June 24-26, 2002, yellowperch (Perca flavescens) were captured from Lake 240 usinga beach seine, marked with a caudal clip, and stocked in themesocosms (40 fish per mesocosm). We only stocked fishwith fork lengths between 50 and 60 mm, which is the typicalsize range for age 1 yellow perch in Lake 240 in early summer(14). Gut contents of yellow perch of a similar size rangestocked in enclosures in an adjacent lake contained, byweight, 66% zooplankton and 31% benthic invertebrates (15).Mesocosms were covered with fine plastic netting (5-cmmesh) to deter predators.
Yellow perch were collected after 5 weeks (July 31-August2, 2002) and 10 weeks (September 4-16, 2002) using minnowtraps and gill nets. A beach seine was also used in Septemberto remove any remaining fish. At each sampling period, 5-10fish were typically sampled from each mesocosm. Fish werekilled by a lethal dose of clove oil, measured (fork length) tothe nearest 1 mm, and weighed to the nearest 0.1 g. A skinlesssection of muscle tissue (ca. 0.2 g) was dissected from eachfish, weighed to the nearest 0.0001 g, transferred to a 20 mLglass vial, and stored frozen until analysis.
Although the mesocosms were fished with minnow trapsand gill nets (6-10 mm mesh) before stocking, some young-of-the-year yellow perch were captured in some mesocosmsin September. Saggital otoliths were examined (as describedin ref 12) to verify the ages of all fish captured during thissampling period.
Mercury Analyses. Concentrations of Hg species weredetermined by inductively coupled plasma mass spectrom-etry (ICP-MS) at Trent University (Peterborough, Ontario,Canada) (16). Methylmercury was measured in compositesamples of zooplankton and benthic invertebrates, and totalHg was measured in muscle samples of individual fish. TotalHg was measured in fish samples because most of the Hg infish muscle is MeHg (17) and this is the standard protocol
in contaminant monitoring programs (e.g., 9). We confirmedthat the large majority of total Hg in muscle tissue of yellowperch was MeHg (mean ( SE: 84 ( 1%; n ) 13). Samplesof zooplankton and benthic invertebrates were digested byadding 5 mL of 4 M HNO3 and heating at 55 °C for 24 h (18).Aliquots of sample digests (50-5000 µL) were added to 100mL of Milli-Q water in gas-wash bottles for purging. Afteradding 0.2 mL of 2 M acetate buffer, solutions wereneutralized with 20% KOH to obtain a pH of 4.9. Methyl-mercury was ethylated with tetraethyl borate (1%, w/v in 1%KOH, w/v), purged and concentrated on Tenax traps,thermodesorbed, and determined by GC-ICP-MS (Micro-mass Platform). Fish muscle samples were digested by adding10 mL of HNO3/H2SO4 (7:3 v/v) and heating at 80 °C untilbrown NOx gases no longer formed. Total Hg of sample digestswas reduced by SnCl2 and determined by ICP-MS (Thermo-Finnigan Element2) using a continuous flow cold vaporgeneration technique.
Method blanks and certified reference materials weremeasured for each batch of samples. Results for MeHg inoyster tissue (measured, 13.5 ( 1.7 ng g-1; certified, 13.2 (0.7 ng g-1) and total Hg in DORM-3 (measured, 4680 ( 240ng g-1; certified, 4640 ( 260 ng g-1) were not statisticallydifferent from certified values. No isotope enrichment wasdetected in samples collected from the control mesocosm.
The detection limit of ambient Hg (i.e., Hg not addedexperimentally) was 0.02-1.0 ng g-1 dry weight (dw) for MeHgin zooplankton and benthic invertebrates (depending on thesample mass available for measurement) and 0.2 ng g-1 wetweight (or 1.0 ng g-1 dw) for total Hg in fish. Mercuryconcentrations in fish were measured in wet muscle tissueand converted to a dry weight basis assuming a moisturecontent of 80% based on the average value determined fora subset of fish (mean ( SE: 80.1 ( 0.2%; n ) 24). In earlySeptember, ambient Hg concentrations across the tenmesocosms that received Hg(II) additions ranged as follows:from 15.4 to 84.0 ng MeHg g-1 dw for zooplankton; from 20.7to 40.4, 56.2 to 81.8, and 14.8 to 104.6 ng MeHg g-1 dw forthe benthic invertebrate taxa Amphipoda, Gomphidae, andHydracarina, respectively; and from 146 to 1057 ng Hg g-1
dw for age 1 yellow perch. The detection limit of spike Hg(i.e., Hg added experimentally) was 0.5% of each sample’sambient Hg concentration. If spike Hg was not detected ina sample, the concentration was estimated as one-half of thedetection limit.
Data Analysis. All data analyses were performed withSTATISTICA 6.1 (StatSoft, Inc.). Model I linear regressionwas used to quantify the relationship between Hg(II) loadingrates and spike MeHg concentrations in biota. For fish data,we used the mean spike Hg concentration of each mesocosmbecause concentrations were not correlated with fish size.All variables were log10-transformed and residuals wereexamined for linearity, homoscedasticity, and normality. Foreach regression model, an F-test was performed to determinewhether the relationship between the variables was significant(Ho: the slope is zero) and a two-tailed t-test was performedto determine whether the relationship between the variableswas directly proportional (Ho: the slope is one). Because adirectly proportional relationship (Y ) aX) also takes theform logY ) loga + 1logX, a slope of one in a log-logrelationship indicates that the relationship between thevariables is directly proportional (i.e., a percent change inthe independent variable results in the same percent changein the dependent variable).
ResultsZooplankton. Two weeks after Hg(II) loading began, zoop-lankton contained detectable levels of spike MeHg inmesocosms receiving higher loading rates (Figure 1). By 4weeks, spike MeHg was detected in zooplankton from all
VOL. 41, NO. 14, 2007 / ENVIRONMENTAL SCIENCE & TECHNOLOGY 9 4953
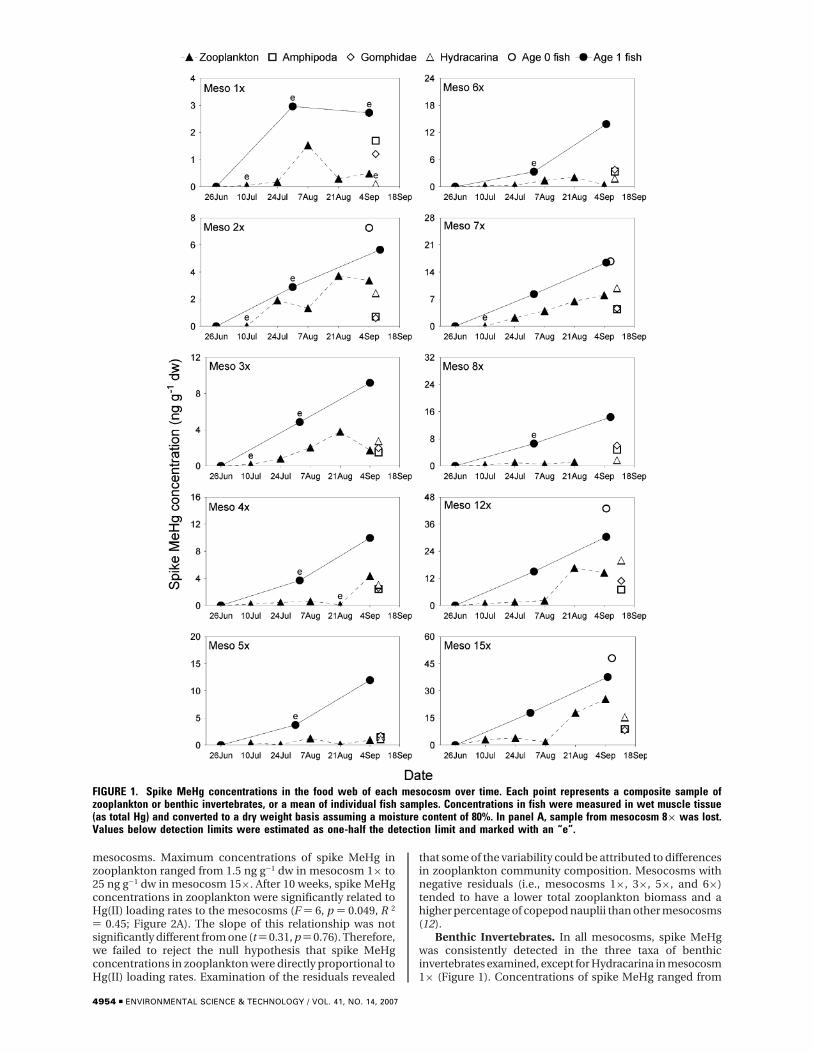
mesocosms. Maximum concentrations of spike MeHg inzooplankton ranged from 1.5 ng g-1 dw in mesocosm 1× to25 ng g-1 dw in mesocosm 15×. After 10 weeks, spike MeHgconcentrations in zooplankton were significantly related toHg(II) loading rates to the mesocosms (F ) 6, p ) 0.049, R 2
) 0.45; Figure 2A). The slope of this relationship was notsignificantly different from one (t ) 0.31, p ) 0.76). Therefore,we failed to reject the null hypothesis that spike MeHgconcentrations in zooplankton were directly proportional toHg(II) loading rates. Examination of the residuals revealed
that some of the variability could be attributed to differencesin zooplankton community composition. Mesocosms withnegative residuals (i.e., mesocosms 1×, 3×, 5×, and 6×)tended to have a lower total zooplankton biomass and ahigher percentage of copepod nauplii than other mesocosms(12).
Benthic Invertebrates. In all mesocosms, spike MeHgwas consistently detected in the three taxa of benthicinvertebrates examined, except for Hydracarina in mesocosm1× (Figure 1). Concentrations of spike MeHg ranged from
FIGURE 1. Spike MeHg concentrations in the food web of each mesocosm over time. Each point represents a composite sample ofzooplankton or benthic invertebrates, or a mean of individual fish samples. Concentrations in fish were measured in wet muscle tissue(as total Hg) and converted to a dry weight basis assuming a moisture content of 80%. In panel A, sample from mesocosm 8× was lost.Values below detection limits were estimated as one-half the detection limit and marked with an “e”.
4954 9 ENVIRONMENTAL SCIENCE & TECHNOLOGY / VOL. 41, NO. 14, 2007

0.7 to 9.0 ng g-1 dw in Amphipoda, from 0.6 to 10.9 ng g-1
dw in Gomphidae, and from <0.2 to 19.9 ng g-1 dw inHydracarina. The relationship between Hg(II) loading rateand spike MeHg concentration across mesocosms was highlysignificant for Amphipoda (F ) 19, p ) 0.002, R 2 ) 0.71;Figure 2B), Gomphidae (F ) 31, p ) 0.0005, R 2 ) 0.79; Figure2C), and Hydracarina (F ) 16, p ) 0.004, R 2 ) 0.66; Figure2D). For Hydracarina, excluding the estimated data pointweakened the relationship (F ) 4.5, p ) 0.07, R 2 ) 0.39). Theslope of the relationship between Hg(II) loading rate andspike MeHg concentration was not significantly different fromone for Amphipoda (t ) -1.0, p ) 0.35), Gomphidae (t )-0.13, p ) 0.90), or Hydracarina (t ) 1.3, p ) 0.22). Hence,we failed to reject the null hypothesis that spike MeHgconcentrations in benthic invertebrates were directly pro-portional to Hg(II) loading rates.
In mesocosms 2×, 5×, and 12×, spike MeHg wasconsistently detected in the eight taxa of benthic invertebrates
examined (Figure 3). Differences in spike MeHg concentra-tions among taxa were between 25- and 50-fold within eachmesocosm. Concentrations in benthic invertebrates rangedfrom 0.05 to 2.4 ng g-1 dw in mesocosm 2×, from 0.1 to 2.6ng g-1 dw in mesocosm 5×, and from 0.4 to 19.9 ng g-1 dwin mesocosm 12×. In general, herbivorous and detritivoroustaxa had the lowest concentrations and predatory taxa hadthe highest concentrations. Spike MeHg concentrations inmost taxa followed the rank order mesocosm 2×< mesocosm5× < mesocosm 12× (Figure 3). Spike MeHg concentrationsof benthic invertebrates in mesocosm 5× were lower thanexpected based on its loading rate, which is consistent withour previous observation that a higher percentage of mercuryevaded from mesocosm 5× than from mesocosms 2× and12× (11).
Fish. For age 1 yellow perch collected after 5 weeks, spikeHg was detected in none of the fish from mesocosm 1×, insome of the fish from mesocosms 2×, 3×, 4×, 5×, 6×, and8×, and in all fish from mesocosms 7×, 12×, and 15× (Figure1). By 10 weeks, all age 1 fish contained detectable levels ofspike Hg, except for some fish from mesocosm 1×. Spike Hgin age 1 fish increased over time, with concentrationsapproximately doubling between 5 and 10 weeks (Figure 1).After 10 weeks, average concentrations of spike Hg in age 1fish ranged from 2.5 ng g-1 dw in mesocosm 1× to 37.5 ngg-1 dw in mesocosm 15×. Age 0 yellow perch inadvertentlytrapped in some mesocosms had spike Hg concentrationssimilar to, or higher than, those of age 1 fish from the samemesocosm. Concentrations in age 0 and age 1 fish wereconsistently higher than those in zooplankton and benthicinvertebrates (Figure 1).
Spike Hg concentrations in yellow perch were stronglyrelated to rates of Hg(II) loading. After 10 weeks, therelationship between Hg(II) loading rate and spike Hgconcentration in age 1 fish was positive, linear, and highlysignificant (F ) 297, p < 0.00001, R 2 ) 0.97; Figure 2E). Themean concentration for mesocosm 1× was calculated usingsome estimated concentrations, but excluding this value fromthe analysis had little influence on the result. Spike Hgconcentrations in age 0 fish collected in four mesocosmsafter 10 weeks were also positively related to Hg(II) loadingrates (Y ) -0.17 + 0.96X, F ) 42, p ) 0.02, R 2 ) 0.95). Theslope of the relationship between Hg(II) loading rate andspike Hg concentration was not significantly different fromone for either age 1 fish (t ) -1.6, p ) 0.14) or age 0 fish (t) -0.26, p ) 0.82). Therefore, we failed to reject the nullhypothesis that spike Hg concentrations in fish were directlyproportional to Hg(II) loading rates.
DiscussionTime Dynamics. Our first objective was to examine the timerequired for inorganic Hg added to mesocosms in a boreallake to be incorporated into the food web as MeHg. Wereported previously that some of the Hg(II) added to themesocosms was rapidly transported to sites of methylationand converted to MeHg (10). Here we show that spike MeHgwas then promptly bioaccumulated by aquatic biota in themesocosms. Spike MeHg was detected in zooplankton after2-4 weeks and in virtually all benthic invertebrates after 10weeks. Furthermore, spike Hg was detected in some fish after5 weeks and in nearly all fish after 10 weeks.
Our finding that newly added Hg(II) was converted to amethylated form and incorporated into the food web withinweeks is consistent with previous experimental studies.Radioactively labeled Hg (203Hg) added to enclosures in ClayLake, Ontario, was measured in zooplankton, crayfish(Orconectes virilis), and pearl dace (Margariscus margarita)within 1 week (19). Similarly, 203Hg added as a single pulseto the epilimnion of Lakes 224 (20) and 226 (21) at the ELAwas detected in various fish species within 1 week. In
FIGURE 2. Relationship between Hg(II) loading rate and spike MeHgconcentration in the food web. Samples were collected September4-16, 2002. Both variables are log transformed, axes are scaled sothat a slope of one has an angle of 45°, and regression lines areshown with 95% confidence bands. Each panel provides the resultsfrom the F-test, which was performed to determine whether therelationship between the variables was significant (Ho: the slopeis zero). Comments in the caption for Figure 1 also apply to thisfigure.
VOL. 41, NO. 14, 2007 / ENVIRONMENTAL SCIENCE & TECHNOLOGY 9 4955

experiments that added Hg enriched in a stable isotope toenclosures (15) or a whole lake (22), the added Hg wasdetected in zooplankton within 2-4 weeks, and in benthicinvertebrates and forage fish within 4-8 weeks.
Our experiment and earlier studies suggest that atmo-spheric Hg deposited to the surface of aquatic ecosystemsmay be quickly incorporated into food webs. This conclusiondoes not imply that MeHg concentrations in aquatic foodwebs will necessarily recover quickly after implementationof Hg emission controls. The response of MeHg levels inbiota to reductions in Hg emissions will depend on how longpreviously deposited Hg stored in lakes and their watershedscontinues to be available to aquatic food webs. This issue ispoorly understood, but previous declines in Hg loadingsuggest that MeHg levels in fish in some aquatic ecosystems
begin to respond within a few years of Hg reductions. Forexample, Hg concentrations in fish decreased significantlywithin 3-5 years after aqueous Hg discharges from chlor-alkali plants or other industries were drastically reduced orstopped (23-28). Mercury levels in fish at some of thesecontaminated sites, however, remained higher than back-ground levels for more than a decade (26-28). Mercuryconcentrations in fish also decreased significantly in less than10 years after Hg emission controls resulted in lower ratesof atmospheric Hg deposition in parts of Sweden and theUnited States (29-33), although simultaneous changes inother environmental factors confound many of these results.
Dose Response. Our second objective was to examinethe relationship between Hg(II) loading rate and accumula-tion of spike MeHg in the aquatic food web. Previously, we
FIGURE 3. Spike MeHg concentrations of benthic invertebrates in mesocosms 2×, 5×, and 12×. Samples were collected September 6-11,2002. Chironomidae includes all chironomids except Tanypodinae. No oligochaetes were collected from mesocosm 12×. Note differentY-axis scales.
4956 9 ENVIRONMENTAL SCIENCE & TECHNOLOGY / VOL. 41, NO. 14, 2007

concluded that production of spike MeHg was significantlyrelated to rates of Hg(II) loading to the mesocosms (10). Here,we extend this finding to the food web. We observed asignificant relationship between rates of Hg(II) loading andconcentrations of spike MeHg in zooplankton, benthicinvertebrates, and fish. Our study provides the first experi-mental evidence of the relationship between loading ratesof Hg(II) to an aquatic system and the bioaccumulation ofMeHg in an aquatic food web.
Our findings corroborate those of previous studies thatexamined whether atmospheric Hg deposition could explainspatial differences in MeHg concentrations in biota (34-39).These studies found that MeHg levels in fish, birds, or insectsacross regions of North America or Europe were correlatedwith rates of atmospheric Hg deposition or other indicatorsof Hg deposition. The results of these spatial analyses,together with the experimental evidence from our study,support the hypothesis that MeHg concentrations in aquaticbiota are related to atmospheric Hg deposition.
In our study, we explicitly tested the hypothesis that spikeMeHg concentrations in the food web were directly propor-tional to Hg(II) loading rates to the mesocosms. For all levelsof the food web, we failed to reject the null hypothesis thatthe log-log relationship between Hg(II) loading rate andspike MeHg concentration in biota had a slope of one. Thus,we conclude that concentrations of spike MeHg in the foodweb of the mesocosms were directly proportional to the ratesof Hg(II) loading. To explore this hypothesis on a broaderscale, we also analyzed data on Hg concentrations inlargemouth bass (Micropterus salmoides) in 25 U.S. states(38). Using Model II regression (40), we determined that thelog-log relationship between the mean annual rate of wetatmospheric Hg deposition and the mean state-wide con-centration of Hg in largemouth bass had a slope of 0.75 (ifthe three outliers omitted by ref 38 were not included in themodel). This slope is not significantly different from one (t) 0.95, p ) 0.3), which agrees with the results of ourexperimental study.
The generalization of our findings relies on severalassumptions, which were discussed in part in refs 10 and 15.Briefly, we assume that (i) the Hg(II) added to the mesocosmsrealistically simulates atmospherically derived Hg after wetdeposition to a lake surface; (ii) the mesocosms are repre-sentative of some aquatic ecosystems (particularly, shallowseepage lakes); and (iii) the dose-response relationshipwould have remained directly proportional had the experi-ment continued to steady-state. The latter is supported bythe fact that we did not observe any saturation effects on Hgmethylation or bioaccumulation, even at the highest loadingrates.
Because this experiment substitutes space for time (i.e.,each mesocosm represents the same lake at different pointsin time), our findings may indicate how MeHg concentrationsin aquatic biota within a single lake respond to changes inatmospheric Hg deposition. We suggest that concentrationsof MeHg in biota should respond in direct proportion tochanges in atmospheric Hg deposition in aquatic ecosystemswhere atmospheric Hg deposition is the primary source ofHg. We suspect, however, that concentrations of MeHg inbiota will respond (at least initially) in less than directproportion to changes in atmospheric Hg deposition inaquatic ecosystems primarily supplied by previously depos-ited Hg stored in lakes and their watersheds.
We conclude that Hg(II) added to littoral mesocosms ina boreal lake was converted to MeHg and bioaccumulatedby the food web in direct proportion to the loading rate. Theloading rates applied in our study simulated a broad rangeof atmospheric Hg deposition (7-107 µg Hg m-2 year-1),which largely encompasses the global range in depositionrates. These experimental results, together with those of
previous studies that have found spatial correlations betweenHg deposition and Hg concentrations in biota, demonstratethat atmospheric Hg deposition affects MeHg bioaccumu-lation in food webs. Consequently, MeHg concentrations infish and other biota should ultimately respond to changesin atmospheric Hg deposition resulting from increases ordecreases in anthropogenic Hg emissions.
AcknowledgmentsThis is contribution number 26 in the METAALICUS seriesof publications. This research was supported by the Col-laborative Mercury Research Network and the Electric PowerResearch Institute. D.O. received scholarships from theNational Science and Engineering Research Council ofCanada, the ELA Graduate Fellowship Fund, and theUniversity of Manitoba. We thank Danielle Godard, JaneOrihel, Marilynn Kullman, Ken Sandilands, and many otherELA students and staff for their help in the field; MichelleDobrin and Cheryl Podemski for advice on benthic inver-tebrate sampling; Rick Wastle and John Babaluk for providingfacilities and training for ageing fish; John Rudd and ReedHarris for their input on the experimental design; and ReedHarris, Carol Kelly, Lisa Loseto, Mariah Mailman, Jane Orihel,Gary Stern, Jill Van Walleghem, and three anonymousreviewers for helpful comments on earlier drafts of thismanuscript.
Literature Cited(1) Mason, R. P.; Fitzgerald, W. F.; Morel, F. M. M. The bio-
geochemical cycling of elemental mercury: Anthropogenicinfluences. Geochim. Cosmochim. Acta 1994, 58, 3191-3198.
(2) Fitzgerald, W. F.; Engstrom, D. R.; Mason, R. P.; Nater, E. A. Thecase for atmospheric mercury contamination in remote areas.Environ. Sci. Technol. 1998, 32, 1-7.
(3) Pacyna, E. G.; Pacyna, J. M.; Steenhuisen, F.; Wilson, S. Globalanthropogenic mercury emission inventory for 2000. Atmos.Environ. 2006, 40, 4048-4063.
(4) Fitzgerald, W. F.; Lamborg, C. H. Geochemistry of Mercury inthe Environment. In Treatise on Geochemistry; Elsevier Ltd.:Amsterdam, 2003; Vol. 9, pp 107-148.
(5) Wiener, J. G.; Krabbenhoft, D. P.; Heinz, G. H.; Scheuhammer,A. M. Ecotoxicology of Mercury. In Handbook of Ecotoxicology;Hoffman, D. J., Rattner, B. A., Burton, G. A., Jr., Cairns, J., Jr.,Eds.; Lewis Publishers: Boca Raton, FL, 2003; pp 409-463.
(6) Cabana, G.; Tremblay, A.; Kalff, J.; Rasmussen, J. B. Pelagic foodchain structure in Ontario lakes: A determinant of mercurylevels in lake trout (Salvelinus namaycush). Can. J. Fish. Aquat.Sci. 1994, 51, 381-389.
(7) Greenfield, B. K.; Hrabik, T. R.; Harvey, C. J.; Carpenter, S. R.Predicting mercury levels in yellow perch: Use of waterchemistry, trophic ecology, and spatial traits. Can. J. Fish. Aquat.Sci. 2001, 58, 1419-1429.
(8) Sorensen, J. A.; Glass, G. E.; Schmidt, K. W.; Huber, J. K.; Rapp,G. R., Jr. Airborne mercury deposition and watershed charac-teristics in relation to mercury concentrations in water, sedi-ments, plankton, and fish of eighty northern Minnesota lakes.Environ. Sci. Technol. 1990, 24, 1716-1727.
(9) Kamman, N. C.; Burgess, N. M.; Driscoll, C. T.; Simonin, H. A.;Goodale, W.; Linehan, J.; Estabrook, R.; Hutcheson, M.; Major,A.; Scheuhammer, A. M.; Scruton, D. A. Mercury in freshwaterfish of northeast North America - A geographic perspective basedon fish tissue monitoring databases. Ecotoxicology 2005, 14,163-180.
(10) Orihel, D. M.; Paterson, M. J.; Gilmour, C. C.; Bodaly, R. A.;Blanchfield, P. J.; Hintelmann, H.; Harris, R. C.; Rudd, J. W. M.Effect of loading rate on the fate of mercury in littoralmesocosms. Environ. Sci. Technol. 2006, 40, 5992-6000.
(11) Poulain, A. J.; Orihel, D. M.; Amyot, M.; Paterson, M. J.;Hintelmann, H.; Southworth, G. R. Relationship between theloading rate of inorganic mercury to aquatic ecosystems anddissolved gaseous mercury production and evasion. Chemo-sphere 2006, 65, 2199-2207.
(12) Orihel, D. M. The effects of changes in atmospheric mercurydeposition on the bioaccumulation of mercury by fish; NaturalResources Institute, University of Manitoba: Winnipeg, MB,2005; p 190.
VOL. 41, NO. 14, 2007 / ENVIRONMENTAL SCIENCE & TECHNOLOGY 9 4957

(13) St. Louis, V. L.; Rudd, J. W. M.; Kelly, C. A.; Hall, B. D.; Rolfhus,K. R.; Scott, K. J.; Lindberg, S. E.; Dong, W. Importance of theforest canopy to fluxes of methyl mercury and total mercury toboreal ecosystems. Environ. Sci. Technol. 2001, 35, 3089-3098.
(14) Blanchfield, P. J. Fisheries and Oceans Canada, Winnipeg, MB,Canada. Personal communication, 2002.
(15) Paterson, M. J.; Blanchfield, P. J.; Podemski, C.; Hintelmann, H.H.; Gilmour, C. C.; Harris, R.; Ogrinc, N.; Rudd, J. W. M.;Sandilands, K. A. Bioaccumulation of newly deposited mercuryby fish and invertebrates: An enclosure study using stablemercury isotopes. Can. J. Fish. Aquat. Sci. 2006, 63, 2213-2224.
(16) Hintelmann, H.; Ogrinc, N. Determination of stable mercuryisotopes by ICP/MS and their application in environmentalstudies. ACS Symp. Ser. 2003, 835, 321-338.
(17) Bloom, N. S. On the chemical form of mercury in edible fishand marine invertebrate tissue. Can. J. Fish. Aquat. Sci. 1992,49, 1010-1017.
(18) Hintelmann, H.; Nguyen, H. T. Extraction of methylmercuryfrom tissue and plant samples using acid leaching. Anal. Bioanal.Chem. 2005, 381, 360-365.
(19) Rudd, J. W. M.; Turner, M. A. The English-Wabigoon Riversystem: V. Mercury and selenium bioaccumulation as a functionof aquatic primary productivity. Can. J. Fish. Aquat. Sci. 1983,40, 2251-2259.
(20) Harrison, S. E.; Klaverkamp, J. F.; Hesslein, R. H. Fates of metalradiotracers added to a whole lake: Accumulation in fatheadminnow (Pimephales promelas) and lake trout (Salvelinusnamaycush). Water, Air, Soil Pollut. 1990, 52, 277-293.
(21) Bird, G. A.; Hesslein, R. H.; Mills, K. H.; Schwartz, W. J.; Turner,M. A. Bioaccumulation of radionuclides in fertilized CanadianShield lake basins. Sci. Total Environ. 1998, 218, 67-83.
(22) Blanchfield, P.; Paterson, M.; Podemski, C.; Hintelmann, H. Themovement of newly deposited mercury through an aquatic foodweb. RMZ - Mater. Geoenviron. 2004, 51, 838-840.
(23) Olsson, M. Mercury level as a function of size and age in northernpike, one and five years after the mercury ban in Sweden. Ambio1976, 5, 73-76.
(24) Armstrong, F. A. J.; Scott, D. P. Decrease in mercury content offishes in Ball Lake, Ontario, since imposition of controls onmercury discharges. J. Fish. Res. Board Can. 1979, 36, 670-672.
(25) Busch, W.-D. N. Decline of mercury in young fishes from westernLake Erie between 1970-71 and 1974. Prog. Fish-Cult. 1983, 45,202-206.
(26) Parks, J. W.; Hamilton, A. L. Accelerating recovery of the mercury-contaminated Wabigoon/English River system. Hydrobiologia1987, 149, 159-188.
(27) Herut, B.; Hornung, H.; Kress, N.; Cohen, Y. Environmentalrelaxation in response to reduced contaminant input: The caseof mercury pollution in Haifa Bay, Isreal. Mar. Pollut. Bull. 1996,32, 366-373.
(28) Francesconi, K. A.; Lenanton, R. C. J.; Caputi, N.; Jones, S. Long-term study of mercury concentrations in fish following cessationof mercury-containing discharge. Mar. Environ. Res. 1997, 43,27-40.
(29) Johansson, K.; Bergback, B.; Tyler, G. Impact of atmosphericlong range transport of lead, mercury and cadmium on theSwedish forest environment. Water, Air, Soil Pollut.: Focus 2001,1, 279-297.
(30) Watras, C. J.; Morrison, K. A.; Hudson, R. J. M.; Frost, T. M.;Kratz, T. K. Decreasing mercury in northern Wisconsin: Tem-poral patterns in bulk precipitation and a precipitation-dominant lake. Environ. Sci. Technol. 2000, 34, 4051-4057.
(31) Hrabik, T. R.; Watras, C. J. Recent declines in mercuryconcentration in a freshwater fishery: Isolating the effects ofde-acidification and decreased atmospheric mercury depositionin Little Rock Lake. Sci. Total Environ. 2002, 297, 229-237.
(32) Atkeson, T. D.; Pollman, C. D.; Axelrad, D. M. Recent Trends inMercury Emissions, Deposition, and Biota in the FloridaEverglades: A Monitoring and Modeling Analysis; In Dynamicsof Mercury Pollution on Regional and Global Scales: AtmosphericProcesses and Human Exposures Around the World; Pirrone, N.,Mahaffey, K. R., Eds.; Springer Science+Business Media, Inc.:New York, 2005; pp 637-653.
(33) Massachusetts Department of Environmental Protection. Masa-chussetts Fish Tissue Mercury Studies: Long-term MonitoringResults, 1999 - 2004; Office of Research and Standards, Boston,MA, and Wall Experimental Station, Lawrence, MA, 2006; http://mass.gov/dep/toxics/stypes/hgtrend.pdf (accessed October 20,2006).
(34) Johnson, M. G. Trace element loadings to sediments of fourteenOntario lakes and correlations with concentrations in fish. Can.J. Fish. Aquat. Sci. 1987, 44, 3-13.
(35) Hakanson, L.; Andersson, T.; Nilsson, A. Mercury in fish inSwedish lakes - Linkages to domestic and European sources ofemission. Water, Air, Soil Pollut. 1990, 50, 171-191.
(36) Fjeld, E.; Rognerud, S. Use of path analysis to investigate mercuryaccumulation in brown trout (Salmo trutta) in Norway and theinfluence of environmental factors. Can. J. Fish. Aquat. Sci. 1993,50, 1158-1167.
(37) Evers, D. C.; Kaplan, J. D.; Meyer, M. W.; Reaman, P. S.; Braselton,W. E.; Major, A.; Burgess, N.; Scheuhammer, A. M. Geographictrend in mercury measured in common loon feathers and blood.Environ. Toxicol. Chem. 1998, 17, 173-183.
(38) Hammerschmidt, C. R.; Fitzgerald, W. F. Methylmercury infreshwater fish linked to atmospheric mercury deposition.Environ. Sci. Technol. 2006, 40, 7764-7770.
(39) Hammerschmidt, C. R.; Fitzgerald, W. F. Methylmercury inmosquitoes related to atmospheric mercury deposition andcontamination. Environ. Sci. Technol. 2005, 39, 3034-3039.
(40) Ricker, W. E. Linear regression in fishery research. J. Fish. Res.Board Can. 1973, 30, 409-434.
Received for review December 22, 2006. Revised manuscriptreceived March 28, 2007. Accepted May 1, 2007.
ES063061R
4958 9 ENVIRONMENTAL SCIENCE & TECHNOLOGY / VOL. 41, NO. 14, 2007