exposureto igd receptors igd in vivo in vitro - pnas.org fileabstract igd is a surface...
TRANSCRIPT

Proc. Natl. Acad. Sci. USAVol. 85, pp. 559-563, January 1988Immunology
Exposure to crosslinked IgD induces receptors for IgD on T cellsin vivo and in vitroRICHARD F. CoIco*, FRED FINKELMANt, CHRISTINA D. SWENSONt, GREGORY W. SISKIND§,AND G. JEANETTE THORBECKEt*City University of New York Medical School, New York, NY 10031; tUniformed Services University of the Health Sciences, Bethesda, MD 20814; tNewYork University School of Medicine, New York, NY 10016; and §Cornell University Medical College, New York, NY 10021
Communicated by Michael Heidelberger, September 16, 1987
ABSTRACT IgD is a surface immunoglobulin, which iscoexpressed with IgM on >90% of mature B cells, but itslevels in serum are extremely low compared to those of IgM.Its role as a surface receptor has been reemphasized by ourrecent findings that IgD receptors are induced on helper Tcells by exposure to IgD and that such cells have immunoaug-menting properties. The present study shows that crosslinkingof soluble IgD or of monomeric cell surface IgD is required andsufficient for the induction ofT cells bearing receptors for IgD,both in vivo and in vitro. Effective IgD crosslinking in thisrespect can be obtained with antigen or with heterologous andimmunogenic as well as nonimmunogenic allotype-specificanti-IgD. These results reinforce the concept that the inductionof T cells bearing receptors for IgD is an integral component ofthe normal immune response.
Recent studies in our laboratory have demonstrated theexistence of helper T cells bearing receptors for IgD (T8) (1).This observation grew out of our earlier studies in whichmice that bore the IgD-secreting TEPC-1017 or TEPC-1033myeloma were found to generate significantly increasedantibody responses to various antigens (2). Investigation ofthe cellular basis for this immunoaugmenting effect of IgDrevealed that the elevated serum IgD levels in such micestimulated the appearance of a significantly increased num-ber of T cells that were capable of forming rosettes withIgD-coated sheep erythrocytes (SRBC) (3). Results from celltransfer studies confirmed that T cells exposed to IgDmyeloma protein either in vivo or in vitro have a significantimmunoaugmenting effect on antibody production in recipi-ents (3, 4).
Various methods of inducing TS cells were investigated. Itwas found that IgD receptors can also be induced in vivo byinjection of antigen (1) or lymphokines, including interleukin2 (IL-2) and interferon y (5). Exposure of helper T cells tosuch lymphokines in vitro also causes the expression of IgDreceptors (5). In addition, resting T-cell clones of helperphenotype (L3T4+, Lyt-1', Lyt-2-), grown in the absenceof IL-2, responded with increased IgD-receptor expressionfollowing overnight exposure to IgD or recombinant IL-2 (5).This effect was also seen when T-cell clones were stimulatedwith antigen in the absence of exogenously supplied IgD orIL-2, presumably as a result of autocrine secretion of lym-phokines in response to antigen (5).Our findings that both T-dependent and T-independent
antigens cause a small but significant increase in the fre-quency of TS cells within 5 days of antigen injection suggestthat crosslinking of B-cell surface IgD by physiologicalstimuli induces TS cells. The present studies were carriedout to explore further the role of IgD crosslinking in IgD-receptor induction. The findings show that T cells are
induced to express receptors for IgD following exposure toIgD aggregated by anti-IgD. In addition, taking advantage ofthe availability of an IgD-secreting B-cell hybridoma, Bi-8.81, we show that soluble monomeric IgD is much lesseffective as a TS-inducing agent in its native than in anaggregated form, such as in antigen-IgD immune complexes.Furthermore, the results clearly demonstrate that crosslink-ing of IgD on the surface of B cells is necessary andsufficient for induction of TS cells in vitro. The biologicalsignificance of these findings is discussed.
MATERIALS AND METHODSMice. BALB/c and CB6F1 mice were obtained from
Charles River Breeding Laboratories. B10.A and C57BL/6mice were purchased from The Jackson Laboratory.
Antigens. Keyhole limpet hemocyanin was purchasedfrom Schwartz/Mann; Ficoll 400 (Mr = 40,000) was fromPharmacia; and trinitrobenzenesulfonic acid was fromSigma. Trinitrophenyl (TNP) conjugates were prepared asdescribed by Little and Eisen (6). SRBC and horse erythro-cytes (HRBC) were purchased from Colorado Serum(Denver).Plaque-Forming Cell (PFC) Assay. Anti-TNP and anti-
HRBC splenic PFC were assayed by the slide modificationof the technique of Jerne et al. (7). IgG-producing cells weredeveloped with rabbit anti-mouse immunoglobulin in thecomplement and goat anti-p in the agar.
Cell Lines. The IgD-secreting plasmacytoma TEPC 1017(8) was maintained i.p. in pristane-primed BALB/c mice.The IgD-secreting B-cell hybridoma B1-8.61 was the kindgift of Klaus Rajewsky (Institute for Genetics, University ofCologne, Cologne, F.R.G.) (9). The B1-8.81 cell line secretesIgD that is specific for the hapten 4-hydroxy-3-nitrophenyl-acetyl (NP). The B1-8.51 cells were maintained i.p. in CB6F1mice. The murine IgM', IgD' B-cell lymphoma CH15 wasobtained from G. Haughton (University of North Carolina,Chapel Hill) and was derived from B10.H-2aH4bp/Wts mice(10) and was maintained by i.p. passage in B10.A mice.
Purification of IgD. TEPC-1017-derived IgD and monoclo-nal B1-8.81 IgD were purified by affinity chromatography ona rabbit anti-IgD-coupled Sepharose 4B column (1, 2).
Antibodies. The following antibodies were generated andpurified as previously described: goat anti-mouse IgD, goatanti-mouse IgM (11), Hba/1 (12) (a monoclonal IgG2a de-rived from a mouse of the b allotype, which is specific forIgD of the a allotype), and rabbit anti-mouse immunoglobu-lin prepared by immunization with mouse IgG in completeFreund's adjuvant. The univalent Fab/Fc fragment of Hba/1was prepared by digestion with elastase (Sigma; enzyme-to-
Abbreviations: T8, helper T cells bearing receptors for IgD; SRBC,sheep erythrocytes; HRBC, horse erythrocytes; RFC, rosette-forming cell(s); PFC, plaque-forming cell(s); NP, 4-hydroxy-3-nitrophenylacetyl; BSA, bovine serum albumin; TNP, trinitro-phenyl; IL-2, -4, interleukin 2, 4.
559
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement"in accordance with 18 U.S.C. §1734 solely to indicate this fact.

Proc. Natl. Acad. Sci. USA 85 (1988)
substrate ratio of 1:100) as described (ref. 13 and D. K.Goroff and F.F., unpublished data). The monoclonal rat Ig2aantibody (2.4G2) to mouse B cell and monocyte Fc'. receptor(14) was isolated from ascites fluid by sequential (NH4)2SO4precipitation, DEAE-cellulose ion-exchange chromatogra-phy, and Sephadex G-200 gel filtration (13).
Purification of Splenic T Cells. Splenic T cells were de-pleted of adherent cells by incubation in tissue-culture-gradePetri dishes (1400-1, Nunclon, Roskilde, Denmark) at 37°C.Splenic T cells were then purified by panning adherent-cell-depleted spleen cells on Petri dishes coated with rabbitanti-mouse immunoglobulin (15). This purification methodresults in <1% contamination with immunoglobulin-positiveB cells, as determined by direct staining with fluorescein-conjugated rabbit anti-mouse immunoglobulin.IgD Rosette-Forming Cell (RFC) Assay. Lymphoid cells
were examined for their expression of receptors for IgD by arosette method (1). Briefly, IgD-coated indicator cells wereprepared by coupling purified TEPC-1017 IgD to SRBCusing the CrCl3-coupling method (16). Two-tenths milliliterof 1% IgD-coated SRBC was mixed with 0.1 ml of assay cells(2.5 x 105) at 370C for 15 min. Cells were then centrifuged at200 x g for 3 min and incubated further at 40C for 45 min.Immediately before scoring RFC, the lymphocytes werestained with toluidine blue. Lymphocytes surrounded by >3indicator cells were scored as rosettes, and the results wereexpressed as percentages of RFC. In each experiment, thepercentage of cells forming rosettes with uncoupled SRBCwas also determined.
RESULTS AND DISCUSSIONRequirement for Antigen Crosslinking of B1-8.81 IgD in the
Induction of TS Cells. The results in Table 1 show that, incontrast with TEPC-1017 IgD, NP-specific B1-8.81 IgD usedat similar concentrations does not induce TS cells. This is notdue to allotype differences in the IgD molecules, sinceTEPC-1017 induces IgD receptors on T cells from bothBALB/c and C57BL/6 mice (i.e., from both a and b allotypestrains). Moreover, B1-8.81 IgD is equally inefficient ininducing IgD receptors on cells from both strains. In com-bination with antigen [NP-bovine serum albumin (BSA)],however, B1-8.31 IgD is as effective as TEPC-1017 IgD in
inducing TS cells. Control monoclonal antibodies of thesame specificity as those from B1-8.81 cells but of differentisotype (IgM and IgG1), whether used with or withoutantigen, fail to induce IgD receptors (Table 1, experiments 1and 2). In additional experiments (not shown), it was foundthat goat anti-IgD could also be used to crosslink B1-8.81IgD (in antigen excess) with similar results. By itself, goatanti-IgD did not induce IgD-RFC in purified T cells (2%),whereas in combination with B1-8.81 IgD, 19% IgD-RFCwere obtained.TEPC-1017 IgD has been reported to be secreted as a 82K2
dimer (8), whereas B1-8.81 is probably secreted as a 82A2monomer (9). This may explain why TEPC-1017 IgD, but notB1-8.81 IgD, induces TS cells without any further aggrega-tion. It is not surprising that dimeric or oligomeric forms ofIgD should be needed for the induction of IgD receptors,since a similar requirement has been described for theinduction of Fc receptors for IgE (17) and IgA (18).
Effects of B1-8.81 IgD on Primary and Secondary Immune
Responses. Since TS cells mediate the immunoaugmentingeffects of IgD from TEPC-1017, we speculated that in vivotreatment of mice with B1-8.81 IgD, which fails to induce TOcells in vitro unless it is crosslinked by antigen, might alsofail to cause augmentation of humoral immune responses.The results in Table 2 demonstrate that this is indeed thecase. When BALB/c mice were pretreated with TEPC-1017-derived IgD, subsequently primed with HRBC, and thenchallenged with this antigen 10 days later, significant in-creases in the IgM and IgG anti-HRBC PFC responses wereobserved (Table 2, experiment 1). Similarly, when mice wereimmunized with TNP-keyhole limpet hemocyanin aftertreatment with TEPC-1017 IgD, their primary antibodyresponse to this antigen was greatly enhanced (Table 2,experiment 3). In contrast, mice pretreated with the B1-8.81-derived IgD mounted primary and secondary antibody re-sponses that were not significantly different from those ofsaline-injected control mice (Table 2). This lack of immu-noaugmenting effect was not due to a failure of mice of the aallotype to be affected by IgD ofthe b allotype, because (a x
b)F1 allotype mice were also unaffected by B1-8.81 IgD.Thus, the failure of the native form of B1-8.81 IgD to induceTO cells correlates with the lack of immunoaugmentingeffects of this monoclonal IgD.
Table 1. Induction of TS cells by B1-8.81 IgD, only when crosslinked with antigen% IgD-RFC, mean ± SD
Addition to T cells Exp. 1; BALB/c Exp. 2; C57BL/6 Exp. 3; BALB/cTEPC-1017 ascites (10%6) 30.5 ± 1.4 21.0 ± 1.9 35.0 ± 2.9TEPC-1017 IgD* (100,g/ml) ND ND 40.5 ± 1.5B1-8.81 ascites (10%o) 0 6.0 ± 3.0 NDB1-8.81 IgD* (100 ug/ml) ND ND 1.0 ± 1.0S58.15 IgM* (100 jug/ml) 0 3.9 ± 3.0 NDB6.55.1 IgGl* (100 Ag/ml) 0 3.0 ± 1.7 NDNP-BSA (50-100 ug/ml) 0 0 NDNP-BSA (500 ,ug/ml) ND 1.5 ± 1.5 0B1-8.81t and NP-BSA (50-100 ,ug/ml) 18.5 ± 1.5 15.5 ± 2.4 NDB1-8.61t and NP-BSA (500 jug/ml) ND 25.5 ± 5.5 40.5 ± 0.5S58.15* and NP-BSA (100,ug/ml) 0 0 NDB6.55.1* and NP-BSA (100 ,tg/ml) 0 0 ND
Splenic T cells (2.5 x 106/ml) were incubated with the appropriate additions for 18 hr at 37C in 5%C02, and the percentages of IgD-RFC were determined. The values given represent the percentage ofIgD-RFC after exposure to indicated substances minus the percentage of IgD-RFC after exposure tomedium only. Subtracted background IgD-RFC varied from 3-7%. ND, not determined.*Affinity purified, 100 jg/ml. S58.15 and B6.55.1 are monoclonal antibodies to NP of IgM and IgG1isotypes, respectively.
tAscites (10%1 in Eagle's minimal essential medium) were used in experiments 1 and 2; the finalconcentration of IgD was 300 ,ug/ml.tAscites (10%6) were used in experiment 2; the final concentration of IgD was 300 ,ug/ml. Affinity-purified IgD (100 ,ug/ml) was used in experiment 3.
560 Immunology: Coico et A

Proc. Natl. Acad. Sci. USA 85 (1988) 561
Table 2. Comparison of the effects of IgD from two different sources on the primary andsecondary immune responses
PFC per spleen,*Source of IgD injected geometric mean x SEM
Exp. Strain TEPC-1017 B1-8.61 IgM IgG n
1 BALB/c - - 15,140 x. 1.2 48,970 x. 1.1 3+ - 31,840 x 1. a 127,050 1.2 4- + 16,740 x 1.3C 47,270 x 1.4c 4
2 CB6F1 - - 10,680 x. 1.3 31,840 x. 1.3 5- + 14,800 x. Lc 54,290 x 1.1C 6
3 BALB/c - - 13,730x13 14,070 x 1.2 3+ - 63,540 69,000 x.1e 3- + 15,800 x1.1c 25,510 x1. lC 3
For experiments 1 and 2, 0.5-1 mg of IgD was injected on days - 1 and 0, and the primary HRBC(107 cells) injection was on day 0. The secondary HRBC injection was given on day 10; anti-HRBCPFC was assayed on day 14. For experiment 3, 0.5-1 mg of IgD was injected on days - 8, - 7, - 6,and -5; TNP-keyhole limpet hemocyanin (100 Iug) was injected on day 0; and anti-TNP PFC wereassayed on day 5.*Significance of differences with control values (Student's t test): a, P = 0.013; b, P = 0.011; c, P notsignificant; d, P = 0.02; e, P < 0.005.
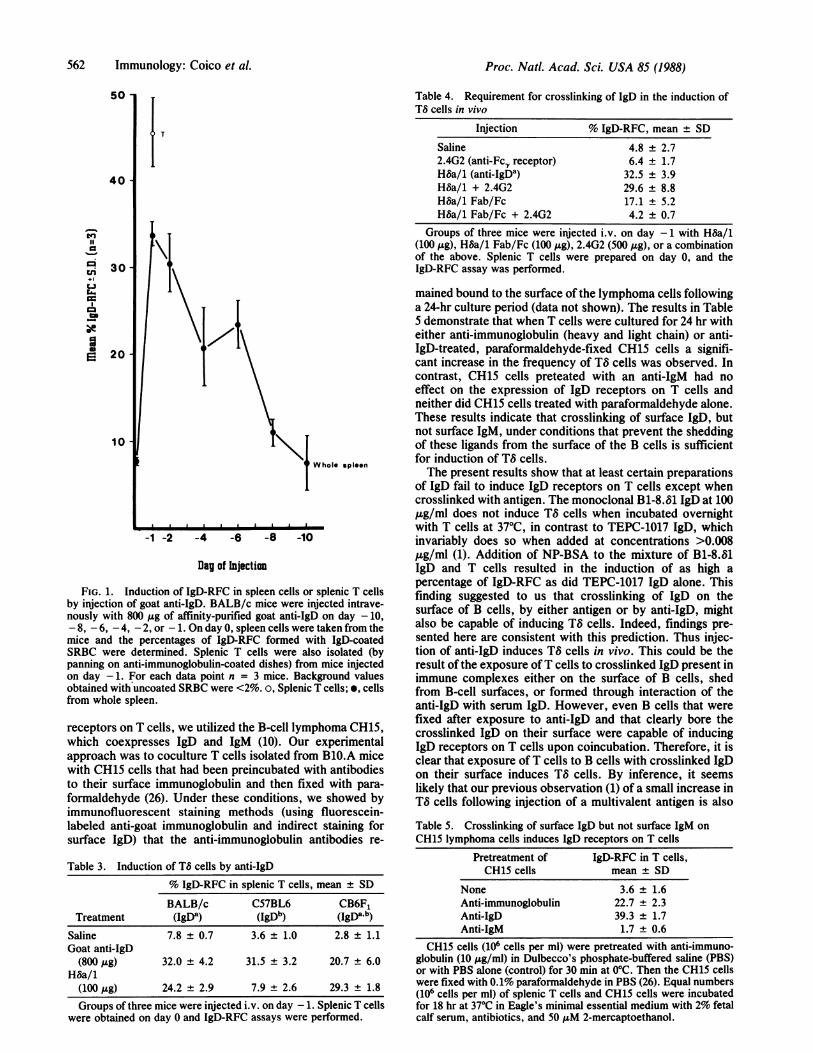
In Vivo Induction of TS Cells by Goat Anti-IgD Antiserum.Our finding that crosslinking of IgD is necessary for induc-tion of TS cells in vitro suggested that a similar stimulusmight also be capable of inducing such cells in vivo. Prelim-inary support for this hypothesis was provided in previouslyreported studies (1), which showed that following injectionwith antigen a small but significant increase in the incidenceof TS cells is observed. In order to confirm if such cross-linking in vivo could also induce T8 cells, we injected micewith affinity-purified goat anti-IgD and followed the kineticsof induction of TS cells in their spleens. The results showthat, 24 hr following injection with goat anti-IgD, there is asignificant increase in the frequency of splenic IgD-RFC(Fig. 1). As expected, cell fractionation studies confirmedthat the splenic T cells were responsible for rosette forma-tion with the IgD-coated indicator cells. It has been reportedthat treatment with goat anti-IgD results in polyclonal acti-vation of the B-cell population (11, 19-21). Analysis of thecellular basis for this effect established that such activationis largely due to B-cell stimulation by the combination ofanti-IgD and lymphokines produced by T cells, which areresponding to the goat immunoglobulin presented to them bythe anti-IgD-coated B cells (22). Under such optimal condi-tions of "antigen" presentation, T cells produce high quan-tities of lymphokines, including interferon y and IL-4 (23).This lymphokine response is first detected three days afterthe injection of the goat anti-IgD (22-24) and is thereforeunlikely to be responsible for the early phase of TS-cellinduction, which is maximal within 24 hr after treatment(Fig. 1). However, since previous studies have establishedthat certain lymphokines such as IL-2, interferon y (5), andIL-4 (25) can also induce TS cells, it is possible that thecontinued presence of increased numbers of TS cells ob-served beyond day 3 is attributable to the stimulatory effectof such factors.
Induction of TS by an Allotype-Specific Monoclonal Anti-IgD Antibody. To distinguish further between IgD crosslink-ing and lymphokine induction as mechanisms of TS induc-tion by anti-IgD antibodies, the effectiveness of polyclonalgoat anti-IgD antibody and a monoclonal antibody specificfor IgD of the a allotype, Hba/1, at inducing TS cells werecompared. Mice of three different strains representing IgDallotypes a (BALB/c), b (C57BL/6), and a x b (CB6F1)were used to examine this question. Goat anti-IgD antibodycrosslinks IgD and induces T-cell help, lymphokine produc-tion, and polyclonal immunoglobulin production in all threestrains used. In contrast, Hba/1 crosslinks IgD in BALB/cand CB6F1 but not in C57BL/6 mice and induces T-cell
activation and polyclonal immunoglobulin production onlyin the BALB/c mice, presumably because T cells in theCB6F1 (a x b allotype hybrid) mice fail to recognize the ballotypic determinants expressed on the Hba/1 antibody asforeign (ref. 19; and F.F., unpublished data). As expected,i.v. injection of goat anti-IgD resulted in the appearance ofincreased numbers of T8 cells in all strains tested (Table 3).However, Hba/1 induced TS cells in both the a allotype and(a x b)F1 allotype strains but not in the homozygous ballotype strain. Thus, IgD crosslinking, rather than inductionof T-cell activation, is sufficient for TS induction by anti-IgDantibody.
Requirements for Crosslinking of IgD in the Induction of TbCells in Vivo. Having established that crosslinking of IgDwith Hba/1 is capable of inducing TS cells in vivo, we thenasked whether a univalent anti-IgD antibody would fail toinduce TS cells. To address the question, we investigatedwhether a monovalent Fab/Fc fragment of HSa/1, whichconsists of a single Fab fragment linked to the Fc fragmentby the hinge region (13), could induce TS in the presence orabsence of 2.4G2, an antibody that blocks the B-cell andmacrophage IgG2b Fc receptor. This approach was takenbecause H~a/1 Fab/Fc has recently been shown to becapable of inducing B cells to increase their surface Iaantigen expression by crosslinking their surface IgD throughan Fcy receptor (FcyR)-dependent mechanism. Antibody2.4G2, by blocking the IgG2b FcR, blocks Hba/1 Fab/Fc-induced B-cell surface IgD crosslinking and activation (13).The results in Table 4 show that whereas the Hba/1 Fab/Fcwas active in inducing TS cells when injected by itself intoBALB/c mice, simultaneous injection of Hba/1 Fab/Fc and2.4G2 failed to induce TS cells. The 2.4G2 antibody, whichdoes not prevent IgD crosslinking or B-cell activation byintact Hba/1, also had no effect on the ability of the intactH~a/1 antibody to induce TS cells in vivo. We conclude thatinduction of IgD receptors using the Fab/Fc fragment ofH~a/1 is mediated by binding of the Fc portion of thisunivalent antibody to FcR' cells, which either crosslinksthe Hba/1 Fab/Fc-bound B-cell surface IgD or mimics theeffect of crosslinking IgD on the B cell surface.
Crosslinking of Surface IgD but not Surface IgM Induces TbCells. Although these studies suggested that direct crosslink-ing of B-cell surface IgD could induce TS cells, we could notexclude the possibility that soluble IgD-anti-IgD immunecomplexes generated in the serum or released from thesurface of the B cells might actually mediate the induction ofTS cells. To determine whether crosslinking of IgD on thesurface of the B cell is sufficient for optimal induction of IgD
Immunology: Coico et A
Proc. Natl. Acad. Sci. USA 85 (1988)
50
40
0-%
ci
Ira
-+l.
I
30
20
10
Whole spleen
-1 -2 -4 -6 -8 -10
fag of Injectian
FIG. 1. Induction of IgD-RFC in spleen cells or splenic T cellsby injection of goat anti-IgD. BALB/c mice were injected intrave-nously with 800 ,g of affinity-purified goat anti-IgD on day -10,- 8, - 6, - 4, - 2, or - 1. On day 0, spleen cells were taken from themice and the percentages of IgD-RFC formed with IgD-coatedSRBC were determined. Splenic T cells were also isolated (bypanning on anti-immunoglobulin-coated dishes) from mice injectedon day -1. For each data point n = 3 mice. Background valuesobtained with uncoated SRBC were <2%. o, Splenic T cells; *, cellsfrom whole spleen.
receptors on T cells, we utilized the B-cell lymphoma CH15,which coexpresses IgD and IgM (10). Our experimentalapproach was to coculture T cells isolated from BMO.A micewith CH15 cells that had been preincubated with antibodiesto their surface immunoglobulin and then fixed with para-formaldehyde (26). Under these conditions, we showed byimmunofluorescent staining methods (using fluorescein-labeled anti-goat immunoglobulin and indirect staining forsurface IgD) that the anti-immunoglobulin antibodies re-
Table 3. Induction of TS cells by anti-IgD% IgD-RFC in splenic T cells, mean SD
BALB/c C57BL6 CB6F1Treatment (IgDa) (IgDb) (IgDaab)
Saline 7.8 0.7 3.6 1.0 2.8 + 1.1Goat anti-IgD
(800,g) 32.0 4.2 31.5 3.2 20.7 6.0H~a/1
(100,g) 24.2 2.9 7.9 2.6 29.3 1.8
Groups of three mice were injected i.v. on day 1. Splenic T cells
were obtained on day 0 and IgD-RFC assays were performed.
Table 4. Requirement for crosslinking of IgD in the induction ofTS cells in vivo
Injection % IgD-RFC, mean + SD
Saline 4.8 ± 2.72.4G2 (anti-Fcr receptor) 6.4 ± 1.7H~a/1 (anti-IgDa) 32.5 ± 3.9H~a/1 + 2.4G2 29.6 ± 8.8HOa/1 Fab/Fc 17.1 ± 5.2H~a/1 Fab/Fc + 2.4G2 4.2 ± 0.7
Groups of three mice were injected i.v. on day -1 with Hba/1(100 jLg), H8a/1 Fab/Fc (100 ,.g), 2.4G2 (500 pug), or a combinationof the above. Splenic T cells were prepared on day 0, and theIgD-RFC assay was performed.
mained bound to the surface of the lymphoma cells followinga 24-hr culture period (data not shown). The results in Table5 demonstrate that when T cells were cultured for 24 hr witheither anti-immunoglobulin (heavy and light chain) or anti-IgD-treated, paraformaldehyde-fixed CH15 cells a signifi-cant increase in the frequency of TS cells was observed. Incontrast, CH15 cells preteated with an anti-IgM had noeffect on the expression of IgD receptors on T cells andneither did CH15 cells treated with paraformaldehyde alone.These results indicate that crosslinking of surface IgD, butnot surface IgM, under conditions that prevent the sheddingof these ligands from the surface of the B cells is sufficientfor induction of TS cells.The present results show that at least certain preparations
of IgD fail to induce IgD receptors on T cells except whencrosslinked with antigen. The monoclonal B1-8.81 IgD at 100,ug/ml does not induce TS cells when incubated overnightwith T cells at 37°C, in contrast to TEPC-1017 IgD, whichinvariably does so when added at concentrations >0.008pgg/ml (1). Addition of NP-BSA to the mixture of B1-8.O1IgD and T cells resulted in the induction of as high apercentage of IgD-RFC as did TEPC-1017 IgD alone. Thisfinding suggested to us that crosslinking of IgD on thesurface of B cells, by either antigen or by anti-IgD, mightalso be capable of inducing TO cells. Indeed, findings pre-sented here are consistent with this prediction. Thus injec-tion of anti-IgD induces TO cells in vivo. This could be theresult of the exposure ofT cells to crosslinked IgD present inimmune complexes either on the surface of B cells, shedfrom B-cell surfaces, or formed through interaction of theanti-IgD with serum IgD. However, even B cells that werefixed after exposure to anti-IgD and that clearly bore thecrosslinked IgD on their surface were capable of inducingIgD receptors on T cells upon coincubation. Therefore, it isclear that exposure ofT cells to B cells with crosslinked IgDon their surface induces TO cells. By inference, it seemslikely that our previous observation (1) of a small increase inTO cells following injection of a multivalent antigen is also
Table 5. Crosslinking of surface IgD but not surface IgM onCH15 lymphoma cells induces IgD receptors on T cells
Pretreatment of IgD-RFC in T cells,CH15 cells mean + SD
None 3.6 ± 1.6Anti-immunoglobulin 22.7 + 2.3Anti-IgD 39.3 + 1.7Anti-IgM 1.7 ± 0.6
CH15 cells (106 cells per ml) were pretreated with anti-immuno-globulin (10 ,ug/ml) in Dulbecco's phosphate-buffered saline (PBS)or with PBS alone (control) for 30 min at 0°C. Then the CH15 cellswere fixed with 0.1% paraformaldehyde in PBS (26). Equal numbers(106 cells per ml) of splenic T cells and CH15 cells were incubatedfor 18 hr at 37°C in Eagle's minimal essential medium with 2% fetalcalf serum, antibiotics, and 50 ,uM 2-mercaptoethanol.
562 Immunology: Coico et al.

Proc. Natl. Acad. Sci. USA 85 (1988) 563
due to exposure of T cells to antigen-crosslinked IgD on Bcells. In other words, the mechanism by which TS cells comeinto play in the normal immune response is through theirinduction by antigen-crosslinked IgD on the surface of Bcells. This is probably a locally occurring phenomenon thathappens, early after the injection of antigen, in the surround-ings of B cells that have specificity for that antigen. Thus,within a local microenvironment, nearby helper T cellsacquire receptors for IgD, which facilitate their interactionwith IgD', antigen-bound B cells. Lymphokines, whichreadily promote the expression of these IgD-specific recep-tors (5), would help to maintain the presence of TS cellsduring the emerging immune response. It is well establishedthat following activation of IgD' B cells with antigen IgDtranslation is rapidly down-regulated (27, 28). Such down-regulation results in the disappearance of IgD from the B-cellsurface within a few days of activation with antigen (28).Indeed, these findings are consistent with the concept thatIgD is primarily a receptor molecule, which, unlike otherimmunoglobulins, is not secreted following triggering of Bcells expressing the 8 heavy chain.The relative role of induction of TS cells in the stimulation
of polyclonal immunoglobulin production, which occursafter injection of anti-IgD antibody, appears to be minor ascompared to the combination of (i) the direct activatingeffect of anti-IgD antibody on B cells and (it) the presenta-tion to T cells of the allo- or xenogeneic determinants onanti-IgD antibodies that are bound to and processed by Bcells and that stimulate T-cell activation and lymphokineproduction (20, 23, 24). This is suggested by the comparisonof our present results on the induction of TS cells with theresults of Finkelman et al. (20). In all cases where thepolyclonal activation of IgG production is induced by injec-tion of anti-IgD, TS cells are also induced. However, inexperiments in which allotype-specific anti-IgD is injectedinto (a x b)F1 hybrid mice, TS cells can be induced withoutthe marked activation of polyclonal immunoglobulin produc-tion seen when the allotypic anti-IgD is antigenic for themice into which it is injected (20). Thus, whereas enhance-ment of antibody production to a totally unrelated antigen isproduced by simultaneous injection or pretreatment withsyngeneic IgD (2), injection of anti-IgD primarily enhancesthe immune response to haptens bound to the anti-IgD or toepitopes of the anti-IgD itself if the anti-IgD antibody isforeign and thus itself immunogenic (20, 29). In view of theTS-mediated immunoaugmenting effect ofTEPC-1017 IgD, itis not immediately clear why the response to simultaneouslyinjected unrelated antigens is not always augmented by TScells induced by allotypic or goat anti-IgD. It seems possiblethat the large excess of anti-IgD used in these experimentscauses such a rapid down-regulation of surface IgD expres-sion that the augmenting effect of TS cells on antibodyproduction is prevented by the absence of IgD-antigencomplexes. However, a less impressive but definite effect onantibody formation to unrelated antigens has been obtainedin (a x b)F1 hybrids with nonimmunogenic allotypic anti-IgD (30). In the absence of additional lymphokine injections,this effect is limited to the allotype not affected by theanti-IgD (30) and may be a consequence of TS-cell induction.Although TS cells are clearly not allotype specific (1), theyapparently augment the responses of those B cells thatexpress surface IgD interacting with antigen more than theresponse by B cells that have had their surface IgD down-modulated by the anti-IgD.
The technical assistance of Marilyn Shapiro and Ariel Soriano isgreatly appreciated. This work was supported by Public HealthService Grants AI-22645, AG-04860, AI-11694, and AI-21328, andby the Uniformed Services University of the Health SciencesResearch Protocol No. R08308.
1. Coico, R. F., Xue, B., Wallace, D., Pernis, B., Siskind, G. W.& Thorbecke, G. J. (1985) Nature (London) 316, 744-746.
2. Xue, B., Coico, R. F., Wallace, D., Siskind, G. W., Pernis, B.& Thorbecke, G. J. (1984) J. Exp. Med. 159, 103-113.
3. Coico, R. F., Xue, B., Wallace, D., Siskind, G. W. & Thor-becke, G. J. (1985) J. Exp. Med. 162, 1852-1861.
4. Coico, R. F., Wallace, D., Xue, B., Rosen, J., Pernis, B.,Thorbecke, G. J. & Siskind, G. W. (1985) in Microenviron-ments in the Lymphoid System, ed. Klaus, G. G. B. (Plenum,New York), pp. 729-738.
5. Coico, R. F., Berzofsky, J. A., York-Jolley, J., Ozaki, S.,Siskind, G. W. & Thorbecke, G. J. (1987) J. Immunol. 138,4-6.
6. Little, J. R. & Eisen, H. N. (1967) Methods Immunol. Immu-nochem. 1, 128-132.
7. Jerne, N. K., Nordin, A. A. & Henry, C. (1983) in Cell BoundAntibody, eds. Amos, B. & Koprowsky, H. (Wistar InstitutePress, Philadelphia), pp. 109-111.
8. Finkelman, F. D., Kessler, S. W., Muchinski, J. F. & Potter,M. (1981) J. Immunol. 126, 680-687.
9. Neuberger, M. S. & Rajewsky, K. (1981) Proc. Natl. Acad.Sci. USA 78, 1138-1142.
10. Pennell, C. A., Arnold, L. W., Lutz, P. M., LoCascio, N. J.,Willoughby, P. B. & Haughton, G. (1985) Proc. Natl. Acad.Sci. USA 82, 3799-3803.
11. Finkelman, F. D., Scher, I., Mond, J. J., Kung, J. I. & Met-calf, E. S. (1982) J. Immunol. 129, 629-637.
12. Zitron, I. M. & Clevinger, B. L. (1980) J. Exp. Med. 152,1135-1146.
13. Parham, P. (1983) J. Immunol. 131, 2895-2902.14. Unkeless, J. C. (1979) J. Exp. Med. 150, 580-596.15. Wysocki, L. J. & Sato, V. L. (1978) Proc. Natl. Acad. Sci.
USA 75, 2844-2848.16. Poston, R. N. (1974) J. Immunol. Methods 5, 91-96.17. Huff, T. F. & Ishizaka, K. (1984) Proc. Nat!. Acad. Sci. USA
81, 1514-1518.18. Yodoi, J., Adachi, M. & Masuda, T. (1982) J. Immunol. 128,
888-894.19. Finkelman, F. D., Smith, J., Villacreses, N. & Metcalf, E. S.
(1985) Eur. J. Immunol. 15, 315-320.20. Finkelman, F. D., Scher, I., Mond, J. J., Kessler, S., Kung,
J. T. & Metcalf, E. S. (1982) J. Immunol. 129, 638-646.21. Finkelman, F. D., Muul, L. M., Yaffe, L., Scher, I., Mond,
J. J., Kessler, S. W., Ryan, I., Kung, J. T. & Metcalf, E. S.(1982) Ann. N. Y. Acad. Sci. 399, 316-328.
22. Finkelman, F. D., Smith, J., Villacreses, N. & Metcalf, E. S.(1984) J. Immunol. 133, 550-555.
23. Finkelman, F. D., Ohara, J., Goroff, D. K., Smith, J., Villa-creses, N., Mond, J. J. & Paul, W. E. (1986) J. Immunol. 137,2878-2885.
24. Finkelman, F. D., Katona, I. M., Mosmann, T. F. & Coff-man, R. L. (1988) J. Immunol., in press.
25. Coico, R. F., Swenson, C. D., Siskind, G. W. & Thorbecke,G. J. (1988) in 9th International Conference on LymphaticTissues and Germinal Centres in Immune Reactions, eds.Fossum, S. & Rolstad, B. (Plenum, New York), in press.
26. Allen, P. M. & Unanue, E. R. (1984) J. Immunol. 132,1077-1079.
27. Yuan, D. (1984) J. Immunol. 132, 1566-1570.28. Kanowith-Klein, S., Vitetta, E. S., Korn, E. L. & Ashman,
R. F. (1979) J. Immunol. 122, 2349-2355.29. Finkelman, F. D., Mond, J. J. & Metcalf, E. S. (1986) in
B-Lymphocyte Differentiation, ed. Cambier, J. (CRC, BocaRaton, FL), pp. 41-71.
30. Jacobson, E. B., Xue, B., O'Neil, M.-A. J., Siskind, G. W.,Pernis, B. & Thorbecke, G. J. (1985) J. Immunol. 135,2293-2298.
Immunology: Coico et al.