fch 532 lecture 10 extra credit posted on website, due on friday (email or typed copy) chapter 29
TRANSCRIPT

FCH 532 Lecture 10
Extra credit posted on website, due on Friday (email or typed copy)
Chapter 29

Sugar Pucker
• Ribose rings will undergo sugar pucker (become slightly nonplanar).
• Most structures show that 4 of the five ring atoms are coplanar, 5th atom is out of the plane in a half-chair conformation.
• Endo conformation-if out of plane atom is the same side of the ring as the C5’
• Exo conformation-if out of plane atom is on the opposite side of the ring as the C5’
• Most nucleoside and nucleotide structures (not in double helices) out of plane atom is either C2’ or C3’.
• C2’-endo most common; C3’-endo and C3’-exo are also common.

Figure 29-9aSugar ring pucker. (a) A planar ribose ring (here viewed down the C3—C4 bond) are all eclipsed.
Pag
e 11
17

Figure 29-9bSugar ring pucker. (b) The steric strain resulting in Part a is partially relieved by ring puckering in a half-chair conformation in which C3 is the out-of-
plane atom (same as C5’ so C3’-endo).
Pag
e 11
17

Sugar Pucker
• Endo conformation-if out of plane atom is the same side of the ring as the C5’
• Exo conformation-if out of plane atom is on the opposite side of the ring as the C5’
• Most nucleoside and nucleotide structures (not in double helices) out of plane atom is either C2’ or C3’.
• C2’-endo most common; C3’-endo and C3’-exo are also common.• This ribose pucker determines the relative orientatation of the phosphates
to the sugar.• To have a regularly repeating model-need C2’-endo (B-DNA) or C3’-endo
(A-DNA, RNA-11)• Z-DNA purines are all C3’-endo, pyrimidines C2’-endo (dinucleotide
repeating unit).• B-DNA has some flexibility (can be observed as C4’-exo, O4’-endo, C1’-
exo and C3’-exo).

Figure 29-10a Nucleotide sugar conformations. (a) The C3-endo conformation (on the same side of the sugar ring as C5), which occurs in A-
RNA and RNA-11.
Pag
e 11
17

Figure 29-10b Nucleotide sugar conformations. (b) The C2-endo conformation, which
occurs in B-DNA.
Pag
e 11
17

Base Pairing
• Watson-Crick pairs most common and only pairing seen in crystal structures of self-complimentary oligonucleotides.
• Other geometries available.

Figure 5-12 Watson-Crick base pairs.P
age
88
Adenine pairs with thymine. Guanine pairs with cytosine.These base pairs are interchangeable in the double helix without altering the positions of the sugar phosphate backbone.
The top edge of each base pair is structurally distinct from the bottom edge.
The deoxyribose residues are asymmetric.
Minor groove exposes the edge from the C1’ atom (open toward bottom)
Major groove exposes the opposite edge of each bp.

Hoogsteen base pairing
• Monomeric A and T derivatives form A-T base pairs with adenine N7 as the hydrogen bonding acceptor (normally N1). This is Hoogsteen base pairing
• Steric influences make Watson-Crick base pairing the norm for double helices.
• Hoogsteen base pairs are biologically relevant. They help stabilize tRNA tertiary structure.
• G-C always form Watson-Crick base pairs in crystallized structures because of their 3 hydrogen bonds.

Figure 29-12a Some non-Watson–Crick base pairs.
(a) The pairing of adenine residues in the crystal structure of 9-methyladenine.
Pag
e 11
18

Figure 29-12b Some non-Watson–Crick base pairs. (b) Hoogsteen pairing between adenine
and thymine residues in the crystal structure of 9-methyladenine·1-methylthymine.
Pag
e 11
08

Figure 29-12c Some non-Watson–Crick base pairs. (c) A hypothetical pairing between cytosine
and thymine residues.
Pag
e 11
18

Watson-Crick pairs have high mutual affinity
• IR spectroscopy in the N-H region of region of guanine and cytosine derivatives separately and in a mixture shows the interaction between base pairs.
• New band near 3500 cm-1 for G+C indicates the interaction.
• The same calculated and observed spectra for G+A indicates the lack of interaction.
• Can be described by association constant K.
B1 + B2 B1 • B2 [B1 • B2]K=
[B1][B2]
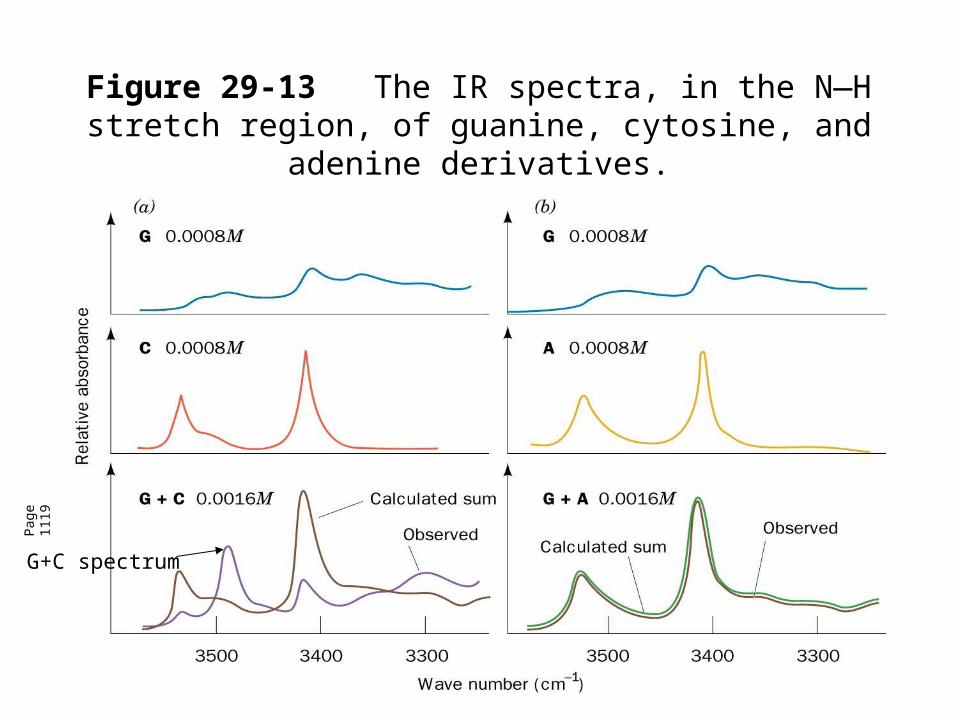
Figure 29-13The IR spectra, in the N—H stretch region, of guanine, cytosine, and adenine derivatives.
Pag
e 11
19
G+C spectrum

Table 29-2 Association Constants for Base Pair Formation.
Pag
e 11
19

Figure 29-14Stacking of adenine rings in the crystal structure of 9-methyladenine.
Pag
e 11
20

Bases stack in aqueous solution
• Bases will aggregate together in aqueous solution.• Interaction can be described by a variation of the van’t Hoff law of osmotic pressure: =RTm is the osmotic pressure• m=molality of solution (mol solute/kg solvent)• R=gas constant• T=temperature• The molecular mass, M, of an ideal solute can be determined from its osmotic pressure. M=c/m, where c=g solute/kg solvent

Bases stack in aqueous solutionFor solutions that aggregate, van’t Hoff law is rewritten as= RTmthe osmotic coefficient: indicates the solute’s degree of association and varies from 1 (no association) to 0 (infinite association)• The variation of with m for nucleic acid bases in solution is consistent with a model where bases aggregate in a step-wise fashion:
A + A A2 + A A3 + A … An
Where n is at least 5 (if the reaction goes to completion, 1/n)
This result is not based on hydrogen bonding since N6, N6-dimethyladenosine cannot form hydrogen bonds to itself.
Aggregation instead results from stacks of planar molecules.Corroborated with NMR studies.Single-stranded polynucleotides show a similar trend.

Figure 29-15Variation of the osmotic coefficient with the molal concentrations m of adenosine derivatives in
H2O.
Pag
e 11
21
molality of solution (mol solute/kg solvent)
No association
∞ association

Figure 29-16 Melting curves for poly(A) and ApA.
Pag
e 11
21
Broad increase in UV abs with increase in temp for poly(A) and dinucleotides
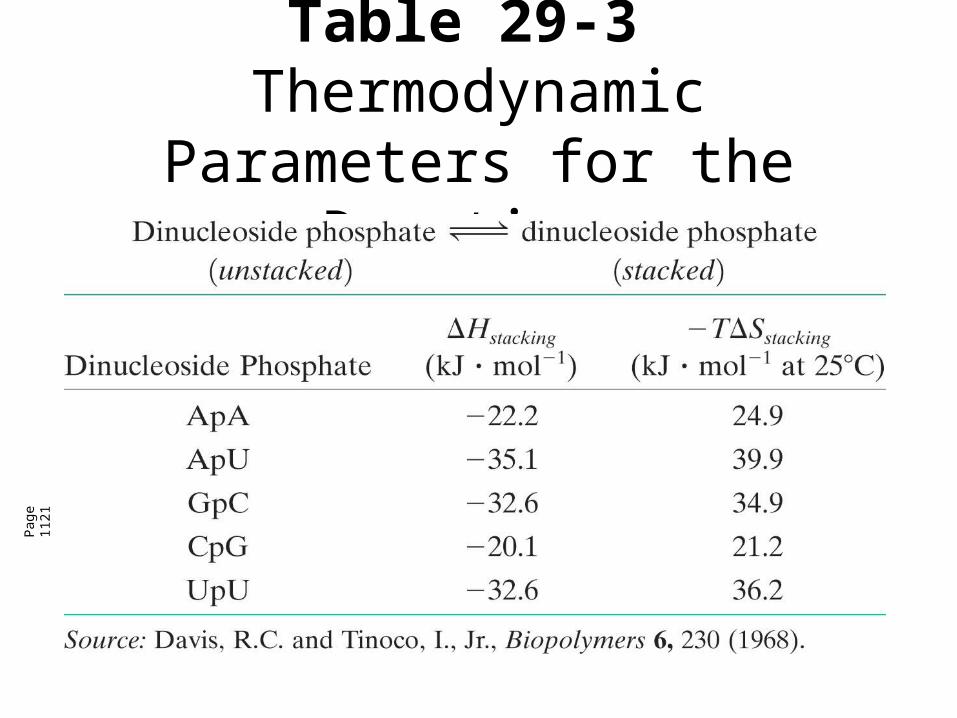
Table 29-3Thermodynamic Parameters for the Reaction.
Pag
e 11
21

Nucleic acids stabilized by hydrophobic forces
• Stacking interactions in aqueous solutions are mostly stabilized by hydrophobic forces.
• Base stacking is enthalpically driven and entropically opposed.
• Hydrophobic intereactions responsible for stability of base stacking nucleic acids are diametrically opposed in character to those that stabilize proteins (enthalpically driven but entropically opposed)

Ionic interactions
• Electrostatic interactions are also important.• Melting temperature of double-stranded DNA increases with the
cation concentration since these ions bind more tightly to duplex DNA(higher anionic charge) than to single-stranded DNA.
• Increased salt shifts equilibrium toward duplex DNA.• Tm = 41.1XG+C + 16.6 log[Na+] +81.5• XG+C = mole fractionof G+C content and is valid if :
0.3< 1XG+C <0.7 and 10-3 M< [Na+]< 1.0 MCan be used for other monovalent cations (K+, Li+)Divalent cations such as Mg2+, Mn2+, Co2+ bind specifically to
phophate groups and are more effective shielding agents.1 Mg2+ ion is the equivalent of 100 to 1000 Na+ ions.
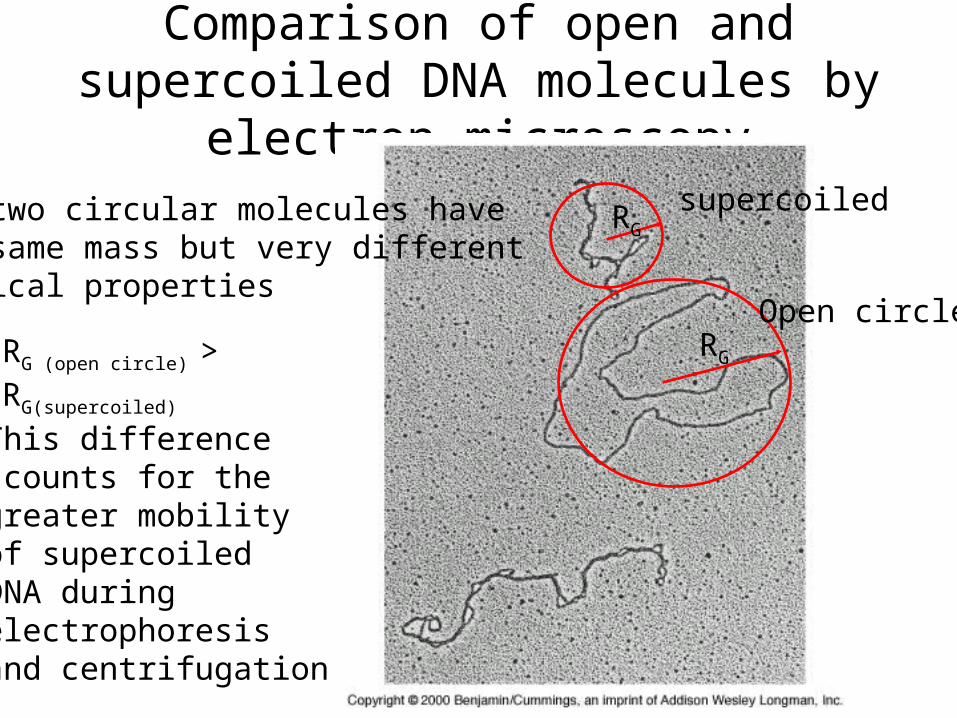
Comparison of open and supercoiled DNA molecules by electron microscopy
RG
RGRG (open circle) >RG(supercoiled)
This differenceccounts for thegreater mobilityof supercoiledDNA during electrophoresisand centrifugation
supercoiled
Open circle
The two circular molecules havethe same mass but very differentphysical properties

DNA topology
• The same double-stranded DNA molecule can have different conformations
Linear“open” Supercoiled
Circular
Relative radius of gyration Linear>circles>supercoils
Frictional coefficient influenceselectrophoretic mobility and sedimentation rate.

Detecting different DNA topoisomers
A. Electrophoretic mobility
B. Sedimentation rate
Which form goes fastest in each technique?

Different topoisomers of circular DNA can be separated by electrophoresis
Supercoiled moleculeswith increasing twists

Detecting different DNA topoisomers by intercalation of dye molecules

Detecting different DNA topoisomers
(Ethidium bromide concentration)
Sedimentation
rate
A
B
C
Supercoiled>open>supercoiled (negative) (______)
Gel electrophoresis of a mixture of linear, open, and supercoiled DNA treated with EB
Top (- electrotrode)
Bottom (+ electrotrode)
Linear/open
topoisomers
To t2hrs

1. Supercoils refer to the DNA structure in which double-stranded circular
DNA twists around each other. Supercoiled DNA contrasts relaxed DNA;
2. In DNA replication, the two strands of DNA have to be separated, which
leads either to overwinding of surrounding regions of DNA or to
supercoiling;
3. A specialized set of enzymes (gyrase, topoisomerases) is present to
introduce supercoils that favor strand separation;
4. The degree of supercoils can be quantitatively described.
Tertiary structure in DNA

Varieties of supercoiled DNA
Toroidal spirals within supercoil