expression of a recombinant lentinula edodes
TRANSCRIPT

Accepted Manuscript
Expression of a recombinant Lentinula edodes cellobiohydrolaseby Pichia pastoris and its effects on in vitro ruminal fermentationof agricultural straws
Lizhi Li, Mingren Qu, Chanjuan Liu, Ke Pan, Lanjiao Xu, KehuiOuYang, Xiaozhen Song, Yanjiao Li, Xianghui Zhao
PII: S0141-8130(19)30672-5DOI: https://doi.org/10.1016/j.ijbiomac.2019.05.043Reference: BIOMAC 12328
To appear in: International Journal of Biological Macromolecules
Received date: 27 January 2019Revised date: 4 April 2019Accepted date: 6 May 2019
Please cite this article as: L. Li, M. Qu, C. Liu, et al., Expression of a recombinantLentinula edodes cellobiohydrolase by Pichia pastoris and its effects on in vitro ruminalfermentation of agricultural straws, International Journal of Biological Macromolecules,https://doi.org/10.1016/j.ijbiomac.2019.05.043
This is a PDF file of an unedited manuscript that has been accepted for publication. Asa service to our customers we are providing this early version of the manuscript. Themanuscript will undergo copyediting, typesetting, and review of the resulting proof beforeit is published in its final form. Please note that during the production process errors maybe discovered which could affect the content, and all legal disclaimers that apply to thejournal pertain.

ACC
EPTE
D M
ANU
SCR
IPT
1
Expression of a recombinant Lentinula edodes cellobiohydrolase by Pichia
pastoris and its effects on in vitro ruminal fermentation of agricultural straws
Lizhi Li§, Mingren Qu§, Chanjuan Liu, Ke Pan, Lanjiao Xu, Kehui OuYang, Xiaozhen
Song, Yanjiao Li, Xianghui Zhao
Jiangxi Province Key Laboratory of Animal Nutrition/Engineering Research Center of
Feed Development, Jiangxi Agricultural University, Nanchang Jiangxi 330045, China
Corresponding author at: Jiangxi Province Key Laboratory of Animal Nutrition/Engineering Research Center
of Feed Development, Jiangxi Agricultural University, Nanchang Jiangxi, China.
E-mail: [email protected].
§These authors contributed equally to this work.
ACCEPTED MANUSCRIPT

ACC
EPTE
D M
ANU
SCR
IPT
2
Abstract
An experiment was conducted to determine the effects of recombinant
cellobiohydrolase on the hydrolysis and in vitro rumen microbial fermentation of
agricultural straws including rice straw, wheat straw, and corn straw. The
cellobiohydrolase from Lentinula edodes (LeCel7A) was produced in Pichia pastoris.
The optimal temperature and pH for LeCel7A were 60°C and 5.0, respectively. The
recombinant protein enhanced the hydrolysis of three straws. During in vitro rumen
fermentation of three straws, the fiber digestibility, concentration of acetate and total
volatile fatty acids, and fermentation liquid microbial protein were increased by
LeCel7A. High throughput sequencing and real-time PCR data showed that the effects
of LeCel7A on ruminal microbial community depended on the fermentation substrates.
The relative abundances of Prevotellaceae_UCG_003 and Saccharofermentans were
increased by LeCel7A regardless of agricultural straws. With rice straw, LeCel7A
increased the relative abundances of Desulfovibrio, Ruminococcaceae and its some
genus. With wheat straw, LeCel7A increased the relative abundances of
Succiniclasticum, Ruminococcus flavefaciens, and Ruminococcus albus. With corn
straw, Succiniclasticum, Christensenellaceae_R_7_group and Desulfovibrio were
increased by LeCel7A. This study demonstrates that LeCel7A could enhance the
hydrolysis and in vitro ruminal fermentation of agricultural straws, showing the
potential of LeCel7A for improving the utilization of agricultural straws in ruminants.
Key words: Recombinant Lentinula edodes cellobiohydrolase; agricultural straw; in
vitro ruminal fermentation
ACCEPTED MANUSCRIPT

ACC
EPTE
D M
ANU
SCR
IPT
3
1. Introduction
Agricultural by-products, for instance rice straw, wheat straw, and maize straw,
are generated in billions of tons per year all around the world [1]. These fibrous
materials contain considerable quantities of cellulose and hemicellulose and have the
potential to be a valuable feed source for ruminants. However, though these straws
could be utilized by ruminants through the ruminal microorganisms [2], their nutritive
feeding value is limited by their high-order molecular packing of lignocelluloses [3].
Consequently, the majority of these agricultural straws are as wastes either left in the
field for natural decay or burnt adding to environmental pollution [4]. In past years,
some physical and chemical technologies such as steaming, alkaline, and acidic
treatments have been investigated to improve the nutritional value of agricultural
straws [5, 6]. However, the application of these methods presents many disadvantages,
including high energy consumption [7] and high risk to the animal and environment,
especially when alkali used [8]. Biological treatments, including the use of white rot
fungi and their enzyme, have the potential to eliminate/reduce the problems associated
with physicochemical methods and appear to be the most promising in improvement
of straws digestibility [4, 9].
As one of white-rot fungi, Lentinula edodes, commonly referred to as the
Shiitake mushroom, produces all the core enzymes essential to the complete
enzymatic hydrolysis of lignocellulose [10]. L. edodes can use its enzymatic
machineries to break down lignocellulose and improve nutritive value of low quality
feeds, such as rape straw, wheat straw, rice straw, corn stover, and sugarcane bagasse,
ACCEPTED MANUSCRIPT

ACC
EPTE
D M
ANU
SCR
IPT
4
but also resulted in the great losses of cellulose and hemicellulose during the
degradation, which limits its practical use [4, 11, 12]. In addition, considering that the
long incubation time of L. edodes and its cultivation products may contain toxic
substances [6], crude enzyme extracts of L. edodes as feed additive may be also not an
attractive option for treatment of straws. However, at least it means that L. edodes is a
good source for fibrolytic enzymes gene pool. Using heterologous expression
technology to obtain the enormous amount of interest enzyme protein within a short
time may be an easy way worthy of consideration.
From the perspective of the best-studied enzyme systems, the heart of
depolymerization of cellulose to glucose comprises cellobiohydrolases,
endoglucanases and β-glucosidase [13]. Cellobiohydrolases (also called
exoglucanases) hydrolyze the crystalline parts of the substrate by initiating their
action from the reducing or non-reducing ends of the cellulose chains, producing
primarily cellobiose and decreasing the substrate polymerization degree very slowly
[14, 15]. Cellobiohydrolases are key components in the multi-enzyme cellulose
complexes. Many studies have been conducted to investigate the functional
characterization of cellobiohydrolases and its potential synergistic role in the
enzymatic hydrolysis of biomass. Some results showed that cellobiohydrolases from
Trichoderma reesei displayed a high synergistic effect with cellulase and xylanase and
enhanced the enzymatic hydrolysis of corn stover, rice straw, and wheat straw [16, 17].
Within the rumen of ruminants, a complex group of anaerobic microorganisms,
including bacteria, archaea and eukaryotes, produces a vast array of lingo-cellulolytic
ACCEPTED MANUSCRIPT

ACC
EPTE
D M
ANU
SCR
IPT
5
enzymes, which means that the rumen can provide an enzyme-containing
environment for the synergistic action and boosting activity of cellobiohydrolases.
Lee et al. (2001) cloned and characterized two cellobiohydrolases genes from L.
edodes, one of which belonged to glycosyl hydrolase family 7 (LeCel7A) [10]. Based
on the effects of L. edodes on agricultural straws degradation aforementioned and
characteristics of rumen, we hypothesize that supplementation of LeCel7A in the
rumen could enhance the hydrolysis and ruminal fermentation of agricultural straws,
however, little information is available. Therefore, this study recombined, expressed
and purified the LeCel7A by P. pastoris, investigated the effects of recombinant
LeCel7A on the hydrolysis of agricultural straws, and evaluated the effects of
recombinant LeCel7A on in vitro ruminal fermentation and microbial community of
agricultural straws and consequently the application possibility of LeCel7A in the
utilization of agricultural straws by ruminants.
2. Materials and Methods
This study was approved by the Animal Care and Use Committee of the College of
Animal Science and Technology of the Jiangxi Agricultural University.
2.1 Strains and reagents
P. pastoris strain X-33, expression vector pPICZαA, and antibiotic zeocin were
purchased from Invitrogen Corporation (Invitrogen, Carlsbad, CA). The competent
Escherichia coli DH5α cells and TIANprep Mini Plasmid Kit were purchased from
Tiangen Biotech (BEIJING) Co., Ltd (Beijing, China). Primers synthesis and DNA
sequencing were performed in Sangon Biotech (Shanghai) Co., Ltd (Shanghai, China).
ACCEPTED MANUSCRIPT

ACC
EPTE
D M
ANU
SCR
IPT
6
The N-glycosidase F (PNGase F), O-glycosidase, and α2-3,6,8,9 neuraminidase were
purchased from NEB Biolabs (Beijing, China). Other reagents were obtained from
standard commercial sources.
2.2 Synthesis of LeCel7A gene and construction of expression vector
The coding sequence of LeCel7A (GenBank accession AF411250), was
synthesized by GenScript Nanjing Co., Ltd. (Nanjing, China) with the following
modifications: Gene sequence was optimized for the preferred codon usage in P.
pastoris without altering the encoded amino acid sequence; the signal sequences
containing 18 codons and the stop codon were removed from the synthesized gene
sequence; and the unique restriction sites including EcoR I and Xba I were used for
plasmid construction, meanwhile, two base pairs (TT/AA) were introduced in the
protein C-terminal domains before the Xba I site to avoiding a translation frame shift
when the vector pPICZαA was used in this study.
The synthetic DNA was ligated into pPICZαA at the EcoR I and Xba I restriction
sites. The resulting plasmid was designated pPICZαA-LeCel7A and transformed into
E. coli DH5α. The pPICZαA-LeCel7A plasmids were extracted from transformed E.
coli and verified by PCR amplification using the α-Factor sequencing primer
(5´-TACTATTGCCAGCATTGCTGC-3´) and 3´ AOX1 sequencing primer
(5´-GCAAATGGCATTCTGACATCC-3´), restriction analysis and sequencing.
2.3. Transformation of P. pastoris and screening of LeCel7A expression stain
Recombinant plasmid pPICZαA-LeCel7A was linearized by restriction digestion
with Sac I and transformed into competent P. pastoris X33 by electroporation
ACCEPTED MANUSCRIPT

ACC
EPTE
D M
ANU
SCR
IPT
7
(MicroPulser; Bio-Rad, CA, USA). The transformants were preliminarily screened on
the YPD agar plates containing zeocin and incubated at 30oC for three days. The
positive colonies were certified for integration of LeCel7A gene into the Pichia
genome by PCR using the primers of α-Factor and 3′AOX1 primers. For their enzyme
activities, the confirmed transformants were further grown in 2 ml of BMGY medium
and shaken overnight at 30°C. The cultures were centrifuged for 5 min at 2,000 rpm.
The pellets were then inoculated into 10 ml of BMMY medium in 100-ml flasks and
shaken at 30°C to induce expression. During the following 1-3 days, methanol was
supplemented every 24 h to the culture to a final concentration of 1.0% (v/v) for
maintaining the induction. On the final day, yeast cultures were collected and
centrifuged at 10,000 rpm for 5 min at 4°C. Supernatants were assayed for the
LeCel7A activity using Avicel (Avicel PH-101, catalog no. 11365; Sigma-Aldrich) as
substrates. The reaction mixture contained supernatants, 1% (w/v) Avicel, and 0.1 M
citrate/disodium hydrogen phosphate buffer. The assay was carried out at 40 °C for 1
h with gentle mixing by measuring the released reducing sugars. Supernatant with the
highest activity was further confirmed by sodium dodecyl sulphate polyacrylamide gel
electrophoresis (SDS-PAGE) staining with Coomassie Blue. The recombinant strain
with the highest activity was selected and kept for further incubation (Data are
included in the Supplemental Material).
2.4. Laboratory scale production of LeCel7A by P. pastoris
According to the Pichia fermentation process guidelines of Invitrogen, a seed
culture of recombinant LeCel7A strain screened was prepared in BMGY medium and
ACCEPTED MANUSCRIPT

ACC
EPTE
D M
ANU
SCR
IPT
8
then inoculated into the batch medium. The production of LeCel7A was carried out in
a 4.0-liter working volume vertical glass bioreactor (Minifors 2, Infors HT,
Switzerland). Culture temperature was controlled at 30°C, pH was controlled at 6.0
with addition of 25% ammonium hydroxide, and the dissolved-oxygen concentration
was maintained above 20% saturation by controlling the stirrer speed or
supplementing pure oxygen, whereas the airflow was kept constant at 4 liters h-1. The
following glycerol fed-batch phase and methanol fed-batch phase were performed
according to the Pichia fermentation process guidelines of Invitrogen. Sample was
taken every 12 hours during the methanol fed-batch phase and the activity was
determined. When the activity no longer increased, the culture was finished.
The yeast culture was collected and centrifuged at 10,000 rpm for 5 min at 4°C.
Supernatants was concentrated and buffer exchanged in a binding buffer (pH 8.0)
consisting of 0.05 M sodium dihydrogen phosphate and 0.3 M sodium chloride using
a membrane separation system (Jinan Bona Biological Technology Co., Ltd., Jinan,
China) and loaded on a 5 ml Bio-Scale Mini Nuvia IMAC Ni-Charged column
(Bio-Rad, CA, USA) connected to a low-pressure chromatographic system (Biologic
LP; Bio-Rad, CA, USA) at 1.0 ml/min. The bound LeCel7A was first washed with the
aforementioned binding buffer containing 0.04 M imidazole (pH 8.0) and then eluted
with the binding buffer containing 0.25 M imidazole (pH 8.0). The flow-through,
which contained LeCel7A, was collected and changed to citrate/disodium hydrogen
phosphate buffer through Amicon Ultra-15 centrifugal filter unit. The protein sample
was verified by Western blot and quantified with Bradford Protein Assay Kit (Sangon
ACCEPTED MANUSCRIPT

ACC
EPTE
D M
ANU
SCR
IPT
9
Biotech, Shanghai, China). SDS-PAGE was carried out to confirm the LeCel7A
protein purity. The N-glycosidase F, O-glycosidase, and α2-3,6,8,9 neuraminidase
were used to investigate the glycosylation of purified LeCel7A according to the
manufacturer's instructions.
2.5 Characteristics of recombinant LeCel7A
The activity of recombinant LeCel7A was measured according to the
aforementioned description. Determining the effects of pH on recombinant LeCel7A
activity was performed by the reaction mixtures containing purified enzyme, 1% (w/v)
Avicel, and 100 mM citrate/disodium hydrogen phosphate buffer (pH 3.0−7.0) at
40°C. The temperature dependency for recombinant LeCel7A activity was determined
by incubating reaction mixtures at 20−80 °C at the optimum pH.
2.6 Enzymatic hydrolysis of agricultural straws
Three agricultural straws including rice straw, wheat straw, and corn straw
(air-dry basis) were used in this study. The hydrolysis was performed using 8 µg
purified recombinant LeCel7A and 20 mg straw substrates in 2 ml 0.1 M
citrate/disodium hydrogen phosphate buffer (pH 5.0). The mixtures were incubated in
10 mL centrifuge tubes and shaken (200 rpm) for 24 h at 60°C as triplicates. The
blanks without recombinant LeCel7A were incubated similarly. The reducing sugars
released in samples were determined using alkaline 3,5-dinitrosalicylic acid reagent.
2.7 In vitro ruminal fermentation
An in vitro study was carried out to investigate the effects of LeCel7A on ruminal
fermentation. Rumen fluid was obtained from three ruminally fistulated beef cattle fed
ACCEPTED MANUSCRIPT

ACC
EPTE
D M
ANU
SCR
IPT
10
a diet consisting of 700 g/kg rice straw and 300 g/kg concentrates before morning
feeding. The rumen liquid collected was filtered through four lays of cheesecloth and
mixed (1:2 v/v) with anaerobic buffer [18]. All manipulations were done under
continuous flushing with CO2. Fermentation was conducted in 120-mL serum bottles
to which 500 mg of agricultural straws and 60 mL of buffered rumen fluid were added.
Two hundred microgram purified LeCel7A was added into the incubation bottles as
LeCel7A treatments, and the same amount of citrate/disodium hydrogen phosphate
buffer instead of LeCel7A were incubated similarly as controls. Bottles were closed
and incubated in a shaking water bath at 39°C for 48 h. All samples were incubated in
triplicate. A blank (rumen fluid without substrate) was incubated in duplicate for
correction of residual dry matter (DM) in samples. The gas produced during
fermentation was expelled per 12 h. Fermentation was terminated by placing the
bottles on the ice, and the residue was filtered using pre-weighted nylon bag (37 µm
pore size) for the determination of in vitro NDF digestibility (IVNDFD). Samples of
filtrate were determined the ruminal pH immediately. One milliliter of ruminal fluid
was preserved by adding 1 ml of deproteinizing solution (100 g/L metaphosphoric
acid and 0.6 g/L crotonic acid) to determine volatile fatty acids (VFA). Ten milliliter
of filtrate was preserved to determine ammonia-N concentration and microbial protein
synthesis. The filtrate samples were also collected and stored at –80°C for DNA
extraction and microbial community determination.
2.6. Analytical procedures
The samples were analyzed for DM by drying at 65°C for 72 h. The NDF content
ACCEPTED MANUSCRIPT

ACC
EPTE
D M
ANU
SCR
IPT
11
in samples was analyzed according to Van Soest et al. (1991) [19]. Ammonia-N in the
samples was analysed according to Weatherburn (1967) [20]. The VFA concentrations
in the filtered samples were determined by a gas chromatography (GC-2014;
Shimadzu Corp., Kyoto, Japan) equipped with a flame ionization detector, fitted with
a 30 m × 0.32 mm I.D. × 0.25 μm thickness film Stabilwax-DA column (Restek Corp.,
Bellefonte, PA, United States). Crotonic acid was used as an internal standard.
Fermentation liquid microbial protein (FLMCP) synthesis was determined according
to Makkar et al. (1982) [21].
The ruminal microbiota was determined by the high-throughput sequencing at Gene
Denovo Co. (Guangzhou, China) using an Illumina Hiseq2500 platform. The raw
Fastq data obtained was processed and analyzed according to our previous report [22].
Based on the operational taxonomic units (OTUs), bacterial richness indices and
bacterial diversity indices were determined. Principal coordinate analysis (PCoA) was
applied on the Bray-Curtis distances to generate two-dimensional plots. Linear
discriminant analysis (LDA) effect size (LEfSe) method was used to identify the most
differentially abundant taxons between groups and LDA ≥ 2.5 was chosen to indicate
significant difference during the analysis of high-throughput sequencing data [23].
The 16S ribosomal deoxyribonucleic acid (rDNA) copy numbers of total bacteria and
three fibrolytic bacterial species (Fibrobacter succinogenes, Ruminococcus
flavefaciens, and Ruminococcus albus) were determined according to our previous
report [22].
2.7 Statistical Analyses
ACCEPTED MANUSCRIPT

ACC
EPTE
D M
ANU
SCR
IPT
12
Statistical analysis was performed using a One-way ANOVA in IBM SPSS
statistics version 20. Significance was declared at P ≤ 0.05.
3. Results and Discussion
3.1 Production and analysis of LeCel7A
For the production of LeCel7A, the LeCel7A gene from L. edodes was optimized
for the preferred codon usage and expressed in P. pastoris in the present study. The
native LeCel7A includes more than 19 % codons, such as GGC (Gly), UGU (Cys),
and AGC (Ser), which share <10 ‰ usage in P. pastoris. These rare codons were
replaced by preferred ones which were more frequently used in P. pastoris. In
addition, an algorithm (GenScript Nanjing Co., Ltd., Nanjing, China) with parameter
settings was used to optimize the mRNA secondary structure and repeat sequences of
LeCel7A, and the G+C content was adjusted from 52.8% in original gene to 40.8% in
optimized gene, which is much closer to the G+C content of P. pastoris (42.7%). The
codon-optimized gene shares 77.1% identity with the original LeCel7A gene.
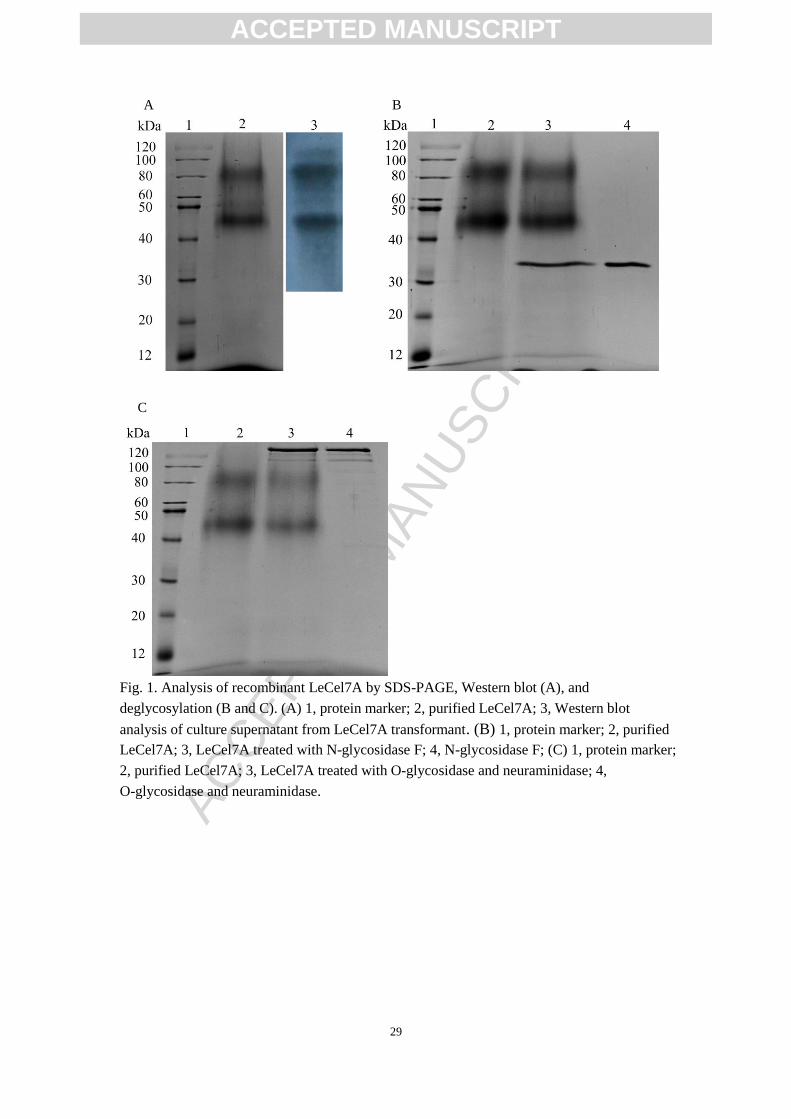
The LeCel7A expressed and purified was verified by SDS-PAGE and Western blot
(Fig. 1A). Two intense protein bands at about 83 kDa and 47 kDa were observed in
the supernatant of the recombinant strain compared with the wild type. Similarly,
Taipakova et al. (2011) also observed two protein bands during the heterogenous
expression of LeCel7A using Escherichia coli [24]. One protein band with the
molecular weight of 83 kDa was larger than the predicted size (51 kDa) based on the
primary sequence of LeCel7A in the present study, which maybe resulted partly from
the increased glycosylation of LeCel7A when expressed in P. pastoris. One potential
ACCEPTED MANUSCRIPT

ACC
EPTE
D M
ANU
SCR
IPT
13
N-glycosylation sites and 40 potential O-β-GlcNAc attachment sites were found in the
LeCel7A sequence using the NetNGlyc 1.0
(http://www.cbs.dtu.dk/services/NetNGlyc/) and YinOYang 1.2 tools
(http://www.cbs.dtu.dk/services/YinOYang/). The glycosylation of purified LeCel7A
was further verified using N-glycosidase and O-glycosidase. However, the
glycosidase treatment did not cause a significant shift in migration (Fig. 1B and Fig.
1C). The results are confusing and need to be elucidated in further studies.
3.2 Characteristics of recombinant LeCel7A
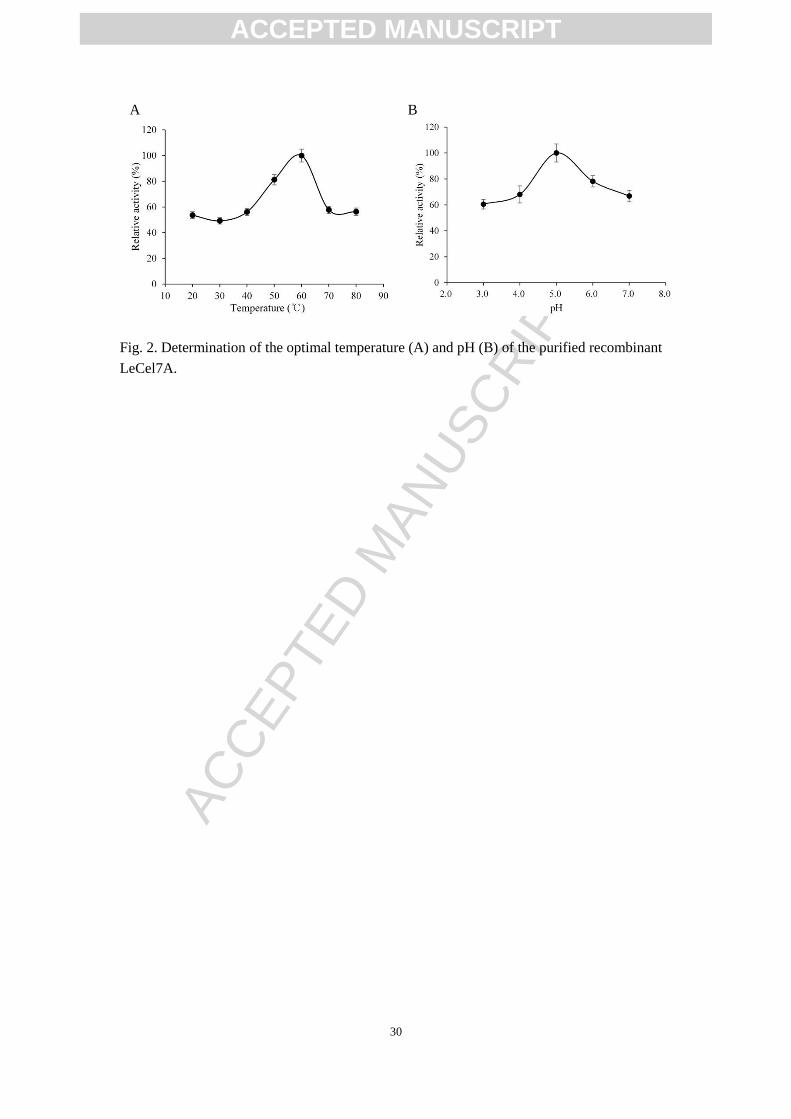
The recombinant LeCel7A exhibited high hydrolytic activity ranging from 50-60℃
and the optimum temperature for enzyme activity was at 60℃ (Fig. 2A). Though the
activity decreased dramatically at 70-80°C, it still kept the 60% of its optimum
activity, which suggested that the LeCel7A in the present study featured resistance to
high temperature. The recombinant LeCel7A showed maximum activity at pH 5.0 and
showed broad pH adaptability (>60% of the maximum activity at pH 3.0-7.0) (Fig.
2B). Current results were partly similar to the report by Bissenbaev et al. (2014), in
which the activity for recombinant LeCel7A by Saccharomyces cerevisiae was shifted
to high values at 60-70°C [25], but were different from the report by Taipakova et al.
(2011), in which the activity for recombinant LeCel7A by E. coli was maximum at
50°C and pH 7.0 [24]. Different expression system used in these studies may be
responsible for the discrepant temperature and pH dependency of LeCel7A [26, 27].
3.3 Enzymatic hydrolysis of agricultural straws
Abilities of LeCel7A to hydrolyze different agricultural straws were analyzed
ACCEPTED MANUSCRIPT

ACC
EPTE
D M
ANU
SCR
IPT
14
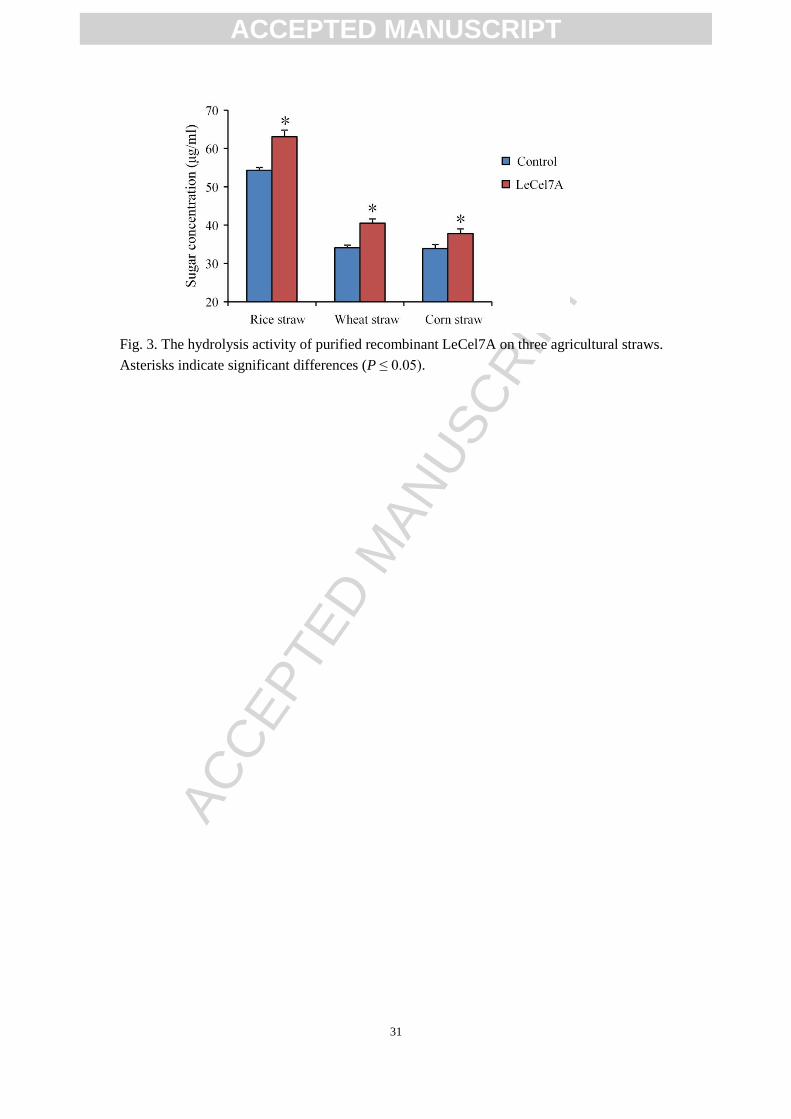
(Fig. 3). The results showed that LeCel7A could significantly accelerated the
hydrolysis of all agricultural straws. After 24 h of incubation, the reducing sugars
content was increased by 5.7%, 23.1%, and 24.0% during the enzymolysis of rice
straw, wheat straw, and corn straw, respectively, relative to the control group.
Similarly, previous studies also found that Cel7A from some fungus exhibited high
hydrolytic efficiency toward agricultural straws [16, 17]. In addition, the dramatical
ability of LeCel7A to hydrolyze agricultural straws may help to explain why
significant cellulose losses were observed in L. edodes cultivation using agricultural
straws [4, 11, 12].
3.4 Effects of LeCel7A on in vitro ruminal fermentation of agricultural straws
Effects of LeCel7A on ruminal fermentation and microbial community in in vitro
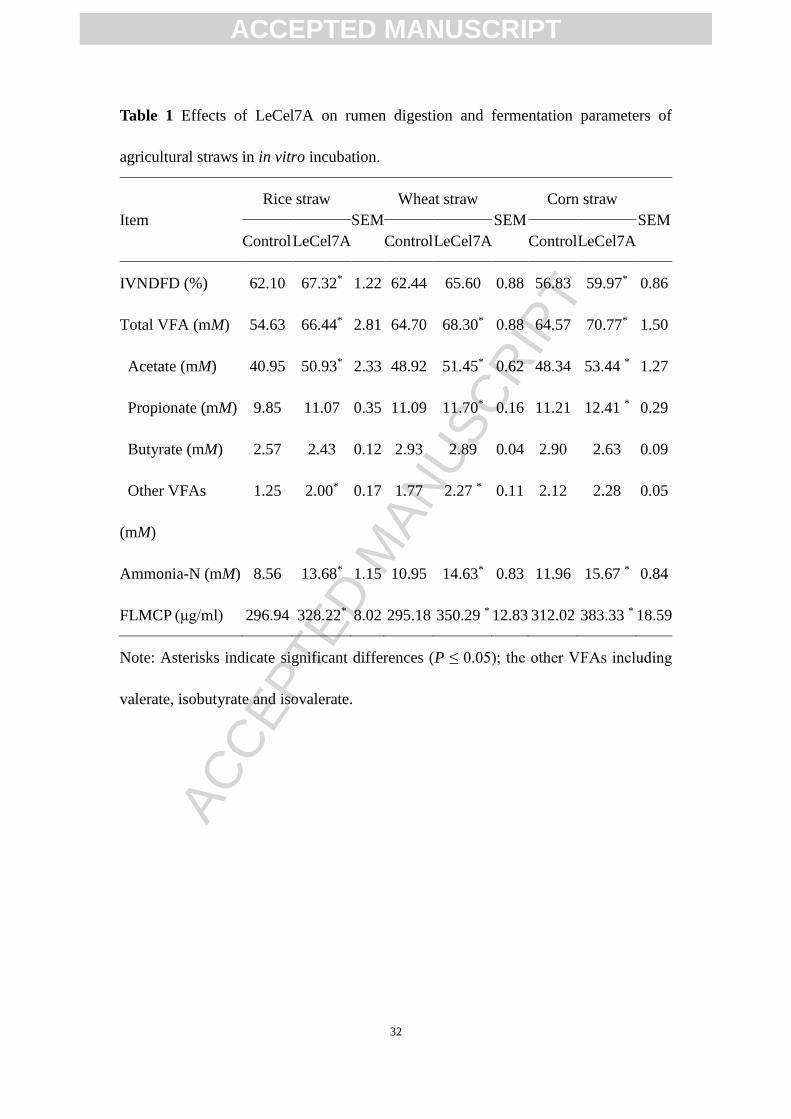
cultures of agricultural straws were investigated (Table 1). The results showed that
LeCel7A increased IVNDFD for rice straw, wheat straw, and corn straw by 8.4%,
5.1%, and 5.5%, respectively, compared with controls. After incubation of 48 h,
regardless of agricultural straws, LeCel7A did not affect the ruminal pH, averaging
6.76 (data not shown). Supplemental LeCel7A increased the total VFA concentration
by 3.9%, 5.5%, 9.3% for rice straw, wheat straw, and corn straw, respectively, relative
to controls. The acetate and propionate concentrations were also greater in
LeCel7A-added agricultural straws than in the controls, which resulted in greater total
VFA production observed in the treatments. Regardless of agricultural straws, the
ammonia-N concentration was increased by added LeCel7A. Supplemental LeCel7A
increased FLMCP synthesis for rice straw, wheat straw, and corn straw by 10.5%,
ACCEPTED MANUSCRIPT

ACC
EPTE
D M
ANU
SCR
IPT
15
18.7%, and 22.9%, respectively.
The high-throughput sequencing technology was used to investigate the effects of
LeCel7A on microbial community in in vitro ruminal incubation of agricultural straws.
To explore the dissimilarities in microbial composition between LeCel7A and control
groups, PCoA analysis based on Bray-Curtis metric was performed (Fig. 4). The
results showed that the microbiota clustered separately and axes accounted for 78.2%,
67.8%, and 74.7% of the total variation detected for rice straw, wheat straw, and corn
straw, respectively, suggesting that certain bacterial species may be affected by
LeCel7A.
Based on the Silva taxonomic database and using the analysis program Uparse, the
bacterial OTUs were classified and confirmed to 31 phyla and 296 genera in this
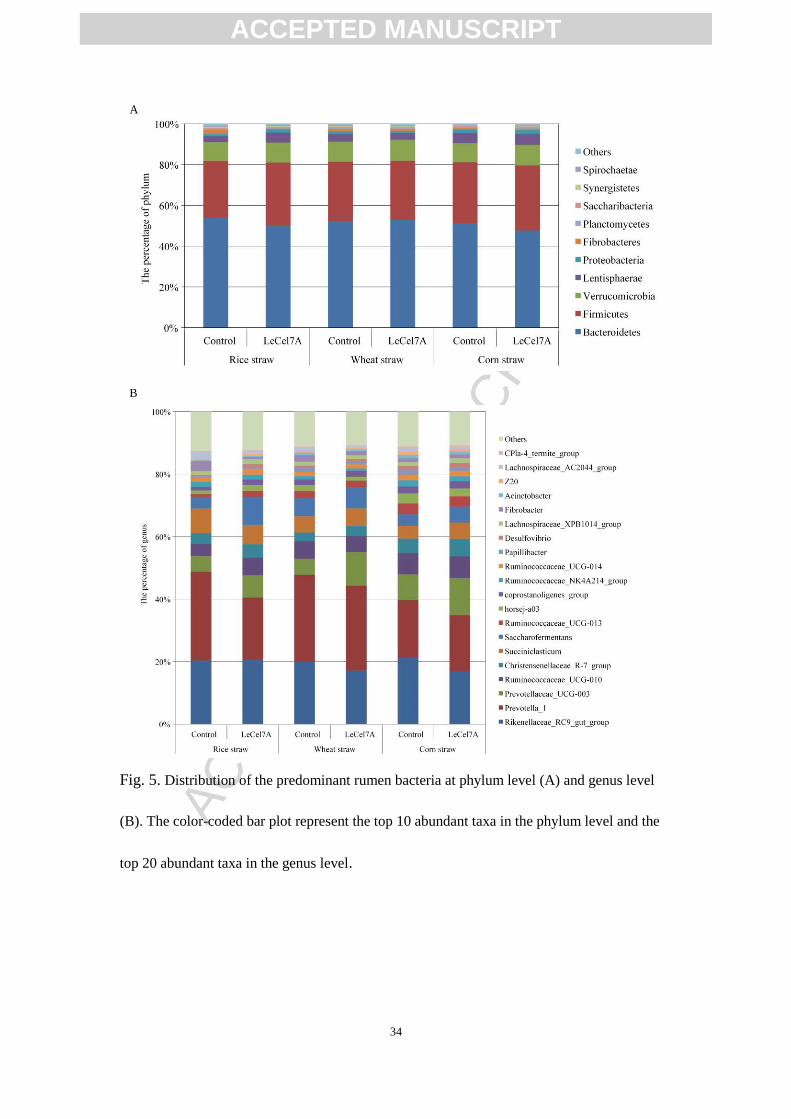
research. The dominant bacterial phylum was Bacteroidetes (47.7-53.9%), followed
by Firmicutes (27.9%-31.9%) and Verrucomicrobia (9.2-10.3%) in all groups (Fig.
5A). At the genus level, Prevotella_1 (9.8-17.9%) and Rikenellaceae_RC9_gut_group
(9.8-12.8%) were the dominant bacteria (Fig. 5B). To identify the taxon had the great
impact on microbial community, LEfSe analysis was performed and biomarkers were
found in both LeCel7A and control groups for three straws (Fig. 6). There were 36, 51
and 94 taxa found as biomarker for rice straw, wheat straw, and corn straw,
respectively. With rice straw, 16 taxa were increased by LeCel7A, among which
Saccharofermentans, Desulfovibrio, Prevotellaceae_UCG_003 and some genus
belonging to Ruminococcaceae, such as Ruminococcaceae_UCG_010,
Ruminococcaceae_UCG_013 and so on, were found to be predominant bacteria; 20
ACCEPTED MANUSCRIPT

ACC
EPTE
D M
ANU
SCR
IPT
16
taxa were decreased by LeCel7A, among which Bacteroidetes,
Lachnospiraceae_AC2044_group, and Fibrobacter were found to be predominant
bacteria. With wheat straw, 23 taxa were increased by LeCel7A, among which
Prevotellaceae_UCG_003, Saccharofermentans, and Succiniclasticum were
predominant bacteria; 28 taxa were decreased by LeCel7A, among which
Fibrobacteres, Lentisphaerae, and Lachnospiraceae_AC2044_group were
predominant bacteria. With corn straw, 53 taxa were increased by LeCel7A, among
which Prevotellaceae_UCG_003, Saccharofermentans, and
Christensenellaceae_R_7_group were found to be predominant bacteria; 41 taxa were
decreased by LeCel7A, among which Bacteroidetes,
Ruminococcaceae_NK4A214_group, and Lachnospiraceae_AC2044_group were
predominant bacteria.
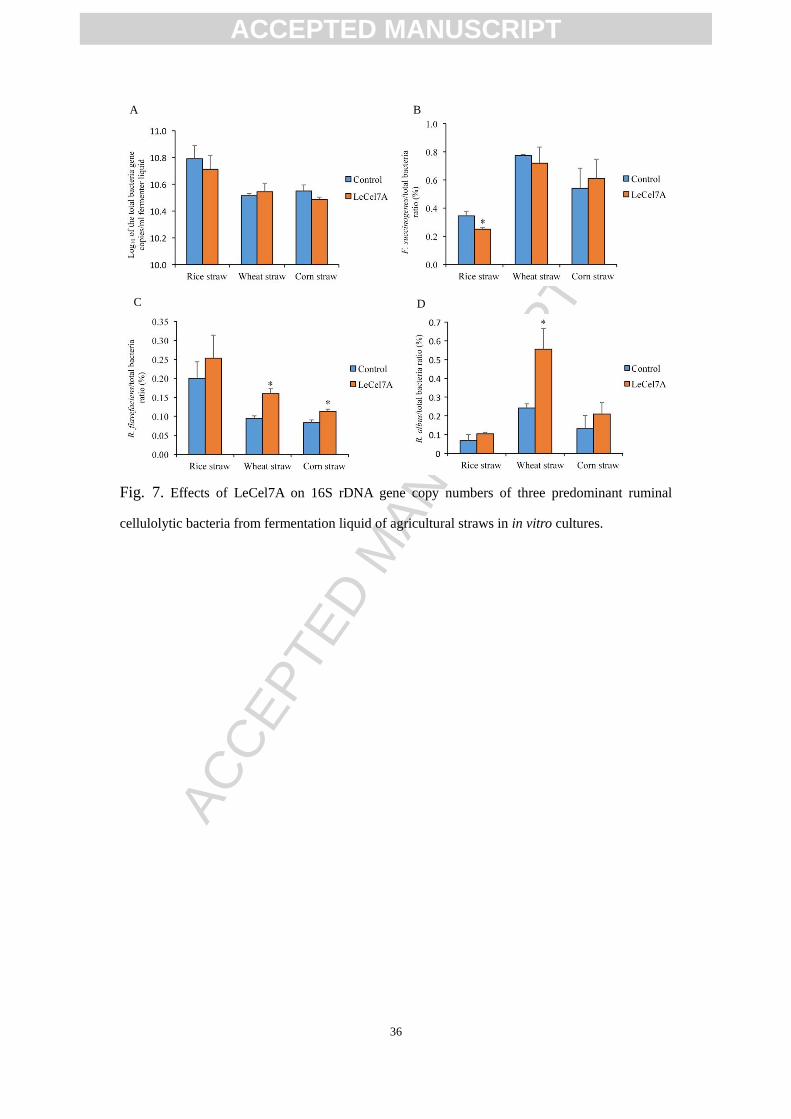
Besides the high-throughput sequencing technology, the quantity of total bacteria
and relative abundance of three representative fibrolytic bacteria species in the
fermentation liquid was determined using real-time PCR (Fig. 7). With rice straw,
LeCel7A tended to increase the relative abundance of R. flavefaciens and R. albus but
significantly decreased the relative abundance of F. succinogenes. With wheat straw,
LeCel7A significantly increased the relative abundance of R. flavefaciens and R. albus.
With corn straw, the relative abundance of R. flavefaciens was significantly increased
by supplemental LeCel7A.
Previous studies have shown that exogenous fibrolytic enzymes could enhance the
in vitro rumen fermentation of agricultural straws, including increased VFA
ACCEPTED MANUSCRIPT

ACC
EPTE
D M
ANU
SCR
IPT
17
production and fiber digestibility [28-30]. Exogenous fibrolytic enzyme increased the
ruminal microorganism colonization of substrate, which accelerated the hydrolysis of
lignocellulose substrate [31]. However, the fibrolytic enzymes used in these studies
was mostly composite enzymes, and little information is available for the effects of
purified cellobiohydrolases on the ruminal fermentation. In current study, LeCel7A
significantly increased the IVNDFD during the incubations of three agricultural
straws, which might be due to the accelerated enzymatic hydrolysis of fibrous
material in the three straws caused by LeCel7A. Accordingly the acetate concentration,
total VFA concentration, and fibrolytic bacteria were significantly increased by
supplemental LeCel7A regardless of straws. In the rumen, VFAs are produced as
end-products of microbial fermentation and dietary carbohydrates including cellulose,
hemicellulose and starch, are the main fermentation substrates [32]. The relative
concentrations of the individual VFAs are determined by the type of dietary
carbohydrate [33]. Fermentation of structural carbohydrates (i.e. cellulose and
hemicellulose), compared to fermentation of starch, encourages the growth of acetate
producing bacterial species and consequently yielded high amount of acetate [32].
The relative abundance of genus Prevotellaceae_UCG_003 and Saccharofermentans
were increased by LeCel7A regardless of straws in the current study. Similarly, the
Prevotellaceae_UCG_003 abundance was increased significantly with increased
dietary NDF content or fiber digestibility of straws in previous studies [22, 34-36].
The genus Prevotellaceae_UCG_003 belongs to the family Prevotellaceae, which has
long been recognized as one of the predominant bacterial group inhabiting the rumen
ACCEPTED MANUSCRIPT

ACC
EPTE
D M
ANU
SCR
IPT
18
[37]. Prevotellaceae could utilize various sugars and produce acetic and succinic acids
as major fermentation end products [38]. Among the family Prevotellacaea, some
genus or species contain endocellulase, β-glucosidase, and endoxylanase and are
involved in utilization of starch and plant cell-wall components digestion [39-41]. As
an uncultured genus, Prevotellaceae_UCG-003 maybe has the similar hydrolysis
activity, which contributes to the increase of IVNDFD for straws, but requires further
study. Saccharofermentans is known to play an important role in fiber degradation and
produces acetate as main end-products [42, 43]. Zhang et al. (2017) found that
Saccharofermentans was linearly increased with increased dietary forage content and
was positively associated with ruminal acetate concentration, which was consistent
with the current study [43]. The relative abundance of genus Desulfovibrio was
increased by LeCel7A for rice straw and corn straw. Desulfovibrio belongs to
sulfate-reducing bacteria and could metabolize lactate and pyruvate into acetate and
CO2 when the latter serves as an electron donor for sulfate reduction [44]. Therefore,
it was understandable that the increased acetate concentration associated with
increased relative abundance of genus Desulfovibrio. Some genus of
Ruminococcaceae, i.e. Ruminococcaceae_UCG_010, Ruminococcaceae_UCG_011,
and Ruminococcaceae_UCG_013 for rice straw, Ruminococcus_1 for wheat straw,
and Ruminococcaceae_UCG_010 and Ruminococcaceae_UCG_005 for corn straw,
were increased by LeCel7A in the present study. The family Ruminococcaceae
belongs to the phylum Firmicutes, which mainly digest cellulose, hemicellulose, and
some simple sugars widely existing dietary forage resulting in increased acetate
ACCEPTED MANUSCRIPT

ACC
EPTE
D M
ANU
SCR
IPT
19
concentration [45, 46]. Though the metabolic characters of members from
Ruminococcaceae were variable, most of its genus are acetate producing bacteria and
could ferment carbohydrates to acetate as the major products [47]. As the
representative fibrolytic bacteria, R. flavefaciens, F. succinogenes, and R. albus were
all the important members of Ruminococcaceae and contribute significantly to fiber
metabolism and acetate production [48]. Previous in vitro studies showed that
proliferation of the three fibrolytic bacteria was often accompanied by the increase in
fiber digestion and acetate production [49, 50]. The IVNDFD, acetate concentration,
and part of the three representative fibrolytic bacteria were increased by LeCel7A in
the present study, which were in agreement with these reports. The relative abundance
of genus Succiniclasticum and Bacteroides were increased by LeCel7A for wheat
straw and corn straw. The genus Succiniclasticum belonging to the family
Acidaminococcaceae could convert succinate to propionate, the most important
precursor of glucose in ruminants [51, 52]. Therefore, the increased Succiniclasticum
may be relative to the increased propionate concentration by LeCel7A for wheat straw
and corn straw. The Bacteroides belongs to the family Bacteroidaceae. Many studies
reported that some Bacteroides species could ferment plant fiber with acetic acid and
succinic acid as major metabolic end products [53, 54], which means that the
Bacteroides maybe plays an important role in ruminal fiber digestibility. Besides the
increased taxa, some taxa including Bacteroidetes, Fibrobacteres, Lentisphaerae, and
so on, were reduced by supplemental LeCel7A. The microbial community was
dominated by Bacteroidetes and Firmicutes regardless of groups in the current study,
ACCEPTED MANUSCRIPT

ACC
EPTE
D M
ANU
SCR
IPT
20
which is consistent with the study by de Oliveira et al. (2013) [55]. The reduction of
Bacteroidetes and other taxa may be due to the competition of faster growing
cellulolytic bacteria belonging to Firmicutes in obtaining nutrients or energy [56, 57].
Supplemental LeCel7A significant increased the ammonia-N concentration for
three straws in the present study. The ammonia-N concentration during fermentation
depends on the extent of dietary nitrogen degradation and nitrogen uptake by ruminal
bacteria. The higher production of ammonia-N for LeCel7A treatments may be related
to higher dietary nitrogen degradation compared with control treatments, and
increased protein or amino-acid degrading bacteria. The genus Desulfovibrio,
Bacteroides, and phylum Synergistetes increased by LeCel7A could degrade peptides
or amino acids and played important role in dietary nitrogen degradation [58-60],
which supported the deduction. The concentrations of ammonia-N exceeded 5 mg/dl
in all groups, which suggests that microbial growth may not be limited by available
ammonia-N in the present study [61]. Increased microbial protein production with
supplemental LeCel7A may relate to the synchrony between protein and carbohydrate
digestion during fermentation. Microbial protein synthesis depends largely on the
available amount and fermentation rate of carbohydrates and N in the rumen. With
enough available ammonia-N, increased NDF digestion by LeCel7A may stimulate
the growth of more ruminal bacteria, which was consistent with the study by Sommart
et al. (2000) [62].
4. Conclusions
The LeCel7A was expressed and produced using P. pastoris and shown
ACCEPTED MANUSCRIPT

ACC
EPTE
D M
ANU
SCR
IPT
21
maximum activity at 60°C and pH 5.0. The LeCel7A could improve the hydrolysis
and in vitro ruminal fermentation of agricultural straws by increasing the digestion of
fiber, changing the microbial community, stimulating the growth of fibrolytic bacteria
and production of VFA, as well as increased rumen microbial protein synthesis. The
LeCel7A might be useful for improving the utilization of agricultural straws as a feed
additive for ruminants.
Funding
This work was supported by National Key R&D Program of China
(2018YFD0501804), the National Natural Science Foundation of China (31760687),
and the National Beef Cattle Industry Technology & System (CARS-Beef Cattle
System: CARS-38).
Notes
The authors declare no conflict of interest.
References
[1] M. Sain, S. Panthapulakkal, Bioprocess preparation of wheat straw fibers and their characterization,
Ind. Crop. Prod. 23 (2006) 1-8.
[2] I.K. Cann, Y. Kobayashi, M. Wakita, S. Hoshino, Effects of 3 chemical treatments on in vitro
fermentation of rice straw by mixed rumen microbes in the presence or absence of anaerobic rumen
fungi, Reprod. Nutr. Dev. 34 (1994) 47-56.
[3] Alborés, S. Pianzzola, M.J. Soubes, M. Cerdeiras, M. Pía, Biodegradation of agroindustrial wastes
by Pleurotus spp for its use as ruminant feed, Electron. J. Biotechn. 9 (2006) 215-220.
ACCEPTED MANUSCRIPT

ACC
EPTE
D M
ANU
SCR
IPT
22
[4] X. Zhao, J. Gong, Z. Shan, K. Ouyang, X. Song, C. Fu, L. Xu, M. Qu, Effect of Fungal Treatments
of Rape Straw on Chemical Composition and in vitro Rumen Fermentation Characteristics,
BioResources 10 (2015) 622-637.
[5] B.W. Alexander, A.H. Gordon, J.A. Lomax, A. Chesson, Composition and rumen degradability of
straw from three varieties of oilseed rape before and after alkali, hydrothermal and oxidative treatment,
J. Sci. Food Agric. 41 (1987) 1-15.
[6] C. Sarnklong, J.W. Cone, W. Pellikaan, W.H. Hendriks, Utilization of Rice Straw and Different
Treatments to Improve Its Feed Value for Ruminants: A Review, Asian-Aust. J. Anim. Sci. 23 (2010)
680-692.
[7] J.X. Liu, E.R. Orskov, X.B. Chen, Optimization of steam treatment as a method for upgrading rice
straw as feeds, Anim. Feed Sci. Tech. 76 (1999) 345-357.
[8] P.J.V. Soest, Rice straw, the role of silica and treatments to improve quality, Anim. Feed Sci.
Tech.130 (2006) 137-171.
[9] O. Loera, Efficiency of lignocellulolytic extracts from thermotolerant strain fomes sp. eum1:
stability and digestibility of agricultural wastes, J. Agr. Sci. Tech-Iran. 15 (2013) 229-240.
[10] C.C. Lee, D.W. Wong, G.H. Robertson, Cloning and characterization of two cellulase genes from
Lentinula edodes, Fems Microbiol. Lett. 205 (2001) 355-360.
[11] V.D. Tuyen, J. Cone, J. Baars, A. Sonnenberg, W. Hendriks, Fungal strain and incubation period
affect chemical composition and nutrient availability of wheat straw for rumen fermentation, Bioresour.
Technol. 111 (2012) 336-342.
[12] D. Tuyen, H. Phuong, J. Cone, J. Baars, A. Sonnenberg, W. Hendriks, Effect of fungal treatments
of fibrous agricultural by-products on chemical composition and in vitro rumen fermentation and
ACCEPTED MANUSCRIPT

ACC
EPTE
D M
ANU
SCR
IPT
23
methane production, Bioresour. Technol. 129 (2013) 256-263.
[13] G. Banerjee, J.S. Scottcraig, J.D. Walton, Improving enzymes for biomass conversion: a basic
research perspective, Bioenerg. Res. 3 (2010) 82-92.
[14] T.T. Teeri, Crystalline cellulose degradation: new insight into the function of cellobiohydrolases,
Trends. Biotechnol. 15 (1997) 160-167.
[15] Y.L. Li, H. Li, A.N. Li, D.C. Li, Cloning of a gene encoding thermostable cellobiohydrolase from
the thermophilic fungus Chaetomium thermophilum and its expression in Pichia pastoris, J. Appl.
Microbiol. 106 (2010) 1867-1875.
[16] F. Hao, L. Xia, Heterologous expression and production of Trichoderma reesei cellobiohydrolase
II in Pichia pastoris and the application in the enzymatic hydrolysis of corn stover and rice straw,
Biomass Bioenerg. 78 (2015) 99-109.
[17] H. Billard, Optimization of a synthetic mixture composed of major Trichoderma reesei enzymes
for the hydrolysis of steam-exploded wheat straw, Biotechnol. Biofuels, 5 (2012) 9.
[18] J.W. Cone, A.H.V. Gelder, G.J.W. Visscher, L. Oudshoorn, Influence of rumen fluid and substrate
concentration on fermentation kinetics measured with a fully automated time related gas production
apparatus, Anim. Feed Sci. Tech. 61 (1996) 113-128.
[19] P.J. Van Soest, J.B. Robertson, B.A. Lewis, Methods for dietary fiber, neutral detergent fiber, and
nonstarch polysaccharides in relation to animal nutrition, J. Dairy Sci. 74 (1991) 3583-3597.
[20] M.W. Weatherburn, Phenol-hypochlorite reaction for determination of ammonia, Anal. Chem. 39
(1967) 971-974.
[21] H.P. Makkar, O.P. Sharma, R.K. Dawra, S.S. Negi, Simple determination of microbial protein in
rumen liquor, J. Dairy Sci. 65 (1982) 2170-2173.
ACCEPTED MANUSCRIPT

ACC
EPTE
D M
ANU
SCR
IPT
24
[22] L. Li, M. Qu, C. Liu, L. Xu, K. Pan, X. Song, K. Ouyang, Y. Li, X. Zhao, Expression of a
recombinant Lentinula edodes xylanase by Pichia pastoris and its effects on ruminal fermentation and
microbial community in in vitro incubation of agricultural straws, Front. Microbiol. 9 (2018) 2944.
[23] N. Segata, J. Izard, L. Waldron, D. Gevers, L. Miropolsky, W.S. Garrett, C. Huttenhower,
Metagenomic biomarker discovery and explanation, Genome Biol. 12 (2011) R60.
[24] S. Taipakova, B. Smailov, G. Stanbekova, A. Bissenbaev, Cloning and expression of Lentinula
edodes cellobiohydrolase gene in E. coli and characterization of the recombinant enzyme, Journal of
Cell and Molecular Biology 9 (2011) 53-61.
[25] A. Bissenbaev, S. Taipakova, I. Smekenov, Expression of cellobiohydrolase CEL7A gene from
Lentinula edodes in Saccharomyces cerevisiae with use of two different promoters, J. Biotechnol. 185
(2014) S69-S69.
[26] L.S. Hsieh, C.S. Yeh, H.C. Pan, C.Y. Cheng, C.C. Yang, P.D. Lee, Cloning and expression of a
phenylalanine ammonia-lyase gene in Escherichia coli and Pichia pastoris, Protein Expres. Purif. 71
(2010) 224-230.
[27] L. Marianne Wittrup, U.T. Bornscheuer, H. Karl, Expression of Candida antarctica lipase B in
Pichia pastoris and various Escherichia coli systems, Protein Expres. Purif. 62 (2008) 90-97.
[28] J.S. Eun, S.H. Hong, K.A. Beauchemin, M.W. Bauer, Exogenous enzymes added to untreated or
ammoniated rice straw: effects on in vitro fermentation characteristics and degradability, Anim. Feed
Sci. Tech. 131 (2006) 87-102.
[29] L.Y. Zhao, Y.J. Peng, J.K. Wang, J.X. Liu, Effects of exogenous fibrolytic enzyme on in vitro
ruminal fiber digestion and methane production of corn stover and corn stover based mixed diets, Life
Sci. J. 12(2015) 1-9.
ACCEPTED MANUSCRIPT

ACC
EPTE
D M
ANU
SCR
IPT
25
[30] L.A. Giraldo, M.D. Carro, M.J. Ranilla, M.L. Tejido, A.H. Mohamed, A. Priolo, L. Biondi, H.B.
Salem, P. Morandfehr, In vitro ruminal fermentation of low-quality forages as influenced by the
treatment with exogenous fibrolytic enzymes, Options Méditerranéennes Series A, 74 (2007) 263-267.
[31] Y. Wang, J.E. Ramirez-Bribiesca, L.J. Yanke, A. Tsang, T.A. Mcallister, Effect of exogenous
fibrolytic enzyme application on the microbial attachment and digestion of barley straw in vitro,
Asian-Aust. J. Anim. Sci. 25 (2012) 66-74.
[32] J. Dijkstra, Production and absorption of volatile fatty acids in the rumen, Livest. Prod. Sci. 39
(1994) 61-69.
[33] J. France, J. Dijkstra, Volatile fatty acid production, in: Dijkstra, J., Forbes, J.M., France, J. (Eds.),
Quantitative aspects of ruminant digestion and metabolism. CABI Publishing/CAB International,
Wallingford, pp: 159.
[34] Q. Zhao, P. Zheng, Y. Tian, H. Huang, Effects of NFC/NDF in diet on rumen fermentation
parameters and microbial flora diversity in calves, Животноводство и кормопроизводство 101 (2018)
7-15.
[35] X. Zhang, L. Wang, Effects of dietary neutral detergent fibre level on structure and composition of
rumen bacteria in goats, Chinese Journal of Animal Nutrition 30 (2018) 1377-1386.
[36] L. Li, M. Qu, C. Liu, L. Xu, K. Pan, K. OuYang, X. Song, Y. Li, H. Liang, Z. Chen, Effects of
recombinant swollenin on the enzymatic hydrolysis, rumen fermentation, and rumen microbiota during
in vitro incubation of agricultural straws, Int. J. Biol. Macromol. 122 (2019) 348-358.
[37] K. Singh, T. Jisha, B. Reddy, N. Parmar, A. Patel, A. Patel, C. Joshi, Microbial profiles of liquid
and solid fraction associated biomaterial in buffalo rumen fed green and dry roughage diets by tagged
16S rRNA gene pyrosequencing, Mol. Biol. Rep. 42 (2015) 95-103.
ACCEPTED MANUSCRIPT

ACC
EPTE
D M
ANU
SCR
IPT
26
[38] N.R. Krieg, J.T. Staley, D.R. Brown, B.P. Hedlund, B.J. Paster, N.L. Ward, W. Ludwig, W.B.
Whitman, Family V. Prevotellaceae fam. nov, Bergey’s Manual of Systematic Bacteriology, 4(2011) 85.
[39] D.O. Krause, S.E. Denman, R.I. Mackie, M. Morrison, A.L. Rae, G.T. Attwood, C.S. McSweeney,
Opportunities to improve fiber degradation in the rumen: microbiology, ecology, and genomics, FEMS
Microbiol. Rev. 27 (2003) 663-693.
[40] M.A. Cotta, Interaction of ruminal bacteria in the production and utilization of
maltooligosaccharides from starch, Appl. Environ. Microb. 58 (1992) 48-54.
[41] H. Matsui, K. Ushida, K. Miyazaki, Y. Kojima, Use of ratio of digested xylan to digested cellulose
(X/C) as an index of fiber digestion in plant cell-wall material by ruminal microorganisms, Anim. Feed
Sci. Tech. 71 (1998) 207-215.
[42] J.H. Liu, M.L. Zhang, R.Y. Zhang, W.Y. Zhu, S.Y. Mao, Comparative studies of the composition
of bacterial microbiota associated with the ruminal content, ruminal epithelium and in the faeces of
lactating dairy cows, Microb. Biotechnol. 9 (2016) 257-268.
[43] J. Zhang, H. Shi, Y. Wang, S. Li, Z. Cao, S. Ji, Y. He, H. Zhang, Effect of dietary forage to
concentrate ratios on dynamic profile changes and interactions of ruminal microbiota and metabolites
in Holstein heifers, Front. Microbiol. 8 (2017) 2206.
[44] G. Voordouw, The genus Desulfovibrio: the centennial, Appl. Environ. Microb. 61 (1995)
2813-2819.
[45] M. Sandri, C. Manfrin, A. Pallavicini, B. Stefanon, Microbial biodiversity of the liquid fraction of
rumen content from lactating cows, Animal 8 (2014) 572-579.
[46] J. Klang, S. Theuerl, U. Szewzyk, M. Huth, R. Tölle, M. Klocke, Dynamic variation of the
microbial community structure during the long‐ time mono‐ fermentation of maize and sugar beet
ACCEPTED MANUSCRIPT

ACC
EPTE
D M
ANU
SCR
IPT
27
silage, Microb. Biotechnol. 8.(2015) 764-775.
[47] F.A. Rainey, Family VIII. Ruminococcaceae fam. nov, Bergey’s Manual of Systematic
Bacteriology 3 (2009) 1016-1018.
[48] S. Koike, Y. Kobayashi, Fibrolytic rumen bacteria: their ecology and functions, Asian-Aust. J.
Anim. Sci 22 (2009) 131-138.
[49] H.G. Sung, Y. Kobayashi, J. Chang, A. Ha, I.H. Hwang, J. Ha, Low ruminal pH reduces dietary
fiber digestion via reduced microbial attachment, Asian-Aust. J. Anim. Sci 20 (2006) 200-207.
[50] Y. Wang, T.W. Alexander, T.A. McAllister, In vitro effects of phlorotannins from Ascophyllum
nodosum (brown seaweed) on rumen bacterial populations and fermentation, J. Sci. Food. Agr. 89(13)
(2009) 2252-2260.
[51] F.A. Rainey, Genus XXIII. Succiniclasticum van Gylswyk 1995, 298VP, Bergey’s Manual of
Systematic Bacteriology 3 (2009) 1116-1117.
[52] M. Hélène, T. Corinne, C. Josiane, J.P. Hélène, R. Frédéric, G. Bernard, C. Jean-Philippe, J.B.
Estelle, Negativicoccus succinicivorans gen. nov., sp. nov., isolated from human clinical samples,
emended description of the family Veillonellaceae and description of Negativicutes classis nov.,
Selenomonadales ord. nov. and Acidaminococcaceae fam. nov. in the bac, Int. J. Syst. Evol. Microbiol.
60 (2010) 1271-1279.
[53] Y. Song, C. Liu, S. Finegold, Genus I. Bacteroides Castellani and Chalmers 1919, 959AL emend.
Shah and Collins 1989, 85, Bergey’s Manual of Systematic Bacteriology 4 (2011) 27-41.
[54] A. Salyers, J. Vercellotti, S. West, T. Wilkins, Fermentation of mucin and plant polysaccharides by
strains of Bacteroides from the human colon, Appl. Environ. Microb. 33 (1977) 319-322.
[55] M.N.V. de Oliveira, K.A. Jewell, F.S. Freitas, L.A. Benjamin, M.R. Tótola, A.C. Borges, C.A.
ACCEPTED MANUSCRIPT

ACC
EPTE
D M
ANU
SCR
IPT
28
Moraes, G. Suen, Characterizing the microbiota across the gastrointestinal tract of a Brazilian Nelore
steer, Vet. Microbiol. 164 (2013) 307-314.
[56] M. Theodorou, J. France, Rumen microorganisms and their interactions, Quantitative aspects of
ruminant digestion and metabolism , CABI, 2005.
[57] G. Witten, F. Richardson, Competition of three aggregated microbial species for four substrates in
the rumen, Ecol. Model. 164 (2003) 121-135.
[58] X. Zhao, Z. Chen, S. Zhou, X. Song, K. Ouyang, K. Pan, L. Xu, C. Liu, M. Qu, Effects of daidzein
on performance, serum metabolites, nutrient digestibility, and fecal bacterial community in bull calves,
Anim. Feed Sci. Tech. 225 (2017) 87-96.
[59] Y. Li, Q. Liao, M. Lin, D. Zhong, L. Wei, B. Han, H. Miao, M. Yao, Z. Xie, An integrated
metabonomics and microbiology analysis of host-microbiota metabolic interactions in rats with Coptis
Chinensis-induced diarrhea, RSC Adv. 5 (2015) 79329-79341.
[60] E. Jumas-Bilak, L. Roudiere, H. Marchandin, Description of 'Synergistetes' phyl. nov. and
emended description of the phylum 'Deferribacteres' and of the family Syntrophomonadaceae, phylum
'Firmicutes', Int. J. Syst. Evol. Micr. 59 (2009) 1028-1035.
[61] L. Satter, L. Slyter, Effect of ammonia concentration on rumen microbial protein production in
vitro, Brit. J. Nutr. 32 (1974) 199-208.
[62] K. Sommart, D. Parker, P. Rowlinson, M. Wanapat, Fermentation characteristics and microbial
protein synthesis in an in vitro system using cassava, rice straw and dried ruzi grass as substrates,
Asian-Aust. J. Anim. Sci. 13 (2000) 1084-1093.
ACCEPTED MANUSCRIPT
ACC
EPTE
D M
ANU
SCR
IPT
29
Fig. 1. Analysis of recombinant LeCel7A by SDS-PAGE, Western blot (A), and
deglycosylation (B and C). (A) 1, protein marker; 2, purified LeCel7A; 3, Western blot
analysis of culture supernatant from LeCel7A transformant. (B) 1, protein marker; 2, purified
LeCel7A; 3, LeCel7A treated with N-glycosidase F; 4, N-glycosidase F; (C) 1, protein marker;
2, purified LeCel7A; 3, LeCel7A treated with O-glycosidase and neuraminidase; 4,
O-glycosidase and neuraminidase.
A B
C
ACCEPTED MANUSCRIPT
ACC
EPTE
D M
ANU
SCR
IPT
30
Fig. 2. Determination of the optimal temperature (A) and pH (B) of the purified recombinant
LeCel7A.
A
B
ACCEPTED MANUSCRIPT
ACC
EPTE
D M
ANU
SCR
IPT
31
Fig. 3. The hydrolysis activity of purified recombinant LeCel7A on three agricultural straws.
Asterisks indicate significant differences (P ≤ 0.05).
ACCEPTED MANUSCRIPT
ACC
EPTE
D M
ANU
SCR
IPT
32
Table 1 Effects of LeCel7A on rumen digestion and fermentation parameters of
agricultural straws in in vitro incubation.
Item
Rice straw
SEM
Wheat straw
SEM
Corn straw
SEM
Control LeCel7A Control LeCel7A Control LeCel7A
IVNDFD (%) 62.10 67.32* 1.22 62.44 65.60 0.88 56.83 59.97* 0.86
Total VFA (mM) 54.63 66.44* 2.81 64.70 68.30* 0.88 64.57 70.77* 1.50
Acetate (mM) 40.95 50.93* 2.33 48.92 51.45* 0.62 48.34 53.44 * 1.27
Propionate (mM) 9.85 11.07 0.35 11.09 11.70* 0.16 11.21 12.41 * 0.29
Butyrate (mM) 2.57 2.43 0.12 2.93 2.89 0.04 2.90 2.63 0.09
Other VFAs
(mM)
1.25 2.00* 0.17 1.77 2.27 * 0.11 2.12 2.28 0.05
Ammonia-N (mM) 8.56 13.68* 1.15 10.95 14.63* 0.83 11.96 15.67 * 0.84
FLMCP (μg/ml) 296.94 328.22* 8.02 295.18 350.29 * 12.83 312.02 383.33 * 18.59
Note: Asterisks indicate significant differences (P ≤ 0.05); the other VFAs including
valerate, isobutyrate and isovalerate.
ACCEPTED MANUSCRIPT

ACC
EPTE
D M
ANU
SCR
IPT
33
Fig. 4. PCoA plot of samples from in vitro ruminal incubation of rice straw (A), wheat
straw (B), and corn straw (C) based on Bray-Curtis distances.
A B
C
ACCEPTED MANUSCRIPT
ACC
EPTE
D M
ANU
SCR
IPT
34
Fig. 5. Distribution of the predominant rumen bacteria at phylum level (A) and genus level
(B). The color-coded bar plot represent the top 10 abundant taxa in the phylum level and the
top 20 abundant taxa in the genus level.
A
B
ACCEPTED MANUSCRIPT

ACC
EPTE
D M
ANU
SCR
IPT
35
Fig. 6. LEfSe analysis displaying the ruminal bacteria change between control group and
LeCel7A group in in vitro ruminal incubation of rice straw (A), wheat straw (B) and corn
straw (C). LDA ≥ 2.5 and P ≤ 0.05 were shown.
A
C
B
ACCEPTED MANUSCRIPT
ACC
EPTE
D M
ANU
SCR
IPT
36
Fig. 7. Effects of LeCel7A on 16S rDNA gene copy numbers of three predominant ruminal
cellulolytic bacteria from fermentation liquid of agricultural straws in in vitro cultures.
A
D C
B
ACCEPTED MANUSCRIPT