fijación biológica del nitrógeno
DESCRIPTION
Ministerio de Agricultura y Tierras, 2006TRANSCRIPT

1la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
la fijación biológica del nitrógeno atmosférico en condiciones tropicales

2 capítulo 1. la fijación biológica del nitrógeno atmosférico como uno de los pilares básicos de la agroecología

3la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
la fijación biológica del nitrógeno atmosfériconitrógeno atmosféricoen condiciones tropicales
rafael martínez viera
marisol lópez
bernardo dibut álvarez
carmen parra zambrano
janet rodríguez sánchez

4 capítulo 1. la fijación biológica del nitrógeno atmosférico como uno de los pilares básicos de la agroecología
HUGO CHÁVEZ FRÍASHUGO CHÁVEZ FRÍASHUGO CHÁVEZ FRÍAS
PRESIDENTE DE LA REPÚBLICA BOLIVARIANA DE VENEZUELAPRESIDENTE DE LA REPÚBLICA BOLIVARIANA DE VENEZUELAPRESIDENTE DE LA REPÚBLICA BOLIVARIANA DE VENEZUELA
ELÍAS JAUA MILANO
MINISTRO DEL PODER POPULAR PARA LA AGRICULTURA Y TIERRAS
ÁNGELA BOLÍVARSERVICIO AUTÓNOMO DE SANIDAD AGOPECUARIA

5la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Contenido
Presentación 9Introducción 11
Capítulo 1 La fijación biológica de nitrógeno atmosféricocomo uno de los pilares básicos de la agroecología 13
La Revolución Verde y su situación actual 13Conceptos agroecológicos 17Importancia de los biofertilizantes dentro de la Agroecología 20Importancia de los biofertilizantes basados en bacteriasfijadoras del N2 atmosférico 23
Capítulo 2 Papel de la rizosfera en la efectividadde los biofertilizantes microbianos 25
Rizosfera y ambiente rizosférico 25Características de la rizosfera 26Influencia de las secreciones de las raíces sobrelos microorganismos 30Quimiotaxismo 31Diferencias entre el efecto de las condiciones de clima templadoy tropical sobre las interacciones planta-microorganismosen la zona rizosférica 33
Capítulo 3 Generalidades sobre la fijacion biológica del nitrógeno atmosférico 37
Importancia del Nitrógeno 37La fijación de N2 por métodos industriales 38La fijación biológica del N2 39Resumen histórico de los estudios sobre fijación biológica del N2 40Mecanismo de la fijación biológica de N2: la nitrogenasa 44Distribución de los sistemas fijadores 46Organismos fijadores de N2 más utilizados como biofertilizantes 49

6 contenido
Influencia de los factores ambientales sobrela fijación biológica de N2 51Importancia del Fósforo para la fijación de N2 52Métodos para medir la fijación de nitrógeno 55Importancia económica de la aplicación de los biofertilizantesa base de bacterias fijadoras de N2 56
Capítulo 4 Estimulación del desarrollo de las plantas por las bacteriasfijadoras de nitrógeno atmosférico 59
Sustancias reguladoras del crecimiento vegetal 59Mecanismos de estimulación del crecimiento porparte de las bacterias fijadoras de Nitrógeno 61Síntesis de auxinas por las bacterias fijadoras 63Síntesis de giberelinas por las bacterias fijadoras 65Síntesis de citoquininas por las bacterias fijadoras 66Sustancias activas sintetizadas por Azotobacter chroococcum 67
Capítulo 5 La fijación simbiótica del nitrógeno 71
Importancia de la fijación simbiótica 71Establecimiento de la simbiosis bacterias fijadoras-leguminosas 73Características de los nódulos 76Relaciones entre los nódulos y la capacidad de fijación de N2 78Clasificación actual de las bacterias que forman nódulosen las plantas leguminosas 80Factores que influyen en el establecimiento de la simbiosis 82Cálculo de los beneficios aportados al suelo porla fijación simbiótica de N2 atmosférico 86Asociación de Rhizobium con cereales 88Fijación simbiótica en plantas no leguminosas 90
Capítulo 6 La fijación asociativa de nitrógeno 95
Mecanismos de protección de la nitrogenasa en la fijación aeróbica 95Microorganismos capaces de fijar N2 en forma asociativa 97

7la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Factores ambientales que afectan la fijación asociativa de N2 98Las bacterias del género Azotobacter 10Azotobacter 10Azotobacter 0Las bacterias del género Azospirillum 110La bacteria endófita Gluconacetobacter diazotrophicus 113La fijación de N2 en la filosfera 116
Capítulo 7 Preparación y aplicación de inoculantes a base de bacteriasfijadoras de nitrógeno atmosfégeno atmosfégeno atmosf rico 119
Método de selección de cepas 120Características de los portadores 121Control de calidad de los biopreparados 123Biofertilizantes mixtos utilizando bacterias fijadoras de N2 126Determinación del efecto antagonista entre las especiesbacterianas aspirantes a ser utilizadas en un producto mixto 132Métodos de inoculación de los biofertilizantes a basede bacterias fijadoras de Nitrógeno atmosférico 133
Bibliografía 137
Índice de tablas 167 Índice de figuras 171

8 contenido

9la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Presentación
Estamos en momento de lucha, pero también de aprendizaje. No podemos negar que la cuestión ambiental ha sido no solo una forma más de expresión de la irracional vo-racidad capitalista, sino también un asunto largamente postergado por los movimientos en lucha contra el capitalismo globalizado.
No se trata de asumir una culpa. Los procesos y las banderas de la resistencia son tam-bién determinados por las circunstancias históricas. Para los que entregamos nuestros más añorados años a la causa de la revolución, circunstancias como las dictaduras sure-ñas, los procesos emancipatorios en Centroamérica, las guerrillas insurgentes de los paí-ses andinos, la represión de nuestras democracias de fachada y las múltiples formas de la dominación y la explotación del hombre no nos permitían desviar nuestros esfuerzos de la cuestión netamente humana. Sin embargo, no por esto somos menos responsables.
Con el advenimiento de la “triste noche neoliberal”, precipitada por la desaparición del bloque socialista, el movimiento mundial por la transformación, en todos sus niveles, tuvo que replegarse para rearticularse. Hay que admitirlo, a pesar de nuestras críticas y nuestros más pesimistas pronósticos, la contrarrevolución conservadora (como algu-nos la han llamado) nos sorprendió y por un tiempo nos abatió.
Sin embargo, pronto nos dimos cuenta de la más irrevocable de las verdades, nuestra causa es una causa invencible. En Venezuela el despertar vino, inimaginablemente, de los substratos de la sociedad. Los marginados despertaron para conmover de un sacu-dón las bases del sistema de dominación y marcarlo con todas las gritas que causarían su posterior desplome, y con él, la apertura de un proceso de emancipación que bien podríamos asumir como continental.
En el mundo, la historia nuevamente comenzó a correr, por todas partes reaparecieron las organizaciones ciudadanas en demanda de un mundo mejor y distinto. Organizacio-nes políticas, sindicales y movimientos sociales levantaron nuevamente las banderas con las más radicales consignas. Y así poco a poco nos hemos encontrado con un mundo que por un lado globaliza la injusticia y por el otro, en conflicto con esta, la esperanza.
Pero en esta marcha por la equidad y la justicia mundial ahora nos acompañaban nue-vos actores y nuevas causas. Juntos con los partidos, los sindicatos, y los movimientos

10 presentación
populares, aparecieron las demandas de otros movimientos. Entre ellos el movimiento mundial por el desarrollo sostenible y la defensa del ambiente.
Al igual que con otros asuntos, hemos tenido que aprender que no podemos supeditar la responsabilidad ética que tenemos con el medio ambiente a la erradicación total del hambre y la explotación humana. De hecho, hemos tenido que darnos cuenta de que estos no son asuntos independientes, de que el hambre y la miseria están estrecha-mente vinculadas a los mecanismos cada vez más ecocidas de aprovechamiento de los recursos naturales.
De la misma forma, nos hemos dado cuenta de que la soberanía tecnológica, parte fundamental de la soberanía nacional va unida a la reivindicación de nuestras potenciali-dades naturales y de nuestras técnicas ancestrales de aprovechamiento de los recursos que nos brinda la madre tierra. Estas, como materia prima del desarrollo de nuevos conocimientos generados con criterios ecológicos, constituyen un factor determinante en el proceso de descolonizaión de nuestras formas de producir y reproducir la vida.
Todo este aprendizaje nos impulsa hoy a producir ciencia liberadora. Conocimientos científicamente adquiridos, como la agroecología, cuyos principios promueven la con-servación y preservación de todos los componentes de los agroecosistemas, iniciándo-se así la promoción, producción y uso de tecnologías como la de los biofertilizantes.
En este contexto se inscribe la iniciativa de publicación de este libro, “La fijación biológi-ca del nitrógeno atmosférico en condiciones tropicales”, que constituye un orgullo para el Ministerio del Poder Popular para la Agricultura y Tierra, en tanto que representa no solo la emancipación tecnológica de nuestros pueblos, sino también la dignidad de científicos cubanos y venezolanos que pese a las dificultades han sumado esfuerzos par hacerlo posible. Dar a conocer, a través de este didáctico texto, los procesos naturales de fijación biológica del nitrógeno atmosférico, y su importantísimo rol en la activación de los sistemas agrícolas sostenibles, así como los beneficios ambientales derivados del mismo, es un acto de desprendimiento y compromiso que agradecemos infinitamente; vista la trascendencia y riqueza del tema descrito, y siguiendo los lineamientos presi-denciales de tercer motor constituyente “Moral y Luces” es más que un deber, una necesidad, proporcionar al pueblo venezolano esta herramienta que los autores nos ofrecen con la humildad que les otorga su propia sabiduría y conciencia planetaria.
Elías Jaua Milano

11la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Introducción
En las últimas décadas se han logrado grandes progresos en el vasto dominio de las Ciencias Biológicas. La investigación multidisciplinaria ha favorecido el desarrollo de la Biología y la Bioquímica y ha aumentado el campo de aplicación de estas ciencias. Al mismo tiempo, muchas búsquedas están originadas por el deseo de ver al hombre y a su entorno ocupando un primer plano, buscando la forma de mejorar la calidad de vida del primero y de sanear y preservar al segundo.
Desde el punto de vista agrícola, se hace cada vez más urgente la necesidad de pro-ducir los más variados alimentos con la utilización de métodos que exijan un consumo menor de energía y que no contaminen los agroecosistemas. Ya los conceptos que rigieron la Revolución Verde se han ido sustituyendo por los que abogan por una Agri-cultura Ecológica, capaz de aprovechar el desarrollo tecnológico, pero que utiliza en lo fundamental elementos naturales, lo cual puede hacerse hoy con notable éxito gracias a los progresos de la Agrobiología.
Entre los temas biológicos que se han estudiado con más profundidad se encuentran los relacionados con los procesos microbiológicos que influyen sobre la nutrición y el desarrollo de las plantas, cuyo conocimiento ha permitido la extensa utilización que hoy tienen en muchos países los biofertilizantes y bioestimuladores. El más estudiado y conocido de estos procesos es la fijación biológica del N2 atmosférico, que aporta a las plantas cantidades importantes de Nitrógeno, el principal elemento nutritivo para los cultivos, lo cual permite racionalizar las aplicaciones de fertilizantes inorgánicos y redu-cir de una forma marcada los niveles de contaminación de los agroecosistemas.
La acción de los microorganismos fijadores se realiza en todos los componentes de la fitosfera, nombre con el que se conoce el ecosistema total suelo-planta-atmósfera, es decir, el ambiente que rodea más estrechamente a las plantas. Por esta razón, puede haber también fijación de Nitrógeno atmosférico en la filosfera, zona que rodea la parte aérea de la planta y que está en contacto con el follaje y la atmósfera, sometida a la acción reguladora de ambas. Pero la mayor actividad ocurre en la rizosfera, zona que rodea a la planta y que está en contacto con las raíces y el suelo.

12 introducción
De lo expuesto se deduce que existe una estrecha interrelación entre el suelo, la planta y los microorganismos, por lo que se hace necesario conocer como influyen cada uno de estos factores sobre los demás y en que forma las variaciones que ocurren en cada uno de ellos modifican el comportamiento general de la asociación. Una vez que se haya adquirido un conocimiento profundo del sistema en su conjunto, será posible sacar los mayores beneficios del uso de los biofertilizantes, representados por mayores aumentos de los rendimientos y la calidad de las cosechas, lo que se consigue con aplicaciones mínimas de fertilizantes inorgánicos y nuevos esquemas de fertilización orgánica.
En el presente libro se ofrece una visión teórica de los distintos procesos fijadores, y se muestran los beneficios que pueden obtenerse en el medio tropical con el uso de los biofertilizantes y bioestimuladores a base de bacterias fijadoras de Nitrógeno atmosférico y de los mejores métodos de aplicación, lo cual permitirá un mayor apro-vechamiento del enorme potencial biológico que se encierra en los suelos tropicales, al mismo tiempo que se reducen los problemas económicos y ecológicos derivados de la aplicación indiscriminada de los fertilizantes industriales.
En la República Bolivariana de Venezuela, la utilización de estos bioproductos constitu-ye una necesidad para el desarrollo agrícola del país, ya que permite los incrementos de los rendimientos agrícolas por medios más económicos y saludables, al mismo tiempo que generan la creación de empleos en una región determinada.
En estos momentos se está desarrollando en Venezuela, con el aporte fundamental del Ministerio de Agricultura y Tierras (MAT), una red de laboratorios que se dedicarán a la fabricación de biofertilizantes y bioestimuladores microbianos para ser utilizados en to-das las regiones del país, lo que incluye la fabricación y aplicación de grandes cantidades de bioproductos a base de bacterias fijadoras de forma simbiótica y asociativa.
El objetivo principal que anima a los autores es suministrar a los profesionales y técnicos venezolanos los elementos necesarios para que puedan obtener los mayores benefi-cios de la utilización de las bacterias fijadoras, con lo que se contribuye de una manera destacada a que plasmen en realidad los esfuerzos que hoy se realizan en el país para garantizar las necesidades alimentarias de la población y preservar el medio ambiente, al mismo tiempo que se ofrece una herramienta para el manejo alternativo y económi-co de los agroecosistemas sin afectar los rendimientos.

13la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Capítulo 1La fijación biológica de nitrógeno atmosféricocomo uno de los pilares básicos de la agroecología
La Revolución Verde y su situación actual
El éxito de la Revolución Industrial provocó la extensión del modelo triunfante a todos los sectores de la economía, generalizándose en su más alto grado en la Agricultura al finalizar la II Guerra Mundial con el nombre de Revolución Verde.
Para alinear a la producción agrícola sobre el modelo industrial fue necesario lograr su estandarización; de esta manera, para facilitar la transformación de los productos agrí-colas y simplificar los circuitos comerciales se redujo el número de especies y varieda-des utilizadas y se aplicó el modelo de separación espacial y temporal de las secuencias de producción. Así, las semillas no se obtienen en los hábitats donde las plantas son cul-tivadas, sino que son creadas algunas zonas especializadas para producirlas y después se comercializan en las más diversas regiones, lo que compromete las posibilidades de adaptación.
Aplicando siempre el modelo industrial, se concentran las producciones agrícolas para rentabilizar el capital. Surge el monocultivo en áreas donde había policultivo y se in-troducen cultivos repetitivos que empobrecen la microfauna y microflora del suelo y facilitan la multiplicación de plagas y organismos patógenos, que disponen de ver-daderos caldos de cultivo. La extensión de superficies uniformes conduce al empleo de máquinas cada vez más pesadas que alteran las propiedades físicas de los suelos (Levins, 1973).
Independientemente de los éxitos productivos, y visualizado la Revolución Verde des-de el punto de vista social, puede decirse que se marginó a gran parte de la población rural en todo el mundo, ya que los beneficios estuvieron centrados en los grupos ricos, los únicos con posibilidades de aplicar las costosas tecnologías intensivas. Fueron des-truidas muchas formas de acceso a la tierra y a los recursos tales como la aparcería, la contratación de mano de obra y la utilización de las aguas de riego y de tierras de pas-

14 capítulo 1. la fijación biológica del nitrógeno atmosférico como uno de los pilares básicos de la agroecología
toreo. Se redujo la diversidad de estrategias de subsistencia disponibles para las familias rurales y aumentó su dependencia del gran empresario agrícola.
Por otra parte, las consecuencias de la quimización han sido nefastas para el ambiente por la elevada contaminación causada por el uso irracional de fertilizantes y plaguicidas, que causa graves daños en la salud del ser humano y de los animales. Sin tomar en consideración a los plaguicidas químicos, los más contaminantes, por no estar relacio-nados con el tema que nos ocupa, el impacto ambiental causado por el uso excesivo de fertilizantes puede resumirse de la siguiente manera, tomando como ejemplo el fertilizante nitrogenado:
1. La fabricación de fertilizantes produce emisiones de CO2 y de NO2 a la atmósfera, lo cual contribuye a incrementar los problemas de la capa de ozono.
2. La aplicación en el campo da lugar a lixiviación de nitratos y a emisiones de N2O y de NH3, especialmente a partir de urea, con la consiguiente contaminación de todos los componentes del agroecosistema.
La abundancia de compuestos nitrogenados en los mantos acuíferos como con-secuencia de la lixiviación hace que en todos los reservorios donde van a desem-bocar estas corrientes subterráneas se produzca el proceso llamado eutroficación, que consiste en el crecimiento anormal de microorganismos que agotan el oxí-geno disuelto en el agua y producen la muerte masiva de los peces. Estos graves problemas de contaminación se han puesto ya de manifiesto en algunos de los ríos más caudalosos del mundo como el Mississipi en Estados Unidos y el Yang Tse en China, en cuyas desembocaduras se aprecia una notable carencia de flora y fauna. Según Altieri y Nicholls (2000), en las aguas afectadas por la eutrificación se estimu-la el desarrollo de Pfiesteria, un organismo que mata a los peces y es dañino para el ser humano.
3. Son muy numerosos los reportes que indican la presencia de altos niveles de nitratos en los productos agrícolas, lo que constituye una importante fuente de toxicidad para el ser humano y ha obligado a un establecimiento más riguroso del control de los niveles críticos de estos compuestos en las actividades de comercia-lización de los alimentos.
Pero, además de la contaminación, el incremento del uso de los fertilizantes industria-les ha estado acompañado por un aumento exponencial en el consumo de formas no

15la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
renovables de energía, las cuales se han convertido en un factor limitante para lograr aumentos de los rendimientos agrícolas. Se necesitan aproximadamente 1.3 t de com-bustible para fijar 1 t de nitrógeno atmosférico con alta presión y temperatura por el proceso industrial de Haber-Bosch.
Los 67 millones de toneladas anuales que se aplicaron en el mundo en 1996 como fertilizante nitrogenado requirieron de 87 millones de toneladas de combustible para su fabricación, lo que correspondió al 1.2 % de todo el combustible consumido en el planeta (Bökman, 1997; Olivares, 2006). Esta insostenibilidad se aumenta si se toma en cuenta que en el año 2020 deben aplicarse 130 millones de toneladas de fertilizante nitrogenado solo para la producción de cereales, de acuerdo con las proyecciones realizadas por Dyson (1996) y Lewis (1996), para cuya fabricación se necesitarán 169 millones de toneladas de petróleo, lo que reduciría a un ritmo alarmante las reservas conocidas en el mundo.
Por otra parte, hay que tomar en cuenta la insostenibilidad económica del uso exclusivo de fertilizantes inorgánicos, cuyos elevados precios los hacen inaccesibles para las ma-yorías campesinas de los países subdesarrollados. Así, en el año 1997, los países desa-rrollados fabricaron 58.202.000 de fertilizantes nitrogenados, mientras que los subde-sarrollados solo fabricaron 11.408.000 t, para una producción mundial de 69.611.000 t. En lo que se refiere a los fertilizantes de todo tipo, el promedio per cápita aplicado en el año 2.000 fue de 145 kg en los países del primer mundo y solo 23 kg en los subdesarrollados, lo que se tradujo en una producción de 927 kg de granos per cápita en el primer grupo de países y 210 kg en el segundo (FAO, 2001).
Como un ejemplo ilustrativo de lo que se exige a los países subdesarrollados para in crementar la producción de alimentos con las tecnologías de la Revolución Verde, puede decirse que en el África sub-sahariana, una de las regiones con más hambre en el mundo, hay que aumentar en 400.000 t anuales el consumo de fertilizantes para lo-grar el modesto aumento de 3% anual en la producción de alimentos sin empobrecer los suelos (FAO, 1995), lo cual es totalmente insostenible.
Por otra parte, el aumento del uso de los fertilizantes inorgánico de origen industrial se ha aproximado con gran rapidez al punto en que no se logran incrementos del rendi-miento proporcionales a su aplicación; así, en Chile recibió el trigo 62% más fertilizante entre 1985-1990 en comparación con la década anterior, pero los rendimientos au-

16 capítulo 1. la fijación biológica del nitrógeno atmosférico como uno de los pilares básicos de la agroecología
mentaron solo 3.1% (Altieri y Rojas, 1999). En el Estado de Georgia (USA) se reportó que el rendimiento de los cultivos se cuadruplicó entre las décadas del 70 y 80, mien-tras que las aplicaciones de nitrógeno crecieron 11 veces (Odum, 1989). Un análisis de los rendimientos del arroz en Filipinas, Indonesia y Tailandia mostró que declinaban, a pesar de los incrementos de fertilizante nitrogenado (Pingali y col., 1992).
De lo expuesto anteriormente se deduce que la producción industrial de fertilizantes no puede satisfacer las necesidades de una población mundial en creciente aumento, sobre todo cuando el exceso conduce a contaminar el agua de consumo, a la eu-trificación de los lagos y a las emisiones de óxido nitroso a la atmósfera, además del incremento considerable de la utilización de fuentes de energía no renovables (Brown, 1994).
Según Hetch (1997), el análisis de la Revolución Verde, hecho desde los puntos de vista de diferentes disciplinas, incluyó críticas ecológicas, tecnológicas y sociales y cons-tituyó la base para la profundización de los estudios posteriores sobre Agroecología y para las investigaciones sobre sistemas de labranza. Se ha llegado a reconocer que las tecnologías de la Revolución Verde solo pueden ser aplicadas en áreas limitadas y las nuevas tendencias de la investigación agrícola están dirigidas a resolver los problemas de los campesinos de bajos recursos.
Hoy se considera que el camino recorrido por la Revolución Verde en Europa y Es-tados Unidos está llegando a sus límites. Su alta eficiencia en términos de rendimiento por hectárea ha sido lograda a expensas de la ineficiencia energética y social, y socava el reciclaje de los recursos renovables con la tala indiscriminada de bosques, la con-taminación de los mantos acuíferos y la reducción de la biodiversidad. Este modelo destruye su propia base productiva con la erosión, la salinización y la compactación de los suelos, aumenta la vulnerabilidad de las plantas frente a las plagas y enfermedades, intoxica a los trabajadores agrícolas, a los consumidores y a la naturaleza entera y, por último, hasta constituye uno de los principales factores de transformación del clima terrestre.
Como se aprecia, los problemas que se van detectando son tantos que surge una situación contradictoria en los países más desarrollados: al mismo tiempo se exaltan las posibilidades de desarrollo de la llamada agricultura “moderna” y, como reacción, apa-rece un movimiento de agricultura “biológica” o alternativa que se fortalece cada vez

17la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
más. Surgen los nuevos conceptos agroecológicos que han dado origen a la moderna Agricultura Sostenible o Sustentable, junto con la crítica al rumbo del desarrollo agrícola mundial, adornado con los rótulos de “moderno”, “progresivo” y “avanzado”. Estos nuevos conceptos se han extendido entre los países desarrollados y subdesarrollados y, desde el punto de vista del suministro de nutrientes a las plantas, abogan por una combinación de cantidades reducidas de fertilizantes inorgánicos con biofertilizantes y abonos orgánicos.
Conceptos agroecológicos
El uso contemporáneo del término Agroecología comenzó en la década de los años 70, pero el conocimiento y la práctica agroecológica son tan antiguos como los orígenes de la Agricultura. La Agroecología incorpora ideas sobre un enfoque de la agricultura más ligado al medio ambiente, es más sensible socialmente y está centrado no solo en la producción, sino también en la sustentabilidad ecológica del sistema de producción.
La unidad ecológica principal en la Agroecología está representada por el agroecosis-tema, que no solo está determinado por los componentes estructurales, bióticos y abióticos, sino también por factores sociales tales como las variaciones de los precios en el mercado o cambios en la tenencia de la tierra, los cuales pueden influir en los sistemas agrícolas de forma tan marcada como la sequía, las plagas o los nutrientes del suelo. Como resultado de la interacción entre las características endógenas (biológicas y ambientales) y los factores exógenos (sociales y económicos) se genera la estructura particular de cada uno de los agroecosistemas.
Un sistema agroecológico es muy complejo y está formado por numerosas variables y parámetros. Estos últimos, ya sean constantes, periódicos o erráticos, están determi-nados externamente al sistema y son de muy difícil control. Las variables están en inte-racción recíproca continua y pertenecen a distintos campos de las Ciencias Naturales y Sociales; forman una red de interacciones por vínculos directos e indirectos y solamen-te se pueden aislar provisionalmente para el estudio, pero después hay que unirlos en los modelos para no cometer errores que pueden ser trascendentales. Ejemplos de errores cometidos por no tener en cuenta las interacciones pueden citarse muchos: plaguicidas que han fomentado nuevas plagas, formas de control de las corrientes de agua que aumentan el daño por las inundaciones, o medidas de modernización agraria que han provocado más desigualdades y más hambre.

18 capítulo 1. la fijación biológica del nitrógeno atmosférico como uno de los pilares básicos de la agroecología
Un sistema ecológico agrícola difiere en aspectos fundamentales de un sistema eco-lógico natural, tanto en su estructura como en su función. Los agroecosistemas son ecosistemas semi-domesticados a los que Odum (1984) les determinó 4 características principales:
1. Requieren fuentes auxiliares de energía, que puede ser humana, animal o combus-tible, para aumentar la productividad de organismos específicos.tible, para aumentar la productividad de organismos específicos.tible, para aumentar la productividad de organismos especí
2. La biodiversidad puede ser muy reducida en comparación con la de otros ecosis-temas.
3. Los animales y plantas que dominan son seleccionados artificialmente y no por selección natural.
4. Los controles del sistema son, en su mayoría, externos y no internos.
Este modelo marca un punto de partida interesante para comprender la agricultu-ra desde una perspectiva de los sistemas ecológicos, pero no abarca la diversidad y complejidad de muchos agroecosistemas que se desarrollan en los países más pobres, especialmente tropicales, y no tiene en cuenta las determinantes sociales que tanto influyen en estos países. Las estrategias agrícolas no solo responden a presiones del medio ambiente, a presiones bióticas y del proceso de cultivo, sino que también refle-jan estrategias humanas de subsistencia y condiciones económicas (Ellen, 1982).
Para complementar las características expresadas por Odum surge la concepción de los agroecosistemas sostenibles o sustentables que, de acuerdo con la definición del Co-mité Internacional sobre Investigación Agrícola (1988), consiste en “el manejo exitoso de los recursos agrícolas para satisfacer las necesidades humanas, mientras se mantiene la calidad del ambiente y se conservan los recursos naturales”.
En 1989, el Consejo Nacional de Investigación de la Academia de Ciencias de los Estados Unidos publicó un significativo reporte sobre “Agricultura Alternativa”, la cual fue definida como “un sistema de producción de alimentos y fibras que aplica tareas de manejo y de información encaminadas a reducir costos, mejorar la eficiencia y mante-ner niveles de producción mediante prácticas de rotación de cultivos, apropiada inte-gración de cultivos y ganado, utilización de leguminosas fijadoras de Nitrógeno, manejo integrado de plagas, prácticas de conservación, reciclaje de residuos agrícolas como acondicionadores de suelos y utilización de biofertilizantes”.

19la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
El reporte propugnaba la adopción colectiva de estas prácticas por los agricultores norteamericanos, como la mejor alternativa frente al uso intensivo y continuado de plaguicidas y fertilizantes químicos, que habían causado la pérdida de calidad de suelos y aguas y contaminado los alimentos.
Los principios básicos que rigen un agroecosistema sustentable son: la conservación de los recursos renovables, la adaptación del cultivo al ambiente y el mantenimiento de un nivel alto de productividad. En este tipo de agricultura, debe ponerse énfasis en la sustentabilidad ecológica a largo plazo más que en la productividad a corto plazo, para lo cual deben cumplirse los siguientes objetivos (Altieri, 1997):
– Reducir el uso de energía y recursos.
– Emplear métodos de producción que restablezcan los mecanismos que conducen a la estabilidad de la comunidad.
– Optimizar el reciclaje de materia y nutrientes y asegurar un flujo eficiente de energía.
– Utilizar al máximo la capacidad multiuso del sistema.
– Disminuir los costos.
Para lograr estos objetivos hay que desarrollar una estrategia que destaca, en primer lugar, la conservación y el manejo de los recursos agrícolas locales, la participación de los agricultores y la adaptación de tecnologías a las condiciones de la región donde se trabaje. En esta estrategia hay que incluir la conservación y regeneración de los recursos naturales, el manejo de los recursos productivos, la regulación biótica como medida de protección de los cultivos y la reutilización de nutrientes y recursos naturales externos e internos del sistema. Para poder cumplir con efectividad esta estrategia, se hace imprescindible la utilización de los biofertilizantes microbianos.
Los conceptos que se han expresado anteriormente han calado muy hondo en las poblaciones de todos los países desarrollados, que han sido los principales impulsores de la Revolución Verde y hoy están convencidos de los graves problemas que se han generado con estos métodos de producción agrícola. Por esta razón, cada año se multiplican los mercados donde se venden exclusivamente productos orgánicos y son mayores las superficies dedicadas a los cultivos agroecológicos en los distintos países. En la Tabla 1 se muestran las superficies de cultivos orgánicos en los países europeos (Hoffman, 2001).

20 capítulo 1. la fijación biológica del nitrógeno atmosférico como uno de los pilares básicos de la agroecología
Importancia de los biofertilizantes dentro de la Agroecología
La sustentabilidad de los sistemas agrícolas a largo plazo debe fomentar el uso y ma-nejo efectivo de los recursos internos de los agroecosistemas. Entre estos recursos, la biota del suelo realiza una serie de funciones que son esenciales para la integridad y productividad de los sistemas agrícolas, por lo que constituye una fracción primordial de la biodiversidad terrestre al contribuir a la descomposición de la materia orgánica, al reciclaje de los nutrientes, a la modificación de la estructura del suelo, a la regulación de la composición atmosférica, al control biológico de plagas y enfermedades y a la degra-dación de compuestos xenobióticos (Palm et al., 2001). La composición de esta biota puede ser manipulada, casi siempre de forma temporal, para mantener e incrementar la productividad de un suelo.
En este sentido, los biofertilizantes y bioestimuladores microbianos representan un componente vital de los sistemas sustentables, ya que constituyen un medio econó-micamente atractivo y ecológicamente aceptable de reducir los insumos externos y mejorar la cantidad y calidad de los recursos internos mediante la utilización de mi-croorganismos del suelo debidamente seleccionados, capaces de aportar a los cultivos Nitrógeno fijado de la atmósfera, Fósforo transformado a partir del que está fijado en el suelo y sustancias fisiológicamente activas que, al interactuar con la planta, desenca-
Tabla 1
Superficie agrícola dedicadaa cultivos orgánicos
en distintos paíseseuropeos
en el año 2001
País SuperficieAlemania 546.023Austria 287.900España 352.164Francia 420.000Gran Bretaña 472.500Holanda 27.820Italia 1.040.373Suecia 174.423Suiza 95.000Total 3.416.203

21la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
denan una mayor activación del metabolismo vegetal (Bauer, 2001; Burdman et al., 2000).
Los biofertilizantes y bioestimuladores microbianos pueden definirse como productos a base de microorganismos que viven normalmente en el suelo, aunque en poblacio-nes bajas, y que, al incrementar sus poblaciones por medio de la inoculación artificial, son capaces de poner a disposición de las plantas, mediante su actividad biológica, una parte importante de los nutrientes que necesitan para su desarrollo, así como de suministrar sustancias hormonales o promotoras del crecimiento. En un sentido am-plio, estos términos pueden usarse también para incluir todos los recursos orgánicos necesarios para el desarrollo de las plantas, los cuales son transformados mediante la acción de los microorganismos. La importancia de estos bioproductos radica en su capacidad para suplementar o movilizar nutrientes con un mínimo uso de recursos no renovables; además, tiene la ventaja de que los procesos microbianos son rápidos y los biopreparados pueden aplicarse en pequeñas unidades para solucionar problemas locales específicos. locales específicos. locales especí
A este respecto, se está imponiendo en el mundo el redimensionamiento de las bio-tecnologías y, entre ellas, los biofertilizantes, con el objetivo de adaptar estas novedosas tecnologías al desarrollo sustentable de las localidades más pobres y necesitadas del planeta, de manera que se ayude a solucionar la problemática productiva del campesi-no pobre y a lograr que éste adquiera una mayor independencia y seguridad.
Los inoculantes microbianos pueden considerarse como biotecnologías “apropiables”, término creado para las herramientas biotecnológicas que contribuyen al desarrollo sustentable por ser técnicamente factibles dentro del nivel científico- técnico de un país y que proveen beneficios tangibles a los destinatarios, son ambientalmente seguras y socioeconómicas y culturalmente aceptables (Izquierdo et al., 1995).
También responden a las exigencias de la Agenda 21, firmada por los Jefes de Estado y Gobierno de todos los países del mundo como parte de la Reunión sobre medio Ambiente y Desarrollo, llamada también “Cumbre de la Tierra”, celebrada en Río de Janeiro (Brasil) en 1992, como se deduce de los siguientes acápites del Capítulo 3 de la citada Agenda 21:
3.1 Encontrar sustitutos o mejoras ecológicamente racionales de los procesos de producción que son nocivos para el medio ambiente.

22 capítulo 1. la fijación biológica del nitrógeno atmosférico como uno de los pilares básicos de la agroecología
3.2 Determinar métodos encaminados a reducir a un mínimo la necesidad de insu-mos químicos sintéticos insostenibles y para utilizar al máximo aquellos productos ecológicamente adecuados, incluidos los naturales.
3.9 Promover la utilización adecuada de los biofertilizantes en todos los programas nacionales de aplicación de fertilizantes.
3.12 Desarrollar nuevas tecnologías que permitan la selección rápida de organismos que puedan tener propiedades biológicamente útiles.
3.16 Elaborar mecanismos para incrementar gradualmente y difundir biotecnologías ecológicamente racionales de gran importancia para el medio ambiente, espe-cialmente a corto plazo, aún cuando estas tecnologías tengan potencial limitado.
En general, el uso de los biofertilizantes microbianos en los sistemas productivos es una alternativa viable y de gran importancia para lograr un desarrollo agrícola ecológi-camente sustentable, ya que permite una producción de bajo costo, no contamina el ambiente y mantiene la conservación del suelo desde el punto de vista de fertilidad y biodiversidad.
A pesar de su inocuidad, el uso masivo de los inoculantes microbianos ha desperta-do algunas preocupaciones acerca del posible impacto ecológico de la liberación de estos microorganismos en el medio ambiente, en especial cuando ha existido una manipulación genética. La evaluación de dicho posible impacto es muy difícil, debido a limitaciones de tipo técnico que impiden hasta el momento determinar si la alteración de algún parámetro medioambiental como consecuencia del uso de inoculantes puede afectar de forma negativa al equilibrio futuro de los ecosistemas del suelo (Sanjuan et al., 2004).
De todas maneras, en las condiciones tropicales de Cuba, y en diversos cultivos, se ha demostrado que las poblaciones de Azotobacter chroococcum, que después de la inoculación alcanzaron niveles de 109 células/g de suelo rizosférico, recuperan su nivel natural (104-105) en un período máximo de 4 meses, según el cultivo. Es decir, que no hay una contaminación a largo plazo, debido al empobrecimiento de las secreciones radiculares por el envejecimiento de las plantas y al antagonismo de otras poblaciones microbianas del suelo (Martínez Viera et al., 2004).

23la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Importancia de los biofertilizantes basadosen bacterias fijadoras del N2 atmosférico
Los primeros estudios que se realizaron acerca de lo que hoy se llama biofertiliza-ción estuvieron relacionados con la fijación biológica de Nitrógeno atmosférico y se remontan a la primera mitad del siglo XIX. Esto es muy importante si se toma en consideración que el Nitrógeno es el mayor nutriente que limita el desarrollo de las especies vegetales, siendo superado solamente por la fotosíntesis en lo que respecta a su influencia sobre el desarrollo. A pesar de que este elemento constituye el 70% de la atmósfera, no puede ser utilizado por las plantas, que se ven obligadas a utilizar las for-mas nitrogenadas combinadas que se encuentran en el suelo, en cantidad insuficiente para soportar cultivos intensivos.
La fijación biológica puede contribuir globalmente con más de la mitad del Nitrógeno necesario para el cultivo de las plantas, siempre que se maneje de forma adecuada. Debido a este importante papel, hoy se concibe el manejo efectivo del Nitrógeno que existe en el ambiente como uno de los pilares básicos de la sustentabilidad agrícola.
Los resultados de los estudios que se han realizado en los últimos 25 años han revo-lucionado el conocimiento sobre la fijación biológica del N2 atmosférico y sobre los beneficios que puede suministrar. A pesar de eso, muy pocos de los conocimientos adquiridos están siendo aplicados por los agricultores en la mayor parte de los países subdesarrollados, a pesar de que en las condiciones tropicales existen temperaturas constantemente favorables y hay una alta fijación de Carbono, que alcanza hasta 20 t/ha/año (Debinstein, 1970), lo que garantiza la presencia de altas poblaciones de mi-croorganismos en la zona rizosférica de las plantas.
Por los motivos que se han expuesto, los únicos biofertilizantes que son utilizados extensamente en muchos países son los que contienen las bacterias fijadoras de los géneros Rhizobium y Bradyrhizobium,y Bradyrhizobium,y que establecen la fijación simbiótica de N2 en las leguminosas. Pero no se está aprovechando la actividad de los fijadores asociativos ni de los organismos solubilizadores de Fósforo o estimuladores del crecimiento y el rendimiento, que tantos beneficios pueden reportar en las regiones tropicales, donde se encuentran los países más necesitados.
Hoy ya se está adquiriendo conciencia de que la aplicación en la práctica agrícola tropi-cal de los conocimientos que hoy existen sobre la fijación biológica de N2 atmosférico

24 capítulo 1. la fijación biológica del nitrógeno atmosférico como uno de los pilares básicos de la agroecología
permite lograr importantes impactos en lo que se refiere a productividad y economía de los cultivos, así como en la reducción de los niveles de contaminación ambiental. Por estas razones, deben elaborarse estrategias encaminadas a lograr la adaptación de las tecnologías existentes a las condiciones agroecológicas de las distintas regiones del país, con la participación de investigadores, docentes, técnicos y estudiantes, cuyos esfuerzos deben coordinarse para que pueda cumplirse este objetivo.
Al elaborar estas estrategias, hay que considerar que la fijación de N2 no puede con-siderarse como una práctica aislada, puesto que la productividad de un cultivo y la productividad a largo plazo de un agroecosistema dependen de múltiples variables, que implica distintos tipos de factores, incluyendo el manejo humano. Por ejemplo, hay que considerar las rotaciones de cultivos o aprovechar las interacciones leguminosas-no leguminosas en cultivos simultáneos, de manera que se optimicen los procesos de fijación y sus beneficios para todos los sistemas productivos; es necesario seleccionar cepas eficientes y competitivas frente a organismos que ya están establecidos en el suelo, así como que sean capaces de adaptarse a distintas condiciones de stress; deben utilizarse métodos de inoculación adecuados para las características de cada agroecosis-tema y para las especies vegetales que quieren cultivarse; las técnicas agronómicas que se apliquen deben facilitar el establecimiento de los inóculos y permitirles desarrollar su actividad fijadora.
Además, hay que considerar muchos aspectos más para los que es necesario preparar al agricultor, organizando talleres, demostrando los efectos de estos microorganismos mediante su aplicación en parcelas demostrativas y con un adecuado trabajo de exten-sión agrícola que permitirá que los agicultores obtengan los mayores beneficios con la utilización de los biopreparados a base de bacterias fijadoras de Nitrógeno atmosférico.
En los capítulos siguientes se ha expuesto la situación actual del conocimiento relacio-nado con la fijación biológica del N2 atmosférico y se incluyen numerosos ejemplos de los beneficios que pueden obtenerse con su utilización, poniéndose de manifiesto la importancia ecológica y económica del uso de los inoculantes a base de bacterias fijadoras. Hoy se están incrementando los niveles de aplicación en muchos países con notable éxito, y los beneficios que se están logrando con la generalización de las nuevas tecnologías se van aproximando con rapidez a las potencialidades que han sido calcu-ladas por distintos autores.

25la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Capítulo 2Papel de la rizosfera en la efectividadde los biofertilizantes microbianos
Rizosfera y ambiente rizosférico
La capacidad de los microorganismos para suministrar nutrientes y estimular el creci-miento de las plantas depende de su exitoso establecimiento sobre las raíces (Zuberer, 1990). Por estas razones, se hace obligado explicar el papel que desempeña la rizosfera y su gran importancia para lograr resultados efectivos con la aplicación de los bioferti-lizantes, sobre todo si se toma en cuenta que la asociación planta-microorganismo es extremadamente compleja, debido a la cantidad de factores que influyen en el estable-cimiento de la asociación y, por ende, en la mayor o menor efectividad, y hasta en la inefectividad, de la inoculación.
La primera definición de rizosfera fue dada por Hiltner en 1904 considerándola como el volumen de suelo que recibe influencia de la raíz. Sus deducciones estaban basadas en los resultados de estudios realizados en macetas y en condiciones de campo, en-caminados a conocer los efectos que la utilización de abonos verdes en cultivos de leguminosas tenían sobre la fertilidad del suelo. Sus observaciones demostraron que ocurrían una serie de procesos en la interfase suelo-raíces, entre los que se encontra-ban los siguientes:
1. Existe un volumen de suelo que está ocupado por raíces y microorganismos.
2. Los materiales exudados por las raíces de distintas leguminosas atraen a organis-mos diferentes a los que son atraídos por plantas no leguminosas.
3. Cada especie de leguminosa atrae a microorganismos que tienen un beneficio específico para esa especie.
A partir del momento en que Hiltner divulgó sus conclusiones se realizaron numerosos estudios que han puesto de manifiesto la diferente influencia de las raíces de distintas especies vegetales sobre la masa del suelo, así como los cambios estacionales en el desarrollo de las raíces (Watson et al., 2000). Por ejemplo, la masa promedio de raí-

26 capítulo 2. papel de la rizosfera en la efectividadde los biofertilizantes microbianos
ces oscila entre 0.2 kg/m2 en las plantas cultivadas y 5 kg/m2 en las especies forestales (Jackson et al., 1996). Pero, además, hay una constante renovación de raíces y de sus secreciones, todo lo cual influye sobre las poblaciones de microorganismos que se encuentran en su zona de influencia. En lo que se refiere a la profundidad de las raíces, se ha reportado para las hortalizas entre 0.65 y 1.6 m (Merril et al., 2002) y entre 2 y 10 m para las especies forestales (Carradell et al., 1996).
Hoy, tomando como base el conocimiento adquirido en las últimas décadas, el con-cepto de rizosfera es mucho más amplio y abarca a la zona de interacción única y diná-mica entre las raíces de las plantas en general y los microorganismos del suelo, lo que origina la formación de una región especializada que se caracteriza por el aumento de la biomasa microbiana y de la microfauna y de su actividad (Steciow, 2004). Tomando como base estos conocimientos, la rizosfera puede definirse como una zona bioló-gicamente activa del suelo con un alto contenido de Carbono, debido a la continua exudación y rizodeposición de las raíces de las plantas. Esta zona puede extenderse entre 1 mm y más de 1 cm, en dependencia de la especie vegetal y de la humedad y textura del suelo (Pinton et al., 2001).
Características de la rizosfera
La zona de la rizosfera es diferente al resto del suelo, debido a los numerosos pro-cesos biológicos, bioquímicos, químicos y físicos que ocurren como consecuencia del crecimiento de las raíces, los movimientos de agua y de nutrientes tomados por ellas, la respiración y la rizodescomposición (Hinsinger et al., 2005). Como consecuencia de estos procesos, las características de esta región son totalmente diferentes de las del resto del suelo: ocurre disminución o aumento de 2 unidades de pH, es 100 veces ma-yor la actividad reductasa y entre 2 y 8 veces más alta la actividad fosfatasa; además, la concentración de azúcares aumenta 2-3 veces y la de aminoácidos 10; la acumulación de Calcio es 10 veces mayor, ocurriendo precipitación, y 6 veces más alta la de NaCl (Römheld y Neumann, 2006).
Lynch (1990) propuso la división de la rizosfera en: 1) endorizosfera, que se extiende desde la superficie de las raíces hasta los primeros estratos celulares internos, y com-prende a tejidos conductores como xilema y floema, a la endodermis, la epidermis y el extremo de la raíz; 2) rizoplano, que comprende la superficie externa de las raíces; 3) ectorizosfera, que consiste en el volumen de suelo en contacto inmediato con las

27la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
raíces y contiene los pelos radicales, muscílagos exudados por la planta y los microorga-nismos y restos de células de la raíz. Se concibe también el término espermosfera para nombrar a la zona que rodea a la semilla en estado de germinación, donde la microflo-ra desarrolla una intensa actividad que afecta el futuro desarrollo de la planta. En los últi-mos tiempos se considera que en esta fase comienza realmente la actividad rizosférica. Es importante señalar que los microorganismos que colonizan la superficie de la semilla pueden tener una notable importancia en la definición de la población microbiana que posteriormente se establecerá en la rizosfera de una planta determinada.
La rizosfera constituye un microambiente complejo y dinámico, cuyo volumen y acti-vidades dependen de las características específicas del suelo y de la especie vegetal, así como de la edad y vigor de esta última. Constituye un ejemplo de equilibrio biológico que se establece gracias a las relaciones de tipo sinérgico o de comensalismo entre la microflora y las raíces de las plantas, y entre ellas y las características inorgánicas del suelo. La causa principal de la influencia de la rizosfera sobre las comunidades que vi-ven en esta zona consiste en la deposición de los compuestos derivados de las raíces, los cuales se clasifican en exudados (procesos pasivos), secreciones (procesos activos), mucigel (mezcla de las raíces con subproductos microbianos) y lisados (contenido de las células que se rompen) (Rovira, 1969).
El estudio del sistema rizosférico es complejo, no solo por el alto número de inte-racciones que en él se dan, sino por la escala espacial y temporal de los elementos y procesos. El ambiente rizosférico es el resultado de las distintas interacciones entre la planta, los microorganismos y el suelo. La planta influye de distintas maneras, en de-pendencia de su estadio, de su estado nutricional y sanitario y de las características de sus secreciones y residuos; el suelo lo hace de acuerdo con su estructura y su conte-nido de nutrientes y de agua, y los microorganismos intervienen según el nivel de sus poblaciones, las relaciones entre las distintas especies y el aporte de sustancias nutritivas (Curt y Truelover 1986; Killham, 1999).
En las plantas que crecen en el suelo, la rizosfera está caracterizada por gradientes que se establecen de forma radial y longitudinal a lo largo de la raíz individual. Estos gradien-se establecen de forma radial y longitudinal a lo largo de la raíz individual. Estos gradien-se establecen de forma radial y longitudinal a lo largo de la raítes pueden existir para los nutrientes minerales, el pH, el potencial redox y los proce-sos reductores, los exudados radicales y la actividad microbiana (Perret et al., 2000).

28 capítulo 2. papel de la rizosfera en la efectividadde los biofertilizantes microbianos
La cantidad y los tipos de sustratos en la rizosfera son sustancialmente distintos de la masa normal del suelo en lo que respecta a las condiciones físico-químicas, como son acidez, humedad, conductividad eléctrica, potencial redox y otros (Cocking, 2003). Estas características, unidas a la liberación de numerosas sustancias orgánicas a través de las secreciones de las raíces, estimulan la llegada y el desarrollo de diferentes pobla-ciones de bacterias, hongos y actinomicetos y de diversos componentes de la micro y mesofauna (protozoos, nematodos, microartrópodos). Las secreciones están forma-das por una gran diversidad de compuestos orgánicos volátiles y solubles de bajo y alto peso molecular, los cuales tienen entre sus funciones la de servir como atrayentes de los organismos citados anteriormente, como reservorio de los nutrimentos necesarios para estos organismos y como fuente de señales genéticas reguladoras.
Como consecuencia de todo lo que se ha expresado, en el suelo que se encuentra muy cerca a las raíces aumenta notablemente la población de organismos, lo que se conoce como efecto rizosfera, el cual tiende a disminuir con la profundidad del suelo debido a que las raíces son más numerosas en las proximidades de la superficie, por lo que puede decirse que el efecto rizosfera constituye en lo esencial un fenómeno de los estratos más superficiales del suelo.
A su vez, la población de microorganismos resultante de este efecto es capaz de ejer-cer una influencia muy positiva sobre las plantas por distintos mecanismos, ya que en esta zona desarrollan una gran variedad de actividades, como son la fijación del Nitró-geno atmosfégeno atmosfégeno atmosf rico, la solubilización del Fósforo retenido en el suelo, la estimulación del crecimiento de las plantas, el establecimiento de simbiosis mutualísticas, el biocontrol, la antibiosis, el mejoramiento de la estructura del suelo, la liberación de enzimas y el incremento de la retención de agua.
Como un ejemplo, puede citarse la síntesis de reguladores del crecimiento, tales como auxinas, giberelinas y citoquininas, que estimulan la actividad metabólica de las raíces (Burdman et al., 2000). Si se toma en cuenta, además, el beneficio recibido por la fijación de Nitrógeno, que es sinergístico con la estimulación del crecimiento, puede comprenderse por qué se incrementa el uso de los biofertilizantes y bioestimuladores en el mundo, con el objetivo de reducir las aplicaciones de fertilizantes inorgánicos (Lambrecht et al., 2000).

29la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Por otra parte, el concepto de colonización microbiana de la rizosfera comprende tres fases (Merckx et al., 1993): una primera etapa, donde los microorganismos migran en dirección a las raíces, atraídos por el efecto de las sustancias que componen los exuda-dos, y comienzan a distribuirse paralelamente al crecimiento de la raíz, con disminución de la población a medida que ésta se aleja de la zona germinada. Esta migración es po-sible porque ocurre un intercambio de señales entre las raíces y los microorganismos
En una segunda fase se produce la supervivencia y la multiplicación de las poblaciones microbianas hasta el límite del nicho ecológico; en este período ocurre una intensa competencia entre los organismos que están siendo atraídos y los que ya se encuentran establecidos, y esta competencia tiene lugar durante y después de la primera etapa.
En una tercera fase ocurre la distribución de la población a lo largo de las raíces, su multiplicación y establecimiento. Pero los microorganismos no están distribuidos de manera uniforme sobre la superficie de la raíz, sino que se localizan en determinados puntos (microambientes) de la misma que poseen mayor actividad segregante de ma-teria orgánica (exudados radicales). En general, se considera que solo entre 4 y 10 % de la superficie de la raíz está colonizada por microorganismos (Metting, 1993).
El sistema radical de las plantas ejerce varios efectos sobre la micropoblación edáfica; así, hay efectos indirectos, producidos por la actividad de las raíces al modificar la es-tructura del suelo, el régimen hídrico, el pH, la concentración de elementos nutritivos y la composición de la atmósfera del suelo. También hay efectos directos causados por la exudación de diversas sustancias (azúcares, fenoles, vitaminas, aminoácidos, ácidos orgánicos y grasos, nucleótidos, esteroides, etc.) que, junto con la decamación de cé-lulas y tejidos, estimulan la microflora al actuar como fuentes de energía, de Carbono, Nitrógeno, Fósforo, etc., o la inhiben por la liberación de sustancias tóxicas.
Entre los factores abióticos, los que más influyen son el suministro de Carbono, la humedad y la aireación. Los microorganismos deben de tener acceso a abundantes fuentes de Carbono para su crecimiento y la producción de energía, sobre todo en el caso de los fijadores de Nitrógeno, ya que la fijación de una molécula de N2 requiere aproximadamente 16 moléculas de ATP, por lo que los organismos deben utilizar con-siderables cantidades de sustratos.
La falta o exceso de humedad limita la vida microbiana en el suelo y, como es natural, también en la zona rizosférica. El exceso influye sobre todo en la capacidad de aireación y es más perjudicial que la falta de humedad, ya que algunos de los organismos rizosfé-

30 capítulo 2. papel de la rizosfera en la efectividadde los biofertilizantes microbianos
ricos son capaces de formar quistes (Azotobacter)ricos son capaces de formar quistes (Azotobacter)ricos son capaces de formar quistes ( o esporas (Bacillus, Clostridium) que les permiten sobrevivir durante largos períodos a la desecación.
La aireación ejerce un efecto muy marcado, ya que las bacterias aeróbicas y microae-rofílicas sufren bajo condiciones anóxicas. A pesar de esto, los fijadores de Nitrógeno funcionan mejor a concentraciones reducidas de Oxígeno, debido a la sensibilidad del complejo nitrogenasa al Oxígeno molecular, el cual inactiva de forma irreversible a la enzima (Postgate,1987). En general, en la rizosfera existen numerosos ecositos con bajo pO2 que proporcionan condiciones favorables para el establecimiento de los dia-zotrofos (Zuberer, 1990).
Influencia de las secreciones de las raícessobre los microorganismos
Como se ha dicho antes, una de las principales características de la rizosfera consiste en lo que se llama rizodeposición, relacionada con el flujo del Carbono fotosintético que se realiza desde las raíces al suelo en forma de exudados de bajo peso molecular (azúcares, ácidos orgánicos, aminoácidos y fenoles), exudados de alto peso molecu-lar (macropolímeros y ectoenzimas) y mucigel (mucílagos mezclados con la población microbiana que puebla el rizoplano, con metabolitos y con células epidérmicas en senescencia).
Los exudados de bajo peso molecular favorecen la toma por la planta de los cationes con valor nutritivo a partir del complejo de intercambio edáfico, para lo que deben cumplir la función de agentes acidificantes; también pueden emitir señales químicas que estimulan la migración de los microorganismos hacia las raíces o facilitar la expresión de determinados genes, como los que inducen la nodulación (Perret et al., 2000). El complejo mucopolisacárido tiene acción lubricante sobre las raíces y mejora de esta forma las interacciones con las partículas del suelo, sobre todo en el caso de suelos se-cos, al mismo tiempo que protege de la deshidratación al área apical de la raíz y regula la toma de nutrientes. Las enzimas que tienen más importancia en estos exudados son las fosfatasas y la polifenoloxidasa, necesarias para la síntesis de las células de las raíces (Merschner, 1999).
La cantidad y tipo de exudados producidos por las plantas están muy estrechamente relacionados con los factores que afectan su propio desarrollo, tales como tempera-

31la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
tura, intensidad y calidad de la luz y contenido de nutrientes en el suelo. Mishustin y Shilnikova (1971) expresaron que la exosmosis puede representar el 5% de la materia orgánica sintetizada por las plantas. Para una cosecha de trigo de 2 t/ha, la biomasa vegetal se calcula en 5 toneladas y, de ella, la exosmosis representa 250 kg/ha. Por su parte, Newman (1985) calculó que los exudados solubles de las raíces estaban en una concentración entre 20-50 mg/g, y contribuyen con 3-15% de la materia orgánica que éstas aportan al suelo, ya que la mayor parte del carbono translocado a las raíces ter-mina como fracciones insolubles, especialmente polisacáridos estructurales (celulosas, hemicelulosas y otras). Pero la profundización de los métodos científicos ha permitido conocer, mediante la incubación de plantas totales con 14CO2, que 20% de los fotosin-tatos son liberados como materia orgánica en la rizosfera (Cocking, 2003).
Como ya se expresó, entre las secreciones de las raíces de cada especie vegetal hay distintos tipos de sustancias orgánicas, algunas de las cuales pueden ayudar al estableci-miento de las bacterias en la zona rizosférica, mientras que otras pueden actuar como repelentes. Así, por ejemplo, en el caso del maíz los exudados que predominan son azúcares (65%), ácidos orgánicos (33%) y aminoácidos (2%), aunque la diversidad es amplia y se han encontrado en este cultivo más de 10 tipos de ácidos orgánicos y 4 tipos de azúcares (Kraffczyk et al., 1984).
Se ha demostrado que las células epidérmicas de las raíces están recubiertas de polisa-cáridos de doble origen, vegetal y microbiano, en los cuales se encuentran asentadas las colonias de microorganismos. De esta manera, los azúcares, aminoácidos, ácidos orgánicos etc., que han sido secretados son responsables del número de microorga-nismos, así como de su composición por especies y variedades. Uno de estos com-ponentes puede ser estimulante del crecimiento de un microorganismo y puede ser neutral o inhibidor para otros. Por ejemplo, los compuestos flavonoides exudados por las raíces de las leguminosas, si se añaden al medio de cultivo, estimulan el crecimien-to de Bradyrhizobium japonicum, pero disminuyen el de Pseudomonas sp. Igualmente, algunas gramíneas, así como algunas sustancias excretadas por los microorganismos, pueden ser inhibidores.
Quimiotaxismo
Debido a esta diversidad, algunas cepas de una bacteria determinada se establecen mejor sobre algunas especies vegetales, e incluso sobre algunas variedades de una mis-

32 capítulo 2. papel de la rizosfera en la efectividadde los biofertilizantes microbianos
ma especie, que sobre otras. Estas diferencias se atribuyen tanto a las características de las plantas hospedantes como a las de las propias cepas microbianas. En este sentido, la cantidad y calidad de las fuentes de carbono que segregan las raíces desempeñan un papel fundamental en el establecimiento de las bacterias (Boddey y Döbereiner, 1988).
Lo expresado anterirmente debe ser comprendido por los investigadores y fabricantes de biofertilizantes, y la no comprensión de este principio fundamental ha sido la causa de la inconsistencia de los resultados obtenidos por distintos investigadores, y hasta del fracaso de una política racional de aplicación de biofertilizantes. Por esta razón, es necesario realizar una correcta selección de la cepa más adecuada para cada especie vegetal, para lo que debe de tomarse en cuenta que la mayor o menor influencia de las secreciones de las raíces sobre las bacterias ocurre por procesos quimiotáxicos.
La quimiotaxis es el movimiento de los organismos vivos hacia o en contra de una sustancia química determinada. Este fenómeno ha sido observado en una amplia gama de animales, vegetales y microorganismos. En el caso de las bacterias, la quimiotaxis se conoce desde finales del siglo XIX, cuando Engelmann, Pfeffer, Beijerinck y otros finales del siglo XIX, cuando Engelmann, Pfeffer, Beijerinck y otros fdescubrieron que esta reacción, positiva o negativa, ocurría cuando algunas bacterias se enfrentaban al Oxígeno, a minerales y a nutrientes orgánicos. Cuando la bacteria tiene una orientación positiva en dirección a la fuente de estimulación el movimiento se llama topotaxis, y cuando es negativa, evitando la fuente, se conoce como fobotaxis (Adler, 1972).
En el caso de las bacterias fijadoras de Nitrógeno atmosférico, se ha demostrado clara-mente la quimiotaxis en varias especies de Rhizobium, aunque ocurre en general en to-das las bacterias que actúan en la rizosfera. En la tabla 2 puede verse el resultado de una selección hecha en una cámara de quimiotaxismo, en la cual se colocaron secreciones de las raíces de cebolla y un cultivo líquido de la cepa de Azotobacter chroococcum que se quería probar (Dibut, 2000).
Como se observa en la tabla, las cepas INIFAT-17 e INIFAT-3 son las más atraídas por los exudados de la cebolla. Al considerar también los resultados del ARA (prueba de reducción del acetileno), que indicaron una mayor eficiencia de fijación de nitrógeno atmosférico en la cepa INIFAT-17, fue seleccionada ésta para su uso comercial en la cebolla, con la garantía de que se establecerá una abundante población de la bacteria

33la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
en la zona rizosférica de las plantas y éstas recibirán los beneficios de la inoculación. Mediante este procedimiento, fue seleccionada la cepa INIFAT-12 para el tomate y otras solanáceas y la INIFAT-9 para las gramíneas.
Diferencias entre el efecto de las condicionesde clima templado y tropical sobre las interaccionesplanta-microorganismos en la zona rizosférica
Todas las interacciones que ocurren entre las plantas y los microorganismos están go-bernadas por las condiciones del ambiente, por la naturaleza, estado fisiológico y vigor de las plantas en desarrollo, por las características del suelo, el régimen hídrico y el manejo agronómico; igualmente, los microorganismos interaccionan entre ellos, dan-do lugar a procesos de naturaleza antagónica o sinérgica, muy importantes para esta relación.
La mayor parte de los conocimientos que se han adquirido sobre estas interacciones son fruto de las investigaciones realizadas en las condiciones de clima templado. Pero, en las regiones tropicales, las excreciones de las raíces y las hojas tienen mayores concentraciones de compuestos carbonados que en las regiones templadas debido a la mayor fijación de Carbono, lo cual trae como consecuencia la presencia de pobla-ciones más altas de microorganismos en la rizosfera y en la filosfera (zona de las hojas) de las plantas en el trópico.
INIFAT -17 INIFAT-15 INIFAT-3 INIFAT-10Variantes Logn Factor Logn Factor Logn Factor Logn Factor
X C X C X C X C
Exudados 7.87 3584 5.03 613 7.84 3287 5.11 660
Control (buffer) 4.32 - 4.25 - 4.30 - 4.24 -
Tabla 2
Efecto quimiotáxico de los exudados totales de las plantas de cebolla sobre distintas cepas de A. chroococcum (expresado en logn de X).
Factor C= Factor quimiotaxis. Relación de células atraídas por los exudados/cé-lulas atraídas por buffer de fosfatoX= Número de células atraídas

34 capítulo 2. papel de la rizosfera en la efectividadde los biofertilizantes microbianos
La baja concentración de compuestos carbonados puede ocasionar que las raíces de las plantas en las regiones templadas tengan menor influencia sobre el desarrollo de algunos organismos fijadores de nitrógeno de forma asociativa, como es el caso de las especies de Azotobacter, ya que estas bacterias se encuentran ausentes, en muchos ca-sos, de la rizosfera de especies vegetales de tanta importancia económica como avena y trigo, y ni siquiera mediante la inoculación es posible lograr que se establezcan en la zona rizosférica de estas especies (Becking, 1977; Knowles, 1981).
La introducción en el suelo de un organismo foráneo que ha demostrado aportar grandes beneficios en condiciones axénicas puede tener problemas en condiciones naturales, y hasta puede no entrar a formar parte de la microflora del suelo. En una revisión de la literatura puede verse una elevada cantidad de fallos en la inoculación. Esto se debe a que existe todavía un conocimiento muy limitado de la rizosfera, lo que impide poder modificar las condiciones de forma adecuada.
Por ejemplo, Rovira (1965) reportó que, en los suelos cultivados de las regiones tem-pladas, se encuentra Azotobacter en niveles muy bajos, y con frecuencia estas bacterias son eliminadas de la rizosfera, especialmente de la superficie de las raíces. También Trolldenier (1977) encontró, mediante el uso de microscopía de fluorescencia, que la rizosfera del trigo contiene solo unas pocas células de estas bacterias.
La pobre incidencia de Azotobacter se atribuye a la falta de sustratos utilizables y a la acumulación de sustancias bacteriostáticas por la presencia de numerosos microorga-nismos antagonistas. En cambio, los reportes de las zonas áridas calientes indican una elevada incidencia, con poblaciones que alcanzan hasta 100 millones de células/g de suelo rizosférico de trigo, algodón y leguminosas (Abd-El-Malek, 1971). Las plantas xerofíticas también pueden estimular las poblaciones de Azotobacter, desde 10 células/ g en los suelos desérticos hasta 1 millón en la rizosfera de algunas plantas cultivadas en esos mismos suelos (Wullstein et al., 1979).
En contraposición a lo expresado por Rovira (1965) y Trolldenier (1977) para los sue-los templados, en un país tropical como Cuba pueden aprovecharse los beneficios de la inoculación del trigo con Azotobacter, como se demuestra en la Tabla 3 (Dibut et al., 1996).
Como puede verse en la tabla, la combinación de una cantidad reducida de fertilizante con el biofertilizante produjo los rendimientos más elevados, lo que indica que hubo

35la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
una marcada influencia de la bacteria, la cual se estableció en la zona rizosférica de las plantas y aportó el nitrógeno fijado y las sustancias activas estimuladoras del crecimiento Frioni (1999) demostró que el efecto rizosférico de plantas de sorgo, maíz, mijo perla (Pennisetum americanum) y mijo común (Panicum miliaceum) influyó marcadamente sobre las distintas poblaciones de Azotobacter, Azospirillum y Clostridium, así como en la actividad nitrogenasa de cada una de ellas. El efecto más marcado ocurrió en la fase de grano al estado lechoso en todos los cultivos.
En los últimos tiempos se realizan numerosos estudios sobre ecología de la rizosfera, enfocados desde una perspectiva dinámica en la que se incluyen las interacciones entre los microorganismos y entre estos y las plantas, así como los mecanismos que rigen estas interacciones, muchos de los cuales están aún por descubrir.
De acuerdo con estos estudios, todo lo que se ha expuesto anteriormente, que se ha ejemplificado con el caso de las bacterias del género Azotobacter, pero que puede aplicarse a todos los microorganismos con características de biofertilizantes o bioes-timuladores en general, demuestra la importancia que tienen las secreciones de las raíces para que pueda establecerse una cepa bacteriana en la zona rizosférica de un determinado cultivo y para que se aprovechen sus efectos benéficos.
En la Tabla 4 (Martínez Viera, 1998) se pone de manifiesto la presencia de poblaciones naturales de Azotobacter en la rizosfera de todos los cultivos situados en diferentes tipos de suelos de la Huasteca Hidalguense (México).
En esta tabla hay que destacar la presencia de estas bacterias en la rizosfera del cafeto en La Puerta, a pH 4.04, lo cual contradice los datos que existen en la bibliografía y que han sido ratificados por nuestros propios datos, ya que nunca se ha podido aislar ni una
Tabla 3
Respuesta del trigoa la inoculación con A. chroococcumen las condicionesde Cuba
Variante Superficie Rendimiento Peso de 1.000 (ha) (t/ha) granos (g)
45 Kg N/ha 7.2 4.39 45.0022.5 Kg N/ha 11.8 3.58 39.00
22.5 Kg N/ha+Azotobacter 8.3 4.71 47.00

36 capítulo 2. papel de la rizosfera en la efectividadde los biofertilizantes microbianos
cepa de Azotobacter en suelos de Cuba con pH inferiores a 5.0, en zonas rizosféricas de tabaco y cítricos, que son los cultivos predominantes en estos suelos. El resultado que se muestra en la tabla sugiere que las secreciones de las raíces de algunos cultivos, en este caso del cafeto, podrían ayudar al establecimiento de estas bacterias aunque las condiciones del suelo sean perjudiciales.
Tabla 4
Poblaciones naturales de bacterias pertenecientes
al género Azotobacteren las muestras de suelos rizosféricos
recogidas en la Huasteca Hidalguense (México)
Lugar Tipo de suelo Cultivo pH del suelo UFC/g de suelo (agua)
Coacuilco 1 Rendzina Plátano átano á 6.59 2 x 104
Coacuilco 2 “ Pasto natural 6.72 3 x 104
Huitzilingo “ Cítricos ítricos í 6.48 2 x 104
Jaltocan “ Maiz 6.97 6 x 104
Tlaica “ Pasto natural 6.30 8 x 103
Ixtle Regosol Maiz 7.44 5 x 104
Tepaco “ Pasto natural 7.35 “ Pasto natural 7.35 “ Pasto na 2 x 104
La Puerta Levisol Cafeto 4.04 3 x 103
Huantla “ Maní 7.34 3 x 104
Coatzonco 1 Feozen Maíz íz í 7.33 9 x 104
Coatzonco 2 “ Maíz 7.42 3 x 104
Coatzonco 3 “ Café 7.65 1 x 105
Coatzonco 4 “ Pasto natural 7.80 8 x 104
Metlatepec “ Pasto natural 7.84 3 x 103

37la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Capítulo 3Generalidades sobre la fijacion biológicadel nitrógeno atmosférico
Importancia del Nitrógeno
El Nitrógeno es uno de los dos mayores factores, junto con el agua, que gobiernan la productividad de los cultivos y procede de tres fuentes principales: las reservas orgá-nicas e inorgánicas del suelo, los fertilizantes minerales y los abonos orgánicos que se aplican y la fijación biológica del Nitrógeno que se encuentra en la atmósfera en forma de N2. Pero las reservas del suelo son muy limitadas y los fertilizantes son económica-mente costosos y fuertes contaminantes del ambiente, por lo que la fijación biológica constituye una alternativa importante para aumentar la producción de alimentos utili-zando métodos económicos y no contaminantes.
En forma gaseosa, este elemento constituye las cuatro quintas partes de la atmósfera terrestre, y representa una reserva prácticamente inextinguible que alcanza a 4. 1015
toneladas. Se ha calculado que en la atmósfera que rodea cada hectárea de la super-ficie terrestre hay unas 70.000 toneladas de Nitrógeno, el cual no puede ser utilizado por los seres vivos porque se encuentra en forma molecular. En esta forma tiene una gran inercia química y se combina muy difícilmente con O2 e H2 para dar compuestos minerales simples como nitratos o amoniaco, que son las formas que asimilan los seres vivos para sintetizar las moléculas biológicas nitrogenadas.
Por estas razones, hace falta que el N2 sea transformado por un grupo reducido de microorganismos llamados fijadores de Nitrógeno atmosférico, o por métodos indus-triales, hasta formar compuestos amoniacales, los cuales son llevados a su vez hasta nitratos por otros microorganismos llamados nitrificantes, que efectúan el proceso de nitrificación. La forma de nitratos constituye la principal fuente nitrogenada para la nu-trición de las plantas.

38 capítulo 3. generalidades sobre la fijaciónbiológica del nitrógeno atmosférico
La fijación de N2 por métodos industriales
Hasta mediados del siglo XIX, el proceso de fijación del Nitrógeno atmosférico por parte de los microorganismos, que se había desarrollado durante millones de años, fue suficiente para suministrar el elemento necesario a los cultivos que debían de mantener a la población del planeta. Pero con el rápido aumento de la población y el crecimiento de las grandes ciudades, la demanda de suministros obligó a buscar nuevas formas de resolver las necesidades de fuentes nitrogenadas para los cultivos. Al principio se importó en Europa el guano de Perú y el salitre de Chile, formas de fertilización que fueron suplementadas con los subproductos amoniacales de la producción de gas a partir del carbón.
La creciente demanda de Nitrógeno condujo a la invención de distintos procesos in-dustriales que permitieran aprovechar, mediante la fijación, la inagotable fuente del elemento que constituía la atmósfera. Así, surgieron las tecnologías de Birkeland-Eyde en 1905, de Frank-Caron en 1906 y de Serpak en 1908, todas con altísimos requeri-mientos de energía que impidieron su desarrollo en gran escala.
Poco después de la Primera Guerra Mundial, en 1919, surgió el proceso de Haber-Bosch, que es el utilizado actualmente, modernizado, en las fábricas de fertilizante nitrogenado y que fue ayudado por el desarrollo simultáneo de un proceso de cata-lización simple para la oxidación de amonio a nitratos en la industria de explosivos. El N2 y el H2 se combinan directamente en condiciones de operación apropiadas para alcanzar una mezcla en equilibrio que contiene 20% de amonio. Cuando se descubrió esta reacción, requería 1.300ºC más que la modificada actualmente, pero, a pesar de todo, para producir 1 t de fertilizante hacen falta 1,3 t de petróleo, además de que se pierden en la atmósfera 6.3x109 t de CO2, metano, NO y NO2 durante la fabricación, lo que contribuye en gran medida al efecto invernadero.
En los sistemas agrícolas tradicionales, este fertilizante constituye el principal insumo externo para optimizar los rendimientos, pero su eficiencia es una de las más bajas entre todos los nutrientes de las plantas, además de su efecto sustancial sobre la con-taminación ambiental mediante la lixiviación de los nitratos a los mantos acuíferos, lo que trae como consecuencia su eutrificación, así como mediante pérdidas gaseosas por la vía de amonio o de N2O, además de que el aumento de la deposición de N en los sistemas naturales influye sobre el balance de nutrientes y la diversidad biológica (Peoples y Craswell,1992).

39la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
A pesar de esto, en los últimos años se utilizan en el mundo 83 millones de toneladas anuales de fertilizante nitrogenado (FAO, 2001b), y la causa principal del aumento desde casi 61 millones en los años 80 ha sido la creciente aplicación en los países asiáticos. Se considera que Asia, Africa y América Latina consumen 46 millones de toneladas (55% del total), según la Asociación Internacional de la Industria de Fertilizantes (2003).
La fijación biológica del N2
En contraste con lo expresado anteriormente, la fijación biológica del Nitrógeno consti-tuye un componente primordial de la sustentabilidad agrícola. En los suelos que tienen escasa disponibilidad del elemento, solamente los cultivos que pueden asociarse con sistemas de fijación biológica de Nitrógeno tienen la posibilidad de sobrevivir (Dôbe-reiner et al., 1995). En contraste con las grandes cantidades de energía fósil utilizadas para desarrollar el proceso de Haber- Bosch, en el proceso biológico la energía se deriva de la fotosíntesis, y se requieren entre 6 y 12 kg de C fotosintético para lograr la fijación de 1 kg de N2 en la simbiosis Rhizobium – leguminosas (Marschmer, 1999). De lo que se ha expuesto se deduce que éste constituye el método de suministrar N a los agroecosistemas más saludable para el ambiente.
El flujo terrestre de Nitrógeno a partir de la fijación biológica ha sido calculado entre 125 y 200 millones de toneladas/año (Paul, 1988), de las cuales 35 millones corres-ponden a la fijación simbiótica por leguminosas, 15 a la fijación simbiótica por no legu-minosas, 7 a los campos de arroz , 80 a otra vegetación y 36 millones de toneladas a la fijación marina. Es decir, que el Nitrógeno aportado por la fijación biológica al suelo se encuentra en el rango de los 137 millones de toneladas y puede ser mayor si se aplican los biofertilizantes en mayor escala. En términos absolutos, estas cifras son pequeñas en comparación con las reservas totales de Nitrógeno en el suelo, que ascienden a 105.000 millones de toneladas (Peoples y Craswell, 1992), pero son más elevadas que los 83 millones de toneladas de fertilizante nitrogenado que se aplican actualmente en el mundo. Si se toma en cuenta que solo a causa de la extracción por los cultivos se pierden del suelo todos los años 100 millones de toneladas del elemento, sin consi-derar las pérdidas por lixiviación, volatilización o por otros procesos, puede deducirse la gran importancia que tiene la fijación biológica para el mantenimiento del ciclo del Nitrógeno.

40 capítulo 3. generalidades sobre la fijaciónbiológica del nitrógeno atmosférico
Resumen histórico de los estudios sobre fijación biológica del N2
El hombre cultivó la tierra de una forma organizada desde hace 8.000 años a.C. Los reportes más antiguos indican que los romanos tenían ya manuales para que los agri-cultores incrementaran la fertilidad del suelo; así, Columella escribió 60 años a.C. un tratado que contiene descripciones de varias prácticas agrícolas, el cual fue usado en el Imperio Romano por muchas generaciones. La cultura árabe floreció en el siglo XII, cuando Ibn-al-Awan escribió un Tratado de Agricultura.
Muchos pueblos conocían el papel de los componentes del suelo en cuanto a la nutri-ción de las plantas. En el siglo XVI, Bernard Pallisy planteó que los residuos vegetales contenían “la sal” o “el principio” que apoyaba el crecimiento de las plantas, mientras que Jan Baptista van Helmont creía que el agua era “el principio”. Mas tarde, se esta-bleció la idea de que ciertos ingredientes del suelo disueltos en el agua intervenían en el crecimiento de las plantas, gracias a los experimentos de John Woodward en Inglaterra. Los trabajos de J.B. Boussingault en Francia, en 1834, revelaron el importante papel de los componentes químicos de las plantas y del suelo. El alemán Justus von Liebig preparó un balance de la nutrición vegetal en relación con el suelo. Uno de los hitos en los estudios sobre fertilidad de los suelos lo constituyó el descubrimiento del super-fosfato, en 1840, por John Bennet Lawes y J. H. Gilbert, en la Estación Experimental de Rothamsted, en Inglaterra. Estudios posteriores en la misma estación condujeron a un mayor conocimiento de la Química del Suelo y de los requerimientos químicos de las plantas.
Los primeros estudios que se realizaron acerca de lo que hoy se llama biofertiliza-ción estuvieron relacionados con la fijación biológica del Nitrógeno atmosférico, y se remontan a la primera mitad del siglo XIX. Así, Boussingault señaló en 1838 que el cultivo del trébol mejoraba la fertilidad de los suelos cuando se rotaba con cereales, aún después de recoger la masa de follaje verde para alimentar el ganado, por lo que encontró razonable pensar “que las partes verdes de las plantas son capaces de asimilar el Nitrógeno de la atmósfera”. Para probarlo, diseñó experimentos con trigo y trébol en un medio totalmente libre de Nitrógeno y demostró que, mientras el trigo tuvo solo una diferencia de 5% en el contenido del elemento, desde la semilla hasta la cosecha, el trébol mostró 36% de aumento. Estos resultados, unidos a los que había obtenido en los experimentos de rotación, le permitieron expresar que “el Nitrógeno de la at-mósfera puede ser asimilado durante la vida de una planta”.

41la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
En 1847, Liebig planteó la “hipótesis del amonio”, y Ville, en 1854, utilizando técnicas muy parecidas a las de Boussingault, reportó una gran acumulación de Nitrógeno en las plantas experimentales. En 1862, Jodin demostró que una solución que contenía azúcar y ácido tartárico, pero ningún Nitrógeno, permitía un vigoroso crecimiento de “micoderma” en forma de velo sobre la superficie del medio, lo que constituye una característica de Azotobacter, que habría de ser descubierto 40 años después. Pero la demostración de Jodin no recibió reconocimiento.
Pronto fue olvidada por los científicos la fijación de N2, hasta que en 1890 Berthelot en-contró que las muestras de suelos sin esterilizar aumentaban su contenido del elemen-to durante el almacenamiento, mientras que las esterilizadas no lo hacían, por lo que sacó la conclusión de que algunas bacterias que vivían libres en el suelo eran capaces de fijar N2 atmosférico, proceso que Berthelot consideró necesario para compensar las pérdidas que ocurren en la naturaleza. En este sentido dijo: “La reserva natural de los compuestos nitrogenados tendería a disminuir si no existieran causas compensadoras. Es preciso que existan acciones inversas capaces de fijar nitrógeno”.
Ya en 1888, Hellriegel y Wilfarth habían descubierto las bacterias simbióticas del género Rhizobium, aisladas de nódulos de raíces de plantas leguminosas. En 1894, Winograds-ky aisló el primer diazotrofo de vida libre, la bacteria anaeróbica Clostridium pasteuri-anum, utilizando un medio de cultivo libre de Nitrógeno enriquecido con manitol. En 1901, Beijerinck aisló la bacteria aerobia Azotobacter chroococcum.
La primera inoculación bacteriana al suelo fue realizada por Nobbe y Hiltner en 1895, quienes prepararon en el laboratorio un inóculo a base de Rhizobium sp sobre un medio que contenía extractos de plantas leguminosas, gelatina, azúcar y asparagina. En pocos años se produjeron 17 inoculantes distintos para diferentes especies de legumi-nosas, todos protegidos por la marca “Nitragin” (Martínez Viera, 1986). Ya en 1920 existían 19 patentes comerciales en Estados Unidos que amparaban la fabricación de inoculantes a base de Rhizobium, y estos bioproductos se utilizan hoy con éxito en todo el mundo.
En relación con las bacterias fijadoras asociativas, comenzó a utilizarse en la URRS y Polonia, en los años 30, un producto sólido llamado “Azotobacterin”, a base de Azo-tobacter chroococcum, y se reportaron incrementos notables en los rendimientos de cereales y hortalizas en algunas regiones y ningún efecto en otras (Mishustin y Silnikova,

42 capítulo 3. generalidades sobre la fijaciónbiológica del nitrógeno atmosférico
1971). Pero el mayor nivel de aplicación efectiva de estas bacterias ha ocurrido en la India (Mehrotra y Lehri, 1971; Joi y Shinde, 1976) y, más recientemente, en Cuba (Di-but et al., 2004; Martínez Viera et al., 2004); en ambos países han alcanzado estos bio-productos altos niveles de aceptación entre los campesinos, debido a que su aplicación en distintos cultivos de plantas no leguminosas promueve la germinación de las semillas y el vigor inicial de las plantas, con importantes incrementos de los rendimientos y la calidad de los cultivos.
En 1976 fueron redescubiertas las bacterias del género Azospirillum, que habían sido descritas por Beijerinck en 1925 con el nombre de Spirillum lipoferum y cayeron en el olvido hasta que Döbereiner y Day (1976) pusieron de manifiesto su capacidad para fijar N2 atmosférico y para sintetizar sustancias activas estimuladoras del crecimiento vegetal. Desde un punto de vista evolucionista, se ha sugerido que estas bacterias son organismos intermedios entre los fijadores libres y los simbióticos. La localización de los puntos de fijación de Nitrógeno dentro de las células de las plantas hospedantes ha apoyado esta hipótesis, porque no se admite hoy que la fijación simbiótica tenga que estar confinada dentro de estructuras visibles como los nódulos cuando existe la posi-bilidad de otras formas de asociación, tales como el establecimiento dentro del tejido cortical de las raíces (Krieg y Tarrand, 1978). Estas bacterias están siendo ampliamente utilizadas como inoculantes para las gramíneas en distintos países.
Más recientemente fue descubierta una bacteria fijadora de nitrógeno atmosférico que vive en el interior del tallo de la caña de azúcar y actúa en asociación con dicha planta, la cual fue bautizada originalmente como Saccharobacter nitrocaptans (Cavalcante y Dobereiner, 1988) y después como Acetobacter diazotrophicus (Gillis et al., 1989) , nombre que cambió en los últimos años por Gluconacetobacter diazotrophicus (Yanada et al., 1997).
Los resultados obtenidos en el estudio de esta nueva bacteria permitieron decir a Dôbereiner (1993): “Todas estas observaciones cambian completamente el concepto de asociación planta-diazotrofo, la cual se pensaba que era una asociación puramente casual con las raíces de las plantas, y abre enteramente nuevas vías para expandir la fijación biológica del nitrógeno en sustitución de los fertilizantes nitrogenados para las gramíneas”. Con esta nueva bacteria se han preparado también inoculantes que se aplican en al-gunos países productores de caña de azúcar. En Cuba se están utilizando con éxito en cultivos tales como la papa, la yuca y la batata, en los cuales se logran importantes in-

43la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
crementos del rendimiento con una escasa aplicación de fertilizante nitrogenado (Dibut et al., 2003).
También existen distintos organismos de alta efectividad en la fijación de N2 pero que no son bacterianos, por lo cual no serán tratados en este espacio, ya que, por su gran importancia, deben de ser objeto de estudios separados. Entre ellos se encuentran el helecho acuático Azolla sp .y las algas verdi-azules. También existen otras bacterias que han sido efectivas en pruebas experimentales, pero que no cuentan aún con tecnolo-gías adecuadas de fabricación y aplicación que les permita alcanzar el nivel de aplicación comercial, por lo cual tampoco se hablará de ellas en este momento.
El más familiar y utilizado de los procesos de fijación es la asociación simbiótica entre las bacterias del género Rhizobium y las raíces de las plantas leguminosas. Pero en las últi-mas décadas se ha demostrado concluyentemente que un gran número de especies de gramíneas, incluyendo cultivos de tanta importancia económica como caña de azúcar, maíz y sorgo, pueden tener asociadas a sus raíces, y hasta viviendo dentro de sus tallos, poblaciones de bacterias fijadoras. Por último, hay bacterias de vida libre que dependen de su asociación con los exudados radicales para obtener la fuente energética que les permite cumplir su función fijadora.
Este proceso puede ser la mayor fuente de Nitrógeno para la agricultura cuando se utilizan los sistemas simbióticos bacterias-leguminosas, los cuales pueden aportar hasta 360 kg de N/ha, como se pone de manifiesto en la Tabla 5, mientras que los sistemas a base de microorganismos fijadores asociativos o de vida libre aportan cantidades mucho menores, por lo que los cultivos requieren aplicaciones suplementarias de fer-tilizante nitrogenado, aunque siempre significativamente menores que las necesarias cuando no se realiza la inoculación. Pero muchos de los conocimientos que se han adquirido sobre el proceso de fijación no están siendo utilizados, especialmente en los países que más necesitan de su aplicación, lo que se debe a la existencia de obstáculos importantes para lograr implementar las tecnologías en la práctica agrícola, los cuales son de orden técnico, socio-económicos o de escasez de talentos humanos.
En la Tabla 5 se presentan los aportes de Nitrógeno a la agricultura, calculados para diferentes sistemas fijadores en asociación con distintos cultivos (Bohlool et al., 1992).

44 capítulo 3. generalidades sobre la fijaciónbiológica del nitrógeno atmosférico
Hay todavía muchos aspectos desconocidos en los procesos de fijación de Nitrógeno atmosférico y, en la medida en que la investigación permita su clarificación, será posible mejorar las prácticas agrícolas que promueven estos procesos.
Mecanismo de la fijación biológica de N2: la nitrogenasa
El sistema enzimático responsable de la fijación se llama nitrogenasa. El primer extracto libre de células fue preparado a partir de Clostridium pasteurianum en 1960. Desde entonces, se han obtenido nitrogenasas parcialmente purificadas a partir de unas 20 especies bacterianas, cinco de las cuales han podido ser totalmente purificadas. Esta enzima puede ser aislada de dos formas distintas: como el complejo nitrogenasa, el cual ha sido bien caracterizado solamente para Azotobacter vinelandii, tiene un peso molecular de 300.000 y contiene una proteína Fe-Mo, una proteína Fe y una ferre-doxina, La segunda forma resulta de la separación de las dos proteínas componentes (Fe-Mo y Fe), que tienen pesos moleculares de 230.000 y 60.000 respectivamente. Ninguna de estas proteínas muestran actividad enzimática cuando están aisladas, pero cuando se recombinan despliegan el mismo nivel de actividad que el complejo (Haaker y Veeger, 1977).
Actualmente se utilizan las designaciones Acl, Avl, Bpl, Cpl, Cal, Kpl, Rll y Rjl para las pro-teínas Fe-Mo de los distintos fijadores que se han estudiado: Azotobacter chroococcum, A. vinelandii, Bacillus polymyxa, Clostridium pasteurianum, Corynebacterium autotrophi-cum, Klebsiella pneumoniae, Rhizobium lupini y Rhizobium japonicum respectivamente.
Tabla 5
Estimados de Nitrógeno aportado por diferentes
sistemas fijadores en la agricultura
Sistema fijador de nitrógeno N2 fijado (kg/ha) por período de cultivo
Sistemas de vida libreArroz-algas verde azules 10-80Arroz-asociación con bacterias 10-60Asociación caña de azúcar-bacterias 20-160
Sistemas simbióticosArroz-Azolla 20-100Leguminosas-Rhizobium 50-360No leguminosas-Frankia 40-60 por año

45la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Estos símbolos deben utilizarse hoy con preferencia al término nitrogenasa, según fue aprobado en la discusión sobre nomenclatura que se realizó en la IV Conferencia sobre Fijación de Nitrógeno, celebrada en Australia en 1980 (Gibson et al., 1982).
Para que se tenga una idea de las variaciones que ocurren entre las distintas nitrogena-sas, pueden verse las variaciones entre los pesos moleculares de las proteínas Fe-Mo de cada una de ellas (Smith, 1983): Avl 216 000, Acl 216 000, Rjl 180 000, Rll 194 000, Cal 235 000, Kpl 220 000 y Cpl 200 000. La nitrogenasa es altamente sensible a la presencia de oxígeno y requiere una fuente de potencial reductor y de adenosina tri-fosfato (ATP) para poder cumplir su función. Cuando existe esta fuente, la enzima cata-liza la reacción de un reductante dependiente de ATP con N2 para formar NH3+ADP (adenosina difosfato) y fosfato. La ecuación que permite la expresión de la reacción nitrogenasa, determinada en el caso de Clostridium pasteurianum, es la siguiente:
N2 + 8 ferredoxinas- + 8H+ + 16 MgATP2- + 18 H2
2 NH4+ + 2 OH- + 8 ferredoxinas + 16 MgADP-+ 16 H2 PO4
- + H2
Esta reacción muestra una clara necesidad de potencial reductor y de energía como ATP. El notable requerimiento de ATP se debe a que actúa como un agente des-hidratante para incrementar la reactividad de los lugares reducidos en el complejo enzimático. En los sistemas microaeróbicos, como son los bacteroides de los nódulos de las raíces de las leguminosas y la mayoría de las bacterias asociativas, la ferredoxina es reemplazada en la reacción anterior por la flavodoxina, un portador de electrones similar que dona electrones al sistema nitrogenasa. Puede decirse con certeza que nin-guna otra reacción bioquímica simple requiere esta cantidad de ATP. Pero todavía más ATP debe utilizarse por las bacterias fijadoras vivas, ya que la utilización incrementada de sacarosa que se necesita para crecer sobre N2 indica que la actividad nitrogenasa requiere 30 moléculas de ATP (Kennedy y Cocking, 1997).
El costo energético del proceso de fijación se considera que es demasiado alto, pero no se ha podido demostrar por los que están en contra de utilizar los biofertilizantes que este costo energético tenga influencia en la reducción de los rendimientos, sino más bien, como se expondrá en otros capítulos, éstos se incrementan, a pesar de que se reduce el fertilizante nitrogenado de origen industrial. En los casos de las legumi-nosas y el arroz no se han observado diferencias en el rendimiento cuando las plantas utilizaban nitratos o Nitrógeno fijado (Lahda y Reddy, 2003), lo que puede ser atribuido

46 capítulo 3. generalidades sobre la fijaciónbiológica del nitrógeno atmosférico
a que los costos energéticos para fijar N2 (687 kJ/mol) son teóricamente similares a los que se necesitan para la reducción de nitratos (605 kJ/mol) por las plantas (Pate et al., 1979).
Aunque desde 1960 se obtuvieron mutantes para la fijación de Nitrógeno, no fue hasta 1971 que se realizaron detalladas investigaciones sobre los genes de fijación (nif). En este año se reportó el agrupamiento de los nif-genes del cromosoma de Klebsiella pneumoniae (Streicher et al., 1971). Hoy se conoce que el número de genes relaciona-dos con la fijación es 17, así como su localización en el caso de Klebsiella pneumoniae, que es la bacteria utilizada como modelo para estos estudios.
Trabajos similares se han realizado con especies pertenecientes a los géneros Azoto-bacter y Rhizobium y algas verdi-azules del género Anabaena, aunque en todos estos casos ocurren complicaciones que le dan mucha lentitud a estas investigaciones; así, por ejemplo, Rhizobium presenta la dificultad de que tiene que ser aplicado a una le-guminosa antes de su estudio, pero ya hoy se conoce bastante acerca del clonaje de los genes-nif de Rhizobium y sobre su presencia en un plásmido y se piensa que estos organismos tengan por lo menos el mismo número de genes que K. pneumoniae.
Distribución de los sistemas fijadores
La fijación de Nitrógeno ocurre en toda la superficie de la tierra, en la parte sólida y en la líquida. Todos los ecosistemas contienen sistemas fijadores, aunque las cantida-des que se fijan del elemento varían con amplitud, en dependencia de la presencia de Nitrógeno en el medio y de la riqueza del ecosistema. La variedad de ecositos donde se ha reconocido la fijación biológica se muestra en la tabla 6 (Kennedy y Cocking, 1997).
Puede causar asombro que, entre los sistemas fijadores de Nitrógeno, se encuentren las trompas de las termitas y, más específicamente, su intestino. En efecto, de 50 es-pecies bacterianas reconocibles como diferentes tipos morfológicos en el intestino de las termitas, fijaban nitrógeno unas 40 especies, las cuales expresaron actividad nitro-genasa aún en presencia de cantidades superiores a 5 mm de amonio en el intestino, y solamente perdían esa expresión cuando se exponían a gas amonio puro (Kennedy y Cocking, 1997).
Aunque se ha recorrido hasta hoy una escala diversa de sistemas fijadores de nitróge-

47la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Ecosistema Naturaleza de la fijación
Tipo de fijación Asociación
Natural
Zonas agrícolas y de bosques
Simbiótica
No simbiótica
Simbióticas
No simbióticas
Nódulos en raícesy tallos por Rhizobiumy Frankia
Musgos, líquenes, Pteridofitas
Simbiosis Gunnera-Nostoc
Insectos
Saprofitosde vida libre
Fotosintética
Igual que ocurre en el medio natural
Leguminosas, plantas actinorrizales
Superficie del suelo, rocas y árboles.
Base de las hojas y raíces de Cycadas
Termitas
Suelo, raíces, materia orgánica, superficie de las plantas.
Algas verdi-azules,bacterias de aguas dulces y del mar.
Cultivos anuales y perennes, rotación, abonos verdes
Plantaciones y áreas de formación de suelos
Azolla, caña de azúcar y otros
Tabla 6
Distribución de sistemas fijadores de Nitrógeno
Leguminosas noduladas
Angiospermas actinorrizales
Misceláneas simbióticas

48 capítulo 3. generalidades sobre la fijaciónbiológica del nitrógeno atmosférico
no, como se pone de manifiesto en dicha tabla, la fijación biológica está restringida a los procariotes y nunca se ha encontrado en organismos eucarióticos. Lo que ocurre es que algunos organismos procariotes fijan Nitrógeno por sí mismos, mientras que otros deben establecer una relación simbiótica con un sistema eucariótico para realizar la fijación.
La escala de microorganismos fijadores de Nitrógeno cubre la mayoría de los dife-rentes sistemas de energía biotrófica, incluyendo bacterias fotosintéticas, anaeróbicas,anaeróbicas,anaeróbicamicroarerobias y aerobias . En cada uno de estos grupos se encuentran numerosos géneros y especies, entre los que se presentan a continuación los más conocidos.
Organismos aeróbicos: Todas las especies de los géneros Azotobacter, Azomonas, Azo-tococcus, Beijerinckia y Derxia, algunas especies de los géneros Aquaspirillum, Metha-nosinus y otras bacterias oxidantes de metano y todas las algas verde-azules hetero-quísticas.
Organismos microaeróbicos: Todas las especies de los géneros Rhizobium, Bradyrhizo-bium, Azospirillum, Gluconacetobacter, Herbaspirillum y Xanthobacter, y Xanthobacter, y la especie Thioba-cillus ferro-oxidans y muchas algas verde-azules no heteroquísticas.
Organismos facultativos: Bacillus azotogensis, B. macerans, B. polymyxa, Klebsiella aero-genes, K. oxytoca, K. pneumoniae, Citrobacter freundii, y algunas especies de los géneros Enterobacter, Rhodopseudomonas y Rhodospirillum.
Organismos anaeróbicos: Clostridium pasteurianum y algunas especies de los géneros Chlorobium, Chromatium, Desulfovibrio, Desulfotomaculum Ectothiospira, Propionibacte-rium y Thiopedia.
Todos estos organismos han sido ubicados como fijadores de Nitrógeno atmosférico a partir de las pruebas de reducción del acetileno a etileno, en las que han demos-trado su capacidad de fijación aunque sea en pequeñas cantidades. Pero de esta larga lista, solo unos pocos géneros se han considerado capaces de aportar beneficios a los cultivos cuando se aplican como biofertilizantes: Azotobacter, Azospirillum, Rhizobium, Bradyrhizobium, Gluconacetobacter, Herbaspirillum y algunas especies de algas verde-azules heteroquísticas.

49la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Organismos fijadores de N2 más utilizados como biofertilizantes
En la tabla 7 se presentan los microorganismos que pueden ser considerados como biofertilizantes principales entre los fijadores de N2 y las mayores limitaciones que exis-ten para su uso (según Ladha, 1997, con algunas modificaciones).
Como se observa en la tabla 7, los únicos biofertilizantes de amplio uso son los prepa-rados a base de Rhizobium y Bradyrhizobium, que se utilizan en la mayor parte de los países en los cultivos de leguminosas para grano y para aceite. Los biofertilizantes que se utilizan en arroz (Azollase utilizan en arroz (Azollase utilizan en arroz ( y cianobacterias) son extensamente aplicados en los países asiáticos, pero no se han extendido a otros continentes porque no se ha tomado en asiáticos, pero no se han extendido a otros continentes porque no se ha tomado en asiácuenta la efectividad demostrada en muchos años de uso en las condiciones de aque-llos países.
En relación con Azospirillum y Azotobacter, aunque se ha alcanzado un buen nivel de desarrollo tecnológico en los últimos años, es poco utilizado todavía por los agricul-tores, a pesar de que, además de su papel como fijadoras, estas bacterias asociativas sintetizan una amplia gama de sustancias promotoras del crecimiento, sideróforos y bacteriocinas, las que no solo incrementan el desarrollo de las plantas, sino que tam-bién aseguran el establecimiento competitivo de estas especies bacterianas en la ri-zosfera (Elmerich et al., 1992). En el caso de Gluconacetobacter, su conocimiento es relativamente reciente, pero su potencialidad es muy grande.
La baja utilización de Azospirillum y Azotobacter y Azotobacter y se debe a las escasas investigaciones en los países tropicales para aprovechar los beneficios que pueden aportar estas bacterias, a pesar de que las condiciones ambientales son más favorables para su establecimiento. Con excepción de Brasil, Cuba y México, muy poco se trabaja con Azospirillum en el mundo tropical, y los estudios sobre Azotobacter se abandonaron prácticamente hace años, con excepción de Cuba e India, como consecuencia de los resultados irregulares que se obtenían en los países templados.
Utilizando la técnica de reducción de acetileno, se han reportado ganancias de Ni-trógeno en los países templados de 34 Kg N/Ha año en el trigo (Jenkinson, 1976), mientras que en el trópico se obtuvieron valores de 876 Kg N/Ha/año en raíces de maíz y de Digitaria decumbens (von Bulow y Döbereiner, 1975; Döbereiner y Day, 1976). Estos datos demuestran la importancia que tiene la aplicación de los procesos de fijación biológica del Nitrógeno atmosférico para la Agricultura Tropical.

50 capítulo 3. generalidades sobre la fijaciónbiológica del nitrógeno atmosférico
Biofertilizante Cultivo Limitaciones técnicas
Uso por los agricultores
RhizobiumyBradyrhizobium
Leguminosas de granosLeguminosas de aceiteLeguminosas forrajeras
Baja calidad del inóculo.Bajo Fósforo en el suelo
Pobre infraestructura.Poco conocimiento por los agricultores
XXX
XXX
X
Azolla Arroz Mantenimiento del inóculo.Bajo Fósforo.Calor, plagas y enfermedades
Pobre infraestructura
X
Cianobacterias Arroz Bajo Fósforo.Malezas
Falta de tecnología
+/-
Azospirillum Gramíneas Pobre inóculo
Limitaciones socio-económ.
Poco conocimiento por los agricultores
+/-
Azotobacter Todos los cultivos
Pobre inóculo Poco conocimiento por los agriculores
+/-
GluconAcetobacter
Caña de azúcar y otros cultivos
Conocimientomuy reciente
Poco conocimientopor los agricultores
+/-
Tabla 7
Biofertilizantes fijadores de N2, limitaciones y uso
por los agricultores
XXX = amplio usoX = uso intermedio+/- = más o menos

51la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Influencia de los factores ambientales sobre la fijación biológica de N2
Un número de factores ambientales gobiernan la magnitud de la fijación, en depen-dencia de las características físicas y químicas del agroecosistema. En primer lugar, los microorganismos fijadores tienen también la habilidad de utilizar amonio, nitratos y algunas otras formas combinadas de Nitrógeno. Las sales de amonio son usadas con preferencia, y muchas veces en mayor cantidad que el N2, por lo que la presencia de amonio inhibe la fijación, es decir, que los microorganismos utilizan el Nitrógeno com-binado en vez del N2 de la atmósfera.
Döbereiner (1987) demostró que la fijación se reduce en suelos brasileños que han sido fertilizados recientemente. En Madagascar se puso de manifiesto la reducción en 60% de la población de Azospirillum en raíces de arroz con la aplicación de fertilizante nitrogenado (Boddey et al., 2006). Pero el estudio más completo, utilizando técnicas de análisis de ADN, fue realizado por Tan et al. (2003), los que reportaron una rápida disminución en la población de diazotrofos en las raíces de arroz 15 días después de la aplicación de fertilizante nitrogenado de origen industrial.
También los compuestos que se transforman en amonio, como urea o nitratos, son efectivos para inhibir la fijación, aunque no todas las aplicaciones de fertilizantes ni-trogenados tienen un efecto negativo. Así, en suelos pobres en Nitrógeno, una dosis de 5-10 kg/ha de fertilizante puede estimular la germinación y el crecimiento de las plántulas y la capacidad de fijación y estimulación, lo que se traduce en incrementos del rendimiento (Ma et al.,1998), y 15 kg N/ha de F-nitrogenado en suelos ácidos de muy baja fertilidad (España et al., 2006); mientras que (España et al., 2000) en un vertisol del Estado Guárico, aplicó 20 Kg N/ha y logró FBN en soya de 47% mejorando la FBN al utilizar labranza conservacionista.
Por estas razones, cuando se quieren aprovechar los beneficios de los organismos fijadores hay que aplicar cantidades reducidas de fertilizantes minerales, por lo menos hasta que se hayan establecido los organismos inoculados y estén cumpliendo con su actividad fijadora.
Aunque muchas sustancias inorgánicas son necesarias para el desarrollo de los micro-organismos, solamente unas pocas de ellas están directamente implicadas en el meta-bolismo del N2, es decir, que son indispensables para que se efectúe el proceso. Así, la falta de Molibdeno, Hierro y Calcio es crítica y puede inhibir la capacidad de fijación.

52 capítulo 3. generalidades sobre la fijaciónbiológica del nitrógeno atmosférico
En el caso de algunos organismos, el Vanadio puede reemplazar al Molibdeno, aunque nunca es igualmente efectivo.
En los suelos ácidos de sabanas hay que aplicar dosis bajas de Mo (0,2 Kg.ha-1 para lograr la fijación de nitrógeno en leguminosas de grano y forrajeras (López et al., 2006), para poder lograr una eficiente fijación.
Algunos fijadores, como los pertenecientes al género Azotobacter, son muy sensibles a las altas concentraciones de hidrogeniones. Por eso, la ausencia de estos fijadores en determinados suelos está asociada directamente con el pH. Investigaciones de carácter ecológico han puesto de manifiesto que el 100% de los suelos con pH entre 7 y 8 contienen Azotobacter mientras que se encuentran en el 80% de los suelos con pH entre 6 y 7, en el 20 % de los que tienen pH entre 5 y 6 y en menos de 10% con pH entre 4.5 y 5. Por debajo de pH 4.5 no se encuentra la bacteria (Alexander, 1977). En un estudio hecho en suelos arenosos de Cuba, con pH inferiores a 5, no fue posible aislar ninguna cepa de Azotobacter (Martínez Viera et al., 1968).
La humedad, en general, limita la vida microbiana del suelo, por lo que su exceso o defecto influye directamente sobre las poblaciones de los diazotrofos. En el caso de exceso de humedad, se reduce la aireación y las poblaciones microbianas disminuyen. Sin embargo, algunas especies de bacterias fijadoras de Nitrógeno pueden resistir pro-longados períodos de sequía y han sido aisladas de muestras que se han conservado secas durante tiempos largos (Moreno et al., 1986). Esta resistencia se debe a que algunos géneros como Azotobacter y Azospirillum son capaces de formar quistes que permiten una larga supervivencia.
La aireación probablemente ejerce un mayor efecto sobre el desarrollo de la mayoría de los diazotrofos, en comparación con otros microorganismos no fijadores. A pesar de esto, la mayor parte de los diazotrofos aeróbicos funcionan mejor a reducidas con-centraciones de O2, debido a la sensibilidad del complejo nitrogenasa (Postgate, 1990). En general, la rizosfera contiene numerosos ecositos con bajo pO2, los que constituyen habitats favorables para las bacterias fijadoras (Zuberer, 1990).
Importancia del Fósforo para la fijación de N2
Después del Nitrógeno, el fósforo es el elemento más necesario para la vida y no es sustituible en los sistemas biológicos. Es uno de los principales constituyentes de los

53la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
ácidos ribonucléico y desoxiribonucléico y del ADN; es el portador de energía de la molécula adenosina trifosfato (ATP) y de sus dos precursores mono y difosfato (AMP y ADP). Su presencia en las moléculas de ADN de los cromosomas permite los procesos de almacenamiento, replicación y, junto con el ARN, la transcripción de información genética.
En el caso de las plantas, el fósforo tiene que existir en el suelo en formas asimilables en cantidades suficientes, ya que, además de los papeles citados en el párrafo anterior, actúa en los procesos de fotosíntesis, respiración, almacenamiento y transferencia de energía, permite la división celular y participa en otros procesos, tales como la fecun-dación de las flores (Wilcox, 1996).
La cantidad total de fósforo en la biomasa vegetal se ha calculado en 1805 millones de toneladas, considerando un contenido de 0.1%; de esta cantidad, se ha estimado una transferencia al suelo como componente orgánico de 136.4 millones de toneladas por año (Bazilevich, 1974). A pesar de estas reservas, gran parte del elemento es inmo-vilizado en formas no accesibles para las plantas, además de que se pierde una parte importante por lixiviación de iones fosfato y por la erosión del suelo. Lo mismo ocurre cuando se aplica fertilizante fosfórico, del cual las plantas aprovechan solo una mínima porción, especialmente en los suelos tropicales.
Un requerimiento fundamental para la fijación de N2 es la presencia de Fósforo asimi-lable, ya que las bacterias deben asimilar por lo menos 1 mg de Fósforo por cada 5 a 10 mg de N2 que se fijan (Alexander, 1977). Debido a esta necesidad, se obtienen los elevados incrementos de los rendimientos que se mostrarán más adelante, en el capí-tulo correspondiente, cuando se hable de los biofertilizantes mixtos a base de fijadores de nitrógeno y solubilizadores de fósforo. La acción limitadora de la falta de fósforo se debe a que este elemento tiene un efecto fundamental en el intercambio del carbono, en la multiplicación de las bacterias y en el mismo proceso de fijación.
Entre los muchos trabajos que se han efectuado para conocer las cantidades de este elemento que deben existir para garantizar un proceso de fijación efectivo, se ha de-mostrado que las leguminosas requieren un mínimo de 30 kg/ha para que pueda efec-tuarse la simbiosis (Weber, 1996). Pero las especies vegetales difieren en su capacidad para asimilar fósforo, especialmente cuando el contenido del elemento asimilable en el suelo es bajo, aunque muchos de ellos contienen considerables reservas que están

54 capítulo 3. generalidades sobre la fijaciónbiológica del nitrógeno atmosférico
fijadas en formas inutilizables. Por eso es importante aplicar la inoculación al suelo de organismos solubilizadores de fósforo, para garantizar que el elemento esté disponible en el proceso de fijación (Sanginga, 2003).
En el caso de la solubilización del fósforo inorgánico, el principal mecanismo microbio-lógico que permite la movilización del compuesto insoluble consiste en la síntesis de ácidos orgánicos en el medio, tales como fórmico, acético, propiónico, glucónico, lácti-co, glicólico, fumárico y succínico. Otros microorganismos sintetizan ácidos inorgánicos que tienen la misma acción, formando quelatos con Calcio o Hierro que traen como consecuencia la efectiva solubilización (Oswald y Bhide, 1977; Gaur y Ostwald, 1977). Por ejemplo, el Ca3 (PO4)2, insoluble, se transforma en fosfato dibásico o monobási-co solubles, lo que trae como consecuencia un aumento de la disponibilidad para las plantas. La cantidad de fósforo solubilizado varía de acuerdo con el consumo de carbo-hidratos por los microorganismos solubilizadores, que deben tener la capacidad para convertir el sustrato carbonatado en ácidos orgánicos.
Algunas bacterias pueden liberar SH2, el cual reacciona con el FePO4 y produce SFe, liberando el ión fosfato. Otra vía, que predomina en los suelos anegados como los arrozales, consiste en reducir el Fe del FePO4 y la liberación de Fe soluble, con la consiguiente liberación del ión fosfato en la solución. Este aumento de la disponibilidad de fósforo en los suelos anegados puede explicar la menor necesidad de fertilizante fosfórico en los campos de arroz bajo agua, en comparación con los secos.
En suelos tropicales gran parte del fósforo se encuentra en forma orgánica (Dalal, 1977), lo cual afecta la producción optima de los cultivos y la fijación eficiente de nitrógeno, debido a que las plantas no pueden utilizar el P-orgánico directamente (Tate, 1984), en estas condiciones la actividad de enzimas fosfatas juegan un importante papel, ya que pueden hidrolizar el P-orgánico a P-inorgánico (Beck et al, 1989) y hacer disponible el fósforo a las plantas, beneficiando el proceso de fijación biológica de nitrógeno (FNB). La actividad de fosfatasa en el suelo proviene de muchas fuentes, incluyendo las plantas (Dinkelaker y Marschner, 1992), los hongos (Tarafdar et al, 1988); hongos micorrizícos (Tarafdar y Marschner, 1994) y bacterias (Tarafdar y Claassen, 1988).
En el caso de la solubilización del fósforo orgánico, el proceso se desarrolla mediante la síntesis de enzimas fosfatasas por los microorganismos, las cuales separan el P de los sustratos orgánicos. Con esta actividad, se puede aportar a las plantas entre 30-60% de sus necesidades del elemento, en dependencia de la capacidad del microorganismo.

55la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Métodos para medir la fijación de nitrógeno
Se han desarrollado muchos métodos para estimar de forma cualitativa y cuantitativa el nitrógeno fijado en condiciones de campo (Unkovich y Pate, 2000; Hardarson y Craig, 2003). Así, en condiciones en que el rendimiento está limitado por la falta del elemento en el suelo, la materia seca producida por una especie vegetal en asociación fijadora está correlacionada estrechamente con la cantidad de nitrógeno fijado. De la misma manera, el número y la masa de los nódulos de las leguminosas pueden suministrar una idea del nivel de fijación. Una apreciación visual del contenido de leghemoglobina de los nódulos mediante el grado de pigmentación roja también da buenos elementos sobre la efectividad de la asociación. Un estimado más directo se obtiene mediante el análisis del contenido de nitrógeno del material cosechado, en comparación con el material procedente de una parcela del mismo suelo donde no ha habido fijación.
Un método más sofisticado consiste en determinar la actividad nitrogenasa mediante la medición de la reducción del Acetileno a Etileno o de la evolución de Hidrógeno, utilizando la cromatografía gaseosa. Estas técnicas se utilizan sobre todo para medir la actividad de fijación en un momento dado, pero son inadecuadas para conocer la can-tidad de nitrógeno fijado en un período de crecimiento del cultivo, si se considera que en este período hay diferentes variables que afectan directamente a la actividad nitro-genasa (Broughton et al., 2003). También se han desarrollado métodos que cuantifican la translocación del Nitrógeno fijado desde los nódulos a otras partes de las plantas, como son análisis colorímétricos de los ureidos (alantoína y ácido alantóico) contenido en el xilema; si se toma en cuenta que el nitrógeno del suelo es asimilado en las raíces como aminoácidos y no como ureidos, la proporción de estos últimos en el xilema representa la contribución del N2 fijado (Herridge y Rose, 2000).
Pero los métodos basados en el uso de isótopos ofrecen las mediciones más exactas de la fijación durante todo el período de crecimiento de un cultivo en condiciones de campo. Se utilizan plantas fijadoras y no fijadoras en suelo que ha sido enriquecido con 15N mediante la adición de fertilizante marcado. La medición de la extensión en la cual el cultivo fijador diluye el 15N tomado del suelo conduce al conocimiento de la cantidad del elemento que se ha fijado, por diferencia en base a los datos obtenidos del cultivo no fijador (Hardarson y Danso, 1993). Este método permite conocer las cantidades de N2 fijado en distintos períodos después de la siembra.

56 capítulo 3. generalidades sobre la fijaciónbiológica del nitrógeno atmosférico
Importancia económica de la aplicaciónde los biofertilizantes a base de bacterias fijadoras de N2
La mayor parte de las biotecnologías que se han desarrollado para los inoculantes bacterianos no están siendo utilizadas en gran escala, debido a una serie de prejuicios basados en la inconsistencia de los resultados obtenidos en las condiciones de clima templado, donde se han realizado la mayor parte de las investigaciones que generan las publicaciones más importantes y de más amplia divulgación. Pero estos prejuicios no se justifican en las condiciones tropicales, con temperaturas constantemente favorables y con una alta fijación de Carbono que alcanza hasta 20 t/ha/año (Debinstein,1970), lo que garantiza la presencia de altas poblaciones de microorganismos en la zona rizosfé-rica de las plantas.
En general, se han generado grandes contradicciones entre los resultados logrados con la aplicación de estos bioproductos en condiciones templadas y tropicales, lo que ha sido manejado de forma negativa por los grandes monopolios que fabrican y distribu-yen los productos agroquímicos, con el fin de aplazar lo más posible la introducción en gran escala de los biofertilizantes.
En tiempos pasados relativamente recientes, los subsidios para comprar fertilizantes industriales, tanto en países desarrollados como subdesarrollados, quitaban el estímulo para utilizar los biofertilizantes y bioestimuladores, por lo que en muchos países las in-vestigaciones aplicadas en este campo eran escasas. Sin embargo, al desaparecer estos subsidios como resultado de las nuevas políticas económicas gubernamentales, estos bioproductos se hacen más atractivos y aceptables para los agricultores.
De acuerdo con los datos que se han presentado en párrafos anteriores, el uso de los productos biológicos constituye una necesidad económica y ecológica. Puede decirse que se está viviendo el momento internacional más favorable para el desarrollo de las investigaciones básicas y aplicadas sobre microorganismos con características como biofertilizantes y bioestimuladores y para su utilización..
Desde un punto de vista económico, la importancia de la aplicación de los biofertilizan-tes y los bioestimuladores es muy grande. En la tabla 8, y a pesar de que los datos no son actualizados, se ponen de manifiesto las grandes posibilidades que están a nuestro alcance para sustituir por medios biológicos el nitrógeno del suelo tomado por los cul-tivos o perdido por lixiviación, volatilización o denitrificación (Martínez Viera, 1986).

57la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Tabla 8
Aporte estimado de nitrógeno a los cultivos agrícolas en distintos países (expresado en millones de toneladas)
En esta tabla, si no se toman en consideración los datos correspondientes a Gran Bre-taña, con una superficie muy inferior a la de otros países, puede verse que, en Australia, donde la fijación biológica es explotada al máximo, hasta el punto de que constituye un modelo para el mundo, hay un aporte total de nitrógeno muy similar al de Estados Unidos, con la diferencia de que, en Australia, el fertilizante químico representa solo 0.7% del total, mientras que en Estados Unidos alcanza 32%. En la India, el aporte de Nitrógeno es muy bajo, pero el 60% se realiza mediante la fijación biológica.
En lo que se refiere a las superficies aplicadas con biofertilizantes, se deduce de la tabla que en Australia se aplican sobre la casi totalidad de la superficie agrícola y en Estados Unidos sobre una parte importante de esta superficie. En la India existen 69 fábricas de biofertilizantes que se aplican sobre leguminosas, oleaginosas, arroz y gramíneas en general, hortalizas y otros cultivos que incluyen viveros forestales; en 1995 se aplica-ron en ese país 1.107 toneladas de inoculantes a base de Rhizobium y Bradyrhizobium(Chandra et al., 1996). En China se inoculan 6 millones de hectáreas de soya, 2.3 millones de maní y 1 millón de no leguminosas (Cheng Tin Wey, 1997).
En Brasil se cultivan 13 millones de hectáreas de soya, con una producción media de 2,4 t/ha; sobre esta superficie no se aplica fertilizante nitrogenado, sino biofertilizante (Hungría et al., 2000). Entre los impactos de la Biotecnología en Cuba, se encuentra la disminución del uso de los fertilizantes de origen industrial, que se han sustituído por fertilizantes biológicos. En ese país se utilizan hoy 11 veces menos fertilizantes inorgá-nicos que en los años 90, lo que constituye un ejemplo de años de labor y resultados del trabajo científico (Borroto, 2005). del trabajo científico (Borroto, 2005). del trabajo cientí
Para que se tenga una idea de los beneficios económicos que pueden lograrse con la aplicación efectiva de los biofertilizantes y bioestimuladores, puede ponerse el ejemplo
Fuente de Nitrógeno Gran Bretaña USA Australia IndiaFijación simbiótica 0.4 8.6 12.8 0.9
Fijación asociativa 0.05 1.4 1.0 0.7
Fertilizantes 0.6 4.9 0.1 1.2
Total 1.05 14.9 13.9 2.8

58 capítulo 3. generalidades sobre la fijaciónbiológica del nitrógeno atmosférico
de la fijación de Nitrógeno atmosférico en la caña de azúcar en Brasil, donde hay unas 2.5 millones de hectáreas dedicadas a este cultivo. Si se calcula un modesto rendi-miento promedio de 70 t/ha y 10% de materia seca con un contenido de 1.4% de Nitrógeno, del cual 15% procede de la fijación atmosférica por las bacterias que viven normalmente en el suelo y por las bacterias endofíticas, resultará que en la superficie cañera de Brasil se fijarán unas 43.000 toneladas de Nitrógeno, que representan un ahorro no menor de 20 millones de USD. Esta fijación es natural y gratuita, y demues-tra que todos los incrementos en los aportes de Nitrógeno que puedan lograrse por la aplicación efectiva de los biofertilizantes tienen un elevado valor económico (Urquiaga et al., 2001).
También en el caso de Brasil, considerando el rendimiento promedio que se obtiene en soya sobre una superficie de 13 millones de hectáreas, que asciende a 2 400 kg/ha y tiene un contenido de Nitrógeno en los granos de 6.5%, puede decirse que se ex-portan anualmente del sistema suelo-planta 169 kg/ha del elemento. Si se asume que en los residuos del cultivo se encuentran 60 kg/ha de Nitrógeno, puede concluirse que la soya acumula en el año 230 kg/ha del nutriente, sin aplicar fertilizante. De estos re-sultados se deduce que la fijación biológica del Nitrógeno, solo en el caso de la soya, le representa a Brasil una economía de fertilizante equivalente a 1 500 millones de USD (Urquiaga et al., 2001).
Vose et al. (1982) calcularon que una ganancia de solo 10% de Nitrógeno por los cultivos mediante la fijación biológica debida a la aplicación de estos inoculantes repre-sentaría 3 mil millones de dólares anualmente si ocurriera en todas las áreas cultivadas del mundo. Igualmente, en la India se ha calculado que si en aquel país se aumentaran solamente 10 kg/ha de Nitrógeno mediante la fijación biológica aportada por estos bioproductos, el valor económico sería de 55 millones de USD (Subba Rao, 1982).
Todo lo que se ha expuesto demuestra la importancia ecológica y económica del uso de los biofertilizantes y bioestimuladores. En un gran número de países se trabaja ac-tualmente por incrementar los niveles de aplicación con notable éxito, y los beneficios económicos que se obtienen con la generalización de las nuevas tecnologías se van aproximando con rapidez a las potencialidades que han sido calculadas por diversos autores.

59la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Capítulo 4Estimulación del desarrollo de las plantaspor las bacterias fijadoras de nitrógeno atmosférico
Sustancias reguladoras del crecimiento vegetal
El desarrollo de los seres vivos se realiza mediante cambios que ocurren en el cre-cimiento y diferenciación de células, tejidos y órganos. En el caso de las plantas, la integración del crecimiento en distintos órganos y la coordinación del desarrollo entre ellos debe ser regulada por un grupo de hormonas, que pueden ser definidas como sustancias orgánicas naturales que influyen sobre los procesos fisiológicos en bajas con-centraciones. La síntesis de estas sustancias puede ser localizada o puede ocurrir den-tro de varios tipos de tejidos y células. Algunas veces pueden ser transportadas, pero en la mayor parte de los casos actúan en los tejidos o células donde son sintetizadas.
A diferencia de las hormonas animales, cuyo efecto es específico, una hormona vegetal puede inducir varias respuestas diferentes, en dependencia del tipo de tejido o célula donde actúa. La magnitud de la respuesta puede depender de muchos factores, tales como la concentración activa, la especie vegetal, la sensibilidad del tejido, la estabilidad y las interacciones con los microorganismos.
Por todas estas razones, el término hormona vegetal ha sido sustituído por el de sustancia del crecimiento vegetal o reguladora del crecimiento vegetal y se identifi-can actualmente como PGRs (Plant Growth Regulators). De acuerdo con Arshad y Frankenberger (1993), cuando estas sustancias son sintetizadas de forma endógena por las plantas deben llamarse hormonas vegetales, mientras que el término PGR in-cluye muchos compuestos naturales y sintéticos, ya que las plantas pueden responder a las aplicaciones exógenas durante ciertas etapas de crecimiento y bajo determinadas condiciones de cultivo.
Según Nieto y Frankenberger (1990), el descubrimiento de las sustancias reguladoras del crecimiento vegetal ocurrió en los trabajos realizados por Darwin sobre el fototro-pismo de los coleóptilos de las gramíneas, en los cuales se determinó que una señal transportada era responsable de la curvatura inducida en la planta por un estílmulo

60 capítulo 4. estimulación del desarrollo de las plantas porlas bacterias fijadores de nitrógeno atmosférico
luminoso. El mensajero químico relacionado con este proceso fue aislado por prime-ra vez por F. W. Went en 1928 y se le llamó auxina, palabra procedente de la griega auxein, que significa crecer.
A partir de entonces, se han descubierto e identificado otras varias sustancias regulado-ras del crecimiento vegetal, entre las cuales han sido las más estudiadas las siguientes: auxinas, giberelinas, citoquininas, ácido abscísico y etileno y, más modernamente, ácido jasmónico. Estas sustancias no actúan solas, sino en conjunción u oposición unas con otras. La acción final sobre el crecimiento y diferenciación de una planta representa el efecto neto de un balance hormonal. También las acciones que pueden ocurrir depen-den de la concentración; así, si ésta es baja, una sustancia puede ser estimuladora, pero si es alta puede ser inhibidora.
Hay microorganismos que son capaces de sintetizar estas sustancias biológicamente activas y, como resultado de la estrecha relación que existe entre los microorganismos rizosféricos y la superficie de las raíces, la producción exógena de PGRs por parte de aquellos afecta el crecimiento de las plantas. Este suministro exógeno puede re-presentar cantidades suplementarias a los niveles endógenos de las plantas o puede alterar la capacidad de síntesis de las plantas y el metabolismo de otros PGRs dentro del sistema.
La productividad de los cultivos puede ser mejorada mediante la manipulación de los microorganismos rizosféricos que tienen la capacidad de sintetizar sustancias activas promotoras del crecimiento vegetal, además de que pueden modificar la fertilidad del suelo y facilitar el establecimiento de las plantas. En el caso de las bacterias que poseen esta capacidad, aunque muchos hongos también la poseen, se las ha incluído en un grupo que se conoce como Rizobacterias Promotoras del Crecimiento Vegetal, más comúnmente llamadas PGPR (Plant Growing Promoting Rhizobacteria), Algunas de PGPR (Plant Growing Promoting Rhizobacteria), Algunas de PGPRestas bacterias han sido llamadas también Rizobacterias Promotoras de la Emergencia o EPR (Emergency Promoting Rhizobacteria), ya que este efecto es de gran valor al estimular la emergencia de las plantas en suelos con pobre estructura, como los de las zonas áridas y semi-áridas (Requena et al., 1997).

61la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Mecanismos de estimulación del crecimientopor parte de las bacterias fijadoras de Nitrógeno
Las bacterias fijadoras de Nitrógeno, especialmente las que lo hacen de forma aso-ciativa, sintetizan sustancias activas reguladoras del crecimiento de las plantas, por lo cual son consideradas también como PGPR, e incluso en numerosas publicaciones se concede mayor importancia a esta actividad que a la fijadora.por parte de estas bacte-rias. Así, en cultivos a los que se aplicó todo el Nitrógeno que necesitaban, Kapulnik et al. (1985) y Sarig et al. (1990) incrementaron el desarrollo y el rendimiento de maíz, sorgo y trigo inoculando Azospirillum y Kloeppper et al. (1994) lograron aumentar 30% el rendimiento del trigo con la inoculación de Azotobacter chroococcum.
Se conocen dos tipos de mecanismos mediante los cuales las bacterias fijadoras de Nitrógeno actúan como PGPR, uno de acción directa y otro de acción indirecta (Glick, 1995). El primero consiste en la síntesis de fitohormonas y de algunas enzimas, tales como la deaminasa ACC, las cuales modulan el nivel de las hormonas en las plantas; el segundo se manifiesta en la reducción de la acción perjudicial de los organismos fito-patógenos mediante la síntesis de sideróforos y antibióticos por parte de las bacterias (Rodríguez y Fraga, 1999), efecto que se demuestra más adelante, en los casos de A. chroococcum y Azospirillum brasilense, en la Tabla 23.
El género bacteriano más importante en lo que se refiere a la síntesis de hormonas ve-getales, entre los fijadores de Nitrógeno, es Azotobacter, como se pone de manifiesto en la Tabla 9, confeccionada a partir de datos de Arshad y Frankenberger (1993). Debi-do a que no se han reportado microorganismos fijadores capaces de sintetizar etileno ni ácido abscísico no se incluyen estos PGRs en la tabla.
El conjunto de estas sustancias, que son asimiladas por las plantas a través de las raíces, permite que cada una de ellas actúe en el momento en que la planta lo requiera; así, algunas estimulan el desarrollo de las raíces o el de la planta entera; otras aumentan la floración o reducen el aborto floral; por último, algunas posibilitan que el fruto se forme y madure en un tiempo menor. Todos estos efectos permiten el desarrollo más precoz de plantas más vigorosas y el incremento del rendimiento en niveles superiores a los que se obtienen en los países de clima templado.

62 capítulo 4. estimulación del desarrollo de las plantas porlas bacterias fijadores de nitrógeno atmosférico
Auxinas
Número de especies bacterianas sintetizadoras: 68Géneros más importantes: Pseudomonas (10 especies)
Bacillus (9)Arthrobacter (8)Arthrobacter (8)ArthrobacterAzotobacter (6)Azotobacter (6)Azotobacter
Número de especies fúmero de especies fúmero de especies f ngicas sintetizadoras: 56Géneros más importantes: Penicillium (12 especies)
Fusarium (7)
Giberelinas
Número de especies bacterianas sintetizadoras: 21Géneros más importantes: Azotobacter (5 especies)
Bacillus (4)Pseudomonas (3)
Número de especies fúmero de especies fúmero de especies f ngicas sintetizadoras: 15Géneros más importantes: Fusarium (5 especies)
Citoquininas
Número de especies bacterianas sintetizadoras: 21Géneros más importantes: Azotobacter (4 especies)
Pseudomonas (3)Número de especies fúmero de especies fúmero de especies f ngicas sintetizadoras: 24Género más importante: Taphrina (4)
Tabla 9
Microorganismos productores de hormonas vegetales

63la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Tabla 10
Auxinas sintetizadas por bacterias fijadoras de Nitrógeno
Síntesis de auxinas por las bacterias fijadoras
En el caso de las auxinas, el ácido indol-3-acético (IAA) es el compuesto más activo y el principal en la mayor parte de las especies vegetales, y es sintetizado a partir de trip-tófano en los tejidos meristemáticos de las hojas jóvenes, flores, semillas y frutos. Las principales funciones que se atribuyen a las auxinas incluyen el alargamiento y la división celular, la iniciación de las raíces, respuestas tropísticas y dominancia apical.
Como se deduce de la Tabla 9, hay un gran número de microorganismos que son capaces de sintetizar auxinas en cultivo puro y en el suelo, y se considera que aquellos que se encuentran en la rizosfera y el rizoplano las sintetizan con más facilidad que los habitantes del suelo libre, aunque todos necesitan la presencia de un precursor como triptófano, el cual se encuentra entre los aminoácidos que son liberados por las raíces.
En la Tabla 10 se muestran las especies de bacterias fijadoras de Nitrógeno que son capaces de sintetizar distintas auxinas. El hecho de que no aparezcan otras especes en la lista no significa que no sean capaces también de prducirlas, sino que no han sido estudiadas.
Auxina Especie bacteriana
Ácido indolacéticoÁcido indolcarboxílicoÁcido indolpirúvicoÁcido indol-lácticoIndolacetaminaHeteroauxina
Azospirillum brasilenseAzospirillum lipoferumAzotobacter beijerinckiiAzotobacter chroococcumAzotobacter paspaliAzotobacter vinelandiiGluconacetobacter diazotrophicusRhizobium leguminosarumRhizobium melilotiRhizobium phaseoliRhizobium trifolii
Frankia sppAzospirillum brasilenseRhizobium leguminosarumRhizobium trifoliiRhizobium leguminosarumRhizobium trifoliiAzospirillum brasilenseAzospirillum lipoferumAzotobacter chroococcumAzotobacter agile

64 capítulo 4. estimulación del desarrollo de las plantas porlas bacterias fijadores de nitrógeno atmosférico
Puede verse la gama de auxinas sintetizadas por las bacterias de los géneros Azospiri-llum, Azotobacter y Azotobacter y Azotobacter Rhizobium, destacándose las especies Azospirillum brasilense, Rhi-zobium leguminosarum y R. trifolii, con tres auxinas cada una, y Azospirillum lipoferum y con tres auxinas cada una, y Azospirillum lipoferum y con tres auxinas cada una, yAzotobacter chroococcum con dos.
En el caso de las especies de Rhizobium, las auxinas que sintetizan promueven la for-mación de raíces laterales en las leguminosas e inhiben su elongación, dando origen a los nódulos. Sin embargo, las auxinas solas no influyen en la diferenciación vascular, por lo que estas bacterias liberan también citoquininas en la superficie de las raíces o en los filamentos de infección que, junto con las auxinas, estimulan la división celular en la corteza. En raíces de leguminosas no inoculadas o en suspensiones de Rhizobium phaseoli sin la presencia de la leguminosa no se han detectado cantidades significativas de citoquininas (Nieto y Frankerberger, 1990). Es decir, que la asociación entre auxinas y citoquininas es una consecuencia de la simbiosis bacterias-plantas.
También se ha observado que la presencia de altas poblaciones de Azospirillum brasilen-se en las raíces de las gramíneas afecta marcadamente el desarrollo de las plantas por la presencia de auxinas sintetizadas por las bacterias (Barbieri et al., 1988).
En la Figura 1 se muestra la influencia de la inoculación con distintas cepas de Azotobac-ter sp.sobre el desarrollo de las raíces de plántulas de tomate.
En la figura se distingue el efecto de las distintas cepas y se pone de manifiesto que algunas de ellas no ejercen influencia sobre el desarrollo de las raíces, lo que puede atribuirse a que no son capaces de sintetizar PGRs. Otras cepas, en cambio, muestran una alta efectividad.
Figura 1
Efecto estimulador e inhibidor de distintas cepas de Azotobacter sp. sobre el
desarrollo de raices de tomate

65la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Tabla 11
Giberelinas sintetizadas por bacterias fijadoras de Nitrógeno
Síntesis de giberelinas por las bacterias fijadoras
Las giberelinas (GAs) son las fitohormonas menos estudiadas como metabolito micro-biano en el sistema suelo-planta. La falta de información se debe a que existen muchas dificultades para su detección analítica.
En las plantas, estas hormonas son sintetizadas a partir del ácido mevalónico en los brotes, semillas en desarrollo y raíces. Estimulan el desarrollo del tallo, inducen la ger-minación de las semillas, el desarrollo de los frutos y la inducción de flores en las plantas dióicas. En el caso de los microorganismos, ha sido reportada su presencia en un nú-mero limitado de bacterias y hongos.
En la Tabla 11 pueden verse las especies de bacterias fijadoras de Nítrógeno que han demostrado su capacidad para sintetizar GAs.
El hecho de que la mayor parte de las bacterias fijadoras aparezcan como capaces de sintetizar solamente GA3 puede deberse a la falta de estudios para detectar otra GA. En el caso de Rhizobium phaseoli se reportan en la tabla 4 giberelinas (Atzorn et al., 1988),
Giberelina Especie bacteriana
GA1 Azospirillum lipoferum Rhizobium phaseoli
Azospirillum brasiliense Azospirillum lipoferum Azotobacter sp. Azotobacter beijerinckii
GA3 Azotobacter chroococcum Azotobacter paspali Azotobacter vinelandii Gluconacetobacter diazotrophicus Rhizobium phaseoli Frankia sp.
GA4 GA4 GA Rhizobium phaseoli
GA9 Rhizobium phaseoli

66 capítulo 4. estimulación del desarrollo de las plantas porlas bacterias fijadores de nitrógeno atmosférico
pero quizás con esta bacteria los estudios fueron más profundos que con las demás debido a las dificultades que se señalaron antes.
Síntesis de citoquininas por las bacterias fijadoras
Las citoquininas constituyen uno de los mayores grupos de hormonas del crecimiento vegetal sintetizadas por las plantas y microorganismos. Se han identificado 50 citoqui-ninas y sus metabolitos y, de ellas, las más activas son los derivados de la adenina. El efecto principal de estas sustancias consiste en promover la formación de flores y frutos con mayor desarrollo y aumentar los rendimientos.
En la Tabla 12 se presentan las bacterias fijadoras de Nitrógeno capaces de sintetizar algunas de las citoquininas más importantes.
Citoquinina Especie bacteriana
Isopentenil adenosina Azospirillum brasiliense Rhizobium japonicum Rhizobium leguminosarum
Isopentenil adenina Azospirillum brasilense Azotobacter beijerinckii
Azotobacter chroococcumAzotobacter paspali
Azotobacter vinelandiiGluconacetobacter diazotrophicus
Rhizobium japonicumRhizobium leguminosarumRhizobium phaseoli
Cis zeatina Azospirillum brasilense Azotobacter chroococcum Rhizobium japonicum
Zeatina ribósido Azotobacter chroococcumTrans zeatina Azotobacter chrococcumDihidrozeatina ribósido Azotobacter chroococcum
Tabla 12
Citoquininas sintetizadas por bacterias fijadoras
de Nitrógeno

67la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Tabla 13
Producción de vitaminas por A. chrococcum cepa INIFAT-12
Como se observa, la mayor parte de las especies que aparecen en esta tabla son también capaces de sintetizar auxinas y giberelinas (Tablas 10 y 11). La especie que mayor número de citoquininas sintetiza es Azotobacter chroococcum (6), por lo cual es necesario poner énfasis en esta bacteria, aunque también Azospirillum brasilense tiene un papel destacado.
Sustancias activas sintetizadas por Azotobacter chroococcum
Estudios realizados con diferentes cepas de A. chroococcum que han sido aisladas en diferentes condiciones edafoclimáticas han permitido conocer que esta bacteria sinteti-za una amplia gama de sustancias biológicamente activas que estimulan el desarrollo y el rendimiento de los cultivos económicos. En las Tablas 13, 14 y 15 se presentan los resultados obenidos en los estudios que se han realizado con cepas aisladas de suelos de Cuba con el fin de conocer su capacidad para sintetizar diversas sustancias activas (Dibut, 2000).
Las concentraciones que aparecen en la tabla 13 pueden considerarse elevadas. Tam-bién en las bacterias del género Azospirillum se han señalado altas concentraciones de vitaminas. Así, Rodelas (2001) reportó una síntesis de tiamina ascendente a 50-100 mg/g de materia celular seca y de ácido nicotínico equivalente a 200-600 mg/g de materia celular seca.
Puede observarse una gama de 14 aminoácidos y una concentración muy alta de mu-chos de ellos.
Vitamina Concentración (mg/100 mL)
Tiamina 5.7Riboflavina 44.0Piridoxina 18.0Ácido Fólico 3.5

68 capítulo 4. estimulación del desarrollo de las plantas porlas bacterias fijadores de nitrógeno atmosférico
Aminoácidos Concentración en proceso de fermentación (n mol./ mL).
Acido aspártico 71.05 Serina 61.05 Glicina 127.35 Valina 38.70 Isoleucina 20.05 Ácido Glutámico 82.15 Ornitina 0.83 Glisina 9.40 Arginia 4.45 Treonina 58.80 Leucina 35.95 Fenilalanina 66.55 Prolina 60.60 Tirosina 2.87
Concentración total 728.90
Tipo de sustancia reguladora Actividad (mg/L)
Actividad Auxínica (Eq. AIA) 14.47Actividad Giberélica (Eq. A3 G) 30.20Actividad Citoquinínica (Eq. Kinetina) 32.50
Los valores que se muestran en la tabla 15 son elevados. Por lo general, en el caso de síntesis de fitohormonas es superior la gama y la concentración que es capaz de producir una cepa aislada de un suelo tropical en comparación con otra cepa de la misma especie aislada de un suelo templado. Así, por ejemplo, en lo que respecta a la actividad citquinínica relativa a Kinetina fue de 4.4 mg/L en el caso de una cepa aislada de un suelo templado (González-López et al., 1986) y de 32.5 mg/L en la aislada del suelo tropical (Tabla 15).
Tabla 14
Producción de aminoácidos por la cepa INIFAT-12 de
Azotobacter chroococcum
Tabla 15
Producción de PGRs por A. chroococcum cepa INIFAT-12

69la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Tabla 16
Porcentaje de plantas de maíz emergidas y tiempo de aparición de la primera hoja verdadera en plantas inoculadas y sin inocular con la cepa Guárico-1 de Azotobacter spAzotobacter spAzotobacter s .
La síntesis de esta amplia variedad de sustancias activas por parte de las bacterias fijado-ras de Nitrógeno hace que tengan un efecto positivo marcado desde las primeras eta-pas del desarrollo de la planta, independientemente del suministro de Nitrógeno que son capaces de aportar. Esto se pone de manifiesto desde el estadío de emergencia de las plántulas, como se demuestra en la Tabla 16, en la que se presenta la actividad ETP de una cepa de Azotobacter sp. aislada de un suelo de Venezuela sobre la emergencia Azotobacter sp. aislada de un suelo de Venezuela sobre la emergencia Azotobacterde plantas de maíz (López et al., 2007; en prensa).
Se observa en la tabla 16 el efecto estimulador de la emergencia por parte del inóculo preparado a partir de una cepa de Azotobacter, fijadora de Nitrógeno de forma aso-ciativa. Ya a los 4 días después de la siembra había emergido en esta variante el 75% de las semillas sembradas, en comparación con 52% de la variante sin inocular. 2 dias después emergió el 96% de las plantas, mientras en la variante sin inocular había germi-nado a los 10 días solo el 66%. En el caso del tiempo necesario para la aparición de la primera hoja verdadera se pone también de manifiesto la estimulación de la precocidad en la variante inoculada.
El efecto EPR que se pone de manifiesto en la tabla 16 puede deberse, según Noel et al. (1996), al mayor desarrollo de las raíces y del crecimiento del hipocotilo en los primeros estadíos de desarrollo de las plantas, como se puso de manifiesto en la Figura 2, lo cual le confiere una ventaja a estas plantas en una situación crítica del desarrollo. 2, lo cual le confiere una ventaja a estas plantas en una situación crítica del desarrollo. 2, lo cual le confiere una ventaja a estas plantas en una situación crí
Emergencia de lasplántulas (%)
Aparición de la primera hoja verdadera (%) en las
plantas emergidas
Variante 4 días 5-10 días 7 días 7-12 días
Sin inóculo 52 66 63 100Inoculadas 75 96 87 100

70 capítulo 4. estimulación del desarrollo de las plantas porlas bacterias fijadores de nitrógeno atmosférico

71la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Capítulo 5La fijación simbiótica del nitrógeno
Importancia de la fijación simbiótica
Entre los distintos sistemas biológicos que son capaces de fijar Nitrógeno, la simbiosis Rhizobium-leguminosas contribuye con el mayor aporte del elemento al ecosistema y a la producción de alimentos. Esta simbiosis constituye la asociación más elaborada y eficiente entre las plantas y las bacterias y, por esta razón, ha sido la más estudiada hasta el momento. Ya en el siglo XVI se dibujaron raíces de plantas leguminosas noduladas, aunque el agente causal permaneció desconocido en los tres siglos siguientes.
En los suelos ácidos de sabanas venezolanas, (España, et al., 2006), utilizando prácticas agrícolas con principios agroecológicos (rotación de cultivos, uso de roca fosforica, cul-tivares tolerantes a la acidez, mezcla de abonos orgánicos e inorgánicos en bajas dosis sin encalar), aplicando técnica isotópica 15N encontraron valores de fijación biológica de nitrógeno alrededor de 79%, generando un balance de nitrógeno positivo en el agrosistema, ya que el índice de cosecha de N fue bajo (40%); igualmente encontraron valores de fijación de N entre: 67% y 75%, utilizando leguminosas usadas como abono verde, éstos investigadores reportaron eficiencia de fijación de nitrógeno entre 62 y 72 mg N/pote y utilizando añil (I. lespediciones) la acumulación de N derivado del aire estuvo entre 29 y 40 mgN/pote cuando se uso C. Juncea.
Menos del 24% de las aproximadamente 16 500 especies de leguminosas conocidas han sido estudiadas en lo que se refiere a su capacidad para formar nódulos fijadores de N. Sin embargo, se ha determinado que más del 75% de las Caesalpinioideae son incapaces de nodular, mientras que 89% de las Mimosoideae y 96% de las Papilionoi-deae lo pueden hacer (Pueppke y Broughton, 1999). Si se toma en cuenta que la gran mayoría de las especies de leguminosas que se utilizan en la agricultura convencional pertenecen a las Papilionoideae es posible deducir los grandes beneficios que pueden obtenerse con el manejo adecuado del proceso de fijación simbiótica de N2.
Como consecuencia del gran número de estudios que se han realizado en el mundo, hoy se conoce que la contribución de la fijación simbiótica al suministro de Nitrógeno

capítulo 5. la fijación simbiótica del nitrógeno72
a las plantas cultivadas depende de la eficiencia de la asociación y de la disponibilidad del elemento en el suelo cuando los factores ambientales son adecuados. La gran importancia de este proceso para los cultivos económicos y el considerable ahorro de fertilizante nitrogenado para los países pobres se pone de manifiesto con los siguientes ejemplos:
En un suelo vertisol del sur de Aragua – Venezuela, (España et al., 2000), encontraron valores de FBN en soya de 54% bajo labranza convencional y de 67% aplicando siem-bra directa mas cincel cada 2 años.
Entre las leguminosas forrajeras, Centrosema pubescens nodula profusamente; en esta especie, apropiada para pastos bajos y resistentes a la sequía, se han reportado fijacio-nes anuales que ascienden a 320 Kg de N/ha. Pero las más elevadas se han encontrado en Leucaena leucocephala, usada extensamente en muchos países, que crece en pe-ríodos prolongados de sequía y suministra forraje con 30% de proteínas todo el año, la cual puede fijar anualmente hasta 800 kg de N/ha aunque, cuando las condiciones ambientales o biológicas no son favorables, esta especie no puede alcanzar más del 13% de la fijación que se logra en ambientes más apropiados (Norris, 1972).
En lo que respecta a las leguminosas de grano y aceite, en la Tabla 17 pueden verse las cantidades de N2 fijado por las principales especies, a partir de datos ofrecidos por Peoples et al. (1995).
Especie de leguminosa Cantidad de N2 fijado (KgN/ha)
Garbanzo (Cicer arietinum) 3-141Lenteja (Lens culinaris) 10-192Haba (Vicia faba) 53-330Lupino (Lupinus angustifolius) 32-288Soya (Glycine max) 0-450Glycine max) 0-450Glycine maxManí (í (í Arachis hypogaea (Arachis hypogaea ( ) 37-206Frijol común (Phaseolus vulgaris) 0-125Frijol mungo (Vigna radiata) 9-112
Tabla 17
Estimados experimentales de la cantidad de N2 derivado de la fijación atmosférica por las más importantes leguminosas
de grano y aceite

73la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Puede observarse en la tabla que el rango de N2 fijado por las leguminosas es muy grande, e incluso hay especies tan importantes como la soya y el frijol que pueden fallar en demostrar su capacidad para establecer la simbiosis en condiciones de producción. Estas variaciones se atribuyen al impacto de factores limitantes que ocurren en el cam-po, los que disminuyen en gran medida la capacidad de fijación.
Aunque en la mayor parte de los suelos del mundo existen bacterias simbióticas de los géneros Rhizobium y Bradyrhizobium, muchos no contienen las específicas para de-terminados hospederos. Por ejemplo, las vastas áreas naturales de suelos “cerrados” de Brasil no poseen cepas capaces de nodular soya, mientras que algunos suelos de Australia poseen estas cepas, pero son inefectivas (Gibson et al., 1982), por lo que es obligatorio aplicar siempre la inoculación con cepas seleccionadas de estas bacterias. Gracias a un trabajo eficiente en este sentido, actualmente se logran en la región de los “cerrados” rendimientos de grano de soya superiores a los 4 t/ha (Alves et al., 2003). Sin embargo, es interesante señalar que en suelos de Africa los cultivares asiáticos de soya nodularon con algunas cepas nativas pertenecientes a la miscelánea cowpea (Jara et al.,1981).
En el caso del frijol, la respuesta a la inoculación en condiciones de campo es variable. Esta es la leguminosa más importante para el consumo humano en América Latina y Brasil es el principal productor, por lo que se utiliza muy extensamente la inoculación con Rhizobium, pero es frecuente encontrar respuestas negativas. Por el contrario, el maní se considera una especie con alta promiscuidad simbiótica, como lo demuestra el hecho de que suspensiones de nódulos colectados de 51 especies de leguminosas silvestres de la India fueron capaces de inducir la nodulación en esta especie ( Martínez Viera,1986). En general, el maní se encuentra siempre bien nodulado en las condicio-nes naturales, por lo que no hay respuesta a la inoculación.
Establecimiento de la simbiosis bacterias fijadoras-leguminosas
Un aspecto fundamental para que se establezca una simbiosis efectiva consiste en la especificidad entre las bacterias y las plantas hospederas, la cual ocurre en tres niveles: la habilidad para formar nódulos, la eficiencia para fijar nitrógeno y el grado en que la fijación simbiótica provee a la planta de sus requerimientos del elemento. Durante muchas décadas, todas las bacterias capaces de inducir la formación de nódulos en las leguminosas fueron clasificadas dentro del género Rhizobium. Este género contenía

capítulo 5. la fijación simbiótica del nitrógeno74
una cantidad de especies, de acuerdo con su especificidad para una o varias especies o grupos de leguminosas.
Hoy, la profundización del conocimiento adquirida por la exitosa utilización de técnicas de investigación de última generación ha permitido descubrir numerosas nuevas espe-cies de bacterias simbióticas, las que se han agrupado en seis géneros: Allorhizobium, Azorhizobium, Bradyrhizobium, Mesorhizobium, Rhizobium y Sinorhizobium, los más im-portantes de los cuales desde una visión económica son Rhizobium y Bradyrhizobium, porque sus especies se asocian con las leguminosas de mayor importancia. Más ade-lante se hablará con más detenimiento acerca de esta moderna clasificación de las bacterias nodulares de las leguminosas. Independientemente de ella, existe consenso en el mundo para seguir llamando comúnmente a las bacterias que forman nódulos en las leguminosas con el nombre de rhizobium o bacterias del grupo rhizobium (Giller, 2001).
Los nódulos radicales son estructuras muy especializadas, adaptadas a la fijación del Nitrógeno para el beneficio de la planta en su conjunto, lo que permite su desarrollo cuando hay deficiencia de este elemento en el suelo. A pesar de que ni las plantas leguminosas ni los microsimbiontes pueden ser fijadores cuando están separados, la morfogénesis de los nódulos crea las condiciones que se requieren para la inducción y el funcionamiento del sistema nitrogenasa. Estos nódulos representan un tipo de crecimiento anormal, pero tan armoniosamente organizado que constituye una nueva estructura, considerada como un órgano sui generis que difiere de forma cualitativa y específica de los que se encuentran normalmente en las plantas. La frecuencia de distri-bución a lo largo del sistema radicular está determinada por factores que controlan los pasos en secuencia para la formación del nódulo:
a. Acumulación de las bacterias en la rizosfera de las plantas.
b. Infección de los pelos de las raíces, que culmina con la formación de los filamentos de infección dentro de la corteza.
c. Inducción del crecimiento meristemático local en la corteza de la raíz y formación del nódulo maduro.
d. Formación de los simbiosomas o bacteroides dentro de las células de las plantas debido a la transformación de las bacterias originales y, en este estadio, comienzo del proceso de fijación de Nitrógeno.

75la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Los procesos señalados en los puntos anteriores ocurren del siguiente modo: las bac-terias se adhieren a los pelos radicales por la acción de una proteína específica llamada ricadesina, que se encuentra en la superficie de la bacteria, y de las lectinas, proteínas que se identificaron en los extremos de los pelos de las raíces y también en la superficie de los microorganismos simbióticos (Sparvoli et al., 2001). Una vez que se establece la unión, los pelos radicales se encurvan y entrampan a las bacterias.
El enroscamiento es causado por ciertas sustancias específicas secretadas por las bacte-rias que se conocen como factores Nod, los cuales poseen características semejantes a las hormonas y son inductores de cambios en las membranas y en la oscilación de de-terminados iones (Ca2
+, Cl- y H-) hasta que se forman los filamentos de infección, den-tro de los cuales se multiplican las bacterias; estos filamentos crecen hasta alcanzar las células adyacentes a la raíz, cuya división se estimula por los factores Nod hasta que se forman los nódulos. Dentro de ellos se transforman las bacterias en bacteroides y co-mienza la fijación de N2. Todos estos cambios en las raíces ocurren por la activación en la planta de fosfolipasa C y proteínas G como respuesta a los factores Nod (Cárdenas et al., 1999, 2000; Irving et al. 2000; Pacios et al., 2001; Kelly e Irving, 2001,2003).
Desde que las bacterias penetran en las raíces se establece un nuevo intercambio de señales entre los simbiontes, lo que incluye la secreción de polisacáridos y proteínas por tres tipos de sistemas de excreción. De estas sustancias, solo se han identificado hasta el momento los flavonoides, los cuales regulan la expresión de genes y se están aplicando artificialmente a las semillas o al suelo para mejorar la nodulación y la fijación. En el caso de los factores Nod, este tipo de inoculación está descartado por el momen-to porque no se ha encontrado beneficio, sino más bien perjuicios (Hogg et al., 2002; Broughton et al., 2003).
Por último, los bacteroides son formaciones hinchadas, ramificadas y deformes, en que se transforman las bacterias, y pueden llegar a ser hasta 40 veces más grandes que los bacilos de los que se originaron. Ellos dependen totalmente de la planta para obtener sus fuentes de energía y, hasta que no se forman, no comienza la fijación de Nitrógeno. Cuando ocurre la senescencia del nódulo, estas formas pasan al suelo y la mayor parte de las veces no se reproducen, pero en los nódulos hay siempre bacterias originales en estado de latencia que se multiplican en el suelo, donde pueden mantenerse en estado libre, aunque no son fijadoras.

capítulo 5. la fijación simbiótica del nitrógeno76
Características de los nódulos
El color interno de los nódulos varía desde rojo hasta blanco, con una amplia escala de colores intermedios. Cuando la simbiosis funciona con efectividad, tienen una región roja relativamente grande, porque existe la presencia del pigmento leghemoglobina; en algunos nódulos puede encontrarse el pigmento rojo antocianina, que está asociado con simbiosis poco efectivas y puede ser confundido con aquel, pero es soluble en agua y se identifica con facilidad introduciendo los nódulos en el agua durante un corto tiempo, con lo que pierden la coloración. La leghemoglobina es similar a la hemoglo-bina de la sangre e integra un grupo de proteínas específicas del nódulo que se llaman nodulinas. Su función consiste en regular los niveles de O2 y aportar este elemento a los bacteroides.
En general, los nódulos de color claro indican poca o ninguna eficiencia en la fijación de nitrógeno y su distinto grado de longevidad refleja usualmente el hábito de crecimiento de la especie vegetal en la que se han formado. Así, los nódulos de las leguminosas herbáceas son frágiles y funcionan durante un período corto del crecimiento vegetativo de las plantas; el corte de éstas, las condiciones desfavorables de humedad, la falta de fotosíntesis y el ataque de nemátodos, insectos y hongos pueden conducir a su desin-tegración. Las especies perennes, en cambio, producen nódulos frescos, que pueden ser temporales o persistentes durante varios años.
Figura 2
Nódulos de frijol (Phaseolus vulgaris)

77la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Figura 3
Nódulos de garbanzo (Cicer arietinum L.)
Figura 4
Nódulos de soya (Glycine max L.)
La forma de los nódulos está determinada, en primer lugar, por la especie de planta leguminosa, aunque puede ser modificada por el grado de efectividad de la cepa bacte-riana y está definida por el material meristemático. Puede ser redonda, oval, alargada, en forma de maza, ramificada, coraloide o en forma de collar. En las figuras 2, 3 y 4 pueden verse algunas de estas formas de nódulos.
Las relaciones entre los nódulos radicales y el resto de la planta están gobernadas por el suministro de carbohidratos a las células bacteroides y por la transferencia de los productos de la fijación. Como se conoce, las sustancias que más comúnmente transporta la savia del floema es la sacarosa. Como productos de la fijación se forman aminoácidos, mediante la ligadura del amonio con los ácidos orgánicos que se produ-

capítulo 5. la fijación simbiótica del nitrógeno78
cen en los nódulos a partir de los azúcares transportados. Hoy se conoce con certeza que la cantidad de fotosintatos utilizables constituye uno de los principales factores que controlan la magnitud de la fijación.
Experimentos en los que se utilizó la reacción de reducción del acetileno no han mos-trado que ocurre una rápida declinación en la actividad nitrogenasa después de colocar las plantas en la oscuridad y después de su defoliación o decapitación; la mayor acti-vidad, en cambio, se encontró durante el día, aún en temperatura constante (Burris, 1978).
Los fotosintatos transportados a los nódulos son metabolizados rápidamente, suminis-trando así el reductor y el ATP requerido para la reducción de N2 a NH4, y también los esqueletos de Carbono necesarios para la formación de aminoácidos. La sacarosa, transportada con rapidez en glucosa y fructuosa por la acción de la invertasa presente en las células nodulares, representa así el principal sustrato para los bacteroides (Neyra, 1978).
Relaciones entre los nódulos y la capacidad de fijación de N2
La fijación de N2 total en la simbiosis bacterias fijadoras-leguminosas depende de dos parámetros: del peso de los nódulos y de su actividad específica de fijación (N2 fijado por g de nódulo). El peso de los nódulos va en aumento durante la etapa de crecimien-to de la planta, ocurriendo el punto máximo durante la segunda mitad del desarrollo, seguido por una disminución. Pueden existir grandes fluctuaciones, causadas por cam-bios de temperatura, intensidad de la luz, humedad o por efectos a largo plazo que relacionan estas variables con el estado de crecimiento de la planta.
La apariencia de la planta, el número, tamaño, peso y la intensidad del color de los nó-dulos, así como el contenido de Nitrógeno de distintas partes de la planta, constituyen características utilizadas para medir la respuesta a la simbiosis. De acuerdo con ellas, la nodulación puede clasificarse como efectiva o inefectiva. Sin embargo, estos dos términos no son suficientemente descriptivos como para distinguir todas las respuestas posibles de la planta a la nodulación, por lo que se elaboró un sistema más completo de clasificación que se muestra en la tabla 18 (Caldwell y Vest, 1977), adecuado para interpretar la mayor parte de las relaciones planta-inoculación que se han observado. Las descripciones se obtuvieron en un medio con Nitrógeno limitado.

79la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Tabla 18
Clasificación y descripción de las respuestas a la nodulación
De acuerdo con esta clasificación, así se establecerá el potencial simbiótico de una leguminosa dada que se asocia con una cepa fijadora. Este potencial se define como la actividad máxima de esta leguminosa cuando nodula eficientemente con una determinada cepa de bacteria fijadora simbiótica que está sometida a las más favorables condiciones ambientales.
Un importante hecho a tomar en cuenta durante la nodulación es el período que transcurre entre la inoculación y la aparición de los nódulos, lo que representa el co-mienzo de la fijación de N2. Este período depende de un número de factores, el más importante de los cuales es la cepa que se utiliza, ya que aquellas cepas que nodulan primero son activas competidoras contra las que lo hacen más tarde. Como se verá
No hay nódulos, ni evidencia externa sobre las raíces de que se ha iniciado su formación. La planta es pe-queña y clorótica, típica de deficiencia nitrogenada
Las raíces muestran que se ha inicia-do la formación de nódulos.
En las raíces se forman proliferaciones corticales y/o pequeñas estructuras semejantes a nódulos, con el in-terior de color blanco, verde o rosado. La planta es pe-queña y clorótica, típica de deficiencia nitrogenada.
Las raíces están noduladas.
El interior de los nódulos es de color verde o blan-co, algunas veces rosado. La planta es peque-ña, con un grado de clorosis que depende de la mayor o menor eficiencia de la fijación.
Los nódulos tienen el interior rosado o rojo. La planta es de color verde oscuro, con apariencia saludable.
Clasificación Descripción
I) No nodulación
II) Nodulación
A) Inefectiva
B) Efectiva
a) Ineficiente
b) Eficiente

capítulo 5. la fijación simbiótica del nitrógeno80
más adelante, el tamaño de la población de Rhizobium, la disponibilidad de humedad, la temperatura, el nivel de Nitrógeno en el suelo son otras determinantes importantes que afectan la velocidad de nodulación.
La fijación de N2 por un nódulo de leguminosa necesita tres fases: lag (durante la inicia-ción del nódulo y el desarrollo temprano), exponencial (rápido crecimiento del nódulo y desarrollo de la actividad nitrogenasa) y senescencia (ruptura del tejido nodular y declinación de la actividad nitrogenasa, la cual puede ser rápida o extenderse por un período considerable). La sequía y el exceso de humedad son dos factores ambientales que promueven esta senescencia. Los nódulos de tipo meristemático (Trifolium, Pisum y Phaseolus) muestran actividades nitrogenasa específicas, significativamente más altas que los nódulos de tipo esférico o de tipo determinado que se encuentran en muchas leguminosas tropicales. Una excepción la constituye el maní, el cual tiene una alta acti-vidad específica (Gibson et al., 1982).
Clasificación actual de las bacterias que formannódulos en las plantas leguminosas
Beijerinck aisló por primera vez, en 1888, un cultivo bacteriano puro proveniente de un nódulo y llamó a esta bacteria Bacillus radicicola. Posteriormente, Frank propuso el nombre de Rhizobium para estos aislados, lo que alcanzó general aceptación. Debido a la especificidad de los aislados bacterianos frente a determinadas especies vegetales hospedantes se crearon distintas especies de este género, y ya en 1929 existían seis de ellas, cada una de las cuales constituía un grupo de nodulación para un conjunto de distintas leguminosas hospedantes: R. japonicum, R. leguminosarum , R. lupini, R. meliloti, R. trifolii y R. phaseoli.
En 1982 se dividió en dos el género Rhizobium, dando origen al género Bradyrhizobium,con una sola especie, B. japonicum, que absorbió a la especie R. lupini, muy similar en muchos de los aspectos de taxonomía numérica y de hibridación ADN-ADN (Jordan, 1982). En los últimos años se desarrolló la taxonomía de las bacterias nodulantes con gran rapidez, y han aparecido muchas especies y géneros nuevos, los que se exponen a continuación, de acuerdo con Tao Wang y Martínez Romero (2003):

81la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Género Allorhizobium, que contiene solo la especie A. undicola, que fue aislada de nó-dulos de raíces de la planta acuática africana Neptunia natans y que es también capaz de nodular otras especies de plantas, entre ellas la alfalfa (Medicago sativanodular otras especies de plantas, entre ellas la alfalfa (Medicago sativanodular otras especies de plantas, entre ellas la alfalfa ( ).
Género Azorhizobium, que también contiene solamente la especie A. caulidonans, ais-lada de nódulos en los tallos de Sesbania rostrata y que puede además nodular las raíces. Esta especie es la única, entre las bacterias fijadoras simbióticas, que tiene la característica de formar nódulos en los tallos (Dreyfus et al., 1988), y solamente en la especie vegetal de que fue aislada, aunque se ha querido utilizar como promotora del crecimiento del arroz con relativo éxito.
Género Bradyrhizobium, que contiene las siguientes cuatro especies: B. elkani, B. ja-ponicum y B. liaoningense, que nodulan la soya, y B. yuanmingense, que solo forma nódulos en Lespedeza cuneata (Jordan, 1982). La especie más importante del género es B. japonicum que, además de la soya, tiene un amplio rango de plantas hospedantes, tanto de regiones tropicales como de zonas templadas.
Género Mesorhizobium, que contiene las siguientes ocho especies, capaces de formar nódulos en distintas especies de leguminosas de las zonas templadas, subtropicales y tropicales: M. amorphae, M. chacoense, M. ciceri, M. huakuii, M. loti, M. mediterra-neum, M. plurifarium y M. tianshanense.
Género Rhizobium, que contiene 13 especies, capaces de nodular diferentes especies de leguminosas en todas las regiones: R. etli, R. galegae, R. gallicum, R. giardinii, R. hai-nanense, R. huautlense, R. indigoferae, R. leguminosarum, R. loessense, R. mongolense, R. sullae, R. tropici y R. yanglingense. La especie más importante para América Latina es R. etli, que predomina en los nódulos del frijol (Phaseolus vulgaris L.).
Género Sinorhizobium, que contiene 9 especies: S. arboris, S. fredii, S. kostiense, S. kum-merowiae, S. medicae., S. meliloti, S. saheli, S. terangae y S. xinjiangense.
La mayor parte de estas especies son capaces de nodular a la soya y a Leucaena leuco-cephala. Recientemente se describieron dos especies no simbióticas en este género: S. morelenses y S. adhaerens. La primera fue aislada de nódulos de L. leucocephala pero no forma nódulos, aunque cuando se inocula puede estimular el crecimiento de la planta. La segunda fue transferida del género Ensifer y es una bacteria predatora de bacterias Ensifer y es una bacteria predatora de bacterias EnsiferGram positivas que viven en el suelo.

capítulo 5. la fijación simbiótica del nitrógeno82
Factores que influyen en el establecimiento de la simbiosis
Son muchos los factores limitantes para el establecimiento de la simbiosis Rhizobium o Bradyrhizobium-leguminosas, por lo cual deben ser discutidos detenidamente, ya que la presencia de algunos de ellos limita el éxito de la inoculación. En la tabla 19 se presen-tan los factores más importantes.
La nodulación y la fijación de Nitrógeno dependen de las interacciones entre una de-terminada leguminosa y las cepas de una especie de Rhizobium. Estas interacciones constituyen el resultado de la variación genética en la planta hospedera y en la bacteria y las relaciones entre los dos sistemas genéticos no son constantes, sino que dependen de los factores ambientales (Martínez Viera, 1986).
Factor Influencia
Relacionados con el macrosimbionteVariedad Sobre la nodulación y fijaciónCantidad de Nodulina Sobre la función nodularUtilización de fotosintatos Sobre la eficiencia de la fijaciónTolerancia al stress Sobre el establecimiento de la simbiosis
Relacionados con el microsimbionteInfertilidad Sobre la formación de nódulosEfectividad Sobre la fijaciónCompetitividad Sobre la ocupación del nóduloCompetencia saprofítica Sobre la persistencia en el suelo
Relacionados con el ambientePresencia de N combinado Sobre la nodulación y la fijaciónLuminosidad Sobre la eficiencia de la fijaciónTemperatura Sobre el crecimiento de la planta y la fijaciónAgua y aireación Sobre la actividad nitrogenasaSalinidad Sobre la viabilidad e infecciónPresencia de agentes bióticos Sobre la viabilidad e infección
Tabla 19
Principales factores que afectan la fijación
simbiótica de Nitrógeno

83la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
La influencia del suelo sobre la simbiosis Rhizobium-leguminosas es muy grande. Existe un balance muy delicado en las relaciones entre macro y microsimbionte, el cual es fácilmente perturbado por las características del suelo. Las bacterias de los géneros Rhizobium y Bradyrhizobium pueden sobrevivir en el suelo en ausencia de leguminosas, aunque su población es siempre más alta cuando se encuentran presentes las plantas hospedantes. Los exudados radicales estimulan la multiplicación, ya que suministran biotina y tiamina, homoserina y ácido glutámico, necesarios para el desarrollo de estas especies, pero son necesarias determinadas condiciones del suelo para que puedan sobrevivir las bacterias (Ledgard y Steele, 1992). Los aumentos más espectaculares de Nitrógeno procedente de la fijación simbiótica son obtenidos manipulando el sistema total suelo-planta-bacteria.
Es muy importante el efecto de las altas temperaturas del suelo sobre la superviven-cia de Rhizobium y Bradyrhizobium en condiciones de campo. El calentamiento bajo condiciones húmedas promueve la muerte de las bacterias de forma más rápida que bajo condiciones secas, por lo que un inoculante húmedo es más susceptible a las altas temperaturas que uno seco. Sin embargo, se han aislado cepas de Rhizobium y Bradyr-hizobium tolerantes a temperaturas elevadas y que forman nódulos efectivos en estas condiciones (Eagleshan et al., 1981).
La temperatura de la superficie del suelo puede limitar la población de las especies simbióticas más susceptibles. Este efecto se ejerce principalmente sobre la cantidad de N2 fijado, aunque cuando las temperaturas son muy altas pueden causar también po-bre nodulación. En el caso del frijol, por ejemplo, la temperatura constituye uno de los principales factores limitantes de la efectividad de la simbiosis en las regiones tropicales y subtropicales; así, en dos localidades situadas a 1.000 m y 1.800 m de altitud, en la zona de mayor altura, con temperatura máxima del suelo de 29°C, la fijación, deter-minada por la reducción de acetileno a etileno (ARA), fue equivalente a la reducción de 20 nmol C2H4/planta/h, mientras que en el lugar más bajo, donde se sobrepasan los 35°C durante varias horas cada día, la fijación máxima conseguida fue menor de 10 nmol C2 H4/planta/h (Pate, 1977).
El stress de humedad, debido a falta o exceso, es perjudicial para la simbiosis, ya que las bacterias muestran una rápida disminución de su viabilidad bajo condiciones de sequía y esto es exacerbado por las condiciones de humedad y sequía (Peña Cabriales y Alexan-der, 1979). La nodulación no ocurre cuando el suelo está seco porque falla la infección,

capítulo 5. la fijación simbiótica del nitrógeno84
y si la sequía ocurre después de la infección disminuye notablemente la fijación, debido a la pérdida de humedad de los nódulos o a que cesa la fotosíntesis. El suelo anegado (sobresaturados), principalmente cuando está asociado a pobre estructura del suelo, retarda la nodulación y la fijación de N2, lo que se debe a bajos niveles de O2.
De la misma manera, los factores nutricionales pueden afectar la fijación de N2 por un efecto directo sobre los procesos de infección y nodulación, ya que pueden influir sobre la actividad de la simbiosis y sobre el metabolismo o el crecimiento de las plantas, independientemente de cualquier efecto sobre la simbiosis. Cuando el pH del suelo es bajo no hay fijación de Nitrógeno o es muy escasa, lo cual se atribuye a la influencia sobre los siguientes aspectos de la simbiosis:
a. Supervivencia de las bacterias en el suelo;
b. Establecimiento en las raíces e inicio del nódulo
c. Eficacia de la simbiosis
d. Nutrición de las plantas
Los suelos ácidos cubren superficies muy extensas en las regiones tropicales. Así, en Brasil constituyen 68% de la superficie; en Colombia 57%; en Perú 44%; en Venezue-la 58%; en Panamá 63% (Lopes,1978). La mayoría de estas áreas están formadas por junglas, pero unas 300.000.000 ha son sabanas, en las que la cantidad y calidad del fo-rraje nativo son inadecuadas, y se ha demostrado que no pueden ser utilizados en estas regiones los resultados de los avanzados programas de inoculación de leguminosas de Australia, Estados Unidos y otras regiones, por lo que se han creado nuevos programas con alta efectividad en algunos países como Brasil, Colombia, India y otros, los cuales están adaptados a las condiciones de estos suelos.
En el oeste de África, Vigna unguiculata y maní son relativamente tolerantes a bajos pH (4-5), pero Leucaena leucocephala no crece ni nodula en estos suelos y responde marcadamente al encalado (Hutton y Andrews, 1978). Con soya en suelos ácidos de Nigeria, Bromfield y Ayanaba (1980) observaron que la inoculación con cepas de Bradyrhizobium japonicum seleccionadas por adaptarse a estas características producía buenos rendimientos (hasta 2 t/ha) y que el encalado no tenía influencia sobre esta respuesta. La acidez, deficiencia de Ca y toxicidad de Mn y Al tienden a ocurrir juntos y, por eso, es difícil señalar la causa exacta que influye sobre la fijación, sin embargo, (Es-

85la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
paña et al., 2006), han encontrado altos % de fijación de N en leguminosas de grano y forrajeras en suelos ácidos encalados.
El Fósforo constituye un importante factor limitante para la nodulación en los suelos tropicales. Los nódulos contienen frecuentemente 2-3 veces más Fósforo por unidad de materia seca que las raíces, por lo que la fertilización fosfórica o la presencia de microorganismos solubilizadores de Fósforo incrementa la nodulación y la fijación. Se ha demostrado que plantas con fuerte asociación micorrízica tienen requerimientos de P externo más bajos que las que tienen micorrización inefectiva (Habte y Manjunath, 1987).
La inhibición de la nodulación por la presencia de Nitrógeno combinado es reconocida por todos los especialistas, pero la naturaleza de la inhibición aún no se ha establecido claramente. Hasta el momento se conoce que se afectan tres fases: la infección de los pelos radicales, el crecimiento del nódulo y la actividad nitrogenasa. Igualmente, se ha demostrado que el nitrato ejerce un efecto sobre las lectinas (glicoproteínas específicas para que las bacterias reconozcan a la planta) sobre la superficie de los pelos radicales, lo cual ocasiona un bajo nivel de infección (Dazzo et al., 1981). El menor crecimiento del nódulo y la reducción de la actividad nitrogenasa parece ser consecuencia de un suministro más bajo de fotosintatos a los nódulos, debido a que estos fotosintatos los utiliza la planta para asimilar el Nitrógeno combinado (Gibson et al., 1982).
Debido a las fluctuaciones de los niveles de nitratos y de amonio en el suelo es difícil es-tablecer con seguridad la significación del Nitrógeno combinado en condiciones natu-rales. Los suelos tropicales son frecuentemente bajos en este elemento, pero pueden existir altos niveles de nitratos en campos no fertilizados por cortos períodos después de la estación seca, lo que puede retardar la nodulación y la fijación, como ha sido observado en maní en Senegal (Gibson et al., 1982).
De los microelementos que son esenciales para que ocurra una nodulación exitosa (Bo, Cu, Mo, Co), el Mo es el más deficiente en los suelos tropicales. En un survey de 41 suelos brasileños, 36 tuvieron deficiencia de Molibdeno (Franco et al., 1978). En Australia, la incorporación de Mo en el Superfosfato ha significado la diferencia entre fracaso y éxito en el positivo establecimiento de la nodulación en muchos suelos ácidos (Gibson et al.,1982); en Venezuela en suelos ácidos debe aplicarse Mo (molibdeno) para lograr una eficiencia en la FBN (López et al.,2006).

capítulo 5. la fijación simbiótica del nitrógeno86
En lo que respecta a la influencia que puede tener la aplicación de herbicidas, pue-de decirse que la susceptibilidad de las bacterias simbióticas frente a estos productos químicos de amplio uso ha sido investigada por métodos variados, que van desde el recuento en placas de las células bacterianas hasta el estudio de la nodulación de las plantas. Los resultados generales han mostrado que la susceptibilidad de la planta hospedante a los herbicidas es más grande que la de las bacterias. Las reducciones en la nodulación y en la fijación de Nitrógeno que se han encontrado en algunos de los trabajos han sido atribuidas a daños sufridos por las plantas, más bien que a efectos directos del herbicida sobre los microorganismos.
Cálculo de los beneficios aportados al suelopor la fijación simbiótica de N2 atmosférico
En general, la siembra de una leguminosa inoculada con una cepa eficiente de Rhizo-bium o Bradyrhizobium aporta importantes beneficios al suelo después de la cosecha, como consecuencia de los aportes de Nitrógeno suministrados por el cultivo y que ha recibido de forma gratuita por la fijación del N2 que se encuentra en la atmósfera. El balance de Nitrógeno que queda en un suelo después de cultivar una leguminosa inoculada puede calcularse aplicando la siguiente fórmula (Woods y Myers, 1987):
Balance neto de N = Nf – NIs
Nf = nitrógeno fijado= P x NI P = proporción de N en la planta derivado de la fijaciónNI = Cantidad total de N en la planta NIs = N extraído en la semilla cosechada
El valor P tiene que ser determinado por técnicas isotópicas, y varía de acuerdo con las condiciones edafo-climáticas de cada país e incluso entre regiones de un mismo país. En la Tabla 20 se presentan algunos valores determinados en diferentes países y en distintos cultivos (Peoples y Craswell, 1992), y en ella se pueden observar las notables oscilaciones que existen en los valores para un mismo cultivo, en dependencia de las condiciones en que se ha desarrollado.
Los abonos verdes vienen siendo utilizados cada vez más en la región, especialmente donde la posibilidad de uso de los fertilizantes nitrogenados está limitada por su alto costo. Dentro de los abonos verdes que tienen mayor potencial de uso en el trópico

87la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Tabla 20
N de la planta derivado de la fijación (P) en distintos cultivos y países
se incluyen no solo las tradicionales especies de Crotalaria, Mucura, Canavalia y Puera-ria, sino también algunas leguminosas de grano como caupí y la propia soya, especial-mente las variedades de rápida producción de biomasa y de alta eficiencia de fijación; la I. lespediciodes leguminosa nativa con alto potencial para ser usada como abono verde, así como el C. Juncea, representaron una alternativa para promover la sustentabilidad de agrosistemas de sabanas venezolanas (López, et al., 2006).
Los residuos de las leguminosas, debido a su baja relación C/N, generalmente presen-tan rápida descomposición, y muchas veces ésta no es una característica muy deseada, por lo cual se está utilizando en algunos lugares el cultivo mixto de leguminosas y gramí-neas para abonos verdes, que presenta una relación C/N intermedia y puede reducir la velocidad de mineralización de los resíduos y favorecer su mayor aprovechamiento por el cultivo principal o subsecuente (Urquiaga et al., 2001; López et al., 2006). De acuerdo con Calegari y Peñalva (1994), dentro de los abonos verdes es posible en-contrar plantas leguminosas que acumulan hasta 300 kg/ha de N y, según Urquiaga y Zapata (2000), hasta 80% de este N es derivado de la fijación.
A modo de ejemplo, es conocido que el cultivo del plátano ocupa un lugar muy re-levante en los países tropicales. Espíndola et al. (2000) determinaron que Pueraria phaseoloides es la especie con mayor poder de adaptación como cobertura viva en este
Cultivo País Valor de P
ManíAustralia
BrasilIndia
0.22-0.650.47-0.730.86-0.92
SoyaBrasil
Hawaii0.70-0.800.80-0.97
FrijolBrasilKeniaKenia
0.16-0.710.16-0.320.16-0.32

capítulo 5. la fijación simbiótica del nitrógeno88
cultivo, y supera hasta en 90% a las demás especies en producción de materia seca. Estos autores indicaron también que el 55% de todo el Nitrógeno acumulado por el plátano se derivó de la fijación por parte de esta leguminosa.
Por otra parte, una de las prácticas más corrientes de la moderna Agricultura Sustenta-ble consiste en la rotación de cultivos, usando siempre en primer lugar una especie de leguminosa inoculada con una cepa eficiente de Rhizobium o Bradyrhizobium. Este culti-vo le suministra al que viene posteriormente una cantidad apreciable de Nitrógeno, lo cual permite una reducción importante del fertilizante nitrogenado en el cultivo subse-cuente, además de que mejora la estructura del suelo y su disponibilidad de nutrientes en general. El valor del Nitrógeno aportado al suelo por la leguminosa que se utilizó en una rotación de cultivos y, por consiguiente, la cantidad de fertilizante que puede reducirse, se calcula de acuerdo con la siguiente fórmula (Myers y Wood, 1987):
Contribución al cultivo subsecuente = NI x P (1-NHi) x Fm x E
Donde: P = proporción de N en la planta derivado de la fijación NI = Cantidad total de N en la planta NHi = índice de N cosechado = NIs/NI NIs = N extraído en la semilla cosechada Fm = proporción del N que se mineraliza de la leguminosa E = Eficiencia de la utilización de este N mineral
Estos dos últimos valores deben ser el resultado de investigaciones encaminadas en esa dirección en distintas regiones.
Asociación de Rhizobium con cereales
En la última década se ha demostrado que Rhizobium puede colonizar de forma endó-fita a los cereales cuando se practica la rotación de cultivos con leguminosas inoculadas con determinadas especies de bacterias fijadoras simbióticas. Así, se ha demostrado que Rhizobium leguminosarum bv trifolii puede encontrarse de forma endófita en el arroz rotado con trébol cuando se practica la rotación en suelos drenados, para su-ministrar las condiciones aerobias que no ocurren en los campos de arroz inundados (Yanni et al., 2001). Después de aislar estas cepas endófitas, se puso de manifiesto que también eran capaces de establecerse dentro de las raíces del arroz cuando eran ino-culadas, sin necesidad de hacer la rotación.

89la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Pruebas efectuadas con rotaciones de trigo con otras leguminosas como trébol, alfalfa, soya, lenteja y caraota ratificaron que la única especie de Rhizobium capaz de estable-cerse dentro de las raíces del trigo fue también Rh. leguminosarum bv trifolii ( Dazzo y Yanni, 2006).
Estudios recientes han puesto de manifiesto que, tanto en el caso del trigo como en el arroz, esta invasión comienza por las raíces laterales y penetra en la raíz principal a través de las células epidérmicas, a lo que sigue una migración ascendente hacia la base del tallo y las hojas, donde desarrollan altas densidades de población que permanecen activas durante períodos relativamente largos (Chi et al., 2005).
Se han determinado los beneficios del establecimiento de esta simbiosis endofítica, y se ha encontrado que ocurre una germinación más rápida, alargamiento de las raíces, incremento del peso del tallo, mayor área foliar, contenido de clorofila, actividad foto-sintética y biomasa total, todo lo cual demuestra una actividad estimuladora del creci-miento por parte de las bacterias. Igualmente, se han encontrado mayores contenidos de Nitrógeno en la paja y en el grano en estado de madurez, aunque la procedencia de este Nitrógeno es el suelo, ya que se ha demostrado mediante pruebas de reduc-ción de acetileno y de 15N que no hay fijación de N2 (Biswas et al., 2000; Yanni et al., 2001).
Se han realizado una cantidad de experimentos en condiciones de campo, tratando las semillas con inóculo en polvo y aplicando solo 70% de fertilizante nitrogenado. Estos experimentos mostraron incrementos del rendimiento entre 18 y 30% en el caso del arroz, y entre 16 y 30% en el trigo (Dazzo y Yanni, 2006).
La capacidad de otras bacterias fijadoras de forma simbiótica para establecerse de for-ma endógena en cultivos cerealeros ha sido demostrada por algunos autores. Así, Gopalaswami et al. (2000) pusieron de manifiesto la colonización del xilema del arroz por Azorhizobium caulidonans , aunque no ha sido todavía bien estudiada.
Como puede verse, falta mucho aún por conocer acerca de estas asociaciones, ya que también se ha reportado que no se han obtenido resultados positivos en otros experi-mentos. La rapidez con que se está trabajando permite pronosticar que, en un tiempo relativamente corto, será posible incorporar las asociaciones Rhizobium-cereales a la gama de biofertilizantes que tantos beneficios aportan a la Agricultura Sustentable.

capítulo 5. la fijación simbiótica del nitrógeno90
Fijación simbiótica en plantas no leguminosas
Hay plantas no leguminosas que tienen también la capacidad de asociarse con deter-minados microorganismos para formar nódulos fijadores en las raíces. Muy pocas de las especies no leguminosas capaces de nodular lo hacen en asociación con Rhizobium y, de ellas, solo se ha demostrado que existe fijación de N2 en especies del género Pa-rasponia, perteneciente a la Familia de las Ulmaceae, que se encuentra extensamente distribuída en Nueva Guinea. Este género agrupa a cinco especies, tres de las cuales son capaces de establecer la simbiosis. El primer reporte sobre esta asociación lo hizo Trinick en 1973 (Trinick, 1988).
También se ha reportado nodulación rhizobiana en algunas especies del género Zygo-phyllum, que pertenece a la Familia Zygophyllaceae, distribuída en el desierto xerofíti-co. Mostafa y Mahmoud (1951) reportaron aislamiento de Rhizobium de nódulos de estas especies, el cual fue capaz de formar nódulos en Arachis hypogaea y Trifolium a lexandrinum. Sin embargo, Becking (1982) no encontró actividad nitrogenasa en estas inoculaciones.
Las plantas no leguminosas con nódulos radicales no formados por Rhizobium y que tienen una importante capacidad de fijación de Nitrógeno están presentes en un nú-mero de familias que no están relacionadas genéticamente, sobre todo en géneros de angiospermas y dicotiledóneas. Las especies noduladas se encuentran en una gran variedad de habitats y muestran una amplia gama de formas morfológicas. Algunas de ellas son herbáceas (Dryas sp.), otras son arbustos (Ceanothus sp y Colletia sp) y otras son especies maderables arbóreas (Alnus sp y Casuarina sp). Ason especies maderables arbóreas (Alnus sp y Casuarina sp). Ason especies maderables arbóreas ( unque estas especies no son de importancia agrícola, todas desempeñan un importante papel en las sucesiones vegetales de los ecosistemas naturales, porque pueden cubrir suelos de áreas pertur-badas.
Todas estas simbiosis son producidas por actinorrizas, y son causadas por actinomi-cetos que pertenecen al género Frankia, de la Familia Frankiaceae. En 1978 se realizó el primer aislamiento del actinomiceto endófito en nódulos de no leguminosas. La taxonomía del género se debe a Becking (1982), quien propuso el nombre de Frankiacomo homenaje al microbiólogo A.B. Frank y determinó 10 especies. Actualmente se considera que dentro del género se encuentran 4 grupos, que son los siguientes (Valdés et al., 2003):

91la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Grupo 1: Cepas infectivas en Myricaceae, Betulaceae y Casuarinaceae.
Grupo 2: Cepas que no pueden ser cultivadas in vitro y que forman nódulos en las familias Rosaceae, Coriariaceae y Datiscaceae.
Grupo 3: Cepas de Eleagnaceae.
Grupo 4: Cepas de Rhamnaceae.
Los nódulos formados en las plantas no leguminosas son diferentes, desde los puntos de vista anatómico y morfológico, de los que se forman en las plantas leguminosas. Pueden distinguirse dos tipos principales: el tipo Alnus, que es de forma coraloide por ramificación dicótoma, se origina en las raíces laterales y los meristemos apicales son inhibidos o con crecimiento muy lento. El segundo tipo es el Myrica-Casuarina, en el que el ápice de cada uno de los lóbulos en los nódulos produce una raíz normal, pero con geotropismo negativo; de esta manera, el nódulo llega a estar encerrado por raici-llas que crecen hacia arriba (Pawlowski y Bisseling, 1996).
El crecimiento inicial de estos nódulos inducidos por los actinomicetos también es di-ferente del que ocurre en las leguminosas. Primero se forma un pre-nódulo como un ligero engrosamiento de la raíz principal. A partir de este estadio se forma el nódulo a partir de un primordio lateral en la raíz que desarrolla por proliferación meristemática del periciclo y de las células corticales de la raíz principal. El estudio de la estructura fina de la endofita ha revelado la presencia de hifas, estructuras vesiculares y partículas semejantes a esporas (Torrey y Callaham, 1979).
En la Tabla 21 se muestran los géneros de plantas capaces de ser nodulados por los actinomicetos.
Como se ve en la Tabla 21, las plantas no leguminosas noduladas por especies de ac-tinomicetos del género Frankia comprenden 7 órdenes, 8 familias, 18 géneros y 175 especies de plantas dicotiledóneas.
La nodulación radicular en los géneros Casuarina, Allocasuarina y Gymnostoma es bien conocida en los trópicos. Sus especies aparecen espontáneamente en el Sudeste de Asia, incluyendo el Sudeste del Pacífico y Australia. Algunas de las 50 especies de los tres géneros se han introducido en el Norte de África, en áreas de América tropical y subtropical y en otras localidades de Asia.

capítulo 5. la fijación simbiótica del nitrógeno92
Orden Familia Género Número de especies noduladas
Casuarina 23 (40) Casuarinales Casuarinaceae Allocasuarina 2 (5) Gymnostoma 2 (5)
Myricales Myricaceae Myrica 26 (35) Comptonia 1 (1)
Fagales Betulaceae Alnus 33 (35)
Eleagnus 17 (45) Elaeagnaceae Hippophae 1 (3) Shepherdia 3 (3) Rhamnales Ceanothus 31 (55) Rhamnaceae Discaria 6 (10) Colletia 3 (17) Trevoa 1 (6)
Coriariales Coriariaceae Coriaria 14 (15)
Rubus 1 (250) Dryas 3 (4) Rosales Rosaceae Purshia 2 (2) Cercocarpus 4 (20)
Cucurbitales Datiscaceae Datisca 2 (2)
Tabla 21
Clasificación de las dicotiledóneas no
leguminosas fijadoras de N2 en simbiosis con actinomicetos del género Frankia
NOTA: Los números entre paréntesis indican el total de especies que
hay en cada género.

93la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
El género Alnus, con 35 especies, se encuentra principalmente en las regiones tem-pladas, aunque importantes representantes del género se encuentran también en los trópicos. Por ejemplo, las especies A. japonica y A. maritima y A. maritima y se han podido introducir en las altas elevaciones de Asia tropical.
El género Eleagnus tiene muchas especies que se encuentran en ecosistemas naturales en el Sudeste de Asia.
Varias especies de montaña que pertenecen al género Myrica se encuentran de forma natural en las regiones altas de Indonesia y Filipinas.
Algunas de las 15 especies del género Coriaria se encuentran en las altas elevaciones de Asia.
La especie Rubus elipticus, la única que nodula entre las 250 especies del género Rubus, se encuentra naturalmente en Asia continental y en algunas islas como Sry Lanka y Luzón.
Las especies de Colletia tienen su distribución principalmente en el trópico. La única especie que nodula de Trevoa vive en la región tropical.
El resto de los géneros que tienen especies capaces de nodular con Frankia spp se encuentran distribuídas en regiones templadas.
La importancia económica de las plantas no leguminosas noduladas radica en su capa-cidad para aumentar el contenido de Nitrógeno en el suelo, por lo que son muy útiles para el mejoramiento de suelos muy pobres en este elemento. Además, desempeñan un papel importante en la estabilización de los suelos después de que han ocurrido disturbios naturales, tales como inundaciones, erupciones volcánicas o grandes incen-dios.
Algunas especies que crecen bien en la zona tropical son de considerable importancia para la reforestación y la producción de madera. Por ejemplo, la Casuarina equisetifolia, de litoral, es particularmente útil en el desarrollo de vegetación en las dunas y en suelos muy arenosos cerca de las costas, mientras que la especie de montaña Casuarina jung-huhniana es buena colonizadora de las áreas erosionadas en las regiones montañosas de los trópicos.
Estas especies tienen un crecimiento relativamente rápido, combinado con un buen potencial de fijación de N2, y muestran habilidad para crecer en climas secos y ca-

capítulo 5. la fijación simbiótica del nitrógeno94
lientes. Así, en la península de Cabo Verde, al norte de Dakar, se encontró que el suelo tenía originalmente 80 kg de N/ha y, después de la reforestación con Casuarina equisetifolia, el mismo suelo tenía 309 kg de N/ha (Dommergues, 1963). Resultados similares que se han obtenido en distintas regiones han permitido sugerir la posibilidad de mezclar pinos tropicales con casuarinas.
En general, la capacidad de adaptación de las plantas actinorrícicas a suelos marginales está relacionada no solo con su capacidad para autoabastecerse de Nitrógeno, sino también a su facilidad para asociarse con hongos endo y ectomicorrízicos, lo que les permite proveerse de otros nutrientes, especialmente Fósforo.

95la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Capítulo 6La fijación asociativa de nitrógeno
La fijación biológica de Nitrógeno atmosférico también puede ser realizada por micro-organismos que no requieren la cooperación de otras formas vivas para cumplir esta función. Sin embargo, la mayor actividad fijadora de estos microorganismos de vida libre ocurre cuando se encuentran asociados a las plantas en las zonas de rizosfera y filosfera, donde aprovechan para utilizar en su desarrollo la excreción por las raíces y las hojas de notables cantidades de compuestos carbonados y otras sustancias.
La asociación entre plantas y bacterias fijadoras de Nitrógeno depende de la interacción entre ambos componentes, con el beneficio de por lo menos uno de los participantes (Dôbereiner, 1992). Estos organismos fijadores no forman simbiosis con especies ve-getales y pueden vivir libremente en el suelo o estableciendo relaciones denominadas asociativas, menos especializadas que la simbiosis, con diversas especies.
Hasta el momento, han sido estudiadas las asociaciones Azotobacter paspali-Paspalum notatum, Derxia y gramíneas, Beijerinckia sp y caña de azúcar, Azospirillum sp (A. brasi-lense, A. lipoferum, A. amazonensis) y monocotiledóneas (gramíneas, cereales y palme-ras), Bacillus sp (B. polymixa y B. azotofixans) con cereales y gramíneas, Pseudomonas sp en arroz, Campylobacter sp y Spartina alterniflora, Herbaspirillum seropedicae y cereales y Gluconacetobacter diazotrophicus y caña de azúcar. Con excepción de Campylobacter, que aparece en pantanos salinos, todas estas especies se encuentran en suelos agríco-las. Además, las especies del género Azotobacter son capaces de establecer asociación Azotobacter son capaces de establecer asociación Azotobactercon una gran cantidad de especies vegetales. Existen otras asociaciones de otros géne-ros de bacterias, pero son mucho menos importantes que las que se han citado.
Mecanismos de protección de la nitrogenasa en la fijación aeróbica
La mayor parte de las especies microbianas involucradas en la fijación asociativa son aerobias y, por ello, tienen que disponer de sistemas de protección de la nitrogenasa frente a O2, los cuales actúan a través de los siguientes mecanismos:

96 capítulo 6. la fijación asociativa de nitrógeno
– Protección respiratoria. Las células de Azotobacter son capaces de ajustar hasta un cierto nivel su coeficiente respiratorio, de manera que se mantenga la concen-tración de O2 en la superficie de la célula igual a cero. Esto significa, por ejemplo, que si se aumentara el potencial de O2 de un cultivo de Azotobacter que está en proceso de fijación, las células pasarían a respirar más rápidamente, procurando consumir todo el O2 a través de la respiración y gastando excesivamente las fuen-tes de carbono.
– Protección conformacional de la nitrogenasa. También en Azotobacter existen for-mas de nitrogenasa en las que la enzima muda a su forma parcial para proteger los puntos sensibles al O2. Es decir, que cuando hay exceso de O2 la enzima se desliga y cuando el O2 cae a una concentración satisfactoria la enzima liga, volviendo en-tonces a su forma activa.
– Producción de polisacáridos extracelulares. Muchas especies fijadoras producen di-versos polisacáridos extracelulares, cuya composición varía entre las especies. Por ejemplo, las células de Derxia spp y de Beijerinckia spp sintetizan un tipo de goma extremadamente elástica alrededor de sus colonias en determinadas condiciones. Se ha demostrado que la producción de polisacáridos sirve para impedir el acceso de O2 al centro de las colonias.
– Variación en el tamaño de las células. El establecimiento de una baja relación su-perficie celular/contenido celular representa un método de impedir el exceso de absorción de O2.
– Movimiento de las células hacia lugares con potencial de O2 adecuados. Debido a que no poseen mecanismos de protección más eficientes, muchas especies de bacterias aerobias son microaerofílicas cuando fijan N2. Las especies de los géne-ros Azospirillum y Herbaspirillum, por ejemplo, tienen un movimiento ambulatorio típico y son capaces de crecer en medio semi-sólido sin Nitrógeno, porque en medios con esta consistencia ocurren distintos gradientes de O2, de modo que las células pueden moverse hacia una zona donde el potencial de O2 sea adecuado. Cuando se multiplican las células, se aglomeran y se mueven hacia la superficie del medio, para formar una película característica en el punto donde haya un potencial de O2 conveniente. Si la película fuera rota por acción mecánica, la actividad nitro-genasa cesará y solo será restablecida cuando aquélla se vuelva a formar.

97la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Con la utilización de alguno de estos mecanismos, el más adecuado para la especie bacteriana de que se trate, es posible lograr el aislamiento selectivo de distintos orga-nismos aerobios fijadores de N2, mediante la utilización de medios de cultivo que no contienen Nitrógeno de forma combinada.
Microorganismos capaces de fijar N2 en forma asociativa
Mediante la utilización del método de reducción del acetileno a etileno se ha confeccio-nado una lista de organismos que son capaces de fijar N2 en condiciones de vida libre o en forma asociativa. Todos estos organismos son procarióticos, y no se conoce ningún eucariótico que sea capaz de realizar la fijación. Los géneros a que pertenecen estos organismos se incluyen a continuación (Bohlool et al., 1992; Elmerich et al., 1992; Boddey et al., 1998).
Organismos aeróbicos: Especies de los géneros Achromobacter, Aerobacter, Aquaspi-rillum, Azomonas, Azotobacter, Azotococcus Beijerinckia, Derxia, Methanosinus y otras bacterias oxidantes de metano, Pseudomonas. Además, se encuentran todas las algas verdi-azules heteroquísticas y algunos actinomicetos y hongos.
Organismos microaeróbicos: Especies de los géneros Azospirillum, Thiobacillus, Xan-thobacter. Además, se encuentran muchas algas verdi-azules no heteroquísticas.
Organismos facultativos: Especies de los géneros Bacillus, Burkholderia, Citrobacter, En-terobacter, Gluconacetobacter, Herbaspirillum, Klebsiella, Rhodomicrobium, Rhodopseu-domonas, Rhodospirillum.
Organismos anaeróbicos: Espécies de los géneros Thiopedia, Chlorobium, Chromatium, Clostridium, Desulfotomaculum, Desulfovibrio, Ectothiospira, Propionibacterium.
Esta amplia gama de organismos heterotróficos indica que la fijación de Nitrógeno ocurre en los más diversos ambientes del suelo, por la capacidad de adaptación de unas u otras especies. Por ejemplo, los campos arroceros constituyen un ambiente muy difícil para el desarrollo de los organismos del suelo. A pesar de esto, Matsuo y Takahashi (1977), en experimentos a largo plazo en Japón, calcularon el balance de Nitrógeno en los suelos arroceros sin aplicación del elemento y estimaron el suminis-tro natural promedio en tales campos entre 49 y 91 kg/ha/año y 70 kg/ha/año como promedio para todos los distritos de Japón. En Hokkaido (Japón), un experimento de

98 capítulo 6. la fijación asociativa de nitrógeno
arroz realizado durante 41 años sin fertilizante nitrogenado mostró un rendimiento promedio de 2.8 t/ha de grano sin que declinara la fertilidad del suelo (Shiga, 1971). La cantidad de Nitrógeno tomado del suelo por el cultivo fue 53 kg/ha/año. El contínuo suministro al suelo, a pesar de su toma por el cultivo, es consecuencia de la fijación.
En general, las algas verdi-azules han recibido más atención que las bacterias en rela-ción con la fijación de N2 en los suelos arroceros. Sin embargo, distintas especies de bacterias fijadoras se encuentran actuando con mucha frecuencia en los campos de arroz, con predominio de los géneros Azotobacter, Azotomonas, Arthrobacter, Bacillus, Beijerinckia, Desulfomaculatum, Desulfovibrio, Flavobacterium, Pseudomonas y bacterias oxidantes de metano. Más recientemente se agregaron a estos géneros Azospirillum, Enterobacter, Derxia y Vibrio (Yoshida y Rinaudo, 1982).
Factores ambientales que afectan la fijación asociativa de N2
Se conoce que algunas cepas se establecen mejor sobre unas especies vegetales que sobre otras, o que en determinados lugares son más efectivas que en otros (Boddey y Dôbereiner, 1988). Estas diferencias se atribuyen tanto a las peculiaridades de cada especie vegetal como a las de las distintas cepas de una misma especie bacteriana pero, sobre todo, se deben a las características edafoclimáticas de cada región.
Entre los numerosos factores ambientales que afectan la supervivencia y el crecimien-to de las bacterias fijadoras en el suelo se encuentra la disponibilidad de nutrientes orgánicos e inorgánicos, principalmente la posibilidad de acceso a fuentes de carbono abundantes para el crecimiento y la producción de energía. Estas bacterias pueden uti-lizar carbohidratos (monosacáridos, disacáridos y determinados polisacáridos), ácidos orgánicos, alcoholes monohídricos y polihídricos y otras sustancias. Incluso pueden uti-lizar algunos compuestos orgánicos volátiles aún en estado gaseoso, como por ejemplo vapores de alcohol etílico, de acetona y de algunos ácidos orgánicos volátiles.
La humedad limita toda la vida microbiana del suelo, así como la producción vegetal. Roper (1983,1984) encontró que condiciones húmedas y calientes incrementaban la actividad nitrogenasa en el suelo cuando se añadía paja y reportó que la mayor actividad nitrogenasa ocurre en el suelo en o por encima de la capacidad de campo. En gene-ral, los sistemas de fijación de Nitrógeno trabajan mejor cuando el suministro de agua llega a ser apropiado (Tchan y Kennedy, 1987). Sin embargo, está claro que algunos

99la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
diazotrofos son capaces de resistir prolongados períodos de sequedad, porque han podido ser aislados de suelos secos al aire que han sido almacenados por largos perío-dos (Moreno et al., 1986). Muchas de estas especies son capaces de formar quistes o células similares a quistes, lo cual ayuda a su supervivencia durante largos períodos de desecación (Lamm y Neyra, 1981).
Un factor muy importante que tiene gran influencia sobre la población de diazotrofos en el suelo consiste en la acción asociativa y antagonista de los organismos que viven normalmente en el suelo. Así, se conocen muchos microorganismos que son capaces de estimular el crecimiento de los organismos fijadores de N2, como por ejemplo los microorganismos celulolíticos, que son capaces de degradar los resíduos vegetales del suelo. Pero también hay otros muchos, como los actinomicetos y hongos, que actúan como antagonistas. Zuberer y Roth (1982) reportan numerosos ejemplos de inhibición de los diazotrofos en la rizosfera de las gramíneas por la acción de los actinomicetos. En general, en dependencia de los grupos microbianos que predominan normalmente en un determinado suelo, así será la efectividad de la cepa que se inocula.
Un requerimiento importante para la fijación de N2 de forma asociativa es la presencia de Fósforo asimilable, ya que las bacterias deben asimilar por lo menos 1 mg de Fós-foro por cada 5 a 10 mg de N2 que se fijan (Alexander, 1977). La acción limitadora de la falta de este elemento se debe a que tiene un efecto fundamental en el intercambio del Carbono, en la multiplicación de las bacterias y en el mismo proceso de fijación. El Potasio también es necesario, pero en cantidades relativamente más pequeñas que el Fósforo, e incluso es suficientemente conocido que altas dosis del elemento impiden el desarrollo de las bacterias fijadoras.
Calcio y Magnesio son muy importantes en el metabolismo de estas bacterias y su presencia es fundamental en el caso de algunos géneros como Azotobacter. La falta de estos elementos conduce a un alargamiento de la fase de crecimiento, así como a la vacualización e hinchamiento de las células, al mismo tiempo que se reduce la actividad metabólica y, en consecuencia, la capacidad de fijación de N2
.
Las bacterias que asimilan N2 pueden utilizar también amonio, nitratos y otras formas combinadas de Nitrógeno. En realidad, las sales de amonio son utilizadas con prefe-rencia, por lo que la presencia de estos compuestos inhiben la fijación, bien sea por su presencia directa o como consecuencia de la transformación de urea o nitratos.

100 capítulo 6. la fijación asociativa de nitrógeno
Muchos otros elementos inorgánicos son necesarios para el desarrollo de los micro-organismos, pero solamente unos pocos están implicados en el metabolismo del N2, siendo los principales Molibdeno, Manganeso, Hierro y Boro, que son críticos para la reacción de fijación, especialmente el Molibdeno, que no solo es fundamental para la fijación del N2, sino también para la utilización de los nitratos, aunque en este último caso en menor grado. En algunos organismos fijadores puede sustituirse el Molibdeno por Vanadio, aunque no es tan efectivo.
Las bacterias del género Azotobacter
Hasta la octava edición del Bergey´s Manual (1974) se consideraba que la Familia Azo-tobacteriaceae contenía cuatro géneros: Azotobacter, Beijerinckia, Derxia y Azomonas. En 1980, De Schmedt et al. sugirieron sacar el género Beijerinckia de la Familia, basán-dose en los análisis de rRNA, que demostraron que esté género no está relacionado con los demás. Algo semejante ocurrió con el género Derxia, ya que, además de los análisis de rRNA, las especies pertenecientes a este género tienen la capacidad de fijar N2 autotroficamente usando H2, propiedad que no tienen los otros géneros. Los análi-sis de taxonomía numérica y los inmunológicos, así como las diferencias morfológicas y fisiológicas ratificaron la separación de estos géneros, por lo que actualmente la Familia Azotobacteriaceae contiene solo los géneros
Azotobacter, Azotococcus y Azomonas, que pueden diferenciarse por la formación de quistes y la composición del DNA (Tchan, 1984).
Hoy se aceptan dentro del género Azotobacter las siguientes especies: A. chroococ-cum, A. vinelandii, A. beijerinckii, A. nigricans, A. armeniacus, A. paspali y A. macro-cytogenes (Crum, 2004).
Las bacterias que pertenecen a este género son Gram negativo, pleomórficas, es de-cir, de distintos tamaños y formas. Su tamaño oscila entre 2-10 x 1-2.5 µ. Las células jóvenes poseen flagelos peritricos que son usados como órganos locomotores. Las po-blaciones viejas incluyen formas encapsuladas resistentes al calor, a la desecación y, en general, a las condiciones adversas. Estos quistes germinan en condiciones favorables para dar células vegetativas. En las figuras 5 y 6, pueden verse colonias caracteristicas en el medio de Ashby y la morfología de células tìpicas de Azotobacter chroococcum.

101la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Figura 6
Tinciòn Gram negativa de células de A. chroococum
Figura 5
Colonias tipicas de Azotobacter sp. en medio de Ashby
Tabla 22
Efectividad de la Azotobacterina preparada en turba en experimentos realizados en Rusia.
En relación con estas bacterias, conocidas y estudiadas por primera vez por Beijerinck en 1901, se realizaron numerosas investigaciones en Rusia durante el siglo pasado, ob-teniéndose resultados que demostraron la efectividad de la inoculación en numerosos cultivos, como se pone de manifiesto en la Tabla 22 (Martínez Viera, 1986).
Como puede verse, la respuesta de todos los cultivos fue positiva, aunque, como se verá más adelante, estas respuestas son muy inferiores a las que se obtienen con la aplicación de cepas aisladas de países tropicales.
Cultivos Número de Promedio de incremento del experimentos rendimieno (%)
Algodón 21 13.7 Avena 77 8.0 Cebada 60 12.8 Centeno 15 6.6 Girasol 12 12.1 Maíz 9 15.5 Papa 86 9.7 Remolacha 126 8.8 Tabaco 3 10.7 Trigo 164 7.7

102 capítulo 6. la fijación asociativa de nitrógeno
Sin embargo, existieron grandes variaciones en la efectividad del mismo biopreparado cuando se aplicó en condiciones de producción en lo que era aquel extenso país, lo que se debió a errores cometidos en la selección de cepas por no haber tomado en cuenta la influencia del quimiotaxismo de las bacterias frente a las secreciones radicales y por haberse seleccionado un grupo muy reducido de cepas que, como es lógico, no podían tener una capacidad igual de adaptación para todos los tipos de suelos existen-tes en aquella gran superficie. Estos errores trajeron como consecuencia la desapari-ción del programa de aplicaciones de biofertilizantes en la antigua URSS.
En general, la capacidad de fijación de N2 por parte de Azotobacter puede variar gran-Azotobacter puede variar gran-Azotobacterdemente de acuerdo con las condiciones del cultivo y con las características del suelo, con su temperatura y el grado de aireación, con la presencia de otras fuentes de Nitró-geno, con el carácter de las fuentes de Carbono y con la presencia de determinados microelementos, así como con la frecuencia de los pasajes de la cepa en el laboratorio, ya que si estos son muy frecuentes puede llegar a perderse la capacidad fijadora. Pero la cantidad de N2 fijado depende, en primer lugar, de las características de la cepa que se utiliza, ya que pueden encontrarse cepas activas y pasivas, siendo estas últimas las que han perdido la capacidad fijadora en condiciones ambientales desfavorables. Una buena cepa activa no asimila más de 10-15 mg de N2 por gramo de carbono consumido, aun-que se han encontrado cepas capaces de asimilar hasta 30 mg (Madigan et al., 1997).
Jensen, en 1940, desarrolló un trabajo monumental encaminado a conocer el papel que desempeñaba Azotobacter en la economía del nitrógeno en las plantaciones de trigo de Australia. Uno de los resultados más importantes consistió en encontrar que las cantidades del elemento fijadas en el suelo por estas bacterias pueden calcularse de acuerdo con el número de células presentes. Así, en el suelo con escaso contenido de Nitrógeno ocurría una fijación de 1 ppm por 5-10 millones de células presentes en 1 g de suelo; cuando se agregaba paja, la fijación era de 0.5-1 mg de nitrógeno/g de paja.
La multiplicación de Azotobacter en el suelo depende en gran medida de la presencia Azotobacter en el suelo depende en gran medida de la presencia Azotobacterde Fósforo y Potasio, y su ausencia reduce los niveles de biomasa y, en consecuencia, disminuye la fijación. Las fuentes de Fósforo pueden ser inorgánicas y orgánicas, puesto que se ha establecido la presencia de fosfatasa activa en los cultivos de estas bacterias (Tchan, 1989). Igualmente, Kumar y Narula (1999) y Narula et al. (2000) reportaron el aislamiento de cepas naturales de Azotobacter chroococcum de suelos de la India capaces de solubilizar Fósforo del suelo a pH altos (8-8.5).

103la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
En términos generales, la fijación del Nitrógeno atmosférico por estas bacterias co-mienza cuando la concentración de Fósforo en el medio es de 0.004% y se inhibe cuando alcanza 0.8% (Mishustin y Silnikova, 1971). En el caso del Potasio, las cantida-des necesarias son menores y cantidades altas impiden la fijación. También la presencia de Calcio y Magnesio es muy importante y la fuente más idónea de Calcio es el car-bonato que, aún en concentraciones elevadas, no impide la asimilación de Nitrógeno. Molibdeno es fundamental para la fijación, en cantidades no mayores de 0.05 ppm, así como trazas de Boro, Manganeso, Zinc, Hierro y Cobalto.
Cuando el pO2 del suelo es muy bajo puede limitarse la fosforilación. Los compuestos de Carbono y algunos reductores como NAD(P)H tienden entonces a acumularse, lo cual conduce a un paso metabólico posterior en el cual la acetil coenzima A es utilizada junto con el NAD(P)H para la síntesis del polímero de almacenamiento poli-beta-hi-droxibutirato (PHB), el que puede alcanzar hasta el 70% del peso de las células y es degradado y usado cuando se restaura el suministro de O2. La presencia de PHB cons-tituye una de las características de las colonias de Azotobacter y también se encuentra Azotobacter y también se encuentra Azotobacteren algunas especies de Azospirillum y de las Rhizobiaceas.
En lo que respecta a la salinidad, se considera que estas bacterias tienen un buen nivel de resistencia, aunque no debe pensarse que son abundantes en los suelos salinos (Mishustin y Silnikova, 1971). La escasez de materia orgánica del suelo constituye un factor limitante (Vanlawe et al., 2000), así como pH inferiores a 5.
Se ha determinado también que Azotobacter produce una sustancia fungistática soluble en éter, termoestable, que inhibe el crecimiento de importantes hongos fitopatóge-nos como Fusarium sp, Helminthosporium sp, Alternaria sp y otros, y se ha logrado disminuir mediante la aplicación de Azotobacter la incidencia de enfermedades virales y bacterianas en papa, el crecimiento de hongos en semillas de trigo en germinación, las infecciones de Fusarium en trigo, de carbón en millo y de roya en trigo (Durkhead et al., 1998).
En la Tabla 23 se muestra el efecto inhibidor in vitro que tienen estas bacterias, así como las del género Azospirillum, sobre algunos importantes hongos fitopatógenos (Lakshmi Kumari et al., 1975).
Como se verá más adelante, estas bacterias también sintetizan una amplia gama de sustancias activas estimuladoras del crecimiento y el rendimiento de los cultivos, por lo

104 capítulo 6. la fijación asociativa de nitrógeno
Sclerotium rolfsii 22 17Sclerotium sclerotiorum 20 18Fusarium moniliforme 18 6Fusarium solani 18 2Fusarium oxysporum 18 2Alternaria brasicola 17 20Colletotrichum falcatum 23 20
Especie de hongo fitopatógeno
Efecto inhibitorio, expresado en mm de la zona de inhibición
Azotobacter chroococcum
Azospirillumbrasilense
que muchos autores le conceden mayor importancia a la capacidad para sintetizar estas sustancias (fitohormonas, vitaminas, aminoácidos) que a la actividad fijadora de nitróge-no. Filtrados libres de células no tienen efecto estimulador, por lo que se ha pensado que una proteína ligada a la célula, no identificada hasta el momento, es responsable por la estimulación (Subba Rao, 1996; Burdman et al., 2000).
Como se deduce de todo lo expuesto, la importancia de las bacterias del género Azotobacter es muy grande, por la doble función de aportar nitrógeno y sustancias Azotobacter es muy grande, por la doble función de aportar nitrógeno y sustancias Azotobacterreguladoras del crecimiento a los cultivos, así como de inhibir el crecimiento de micro-organismos fitopatógenos y hasta de solubilizar en determinadas condiciones el Fósforo del suelo.
También se han citado una serie de factores que limitan las poblaciones de estas bac-terias en el suelo. Sin embargo, no deben ser manejados estrictamente los datos que se han expuesto a la hora de aplicar en la práctica agrícola los microorganismos con características de biofertilizantes. Por ejemplo, en los suelos pobres en Azotobacterque fueron inoculados con estas bacterias se logró la colonización de la rizosfera de las plantas hospedantes y se produjo un efecto positivo en cereales y otros vegetales (Ar-shald y Frankerberger, 1993). De acuerdo a la experiencia cubana, es verdad que no se han podido aislar cepas de Azotobacter en suelos con pH 4.5, pero se han logrado
Tabla 23
Efecto antagonista de Azotobacter y Azotobacter y Azotobacter Azospirillum
frente a hongos fitopatógenos que atacan a las raíces de las plantas

105la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Tabla 24
Resultados obtenidos en la India con la inoculación de Azotobacter chroococcum en distintos cultivos
importantes resultados efectivos al realizar inoculaciones en suelos similares cultivados de tabaco (Martínez Viera et al., 1968).
Es decir, que el ambiente rizosférico crea determinadas condiciones que facilitan la vida de estas bacterias, las cuales se comportan como si estuvieran en condiciones norma-les. Pero al envejecer el cultivo o una vez recogida la cosecha, la bacteria no puede permanecer en el suelo y desaparece. Mientras tanto, cumplió con su función de bio-fertilizante y bioestimuladora y debe ser inoculada de nuevo con el próximo cultivo, con la seguridad de que se obtendrán beneficios similares.
Los países donde más se aplican estas bacterias en la práctica agrícola son India y Cuba. En la India existen 75 fábricas dedicadas a la producción de estos biofertilizantes. En Cuba se desarrolla, desde 1990, un programa de fabricación y aplicación de Azotobac-ter a base de cepas seleccionadas que son capaces de suministrar entre 30 y 50% de los requerimientos de Nitrógeno de las plantas, y se determinó la capacidad fijadora por parte de 15 cepas de A. chroococcum, cuyos valores oscilaron entre 12.2 y 25 nmol de C2H2/h. De todas las cepas se destacó la INIFAT-8, con 24.46 nmol C2H2 /h (Dibut et al., 2004).
En la tabla 24 se presentan algunos resultados obtenidos en diferentes evaluaciones en condiciones de producción en la India, utilizando como procedimiento de inoculación la introducción de las raíces en el momento del trasplante (en el caso de las hortalizas) o de las semillas (en el caso de los cereales) en un inoculante semi-líquido.
Cultivo Incremento Autoresdel rendimiento (%)
Cebolla Entre 18 y 22 Joi y Shinde, 1976Berenjena Entre 20 y 24 Mehrotra y Lehri, 1971Tomate Entre 15 y 29 Lehri y Mehrotra, 1972Col Entre 25 y 50 Lehri y Mehrotra, 1972Arroz Entre 15 y 22 Mehrotra y Lehri, 1971Trigo Entre 15 y 30 Sundara Rao et al.,1973Trigo Entre 10 y 20 Zambre et al., 1984Maíz Entre 15 y 30 Mishram y Shende,1982Maíz Entre 20 y 35 Zambre et al., 1984

106 capítulo 6. la fijación asociativa de nitrógeno
Pueden observarse los importantes incrementos que se obtuvieron en todos los culti-vos evaluados, muy superiores a los que aparecen en la Tabla 22 como reportados en condiciones templadas, a pesar de que estos últimos fueron logrados en condiciones experimentales, en las que se ajustan todos los factores que pueden influir en los resul-tados. Se pone así de manifiesto la superior respuesta a la inoculación que se logra en los países tropicales en comparación con los países templados, debido a las diferencias de condiciones agroecológicas que se ha comentado en varias oportunidades en el transcurso de este libro.
En lo que se refiere a las hortalizas, el efecto de la inoculación con Azotobacter en Cuba Azotobacter en Cuba Azotobacterpuede resumirse con el ejemplo del tomate (Martínez Viera y Dibut,1996):
1. En los semilleros se obtiene un aumento en la población de plántulas entre 30-40% más, lo que permite que se logre un mayor número de plantas viables por Kg de semillas, con la posibilidad de reducir la superficie necesaria para producir la cantidad de plántulas que deben ser trasplantadas posteriormente.
2. Las sustancias activas producidas por las bacterias aceleran el desarrollo de las plan-tas en el semillero: es mayor la altura en comparación con las plantas no tratadas (30% como promedio),el número de hojas (20%), el diámetro del tallo (40%) y la masa seca de las plantas completas (52%); estos resultados indican la posibilidad de acortar el período que transcurre entre la siembra del semillero y el momento en que las plantas están listas para ser transplantadas , ya que el desarrollo es más rápido y puede transplantarse entre 7 y10 días antes, con el consiguiente ahorro de agua, combustible, plaguicidas y mano de obra, al mismo tiempo que se reduce el ciclo total del cultivo.
3. El número de flores por planta es mayor en los campos tratados.
4. La fructificación de las plantas que fueron tratadas ocurre más temprano y el nú-mero de frutos por planta es 35% superior en la época normal y 60% fuera de época.
5. Se obtiene un incremento promedio de rendimiento de 25% en época normal y 40% fuera de época, lo que permite rendimientos altamente rentables en las siembras de períodos anormales.

107la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Figura 8
Efecto de la aplicación de A. chroococcum en semilleros de pimiento
Izquierda: inoculadaDere cha: sin inocular
Figura 7
Efecto de la aplicación de A. chroococcum en semilleros de tomate
Izquierda: inoculadaDerecha: sin inocular
6. La calidad de los frutos es superior, con pesos y diámetros promedios que son significativamente más altos en las variantes tratadas. En lo que respecta al valor comercial, en la época normal son de primera calidad entre 80 y 85% de los frutos en comparación con 60-70% en las siembras no tratadas.
7. Estos resultados se obtienen eliminando el 30% del fertilizante nitrogenado.
En las figuras 7 y 8 se presenta el efecto de la aplicación de esta bacteria fijadora de nitrógeno y estimuladora del crecimiento sobre las plantas de tomate y pimiento en semillero. Puede observarse la diferencia en el vigor de las plantas tratadas en compa-ración con las que no recibieron biofertilizante.

108 capítulo 6. la fijación asociativa de nitrógeno
En las figuras 9 y 10 se presenta el efecto de la aplicación de A. chroococcum sobre el desarrollo de los frutos de tomate y pepino en condiciones de plantación. Puede observarse la más rápida maduración y el mayor número y diámetro de los frutos de tomate en la variante tratada. En el caso del pepino, fue más alto el rendimiento por el mayor número y peso de los frutos inoculados.
En la figura 11 se presenta el efecto de la aplicación de A chroococcum en semilleros de cebolla. En ambos racimos hay igual número de plantas y puede verse el mayor desa-rrollo y el vigor de las plantas inoculadas. Se observa que cuando las plantas inoculadas se van a trasplantar ya tienen los bulbos desarrollados, lo cual incide en el acortamiento del ciclo del cultivo.
Figura 9
Efecto de la aplicación de Azotobacter sobre
el desarrollo de los frutos de tomate
Izquierda: Planta inoculadaDerecha: Planta sin inocular
Figura 10
Efecto de la aplicación de A. chroococcum sobre los
frutos de pepino cv. Ashley
Izquierda: Plantas inoculadasDerecha: Plantas sin inocular

109la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Figura 11
Efecto de la aplicación de A. chroococcum sobre semilleros de cebolla
Izquierda: plantas sin inocular Derecha: plantas inoculadas
Tabla 25
Efecto de la aplicación de A.chroococcum sobre el rendimiento de yuca y batata(Tm/ha).
En el caso de la yuca y la batata se aprovecha, además de la actividad fijadora de Ni-trógeno, la capacidad que tienen las sustancias activas sintetizadas por las bacterias de estimular la fotosíntesis (acumulación de compuestos) y reducir la respiración (gasto de compuestos) de las plantas, lo que permite el almacenamiento de fotosintatos, que constituye la base de la formación de tubérculos y raíces, constituidas por material de reserva (Martínez Viera et al., 2004). En la tabla 25 se ofrecen los rendimientos obteni-dos con la aplicación de biopreparados a base de A. chroococcum sobre dos clones de yuca (Manihot sculentayuca (Manihot sculentayuca ( ) y dos de batata (Ipomoea batatas) (Martínez Viera, 1998).
Como se observa, la aplicación no solo compensa el 50% del Nitrógeno necesario para estos cultivos, sino que además incrementa los rendimientos por la acción de las sustancias activas sintetizadas por las bacterias.
Variante Clones de yuca Clones de batata CMC-90 Señorita Señorita CEMSA 78354
100 % N 31.5 32.9 25.0 30.050 % N 20.0 20.6 18.7 20.650 % N + 32.6 35.4 30.6 34.3Azotobacter

110 capítulo 6. la fijación asociativa de nitrógeno
Las bacterias del género Azospirillum
Estas bacterias fueron descritas por Beijerinck en 1925 con el nombre de Spirillum lipoferum y cayeron en el olvido hasta que fueron redescubiertas por Döbereiner y Day en 1976. Los estudios taxonómicos permitieron su reclasificación en el nuevo gé-nero Azospirillum. Inicialmente se describieron las especies A. lipoferum y A. brasiliensey actualmente se consideran también dentro del género las especies A. amazonense, A. halopraeferans, A. irakense, A. largimobile y A. doebereinerae (Caballero-Mellado, 2003). El aislamiento de estas bacterias puede hacerse a partir del suelo rizosférico, de la superficie de las raíces (rizoplano) o del interior de las raíces y tallos de numerosas plantas hospederas.
Azospirillum coloniza la superficie y el interior de la raíz. En algunas gramíneas, como el millo, se han encontrado estas bacterias en la epidermis de la raíz, cubierta con una capa mucilaginosa; en otras, como trigo, sorgo, maíz y diferentes pastos proliferan en la zona de alargamiento de las raíces, principalmente en áreas de contacto longitudinal entre las células epidérmicas y la base de los pelos radicales, embebidas en capas mu-cilaginosas (Ramirez y Luna, 1995). Mediante técnicas inmunológicas, Levanony et al. (1998) encontraron que A. brasilense inoculado en trigo se asocia internamente con células corticales de las raíces, y localizaron a la bacteria en los espacios intercelulares de las células corticales en las zonas de alargamiento y de formación de pelos radicales, pero no en la endodermis y el sistema vascular.
Desde un punto de vista evolucionista, se ha sugerido que estas bacterias son organis-mos intermedios entre los fijadores libres y los simbióticos; la localización de los puntos de fijación de Nitrógeno dentro de las células de las plantas hospedantes ha apoyado esta hipótesis, porque no se admite hoy que la fijación simbiótica tenga que estar con-finada dentro de estructuras visibles como los nódulos cuando existe la posibilidad de otras formas de asociación, tales como el establecimiento dentro del tejido cortical de las raíces. La fijación de O2 por estas bacterias ocurre en condiciones microaerofílicas y pueden cumplir esta función en condiciones extremas, entre 0.05 y 0.1 atmósferas de O2 (Moens, 1996; Dommelen, 1998).
Los requerimientos de temperatura para Azospirillum son altos y su mayor crecimiento ocurre entre 32 y 36º C. La fijación máxima de Nitrógeno se logra entre 33 y 40º C y disminuye de forma pronunciada por debajo de 33º C. La incidencia de la bacteria en el suelo está controlada en alto grado por el pH, con un punto óptimo entre 6.8 y

111la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Tabla 26
Distribución geográfica de Azospirillum en raíces y suelos colectados en varias regiones del mundo.
7.8; por debajo de pH 5 no es posible lograr su aislamiento. Factores abióticos tales como alto contenido de arcilla y de materia orgánica y buena capacidad de retención de agua afectan positivamente la presencia de Azospirillum en el suelo, mientras que alto contenido de partículas arenosas y elevada concentración de Carbonato de Calcio afectan negativamente su supervivencia.
A pesar de la influencia de estos factores, es válido para estas bacterias lo que se ex-presó anteriormente en el caso de Azotobacter acerca de la inoculación en suelos con Azotobacter acerca de la inoculación en suelos con Azotobacterdificultades, ya que el ambiente rizosférico tiene determinadas condiciones que favo-recen el desarrollo de Azospirillum independientemente de las características del suelo, aunque el envejecimiento o la cosecha del cultivo causará la desaparición de estas bacterias después de que aportaron los beneficios que se esperaban.
Los azúcares no constituyen sustratos apropiados y la actividad máxima de fijación se obtiene con malato, succinato, lactato y piruvato. Las bacterias del género Azospirillumproducen bacteriocinas y sideróforos (agentes quelantes del hierro), los que tienen un efecto antagonista sobre agentes fitopatógenos de las raíces. En la Tabla 23 pudieron verse los resultados de esta actividad frente a un grupo de hongos fitopatógenos.
En la Tabla 26 se presenta la distribución geográfica de Azospiriillum (Sarro da Silva y Döbereiner, 1977).
Puede verse la abundancia de muestras positivas en las regiones tropicales en compa-ración con los bajos resultados obtenidos en las regiones templadas, lo que indica que los mayores beneficios de la aplicación de estas bacterias al suelo pueden ser logrados
Origen Raíces de pastos Suelode las Número % Positivas Número % Positivasmuestras de muestras de muestras
Europa - - 48 8EE.UU. 62 11 6 17África 58 45 79 53Colombia 54 43 - -Brasil Tropical 926 61 192 62Brasil Subtrop. 226 22 - -

112 capítulo 6. la fijación asociativa de nitrógeno
en el trópico. De este estudio se pudo deducir que los factores que más limitan la pre-sencia de Azospirillum son el pH y el tipo de vegetación; así, los suelos de bosques tro-picales y de sabanas muestran las bacterias esporádicamente pero, cuando la tierra es cultivada, aumenta su presencia de manera considerable; también pueden encontrarse en suelos semi-desérticos (Bashan y Holguin, 1997). Muestras con pH bajo fueron positivas cuando el contenido de Aluminio era también bajo.
Se ha demostrado que existe una gran variabilidad genotípica entre cultivares de trigo, maíz, y otros cereales en cuanto a lograr una eficiente fijación de Nitrógeno en asocia-ción con Azospirillum y también se ha reportado la presencia de quimiotaxismo, lo que es causa de que plantas C4, como maíz, se asocien principalmente con A. lipoferum, mientras que las plantas C3 se asocian con A. brasilense (Döbereiner, 1983). Estos datos deben ser manejados cuidadosamente por los investigadores y fabricantes de inoculantes para seleccionar las cepas adecuadas con el fin de inocular diferentes espe-cies y variedades de gramíneas.
Desde los hallazgos iniciales sobre Azospirillum se creó una expectativa en torno a los resultados prácticos de la introducción de estas bacterias en la agricultura. En compara-ciones de inoculación de distintas bacterias fijadoras (Azotobacter, Beijerinckia y Azospi-ciones de inoculación de distintas bacterias fijadoras (Azotobacter, Beijerinckia y Azospi-ciones de inoculación de distintas bacterias fijadoras (rillum) sobre maíz, los mejores resultados se obtuvieron con Azospirillum (Jaime et al., 1999; Zanier et al., 2001). La inoculación de la caña de azúcar tratada con bajos niveles de fertilizante nitrógenado produjo un aumento en el rendimiento, igual que ocurrió con sorgo y maíz (Macalintal y Urgel, 1992). Stolfus et al. (1997) informaron que la fijación en suelos inundados aporta hasta 50 kg/ha/ciclo de cultivo.
Los resultados positivos obtenidos en Israel en aplicaciones de campos de trigo han conducido a establecer el uso de Azospirillum en aquel país como una práctica agronó-mica (Okon et al., 1993), aunque aún debe verificarse más profundamente el éxito de las aplicaciones. La causa de que los resultados positivos sean inconsistentes en algunos lugares se atribuye a que no se conoce cual es el número necesario de bacterias ni la cantidad de sustratos carbonados que deben existir para que se logre un aporte signifi-cativamente adecuado a las necesidades de Nitrógeno de las plantas de trigo (Kennedy y Tchan, 1992).
En la Tabla 27 se muestran algunos resultados obtenidos por diferentes autores con la inoculación de dos especies de Azospirillum en distintos países, inoculando las semillas

113la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Tabla 27
Resultados obtenidos en distintos países con la inoculación de Azospirillum brasiliense y A. lipoferum en diferentes especies de gramíneas
mediante su mezcla con producto en polvo que contenía las bacterias (Martínez Viera, 1986).
En México se llevó a cabo en 1999 la inoculación de 450 000 ha de maíz y 150 000 ha de sorgo, cebada y trigo, empleando cepas seleccionadas de Azospirillum. La evalua-ción del rendimiento en grano se efectuó sobre 675 ha en 170 lugares de México. El éxito de la inoculación estuvo en el rango entre 62 y 95%, de acuerdo con el cultivo, y los incrementos del rendimiento fluctuaron entre 6 y 98%, de acuerdo con la región evaluada. El incremento promedio de la producción en todos los lugares evaluados fue de 26%. Las variaciones en los resultados dependieron de la variedad y cultivo, tipo de suelo, uso y nivel de fertilizantes (Caballero-Mellado, 2003).
La bacteria endófita Gluconacetobacter diazotrophicus
Los microorganismos endófitos comprenden a los hongos y bacterias que viven sin causar daño en el interior de células y tejidos de plantas superiores durante una parte considerable de su ciclo de vida, y pueden localizarse en espacios intracelulares, inter-celulares o en el tejido vascular. Existe una extensa diversidad de endófitos, y los que se han aislado con más frecuencia pertenecen a los géneros Pseudomonas, Enterobacter, Bacillus, Erwinia y Xantomonas (Muñoz-Rojas y Caballero-Mellado, 2003).
Cultivo País Bacteria Incremento del rendimiento (%)rendimiento (%)
Maízízí Brasil A. lipoferum 25Maízízí Israel A. brasilense 34Sorgo Egipto A. lipoferum Entre 60 y 100Sorgo India A. brasilense Entre 50 y 80Trigo India A. brasilense Entre 15 y 26Trigo India A. lipoferum Entre 40 y 60Trigo Israel A. brasilense Entre 10 y 25Trigo Bélgica A. brasilense 28
TrigoTrigo Alemania A. lipoferumA. lipoferum Entre 35 y 60Entre 35 y 60

114 capítulo 6. la fijación asociativa de nitrógeno
La más importante de las bacterias endófitas fijadoras de Nitrógeno atmosférico, por su gran importancia práctica, es la que hoy se conoce como Gluconacetobacter diazotro-phicus, que fue descubierta por Cavalcante y Döbereiner (1988) en raíces y tallos de caña de azúcar cultivada en Brasil y bautizada como Saccharobacter nitrocaptans, nom-bre que cambió al poco tiempo por Acetobacter nitrocaptans y, pocos meses después, por Acetobacter diazotrophicus (Gillis et al., 1989). Yamada et al. (1997) propusieron, y así fue aceptado, el nombre actual de Gluconacetobacter diazotrophicus. El género Glu-conacetobacter comprende además a las especies conacetobacter comprende además a las especies conacetobacter G.liquefaciens, G. xylinus, G. hansenii, G. europeaus, G. obodiens, G. pomorum, G. sacchari, G. johannae y G. azotocampans(Fuentes et al., 2001; Muñoz-Rojas y Caballero-Mellado, 2003).
G. diazotrophicus produce sustancias estimuladoras del crecimiento vegetal y es capaz de excretar más del 50% del Nitrógeno fijado en una forma asimilable por la planta (Cojho et al., 1993). Se ha calculado que es capaz de entregar entre 11 y 270 kg de Nitrógeno/ha a la caña de azúcar (Döbereiner, 1993), y las cualidades excepcionales de esta bacteria se ponen además de manifiesto porque su pH óptimo es 5.5 y puede vivir en concentraciones de sacarosa tan altas como 30%. Se encuentra en los vasos xilemáticos del tallo de plantas maduras y en el espacio apoplástico del tallo. Las po-blaciones de las bacterias diazotrofas endofíticas se ven disminuídas cuando se aplica una alta fertilización nitrogenada, lo que ha sido demostrado en México, India y Brasil (Fuentes-Ramirez et al.,1999; Muthukumarasamy et al.,1999; Reis et al., 2000).
Los aislamientos de la bacteria se han realizado en su mayoría a partir de especies ve-getales que se reproducen de forma asexual y se ha propuesto que su dispersión geo-gráfica a gran distancia se lleva a cabo durante la reproducción del material vegetativo que alberga a la bacteria de forma natural; por ejemplo, se ha aislado la bacteria a partir de esquejes de caña de azúcar cultivados en Cuba, que fueron transportados a Canadá y propagados bajo condiciones de invernadero (Muñoz-Rojas y Caballero-Mellado, 2003). También ha sido aislada de insectos de la caña de azúcar, como las cochinillas, los cuales pueden actuar como agentes de transmisión de la bacteria (Franke et al., 1999). En la Figura 12 se muestra el crecimiento de G. diazothrophicus en forma de colonias amarillas sobre el medio cultivo LGI-P.
Las especies vegetales de las que ha sido aislada esta bacteria endofítica han sido las siguientes: en primer lugar, la caña de azúcar, donde se descubrió su existencia; pos-teriormente fue aislada de batata (Ipomoea batatas) (Paula et al., 1993) y de hierba

115la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Figura 12
Colonias de G. diazothrophicus en medio LGI-P
elefante (Pennisetum purpureum) (Reis et al., 1994). Mas recientemente fue aislada de cafeto (Coffea arabica) (Jiménez-Salgado et al., 1997), así como de plantas adultas e hijuelos de piña (Ananas comosushijuelos de piña (Ananas comosushijuelos de piña ( ) (Tapia-Hernández et al., 2000), de la gramínea Eleu-sine coracana y de maíz (Zea mays), melón (Cucumis melo) y remolacha (Beta vulgaris) (González y Barraquio, 2000). En los últimos tiempos, se ha reportado el aislamiento en Cuba a partir de auyama (Cucúrbita pepo), ocumo (Xanthosoma sp), ocumo (Xanthosoma sp), ocumo ( papaya (Carica papaya), yuca (Manihot sculenta) y rosa (Rosa montezumae) (Dibut et al., 2005). Como se observa, con excepción de cafeto y rosa, la tendencia de la bacteria es habitar dentro de especies vegetales de alto contenido en azúcares.
La distribución de las poblaciones en los diferentes órganos de las plantas es variable de acuerdo con la especie vegetal; así, en caña de azúcar, en Brasil, los estudios arro-jaron un título entre 103 y 107 UFC/g de tejido fresco y en maíz en Estados Unidos se encontraron poblaciones de 105 UFC/g de tejido fresco (Döbereiner et al., 1993). En Cuba se reportaron poblaciones entre 103-105 en maíz, batata y ocumo, entre 102-104
en yuca y entre 104-107 en caña de azúcar, dependiendo las variaciones del órgano vegetal. En papa no se ha conseguido aislar la bacteria en condiciones naturales, pero ésta se encuentra en concentraciones elevadas (106 UFC/ g de tejido fresco) después de la inoculación y produce un marcado efecto sobre el cultivo.
Las perspectivas que presentan las bacterias diazotróficas endofíticas son muy grandes, aunque es necesario todavía hacer muchas investigaciones para lograr el mayor pro-vecho con su utilización. Estas bacterias no se han encontrado en la zona rizosférica y muy raramente sobreviven en el suelo. No infectan fácilmente plantas intactas de caña o de otros cultivos, por lo que se están buscando métodos que faciliten la penetración de la bacteria. Hoy existe la posibilidad de introducir estos organismos diazotróficos en

116 capítulo 6. la fijación asociativa de nitrógeno
las raíces de las plantas por la vía de las esporas de micorrizas vesículo-arbusculares, lo que abre un campo totalmente nuevo para la Biotecnología.
En caña de azúcar, batata y sorgo dulce se han logrado infecciones exitosas con G. dia-zotrophicus mediante la coinoculación con el hongo micorrízico Glomus clarum (Coc-king, 2003). En el caso de la caña de azúcar, sobre todo en Brasil, se ha considerado prematuro utilizar inoculantes a base de G. diazotrophicus, porque las bacterias parece que están bien establecidas en el cultivo y se considera necesario obtener mayores conocimientos que permitan lograr la mayor efectividad con las inoculaciones con otras cepas.
Las especiales cualidades de G. diazotrophicus permitieron decir a la Profesora Johanna Döbereiner (1993): “Todas estas observaciones cambian completamente el concepto de asociación planta-diazotrofo, la cual se pensaba que era una asociación solamente casual de las raíces de las plantas, y abre enteramente nuevas vías para expandir la fijación biológica del nitrógeno como una sustitución a los fertilizantes nitrogenados por las gramí-neas”.
La fijación de N2 en la filosfera
En el medio tropical ocurre también fijación de nitrógeno en la filosfera, zona que está en contacto con la hoja y la atmósfera, sometida a la actividad reguladora de ambas. Los microorganismos que viven en las hojas, entre los cuales se encuentran las bacterias del género Azotobacter, toman el agua y los gases disueltos de la atmósfera y los nutrientes a partir de los exudados de las hojas vivas, los cuales están regulados por el estado nutricional de las plantas. El follaje funciona como soporte, trampa de agua y centro de producción de nutrientes y condiciona el medio para el crecimiento microbiano; además, procesa y distribuye compuestos nitrogenados en flujo a través de la planta hacia las partes más jóvenes. Esta propiedad, y la habilidad para concentrar materia re-suspendida o disuelta en la atmósfera con gran rapidez, hace que las hojas tengan una gran importancia en los ecosistemas agrícolas (Bhat et al., 1971).
La abundante población microbiana que se encuentra en las hojas es, por sí misma, prueba de un ambiente que posee considerable valor nutritivo; la humedad contribuye también al desarrollo y supervivencia de ésta población, ofreciéndole espacio y estimu-lando el intercambio de productos metabólicos. El importante papel que desempeña

117la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
la filosfera en la economía del Nitrógeno de un ecosistema natural fue sugerido inicial-mente en bosques semidecíduos de las regiones tropicales, basado en el conocimiento de que, en ellos, la biomasa alcanza valores de 500 a 1700 Kg/Ha, muy superiores a los encontrados en los bosques subtropicales y templados, en los que no excede de 400 Kg/Ha; además, la fracción verde asimiladora es 10 a 20 veces superior en el trópico a la de la zona templada y 4 veces la de los bosques subtropicales (Debinstein, 1970).
Ruinen (1974) reportó desde 1954 la presencia permanente de las bacterias fijadoras de nitrógeno sobre la filosfera de distintas plantas de Java y Sumatra, donde aisló estas bacterias en 192 y 198 muestras de hojas de árboles, arbustos, enredaderas, epífitas y vegetación costera. Recuentos de bacterias pertenecientes al género Azotobactersobre hojas maduras de cacao, en Surinam, dieron un promedio de 1.3 x107/cm2. En Egipto se estudiaron 58 muestras de árboles, cultivos de campo, vegetales, plantas ornamentales, silvestres y acuáticas y solamente 2 de las muestras no contenían Azo-tobacter (Abd-El-Malek,1971). En la India se encontró una fijación de 10 mg N/g de manitol oxidado sobre hojas de jacinto de agua (Eichhornia crassipes), con un abundan-te crecimiento de A. chroococcum, incluso en el tejido interior de las hojas (Iswaran et al., 1973). Se ha demostrado la presencia de la enzima nitrogenasa en el interior de las hojas de caña en Brasil (Olivares et al., 1997), lo cual constituye una confirmación irrebatible de que ocurre la fijación en este medio.
Al determinar los métodos de aplicación de biofertilizantes a base de A. chroococcumen Cuba se aprovecharon los conocimientos existentes acerca del papel de la filosfera, y se comprobó que las aplicaciones foliares en las extensas plantaciones de cítricos del país, utilizando el avión, eran de gran efectividad, como se demuestra en la tabla 28 (Martínez Viera y Dibut, 2006).
Puede verse en la tabla que, por el método de aplicación aérea, se consigue no solo la sustitución del fertilizante, sino que permite una alta expresión de las sustancias activas sintetizadas por las bacterias, que estimulan la floración y la fructificación, lo que se traduce en rendimientos más altos. En este sentido, la inoculación actúa en sustitución de productos comerciales a base de aminoácidos, hormonas vegetales y otras sustan-cias que se ofrecen para estos mismos fines y tienen un precio elevado. En la Tabla 29 puede verse también el efecto de las aplicaciones foliares sobre el mango en el Estado Sinaloa (México) (Martínez Viera et al., 2003).
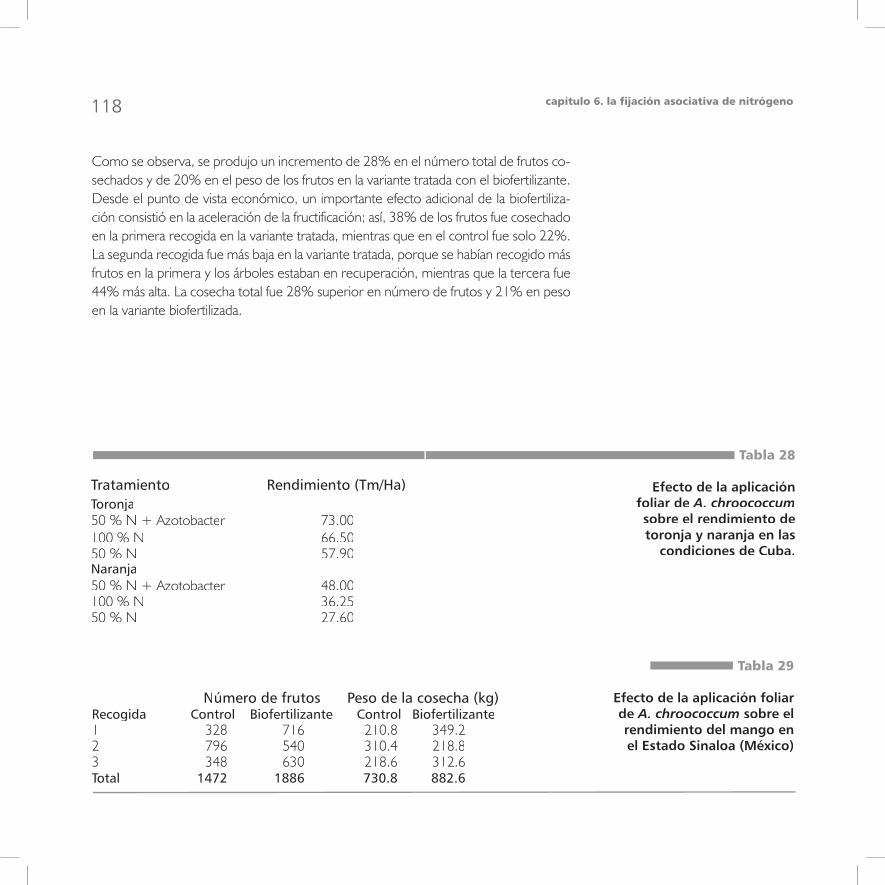
118 capítulo 6. la fijación asociativa de nitrógeno
Como se observa, se produjo un incremento de 28% en el número total de frutos co-sechados y de 20% en el peso de los frutos en la variante tratada con el biofertilizante. Desde el punto de vista económico, un importante efecto adicional de la biofertiliza-ción consistió en la aceleración de la fructificación; así, 38% de los frutos fue cosechado en la primera recogida en la variante tratada, mientras que en el control fue solo 22%. La segunda recogida fue más baja en la variante tratada, porque se habían recogido más frutos en la primera y los árboles estaban en recuperación, mientras que la tercera fue 44% más alta. La cosecha total fue 28% superior en número de frutos y 21% en peso en la variante biofertilizada.
Tratamiento Rendimiento (Tm/Ha)Toronja50 % N + Azotobacter 73.00100 % N 66.5050 % N 57.90Naranja50 % N + Azotobacter 48.00100 % N 36.2550 % N 27.60
Número de frutos Peso de la cosecha (kg)Recogida Control Biofertilizante Control Biofertilizante1 328 716 210.8 349.22 796 540 310.4 218.83 348 630 218.6 312.6Total 1472 1886 730.8 882.6
Tabla 28
Efecto de la aplicación foliar de A. chroococcumsobre el rendimiento de toronja y naranja en las
condiciones de Cuba.
Tabla 29
Efecto de la aplicación foliar de A. chroococcum sobre el rendimiento del mango en el Estado Sinaloa (México)

119la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Capítulo 7Preparación y aplicación de inoculantesa base de bacterias fijadorasde nitrógeno atmosférico
La práctica agrícola de inocular las plantas con bacterias beneficiosas es muy antigua. En el último tercio del siglo XIX ya era un método muy extendido en los Estados Unidos el mezclar las semillas de leguminosas con suelo inoculado naturalmente. En 1896, Nob-be y Hiltner patentaron por primera vez un producto para inocular las leguminosas con Rhizobium, al que dieron el nombre de “Nitragin”. Los inoculantes que se utilizaban en aquella lejana época para aplicar al suelo las bacterias fijadoras de Nitrógeno atmosféri-co fueron cultivos puros de las bacterias propagados en medios sólidos a base de agar y envasados en botellas de cristal con un volumen entre 250 y 300 mL.
Después de una larga evolución encaminada a satisfacer las demandas del mercado, hoy se presentan estos biopreparados en forma líquida o como materiales sólidos ab-sorbentes de distintos tipos, llamados portadores, que son impregnados con cultivos líquidos de la bacteria que se quiere inocular. Una vez que se demostró la efectividad de estas nuevas tecnologías, durante muchas décadas se han estado introduciendo bacterias en el suelo para mejorar el desarrollo de los cultivos de mayor importancia económica, mediante la inoculación directa del suelo o la mezcla de preparados bacte-rianos en polvo con semillas, raíces o tubérculos.
El efecto de la inoculación con bacterias fijadoras de Nitrógeno atmosférico de forma simbiótica o asociativa, que al mismo tiempo son estimuladoras del crecimiento, puede variar en dependencia de la cepa bacteriana, de la especie vegetal, del tipo de suelo, de la densidad del inoculante y de las condiciones ambientales. En general, la población de un diazotrofo que se ha inoculado al suelo comienza a disminuir a los pocos días después de la inoculación, debido a que tiene que competir con una extensa gama de microorganismos rizosféricos bien adaptados a las condiciones de todos los facto-res ambientales del lugar. Por otra parte, al envejecer las plantas se empobrecen las secreciones de las raíces y se hace más difícil la adquisición del alimento, por lo cual disminuyen las poblaciones.

120 capítulo 7. preparación y aplicación de inoculantes a base de bacterias fijadoras de nitrógeno atmosférico
Por las razones expuestas, la formulación del inóculo debe suministrar a los microorga-nismos que se quieren inocular las condiciones necesarias para sobrevivir hasta que se adaptan a las condiciones del ambiente rizosférico y se hacen competitivas durante el tiempo necesario para que puedan aportar a las plantas los beneficios esperados.
De acuerdo con todo lo expuesto, la producción de inóculos comerciales es el paso por el cual las cepas seleccionadas de una determinada especie microbiana son multi-plicadas en masa y comercializadas en formulaciones que permiten su aplicación a los cultivos de importancia económica.
Método de selección de cepas
El primer objetivo cuando se quiere preparar un inóculo es utilizar la cepa bacteriana más activa y efectiva. La optimización de una formulación inoculante es independiente de la cepa utilizada porque, en general, la mayoría de las cepas de una misma especie microbiana tienen características fisiológicas comunes. Esto generalmente no es aplica-ble cuando se trata de diferentes géneros bacterianos (Bashan, 1998).
Tomando en cuenta estos conceptos, se han elaborado esquemas para seleccionar cepas de microorganismos con mayor eficiencia y efectividad como biofertilizantes y bioestimuladores para cada una de las especies vegetales, incluyéndose en ellos los aspectos siguientes:
– Estudios quimiotáxicos utilizando secreciones de las raíces de diferentes especies vegetales, con el fin de seleccionar las cepas que más pueden ser atraídas por las secreciones radicales de cada una de las especies.
– A las cepas que han sido seleccionadas en las pruebas de quimiotaxismo se les hace la prueba de reducción del acetileno (ARA) y de solubilización del fósforo, para seleccionar las que tienen mayor capacidad de fijación de nitrógeno atmosférico o de solubilización del fósforo del suelo.
– A las que presentan valores más altos para la fijación y la solubilización se les hacen pruebas en bandejas en las que se han sembrado semillas de la especie vegetal que se desea probar, para seleccionar las que presentan mayor actividad estimuladora del crecimiento vegetal.

121la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Figura 13
Efecto de la inoculación de distintas cepas de bacterias fijadoras de N sobre el desarrollo de plantulas de tomate
– Terminada la selección, las cepas se cultivan in vitro en medios que contienen los plaguicidas que más se usan en la región, incluyendo los fungicidas para tratar las semillas, con el fin de conocer si alguno de ellos puede hacer fracasar la inocula-ción.
– Las cepas seleccionadas son sometidas a los estudios taxonómicos correspondien-tes para su correcta identificación y se les realizan las pruebas toxicológicas para demostrar su inocuidad.
En la figura 13 se muestran algunos de los resultados que pueden obtenerse con la inoculación de semillas de tomate con distintas cepas, con el fin de seleccionar las más efectivas.
Características de los portadores
Los portadores son los vehículos que sirven para transportar los microorganismos vi-vos desde la fábrica hasta el campo. Un buen portador debe tener la capacidad de

122 capítulo 7. preparación y aplicación de inoculantes a base de bacterias fijadoras de nitrógeno atmosférico
mantener una población elevada de células viables de la cepa bacteriana que ha sido seleccionada, en buenas condiciones fisiológicas, durante el mayor tiempo posible. Se-gún Bashan (1998), el portador debe ser un material asequible y disponible, fácilmente esterilizable en el autoclave o mediante rayos gamma, uniforme desde los puntos de vista físico y químico, no tóxico, biodegradable, con alta capacidad de hidratación y utilizable por el mayor número posible de especies y cepas bacterianas.
También tiene que ser fácilmente manufacturado y mezclado, manteniendo sus ca-racterísticas durante un tiempo largo de almacenamiento, al mismo tiempo que debe tener propiedades adhesivas para que pueda recubrir las semillas y permitir una rápida liberación de las bacterias en el suelo.
Hay muchos materiales que pueden ser utilizados como portadores, y entre ellos se encuentran turba, arcilla, suelo inorgánico, compost, abono de establo, humus de lom-briz, harina de soya, aceite de soya o maní, cachaza y otros residuos agrícolas, materia-les inertes como vermiculita, perlita, zeolita, geles de poliacrilamida, alginato y otros.
En los últimos años se ha utilizado el método de encapsular las bacterias con portadores sintéticos con el objetivo de incrementar la supervivencia durante el almacenamiento. Al bajar el contenido de agua, las bacterias permanecen inactivas, resistentes a las con-diciones ambientales e insensibles a la contaminación (Paau, 1988). Los inoculantes que han sido sometidos al proceso de deshidratación tienen un período muy largo de viabi-lidad, aunque se ha demostrado que una importante proporción de células bacterianas son destruidas durante el proceso (Paul et al., 1993); sin embargo, las supervivientes son suficientes para que pueda hacerse una inoculación efectiva.
Para preparar el inoculante se introduce en el portador una cantidad determinada de cultivo líquido del microorganismo que se quiere aplicar al suelo, procurando que la humedad del sustrato no sea mayor de 60%. El portador se puede utilizar esterilizado previamente o sin esterilizar; el primero es más caro debido al costo del proceso de esterilización, pero presenta numerosas ventajas que se ponen de manifiesto en la Tabla 30, según Bashan (1998).
Se ha demostrado en muchos países que, en el caso de los inoculantes a base de Rhi-zobium, las poblaciones contaminantes superan varias veces a las de bacterias benéficas cuando se utilizan portadores sin esterilizar. Los altos niveles de contaminantes hacen

123la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Tabla 30
Comparación de inoculantes a base de turba estéril y no estéril
que las bacterias que se desea inocular pierdan su efectividad, pero a la vez representan un peligro potencial para la salud del hombre, los animales y las plantas.
Control de calidad de los biopreparados
El éxito de la aplicación de los biofertilizantes depende de la calidad del inoculante uti-lizado, y es necesario que contenga un número adecuado de células viables efectivas, con el fin de asegurar la infección de las plantas. Este requisito es fundamental, y debe
Parámetros del inoculante Estéril No estéril
Población de bacterias benéficas
Alta Variable
Longevidad Alta Relativamente baja
Adición de nutrientes para aumentar la población final
Posible sin sacrificar la calidad final del inoculante
Puede ocurrir desarrollo más rápido de contaminantes
Elección de materiales para ser usados como portadores
Muchos materiales no son esterilizables o cambian su composición con el proceso
Casi ilimitada
Requerimientos de trabajo Muchos y caros Pocos y baratos
Equipamiento requerido Grandes y costosos autoclaves
No necesita
Espacio necesario estéril Grande No necesita
Monitoreo de contaminación Esencial para el control de calidad
Esencial para el control de calidad
Costo total de producción Alto Mucho más bajo

124 capítulo 7. preparación y aplicación de inoculantes a base de bacterias fijadoras de nitrógeno atmosférico
organizarse un mecanismo adecuado para garantizar que no se engañe a los agriculto-res, ya que ellos no tienen medios para determinar la calidad del producto que están recibiendo.
Algunos países desarrollados, entre los que no se cuenta Estados Unidos, tienen regu-laciones oficiales para controlar la calidad de los inoculantes, pero en la mayor parte de los países latinoamericanos esto no ocurre. En algunos lugares como Canadá y Francia ocurrió un mejoramiento en la calidad de los inoculantes después de estas regulaciones (Brockwell et al., 1988), pero en gran parte de ellos no se regula el nivel de contami-nación ni la población de organismos benéficos que debe existir en el inóculo, a pesar de que en casi todos se utilizan portadores sin esterilizar.
En América del Sur solo Brasil y Uruguay y en América del Norte solo Canadá tienen autoridades regulatorias apoyadas por la legislación. En Cuba existe un Registro en el cual hay que inscribir los diversos bioproductos exponiendo las características que deben poseer y los resultados de las pruebas de campo realizadas por instituciones oficialmente autorizadas y se comienzan a implantar oficialmente las normas d e control de calidad, igual que ocurre en Venezuela. Australia, África del Sur, Nueva Zelandia e India también tienen importantes programas de control. Pero los más exigentes re-quisitos para el control de calidad de los inoculantes los ha puesto Francia, donde está prohibido el más mínimo nivel de contaminación y se exigen pruebas de campo antes de aceptar nuevas formulaciones (Cantroux, 1991). En Estados Unidos e Inglaterra también existen las normas, pero su cumplimiento se deja a discreción de los dueños de empresas.
En general, no existen normas internacionales para el control de calidad de los biofer-tilizantes y bioestimuladores microbianos, por lo que la calidad de los inoculantes varía de país en país, debido a que la falta de estos mecanismos de control no incentiva a los fabricantes a establecer sus propios programas de calidad, como ha sido reconocido por NiffTAL, el organismo más importante que asesora, coordina y divulga en el mun-do la producción y el uso de los inoculantes para las leguminosas.
Hace pocos años, en el año 2001, el Buró de Standards de la India (BIS), país donde hay unas 75 fábricas de biofertilizantes, publicó las especificaciones para los inoculantes a base de Rhizobium, Azospirillum, Azotobacter y solubilizadores de fósforo. En la Tabla 31 se exponen estas especificaciones (Kannaiyan, 2003).

125la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Tabla 31
Especificacionesde control de calidaddel Buró de Standardsde la India para los biofertilizantes y bioestimuladores
En estas especificaciones se ha sido muy conservador en algunos de los parámetros, lo cual atenta contra la calidad y la efectividad que debe tener el biopreparado. Por ejemplo, el número de células/g está muy bajo, de acuerdo con las normas que ha recomendado la FAO para Rhízobium, que no debe ser inferior a 108, y de acuerdo a la experiencia con Azotobacter o Azotobacter o Azotobacter Azospirillum, ya que las poblaciones señaladas en la tabla no permiten lograr la debida efectividad, no solo en la fijación de Nitrógeno sino también en la estimulación del crecimiento vegetal. Estas poblaciones no deben ser menores de 108-9 UFC/g de suelo y pueden lograrse con las tecnologías adecuadas.
Como se ha explicado en uno de los capítulos anteriores, Australia es el país que mejor maneja la inoculación de las leguminosas. Según las especificaciones de la Aus-tralian Inoculant Research and Control Service (AICRS), un buen inoculante a base de Rhizobium o Bradyrhizobium debe presentar las siguientes características (Subba Rao, 1996):
Parámetros Rhizobium Azotobacter AzospirillumAzospirillum Solubilizadores
Base Portador Portador Portador Portador
Células/g I07 106 I07 108
Contaminación 0 a dilución 10-5 0 a dilución 10-5 0 a dilución10-5 0 a dilución 10-7
Expiración 6 meses 6 meses 6 meses 6 meses
pH 6.5-7.5 6.5-7.5 6.5-7.5 6.5-7.5
Cepa Chequeada serológicamente
No específicaíficaí No específicaíficaí No específicaíficaí
Partícula del portador
150-212 micras 106 micras 106 micras 106 micras
Otros Nodulación positiva. Aumento de materia seca superior a 50%superior a 50%
Fijación de N mayor de 5 mg/g de sacarosa
Película blanca en medio NFB a dilución 10-7
Zona de solubilización mayor de 8 mm a dilución 10-5

126 capítulo 7. preparación y aplicación de inoculantes a base de bacterias fijadoras de nitrógeno atmosférico
1. Tener un solo portador.
2. Contener un mínimo de 108 células viables dentro de los 15 días después de su fabricación y 107 dentro de los15 días antes de la fecha de expiración cuando se almacena a 6-100C.
3. El período máximo de expiración no debe ser mayor de 6 meses desde la fecha de fabricación.
4. No debe tener contaminación con otros microorganismos.
5. El pH del inoculante debe estar entre 6.0 y 7.5.
6. Tiene que ocasionar buena nodulación antes de la fecha de expiración en todas las especies vegetales que aparezcan listadas en el paquete.
7. El total de masa seca de las plantas inoculadas debe ser más alto, de una forma significativa, en comparación con los controles no inoculados.
8. El material portador debe pasar por malla 75-106 micras.
Biofertilizantes mixtos utilizando bacterias fijadoras de N2
Hasta mediados de la década de los 90, en el mundo se habían realizado estudios relati-vamente escasos en relación con la inoculación simultánea de varios microorganismos; en su mayor parte, los resultados que se han mostrado no habían rebasado el nivel de macetas con suelos esterilizados, lo que aleja a estos estudios de la práctica agrícola. A pesar de eso, y tomando en cuenta las grandes posibilidades que mostraban las asocia-ciones, Subba Rao (1996) enfatizó que el efecto asociativo de otros microorganismos con fijadores de Nitrógeno, en el suelo y en las raíces debe ser estimulado mediante la fabricación de inoculantes mixtos basados en portadores. También Parr et al. (1994) y Higa (1995) plantearon que la mejor probabilidad de lograr importantes ventajas de la aplicación de microorganismos beneficiosos al suelo es la inoculación como parte de un cultivo mixto, con una densidad de inóculo suficientemente grande para permitir maximizar la posibilidad de su adaptación a las condiciones ecológicas del lugar.
A partir de 1995, las investigaciones sobre inoculantes mixtos se intensificaron, lográn-dose su aplicación con grandes beneficios en la producción agrícola. Los primeros es-tudios sobre la utilización de biofertilizantes mixtos fueron realizados con la asociación

127la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Tabla 32
Efecto de la aplicación de inoculante mixto a base de A. chroococcum y B. megatherium var. phosphaticum sobre rosales de seis años de edad
NOTA: PC-Periodo de corte.
de bacterias fijadoras de Nitrógeno y solubilizadoras del Fósforo del suelo. Así, Cantir (1962), tomando en cuenta la necesidad de Fósforo para lograr una fijación eficiente, estudió el efecto de la inoculación simultánea de Rhizobium, Azotobacter y Bacillus me-gatherium var. phosphaticum sobre el rendimiento de las leguminosas, reportando un incremento del rendimiento superior a los obtenidos con la inoculación independiente de cada uno de ellos.
Kunda y Gaur (1980, 1982, 1984) demostraron en la India importantes incrementos del rendimiento en trigo, papa, algodón y arroz con inoculaciones mixtas de Azotobac-ter chroococcum y diferentes bacterias solubilizadoras del Fósforo; estos incrementos fueron superiores en comparación con los logrados con la inoculación separada de es-tas bacterias. Alagwadi y Baur (1988) también lograron resultados altamente positivos con la inoculación conjunta de Rhizobium y B. megatherium, y B. megatherium, y en comparación con los que obtuvieron con inoculaciones separadas. En Cuba se han desarrollado productos a base de A. chroococcum y Pseudomonas. fluorescens y Rhizobium sp. y Ps. fluorescens (Bach et al., 2000), A. chroococcum y B. megatherium var. phosphaticum (Dibut et al., 2003) y A. chroococcum y el hongo Penicillium bilaii (García et al., 2004).
En la Tabla 32 pueden verse los resultados obtenidos en el cultivo de la rosa con la aplicación de un inoculante mixto a base de Azotobacter chroococcum y Bacillus mega-therium var. phosphaticum (Dibut et al., 2004).
Los resultados expresados en la tabla anterior se reflejan en las figuras 14 y 15, donde puede observarse la diferencia de densidad en los campos de rosas inoculados con el biofertilizante mixto y las diferencias entre el tamaño de los racimos de rosas obtenidos de plantas inoculadas y sin inocular.
Variante Numero de ramas primarias
Peso de los tallos g.(25 cm)
Numero de retoños
Numero de mazos /plantaPC.
Peso fresco botón (g)
Testigo 2.20 b 1.89 b 4.24 b 1.10 b 10.28 bInoculado 9.53 a 2.36 a 7.50 a 2.13 a 15.36 aEsx 0.48 0.50 2.51 1.00 0.42CV (%)CV (%) 31.9131.91 11.1011.10 16.7016.70 17.717.7 4.204.20

128 capítulo 7. preparación y aplicación de inoculantes a base de bacterias fijadoras de nitrógeno atmosférico
En lo que respecta a la asociación entre las bacterias fijadoras de Nitrógeno, se han puesto de manifiesto los beneficios obtenidos con la aplicación de inoculante mixto a base de Rhizobium y Azotobacter, lográndose incrementos de la eficiencia fijadora de Rhizobium en las leguminosas y el aumento de los rendimientos agrícolas por la acción de las sustancias activas sintetizadas por Azotobacter (Elmerich, 1984; Lynch, 1990; Azotobacter (Elmerich, 1984; Lynch, 1990; AzotobacterPeoples y Craswell, 1992). Igualmente, la asociación entre Bradyrhizobium japonicum y A. chroococcum en sustrato a base de turba ha demostrado también que incrementa la nodulación y el rendimiento de la soya (Katayama y col., 1996). Rodelas et al. (1999)
Figura 14
Efecto de la aplicación de inoculante mixto, a
base de A. chroococcumy B. megatherium var.
phosphaticum, sobre la densidad y la velocidad de floración de un campo de
rosas inoculado (derecha) en comparación con un campo
vecino sin inocular (izquierda)

129la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Figura 15
Efecto de la aplicación de inoculo mixto a base de A. chroococcum y B. megatherium var. phosphaticum sobre la producción de rosas
Izquierda: Plantas inoculadasDerecha: plantas sin inocular
encontraron un incremento del crecimiento de las plantas, de la nodulación y del con-tenido de Nitrógeno en haba (Vicia faba) con la inoculación mixta de Rhizobium legumi-nosarum bv. viceae y Azotobacter chroococcum, así como un aumento en el contenido total de Fósforo, Potasio, Calcio, Magnesio y microelementos en la parte aérea de las plantas. Resultados similares fueron obtenidos en frijol por Torres et al. (2001) con la inoculación mixta de R. leguminosarum bv phaseoli y A. chroococcum.
También la asociación de Azospirillum y Bradyrhizobium japonicum en soya ha demos-trado el incremento de la nodulación y el rendimiento (Singh y Subba Rao, 1979), así como la de Azospirillum y Rhizobium en alfalfa y garbanzo (Tilak et al., 1981) y en haba (Rodelas, 2001). La estimulación de la nodulación posterior a la inoculación con Azospirillum puede estar dada por un incremento en la inducción de raíces laterales, la densidad de los pelos radicales y las ramificaciones de dichos pelos; pero también por la diferenciación de un número mayor de células epidérmicas dentro del pelo radical infectado (Burdman et al., 2000).
Hasta 1999, más del 15% del total de 150 000 ha de leguminosas inoculadas con Rhizobium sp. en Africa del Sur habían sido coinoculadas con Azospirillum brasilense. Debido al éxito alcanzado con esta co-inoculación, en el periodo 1999-2000 este porcentaje se ha incrementado alrededor del 50% (Burdman et al., 2000). Según estos autores, se han demostrado incrementos en los rendimientos de 15-30 % en legumi-nosas coinoculadas, valores superiores a los obtenidos con la inoculación de Rhizobium

130 capítulo 7. preparación y aplicación de inoculantes a base de bacterias fijadoras de nitrógeno atmosférico
solo. Estos resultados coinciden con los citados por Okon y Vanderleyden (1997), los cuales, en investigaciones realizadas en relación con la inoculación conjunta de Rhizo-bium y Azospirillum, reportan un efecto positivo para diversos tipos de leguminosas, incluyendo una temprana nodulación, incrementos en el número de nódulos, altas cantidades de fijación de N2 y un mejoramiento en general del desarrollo del sistema radical. En Cuba se han desarrollado productos a base de Azotobacter chroococcum y Azospirillum brasiliense. En las tablas 33 y 34 se presentan algunos resultados obtenidos con la aplicación de estos inoculantes mixtos (Martínez Viera et al., 2001).
Un mayor número de estudios se han ejecutado para demostrar la efectividad de la asociación entre las micorrizas y Azotobacter, Rhizobium, Azospirillum y bacterias so-lubilizadoras del Fósforo. Los resultados han demostrado la gran efectividad de estas asociaciones en todos los casos ( Barea et al., 1977; Ocampo et al., 1975; Bagyaraj y Menge, 1978; Brown y Carr, 1984; Young, 1990; Rivera et al., 1997).
Rendimiento de campos testigo
(kg/ha) 5000-5900 6000-6900 7000-7900 8000-8900
Incremento por la aplicación del
inóóculo (%)culo (%)culo (%)13 (4) 12.5 (8) 12 (7) 6 (6)
Variante Granos/espiga
Granos vanos (%)
Rendimiento (Tm/ha)
Incremento(%)
ControlInoculanteInoculante
145153153
64
7.818.968.96
-1515
Tabla 33
Incremento del rendimiento de arroz producido por la
aplicación de biofertilizante mixto a base de A.
chroococcum y Az. brasilienseen 25 haciendas arroceras
Nota: Números entre parén-tesis en la Tabla indican el nú-
mero de campos evaluados
Tabla 34
Efecto de la aplicación del biofertilizante mixto a
base de A. chroococcum y Az. brasiliense sobre el rendimiento del arroz en
condiciones de producción

131la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Como puede verse, la información existente demuestra que existe una total compatibi-lidad entre las más importantes bacterias fijadoras de Nitrógeno atmosférico de forma simbiótica o asociativa y entre ellas y las bacterias solubilizadoras del Fósforo del suelo. Pero cada uno de los inóculos a base de estos microorganismos se fabrican y aplican de forma individual, en una secuencia que encarece los bioproductos y que complica la agrotecnia de un determinado cultivo, por lo que muchas veces los productores agrí-colas renuncian a su aplicación, a pesar de los evidentes beneficios que reciben de esta práctica. Por estas razones, se hace necesario encaminar los esfuerzos en el sentido de obtener bioproductos mixtos, con lo cual se abarata su costo y con cuya aplicación pueden aportarse a los cultivos los beneficios derivados de la actividad de cada uno de los microorganismos que integran la mezcla.
Para lograr esto, es necesario determinar el grado de compatibilidad que existe en un mismo sustrato entre los distintos microorganismos que pueden utilizarse, así como la viabilidad de cada microorganismo en el sustrato que se va a emplear en distintas condiciones ambientales. Hay que tomar en cuenta que, durante los últimos 30 años, se han publicado gran número de trabajos en relación con los efectos de las sustancias antagonistas sintetizadas por algunos microorganismos del suelo sobre las bacterias fijadoras de N2 atmosférico, y se han reportado numerosos microorganismos que son fuertes antagonistas para estas bacterias y que disminuyen de manera considerable la cantidad de N2 fijado cuando crecen en cultivos mixtos. Lo mismo ocurre en relación con las bacterias solubilizadoras del fósforo del suelo.
El mayor número de microorganismos productores de sustancias inhibidoras del cre-cimiento de estos dos grupos de bacterias son actinomicetos y hongos y, en mucha menor relación, bacterias, especialmente las que pertenecen al género Bacillus (Mar-tínez Viera, 1986). Por su parte, las bacterias con características de biofertilizantes tie-nen también una apreciable actividad antagonista, como se expuso en los casos de A. chroococcum y Az. brasiliense.
Debido a la existencia de estas interrelaciones negativas, que afectan tanto a la multi-plicación de algunas especies como a sus funciones biológicas, se hace necesario de-terminar, en un primer paso, los posibles efectos antagonistas que pueden producirse entre las asociaciones microbianas que se pretende establecer artificialmente, para se-leccionar solo aquellas especies y cepas que son totalmente compatibles o que tienen

132 capítulo 7. preparación y aplicación de inoculantes a base de bacterias fijadoras de nitrógeno atmosférico
un grado muy ligero de incompatibilidad. Después de hacer esta selección, pueden empezar a prepararse los biofertilizantes mixtos.
Determinación del efecto antagonista entre las especies bacterianas aspirantes a ser utilizadas en un producto mixto
Sobre la superficie del medio de cultivo agarizado más adecuado para el desarrollo de los microorganismos que se van a probar se abren concavidades en las cuales se coloca cultivo líquido de las especies cuyo antagonismo se quiere determinar. Sobre el césped de la placa se realiza una siembra extensa de la especie bacteriana que se va a probar. En caso de existir antagonismo, se produce un halo de no crecimiento alrededor de las concavidades, debido a la difusión de las sustancias antagónicas en el medio de cultivo. El diámetro de este halo se mide con una regla.
Se ha establecido una escala para determinar la compatibilidad interespecífica (Martínez Viera, 1986):
Halo de 0 – 2.5 mm se consideran especies compatiblesHalo de 2.6 – 5.0 mm son especies con compatibilidad aceptableHalo superior a 5.0 mm se consideran especies incompatibles.
En la tabla 35 se presentan, a modo de ejemplo, algunos resultados que se han obte-nido en estas pruebas, expresados en cm.
Césped INIFAT-9 INIFAT-12 INIFAT-17
Bacillus megatherium 0.16 0.11 0.13Pseudomonas fluorescens 0.15 0.14 0.10Bacillus subtilis 0.15 0.14 0.18Azospirillum brasilense 0.10 0.13 0.08Rhizobium etlii 0.16 0.20 0.16Rhizobium ciceri 0.14 0.16 0.14
Concavidades
Tabla 35
Acción antagonista de 3 cepas comerciales de
Azotobacter chroococcum frente a otras 6 especies de bacterias con características
de biofertilizantes

133la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Métodos de inoculación de los biofertilizantesa base de bacterias fijadoras de Nitrógeno atmosférico
En lo que respecta a los métodos de inoculación, existen muchos que se aplican en distintos países, unos más seguros y efectivos que otros, aunque siempre hay que aplicar aquel que le ha dado mejores resultados a cada productor, de acuerdo con su propia experiencia. Uno de los aspectos más importantes que deben ser manejados para tener éxito en la inoculación tiene que ser la calidad del inóculo, sobre lo cual se habla extensamente en párrafos anteriores. Los métodos de inoculación más utilizados son los siguientes:
1. Polvo: seco, en base a portadores. Se aplica el polvo sobre las semillas. Es rápido, simple, pero se pierde inoculante antes de alcanzar el suelo y no es muy durade-ro.
2. Suspensión: igual que el anterior, pero suspendido en agua o solución adhesiva. Se aplica a las semillas y se deja secar. No requiere equipo especial. Las semillas se secan al abrigo de la luz solar. El procedimiento es algo lento y a veces hay que reinocular. No puede utilizarse cuando las semillas están tratadas con fungicidas.
3. Sólido: Mezcla de gránulos de polietileno, polvo de mármol, zeolita u otro produc-to. Se mezcla con inoculante con base de portador y se aplica en el surco con las semillas. Mantiene la viabilidad y puede aplicarse con semillas tratadas con fungui-cidas.
4. Peletización: Inoculante con base de turba u otro portador, aplicado a la semilla y mezclado por agitación con un material peletizador. La semilla se seca. Protege al inoculante contra factores ambientales perjudiciales. La preparación consume tiempo y se observa a veces alta mortalidad en las bacterias.
5. Aspersión: Inoculante basado en un portador, suspendido en agua y aplicado a los surcos. No hay problemas para la conservación ni la preparación. Es adecuado para semillas tratadas con funguicidas.
6. Pre-inoculación: Inoculación de las semillas antes de venderlas, lo que permite un período de almacenamiento de la semilla inoculada antes de la siembra. Si no está bien preparado el inóculo puede ocurrir un alto nivel de mortalidad de bacterias.

134 capítulo 7. preparación y aplicación de inoculantes a base de bacterias fijadoras de nitrógeno atmosférico
7. Aplicación del inoculante líquido, sin portador, utilizando una asperjadora de es-palda o una mecánica si la superficie que debe aplicarse es extensa. Se utiliza en el caso de semillas muy pequeñas, como las de tabaco, lechuga, cebolla o tomate. La aplicación se hace a la superficie del suelo, inmediatamente después de la siembra y antes del riego, para que el inoculante penetre en el suelo con el agua de riego.
8. Aplicación del inoculante líquido por aspersión aérea, cuando se trata de extensas superficies de frutales, de arrozales o de otros cultivos, aprovechando la facilidad que tienen estas bacterias para establecerse sobre las hojas, como ya se explicó en el Capítulo 3.
9. Aplicación del inoculante líquido por el sistema de riego por aspersión o por go-teo.
De todos estos métodos, el más simple y el más generalmente utilizado en todo el mundo consiste en la mezcla del inoculante en polvo con las semillas de leguminosas antes de la siembra. Este proceso se realiza con las semillas correspondientes a 1 ha en un lugar sombreado, para evitar el efecto perjudicial de los rayos del sol. Las semillas se colocan sobre un piso de cemento limpio o sobre una manta plástica limpia.
Se vacía el contenido de una bolsa de inoculante (500 g) en un tobo plástico que con-tiene 1 L de agua en la que se han disuelto 100 g de azúcar. Se mezcla bien hasta que se forme una masa fluida, deshaciendo con los dedos los grumos que aparezcan.
La mezcla se vierte sobre las semillas y se revuelve varias veces con una pala pequeña hasta que todas las semillas se cubran con la mezcla preparada que se aplicó. Se dejan secar durante media hora, para que el inoculante se adhiera a la superficie de las semi-llas y se procede a la siembra.
Es esencial la selección del inoculante más apropiado para una determinada empresa agrícola, especialmente en aquellos suelos donde existen cepas autóctonas inefectivas que van a competir con las inoculadas por la formación y ocupación de los nódulos. Por ello, para comprobar la calidad del inoculante y conocer si será efectivo y aportará beneficios al cultivo que se ha programado en una región, se recomienda a los pro-ductores efectuar una pequeña prueba un mes antes del momento en que ha decidido efectuar la siembra de la leguminosa.

135la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
La prueba consiste en preparar tres pequeñas parcelas de 12 m2 cada una, en las que se va a sembrar la leguminosa que se quiere probar. En la primera parcela se aplica el fertilizante nitrogenado y no se hace inoculación. En la segunda ni se aplica fertilizante ni se hace inoculación y en la tercera se hace la inoculación y no se aplica fertilizante. Después de un mes se arrancan las plantas y se comprueba el resultado en las tres parcelas.
Se pueden presentar cualquiera de las siguientes 7 situaciones, las cuales permitirán definir si debe ser aplicado el inoculante comercial en ese campo o si la aplicación no va a tener efectividad:
Situación 1: La variante no inoculada no forma nódulos y el crecimiento es pobre. Igual ocurre con las variantes inoculada y con aplicación de N. La explicación es que influye otro factor limitante distinto del N ( por ejemplo falta de Fósforo).
Situación 2: Las variantes no inoculada e inoculada no forman nódulos y el crecimiento es pobre. La variante con N no forma nódulos y el crecimiento es bueno. La explica-ción es que la cepa utilizada no es satisfactoria o que hay condiciones adversas en el suelo para la infección.
Situación 3: La variante no inoculada no forma nódulos y el crecimiento es pobre; la variante inoculada forma nódulos efectivos y el crecimiento es bueno. La variante con N puede o no formar nódulos y el crecimiento es bueno. La explicación es que la cepa es efectiva para la nodulación y la fijación.
Situación 4: La variante no inoculada no forma nódulos y el crecimiento es pobre; la variante inoculada presenta nódulos inefectivos y el crecimiento es pobre; la variante con N puede o no formar nódulos y el crecimiento es bueno. La explicación es que la cepa no es apropiada.
Situación 5. La variante no inoculada forma nódulos inefectivos y su desarrollo es po-bre; la variante inoculada forma nódulos efectivos y su crecimiento es bueno; la varian-te con N puede o no formar nódulos y su crecimiento es bueno. La explicación es que la cepa es competitiva frente a las cepas nativas.
Situación 6: La variante no inoculada forma nódulos inefectivos y su desarrollo es po-bre; la variante inoculada forma nódulos inefectivos y su crecimiento es pobre; la va-riante con N puede o no formar nódulos y su crecimiento es bueno. La explicación es que la cepa no es competitiva o es inefectiva.

136 capítulo 7. preparación y aplicación de inoculantes a base de bacterias fijadoras de nitrógeno atmosférico
Situación 7. Las variantes no inoculada e inoculada forman nódulos efectivos y el cre-cimiento es bueno; la variante con N puede o no formar nódulos y el crecimiento es bueno. La explicación es que no hace falta aplicar inóculo.

137la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Bibliografía
Abd-El-Malek, Y., (1971): “Free living nitrogen-fixing bacteria in Egyptian soils and their posible contributions to soil fertility”. Plant and Soil, Special Vol.: 423-442.
Adler, J. (1972): “A method for mesuring chemotaxis and use of the method to deter-mine optimum conditions for chemotaxis by Escherichia coli”. J. of General Micro-biology, 74: 77-91.
Alagwady R. y G. Baur (1988): “Effects of Rhizobium and phosphate solubilizing bacte-ria on the yield and nutrient uptake of chickpea”. Plant and Soil, 105 (2): 241-246.
Alexander, M. (1977): Introduction to Soil Microbiology. John Wiley and sons, Nueva York, 460 pp.
Altieri, M.I. y A. Yukevich (1991): “La agroecología y el desarrollo rural sostenible en América Latina”. Agroecología y Desarrollo, 1: 25-36.
Altieri, M.A. y A. Rojas (1999): “Ecological impacts of Chile´s neoliberal policies, with special emphasis in agroecosystems”. Environment, Development and Sustainability, 1: 55-72.
Altieri, M.A. y C.I. Nicholls (2000): “Ecological impacts of modern Agriculture in the United States and Latin America”. En Globalization and the Rural Environment, pp. 121-135.
Altieri, M.A., S. Hetch, M. Liebman, F. Magdoff, R. Norgaard y T. O. Sirkor (1999): Agroecología: Bases científicas para una Agricultura Sustentable. 201 pp.
Alves, B.J.R., L. Zotarelli, W.A.R. Lara, E. Torres, M. Hungria, S. Urquiaga y R.M. Bod-dey (2000): “Benefit of legume fixed N in crop rotation under zero tillage”. En Nitrogen Fixation: From Molecules to Crop Productivity. Kluwer Acad. Publ., Dor-drecht, pp. 523-534.

138 bibliografía
Alves, B.J.R., R.M. Boddey y S. Urquiaga (2003): “The success of BNF in soybean in Brazil”. En Maximizing the Use of Biological Nitrogen Fixation in Agriculture. Kluwer Academic Publishers, Londres, pp. 1-9.
Anónimo (2004): Agricultural Tecnologies: Manures and Fertilizers. (//A:/Medios%20de%20cultivo/azotobacter.htm)
App, A., T. Santiago, C. Ventura y J. Watanabe (1984): “Estimation of the nitrogen ba-lance for irrigated rice and the contribution of phototrophic nitrogen fixation”. Field Crop Res., 9: 17-27.
Arshad, M. y W.T. Frankenberger (1990): “Microbial production of plant hormones”. Plant Soil, 133: 1-8.
Arshad, M. y W.T. Frankenberger (1993): “Microbial production of plant growth regula-tors”. En Soil Microbial Ecology, Marcel Dekker Ed., Nueva York, pp. 307-344.
Asociación Internacional de la Industria de Fertilizantes (2003): Consumo de fertilizante nitrogenado en el mundo. Informe Anual, 87 pp.
Atzorn, R., A. Crozier, C.T. Wheeler y G. Sandberg (1988): “Production of gibberellins and indole-3 acetic acid by Rhizobium phaseoli in relation to nodulation of Phaseo-lus vulgaris roots”. Planta, 175: 532-538.
Azcon, R. y J.M. Barea (1975): “Synthesis of auxins, gibberellins and cytokinins by Azotobacter vinelandii and A. beijerinckia related to effects produced on tomato plants”. Plant Soil, 43: 609-619.
Azcón, R., M. Gómez y J.M. Barea (1975): “Efectos de la aplicación conjunta de ferti-lizantes químicos y microbianos en cultivos enarenados”. An. de Edafología y Agro-biología, 33 (9-10): 863-870.
Bach, T., J. Ferrán y A. Delgado (2000): Obtención de un biofertilizante multipropósito a partir de la Fosforina. Informe presentado al Comité de Expertos del CITMA, PMCT Biotecnología Agrícola, 81 pp.

139la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Bagyraj, D.J. y J.A. Menge (1978): “Interaction between VA-mycorhyza and Azoto-bacter and their effects on rhizosphere microflora and plant growth”. New Phytol., 80 (3): 567-573.
Barea, J.M., E. Navarro y E. Montoya (1976): “Production of plant growth regulators by rizosphere phosphate-solubilizing bacteria”. J. Appl. Bacteriol., 40: 129-134.
Barea, J.M., J.A. Ocampo y E. Montoya (1977): “Estudio crítico sobre la utilización de Azotobacter y fosfobacterias como fertilizantes microbianos”. An. de Edafología y Agrobiología, 36 (11-12): 1198-1208.
Bashan,Y. (1998): “Inoculants of plant growth-promoting bacteria for use in Agricul-ture”. Biotechnology Advances, 16 (4): 729-770.
Bashan, Y. y G. Holguin (1997): “Azospirillum-plant relationship: environmental and physiological advances”. Canadian J. Microbiol., 103-121.
Bauer, T. (2001): Microorganismos fijadores de nitrógeno. (http//www microbiologia.com/nf/suelo/rhizobium.html)
Bazilevich, N.I. (1974): “Energy flor and biogeochemical regularities of the main world ecosystems”. En Proceed. of I Int. Cong. of Ecology, Wagenigen, pp. 182-186.
Beck, E., Fubeder a., Graus M., 1989. “The maize root system in situ: evaluation of structure and capability of utilisation of phytate and inorganic soil phosphates”. Zeitschrift füZeitschrift füZeitschrift f r pflanzenerährung und bodenkunde 152, 159-167.
Becking, J.H. (1977): “Endophyte and associations establishment in non-leguminous ni-trogen-fixing plants”. En Recent Developments in Nitrogen Fixation, Academic Press, Nueva York, pp. 559-567.
Becking, J.H. (1982): “N-fixing tropical non-legumes”. En Microbiology of Tropical Soils and Plant Productivity. Martinus Wijhoff y W. Junk Publishers, La Haya, pp. 109-146.
Bhat, J.B., E.S. Limayev y B.L. Vasantharajan (1971): Ecology of the leaf surface micro-organisms, 322 pp.

140 bibliografía
Biswas, J., J.K. Lhada y F.B. Dazzo (2000): “Rhizobia inoculation improves nutrient up-take and growth in lowland rice”. Soil Sci. Soc. Am. J., 64: 1644-1650.
Böckman O. (1997): “Non biological nitrogen fixation”. En Biological Nitrogen Fixation. The Global Challenge and Future Needs. Roma, pp. 24-26.
Boddey, R.M. y J. Dobereiner (1988): “Nitrogen fixation associated with grasses and cereals: recent results and perspectives”. Plant and Soil, 108: 53-65.
Boddey, R.M., K.E. Giller, B.J. Alves y S. Urquiaga (1998): “Contribution of biological nitrogen fixation to tropical agriculture: actual potential”. En Biological Nitrogen Fixa-tion for the 21st Century. Kluwer Academic Publ., Dordrecht, pp. 599-604.
Boddey, R.M., B.J.R. Alves, O.C. Oliveira y S. Urquiaga (2000): “A degradaçao das pastogens e o ciclo do nitrogenio”. En Workshop nitrogeniona sustentabilidade de sistemas intensivos de produçao agropecuaria .Eds. F.M. Mercante y M.S. Dourados, pp. 110-124.
Boddey, R.M., S. Urquiaga, B.J. Alves y V. Reis (2003): “Endophytic nitrogen fixation in sugar cane: present knowledge and future applications”. Plant and Soil, 252: 139-149.
Boddey, R. M., B.J.R. Alves, V.M. Reis y S. Urquiaga (2006): “Biological Nitrogen Fixa-tion in Agroecosystems and in Plant Roots”. En Biological Approaches to Sustainable Soil Systems, Taylor and Francis Ed., Boca Ratón.
Bohlool, B.B., J.K.Ladha, D.P. Garrity y T. George (1992): “Biological nitrogen fixation for sustainable agriculture: a perspective”. Plant and Soil, 141: 1-11.
Bohorquez, P. (2001): Aislamiento y caracterización de bacterias solubilizadores de fosfa-tos. Tesis de Grado, Universidad Nacional de Colombia, 52 pp.
Borroto, C. (2005): Palabras de inauguración del Congreso Biotecnología Habana 2005. (http:// w.w.w. prensalatina.com.mx/print.asp?ID
Brockwell, J., R. R. Gault, D.F. Herridge y L.J. Morthorpe (1988): “Studies on alter-native means of legume inoculation: microbiological and agronomic appraisals of

141la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
commercial producers for inoculating soybeans with Bradyrhizobium japonicum”. Aust. J. Agric. Res., 39: 965-972.
Bromfield, E.S.B. y A. Ayanaba (1980): “The efficacy of soybean inoculation on acid soil in tropical Africa”. Plant and Soil, 54: 95-106.
Broughton, W.J., F. Zhang, X. Perret y C. Stachelin (2003): “Signals exchanged bet-ween legumes and Rhizobium: Agricultural uses and perspectives”. Plant and Soil, 249: 365-373.
Brown, M.E. y S.K. Burlingham (1968): “Production of plant growth substances by Azotobacter chroococcum”. J. Gen. Microbiol., 53: 135-144.
Brown, M.E. y G.R. Carr (1984): “Interaction between Azotobacter chroococcum and vesicular – arbuscular mycorhiza and their effects on plant growth”. J. Appl. Bacteriol., 56 (4): 429-437.
Brown L.R. (1994): “Facing food inecurity”. En State of the World, Earthcan Publications Ltd, Londres, pp. 110-131.
Bûll, L., S. Lacerda y J. Nakagawa (1997): “Fertilidades do solo e nutriçao de plantas. Termofosfatos: Alteraçoes em propiedades químicas em um latossolo vermelho-oscuro e eficiencia agronómica”. Bragantia, 56(1): 169-179.
Bullow, .W. von y J. Dobereiner (1975): “Potential for nitrogen fixation in maize geno-types in Brazil”. Proc. Natl. Acad. Sci. USA, 72: 2389-2393.
Burdman S., E. Jarkevicht e Y. Okón (2000): “Recent advances in the use of PGPR in Agriculture”. En Microbial Interactions in Agriculture and Forestry, . Science Publis-hers Inc., Plymouth, UK, pp. 29-250.
Burdman, S., B. Hamnoni e Y. Okon (2000): Improvement of legume crop yields by co-inoculation with Azospirillum and Rhizobium. Jerusalem. Center for Agricultural Biotechnology, Israel.
Burris, R.H., T.L. Jones y D.W. Elmerich (1978): “Nitrogenase systems”. En Limita-tions and Potentials for Biological Nitrogen Fixation in the Tropics, Plenum Press Ed., Nueva York, pp. 191-209.

142 bibliografía
Caballero-Mellado, J. (2003): “El género Azospirillum”. En (http//biblioweb.dgsca.unam.mx/Libros/Microbios)
Caldwell, E.E. y N.G. Vest (1977): “Genetic aspects of nodulation and dinitrogen fixa-tion by legumes”. En A Treatise on Dinitrogen Fixation. John Wiley and sons Ed., Nueva York, pp. 557-577.
Calegari, A. (2000): “Coberturas verdes em sistemas intensivos de produçao”. En Wor-kshop nitrogeniona sustentabilidade de sistemas intensivos de produçao agropecuaria.Eds. F.M. Mercante y M.S. Dourados, pp. 141-153.
Cárdenas, L., J, A, Feijóo, J. K. Kunkel, F. Sánchez y P. Hepler (1999): “Rhizobium Nod factors induce increases in intracellular free calcium and extracellular calcium influxes in bean root hairs”. Plant J., 19: 347-352.
Cárdenas, L., T.L. Holdaway, F. Sánchez y P. Hepler (2000): “Ion changes in legume root hairs responding to Nod factors”. Plant Physiology, 123: 443-451.
Carradell, J. y cinco colaboradores (1996): “Maximum rooting depth of vegetation types at the global scale”. Oecologia, 108: 583-595.
Casanova, E. (1993): Las rocas fosfóricas y su uso industrial en Venezuela. Apuntes técni-cos de Palmaven, Caracas, 124 pp.
Catroux, G. (1991): “Inoculant quality standars and controls in France”. En Report of the expert consultation on legume inoculant production and quality control, FAO, Roma, pp. 113-120.
Cavalcante, V:A: y J. Dobereiner (1988): “A new acid-tolerant nitrogen-fixing bacterium “A new acid-tolerant nitrogen-fixing bacterium “associated with sugarcane”. Plant and Soil, 108: 23-31.
Chen Ting Wey (1997): “Development and application of biofertilizers in China. En Biological Nitrogen Fixation: The Global Challenge and Future Needs. Roma, pp. 24-26.
Chi, F., S.H. Shen, H.P. Chen, Y.G. Yanni y F.B. Dazzo (2005): “Ascending migration of “Ascending migration of “endophytic Rhizobia from roots to leaves inside rice plants and assessments of their benefits to the growth physiology of rice”. Appl. Envir. Microbiol., 71: 7271-7278.

143la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Chowdhary, M., L. Bailey y T. Peck (1996): “Effect of rock phosphate and superphos-phate on crop yield and soil phosphorus test in long-term fertility plots”. Commun. Soil Sci. Plant Anal., 27(18-20): 3085-3099.
Chhoucar, P.K. y N.S. Subba Rao (1967): “Phosphate solubilization by fungi associated with legume root nodules”. Canada J. Microbiol., 13: 749-753.
Chude, V.O. (1994): “Response of tomato to nitrogen fertilization and irrigation fre-quencies in a semiarid tropical soil”. Fertilizer Research, 4(20): 85-88.
Cleveland, C.C. A.R. Townsend y A.R. Schinel (1999): “Global patterns of terres trial biological nitrogen fixation in natural ecosystems”. Global Biogeochem. Cycles, 13: 623-645.
Cocking, E.C. (2003): “Endophytic colonization of plant roots by nitrogen- fixing bacte-ria”. Plant and Soil, 252: 169-175.
Comité Internacional de Investigación Agrícola, TAC-CGIAR (1988): Sustainable Agri-cultural Production: Implications for International Agricultural Research, Berlín, 45 pp.
Curl E.A. y A. Truelover (1986): The Rizosphere, Springler-Verlag, Nueva York, 288 pp.
Crum, A. (2004): “Azotobacter”. En File//A:/BIOL-CSES 4684-Azotobacter.htm
Dalal, R.C. (1977): “Organic phosphorus”. Dvances in agronomy 29, 83-117.Dvances in agronomy 29, 83-117.Dvances in agronomy
Dazzo, F.B., M.R. Urbano y G. Truchot (1981): “Regulation of recognition in the Rhi-zobium-clover symbiosis”. En Current Perspectives in Nitrogen Fixation, Australian Academy of Sciences.
Dazzo, F.B. e Y.G. Yanni (2006): “The Natural Rhizobium - Cereal Crop Association as an Example of Plant - Bacteria Interaction”. En Biological Approaches to Sustainable Soil Systems. Taylor and Francis Ed., Boca Ratón.
Debinstein J. (1970): A Tropical Rain Forest, John Wiley and sons Ed., Nueva York, 230 pp.

144 bibliografía
Diario “El Nacional”, Caracas (2006): El Mundo de la Agricultura Orgánica. Edición 4 de marzo, pp. 6.
Dibut, B. (2000): Obtención de un bioestimulador del crecimiento y el rendimiento para el beneficio de la cebolla. Tesis de Doctorado, La Habana, 104 pp.
Dibut, B. y R. Martínez Viera (2003): “Biofertilizantes y bioestimuladores: Métodos de inoculación”. En: Manual de Agricultura Orgánica Sostenible, FAO, La Habana, pp. 17-22.
Dibut B, R. Seoane y R. Martínez Viera (1996): “Efecto de la doble función de A. chro-ococcum sobre el cultivo del trigo en suelos Ferraliticos Rojos”. En Resúmenes del III Congreso Cubano de Ciencias del Suelo, La Habana, pp. 57-59.
Dibut, B., A. Rodríguez y R. Martínez Viera (1996): “Efecto de la doble función de Azotoryza sobre el cultivo del plátano”. Infomusa, 5 (1): 20-23.
Dibut, B., R. Martínez Viera, R. García, T. Yemischi y G. Kaya (2001): “Efecto de la aplicación de DIMARGON sobre diferentes cultivos económicos en Turquía”. En Memorias del XV Congreso Latinoamericano de la Ciencia del Suelo, vol. III, Varadero, Cuba.
Dibut, B., R. Martínez Viera, Y. Ríos y M. Ortega (2003): “DIMARGON-M, nueva va-riante nutritiva para la producción de biofertilizantes y bioestimuladores a base de Azotobacter”. En Resúmenes del V Encuentro de Agricultura Orgánica, La Habana, pp. 36.
Dibut, B., R. Martínez Viera, M. Ortega (2004): “Desarrollo y aplicación de bioestimu-ladores microbianos”. En Memorias del Congreso “TróMemorias del Congreso “TróMemorias del Congreso “Tr pico 2004”, La Habana, CD.
Dibut, B., M. Ortega, R. Martinez Viera, L. Fey y Y. Rios (2005): “Aislamiento y distribu-Dibut, B., M. Ortega, R. Martinez Viera, L. Fey y Y. Rios (2005): “Aislamiento y distribu-Dibut, B., M. Ortega, R. Martinez Viera, L. Fey y Y. Rios (2005): “ción de Gluconacetobacter diazotrophicus en cultivos de importancia económica para Cuba”. Cultivos Tropicales.
Dinkelaker, B., Marschner, H. (1992): “In vivo demonstration of acid phosphatase activ-ity in the rhizosphere of soil grown plants”. Plant and Soil, 144, 199-205.

145la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Dôbereiner, J. (1983): “Dinitrogen fixation in rhizosphere and phyllosphere associa-tions”. En Encyclopedia of Plant Physiology, Springer Verlag Ed., Berlin, pp. 332-350.
Döbereiner, J. (1987): Nitrogen- Fixing Bacteria in Non-Leguminous Crop Plants. Sprin-gler Verlag Ed., Berlín.
Dôbereiner, J., (1992): “History and new perspectives of diazotrophs in association with non-leguminous plants”. Symbiosis, 13: 1-13.
Dôbereiner, J. (1993): Fixaçao do nitrogenio em associaçao com gramineas. Ed. Cardo-zo, Rio de Janeiro, 320 pp.
Dôbereiner, J. (1993): “Recent changes in concepts of plant-bacteria interactions: en-dophytic N2-fixing bacterias”. En Ciencia e Cultura (Brasil), pp. 1-16.
Dôbereiner,J., (1998): “The genera Azospirillum and Herbaspirillum”. En Biological Ni-trogen Fixation. Marcel Dekker Inc., Nueva York, pp. 235-267.
Döbereiner J. y J.L. Day (1976): “Associative symbiosis in tropical grasses. Character-“Associative symbiosis in tropical grasses. Character-“ization of microorganisms and dinitrogen-fixing sites”. En Simposium on Nitrogen Fixation, Vol. 2. Washington States University Press, pp. 516-538.
Dôbereiner, J., E. Marriel y M. Nery (1976): “Ecological distribution of Spirillum lipo-ferum Beijerinck”. Can. J. Microbiol., 22 : 1464-1473.
Dôbereiner, J.,V.M. Reis y F. Olivares (1993): “Endophitic diazotroph in cereals, sugar-cane and tuber plants”. En New Horizons in Nitrogen Fixation. Kluwer Academic Publ., Dordrecht, pp. 671-676.
Dôbereiner, J., V.I. Baldani y F. Olivares (1995): “Endopdhyte Diazotrophs. The key to BNF in gramineous plants”. En Nitrogen Fixation with non Legumes. The American University of Cairo Press, pp. 395-408.
Dobbelaere, S., A. Croonenborghs y A. Tiss (2001): “Azospirillum“Azospirillum“ ”. Aust. J.Plant Phy-siol., 28: 871-879.
Dommergues, Y. (1963) : “Evaluation du taux de fixation de l´azote dans un sol dunaire reboisié en filao (Casuarina equisetifolia) ”. Agrochimica, 7: 335-340.

146 bibliografía
Dommelen, A.V. (1998): “Ammonium transports in Azospirillum brasilense”. Disserta-tione de agriculture, 7: 1-9.
Dreyfus, B.L., H.G. Diem e Y. Dommergues (1988): “Future directions for biological nitrogen fixation research”. Plant and Soil, 108: 190-191.
Durkhead, P., A. David y P. Suninger (1998): “Bioautography shows antibiotic produc-tion by soil bacterial isolated antagonistic to fungal dry rot of potatoes”. Soil Biology and Biochemistry, 27(12): 1611-1616.
Dyson T. (1996): Population and Foods. Global Trends and Future Prospects. Global En-vironmental Programme, Rutledge, ISBN. 0-415-11975-8, 45 pp.
Eaglesham, A., B. Seaman y K. Mulongoy (1981): “High-temperature tolerant cow-pea rhizobia”. En Current Perspectives in Nitrogen Fixation, Australian Academy of Sciences, Camberra, pp. 436-441.
Ellen, R. (1982): Environment, Subsistence and Systems, Londres, Cambridge University Press, 235 pp.
Elmerich, C., (1984): “Molecular Biology and Ecology of diaoztrophicus associations with non-leguminous plants”. Biotechnology, 1: 967-970.
Elmerich, C., W. Zimmer y C. Vieille (1992): “Associative nitrogen-fixing bacteria“Associative nitrogen-fixing bacteria“ ”. En Biological Nitrogen Fixation, Chapman and Hall, Nueva York, pp. 212-258.
España, M; E. Cabrera-Bisbal; M López (2006): “Study of nitrogen fixation by tropical legumes in aced soil from Venezuelan Savannas using 15N”. Terciencia. 31 (3): 197-201.
España M; E Cabrera de B y B. Rodríguez (2000): “Evaluaciones de la fijación biológica de nitrógeno de la soya en dos sistemas de labranaza mediante la utilización de ISN. En la fijación biológica del nitrógeno en América Latina. El aporte de las técni-cas isotópicas”. Arcal México, 117-120.
Espindola J.A.A., Guerra J.G.M., Almeida D.L., Teixeira M.G., Urquiaga S. & Busquet R.N.B. (2000). “Estimativa da fixação biológica de nitrogênio em três leguminosas herbáceas perenes e sua contribuição no fornecimento de nitrogênio para a ba-

147la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
naneira”. CD-ROM, XIII Reunião Brasileira de Manejo e conservação do solo e da água. água. á Ilhéus, Bahia.
FAO (1991): Expert consultation on legume inoculant production and quality control, Roma, 144 pp.
FAO (1995): Manual Técnico de la fijación biológica del nitrógeno, Roma, 160 pp.
FAO (2001a): Production Yearbook 2000, Roma, 243 pp.
FAO (2001b): w.w.w.fao.org.
Fernández, S.M., L.M. Meléndez, C. Meza y R. Noguera (2001): “Evaluación agronómi-ca de rocas fosfóricas modificadas en tres condiciones de pH, utilizando maíz como cultivo indicador”. En Memorias del XV Congreso Latinoamericano de Ciencia del Suelo, Comisión III, pp. 69-73.
Franco, A.A., J.R.R. Peres y M. Nery (1978): “The use of Azotobacter paspali N2-ase to measure molybdenum deficiency in soils”. Plant and Soil, 50: 1-11.
Franke, I.H., M. Fegan, C. Hayward, G. Leonard, E. Strackebrandt y L.I. Sly (1999): “Description of Gluconacetobacter sacchari sp.nov., a new species of acetic acid bacterium isolated from the leaf sheath of sugar-cane and from the pink sugar-cane mealy bug”. Int. J. of Syst. Bacteriol., 49: 1681-1693.
Frioni, L. (1999): Procesos microbianos. Universidad Nacional de Río Cuarto, Córdoba, 286 pp.
Fuentes-Ramirez, L.E., J. Caballero-Mellado y E. Martínez-Romero (1999): “Coloni-zation of sugarcane by Gluconacetobacter diazotrophicus is inhibited by high N-fertilization”. FEMS Microbiol. Ecol., 29: 117-128.
Fuentes-Ramírez, L.E., R. Bustillos, A. Tapia, T. Jiménez y J. Caballero-Mellado (2001): “Novel nitrogen-fixing acetic acid bacteria, G. johannae sp.nov. y G. azotocampans sp. Nov., associated with coffee plants”. Int. J. Syst. Evol. Microbiol., 1(5): 43-49.
Galal, Y.G.M. (1997): “Dual inoculation with strains of Bradyrhizobium japonicum and Azospirillum brasilense to improve growth and biological fixation in soybean”. Biol. Fert. Soils, 24 (3): 317-322.

148 bibliografía
García, R., R. Martínez Viera, B. Dibut y G. Tejeda (2004): “Desarrollo de un biofertili-zante solubilizador del fósforo y estimulador del crecimiento vegetal”. En Memorias del Congreso “Trópico 2004”, La Habana.
Garrity, G.M. y J.G. Holt (2001): “The road map to the manual”. En Bergey´s Manual of Systematic Bacteriology, vol 1, pp. 119-166.
Gaur, A.C. y K.P. Ostwald (1977): “Influence of phosphate dissolving bacilli on yield and phosphate uptake of wheat crop”. Indian J. Exptl. Biol., 10: 393-398.
Gibsons, A.H., B.L. Dreyfus e Y.R. Dommergues (1982): “Nitrogen fixation by legumes in the tropics”. En Microbiology of Tropical Soils and Plant Productivity, M. Nithoff y W. Jung Publishers, La Haya, pp. 37-64.
Giller, K.E. (2001): Nitrogen Fixation in Tropical Cropping Systems, CABI, Wallingford, UK.
Gleddie S.C, D.Schlechte y G. Turnbull (1993): “Effect of inoculation with P. bilaii on phosphate uptake and yield of canola in Western Canada”. En Proceed. Alberta Soil Sciencie Workshop, Edmonton, pp. 155-160.
Glick, B.R. (1995): “The enhancement of plant growth by free-living bacteria”. Cana-dian J. Microbiol., 41: 109-117.
González, J. y C. Lluch (1992): Biología del nitrógeno. Interacción planta-microorganis-mo. Ed. Rueda, Madrid, 326 pp.
Gonzalez, M.S. y W.L. Barraquio (2000): “Isolation and characterization of Glucona-cetobacter diazotrophicus in Saccharum officinarum, S. spontaneum and Erianthus sp”. The Philippine Agricultural Scientist, 83: 173-181.
Gopalaswamy, G. et al., (2000): “The xylem of rice (Oryza sativa) is colonized by Azor-hizobium caulinodans”. Proc. R. Soc. London, Ser. B, 267: 103-107.
Grierson, P.F., P. Smithson y N.B. Comerford (2004): “Phosphorus dynamics and mo-bilization by plants”. En Below-ground Interactions in Tropical Agroecosystems. CABI Publishers, Cambridge, pp. 127-140.

149la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Haaker, H. y C. Veeger (1977): “Involvement of the cytoplasmic membrane in nitrogen fixation by Azotobacter vinelandii”. Eur. J. Biochem., 77: 1-12.
Habte, M. y P. Manjunath (1987): “Soil solution P status and mycorrizhal dependency in Leucaena leucocephala”. Appl. Environm. Microbiol., 53: 797-801.
Hardarson, G. y S. K. A. Danso (1993): “Methods for measuring biological nitrogen fixation in grain legumes”. Plant and Soil, 152: 19-32.
Hardarson, G. y A. Craig (2003): “Optimising biological N2 fixation by legumes in far-ming systems”. Plant and Soil, 252: 41-54.
He, Z., V. Baligar, D. Martens y W. Kemper (1996): “Kinetics of phosphate rock disso-lution in acidic soil amended with liming materials and cellulose”. J. of Soil Sci. Soc. Am., 60: 1589-1595.
Hernández, A. (2002): Obtención de un biopreparado a partir de rizobacterias asociadas al cultivo del maíz. Tesis de Doctorado, La Habana, 96 pp.
Herridge, D.F. e I. Rose (2000): “Breeding for enhanced nitrogen fixation in crop legu-mes”. Field Crop Res., 65: 229-248.
Hetch, S.B. (1997): “La evolución del pensamiento agroecológico”. En: Agroecología: Bases científicas para una agricultura sustentable. CLADES-ACAO, La Habana.
Higa, T. (1995): “Effective microorganisms: Their role in Kusei Nature Farming and sus-tainable Agriculture”. En Proceedings of the Third International Conference on Kusei Nature Farming, Washington, pp. 161-184.
Hinsinger, P., G.R. Gobran, P.J. Gregory y W.W. Wennel (2005): “Rhizosphere ge-ometry and heterogeneity arising from root-mediated physical and chemical pro-cesses”. New Phytologist, 168: 293-303.
Hoffman, G. (2001): Organic Agriculture Worldwide. “Statistics and Further Prospects”. Stiftung Ökologie and Landbau, 76: 43-49.
Hogg, B., A.E. Davies, K.E. Wilson y J.A. Downie (2002): “Competitive nodulation blocking of cv Afganistan pea is related to high levels of Nod factors”. Mol. Plant Microbe Interact., 15: 60-68.

150 bibliografía
Huerta, J., I.A. Escalante, J.Z. Castellanos y J.A. Flores (2001): “Producción de biomasa y grano en frijol común en función de la fertilización nitrogenada y la inoculación con Rhizobium leguminosarum var.phaseoli”. (http:Ilesa.chapingo. mx//sometil RFM/20-1-es.html. Art. 5)
Hungria, M., R.J. Campo e I.C. Mendes (2000): “Fixacao biologica do nitrogenio com a cultura da soja”. En: Workshop nitrogenio na sustentabilidade de sistemas intensivos de producao agropecuaria, Ed. Mercante, Rio de Janeiro, pp. 51-71.
Hutton, E.M. y C.S. Andrews (1978): “Comparative effects of calcium carbonate on growth, nodulation and chemical composition of four Leucaena leucocephala lines”. Austr. J. of Austr. J. of Austr. J. of Exp. Agric. And An. Husb., 18: 81-88.
IFOAM (2001): What is IFOAM? (http://ecoweb.dk/ifoam)? (http://ecoweb.dk/ifoam)? (http://ecoweb.dk/ifoam
Irving, H.I., N.M. Boukli, M.N. Kelly y W. J. Broughton (2000): “Nod factors, in sym-biotic development of root-hairs”. En Root-Hairs Cell and Molecular Biology. Molecular Biology. Molecular Sprin-gler-Verlag, Tokyo, pp. 241-265.
Itzigsohn, R., S. Burdman e Y. Okon (2000): “Plant growth promotion in pastures by inoculation with Azospirillum brasilense under suboptimal growth conditions”. Arid Soil Research and Rehabilitation, 13: 151-158.
Izquierdo J. L. Ciampi y E. de García (1995): Biotecnología apropiable: racionalidad de su desarrollo y aplicación en América Latina y el Caribe, FAO, 65 pp.
Jackson, R.B. y cinco colaboradores (1996): “A global an“A global an“ álisis of root distributions for terrestrial biomes”. Oecologia, 108: 389-411.
Jara, R., B. Dreyfus e Y. Dommergues (1981): “Is it necessary to inoculate soybean in West Africa soils? ”. En Proceed. 8th North American Rhizobium Conference, Winni-peg, pp. 165-169.
Jarvis, B:D:W:, P. van Berkum y W:X: Chen (1997): “Transfer of Rhizobium loti, Rh. huakuii, Rh. ciceri, Rh. mediterraneum and Rh. thianshanense to Mesorhizobium gen. nov”. Int. J. Syst. Bacteriol., 47: 895-898.

151la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Jenkinson, D.A. (2001): “The impacts of humans on the nitrogen-cycle, with focus on temperate arable agriculture”. Plant and Soil, 228: 3-15.
Jensen, H.L. (1940): “Contributions to the nitrogen economy of australian wheat soils”. Proc. Linn. Soc. NSW, 65: 1-15.
Jimenez-Salgado T., L.E. Fuentes-Ramirez, A. Tapia-Hernández, E. Martínez-Romero y J. Caballero-Mellado (1997): “Coffea arabica L., a new host plant for Gluconace-tobacter diazotrophicus and isolation of other nitrogen-fixing acetobacteria”. Appl. Environ. Microbiol., 63: 3676-3683.
Joi, M.B. y P.A. Shinde (1976): “Response of onion crops to azotobacterization”. J. of Maharastra Agricultural University, 1: 161-162.
Jordan, D.C. (1982): “Transfer of Rhizobium japonicum to Bradyrhizobium gen”. nov. Int. J. Syst. Bacteriol., 32: 136-139.
Kalinska, D. y M. Drosdowic (1983): “Occurrence of microorganisms amtagonistic to Azospirillum sp”. Zentralbl. Mikrobiol., 138: 585-594.
Kamnas, K.M. y P. Kaiser (1997): “Diazotrophic mixed cultures of Azospirillum and Bacillus sp using pectin on rice plantlets as substrates”. En Proceed. XI Int. Cong. of Nitrogen Fixation, Paris, pp. 165-171.
Kannaiyan, J. (2003): “Inoculant production in developing countries. Problems, poten-tials and succes”. En Maximising the use of biological nitrogen fixation in agriculture, Kluwer Academic Press, La Haya, pp. 187-198.
Kapulnik, J., R. Gafuy e Y. Okon (1985): “Effect of Azospirillum inoculation on root de-velopment and nitrate uptake in wheat”. Can. J. Bot., 63: 627-631.
Katayama, A., T.P. Rao y O. Loto (1996): “Root system development of components in intercropping”. En: Dynamic od roots and nitrogen in cropping systems of the semi-arid tropics, SIRKAS,Tokyo, pp. 619-623.
Kelly, M.N., y H.R. Irving (2001): “Nod factors stimulate plasma membrane delimited phospholipase activity in vitro”. Physiol. Plant., 113: 461-468.

152 bibliografía
Kelly, M.N., y H.R. Irving (2003): “Nod factors activate both heterotrimetic and mo-nomeric G-proteins in Vigna unguiculata”. Planta, 216: 674-895.
Kennedy, I.R. (1997): “BNF: An energy-costly process? ”. En Biological Nitrogen Fixa-tion: The Global Challenge and Future Needs, Rockefeller Center, pp. 15-17.
Kennedy, I. R. e Y.T. Tchan (1992): “Biological nitrogen fixation in non-leguleguminous field crops: recent advances”. Plant and Soil, 141: 93-118.
Kennedy, I.R. y E. C. Cocking (1997): “Current status of BNF in global ecosystems”. En Biological Nitrogen Fixation: The Global Challenge and Future Needs, Rockefeller Center, pp. 13-14.
Killham, K. (1999): Soil Ecology, Cambridge University Press, Cambridge, 465 pp.
Kloepper, J. W. (1994): “Plant-growth promoting bacterias”. En Azospirillum/Plant Asso-ciations. CRC Press, Boca Ratón, pp. 137-154.
Knowles, R. (1981): “Nitrogen fixation in natural plant communities and soils”. En Me-thods for Evaluating Biological Nitrogen Fixation, John Wiley and sons, Nueva York, pp. 557-582.
Kraffczyk I., G Tolldeiner y H. Beringer (1984): “Soluble roots exudates of maize: in-fluence of potasium supply and rhyzosphere microorganims”. Soil Biol. Biochem, 16: 315-322.
Krieg N.R. y J.J. Tarrand (1978): “Taxonomy of the root associated nitrogen fixer Spi-rillum lipoferum”. En Limitations and Potentials for Biological Nitrogen in the Tropics. Plenum Press Ed., Nueva York, pp. 317-335.
Kumar, V. y N. Narula (1999): “Solubilization of inorganic phosphates and growth emergence of wheat as affected by Azotobacter chroococcum”. Biol. Fertil. Soil, 27: 301-305.
Kunda, B.S. y A. C. Gaur (1980): “Establishment of nitrogen fixing and phosphate so-lubilizing bacteria in rhizosphere and their effect on yields and nutrient uptake by wheat”. Plant and Soil, 57: 223-230.

153la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Kunda, B.S. y A. C. Gaur (1980a): “Effect of nitrogen fixing and phosphate solubilizing microorganisms as single and composite inoculants on cotton”. Indian J. Microbiol., 20: 225-229.
Kunda, B.S. y A. C. Gaur (1982): “Yield increase of wheat after inoculation with A. chroococcum and phosphobacterias”. Curr. Sci., 51: 291-293.
Kunda, B.S. y A. C. Gaur (1984): “Rice response to inoculation with N-fixing and P-solubilizing microorganisms”. Plant and Soil, 79: 227-234.
Kuykendall, L.D. y F.B. Dazzo (2003): “Genus Allorhizobium. En Bergey´s Manual of Determinative Bacteriology”, vol. 2, 124-127.
Ladha, J.K. (1997): “Biofertilizers: The way ahead?”. En Biological Nitrogen Fixation: The Global Challenge and Future Needs, Rockefeller Center, pp. 67-69.
Ladha, J.K. y P.M. Reddy (2003): “Nitrogen fixation in rice systems: state of knowledge and future prospects”. Plant and Soil, 252: 151-167.
Lakshmi-Kumari, M., K. Vijayalaksmi y N. S. Subba Rao (1975): “Interaction between Azotobacter sp and fungi. I. In vitro studies with Fusarium moniliforme”. Phytopath. Z, 75: 27-30.
Lambrecht, M., Y. Okón, A. Vanderbrock y J. Vanderleyden (2000): “Indole-3-acetic acid. A reciprocal signaling molecule in bacteria-plant interactions”. Trends Micro-biol., 8: 298-309.
Lamm, R.B. y C.B. Neyra (1981): “Characterization and cysts production of azospirilla isolated from selected grasses growing in New Jersey and New York”. Can. J. Mi-crobiol., 27: 1320-1325.
Ledgard, S.F. y K.W. Steele (1992): “Biological nitrogen fixation in mixed legume/grass pastures”. Plant and Soil, 141: 137-153.
Lehri, L.K. y C.L. Mehrotra (1968): “Use of bacterial fertilizer in crop production in U.H. Pradesh”. Curr. Sci., 37: 494-495.
Lehri, L.K. y C.L. Mehrotra (1972): “Effect of Azotobacter inoculation on the yield of vegetable crops”. Indian J. Agric. Res., 9 (3): 201-204.

154 bibliografía
Leigh, G.H. (2002): Nitrogen fixation at the millenium. Elsevier Science, Londres, 354 pp.
Levanoni, H., Y. Bashen, D. Romand y E. Klein (1998): “Ultrastructural localization and identification of Azospirillum brasilense Cd on an wheat root by Inmuno-gold labelling”. Plant and Soil, 117: 207-218.
Levins, R. (1973): “Fundamental and applied research in agriculture”. Science, 181: 523-524.
Lopes, E.S. (1978): “Ecology of legume-Rhizobium symbiosis”. En Limitations and Po-tentials for Biological Nitrogen Fixation in the Tropics, Plenum Press Ed., Nueva York, pp. 178-191.
Lynch,J.M. (1990): The Rhizosphere. John Wiley and sons Ed., Nueva York, 521 pp.
Ma, Q., N. Longnecker, R.J.W. Emery y C.A. Atkins (1998): “Growth and yield of Lupinus angustifolius are depressed by early transient nitrogen deficiency”. Austr. J. Agric. Res., 49: 811-819.
Madigan, M.T., y J.M. Martinko (1997): Brock Biology of Microorganisms. Prentice Hall, New Jersey, 860 pp.
Martínez Cruz, A., T. Bach, A. Delgado y V. Martínez (1993): Fosforina, un nuevo biopre-parado para la Agricultura. VII Forum de Ciencia y Técnica, La Habana, 68 pp.
Martínez-Romero, E. (1997): “Matching rhizobia to host plants and the environment”. En Biological Nitrogen Fixation: The Global Challenge and Future Needs, Rockefeller Center, pp. 67-69.
Martínez Viera R. (1986): Ciclo Biológico del Nitrógeno en el Suelo. Ed. Cientifico-Téc-nica, La Habana, 135 pp.
Martínez Viera R.(1998): “Los biofertilizantes como factores de economía y produc-tividad en la Agricultura Tropical”. En Curso-Taller sobre Agricultura Sostenible en el Trópico, La Habana, pp. 26-41.
Martínez Viera, R. (2003): Beneficios de la aplicación de los biofertilizantes para la Agri-cultura Sostenible. Centro de Desarrollo Agropecuario del FIRA, México, 30 pp.

155la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Martínez Viera, R. (2006): Los biofertilizantes y bioestimuladores bacterianos como pilares básicos de la Agroecología. Ed. CIARA, Caracas, 35 pp.
Martinez Viera, R. y B. Dibut (1996): “Los biofertilizantes como pilares básicos de la Agricultura Sostenible”. En Curso-Taller Gestión Medioambiental de Desarrollo Rural, INIFAT, La Habana, pp. 63-81.
Martínez Viera, R., y B. Dibut (2006): “Practical applications of Bacterial Biofertilizers and Biostimulators”. En Biological Approachs to Sustainable Soil Systems, Francis and Taylor Publ., Nueva York, pp. 467-477.
Martínez Viera, R., J. Szegi y L. Felipe (1968): Estudio microbiológico de los suelos de Isla de Pinos. Academia de Ciencias de Cuba, La Habana, 46 pp.
Martínez Viera, R., B. Dibut, I. Casanova y M. Ortega (1997): “Acción estimuladora de “Acción estimuladora de “Azotobacter chroococcum sobre el cultivo del tomate en suelo Ferralítico Rojo . Efecto sobre el semillero”. Agrotecnia de Cuba, 27(1): 23-30.
Martínez Viera, R., N. Toledo y C. Argûelles (1999): Introducción al conocimiento de los biofertilizantes . Universidad Tecnológica de la Huasteca Hidalguense, México, 43 pp.
Martínez Viera, R., B. Dibut, G. Tejeda, R. García (2001): “Trascendencia internacional de los biofertilizantes cubanos”. En: Memorias del XV Congreso Latinoamericano de la Ciencia del Suelo, Varadero. CD.
Martínez Viera, R., B. Dibut, R. García y G. Tejeda (2003): Informe Final del Proyecto “Obtención de un biofertilizante multipropósito para uso de la Agricultura Urbana”, Ministerio de la Agricultura, La Habana, 75 pp.
Martínez Viera, R., B. Dibut, G. Tejeda y R. García (2004): “Reducción de la fertilización nitrogenada en distintos cultivos económicos mediante la aplicación de biofertili-zantes”. En: Memorias del Congreso “Trópico 2004”, La Habana, CD.
Matsuo Y. y E. Takahashi (1977): “Statistical análisis of Soil Science data. II Quantitative estimate of natural supply of nitrogen in paddy fields over Japan”. Abst. Soil Sci. Soil Manure Meeting, Tokyo, pp. 88.

156 bibliografía
Mehrotra, C.C. y C.K. Lehri (1971): “Effect of Azotobacter inoculation on crop yields”. J. Indian Soc. Soil Sci., 191 (3): 243-248.
Merckx R, J.H. van Ginkell, J. Sinnaeve y A. Cremens (1993): “Plant induce changes in the rhyzosphere of maizes and wheat”. Plant and Soil, 96: 85-93.
Merrill, S.D., D.L. Tanaka y J. D. Hansen (2002): “Root length growth of eight crops species in Haplustoll soils”. Soil Sci. Soc. of Am. J., 66: 913-923.
Merschner H. (1999): Mineral Nutrition of Higher Plants, Academic Press, Londres, 889 pp.
Metting F.B. (1993): Soil Microbial Ecology. Application in Agricultural and Environmental Management. Marcel Dekker Ink, Nueva York, 450 pp.
Mishustin E.N. y E.K. Silnikova (1971): Biological fixation of atmospheric nitrogen. Mc. Millan Ed. Londres 675 pp.
Moens, S. (1996): “Genetic and biochemical analysis of the flagellation of Azospirillum brasilense”. Dissertations de Agriculture, 1: 5-8.
Montecinos, C. (2003): Manejo biológico del fósforo en el suelo. Sitio Internet Agroeco-logía y Desarrollo, 4 pp.
Moreno, J., J. González-López y G.R. Vela (1986): “Survival of Azotobacter spp in dry soils”. Appl. Environm. Microbiol., 51: 123-125.
Morot-Gaudry, J.F. (2001): Nitrogen assimilation by plants. Physiological, biochemical and molecular aspects. Science Publisher Inc.,Plymouth, 270 pp.
Mostafa, M.A. y M.Z. Mahmoud (1951): “Bacterial isolates from root nodules of Zygo-phyllaceae”. Nature, 167: 446-447.
Muñoz-Rojas, J. y J. Caballero-Mellado (2003): Gluconacetobacter diazotrophicus, mo-delo de bacteria endófita. En http//biblioweb.dgsca.unam.mx/Libros/Microbios/cap. 10.

157la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Muthukumarasamy, R., G. Rebathi y C. Lakshminarasimham (1999): “Diazotrophic associations in sugar cane cultivation in South India”. Tropical Agriculture, 76 (3): 171-178.
Myers, R.J. e I.M. Wood (1987): “Food legumes in the nytrogen cycle of farming sys-tems”. En Food Legume Improvement for Asian Farming Systems, ACIAR Proceeding N° 18, Camberra, pp. 34-45.
Narula, N., V. Kumar, R. Behl, A. Gransea y W. Merbach (2000): “Effect of P-solubili-zing Azotobacter chroococcum on N,P,K uptake in P-responsive wheat genotypes grown under greenhouse conditions”. J. Plant Nutr. Soil Sci., 163: 393-398.
Naumova, A.N., E.N. Mishustin y V.G. Marenko (1962): “Nature of the action of bac-terial fertilizers”. Izv. Akad. Nauk SSSR, Ser. Biol., 5: 709-717.
New, P.B., H.I. Sukiman e I. R. Kennedy (1991): “Host-plant bacterial strain specificity in Azospirillum”. En Proceed. IX Austr, Nitrogen Fixation Conf., Sydney, pp. 76-77.
Newman E.I. (1985): “The rhyzosphere: carbon sources and microbial populations”. En Ecological Interactions in Soil: Plants, Microbes and Animals, Blackwell Scientific Publications, Oxford, pp 107-121.
Neyra, C.A. (1978): “Interactions of plant photosyntesis with dinitrogen fixation and nitrate assimilation”. En Limitations and Potentials for Biological Nitrogen Fixation in the Tropics, Plenum Press Ed., Nueva York, pp. 111-121.
Nieto, K.F. y W.T. Frankenberger (1990): “Microbial production of cytokinins”. En Soil Biochemistry, vol. 6, Marcel Dekker Ed., Nueva York, pp. 195-248.
Norris, D.O. (1972): “Leguminous plants in tropical pastures”. Tropical Grasslands, 6: 159-165.
Ocampo, J.A., J.M. Barea y E.Montoya (1975): “Interaction between Azotobacter and phosphobacteria and their stablishment in the rizosphere is affected by soil fertili-ty”. Canadian J. of Microbiology, 21 (8): 1160-1165.
Odum,L..P. (1984): “Properties of Agroecosystems”. En Agricultural Ecosystems, J. Wiley and sons Ed., New York.

158 bibliografía
Odum, L.P. (1989): “Input management of production systems”. Science, 243: 177-182.
Okon, Y. y J. Vanderleyden (1997): “Root associated Azospirillum species can stimulate plants”. ASM News, 63 (7): 364-370.
Okon,Y., Y. Kapulnik y S. Varig (1993): “Field inoculation studies with Azospirillum in Israel”. En Biological Nitrogen Fixation. Oxford and IBH Publishing, Nueva Delhi, pp. 175-196.
Olivares, F.L., E.K. James, J.I. Baldani y J. Dôbereiner (1997): “Injection of mottled stripe disease in susceptible and resistent sugar-cane varieties by the endophytic diazotroph Herbaspirillum”. New Phytol., 135: 723-737.
Olivares, J. (2006): Fijación biológica de Nitrógeno. http//w.w.w.eez.csic.es-olivares/ciencia/fijación.
Ostwald, K.P. y V.P. Bhide (1977): “Solubilization of tricalcium phosphate by soil Pseu-domonas”. Indian J. Exptl. Biol., 10: 393-398.
Paau, A.S. (1988): “Formulations useful in applying beneficial microorganims to seeds”. Trends Biotechnology, 6: 276-279.
Pacios, C., H.P. Spaink y N. Sturma (2001): “Structure and function of Nod-factors”. En Prokaryotic Nitrogen Fixation: A Model System for Analysis of a Biological Process. Horizon Scientific Press, Londres, pp. 365-383.
Pacovsky, R.S. (1986): “Micronutrient uptake and distribution in mycorhizal or phos-phorus-fertilized soybeans”. Plant and Soil, 95 (3): 379-388.
Palazzo, D.G., G. Capotorti, F. Montemurro y F. Sunser (1997): “Riposte productive di culture erbacee all inoculo con Azospirillo”. L´ Informatore Agrario, 53 (12): 53-55.
Palm Ch., M. Swift e I. Barois (2001): “Un enfoque integrado para el manejo biológico de los suelos”. En: Memorias del XV Congreso Latinoamericano de la Ciencia del Suelo, Varadero. CD.

159la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Parr,J.F., S.B. Hornick y D.D. Kaufman (1994): Use of microbial inoculants and organic fertilizers in agricultural production. Food and Fertilizer Technology Center, Taipei, Taiwan.
Pate, J.S. (1977): “Functional biology of dinitrogen fixation by legumes”. En A Treatise on Dinitrogen Fixation. John Wiley and sons Ed., Nueva York, pp. 220-232.
Pate, J.S., D.B. Layzell y C.A. Atkins (1979): “Economy of carbon and nitrogen in a nodulated and non-nodulated legume”. Plant Physiol., 64: 1083-1088.
Paul, E:A. (1988): “Towards the year 2000: directions for future nitrogen research”. En Advances in Nitrogen Cycling in Agricultural Ecosystems, CAB International, Londres, pp. 417-425.
Paul, E.A. y F.E. Clark (1992): “Phosphorus transformation in soil”. En Microbiology and Biochemistry. Marcel Dekker Inc., Nueva York, pp. 235-267.
Paul, E., J. Fages, P. Blanc, G. Goma y A. Pareilleux (1993): “Survival of alginated–entra-pped cells of Azospirillum lipoferum during dehydratation and storage in relation to water properties”. Appl. Microbiol. Biotechnol. , 40: 34-39.
Paula, M.A. de, J. Dôbereiner y J.O. Siquiera (1993): “Ocorrência de fungos micorri-zicos vesiculo-arbusculares e de bactérias diazotróficas na cultura de batata doce”. Revista Brasileira de Ciência do Solo, 17: 349-356.
Pawlovsky, K. y T. Bisseling (1996): “Rhizobial and actinorhizal symbioses: What are the shared features? ”. Plant Cell, 8: 1899-1913.
Peña Cabriales, J.J. y M. Alexander (1979): “Effect of moisture on the survival of Rhizo-bium in soil”. J. of Soil Sci. Soc. of America, 43: 925-930.
Peoples M.B. y E.T. Craswell (1992): “Biological Nitrogen Fixation: Investments, expec-tations and actual contributions to Agriculture”. Plant and Soil, 141: 13-39.
Peoples, M.B., D. Herridge y J.K. Ladha (1995): “Biological nitrogen fixation: an efficient source of nitrogen for sustainable agricultural production? ”. Plant and Soil, 174: 3-28.

160 bibliografía
Pérez, A., C.R. Pérez y G. Cardona (2005): “Hongos solubilizadores de fosfatos en suelos arroceros de La Mojana, Sucre (Colombia) ”. Revista Arroz, 53 (458): 355-345.
Perret, X., C. Staehelin y W.J. Broughton (2000): “Molecular basis of symbiotic promis-cuity”. Microbiol Mol. Biol. Rev., 64: 180-201.
Piceno, Y.M. y C.R. Lowell (2000): “Stability in natural bacterial communities. I. Nu-trients addition effects on rhizosphere diazotroph assemblage composition”. Mi-crob. Ecology, 39 (1): 30-40.
Pingali, P.L., P.F. Moya y L.E. Velasco (1992): The post-green revolution blues in asian rice production. IRRI Social Science Division, paper N° 90-01.
Pinton, R., Z. Varanini y P. Nannipieri (2001): The Rizhosphere: Biochemistry and Chemi-cal Substances at the Soil-Plant Interface. Marccel Dekker Ed., Nueva York.
Postgate J. R.(1987): Nitrogen Fixation. Postgate J. R.(1987): Nitrogen Fixation. Postgate J. R.(1987 Edward Arnold Ed., Londres, 265 pp.
Postgate, J. (1990): “Fixing the nitrogen fixers”. New Scientist, 36: 57-61.
Pueppke, S.G. y W.J. Broughton (1999): “Rhizobium sp NGR 234 and R. fredii USDA257 share exceptionally broad, nested host-ranges”. Mol. Plant-Microbe In-teract., 12: 293-318.
Purcino, A.A.C., C. Lurlarp y J.C. Lind (1986): “Mycorhiza and soil fertility effects with growth, nodulation and nitrogen fixation on Leucaena growth on a typic eutrus-tox”. Commun. in Soil Sci. Plant Anal., 17 (5): 473-489.
Ramírez Gama, R.M. y B. Luna Millán (1995): “Simbiosis asociativas”. En Agromicrobio-logia, elemento útil en la Agricultura Sustentable. Ed. Colegio de Postgraduados en Ciencias Agrícolas, Ciudad México, pp. 124-143.
Requena, N., I. Jimenez, M. Toro y J.M. Barea (1997): “Interactions between plant-growth promoting rhizobacteria (PGPR), arbuscular mycorrhizal fungi and Rhizo-bium spp”. New Phytol., 136: 667-677.

161la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Reis, V.M., F.L. Olivares y J. Dôbereiner (1994): “Improved methodology for isolation of Azotobacter diazotrophicus and confirmation of its endophytic habitat”. World J. of Microbiol. and Biotech., 10: 401-405.
Reis, F.B., V.M. Reis, S. Urquiaga y J. Dôbereiner (2000): “Influence of nitrogen fertiliza-tion on the populations of diazotrophic Herbaspirillum sp and G. diazotrophicus in sugar cane”. Plant and Soil, 219: 153-159.
Riggs, P.J., M.K. Chelius, S.M. Kaepler y E.W. Triplett (2001): “Enhanced maize pro-ductivity by inoculation with diazotrophic bacterias”. Austr. J. Plant Physiol., 28 (9): 829-836.
Rivera, R., F. Fernández, C. Sánchez, C. Bustamante y M. Ochoa (1997): “Efecto de la inoculación con hongos micorrizógenos VA y bacterias rizosféricas sobre el creci-miento de posturas de cafeto”. Cultivos Tropicales, 18(3): 15-23.
Rodelas, M.B. (2001): “Interacción Rhizobium-Azospirillum y Rhizobium-Azotobacter. Efecto sobre la nodulación y la fijación simbiótica de dinitrógeno en Vicia faba”. En (http://193.146.205.198/sefin/Ecología/Rodelas.html)l)l
Rodelas, M.B., J. González, M.V. Martínez y V. Salmerón (1999): “Influence of Rhizo-bium-Azospirillum and Rhizobium-Azotobacter combined inoculation on mineral composition of faba bean”. En (http://link.springer-ny.com/link/service/journals)s)s
Rodriguez, H. y R. Fraga (1999): “Phosphate solubilizing bacteria and their role in plant growth promotion”. Biotechnology Advances, 17: 319-339.
Römheld, V. y G. Neumann (2006): “The Rizosphere: Contributions of the Soil-Root Interface to Sustainable Soil Systems”. En Biological Approaches to Sustainable Soil Systems, pp. 91-109, Taylor and Francis Ed., Boca Ratón, USA.
Roper, M.M. (1983): “Field measurements of nitrogenase activity in soils amended with wheat straw”. Austr. J. Agric. Res., 34: 725-731.
Roper, M.M. (1984): “Straw decomposition and nitrogenase activity: effects of soil moisture and temperature”. Soil Biol. Biochem., 17: 65-72.
Rovira, A.D. (1969): “Plant root exudates”. Bot. Rev., 35: 35-58.

162 bibliografía
Rovira A. D. (1965): Interaction between plant roots and soil microorganisms”. Ann. Rev. Microbiol., 19: 241-266.
Ruinen, J., (1974): “Foliar associations in higher plants”. En The Biology of Nitrogen Fixa-tion. North -Holland Ed., La Haya, pp. 210-246.
Sanginga, R.(2003): “Role of biological nitrogen fixation in legume based cropping sys-tems”. Plant and Soil, 252: 25-39.
Sanjuan, J., S.A: Herrera-Cervera, A.I. Rodriguez-Alonso y J. Olivares (2004): Trans-formación genética en el suelo y biodiversidad (http://nostoc.usal.es/sefin/sanjuan.html).
Sarig, G., Y. Okon y A. Blue (1990): “Promotion of leaf area development and field in Sorghum bicolor inoculated with Azospirillum brasilense”. Symbiosis, 9: 235-245.
Sarro da Silva, M.F. y J. Dôbereiner (1977): “Occurrence of Azospirillum sp in soils and roots”. En Proceed. of Int. Symposium on Biological Fixation in the Tropics, Brasilia, pp. 136-142.
Sasson, A., (2000): “La contribución de las tecnologías a la alimentación”. BiotecnologíBiotecnologíBiotecnolog a ía íAplicada, 17 (1): 2-6.
Shiga, H. (1971): “41 years three elements experiments for rice”. Hort. and Agric., 26: 523-525.
Shing, T. (1977): Studies on interaction between Azotobacter chroococcum and some plant pathogens. IARI Ph. D. Thesis, Nueva Dehli, 115 pp.
Singh, S.C. y N.S. Subba Rao (1979): “Associative effect of Azospirillum brasiliense and Rhizobium japonicum on nodulation and yield of soybean”. Plant and Soil, 53: 387-392.
Smithson, P.C. y P.A. Sánchez (2001): “Plant nutricional problems in marginal soils of developing countries”. En Plant Nutrient Acquisition and Perspectives. Springler-Ver-lag, Tokyo, pp. 32-68.

163la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Sparvoli, F., C. Lanave, R. Bollini y L. Lioi (2001): “Lectin and lectin-related proteins in Lima bean seeds: biochemical and evolutionary studies”. Plant Mol. Biol.,45: 587-597.
Steciow, M. (2003): Rizosfera. En w.w.w.cricyt.edu.ar
Subba Rao, N.S. (1990): Biofertilizers in Agriculture. Oxford and IBH Publishing Co., New Dehli, 186 pp.
Subba Rao, N.S. (1996): “Interaction of nitrogen-fixing microorganisms with other soil microorganisms”. En: Biological Nitrogen Fixation, Ed. Marcel Dekker, Nueva York, pp. 37-63.
Tarafdar J. C., Claassen, N. (1988): “Organic phosphorus compounds as a phosphorus source for higher plants through the avtivity of phosphatase produce by plants roots and microorganisms”. Biology and Fertility of Soils, 5, 308-312.
Tarafdar J.C., Marschner, H. (1994): “Phosphatase activity in the rhizosphere and hyphosphere of VA mycorrhizal wheat supplied with inorganic and organic phos-phorus”. Soil Biology and Biochemistry, 26, 387-395.
Tarafdar J. C., Rao A.V. Bala K. (1988): “Production of phosphatase by fungi isolated from desert soils”. Folia Microbiologia, 33, 453-457.
Tan, Z., T. Hurek y B. Reinhold-Hurek (2003): “Effect of N-fertilization, plant genotypes and environmental conditions on nifH gene pools in roots of rice”. Environ. Micro-biol., 5: 1009-1015.
Tao Wang, E. y J. Martínez Romero (2003): “Taxonomía de Rhizobium”. En (http// biblioweb.dgsca.unam.mx/Libros/Microbios/cap. 8).
Tapia-Hernández, A., M. R. Bustillos, T. Jiménez, J. Caballero-Mellado y L.E. Fuentes (2000): “Natural endophytic occurrence of Acetobacter diazotrophicus in pine-apple plants”, Microbial Ecology, 39 (1): 49-55.
Tchan, Y. T. (1984): “Family II Azotobacteriaceae Pribram 1933”. En Bergey´s Manual of Determinative Bacteriology.of Determinative Bacteriology.of Determinative Bacteriology

164 bibliografía
Tchan, Y.T. (1989): “Some aspects of non-rhizobial diazotrophs: their past and their future”. En Microbiology in Action. Research Studies Press Ltd., Camberra, pp. 193-207.
Tchan, Y.T. e I.R. Kennedy (1987): “Asymbiotic nitrogen fixation”. En Nitrogen Cycling in Agriculture Systems of Temperate Australia. Soil Science Society of Australia Press, Sydney, pp. 243-262.
Tejeda, G., J. Rodríguez, R. García, R. Martínez Viera (2002): “Obtención de un bio-preparado con efecto antagonista y estimulador del crecimiento vegetal, mediante el cocultivo de Bacillus subtilis y Azotobacter chroococcum”. En Resúmenes del XII Congreso del INCA, La Habana, pp. 38.
Tilak, K.V.B.R., C.S. Singh, N.K. Roy y N.S. Subba Rao (1981) : “A. brasilense and A. “A. brasilense and A. “chroococcum: effect on yield of maize and sorghum. Soil Biology and Biochemistry, 14: 417-418.
Torres, R., (2000): “Incrementos de la fijación biológica de N mediante la inoculación combinada de bacterias fijadoras. En http://www.ilustrados. com/publicaciones.
Torrey, J.G. y D. Callaham (1979): Early nodule development in Myrica gale. Bot. Gaz., 140: 10-14.
Trinick, M.J. (1988): “Bradyrhizobium of the non-legume Parasponia. En Microbiology in Action, Research Studies Press/Wiley, Londres, pp. 107-118.
Triplett, E.W. (2000): “Prokaryotic Nitrogen Fixation. Horizon Scientific Press, Norfolk, 340 pp.
Trolldenier, G. (1977): “Influence of some environment on rhizosphere of rice. Plant and Soil, 47: 203-217.
Unkovich, M.J. y J. S. Pate (2000): An appraisal of recent field measurements of sym-biotic N2 fixation by annual legumes. Field Crop Res., 65: 211-228.
Urquiaga S. & Zapata F. 2000. Manejo eficiente de la fertilización nitrogenada de cultivos anuales en América Latina y el Caribe. Porto Alegre: Ed. Gênesis. 110 pp.

165la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Urquiaga, S., A.S. de Resende, B.J. Alves y R. Boddey (2001): “Potencial de fijación bio-lógica de nitrógeno en la productividad de sistemas agrícolas de América Latina. En:Memorias del XV Congreso Latinoamericano de la Ciencia del Suelo, Varadero. CD.
Valdés, M., N.O. Pérez y L. Vázquez (2003): La bacteria filamentosa Frankia. (http//biblioweb.dgsca.unam.mx/Libros/Microbios/cap.11).
Vanlawa, B., K. Aihou; S. Aman y S. Deekers (2000): Agronomic evaluation of interac-tions between organic and inorganic inputs of N in a multinational trial in the moist savanna of West-Africa. IITA Annual Report.
Vose, P.B., A.P. Ruschel, R.L. Victoria, S.M. Tsai Saito y E. Matsui (1982): “15-N as a tool in biological nitrogen fixation research”. En Biological nitrogen fixation technology for tropical agriculture, CIAT, Cali.
Watson, C.A., J.M. Ross, G. F. Minotta y F. Roffi (2000): “Environment-induced modifi-cations to root longevity in Lolium perenne and Trifolium repens”. Annals of Botany, 85: 397-401.
Weber,G. (1996): “Legume-based technologies for Africa savannas: Challenges for re-search and development”. Biol. Agric. Hort., 13: 309-333.
Wilcox, G.E. (1996): “Tomato”. En Nutrient Deficiences and Toxicities in Crop Plants. APS Press, Nueva York, pp. 137-141.
Wood, I.M. y R.J.K. Myers (1987): “Food legumes in farming systems in the tropics and subtropics”. En Food Legumes Improvement for Asian Farming Systems, ACIAR Proceeding N° 18, Camberra, pp. 34-45.
Wullstein L. H., M. L. Bruening y W. B. Bollen (1979): “Nitrogen fixation associated with sand grain root sheaths of certain xeric grasses”. Physiol. Plant., 46: 1-4.
Yanni, Y.G.(2001): “The beneficial plant growth promoting association of Rhizobium leguminosarum bv. trifolii with rice roots”. Aust. J. Plant Physiol., 28: 845-870.
Yao, Z.J., F.I. Khan, E.T. Wang y G.H. Wei (2002): “Characterizationof rhizobia that no-dulate legume species of the genus Lespedeza and description of Bradyrhizobium yuanmingense sp nov”. Int. J. Syst. Evol. Microbiol., 52: 2219-2230.

166 bibliografía
Yoshida, T. y G. Rinaudo (1982): “Heterotrophic N2 fixation in paddy soils”. En Micro-biology of tropical soils and plant productivity. Martinus Nijhoff y W. Junk Publ., La Haya, pp. 75-107.
Young, C.C. (1990): “Effect of phosphorus solubilizing bacteria and vesicular arbuscular mycorrhizal fungi on the growth of three species in subtropical-tropical soils”. Soil Sci. and Plant Nutrition, 36 (2): 225-231.
Zapata, F. y A.N. Roy (2004): Use of phosphate rocks for sustainable agriculture, FAO, Roma , 140 pp.
Zuberer D. A. (1990): “Soil rhizosphere aspects of N2-fixing plant-microbe associatio-ns”. En The Rhizosphere, John Wiley and sons Ed., Nueva York, pp. 317-352.
Zuberer, D.A. y M. Roth (1982): “In vitro inhibition of non symbiotic nitrogen-fixing bacteria by rizosphere actinomycetes associated with grass”. Can. J. Microbiol., 29: 705-709.

167la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Índice de tablas
Tabla 1 Superficie agrícola dedicada a cultivos orgánicos en distintospaíses europeos en el año 2001 20
Tabla 2 Efecto quimiotáxico de los exudados totales de las plantas decebolla sobre distintas cepas de A. chroococcum(expresado en logn de X). 33
Tabla 3 Respuesta del trigoa la inoculación con A. chroococcumenlas condicionesde Cuba 35
Tabla 4 Poblaciones naturales de bacterias pertenecientes al géneroAzotobacter en las muestras de suelos rizosféricos recogidasAzotobacter en las muestras de suelos rizosféricos recogidasAzotobacteren la Huasteca Hidalguense (México) 36
Tabla 5 Estimados de Nitrógeno aportado por diferentes sistemasfijadores en la agricultura 44
Tabla 6 Distribución de sistemas fijadores de Nitrógeno 47
Tabla 7 Biofertilizantes fijadores de N2, limitaciones y uso por los agricultores 50
Tabla 8 Aporte estimado de nitrógeno a los cultivos agrícolasen distintos países (expresado en millones de toneladas) 57
Tabla 9 Microorganismos productores de hormonas vegetales 62
Tabla 10 Auxinas sintetizadas por bacterias fijadoras de Nitrógeno 63
Tabla 11 Giberelinas sintetizadas por bacterias fijadoras de Nitrógeno 65

168 índice de tablas
Tabla 12 Citoquininas sintetizadas por bacterias fijadoras de Nitrógeno 66
Tabla 13 Producción de vitaminas por A. chrococcum cepa INIFAT-12 67
Tabla 14 Producción de aminoácidos por la cepa INIFAT-12 deAzotobacter chroococcum 68
Tabla 15 Producción de PGRs por A. chroococcum cepa INIFAT-12 68
Tabla 16 Porcentaje de plantas de maíz emergidas y tiempo de apariciónde la primera hoja verdadera en plantas inoculadas y sin inocularcon la cepa Guárico-1 de Azotobacter sAzotobacter sAzotobacter p. 69
Tabla 17 Estimados experimentales de la cantidad de N2 derivadode la fijación atmosférica por las más importantes leguminosasde grano y aceite 72
Tabla 18 Clasificación y descripción de las respuestas a la nodulación 79
Tabla 19 Principales factores que afectan la fijación simbiótica de Nitrógeno 82
Tabla 20 N de la planta derivado de la fijación (P) en distintos cultivos y países 87
Tabla 21 Clasificación de las dicotiledóneas no leguminosasfijadoras de N2 en simbiosis con actinomicetos del género Frankia 92
Tabla 22 Efectividad de la Azotobacterina preparada en turbaen experimentos realizados en Rusia. 101
Tabla 23 Efecto antagonista de Azotobacter y Azotobacter y Azotobacter Azospirillum frentea hongos fitopatógenos que atacan a las raíces de las plantas 104
Tabla 24 Resultados obtenidos en la India con la inoculación deAzotobacter chroococcum en distintos cultivos 105

169la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Tabla 25 Efecto de la aplicación de A.chroococcum sobre el rendimientode yuca y batata(Tm/ha). 109
Tabla 26 Distribución geográfica de Azospirillum en raíces y sueloscolectados en varias regiones del mundo. 111
Tabla 27 Resultados obtenidos en distintos países con la inoculaciónde Azospirillum brasiliense y A. lipoferum en diferentesespecies de gramíneas 113
Tabla 28 Efecto de la aplicación foliar de A. chroococcum sobreel rendimiento de toronja y naranja en las condiciones de Cuba. 118
Tabla 29 Efecto de la aplicación foliar de A. chroococcum sobre el rendimientodel mango en el Estado Sinaloa (México) 118
Tabla 30 Comparación de inoculantes a base de turba estéril y no estéril 123
Tabla 31 Especificaciones de control de calidaddel Buró de Standardsde la India para los biofertilizantes y bioestimuladores 125
Tabla 32 Efecto de la aplicación de inoculante mixto a base deA. chroococcum y B. megatherium var. phosphaticum sobre rosalesde seis años de edad 127
Tabla 33 Incremento del rendimiento de arroz producido por la aplicaciónde biofertilizante mixto a base de A. chroococcum y Az. brasilienseen 25 haciendas arroceras 130
Tabla 34 Efecto de la aplicación del biofertilizante mixto a base deA. chroococcum y Az. brasiliense sobre el rendimiento del arrozen condiciones de producción 130
Tabla 35 Acción antagonista de 3 cepas comerciales de Azotobacterchroococcum frente a otras 6 especies de bacterias concaracterísticas de biofertilizantes 132

170 índice de tablas

171la fijación biológica del nitrógeno atmosféricoen condiciones tropicales
Índice de figuras
Figura 1 Efecto estimulador e inhibidor de distintas cepas de Azotobacter sp.sobre el desarrollo de raices de tomate 64
Figura 2 Nódulos de frijol (Phaseolus vulgaris) 76
Figura 3 Nódulos de garbanzo (Cicer arietinum L.) 77
Figura 4 Nódulos de soya (Glycine max L.) 77
Figura 5 Colonias tipicas de Azotobacter spAzotobacter spA . en medio de Ashby 101
Figura 6 Tinciòn Gram negativa de células de A. chroococum 101
Figura 7 Efecto de la aplicación de A. chroococcum en semilleros de tomate 107
Figura 8 Efecto de la aplicación de A. chroococcum en semilleros de pimiento 107
Figura 9 Efecto de la aplicación de Azotobacter sobre el desarrolloAzotobacter sobre el desarrolloAzotobacterde los frutos de tomate 108
Figura 10 Efecto de la aplicación de A. chroococcum sobre los frutos depepino cv. Ashley 108
Figura 11 Efecto de la aplicación de A. chroococcum sobre semilleros de cebolla 109
Figura 12 Colonias de G. diazothrophicus en medio LGI-P 115
Figura 13 Efecto de la inoculación de distintas cepas de bacterias fijadorasde N sobre el desarrollo de plantulas de tomate 121

172 índice de figuras
Figura 14 Efecto de la aplicación de inoculante mixto, a base de A. chroococcumy B. megatherium var. phosphaticum, sobre la densidad y la velocidadde floración de un campo de rosas inoculado (derecha)en comparación con un campo vecino sin inocular (izquierda) 128
Figura 15 Efecto de la aplicación de inoculo mixto a base de A. chroococcum y B. megatherium var. phosphaticumy B. megatherium var. phosphaticumy sobre la producción de rosas 129