fatty acid metabolism
DESCRIPTION
FATTY ACID METABOLISM. FATTY ACID METABOLISM. A fatty acid is a carboxylic acid. often with a long unbranched aliphatic chain, which is either saturated or unsaturated. Carboxylic acids as short as butyric acid (4 carbon atoms) are considered to be fatty acids, - PowerPoint PPT PresentationTRANSCRIPT

FATTY ACID METABOLISM
1

2
FATTY ACID METABOLISM
A fatty acid is a carboxylic acid. often with a long unbranched aliphatic chain, which is either saturated or unsaturated. Carboxylic acids as short as butyric acid (4 carbon
atoms) are considered to be fatty acids,Fatty acids are produced by the hydrolysis of the ester linkages in a fat or biological oil (both of which are triglycerides), with the removal of glycerol.The most abundant natural fatty acids have an even number of carbon atoms.

3
Major Physiological Roles oF Fatty Acids
1. Fatty Acids (FAs) are building blocks of phospholipids and glycolipids.
2. Many proteins are modified by the covalent attachment of FAs, which targets them to membrane locations.
3. FAs are fuel molecules that are stored as triacylglycerols (or triglycerides) (TGs).
4. FAs derivatives serve as hormones and intracellular messengers.

4
In mammals, the major site of accumulation of triacylglycerols is the cytoplasm of adipose cells (fat cells)

5
• Fatty acid degradation and synthesis are relatively simple processes that are essentially the reverse of each other.

6
TGs Are Highly Concentrated Energy Stores
• 9 Kcal/g VS 4 Kcal/g for protein/carbohydrates.• Anhydrous storage in adipose VS hydrated glycogen in
liver/muscle• A typical 70kg man would have fuel reserves:
100,000 kcal in TGs, 25,000 kcal in protein (mostly in muscle), 600 kcal in glycogen 40 kcal in glucose.
• TGs constitute about 11 kg of his total body weight.• If the 11kg TGs to be stored in Glycogen form, his total
body weight would be 55kg greater (11+55 kg) + 59.• Storing energy in TGs saves 55Kg of body weight.• Glycogen & Glucose provide energy for short-term (a day
or so), while TGs support the body with energy for several weeks.

7
Dietary Lipids Are Digested by Pancreatic Lipases
• TGs in the intestinal lumen are incorporated into micelles formed with the aid of bile salts (e.g. glycocholate)
• Amphipathic molecules synthesized from cholesterol in the liver and secreted from the gall bladder.
8
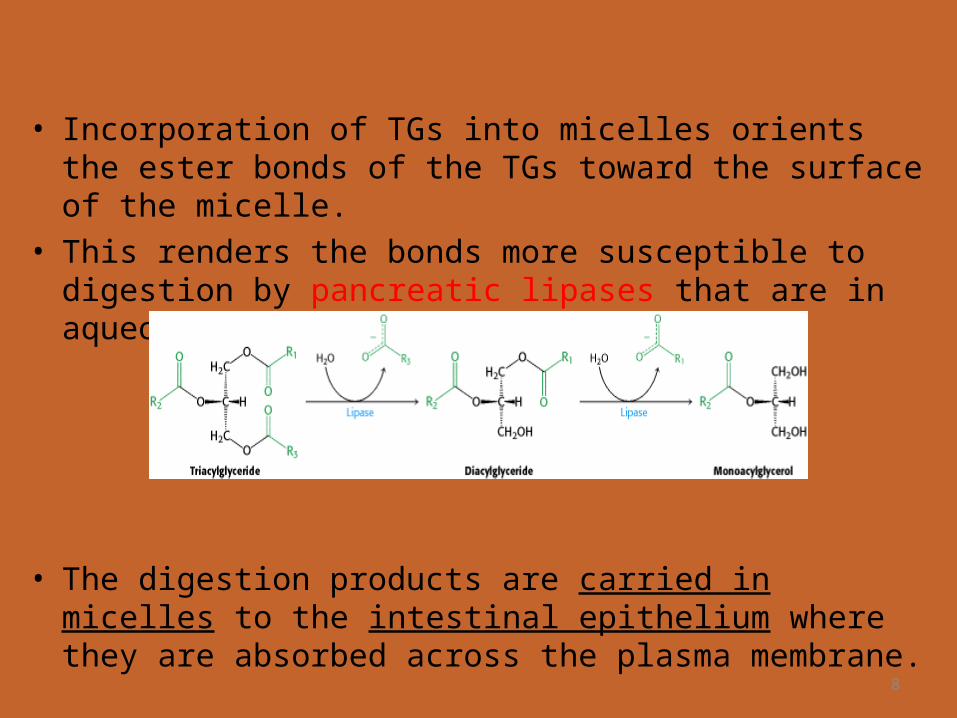
• Incorporation of TGs into micelles orients the ester bonds of the TGs toward the surface of the micelle.
• This renders the bonds more susceptible to digestion by pancreatic lipases that are in aqueous solution.
• The digestion products are carried in micelles to the intestinal epithelium where they are absorbed across the plasma membrane.

9
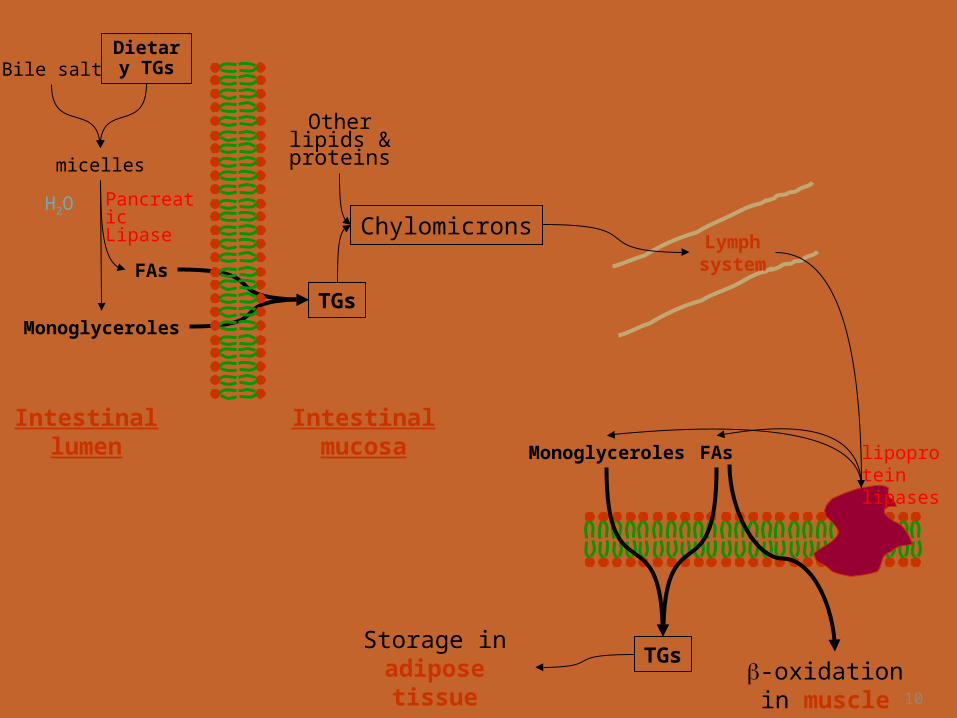
Dietary Lipids Are Transported in Chylomicrons
• In the intestinal mucosal cells, the TGs are re-synthesized from FAs and monoacylglycerols.
• Then packaged into chylomicrons.
10
Dietary TGsBile salt
micelles
Pancreatic Lipase
FAs
MonoglycerolesTGs
H2OChylomicrons
Other lipids & proteins
Lymph system
lipoprotein lipases
FAsMonoglyceroles
TGsStorage in adipose
tissue -oxidation in muscle
Intestinal lumen
Intestinal mucosa

11
The Utilization of Fatty Acids as Fuel Requires Three Stages of Processing
1. The lipids must be mobilized:– TGs are degraded to FAs and glycerol– They are released from the adipose tissue and transported
to the energy-requiring tissues. 2. At these tissues, the fatty acids must be activated and
transported into mitochondria for degradation.3. The fatty acids are broken down step-by-step into
acetyl CoA, which is then processed in the citric acid cycle.

12
1. Triacylglycerols Are Hydrolyzed by Cyclic AMP-Regulated Lipases
epinephrine, norepinephrine, glucagon, adrenocorticotropic hormone

13
Glycerol formed by lipolysis is absorbed by the liver where it enters glycolysis or gluconeogenesis.
phosphatase

14
2. Fatty Acids Are Linked to CoA Before They Are Oxidized
• Fatty acids are oxidized in mitochondria.• They are activated before they enter the mitochondrial
matrix.• ATP drives the formation of a thioester linkage between
the carboxyl group of a FA and the sulfhydryl group of CoA.
• This activation reaction takes place on the outer mitochondrial membrane, where it is catalyzed by acyl CoA synthase (also called fatty acid thiokinase).
15
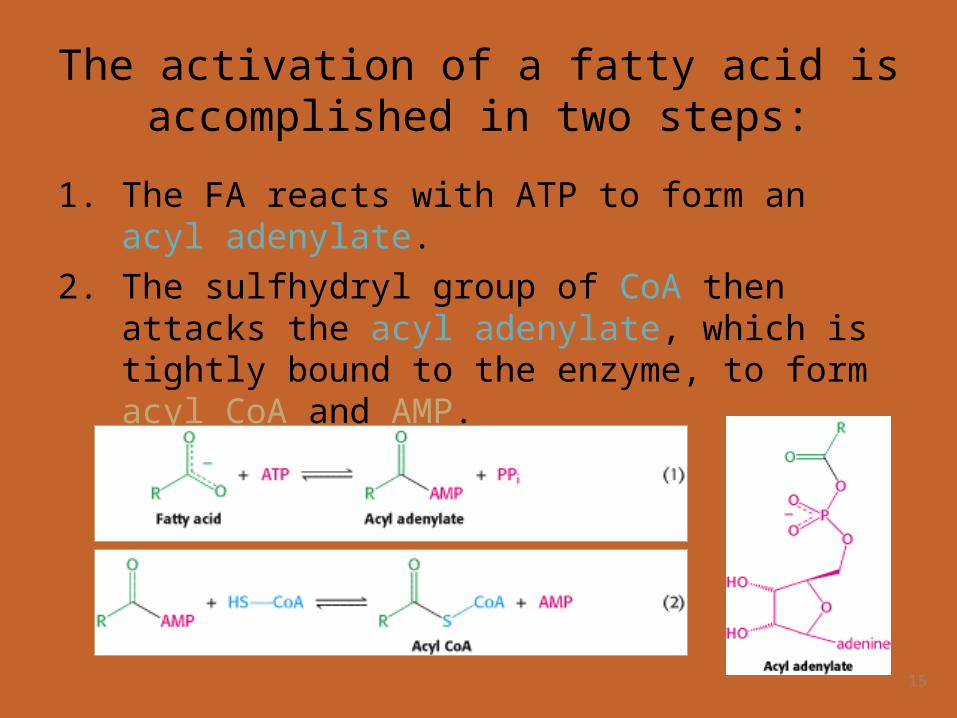
The activation of a fatty acid is accomplished in two steps:
1. The FA reacts with ATP to form an acyl adenylate.2. The sulfhydryl group of CoA then attacks the acyl
adenylate, which is tightly bound to the enzyme, to form acyl CoA and AMP.

16
• These partial reactions are freely reversible• One high-transfer-potential compound is cleaved (between
PPi and AMP) and one is formed (the thioester acyl CoA).• How is the overall reaction driven forward?• The answer is that PPi is rapidly hydrolyzed by a
pyrophosphatase• The equivalent of two molecules of ATP are consumed.
17
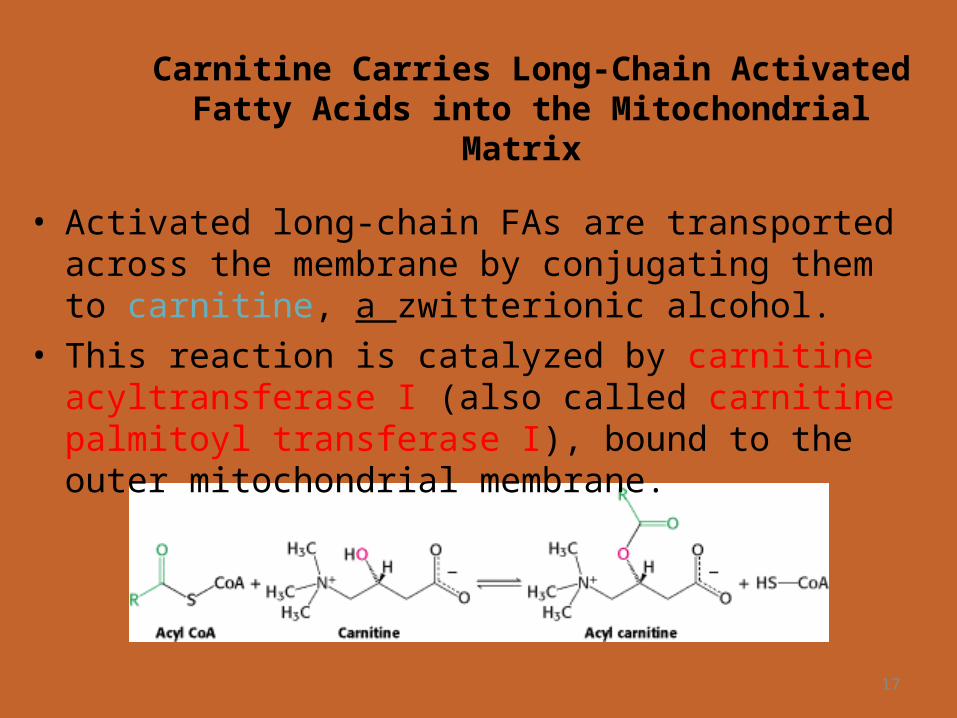
Carnitine Carries Long-Chain Activated Fatty Acids into the Mitochondrial Matrix
• Activated long-chain FAs are transported across the membrane by conjugating them to carnitine, a zwitterionic alcohol.
• This reaction is catalyzed by carnitine acyltransferase I (also called carnitine palmitoyl transferase I), bound to the outer mitochondrial membrane.

18
• Acyl carnitine is shuttled across the inner mitochondrial membrane by a translocase
• The acyl group is transferred back to CoA on the matrix side of the membrane.
• This reaction, which is catalyzed by carnitine acyltransferase II (carnitine palmitoyl transferase II), is simply the reverse of the reaction that takes place in the cytosol.

19
• Normally, the transfer of an acyl group from an alcohol to a sulfhydryl group is thermodynamically unfavorable.
• However equilibrium constant for this reaction for carnitine is near 1.
• Apparently because carnatine and its esters are solvated differently from most other alcohols and their esters due to the zwitterionic nature of carnitine.
• As a result, the O-acyl link in carnitine has a high group-transfer potential.
• Finally, the translocase returns carnitine to the cytosolic side in exchange for an incoming acyl carnitine.

20
3. Fatty Acid Oxidation
• Through a sequence of 4 reactions:1. Oxidation by FAD2. Hydration by H2O
3. Oxidation by NAD+4. Thiolysis by CoA

21
1. oxidation of acyl CoA by an acyl CoA dehydrogenase to give an enoyl CoA with a trans C2=C3.
electron-transferring flavoprotein
ETF:ubiquinone reductase, an iron-sulfur protein

22
2. Stereospecific hydration of the C2=C3 by enoyl CoA hydratase.

23
3. A second oxidation reaction converts the hydroxyl group at C3 into a keto group and generates NADH.
This oxidation is catalyzed by L-3-hydroxyacyl CoA dehydrogenase, which is specific for the L isomer of the hydroxyacyl substrate

24
4. Thiolysis of 3-ketoacyl CoA by the thiol group of a second molecule of CoA yields acetyl CoA and an acyl CoA shortened by two carbon atoms.
This thiolytic cleavage is catalyzed by -ketothiolase.

25
• The shortened acyl CoA then undergoes another cycle of oxidation, starting with the reaction catalyzed by acyl CoA dehydrogenase

26
-Ketothiolase (also called thiolase)3-ketoacyl CoA + CoA acetyl CoA + acyl CoA (shortened by C2)
6
L-3-Hydroxyacyl CoA dehydrogenaseL-3-Hydroxyacyl CoA + NAD+
3-ketoacyl CoA + NADH + H+ 5
Enoyl CoA hydratase (also called crotonase or 3-hydroxyacyl CoA hydrolyase)
trans-2 -Enoyl CoA +H2OL-3-hydroxyacyl CoA
4
Acyl CoA dehydrogenases (several isozymes having different chain-length specificity)
Acyl CoA + E-FAD trans- 2 -enoyl CoA + E-FADH2
3
Carnitine acyltransferase (also called carnitine palmitoyl transferase)
Carnitine + acyl CoA acyl carnitine + CoA
2
Acyl CoA synthetase [also called fatty acid thiokinase and fatty acid:CoA ligase (AMP)]
Fatty acid + CoA + ATP acyl CoA + AMP + PPi
1
EnzymeReactionStep

27
The Complete Oxidation of Palmitate Yields 106 Molecules of ATP
• The degradation of palmitoyl CoA (C16-acyl CoA) requires seven reaction cycles.
• In seventh cycle, C4-ketoacyl CoA is thiolyzed to two molecules of acetyl CoA.
• Hence, the stoichiometry of oxidation of palmitoyl CoA is:

28
• Approximately 2.5 of ATP are generated per each of the 7 molecules of NADH.
• 1.5 of ATP are formed for each of the 7 molecules of FADH2.• Recall that the oxidation of acetyl CoA by the citric acid cycle
yields 10 molecules of ATP.• Hence, the number of ATP molecules formed in the oxidation
of palmitoyl CoA is:– 10.5 from the 7 molecules of FADH2– 17.5 from the 7 molecules of NADH– 80 from the 8 molecules of acetyl CoA– ~2 ATP are consumed in the activation of palmitate
• The total is 106 ATP.

29
Certain Fatty Acids Require Additional Steps for Degradation
1.The oxidation of unsaturated fatty acids requires additional steps.
2.Fatty acids containing an odd number of carbon atoms yield a propionyl CoA at the final thiolysis step.
3.Must be converted into an easily usable form by additional enzyme reactions.

30
1. Unsaturated fatty acids
• Palmitoleate, C16 unsaturated fatty acid has one double bond between C-9 and C-10, (9).
• It is activated and transported across the inner mitochondrial membrane in the same way as saturated fatty acids.
• Palmitoleoyl CoA then undergoes 3 cycles of degradation; carried out by the same enzymes as in saturated ones.
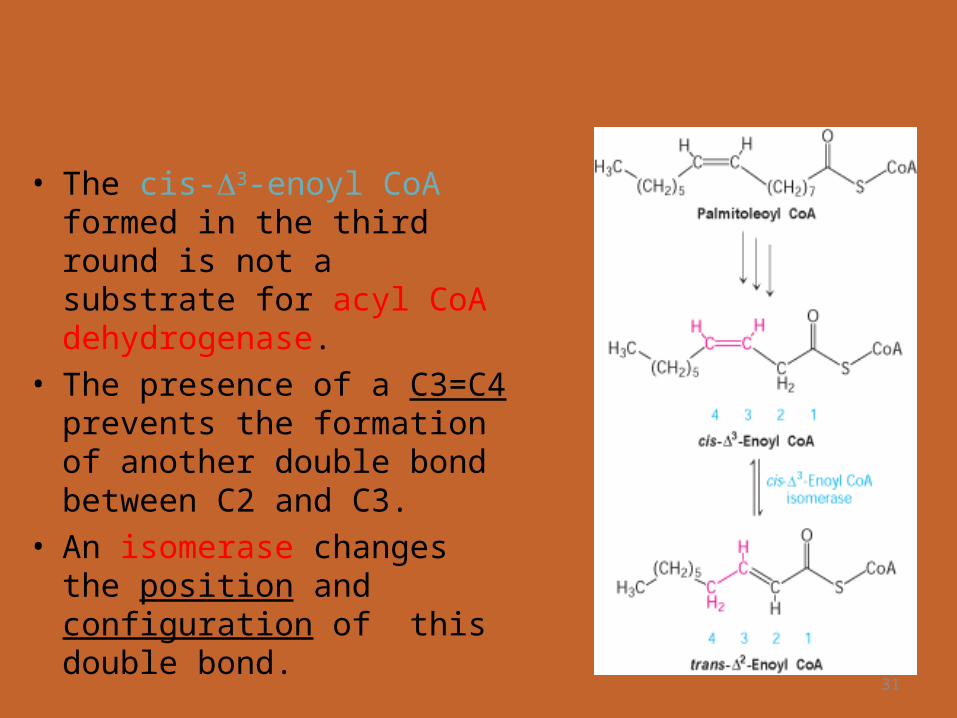
31
• The cis-3-enoyl CoA formed in the third round is not a substrate for acyl CoA dehydrogenase.
• The presence of a C3=C4 prevents the formation of another double bond between C2 and C3.
• An isomerase changes the position and configuration of this double bond.
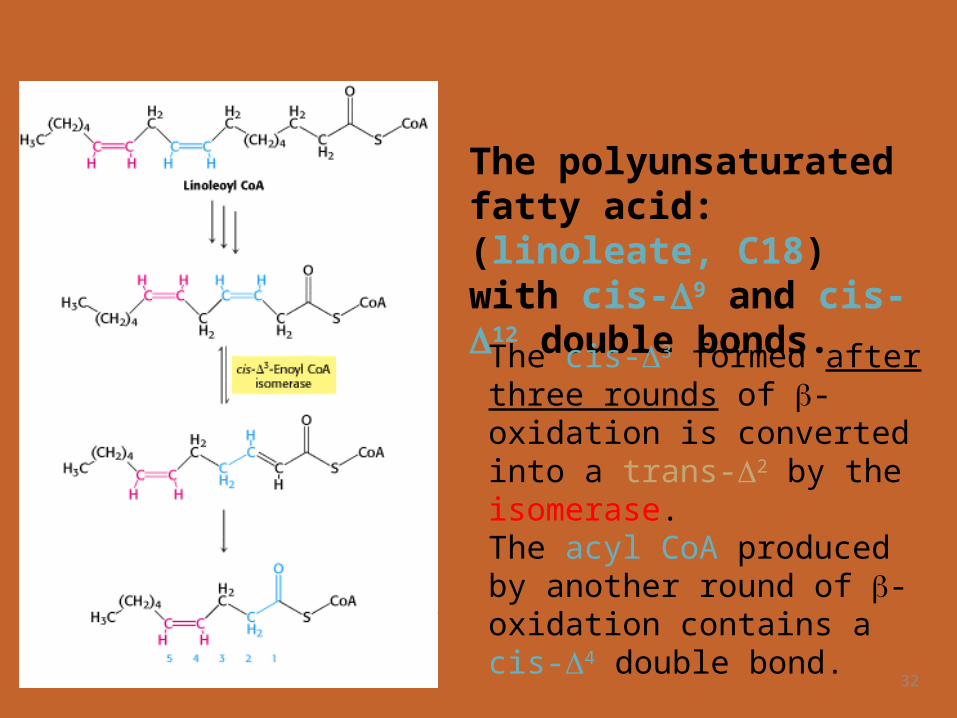
32
The polyunsaturated fatty acid: (linoleate, C18) with cis-9 and cis-12 double bonds.
The cis-3 formed after three rounds of -oxidation is converted into a trans-2 by the isomerase.
The acyl CoA produced by another round of -oxidation contains a cis-4 double bond.

33
2,4-dienoyl CoA reductase, uses NADPH to reduce the 2,4-dienoyl intermediate to trans-3-enoyl CoA.Cis-3-Enoyl CoA isomerase then converts trans-3-enoyl CoA into the trans-2 form, a customary intermediate in the -oxidation pathway
Dehydrogenation of this species by acyl CoA dehydrogenase yields a 2,4-dienoyl intermediate, which is not a substrate for the next enzyme in the -oxidation pathway.

34
• Only two extra enzymes needed for oxidation of any polyunsaturated fatty acid:– Odd-numbered double bonds are handled by the
isomerase.– Even-numbered ones by both reductase and
isomerase.

35
Odd-Chain Fatty Acids Yield Propionyl Coenzyme A in the Final
Thiolysis Step
• Fatty acids having an odd number of carbon atoms are minor species.
• They’re oxidized in same way as those having an even number of C.
• Propionyl CoA and acetyl CoA, rather than two molecules of acetyl CoA, are produced in the final round of degradation.

36
• Propionyl CoA is carboxylated by propionyl CoA carboxylase yielding D-isomer of methylmalonyl CoA at the expense of ATP hydrolysis.– This enzyme is a biotin enzyme that is homologous to
and has a catalytic mechanism like that of pyruvate carboxylase.
• The D-isomer is racemized into the L-isomer.• The L-isomer of methylmalonyl CoA is converted by a mutase into
succinyl CoA – The (-CO-S-CoA) group migrates from C2 to C3 in
exchange for a hydrogen atom.

37
Fatty Acids Are Also Oxidized in Peroxisomes
• Some oxidation takes place in cellular organelles called peroxisomes that have high concentrations of catalase.
• Fatty acid oxidation in these organelles, stops at octanyl CoA.
• It may serve to shorten long chains to make them better substrates of oxidation in mitochondria.

38
• Peroxisomal oxidation differs from oxidation in the initial dehydrogenation reaction.– In peroxisomes, a flavoprotein dehydrogenase transfers electrons to O2 to
yield H2O2
– Catalase is needed to convert the H2O2 produced in the initial reaction into H2O and O2 .
• Subsequent steps are identical with their mitochondrial counterparts, although they are carried out by different isoforms of the enzymes.

39
Fats burn in the flame of carbohydrates
• The entry of acetyl CoA into the citric acid cycle depends on the availability of oxaloacetate for the formation of citrate.
• Oxaloacetate is normally formed from pyruvate, the product of glycolysis, by pyruvate carboxylase
• The concentration of oxaloacetate is lowered if carbohydrate is unavailable or improperly utilized.

40
In fasting or diabetes
• Oxaloacetate is consumed to form glucose by the gluconeogenesis.
• Under these conditions, acetyl CoA is diverted to the formation of acetoacetate and D-3-hydroxybutyrate.
• Acetoacetate, D-3-hydroxybutyrate, and acetone are often referred to as ketone bodies.

41
Thiolase
HydroxymethylglutarylCoA synthase
HydroxymethylglutarylCoA cleavage enzyme
D-3-Hydroxybutyratedehydrogenase

42
Ketone Bodies Are a Major Fuel in Some Tissues
• Acetoacetate and 3-hydroxybutyrate are produced mainly in the liver mitochondria.
• Acetoacetate and 3-hydroxybutyrate are normal fuels of respiration and are quantitatively important as sources of energy.– Heart muscle and the renal cortex use acetoacetate in
preference to glucose.– In contrast, glucose is the major fuel for the brain and red
blood cells in normal conditions.– However, the brain adapts to the utilization of acetoacetate
during starvation and diabetes

43
• Acetoacetate can be activated by the transfer of CoA from succinyl CoA in a reaction catalyzed by a specific CoA transferase.
• Acetoacetyl CoA is then cleaved by thiolase to yield two molecules of acetyl CoA, which can then enter the citric acid cycle.
• The liver has acetoacetate available to supply other organs because it lacks this particular CoA transferase.
• Acetoacetate also has a regulatory role:– High levels of acetoacetate in the blood lead to a
decrease in the rate of lipolysis in adipose tissue.

44
Animals Cannot Convert Fatty Acids into Glucose
• Acetyl CoA cannot be converted into pyruvate or oxaloacetate in animals.
• Oxaloacetate is regenerated, but it is not formed de novo when the acetyl unit of acetyl CoA is oxidized by the citric acid cycle.
• In contrast, plants have two additional enzymes enabling them to convert the carbon atoms of acetyl CoA into oxaloacetate.

45
Fatty Acids Synthesis

46
Fatty Acids Are Synthesized and Degraded by Different Pathways
• Some important differences between the pathways are:1.Synthesis takes place in the cytosol, in contrast with
degradation, which takes place primarily in the mitochondrial matrix.
2. Intermediates in fatty acid synthesis are covalently linked to the sulfhydryl groups of an acyl carrier protein (ACP), whereas intermediates in fatty acid breakdown are covalently attached to the sulfhydryl group of coenzyme A.
3.The enzymes of fatty acid synthesis in higher organisms are joined in a single polypeptide chain called fatty acid synthase. In contrast, the degradative enzymes do not seem to be associated.

47
4. The growing fatty acid chain is elongated by the sequential addition of two-carbon units derived from acetyl CoA.
The activated donor of two carbon units in the elongation step is malonyl ACP. The elongation reaction is driven by the release of CO2.
5. The reductant in fatty acid synthesis is NADPH, whereas the oxidants in fatty acid degradation are NAD+ and FAD.
6. Elongation by the fatty acid synthase complex stops on formation of palmitate (C16).
Further elongation and the insertion of double bonds are carried out by other enzyme systems.

48
The Formation of Malonyl Coenzyme A Is the Committed Step in Fatty Acid
Synthesis
• Fatty acid synthesis starts with the carboxylation of acetyl CoA to malonyl CoA by acetyl CoA carboxylase, which contains a biotin prosthetic group.

49
• The carboxyl group of biotin is covalently attached to the -amino group of a lysine residue, as in pyruvate carboxylase.
• A carboxybiotin intermediate is formed at the expense of the hydrolysis of a molecule of ATP.
• The activated CO2 group in this intermediate is then transferred to acetyl CoA to form malonyl CoA.
• Acetyl CoA carboxylase is also the essential regulatory enzyme for fatty acid metabolism.
50
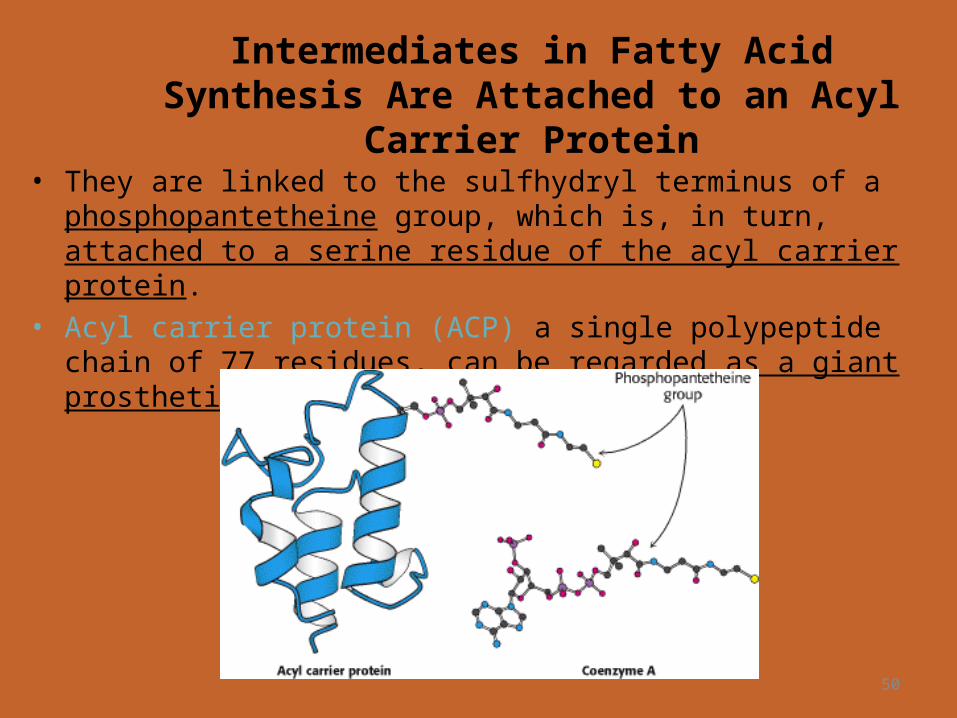
Intermediates in Fatty Acid Synthesis Are Attached to an Acyl Carrier
Protein• They are linked to the sulfhydryl terminus of a phosphopantetheine
group, which is, in turn, attached to a serine residue of the acyl carrier protein.
• Acyl carrier protein (ACP) a single polypeptide chain of 77 residues, can be regarded as a giant prosthetic group, a "macro CoA."

51
The Elongation Cycle in Fatty Acid Synthesis
• The enzyme system that catalyzes the synthesis of saturated long-chain fatty acids from acetyl CoA, malonyl CoA, and NADPH is called the fatty acid synthase.
• The elongation phase of fatty acid synthesis starts with the formation of acetyl ACP and malonyl ACP.
• Acetyl transacylase (nonspecific) and malonyl transacylase (specific) catalyze these reactions.

52
1
The decarboxylation of malonyl ACP contributes a substantial decrease in free energy.
• In effect, ATP drives the condensation reaction:– ATP is used to carboxylate acetyl CoA to
malonyl CoA.– The free energy stored in malonyl CoA is
released in the decarboxylation accompanying the formation of acetoacetyl ACP.

53
Acetoacetyl ACP is reduced to D-3-hydroxybutyryl ACP.
• This reaction differs from the corresponding one in fatty acid degradation in two respects: – The D rather than the L isomer is
formed.– NADPH is the reducing agent, whereas
NAD+ is the oxidizing agent in oxidation.
2

54
3
Then D-3-hydroxybutyryl ACP is dehydrated to form crotonyl ACP, which is a trans-2-enoyl ACP.
Crotonyl ACP is reduced to butyryl ACP. • NADPH is again the reductant, whereas FAD is
the oxidant in the corresponding reaction in -oxidation.
• The enzyme that catalyzes this step is enoyl ACP reductase.
4

55
In the second round of FA synthesis, butyryl ACP condenses with malonyl ACP to form a C6--ketoacyl ACP.
• The elongation cycles continue until C16-acyl ACP is formed.
This intermediate is a good substrate for a thioesterase that hydrolyzes C16-acyl ACP to yield palmitate and ACP.
FAs with an odd number of carbon atoms are synthesized starting with propionyl ACP, which is formed from propionyl CoA by acetyl transacylase.

56
Fatty Acids Are Synthesized by a Multifunctional Enzyme Complex in Eukaryotes
• Many eukaryotic multienzyme complexes are covalently linked enzymes in multifunctional proteins:
1. An advantage of this arrangement is that the synthetic activity of different enzymes is coordinated.
2. A multienzyme complex consisting of covalently joined enzymes is more stable than one formed by noncovalent attractions.
3. Intermediates can be efficiently handed from one active site to another without leaving the assembly.

57
Schematic Representation of Animal Fatty Acid Synthase: Each of the identical chains in the dimer contains three domains
Acetyl transferase
Condensing enzyme-ketoacyl synthase
Malonyl transferase
acyl carrier protein
-ketoacyl reductase
enoyl reductase
dehydratase
Thioesterase
1
2
3

58
Condensation Reduction Dehydration
Reduction
TranslocationNew Malonyl unit

59
• Five more rounds of condensation and reduction produce a palmitoyl (C16) chain on the condensing enzyme.
• Palmitoyl is hydrolyzed to palmitate by the thioesterase on domain 3 of the opposite chain.
• The flexibility and 20-Å maximal length of the phosphopantetheinyl moiety are critical for the function of this multienzyme complex.– The enzyme subunits need not undergo large structural
rearrangements to interact with the substrate.– Instead, the substrate is on a long, flexible arm that can
reach each of the numerous active sites.

60
The Stoichiometry of Fatty Acid Synthesis:
The stoichiometry of the synthesis of palmitate is:
The equation for the synthesis of the malonyl CoA used in the preceding reaction is:
Hence, the overall stoichiometry for the synthesis of palmitate is:

61
Citrate Carries Acetyl Groups from Mitochondria to the Cytosol for Fatty Acid Synthesis
• Fatty acids are synthesized in the cytosol, whereas acetyl CoA is formed from pyruvate in mitochondria.
• Carnitine carries only long-chain fatty acids.• The barrier to acetyl CoA is bypassed by citrate, which carries
acetyl groups across the inner mitochondrial membrane.

62
First TCA reaction
When present at high conc. ATP-citrate lyase
Malate dehydrogenase
Malic enzyme
Pyruvate carboxylase
The sum

63
• One molecule of NADPH is generated for each molecule of acetyl CoA that is transferred from mitochondria to the cytosol.
• Hence, eight molecules of NADPH are formed when eight molecules of acetyl CoA are transferred to the cytosol for the synthesis of palmitate (C16).
• The additional six molecules of NADPH required for this process come from the pentose phosphate pathway.

64
Acetyl Coenzyme A Carboxylase Plays a Key Role in Controlling Fatty Acid Metabolism
1. Global Regulation.– Global regulation is carried out by means of
reversible phosphorylation.– Acetyl CoA carboxylase is switched off by
phosphorylation and activated by dephosphorylation.

65
ATP
(-)
AMP
(+)
Inactivated at low energy charge
(-) PKA
P
Epinephrine
Glucagon
Phosphatases ?
Insulin
Activated by insulinAnd inactivated by catablic hormons (Epinephrin and Glucagon)

66
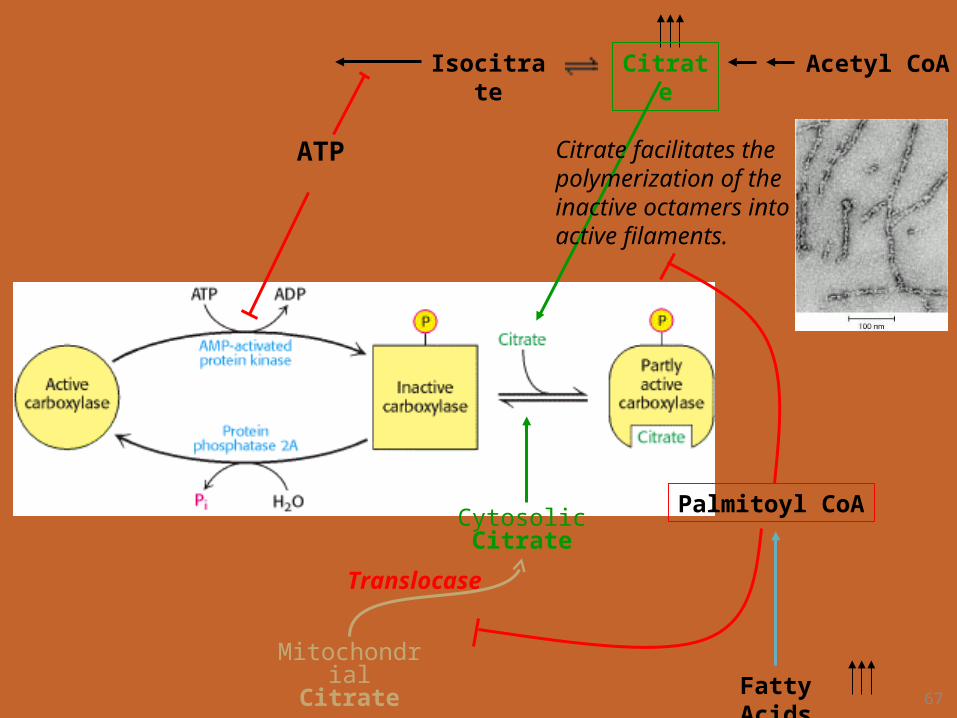
2. Local Regulation.– Acetyl CoA carboxylase is allosterically
stimulated by citrate.– Citrate partly reverses the inhibition produced
by phosphorylation.
67
ATP
(-)
CitrateIsocitrate Acetyl CoA
Citrate facilitates the polymerization of the inactive octamers into active filaments.
Palmitoyl CoA
Fatty Acids
Translocase
Mitochondrial Citrate
Cytosolic Citrate

68
Elongation of Fatty Acids• The major product of fatty acid synthase is palmitate.• In eukaryotes, longer fatty acids are formed by reactions catalyzed by
enzymes on the cytosolic face of the endoplasmic reticulum membrane.
• These reactions add two-carbon units sequentially to the carboxyl ends of both saturated and unsaturated fatty acyl CoA substrates.
• Malonyl CoA is the two-carbon donor in the elongation of fatty acyl CoAs.
• Condensation is driven by the decarboxylation of malonyl CoA.

69
Membrane-Bound Enzymes Generate Unsaturated Fatty Acids
• Double bonds are introduced into long-chain acyl CoAs in endoplasmic reticulum systems.
• For example: in the conversion of stearoyl CoA into oleoyl CoA, a cis-9 double bond is inserted by an oxidase.

70
• Mammals lack the enzymes to introduce double bonds at carbon atoms beyond C-9 in the fatty acid chain.
• Hence, mammals cannot synthesize linoleate (18:2 cis-9, 12) and linolenate (18:3 cis-9, 12, 15).
• The two essential fatty acids linoleate and linolenate furnished by the diet are the starting points for the synthesis of a variety of other unsaturated fatty acids.