factors affecting epizootics of winter ticks and … · 2014-10-15 · factors affecting epizootics...
TRANSCRIPT

FACTORS AFFECTING EPIZOOTICS OF WINTER TICKS ANDMORTALITY OF MOOSE
W.M. Samuel
Department of Biological Sciences, University of Alberta, Edmonton, AB, Canada T6G 2E9
ABSTRACT: Die-olT's ofmoose (Alces alces) associated wilh, or attributed to, winter ticks [Dermaccrt-/o/'iï//ï//j/c7iis) are widespread and have been reported since iheearly part of ihelasl century. Extrinsicfactors such JUS weather and vegetative structure, and host factors such as moose density and. indirectly,tick-induced damage to the hair coat, were examined in an attempt to predict related problems lormoose. The proposal that warmer and shorier winters result in increased stirvival i)f adult female ticksdropping off moose in March and April, and increased tick populations on moose the following winter,was generally confirmed. Annual changes in hair damage and loss on moose, which are documentedfrom the air, coincided with annual changes in numbers of ticks on moose, providing managers wilha survey tool to monitor and estimate changing numbers of ticks. Tick numbers lagged 1 year behindmoose numbers in Elk Island National Park over a 12-year period, and many moose died when numbersof both were high. Several widespread, concurrent die-ofis suggest extrinsic influences play a role,pos.sibly independent ofmoose density. The lack of objective and continuous data sets should guidefuture research etTorts.
ALCES VOL. 43: 39-48 (2007)
Key words: Alces, density, Dermacentor albipictus,. epizootics, hair, moose, mortality, ticks, transmis-sion, weather
Die-offs of moose associated with, orattributed to, winter ticks are numerous andwidespread across North America, having oc-curred since the early part ofthe last century(summarized by Samuel 2004). Almost allpublished and unpublished reports of suchevents indicate finding a certain number,sometimes few, often many, dead or dyingmoose covered with ticks. Tick-associateddie-offs often are widespread and concur-rent, involving many populations ofmoose inscvcral-to-many jurisdictions. For example,the most recent widespread outbreaks of winterticks, accompanied by losses of many moose,occurred in late winter and spring, 2002. Therewere reports from Isle Royale National Park.Minnesota, Maine, New Hampshire, and Ver-mont in the United States, and Alberta, Mani-toba. Ontario, and Saskatchewan in Canada(Samuel et al. 2002, Peterson and Vucetieh2003, Samuel and Crichton 2003). Earlier
concuirent widespread die-offs occurred inlate winter-spring, 1999 and 1992, and oneacross Alberta in 1982 (Samuel 2004).
Die-offs are often attributed to winterticks, perhaps because ticks are obvious andnumerous on dead and dying moose. Unlbr-tunalcly, direct evidence ofthe lethal elTeet ofwinter ticks on moose populations is lacking.There is, however, good information fromexperimental and field studies (reviewed bySamuel et al. 2000) that, at the least, sug-gests that winter ticks are a significant faetorin rapid declines of tnoose numbers. Otherfaetors, such as moose numbers or density,habitai, weather, and prédation, likely alsoplay a role.
The objective here is to review factorspotentially eontributing to tick-related die-offsofmoose using published literature, reports,andobservationsfrom central Alberta, particu-larly Elk Island National Park and vicinity;
39

WINTER TICKS AND DIE-OFFS OF MOOSE - SAMUEL ALCES VOL. 43. 2007
then, to assess specific factors or infiueneesthat might be used to predict epizootics ofticks and dic-ofTs of moose. In an attempt todetermine ifvarious weather parameters mightbe predictive of moose die-ofTs. I examinedparameters of snow cover and temperature atthe time of 5 die-offs in Alberta, 2 of whieh(late winter-springs of 1982 and 1999) killedmany moose. 1 also examined the feasibilityof using tick-caused damage to the winterhair coat of moose as an indication of ticknumbers asdone by Wilton and Garner ( 1993)and others.
WEATHERSeveral aspects ofweather are potentially
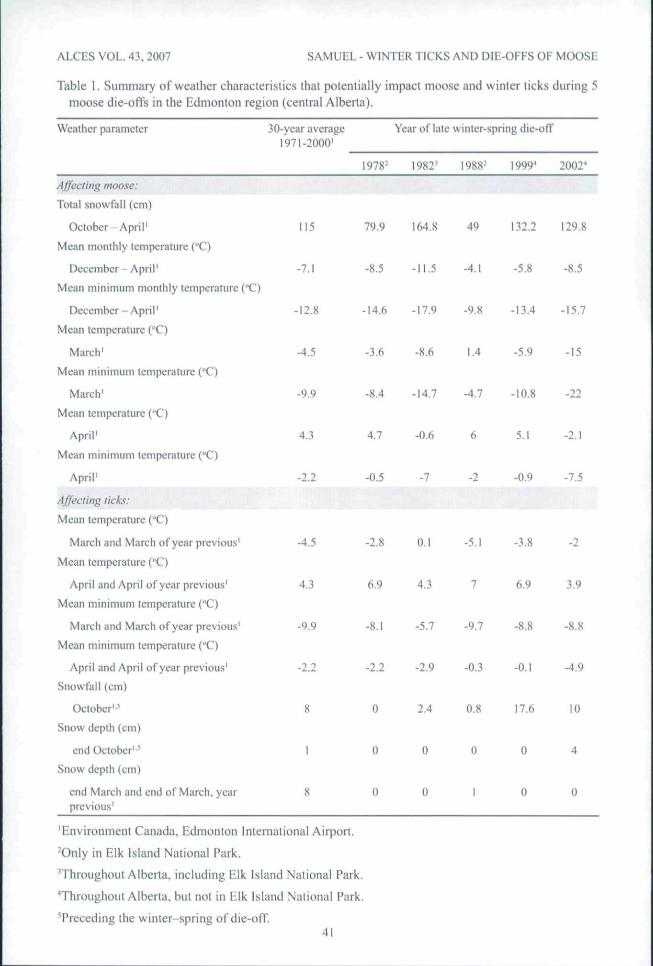
harmful to moose and ticks. For moose,depth of snow and length of severe cold, of-ten in concert with condition of habitat, canbe important (Mech et al. 1987, see logic inVuectich and Peterson 2004). Snowfall wasmuch above normal during 3 of 5 years oftick-related moose die-ofts in eentral Alberta,ineiuding 50 cm above nonnal during themajor die-off of moose in late winter-spring1982 (Table 1). The winter-spring in 4 of5 die-off years was colder ihan the 30-yearaverage (Table 1 ). In particular, March andApril 1982 and 2002, were mueh colder thanthe 30-year average.
For tieks, weather in autumn, when youngticks are on vegetation ambushing moose, andearly spring, when female ticks are droppingfrom moose to lay eggs for the next gen-eration of ticks, is critical for survival andtransmission. Significant snowfall in early-to-mid October could bury tick larvae, thuspotentially curtailing transmission to moose.That happened the week of Oetober 14, 1991at Elk Island National Park, near EdmontonAlberta, when approximately 45 cm of snowfell, burying most low vegetation (Aalangdong1994). In autumn, particularly Oetober. tieklarvae tend to quest on low vegetation andambush passing ungulate hosts (Drew andSamuel 1985). In 1991, minimum daily tem-
peratures at the park were below -]0"C thelast 10 days of Oetober. The combination ofsnow that likely buried tick larvae and eoldthat inactivated or killed larvae (Drew andSamuel 1985, Aalangdong 1994), deereasedpotential transmission of tieks to moose byalmost half in 1991 (Fig. 1). Unfortunately,it is impossible to estimate the effeet of thissnowfall on subsequent tiek numbers onmoose, because no moose were examinedfor tieks or tiek-eaused damage to the wintercoat of hair (see below). There was no die-ofïof moose in the park or other parts of centralAlberta that winter. There was also no moosedie-ofi'in winter 1991 -1992 in the Peace Rivercountry of northwest central Alberta, wheretieks often cause problems for moose (Pybus1999). There was also no major snowfall orsevere cold there in Oetober 1991.
Weather in late winter-early spring, par-ticularly related to snowfall and temperaturethe year betöre a moose die-otT, is mentionedin literature as influencing tick survival andfuture numbers of ticks (e.g., Timmermannand Whitlaw 1992, Wilton and Gamer 1993,
DATE
Fig. I. Mean daily niimberof lick larvae recoveredfrom vegetation in ilie Main Park area of ElkIsland National Park, central Alberta. Larvaewere collected by dragging l-m- white flannelcloths (599 samples in 1991. 1,556 in 1992)along transects on vegetation 1 - 1.5 m in height4 days each week at 4 sites. Transects were 2km long and divided inio eighty 25-m unitsof which 10 units were selected randomly torsampling (details in Aalangdong 1994; data fromO. Aalangdong).
4U
ALCES VOL. 43,2007 SAMUEL - WTNTER TICKS AND DIE-OFFS OF MOOSE
Table I. Summary of weather characteristics that potentially impact moose and winter ticks during 5moose die-ofts in the Edmonton region {central Alberta).
Weather parameter 30-year aventgu1971-2000'
Year of late winter-spring die-otî
1978^ 1982' 19SÍÍ- 1999^ 2002^
Affecting moose:
Total snowfall (cm)
October-April' 115
Mean monthly temperature ("C)
December - April' -7.1
Mean minimiim monthly temperature C^̂ )
December-April' -12.8
Mean temperature ("C)
Mareh' -4.5
Mean minimum temperature ("C)
March' -9.9
Mean temperature ("C)
April' 4,3
Mean minimum tcmperaUire ("C)
April' -2.2
Affecting licks: ,,,
Mean temperature ("C)
March and March of year previous' -4.5
Mean temperature ("C)
April and April of year previous' 4.3
Mean minimum temperature ("C)
March and Mareh of year previous' -9.9
Mean minimum temperature ("C)
April and April of year previous' -2.2
Snowfall (em)
October'-' 8
Snow depth (cm)
end October" 1
Snow depth (cm)
end Mareh and end of March, year 8previous'
79.9 164.8 49 132,2 129.Í
-8.5 -II.5 -4.1 -5.8 -8.5
-14.6 -17.9 -9.8 -13.4 -15.7
-3.6 -8.6 1.4 -5.9 -15
-84 -14.7 -4.7 -10.8 -22
4.7 -0.6
-0.5 -7
6,9 43
-8.1
5.1 -2.1
-2 -0.9 -7.5
-2.8 0.1 -5.1 -3.Í
6.9 3.9
-5.7 -9.7 -8.8 -8.8
-2.2 -2.9 -0.3 -0.1 -4.9
2.4 0.8 17.6 10
'Environment Canada. Edmonton Intemalioniil Airport.
^Only in Elk Island National Park.
•Throughout Alberta, including Elk Island National Park.
^Throughout Alberta, but not in Elk Island National Park.
-^Preceding the winter-spring of die-off.41

WINTER TICKS AND DIE-OFFS OF MOOSE - SAMUEL ALCES VOL. 43, 2007
DclGiudiccetal. 1997). In general, the logicis that a warm late winter-eady spring withlow preeipitation is conducive to survivaland egg-laying of female ticks that drop frommoose at that time. This results in abovenonnal numbers of young ticks on moose thefollowingautumn, possibly lcadingtodic-oifsof moose the subsequent winter. Data fromDrew and Samuel ( 1986) appear to be the gen-esis of this logic. They put blood-fed, adult,female winter tieks in outdoor, screen-wireeages at Elk Island National Park at biweeklyintervals from early Mareh to late April-midMay, 1982 and 1983, and monitored lor sur-vival. Only 11% ol" ticks (same pereentagc,both years) put in cages on snow (i.e., beforesnowmcit) survived to lay eggs, while 73 and55% of ticks put in cages on leaf litter {i.e.,after snowmelt) in 1982{- 10April)and 1983(- 24 April), respectively, survived. Drew andSamuel (1986) suggested that low survivalprior to snowmelt in spring might be due toprolonged exposure to ambient temperaturesat the snow surface that are below the tick'sthreshold for survival {- minus 17"C). Anextension of this idea is that snow depth orcrusting of snow, which forms during thefluctuating temperatures in late March andApril, prevents ticks from getting to the dufflayer, and they die before laying eggs.
Drew and Samuel ( 1989) found that blood-fed adult female ticks dropped off moose fromiate Februaiy to early May, but the peak dropperiod was late Maieh. So, what do the dataindicate for late March preeeding each ofthe 5 tick-related moose die-offs in the last30 years; i.e., did female ticks drop on snowor leaf litter? The answer is that most ticksdropped on leaf litter (see bottom. Table 1 ).However, the eontext of these results is that,even though the 30-year average amount ofsnow on Ihe ground at end March was 8.0cm, there has been no snow on the groundat the end of March in 17 of 29 (65%) years,1977-2005, in the Edmonton region. No snowtends to be "normal."
Warmer temperatures in Mareh and Aprilshould result in highcrsurvival of adult femaletieks that have dropped trom moose and. ingeneral, it was wanner In March and Aprilin years preceding die-oíTs (Table 1 ). MeanMarch and April temperatures were at orabove ihe 30-year average 4 of 5 die-ofîycars.Mean minimum temperatures in March werewanner than the 30-year average all 5 years,and minimum temperatures in April were ator above average 4 years; eonditions good fortiek survival and reproduction. In Ontario,Wilton and Gamer (1993) saw more severetick-induced hair damage and loss to moosein years following mean April temperaturesabove 3*'C', suggesting that survival of tieks wasgood above and poor below this temperature.In the Edmonton region the 30-ycar, dailymean April temperature was 4.3"C.
Thus, these data indicate that eolder-than-averagc winters in 4 of 5 die-off years,and extensive snow cover in 3 of 5 die-offyears might have adversely affected moosein central Alberta. Further, tieks might havebenefited from little orno snow on the groundand above normal temperatures during thedrop-on" period in March-April in all yearspreceding die-offs.
TICK NUMBERS ANDHAIR DAMAGE AND LOSS
In studies of moose and ticks at Elk IslandNational Park, hair loss correlated with rateof tick-induced self-grooming; i.e.. moosethat groomed more lost more hair (Mooringand Samuel 1999). ln general, annual meanpercent hair damage and loss coincided withannual mean number ofwinter tieks on moose(Sanuicl 2004) indicating that grooming wasrelated to the number of ticks on moose. In a10-year study in Algonquin Provincial Park,Ontario, damage to the winter hair coat ofmoose was used as an index of tick numberson moose. Years with high numbers of moosecarcasses generally coincided with yearswhen hair damage was highest and vice versa
42

ALCES VOL. 43, 2007 SAMUEL - WINTER TICKS AND DIE-OFFS OF MOOSE
(Gamerand Wilton 1993, Wilton and Gamer1993). However, a manager has few optionsto estimate trends in tick numbers. Estimat-ing numbers of ticks on a sedated live mooseis not reliable beeause there ean be manythousands of tieks on this large mammal witha thick winter eoat of hair, making countingimpossible (personal observation). Countingticks on dead moose is not an option becausethe digestion technique used to recover ticks,though accurate (Welch and Samuel 1989) islaborious.
An important management issue iswhether data on annual hair damage/loss eanprovide an early warning of impending die-offs of moose. The answer might hinge onthe accuracy of teehniques used during aerialand ground surveys ofmoose to record tick-induced hair damage and loss. Hair damageand loss on moose ean be quantified by groundor aerial survey in 2 ways; ( I ) use a digitizerto detemiine the percentage of the lateralsilhouette of the torso with hair damage orloss from photographs or diagrams of lateralviews ofmoose (see description of techniquein Welch et al. 1990, and Samuel and Welch1991); or (2) group moose subjectively intoseveral eategorics of hair-loss severity (seephotographs of categories in Samuel 1989,2004). The first method takes more time be-cause it demands either taking photographs,or making diagrams of hair damage of thelateral torso ofmoose. With some experienceit is relatively easy to tise the second methodand assign moose to 1 of 5 ealegories of hairdamage or loss; no damage to hair eoat, slight(wliieh is approximately 5 - 20%of wintcrhairbroken or lost), moderate (~ 20-40%), severe(~ 40 - 80%), and ghost moose (> 80%).
Because grooming against ticks, and theresulting hair damage and loss, continues tolate April (Welch ctal. 1990). it is best to dosurveys for hair damage as late in the seasonas possible. Unfortunately, moose managersmust do their surveys well before mid-Apriltaking advantage ofsnow and moose behavior.
However, annual surveys done earlier, prefer-ably late Febmary, ean provide comparativetrends in hairdamage/loss, as long as they aredone at approximately the same time each year(see Welch et al. 1990 and Samuel and Welch1991 for temporal pattems ofthe progressionof hair loss).
Hairdamage and lossshould coincide withtick numbers given that grooming against ticksby moose is proportional to tick bite; i.e.. themore ticks present, the more blood-feedingby ticks, whieh results in more groomingand subsequent hair damaged or lost (Moor-ing and Samuel 1998). Wilton and Garner(1993) calculated a hair-loss severity index(HSI) to summarize category-type data (Fig.2). The two methods, mean % hair loss andHSI, were compared using hair-loss data formoose from Elk Island National Park (Fig.2). Yearly changes were parallel, in general,partieularlysoafterthemajordie-oiTof moosein late winter-spring of 1982. The same wastme tbryearly change in HSI and tick numbers(Fig. 3) and yearly change in mean % damageto the winter hair coat and mean nutnber ofticks (Fig. 4).
In summary, anntial surveys oftick-causeddamage to winterhairof moose, done as late aspossible in the winter-spring season, providegood indication of numbers of ticks on moose.Unfortunately, more repetitive annual surveysare needed to detennine if hair loss surveys areuseful to prediet moose population response insubsequent years (but see next paragraph ondata currently being collected at Isle RoyaleNational Park).
NUMBERS OF MOOSESamuel (2004) summarized a cascade of
events, starting with increasing moose num-bers that led to lethargic, ill, or dead moose.The eascadc goes as follows: moose numbersincrease in a local area; tick numbers inerease;tick bite increases, which eauses more itehingand grooming; hairdaniage and loss increases;energy expended to grooming iiicreascs; blood
43

WINTER TICKS AND DIE-OFFS OF MOOSE - SAMUEL ALCES VOL. 43. 2007
50
Fig. 2. Comparison of mean % hair damage/lossand hair-loss severity index, two methods used lodetennine yearly changes in lick-caused damageto the winter hair of moose in Elk Island NationalPark. Categories of hair damage on 327 moose(variable numbers per year), observed from theair and ground, were used to calculate a Hair-lossSeverity Index (HSl). Moose in categories weregrouped subjectively as follows: no damage tohair coat of lateral torso (HSl class value = 1 );slight damage (~ 5 - 20% of winter hair brokenorlost)(HSl-2); modérale damage(~ 20-40%broken/lost) (HSl = 3); severe damage (- 40 -80% broken/lost) (HSl = 4); and ghost moose(> 80% broken/lost) (HSl = 5). The frequencyof moose in each calegoiy was multiplied by theappropriate class value and the sum divided bythe total number of moose sampled for that timeperiod. The second method involved diagramsof the hair damage on the lateral torso of 302moose (variable numbers per year), surveyedfrom the air and ground, that were digitizedfor hair damage using methods in Samuel andWelch (1991).
loss increases; appetite and feeding by moosepossibly suppressed; smaller energy reservesavai lable for blood replacement and to supporthigh rates of grooming; and increased cost ofreplacing heat energy lost through a darnagedhair coat. AU of this results in léthargie, ill,or dead moose. This easeade assumes thatnumbers of ticks track changes in moosenumbers. Unfortunately, long-term data setsto test this assumption are scant because of thedifficulty in monitoring a moose populationfor both moose numbers and tick numbers onan annual basis for many years; 12 years datawere eolleeted at Elk Island National Park
(Fig. 5). Currently, similar data are collectedonly at Isle Royale National Park where hairloss is monitored as an index of tick numbers(Delguidice et al. 1997, Peterson and Vueetich2006).
Samuel and eolleagues (see Samuel 2004)studied winter ticks in a closed system atElk Island National Park in central Alberta,1978-1996. The park is 195 km- in size, andsurrounded by fences that prevent the largeungulates from leaving. It is dotted with nu-merous wetlands with aspen (PÎ;/ÏW/Î/.V spp.) thedominant tree. Coyotes (CÍ/H/.V/CÍÍ/WÍ.S) are theonly resident large carnivore. The dynamicsofthe local ungulates were described by Blythand Hudson (1987) atid Blyth (1995). Somemoose die-offs in the park were concurrentwith die-offs of moose populations acrosscentral Alberta (Table 1 ), most reeently in latewinter-spring 1982, when tick-related die-offs occurred throughout Alberta. However,there was no major tick-mediated die-off inthe park in 1999 (N. L. Cool, Parks Canada,Elk Island National Park, personal commu-tiication) when the province was experienc-ing major losses everywhere (Pybus 1999).Similarly, though die-otïs occurred in localpopulations throughout much of Alberta in2002, including central Alberta, there was no
70000
10000
Fig. 3. Annual changes in mean number of wintertickscollected from digested hides and Hair-lossSeverity Index (HSl) fornioose trom Elk IslandNational Park. Hides of 118 moose (variableluimbers per year) were digested and ticks col-lected tising techniques in Welch and Samuel(1989). See Figure 2 for infomiation on HSl.
44

ALCES VOL. 43, 2007 SAMUEL - WINTER TICKS AND DIE-OFFS OF MOOSE
70000
Op 55000
40000 -
25000
10000
Fig. 4. Antuial changes in mean nutnber of tickscollected from digested hides and extent of hairdamage and loss in spring on moose, from ElkIsland National Park. See Figures 2 and 3 forrelevant infonnation. Figure modified fromSamuel (2004).
die-off in Elk Island National Park, thottgh adic-oiï of about 150 moose occurred the nextyear in the Main Park area (N. L. Cool, ParksCanada, Elk Island National Park, personalcommunication).
Mooseatid tick numbers (and hairdamageand loss, Fig. 4) were monitored in the northempart ofthe park {Maiti Park area, 136 km-) in1978-1990. Jn general, there was a lag of 1year in tick numbers traeking moose numbers(Fig. 5). Die-offsoceurred when moose densi-ties approached 3 moose per km- and meannufnbers of ticks on moose approaehed 50,000- 60,000. Data for estitiiates of numbers ofmoose dying iti sotne years are somewhatmisleading beeause various forms of moosemanagement were itnplemenled when tnoosedensities were high. For example, the parkeuUed tnoose in some yeat^(BIyth and Hudsoti1987, BIyth 1995). and culls are iticluded inthe annual declines in tnoose density (Fig. 5).The tuimber of eulled moose in Main Park,where the tiek researeh was done, cannot beseparated from the estimates of moose dyingnaturally.
Culls were done in December 1977 and1980 in an attempt to balance the moosepopulation with estitnated food rcsotirees.Nonetheless, Main Park suffered a tuimberof natural, tiek-mediated losses, including anestimated 100 tnoose in winter 1977 (Satiiuel
and Barker J979), "extreme levels oftnortal-ity" (Blyth atid Hudson 1987) iti winter 1982,and smaller natural losses of moose in 1978,1984.1988.and 1989(BIyth 1995). Chemicalreptoductivc inhibitioti was administered to59 adult females in Main Park in 1987, almost25%ofthe mature eows(BIyth 1995), and 108moose were shot from 1978-1990 for researehon winter tieks.
Pybus (1999) summarized 1,130 oceur-renees involving moose in trouble or founddead (H = 311) during a winter (1999) ofsignificant moose moi1a!ity inAlberta. Manyreports («= 1,035) itwolvcdwititer tick-causedhair loss and itidicated increasing oeeur-rence with increasing density of moose. Insummary, in spite ofa mixture of somewhatsubjective data sets, there appears to be ahost-density component to tick numbers onmoose. However, more reliable long-tenn dataand objeetive studies arc needed to aseertainthis relationship,
VEGETATION STRUCTUREWhere a blood-fed, adult female winter
tiek drops trom a tnoose in late winter-earlyspritig, is where she must survive and produceoffspring. Drew and Satnuel (1986), Aa-Iangdotig(l 994), atid Aalangdong etal. (2001)found that different habitats iti Elk IslandPark,withdifferentmieroelimatieeonditions,
80000
0.5
Fig. 5. Estimateci density oí iiioose (late fall, priorto itnplenientation of management options; seetext) and mean number of ticks collected fromdigested hides in Main Park area of Elk IslandNational Park. Figure modifted from Samuel(2004).
45

WINTER TICKS AND DIE-OFFS OF MOOSE - SAMUEL ALCES VOL. 43. 2007
influence survival and reproduction of winterticks. Aalangdong et al. (2001) put ticks ingauze bags - 2 cm beneath the litter in varioushabitat types in Elk Island National Park. AH3 habitat types with open canopies were moresuitable tbrwintcrtick survival and productionof larvae than the 4 habitat types with closedcanopies. Thus, more ticks survived to produceeggs, and more eggs hatched to larvae, andthe larvae survived longer, in habitat typeswith open canopies. The authors attribtitedthis to mean monthly temperatures at groundlevel, which in summer were several degreeslower in all 4 'elosed' habitats, than in 'open'habitats. These restilts might have broaderimplications and help explain why moose inparts of Alberta with open canopies, sueh asthe aspen-rich Peace River country of north-west central Alberta, and the aspen parklandsand southern edges of the boreal mixed woodforests of central Alberta, appear to sutlermore trom winter ticks than moose in moretiortherly spruce-dominated forests.
SUMMARYDie-otTs of moose are complex events
that arc probably mediated by several factorsincluding winter weather, habitat conditions,detisity of moose, and winter ticks. The factthat some tick-associated declines in moose areconcurrent in many populations in a ntmiberof areas suggests an "extrinsic influence" isinvoIved(Dclguidiecetal. 1997). Thehypoth-esis that weather in the form of shorter, wannerwinters with less preeipitation than usualresults in more ticks ihe following year wasgenerally confinncd. Intuitively, the scenariomakes sense, but it is probably more com-plex than presented here. As Holmes (1995)states, "finding simple, consistent causes forcomplex biological phenomena" is wishfulthinking. More rigorous analysis of climaticvariables important to winter ticks in regionswith moose dic-ofTs (Deiguidiec ct al. 1997,Vucctich and Peterson 2004) is needed, alongwith more long-term study of moose and tick
numbers via monitoring of damage and loss towinter hair coat of moose; such monitoring isnow done only at Isle Royale National Park.Assuming future climate change, minimumwinter temperatures at high northern latitudesare expected to rise more than temperaturesin other seasons (Intergovernmental Goverti-mental Panel on Climate Change 1996). Thiswill most certainly impact late winter-springsurvival and development of off-host stagesof ticks (Lindgren ct al. 2000), in this ease,winter ticks.
ACKNOWLEDGEMENTSThe long-term contribution of many
colleagues at Elk Island National Park isappreeiatcd. The Natural Seiences and En-gineering Research Council of Canada sup-ported much of the work done in Elk IslandNational Park.
REFERENCESAALANGDONG,0.1. 1994. Wintertiek(Dc/-ffií7-
cetitor albipictus) ecology and transmis-sion in Elk Island National Park, Alberta.M.Se. Thesis, Department of BiologicalSciences, University of Alberta, Edmon-ton, Alberta, Canada.
,̂ W. M. SAMUFL, and A. W. SHOSTAK.
2001. Off-host survival and reproduc-tive success of adult female winter ticks,Dermacentor albipictus in seven habitattypes of Central Alberta. Journal of theGhana Science Association 3:109-116.
BLYTH, C. B. 1995. Dynamics of ungulatepopulations in Elk Island National Park.M.Sc. Thesis. Department of Agricultural,Food and Nutritional Science, Universityof Alberta, Edmonton, Alberta, Canada.
^ and R. J. HUDSON. 1987. A plan forthe management of vegetation and un-gulates. Elk Island National Park. ElkIsland National Park and Department ofAnimal Science (now, Agricultural, Foodand Ntitritional Science), University ofAlbeila, Edmonton, Alberta, Canada.
46

ALCES VOL. 43,2007 SAMUEL - WINTER TICKS AND DIE-OFFS OF MOOSE
DELGUIDICH, G. D., R. O. PETERSON, and W.
M. SAMUHL. 1997. Tretids ofwititcrnu-tritional restriction, ticks, and numbers ofmooseon Isle Royale. Jottmal of WildlifeManagement 61:895-9O3.
DiíFAV,M.L.. and W.M.SAMUEL. 1985. Fac-tors affecting Iransmission of larval winterticks, Dermacentoralhipictus (Packard),to moose, Alces alces (L.), in Albetia.Canada. Jotimal of Wildlife Diseases21:274-282.
and . 1986. Reproduction ofthewintertick, of rmaí:í:'í7/w¿///í//?/íV//,v, underfield eonditions in Alberta, Canada. Cana-dian Joumal of Zoology 64:714-721.
and . 1989. Instar developtnent
and . 1999. Prematurcwinterhair
and disengagetncnt rate of engorged TQ-ma[cwlnlCTÚcks,Denmicentora¡hÍpictus(Acari: Ixodidae), following sitigle- andtriekle-exposure of tnoose (Alces alces).Experimental and Applied Aearology6:189-196.
GARNHR, D. L., and M. L. WILTON. 1993. The
potential role ofwinter tiek (Dermacen-loralhipictus) in the dynamics of a southcentral Ontario moose population. Alces29:169-173.
HoLM!-s,J.C. 1995. Population regulation: adynamic complex of interaetions. WildlifeReseareh 22:11-19.
LiNDGREN, E., L. TALLEKLINT, and T. POLFELDT.
2000. Impaet of climatic change on thenothern latilude limit and populationdensity of the disease-transmitting Euro-pean tick Ixodes ricinus. EnvironmentalHealth Perspeelives 108:119-123.
Man, L. D., R. E. MCRORERTS, R. O. PETERSON,
and R. E. PAGE. 1987. Relationship ofdeer and moose populations to previouswinters' snow. Joumal ofAnitnal Eeology56:615-627.
MOORING, M. S., and W. M. SAMUEL. 1998.
The biological basis of grooming inmoose (Alces alces): programmed ver-sus stitnulus-driven grooming. AnimalBehaviour 56:1561-1570.
loss in free-ranging moose {Alces alces)infested with winter ticks (Dennacentoralbipictus) is correlated with groom-ing rate. Canadian Joumal of Zoology77:148-156.
PETERSON, R. O., and J. A. VUCETICH. 2003.Ecological studies of wolves on Isle Roy-ale. Annual Report 2002-2003. MichiganTechnological University, Houghton,Michigan, USA.
and . 2006. Ecological studiesof wolves on Isle Royale. Annual Report2005-2006. MiehiganTechnological Uni-versity, Houghton, Michigan, USA.
PYBUS, M.J. 1999. Mooseand tieks in Alberta:adieotïin 1998/99. Occasional Paper No.20. Fisheries and Wildlife ManagementDivision, Edtnonton, Alberta, Canada.
SAMUEL, B. 2004. White as a Ghost: WinterTieks and Moose. Natural Hisloiy Series,Volume 1, Federation of Alberta Natural-ists, Edmonton, Alberta, Canada.
andV.CRICHTON. 2003. Winterticksandwinter-spring los.ses of tnoose in vvesteniCatiada. TheMooseCall 16:15-16.
SAMUEL, W.M. 1989. Loeationsofmoose innorthwestem Canada with hair loss prob-ably eaused by the winter tick, Dernuicen-tor alhipictus {Acari: Ixodidae). Joumalof Wildlife Diseases 25:436-439.
and M. BARKER. 1979. The wintertick, Dermacentor alhipictus (Packard,1869), on tnoose, Alces alces (L.), ofcentral Alberta. Proceedings ofthe NorthAmerican Moose Conference and Work-shop 15:303-348.
, . and T. LEIGHTON. 2002. Highmoose mortality from winter tick - spring2002. Canadian Cooperative WildlifeHealth Centre Newsletter 9:9.
. M. S. MOORING, andO. 1. AALANGDONG.
2000. Adaptations ofwinter tieks (Der-macentor alhipictus) to invade mooseand moose to evade ticks. Alces 36:183-195.
47

WINTER TICKS AND DIE-OFFS OF MOOSE - SAMUEL ALCES VOL. 43, 2007
and D. A. WELCH. 1991. Winter tickson moose and other ungulates: factorsinfluencing their population size. Alces27:169-182.
TiMMHRMANM, H. R., and H. A. WHITLAW.
1992. Selective moose harvest in NorthCentral Ontario-a progress report. Alces28:1-7.
VucETiCH, J. A., and R. O. PFTRRSON. 2004.The influence of top-down, bottom-up,and abiotic factors on the moose (Ak-esalces) population of Isle Royale. Pro-ceedings of the Royal Society of London.Scries B, 271:183-189.
WF.LCH,D.A.,andW.M.SAMur:L. 1989. Evalu-ation of random sampling for estimatingdensity of winter ticks (Dermacenioralbipictus) on moose {Alces alces) hides.International Journal of Parasitology19:691-693.
, , and R.J.HUDSON. 1990. Bio-
energetic consequences of tick-indueedalopecia on moose. Journal of MedicalEntomology 27:656-660.
WILTON, M, L., and D. L. GARNER. 1993.
Preliminary observations regarding meanApril temperature as a possible prcdie-tor of tick-induced hair-loss on moosein south central Ontario, Canada. Alces29:197-200.
48
