fixational eye movements across vertebrates: comparative...
TRANSCRIPT

Fixational eye movements across vertebratesComparative dynamics physiology and perception
Barrow Neurological Institute Phoenix AZ USASusana Martinez-Conde
Barrow Neurological Institute Phoenix AZ USAStephen L Macknik
During visual fixation human eyes are never still Instead they constantly produce involuntary ldquofixational eye movementsrdquoFixational eye movements overcome neural adaptation and prevent visual fading thus they are an important tool tounderstand how the brain makes the environment visible The last decade has seen a growing interest in the analysis offixational eye movements in humans and primates as well as in their perceptual and physiological consequencesHowever no comprehensive comparison of fixational eye movements across species has been offered Here we review fivedecades of fixational eye movement studies in non-human vertebrates and we discuss the existing evidence concerningtheir physiological and perceptual effects We also provide a table that summarizes the physical parameters of the differenttypes of fixational eye movements described in non-human vertebrates
Keywords fixation comparative physiology non-human primates mammals monkey cat rabbit birds reptiles turtleamphibians salamander fish evolution oculomotor microsaccades drifts tremor
Citation Martinez-Conde S amp Macknik S L (2008) Fixational eye movements across vertebrates Comparative dynamicsphysiology and perception Journal of Vision 8(14)28 1ndash16 httpjournalofvisionorg81428 doi10116781428
Introduction
When we fixate our gaze on an object of interest oureyes are never still Instead we constantly produce smallinvoluntary eye movements generally called ldquofixationaleye movementsrdquo If these eye movements are eliminatedour perception of stationary objects fades due to neuraladaptation (Ditchburn amp Ginsborg 1952 Martinez-Conde Macknik Troncoso amp Dyar 2006 Riggs ampRatliff 1952 Troncoso Macknik amp Martinez-Conde2008) When our eyes are free to move across the imageonce again visual perception reappears (Yarbus 1967)Due to their role in counteracting visual adaptationfixational eye movements are an important tool to under-stand how the brain makes the environment visible bothin normal and pathological vision (Martinez-Conde2006) Further because we fixate our gaze most of thetime during visual exploration (Martinez-Conde 2006Otero-Millan Troncoso Macknik Serrano-Pedraza ampMartinez-Conde 2008) fixational eye movements areoften responsible for driving our visual experienceFixational eye movements can help us understand the
underpinnings of visual awareness (Martinez-Conde ampMacknik 2007b) in a number of ways
1 Fixational eye movements drive the visibility (andcounteract the fading of) stationary objects duringfixation Thus fixational eye movements can helpconstrain the spatiotemporal characteristics of visi-ble stimuli Moreover the neural responses trig-gered by fixational eye movements along the visual
pathway must encompass the neural code forvisibility (Martinez-Conde 2006 Martinez-CondeMacknik amp Hubel 2000 2002)
2 Fixational microsaccades in human subjects maydrive perceptual alternations for a variety of multi-stable stimuli (Martinez-Conde 2006 Martinez-Conde amp Macknik 2007b Martinez-Conde et al2006 Troncoso Macknik amp Martinez-Conde2008 Troncoso Macknik Otero-Millan ampMartinez-Conde 2008 van Dam amp van Ee 2006)
3 Fixational microsaccades may indicate attentional orcognitive engagement (Engbert amp Kliegl 2003bGalfano Betta amp Turatto 2004 Hafed amp Clark2002 Martinez-Conde amp Macknik 2007b Otero-Millan et al 2008)
4 Human subjects are unaware of their fixational eyemovements (Martinez-Conde Macknik amp Hubel2004) That is despite the continuous motion causedby fixational eye movements the world remainsperceptually stable when we fixate This perceptualstability can be foiled the ldquovisual jitterrdquo illusionshows that in the absence of fixational eye move-ment compensation the world is seen as unstableand jittery (Murakami amp Cavanagh 1998) Thus theneuronal circuits populations etc that constitutethe neural correlates of visual awareness mustsustain perceptual stability during fixation
Research in fixational eye movements is one of thefastest moving fields of vision research today The lastdecade has seen a proliferation of fixational eye movementstudies including a few reviews of the published literature
Journal of Vision (2008) 8(14)28 1ndash16 httpjournalofvisionorg81428 1
doi 10 1167 8 14 28 Received February 7 2008 published December 24 2008 ISSN 1534-7362 ARVO
Typeofeye
move
ment
Amplitude
Frequency
intersacc
adic
interval
Duratio
nMax
speed
Mean
speed
Conjugate
Study
Mammals
Primate
Microsaccade
84ndash162
min
(meansfor
2monke
ys)
23ndash25
Hz(m
ean
frequenciesfor
2monke
ys)
atleast
8mse
cndash
ndashndash
Horw
itzand
Albrig
ht(2003)
Microsaccade
~40min
(mean)
ndashndash
ndash~30degsec
ndashSnodderly
etal(2001)
Microsaccade
~20min
(mean)
~3ndash5Hz
29mse
c(m
ean)
ndash~30degsec
ndashMartinez-Conde
etal(2000)
Microsaccade
48min
(mean)
03ndash14
Hz
(meansfor
2monke
ys)
25ms(m
ean)
9ndash11
0degsec
(median
40degsec)
~30degsec
ndashBairandOrsquoKeefe
(1998)
Microsaccade
101
min
(median)
0597se
c(m
edian)
20mse
c(m
ean)
ndashndash
ndashLeopold
and
Logothetis
(1998)
Microsaccade
99ndash403
min
(mediansfor
4monke
ys)
08ndash74
sec
(mediansfor
4monke
ys)
ndashndash
ndashndash
Ska
vensk
ietal(1975)
Microsaccade
40min
(mean
minim
um
amplitude
23min)
ndashndash
ndashndash
ndashSteinmanetal(1973)
Drift
ndashndash
ndashndash
6minsec
(meanfor
1monke
y)
ndashBairandOrsquoKeefe
(1998)
Drift
ndashndash
ndashndash
042ndash
1191minsec
(meansfor
4monke
ys)a
ndashSka
vensk
ietal(1975)
Cat
Microsaccade
ldquosomewhat
smallerrdquo
thanin
humans
ldquofarfewerrdquothan
inhumans
ndashndash
ndashNo(m
ostly
monocu
lar
andso
me
binocu
lar)
Hebbard
andMarg
(1960)
Microsaccade
35min
bndash
ndashndash
ndashndash
Pritch
ard
and
Heron(1960)
Drift
ndashndash
ndashndash
147
minsec
(meanfor
3ca
ts)
ndashWintersonand
Robinso
n(1975)
Drift
ndashndash
ndashndash
ndashndash
Hebbard
and
Marg
(1960)
Drift
ldquoUsu
ally
~25min
butca
nexc
eed
2degrdquob
ndashndash
ndash30minsec
ndashPritch
ard
and
Heron(1960)
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 2
Typeofeye
move
ment
Amplitude
Frequency
intersacc
adic
interval
Duratio
nMax
speed
Mean
speed
Conjugate
Study
Tremor
56ndash733
s(ave
rage31s)
b35ndash
65Hz
(ave
rage50Hz)
ndashndash
ndashNo
Hebbard
and
Marg
(1960)
Tremor
04
min
b2ndash
40Hz
(unable
toreco
rdfasterrates)
ndashndash
ndashndash
Pritch
ard
and
Heron(1960)
Rabbit
Drift
ndashndash
ndashndash
25ndash
50minsec
ndashVanderSteenand
Collewijn
(1984)
Drift
e4degb
ndashndash
Q1degsec
(unable
toreco
rdfasterrates)
15
minsec
ndashCollewijn
and
vanderMark
(1972)
Tremor
ldquosmallamplituderdquo
ndashndash
ndashndash
ndashCollewijn
and
vanderMark
(1972)
Birds
Pigeon
Microsaccade
ndashldquovery
lowrdquo
G20mse
cndash
ndashndash
Nye
(1969)
Impulse
Upto
2deg
ndashndash
ndashndash
ndashNye
(1969)
Drift
3ndash5deg
(occ
asionally
exc
eedingthe
limitofthereco
rding
system75
deg)
ndashndash
ndash1ndash
5degsec
ndashNye
(1969)
Oscillation
Upto
seve
rald
eg
(peak-to-peak)
Shortbursts
with
afrequency
of
28ndash35Hz
usu
ally
separatedby
intervals
of1ndash
2se
c
Upto
08
sec
ndashndash
ndashNye
(1969)
Owl
Microsaccade
e1deg
ndashndash
10degsec
ndashndash
Steinbach
and
Money(1973)
Drift
e1deg
ndashndash
ndashndash
ndashSteinbach
and
Money(1973)
Tremor
1min
20Hz
ndashndash
ndashndash
Steinbach
and
Money(1973)
Oscillation
15
deg
Shortbursts
with
afrequency
of20Hz
about
4tim
esaminute
025se
cndash
ndashndash
Steinbach
and
Money(1973)
Reptiles
Turtle
Drift-like
ndashndash
ndashndash
48
T2minsec
ndashGresc
hneretal(2002)
Periodic
Upto
5min
5Hz
ndashUpto
214
degsec
ndashndash
Gresc
hneretal(2002)
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 3
(Engbert 2006 Martinez-Conde 2006 Martinez-Condeet al 2004) However the differences and similarities infixational eye movements across species as well as theirpotential significance have been neglected so far Despitewide-reaching interest in this topic (Martinez-Conde ampMacknik 2007a) a comprehensive comparative reviewhas not been published Here we review the comparativedynamics the physiology and the perceptual consequen-ces of fixational eye movements across all species studiedto date To the best of our knowledge our review includesall Medline-indexed studies (as well as several bookchapters and other non-indexed publications) of fixationaleye movements in non-human vertebrates These span48 years of research from the first studies in the catpublished in 1960 (Hebbard amp Marg 1960 Pritchardamp Heron 1960) to the most recent studies in thesalamander and archer fish published in 2007 and 2008(Baccus Olveczky Manu amp Meister 2008 OlveczkyBaccus amp Meister 2007 Segev Schneidman Goodhouseamp Berry 2007) As a result of our analysis we havegenerated a table (Table 1) that summarizes the physicalparameters of the different types of fixational eye move-ment acrossnon-humanvertebratesWewill relatewheneverpossible fixational eye movements in non-humanvertebrates to human fixational eye movements
Fixational eye movements inmammals
Fixational eye movements in the primate
Fixational eye movements were originally discoveredand characterized in human subjects (Ditchburn ampGinsborg 1952 1953 Ratliff amp Riggs 1950) Humanfixational eye movements comprise three main typesmicrosaccades (called ldquoflicksrdquo in early studies) drift andtremor (Figure 1) Microsaccades are the fastest andlargest of the three types of fixational eye movements(Supplementary Video 1) They travel in a straighttrajectory carrying the retinal image across a range ofseveral dozen to several hundred photoreceptor widthsDrifts are slow curvy motions that occur between micro-saccades and tremor is a very fast (Egrave90 Hz) andextremely small oscillation (about the diameter of a fovealcone) superimposed on drifts (see Tables 1ndash3 of Martinez-Conde et al 2004 for a review of fixational eye move-ment parameters in humans)Fixational eye movements in old-world monkeys are
very similar to human fixational eye movements (Martinez-Conde 2006 Martinez-Conde et al 2000 2002Skavenski Robinson Steinman amp Timberlake 1975Snodderly 1987 Snodderly amp Kurtz 1985) Severalmacaque species have been studied to date Macaca
Typeofeye
move
ment
Amplitude
Frequency
intersacc
adic
interval
Duratio
nMax
speed
Mean
speed
Conjugate
Study
Amphibians
Salamander
Tremor-lik
eAbout
125
min
ndashndash
ndashndash
ndashManteuffe
letal(1977)
Respiratory
Upto
30min
ndashndash
ndashndash
ndashManteuffe
letal(1977)
Fish
Goldfish
Drift
ndashndash
01ndash55
sec
e1degsec
036degsec
NoSim
ulta
neous
inboth
eye
sbut
conve
rgentin
thelight
Mensh
etal(2004)
Drift
Typ
ically
G5deg
ndashndash
ndashndash
NoSim
ulta
neous
inboth
eye
sbut
conve
rgent
Easter(1971)
ArcherFish
Microsaccade
ndashldquorarelyrdquofound
ndashndash
ndashndash
Segevetal(2007)
Drift
ndash02ndash1Hz
133se
ce0
4degsec
ndashndash
Segevetal(2007)
Tremor
01ndash02
deg
5Hz
ndash4degsec
~1degsec
ndashSegevetal(2007)
Table
1Characteris
ticsoffixatio
naleye
move
ments
innon-humanve
rtebrates
acc
ordingto
differentstudies
NoteaCalculatedfrom
horiz
ontalandve
rticalco
mponents
bVectorva
luesfrom
H-componentsaco
nve
rsionfactoroffrac342
hasbeenass
umed
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 4
mulatta (Goffart Quinet Chavane amp Masson 2006Horwitz amp Albright 2003 Leopold amp Logothetis1998 Martinez-Conde 2006 Martinez-Conde et al 20002002 Motter amp Poggio 1984 Skavenski et al 1975Snodderly Kagan amp Gur 2001 Steinman HaddadSkavenski amp Wyman 1973) Macaca fascicularis(Snodderly 1987 Snodderly et al 2001 Snodderly ampKurtz 1985) and Macaca nemestrina (Bair amp OrsquoKeefe1998) with no substantial differences in the spatiotemporalcharacteristics of fixational eye movements (rate velocityand magnitude profiles etc) between them Microsaccadevelocities are parametrically related to microsaccadeamplitudes following the ldquomain sequencerdquo in both ma-caques (Martinez-Conde et al 2000) and humans (Engbert2006 Martinez-Conde 2006 Martinez-Conde et al 2006Zuber amp Stark 1965) Fixational microsaccades have alsobeen observed in the baboon (Papio papio) with ampli-tudes up to 25 degrees (Marchetti Gauthier amp Pellet1983)It is important to note that microsaccades cannot be
defined according to their amplitude alone because smallexploratory or voluntary saccades can be the same size asmicrosaccades Microsaccades can be defined only opera-tionally as the involuntary saccades that are producedwhile the subject attempts to fixate (Martinez-Conde2006) This ldquoattempt to fixaterdquo includes both holding theeyes still and static while foveating the visual target andalso orienting the fovea toward a target It is worth notingthat microsaccade production may require the presence ofa visual andor attentional target (Otero-Millan et al2008)Thus there is no known physical parameter (or
combination of parameters) that separates saccades frommicrosaccades Mounting evidence points toward a com-mon neural generator of saccades and microsaccades(Engbert 2006 Martinez-Conde et al 2004 Otero-Millan et al 2008 Rolfs Kliegl amp Engbert 2008 RolfsLaubrock amp Kliegl 2006 Zuber amp Stark 1965)However one should note that most of the available datain support of a shared microsaccadendashsaccade generatorare indirect To date only two physiological studies havedirectly addressed the question of the oculomotor
mechanisms leading to the generation of microsaccadesIn those studies Van Gisbergen and colleagues foundthat putative motoneurons in the primate abducensnucleus and burst neurons in the nearby pontomedullaryreticular formation were similarly active during saccadesand microsaccades (Van Gisbergen Robinson amp Gielen1981 Van Gisbergen amp Robinson 1977) Future researchshould further explore the neural bases underlying thegeneration of microsaccades and other fixational eyemovementsSimultaneous recordings of microsaccades and neural
responses have been conducted at multiple levels of theprimate visual pathway including the lateral geniculatenucleus (LGN) area V1 and the extrastriate visual cortex(Bair amp OrsquoKeefe 1998 Leopold amp Logothetis 1998Martinez-Conde 2006 Martinez-Conde et al 20002002 Reppas Usrey amp Reid 2002 Snodderly et al2001) Microsaccades are predominantly excitatory at allthese levels leading to firing rate increases in both earlyand higher visual areas The firing rate increases followingmicrosaccades in the LGN and V1 are of visual origin(Figure 2) and result from the microsaccade-induceddisplacement of visual receptive fields over stationarystimuli Microsaccade-driven increases in firing rate tendto be clustered in tight bursts of spikes (Martinez-Conde2006 Martinez-Conde et al 2000 2002) A recent studyhas suggested that primate microsaccades may improvethe efficient sampling of fine spatial detail (Donner ampHemila 2007)Microsaccades counteract visual fading and increase
visibility during fixation in human subjects (Martinez-Conde et al 2006 Troncoso Macknik amp Martinez-Conde 2008) Numerous studies have also reported thathuman microsaccade rates andor directions are modulatedby cognitive processes such as the allocation of spatialattention (Betta amp Turatto 2006 Engbert amp Kliegl2003a 2003b 2004 Galfano et al 2004 Hafed amp Clark2002 Laubrock Engbert Rolfs amp Kliegl 2007 Otero-Millan et al 2008 Turatto Valsecchi Tame Betta 2007Valsecchi Betta amp Turatto 2007 but see also HorowitzFine Fencsik Yurgenson amp Wolfe 2007 and TseSheinberg amp Logothetis 2004)
Figure 1 Cartoon representation of fixational eye movements in humans and primates Microsaccades (straight and fast movements)drifts (curvy slow movements) and tremor (oscillations superimposed on drifts) transport the visual image across the retinal photoreceptormosaic From Martinez-Conde and Macknik (2007b)
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 5
Neural responses to drifts have received considerablyless attention than neural responses to microsaccades Thismay be due to the fact that drifts are more difficult tocharacterize objectively than microsaccades (which aremore easily detected by automatic algorithms that com-bine amplitude and velocity thresholds) Thus drifts areusually identified indirectly as the eye position changesthat occur between microsaccades (Snodderly et al2001) However this method has the potential flaw thatone may unintentionally attribute non-drift-related activity(for instance undetected tremor) to drifts Drifts mayincrease firing in a subset of primate V1 neurons(Snodderly et al 2001) However they generate lessvariability in neuronal responses than a combination ofdrifts and microsaccades (Gur Beylin amp Snodderly1997) No studies to date have recorded primate neuronalresponses in correlation to tremor (but see Hennig ampWorgotter 2007 for the responses of a model macaqueretina to simulated drift and tremor)
Fixational eye movements in the cat
The catrsquos fixational eye movements have been inves-tigated in a handful of studies and their conclusions
remain somewhat controversial In 1960 Hebbard andMarg (1960) found fixational eye movements in the cat tobe quite comparable to those in humans although catmicrosaccades were scarcer and smaller than in humansPritchard and Heron (1960) also found that the catrsquosfixational eye movements included microsaccades driftsand tremor Microsaccades were much rarer in the cat thanin humans however possibly related to the fact that thecat lacks a well-developed fovea Delgado-Garcia delPozo and Baker (1986) also observed some putativemicrosaccades in the alert behaving cat Other studiesfound no microsaccades in the cat (Conway Timberlakeamp Skavenski 1981 Winterson amp Robinson 1975)However in these last studies microsaccades were definedas ldquosaccades smaller than 10 minutes of arcrdquo which iswell below the average microsaccade amplitude found inrecent human and primate studies (see Martinez-Condeet al 2004 for a review of microsaccade parameters inhumans and primates) More recently several studies haveaddressed the contribution of fixational eye movements toneural responses in the catrsquos visual system HennigKerscher Funke and Worgotter (2002) found increasedresponses from neurons in cat areas 17 and 18 duringstimulus motions that mimicked fixational eye move-ments Using a model retina they also found that
1 simulated fixational eye movements increased neu-ral responses and
2 simulated tremor improved spatial acuity
Hennig and Worgotter (2004) later reported that acombination of tremor and microsaccades improved theperformance of simulated ganglion cells in a hyperacuitytask in the retinal periphery (but not in the central retina)Miller Denning George Marshak and Kenyon (2006)also investigated the effects of simulated tremor on amodel of the catrsquos retina and suggested that tremor mayselectively enhance the processing of large stimuli
Fixational eye movements in the rabbit
In contrast to foveate species such as primates (includ-ing humans) and cats (which have an area centralis if nota proper fovea) the rabbit (Oryctolagus cuniculus) dis-plays few spontaneous eye movements especially in theabsence of head motion (Collewijn 1977 Collewijn ampvan der Mark 1972 Fuller 1980 1981 Van der Steen ampCollewijn 1984) In normal visual conditions the rabbitrsquoseyes are very stable but not perfectly immobile Smallamplitude tremor can be observed as well as very slowdrift Microsaccades have not been observed Howeverdrift is occasionally corrected by fast saccadic movements(Collewijn 1977 Collewijn amp van der Mark 1972 Vander Steen amp Collewijn 1984) These differences may berelated to the fact that the rabbit does not have a fovea butan elongated horizontal streak with elevated ganglion cell
Figure 2 Microsaccades drive neural activity in primate area V1Microsaccades lead to firing rate increases in V1 neurons (n = 308)of the awake primate In the absence of visual stimulationmicrosaccades do not increase neural activity in area V1 (n = 37neurons) The resultsVshown here for the average of all neuronsVwere consistent with responses from individual neurons (data notshown) Modified from Martinez-Conde et al (2000 2002)
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 6
counts Olveczky Baccus and Meister (2003) recordedfrom ganglion cells in the rabbit and salamander retinaswhile presenting visual stimuli that were jittered tosimulate fixational eye movements (based on parametersfrom previously published studies by Manteuffel PlasaSommer amp Wess 1977 and Van der Steen amp Collewijn1984) In both species fixational eye movements helpedsegregate a moving foreground from a complex back-ground in a population of ganglion cells
Fixational eye movements inbirds
Fixational eye movements in the pigeon
There are some remarkable differences between fixa-tional eye movements in birds and mammals Nye (1969)identified four types of fixational eye movements in thepigeon (Columba livia) flicks (ie microsaccades)impulse movements drift and oscillations No tremorwas detected Microsaccade frequency was very lowrelative to humans Impulse movements occurred morefrequently usually between bursts of oscillations withamplitudes ranging up to 2 degrees Drift velocitiesranged from 1 to 5 degrees per second Both micro-saccades and drifts occurred in response to movingstimuli Oscillations occurred in short bursts at rates ofabout 30 Hz and could be quite large (up to severaldegrees of amplitude) Bloch Rivaud and Martinoya(1984) reported microsaccades drifts and oscillationsldquosimilar to those described by Nyerdquo but none of thesewere analyzed in detailCyclotorsional oscillations are an integral part of avian
saccades (Wallman Pettigrew amp Letelier 1994) Thusone might speculate that oscillations are associated withavian microsaccades as well especially if saccades andmicrosaccades share a common generator (Engbert 2006Otero-Millan et al 2008 Rolfs et al 2008 2006 Zuberamp Stark 1965) However this possibility has not yet beenaddressed in the literature The function of oscillations isunknown (Carpenter 1988) but one hypothesis is thatthey serve to facilitate the delivery of arterial blood to theocular capillary bed (Pettigrew Wallman amp Wildsoet1990 Wallman et al 1994)
Fixational eye movements in the owl
Steinbach and Money (1973) recorded fixational eyemovements in the great horned owl (Bubo virginianus)a species previously thought incapable of moving itseyes The owlrsquos fixational eye movements includeflicks or microsaccades drift tremor and oscillations
Microsaccades seemed correlated to visual events ofinterest to the owl they were 1 deg or less in amplitudeand had peak velocities of about 10 degsec Drifts wereslow motions of the eye in between fixations withamplitudes of 1 degree or less Tremor amplitude wasabout 1 arcmin with frequencies around 20 Hz High-frequency oscillations (25 Hz) of short duration (025 sec)were very similar to those previously found in the pigeon(Nye 1969) No studies to date have examined theperceptual or physiological effects of fixational eyemovements in any bird species
Fixational eye movements inreptiles
Fixational eye movements in the turtle
Greschner Bongard Rujan and Ammermuller (2002)identified two types of fixational eye movements in theturtle (Pseudemys scripta elegans) which they calledldquodrift-likerdquo and ldquoperiodicrdquo Drift-like motions were smalland slow (with velocities comparable to human drift) andwere superimposed by larger and faster periodic componentsPeriodic motion had retinal amplitudes on the order of thediameter of a photoreceptor which are comparable to thoseof human tremor On the other hand periodic motionrsquosfrequency (5 Hz) velocity and orderly relationship betweenvelocity and amplitude are closer to those of human micro-saccades (see Martinez-Conde et al 2004 for a review oftremor and microsaccade parameters in humans)Simulated periodic motion generated strong synchronous
firing in the turtlersquos retina whereas simulated drift-likemotion had little effect (Figure 3) These results agree withprevious physiological and modeling studies in the primatevisual system in which strong neural transients wereobserved in response to microsaccades (Donner amp Hemila2007 Martinez-Conde 2006 Martinez-Conde et al 20002002) Such neural transients may underlie the behavior ofcortical neurons as coincidence detectors (ShelleyMcLaughlin Shapley amp Wielaard 2002 Williams ampShapley 2007) Moreover neural transients to stimulionsets and terminations (similar to those produced bymicrosaccades in the primate visual system Martinez-Conde 2006 Martinez-Conde et al 2000 2002) havebeen related to target visibility in visual masking para-digms (Macknik amp Livingstone 1998 Macknik ampMartinez-Conde 2004 Macknik Martinez-Conde ampHaglund 2000)In the turtle retina periodic motion-driven neurons with
receptive fields located along contrast borders weremoreover synchronized and reliably indicated the preced-ing motions The authors proposed that this synchroniza-tion of retinal activity could be used by the brain to
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 7
improve the estimation of stimulus features such asspatial frequency (Greschner et al 2002) This findingcould have a parallel in the primate visual system Donnerand Hemila (2007) recently proposed that microsaccadessignificantly ldquore-sharpenrdquo the image and improve spatialresolution in primates Rucci Iovin Poletti and Santini(2007) also found that fixational eye movements enhancefine spatial detail in human subjects
Fixational eye movements in the chameleon
Fixational eye movements in the chameleon have notbeen studied in detail However Gioanni and colleaguesfound that the chameleon (Chamaleo chamaleo) makessaccades during fixation The authors speculated that theseprobably correspond to fixational microsaccades found inother foveate species (Gioanni Bennis amp Sansonetti1993)
Fixational eye movements inamphibians
Fixational (and more generally spontaneous) eye move-ments vary widely among amphibians The frog (anafoveate amphibian) shows a general lack of eye andhead movements except for those that stabilize the retinalimage Dieringer and colleagues (Dieringer amp Daunicht1986 Dieringer amp Precht 1982 Dieringer Precht ampBlight 1982) found that the grass frog (Rana temporaria)produces no spontaneous eye movements in the absenceof head movements with the possible exception of tremor(similarly to the rabbit another afoveate species Van derSteen amp Collewijn 1984) It is likely that the frogrsquos lack ofeye movements leads to fast visual fading of all stationaryobjects in its field of view (Ditchburn amp Ginsborg 1952Yarbus 1967) Lettvin Maturana MsCulloch and Pitts
Figure 3 Fixational eye movements increase retinal activity in the turtle (A) Continuous recording from 54 retinal ganglion cells in theturtle during darkness after switching on a stationary grating and during wobbling of this grating The grating was wobbled to simulateperiodic (ie tremor-like) eye movements (B) Responses of a single ganglion cell to a drifting contrast border of velocity comparable toocular drifts (C) When the drift is superimposed with wobbling the responses of the cell shown in (B) increase dramatically FromGreschner et al (2002) and modified by Martinez-Conde et al (2004)
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 8
(1968) stated that a frog ldquowill starve to death surroundedby food if it is not movingrdquo (but see Dieringer amp Daunicht1986 for a counterpoint) Small head motions or eyemovements correlated with respiratory motions maydiminish fading to some extent (Dieringer amp Precht1982 Schipperheyn 1963 1965 1967)As a side note the frogrsquos respiratory eye movements
play no role in the respiratory function ldquoin the course ofeach respiratory cycle increase of pressure in the frogrsquosbuccal cavity protrudes the eye and causes the opticalaxis to deviate upwards over 25ndash35 minutes of arcrdquo(Schipperheyn 1963) That is respiratory eye movementsare merely synchronized with respirationAutrum (1959) found no evidence of respiratory eye
movements in the frog Ewert andBorchers (1974) observedthat stationary stimuli elicit no responses from visualneurons in the common toad (Bufo Bufo L) regardless ofrespiratory pressure changes in the buccal cavity
Fixational eye movements in the salamander
Manteuffel et al (1977) identified two types of fixa-tional eye movements in salamanders tremor-like andrespiratory eye movements Tremor-like is a rapid move-ment of small amplitude Torsional eye movements resultfrom the animalrsquos breathing (also found in the frog bySchipperheyn 1963) Additionally flicks are occasionallyobserved which rapidly change the position of gaze It isunclear whether such flicks may generally correspond towhat we now call microsaccades andor to voluntarysaccades Based on these observations the authors pro-posed that unlike frogs and toads (Autrum 1959 Ewert ampBorchers 1974 Lettvin et al 1968) salamanders cancontinuously perceive stationary objects (Manteuffel et al1977) Fixational eye movements in the salamander (Tigersalamander Ambystoma tigrinum) may also help segre-gate object and background motion in the retina(Olveczky et al 2003 see also Fixational eye movementsin the rabbit section) and even work to emphasize novelvisual features (Olveczky et al 2007 see Baccus et al2008 for details on the retinal neurons and synapses thatmay underlie the computation of differential motionbetween an object and its background)
Fixational eye movements in fish
Spontaneous eye movements in the goldfish
Eye movements during fixation have been studied inboth afoveate (goldfish Carassius auratus) and foveate(archer fish Toxotes chatareus) fish species The goldfisha long-standing model organism in vision and oculomotor
research (Aksay et al 2003 2007 Cabrera Torres PasaroPastor amp Delgado-Garcia 1992 Pastor Torres Delgado-Garcia amp Baker 1991 Salas Navarro Torres ampDelgado-Garcia 1992 Torres Pastor Cabrera Salas ampDelgado-Garcia 1992) produces three types of sponta-neous eye movements saccades stretch and drift (Easter1971 Mensh Aksay Lee Seung amp Tank 2004) Weshould note that in the motionless goldfish eye move-ments are very infrequent Those that do occur consistmainly of saccades slow drifts are very rare (Easter Johnsamp Heckenlively 1974) During saccades (identified bythresholding the eye velocity at 5 degsec) both eyes moverapidly simultaneously and usually in the same direction(Mensh et al 2004) Stretches occur every 2ndash3 min and arecharacterized by a brief simultaneous large excursion ofboth eyes to extreme temporal positions (Easter 1971Mensh et al 2004) The function of stretches is not known(Mensh et al 2004) Because fixations were defined asthose periods that were free of saccadic and stretchmovements (Easter 1971) only drifts (from the threetypes of spontaneous movements) can technically beconsidered ldquofixational eye movementsrdquo However thegoldfish saccadic main sequence includes saccades withequivalent amplitudes and velocities to those found inhuman microsaccades (Mensh et al 2004 Zuber amp Stark1965) Indeed microsaccades have been identified in other(foveate) fish species although they are rarer than inhumans (Segev et al 2007) Small saccades in thegoldfish tend to be divergent whereas large saccadestend to be convergent (Mensh et al 2004) Drift occurssimultaneously in both eyes usually in the samedirection Drift velocities are almost exclusively e1 degsec(Mensh et al 2004) No tremor has been identified in thegoldfish
Spontaneous eye movements in teleost fish
Spontaneous eye movements have been studied in theNew Zealand parore (Girella tricuspidata MontgomeryMcVean amp McCarthy 1983) The pattern of eye move-ments was similar to that previously described in goldfish(Easter 1971 Hermann amp Constantine 1971)
Fixational eye movements in the archer fish
A recent study in the archer fish (Segev et al 2007) hasprovided the first perceptual and physiological correlatesof fixational eye movements in any fish species Archerfish have remarkable visual abilities they shoot downinsects resting on foliageVas well as stationary targetscovering a single retinal photoreceptorVwith squirts ofwater from their mouths they predict the location atwhich the insect will splash down in G100 msec and theydeduce the absolute size of an object presented at differentviewing distances (Supplementary Videos 2 and 3) Segev
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 9
et al (2007) identified three types of fixational eyemovements in the archer fish microsaccades drift andtremor Microsaccades were rare and were not analyzed indetail Drift velocities were e040 degsec Tremor was ahigh-frequency oscillatory movement (the primary com-ponent was 5 Hz) with peak-to-peak amplitude of 01ndash02 deg (corresponding to an image displacement of 1ndash2photoreceptors) and velocity of about 1 degsec (Figure 4)Whereas the amplitude and velocity of tremor in thearcher fish is comparable to those in humans its frequencyis much lower (5 Hz vs 940 Hz) Interestingly archer fishtremor is very similar both in amplitude (photoreceptorwidths covered) as well as in frequency (5 Hz) to thetremor-like periodic motion in the turtle (discussedearlier Greschner et al 2002)Segev et al (2007) trained archer fish to perform a size
discrimination task the fish had to distinguish a medium-sized target from larger and smaller distracters (theyindicated their choice by shooting a jet of water) To relatethis behavior to responses from retinal neurons theauthors recorded from populations of retinal ganglioncells while presenting objects of different sizes moved tosimulate saccades drifts and tremor Drifts and tremorelicited weak neuronal responses that were not veryinformative about the spatial structure of the stimulus(although they did provide some information about target
size in agreement with the results of Greschner et al2002 in the turtle) Moreover performance was similar fortremor alone and for tremor plus drift indicating that driftplays a negligible role in revealing spatial structure (alsoin agreement with Greschner et al 2002)When present the firing rate was phase-locked to
tremor Saccades were more effective than drifts or tremorin driving informative responses from the ganglion cells(consistent with theoretical and physiological studies ofmicrosaccade-driven neuronal activity in the primateretina and LGN Donner amp Hemila 2007 Martinez-Conde et al 2002) Segev et al (2007) proposed that inprimates both saccades and microsaccades (which arerare in the archer fish) may contribute to size discrim-ination tasks as well as enhance spatial detail whereasdrifts and tremor by themselves may not be sufficient toimprove visual perception However it is important toremember that central vision in humans can be maintainedby tremor andor drift even when microsaccades aresuppressed during strict fixation (Martinez-Conde 2006Martinez-Conde et al 2004 2006) On the other handone cannot rule out the possibility that if one couldcompletely eliminate drifts and tremor microsaccadesmight then suffice to sustain both central and peripheralvision in primates and humans Future experiments areneeded to further explore these ideas
Figure 4 Archer fish eye movements (A) Example trace of eye position as a function of time as measured by search coil (B) Example ofa single 15-s fixation superimposed on a grid representing the photoreceptor mosaic The back and forth movement is caused by tremor(C) Distribution of fixation durations (taken as time interval between saccades) (D) Power spectrum of horizontal fixational eyemovements (E) Distribution of tremor amplitude (data taken from 1 animal in a 15-min session) Modified from Segev et al (2007)
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 10
Discussion
The vast majority of eye movement studies conductedin non-human species to date have not analyzed eyeposition data in the range of fixational eye movementsThus eye movements during fixation have been reportedin only a handful of vertebrates Most of these studieshave concentrated solely on the physical parameters ofthe fixational eye movements without addressing theirphysiological and perceptual consequences Fortunatelythis trend has started to reverse in recent years with agrowing list of papers addressing the effects of non-human fixational eye movements in physiology andperception This is an important development A largeamount of all psychophysical and physiological visualresearch has been conducted during conditions of visualfixation Therefore understanding the precise physiolog-ical and perceptual contributions of fixational eye move-ments may be critical to the interpretation of previous andfuture vision researchThere are some interesting differences and similarities
in the types of fixational eye movements produced bydifferent species For instance ocular tremor is possiblyubiquitous to all vertebrates The fact that tremor has notbeen observed in all species studied could be due to thetechnical difficulty of recording tremor accurately (ietremor amplitudes and frequencies are usually in the rangeof the recording systemrsquos noise) Drift has also beenrecorded in most vertebrates and it may be as omnipre-sent as tremor Conversely microsaccades seem mostimportant in species with foveal vision (ie primates andhumans) whereas they are scarce or absent in afoveatespecies (ie rabbits frogs goldfish) A link betweenmicrosaccades and foveal vision may reflect the two-pronged functional significance of microsaccades inhumans and primates first microsaccades may benecessary to correct the slow drift of the eye and bringback the objectregion of interest to the retinal area wherephotoreceptor density is highest (Cornsweet 1956) Thecorrective role of human microsaccades has been demon-strated in recent studies (Engbert amp Kliegl 2004 Engbertamp Mergenthaler 2006) Second the dynamics of micro-saccades themselves may improve spatial resolution(Donner amp Hemila 2005)In reviewing the patterns of fixational eye movements
across vertebrates it is tempting to draw parallels withother sensory modalities Fixational eye movementsrefresh the retinal images during fixation and thus preventneural adaptation and visual fading But this mechanism isnot circumscribed to the visual system Sniffs in rodentolfaction discretely sample sensory information every200ndash300 msec and thus are similar in their temporaldynamics to primate saccades (Otero-Millan et al 2008Uchida Kepecs amp Mainen 2006) and microsaccades(Otero-Millan et al 2008) A similar mode of discretesampling may also be at play when objects are recognized
through tactile information for instance if we sweep ourfingertips over an objectrsquos surface with our eyes closed orwhen blind individuals read Braille scriptOne intriguing possibility that merits further experi-
ments and analyses is the potential correlation between thespatiotemporal parameters of fixational eye movements(such as those summarized in Table 1) and the adaptationproperties of visual neurons across species For instancethe greater ocular stability of some species (rabbitgoldfish frog etc) versus others (primates) could beassociated with some particular adaptive properties oftheir visual cells Thus future comparative studies shouldpay attention to the response properties of visual cellstriggered by fixational eye movements in the presence ofstationary stimuli Other potential factors underlying thevariation in fixational eye movements across species couldbe differences in oculomotor control at the level of the eyemuscles or perhaps differences in retinal organizationFuture research will further improve our understanding
of fixational eye movements and the neural activity thatmaintains the visibility of stationary visual scenes inhuman and non-human species One interesting questionthat remains to be addressed concerns the role of tremor invision and visual physiology Because tremor frequenciesare well over the flicker fusion frequencies in humans ithas been argued that tremor of the visual image would beineffective as a stimulus (Ditchburn 1955 Gerrits ampVendrik 1970 Sharpe 1972) However recent studieshave found simulated tremor to enhance visual processingand improve visual acuity in a model retina (Hennig et al2002 Miller et al 2006) Follow-up research should beaimed to replicate these findings in actual neurons withcorrelated perceptual reports One should also note thatearly visual neurons can follow high-frequency flickeringthat surpasses the perceptual threshold for flicker fusion(Martinez-Conde et al 2002) Thus high-frequencytremor could be adequate to maintain activity in the earlyvisual system which then may lead to visual perception inhigher visual areas Another significant question thatremains unanswered concerns the oculomotor bases ofthe various types of fixational eye movements acrossspecies To date there is too little information on thisimportant topic to draw a conclusion
Conclusions
We have reviewed the parameters of fixational eyemovements in non-human mammals birds reptilesamphibians and fish as well as their consequences forphysiology and perception Fixational eye movementsand microsaccades in particular appear to be mostimportant in foveate vs afoveate species Animals lackingfixational eye movements such as afoveate frogs andtoads may be incapable of seeing stationary objects
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 11
during visual fixation Future research should furtherexplore the perceptual effects of fixational eye movementsand determine the oculomotor mechanisms responsible fortheir generation across species
Acknowledgments
We thank Mona Stewart and Isabel Gomez-Caraballo fortechnical assistance Jorge Otero-Millan for programmingassistance and Dr Xoana G Troncoso for comments on themanuscript This study was funded through grants fromthe Barrow Neurological Foundation (to SMC and SLM)the National Science Foundation (Award 0643306 toSMC) the Arizona Biomedical Research Commission(Award 07-102 to SMC) and Science Foundation Arizona(SLM)
Commercial relationships noneCorresponding author Susana Martinez-Conde PhDEmail smartneuralcorrelatecomAddress Barrow Neurological Institute 350 W ThomasRd Phoenix AZ 85013 USA
References
Aksay E Major G Goldman M S Baker R SeungH S amp Tank D W (2003) History dependence ofrate covariation between neurons during persistentactivity in an oculomotor integrator Cerebral Cortex13 1173ndash1184 [PubMed] [Article]
Aksay E Olasagasti I Mensh B D Baker RGoldman M S amp Tank D W (2007) Functionaldissection of circuitry in a neural integrator NatureNeuroscience 10 494ndash504 [PubMed]
Autrum H (1959) Das Fehlen unwillkurlicher Augenbe-wegungen beim Frosch Naturwissenschaften 46435
Baccus S A Olveczky B P Manu M amp Meister M(2008) A retinal circuit that computes object motionJournal of Neuroscience 28 6807ndash6817 [PubMed]
Bair W amp OrsquoKeefe L P (1998) The influence offixational eye movements on the response of neuronsin area MT of the macaque Visual Neuroscience 15779ndash786 [PubMed]
Betta E amp Turatto M (2006) Are you ready I can tellby looking at your microsaccades Neuroreport 171001ndash1004 [PubMed]
Bloch S Rivaud S amp Martinoya C (1984) Compar-ing frontal and lateral viewing in the pigeon IIIDifferent patterns of eye movements for binocularand monocular fixation Behavioural Brain Research13 173ndash182 [PubMed]
Cabrera B Torres B Pasaro R Pastor A M ampDelgado-Garcia J M (1992) A morphological studyof abducens nucleus motoneurons and internuclearneurons in the goldfish (Carassius auratus) BrainResearch Bulletin 28 137ndash144 [PubMed]
Carpenter R H S (1988) Movements of the eyes(2nd ed) Pion London
Collewijn H (1977) Eye- and head movements infreely moving rabbits The Journal of Physiology266 471ndash498 [PubMed] [Article]
Collewijn H amp van der Mark F (1972) Ocular stabilityin variable visual feedback conditions in the rabbitBrain Research 36 47ndash57 [PubMed]
Conway J L Timberlake G T amp Skavenski A A(1981) Oculomotor changes in cats reared withoutexperiencing continuous retinal image motion Exper-imental Brain Research 43 229ndash232 [PubMed]
Cornsweet T N (1956) Determination of the stimuli forinvoluntary drifts and saccadic eye movements Jour-nal of the Optical Society of America 46 987ndash993[PubMed]
Delgado-Garcia J M del Pozo F amp Baker R (1986)Behavior of neurons in the abducens nucleus of thealert catVII Internuclear neurons Neuroscience 17953ndash973 [PubMed]
Dieringer N amp Daunicht W J (1986) Image fadingVAproblem for frogs Naturwissenschaften 73 330ndash332[PubMed]
Dieringer N amp Precht W (1982) Compensatory headand eye movements in the frog and their contributionto stabilization of gaze Experimental BrainResearch 47 394ndash406 [PubMed]
Dieringer N Precht W amp Blight A R (1982)Resetting fast phases of head and eye and theirlinkage in the frog Experimental Brain Research 47407ndash416 [PubMed]
Ditchburn R W (1955) Eye-movements in relation toretinal action Optica Acta 1 171ndash176
Ditchburn R W amp Ginsborg B L (1952) Vision with astabilized retinal image Nature 170 36ndash37[PubMed]
Ditchburn R W amp Ginsborg B L (1953) Involuntaryeye movements during fixation The Journal ofPhysiology 119 1ndash17 [PubMed] [Article]
Donner K amp Hemila S (2007) Modelling the effect ofmicrosaccades on retinal responses to stationarycontrast patterns Vision Research 47 1166ndash1177[PubMed]
Donner K K amp Hemila S (2005) The possible purposeof microsaccades Paper presented at the EuropeanConference on Visual Perception A Coruna Spain
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 12
Easter S S Jr (1971) Spontaneous eye movements inrestrained goldfish Vision Research 11 333ndash342[PubMed]
Easter S S Jr Johns P R amp Heckenlively D R(1974) Horizontal compensatory eye movements ingoldfish (Carassius auratus) I The normal animalJournal of Comparative Physiology 92 23ndash35
Engbert R (2006) Microsaccades A microcosm forresearch on oculomotor control attention and visualperception Progress Brain Research 154 177ndash192[PubMed]
Engbert R amp Kliegl R (2003a) Binocular coordinationin microsaccades In R R J Hyona amp H Deubel(Eds) The mindrsquos eyes Cognitive and appliedaspects of eye movements (pp 103ndash117) OxfordUK Elsevier
Engbert R amp Kliegl R (2003b) Microsaccades uncoverthe orientation of covert attention Vision Research43 1035ndash1045 [PubMed]
Engbert R amp Kliegl R (2004) Microsaccades keep theeyesrsquo balance during fixation Psychological Science15 431ndash436 [PubMed]
Engbert R amp Mergenthaler K (2006) Microsaccadesare triggered by low retinal image slip Proceedingsof the National Academy of Sciences of the UnitedStates of America 103 7192ndash7197 [PubMed][Article]
Ewert J P amp Borchers H W (1974) Antwort vonretinalen Ganglienzellen bei freibeweglichen Kroten(Bufo Bufo L) Journal of Comparative and Physi-ology 92 117ndash130
Fuller J H (1980) Linkage of eye and head movementsin the alert rabbit Brain Research 194 219ndash222[PubMed]
Fuller J H (1981) Eye and head movements duringvestibular stimulation in the alert rabbit BrainResearch 205 363ndash381 [PubMed]
Galfano G Betta E amp Turatto M (2004) Inhibition ofreturn in microsaccades Experimental BrainResearch 159 400ndash404 [PubMed]
Gerrits H J amp Vendrik A J (1970) Artificial move-ments of a stabilized image Vision Research 101443ndash1456 [PubMed]
Gioanni H Bennis M amp Sansonetti A (1993) Visualand vestibular reflexes that stabilize gaze in thechameleon Visual Neuroscience 10 947ndash956[PubMed]
Goffart L Quinet J Chavane F amp Masson G S(2006) Influence of background illumination onfixation and visually guided saccades in the rhesusmonkey Vision Research 46 149ndash162 [PubMed]
Greschner M Bongard M Rujan P amp Ammermuller J(2002) Retinal ganglion cell synchronization by fixa-tional eye movements improves feature estimationNature Neuroscience 5 341ndash347 [PubMed]
Gur M Beylin A amp Snodderly D M (1997)Response variability of neurons in primary visualcortex (V1) of alert monkeys Journal of Neuro-science 17 2914ndash2920 [PubMed] [Article]
Hafed Z M amp Clark J J (2002) Microsaccades as anovert measure of covert attention shifts VisionResearch 42 2533ndash2545 [PubMed]
Hebbard F W amp Marg E (1960) Physiologicalnystagmus in the cat Journal of the Optical Societyof America 50 151ndash155 [PubMed]
Hennig M A Kerscher N J Funke K amp Worgotter F(2002) Stochastic resonance in visual cortical neuronsDoes the eye-tremor actually improve visual acuityNeurocomputing 44ndash46 115ndash120
Hennig M H amp Worgotter F (2004) Eye micro-movements improve stimulus detection beyond theNyquist limit in the peripheral retina In Advances inNeural Information Processing Systems (NIPS2003)(vol 16) Denver CO MIT Press
Hennig M H amp Worgotter F (2007) Effects offixational eye movements on retinal ganglion cellresponses A modelling study Frontiers in Computa-tional Neuroscience 1 2 [PubMed] [Article]
Hermann H T amp Constantine M M (1971) Eyemovements in the goldfish Vision Research 11313ndash331 [PubMed]
Horowitz T S Fine E M Fencsik D E Yurgenson Samp Wolfe J M (2007) Fixational eye movements arenot an index of covert attention PsychologicalScience 18 356ndash363 [PubMed]
Horwitz G D amp Albright T D (2003) Short-latencyfixational saccades induced by luminance incrementsJournal of Neurophysiology 90 1333ndash1339[PubMed] [Article]
Laubrock J Engbert R Rolfs M amp Kliegl R(2007) Microsaccades are an index of covertattention Commentary on Horowitz Fine FencsikYurgenson and Wolfe (2007) Psychological Sci-ence 18 364ndash366 discussion 367ndash368 [PubMed]
Leopold D A amp Logothetis N K (1998) Micro-saccades differentially modulate neural activity in thestriate and extrastriate visual cortex ExperimentalBrain Research 123 341ndash345 [PubMed]
Lettvin J Y Maturana H R MsCulloch W S amp PittsW H (1968) What the frogrsquos eye tells the frogrsquosbrain In W C C a M Balaban (Ed) The mind
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 13
Biological approaches to its functions (pp 233ndash258)John Wiley and Sons London
Macknik S L amp Livingstone M S (1998) Neuronalcorrelates of visibility and invisibility in the primatevisual system Nature Neuroscience 1 144ndash149[PubMed]
Macknik S L amp Martinez-Conde S (2004) The spatialand temporal effects of lateral inhibitory networksand their relevance to the visibility of spatiotemporaledges Neurocomputing 58ndash60 775ndash782
Macknik S L Martinez-Conde S amp Haglund M M(2000) The role of spatiotemporal edges in visibilityand visual masking Proceedings of the NationalAcademy of Sciences of the United States of America97 7556ndash7560 [PubMed] [Article]
Manteuffel G Plasa L Sommer T J amp Wess O(1977) Involuntary eye movements in salamandersNaturwissenschaften 64 533ndash534 [PubMed]
Marchetti E Gauthier G M amp Pellet J (1983)Cerebellar control of eye movements studied withinjection of harmaline in the trained baboon ArchivesItaliennes de Biologies 121 1ndash17 [PubMed]
Martinez-Conde S (2006) Fixational eye movements innormal and pathological vision Progress in BrainResearch 154 151ndash176 [PubMed]
Martinez-Conde S amp Macknik S L (2007a) Bobbingand perceiving Scientific American 297 16
Martinez-Conde S amp Macknik S L (2007b) Windowson the mind Scientific American 297 56ndash63[PubMed]
Martinez-Conde S Macknik S L amp Hubel D H(2000) Microsaccadic eye movements and firing ofsingle cells in the striate cortex of macaque monkeysNature Neuroscience 3 251ndash258 [PubMed]
Martinez-Conde S Macknik S L amp Hubel D H(2002) The function of bursts of spikes during visualfixation in the awake primate lateral geniculate nucleusand primary visual cortex Proceedings of theNational Academy of Sciences of the United Statesof America 99 13920ndash13925 [PubMed] [Article]
Martinez-Conde S Macknik S L amp Hubel D H(2004) The role of fixational eye movements invisual perception Nature Reviews Neuroscience 5229ndash240 [PubMed]
Martinez-Conde S Macknik S L Troncoso X G ampDyar T A (2006) Microsaccades counteract visualfading during fixation Neuron 49 297ndash305[PubMed] [Article]
Mensh B D Aksay E Lee D D Seung H S ampTank D W (2004) Spontaneous eye movements ingoldfish Oculomotor integrator performance plasti-
city and dependence on visual feedback VisionResearch 44 711ndash726 [PubMed]
Miller J A Denning K S George J S MarshakD W amp Kenyon G T (2006) A high frequencyresonance in the responses of retinal ganglion cells torapidly modulated stimuli A computer model VisualNeuroscience 23 779ndash794 [PubMed]
Montgomery J C McVean A R amp McCarthy D(1983) The effects of lowered temperature onspontaneous eye movements in a teleost fish Com-parative Biochemistry and Physiology A Compara-tive Physiology 75 363ndash368 [PubMed]
Motter B C amp Poggio G F (1984) Binocularfixation in the rhesus monkey Spatial and temporalcharacteristics Experimental Brain Research 54304ndash314 [PubMed]
Murakami I amp Cavanagh P (1998) A jitter after-effectreveals motion-based stabilization of vision Nature395 798ndash801 [PubMed]
Nye P W (1969) The monocular eye movements of thepigeon Vision Research 9 133ndash144 [PubMed]
Olveczky B P Baccus S A amp Meister M (2003)Segregation of object and background motion in theretina Nature 423 401ndash408 [PubMed]
Olveczky B P Baccus S A amp Meister M (2007)Retinal adaptation to object motion Neuron 56689ndash700 [PubMed] [Article]
Otero-Millan J Troncoso X Macknik S L Serrano-Pedraza I amp Martinez-Conde S (2008) Saccadesand microsaccades during visual fixation explorationand search Foundations for a common saccadicgenerator Journal of Vision 8(14)21 1ndash18 httpjournalofvisionorg81421 doi10116781421[PubMed] [Article]
Pastor A M Torres B Delgado-Garcia J M ampBaker R (1991) Discharge characteristics of medialrectus and abducens motoneurons in the goldfishJournal of Neurophysiology 66 2125ndash2140 [PubMed]
Pettigrew J D Wallman J amp Wildsoet C F (1990)Saccadic oscillations facilitate ocular perfusion fromthe avian pecten Nature 343 362ndash363 [PubMed]
Pritchard R M amp Heron W (1960) Small eye move-ments of the cat Canadian Journal of Psychology14 131ndash137 [PubMed]
Ratliff F amp Riggs L A (1950) Involuntary motions ofthe eye during monocular fixation Journal of Exper-imental Psychology 40 687ndash701 [PubMed]
Reppas J B UsreyWM amp Reid R C (2002) Saccadiceye movements modulate visual responses in thelateral geniculate nucleus Neuron 35 961ndash974[PubMed] [Article]
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 14
Riggs L A amp Ratliff F (1952) The effects of counter-acting the normal movements of the eye Journal ofthe Optical Society of America 42 872ndash873
Rolfs M Kliegl R amp Engbert R (2008) Toward amodel of microsaccade generation The case ofmicrosaccadic inhibition Journal of Vision 8(11)51ndash23 httpjournalofvisionorg8115 doi1011678115 [PubMed] [Article]
Rolfs M Laubrock J amp Kliegl R (2006) Shorteningand prolongation of saccade latencies followingmicrosaccades Experimental Brain Research 169369ndash376 [PubMed]
Rucci M Iovin R Poletti M amp Santini F (2007)Miniature eye movements enhance fine spatial detailNature 447 851ndash854 [PubMed]
Salas C Navarro F Torres B amp Delgado-Garcia J M(1992) Effects of diazepam and D-amphetamine onrhythmic pattern of eye movements in goldfishNeuroreport 3 131ndash134 [PubMed]
Schipperheyn J J (1963) Respiratory eye movementsand perception of stationary objects in the frog ActaPhysiologica et Pharmacologica Neerlandica 12157ndash159 [PubMed]
Schipperheyn J J (1965) Contrast detection in frogrsquosretina Acta Physiologica et Pharmacologica Neer-landica 13 231ndash277 [PubMed]
Schipperheyn J J (1967) Influence of eye movements onretinal ganglion cell activity in frogs Acta Physi-ologica et Pharmacologica Neerlandica 14 506[PubMed]
Segev R Schneidman E Goodhouse J amp Berry M J2nd (2007) Role of eye movements in the retinalcode for a size discrimination task Journal ofNeurophysiology 98 1380ndash1391 [PubMed] [Article]
Sharpe C R (1972) The visibility and fading of thinlines visualized by their controlled movement acrossthe retina The Journal of Physiology 222 113ndash134[PubMed] [Article]
Shelley M McLaughlin D Shapley R amp Wielaard J(2002) States of high conductance in a large-scalemodel of the visual cortex Journal of ComputationalNeuroscience 13 93ndash109 [PubMed]
Skavenski A A Robinson D A Steinman R M ampTimberlake G T (1975) Miniature eye movementsof fixation in rhesus monkey Vision Research 151269ndash1273 [PubMed]
Snodderly D M (1987) Effects of light and darkenvironments on macaque and human fixational eyemovements Vision Research 27 401ndash415 [PubMed]
Snodderly D M Kagan I amp Gur M (2001) Selectiveactivation of visual cortex neurons by fixational eyemovements Implications for neural coding VisualNeuroscience 18 259ndash277 [PubMed]
Snodderly D M amp Kurtz D (1985) Eye position duringfixation tasks Comparison of macaque and humanVision Research 25 83ndash98 [PubMed]
Steinbach M J amp Money K E (1973) Eye movementsof the owl Vision Research 13 889ndash891 [PubMed]
Steinman R M Haddad G M Skavenski A A ampWyman D (1973) Miniature eye movement Science181 810ndash819 [PubMed]
Torres B Pastor A M Cabrera B Salas C ampDelgado-Garcia J M (1992) Afferents to theoculomotor nucleus in the goldfish (Carassius aur-atus) as revealed by retrograde labeling with horse-radish peroxidase Journal of Comparative Neurology324 449ndash461 [PubMed]
Troncoso X G Macknik S L amp Martinez-Conde S(2008) Microsaccades counteract perceptual filling-inJournal of Vision 8(14)15 1ndash9 httpjournalofvisionorg81415 doi10116781415 [Article]
Troncoso X G Macknik S L Otero-Millan J ampMartinez-Conde S (2008) Microsaccades driveillusory motion in the Enigma illusion Proceedingsof the National Academy of Sciences of the UnitedStates of America 105 16033ndash16038 [PubMed]
Tse P U Sheinberg D S amp Logothetis N K (2004)The distribution of microsaccade directions need notreveal the location of attention Psychological Science15 708ndash710
Turatto M Valsecchi M Tame L amp Betta E (2007)Microsaccades distinguish between global and localvisual processing Neuroreport 18 1015ndash1018[PubMed]
Uchida N Kepecs A amp Mainen Z F (2006) Seeing ata glance smelling in a whiff Rapid forms ofperceptual decision making Nature Reviews Neuro-science 7 485ndash491 [PubMed]
Valsecchi M Betta E amp Turatto M (2007) Visualoddballs induce prolonged microsaccadic inhibitionExperimental Brain Research 177 196ndash208[PubMed]
van Dam L C amp van Ee R (2006) Retinal image shiftsbut not eye movements per se cause alternations inawareness during binocular rivalry Journal of Vision6(11)3 1172ndash1179 httpjournalofvisionorg6113doi1011676113 [PubMed] [Article]
Van der Steen J amp Collewijn H (1984) Ocular stabilityin the horizontal frontal and sagittal planes in therabbit Experimental Brain Research 56 263ndash274[PubMed]
Van Gisbergen J A Robinson D A amp Gielen S(1981) A quantitative analysis of generation ofsaccadic eye movements by burst neurons Journalof Neurophysiology 45 417ndash442 [PubMed]
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 15
Van Gisbergen J A M amp Robinson D A (1977)Generation of micro and macrosaccades by burstneurons in the monkey In R Baker amp A Berthoz(Eds) Control of gaze by brain stem neurons (vol 1pp 301ndash308) New York ElsevierNorth-Holland
Wallman J Pettigrew J D amp Letelier J C (1994)The oscillatory saccades of birds Motoneural activ-ity and possible functions In B T Fuchs A FV Buttner amp D Zee (Eds) Contemporary ocularmotor and vestibular research A tribute to David ARobinson (pp 480ndash487) Stuttgart Germany ThiemePublishing
Williams P E amp Shapley R M (2007) A dynamicnonlinearity and spatial phase specificity in macaque
V1 neurons Journal of Neuroscience 27 5706ndash5718[PubMed] [Article]
Winterson B J amp Robinson D A (1975) Fixationby the alert but solitary cat Vision Research 151349ndash1352 [PubMed]
Yarbus A L (1967) Eye movements and vision(B Haigh Trans) New York Plenum Press
Zuber B L amp Stark L (1965) Microsaccades and thevelocityltamplitude relationship for saccadic eyemovements Science 150 1459ndash1460 [PubMed]
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 16

Typeofeye
move
ment
Amplitude
Frequency
intersacc
adic
interval
Duratio
nMax
speed
Mean
speed
Conjugate
Study
Mammals
Primate
Microsaccade
84ndash162
min
(meansfor
2monke
ys)
23ndash25
Hz(m
ean
frequenciesfor
2monke
ys)
atleast
8mse
cndash
ndashndash
Horw
itzand
Albrig
ht(2003)
Microsaccade
~40min
(mean)
ndashndash
ndash~30degsec
ndashSnodderly
etal(2001)
Microsaccade
~20min
(mean)
~3ndash5Hz
29mse
c(m
ean)
ndash~30degsec
ndashMartinez-Conde
etal(2000)
Microsaccade
48min
(mean)
03ndash14
Hz
(meansfor
2monke
ys)
25ms(m
ean)
9ndash11
0degsec
(median
40degsec)
~30degsec
ndashBairandOrsquoKeefe
(1998)
Microsaccade
101
min
(median)
0597se
c(m
edian)
20mse
c(m
ean)
ndashndash
ndashLeopold
and
Logothetis
(1998)
Microsaccade
99ndash403
min
(mediansfor
4monke
ys)
08ndash74
sec
(mediansfor
4monke
ys)
ndashndash
ndashndash
Ska
vensk
ietal(1975)
Microsaccade
40min
(mean
minim
um
amplitude
23min)
ndashndash
ndashndash
ndashSteinmanetal(1973)
Drift
ndashndash
ndashndash
6minsec
(meanfor
1monke
y)
ndashBairandOrsquoKeefe
(1998)
Drift
ndashndash
ndashndash
042ndash
1191minsec
(meansfor
4monke
ys)a
ndashSka
vensk
ietal(1975)
Cat
Microsaccade
ldquosomewhat
smallerrdquo
thanin
humans
ldquofarfewerrdquothan
inhumans
ndashndash
ndashNo(m
ostly
monocu
lar
andso
me
binocu
lar)
Hebbard
andMarg
(1960)
Microsaccade
35min
bndash
ndashndash
ndashndash
Pritch
ard
and
Heron(1960)
Drift
ndashndash
ndashndash
147
minsec
(meanfor
3ca
ts)
ndashWintersonand
Robinso
n(1975)
Drift
ndashndash
ndashndash
ndashndash
Hebbard
and
Marg
(1960)
Drift
ldquoUsu
ally
~25min
butca
nexc
eed
2degrdquob
ndashndash
ndash30minsec
ndashPritch
ard
and
Heron(1960)
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 2
Typeofeye
move
ment
Amplitude
Frequency
intersacc
adic
interval
Duratio
nMax
speed
Mean
speed
Conjugate
Study
Tremor
56ndash733
s(ave
rage31s)
b35ndash
65Hz
(ave
rage50Hz)
ndashndash
ndashNo
Hebbard
and
Marg
(1960)
Tremor
04
min
b2ndash
40Hz
(unable
toreco
rdfasterrates)
ndashndash
ndashndash
Pritch
ard
and
Heron(1960)
Rabbit
Drift
ndashndash
ndashndash
25ndash
50minsec
ndashVanderSteenand
Collewijn
(1984)
Drift
e4degb
ndashndash
Q1degsec
(unable
toreco
rdfasterrates)
15
minsec
ndashCollewijn
and
vanderMark
(1972)
Tremor
ldquosmallamplituderdquo
ndashndash
ndashndash
ndashCollewijn
and
vanderMark
(1972)
Birds
Pigeon
Microsaccade
ndashldquovery
lowrdquo
G20mse
cndash
ndashndash
Nye
(1969)
Impulse
Upto
2deg
ndashndash
ndashndash
ndashNye
(1969)
Drift
3ndash5deg
(occ
asionally
exc
eedingthe
limitofthereco
rding
system75
deg)
ndashndash
ndash1ndash
5degsec
ndashNye
(1969)
Oscillation
Upto
seve
rald
eg
(peak-to-peak)
Shortbursts
with
afrequency
of
28ndash35Hz
usu
ally
separatedby
intervals
of1ndash
2se
c
Upto
08
sec
ndashndash
ndashNye
(1969)
Owl
Microsaccade
e1deg
ndashndash
10degsec
ndashndash
Steinbach
and
Money(1973)
Drift
e1deg
ndashndash
ndashndash
ndashSteinbach
and
Money(1973)
Tremor
1min
20Hz
ndashndash
ndashndash
Steinbach
and
Money(1973)
Oscillation
15
deg
Shortbursts
with
afrequency
of20Hz
about
4tim
esaminute
025se
cndash
ndashndash
Steinbach
and
Money(1973)
Reptiles
Turtle
Drift-like
ndashndash
ndashndash
48
T2minsec
ndashGresc
hneretal(2002)
Periodic
Upto
5min
5Hz
ndashUpto
214
degsec
ndashndash
Gresc
hneretal(2002)
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 3
(Engbert 2006 Martinez-Conde 2006 Martinez-Condeet al 2004) However the differences and similarities infixational eye movements across species as well as theirpotential significance have been neglected so far Despitewide-reaching interest in this topic (Martinez-Conde ampMacknik 2007a) a comprehensive comparative reviewhas not been published Here we review the comparativedynamics the physiology and the perceptual consequen-ces of fixational eye movements across all species studiedto date To the best of our knowledge our review includesall Medline-indexed studies (as well as several bookchapters and other non-indexed publications) of fixationaleye movements in non-human vertebrates These span48 years of research from the first studies in the catpublished in 1960 (Hebbard amp Marg 1960 Pritchardamp Heron 1960) to the most recent studies in thesalamander and archer fish published in 2007 and 2008(Baccus Olveczky Manu amp Meister 2008 OlveczkyBaccus amp Meister 2007 Segev Schneidman Goodhouseamp Berry 2007) As a result of our analysis we havegenerated a table (Table 1) that summarizes the physicalparameters of the different types of fixational eye move-ment acrossnon-humanvertebratesWewill relatewheneverpossible fixational eye movements in non-humanvertebrates to human fixational eye movements
Fixational eye movements inmammals
Fixational eye movements in the primate
Fixational eye movements were originally discoveredand characterized in human subjects (Ditchburn ampGinsborg 1952 1953 Ratliff amp Riggs 1950) Humanfixational eye movements comprise three main typesmicrosaccades (called ldquoflicksrdquo in early studies) drift andtremor (Figure 1) Microsaccades are the fastest andlargest of the three types of fixational eye movements(Supplementary Video 1) They travel in a straighttrajectory carrying the retinal image across a range ofseveral dozen to several hundred photoreceptor widthsDrifts are slow curvy motions that occur between micro-saccades and tremor is a very fast (Egrave90 Hz) andextremely small oscillation (about the diameter of a fovealcone) superimposed on drifts (see Tables 1ndash3 of Martinez-Conde et al 2004 for a review of fixational eye move-ment parameters in humans)Fixational eye movements in old-world monkeys are
very similar to human fixational eye movements (Martinez-Conde 2006 Martinez-Conde et al 2000 2002Skavenski Robinson Steinman amp Timberlake 1975Snodderly 1987 Snodderly amp Kurtz 1985) Severalmacaque species have been studied to date Macaca
Typeofeye
move
ment
Amplitude
Frequency
intersacc
adic
interval
Duratio
nMax
speed
Mean
speed
Conjugate
Study
Amphibians
Salamander
Tremor-lik
eAbout
125
min
ndashndash
ndashndash
ndashManteuffe
letal(1977)
Respiratory
Upto
30min
ndashndash
ndashndash
ndashManteuffe
letal(1977)
Fish
Goldfish
Drift
ndashndash
01ndash55
sec
e1degsec
036degsec
NoSim
ulta
neous
inboth
eye
sbut
conve
rgentin
thelight
Mensh
etal(2004)
Drift
Typ
ically
G5deg
ndashndash
ndashndash
NoSim
ulta
neous
inboth
eye
sbut
conve
rgent
Easter(1971)
ArcherFish
Microsaccade
ndashldquorarelyrdquofound
ndashndash
ndashndash
Segevetal(2007)
Drift
ndash02ndash1Hz
133se
ce0
4degsec
ndashndash
Segevetal(2007)
Tremor
01ndash02
deg
5Hz
ndash4degsec
~1degsec
ndashSegevetal(2007)
Table
1Characteris
ticsoffixatio
naleye
move
ments
innon-humanve
rtebrates
acc
ordingto
differentstudies
NoteaCalculatedfrom
horiz
ontalandve
rticalco
mponents
bVectorva
luesfrom
H-componentsaco
nve
rsionfactoroffrac342
hasbeenass
umed
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 4
mulatta (Goffart Quinet Chavane amp Masson 2006Horwitz amp Albright 2003 Leopold amp Logothetis1998 Martinez-Conde 2006 Martinez-Conde et al 20002002 Motter amp Poggio 1984 Skavenski et al 1975Snodderly Kagan amp Gur 2001 Steinman HaddadSkavenski amp Wyman 1973) Macaca fascicularis(Snodderly 1987 Snodderly et al 2001 Snodderly ampKurtz 1985) and Macaca nemestrina (Bair amp OrsquoKeefe1998) with no substantial differences in the spatiotemporalcharacteristics of fixational eye movements (rate velocityand magnitude profiles etc) between them Microsaccadevelocities are parametrically related to microsaccadeamplitudes following the ldquomain sequencerdquo in both ma-caques (Martinez-Conde et al 2000) and humans (Engbert2006 Martinez-Conde 2006 Martinez-Conde et al 2006Zuber amp Stark 1965) Fixational microsaccades have alsobeen observed in the baboon (Papio papio) with ampli-tudes up to 25 degrees (Marchetti Gauthier amp Pellet1983)It is important to note that microsaccades cannot be
defined according to their amplitude alone because smallexploratory or voluntary saccades can be the same size asmicrosaccades Microsaccades can be defined only opera-tionally as the involuntary saccades that are producedwhile the subject attempts to fixate (Martinez-Conde2006) This ldquoattempt to fixaterdquo includes both holding theeyes still and static while foveating the visual target andalso orienting the fovea toward a target It is worth notingthat microsaccade production may require the presence ofa visual andor attentional target (Otero-Millan et al2008)Thus there is no known physical parameter (or
combination of parameters) that separates saccades frommicrosaccades Mounting evidence points toward a com-mon neural generator of saccades and microsaccades(Engbert 2006 Martinez-Conde et al 2004 Otero-Millan et al 2008 Rolfs Kliegl amp Engbert 2008 RolfsLaubrock amp Kliegl 2006 Zuber amp Stark 1965)However one should note that most of the available datain support of a shared microsaccadendashsaccade generatorare indirect To date only two physiological studies havedirectly addressed the question of the oculomotor
mechanisms leading to the generation of microsaccadesIn those studies Van Gisbergen and colleagues foundthat putative motoneurons in the primate abducensnucleus and burst neurons in the nearby pontomedullaryreticular formation were similarly active during saccadesand microsaccades (Van Gisbergen Robinson amp Gielen1981 Van Gisbergen amp Robinson 1977) Future researchshould further explore the neural bases underlying thegeneration of microsaccades and other fixational eyemovementsSimultaneous recordings of microsaccades and neural
responses have been conducted at multiple levels of theprimate visual pathway including the lateral geniculatenucleus (LGN) area V1 and the extrastriate visual cortex(Bair amp OrsquoKeefe 1998 Leopold amp Logothetis 1998Martinez-Conde 2006 Martinez-Conde et al 20002002 Reppas Usrey amp Reid 2002 Snodderly et al2001) Microsaccades are predominantly excitatory at allthese levels leading to firing rate increases in both earlyand higher visual areas The firing rate increases followingmicrosaccades in the LGN and V1 are of visual origin(Figure 2) and result from the microsaccade-induceddisplacement of visual receptive fields over stationarystimuli Microsaccade-driven increases in firing rate tendto be clustered in tight bursts of spikes (Martinez-Conde2006 Martinez-Conde et al 2000 2002) A recent studyhas suggested that primate microsaccades may improvethe efficient sampling of fine spatial detail (Donner ampHemila 2007)Microsaccades counteract visual fading and increase
visibility during fixation in human subjects (Martinez-Conde et al 2006 Troncoso Macknik amp Martinez-Conde 2008) Numerous studies have also reported thathuman microsaccade rates andor directions are modulatedby cognitive processes such as the allocation of spatialattention (Betta amp Turatto 2006 Engbert amp Kliegl2003a 2003b 2004 Galfano et al 2004 Hafed amp Clark2002 Laubrock Engbert Rolfs amp Kliegl 2007 Otero-Millan et al 2008 Turatto Valsecchi Tame Betta 2007Valsecchi Betta amp Turatto 2007 but see also HorowitzFine Fencsik Yurgenson amp Wolfe 2007 and TseSheinberg amp Logothetis 2004)
Figure 1 Cartoon representation of fixational eye movements in humans and primates Microsaccades (straight and fast movements)drifts (curvy slow movements) and tremor (oscillations superimposed on drifts) transport the visual image across the retinal photoreceptormosaic From Martinez-Conde and Macknik (2007b)
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 5
Neural responses to drifts have received considerablyless attention than neural responses to microsaccades Thismay be due to the fact that drifts are more difficult tocharacterize objectively than microsaccades (which aremore easily detected by automatic algorithms that com-bine amplitude and velocity thresholds) Thus drifts areusually identified indirectly as the eye position changesthat occur between microsaccades (Snodderly et al2001) However this method has the potential flaw thatone may unintentionally attribute non-drift-related activity(for instance undetected tremor) to drifts Drifts mayincrease firing in a subset of primate V1 neurons(Snodderly et al 2001) However they generate lessvariability in neuronal responses than a combination ofdrifts and microsaccades (Gur Beylin amp Snodderly1997) No studies to date have recorded primate neuronalresponses in correlation to tremor (but see Hennig ampWorgotter 2007 for the responses of a model macaqueretina to simulated drift and tremor)
Fixational eye movements in the cat
The catrsquos fixational eye movements have been inves-tigated in a handful of studies and their conclusions
remain somewhat controversial In 1960 Hebbard andMarg (1960) found fixational eye movements in the cat tobe quite comparable to those in humans although catmicrosaccades were scarcer and smaller than in humansPritchard and Heron (1960) also found that the catrsquosfixational eye movements included microsaccades driftsand tremor Microsaccades were much rarer in the cat thanin humans however possibly related to the fact that thecat lacks a well-developed fovea Delgado-Garcia delPozo and Baker (1986) also observed some putativemicrosaccades in the alert behaving cat Other studiesfound no microsaccades in the cat (Conway Timberlakeamp Skavenski 1981 Winterson amp Robinson 1975)However in these last studies microsaccades were definedas ldquosaccades smaller than 10 minutes of arcrdquo which iswell below the average microsaccade amplitude found inrecent human and primate studies (see Martinez-Condeet al 2004 for a review of microsaccade parameters inhumans and primates) More recently several studies haveaddressed the contribution of fixational eye movements toneural responses in the catrsquos visual system HennigKerscher Funke and Worgotter (2002) found increasedresponses from neurons in cat areas 17 and 18 duringstimulus motions that mimicked fixational eye move-ments Using a model retina they also found that
1 simulated fixational eye movements increased neu-ral responses and
2 simulated tremor improved spatial acuity
Hennig and Worgotter (2004) later reported that acombination of tremor and microsaccades improved theperformance of simulated ganglion cells in a hyperacuitytask in the retinal periphery (but not in the central retina)Miller Denning George Marshak and Kenyon (2006)also investigated the effects of simulated tremor on amodel of the catrsquos retina and suggested that tremor mayselectively enhance the processing of large stimuli
Fixational eye movements in the rabbit
In contrast to foveate species such as primates (includ-ing humans) and cats (which have an area centralis if nota proper fovea) the rabbit (Oryctolagus cuniculus) dis-plays few spontaneous eye movements especially in theabsence of head motion (Collewijn 1977 Collewijn ampvan der Mark 1972 Fuller 1980 1981 Van der Steen ampCollewijn 1984) In normal visual conditions the rabbitrsquoseyes are very stable but not perfectly immobile Smallamplitude tremor can be observed as well as very slowdrift Microsaccades have not been observed Howeverdrift is occasionally corrected by fast saccadic movements(Collewijn 1977 Collewijn amp van der Mark 1972 Vander Steen amp Collewijn 1984) These differences may berelated to the fact that the rabbit does not have a fovea butan elongated horizontal streak with elevated ganglion cell
Figure 2 Microsaccades drive neural activity in primate area V1Microsaccades lead to firing rate increases in V1 neurons (n = 308)of the awake primate In the absence of visual stimulationmicrosaccades do not increase neural activity in area V1 (n = 37neurons) The resultsVshown here for the average of all neuronsVwere consistent with responses from individual neurons (data notshown) Modified from Martinez-Conde et al (2000 2002)
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 6
counts Olveczky Baccus and Meister (2003) recordedfrom ganglion cells in the rabbit and salamander retinaswhile presenting visual stimuli that were jittered tosimulate fixational eye movements (based on parametersfrom previously published studies by Manteuffel PlasaSommer amp Wess 1977 and Van der Steen amp Collewijn1984) In both species fixational eye movements helpedsegregate a moving foreground from a complex back-ground in a population of ganglion cells
Fixational eye movements inbirds
Fixational eye movements in the pigeon
There are some remarkable differences between fixa-tional eye movements in birds and mammals Nye (1969)identified four types of fixational eye movements in thepigeon (Columba livia) flicks (ie microsaccades)impulse movements drift and oscillations No tremorwas detected Microsaccade frequency was very lowrelative to humans Impulse movements occurred morefrequently usually between bursts of oscillations withamplitudes ranging up to 2 degrees Drift velocitiesranged from 1 to 5 degrees per second Both micro-saccades and drifts occurred in response to movingstimuli Oscillations occurred in short bursts at rates ofabout 30 Hz and could be quite large (up to severaldegrees of amplitude) Bloch Rivaud and Martinoya(1984) reported microsaccades drifts and oscillationsldquosimilar to those described by Nyerdquo but none of thesewere analyzed in detailCyclotorsional oscillations are an integral part of avian
saccades (Wallman Pettigrew amp Letelier 1994) Thusone might speculate that oscillations are associated withavian microsaccades as well especially if saccades andmicrosaccades share a common generator (Engbert 2006Otero-Millan et al 2008 Rolfs et al 2008 2006 Zuberamp Stark 1965) However this possibility has not yet beenaddressed in the literature The function of oscillations isunknown (Carpenter 1988) but one hypothesis is thatthey serve to facilitate the delivery of arterial blood to theocular capillary bed (Pettigrew Wallman amp Wildsoet1990 Wallman et al 1994)
Fixational eye movements in the owl
Steinbach and Money (1973) recorded fixational eyemovements in the great horned owl (Bubo virginianus)a species previously thought incapable of moving itseyes The owlrsquos fixational eye movements includeflicks or microsaccades drift tremor and oscillations
Microsaccades seemed correlated to visual events ofinterest to the owl they were 1 deg or less in amplitudeand had peak velocities of about 10 degsec Drifts wereslow motions of the eye in between fixations withamplitudes of 1 degree or less Tremor amplitude wasabout 1 arcmin with frequencies around 20 Hz High-frequency oscillations (25 Hz) of short duration (025 sec)were very similar to those previously found in the pigeon(Nye 1969) No studies to date have examined theperceptual or physiological effects of fixational eyemovements in any bird species
Fixational eye movements inreptiles
Fixational eye movements in the turtle
Greschner Bongard Rujan and Ammermuller (2002)identified two types of fixational eye movements in theturtle (Pseudemys scripta elegans) which they calledldquodrift-likerdquo and ldquoperiodicrdquo Drift-like motions were smalland slow (with velocities comparable to human drift) andwere superimposed by larger and faster periodic componentsPeriodic motion had retinal amplitudes on the order of thediameter of a photoreceptor which are comparable to thoseof human tremor On the other hand periodic motionrsquosfrequency (5 Hz) velocity and orderly relationship betweenvelocity and amplitude are closer to those of human micro-saccades (see Martinez-Conde et al 2004 for a review oftremor and microsaccade parameters in humans)Simulated periodic motion generated strong synchronous
firing in the turtlersquos retina whereas simulated drift-likemotion had little effect (Figure 3) These results agree withprevious physiological and modeling studies in the primatevisual system in which strong neural transients wereobserved in response to microsaccades (Donner amp Hemila2007 Martinez-Conde 2006 Martinez-Conde et al 20002002) Such neural transients may underlie the behavior ofcortical neurons as coincidence detectors (ShelleyMcLaughlin Shapley amp Wielaard 2002 Williams ampShapley 2007) Moreover neural transients to stimulionsets and terminations (similar to those produced bymicrosaccades in the primate visual system Martinez-Conde 2006 Martinez-Conde et al 2000 2002) havebeen related to target visibility in visual masking para-digms (Macknik amp Livingstone 1998 Macknik ampMartinez-Conde 2004 Macknik Martinez-Conde ampHaglund 2000)In the turtle retina periodic motion-driven neurons with
receptive fields located along contrast borders weremoreover synchronized and reliably indicated the preced-ing motions The authors proposed that this synchroniza-tion of retinal activity could be used by the brain to
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 7
improve the estimation of stimulus features such asspatial frequency (Greschner et al 2002) This findingcould have a parallel in the primate visual system Donnerand Hemila (2007) recently proposed that microsaccadessignificantly ldquore-sharpenrdquo the image and improve spatialresolution in primates Rucci Iovin Poletti and Santini(2007) also found that fixational eye movements enhancefine spatial detail in human subjects
Fixational eye movements in the chameleon
Fixational eye movements in the chameleon have notbeen studied in detail However Gioanni and colleaguesfound that the chameleon (Chamaleo chamaleo) makessaccades during fixation The authors speculated that theseprobably correspond to fixational microsaccades found inother foveate species (Gioanni Bennis amp Sansonetti1993)
Fixational eye movements inamphibians
Fixational (and more generally spontaneous) eye move-ments vary widely among amphibians The frog (anafoveate amphibian) shows a general lack of eye andhead movements except for those that stabilize the retinalimage Dieringer and colleagues (Dieringer amp Daunicht1986 Dieringer amp Precht 1982 Dieringer Precht ampBlight 1982) found that the grass frog (Rana temporaria)produces no spontaneous eye movements in the absenceof head movements with the possible exception of tremor(similarly to the rabbit another afoveate species Van derSteen amp Collewijn 1984) It is likely that the frogrsquos lack ofeye movements leads to fast visual fading of all stationaryobjects in its field of view (Ditchburn amp Ginsborg 1952Yarbus 1967) Lettvin Maturana MsCulloch and Pitts
Figure 3 Fixational eye movements increase retinal activity in the turtle (A) Continuous recording from 54 retinal ganglion cells in theturtle during darkness after switching on a stationary grating and during wobbling of this grating The grating was wobbled to simulateperiodic (ie tremor-like) eye movements (B) Responses of a single ganglion cell to a drifting contrast border of velocity comparable toocular drifts (C) When the drift is superimposed with wobbling the responses of the cell shown in (B) increase dramatically FromGreschner et al (2002) and modified by Martinez-Conde et al (2004)
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 8
(1968) stated that a frog ldquowill starve to death surroundedby food if it is not movingrdquo (but see Dieringer amp Daunicht1986 for a counterpoint) Small head motions or eyemovements correlated with respiratory motions maydiminish fading to some extent (Dieringer amp Precht1982 Schipperheyn 1963 1965 1967)As a side note the frogrsquos respiratory eye movements
play no role in the respiratory function ldquoin the course ofeach respiratory cycle increase of pressure in the frogrsquosbuccal cavity protrudes the eye and causes the opticalaxis to deviate upwards over 25ndash35 minutes of arcrdquo(Schipperheyn 1963) That is respiratory eye movementsare merely synchronized with respirationAutrum (1959) found no evidence of respiratory eye
movements in the frog Ewert andBorchers (1974) observedthat stationary stimuli elicit no responses from visualneurons in the common toad (Bufo Bufo L) regardless ofrespiratory pressure changes in the buccal cavity
Fixational eye movements in the salamander
Manteuffel et al (1977) identified two types of fixa-tional eye movements in salamanders tremor-like andrespiratory eye movements Tremor-like is a rapid move-ment of small amplitude Torsional eye movements resultfrom the animalrsquos breathing (also found in the frog bySchipperheyn 1963) Additionally flicks are occasionallyobserved which rapidly change the position of gaze It isunclear whether such flicks may generally correspond towhat we now call microsaccades andor to voluntarysaccades Based on these observations the authors pro-posed that unlike frogs and toads (Autrum 1959 Ewert ampBorchers 1974 Lettvin et al 1968) salamanders cancontinuously perceive stationary objects (Manteuffel et al1977) Fixational eye movements in the salamander (Tigersalamander Ambystoma tigrinum) may also help segre-gate object and background motion in the retina(Olveczky et al 2003 see also Fixational eye movementsin the rabbit section) and even work to emphasize novelvisual features (Olveczky et al 2007 see Baccus et al2008 for details on the retinal neurons and synapses thatmay underlie the computation of differential motionbetween an object and its background)
Fixational eye movements in fish
Spontaneous eye movements in the goldfish
Eye movements during fixation have been studied inboth afoveate (goldfish Carassius auratus) and foveate(archer fish Toxotes chatareus) fish species The goldfisha long-standing model organism in vision and oculomotor
research (Aksay et al 2003 2007 Cabrera Torres PasaroPastor amp Delgado-Garcia 1992 Pastor Torres Delgado-Garcia amp Baker 1991 Salas Navarro Torres ampDelgado-Garcia 1992 Torres Pastor Cabrera Salas ampDelgado-Garcia 1992) produces three types of sponta-neous eye movements saccades stretch and drift (Easter1971 Mensh Aksay Lee Seung amp Tank 2004) Weshould note that in the motionless goldfish eye move-ments are very infrequent Those that do occur consistmainly of saccades slow drifts are very rare (Easter Johnsamp Heckenlively 1974) During saccades (identified bythresholding the eye velocity at 5 degsec) both eyes moverapidly simultaneously and usually in the same direction(Mensh et al 2004) Stretches occur every 2ndash3 min and arecharacterized by a brief simultaneous large excursion ofboth eyes to extreme temporal positions (Easter 1971Mensh et al 2004) The function of stretches is not known(Mensh et al 2004) Because fixations were defined asthose periods that were free of saccadic and stretchmovements (Easter 1971) only drifts (from the threetypes of spontaneous movements) can technically beconsidered ldquofixational eye movementsrdquo However thegoldfish saccadic main sequence includes saccades withequivalent amplitudes and velocities to those found inhuman microsaccades (Mensh et al 2004 Zuber amp Stark1965) Indeed microsaccades have been identified in other(foveate) fish species although they are rarer than inhumans (Segev et al 2007) Small saccades in thegoldfish tend to be divergent whereas large saccadestend to be convergent (Mensh et al 2004) Drift occurssimultaneously in both eyes usually in the samedirection Drift velocities are almost exclusively e1 degsec(Mensh et al 2004) No tremor has been identified in thegoldfish
Spontaneous eye movements in teleost fish
Spontaneous eye movements have been studied in theNew Zealand parore (Girella tricuspidata MontgomeryMcVean amp McCarthy 1983) The pattern of eye move-ments was similar to that previously described in goldfish(Easter 1971 Hermann amp Constantine 1971)
Fixational eye movements in the archer fish
A recent study in the archer fish (Segev et al 2007) hasprovided the first perceptual and physiological correlatesof fixational eye movements in any fish species Archerfish have remarkable visual abilities they shoot downinsects resting on foliageVas well as stationary targetscovering a single retinal photoreceptorVwith squirts ofwater from their mouths they predict the location atwhich the insect will splash down in G100 msec and theydeduce the absolute size of an object presented at differentviewing distances (Supplementary Videos 2 and 3) Segev
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 9
et al (2007) identified three types of fixational eyemovements in the archer fish microsaccades drift andtremor Microsaccades were rare and were not analyzed indetail Drift velocities were e040 degsec Tremor was ahigh-frequency oscillatory movement (the primary com-ponent was 5 Hz) with peak-to-peak amplitude of 01ndash02 deg (corresponding to an image displacement of 1ndash2photoreceptors) and velocity of about 1 degsec (Figure 4)Whereas the amplitude and velocity of tremor in thearcher fish is comparable to those in humans its frequencyis much lower (5 Hz vs 940 Hz) Interestingly archer fishtremor is very similar both in amplitude (photoreceptorwidths covered) as well as in frequency (5 Hz) to thetremor-like periodic motion in the turtle (discussedearlier Greschner et al 2002)Segev et al (2007) trained archer fish to perform a size
discrimination task the fish had to distinguish a medium-sized target from larger and smaller distracters (theyindicated their choice by shooting a jet of water) To relatethis behavior to responses from retinal neurons theauthors recorded from populations of retinal ganglioncells while presenting objects of different sizes moved tosimulate saccades drifts and tremor Drifts and tremorelicited weak neuronal responses that were not veryinformative about the spatial structure of the stimulus(although they did provide some information about target
size in agreement with the results of Greschner et al2002 in the turtle) Moreover performance was similar fortremor alone and for tremor plus drift indicating that driftplays a negligible role in revealing spatial structure (alsoin agreement with Greschner et al 2002)When present the firing rate was phase-locked to
tremor Saccades were more effective than drifts or tremorin driving informative responses from the ganglion cells(consistent with theoretical and physiological studies ofmicrosaccade-driven neuronal activity in the primateretina and LGN Donner amp Hemila 2007 Martinez-Conde et al 2002) Segev et al (2007) proposed that inprimates both saccades and microsaccades (which arerare in the archer fish) may contribute to size discrim-ination tasks as well as enhance spatial detail whereasdrifts and tremor by themselves may not be sufficient toimprove visual perception However it is important toremember that central vision in humans can be maintainedby tremor andor drift even when microsaccades aresuppressed during strict fixation (Martinez-Conde 2006Martinez-Conde et al 2004 2006) On the other handone cannot rule out the possibility that if one couldcompletely eliminate drifts and tremor microsaccadesmight then suffice to sustain both central and peripheralvision in primates and humans Future experiments areneeded to further explore these ideas
Figure 4 Archer fish eye movements (A) Example trace of eye position as a function of time as measured by search coil (B) Example ofa single 15-s fixation superimposed on a grid representing the photoreceptor mosaic The back and forth movement is caused by tremor(C) Distribution of fixation durations (taken as time interval between saccades) (D) Power spectrum of horizontal fixational eyemovements (E) Distribution of tremor amplitude (data taken from 1 animal in a 15-min session) Modified from Segev et al (2007)
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 10
Discussion
The vast majority of eye movement studies conductedin non-human species to date have not analyzed eyeposition data in the range of fixational eye movementsThus eye movements during fixation have been reportedin only a handful of vertebrates Most of these studieshave concentrated solely on the physical parameters ofthe fixational eye movements without addressing theirphysiological and perceptual consequences Fortunatelythis trend has started to reverse in recent years with agrowing list of papers addressing the effects of non-human fixational eye movements in physiology andperception This is an important development A largeamount of all psychophysical and physiological visualresearch has been conducted during conditions of visualfixation Therefore understanding the precise physiolog-ical and perceptual contributions of fixational eye move-ments may be critical to the interpretation of previous andfuture vision researchThere are some interesting differences and similarities
in the types of fixational eye movements produced bydifferent species For instance ocular tremor is possiblyubiquitous to all vertebrates The fact that tremor has notbeen observed in all species studied could be due to thetechnical difficulty of recording tremor accurately (ietremor amplitudes and frequencies are usually in the rangeof the recording systemrsquos noise) Drift has also beenrecorded in most vertebrates and it may be as omnipre-sent as tremor Conversely microsaccades seem mostimportant in species with foveal vision (ie primates andhumans) whereas they are scarce or absent in afoveatespecies (ie rabbits frogs goldfish) A link betweenmicrosaccades and foveal vision may reflect the two-pronged functional significance of microsaccades inhumans and primates first microsaccades may benecessary to correct the slow drift of the eye and bringback the objectregion of interest to the retinal area wherephotoreceptor density is highest (Cornsweet 1956) Thecorrective role of human microsaccades has been demon-strated in recent studies (Engbert amp Kliegl 2004 Engbertamp Mergenthaler 2006) Second the dynamics of micro-saccades themselves may improve spatial resolution(Donner amp Hemila 2005)In reviewing the patterns of fixational eye movements
across vertebrates it is tempting to draw parallels withother sensory modalities Fixational eye movementsrefresh the retinal images during fixation and thus preventneural adaptation and visual fading But this mechanism isnot circumscribed to the visual system Sniffs in rodentolfaction discretely sample sensory information every200ndash300 msec and thus are similar in their temporaldynamics to primate saccades (Otero-Millan et al 2008Uchida Kepecs amp Mainen 2006) and microsaccades(Otero-Millan et al 2008) A similar mode of discretesampling may also be at play when objects are recognized
through tactile information for instance if we sweep ourfingertips over an objectrsquos surface with our eyes closed orwhen blind individuals read Braille scriptOne intriguing possibility that merits further experi-
ments and analyses is the potential correlation between thespatiotemporal parameters of fixational eye movements(such as those summarized in Table 1) and the adaptationproperties of visual neurons across species For instancethe greater ocular stability of some species (rabbitgoldfish frog etc) versus others (primates) could beassociated with some particular adaptive properties oftheir visual cells Thus future comparative studies shouldpay attention to the response properties of visual cellstriggered by fixational eye movements in the presence ofstationary stimuli Other potential factors underlying thevariation in fixational eye movements across species couldbe differences in oculomotor control at the level of the eyemuscles or perhaps differences in retinal organizationFuture research will further improve our understanding
of fixational eye movements and the neural activity thatmaintains the visibility of stationary visual scenes inhuman and non-human species One interesting questionthat remains to be addressed concerns the role of tremor invision and visual physiology Because tremor frequenciesare well over the flicker fusion frequencies in humans ithas been argued that tremor of the visual image would beineffective as a stimulus (Ditchburn 1955 Gerrits ampVendrik 1970 Sharpe 1972) However recent studieshave found simulated tremor to enhance visual processingand improve visual acuity in a model retina (Hennig et al2002 Miller et al 2006) Follow-up research should beaimed to replicate these findings in actual neurons withcorrelated perceptual reports One should also note thatearly visual neurons can follow high-frequency flickeringthat surpasses the perceptual threshold for flicker fusion(Martinez-Conde et al 2002) Thus high-frequencytremor could be adequate to maintain activity in the earlyvisual system which then may lead to visual perception inhigher visual areas Another significant question thatremains unanswered concerns the oculomotor bases ofthe various types of fixational eye movements acrossspecies To date there is too little information on thisimportant topic to draw a conclusion
Conclusions
We have reviewed the parameters of fixational eyemovements in non-human mammals birds reptilesamphibians and fish as well as their consequences forphysiology and perception Fixational eye movementsand microsaccades in particular appear to be mostimportant in foveate vs afoveate species Animals lackingfixational eye movements such as afoveate frogs andtoads may be incapable of seeing stationary objects
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 11
during visual fixation Future research should furtherexplore the perceptual effects of fixational eye movementsand determine the oculomotor mechanisms responsible fortheir generation across species
Acknowledgments
We thank Mona Stewart and Isabel Gomez-Caraballo fortechnical assistance Jorge Otero-Millan for programmingassistance and Dr Xoana G Troncoso for comments on themanuscript This study was funded through grants fromthe Barrow Neurological Foundation (to SMC and SLM)the National Science Foundation (Award 0643306 toSMC) the Arizona Biomedical Research Commission(Award 07-102 to SMC) and Science Foundation Arizona(SLM)
Commercial relationships noneCorresponding author Susana Martinez-Conde PhDEmail smartneuralcorrelatecomAddress Barrow Neurological Institute 350 W ThomasRd Phoenix AZ 85013 USA
References
Aksay E Major G Goldman M S Baker R SeungH S amp Tank D W (2003) History dependence ofrate covariation between neurons during persistentactivity in an oculomotor integrator Cerebral Cortex13 1173ndash1184 [PubMed] [Article]
Aksay E Olasagasti I Mensh B D Baker RGoldman M S amp Tank D W (2007) Functionaldissection of circuitry in a neural integrator NatureNeuroscience 10 494ndash504 [PubMed]
Autrum H (1959) Das Fehlen unwillkurlicher Augenbe-wegungen beim Frosch Naturwissenschaften 46435
Baccus S A Olveczky B P Manu M amp Meister M(2008) A retinal circuit that computes object motionJournal of Neuroscience 28 6807ndash6817 [PubMed]
Bair W amp OrsquoKeefe L P (1998) The influence offixational eye movements on the response of neuronsin area MT of the macaque Visual Neuroscience 15779ndash786 [PubMed]
Betta E amp Turatto M (2006) Are you ready I can tellby looking at your microsaccades Neuroreport 171001ndash1004 [PubMed]
Bloch S Rivaud S amp Martinoya C (1984) Compar-ing frontal and lateral viewing in the pigeon IIIDifferent patterns of eye movements for binocularand monocular fixation Behavioural Brain Research13 173ndash182 [PubMed]
Cabrera B Torres B Pasaro R Pastor A M ampDelgado-Garcia J M (1992) A morphological studyof abducens nucleus motoneurons and internuclearneurons in the goldfish (Carassius auratus) BrainResearch Bulletin 28 137ndash144 [PubMed]
Carpenter R H S (1988) Movements of the eyes(2nd ed) Pion London
Collewijn H (1977) Eye- and head movements infreely moving rabbits The Journal of Physiology266 471ndash498 [PubMed] [Article]
Collewijn H amp van der Mark F (1972) Ocular stabilityin variable visual feedback conditions in the rabbitBrain Research 36 47ndash57 [PubMed]
Conway J L Timberlake G T amp Skavenski A A(1981) Oculomotor changes in cats reared withoutexperiencing continuous retinal image motion Exper-imental Brain Research 43 229ndash232 [PubMed]
Cornsweet T N (1956) Determination of the stimuli forinvoluntary drifts and saccadic eye movements Jour-nal of the Optical Society of America 46 987ndash993[PubMed]
Delgado-Garcia J M del Pozo F amp Baker R (1986)Behavior of neurons in the abducens nucleus of thealert catVII Internuclear neurons Neuroscience 17953ndash973 [PubMed]
Dieringer N amp Daunicht W J (1986) Image fadingVAproblem for frogs Naturwissenschaften 73 330ndash332[PubMed]
Dieringer N amp Precht W (1982) Compensatory headand eye movements in the frog and their contributionto stabilization of gaze Experimental BrainResearch 47 394ndash406 [PubMed]
Dieringer N Precht W amp Blight A R (1982)Resetting fast phases of head and eye and theirlinkage in the frog Experimental Brain Research 47407ndash416 [PubMed]
Ditchburn R W (1955) Eye-movements in relation toretinal action Optica Acta 1 171ndash176
Ditchburn R W amp Ginsborg B L (1952) Vision with astabilized retinal image Nature 170 36ndash37[PubMed]
Ditchburn R W amp Ginsborg B L (1953) Involuntaryeye movements during fixation The Journal ofPhysiology 119 1ndash17 [PubMed] [Article]
Donner K amp Hemila S (2007) Modelling the effect ofmicrosaccades on retinal responses to stationarycontrast patterns Vision Research 47 1166ndash1177[PubMed]
Donner K K amp Hemila S (2005) The possible purposeof microsaccades Paper presented at the EuropeanConference on Visual Perception A Coruna Spain
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 12
Easter S S Jr (1971) Spontaneous eye movements inrestrained goldfish Vision Research 11 333ndash342[PubMed]
Easter S S Jr Johns P R amp Heckenlively D R(1974) Horizontal compensatory eye movements ingoldfish (Carassius auratus) I The normal animalJournal of Comparative Physiology 92 23ndash35
Engbert R (2006) Microsaccades A microcosm forresearch on oculomotor control attention and visualperception Progress Brain Research 154 177ndash192[PubMed]
Engbert R amp Kliegl R (2003a) Binocular coordinationin microsaccades In R R J Hyona amp H Deubel(Eds) The mindrsquos eyes Cognitive and appliedaspects of eye movements (pp 103ndash117) OxfordUK Elsevier
Engbert R amp Kliegl R (2003b) Microsaccades uncoverthe orientation of covert attention Vision Research43 1035ndash1045 [PubMed]
Engbert R amp Kliegl R (2004) Microsaccades keep theeyesrsquo balance during fixation Psychological Science15 431ndash436 [PubMed]
Engbert R amp Mergenthaler K (2006) Microsaccadesare triggered by low retinal image slip Proceedingsof the National Academy of Sciences of the UnitedStates of America 103 7192ndash7197 [PubMed][Article]
Ewert J P amp Borchers H W (1974) Antwort vonretinalen Ganglienzellen bei freibeweglichen Kroten(Bufo Bufo L) Journal of Comparative and Physi-ology 92 117ndash130
Fuller J H (1980) Linkage of eye and head movementsin the alert rabbit Brain Research 194 219ndash222[PubMed]
Fuller J H (1981) Eye and head movements duringvestibular stimulation in the alert rabbit BrainResearch 205 363ndash381 [PubMed]
Galfano G Betta E amp Turatto M (2004) Inhibition ofreturn in microsaccades Experimental BrainResearch 159 400ndash404 [PubMed]
Gerrits H J amp Vendrik A J (1970) Artificial move-ments of a stabilized image Vision Research 101443ndash1456 [PubMed]
Gioanni H Bennis M amp Sansonetti A (1993) Visualand vestibular reflexes that stabilize gaze in thechameleon Visual Neuroscience 10 947ndash956[PubMed]
Goffart L Quinet J Chavane F amp Masson G S(2006) Influence of background illumination onfixation and visually guided saccades in the rhesusmonkey Vision Research 46 149ndash162 [PubMed]
Greschner M Bongard M Rujan P amp Ammermuller J(2002) Retinal ganglion cell synchronization by fixa-tional eye movements improves feature estimationNature Neuroscience 5 341ndash347 [PubMed]
Gur M Beylin A amp Snodderly D M (1997)Response variability of neurons in primary visualcortex (V1) of alert monkeys Journal of Neuro-science 17 2914ndash2920 [PubMed] [Article]
Hafed Z M amp Clark J J (2002) Microsaccades as anovert measure of covert attention shifts VisionResearch 42 2533ndash2545 [PubMed]
Hebbard F W amp Marg E (1960) Physiologicalnystagmus in the cat Journal of the Optical Societyof America 50 151ndash155 [PubMed]
Hennig M A Kerscher N J Funke K amp Worgotter F(2002) Stochastic resonance in visual cortical neuronsDoes the eye-tremor actually improve visual acuityNeurocomputing 44ndash46 115ndash120
Hennig M H amp Worgotter F (2004) Eye micro-movements improve stimulus detection beyond theNyquist limit in the peripheral retina In Advances inNeural Information Processing Systems (NIPS2003)(vol 16) Denver CO MIT Press
Hennig M H amp Worgotter F (2007) Effects offixational eye movements on retinal ganglion cellresponses A modelling study Frontiers in Computa-tional Neuroscience 1 2 [PubMed] [Article]
Hermann H T amp Constantine M M (1971) Eyemovements in the goldfish Vision Research 11313ndash331 [PubMed]
Horowitz T S Fine E M Fencsik D E Yurgenson Samp Wolfe J M (2007) Fixational eye movements arenot an index of covert attention PsychologicalScience 18 356ndash363 [PubMed]
Horwitz G D amp Albright T D (2003) Short-latencyfixational saccades induced by luminance incrementsJournal of Neurophysiology 90 1333ndash1339[PubMed] [Article]
Laubrock J Engbert R Rolfs M amp Kliegl R(2007) Microsaccades are an index of covertattention Commentary on Horowitz Fine FencsikYurgenson and Wolfe (2007) Psychological Sci-ence 18 364ndash366 discussion 367ndash368 [PubMed]
Leopold D A amp Logothetis N K (1998) Micro-saccades differentially modulate neural activity in thestriate and extrastriate visual cortex ExperimentalBrain Research 123 341ndash345 [PubMed]
Lettvin J Y Maturana H R MsCulloch W S amp PittsW H (1968) What the frogrsquos eye tells the frogrsquosbrain In W C C a M Balaban (Ed) The mind
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 13
Biological approaches to its functions (pp 233ndash258)John Wiley and Sons London
Macknik S L amp Livingstone M S (1998) Neuronalcorrelates of visibility and invisibility in the primatevisual system Nature Neuroscience 1 144ndash149[PubMed]
Macknik S L amp Martinez-Conde S (2004) The spatialand temporal effects of lateral inhibitory networksand their relevance to the visibility of spatiotemporaledges Neurocomputing 58ndash60 775ndash782
Macknik S L Martinez-Conde S amp Haglund M M(2000) The role of spatiotemporal edges in visibilityand visual masking Proceedings of the NationalAcademy of Sciences of the United States of America97 7556ndash7560 [PubMed] [Article]
Manteuffel G Plasa L Sommer T J amp Wess O(1977) Involuntary eye movements in salamandersNaturwissenschaften 64 533ndash534 [PubMed]
Marchetti E Gauthier G M amp Pellet J (1983)Cerebellar control of eye movements studied withinjection of harmaline in the trained baboon ArchivesItaliennes de Biologies 121 1ndash17 [PubMed]
Martinez-Conde S (2006) Fixational eye movements innormal and pathological vision Progress in BrainResearch 154 151ndash176 [PubMed]
Martinez-Conde S amp Macknik S L (2007a) Bobbingand perceiving Scientific American 297 16
Martinez-Conde S amp Macknik S L (2007b) Windowson the mind Scientific American 297 56ndash63[PubMed]
Martinez-Conde S Macknik S L amp Hubel D H(2000) Microsaccadic eye movements and firing ofsingle cells in the striate cortex of macaque monkeysNature Neuroscience 3 251ndash258 [PubMed]
Martinez-Conde S Macknik S L amp Hubel D H(2002) The function of bursts of spikes during visualfixation in the awake primate lateral geniculate nucleusand primary visual cortex Proceedings of theNational Academy of Sciences of the United Statesof America 99 13920ndash13925 [PubMed] [Article]
Martinez-Conde S Macknik S L amp Hubel D H(2004) The role of fixational eye movements invisual perception Nature Reviews Neuroscience 5229ndash240 [PubMed]
Martinez-Conde S Macknik S L Troncoso X G ampDyar T A (2006) Microsaccades counteract visualfading during fixation Neuron 49 297ndash305[PubMed] [Article]
Mensh B D Aksay E Lee D D Seung H S ampTank D W (2004) Spontaneous eye movements ingoldfish Oculomotor integrator performance plasti-
city and dependence on visual feedback VisionResearch 44 711ndash726 [PubMed]
Miller J A Denning K S George J S MarshakD W amp Kenyon G T (2006) A high frequencyresonance in the responses of retinal ganglion cells torapidly modulated stimuli A computer model VisualNeuroscience 23 779ndash794 [PubMed]
Montgomery J C McVean A R amp McCarthy D(1983) The effects of lowered temperature onspontaneous eye movements in a teleost fish Com-parative Biochemistry and Physiology A Compara-tive Physiology 75 363ndash368 [PubMed]
Motter B C amp Poggio G F (1984) Binocularfixation in the rhesus monkey Spatial and temporalcharacteristics Experimental Brain Research 54304ndash314 [PubMed]
Murakami I amp Cavanagh P (1998) A jitter after-effectreveals motion-based stabilization of vision Nature395 798ndash801 [PubMed]
Nye P W (1969) The monocular eye movements of thepigeon Vision Research 9 133ndash144 [PubMed]
Olveczky B P Baccus S A amp Meister M (2003)Segregation of object and background motion in theretina Nature 423 401ndash408 [PubMed]
Olveczky B P Baccus S A amp Meister M (2007)Retinal adaptation to object motion Neuron 56689ndash700 [PubMed] [Article]
Otero-Millan J Troncoso X Macknik S L Serrano-Pedraza I amp Martinez-Conde S (2008) Saccadesand microsaccades during visual fixation explorationand search Foundations for a common saccadicgenerator Journal of Vision 8(14)21 1ndash18 httpjournalofvisionorg81421 doi10116781421[PubMed] [Article]
Pastor A M Torres B Delgado-Garcia J M ampBaker R (1991) Discharge characteristics of medialrectus and abducens motoneurons in the goldfishJournal of Neurophysiology 66 2125ndash2140 [PubMed]
Pettigrew J D Wallman J amp Wildsoet C F (1990)Saccadic oscillations facilitate ocular perfusion fromthe avian pecten Nature 343 362ndash363 [PubMed]
Pritchard R M amp Heron W (1960) Small eye move-ments of the cat Canadian Journal of Psychology14 131ndash137 [PubMed]
Ratliff F amp Riggs L A (1950) Involuntary motions ofthe eye during monocular fixation Journal of Exper-imental Psychology 40 687ndash701 [PubMed]
Reppas J B UsreyWM amp Reid R C (2002) Saccadiceye movements modulate visual responses in thelateral geniculate nucleus Neuron 35 961ndash974[PubMed] [Article]
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 14
Riggs L A amp Ratliff F (1952) The effects of counter-acting the normal movements of the eye Journal ofthe Optical Society of America 42 872ndash873
Rolfs M Kliegl R amp Engbert R (2008) Toward amodel of microsaccade generation The case ofmicrosaccadic inhibition Journal of Vision 8(11)51ndash23 httpjournalofvisionorg8115 doi1011678115 [PubMed] [Article]
Rolfs M Laubrock J amp Kliegl R (2006) Shorteningand prolongation of saccade latencies followingmicrosaccades Experimental Brain Research 169369ndash376 [PubMed]
Rucci M Iovin R Poletti M amp Santini F (2007)Miniature eye movements enhance fine spatial detailNature 447 851ndash854 [PubMed]
Salas C Navarro F Torres B amp Delgado-Garcia J M(1992) Effects of diazepam and D-amphetamine onrhythmic pattern of eye movements in goldfishNeuroreport 3 131ndash134 [PubMed]
Schipperheyn J J (1963) Respiratory eye movementsand perception of stationary objects in the frog ActaPhysiologica et Pharmacologica Neerlandica 12157ndash159 [PubMed]
Schipperheyn J J (1965) Contrast detection in frogrsquosretina Acta Physiologica et Pharmacologica Neer-landica 13 231ndash277 [PubMed]
Schipperheyn J J (1967) Influence of eye movements onretinal ganglion cell activity in frogs Acta Physi-ologica et Pharmacologica Neerlandica 14 506[PubMed]
Segev R Schneidman E Goodhouse J amp Berry M J2nd (2007) Role of eye movements in the retinalcode for a size discrimination task Journal ofNeurophysiology 98 1380ndash1391 [PubMed] [Article]
Sharpe C R (1972) The visibility and fading of thinlines visualized by their controlled movement acrossthe retina The Journal of Physiology 222 113ndash134[PubMed] [Article]
Shelley M McLaughlin D Shapley R amp Wielaard J(2002) States of high conductance in a large-scalemodel of the visual cortex Journal of ComputationalNeuroscience 13 93ndash109 [PubMed]
Skavenski A A Robinson D A Steinman R M ampTimberlake G T (1975) Miniature eye movementsof fixation in rhesus monkey Vision Research 151269ndash1273 [PubMed]
Snodderly D M (1987) Effects of light and darkenvironments on macaque and human fixational eyemovements Vision Research 27 401ndash415 [PubMed]
Snodderly D M Kagan I amp Gur M (2001) Selectiveactivation of visual cortex neurons by fixational eyemovements Implications for neural coding VisualNeuroscience 18 259ndash277 [PubMed]
Snodderly D M amp Kurtz D (1985) Eye position duringfixation tasks Comparison of macaque and humanVision Research 25 83ndash98 [PubMed]
Steinbach M J amp Money K E (1973) Eye movementsof the owl Vision Research 13 889ndash891 [PubMed]
Steinman R M Haddad G M Skavenski A A ampWyman D (1973) Miniature eye movement Science181 810ndash819 [PubMed]
Torres B Pastor A M Cabrera B Salas C ampDelgado-Garcia J M (1992) Afferents to theoculomotor nucleus in the goldfish (Carassius aur-atus) as revealed by retrograde labeling with horse-radish peroxidase Journal of Comparative Neurology324 449ndash461 [PubMed]
Troncoso X G Macknik S L amp Martinez-Conde S(2008) Microsaccades counteract perceptual filling-inJournal of Vision 8(14)15 1ndash9 httpjournalofvisionorg81415 doi10116781415 [Article]
Troncoso X G Macknik S L Otero-Millan J ampMartinez-Conde S (2008) Microsaccades driveillusory motion in the Enigma illusion Proceedingsof the National Academy of Sciences of the UnitedStates of America 105 16033ndash16038 [PubMed]
Tse P U Sheinberg D S amp Logothetis N K (2004)The distribution of microsaccade directions need notreveal the location of attention Psychological Science15 708ndash710
Turatto M Valsecchi M Tame L amp Betta E (2007)Microsaccades distinguish between global and localvisual processing Neuroreport 18 1015ndash1018[PubMed]
Uchida N Kepecs A amp Mainen Z F (2006) Seeing ata glance smelling in a whiff Rapid forms ofperceptual decision making Nature Reviews Neuro-science 7 485ndash491 [PubMed]
Valsecchi M Betta E amp Turatto M (2007) Visualoddballs induce prolonged microsaccadic inhibitionExperimental Brain Research 177 196ndash208[PubMed]
van Dam L C amp van Ee R (2006) Retinal image shiftsbut not eye movements per se cause alternations inawareness during binocular rivalry Journal of Vision6(11)3 1172ndash1179 httpjournalofvisionorg6113doi1011676113 [PubMed] [Article]
Van der Steen J amp Collewijn H (1984) Ocular stabilityin the horizontal frontal and sagittal planes in therabbit Experimental Brain Research 56 263ndash274[PubMed]
Van Gisbergen J A Robinson D A amp Gielen S(1981) A quantitative analysis of generation ofsaccadic eye movements by burst neurons Journalof Neurophysiology 45 417ndash442 [PubMed]
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 15
Van Gisbergen J A M amp Robinson D A (1977)Generation of micro and macrosaccades by burstneurons in the monkey In R Baker amp A Berthoz(Eds) Control of gaze by brain stem neurons (vol 1pp 301ndash308) New York ElsevierNorth-Holland
Wallman J Pettigrew J D amp Letelier J C (1994)The oscillatory saccades of birds Motoneural activ-ity and possible functions In B T Fuchs A FV Buttner amp D Zee (Eds) Contemporary ocularmotor and vestibular research A tribute to David ARobinson (pp 480ndash487) Stuttgart Germany ThiemePublishing
Williams P E amp Shapley R M (2007) A dynamicnonlinearity and spatial phase specificity in macaque
V1 neurons Journal of Neuroscience 27 5706ndash5718[PubMed] [Article]
Winterson B J amp Robinson D A (1975) Fixationby the alert but solitary cat Vision Research 151349ndash1352 [PubMed]
Yarbus A L (1967) Eye movements and vision(B Haigh Trans) New York Plenum Press
Zuber B L amp Stark L (1965) Microsaccades and thevelocityltamplitude relationship for saccadic eyemovements Science 150 1459ndash1460 [PubMed]
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 16
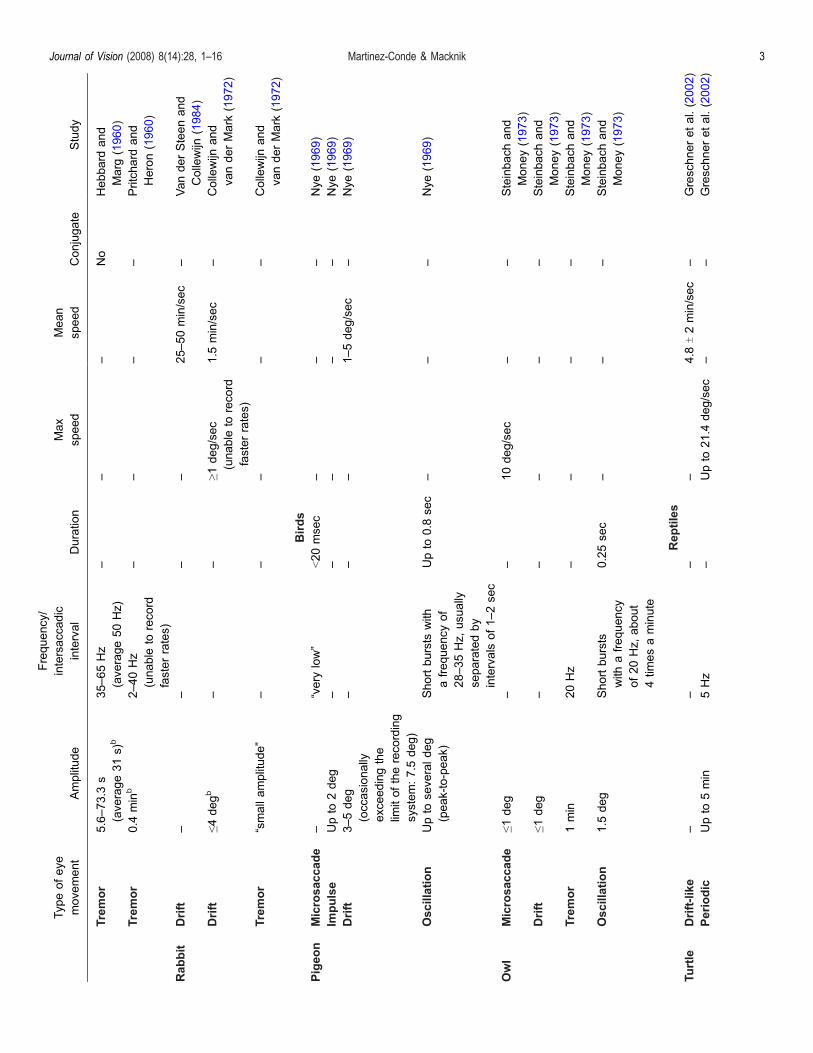
Typeofeye
move
ment
Amplitude
Frequency
intersacc
adic
interval
Duratio
nMax
speed
Mean
speed
Conjugate
Study
Tremor
56ndash733
s(ave
rage31s)
b35ndash
65Hz
(ave
rage50Hz)
ndashndash
ndashNo
Hebbard
and
Marg
(1960)
Tremor
04
min
b2ndash
40Hz
(unable
toreco
rdfasterrates)
ndashndash
ndashndash
Pritch
ard
and
Heron(1960)
Rabbit
Drift
ndashndash
ndashndash
25ndash
50minsec
ndashVanderSteenand
Collewijn
(1984)
Drift
e4degb
ndashndash
Q1degsec
(unable
toreco
rdfasterrates)
15
minsec
ndashCollewijn
and
vanderMark
(1972)
Tremor
ldquosmallamplituderdquo
ndashndash
ndashndash
ndashCollewijn
and
vanderMark
(1972)
Birds
Pigeon
Microsaccade
ndashldquovery
lowrdquo
G20mse
cndash
ndashndash
Nye
(1969)
Impulse
Upto
2deg
ndashndash
ndashndash
ndashNye
(1969)
Drift
3ndash5deg
(occ
asionally
exc
eedingthe
limitofthereco
rding
system75
deg)
ndashndash
ndash1ndash
5degsec
ndashNye
(1969)
Oscillation
Upto
seve
rald
eg
(peak-to-peak)
Shortbursts
with
afrequency
of
28ndash35Hz
usu
ally
separatedby
intervals
of1ndash
2se
c
Upto
08
sec
ndashndash
ndashNye
(1969)
Owl
Microsaccade
e1deg
ndashndash
10degsec
ndashndash
Steinbach
and
Money(1973)
Drift
e1deg
ndashndash
ndashndash
ndashSteinbach
and
Money(1973)
Tremor
1min
20Hz
ndashndash
ndashndash
Steinbach
and
Money(1973)
Oscillation
15
deg
Shortbursts
with
afrequency
of20Hz
about
4tim
esaminute
025se
cndash
ndashndash
Steinbach
and
Money(1973)
Reptiles
Turtle
Drift-like
ndashndash
ndashndash
48
T2minsec
ndashGresc
hneretal(2002)
Periodic
Upto
5min
5Hz
ndashUpto
214
degsec
ndashndash
Gresc
hneretal(2002)
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 3
(Engbert 2006 Martinez-Conde 2006 Martinez-Condeet al 2004) However the differences and similarities infixational eye movements across species as well as theirpotential significance have been neglected so far Despitewide-reaching interest in this topic (Martinez-Conde ampMacknik 2007a) a comprehensive comparative reviewhas not been published Here we review the comparativedynamics the physiology and the perceptual consequen-ces of fixational eye movements across all species studiedto date To the best of our knowledge our review includesall Medline-indexed studies (as well as several bookchapters and other non-indexed publications) of fixationaleye movements in non-human vertebrates These span48 years of research from the first studies in the catpublished in 1960 (Hebbard amp Marg 1960 Pritchardamp Heron 1960) to the most recent studies in thesalamander and archer fish published in 2007 and 2008(Baccus Olveczky Manu amp Meister 2008 OlveczkyBaccus amp Meister 2007 Segev Schneidman Goodhouseamp Berry 2007) As a result of our analysis we havegenerated a table (Table 1) that summarizes the physicalparameters of the different types of fixational eye move-ment acrossnon-humanvertebratesWewill relatewheneverpossible fixational eye movements in non-humanvertebrates to human fixational eye movements
Fixational eye movements inmammals
Fixational eye movements in the primate
Fixational eye movements were originally discoveredand characterized in human subjects (Ditchburn ampGinsborg 1952 1953 Ratliff amp Riggs 1950) Humanfixational eye movements comprise three main typesmicrosaccades (called ldquoflicksrdquo in early studies) drift andtremor (Figure 1) Microsaccades are the fastest andlargest of the three types of fixational eye movements(Supplementary Video 1) They travel in a straighttrajectory carrying the retinal image across a range ofseveral dozen to several hundred photoreceptor widthsDrifts are slow curvy motions that occur between micro-saccades and tremor is a very fast (Egrave90 Hz) andextremely small oscillation (about the diameter of a fovealcone) superimposed on drifts (see Tables 1ndash3 of Martinez-Conde et al 2004 for a review of fixational eye move-ment parameters in humans)Fixational eye movements in old-world monkeys are
very similar to human fixational eye movements (Martinez-Conde 2006 Martinez-Conde et al 2000 2002Skavenski Robinson Steinman amp Timberlake 1975Snodderly 1987 Snodderly amp Kurtz 1985) Severalmacaque species have been studied to date Macaca
Typeofeye
move
ment
Amplitude
Frequency
intersacc
adic
interval
Duratio
nMax
speed
Mean
speed
Conjugate
Study
Amphibians
Salamander
Tremor-lik
eAbout
125
min
ndashndash
ndashndash
ndashManteuffe
letal(1977)
Respiratory
Upto
30min
ndashndash
ndashndash
ndashManteuffe
letal(1977)
Fish
Goldfish
Drift
ndashndash
01ndash55
sec
e1degsec
036degsec
NoSim
ulta
neous
inboth
eye
sbut
conve
rgentin
thelight
Mensh
etal(2004)
Drift
Typ
ically
G5deg
ndashndash
ndashndash
NoSim
ulta
neous
inboth
eye
sbut
conve
rgent
Easter(1971)
ArcherFish
Microsaccade
ndashldquorarelyrdquofound
ndashndash
ndashndash
Segevetal(2007)
Drift
ndash02ndash1Hz
133se
ce0
4degsec
ndashndash
Segevetal(2007)
Tremor
01ndash02
deg
5Hz
ndash4degsec
~1degsec
ndashSegevetal(2007)
Table
1Characteris
ticsoffixatio
naleye
move
ments
innon-humanve
rtebrates
acc
ordingto
differentstudies
NoteaCalculatedfrom
horiz
ontalandve
rticalco
mponents
bVectorva
luesfrom
H-componentsaco
nve
rsionfactoroffrac342
hasbeenass
umed
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 4
mulatta (Goffart Quinet Chavane amp Masson 2006Horwitz amp Albright 2003 Leopold amp Logothetis1998 Martinez-Conde 2006 Martinez-Conde et al 20002002 Motter amp Poggio 1984 Skavenski et al 1975Snodderly Kagan amp Gur 2001 Steinman HaddadSkavenski amp Wyman 1973) Macaca fascicularis(Snodderly 1987 Snodderly et al 2001 Snodderly ampKurtz 1985) and Macaca nemestrina (Bair amp OrsquoKeefe1998) with no substantial differences in the spatiotemporalcharacteristics of fixational eye movements (rate velocityand magnitude profiles etc) between them Microsaccadevelocities are parametrically related to microsaccadeamplitudes following the ldquomain sequencerdquo in both ma-caques (Martinez-Conde et al 2000) and humans (Engbert2006 Martinez-Conde 2006 Martinez-Conde et al 2006Zuber amp Stark 1965) Fixational microsaccades have alsobeen observed in the baboon (Papio papio) with ampli-tudes up to 25 degrees (Marchetti Gauthier amp Pellet1983)It is important to note that microsaccades cannot be
defined according to their amplitude alone because smallexploratory or voluntary saccades can be the same size asmicrosaccades Microsaccades can be defined only opera-tionally as the involuntary saccades that are producedwhile the subject attempts to fixate (Martinez-Conde2006) This ldquoattempt to fixaterdquo includes both holding theeyes still and static while foveating the visual target andalso orienting the fovea toward a target It is worth notingthat microsaccade production may require the presence ofa visual andor attentional target (Otero-Millan et al2008)Thus there is no known physical parameter (or
combination of parameters) that separates saccades frommicrosaccades Mounting evidence points toward a com-mon neural generator of saccades and microsaccades(Engbert 2006 Martinez-Conde et al 2004 Otero-Millan et al 2008 Rolfs Kliegl amp Engbert 2008 RolfsLaubrock amp Kliegl 2006 Zuber amp Stark 1965)However one should note that most of the available datain support of a shared microsaccadendashsaccade generatorare indirect To date only two physiological studies havedirectly addressed the question of the oculomotor
mechanisms leading to the generation of microsaccadesIn those studies Van Gisbergen and colleagues foundthat putative motoneurons in the primate abducensnucleus and burst neurons in the nearby pontomedullaryreticular formation were similarly active during saccadesand microsaccades (Van Gisbergen Robinson amp Gielen1981 Van Gisbergen amp Robinson 1977) Future researchshould further explore the neural bases underlying thegeneration of microsaccades and other fixational eyemovementsSimultaneous recordings of microsaccades and neural
responses have been conducted at multiple levels of theprimate visual pathway including the lateral geniculatenucleus (LGN) area V1 and the extrastriate visual cortex(Bair amp OrsquoKeefe 1998 Leopold amp Logothetis 1998Martinez-Conde 2006 Martinez-Conde et al 20002002 Reppas Usrey amp Reid 2002 Snodderly et al2001) Microsaccades are predominantly excitatory at allthese levels leading to firing rate increases in both earlyand higher visual areas The firing rate increases followingmicrosaccades in the LGN and V1 are of visual origin(Figure 2) and result from the microsaccade-induceddisplacement of visual receptive fields over stationarystimuli Microsaccade-driven increases in firing rate tendto be clustered in tight bursts of spikes (Martinez-Conde2006 Martinez-Conde et al 2000 2002) A recent studyhas suggested that primate microsaccades may improvethe efficient sampling of fine spatial detail (Donner ampHemila 2007)Microsaccades counteract visual fading and increase
visibility during fixation in human subjects (Martinez-Conde et al 2006 Troncoso Macknik amp Martinez-Conde 2008) Numerous studies have also reported thathuman microsaccade rates andor directions are modulatedby cognitive processes such as the allocation of spatialattention (Betta amp Turatto 2006 Engbert amp Kliegl2003a 2003b 2004 Galfano et al 2004 Hafed amp Clark2002 Laubrock Engbert Rolfs amp Kliegl 2007 Otero-Millan et al 2008 Turatto Valsecchi Tame Betta 2007Valsecchi Betta amp Turatto 2007 but see also HorowitzFine Fencsik Yurgenson amp Wolfe 2007 and TseSheinberg amp Logothetis 2004)
Figure 1 Cartoon representation of fixational eye movements in humans and primates Microsaccades (straight and fast movements)drifts (curvy slow movements) and tremor (oscillations superimposed on drifts) transport the visual image across the retinal photoreceptormosaic From Martinez-Conde and Macknik (2007b)
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 5
Neural responses to drifts have received considerablyless attention than neural responses to microsaccades Thismay be due to the fact that drifts are more difficult tocharacterize objectively than microsaccades (which aremore easily detected by automatic algorithms that com-bine amplitude and velocity thresholds) Thus drifts areusually identified indirectly as the eye position changesthat occur between microsaccades (Snodderly et al2001) However this method has the potential flaw thatone may unintentionally attribute non-drift-related activity(for instance undetected tremor) to drifts Drifts mayincrease firing in a subset of primate V1 neurons(Snodderly et al 2001) However they generate lessvariability in neuronal responses than a combination ofdrifts and microsaccades (Gur Beylin amp Snodderly1997) No studies to date have recorded primate neuronalresponses in correlation to tremor (but see Hennig ampWorgotter 2007 for the responses of a model macaqueretina to simulated drift and tremor)
Fixational eye movements in the cat
The catrsquos fixational eye movements have been inves-tigated in a handful of studies and their conclusions
remain somewhat controversial In 1960 Hebbard andMarg (1960) found fixational eye movements in the cat tobe quite comparable to those in humans although catmicrosaccades were scarcer and smaller than in humansPritchard and Heron (1960) also found that the catrsquosfixational eye movements included microsaccades driftsand tremor Microsaccades were much rarer in the cat thanin humans however possibly related to the fact that thecat lacks a well-developed fovea Delgado-Garcia delPozo and Baker (1986) also observed some putativemicrosaccades in the alert behaving cat Other studiesfound no microsaccades in the cat (Conway Timberlakeamp Skavenski 1981 Winterson amp Robinson 1975)However in these last studies microsaccades were definedas ldquosaccades smaller than 10 minutes of arcrdquo which iswell below the average microsaccade amplitude found inrecent human and primate studies (see Martinez-Condeet al 2004 for a review of microsaccade parameters inhumans and primates) More recently several studies haveaddressed the contribution of fixational eye movements toneural responses in the catrsquos visual system HennigKerscher Funke and Worgotter (2002) found increasedresponses from neurons in cat areas 17 and 18 duringstimulus motions that mimicked fixational eye move-ments Using a model retina they also found that
1 simulated fixational eye movements increased neu-ral responses and
2 simulated tremor improved spatial acuity
Hennig and Worgotter (2004) later reported that acombination of tremor and microsaccades improved theperformance of simulated ganglion cells in a hyperacuitytask in the retinal periphery (but not in the central retina)Miller Denning George Marshak and Kenyon (2006)also investigated the effects of simulated tremor on amodel of the catrsquos retina and suggested that tremor mayselectively enhance the processing of large stimuli
Fixational eye movements in the rabbit
In contrast to foveate species such as primates (includ-ing humans) and cats (which have an area centralis if nota proper fovea) the rabbit (Oryctolagus cuniculus) dis-plays few spontaneous eye movements especially in theabsence of head motion (Collewijn 1977 Collewijn ampvan der Mark 1972 Fuller 1980 1981 Van der Steen ampCollewijn 1984) In normal visual conditions the rabbitrsquoseyes are very stable but not perfectly immobile Smallamplitude tremor can be observed as well as very slowdrift Microsaccades have not been observed Howeverdrift is occasionally corrected by fast saccadic movements(Collewijn 1977 Collewijn amp van der Mark 1972 Vander Steen amp Collewijn 1984) These differences may berelated to the fact that the rabbit does not have a fovea butan elongated horizontal streak with elevated ganglion cell
Figure 2 Microsaccades drive neural activity in primate area V1Microsaccades lead to firing rate increases in V1 neurons (n = 308)of the awake primate In the absence of visual stimulationmicrosaccades do not increase neural activity in area V1 (n = 37neurons) The resultsVshown here for the average of all neuronsVwere consistent with responses from individual neurons (data notshown) Modified from Martinez-Conde et al (2000 2002)
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 6
counts Olveczky Baccus and Meister (2003) recordedfrom ganglion cells in the rabbit and salamander retinaswhile presenting visual stimuli that were jittered tosimulate fixational eye movements (based on parametersfrom previously published studies by Manteuffel PlasaSommer amp Wess 1977 and Van der Steen amp Collewijn1984) In both species fixational eye movements helpedsegregate a moving foreground from a complex back-ground in a population of ganglion cells
Fixational eye movements inbirds
Fixational eye movements in the pigeon
There are some remarkable differences between fixa-tional eye movements in birds and mammals Nye (1969)identified four types of fixational eye movements in thepigeon (Columba livia) flicks (ie microsaccades)impulse movements drift and oscillations No tremorwas detected Microsaccade frequency was very lowrelative to humans Impulse movements occurred morefrequently usually between bursts of oscillations withamplitudes ranging up to 2 degrees Drift velocitiesranged from 1 to 5 degrees per second Both micro-saccades and drifts occurred in response to movingstimuli Oscillations occurred in short bursts at rates ofabout 30 Hz and could be quite large (up to severaldegrees of amplitude) Bloch Rivaud and Martinoya(1984) reported microsaccades drifts and oscillationsldquosimilar to those described by Nyerdquo but none of thesewere analyzed in detailCyclotorsional oscillations are an integral part of avian
saccades (Wallman Pettigrew amp Letelier 1994) Thusone might speculate that oscillations are associated withavian microsaccades as well especially if saccades andmicrosaccades share a common generator (Engbert 2006Otero-Millan et al 2008 Rolfs et al 2008 2006 Zuberamp Stark 1965) However this possibility has not yet beenaddressed in the literature The function of oscillations isunknown (Carpenter 1988) but one hypothesis is thatthey serve to facilitate the delivery of arterial blood to theocular capillary bed (Pettigrew Wallman amp Wildsoet1990 Wallman et al 1994)
Fixational eye movements in the owl
Steinbach and Money (1973) recorded fixational eyemovements in the great horned owl (Bubo virginianus)a species previously thought incapable of moving itseyes The owlrsquos fixational eye movements includeflicks or microsaccades drift tremor and oscillations
Microsaccades seemed correlated to visual events ofinterest to the owl they were 1 deg or less in amplitudeand had peak velocities of about 10 degsec Drifts wereslow motions of the eye in between fixations withamplitudes of 1 degree or less Tremor amplitude wasabout 1 arcmin with frequencies around 20 Hz High-frequency oscillations (25 Hz) of short duration (025 sec)were very similar to those previously found in the pigeon(Nye 1969) No studies to date have examined theperceptual or physiological effects of fixational eyemovements in any bird species
Fixational eye movements inreptiles
Fixational eye movements in the turtle
Greschner Bongard Rujan and Ammermuller (2002)identified two types of fixational eye movements in theturtle (Pseudemys scripta elegans) which they calledldquodrift-likerdquo and ldquoperiodicrdquo Drift-like motions were smalland slow (with velocities comparable to human drift) andwere superimposed by larger and faster periodic componentsPeriodic motion had retinal amplitudes on the order of thediameter of a photoreceptor which are comparable to thoseof human tremor On the other hand periodic motionrsquosfrequency (5 Hz) velocity and orderly relationship betweenvelocity and amplitude are closer to those of human micro-saccades (see Martinez-Conde et al 2004 for a review oftremor and microsaccade parameters in humans)Simulated periodic motion generated strong synchronous
firing in the turtlersquos retina whereas simulated drift-likemotion had little effect (Figure 3) These results agree withprevious physiological and modeling studies in the primatevisual system in which strong neural transients wereobserved in response to microsaccades (Donner amp Hemila2007 Martinez-Conde 2006 Martinez-Conde et al 20002002) Such neural transients may underlie the behavior ofcortical neurons as coincidence detectors (ShelleyMcLaughlin Shapley amp Wielaard 2002 Williams ampShapley 2007) Moreover neural transients to stimulionsets and terminations (similar to those produced bymicrosaccades in the primate visual system Martinez-Conde 2006 Martinez-Conde et al 2000 2002) havebeen related to target visibility in visual masking para-digms (Macknik amp Livingstone 1998 Macknik ampMartinez-Conde 2004 Macknik Martinez-Conde ampHaglund 2000)In the turtle retina periodic motion-driven neurons with
receptive fields located along contrast borders weremoreover synchronized and reliably indicated the preced-ing motions The authors proposed that this synchroniza-tion of retinal activity could be used by the brain to
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 7
improve the estimation of stimulus features such asspatial frequency (Greschner et al 2002) This findingcould have a parallel in the primate visual system Donnerand Hemila (2007) recently proposed that microsaccadessignificantly ldquore-sharpenrdquo the image and improve spatialresolution in primates Rucci Iovin Poletti and Santini(2007) also found that fixational eye movements enhancefine spatial detail in human subjects
Fixational eye movements in the chameleon
Fixational eye movements in the chameleon have notbeen studied in detail However Gioanni and colleaguesfound that the chameleon (Chamaleo chamaleo) makessaccades during fixation The authors speculated that theseprobably correspond to fixational microsaccades found inother foveate species (Gioanni Bennis amp Sansonetti1993)
Fixational eye movements inamphibians
Fixational (and more generally spontaneous) eye move-ments vary widely among amphibians The frog (anafoveate amphibian) shows a general lack of eye andhead movements except for those that stabilize the retinalimage Dieringer and colleagues (Dieringer amp Daunicht1986 Dieringer amp Precht 1982 Dieringer Precht ampBlight 1982) found that the grass frog (Rana temporaria)produces no spontaneous eye movements in the absenceof head movements with the possible exception of tremor(similarly to the rabbit another afoveate species Van derSteen amp Collewijn 1984) It is likely that the frogrsquos lack ofeye movements leads to fast visual fading of all stationaryobjects in its field of view (Ditchburn amp Ginsborg 1952Yarbus 1967) Lettvin Maturana MsCulloch and Pitts
Figure 3 Fixational eye movements increase retinal activity in the turtle (A) Continuous recording from 54 retinal ganglion cells in theturtle during darkness after switching on a stationary grating and during wobbling of this grating The grating was wobbled to simulateperiodic (ie tremor-like) eye movements (B) Responses of a single ganglion cell to a drifting contrast border of velocity comparable toocular drifts (C) When the drift is superimposed with wobbling the responses of the cell shown in (B) increase dramatically FromGreschner et al (2002) and modified by Martinez-Conde et al (2004)
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 8
(1968) stated that a frog ldquowill starve to death surroundedby food if it is not movingrdquo (but see Dieringer amp Daunicht1986 for a counterpoint) Small head motions or eyemovements correlated with respiratory motions maydiminish fading to some extent (Dieringer amp Precht1982 Schipperheyn 1963 1965 1967)As a side note the frogrsquos respiratory eye movements
play no role in the respiratory function ldquoin the course ofeach respiratory cycle increase of pressure in the frogrsquosbuccal cavity protrudes the eye and causes the opticalaxis to deviate upwards over 25ndash35 minutes of arcrdquo(Schipperheyn 1963) That is respiratory eye movementsare merely synchronized with respirationAutrum (1959) found no evidence of respiratory eye
movements in the frog Ewert andBorchers (1974) observedthat stationary stimuli elicit no responses from visualneurons in the common toad (Bufo Bufo L) regardless ofrespiratory pressure changes in the buccal cavity
Fixational eye movements in the salamander
Manteuffel et al (1977) identified two types of fixa-tional eye movements in salamanders tremor-like andrespiratory eye movements Tremor-like is a rapid move-ment of small amplitude Torsional eye movements resultfrom the animalrsquos breathing (also found in the frog bySchipperheyn 1963) Additionally flicks are occasionallyobserved which rapidly change the position of gaze It isunclear whether such flicks may generally correspond towhat we now call microsaccades andor to voluntarysaccades Based on these observations the authors pro-posed that unlike frogs and toads (Autrum 1959 Ewert ampBorchers 1974 Lettvin et al 1968) salamanders cancontinuously perceive stationary objects (Manteuffel et al1977) Fixational eye movements in the salamander (Tigersalamander Ambystoma tigrinum) may also help segre-gate object and background motion in the retina(Olveczky et al 2003 see also Fixational eye movementsin the rabbit section) and even work to emphasize novelvisual features (Olveczky et al 2007 see Baccus et al2008 for details on the retinal neurons and synapses thatmay underlie the computation of differential motionbetween an object and its background)
Fixational eye movements in fish
Spontaneous eye movements in the goldfish
Eye movements during fixation have been studied inboth afoveate (goldfish Carassius auratus) and foveate(archer fish Toxotes chatareus) fish species The goldfisha long-standing model organism in vision and oculomotor
research (Aksay et al 2003 2007 Cabrera Torres PasaroPastor amp Delgado-Garcia 1992 Pastor Torres Delgado-Garcia amp Baker 1991 Salas Navarro Torres ampDelgado-Garcia 1992 Torres Pastor Cabrera Salas ampDelgado-Garcia 1992) produces three types of sponta-neous eye movements saccades stretch and drift (Easter1971 Mensh Aksay Lee Seung amp Tank 2004) Weshould note that in the motionless goldfish eye move-ments are very infrequent Those that do occur consistmainly of saccades slow drifts are very rare (Easter Johnsamp Heckenlively 1974) During saccades (identified bythresholding the eye velocity at 5 degsec) both eyes moverapidly simultaneously and usually in the same direction(Mensh et al 2004) Stretches occur every 2ndash3 min and arecharacterized by a brief simultaneous large excursion ofboth eyes to extreme temporal positions (Easter 1971Mensh et al 2004) The function of stretches is not known(Mensh et al 2004) Because fixations were defined asthose periods that were free of saccadic and stretchmovements (Easter 1971) only drifts (from the threetypes of spontaneous movements) can technically beconsidered ldquofixational eye movementsrdquo However thegoldfish saccadic main sequence includes saccades withequivalent amplitudes and velocities to those found inhuman microsaccades (Mensh et al 2004 Zuber amp Stark1965) Indeed microsaccades have been identified in other(foveate) fish species although they are rarer than inhumans (Segev et al 2007) Small saccades in thegoldfish tend to be divergent whereas large saccadestend to be convergent (Mensh et al 2004) Drift occurssimultaneously in both eyes usually in the samedirection Drift velocities are almost exclusively e1 degsec(Mensh et al 2004) No tremor has been identified in thegoldfish
Spontaneous eye movements in teleost fish
Spontaneous eye movements have been studied in theNew Zealand parore (Girella tricuspidata MontgomeryMcVean amp McCarthy 1983) The pattern of eye move-ments was similar to that previously described in goldfish(Easter 1971 Hermann amp Constantine 1971)
Fixational eye movements in the archer fish
A recent study in the archer fish (Segev et al 2007) hasprovided the first perceptual and physiological correlatesof fixational eye movements in any fish species Archerfish have remarkable visual abilities they shoot downinsects resting on foliageVas well as stationary targetscovering a single retinal photoreceptorVwith squirts ofwater from their mouths they predict the location atwhich the insect will splash down in G100 msec and theydeduce the absolute size of an object presented at differentviewing distances (Supplementary Videos 2 and 3) Segev
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 9
et al (2007) identified three types of fixational eyemovements in the archer fish microsaccades drift andtremor Microsaccades were rare and were not analyzed indetail Drift velocities were e040 degsec Tremor was ahigh-frequency oscillatory movement (the primary com-ponent was 5 Hz) with peak-to-peak amplitude of 01ndash02 deg (corresponding to an image displacement of 1ndash2photoreceptors) and velocity of about 1 degsec (Figure 4)Whereas the amplitude and velocity of tremor in thearcher fish is comparable to those in humans its frequencyis much lower (5 Hz vs 940 Hz) Interestingly archer fishtremor is very similar both in amplitude (photoreceptorwidths covered) as well as in frequency (5 Hz) to thetremor-like periodic motion in the turtle (discussedearlier Greschner et al 2002)Segev et al (2007) trained archer fish to perform a size
discrimination task the fish had to distinguish a medium-sized target from larger and smaller distracters (theyindicated their choice by shooting a jet of water) To relatethis behavior to responses from retinal neurons theauthors recorded from populations of retinal ganglioncells while presenting objects of different sizes moved tosimulate saccades drifts and tremor Drifts and tremorelicited weak neuronal responses that were not veryinformative about the spatial structure of the stimulus(although they did provide some information about target
size in agreement with the results of Greschner et al2002 in the turtle) Moreover performance was similar fortremor alone and for tremor plus drift indicating that driftplays a negligible role in revealing spatial structure (alsoin agreement with Greschner et al 2002)When present the firing rate was phase-locked to
tremor Saccades were more effective than drifts or tremorin driving informative responses from the ganglion cells(consistent with theoretical and physiological studies ofmicrosaccade-driven neuronal activity in the primateretina and LGN Donner amp Hemila 2007 Martinez-Conde et al 2002) Segev et al (2007) proposed that inprimates both saccades and microsaccades (which arerare in the archer fish) may contribute to size discrim-ination tasks as well as enhance spatial detail whereasdrifts and tremor by themselves may not be sufficient toimprove visual perception However it is important toremember that central vision in humans can be maintainedby tremor andor drift even when microsaccades aresuppressed during strict fixation (Martinez-Conde 2006Martinez-Conde et al 2004 2006) On the other handone cannot rule out the possibility that if one couldcompletely eliminate drifts and tremor microsaccadesmight then suffice to sustain both central and peripheralvision in primates and humans Future experiments areneeded to further explore these ideas
Figure 4 Archer fish eye movements (A) Example trace of eye position as a function of time as measured by search coil (B) Example ofa single 15-s fixation superimposed on a grid representing the photoreceptor mosaic The back and forth movement is caused by tremor(C) Distribution of fixation durations (taken as time interval between saccades) (D) Power spectrum of horizontal fixational eyemovements (E) Distribution of tremor amplitude (data taken from 1 animal in a 15-min session) Modified from Segev et al (2007)
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 10
Discussion
The vast majority of eye movement studies conductedin non-human species to date have not analyzed eyeposition data in the range of fixational eye movementsThus eye movements during fixation have been reportedin only a handful of vertebrates Most of these studieshave concentrated solely on the physical parameters ofthe fixational eye movements without addressing theirphysiological and perceptual consequences Fortunatelythis trend has started to reverse in recent years with agrowing list of papers addressing the effects of non-human fixational eye movements in physiology andperception This is an important development A largeamount of all psychophysical and physiological visualresearch has been conducted during conditions of visualfixation Therefore understanding the precise physiolog-ical and perceptual contributions of fixational eye move-ments may be critical to the interpretation of previous andfuture vision researchThere are some interesting differences and similarities
in the types of fixational eye movements produced bydifferent species For instance ocular tremor is possiblyubiquitous to all vertebrates The fact that tremor has notbeen observed in all species studied could be due to thetechnical difficulty of recording tremor accurately (ietremor amplitudes and frequencies are usually in the rangeof the recording systemrsquos noise) Drift has also beenrecorded in most vertebrates and it may be as omnipre-sent as tremor Conversely microsaccades seem mostimportant in species with foveal vision (ie primates andhumans) whereas they are scarce or absent in afoveatespecies (ie rabbits frogs goldfish) A link betweenmicrosaccades and foveal vision may reflect the two-pronged functional significance of microsaccades inhumans and primates first microsaccades may benecessary to correct the slow drift of the eye and bringback the objectregion of interest to the retinal area wherephotoreceptor density is highest (Cornsweet 1956) Thecorrective role of human microsaccades has been demon-strated in recent studies (Engbert amp Kliegl 2004 Engbertamp Mergenthaler 2006) Second the dynamics of micro-saccades themselves may improve spatial resolution(Donner amp Hemila 2005)In reviewing the patterns of fixational eye movements
across vertebrates it is tempting to draw parallels withother sensory modalities Fixational eye movementsrefresh the retinal images during fixation and thus preventneural adaptation and visual fading But this mechanism isnot circumscribed to the visual system Sniffs in rodentolfaction discretely sample sensory information every200ndash300 msec and thus are similar in their temporaldynamics to primate saccades (Otero-Millan et al 2008Uchida Kepecs amp Mainen 2006) and microsaccades(Otero-Millan et al 2008) A similar mode of discretesampling may also be at play when objects are recognized
through tactile information for instance if we sweep ourfingertips over an objectrsquos surface with our eyes closed orwhen blind individuals read Braille scriptOne intriguing possibility that merits further experi-
ments and analyses is the potential correlation between thespatiotemporal parameters of fixational eye movements(such as those summarized in Table 1) and the adaptationproperties of visual neurons across species For instancethe greater ocular stability of some species (rabbitgoldfish frog etc) versus others (primates) could beassociated with some particular adaptive properties oftheir visual cells Thus future comparative studies shouldpay attention to the response properties of visual cellstriggered by fixational eye movements in the presence ofstationary stimuli Other potential factors underlying thevariation in fixational eye movements across species couldbe differences in oculomotor control at the level of the eyemuscles or perhaps differences in retinal organizationFuture research will further improve our understanding
of fixational eye movements and the neural activity thatmaintains the visibility of stationary visual scenes inhuman and non-human species One interesting questionthat remains to be addressed concerns the role of tremor invision and visual physiology Because tremor frequenciesare well over the flicker fusion frequencies in humans ithas been argued that tremor of the visual image would beineffective as a stimulus (Ditchburn 1955 Gerrits ampVendrik 1970 Sharpe 1972) However recent studieshave found simulated tremor to enhance visual processingand improve visual acuity in a model retina (Hennig et al2002 Miller et al 2006) Follow-up research should beaimed to replicate these findings in actual neurons withcorrelated perceptual reports One should also note thatearly visual neurons can follow high-frequency flickeringthat surpasses the perceptual threshold for flicker fusion(Martinez-Conde et al 2002) Thus high-frequencytremor could be adequate to maintain activity in the earlyvisual system which then may lead to visual perception inhigher visual areas Another significant question thatremains unanswered concerns the oculomotor bases ofthe various types of fixational eye movements acrossspecies To date there is too little information on thisimportant topic to draw a conclusion
Conclusions
We have reviewed the parameters of fixational eyemovements in non-human mammals birds reptilesamphibians and fish as well as their consequences forphysiology and perception Fixational eye movementsand microsaccades in particular appear to be mostimportant in foveate vs afoveate species Animals lackingfixational eye movements such as afoveate frogs andtoads may be incapable of seeing stationary objects
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 11
during visual fixation Future research should furtherexplore the perceptual effects of fixational eye movementsand determine the oculomotor mechanisms responsible fortheir generation across species
Acknowledgments
We thank Mona Stewart and Isabel Gomez-Caraballo fortechnical assistance Jorge Otero-Millan for programmingassistance and Dr Xoana G Troncoso for comments on themanuscript This study was funded through grants fromthe Barrow Neurological Foundation (to SMC and SLM)the National Science Foundation (Award 0643306 toSMC) the Arizona Biomedical Research Commission(Award 07-102 to SMC) and Science Foundation Arizona(SLM)
Commercial relationships noneCorresponding author Susana Martinez-Conde PhDEmail smartneuralcorrelatecomAddress Barrow Neurological Institute 350 W ThomasRd Phoenix AZ 85013 USA
References
Aksay E Major G Goldman M S Baker R SeungH S amp Tank D W (2003) History dependence ofrate covariation between neurons during persistentactivity in an oculomotor integrator Cerebral Cortex13 1173ndash1184 [PubMed] [Article]
Aksay E Olasagasti I Mensh B D Baker RGoldman M S amp Tank D W (2007) Functionaldissection of circuitry in a neural integrator NatureNeuroscience 10 494ndash504 [PubMed]
Autrum H (1959) Das Fehlen unwillkurlicher Augenbe-wegungen beim Frosch Naturwissenschaften 46435
Baccus S A Olveczky B P Manu M amp Meister M(2008) A retinal circuit that computes object motionJournal of Neuroscience 28 6807ndash6817 [PubMed]
Bair W amp OrsquoKeefe L P (1998) The influence offixational eye movements on the response of neuronsin area MT of the macaque Visual Neuroscience 15779ndash786 [PubMed]
Betta E amp Turatto M (2006) Are you ready I can tellby looking at your microsaccades Neuroreport 171001ndash1004 [PubMed]
Bloch S Rivaud S amp Martinoya C (1984) Compar-ing frontal and lateral viewing in the pigeon IIIDifferent patterns of eye movements for binocularand monocular fixation Behavioural Brain Research13 173ndash182 [PubMed]
Cabrera B Torres B Pasaro R Pastor A M ampDelgado-Garcia J M (1992) A morphological studyof abducens nucleus motoneurons and internuclearneurons in the goldfish (Carassius auratus) BrainResearch Bulletin 28 137ndash144 [PubMed]
Carpenter R H S (1988) Movements of the eyes(2nd ed) Pion London
Collewijn H (1977) Eye- and head movements infreely moving rabbits The Journal of Physiology266 471ndash498 [PubMed] [Article]
Collewijn H amp van der Mark F (1972) Ocular stabilityin variable visual feedback conditions in the rabbitBrain Research 36 47ndash57 [PubMed]
Conway J L Timberlake G T amp Skavenski A A(1981) Oculomotor changes in cats reared withoutexperiencing continuous retinal image motion Exper-imental Brain Research 43 229ndash232 [PubMed]
Cornsweet T N (1956) Determination of the stimuli forinvoluntary drifts and saccadic eye movements Jour-nal of the Optical Society of America 46 987ndash993[PubMed]
Delgado-Garcia J M del Pozo F amp Baker R (1986)Behavior of neurons in the abducens nucleus of thealert catVII Internuclear neurons Neuroscience 17953ndash973 [PubMed]
Dieringer N amp Daunicht W J (1986) Image fadingVAproblem for frogs Naturwissenschaften 73 330ndash332[PubMed]
Dieringer N amp Precht W (1982) Compensatory headand eye movements in the frog and their contributionto stabilization of gaze Experimental BrainResearch 47 394ndash406 [PubMed]
Dieringer N Precht W amp Blight A R (1982)Resetting fast phases of head and eye and theirlinkage in the frog Experimental Brain Research 47407ndash416 [PubMed]
Ditchburn R W (1955) Eye-movements in relation toretinal action Optica Acta 1 171ndash176
Ditchburn R W amp Ginsborg B L (1952) Vision with astabilized retinal image Nature 170 36ndash37[PubMed]
Ditchburn R W amp Ginsborg B L (1953) Involuntaryeye movements during fixation The Journal ofPhysiology 119 1ndash17 [PubMed] [Article]
Donner K amp Hemila S (2007) Modelling the effect ofmicrosaccades on retinal responses to stationarycontrast patterns Vision Research 47 1166ndash1177[PubMed]
Donner K K amp Hemila S (2005) The possible purposeof microsaccades Paper presented at the EuropeanConference on Visual Perception A Coruna Spain
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 12
Easter S S Jr (1971) Spontaneous eye movements inrestrained goldfish Vision Research 11 333ndash342[PubMed]
Easter S S Jr Johns P R amp Heckenlively D R(1974) Horizontal compensatory eye movements ingoldfish (Carassius auratus) I The normal animalJournal of Comparative Physiology 92 23ndash35
Engbert R (2006) Microsaccades A microcosm forresearch on oculomotor control attention and visualperception Progress Brain Research 154 177ndash192[PubMed]
Engbert R amp Kliegl R (2003a) Binocular coordinationin microsaccades In R R J Hyona amp H Deubel(Eds) The mindrsquos eyes Cognitive and appliedaspects of eye movements (pp 103ndash117) OxfordUK Elsevier
Engbert R amp Kliegl R (2003b) Microsaccades uncoverthe orientation of covert attention Vision Research43 1035ndash1045 [PubMed]
Engbert R amp Kliegl R (2004) Microsaccades keep theeyesrsquo balance during fixation Psychological Science15 431ndash436 [PubMed]
Engbert R amp Mergenthaler K (2006) Microsaccadesare triggered by low retinal image slip Proceedingsof the National Academy of Sciences of the UnitedStates of America 103 7192ndash7197 [PubMed][Article]
Ewert J P amp Borchers H W (1974) Antwort vonretinalen Ganglienzellen bei freibeweglichen Kroten(Bufo Bufo L) Journal of Comparative and Physi-ology 92 117ndash130
Fuller J H (1980) Linkage of eye and head movementsin the alert rabbit Brain Research 194 219ndash222[PubMed]
Fuller J H (1981) Eye and head movements duringvestibular stimulation in the alert rabbit BrainResearch 205 363ndash381 [PubMed]
Galfano G Betta E amp Turatto M (2004) Inhibition ofreturn in microsaccades Experimental BrainResearch 159 400ndash404 [PubMed]
Gerrits H J amp Vendrik A J (1970) Artificial move-ments of a stabilized image Vision Research 101443ndash1456 [PubMed]
Gioanni H Bennis M amp Sansonetti A (1993) Visualand vestibular reflexes that stabilize gaze in thechameleon Visual Neuroscience 10 947ndash956[PubMed]
Goffart L Quinet J Chavane F amp Masson G S(2006) Influence of background illumination onfixation and visually guided saccades in the rhesusmonkey Vision Research 46 149ndash162 [PubMed]
Greschner M Bongard M Rujan P amp Ammermuller J(2002) Retinal ganglion cell synchronization by fixa-tional eye movements improves feature estimationNature Neuroscience 5 341ndash347 [PubMed]
Gur M Beylin A amp Snodderly D M (1997)Response variability of neurons in primary visualcortex (V1) of alert monkeys Journal of Neuro-science 17 2914ndash2920 [PubMed] [Article]
Hafed Z M amp Clark J J (2002) Microsaccades as anovert measure of covert attention shifts VisionResearch 42 2533ndash2545 [PubMed]
Hebbard F W amp Marg E (1960) Physiologicalnystagmus in the cat Journal of the Optical Societyof America 50 151ndash155 [PubMed]
Hennig M A Kerscher N J Funke K amp Worgotter F(2002) Stochastic resonance in visual cortical neuronsDoes the eye-tremor actually improve visual acuityNeurocomputing 44ndash46 115ndash120
Hennig M H amp Worgotter F (2004) Eye micro-movements improve stimulus detection beyond theNyquist limit in the peripheral retina In Advances inNeural Information Processing Systems (NIPS2003)(vol 16) Denver CO MIT Press
Hennig M H amp Worgotter F (2007) Effects offixational eye movements on retinal ganglion cellresponses A modelling study Frontiers in Computa-tional Neuroscience 1 2 [PubMed] [Article]
Hermann H T amp Constantine M M (1971) Eyemovements in the goldfish Vision Research 11313ndash331 [PubMed]
Horowitz T S Fine E M Fencsik D E Yurgenson Samp Wolfe J M (2007) Fixational eye movements arenot an index of covert attention PsychologicalScience 18 356ndash363 [PubMed]
Horwitz G D amp Albright T D (2003) Short-latencyfixational saccades induced by luminance incrementsJournal of Neurophysiology 90 1333ndash1339[PubMed] [Article]
Laubrock J Engbert R Rolfs M amp Kliegl R(2007) Microsaccades are an index of covertattention Commentary on Horowitz Fine FencsikYurgenson and Wolfe (2007) Psychological Sci-ence 18 364ndash366 discussion 367ndash368 [PubMed]
Leopold D A amp Logothetis N K (1998) Micro-saccades differentially modulate neural activity in thestriate and extrastriate visual cortex ExperimentalBrain Research 123 341ndash345 [PubMed]
Lettvin J Y Maturana H R MsCulloch W S amp PittsW H (1968) What the frogrsquos eye tells the frogrsquosbrain In W C C a M Balaban (Ed) The mind
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 13
Biological approaches to its functions (pp 233ndash258)John Wiley and Sons London
Macknik S L amp Livingstone M S (1998) Neuronalcorrelates of visibility and invisibility in the primatevisual system Nature Neuroscience 1 144ndash149[PubMed]
Macknik S L amp Martinez-Conde S (2004) The spatialand temporal effects of lateral inhibitory networksand their relevance to the visibility of spatiotemporaledges Neurocomputing 58ndash60 775ndash782
Macknik S L Martinez-Conde S amp Haglund M M(2000) The role of spatiotemporal edges in visibilityand visual masking Proceedings of the NationalAcademy of Sciences of the United States of America97 7556ndash7560 [PubMed] [Article]
Manteuffel G Plasa L Sommer T J amp Wess O(1977) Involuntary eye movements in salamandersNaturwissenschaften 64 533ndash534 [PubMed]
Marchetti E Gauthier G M amp Pellet J (1983)Cerebellar control of eye movements studied withinjection of harmaline in the trained baboon ArchivesItaliennes de Biologies 121 1ndash17 [PubMed]
Martinez-Conde S (2006) Fixational eye movements innormal and pathological vision Progress in BrainResearch 154 151ndash176 [PubMed]
Martinez-Conde S amp Macknik S L (2007a) Bobbingand perceiving Scientific American 297 16
Martinez-Conde S amp Macknik S L (2007b) Windowson the mind Scientific American 297 56ndash63[PubMed]
Martinez-Conde S Macknik S L amp Hubel D H(2000) Microsaccadic eye movements and firing ofsingle cells in the striate cortex of macaque monkeysNature Neuroscience 3 251ndash258 [PubMed]
Martinez-Conde S Macknik S L amp Hubel D H(2002) The function of bursts of spikes during visualfixation in the awake primate lateral geniculate nucleusand primary visual cortex Proceedings of theNational Academy of Sciences of the United Statesof America 99 13920ndash13925 [PubMed] [Article]
Martinez-Conde S Macknik S L amp Hubel D H(2004) The role of fixational eye movements invisual perception Nature Reviews Neuroscience 5229ndash240 [PubMed]
Martinez-Conde S Macknik S L Troncoso X G ampDyar T A (2006) Microsaccades counteract visualfading during fixation Neuron 49 297ndash305[PubMed] [Article]
Mensh B D Aksay E Lee D D Seung H S ampTank D W (2004) Spontaneous eye movements ingoldfish Oculomotor integrator performance plasti-
city and dependence on visual feedback VisionResearch 44 711ndash726 [PubMed]
Miller J A Denning K S George J S MarshakD W amp Kenyon G T (2006) A high frequencyresonance in the responses of retinal ganglion cells torapidly modulated stimuli A computer model VisualNeuroscience 23 779ndash794 [PubMed]
Montgomery J C McVean A R amp McCarthy D(1983) The effects of lowered temperature onspontaneous eye movements in a teleost fish Com-parative Biochemistry and Physiology A Compara-tive Physiology 75 363ndash368 [PubMed]
Motter B C amp Poggio G F (1984) Binocularfixation in the rhesus monkey Spatial and temporalcharacteristics Experimental Brain Research 54304ndash314 [PubMed]
Murakami I amp Cavanagh P (1998) A jitter after-effectreveals motion-based stabilization of vision Nature395 798ndash801 [PubMed]
Nye P W (1969) The monocular eye movements of thepigeon Vision Research 9 133ndash144 [PubMed]
Olveczky B P Baccus S A amp Meister M (2003)Segregation of object and background motion in theretina Nature 423 401ndash408 [PubMed]
Olveczky B P Baccus S A amp Meister M (2007)Retinal adaptation to object motion Neuron 56689ndash700 [PubMed] [Article]
Otero-Millan J Troncoso X Macknik S L Serrano-Pedraza I amp Martinez-Conde S (2008) Saccadesand microsaccades during visual fixation explorationand search Foundations for a common saccadicgenerator Journal of Vision 8(14)21 1ndash18 httpjournalofvisionorg81421 doi10116781421[PubMed] [Article]
Pastor A M Torres B Delgado-Garcia J M ampBaker R (1991) Discharge characteristics of medialrectus and abducens motoneurons in the goldfishJournal of Neurophysiology 66 2125ndash2140 [PubMed]
Pettigrew J D Wallman J amp Wildsoet C F (1990)Saccadic oscillations facilitate ocular perfusion fromthe avian pecten Nature 343 362ndash363 [PubMed]
Pritchard R M amp Heron W (1960) Small eye move-ments of the cat Canadian Journal of Psychology14 131ndash137 [PubMed]
Ratliff F amp Riggs L A (1950) Involuntary motions ofthe eye during monocular fixation Journal of Exper-imental Psychology 40 687ndash701 [PubMed]
Reppas J B UsreyWM amp Reid R C (2002) Saccadiceye movements modulate visual responses in thelateral geniculate nucleus Neuron 35 961ndash974[PubMed] [Article]
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 14
Riggs L A amp Ratliff F (1952) The effects of counter-acting the normal movements of the eye Journal ofthe Optical Society of America 42 872ndash873
Rolfs M Kliegl R amp Engbert R (2008) Toward amodel of microsaccade generation The case ofmicrosaccadic inhibition Journal of Vision 8(11)51ndash23 httpjournalofvisionorg8115 doi1011678115 [PubMed] [Article]
Rolfs M Laubrock J amp Kliegl R (2006) Shorteningand prolongation of saccade latencies followingmicrosaccades Experimental Brain Research 169369ndash376 [PubMed]
Rucci M Iovin R Poletti M amp Santini F (2007)Miniature eye movements enhance fine spatial detailNature 447 851ndash854 [PubMed]
Salas C Navarro F Torres B amp Delgado-Garcia J M(1992) Effects of diazepam and D-amphetamine onrhythmic pattern of eye movements in goldfishNeuroreport 3 131ndash134 [PubMed]
Schipperheyn J J (1963) Respiratory eye movementsand perception of stationary objects in the frog ActaPhysiologica et Pharmacologica Neerlandica 12157ndash159 [PubMed]
Schipperheyn J J (1965) Contrast detection in frogrsquosretina Acta Physiologica et Pharmacologica Neer-landica 13 231ndash277 [PubMed]
Schipperheyn J J (1967) Influence of eye movements onretinal ganglion cell activity in frogs Acta Physi-ologica et Pharmacologica Neerlandica 14 506[PubMed]
Segev R Schneidman E Goodhouse J amp Berry M J2nd (2007) Role of eye movements in the retinalcode for a size discrimination task Journal ofNeurophysiology 98 1380ndash1391 [PubMed] [Article]
Sharpe C R (1972) The visibility and fading of thinlines visualized by their controlled movement acrossthe retina The Journal of Physiology 222 113ndash134[PubMed] [Article]
Shelley M McLaughlin D Shapley R amp Wielaard J(2002) States of high conductance in a large-scalemodel of the visual cortex Journal of ComputationalNeuroscience 13 93ndash109 [PubMed]
Skavenski A A Robinson D A Steinman R M ampTimberlake G T (1975) Miniature eye movementsof fixation in rhesus monkey Vision Research 151269ndash1273 [PubMed]
Snodderly D M (1987) Effects of light and darkenvironments on macaque and human fixational eyemovements Vision Research 27 401ndash415 [PubMed]
Snodderly D M Kagan I amp Gur M (2001) Selectiveactivation of visual cortex neurons by fixational eyemovements Implications for neural coding VisualNeuroscience 18 259ndash277 [PubMed]
Snodderly D M amp Kurtz D (1985) Eye position duringfixation tasks Comparison of macaque and humanVision Research 25 83ndash98 [PubMed]
Steinbach M J amp Money K E (1973) Eye movementsof the owl Vision Research 13 889ndash891 [PubMed]
Steinman R M Haddad G M Skavenski A A ampWyman D (1973) Miniature eye movement Science181 810ndash819 [PubMed]
Torres B Pastor A M Cabrera B Salas C ampDelgado-Garcia J M (1992) Afferents to theoculomotor nucleus in the goldfish (Carassius aur-atus) as revealed by retrograde labeling with horse-radish peroxidase Journal of Comparative Neurology324 449ndash461 [PubMed]
Troncoso X G Macknik S L amp Martinez-Conde S(2008) Microsaccades counteract perceptual filling-inJournal of Vision 8(14)15 1ndash9 httpjournalofvisionorg81415 doi10116781415 [Article]
Troncoso X G Macknik S L Otero-Millan J ampMartinez-Conde S (2008) Microsaccades driveillusory motion in the Enigma illusion Proceedingsof the National Academy of Sciences of the UnitedStates of America 105 16033ndash16038 [PubMed]
Tse P U Sheinberg D S amp Logothetis N K (2004)The distribution of microsaccade directions need notreveal the location of attention Psychological Science15 708ndash710
Turatto M Valsecchi M Tame L amp Betta E (2007)Microsaccades distinguish between global and localvisual processing Neuroreport 18 1015ndash1018[PubMed]
Uchida N Kepecs A amp Mainen Z F (2006) Seeing ata glance smelling in a whiff Rapid forms ofperceptual decision making Nature Reviews Neuro-science 7 485ndash491 [PubMed]
Valsecchi M Betta E amp Turatto M (2007) Visualoddballs induce prolonged microsaccadic inhibitionExperimental Brain Research 177 196ndash208[PubMed]
van Dam L C amp van Ee R (2006) Retinal image shiftsbut not eye movements per se cause alternations inawareness during binocular rivalry Journal of Vision6(11)3 1172ndash1179 httpjournalofvisionorg6113doi1011676113 [PubMed] [Article]
Van der Steen J amp Collewijn H (1984) Ocular stabilityin the horizontal frontal and sagittal planes in therabbit Experimental Brain Research 56 263ndash274[PubMed]
Van Gisbergen J A Robinson D A amp Gielen S(1981) A quantitative analysis of generation ofsaccadic eye movements by burst neurons Journalof Neurophysiology 45 417ndash442 [PubMed]
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 15
Van Gisbergen J A M amp Robinson D A (1977)Generation of micro and macrosaccades by burstneurons in the monkey In R Baker amp A Berthoz(Eds) Control of gaze by brain stem neurons (vol 1pp 301ndash308) New York ElsevierNorth-Holland
Wallman J Pettigrew J D amp Letelier J C (1994)The oscillatory saccades of birds Motoneural activ-ity and possible functions In B T Fuchs A FV Buttner amp D Zee (Eds) Contemporary ocularmotor and vestibular research A tribute to David ARobinson (pp 480ndash487) Stuttgart Germany ThiemePublishing
Williams P E amp Shapley R M (2007) A dynamicnonlinearity and spatial phase specificity in macaque
V1 neurons Journal of Neuroscience 27 5706ndash5718[PubMed] [Article]
Winterson B J amp Robinson D A (1975) Fixationby the alert but solitary cat Vision Research 151349ndash1352 [PubMed]
Yarbus A L (1967) Eye movements and vision(B Haigh Trans) New York Plenum Press
Zuber B L amp Stark L (1965) Microsaccades and thevelocityltamplitude relationship for saccadic eyemovements Science 150 1459ndash1460 [PubMed]
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 16

(Engbert 2006 Martinez-Conde 2006 Martinez-Condeet al 2004) However the differences and similarities infixational eye movements across species as well as theirpotential significance have been neglected so far Despitewide-reaching interest in this topic (Martinez-Conde ampMacknik 2007a) a comprehensive comparative reviewhas not been published Here we review the comparativedynamics the physiology and the perceptual consequen-ces of fixational eye movements across all species studiedto date To the best of our knowledge our review includesall Medline-indexed studies (as well as several bookchapters and other non-indexed publications) of fixationaleye movements in non-human vertebrates These span48 years of research from the first studies in the catpublished in 1960 (Hebbard amp Marg 1960 Pritchardamp Heron 1960) to the most recent studies in thesalamander and archer fish published in 2007 and 2008(Baccus Olveczky Manu amp Meister 2008 OlveczkyBaccus amp Meister 2007 Segev Schneidman Goodhouseamp Berry 2007) As a result of our analysis we havegenerated a table (Table 1) that summarizes the physicalparameters of the different types of fixational eye move-ment acrossnon-humanvertebratesWewill relatewheneverpossible fixational eye movements in non-humanvertebrates to human fixational eye movements
Fixational eye movements inmammals
Fixational eye movements in the primate
Fixational eye movements were originally discoveredand characterized in human subjects (Ditchburn ampGinsborg 1952 1953 Ratliff amp Riggs 1950) Humanfixational eye movements comprise three main typesmicrosaccades (called ldquoflicksrdquo in early studies) drift andtremor (Figure 1) Microsaccades are the fastest andlargest of the three types of fixational eye movements(Supplementary Video 1) They travel in a straighttrajectory carrying the retinal image across a range ofseveral dozen to several hundred photoreceptor widthsDrifts are slow curvy motions that occur between micro-saccades and tremor is a very fast (Egrave90 Hz) andextremely small oscillation (about the diameter of a fovealcone) superimposed on drifts (see Tables 1ndash3 of Martinez-Conde et al 2004 for a review of fixational eye move-ment parameters in humans)Fixational eye movements in old-world monkeys are
very similar to human fixational eye movements (Martinez-Conde 2006 Martinez-Conde et al 2000 2002Skavenski Robinson Steinman amp Timberlake 1975Snodderly 1987 Snodderly amp Kurtz 1985) Severalmacaque species have been studied to date Macaca
Typeofeye
move
ment
Amplitude
Frequency
intersacc
adic
interval
Duratio
nMax
speed
Mean
speed
Conjugate
Study
Amphibians
Salamander
Tremor-lik
eAbout
125
min
ndashndash
ndashndash
ndashManteuffe
letal(1977)
Respiratory
Upto
30min
ndashndash
ndashndash
ndashManteuffe
letal(1977)
Fish
Goldfish
Drift
ndashndash
01ndash55
sec
e1degsec
036degsec
NoSim
ulta
neous
inboth
eye
sbut
conve
rgentin
thelight
Mensh
etal(2004)
Drift
Typ
ically
G5deg
ndashndash
ndashndash
NoSim
ulta
neous
inboth
eye
sbut
conve
rgent
Easter(1971)
ArcherFish
Microsaccade
ndashldquorarelyrdquofound
ndashndash
ndashndash
Segevetal(2007)
Drift
ndash02ndash1Hz
133se
ce0
4degsec
ndashndash
Segevetal(2007)
Tremor
01ndash02
deg
5Hz
ndash4degsec
~1degsec
ndashSegevetal(2007)
Table
1Characteris
ticsoffixatio
naleye
move
ments
innon-humanve
rtebrates
acc
ordingto
differentstudies
NoteaCalculatedfrom
horiz
ontalandve
rticalco
mponents
bVectorva
luesfrom
H-componentsaco
nve
rsionfactoroffrac342
hasbeenass
umed
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 4
mulatta (Goffart Quinet Chavane amp Masson 2006Horwitz amp Albright 2003 Leopold amp Logothetis1998 Martinez-Conde 2006 Martinez-Conde et al 20002002 Motter amp Poggio 1984 Skavenski et al 1975Snodderly Kagan amp Gur 2001 Steinman HaddadSkavenski amp Wyman 1973) Macaca fascicularis(Snodderly 1987 Snodderly et al 2001 Snodderly ampKurtz 1985) and Macaca nemestrina (Bair amp OrsquoKeefe1998) with no substantial differences in the spatiotemporalcharacteristics of fixational eye movements (rate velocityand magnitude profiles etc) between them Microsaccadevelocities are parametrically related to microsaccadeamplitudes following the ldquomain sequencerdquo in both ma-caques (Martinez-Conde et al 2000) and humans (Engbert2006 Martinez-Conde 2006 Martinez-Conde et al 2006Zuber amp Stark 1965) Fixational microsaccades have alsobeen observed in the baboon (Papio papio) with ampli-tudes up to 25 degrees (Marchetti Gauthier amp Pellet1983)It is important to note that microsaccades cannot be
defined according to their amplitude alone because smallexploratory or voluntary saccades can be the same size asmicrosaccades Microsaccades can be defined only opera-tionally as the involuntary saccades that are producedwhile the subject attempts to fixate (Martinez-Conde2006) This ldquoattempt to fixaterdquo includes both holding theeyes still and static while foveating the visual target andalso orienting the fovea toward a target It is worth notingthat microsaccade production may require the presence ofa visual andor attentional target (Otero-Millan et al2008)Thus there is no known physical parameter (or
combination of parameters) that separates saccades frommicrosaccades Mounting evidence points toward a com-mon neural generator of saccades and microsaccades(Engbert 2006 Martinez-Conde et al 2004 Otero-Millan et al 2008 Rolfs Kliegl amp Engbert 2008 RolfsLaubrock amp Kliegl 2006 Zuber amp Stark 1965)However one should note that most of the available datain support of a shared microsaccadendashsaccade generatorare indirect To date only two physiological studies havedirectly addressed the question of the oculomotor
mechanisms leading to the generation of microsaccadesIn those studies Van Gisbergen and colleagues foundthat putative motoneurons in the primate abducensnucleus and burst neurons in the nearby pontomedullaryreticular formation were similarly active during saccadesand microsaccades (Van Gisbergen Robinson amp Gielen1981 Van Gisbergen amp Robinson 1977) Future researchshould further explore the neural bases underlying thegeneration of microsaccades and other fixational eyemovementsSimultaneous recordings of microsaccades and neural
responses have been conducted at multiple levels of theprimate visual pathway including the lateral geniculatenucleus (LGN) area V1 and the extrastriate visual cortex(Bair amp OrsquoKeefe 1998 Leopold amp Logothetis 1998Martinez-Conde 2006 Martinez-Conde et al 20002002 Reppas Usrey amp Reid 2002 Snodderly et al2001) Microsaccades are predominantly excitatory at allthese levels leading to firing rate increases in both earlyand higher visual areas The firing rate increases followingmicrosaccades in the LGN and V1 are of visual origin(Figure 2) and result from the microsaccade-induceddisplacement of visual receptive fields over stationarystimuli Microsaccade-driven increases in firing rate tendto be clustered in tight bursts of spikes (Martinez-Conde2006 Martinez-Conde et al 2000 2002) A recent studyhas suggested that primate microsaccades may improvethe efficient sampling of fine spatial detail (Donner ampHemila 2007)Microsaccades counteract visual fading and increase
visibility during fixation in human subjects (Martinez-Conde et al 2006 Troncoso Macknik amp Martinez-Conde 2008) Numerous studies have also reported thathuman microsaccade rates andor directions are modulatedby cognitive processes such as the allocation of spatialattention (Betta amp Turatto 2006 Engbert amp Kliegl2003a 2003b 2004 Galfano et al 2004 Hafed amp Clark2002 Laubrock Engbert Rolfs amp Kliegl 2007 Otero-Millan et al 2008 Turatto Valsecchi Tame Betta 2007Valsecchi Betta amp Turatto 2007 but see also HorowitzFine Fencsik Yurgenson amp Wolfe 2007 and TseSheinberg amp Logothetis 2004)
Figure 1 Cartoon representation of fixational eye movements in humans and primates Microsaccades (straight and fast movements)drifts (curvy slow movements) and tremor (oscillations superimposed on drifts) transport the visual image across the retinal photoreceptormosaic From Martinez-Conde and Macknik (2007b)
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 5
Neural responses to drifts have received considerablyless attention than neural responses to microsaccades Thismay be due to the fact that drifts are more difficult tocharacterize objectively than microsaccades (which aremore easily detected by automatic algorithms that com-bine amplitude and velocity thresholds) Thus drifts areusually identified indirectly as the eye position changesthat occur between microsaccades (Snodderly et al2001) However this method has the potential flaw thatone may unintentionally attribute non-drift-related activity(for instance undetected tremor) to drifts Drifts mayincrease firing in a subset of primate V1 neurons(Snodderly et al 2001) However they generate lessvariability in neuronal responses than a combination ofdrifts and microsaccades (Gur Beylin amp Snodderly1997) No studies to date have recorded primate neuronalresponses in correlation to tremor (but see Hennig ampWorgotter 2007 for the responses of a model macaqueretina to simulated drift and tremor)
Fixational eye movements in the cat
The catrsquos fixational eye movements have been inves-tigated in a handful of studies and their conclusions
remain somewhat controversial In 1960 Hebbard andMarg (1960) found fixational eye movements in the cat tobe quite comparable to those in humans although catmicrosaccades were scarcer and smaller than in humansPritchard and Heron (1960) also found that the catrsquosfixational eye movements included microsaccades driftsand tremor Microsaccades were much rarer in the cat thanin humans however possibly related to the fact that thecat lacks a well-developed fovea Delgado-Garcia delPozo and Baker (1986) also observed some putativemicrosaccades in the alert behaving cat Other studiesfound no microsaccades in the cat (Conway Timberlakeamp Skavenski 1981 Winterson amp Robinson 1975)However in these last studies microsaccades were definedas ldquosaccades smaller than 10 minutes of arcrdquo which iswell below the average microsaccade amplitude found inrecent human and primate studies (see Martinez-Condeet al 2004 for a review of microsaccade parameters inhumans and primates) More recently several studies haveaddressed the contribution of fixational eye movements toneural responses in the catrsquos visual system HennigKerscher Funke and Worgotter (2002) found increasedresponses from neurons in cat areas 17 and 18 duringstimulus motions that mimicked fixational eye move-ments Using a model retina they also found that
1 simulated fixational eye movements increased neu-ral responses and
2 simulated tremor improved spatial acuity
Hennig and Worgotter (2004) later reported that acombination of tremor and microsaccades improved theperformance of simulated ganglion cells in a hyperacuitytask in the retinal periphery (but not in the central retina)Miller Denning George Marshak and Kenyon (2006)also investigated the effects of simulated tremor on amodel of the catrsquos retina and suggested that tremor mayselectively enhance the processing of large stimuli
Fixational eye movements in the rabbit
In contrast to foveate species such as primates (includ-ing humans) and cats (which have an area centralis if nota proper fovea) the rabbit (Oryctolagus cuniculus) dis-plays few spontaneous eye movements especially in theabsence of head motion (Collewijn 1977 Collewijn ampvan der Mark 1972 Fuller 1980 1981 Van der Steen ampCollewijn 1984) In normal visual conditions the rabbitrsquoseyes are very stable but not perfectly immobile Smallamplitude tremor can be observed as well as very slowdrift Microsaccades have not been observed Howeverdrift is occasionally corrected by fast saccadic movements(Collewijn 1977 Collewijn amp van der Mark 1972 Vander Steen amp Collewijn 1984) These differences may berelated to the fact that the rabbit does not have a fovea butan elongated horizontal streak with elevated ganglion cell
Figure 2 Microsaccades drive neural activity in primate area V1Microsaccades lead to firing rate increases in V1 neurons (n = 308)of the awake primate In the absence of visual stimulationmicrosaccades do not increase neural activity in area V1 (n = 37neurons) The resultsVshown here for the average of all neuronsVwere consistent with responses from individual neurons (data notshown) Modified from Martinez-Conde et al (2000 2002)
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 6
counts Olveczky Baccus and Meister (2003) recordedfrom ganglion cells in the rabbit and salamander retinaswhile presenting visual stimuli that were jittered tosimulate fixational eye movements (based on parametersfrom previously published studies by Manteuffel PlasaSommer amp Wess 1977 and Van der Steen amp Collewijn1984) In both species fixational eye movements helpedsegregate a moving foreground from a complex back-ground in a population of ganglion cells
Fixational eye movements inbirds
Fixational eye movements in the pigeon
There are some remarkable differences between fixa-tional eye movements in birds and mammals Nye (1969)identified four types of fixational eye movements in thepigeon (Columba livia) flicks (ie microsaccades)impulse movements drift and oscillations No tremorwas detected Microsaccade frequency was very lowrelative to humans Impulse movements occurred morefrequently usually between bursts of oscillations withamplitudes ranging up to 2 degrees Drift velocitiesranged from 1 to 5 degrees per second Both micro-saccades and drifts occurred in response to movingstimuli Oscillations occurred in short bursts at rates ofabout 30 Hz and could be quite large (up to severaldegrees of amplitude) Bloch Rivaud and Martinoya(1984) reported microsaccades drifts and oscillationsldquosimilar to those described by Nyerdquo but none of thesewere analyzed in detailCyclotorsional oscillations are an integral part of avian
saccades (Wallman Pettigrew amp Letelier 1994) Thusone might speculate that oscillations are associated withavian microsaccades as well especially if saccades andmicrosaccades share a common generator (Engbert 2006Otero-Millan et al 2008 Rolfs et al 2008 2006 Zuberamp Stark 1965) However this possibility has not yet beenaddressed in the literature The function of oscillations isunknown (Carpenter 1988) but one hypothesis is thatthey serve to facilitate the delivery of arterial blood to theocular capillary bed (Pettigrew Wallman amp Wildsoet1990 Wallman et al 1994)
Fixational eye movements in the owl
Steinbach and Money (1973) recorded fixational eyemovements in the great horned owl (Bubo virginianus)a species previously thought incapable of moving itseyes The owlrsquos fixational eye movements includeflicks or microsaccades drift tremor and oscillations
Microsaccades seemed correlated to visual events ofinterest to the owl they were 1 deg or less in amplitudeand had peak velocities of about 10 degsec Drifts wereslow motions of the eye in between fixations withamplitudes of 1 degree or less Tremor amplitude wasabout 1 arcmin with frequencies around 20 Hz High-frequency oscillations (25 Hz) of short duration (025 sec)were very similar to those previously found in the pigeon(Nye 1969) No studies to date have examined theperceptual or physiological effects of fixational eyemovements in any bird species
Fixational eye movements inreptiles
Fixational eye movements in the turtle
Greschner Bongard Rujan and Ammermuller (2002)identified two types of fixational eye movements in theturtle (Pseudemys scripta elegans) which they calledldquodrift-likerdquo and ldquoperiodicrdquo Drift-like motions were smalland slow (with velocities comparable to human drift) andwere superimposed by larger and faster periodic componentsPeriodic motion had retinal amplitudes on the order of thediameter of a photoreceptor which are comparable to thoseof human tremor On the other hand periodic motionrsquosfrequency (5 Hz) velocity and orderly relationship betweenvelocity and amplitude are closer to those of human micro-saccades (see Martinez-Conde et al 2004 for a review oftremor and microsaccade parameters in humans)Simulated periodic motion generated strong synchronous
firing in the turtlersquos retina whereas simulated drift-likemotion had little effect (Figure 3) These results agree withprevious physiological and modeling studies in the primatevisual system in which strong neural transients wereobserved in response to microsaccades (Donner amp Hemila2007 Martinez-Conde 2006 Martinez-Conde et al 20002002) Such neural transients may underlie the behavior ofcortical neurons as coincidence detectors (ShelleyMcLaughlin Shapley amp Wielaard 2002 Williams ampShapley 2007) Moreover neural transients to stimulionsets and terminations (similar to those produced bymicrosaccades in the primate visual system Martinez-Conde 2006 Martinez-Conde et al 2000 2002) havebeen related to target visibility in visual masking para-digms (Macknik amp Livingstone 1998 Macknik ampMartinez-Conde 2004 Macknik Martinez-Conde ampHaglund 2000)In the turtle retina periodic motion-driven neurons with
receptive fields located along contrast borders weremoreover synchronized and reliably indicated the preced-ing motions The authors proposed that this synchroniza-tion of retinal activity could be used by the brain to
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 7
improve the estimation of stimulus features such asspatial frequency (Greschner et al 2002) This findingcould have a parallel in the primate visual system Donnerand Hemila (2007) recently proposed that microsaccadessignificantly ldquore-sharpenrdquo the image and improve spatialresolution in primates Rucci Iovin Poletti and Santini(2007) also found that fixational eye movements enhancefine spatial detail in human subjects
Fixational eye movements in the chameleon
Fixational eye movements in the chameleon have notbeen studied in detail However Gioanni and colleaguesfound that the chameleon (Chamaleo chamaleo) makessaccades during fixation The authors speculated that theseprobably correspond to fixational microsaccades found inother foveate species (Gioanni Bennis amp Sansonetti1993)
Fixational eye movements inamphibians
Fixational (and more generally spontaneous) eye move-ments vary widely among amphibians The frog (anafoveate amphibian) shows a general lack of eye andhead movements except for those that stabilize the retinalimage Dieringer and colleagues (Dieringer amp Daunicht1986 Dieringer amp Precht 1982 Dieringer Precht ampBlight 1982) found that the grass frog (Rana temporaria)produces no spontaneous eye movements in the absenceof head movements with the possible exception of tremor(similarly to the rabbit another afoveate species Van derSteen amp Collewijn 1984) It is likely that the frogrsquos lack ofeye movements leads to fast visual fading of all stationaryobjects in its field of view (Ditchburn amp Ginsborg 1952Yarbus 1967) Lettvin Maturana MsCulloch and Pitts
Figure 3 Fixational eye movements increase retinal activity in the turtle (A) Continuous recording from 54 retinal ganglion cells in theturtle during darkness after switching on a stationary grating and during wobbling of this grating The grating was wobbled to simulateperiodic (ie tremor-like) eye movements (B) Responses of a single ganglion cell to a drifting contrast border of velocity comparable toocular drifts (C) When the drift is superimposed with wobbling the responses of the cell shown in (B) increase dramatically FromGreschner et al (2002) and modified by Martinez-Conde et al (2004)
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 8
(1968) stated that a frog ldquowill starve to death surroundedby food if it is not movingrdquo (but see Dieringer amp Daunicht1986 for a counterpoint) Small head motions or eyemovements correlated with respiratory motions maydiminish fading to some extent (Dieringer amp Precht1982 Schipperheyn 1963 1965 1967)As a side note the frogrsquos respiratory eye movements
play no role in the respiratory function ldquoin the course ofeach respiratory cycle increase of pressure in the frogrsquosbuccal cavity protrudes the eye and causes the opticalaxis to deviate upwards over 25ndash35 minutes of arcrdquo(Schipperheyn 1963) That is respiratory eye movementsare merely synchronized with respirationAutrum (1959) found no evidence of respiratory eye
movements in the frog Ewert andBorchers (1974) observedthat stationary stimuli elicit no responses from visualneurons in the common toad (Bufo Bufo L) regardless ofrespiratory pressure changes in the buccal cavity
Fixational eye movements in the salamander
Manteuffel et al (1977) identified two types of fixa-tional eye movements in salamanders tremor-like andrespiratory eye movements Tremor-like is a rapid move-ment of small amplitude Torsional eye movements resultfrom the animalrsquos breathing (also found in the frog bySchipperheyn 1963) Additionally flicks are occasionallyobserved which rapidly change the position of gaze It isunclear whether such flicks may generally correspond towhat we now call microsaccades andor to voluntarysaccades Based on these observations the authors pro-posed that unlike frogs and toads (Autrum 1959 Ewert ampBorchers 1974 Lettvin et al 1968) salamanders cancontinuously perceive stationary objects (Manteuffel et al1977) Fixational eye movements in the salamander (Tigersalamander Ambystoma tigrinum) may also help segre-gate object and background motion in the retina(Olveczky et al 2003 see also Fixational eye movementsin the rabbit section) and even work to emphasize novelvisual features (Olveczky et al 2007 see Baccus et al2008 for details on the retinal neurons and synapses thatmay underlie the computation of differential motionbetween an object and its background)
Fixational eye movements in fish
Spontaneous eye movements in the goldfish
Eye movements during fixation have been studied inboth afoveate (goldfish Carassius auratus) and foveate(archer fish Toxotes chatareus) fish species The goldfisha long-standing model organism in vision and oculomotor
research (Aksay et al 2003 2007 Cabrera Torres PasaroPastor amp Delgado-Garcia 1992 Pastor Torres Delgado-Garcia amp Baker 1991 Salas Navarro Torres ampDelgado-Garcia 1992 Torres Pastor Cabrera Salas ampDelgado-Garcia 1992) produces three types of sponta-neous eye movements saccades stretch and drift (Easter1971 Mensh Aksay Lee Seung amp Tank 2004) Weshould note that in the motionless goldfish eye move-ments are very infrequent Those that do occur consistmainly of saccades slow drifts are very rare (Easter Johnsamp Heckenlively 1974) During saccades (identified bythresholding the eye velocity at 5 degsec) both eyes moverapidly simultaneously and usually in the same direction(Mensh et al 2004) Stretches occur every 2ndash3 min and arecharacterized by a brief simultaneous large excursion ofboth eyes to extreme temporal positions (Easter 1971Mensh et al 2004) The function of stretches is not known(Mensh et al 2004) Because fixations were defined asthose periods that were free of saccadic and stretchmovements (Easter 1971) only drifts (from the threetypes of spontaneous movements) can technically beconsidered ldquofixational eye movementsrdquo However thegoldfish saccadic main sequence includes saccades withequivalent amplitudes and velocities to those found inhuman microsaccades (Mensh et al 2004 Zuber amp Stark1965) Indeed microsaccades have been identified in other(foveate) fish species although they are rarer than inhumans (Segev et al 2007) Small saccades in thegoldfish tend to be divergent whereas large saccadestend to be convergent (Mensh et al 2004) Drift occurssimultaneously in both eyes usually in the samedirection Drift velocities are almost exclusively e1 degsec(Mensh et al 2004) No tremor has been identified in thegoldfish
Spontaneous eye movements in teleost fish
Spontaneous eye movements have been studied in theNew Zealand parore (Girella tricuspidata MontgomeryMcVean amp McCarthy 1983) The pattern of eye move-ments was similar to that previously described in goldfish(Easter 1971 Hermann amp Constantine 1971)
Fixational eye movements in the archer fish
A recent study in the archer fish (Segev et al 2007) hasprovided the first perceptual and physiological correlatesof fixational eye movements in any fish species Archerfish have remarkable visual abilities they shoot downinsects resting on foliageVas well as stationary targetscovering a single retinal photoreceptorVwith squirts ofwater from their mouths they predict the location atwhich the insect will splash down in G100 msec and theydeduce the absolute size of an object presented at differentviewing distances (Supplementary Videos 2 and 3) Segev
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 9
et al (2007) identified three types of fixational eyemovements in the archer fish microsaccades drift andtremor Microsaccades were rare and were not analyzed indetail Drift velocities were e040 degsec Tremor was ahigh-frequency oscillatory movement (the primary com-ponent was 5 Hz) with peak-to-peak amplitude of 01ndash02 deg (corresponding to an image displacement of 1ndash2photoreceptors) and velocity of about 1 degsec (Figure 4)Whereas the amplitude and velocity of tremor in thearcher fish is comparable to those in humans its frequencyis much lower (5 Hz vs 940 Hz) Interestingly archer fishtremor is very similar both in amplitude (photoreceptorwidths covered) as well as in frequency (5 Hz) to thetremor-like periodic motion in the turtle (discussedearlier Greschner et al 2002)Segev et al (2007) trained archer fish to perform a size
discrimination task the fish had to distinguish a medium-sized target from larger and smaller distracters (theyindicated their choice by shooting a jet of water) To relatethis behavior to responses from retinal neurons theauthors recorded from populations of retinal ganglioncells while presenting objects of different sizes moved tosimulate saccades drifts and tremor Drifts and tremorelicited weak neuronal responses that were not veryinformative about the spatial structure of the stimulus(although they did provide some information about target
size in agreement with the results of Greschner et al2002 in the turtle) Moreover performance was similar fortremor alone and for tremor plus drift indicating that driftplays a negligible role in revealing spatial structure (alsoin agreement with Greschner et al 2002)When present the firing rate was phase-locked to
tremor Saccades were more effective than drifts or tremorin driving informative responses from the ganglion cells(consistent with theoretical and physiological studies ofmicrosaccade-driven neuronal activity in the primateretina and LGN Donner amp Hemila 2007 Martinez-Conde et al 2002) Segev et al (2007) proposed that inprimates both saccades and microsaccades (which arerare in the archer fish) may contribute to size discrim-ination tasks as well as enhance spatial detail whereasdrifts and tremor by themselves may not be sufficient toimprove visual perception However it is important toremember that central vision in humans can be maintainedby tremor andor drift even when microsaccades aresuppressed during strict fixation (Martinez-Conde 2006Martinez-Conde et al 2004 2006) On the other handone cannot rule out the possibility that if one couldcompletely eliminate drifts and tremor microsaccadesmight then suffice to sustain both central and peripheralvision in primates and humans Future experiments areneeded to further explore these ideas
Figure 4 Archer fish eye movements (A) Example trace of eye position as a function of time as measured by search coil (B) Example ofa single 15-s fixation superimposed on a grid representing the photoreceptor mosaic The back and forth movement is caused by tremor(C) Distribution of fixation durations (taken as time interval between saccades) (D) Power spectrum of horizontal fixational eyemovements (E) Distribution of tremor amplitude (data taken from 1 animal in a 15-min session) Modified from Segev et al (2007)
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 10
Discussion
The vast majority of eye movement studies conductedin non-human species to date have not analyzed eyeposition data in the range of fixational eye movementsThus eye movements during fixation have been reportedin only a handful of vertebrates Most of these studieshave concentrated solely on the physical parameters ofthe fixational eye movements without addressing theirphysiological and perceptual consequences Fortunatelythis trend has started to reverse in recent years with agrowing list of papers addressing the effects of non-human fixational eye movements in physiology andperception This is an important development A largeamount of all psychophysical and physiological visualresearch has been conducted during conditions of visualfixation Therefore understanding the precise physiolog-ical and perceptual contributions of fixational eye move-ments may be critical to the interpretation of previous andfuture vision researchThere are some interesting differences and similarities
in the types of fixational eye movements produced bydifferent species For instance ocular tremor is possiblyubiquitous to all vertebrates The fact that tremor has notbeen observed in all species studied could be due to thetechnical difficulty of recording tremor accurately (ietremor amplitudes and frequencies are usually in the rangeof the recording systemrsquos noise) Drift has also beenrecorded in most vertebrates and it may be as omnipre-sent as tremor Conversely microsaccades seem mostimportant in species with foveal vision (ie primates andhumans) whereas they are scarce or absent in afoveatespecies (ie rabbits frogs goldfish) A link betweenmicrosaccades and foveal vision may reflect the two-pronged functional significance of microsaccades inhumans and primates first microsaccades may benecessary to correct the slow drift of the eye and bringback the objectregion of interest to the retinal area wherephotoreceptor density is highest (Cornsweet 1956) Thecorrective role of human microsaccades has been demon-strated in recent studies (Engbert amp Kliegl 2004 Engbertamp Mergenthaler 2006) Second the dynamics of micro-saccades themselves may improve spatial resolution(Donner amp Hemila 2005)In reviewing the patterns of fixational eye movements
across vertebrates it is tempting to draw parallels withother sensory modalities Fixational eye movementsrefresh the retinal images during fixation and thus preventneural adaptation and visual fading But this mechanism isnot circumscribed to the visual system Sniffs in rodentolfaction discretely sample sensory information every200ndash300 msec and thus are similar in their temporaldynamics to primate saccades (Otero-Millan et al 2008Uchida Kepecs amp Mainen 2006) and microsaccades(Otero-Millan et al 2008) A similar mode of discretesampling may also be at play when objects are recognized
through tactile information for instance if we sweep ourfingertips over an objectrsquos surface with our eyes closed orwhen blind individuals read Braille scriptOne intriguing possibility that merits further experi-
ments and analyses is the potential correlation between thespatiotemporal parameters of fixational eye movements(such as those summarized in Table 1) and the adaptationproperties of visual neurons across species For instancethe greater ocular stability of some species (rabbitgoldfish frog etc) versus others (primates) could beassociated with some particular adaptive properties oftheir visual cells Thus future comparative studies shouldpay attention to the response properties of visual cellstriggered by fixational eye movements in the presence ofstationary stimuli Other potential factors underlying thevariation in fixational eye movements across species couldbe differences in oculomotor control at the level of the eyemuscles or perhaps differences in retinal organizationFuture research will further improve our understanding
of fixational eye movements and the neural activity thatmaintains the visibility of stationary visual scenes inhuman and non-human species One interesting questionthat remains to be addressed concerns the role of tremor invision and visual physiology Because tremor frequenciesare well over the flicker fusion frequencies in humans ithas been argued that tremor of the visual image would beineffective as a stimulus (Ditchburn 1955 Gerrits ampVendrik 1970 Sharpe 1972) However recent studieshave found simulated tremor to enhance visual processingand improve visual acuity in a model retina (Hennig et al2002 Miller et al 2006) Follow-up research should beaimed to replicate these findings in actual neurons withcorrelated perceptual reports One should also note thatearly visual neurons can follow high-frequency flickeringthat surpasses the perceptual threshold for flicker fusion(Martinez-Conde et al 2002) Thus high-frequencytremor could be adequate to maintain activity in the earlyvisual system which then may lead to visual perception inhigher visual areas Another significant question thatremains unanswered concerns the oculomotor bases ofthe various types of fixational eye movements acrossspecies To date there is too little information on thisimportant topic to draw a conclusion
Conclusions
We have reviewed the parameters of fixational eyemovements in non-human mammals birds reptilesamphibians and fish as well as their consequences forphysiology and perception Fixational eye movementsand microsaccades in particular appear to be mostimportant in foveate vs afoveate species Animals lackingfixational eye movements such as afoveate frogs andtoads may be incapable of seeing stationary objects
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 11
during visual fixation Future research should furtherexplore the perceptual effects of fixational eye movementsand determine the oculomotor mechanisms responsible fortheir generation across species
Acknowledgments
We thank Mona Stewart and Isabel Gomez-Caraballo fortechnical assistance Jorge Otero-Millan for programmingassistance and Dr Xoana G Troncoso for comments on themanuscript This study was funded through grants fromthe Barrow Neurological Foundation (to SMC and SLM)the National Science Foundation (Award 0643306 toSMC) the Arizona Biomedical Research Commission(Award 07-102 to SMC) and Science Foundation Arizona(SLM)
Commercial relationships noneCorresponding author Susana Martinez-Conde PhDEmail smartneuralcorrelatecomAddress Barrow Neurological Institute 350 W ThomasRd Phoenix AZ 85013 USA
References
Aksay E Major G Goldman M S Baker R SeungH S amp Tank D W (2003) History dependence ofrate covariation between neurons during persistentactivity in an oculomotor integrator Cerebral Cortex13 1173ndash1184 [PubMed] [Article]
Aksay E Olasagasti I Mensh B D Baker RGoldman M S amp Tank D W (2007) Functionaldissection of circuitry in a neural integrator NatureNeuroscience 10 494ndash504 [PubMed]
Autrum H (1959) Das Fehlen unwillkurlicher Augenbe-wegungen beim Frosch Naturwissenschaften 46435
Baccus S A Olveczky B P Manu M amp Meister M(2008) A retinal circuit that computes object motionJournal of Neuroscience 28 6807ndash6817 [PubMed]
Bair W amp OrsquoKeefe L P (1998) The influence offixational eye movements on the response of neuronsin area MT of the macaque Visual Neuroscience 15779ndash786 [PubMed]
Betta E amp Turatto M (2006) Are you ready I can tellby looking at your microsaccades Neuroreport 171001ndash1004 [PubMed]
Bloch S Rivaud S amp Martinoya C (1984) Compar-ing frontal and lateral viewing in the pigeon IIIDifferent patterns of eye movements for binocularand monocular fixation Behavioural Brain Research13 173ndash182 [PubMed]
Cabrera B Torres B Pasaro R Pastor A M ampDelgado-Garcia J M (1992) A morphological studyof abducens nucleus motoneurons and internuclearneurons in the goldfish (Carassius auratus) BrainResearch Bulletin 28 137ndash144 [PubMed]
Carpenter R H S (1988) Movements of the eyes(2nd ed) Pion London
Collewijn H (1977) Eye- and head movements infreely moving rabbits The Journal of Physiology266 471ndash498 [PubMed] [Article]
Collewijn H amp van der Mark F (1972) Ocular stabilityin variable visual feedback conditions in the rabbitBrain Research 36 47ndash57 [PubMed]
Conway J L Timberlake G T amp Skavenski A A(1981) Oculomotor changes in cats reared withoutexperiencing continuous retinal image motion Exper-imental Brain Research 43 229ndash232 [PubMed]
Cornsweet T N (1956) Determination of the stimuli forinvoluntary drifts and saccadic eye movements Jour-nal of the Optical Society of America 46 987ndash993[PubMed]
Delgado-Garcia J M del Pozo F amp Baker R (1986)Behavior of neurons in the abducens nucleus of thealert catVII Internuclear neurons Neuroscience 17953ndash973 [PubMed]
Dieringer N amp Daunicht W J (1986) Image fadingVAproblem for frogs Naturwissenschaften 73 330ndash332[PubMed]
Dieringer N amp Precht W (1982) Compensatory headand eye movements in the frog and their contributionto stabilization of gaze Experimental BrainResearch 47 394ndash406 [PubMed]
Dieringer N Precht W amp Blight A R (1982)Resetting fast phases of head and eye and theirlinkage in the frog Experimental Brain Research 47407ndash416 [PubMed]
Ditchburn R W (1955) Eye-movements in relation toretinal action Optica Acta 1 171ndash176
Ditchburn R W amp Ginsborg B L (1952) Vision with astabilized retinal image Nature 170 36ndash37[PubMed]
Ditchburn R W amp Ginsborg B L (1953) Involuntaryeye movements during fixation The Journal ofPhysiology 119 1ndash17 [PubMed] [Article]
Donner K amp Hemila S (2007) Modelling the effect ofmicrosaccades on retinal responses to stationarycontrast patterns Vision Research 47 1166ndash1177[PubMed]
Donner K K amp Hemila S (2005) The possible purposeof microsaccades Paper presented at the EuropeanConference on Visual Perception A Coruna Spain
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 12
Easter S S Jr (1971) Spontaneous eye movements inrestrained goldfish Vision Research 11 333ndash342[PubMed]
Easter S S Jr Johns P R amp Heckenlively D R(1974) Horizontal compensatory eye movements ingoldfish (Carassius auratus) I The normal animalJournal of Comparative Physiology 92 23ndash35
Engbert R (2006) Microsaccades A microcosm forresearch on oculomotor control attention and visualperception Progress Brain Research 154 177ndash192[PubMed]
Engbert R amp Kliegl R (2003a) Binocular coordinationin microsaccades In R R J Hyona amp H Deubel(Eds) The mindrsquos eyes Cognitive and appliedaspects of eye movements (pp 103ndash117) OxfordUK Elsevier
Engbert R amp Kliegl R (2003b) Microsaccades uncoverthe orientation of covert attention Vision Research43 1035ndash1045 [PubMed]
Engbert R amp Kliegl R (2004) Microsaccades keep theeyesrsquo balance during fixation Psychological Science15 431ndash436 [PubMed]
Engbert R amp Mergenthaler K (2006) Microsaccadesare triggered by low retinal image slip Proceedingsof the National Academy of Sciences of the UnitedStates of America 103 7192ndash7197 [PubMed][Article]
Ewert J P amp Borchers H W (1974) Antwort vonretinalen Ganglienzellen bei freibeweglichen Kroten(Bufo Bufo L) Journal of Comparative and Physi-ology 92 117ndash130
Fuller J H (1980) Linkage of eye and head movementsin the alert rabbit Brain Research 194 219ndash222[PubMed]
Fuller J H (1981) Eye and head movements duringvestibular stimulation in the alert rabbit BrainResearch 205 363ndash381 [PubMed]
Galfano G Betta E amp Turatto M (2004) Inhibition ofreturn in microsaccades Experimental BrainResearch 159 400ndash404 [PubMed]
Gerrits H J amp Vendrik A J (1970) Artificial move-ments of a stabilized image Vision Research 101443ndash1456 [PubMed]
Gioanni H Bennis M amp Sansonetti A (1993) Visualand vestibular reflexes that stabilize gaze in thechameleon Visual Neuroscience 10 947ndash956[PubMed]
Goffart L Quinet J Chavane F amp Masson G S(2006) Influence of background illumination onfixation and visually guided saccades in the rhesusmonkey Vision Research 46 149ndash162 [PubMed]
Greschner M Bongard M Rujan P amp Ammermuller J(2002) Retinal ganglion cell synchronization by fixa-tional eye movements improves feature estimationNature Neuroscience 5 341ndash347 [PubMed]
Gur M Beylin A amp Snodderly D M (1997)Response variability of neurons in primary visualcortex (V1) of alert monkeys Journal of Neuro-science 17 2914ndash2920 [PubMed] [Article]
Hafed Z M amp Clark J J (2002) Microsaccades as anovert measure of covert attention shifts VisionResearch 42 2533ndash2545 [PubMed]
Hebbard F W amp Marg E (1960) Physiologicalnystagmus in the cat Journal of the Optical Societyof America 50 151ndash155 [PubMed]
Hennig M A Kerscher N J Funke K amp Worgotter F(2002) Stochastic resonance in visual cortical neuronsDoes the eye-tremor actually improve visual acuityNeurocomputing 44ndash46 115ndash120
Hennig M H amp Worgotter F (2004) Eye micro-movements improve stimulus detection beyond theNyquist limit in the peripheral retina In Advances inNeural Information Processing Systems (NIPS2003)(vol 16) Denver CO MIT Press
Hennig M H amp Worgotter F (2007) Effects offixational eye movements on retinal ganglion cellresponses A modelling study Frontiers in Computa-tional Neuroscience 1 2 [PubMed] [Article]
Hermann H T amp Constantine M M (1971) Eyemovements in the goldfish Vision Research 11313ndash331 [PubMed]
Horowitz T S Fine E M Fencsik D E Yurgenson Samp Wolfe J M (2007) Fixational eye movements arenot an index of covert attention PsychologicalScience 18 356ndash363 [PubMed]
Horwitz G D amp Albright T D (2003) Short-latencyfixational saccades induced by luminance incrementsJournal of Neurophysiology 90 1333ndash1339[PubMed] [Article]
Laubrock J Engbert R Rolfs M amp Kliegl R(2007) Microsaccades are an index of covertattention Commentary on Horowitz Fine FencsikYurgenson and Wolfe (2007) Psychological Sci-ence 18 364ndash366 discussion 367ndash368 [PubMed]
Leopold D A amp Logothetis N K (1998) Micro-saccades differentially modulate neural activity in thestriate and extrastriate visual cortex ExperimentalBrain Research 123 341ndash345 [PubMed]
Lettvin J Y Maturana H R MsCulloch W S amp PittsW H (1968) What the frogrsquos eye tells the frogrsquosbrain In W C C a M Balaban (Ed) The mind
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 13
Biological approaches to its functions (pp 233ndash258)John Wiley and Sons London
Macknik S L amp Livingstone M S (1998) Neuronalcorrelates of visibility and invisibility in the primatevisual system Nature Neuroscience 1 144ndash149[PubMed]
Macknik S L amp Martinez-Conde S (2004) The spatialand temporal effects of lateral inhibitory networksand their relevance to the visibility of spatiotemporaledges Neurocomputing 58ndash60 775ndash782
Macknik S L Martinez-Conde S amp Haglund M M(2000) The role of spatiotemporal edges in visibilityand visual masking Proceedings of the NationalAcademy of Sciences of the United States of America97 7556ndash7560 [PubMed] [Article]
Manteuffel G Plasa L Sommer T J amp Wess O(1977) Involuntary eye movements in salamandersNaturwissenschaften 64 533ndash534 [PubMed]
Marchetti E Gauthier G M amp Pellet J (1983)Cerebellar control of eye movements studied withinjection of harmaline in the trained baboon ArchivesItaliennes de Biologies 121 1ndash17 [PubMed]
Martinez-Conde S (2006) Fixational eye movements innormal and pathological vision Progress in BrainResearch 154 151ndash176 [PubMed]
Martinez-Conde S amp Macknik S L (2007a) Bobbingand perceiving Scientific American 297 16
Martinez-Conde S amp Macknik S L (2007b) Windowson the mind Scientific American 297 56ndash63[PubMed]
Martinez-Conde S Macknik S L amp Hubel D H(2000) Microsaccadic eye movements and firing ofsingle cells in the striate cortex of macaque monkeysNature Neuroscience 3 251ndash258 [PubMed]
Martinez-Conde S Macknik S L amp Hubel D H(2002) The function of bursts of spikes during visualfixation in the awake primate lateral geniculate nucleusand primary visual cortex Proceedings of theNational Academy of Sciences of the United Statesof America 99 13920ndash13925 [PubMed] [Article]
Martinez-Conde S Macknik S L amp Hubel D H(2004) The role of fixational eye movements invisual perception Nature Reviews Neuroscience 5229ndash240 [PubMed]
Martinez-Conde S Macknik S L Troncoso X G ampDyar T A (2006) Microsaccades counteract visualfading during fixation Neuron 49 297ndash305[PubMed] [Article]
Mensh B D Aksay E Lee D D Seung H S ampTank D W (2004) Spontaneous eye movements ingoldfish Oculomotor integrator performance plasti-
city and dependence on visual feedback VisionResearch 44 711ndash726 [PubMed]
Miller J A Denning K S George J S MarshakD W amp Kenyon G T (2006) A high frequencyresonance in the responses of retinal ganglion cells torapidly modulated stimuli A computer model VisualNeuroscience 23 779ndash794 [PubMed]
Montgomery J C McVean A R amp McCarthy D(1983) The effects of lowered temperature onspontaneous eye movements in a teleost fish Com-parative Biochemistry and Physiology A Compara-tive Physiology 75 363ndash368 [PubMed]
Motter B C amp Poggio G F (1984) Binocularfixation in the rhesus monkey Spatial and temporalcharacteristics Experimental Brain Research 54304ndash314 [PubMed]
Murakami I amp Cavanagh P (1998) A jitter after-effectreveals motion-based stabilization of vision Nature395 798ndash801 [PubMed]
Nye P W (1969) The monocular eye movements of thepigeon Vision Research 9 133ndash144 [PubMed]
Olveczky B P Baccus S A amp Meister M (2003)Segregation of object and background motion in theretina Nature 423 401ndash408 [PubMed]
Olveczky B P Baccus S A amp Meister M (2007)Retinal adaptation to object motion Neuron 56689ndash700 [PubMed] [Article]
Otero-Millan J Troncoso X Macknik S L Serrano-Pedraza I amp Martinez-Conde S (2008) Saccadesand microsaccades during visual fixation explorationand search Foundations for a common saccadicgenerator Journal of Vision 8(14)21 1ndash18 httpjournalofvisionorg81421 doi10116781421[PubMed] [Article]
Pastor A M Torres B Delgado-Garcia J M ampBaker R (1991) Discharge characteristics of medialrectus and abducens motoneurons in the goldfishJournal of Neurophysiology 66 2125ndash2140 [PubMed]
Pettigrew J D Wallman J amp Wildsoet C F (1990)Saccadic oscillations facilitate ocular perfusion fromthe avian pecten Nature 343 362ndash363 [PubMed]
Pritchard R M amp Heron W (1960) Small eye move-ments of the cat Canadian Journal of Psychology14 131ndash137 [PubMed]
Ratliff F amp Riggs L A (1950) Involuntary motions ofthe eye during monocular fixation Journal of Exper-imental Psychology 40 687ndash701 [PubMed]
Reppas J B UsreyWM amp Reid R C (2002) Saccadiceye movements modulate visual responses in thelateral geniculate nucleus Neuron 35 961ndash974[PubMed] [Article]
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 14
Riggs L A amp Ratliff F (1952) The effects of counter-acting the normal movements of the eye Journal ofthe Optical Society of America 42 872ndash873
Rolfs M Kliegl R amp Engbert R (2008) Toward amodel of microsaccade generation The case ofmicrosaccadic inhibition Journal of Vision 8(11)51ndash23 httpjournalofvisionorg8115 doi1011678115 [PubMed] [Article]
Rolfs M Laubrock J amp Kliegl R (2006) Shorteningand prolongation of saccade latencies followingmicrosaccades Experimental Brain Research 169369ndash376 [PubMed]
Rucci M Iovin R Poletti M amp Santini F (2007)Miniature eye movements enhance fine spatial detailNature 447 851ndash854 [PubMed]
Salas C Navarro F Torres B amp Delgado-Garcia J M(1992) Effects of diazepam and D-amphetamine onrhythmic pattern of eye movements in goldfishNeuroreport 3 131ndash134 [PubMed]
Schipperheyn J J (1963) Respiratory eye movementsand perception of stationary objects in the frog ActaPhysiologica et Pharmacologica Neerlandica 12157ndash159 [PubMed]
Schipperheyn J J (1965) Contrast detection in frogrsquosretina Acta Physiologica et Pharmacologica Neer-landica 13 231ndash277 [PubMed]
Schipperheyn J J (1967) Influence of eye movements onretinal ganglion cell activity in frogs Acta Physi-ologica et Pharmacologica Neerlandica 14 506[PubMed]
Segev R Schneidman E Goodhouse J amp Berry M J2nd (2007) Role of eye movements in the retinalcode for a size discrimination task Journal ofNeurophysiology 98 1380ndash1391 [PubMed] [Article]
Sharpe C R (1972) The visibility and fading of thinlines visualized by their controlled movement acrossthe retina The Journal of Physiology 222 113ndash134[PubMed] [Article]
Shelley M McLaughlin D Shapley R amp Wielaard J(2002) States of high conductance in a large-scalemodel of the visual cortex Journal of ComputationalNeuroscience 13 93ndash109 [PubMed]
Skavenski A A Robinson D A Steinman R M ampTimberlake G T (1975) Miniature eye movementsof fixation in rhesus monkey Vision Research 151269ndash1273 [PubMed]
Snodderly D M (1987) Effects of light and darkenvironments on macaque and human fixational eyemovements Vision Research 27 401ndash415 [PubMed]
Snodderly D M Kagan I amp Gur M (2001) Selectiveactivation of visual cortex neurons by fixational eyemovements Implications for neural coding VisualNeuroscience 18 259ndash277 [PubMed]
Snodderly D M amp Kurtz D (1985) Eye position duringfixation tasks Comparison of macaque and humanVision Research 25 83ndash98 [PubMed]
Steinbach M J amp Money K E (1973) Eye movementsof the owl Vision Research 13 889ndash891 [PubMed]
Steinman R M Haddad G M Skavenski A A ampWyman D (1973) Miniature eye movement Science181 810ndash819 [PubMed]
Torres B Pastor A M Cabrera B Salas C ampDelgado-Garcia J M (1992) Afferents to theoculomotor nucleus in the goldfish (Carassius aur-atus) as revealed by retrograde labeling with horse-radish peroxidase Journal of Comparative Neurology324 449ndash461 [PubMed]
Troncoso X G Macknik S L amp Martinez-Conde S(2008) Microsaccades counteract perceptual filling-inJournal of Vision 8(14)15 1ndash9 httpjournalofvisionorg81415 doi10116781415 [Article]
Troncoso X G Macknik S L Otero-Millan J ampMartinez-Conde S (2008) Microsaccades driveillusory motion in the Enigma illusion Proceedingsof the National Academy of Sciences of the UnitedStates of America 105 16033ndash16038 [PubMed]
Tse P U Sheinberg D S amp Logothetis N K (2004)The distribution of microsaccade directions need notreveal the location of attention Psychological Science15 708ndash710
Turatto M Valsecchi M Tame L amp Betta E (2007)Microsaccades distinguish between global and localvisual processing Neuroreport 18 1015ndash1018[PubMed]
Uchida N Kepecs A amp Mainen Z F (2006) Seeing ata glance smelling in a whiff Rapid forms ofperceptual decision making Nature Reviews Neuro-science 7 485ndash491 [PubMed]
Valsecchi M Betta E amp Turatto M (2007) Visualoddballs induce prolonged microsaccadic inhibitionExperimental Brain Research 177 196ndash208[PubMed]
van Dam L C amp van Ee R (2006) Retinal image shiftsbut not eye movements per se cause alternations inawareness during binocular rivalry Journal of Vision6(11)3 1172ndash1179 httpjournalofvisionorg6113doi1011676113 [PubMed] [Article]
Van der Steen J amp Collewijn H (1984) Ocular stabilityin the horizontal frontal and sagittal planes in therabbit Experimental Brain Research 56 263ndash274[PubMed]
Van Gisbergen J A Robinson D A amp Gielen S(1981) A quantitative analysis of generation ofsaccadic eye movements by burst neurons Journalof Neurophysiology 45 417ndash442 [PubMed]
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 15
Van Gisbergen J A M amp Robinson D A (1977)Generation of micro and macrosaccades by burstneurons in the monkey In R Baker amp A Berthoz(Eds) Control of gaze by brain stem neurons (vol 1pp 301ndash308) New York ElsevierNorth-Holland
Wallman J Pettigrew J D amp Letelier J C (1994)The oscillatory saccades of birds Motoneural activ-ity and possible functions In B T Fuchs A FV Buttner amp D Zee (Eds) Contemporary ocularmotor and vestibular research A tribute to David ARobinson (pp 480ndash487) Stuttgart Germany ThiemePublishing
Williams P E amp Shapley R M (2007) A dynamicnonlinearity and spatial phase specificity in macaque
V1 neurons Journal of Neuroscience 27 5706ndash5718[PubMed] [Article]
Winterson B J amp Robinson D A (1975) Fixationby the alert but solitary cat Vision Research 151349ndash1352 [PubMed]
Yarbus A L (1967) Eye movements and vision(B Haigh Trans) New York Plenum Press
Zuber B L amp Stark L (1965) Microsaccades and thevelocityltamplitude relationship for saccadic eyemovements Science 150 1459ndash1460 [PubMed]
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 16

mulatta (Goffart Quinet Chavane amp Masson 2006Horwitz amp Albright 2003 Leopold amp Logothetis1998 Martinez-Conde 2006 Martinez-Conde et al 20002002 Motter amp Poggio 1984 Skavenski et al 1975Snodderly Kagan amp Gur 2001 Steinman HaddadSkavenski amp Wyman 1973) Macaca fascicularis(Snodderly 1987 Snodderly et al 2001 Snodderly ampKurtz 1985) and Macaca nemestrina (Bair amp OrsquoKeefe1998) with no substantial differences in the spatiotemporalcharacteristics of fixational eye movements (rate velocityand magnitude profiles etc) between them Microsaccadevelocities are parametrically related to microsaccadeamplitudes following the ldquomain sequencerdquo in both ma-caques (Martinez-Conde et al 2000) and humans (Engbert2006 Martinez-Conde 2006 Martinez-Conde et al 2006Zuber amp Stark 1965) Fixational microsaccades have alsobeen observed in the baboon (Papio papio) with ampli-tudes up to 25 degrees (Marchetti Gauthier amp Pellet1983)It is important to note that microsaccades cannot be
defined according to their amplitude alone because smallexploratory or voluntary saccades can be the same size asmicrosaccades Microsaccades can be defined only opera-tionally as the involuntary saccades that are producedwhile the subject attempts to fixate (Martinez-Conde2006) This ldquoattempt to fixaterdquo includes both holding theeyes still and static while foveating the visual target andalso orienting the fovea toward a target It is worth notingthat microsaccade production may require the presence ofa visual andor attentional target (Otero-Millan et al2008)Thus there is no known physical parameter (or
combination of parameters) that separates saccades frommicrosaccades Mounting evidence points toward a com-mon neural generator of saccades and microsaccades(Engbert 2006 Martinez-Conde et al 2004 Otero-Millan et al 2008 Rolfs Kliegl amp Engbert 2008 RolfsLaubrock amp Kliegl 2006 Zuber amp Stark 1965)However one should note that most of the available datain support of a shared microsaccadendashsaccade generatorare indirect To date only two physiological studies havedirectly addressed the question of the oculomotor
mechanisms leading to the generation of microsaccadesIn those studies Van Gisbergen and colleagues foundthat putative motoneurons in the primate abducensnucleus and burst neurons in the nearby pontomedullaryreticular formation were similarly active during saccadesand microsaccades (Van Gisbergen Robinson amp Gielen1981 Van Gisbergen amp Robinson 1977) Future researchshould further explore the neural bases underlying thegeneration of microsaccades and other fixational eyemovementsSimultaneous recordings of microsaccades and neural
responses have been conducted at multiple levels of theprimate visual pathway including the lateral geniculatenucleus (LGN) area V1 and the extrastriate visual cortex(Bair amp OrsquoKeefe 1998 Leopold amp Logothetis 1998Martinez-Conde 2006 Martinez-Conde et al 20002002 Reppas Usrey amp Reid 2002 Snodderly et al2001) Microsaccades are predominantly excitatory at allthese levels leading to firing rate increases in both earlyand higher visual areas The firing rate increases followingmicrosaccades in the LGN and V1 are of visual origin(Figure 2) and result from the microsaccade-induceddisplacement of visual receptive fields over stationarystimuli Microsaccade-driven increases in firing rate tendto be clustered in tight bursts of spikes (Martinez-Conde2006 Martinez-Conde et al 2000 2002) A recent studyhas suggested that primate microsaccades may improvethe efficient sampling of fine spatial detail (Donner ampHemila 2007)Microsaccades counteract visual fading and increase
visibility during fixation in human subjects (Martinez-Conde et al 2006 Troncoso Macknik amp Martinez-Conde 2008) Numerous studies have also reported thathuman microsaccade rates andor directions are modulatedby cognitive processes such as the allocation of spatialattention (Betta amp Turatto 2006 Engbert amp Kliegl2003a 2003b 2004 Galfano et al 2004 Hafed amp Clark2002 Laubrock Engbert Rolfs amp Kliegl 2007 Otero-Millan et al 2008 Turatto Valsecchi Tame Betta 2007Valsecchi Betta amp Turatto 2007 but see also HorowitzFine Fencsik Yurgenson amp Wolfe 2007 and TseSheinberg amp Logothetis 2004)
Figure 1 Cartoon representation of fixational eye movements in humans and primates Microsaccades (straight and fast movements)drifts (curvy slow movements) and tremor (oscillations superimposed on drifts) transport the visual image across the retinal photoreceptormosaic From Martinez-Conde and Macknik (2007b)
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 5
Neural responses to drifts have received considerablyless attention than neural responses to microsaccades Thismay be due to the fact that drifts are more difficult tocharacterize objectively than microsaccades (which aremore easily detected by automatic algorithms that com-bine amplitude and velocity thresholds) Thus drifts areusually identified indirectly as the eye position changesthat occur between microsaccades (Snodderly et al2001) However this method has the potential flaw thatone may unintentionally attribute non-drift-related activity(for instance undetected tremor) to drifts Drifts mayincrease firing in a subset of primate V1 neurons(Snodderly et al 2001) However they generate lessvariability in neuronal responses than a combination ofdrifts and microsaccades (Gur Beylin amp Snodderly1997) No studies to date have recorded primate neuronalresponses in correlation to tremor (but see Hennig ampWorgotter 2007 for the responses of a model macaqueretina to simulated drift and tremor)
Fixational eye movements in the cat
The catrsquos fixational eye movements have been inves-tigated in a handful of studies and their conclusions
remain somewhat controversial In 1960 Hebbard andMarg (1960) found fixational eye movements in the cat tobe quite comparable to those in humans although catmicrosaccades were scarcer and smaller than in humansPritchard and Heron (1960) also found that the catrsquosfixational eye movements included microsaccades driftsand tremor Microsaccades were much rarer in the cat thanin humans however possibly related to the fact that thecat lacks a well-developed fovea Delgado-Garcia delPozo and Baker (1986) also observed some putativemicrosaccades in the alert behaving cat Other studiesfound no microsaccades in the cat (Conway Timberlakeamp Skavenski 1981 Winterson amp Robinson 1975)However in these last studies microsaccades were definedas ldquosaccades smaller than 10 minutes of arcrdquo which iswell below the average microsaccade amplitude found inrecent human and primate studies (see Martinez-Condeet al 2004 for a review of microsaccade parameters inhumans and primates) More recently several studies haveaddressed the contribution of fixational eye movements toneural responses in the catrsquos visual system HennigKerscher Funke and Worgotter (2002) found increasedresponses from neurons in cat areas 17 and 18 duringstimulus motions that mimicked fixational eye move-ments Using a model retina they also found that
1 simulated fixational eye movements increased neu-ral responses and
2 simulated tremor improved spatial acuity
Hennig and Worgotter (2004) later reported that acombination of tremor and microsaccades improved theperformance of simulated ganglion cells in a hyperacuitytask in the retinal periphery (but not in the central retina)Miller Denning George Marshak and Kenyon (2006)also investigated the effects of simulated tremor on amodel of the catrsquos retina and suggested that tremor mayselectively enhance the processing of large stimuli
Fixational eye movements in the rabbit
In contrast to foveate species such as primates (includ-ing humans) and cats (which have an area centralis if nota proper fovea) the rabbit (Oryctolagus cuniculus) dis-plays few spontaneous eye movements especially in theabsence of head motion (Collewijn 1977 Collewijn ampvan der Mark 1972 Fuller 1980 1981 Van der Steen ampCollewijn 1984) In normal visual conditions the rabbitrsquoseyes are very stable but not perfectly immobile Smallamplitude tremor can be observed as well as very slowdrift Microsaccades have not been observed Howeverdrift is occasionally corrected by fast saccadic movements(Collewijn 1977 Collewijn amp van der Mark 1972 Vander Steen amp Collewijn 1984) These differences may berelated to the fact that the rabbit does not have a fovea butan elongated horizontal streak with elevated ganglion cell
Figure 2 Microsaccades drive neural activity in primate area V1Microsaccades lead to firing rate increases in V1 neurons (n = 308)of the awake primate In the absence of visual stimulationmicrosaccades do not increase neural activity in area V1 (n = 37neurons) The resultsVshown here for the average of all neuronsVwere consistent with responses from individual neurons (data notshown) Modified from Martinez-Conde et al (2000 2002)
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 6
counts Olveczky Baccus and Meister (2003) recordedfrom ganglion cells in the rabbit and salamander retinaswhile presenting visual stimuli that were jittered tosimulate fixational eye movements (based on parametersfrom previously published studies by Manteuffel PlasaSommer amp Wess 1977 and Van der Steen amp Collewijn1984) In both species fixational eye movements helpedsegregate a moving foreground from a complex back-ground in a population of ganglion cells
Fixational eye movements inbirds
Fixational eye movements in the pigeon
There are some remarkable differences between fixa-tional eye movements in birds and mammals Nye (1969)identified four types of fixational eye movements in thepigeon (Columba livia) flicks (ie microsaccades)impulse movements drift and oscillations No tremorwas detected Microsaccade frequency was very lowrelative to humans Impulse movements occurred morefrequently usually between bursts of oscillations withamplitudes ranging up to 2 degrees Drift velocitiesranged from 1 to 5 degrees per second Both micro-saccades and drifts occurred in response to movingstimuli Oscillations occurred in short bursts at rates ofabout 30 Hz and could be quite large (up to severaldegrees of amplitude) Bloch Rivaud and Martinoya(1984) reported microsaccades drifts and oscillationsldquosimilar to those described by Nyerdquo but none of thesewere analyzed in detailCyclotorsional oscillations are an integral part of avian
saccades (Wallman Pettigrew amp Letelier 1994) Thusone might speculate that oscillations are associated withavian microsaccades as well especially if saccades andmicrosaccades share a common generator (Engbert 2006Otero-Millan et al 2008 Rolfs et al 2008 2006 Zuberamp Stark 1965) However this possibility has not yet beenaddressed in the literature The function of oscillations isunknown (Carpenter 1988) but one hypothesis is thatthey serve to facilitate the delivery of arterial blood to theocular capillary bed (Pettigrew Wallman amp Wildsoet1990 Wallman et al 1994)
Fixational eye movements in the owl
Steinbach and Money (1973) recorded fixational eyemovements in the great horned owl (Bubo virginianus)a species previously thought incapable of moving itseyes The owlrsquos fixational eye movements includeflicks or microsaccades drift tremor and oscillations
Microsaccades seemed correlated to visual events ofinterest to the owl they were 1 deg or less in amplitudeand had peak velocities of about 10 degsec Drifts wereslow motions of the eye in between fixations withamplitudes of 1 degree or less Tremor amplitude wasabout 1 arcmin with frequencies around 20 Hz High-frequency oscillations (25 Hz) of short duration (025 sec)were very similar to those previously found in the pigeon(Nye 1969) No studies to date have examined theperceptual or physiological effects of fixational eyemovements in any bird species
Fixational eye movements inreptiles
Fixational eye movements in the turtle
Greschner Bongard Rujan and Ammermuller (2002)identified two types of fixational eye movements in theturtle (Pseudemys scripta elegans) which they calledldquodrift-likerdquo and ldquoperiodicrdquo Drift-like motions were smalland slow (with velocities comparable to human drift) andwere superimposed by larger and faster periodic componentsPeriodic motion had retinal amplitudes on the order of thediameter of a photoreceptor which are comparable to thoseof human tremor On the other hand periodic motionrsquosfrequency (5 Hz) velocity and orderly relationship betweenvelocity and amplitude are closer to those of human micro-saccades (see Martinez-Conde et al 2004 for a review oftremor and microsaccade parameters in humans)Simulated periodic motion generated strong synchronous
firing in the turtlersquos retina whereas simulated drift-likemotion had little effect (Figure 3) These results agree withprevious physiological and modeling studies in the primatevisual system in which strong neural transients wereobserved in response to microsaccades (Donner amp Hemila2007 Martinez-Conde 2006 Martinez-Conde et al 20002002) Such neural transients may underlie the behavior ofcortical neurons as coincidence detectors (ShelleyMcLaughlin Shapley amp Wielaard 2002 Williams ampShapley 2007) Moreover neural transients to stimulionsets and terminations (similar to those produced bymicrosaccades in the primate visual system Martinez-Conde 2006 Martinez-Conde et al 2000 2002) havebeen related to target visibility in visual masking para-digms (Macknik amp Livingstone 1998 Macknik ampMartinez-Conde 2004 Macknik Martinez-Conde ampHaglund 2000)In the turtle retina periodic motion-driven neurons with
receptive fields located along contrast borders weremoreover synchronized and reliably indicated the preced-ing motions The authors proposed that this synchroniza-tion of retinal activity could be used by the brain to
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 7
improve the estimation of stimulus features such asspatial frequency (Greschner et al 2002) This findingcould have a parallel in the primate visual system Donnerand Hemila (2007) recently proposed that microsaccadessignificantly ldquore-sharpenrdquo the image and improve spatialresolution in primates Rucci Iovin Poletti and Santini(2007) also found that fixational eye movements enhancefine spatial detail in human subjects
Fixational eye movements in the chameleon
Fixational eye movements in the chameleon have notbeen studied in detail However Gioanni and colleaguesfound that the chameleon (Chamaleo chamaleo) makessaccades during fixation The authors speculated that theseprobably correspond to fixational microsaccades found inother foveate species (Gioanni Bennis amp Sansonetti1993)
Fixational eye movements inamphibians
Fixational (and more generally spontaneous) eye move-ments vary widely among amphibians The frog (anafoveate amphibian) shows a general lack of eye andhead movements except for those that stabilize the retinalimage Dieringer and colleagues (Dieringer amp Daunicht1986 Dieringer amp Precht 1982 Dieringer Precht ampBlight 1982) found that the grass frog (Rana temporaria)produces no spontaneous eye movements in the absenceof head movements with the possible exception of tremor(similarly to the rabbit another afoveate species Van derSteen amp Collewijn 1984) It is likely that the frogrsquos lack ofeye movements leads to fast visual fading of all stationaryobjects in its field of view (Ditchburn amp Ginsborg 1952Yarbus 1967) Lettvin Maturana MsCulloch and Pitts
Figure 3 Fixational eye movements increase retinal activity in the turtle (A) Continuous recording from 54 retinal ganglion cells in theturtle during darkness after switching on a stationary grating and during wobbling of this grating The grating was wobbled to simulateperiodic (ie tremor-like) eye movements (B) Responses of a single ganglion cell to a drifting contrast border of velocity comparable toocular drifts (C) When the drift is superimposed with wobbling the responses of the cell shown in (B) increase dramatically FromGreschner et al (2002) and modified by Martinez-Conde et al (2004)
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 8
(1968) stated that a frog ldquowill starve to death surroundedby food if it is not movingrdquo (but see Dieringer amp Daunicht1986 for a counterpoint) Small head motions or eyemovements correlated with respiratory motions maydiminish fading to some extent (Dieringer amp Precht1982 Schipperheyn 1963 1965 1967)As a side note the frogrsquos respiratory eye movements
play no role in the respiratory function ldquoin the course ofeach respiratory cycle increase of pressure in the frogrsquosbuccal cavity protrudes the eye and causes the opticalaxis to deviate upwards over 25ndash35 minutes of arcrdquo(Schipperheyn 1963) That is respiratory eye movementsare merely synchronized with respirationAutrum (1959) found no evidence of respiratory eye
movements in the frog Ewert andBorchers (1974) observedthat stationary stimuli elicit no responses from visualneurons in the common toad (Bufo Bufo L) regardless ofrespiratory pressure changes in the buccal cavity
Fixational eye movements in the salamander
Manteuffel et al (1977) identified two types of fixa-tional eye movements in salamanders tremor-like andrespiratory eye movements Tremor-like is a rapid move-ment of small amplitude Torsional eye movements resultfrom the animalrsquos breathing (also found in the frog bySchipperheyn 1963) Additionally flicks are occasionallyobserved which rapidly change the position of gaze It isunclear whether such flicks may generally correspond towhat we now call microsaccades andor to voluntarysaccades Based on these observations the authors pro-posed that unlike frogs and toads (Autrum 1959 Ewert ampBorchers 1974 Lettvin et al 1968) salamanders cancontinuously perceive stationary objects (Manteuffel et al1977) Fixational eye movements in the salamander (Tigersalamander Ambystoma tigrinum) may also help segre-gate object and background motion in the retina(Olveczky et al 2003 see also Fixational eye movementsin the rabbit section) and even work to emphasize novelvisual features (Olveczky et al 2007 see Baccus et al2008 for details on the retinal neurons and synapses thatmay underlie the computation of differential motionbetween an object and its background)
Fixational eye movements in fish
Spontaneous eye movements in the goldfish
Eye movements during fixation have been studied inboth afoveate (goldfish Carassius auratus) and foveate(archer fish Toxotes chatareus) fish species The goldfisha long-standing model organism in vision and oculomotor
research (Aksay et al 2003 2007 Cabrera Torres PasaroPastor amp Delgado-Garcia 1992 Pastor Torres Delgado-Garcia amp Baker 1991 Salas Navarro Torres ampDelgado-Garcia 1992 Torres Pastor Cabrera Salas ampDelgado-Garcia 1992) produces three types of sponta-neous eye movements saccades stretch and drift (Easter1971 Mensh Aksay Lee Seung amp Tank 2004) Weshould note that in the motionless goldfish eye move-ments are very infrequent Those that do occur consistmainly of saccades slow drifts are very rare (Easter Johnsamp Heckenlively 1974) During saccades (identified bythresholding the eye velocity at 5 degsec) both eyes moverapidly simultaneously and usually in the same direction(Mensh et al 2004) Stretches occur every 2ndash3 min and arecharacterized by a brief simultaneous large excursion ofboth eyes to extreme temporal positions (Easter 1971Mensh et al 2004) The function of stretches is not known(Mensh et al 2004) Because fixations were defined asthose periods that were free of saccadic and stretchmovements (Easter 1971) only drifts (from the threetypes of spontaneous movements) can technically beconsidered ldquofixational eye movementsrdquo However thegoldfish saccadic main sequence includes saccades withequivalent amplitudes and velocities to those found inhuman microsaccades (Mensh et al 2004 Zuber amp Stark1965) Indeed microsaccades have been identified in other(foveate) fish species although they are rarer than inhumans (Segev et al 2007) Small saccades in thegoldfish tend to be divergent whereas large saccadestend to be convergent (Mensh et al 2004) Drift occurssimultaneously in both eyes usually in the samedirection Drift velocities are almost exclusively e1 degsec(Mensh et al 2004) No tremor has been identified in thegoldfish
Spontaneous eye movements in teleost fish
Spontaneous eye movements have been studied in theNew Zealand parore (Girella tricuspidata MontgomeryMcVean amp McCarthy 1983) The pattern of eye move-ments was similar to that previously described in goldfish(Easter 1971 Hermann amp Constantine 1971)
Fixational eye movements in the archer fish
A recent study in the archer fish (Segev et al 2007) hasprovided the first perceptual and physiological correlatesof fixational eye movements in any fish species Archerfish have remarkable visual abilities they shoot downinsects resting on foliageVas well as stationary targetscovering a single retinal photoreceptorVwith squirts ofwater from their mouths they predict the location atwhich the insect will splash down in G100 msec and theydeduce the absolute size of an object presented at differentviewing distances (Supplementary Videos 2 and 3) Segev
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 9
et al (2007) identified three types of fixational eyemovements in the archer fish microsaccades drift andtremor Microsaccades were rare and were not analyzed indetail Drift velocities were e040 degsec Tremor was ahigh-frequency oscillatory movement (the primary com-ponent was 5 Hz) with peak-to-peak amplitude of 01ndash02 deg (corresponding to an image displacement of 1ndash2photoreceptors) and velocity of about 1 degsec (Figure 4)Whereas the amplitude and velocity of tremor in thearcher fish is comparable to those in humans its frequencyis much lower (5 Hz vs 940 Hz) Interestingly archer fishtremor is very similar both in amplitude (photoreceptorwidths covered) as well as in frequency (5 Hz) to thetremor-like periodic motion in the turtle (discussedearlier Greschner et al 2002)Segev et al (2007) trained archer fish to perform a size
discrimination task the fish had to distinguish a medium-sized target from larger and smaller distracters (theyindicated their choice by shooting a jet of water) To relatethis behavior to responses from retinal neurons theauthors recorded from populations of retinal ganglioncells while presenting objects of different sizes moved tosimulate saccades drifts and tremor Drifts and tremorelicited weak neuronal responses that were not veryinformative about the spatial structure of the stimulus(although they did provide some information about target
size in agreement with the results of Greschner et al2002 in the turtle) Moreover performance was similar fortremor alone and for tremor plus drift indicating that driftplays a negligible role in revealing spatial structure (alsoin agreement with Greschner et al 2002)When present the firing rate was phase-locked to
tremor Saccades were more effective than drifts or tremorin driving informative responses from the ganglion cells(consistent with theoretical and physiological studies ofmicrosaccade-driven neuronal activity in the primateretina and LGN Donner amp Hemila 2007 Martinez-Conde et al 2002) Segev et al (2007) proposed that inprimates both saccades and microsaccades (which arerare in the archer fish) may contribute to size discrim-ination tasks as well as enhance spatial detail whereasdrifts and tremor by themselves may not be sufficient toimprove visual perception However it is important toremember that central vision in humans can be maintainedby tremor andor drift even when microsaccades aresuppressed during strict fixation (Martinez-Conde 2006Martinez-Conde et al 2004 2006) On the other handone cannot rule out the possibility that if one couldcompletely eliminate drifts and tremor microsaccadesmight then suffice to sustain both central and peripheralvision in primates and humans Future experiments areneeded to further explore these ideas
Figure 4 Archer fish eye movements (A) Example trace of eye position as a function of time as measured by search coil (B) Example ofa single 15-s fixation superimposed on a grid representing the photoreceptor mosaic The back and forth movement is caused by tremor(C) Distribution of fixation durations (taken as time interval between saccades) (D) Power spectrum of horizontal fixational eyemovements (E) Distribution of tremor amplitude (data taken from 1 animal in a 15-min session) Modified from Segev et al (2007)
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 10
Discussion
The vast majority of eye movement studies conductedin non-human species to date have not analyzed eyeposition data in the range of fixational eye movementsThus eye movements during fixation have been reportedin only a handful of vertebrates Most of these studieshave concentrated solely on the physical parameters ofthe fixational eye movements without addressing theirphysiological and perceptual consequences Fortunatelythis trend has started to reverse in recent years with agrowing list of papers addressing the effects of non-human fixational eye movements in physiology andperception This is an important development A largeamount of all psychophysical and physiological visualresearch has been conducted during conditions of visualfixation Therefore understanding the precise physiolog-ical and perceptual contributions of fixational eye move-ments may be critical to the interpretation of previous andfuture vision researchThere are some interesting differences and similarities
in the types of fixational eye movements produced bydifferent species For instance ocular tremor is possiblyubiquitous to all vertebrates The fact that tremor has notbeen observed in all species studied could be due to thetechnical difficulty of recording tremor accurately (ietremor amplitudes and frequencies are usually in the rangeof the recording systemrsquos noise) Drift has also beenrecorded in most vertebrates and it may be as omnipre-sent as tremor Conversely microsaccades seem mostimportant in species with foveal vision (ie primates andhumans) whereas they are scarce or absent in afoveatespecies (ie rabbits frogs goldfish) A link betweenmicrosaccades and foveal vision may reflect the two-pronged functional significance of microsaccades inhumans and primates first microsaccades may benecessary to correct the slow drift of the eye and bringback the objectregion of interest to the retinal area wherephotoreceptor density is highest (Cornsweet 1956) Thecorrective role of human microsaccades has been demon-strated in recent studies (Engbert amp Kliegl 2004 Engbertamp Mergenthaler 2006) Second the dynamics of micro-saccades themselves may improve spatial resolution(Donner amp Hemila 2005)In reviewing the patterns of fixational eye movements
across vertebrates it is tempting to draw parallels withother sensory modalities Fixational eye movementsrefresh the retinal images during fixation and thus preventneural adaptation and visual fading But this mechanism isnot circumscribed to the visual system Sniffs in rodentolfaction discretely sample sensory information every200ndash300 msec and thus are similar in their temporaldynamics to primate saccades (Otero-Millan et al 2008Uchida Kepecs amp Mainen 2006) and microsaccades(Otero-Millan et al 2008) A similar mode of discretesampling may also be at play when objects are recognized
through tactile information for instance if we sweep ourfingertips over an objectrsquos surface with our eyes closed orwhen blind individuals read Braille scriptOne intriguing possibility that merits further experi-
ments and analyses is the potential correlation between thespatiotemporal parameters of fixational eye movements(such as those summarized in Table 1) and the adaptationproperties of visual neurons across species For instancethe greater ocular stability of some species (rabbitgoldfish frog etc) versus others (primates) could beassociated with some particular adaptive properties oftheir visual cells Thus future comparative studies shouldpay attention to the response properties of visual cellstriggered by fixational eye movements in the presence ofstationary stimuli Other potential factors underlying thevariation in fixational eye movements across species couldbe differences in oculomotor control at the level of the eyemuscles or perhaps differences in retinal organizationFuture research will further improve our understanding
of fixational eye movements and the neural activity thatmaintains the visibility of stationary visual scenes inhuman and non-human species One interesting questionthat remains to be addressed concerns the role of tremor invision and visual physiology Because tremor frequenciesare well over the flicker fusion frequencies in humans ithas been argued that tremor of the visual image would beineffective as a stimulus (Ditchburn 1955 Gerrits ampVendrik 1970 Sharpe 1972) However recent studieshave found simulated tremor to enhance visual processingand improve visual acuity in a model retina (Hennig et al2002 Miller et al 2006) Follow-up research should beaimed to replicate these findings in actual neurons withcorrelated perceptual reports One should also note thatearly visual neurons can follow high-frequency flickeringthat surpasses the perceptual threshold for flicker fusion(Martinez-Conde et al 2002) Thus high-frequencytremor could be adequate to maintain activity in the earlyvisual system which then may lead to visual perception inhigher visual areas Another significant question thatremains unanswered concerns the oculomotor bases ofthe various types of fixational eye movements acrossspecies To date there is too little information on thisimportant topic to draw a conclusion
Conclusions
We have reviewed the parameters of fixational eyemovements in non-human mammals birds reptilesamphibians and fish as well as their consequences forphysiology and perception Fixational eye movementsand microsaccades in particular appear to be mostimportant in foveate vs afoveate species Animals lackingfixational eye movements such as afoveate frogs andtoads may be incapable of seeing stationary objects
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 11
during visual fixation Future research should furtherexplore the perceptual effects of fixational eye movementsand determine the oculomotor mechanisms responsible fortheir generation across species
Acknowledgments
We thank Mona Stewart and Isabel Gomez-Caraballo fortechnical assistance Jorge Otero-Millan for programmingassistance and Dr Xoana G Troncoso for comments on themanuscript This study was funded through grants fromthe Barrow Neurological Foundation (to SMC and SLM)the National Science Foundation (Award 0643306 toSMC) the Arizona Biomedical Research Commission(Award 07-102 to SMC) and Science Foundation Arizona(SLM)
Commercial relationships noneCorresponding author Susana Martinez-Conde PhDEmail smartneuralcorrelatecomAddress Barrow Neurological Institute 350 W ThomasRd Phoenix AZ 85013 USA
References
Aksay E Major G Goldman M S Baker R SeungH S amp Tank D W (2003) History dependence ofrate covariation between neurons during persistentactivity in an oculomotor integrator Cerebral Cortex13 1173ndash1184 [PubMed] [Article]
Aksay E Olasagasti I Mensh B D Baker RGoldman M S amp Tank D W (2007) Functionaldissection of circuitry in a neural integrator NatureNeuroscience 10 494ndash504 [PubMed]
Autrum H (1959) Das Fehlen unwillkurlicher Augenbe-wegungen beim Frosch Naturwissenschaften 46435
Baccus S A Olveczky B P Manu M amp Meister M(2008) A retinal circuit that computes object motionJournal of Neuroscience 28 6807ndash6817 [PubMed]
Bair W amp OrsquoKeefe L P (1998) The influence offixational eye movements on the response of neuronsin area MT of the macaque Visual Neuroscience 15779ndash786 [PubMed]
Betta E amp Turatto M (2006) Are you ready I can tellby looking at your microsaccades Neuroreport 171001ndash1004 [PubMed]
Bloch S Rivaud S amp Martinoya C (1984) Compar-ing frontal and lateral viewing in the pigeon IIIDifferent patterns of eye movements for binocularand monocular fixation Behavioural Brain Research13 173ndash182 [PubMed]
Cabrera B Torres B Pasaro R Pastor A M ampDelgado-Garcia J M (1992) A morphological studyof abducens nucleus motoneurons and internuclearneurons in the goldfish (Carassius auratus) BrainResearch Bulletin 28 137ndash144 [PubMed]
Carpenter R H S (1988) Movements of the eyes(2nd ed) Pion London
Collewijn H (1977) Eye- and head movements infreely moving rabbits The Journal of Physiology266 471ndash498 [PubMed] [Article]
Collewijn H amp van der Mark F (1972) Ocular stabilityin variable visual feedback conditions in the rabbitBrain Research 36 47ndash57 [PubMed]
Conway J L Timberlake G T amp Skavenski A A(1981) Oculomotor changes in cats reared withoutexperiencing continuous retinal image motion Exper-imental Brain Research 43 229ndash232 [PubMed]
Cornsweet T N (1956) Determination of the stimuli forinvoluntary drifts and saccadic eye movements Jour-nal of the Optical Society of America 46 987ndash993[PubMed]
Delgado-Garcia J M del Pozo F amp Baker R (1986)Behavior of neurons in the abducens nucleus of thealert catVII Internuclear neurons Neuroscience 17953ndash973 [PubMed]
Dieringer N amp Daunicht W J (1986) Image fadingVAproblem for frogs Naturwissenschaften 73 330ndash332[PubMed]
Dieringer N amp Precht W (1982) Compensatory headand eye movements in the frog and their contributionto stabilization of gaze Experimental BrainResearch 47 394ndash406 [PubMed]
Dieringer N Precht W amp Blight A R (1982)Resetting fast phases of head and eye and theirlinkage in the frog Experimental Brain Research 47407ndash416 [PubMed]
Ditchburn R W (1955) Eye-movements in relation toretinal action Optica Acta 1 171ndash176
Ditchburn R W amp Ginsborg B L (1952) Vision with astabilized retinal image Nature 170 36ndash37[PubMed]
Ditchburn R W amp Ginsborg B L (1953) Involuntaryeye movements during fixation The Journal ofPhysiology 119 1ndash17 [PubMed] [Article]
Donner K amp Hemila S (2007) Modelling the effect ofmicrosaccades on retinal responses to stationarycontrast patterns Vision Research 47 1166ndash1177[PubMed]
Donner K K amp Hemila S (2005) The possible purposeof microsaccades Paper presented at the EuropeanConference on Visual Perception A Coruna Spain
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 12
Easter S S Jr (1971) Spontaneous eye movements inrestrained goldfish Vision Research 11 333ndash342[PubMed]
Easter S S Jr Johns P R amp Heckenlively D R(1974) Horizontal compensatory eye movements ingoldfish (Carassius auratus) I The normal animalJournal of Comparative Physiology 92 23ndash35
Engbert R (2006) Microsaccades A microcosm forresearch on oculomotor control attention and visualperception Progress Brain Research 154 177ndash192[PubMed]
Engbert R amp Kliegl R (2003a) Binocular coordinationin microsaccades In R R J Hyona amp H Deubel(Eds) The mindrsquos eyes Cognitive and appliedaspects of eye movements (pp 103ndash117) OxfordUK Elsevier
Engbert R amp Kliegl R (2003b) Microsaccades uncoverthe orientation of covert attention Vision Research43 1035ndash1045 [PubMed]
Engbert R amp Kliegl R (2004) Microsaccades keep theeyesrsquo balance during fixation Psychological Science15 431ndash436 [PubMed]
Engbert R amp Mergenthaler K (2006) Microsaccadesare triggered by low retinal image slip Proceedingsof the National Academy of Sciences of the UnitedStates of America 103 7192ndash7197 [PubMed][Article]
Ewert J P amp Borchers H W (1974) Antwort vonretinalen Ganglienzellen bei freibeweglichen Kroten(Bufo Bufo L) Journal of Comparative and Physi-ology 92 117ndash130
Fuller J H (1980) Linkage of eye and head movementsin the alert rabbit Brain Research 194 219ndash222[PubMed]
Fuller J H (1981) Eye and head movements duringvestibular stimulation in the alert rabbit BrainResearch 205 363ndash381 [PubMed]
Galfano G Betta E amp Turatto M (2004) Inhibition ofreturn in microsaccades Experimental BrainResearch 159 400ndash404 [PubMed]
Gerrits H J amp Vendrik A J (1970) Artificial move-ments of a stabilized image Vision Research 101443ndash1456 [PubMed]
Gioanni H Bennis M amp Sansonetti A (1993) Visualand vestibular reflexes that stabilize gaze in thechameleon Visual Neuroscience 10 947ndash956[PubMed]
Goffart L Quinet J Chavane F amp Masson G S(2006) Influence of background illumination onfixation and visually guided saccades in the rhesusmonkey Vision Research 46 149ndash162 [PubMed]
Greschner M Bongard M Rujan P amp Ammermuller J(2002) Retinal ganglion cell synchronization by fixa-tional eye movements improves feature estimationNature Neuroscience 5 341ndash347 [PubMed]
Gur M Beylin A amp Snodderly D M (1997)Response variability of neurons in primary visualcortex (V1) of alert monkeys Journal of Neuro-science 17 2914ndash2920 [PubMed] [Article]
Hafed Z M amp Clark J J (2002) Microsaccades as anovert measure of covert attention shifts VisionResearch 42 2533ndash2545 [PubMed]
Hebbard F W amp Marg E (1960) Physiologicalnystagmus in the cat Journal of the Optical Societyof America 50 151ndash155 [PubMed]
Hennig M A Kerscher N J Funke K amp Worgotter F(2002) Stochastic resonance in visual cortical neuronsDoes the eye-tremor actually improve visual acuityNeurocomputing 44ndash46 115ndash120
Hennig M H amp Worgotter F (2004) Eye micro-movements improve stimulus detection beyond theNyquist limit in the peripheral retina In Advances inNeural Information Processing Systems (NIPS2003)(vol 16) Denver CO MIT Press
Hennig M H amp Worgotter F (2007) Effects offixational eye movements on retinal ganglion cellresponses A modelling study Frontiers in Computa-tional Neuroscience 1 2 [PubMed] [Article]
Hermann H T amp Constantine M M (1971) Eyemovements in the goldfish Vision Research 11313ndash331 [PubMed]
Horowitz T S Fine E M Fencsik D E Yurgenson Samp Wolfe J M (2007) Fixational eye movements arenot an index of covert attention PsychologicalScience 18 356ndash363 [PubMed]
Horwitz G D amp Albright T D (2003) Short-latencyfixational saccades induced by luminance incrementsJournal of Neurophysiology 90 1333ndash1339[PubMed] [Article]
Laubrock J Engbert R Rolfs M amp Kliegl R(2007) Microsaccades are an index of covertattention Commentary on Horowitz Fine FencsikYurgenson and Wolfe (2007) Psychological Sci-ence 18 364ndash366 discussion 367ndash368 [PubMed]
Leopold D A amp Logothetis N K (1998) Micro-saccades differentially modulate neural activity in thestriate and extrastriate visual cortex ExperimentalBrain Research 123 341ndash345 [PubMed]
Lettvin J Y Maturana H R MsCulloch W S amp PittsW H (1968) What the frogrsquos eye tells the frogrsquosbrain In W C C a M Balaban (Ed) The mind
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 13
Biological approaches to its functions (pp 233ndash258)John Wiley and Sons London
Macknik S L amp Livingstone M S (1998) Neuronalcorrelates of visibility and invisibility in the primatevisual system Nature Neuroscience 1 144ndash149[PubMed]
Macknik S L amp Martinez-Conde S (2004) The spatialand temporal effects of lateral inhibitory networksand their relevance to the visibility of spatiotemporaledges Neurocomputing 58ndash60 775ndash782
Macknik S L Martinez-Conde S amp Haglund M M(2000) The role of spatiotemporal edges in visibilityand visual masking Proceedings of the NationalAcademy of Sciences of the United States of America97 7556ndash7560 [PubMed] [Article]
Manteuffel G Plasa L Sommer T J amp Wess O(1977) Involuntary eye movements in salamandersNaturwissenschaften 64 533ndash534 [PubMed]
Marchetti E Gauthier G M amp Pellet J (1983)Cerebellar control of eye movements studied withinjection of harmaline in the trained baboon ArchivesItaliennes de Biologies 121 1ndash17 [PubMed]
Martinez-Conde S (2006) Fixational eye movements innormal and pathological vision Progress in BrainResearch 154 151ndash176 [PubMed]
Martinez-Conde S amp Macknik S L (2007a) Bobbingand perceiving Scientific American 297 16
Martinez-Conde S amp Macknik S L (2007b) Windowson the mind Scientific American 297 56ndash63[PubMed]
Martinez-Conde S Macknik S L amp Hubel D H(2000) Microsaccadic eye movements and firing ofsingle cells in the striate cortex of macaque monkeysNature Neuroscience 3 251ndash258 [PubMed]
Martinez-Conde S Macknik S L amp Hubel D H(2002) The function of bursts of spikes during visualfixation in the awake primate lateral geniculate nucleusand primary visual cortex Proceedings of theNational Academy of Sciences of the United Statesof America 99 13920ndash13925 [PubMed] [Article]
Martinez-Conde S Macknik S L amp Hubel D H(2004) The role of fixational eye movements invisual perception Nature Reviews Neuroscience 5229ndash240 [PubMed]
Martinez-Conde S Macknik S L Troncoso X G ampDyar T A (2006) Microsaccades counteract visualfading during fixation Neuron 49 297ndash305[PubMed] [Article]
Mensh B D Aksay E Lee D D Seung H S ampTank D W (2004) Spontaneous eye movements ingoldfish Oculomotor integrator performance plasti-
city and dependence on visual feedback VisionResearch 44 711ndash726 [PubMed]
Miller J A Denning K S George J S MarshakD W amp Kenyon G T (2006) A high frequencyresonance in the responses of retinal ganglion cells torapidly modulated stimuli A computer model VisualNeuroscience 23 779ndash794 [PubMed]
Montgomery J C McVean A R amp McCarthy D(1983) The effects of lowered temperature onspontaneous eye movements in a teleost fish Com-parative Biochemistry and Physiology A Compara-tive Physiology 75 363ndash368 [PubMed]
Motter B C amp Poggio G F (1984) Binocularfixation in the rhesus monkey Spatial and temporalcharacteristics Experimental Brain Research 54304ndash314 [PubMed]
Murakami I amp Cavanagh P (1998) A jitter after-effectreveals motion-based stabilization of vision Nature395 798ndash801 [PubMed]
Nye P W (1969) The monocular eye movements of thepigeon Vision Research 9 133ndash144 [PubMed]
Olveczky B P Baccus S A amp Meister M (2003)Segregation of object and background motion in theretina Nature 423 401ndash408 [PubMed]
Olveczky B P Baccus S A amp Meister M (2007)Retinal adaptation to object motion Neuron 56689ndash700 [PubMed] [Article]
Otero-Millan J Troncoso X Macknik S L Serrano-Pedraza I amp Martinez-Conde S (2008) Saccadesand microsaccades during visual fixation explorationand search Foundations for a common saccadicgenerator Journal of Vision 8(14)21 1ndash18 httpjournalofvisionorg81421 doi10116781421[PubMed] [Article]
Pastor A M Torres B Delgado-Garcia J M ampBaker R (1991) Discharge characteristics of medialrectus and abducens motoneurons in the goldfishJournal of Neurophysiology 66 2125ndash2140 [PubMed]
Pettigrew J D Wallman J amp Wildsoet C F (1990)Saccadic oscillations facilitate ocular perfusion fromthe avian pecten Nature 343 362ndash363 [PubMed]
Pritchard R M amp Heron W (1960) Small eye move-ments of the cat Canadian Journal of Psychology14 131ndash137 [PubMed]
Ratliff F amp Riggs L A (1950) Involuntary motions ofthe eye during monocular fixation Journal of Exper-imental Psychology 40 687ndash701 [PubMed]
Reppas J B UsreyWM amp Reid R C (2002) Saccadiceye movements modulate visual responses in thelateral geniculate nucleus Neuron 35 961ndash974[PubMed] [Article]
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 14
Riggs L A amp Ratliff F (1952) The effects of counter-acting the normal movements of the eye Journal ofthe Optical Society of America 42 872ndash873
Rolfs M Kliegl R amp Engbert R (2008) Toward amodel of microsaccade generation The case ofmicrosaccadic inhibition Journal of Vision 8(11)51ndash23 httpjournalofvisionorg8115 doi1011678115 [PubMed] [Article]
Rolfs M Laubrock J amp Kliegl R (2006) Shorteningand prolongation of saccade latencies followingmicrosaccades Experimental Brain Research 169369ndash376 [PubMed]
Rucci M Iovin R Poletti M amp Santini F (2007)Miniature eye movements enhance fine spatial detailNature 447 851ndash854 [PubMed]
Salas C Navarro F Torres B amp Delgado-Garcia J M(1992) Effects of diazepam and D-amphetamine onrhythmic pattern of eye movements in goldfishNeuroreport 3 131ndash134 [PubMed]
Schipperheyn J J (1963) Respiratory eye movementsand perception of stationary objects in the frog ActaPhysiologica et Pharmacologica Neerlandica 12157ndash159 [PubMed]
Schipperheyn J J (1965) Contrast detection in frogrsquosretina Acta Physiologica et Pharmacologica Neer-landica 13 231ndash277 [PubMed]
Schipperheyn J J (1967) Influence of eye movements onretinal ganglion cell activity in frogs Acta Physi-ologica et Pharmacologica Neerlandica 14 506[PubMed]
Segev R Schneidman E Goodhouse J amp Berry M J2nd (2007) Role of eye movements in the retinalcode for a size discrimination task Journal ofNeurophysiology 98 1380ndash1391 [PubMed] [Article]
Sharpe C R (1972) The visibility and fading of thinlines visualized by their controlled movement acrossthe retina The Journal of Physiology 222 113ndash134[PubMed] [Article]
Shelley M McLaughlin D Shapley R amp Wielaard J(2002) States of high conductance in a large-scalemodel of the visual cortex Journal of ComputationalNeuroscience 13 93ndash109 [PubMed]
Skavenski A A Robinson D A Steinman R M ampTimberlake G T (1975) Miniature eye movementsof fixation in rhesus monkey Vision Research 151269ndash1273 [PubMed]
Snodderly D M (1987) Effects of light and darkenvironments on macaque and human fixational eyemovements Vision Research 27 401ndash415 [PubMed]
Snodderly D M Kagan I amp Gur M (2001) Selectiveactivation of visual cortex neurons by fixational eyemovements Implications for neural coding VisualNeuroscience 18 259ndash277 [PubMed]
Snodderly D M amp Kurtz D (1985) Eye position duringfixation tasks Comparison of macaque and humanVision Research 25 83ndash98 [PubMed]
Steinbach M J amp Money K E (1973) Eye movementsof the owl Vision Research 13 889ndash891 [PubMed]
Steinman R M Haddad G M Skavenski A A ampWyman D (1973) Miniature eye movement Science181 810ndash819 [PubMed]
Torres B Pastor A M Cabrera B Salas C ampDelgado-Garcia J M (1992) Afferents to theoculomotor nucleus in the goldfish (Carassius aur-atus) as revealed by retrograde labeling with horse-radish peroxidase Journal of Comparative Neurology324 449ndash461 [PubMed]
Troncoso X G Macknik S L amp Martinez-Conde S(2008) Microsaccades counteract perceptual filling-inJournal of Vision 8(14)15 1ndash9 httpjournalofvisionorg81415 doi10116781415 [Article]
Troncoso X G Macknik S L Otero-Millan J ampMartinez-Conde S (2008) Microsaccades driveillusory motion in the Enigma illusion Proceedingsof the National Academy of Sciences of the UnitedStates of America 105 16033ndash16038 [PubMed]
Tse P U Sheinberg D S amp Logothetis N K (2004)The distribution of microsaccade directions need notreveal the location of attention Psychological Science15 708ndash710
Turatto M Valsecchi M Tame L amp Betta E (2007)Microsaccades distinguish between global and localvisual processing Neuroreport 18 1015ndash1018[PubMed]
Uchida N Kepecs A amp Mainen Z F (2006) Seeing ata glance smelling in a whiff Rapid forms ofperceptual decision making Nature Reviews Neuro-science 7 485ndash491 [PubMed]
Valsecchi M Betta E amp Turatto M (2007) Visualoddballs induce prolonged microsaccadic inhibitionExperimental Brain Research 177 196ndash208[PubMed]
van Dam L C amp van Ee R (2006) Retinal image shiftsbut not eye movements per se cause alternations inawareness during binocular rivalry Journal of Vision6(11)3 1172ndash1179 httpjournalofvisionorg6113doi1011676113 [PubMed] [Article]
Van der Steen J amp Collewijn H (1984) Ocular stabilityin the horizontal frontal and sagittal planes in therabbit Experimental Brain Research 56 263ndash274[PubMed]
Van Gisbergen J A Robinson D A amp Gielen S(1981) A quantitative analysis of generation ofsaccadic eye movements by burst neurons Journalof Neurophysiology 45 417ndash442 [PubMed]
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 15
Van Gisbergen J A M amp Robinson D A (1977)Generation of micro and macrosaccades by burstneurons in the monkey In R Baker amp A Berthoz(Eds) Control of gaze by brain stem neurons (vol 1pp 301ndash308) New York ElsevierNorth-Holland
Wallman J Pettigrew J D amp Letelier J C (1994)The oscillatory saccades of birds Motoneural activ-ity and possible functions In B T Fuchs A FV Buttner amp D Zee (Eds) Contemporary ocularmotor and vestibular research A tribute to David ARobinson (pp 480ndash487) Stuttgart Germany ThiemePublishing
Williams P E amp Shapley R M (2007) A dynamicnonlinearity and spatial phase specificity in macaque
V1 neurons Journal of Neuroscience 27 5706ndash5718[PubMed] [Article]
Winterson B J amp Robinson D A (1975) Fixationby the alert but solitary cat Vision Research 151349ndash1352 [PubMed]
Yarbus A L (1967) Eye movements and vision(B Haigh Trans) New York Plenum Press
Zuber B L amp Stark L (1965) Microsaccades and thevelocityltamplitude relationship for saccadic eyemovements Science 150 1459ndash1460 [PubMed]
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 16
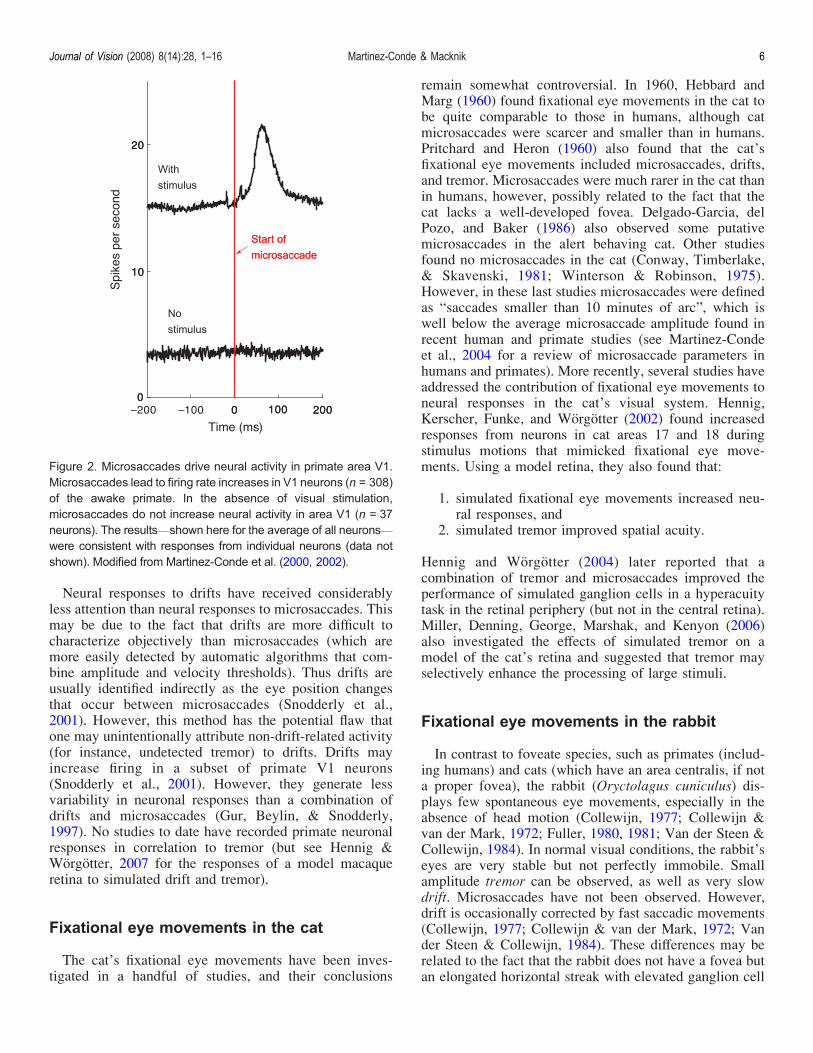
Neural responses to drifts have received considerablyless attention than neural responses to microsaccades Thismay be due to the fact that drifts are more difficult tocharacterize objectively than microsaccades (which aremore easily detected by automatic algorithms that com-bine amplitude and velocity thresholds) Thus drifts areusually identified indirectly as the eye position changesthat occur between microsaccades (Snodderly et al2001) However this method has the potential flaw thatone may unintentionally attribute non-drift-related activity(for instance undetected tremor) to drifts Drifts mayincrease firing in a subset of primate V1 neurons(Snodderly et al 2001) However they generate lessvariability in neuronal responses than a combination ofdrifts and microsaccades (Gur Beylin amp Snodderly1997) No studies to date have recorded primate neuronalresponses in correlation to tremor (but see Hennig ampWorgotter 2007 for the responses of a model macaqueretina to simulated drift and tremor)
Fixational eye movements in the cat
The catrsquos fixational eye movements have been inves-tigated in a handful of studies and their conclusions
remain somewhat controversial In 1960 Hebbard andMarg (1960) found fixational eye movements in the cat tobe quite comparable to those in humans although catmicrosaccades were scarcer and smaller than in humansPritchard and Heron (1960) also found that the catrsquosfixational eye movements included microsaccades driftsand tremor Microsaccades were much rarer in the cat thanin humans however possibly related to the fact that thecat lacks a well-developed fovea Delgado-Garcia delPozo and Baker (1986) also observed some putativemicrosaccades in the alert behaving cat Other studiesfound no microsaccades in the cat (Conway Timberlakeamp Skavenski 1981 Winterson amp Robinson 1975)However in these last studies microsaccades were definedas ldquosaccades smaller than 10 minutes of arcrdquo which iswell below the average microsaccade amplitude found inrecent human and primate studies (see Martinez-Condeet al 2004 for a review of microsaccade parameters inhumans and primates) More recently several studies haveaddressed the contribution of fixational eye movements toneural responses in the catrsquos visual system HennigKerscher Funke and Worgotter (2002) found increasedresponses from neurons in cat areas 17 and 18 duringstimulus motions that mimicked fixational eye move-ments Using a model retina they also found that
1 simulated fixational eye movements increased neu-ral responses and
2 simulated tremor improved spatial acuity
Hennig and Worgotter (2004) later reported that acombination of tremor and microsaccades improved theperformance of simulated ganglion cells in a hyperacuitytask in the retinal periphery (but not in the central retina)Miller Denning George Marshak and Kenyon (2006)also investigated the effects of simulated tremor on amodel of the catrsquos retina and suggested that tremor mayselectively enhance the processing of large stimuli
Fixational eye movements in the rabbit
In contrast to foveate species such as primates (includ-ing humans) and cats (which have an area centralis if nota proper fovea) the rabbit (Oryctolagus cuniculus) dis-plays few spontaneous eye movements especially in theabsence of head motion (Collewijn 1977 Collewijn ampvan der Mark 1972 Fuller 1980 1981 Van der Steen ampCollewijn 1984) In normal visual conditions the rabbitrsquoseyes are very stable but not perfectly immobile Smallamplitude tremor can be observed as well as very slowdrift Microsaccades have not been observed Howeverdrift is occasionally corrected by fast saccadic movements(Collewijn 1977 Collewijn amp van der Mark 1972 Vander Steen amp Collewijn 1984) These differences may berelated to the fact that the rabbit does not have a fovea butan elongated horizontal streak with elevated ganglion cell
Figure 2 Microsaccades drive neural activity in primate area V1Microsaccades lead to firing rate increases in V1 neurons (n = 308)of the awake primate In the absence of visual stimulationmicrosaccades do not increase neural activity in area V1 (n = 37neurons) The resultsVshown here for the average of all neuronsVwere consistent with responses from individual neurons (data notshown) Modified from Martinez-Conde et al (2000 2002)
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 6
counts Olveczky Baccus and Meister (2003) recordedfrom ganglion cells in the rabbit and salamander retinaswhile presenting visual stimuli that were jittered tosimulate fixational eye movements (based on parametersfrom previously published studies by Manteuffel PlasaSommer amp Wess 1977 and Van der Steen amp Collewijn1984) In both species fixational eye movements helpedsegregate a moving foreground from a complex back-ground in a population of ganglion cells
Fixational eye movements inbirds
Fixational eye movements in the pigeon
There are some remarkable differences between fixa-tional eye movements in birds and mammals Nye (1969)identified four types of fixational eye movements in thepigeon (Columba livia) flicks (ie microsaccades)impulse movements drift and oscillations No tremorwas detected Microsaccade frequency was very lowrelative to humans Impulse movements occurred morefrequently usually between bursts of oscillations withamplitudes ranging up to 2 degrees Drift velocitiesranged from 1 to 5 degrees per second Both micro-saccades and drifts occurred in response to movingstimuli Oscillations occurred in short bursts at rates ofabout 30 Hz and could be quite large (up to severaldegrees of amplitude) Bloch Rivaud and Martinoya(1984) reported microsaccades drifts and oscillationsldquosimilar to those described by Nyerdquo but none of thesewere analyzed in detailCyclotorsional oscillations are an integral part of avian
saccades (Wallman Pettigrew amp Letelier 1994) Thusone might speculate that oscillations are associated withavian microsaccades as well especially if saccades andmicrosaccades share a common generator (Engbert 2006Otero-Millan et al 2008 Rolfs et al 2008 2006 Zuberamp Stark 1965) However this possibility has not yet beenaddressed in the literature The function of oscillations isunknown (Carpenter 1988) but one hypothesis is thatthey serve to facilitate the delivery of arterial blood to theocular capillary bed (Pettigrew Wallman amp Wildsoet1990 Wallman et al 1994)
Fixational eye movements in the owl
Steinbach and Money (1973) recorded fixational eyemovements in the great horned owl (Bubo virginianus)a species previously thought incapable of moving itseyes The owlrsquos fixational eye movements includeflicks or microsaccades drift tremor and oscillations
Microsaccades seemed correlated to visual events ofinterest to the owl they were 1 deg or less in amplitudeand had peak velocities of about 10 degsec Drifts wereslow motions of the eye in between fixations withamplitudes of 1 degree or less Tremor amplitude wasabout 1 arcmin with frequencies around 20 Hz High-frequency oscillations (25 Hz) of short duration (025 sec)were very similar to those previously found in the pigeon(Nye 1969) No studies to date have examined theperceptual or physiological effects of fixational eyemovements in any bird species
Fixational eye movements inreptiles
Fixational eye movements in the turtle
Greschner Bongard Rujan and Ammermuller (2002)identified two types of fixational eye movements in theturtle (Pseudemys scripta elegans) which they calledldquodrift-likerdquo and ldquoperiodicrdquo Drift-like motions were smalland slow (with velocities comparable to human drift) andwere superimposed by larger and faster periodic componentsPeriodic motion had retinal amplitudes on the order of thediameter of a photoreceptor which are comparable to thoseof human tremor On the other hand periodic motionrsquosfrequency (5 Hz) velocity and orderly relationship betweenvelocity and amplitude are closer to those of human micro-saccades (see Martinez-Conde et al 2004 for a review oftremor and microsaccade parameters in humans)Simulated periodic motion generated strong synchronous
firing in the turtlersquos retina whereas simulated drift-likemotion had little effect (Figure 3) These results agree withprevious physiological and modeling studies in the primatevisual system in which strong neural transients wereobserved in response to microsaccades (Donner amp Hemila2007 Martinez-Conde 2006 Martinez-Conde et al 20002002) Such neural transients may underlie the behavior ofcortical neurons as coincidence detectors (ShelleyMcLaughlin Shapley amp Wielaard 2002 Williams ampShapley 2007) Moreover neural transients to stimulionsets and terminations (similar to those produced bymicrosaccades in the primate visual system Martinez-Conde 2006 Martinez-Conde et al 2000 2002) havebeen related to target visibility in visual masking para-digms (Macknik amp Livingstone 1998 Macknik ampMartinez-Conde 2004 Macknik Martinez-Conde ampHaglund 2000)In the turtle retina periodic motion-driven neurons with
receptive fields located along contrast borders weremoreover synchronized and reliably indicated the preced-ing motions The authors proposed that this synchroniza-tion of retinal activity could be used by the brain to
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 7
improve the estimation of stimulus features such asspatial frequency (Greschner et al 2002) This findingcould have a parallel in the primate visual system Donnerand Hemila (2007) recently proposed that microsaccadessignificantly ldquore-sharpenrdquo the image and improve spatialresolution in primates Rucci Iovin Poletti and Santini(2007) also found that fixational eye movements enhancefine spatial detail in human subjects
Fixational eye movements in the chameleon
Fixational eye movements in the chameleon have notbeen studied in detail However Gioanni and colleaguesfound that the chameleon (Chamaleo chamaleo) makessaccades during fixation The authors speculated that theseprobably correspond to fixational microsaccades found inother foveate species (Gioanni Bennis amp Sansonetti1993)
Fixational eye movements inamphibians
Fixational (and more generally spontaneous) eye move-ments vary widely among amphibians The frog (anafoveate amphibian) shows a general lack of eye andhead movements except for those that stabilize the retinalimage Dieringer and colleagues (Dieringer amp Daunicht1986 Dieringer amp Precht 1982 Dieringer Precht ampBlight 1982) found that the grass frog (Rana temporaria)produces no spontaneous eye movements in the absenceof head movements with the possible exception of tremor(similarly to the rabbit another afoveate species Van derSteen amp Collewijn 1984) It is likely that the frogrsquos lack ofeye movements leads to fast visual fading of all stationaryobjects in its field of view (Ditchburn amp Ginsborg 1952Yarbus 1967) Lettvin Maturana MsCulloch and Pitts
Figure 3 Fixational eye movements increase retinal activity in the turtle (A) Continuous recording from 54 retinal ganglion cells in theturtle during darkness after switching on a stationary grating and during wobbling of this grating The grating was wobbled to simulateperiodic (ie tremor-like) eye movements (B) Responses of a single ganglion cell to a drifting contrast border of velocity comparable toocular drifts (C) When the drift is superimposed with wobbling the responses of the cell shown in (B) increase dramatically FromGreschner et al (2002) and modified by Martinez-Conde et al (2004)
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 8
(1968) stated that a frog ldquowill starve to death surroundedby food if it is not movingrdquo (but see Dieringer amp Daunicht1986 for a counterpoint) Small head motions or eyemovements correlated with respiratory motions maydiminish fading to some extent (Dieringer amp Precht1982 Schipperheyn 1963 1965 1967)As a side note the frogrsquos respiratory eye movements
play no role in the respiratory function ldquoin the course ofeach respiratory cycle increase of pressure in the frogrsquosbuccal cavity protrudes the eye and causes the opticalaxis to deviate upwards over 25ndash35 minutes of arcrdquo(Schipperheyn 1963) That is respiratory eye movementsare merely synchronized with respirationAutrum (1959) found no evidence of respiratory eye
movements in the frog Ewert andBorchers (1974) observedthat stationary stimuli elicit no responses from visualneurons in the common toad (Bufo Bufo L) regardless ofrespiratory pressure changes in the buccal cavity
Fixational eye movements in the salamander
Manteuffel et al (1977) identified two types of fixa-tional eye movements in salamanders tremor-like andrespiratory eye movements Tremor-like is a rapid move-ment of small amplitude Torsional eye movements resultfrom the animalrsquos breathing (also found in the frog bySchipperheyn 1963) Additionally flicks are occasionallyobserved which rapidly change the position of gaze It isunclear whether such flicks may generally correspond towhat we now call microsaccades andor to voluntarysaccades Based on these observations the authors pro-posed that unlike frogs and toads (Autrum 1959 Ewert ampBorchers 1974 Lettvin et al 1968) salamanders cancontinuously perceive stationary objects (Manteuffel et al1977) Fixational eye movements in the salamander (Tigersalamander Ambystoma tigrinum) may also help segre-gate object and background motion in the retina(Olveczky et al 2003 see also Fixational eye movementsin the rabbit section) and even work to emphasize novelvisual features (Olveczky et al 2007 see Baccus et al2008 for details on the retinal neurons and synapses thatmay underlie the computation of differential motionbetween an object and its background)
Fixational eye movements in fish
Spontaneous eye movements in the goldfish
Eye movements during fixation have been studied inboth afoveate (goldfish Carassius auratus) and foveate(archer fish Toxotes chatareus) fish species The goldfisha long-standing model organism in vision and oculomotor
research (Aksay et al 2003 2007 Cabrera Torres PasaroPastor amp Delgado-Garcia 1992 Pastor Torres Delgado-Garcia amp Baker 1991 Salas Navarro Torres ampDelgado-Garcia 1992 Torres Pastor Cabrera Salas ampDelgado-Garcia 1992) produces three types of sponta-neous eye movements saccades stretch and drift (Easter1971 Mensh Aksay Lee Seung amp Tank 2004) Weshould note that in the motionless goldfish eye move-ments are very infrequent Those that do occur consistmainly of saccades slow drifts are very rare (Easter Johnsamp Heckenlively 1974) During saccades (identified bythresholding the eye velocity at 5 degsec) both eyes moverapidly simultaneously and usually in the same direction(Mensh et al 2004) Stretches occur every 2ndash3 min and arecharacterized by a brief simultaneous large excursion ofboth eyes to extreme temporal positions (Easter 1971Mensh et al 2004) The function of stretches is not known(Mensh et al 2004) Because fixations were defined asthose periods that were free of saccadic and stretchmovements (Easter 1971) only drifts (from the threetypes of spontaneous movements) can technically beconsidered ldquofixational eye movementsrdquo However thegoldfish saccadic main sequence includes saccades withequivalent amplitudes and velocities to those found inhuman microsaccades (Mensh et al 2004 Zuber amp Stark1965) Indeed microsaccades have been identified in other(foveate) fish species although they are rarer than inhumans (Segev et al 2007) Small saccades in thegoldfish tend to be divergent whereas large saccadestend to be convergent (Mensh et al 2004) Drift occurssimultaneously in both eyes usually in the samedirection Drift velocities are almost exclusively e1 degsec(Mensh et al 2004) No tremor has been identified in thegoldfish
Spontaneous eye movements in teleost fish
Spontaneous eye movements have been studied in theNew Zealand parore (Girella tricuspidata MontgomeryMcVean amp McCarthy 1983) The pattern of eye move-ments was similar to that previously described in goldfish(Easter 1971 Hermann amp Constantine 1971)
Fixational eye movements in the archer fish
A recent study in the archer fish (Segev et al 2007) hasprovided the first perceptual and physiological correlatesof fixational eye movements in any fish species Archerfish have remarkable visual abilities they shoot downinsects resting on foliageVas well as stationary targetscovering a single retinal photoreceptorVwith squirts ofwater from their mouths they predict the location atwhich the insect will splash down in G100 msec and theydeduce the absolute size of an object presented at differentviewing distances (Supplementary Videos 2 and 3) Segev
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 9
et al (2007) identified three types of fixational eyemovements in the archer fish microsaccades drift andtremor Microsaccades were rare and were not analyzed indetail Drift velocities were e040 degsec Tremor was ahigh-frequency oscillatory movement (the primary com-ponent was 5 Hz) with peak-to-peak amplitude of 01ndash02 deg (corresponding to an image displacement of 1ndash2photoreceptors) and velocity of about 1 degsec (Figure 4)Whereas the amplitude and velocity of tremor in thearcher fish is comparable to those in humans its frequencyis much lower (5 Hz vs 940 Hz) Interestingly archer fishtremor is very similar both in amplitude (photoreceptorwidths covered) as well as in frequency (5 Hz) to thetremor-like periodic motion in the turtle (discussedearlier Greschner et al 2002)Segev et al (2007) trained archer fish to perform a size
discrimination task the fish had to distinguish a medium-sized target from larger and smaller distracters (theyindicated their choice by shooting a jet of water) To relatethis behavior to responses from retinal neurons theauthors recorded from populations of retinal ganglioncells while presenting objects of different sizes moved tosimulate saccades drifts and tremor Drifts and tremorelicited weak neuronal responses that were not veryinformative about the spatial structure of the stimulus(although they did provide some information about target
size in agreement with the results of Greschner et al2002 in the turtle) Moreover performance was similar fortremor alone and for tremor plus drift indicating that driftplays a negligible role in revealing spatial structure (alsoin agreement with Greschner et al 2002)When present the firing rate was phase-locked to
tremor Saccades were more effective than drifts or tremorin driving informative responses from the ganglion cells(consistent with theoretical and physiological studies ofmicrosaccade-driven neuronal activity in the primateretina and LGN Donner amp Hemila 2007 Martinez-Conde et al 2002) Segev et al (2007) proposed that inprimates both saccades and microsaccades (which arerare in the archer fish) may contribute to size discrim-ination tasks as well as enhance spatial detail whereasdrifts and tremor by themselves may not be sufficient toimprove visual perception However it is important toremember that central vision in humans can be maintainedby tremor andor drift even when microsaccades aresuppressed during strict fixation (Martinez-Conde 2006Martinez-Conde et al 2004 2006) On the other handone cannot rule out the possibility that if one couldcompletely eliminate drifts and tremor microsaccadesmight then suffice to sustain both central and peripheralvision in primates and humans Future experiments areneeded to further explore these ideas
Figure 4 Archer fish eye movements (A) Example trace of eye position as a function of time as measured by search coil (B) Example ofa single 15-s fixation superimposed on a grid representing the photoreceptor mosaic The back and forth movement is caused by tremor(C) Distribution of fixation durations (taken as time interval between saccades) (D) Power spectrum of horizontal fixational eyemovements (E) Distribution of tremor amplitude (data taken from 1 animal in a 15-min session) Modified from Segev et al (2007)
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 10
Discussion
The vast majority of eye movement studies conductedin non-human species to date have not analyzed eyeposition data in the range of fixational eye movementsThus eye movements during fixation have been reportedin only a handful of vertebrates Most of these studieshave concentrated solely on the physical parameters ofthe fixational eye movements without addressing theirphysiological and perceptual consequences Fortunatelythis trend has started to reverse in recent years with agrowing list of papers addressing the effects of non-human fixational eye movements in physiology andperception This is an important development A largeamount of all psychophysical and physiological visualresearch has been conducted during conditions of visualfixation Therefore understanding the precise physiolog-ical and perceptual contributions of fixational eye move-ments may be critical to the interpretation of previous andfuture vision researchThere are some interesting differences and similarities
in the types of fixational eye movements produced bydifferent species For instance ocular tremor is possiblyubiquitous to all vertebrates The fact that tremor has notbeen observed in all species studied could be due to thetechnical difficulty of recording tremor accurately (ietremor amplitudes and frequencies are usually in the rangeof the recording systemrsquos noise) Drift has also beenrecorded in most vertebrates and it may be as omnipre-sent as tremor Conversely microsaccades seem mostimportant in species with foveal vision (ie primates andhumans) whereas they are scarce or absent in afoveatespecies (ie rabbits frogs goldfish) A link betweenmicrosaccades and foveal vision may reflect the two-pronged functional significance of microsaccades inhumans and primates first microsaccades may benecessary to correct the slow drift of the eye and bringback the objectregion of interest to the retinal area wherephotoreceptor density is highest (Cornsweet 1956) Thecorrective role of human microsaccades has been demon-strated in recent studies (Engbert amp Kliegl 2004 Engbertamp Mergenthaler 2006) Second the dynamics of micro-saccades themselves may improve spatial resolution(Donner amp Hemila 2005)In reviewing the patterns of fixational eye movements
across vertebrates it is tempting to draw parallels withother sensory modalities Fixational eye movementsrefresh the retinal images during fixation and thus preventneural adaptation and visual fading But this mechanism isnot circumscribed to the visual system Sniffs in rodentolfaction discretely sample sensory information every200ndash300 msec and thus are similar in their temporaldynamics to primate saccades (Otero-Millan et al 2008Uchida Kepecs amp Mainen 2006) and microsaccades(Otero-Millan et al 2008) A similar mode of discretesampling may also be at play when objects are recognized
through tactile information for instance if we sweep ourfingertips over an objectrsquos surface with our eyes closed orwhen blind individuals read Braille scriptOne intriguing possibility that merits further experi-
ments and analyses is the potential correlation between thespatiotemporal parameters of fixational eye movements(such as those summarized in Table 1) and the adaptationproperties of visual neurons across species For instancethe greater ocular stability of some species (rabbitgoldfish frog etc) versus others (primates) could beassociated with some particular adaptive properties oftheir visual cells Thus future comparative studies shouldpay attention to the response properties of visual cellstriggered by fixational eye movements in the presence ofstationary stimuli Other potential factors underlying thevariation in fixational eye movements across species couldbe differences in oculomotor control at the level of the eyemuscles or perhaps differences in retinal organizationFuture research will further improve our understanding
of fixational eye movements and the neural activity thatmaintains the visibility of stationary visual scenes inhuman and non-human species One interesting questionthat remains to be addressed concerns the role of tremor invision and visual physiology Because tremor frequenciesare well over the flicker fusion frequencies in humans ithas been argued that tremor of the visual image would beineffective as a stimulus (Ditchburn 1955 Gerrits ampVendrik 1970 Sharpe 1972) However recent studieshave found simulated tremor to enhance visual processingand improve visual acuity in a model retina (Hennig et al2002 Miller et al 2006) Follow-up research should beaimed to replicate these findings in actual neurons withcorrelated perceptual reports One should also note thatearly visual neurons can follow high-frequency flickeringthat surpasses the perceptual threshold for flicker fusion(Martinez-Conde et al 2002) Thus high-frequencytremor could be adequate to maintain activity in the earlyvisual system which then may lead to visual perception inhigher visual areas Another significant question thatremains unanswered concerns the oculomotor bases ofthe various types of fixational eye movements acrossspecies To date there is too little information on thisimportant topic to draw a conclusion
Conclusions
We have reviewed the parameters of fixational eyemovements in non-human mammals birds reptilesamphibians and fish as well as their consequences forphysiology and perception Fixational eye movementsand microsaccades in particular appear to be mostimportant in foveate vs afoveate species Animals lackingfixational eye movements such as afoveate frogs andtoads may be incapable of seeing stationary objects
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 11
during visual fixation Future research should furtherexplore the perceptual effects of fixational eye movementsand determine the oculomotor mechanisms responsible fortheir generation across species
Acknowledgments
We thank Mona Stewart and Isabel Gomez-Caraballo fortechnical assistance Jorge Otero-Millan for programmingassistance and Dr Xoana G Troncoso for comments on themanuscript This study was funded through grants fromthe Barrow Neurological Foundation (to SMC and SLM)the National Science Foundation (Award 0643306 toSMC) the Arizona Biomedical Research Commission(Award 07-102 to SMC) and Science Foundation Arizona(SLM)
Commercial relationships noneCorresponding author Susana Martinez-Conde PhDEmail smartneuralcorrelatecomAddress Barrow Neurological Institute 350 W ThomasRd Phoenix AZ 85013 USA
References
Aksay E Major G Goldman M S Baker R SeungH S amp Tank D W (2003) History dependence ofrate covariation between neurons during persistentactivity in an oculomotor integrator Cerebral Cortex13 1173ndash1184 [PubMed] [Article]
Aksay E Olasagasti I Mensh B D Baker RGoldman M S amp Tank D W (2007) Functionaldissection of circuitry in a neural integrator NatureNeuroscience 10 494ndash504 [PubMed]
Autrum H (1959) Das Fehlen unwillkurlicher Augenbe-wegungen beim Frosch Naturwissenschaften 46435
Baccus S A Olveczky B P Manu M amp Meister M(2008) A retinal circuit that computes object motionJournal of Neuroscience 28 6807ndash6817 [PubMed]
Bair W amp OrsquoKeefe L P (1998) The influence offixational eye movements on the response of neuronsin area MT of the macaque Visual Neuroscience 15779ndash786 [PubMed]
Betta E amp Turatto M (2006) Are you ready I can tellby looking at your microsaccades Neuroreport 171001ndash1004 [PubMed]
Bloch S Rivaud S amp Martinoya C (1984) Compar-ing frontal and lateral viewing in the pigeon IIIDifferent patterns of eye movements for binocularand monocular fixation Behavioural Brain Research13 173ndash182 [PubMed]
Cabrera B Torres B Pasaro R Pastor A M ampDelgado-Garcia J M (1992) A morphological studyof abducens nucleus motoneurons and internuclearneurons in the goldfish (Carassius auratus) BrainResearch Bulletin 28 137ndash144 [PubMed]
Carpenter R H S (1988) Movements of the eyes(2nd ed) Pion London
Collewijn H (1977) Eye- and head movements infreely moving rabbits The Journal of Physiology266 471ndash498 [PubMed] [Article]
Collewijn H amp van der Mark F (1972) Ocular stabilityin variable visual feedback conditions in the rabbitBrain Research 36 47ndash57 [PubMed]
Conway J L Timberlake G T amp Skavenski A A(1981) Oculomotor changes in cats reared withoutexperiencing continuous retinal image motion Exper-imental Brain Research 43 229ndash232 [PubMed]
Cornsweet T N (1956) Determination of the stimuli forinvoluntary drifts and saccadic eye movements Jour-nal of the Optical Society of America 46 987ndash993[PubMed]
Delgado-Garcia J M del Pozo F amp Baker R (1986)Behavior of neurons in the abducens nucleus of thealert catVII Internuclear neurons Neuroscience 17953ndash973 [PubMed]
Dieringer N amp Daunicht W J (1986) Image fadingVAproblem for frogs Naturwissenschaften 73 330ndash332[PubMed]
Dieringer N amp Precht W (1982) Compensatory headand eye movements in the frog and their contributionto stabilization of gaze Experimental BrainResearch 47 394ndash406 [PubMed]
Dieringer N Precht W amp Blight A R (1982)Resetting fast phases of head and eye and theirlinkage in the frog Experimental Brain Research 47407ndash416 [PubMed]
Ditchburn R W (1955) Eye-movements in relation toretinal action Optica Acta 1 171ndash176
Ditchburn R W amp Ginsborg B L (1952) Vision with astabilized retinal image Nature 170 36ndash37[PubMed]
Ditchburn R W amp Ginsborg B L (1953) Involuntaryeye movements during fixation The Journal ofPhysiology 119 1ndash17 [PubMed] [Article]
Donner K amp Hemila S (2007) Modelling the effect ofmicrosaccades on retinal responses to stationarycontrast patterns Vision Research 47 1166ndash1177[PubMed]
Donner K K amp Hemila S (2005) The possible purposeof microsaccades Paper presented at the EuropeanConference on Visual Perception A Coruna Spain
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 12
Easter S S Jr (1971) Spontaneous eye movements inrestrained goldfish Vision Research 11 333ndash342[PubMed]
Easter S S Jr Johns P R amp Heckenlively D R(1974) Horizontal compensatory eye movements ingoldfish (Carassius auratus) I The normal animalJournal of Comparative Physiology 92 23ndash35
Engbert R (2006) Microsaccades A microcosm forresearch on oculomotor control attention and visualperception Progress Brain Research 154 177ndash192[PubMed]
Engbert R amp Kliegl R (2003a) Binocular coordinationin microsaccades In R R J Hyona amp H Deubel(Eds) The mindrsquos eyes Cognitive and appliedaspects of eye movements (pp 103ndash117) OxfordUK Elsevier
Engbert R amp Kliegl R (2003b) Microsaccades uncoverthe orientation of covert attention Vision Research43 1035ndash1045 [PubMed]
Engbert R amp Kliegl R (2004) Microsaccades keep theeyesrsquo balance during fixation Psychological Science15 431ndash436 [PubMed]
Engbert R amp Mergenthaler K (2006) Microsaccadesare triggered by low retinal image slip Proceedingsof the National Academy of Sciences of the UnitedStates of America 103 7192ndash7197 [PubMed][Article]
Ewert J P amp Borchers H W (1974) Antwort vonretinalen Ganglienzellen bei freibeweglichen Kroten(Bufo Bufo L) Journal of Comparative and Physi-ology 92 117ndash130
Fuller J H (1980) Linkage of eye and head movementsin the alert rabbit Brain Research 194 219ndash222[PubMed]
Fuller J H (1981) Eye and head movements duringvestibular stimulation in the alert rabbit BrainResearch 205 363ndash381 [PubMed]
Galfano G Betta E amp Turatto M (2004) Inhibition ofreturn in microsaccades Experimental BrainResearch 159 400ndash404 [PubMed]
Gerrits H J amp Vendrik A J (1970) Artificial move-ments of a stabilized image Vision Research 101443ndash1456 [PubMed]
Gioanni H Bennis M amp Sansonetti A (1993) Visualand vestibular reflexes that stabilize gaze in thechameleon Visual Neuroscience 10 947ndash956[PubMed]
Goffart L Quinet J Chavane F amp Masson G S(2006) Influence of background illumination onfixation and visually guided saccades in the rhesusmonkey Vision Research 46 149ndash162 [PubMed]
Greschner M Bongard M Rujan P amp Ammermuller J(2002) Retinal ganglion cell synchronization by fixa-tional eye movements improves feature estimationNature Neuroscience 5 341ndash347 [PubMed]
Gur M Beylin A amp Snodderly D M (1997)Response variability of neurons in primary visualcortex (V1) of alert monkeys Journal of Neuro-science 17 2914ndash2920 [PubMed] [Article]
Hafed Z M amp Clark J J (2002) Microsaccades as anovert measure of covert attention shifts VisionResearch 42 2533ndash2545 [PubMed]
Hebbard F W amp Marg E (1960) Physiologicalnystagmus in the cat Journal of the Optical Societyof America 50 151ndash155 [PubMed]
Hennig M A Kerscher N J Funke K amp Worgotter F(2002) Stochastic resonance in visual cortical neuronsDoes the eye-tremor actually improve visual acuityNeurocomputing 44ndash46 115ndash120
Hennig M H amp Worgotter F (2004) Eye micro-movements improve stimulus detection beyond theNyquist limit in the peripheral retina In Advances inNeural Information Processing Systems (NIPS2003)(vol 16) Denver CO MIT Press
Hennig M H amp Worgotter F (2007) Effects offixational eye movements on retinal ganglion cellresponses A modelling study Frontiers in Computa-tional Neuroscience 1 2 [PubMed] [Article]
Hermann H T amp Constantine M M (1971) Eyemovements in the goldfish Vision Research 11313ndash331 [PubMed]
Horowitz T S Fine E M Fencsik D E Yurgenson Samp Wolfe J M (2007) Fixational eye movements arenot an index of covert attention PsychologicalScience 18 356ndash363 [PubMed]
Horwitz G D amp Albright T D (2003) Short-latencyfixational saccades induced by luminance incrementsJournal of Neurophysiology 90 1333ndash1339[PubMed] [Article]
Laubrock J Engbert R Rolfs M amp Kliegl R(2007) Microsaccades are an index of covertattention Commentary on Horowitz Fine FencsikYurgenson and Wolfe (2007) Psychological Sci-ence 18 364ndash366 discussion 367ndash368 [PubMed]
Leopold D A amp Logothetis N K (1998) Micro-saccades differentially modulate neural activity in thestriate and extrastriate visual cortex ExperimentalBrain Research 123 341ndash345 [PubMed]
Lettvin J Y Maturana H R MsCulloch W S amp PittsW H (1968) What the frogrsquos eye tells the frogrsquosbrain In W C C a M Balaban (Ed) The mind
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 13
Biological approaches to its functions (pp 233ndash258)John Wiley and Sons London
Macknik S L amp Livingstone M S (1998) Neuronalcorrelates of visibility and invisibility in the primatevisual system Nature Neuroscience 1 144ndash149[PubMed]
Macknik S L amp Martinez-Conde S (2004) The spatialand temporal effects of lateral inhibitory networksand their relevance to the visibility of spatiotemporaledges Neurocomputing 58ndash60 775ndash782
Macknik S L Martinez-Conde S amp Haglund M M(2000) The role of spatiotemporal edges in visibilityand visual masking Proceedings of the NationalAcademy of Sciences of the United States of America97 7556ndash7560 [PubMed] [Article]
Manteuffel G Plasa L Sommer T J amp Wess O(1977) Involuntary eye movements in salamandersNaturwissenschaften 64 533ndash534 [PubMed]
Marchetti E Gauthier G M amp Pellet J (1983)Cerebellar control of eye movements studied withinjection of harmaline in the trained baboon ArchivesItaliennes de Biologies 121 1ndash17 [PubMed]
Martinez-Conde S (2006) Fixational eye movements innormal and pathological vision Progress in BrainResearch 154 151ndash176 [PubMed]
Martinez-Conde S amp Macknik S L (2007a) Bobbingand perceiving Scientific American 297 16
Martinez-Conde S amp Macknik S L (2007b) Windowson the mind Scientific American 297 56ndash63[PubMed]
Martinez-Conde S Macknik S L amp Hubel D H(2000) Microsaccadic eye movements and firing ofsingle cells in the striate cortex of macaque monkeysNature Neuroscience 3 251ndash258 [PubMed]
Martinez-Conde S Macknik S L amp Hubel D H(2002) The function of bursts of spikes during visualfixation in the awake primate lateral geniculate nucleusand primary visual cortex Proceedings of theNational Academy of Sciences of the United Statesof America 99 13920ndash13925 [PubMed] [Article]
Martinez-Conde S Macknik S L amp Hubel D H(2004) The role of fixational eye movements invisual perception Nature Reviews Neuroscience 5229ndash240 [PubMed]
Martinez-Conde S Macknik S L Troncoso X G ampDyar T A (2006) Microsaccades counteract visualfading during fixation Neuron 49 297ndash305[PubMed] [Article]
Mensh B D Aksay E Lee D D Seung H S ampTank D W (2004) Spontaneous eye movements ingoldfish Oculomotor integrator performance plasti-
city and dependence on visual feedback VisionResearch 44 711ndash726 [PubMed]
Miller J A Denning K S George J S MarshakD W amp Kenyon G T (2006) A high frequencyresonance in the responses of retinal ganglion cells torapidly modulated stimuli A computer model VisualNeuroscience 23 779ndash794 [PubMed]
Montgomery J C McVean A R amp McCarthy D(1983) The effects of lowered temperature onspontaneous eye movements in a teleost fish Com-parative Biochemistry and Physiology A Compara-tive Physiology 75 363ndash368 [PubMed]
Motter B C amp Poggio G F (1984) Binocularfixation in the rhesus monkey Spatial and temporalcharacteristics Experimental Brain Research 54304ndash314 [PubMed]
Murakami I amp Cavanagh P (1998) A jitter after-effectreveals motion-based stabilization of vision Nature395 798ndash801 [PubMed]
Nye P W (1969) The monocular eye movements of thepigeon Vision Research 9 133ndash144 [PubMed]
Olveczky B P Baccus S A amp Meister M (2003)Segregation of object and background motion in theretina Nature 423 401ndash408 [PubMed]
Olveczky B P Baccus S A amp Meister M (2007)Retinal adaptation to object motion Neuron 56689ndash700 [PubMed] [Article]
Otero-Millan J Troncoso X Macknik S L Serrano-Pedraza I amp Martinez-Conde S (2008) Saccadesand microsaccades during visual fixation explorationand search Foundations for a common saccadicgenerator Journal of Vision 8(14)21 1ndash18 httpjournalofvisionorg81421 doi10116781421[PubMed] [Article]
Pastor A M Torres B Delgado-Garcia J M ampBaker R (1991) Discharge characteristics of medialrectus and abducens motoneurons in the goldfishJournal of Neurophysiology 66 2125ndash2140 [PubMed]
Pettigrew J D Wallman J amp Wildsoet C F (1990)Saccadic oscillations facilitate ocular perfusion fromthe avian pecten Nature 343 362ndash363 [PubMed]
Pritchard R M amp Heron W (1960) Small eye move-ments of the cat Canadian Journal of Psychology14 131ndash137 [PubMed]
Ratliff F amp Riggs L A (1950) Involuntary motions ofthe eye during monocular fixation Journal of Exper-imental Psychology 40 687ndash701 [PubMed]
Reppas J B UsreyWM amp Reid R C (2002) Saccadiceye movements modulate visual responses in thelateral geniculate nucleus Neuron 35 961ndash974[PubMed] [Article]
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 14
Riggs L A amp Ratliff F (1952) The effects of counter-acting the normal movements of the eye Journal ofthe Optical Society of America 42 872ndash873
Rolfs M Kliegl R amp Engbert R (2008) Toward amodel of microsaccade generation The case ofmicrosaccadic inhibition Journal of Vision 8(11)51ndash23 httpjournalofvisionorg8115 doi1011678115 [PubMed] [Article]
Rolfs M Laubrock J amp Kliegl R (2006) Shorteningand prolongation of saccade latencies followingmicrosaccades Experimental Brain Research 169369ndash376 [PubMed]
Rucci M Iovin R Poletti M amp Santini F (2007)Miniature eye movements enhance fine spatial detailNature 447 851ndash854 [PubMed]
Salas C Navarro F Torres B amp Delgado-Garcia J M(1992) Effects of diazepam and D-amphetamine onrhythmic pattern of eye movements in goldfishNeuroreport 3 131ndash134 [PubMed]
Schipperheyn J J (1963) Respiratory eye movementsand perception of stationary objects in the frog ActaPhysiologica et Pharmacologica Neerlandica 12157ndash159 [PubMed]
Schipperheyn J J (1965) Contrast detection in frogrsquosretina Acta Physiologica et Pharmacologica Neer-landica 13 231ndash277 [PubMed]
Schipperheyn J J (1967) Influence of eye movements onretinal ganglion cell activity in frogs Acta Physi-ologica et Pharmacologica Neerlandica 14 506[PubMed]
Segev R Schneidman E Goodhouse J amp Berry M J2nd (2007) Role of eye movements in the retinalcode for a size discrimination task Journal ofNeurophysiology 98 1380ndash1391 [PubMed] [Article]
Sharpe C R (1972) The visibility and fading of thinlines visualized by their controlled movement acrossthe retina The Journal of Physiology 222 113ndash134[PubMed] [Article]
Shelley M McLaughlin D Shapley R amp Wielaard J(2002) States of high conductance in a large-scalemodel of the visual cortex Journal of ComputationalNeuroscience 13 93ndash109 [PubMed]
Skavenski A A Robinson D A Steinman R M ampTimberlake G T (1975) Miniature eye movementsof fixation in rhesus monkey Vision Research 151269ndash1273 [PubMed]
Snodderly D M (1987) Effects of light and darkenvironments on macaque and human fixational eyemovements Vision Research 27 401ndash415 [PubMed]
Snodderly D M Kagan I amp Gur M (2001) Selectiveactivation of visual cortex neurons by fixational eyemovements Implications for neural coding VisualNeuroscience 18 259ndash277 [PubMed]
Snodderly D M amp Kurtz D (1985) Eye position duringfixation tasks Comparison of macaque and humanVision Research 25 83ndash98 [PubMed]
Steinbach M J amp Money K E (1973) Eye movementsof the owl Vision Research 13 889ndash891 [PubMed]
Steinman R M Haddad G M Skavenski A A ampWyman D (1973) Miniature eye movement Science181 810ndash819 [PubMed]
Torres B Pastor A M Cabrera B Salas C ampDelgado-Garcia J M (1992) Afferents to theoculomotor nucleus in the goldfish (Carassius aur-atus) as revealed by retrograde labeling with horse-radish peroxidase Journal of Comparative Neurology324 449ndash461 [PubMed]
Troncoso X G Macknik S L amp Martinez-Conde S(2008) Microsaccades counteract perceptual filling-inJournal of Vision 8(14)15 1ndash9 httpjournalofvisionorg81415 doi10116781415 [Article]
Troncoso X G Macknik S L Otero-Millan J ampMartinez-Conde S (2008) Microsaccades driveillusory motion in the Enigma illusion Proceedingsof the National Academy of Sciences of the UnitedStates of America 105 16033ndash16038 [PubMed]
Tse P U Sheinberg D S amp Logothetis N K (2004)The distribution of microsaccade directions need notreveal the location of attention Psychological Science15 708ndash710
Turatto M Valsecchi M Tame L amp Betta E (2007)Microsaccades distinguish between global and localvisual processing Neuroreport 18 1015ndash1018[PubMed]
Uchida N Kepecs A amp Mainen Z F (2006) Seeing ata glance smelling in a whiff Rapid forms ofperceptual decision making Nature Reviews Neuro-science 7 485ndash491 [PubMed]
Valsecchi M Betta E amp Turatto M (2007) Visualoddballs induce prolonged microsaccadic inhibitionExperimental Brain Research 177 196ndash208[PubMed]
van Dam L C amp van Ee R (2006) Retinal image shiftsbut not eye movements per se cause alternations inawareness during binocular rivalry Journal of Vision6(11)3 1172ndash1179 httpjournalofvisionorg6113doi1011676113 [PubMed] [Article]
Van der Steen J amp Collewijn H (1984) Ocular stabilityin the horizontal frontal and sagittal planes in therabbit Experimental Brain Research 56 263ndash274[PubMed]
Van Gisbergen J A Robinson D A amp Gielen S(1981) A quantitative analysis of generation ofsaccadic eye movements by burst neurons Journalof Neurophysiology 45 417ndash442 [PubMed]
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 15
Van Gisbergen J A M amp Robinson D A (1977)Generation of micro and macrosaccades by burstneurons in the monkey In R Baker amp A Berthoz(Eds) Control of gaze by brain stem neurons (vol 1pp 301ndash308) New York ElsevierNorth-Holland
Wallman J Pettigrew J D amp Letelier J C (1994)The oscillatory saccades of birds Motoneural activ-ity and possible functions In B T Fuchs A FV Buttner amp D Zee (Eds) Contemporary ocularmotor and vestibular research A tribute to David ARobinson (pp 480ndash487) Stuttgart Germany ThiemePublishing
Williams P E amp Shapley R M (2007) A dynamicnonlinearity and spatial phase specificity in macaque
V1 neurons Journal of Neuroscience 27 5706ndash5718[PubMed] [Article]
Winterson B J amp Robinson D A (1975) Fixationby the alert but solitary cat Vision Research 151349ndash1352 [PubMed]
Yarbus A L (1967) Eye movements and vision(B Haigh Trans) New York Plenum Press
Zuber B L amp Stark L (1965) Microsaccades and thevelocityltamplitude relationship for saccadic eyemovements Science 150 1459ndash1460 [PubMed]
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 16

counts Olveczky Baccus and Meister (2003) recordedfrom ganglion cells in the rabbit and salamander retinaswhile presenting visual stimuli that were jittered tosimulate fixational eye movements (based on parametersfrom previously published studies by Manteuffel PlasaSommer amp Wess 1977 and Van der Steen amp Collewijn1984) In both species fixational eye movements helpedsegregate a moving foreground from a complex back-ground in a population of ganglion cells
Fixational eye movements inbirds
Fixational eye movements in the pigeon
There are some remarkable differences between fixa-tional eye movements in birds and mammals Nye (1969)identified four types of fixational eye movements in thepigeon (Columba livia) flicks (ie microsaccades)impulse movements drift and oscillations No tremorwas detected Microsaccade frequency was very lowrelative to humans Impulse movements occurred morefrequently usually between bursts of oscillations withamplitudes ranging up to 2 degrees Drift velocitiesranged from 1 to 5 degrees per second Both micro-saccades and drifts occurred in response to movingstimuli Oscillations occurred in short bursts at rates ofabout 30 Hz and could be quite large (up to severaldegrees of amplitude) Bloch Rivaud and Martinoya(1984) reported microsaccades drifts and oscillationsldquosimilar to those described by Nyerdquo but none of thesewere analyzed in detailCyclotorsional oscillations are an integral part of avian
saccades (Wallman Pettigrew amp Letelier 1994) Thusone might speculate that oscillations are associated withavian microsaccades as well especially if saccades andmicrosaccades share a common generator (Engbert 2006Otero-Millan et al 2008 Rolfs et al 2008 2006 Zuberamp Stark 1965) However this possibility has not yet beenaddressed in the literature The function of oscillations isunknown (Carpenter 1988) but one hypothesis is thatthey serve to facilitate the delivery of arterial blood to theocular capillary bed (Pettigrew Wallman amp Wildsoet1990 Wallman et al 1994)
Fixational eye movements in the owl
Steinbach and Money (1973) recorded fixational eyemovements in the great horned owl (Bubo virginianus)a species previously thought incapable of moving itseyes The owlrsquos fixational eye movements includeflicks or microsaccades drift tremor and oscillations
Microsaccades seemed correlated to visual events ofinterest to the owl they were 1 deg or less in amplitudeand had peak velocities of about 10 degsec Drifts wereslow motions of the eye in between fixations withamplitudes of 1 degree or less Tremor amplitude wasabout 1 arcmin with frequencies around 20 Hz High-frequency oscillations (25 Hz) of short duration (025 sec)were very similar to those previously found in the pigeon(Nye 1969) No studies to date have examined theperceptual or physiological effects of fixational eyemovements in any bird species
Fixational eye movements inreptiles
Fixational eye movements in the turtle
Greschner Bongard Rujan and Ammermuller (2002)identified two types of fixational eye movements in theturtle (Pseudemys scripta elegans) which they calledldquodrift-likerdquo and ldquoperiodicrdquo Drift-like motions were smalland slow (with velocities comparable to human drift) andwere superimposed by larger and faster periodic componentsPeriodic motion had retinal amplitudes on the order of thediameter of a photoreceptor which are comparable to thoseof human tremor On the other hand periodic motionrsquosfrequency (5 Hz) velocity and orderly relationship betweenvelocity and amplitude are closer to those of human micro-saccades (see Martinez-Conde et al 2004 for a review oftremor and microsaccade parameters in humans)Simulated periodic motion generated strong synchronous
firing in the turtlersquos retina whereas simulated drift-likemotion had little effect (Figure 3) These results agree withprevious physiological and modeling studies in the primatevisual system in which strong neural transients wereobserved in response to microsaccades (Donner amp Hemila2007 Martinez-Conde 2006 Martinez-Conde et al 20002002) Such neural transients may underlie the behavior ofcortical neurons as coincidence detectors (ShelleyMcLaughlin Shapley amp Wielaard 2002 Williams ampShapley 2007) Moreover neural transients to stimulionsets and terminations (similar to those produced bymicrosaccades in the primate visual system Martinez-Conde 2006 Martinez-Conde et al 2000 2002) havebeen related to target visibility in visual masking para-digms (Macknik amp Livingstone 1998 Macknik ampMartinez-Conde 2004 Macknik Martinez-Conde ampHaglund 2000)In the turtle retina periodic motion-driven neurons with
receptive fields located along contrast borders weremoreover synchronized and reliably indicated the preced-ing motions The authors proposed that this synchroniza-tion of retinal activity could be used by the brain to
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 7
improve the estimation of stimulus features such asspatial frequency (Greschner et al 2002) This findingcould have a parallel in the primate visual system Donnerand Hemila (2007) recently proposed that microsaccadessignificantly ldquore-sharpenrdquo the image and improve spatialresolution in primates Rucci Iovin Poletti and Santini(2007) also found that fixational eye movements enhancefine spatial detail in human subjects
Fixational eye movements in the chameleon
Fixational eye movements in the chameleon have notbeen studied in detail However Gioanni and colleaguesfound that the chameleon (Chamaleo chamaleo) makessaccades during fixation The authors speculated that theseprobably correspond to fixational microsaccades found inother foveate species (Gioanni Bennis amp Sansonetti1993)
Fixational eye movements inamphibians
Fixational (and more generally spontaneous) eye move-ments vary widely among amphibians The frog (anafoveate amphibian) shows a general lack of eye andhead movements except for those that stabilize the retinalimage Dieringer and colleagues (Dieringer amp Daunicht1986 Dieringer amp Precht 1982 Dieringer Precht ampBlight 1982) found that the grass frog (Rana temporaria)produces no spontaneous eye movements in the absenceof head movements with the possible exception of tremor(similarly to the rabbit another afoveate species Van derSteen amp Collewijn 1984) It is likely that the frogrsquos lack ofeye movements leads to fast visual fading of all stationaryobjects in its field of view (Ditchburn amp Ginsborg 1952Yarbus 1967) Lettvin Maturana MsCulloch and Pitts
Figure 3 Fixational eye movements increase retinal activity in the turtle (A) Continuous recording from 54 retinal ganglion cells in theturtle during darkness after switching on a stationary grating and during wobbling of this grating The grating was wobbled to simulateperiodic (ie tremor-like) eye movements (B) Responses of a single ganglion cell to a drifting contrast border of velocity comparable toocular drifts (C) When the drift is superimposed with wobbling the responses of the cell shown in (B) increase dramatically FromGreschner et al (2002) and modified by Martinez-Conde et al (2004)
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 8
(1968) stated that a frog ldquowill starve to death surroundedby food if it is not movingrdquo (but see Dieringer amp Daunicht1986 for a counterpoint) Small head motions or eyemovements correlated with respiratory motions maydiminish fading to some extent (Dieringer amp Precht1982 Schipperheyn 1963 1965 1967)As a side note the frogrsquos respiratory eye movements
play no role in the respiratory function ldquoin the course ofeach respiratory cycle increase of pressure in the frogrsquosbuccal cavity protrudes the eye and causes the opticalaxis to deviate upwards over 25ndash35 minutes of arcrdquo(Schipperheyn 1963) That is respiratory eye movementsare merely synchronized with respirationAutrum (1959) found no evidence of respiratory eye
movements in the frog Ewert andBorchers (1974) observedthat stationary stimuli elicit no responses from visualneurons in the common toad (Bufo Bufo L) regardless ofrespiratory pressure changes in the buccal cavity
Fixational eye movements in the salamander
Manteuffel et al (1977) identified two types of fixa-tional eye movements in salamanders tremor-like andrespiratory eye movements Tremor-like is a rapid move-ment of small amplitude Torsional eye movements resultfrom the animalrsquos breathing (also found in the frog bySchipperheyn 1963) Additionally flicks are occasionallyobserved which rapidly change the position of gaze It isunclear whether such flicks may generally correspond towhat we now call microsaccades andor to voluntarysaccades Based on these observations the authors pro-posed that unlike frogs and toads (Autrum 1959 Ewert ampBorchers 1974 Lettvin et al 1968) salamanders cancontinuously perceive stationary objects (Manteuffel et al1977) Fixational eye movements in the salamander (Tigersalamander Ambystoma tigrinum) may also help segre-gate object and background motion in the retina(Olveczky et al 2003 see also Fixational eye movementsin the rabbit section) and even work to emphasize novelvisual features (Olveczky et al 2007 see Baccus et al2008 for details on the retinal neurons and synapses thatmay underlie the computation of differential motionbetween an object and its background)
Fixational eye movements in fish
Spontaneous eye movements in the goldfish
Eye movements during fixation have been studied inboth afoveate (goldfish Carassius auratus) and foveate(archer fish Toxotes chatareus) fish species The goldfisha long-standing model organism in vision and oculomotor
research (Aksay et al 2003 2007 Cabrera Torres PasaroPastor amp Delgado-Garcia 1992 Pastor Torres Delgado-Garcia amp Baker 1991 Salas Navarro Torres ampDelgado-Garcia 1992 Torres Pastor Cabrera Salas ampDelgado-Garcia 1992) produces three types of sponta-neous eye movements saccades stretch and drift (Easter1971 Mensh Aksay Lee Seung amp Tank 2004) Weshould note that in the motionless goldfish eye move-ments are very infrequent Those that do occur consistmainly of saccades slow drifts are very rare (Easter Johnsamp Heckenlively 1974) During saccades (identified bythresholding the eye velocity at 5 degsec) both eyes moverapidly simultaneously and usually in the same direction(Mensh et al 2004) Stretches occur every 2ndash3 min and arecharacterized by a brief simultaneous large excursion ofboth eyes to extreme temporal positions (Easter 1971Mensh et al 2004) The function of stretches is not known(Mensh et al 2004) Because fixations were defined asthose periods that were free of saccadic and stretchmovements (Easter 1971) only drifts (from the threetypes of spontaneous movements) can technically beconsidered ldquofixational eye movementsrdquo However thegoldfish saccadic main sequence includes saccades withequivalent amplitudes and velocities to those found inhuman microsaccades (Mensh et al 2004 Zuber amp Stark1965) Indeed microsaccades have been identified in other(foveate) fish species although they are rarer than inhumans (Segev et al 2007) Small saccades in thegoldfish tend to be divergent whereas large saccadestend to be convergent (Mensh et al 2004) Drift occurssimultaneously in both eyes usually in the samedirection Drift velocities are almost exclusively e1 degsec(Mensh et al 2004) No tremor has been identified in thegoldfish
Spontaneous eye movements in teleost fish
Spontaneous eye movements have been studied in theNew Zealand parore (Girella tricuspidata MontgomeryMcVean amp McCarthy 1983) The pattern of eye move-ments was similar to that previously described in goldfish(Easter 1971 Hermann amp Constantine 1971)
Fixational eye movements in the archer fish
A recent study in the archer fish (Segev et al 2007) hasprovided the first perceptual and physiological correlatesof fixational eye movements in any fish species Archerfish have remarkable visual abilities they shoot downinsects resting on foliageVas well as stationary targetscovering a single retinal photoreceptorVwith squirts ofwater from their mouths they predict the location atwhich the insect will splash down in G100 msec and theydeduce the absolute size of an object presented at differentviewing distances (Supplementary Videos 2 and 3) Segev
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 9
et al (2007) identified three types of fixational eyemovements in the archer fish microsaccades drift andtremor Microsaccades were rare and were not analyzed indetail Drift velocities were e040 degsec Tremor was ahigh-frequency oscillatory movement (the primary com-ponent was 5 Hz) with peak-to-peak amplitude of 01ndash02 deg (corresponding to an image displacement of 1ndash2photoreceptors) and velocity of about 1 degsec (Figure 4)Whereas the amplitude and velocity of tremor in thearcher fish is comparable to those in humans its frequencyis much lower (5 Hz vs 940 Hz) Interestingly archer fishtremor is very similar both in amplitude (photoreceptorwidths covered) as well as in frequency (5 Hz) to thetremor-like periodic motion in the turtle (discussedearlier Greschner et al 2002)Segev et al (2007) trained archer fish to perform a size
discrimination task the fish had to distinguish a medium-sized target from larger and smaller distracters (theyindicated their choice by shooting a jet of water) To relatethis behavior to responses from retinal neurons theauthors recorded from populations of retinal ganglioncells while presenting objects of different sizes moved tosimulate saccades drifts and tremor Drifts and tremorelicited weak neuronal responses that were not veryinformative about the spatial structure of the stimulus(although they did provide some information about target
size in agreement with the results of Greschner et al2002 in the turtle) Moreover performance was similar fortremor alone and for tremor plus drift indicating that driftplays a negligible role in revealing spatial structure (alsoin agreement with Greschner et al 2002)When present the firing rate was phase-locked to
tremor Saccades were more effective than drifts or tremorin driving informative responses from the ganglion cells(consistent with theoretical and physiological studies ofmicrosaccade-driven neuronal activity in the primateretina and LGN Donner amp Hemila 2007 Martinez-Conde et al 2002) Segev et al (2007) proposed that inprimates both saccades and microsaccades (which arerare in the archer fish) may contribute to size discrim-ination tasks as well as enhance spatial detail whereasdrifts and tremor by themselves may not be sufficient toimprove visual perception However it is important toremember that central vision in humans can be maintainedby tremor andor drift even when microsaccades aresuppressed during strict fixation (Martinez-Conde 2006Martinez-Conde et al 2004 2006) On the other handone cannot rule out the possibility that if one couldcompletely eliminate drifts and tremor microsaccadesmight then suffice to sustain both central and peripheralvision in primates and humans Future experiments areneeded to further explore these ideas
Figure 4 Archer fish eye movements (A) Example trace of eye position as a function of time as measured by search coil (B) Example ofa single 15-s fixation superimposed on a grid representing the photoreceptor mosaic The back and forth movement is caused by tremor(C) Distribution of fixation durations (taken as time interval between saccades) (D) Power spectrum of horizontal fixational eyemovements (E) Distribution of tremor amplitude (data taken from 1 animal in a 15-min session) Modified from Segev et al (2007)
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 10
Discussion
The vast majority of eye movement studies conductedin non-human species to date have not analyzed eyeposition data in the range of fixational eye movementsThus eye movements during fixation have been reportedin only a handful of vertebrates Most of these studieshave concentrated solely on the physical parameters ofthe fixational eye movements without addressing theirphysiological and perceptual consequences Fortunatelythis trend has started to reverse in recent years with agrowing list of papers addressing the effects of non-human fixational eye movements in physiology andperception This is an important development A largeamount of all psychophysical and physiological visualresearch has been conducted during conditions of visualfixation Therefore understanding the precise physiolog-ical and perceptual contributions of fixational eye move-ments may be critical to the interpretation of previous andfuture vision researchThere are some interesting differences and similarities
in the types of fixational eye movements produced bydifferent species For instance ocular tremor is possiblyubiquitous to all vertebrates The fact that tremor has notbeen observed in all species studied could be due to thetechnical difficulty of recording tremor accurately (ietremor amplitudes and frequencies are usually in the rangeof the recording systemrsquos noise) Drift has also beenrecorded in most vertebrates and it may be as omnipre-sent as tremor Conversely microsaccades seem mostimportant in species with foveal vision (ie primates andhumans) whereas they are scarce or absent in afoveatespecies (ie rabbits frogs goldfish) A link betweenmicrosaccades and foveal vision may reflect the two-pronged functional significance of microsaccades inhumans and primates first microsaccades may benecessary to correct the slow drift of the eye and bringback the objectregion of interest to the retinal area wherephotoreceptor density is highest (Cornsweet 1956) Thecorrective role of human microsaccades has been demon-strated in recent studies (Engbert amp Kliegl 2004 Engbertamp Mergenthaler 2006) Second the dynamics of micro-saccades themselves may improve spatial resolution(Donner amp Hemila 2005)In reviewing the patterns of fixational eye movements
across vertebrates it is tempting to draw parallels withother sensory modalities Fixational eye movementsrefresh the retinal images during fixation and thus preventneural adaptation and visual fading But this mechanism isnot circumscribed to the visual system Sniffs in rodentolfaction discretely sample sensory information every200ndash300 msec and thus are similar in their temporaldynamics to primate saccades (Otero-Millan et al 2008Uchida Kepecs amp Mainen 2006) and microsaccades(Otero-Millan et al 2008) A similar mode of discretesampling may also be at play when objects are recognized
through tactile information for instance if we sweep ourfingertips over an objectrsquos surface with our eyes closed orwhen blind individuals read Braille scriptOne intriguing possibility that merits further experi-
ments and analyses is the potential correlation between thespatiotemporal parameters of fixational eye movements(such as those summarized in Table 1) and the adaptationproperties of visual neurons across species For instancethe greater ocular stability of some species (rabbitgoldfish frog etc) versus others (primates) could beassociated with some particular adaptive properties oftheir visual cells Thus future comparative studies shouldpay attention to the response properties of visual cellstriggered by fixational eye movements in the presence ofstationary stimuli Other potential factors underlying thevariation in fixational eye movements across species couldbe differences in oculomotor control at the level of the eyemuscles or perhaps differences in retinal organizationFuture research will further improve our understanding
of fixational eye movements and the neural activity thatmaintains the visibility of stationary visual scenes inhuman and non-human species One interesting questionthat remains to be addressed concerns the role of tremor invision and visual physiology Because tremor frequenciesare well over the flicker fusion frequencies in humans ithas been argued that tremor of the visual image would beineffective as a stimulus (Ditchburn 1955 Gerrits ampVendrik 1970 Sharpe 1972) However recent studieshave found simulated tremor to enhance visual processingand improve visual acuity in a model retina (Hennig et al2002 Miller et al 2006) Follow-up research should beaimed to replicate these findings in actual neurons withcorrelated perceptual reports One should also note thatearly visual neurons can follow high-frequency flickeringthat surpasses the perceptual threshold for flicker fusion(Martinez-Conde et al 2002) Thus high-frequencytremor could be adequate to maintain activity in the earlyvisual system which then may lead to visual perception inhigher visual areas Another significant question thatremains unanswered concerns the oculomotor bases ofthe various types of fixational eye movements acrossspecies To date there is too little information on thisimportant topic to draw a conclusion
Conclusions
We have reviewed the parameters of fixational eyemovements in non-human mammals birds reptilesamphibians and fish as well as their consequences forphysiology and perception Fixational eye movementsand microsaccades in particular appear to be mostimportant in foveate vs afoveate species Animals lackingfixational eye movements such as afoveate frogs andtoads may be incapable of seeing stationary objects
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 11
during visual fixation Future research should furtherexplore the perceptual effects of fixational eye movementsand determine the oculomotor mechanisms responsible fortheir generation across species
Acknowledgments
We thank Mona Stewart and Isabel Gomez-Caraballo fortechnical assistance Jorge Otero-Millan for programmingassistance and Dr Xoana G Troncoso for comments on themanuscript This study was funded through grants fromthe Barrow Neurological Foundation (to SMC and SLM)the National Science Foundation (Award 0643306 toSMC) the Arizona Biomedical Research Commission(Award 07-102 to SMC) and Science Foundation Arizona(SLM)
Commercial relationships noneCorresponding author Susana Martinez-Conde PhDEmail smartneuralcorrelatecomAddress Barrow Neurological Institute 350 W ThomasRd Phoenix AZ 85013 USA
References
Aksay E Major G Goldman M S Baker R SeungH S amp Tank D W (2003) History dependence ofrate covariation between neurons during persistentactivity in an oculomotor integrator Cerebral Cortex13 1173ndash1184 [PubMed] [Article]
Aksay E Olasagasti I Mensh B D Baker RGoldman M S amp Tank D W (2007) Functionaldissection of circuitry in a neural integrator NatureNeuroscience 10 494ndash504 [PubMed]
Autrum H (1959) Das Fehlen unwillkurlicher Augenbe-wegungen beim Frosch Naturwissenschaften 46435
Baccus S A Olveczky B P Manu M amp Meister M(2008) A retinal circuit that computes object motionJournal of Neuroscience 28 6807ndash6817 [PubMed]
Bair W amp OrsquoKeefe L P (1998) The influence offixational eye movements on the response of neuronsin area MT of the macaque Visual Neuroscience 15779ndash786 [PubMed]
Betta E amp Turatto M (2006) Are you ready I can tellby looking at your microsaccades Neuroreport 171001ndash1004 [PubMed]
Bloch S Rivaud S amp Martinoya C (1984) Compar-ing frontal and lateral viewing in the pigeon IIIDifferent patterns of eye movements for binocularand monocular fixation Behavioural Brain Research13 173ndash182 [PubMed]
Cabrera B Torres B Pasaro R Pastor A M ampDelgado-Garcia J M (1992) A morphological studyof abducens nucleus motoneurons and internuclearneurons in the goldfish (Carassius auratus) BrainResearch Bulletin 28 137ndash144 [PubMed]
Carpenter R H S (1988) Movements of the eyes(2nd ed) Pion London
Collewijn H (1977) Eye- and head movements infreely moving rabbits The Journal of Physiology266 471ndash498 [PubMed] [Article]
Collewijn H amp van der Mark F (1972) Ocular stabilityin variable visual feedback conditions in the rabbitBrain Research 36 47ndash57 [PubMed]
Conway J L Timberlake G T amp Skavenski A A(1981) Oculomotor changes in cats reared withoutexperiencing continuous retinal image motion Exper-imental Brain Research 43 229ndash232 [PubMed]
Cornsweet T N (1956) Determination of the stimuli forinvoluntary drifts and saccadic eye movements Jour-nal of the Optical Society of America 46 987ndash993[PubMed]
Delgado-Garcia J M del Pozo F amp Baker R (1986)Behavior of neurons in the abducens nucleus of thealert catVII Internuclear neurons Neuroscience 17953ndash973 [PubMed]
Dieringer N amp Daunicht W J (1986) Image fadingVAproblem for frogs Naturwissenschaften 73 330ndash332[PubMed]
Dieringer N amp Precht W (1982) Compensatory headand eye movements in the frog and their contributionto stabilization of gaze Experimental BrainResearch 47 394ndash406 [PubMed]
Dieringer N Precht W amp Blight A R (1982)Resetting fast phases of head and eye and theirlinkage in the frog Experimental Brain Research 47407ndash416 [PubMed]
Ditchburn R W (1955) Eye-movements in relation toretinal action Optica Acta 1 171ndash176
Ditchburn R W amp Ginsborg B L (1952) Vision with astabilized retinal image Nature 170 36ndash37[PubMed]
Ditchburn R W amp Ginsborg B L (1953) Involuntaryeye movements during fixation The Journal ofPhysiology 119 1ndash17 [PubMed] [Article]
Donner K amp Hemila S (2007) Modelling the effect ofmicrosaccades on retinal responses to stationarycontrast patterns Vision Research 47 1166ndash1177[PubMed]
Donner K K amp Hemila S (2005) The possible purposeof microsaccades Paper presented at the EuropeanConference on Visual Perception A Coruna Spain
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 12
Easter S S Jr (1971) Spontaneous eye movements inrestrained goldfish Vision Research 11 333ndash342[PubMed]
Easter S S Jr Johns P R amp Heckenlively D R(1974) Horizontal compensatory eye movements ingoldfish (Carassius auratus) I The normal animalJournal of Comparative Physiology 92 23ndash35
Engbert R (2006) Microsaccades A microcosm forresearch on oculomotor control attention and visualperception Progress Brain Research 154 177ndash192[PubMed]
Engbert R amp Kliegl R (2003a) Binocular coordinationin microsaccades In R R J Hyona amp H Deubel(Eds) The mindrsquos eyes Cognitive and appliedaspects of eye movements (pp 103ndash117) OxfordUK Elsevier
Engbert R amp Kliegl R (2003b) Microsaccades uncoverthe orientation of covert attention Vision Research43 1035ndash1045 [PubMed]
Engbert R amp Kliegl R (2004) Microsaccades keep theeyesrsquo balance during fixation Psychological Science15 431ndash436 [PubMed]
Engbert R amp Mergenthaler K (2006) Microsaccadesare triggered by low retinal image slip Proceedingsof the National Academy of Sciences of the UnitedStates of America 103 7192ndash7197 [PubMed][Article]
Ewert J P amp Borchers H W (1974) Antwort vonretinalen Ganglienzellen bei freibeweglichen Kroten(Bufo Bufo L) Journal of Comparative and Physi-ology 92 117ndash130
Fuller J H (1980) Linkage of eye and head movementsin the alert rabbit Brain Research 194 219ndash222[PubMed]
Fuller J H (1981) Eye and head movements duringvestibular stimulation in the alert rabbit BrainResearch 205 363ndash381 [PubMed]
Galfano G Betta E amp Turatto M (2004) Inhibition ofreturn in microsaccades Experimental BrainResearch 159 400ndash404 [PubMed]
Gerrits H J amp Vendrik A J (1970) Artificial move-ments of a stabilized image Vision Research 101443ndash1456 [PubMed]
Gioanni H Bennis M amp Sansonetti A (1993) Visualand vestibular reflexes that stabilize gaze in thechameleon Visual Neuroscience 10 947ndash956[PubMed]
Goffart L Quinet J Chavane F amp Masson G S(2006) Influence of background illumination onfixation and visually guided saccades in the rhesusmonkey Vision Research 46 149ndash162 [PubMed]
Greschner M Bongard M Rujan P amp Ammermuller J(2002) Retinal ganglion cell synchronization by fixa-tional eye movements improves feature estimationNature Neuroscience 5 341ndash347 [PubMed]
Gur M Beylin A amp Snodderly D M (1997)Response variability of neurons in primary visualcortex (V1) of alert monkeys Journal of Neuro-science 17 2914ndash2920 [PubMed] [Article]
Hafed Z M amp Clark J J (2002) Microsaccades as anovert measure of covert attention shifts VisionResearch 42 2533ndash2545 [PubMed]
Hebbard F W amp Marg E (1960) Physiologicalnystagmus in the cat Journal of the Optical Societyof America 50 151ndash155 [PubMed]
Hennig M A Kerscher N J Funke K amp Worgotter F(2002) Stochastic resonance in visual cortical neuronsDoes the eye-tremor actually improve visual acuityNeurocomputing 44ndash46 115ndash120
Hennig M H amp Worgotter F (2004) Eye micro-movements improve stimulus detection beyond theNyquist limit in the peripheral retina In Advances inNeural Information Processing Systems (NIPS2003)(vol 16) Denver CO MIT Press
Hennig M H amp Worgotter F (2007) Effects offixational eye movements on retinal ganglion cellresponses A modelling study Frontiers in Computa-tional Neuroscience 1 2 [PubMed] [Article]
Hermann H T amp Constantine M M (1971) Eyemovements in the goldfish Vision Research 11313ndash331 [PubMed]
Horowitz T S Fine E M Fencsik D E Yurgenson Samp Wolfe J M (2007) Fixational eye movements arenot an index of covert attention PsychologicalScience 18 356ndash363 [PubMed]
Horwitz G D amp Albright T D (2003) Short-latencyfixational saccades induced by luminance incrementsJournal of Neurophysiology 90 1333ndash1339[PubMed] [Article]
Laubrock J Engbert R Rolfs M amp Kliegl R(2007) Microsaccades are an index of covertattention Commentary on Horowitz Fine FencsikYurgenson and Wolfe (2007) Psychological Sci-ence 18 364ndash366 discussion 367ndash368 [PubMed]
Leopold D A amp Logothetis N K (1998) Micro-saccades differentially modulate neural activity in thestriate and extrastriate visual cortex ExperimentalBrain Research 123 341ndash345 [PubMed]
Lettvin J Y Maturana H R MsCulloch W S amp PittsW H (1968) What the frogrsquos eye tells the frogrsquosbrain In W C C a M Balaban (Ed) The mind
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 13
Biological approaches to its functions (pp 233ndash258)John Wiley and Sons London
Macknik S L amp Livingstone M S (1998) Neuronalcorrelates of visibility and invisibility in the primatevisual system Nature Neuroscience 1 144ndash149[PubMed]
Macknik S L amp Martinez-Conde S (2004) The spatialand temporal effects of lateral inhibitory networksand their relevance to the visibility of spatiotemporaledges Neurocomputing 58ndash60 775ndash782
Macknik S L Martinez-Conde S amp Haglund M M(2000) The role of spatiotemporal edges in visibilityand visual masking Proceedings of the NationalAcademy of Sciences of the United States of America97 7556ndash7560 [PubMed] [Article]
Manteuffel G Plasa L Sommer T J amp Wess O(1977) Involuntary eye movements in salamandersNaturwissenschaften 64 533ndash534 [PubMed]
Marchetti E Gauthier G M amp Pellet J (1983)Cerebellar control of eye movements studied withinjection of harmaline in the trained baboon ArchivesItaliennes de Biologies 121 1ndash17 [PubMed]
Martinez-Conde S (2006) Fixational eye movements innormal and pathological vision Progress in BrainResearch 154 151ndash176 [PubMed]
Martinez-Conde S amp Macknik S L (2007a) Bobbingand perceiving Scientific American 297 16
Martinez-Conde S amp Macknik S L (2007b) Windowson the mind Scientific American 297 56ndash63[PubMed]
Martinez-Conde S Macknik S L amp Hubel D H(2000) Microsaccadic eye movements and firing ofsingle cells in the striate cortex of macaque monkeysNature Neuroscience 3 251ndash258 [PubMed]
Martinez-Conde S Macknik S L amp Hubel D H(2002) The function of bursts of spikes during visualfixation in the awake primate lateral geniculate nucleusand primary visual cortex Proceedings of theNational Academy of Sciences of the United Statesof America 99 13920ndash13925 [PubMed] [Article]
Martinez-Conde S Macknik S L amp Hubel D H(2004) The role of fixational eye movements invisual perception Nature Reviews Neuroscience 5229ndash240 [PubMed]
Martinez-Conde S Macknik S L Troncoso X G ampDyar T A (2006) Microsaccades counteract visualfading during fixation Neuron 49 297ndash305[PubMed] [Article]
Mensh B D Aksay E Lee D D Seung H S ampTank D W (2004) Spontaneous eye movements ingoldfish Oculomotor integrator performance plasti-
city and dependence on visual feedback VisionResearch 44 711ndash726 [PubMed]
Miller J A Denning K S George J S MarshakD W amp Kenyon G T (2006) A high frequencyresonance in the responses of retinal ganglion cells torapidly modulated stimuli A computer model VisualNeuroscience 23 779ndash794 [PubMed]
Montgomery J C McVean A R amp McCarthy D(1983) The effects of lowered temperature onspontaneous eye movements in a teleost fish Com-parative Biochemistry and Physiology A Compara-tive Physiology 75 363ndash368 [PubMed]
Motter B C amp Poggio G F (1984) Binocularfixation in the rhesus monkey Spatial and temporalcharacteristics Experimental Brain Research 54304ndash314 [PubMed]
Murakami I amp Cavanagh P (1998) A jitter after-effectreveals motion-based stabilization of vision Nature395 798ndash801 [PubMed]
Nye P W (1969) The monocular eye movements of thepigeon Vision Research 9 133ndash144 [PubMed]
Olveczky B P Baccus S A amp Meister M (2003)Segregation of object and background motion in theretina Nature 423 401ndash408 [PubMed]
Olveczky B P Baccus S A amp Meister M (2007)Retinal adaptation to object motion Neuron 56689ndash700 [PubMed] [Article]
Otero-Millan J Troncoso X Macknik S L Serrano-Pedraza I amp Martinez-Conde S (2008) Saccadesand microsaccades during visual fixation explorationand search Foundations for a common saccadicgenerator Journal of Vision 8(14)21 1ndash18 httpjournalofvisionorg81421 doi10116781421[PubMed] [Article]
Pastor A M Torres B Delgado-Garcia J M ampBaker R (1991) Discharge characteristics of medialrectus and abducens motoneurons in the goldfishJournal of Neurophysiology 66 2125ndash2140 [PubMed]
Pettigrew J D Wallman J amp Wildsoet C F (1990)Saccadic oscillations facilitate ocular perfusion fromthe avian pecten Nature 343 362ndash363 [PubMed]
Pritchard R M amp Heron W (1960) Small eye move-ments of the cat Canadian Journal of Psychology14 131ndash137 [PubMed]
Ratliff F amp Riggs L A (1950) Involuntary motions ofthe eye during monocular fixation Journal of Exper-imental Psychology 40 687ndash701 [PubMed]
Reppas J B UsreyWM amp Reid R C (2002) Saccadiceye movements modulate visual responses in thelateral geniculate nucleus Neuron 35 961ndash974[PubMed] [Article]
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 14
Riggs L A amp Ratliff F (1952) The effects of counter-acting the normal movements of the eye Journal ofthe Optical Society of America 42 872ndash873
Rolfs M Kliegl R amp Engbert R (2008) Toward amodel of microsaccade generation The case ofmicrosaccadic inhibition Journal of Vision 8(11)51ndash23 httpjournalofvisionorg8115 doi1011678115 [PubMed] [Article]
Rolfs M Laubrock J amp Kliegl R (2006) Shorteningand prolongation of saccade latencies followingmicrosaccades Experimental Brain Research 169369ndash376 [PubMed]
Rucci M Iovin R Poletti M amp Santini F (2007)Miniature eye movements enhance fine spatial detailNature 447 851ndash854 [PubMed]
Salas C Navarro F Torres B amp Delgado-Garcia J M(1992) Effects of diazepam and D-amphetamine onrhythmic pattern of eye movements in goldfishNeuroreport 3 131ndash134 [PubMed]
Schipperheyn J J (1963) Respiratory eye movementsand perception of stationary objects in the frog ActaPhysiologica et Pharmacologica Neerlandica 12157ndash159 [PubMed]
Schipperheyn J J (1965) Contrast detection in frogrsquosretina Acta Physiologica et Pharmacologica Neer-landica 13 231ndash277 [PubMed]
Schipperheyn J J (1967) Influence of eye movements onretinal ganglion cell activity in frogs Acta Physi-ologica et Pharmacologica Neerlandica 14 506[PubMed]
Segev R Schneidman E Goodhouse J amp Berry M J2nd (2007) Role of eye movements in the retinalcode for a size discrimination task Journal ofNeurophysiology 98 1380ndash1391 [PubMed] [Article]
Sharpe C R (1972) The visibility and fading of thinlines visualized by their controlled movement acrossthe retina The Journal of Physiology 222 113ndash134[PubMed] [Article]
Shelley M McLaughlin D Shapley R amp Wielaard J(2002) States of high conductance in a large-scalemodel of the visual cortex Journal of ComputationalNeuroscience 13 93ndash109 [PubMed]
Skavenski A A Robinson D A Steinman R M ampTimberlake G T (1975) Miniature eye movementsof fixation in rhesus monkey Vision Research 151269ndash1273 [PubMed]
Snodderly D M (1987) Effects of light and darkenvironments on macaque and human fixational eyemovements Vision Research 27 401ndash415 [PubMed]
Snodderly D M Kagan I amp Gur M (2001) Selectiveactivation of visual cortex neurons by fixational eyemovements Implications for neural coding VisualNeuroscience 18 259ndash277 [PubMed]
Snodderly D M amp Kurtz D (1985) Eye position duringfixation tasks Comparison of macaque and humanVision Research 25 83ndash98 [PubMed]
Steinbach M J amp Money K E (1973) Eye movementsof the owl Vision Research 13 889ndash891 [PubMed]
Steinman R M Haddad G M Skavenski A A ampWyman D (1973) Miniature eye movement Science181 810ndash819 [PubMed]
Torres B Pastor A M Cabrera B Salas C ampDelgado-Garcia J M (1992) Afferents to theoculomotor nucleus in the goldfish (Carassius aur-atus) as revealed by retrograde labeling with horse-radish peroxidase Journal of Comparative Neurology324 449ndash461 [PubMed]
Troncoso X G Macknik S L amp Martinez-Conde S(2008) Microsaccades counteract perceptual filling-inJournal of Vision 8(14)15 1ndash9 httpjournalofvisionorg81415 doi10116781415 [Article]
Troncoso X G Macknik S L Otero-Millan J ampMartinez-Conde S (2008) Microsaccades driveillusory motion in the Enigma illusion Proceedingsof the National Academy of Sciences of the UnitedStates of America 105 16033ndash16038 [PubMed]
Tse P U Sheinberg D S amp Logothetis N K (2004)The distribution of microsaccade directions need notreveal the location of attention Psychological Science15 708ndash710
Turatto M Valsecchi M Tame L amp Betta E (2007)Microsaccades distinguish between global and localvisual processing Neuroreport 18 1015ndash1018[PubMed]
Uchida N Kepecs A amp Mainen Z F (2006) Seeing ata glance smelling in a whiff Rapid forms ofperceptual decision making Nature Reviews Neuro-science 7 485ndash491 [PubMed]
Valsecchi M Betta E amp Turatto M (2007) Visualoddballs induce prolonged microsaccadic inhibitionExperimental Brain Research 177 196ndash208[PubMed]
van Dam L C amp van Ee R (2006) Retinal image shiftsbut not eye movements per se cause alternations inawareness during binocular rivalry Journal of Vision6(11)3 1172ndash1179 httpjournalofvisionorg6113doi1011676113 [PubMed] [Article]
Van der Steen J amp Collewijn H (1984) Ocular stabilityin the horizontal frontal and sagittal planes in therabbit Experimental Brain Research 56 263ndash274[PubMed]
Van Gisbergen J A Robinson D A amp Gielen S(1981) A quantitative analysis of generation ofsaccadic eye movements by burst neurons Journalof Neurophysiology 45 417ndash442 [PubMed]
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 15
Van Gisbergen J A M amp Robinson D A (1977)Generation of micro and macrosaccades by burstneurons in the monkey In R Baker amp A Berthoz(Eds) Control of gaze by brain stem neurons (vol 1pp 301ndash308) New York ElsevierNorth-Holland
Wallman J Pettigrew J D amp Letelier J C (1994)The oscillatory saccades of birds Motoneural activ-ity and possible functions In B T Fuchs A FV Buttner amp D Zee (Eds) Contemporary ocularmotor and vestibular research A tribute to David ARobinson (pp 480ndash487) Stuttgart Germany ThiemePublishing
Williams P E amp Shapley R M (2007) A dynamicnonlinearity and spatial phase specificity in macaque
V1 neurons Journal of Neuroscience 27 5706ndash5718[PubMed] [Article]
Winterson B J amp Robinson D A (1975) Fixationby the alert but solitary cat Vision Research 151349ndash1352 [PubMed]
Yarbus A L (1967) Eye movements and vision(B Haigh Trans) New York Plenum Press
Zuber B L amp Stark L (1965) Microsaccades and thevelocityltamplitude relationship for saccadic eyemovements Science 150 1459ndash1460 [PubMed]
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 16
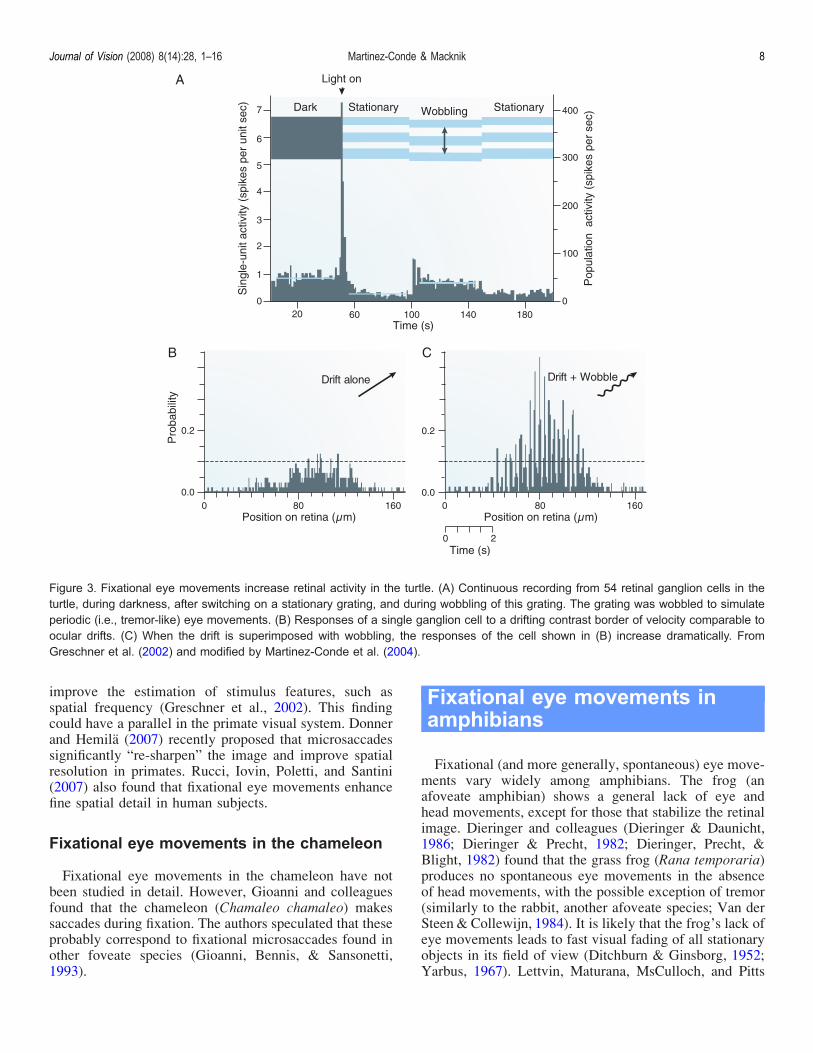
improve the estimation of stimulus features such asspatial frequency (Greschner et al 2002) This findingcould have a parallel in the primate visual system Donnerand Hemila (2007) recently proposed that microsaccadessignificantly ldquore-sharpenrdquo the image and improve spatialresolution in primates Rucci Iovin Poletti and Santini(2007) also found that fixational eye movements enhancefine spatial detail in human subjects
Fixational eye movements in the chameleon
Fixational eye movements in the chameleon have notbeen studied in detail However Gioanni and colleaguesfound that the chameleon (Chamaleo chamaleo) makessaccades during fixation The authors speculated that theseprobably correspond to fixational microsaccades found inother foveate species (Gioanni Bennis amp Sansonetti1993)
Fixational eye movements inamphibians
Fixational (and more generally spontaneous) eye move-ments vary widely among amphibians The frog (anafoveate amphibian) shows a general lack of eye andhead movements except for those that stabilize the retinalimage Dieringer and colleagues (Dieringer amp Daunicht1986 Dieringer amp Precht 1982 Dieringer Precht ampBlight 1982) found that the grass frog (Rana temporaria)produces no spontaneous eye movements in the absenceof head movements with the possible exception of tremor(similarly to the rabbit another afoveate species Van derSteen amp Collewijn 1984) It is likely that the frogrsquos lack ofeye movements leads to fast visual fading of all stationaryobjects in its field of view (Ditchburn amp Ginsborg 1952Yarbus 1967) Lettvin Maturana MsCulloch and Pitts
Figure 3 Fixational eye movements increase retinal activity in the turtle (A) Continuous recording from 54 retinal ganglion cells in theturtle during darkness after switching on a stationary grating and during wobbling of this grating The grating was wobbled to simulateperiodic (ie tremor-like) eye movements (B) Responses of a single ganglion cell to a drifting contrast border of velocity comparable toocular drifts (C) When the drift is superimposed with wobbling the responses of the cell shown in (B) increase dramatically FromGreschner et al (2002) and modified by Martinez-Conde et al (2004)
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 8
(1968) stated that a frog ldquowill starve to death surroundedby food if it is not movingrdquo (but see Dieringer amp Daunicht1986 for a counterpoint) Small head motions or eyemovements correlated with respiratory motions maydiminish fading to some extent (Dieringer amp Precht1982 Schipperheyn 1963 1965 1967)As a side note the frogrsquos respiratory eye movements
play no role in the respiratory function ldquoin the course ofeach respiratory cycle increase of pressure in the frogrsquosbuccal cavity protrudes the eye and causes the opticalaxis to deviate upwards over 25ndash35 minutes of arcrdquo(Schipperheyn 1963) That is respiratory eye movementsare merely synchronized with respirationAutrum (1959) found no evidence of respiratory eye
movements in the frog Ewert andBorchers (1974) observedthat stationary stimuli elicit no responses from visualneurons in the common toad (Bufo Bufo L) regardless ofrespiratory pressure changes in the buccal cavity
Fixational eye movements in the salamander
Manteuffel et al (1977) identified two types of fixa-tional eye movements in salamanders tremor-like andrespiratory eye movements Tremor-like is a rapid move-ment of small amplitude Torsional eye movements resultfrom the animalrsquos breathing (also found in the frog bySchipperheyn 1963) Additionally flicks are occasionallyobserved which rapidly change the position of gaze It isunclear whether such flicks may generally correspond towhat we now call microsaccades andor to voluntarysaccades Based on these observations the authors pro-posed that unlike frogs and toads (Autrum 1959 Ewert ampBorchers 1974 Lettvin et al 1968) salamanders cancontinuously perceive stationary objects (Manteuffel et al1977) Fixational eye movements in the salamander (Tigersalamander Ambystoma tigrinum) may also help segre-gate object and background motion in the retina(Olveczky et al 2003 see also Fixational eye movementsin the rabbit section) and even work to emphasize novelvisual features (Olveczky et al 2007 see Baccus et al2008 for details on the retinal neurons and synapses thatmay underlie the computation of differential motionbetween an object and its background)
Fixational eye movements in fish
Spontaneous eye movements in the goldfish
Eye movements during fixation have been studied inboth afoveate (goldfish Carassius auratus) and foveate(archer fish Toxotes chatareus) fish species The goldfisha long-standing model organism in vision and oculomotor
research (Aksay et al 2003 2007 Cabrera Torres PasaroPastor amp Delgado-Garcia 1992 Pastor Torres Delgado-Garcia amp Baker 1991 Salas Navarro Torres ampDelgado-Garcia 1992 Torres Pastor Cabrera Salas ampDelgado-Garcia 1992) produces three types of sponta-neous eye movements saccades stretch and drift (Easter1971 Mensh Aksay Lee Seung amp Tank 2004) Weshould note that in the motionless goldfish eye move-ments are very infrequent Those that do occur consistmainly of saccades slow drifts are very rare (Easter Johnsamp Heckenlively 1974) During saccades (identified bythresholding the eye velocity at 5 degsec) both eyes moverapidly simultaneously and usually in the same direction(Mensh et al 2004) Stretches occur every 2ndash3 min and arecharacterized by a brief simultaneous large excursion ofboth eyes to extreme temporal positions (Easter 1971Mensh et al 2004) The function of stretches is not known(Mensh et al 2004) Because fixations were defined asthose periods that were free of saccadic and stretchmovements (Easter 1971) only drifts (from the threetypes of spontaneous movements) can technically beconsidered ldquofixational eye movementsrdquo However thegoldfish saccadic main sequence includes saccades withequivalent amplitudes and velocities to those found inhuman microsaccades (Mensh et al 2004 Zuber amp Stark1965) Indeed microsaccades have been identified in other(foveate) fish species although they are rarer than inhumans (Segev et al 2007) Small saccades in thegoldfish tend to be divergent whereas large saccadestend to be convergent (Mensh et al 2004) Drift occurssimultaneously in both eyes usually in the samedirection Drift velocities are almost exclusively e1 degsec(Mensh et al 2004) No tremor has been identified in thegoldfish
Spontaneous eye movements in teleost fish
Spontaneous eye movements have been studied in theNew Zealand parore (Girella tricuspidata MontgomeryMcVean amp McCarthy 1983) The pattern of eye move-ments was similar to that previously described in goldfish(Easter 1971 Hermann amp Constantine 1971)
Fixational eye movements in the archer fish
A recent study in the archer fish (Segev et al 2007) hasprovided the first perceptual and physiological correlatesof fixational eye movements in any fish species Archerfish have remarkable visual abilities they shoot downinsects resting on foliageVas well as stationary targetscovering a single retinal photoreceptorVwith squirts ofwater from their mouths they predict the location atwhich the insect will splash down in G100 msec and theydeduce the absolute size of an object presented at differentviewing distances (Supplementary Videos 2 and 3) Segev
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 9
et al (2007) identified three types of fixational eyemovements in the archer fish microsaccades drift andtremor Microsaccades were rare and were not analyzed indetail Drift velocities were e040 degsec Tremor was ahigh-frequency oscillatory movement (the primary com-ponent was 5 Hz) with peak-to-peak amplitude of 01ndash02 deg (corresponding to an image displacement of 1ndash2photoreceptors) and velocity of about 1 degsec (Figure 4)Whereas the amplitude and velocity of tremor in thearcher fish is comparable to those in humans its frequencyis much lower (5 Hz vs 940 Hz) Interestingly archer fishtremor is very similar both in amplitude (photoreceptorwidths covered) as well as in frequency (5 Hz) to thetremor-like periodic motion in the turtle (discussedearlier Greschner et al 2002)Segev et al (2007) trained archer fish to perform a size
discrimination task the fish had to distinguish a medium-sized target from larger and smaller distracters (theyindicated their choice by shooting a jet of water) To relatethis behavior to responses from retinal neurons theauthors recorded from populations of retinal ganglioncells while presenting objects of different sizes moved tosimulate saccades drifts and tremor Drifts and tremorelicited weak neuronal responses that were not veryinformative about the spatial structure of the stimulus(although they did provide some information about target
size in agreement with the results of Greschner et al2002 in the turtle) Moreover performance was similar fortremor alone and for tremor plus drift indicating that driftplays a negligible role in revealing spatial structure (alsoin agreement with Greschner et al 2002)When present the firing rate was phase-locked to
tremor Saccades were more effective than drifts or tremorin driving informative responses from the ganglion cells(consistent with theoretical and physiological studies ofmicrosaccade-driven neuronal activity in the primateretina and LGN Donner amp Hemila 2007 Martinez-Conde et al 2002) Segev et al (2007) proposed that inprimates both saccades and microsaccades (which arerare in the archer fish) may contribute to size discrim-ination tasks as well as enhance spatial detail whereasdrifts and tremor by themselves may not be sufficient toimprove visual perception However it is important toremember that central vision in humans can be maintainedby tremor andor drift even when microsaccades aresuppressed during strict fixation (Martinez-Conde 2006Martinez-Conde et al 2004 2006) On the other handone cannot rule out the possibility that if one couldcompletely eliminate drifts and tremor microsaccadesmight then suffice to sustain both central and peripheralvision in primates and humans Future experiments areneeded to further explore these ideas
Figure 4 Archer fish eye movements (A) Example trace of eye position as a function of time as measured by search coil (B) Example ofa single 15-s fixation superimposed on a grid representing the photoreceptor mosaic The back and forth movement is caused by tremor(C) Distribution of fixation durations (taken as time interval between saccades) (D) Power spectrum of horizontal fixational eyemovements (E) Distribution of tremor amplitude (data taken from 1 animal in a 15-min session) Modified from Segev et al (2007)
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 10
Discussion
The vast majority of eye movement studies conductedin non-human species to date have not analyzed eyeposition data in the range of fixational eye movementsThus eye movements during fixation have been reportedin only a handful of vertebrates Most of these studieshave concentrated solely on the physical parameters ofthe fixational eye movements without addressing theirphysiological and perceptual consequences Fortunatelythis trend has started to reverse in recent years with agrowing list of papers addressing the effects of non-human fixational eye movements in physiology andperception This is an important development A largeamount of all psychophysical and physiological visualresearch has been conducted during conditions of visualfixation Therefore understanding the precise physiolog-ical and perceptual contributions of fixational eye move-ments may be critical to the interpretation of previous andfuture vision researchThere are some interesting differences and similarities
in the types of fixational eye movements produced bydifferent species For instance ocular tremor is possiblyubiquitous to all vertebrates The fact that tremor has notbeen observed in all species studied could be due to thetechnical difficulty of recording tremor accurately (ietremor amplitudes and frequencies are usually in the rangeof the recording systemrsquos noise) Drift has also beenrecorded in most vertebrates and it may be as omnipre-sent as tremor Conversely microsaccades seem mostimportant in species with foveal vision (ie primates andhumans) whereas they are scarce or absent in afoveatespecies (ie rabbits frogs goldfish) A link betweenmicrosaccades and foveal vision may reflect the two-pronged functional significance of microsaccades inhumans and primates first microsaccades may benecessary to correct the slow drift of the eye and bringback the objectregion of interest to the retinal area wherephotoreceptor density is highest (Cornsweet 1956) Thecorrective role of human microsaccades has been demon-strated in recent studies (Engbert amp Kliegl 2004 Engbertamp Mergenthaler 2006) Second the dynamics of micro-saccades themselves may improve spatial resolution(Donner amp Hemila 2005)In reviewing the patterns of fixational eye movements
across vertebrates it is tempting to draw parallels withother sensory modalities Fixational eye movementsrefresh the retinal images during fixation and thus preventneural adaptation and visual fading But this mechanism isnot circumscribed to the visual system Sniffs in rodentolfaction discretely sample sensory information every200ndash300 msec and thus are similar in their temporaldynamics to primate saccades (Otero-Millan et al 2008Uchida Kepecs amp Mainen 2006) and microsaccades(Otero-Millan et al 2008) A similar mode of discretesampling may also be at play when objects are recognized
through tactile information for instance if we sweep ourfingertips over an objectrsquos surface with our eyes closed orwhen blind individuals read Braille scriptOne intriguing possibility that merits further experi-
ments and analyses is the potential correlation between thespatiotemporal parameters of fixational eye movements(such as those summarized in Table 1) and the adaptationproperties of visual neurons across species For instancethe greater ocular stability of some species (rabbitgoldfish frog etc) versus others (primates) could beassociated with some particular adaptive properties oftheir visual cells Thus future comparative studies shouldpay attention to the response properties of visual cellstriggered by fixational eye movements in the presence ofstationary stimuli Other potential factors underlying thevariation in fixational eye movements across species couldbe differences in oculomotor control at the level of the eyemuscles or perhaps differences in retinal organizationFuture research will further improve our understanding
of fixational eye movements and the neural activity thatmaintains the visibility of stationary visual scenes inhuman and non-human species One interesting questionthat remains to be addressed concerns the role of tremor invision and visual physiology Because tremor frequenciesare well over the flicker fusion frequencies in humans ithas been argued that tremor of the visual image would beineffective as a stimulus (Ditchburn 1955 Gerrits ampVendrik 1970 Sharpe 1972) However recent studieshave found simulated tremor to enhance visual processingand improve visual acuity in a model retina (Hennig et al2002 Miller et al 2006) Follow-up research should beaimed to replicate these findings in actual neurons withcorrelated perceptual reports One should also note thatearly visual neurons can follow high-frequency flickeringthat surpasses the perceptual threshold for flicker fusion(Martinez-Conde et al 2002) Thus high-frequencytremor could be adequate to maintain activity in the earlyvisual system which then may lead to visual perception inhigher visual areas Another significant question thatremains unanswered concerns the oculomotor bases ofthe various types of fixational eye movements acrossspecies To date there is too little information on thisimportant topic to draw a conclusion
Conclusions
We have reviewed the parameters of fixational eyemovements in non-human mammals birds reptilesamphibians and fish as well as their consequences forphysiology and perception Fixational eye movementsand microsaccades in particular appear to be mostimportant in foveate vs afoveate species Animals lackingfixational eye movements such as afoveate frogs andtoads may be incapable of seeing stationary objects
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 11
during visual fixation Future research should furtherexplore the perceptual effects of fixational eye movementsand determine the oculomotor mechanisms responsible fortheir generation across species
Acknowledgments
We thank Mona Stewart and Isabel Gomez-Caraballo fortechnical assistance Jorge Otero-Millan for programmingassistance and Dr Xoana G Troncoso for comments on themanuscript This study was funded through grants fromthe Barrow Neurological Foundation (to SMC and SLM)the National Science Foundation (Award 0643306 toSMC) the Arizona Biomedical Research Commission(Award 07-102 to SMC) and Science Foundation Arizona(SLM)
Commercial relationships noneCorresponding author Susana Martinez-Conde PhDEmail smartneuralcorrelatecomAddress Barrow Neurological Institute 350 W ThomasRd Phoenix AZ 85013 USA
References
Aksay E Major G Goldman M S Baker R SeungH S amp Tank D W (2003) History dependence ofrate covariation between neurons during persistentactivity in an oculomotor integrator Cerebral Cortex13 1173ndash1184 [PubMed] [Article]
Aksay E Olasagasti I Mensh B D Baker RGoldman M S amp Tank D W (2007) Functionaldissection of circuitry in a neural integrator NatureNeuroscience 10 494ndash504 [PubMed]
Autrum H (1959) Das Fehlen unwillkurlicher Augenbe-wegungen beim Frosch Naturwissenschaften 46435
Baccus S A Olveczky B P Manu M amp Meister M(2008) A retinal circuit that computes object motionJournal of Neuroscience 28 6807ndash6817 [PubMed]
Bair W amp OrsquoKeefe L P (1998) The influence offixational eye movements on the response of neuronsin area MT of the macaque Visual Neuroscience 15779ndash786 [PubMed]
Betta E amp Turatto M (2006) Are you ready I can tellby looking at your microsaccades Neuroreport 171001ndash1004 [PubMed]
Bloch S Rivaud S amp Martinoya C (1984) Compar-ing frontal and lateral viewing in the pigeon IIIDifferent patterns of eye movements for binocularand monocular fixation Behavioural Brain Research13 173ndash182 [PubMed]
Cabrera B Torres B Pasaro R Pastor A M ampDelgado-Garcia J M (1992) A morphological studyof abducens nucleus motoneurons and internuclearneurons in the goldfish (Carassius auratus) BrainResearch Bulletin 28 137ndash144 [PubMed]
Carpenter R H S (1988) Movements of the eyes(2nd ed) Pion London
Collewijn H (1977) Eye- and head movements infreely moving rabbits The Journal of Physiology266 471ndash498 [PubMed] [Article]
Collewijn H amp van der Mark F (1972) Ocular stabilityin variable visual feedback conditions in the rabbitBrain Research 36 47ndash57 [PubMed]
Conway J L Timberlake G T amp Skavenski A A(1981) Oculomotor changes in cats reared withoutexperiencing continuous retinal image motion Exper-imental Brain Research 43 229ndash232 [PubMed]
Cornsweet T N (1956) Determination of the stimuli forinvoluntary drifts and saccadic eye movements Jour-nal of the Optical Society of America 46 987ndash993[PubMed]
Delgado-Garcia J M del Pozo F amp Baker R (1986)Behavior of neurons in the abducens nucleus of thealert catVII Internuclear neurons Neuroscience 17953ndash973 [PubMed]
Dieringer N amp Daunicht W J (1986) Image fadingVAproblem for frogs Naturwissenschaften 73 330ndash332[PubMed]
Dieringer N amp Precht W (1982) Compensatory headand eye movements in the frog and their contributionto stabilization of gaze Experimental BrainResearch 47 394ndash406 [PubMed]
Dieringer N Precht W amp Blight A R (1982)Resetting fast phases of head and eye and theirlinkage in the frog Experimental Brain Research 47407ndash416 [PubMed]
Ditchburn R W (1955) Eye-movements in relation toretinal action Optica Acta 1 171ndash176
Ditchburn R W amp Ginsborg B L (1952) Vision with astabilized retinal image Nature 170 36ndash37[PubMed]
Ditchburn R W amp Ginsborg B L (1953) Involuntaryeye movements during fixation The Journal ofPhysiology 119 1ndash17 [PubMed] [Article]
Donner K amp Hemila S (2007) Modelling the effect ofmicrosaccades on retinal responses to stationarycontrast patterns Vision Research 47 1166ndash1177[PubMed]
Donner K K amp Hemila S (2005) The possible purposeof microsaccades Paper presented at the EuropeanConference on Visual Perception A Coruna Spain
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 12
Easter S S Jr (1971) Spontaneous eye movements inrestrained goldfish Vision Research 11 333ndash342[PubMed]
Easter S S Jr Johns P R amp Heckenlively D R(1974) Horizontal compensatory eye movements ingoldfish (Carassius auratus) I The normal animalJournal of Comparative Physiology 92 23ndash35
Engbert R (2006) Microsaccades A microcosm forresearch on oculomotor control attention and visualperception Progress Brain Research 154 177ndash192[PubMed]
Engbert R amp Kliegl R (2003a) Binocular coordinationin microsaccades In R R J Hyona amp H Deubel(Eds) The mindrsquos eyes Cognitive and appliedaspects of eye movements (pp 103ndash117) OxfordUK Elsevier
Engbert R amp Kliegl R (2003b) Microsaccades uncoverthe orientation of covert attention Vision Research43 1035ndash1045 [PubMed]
Engbert R amp Kliegl R (2004) Microsaccades keep theeyesrsquo balance during fixation Psychological Science15 431ndash436 [PubMed]
Engbert R amp Mergenthaler K (2006) Microsaccadesare triggered by low retinal image slip Proceedingsof the National Academy of Sciences of the UnitedStates of America 103 7192ndash7197 [PubMed][Article]
Ewert J P amp Borchers H W (1974) Antwort vonretinalen Ganglienzellen bei freibeweglichen Kroten(Bufo Bufo L) Journal of Comparative and Physi-ology 92 117ndash130
Fuller J H (1980) Linkage of eye and head movementsin the alert rabbit Brain Research 194 219ndash222[PubMed]
Fuller J H (1981) Eye and head movements duringvestibular stimulation in the alert rabbit BrainResearch 205 363ndash381 [PubMed]
Galfano G Betta E amp Turatto M (2004) Inhibition ofreturn in microsaccades Experimental BrainResearch 159 400ndash404 [PubMed]
Gerrits H J amp Vendrik A J (1970) Artificial move-ments of a stabilized image Vision Research 101443ndash1456 [PubMed]
Gioanni H Bennis M amp Sansonetti A (1993) Visualand vestibular reflexes that stabilize gaze in thechameleon Visual Neuroscience 10 947ndash956[PubMed]
Goffart L Quinet J Chavane F amp Masson G S(2006) Influence of background illumination onfixation and visually guided saccades in the rhesusmonkey Vision Research 46 149ndash162 [PubMed]
Greschner M Bongard M Rujan P amp Ammermuller J(2002) Retinal ganglion cell synchronization by fixa-tional eye movements improves feature estimationNature Neuroscience 5 341ndash347 [PubMed]
Gur M Beylin A amp Snodderly D M (1997)Response variability of neurons in primary visualcortex (V1) of alert monkeys Journal of Neuro-science 17 2914ndash2920 [PubMed] [Article]
Hafed Z M amp Clark J J (2002) Microsaccades as anovert measure of covert attention shifts VisionResearch 42 2533ndash2545 [PubMed]
Hebbard F W amp Marg E (1960) Physiologicalnystagmus in the cat Journal of the Optical Societyof America 50 151ndash155 [PubMed]
Hennig M A Kerscher N J Funke K amp Worgotter F(2002) Stochastic resonance in visual cortical neuronsDoes the eye-tremor actually improve visual acuityNeurocomputing 44ndash46 115ndash120
Hennig M H amp Worgotter F (2004) Eye micro-movements improve stimulus detection beyond theNyquist limit in the peripheral retina In Advances inNeural Information Processing Systems (NIPS2003)(vol 16) Denver CO MIT Press
Hennig M H amp Worgotter F (2007) Effects offixational eye movements on retinal ganglion cellresponses A modelling study Frontiers in Computa-tional Neuroscience 1 2 [PubMed] [Article]
Hermann H T amp Constantine M M (1971) Eyemovements in the goldfish Vision Research 11313ndash331 [PubMed]
Horowitz T S Fine E M Fencsik D E Yurgenson Samp Wolfe J M (2007) Fixational eye movements arenot an index of covert attention PsychologicalScience 18 356ndash363 [PubMed]
Horwitz G D amp Albright T D (2003) Short-latencyfixational saccades induced by luminance incrementsJournal of Neurophysiology 90 1333ndash1339[PubMed] [Article]
Laubrock J Engbert R Rolfs M amp Kliegl R(2007) Microsaccades are an index of covertattention Commentary on Horowitz Fine FencsikYurgenson and Wolfe (2007) Psychological Sci-ence 18 364ndash366 discussion 367ndash368 [PubMed]
Leopold D A amp Logothetis N K (1998) Micro-saccades differentially modulate neural activity in thestriate and extrastriate visual cortex ExperimentalBrain Research 123 341ndash345 [PubMed]
Lettvin J Y Maturana H R MsCulloch W S amp PittsW H (1968) What the frogrsquos eye tells the frogrsquosbrain In W C C a M Balaban (Ed) The mind
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 13
Biological approaches to its functions (pp 233ndash258)John Wiley and Sons London
Macknik S L amp Livingstone M S (1998) Neuronalcorrelates of visibility and invisibility in the primatevisual system Nature Neuroscience 1 144ndash149[PubMed]
Macknik S L amp Martinez-Conde S (2004) The spatialand temporal effects of lateral inhibitory networksand their relevance to the visibility of spatiotemporaledges Neurocomputing 58ndash60 775ndash782
Macknik S L Martinez-Conde S amp Haglund M M(2000) The role of spatiotemporal edges in visibilityand visual masking Proceedings of the NationalAcademy of Sciences of the United States of America97 7556ndash7560 [PubMed] [Article]
Manteuffel G Plasa L Sommer T J amp Wess O(1977) Involuntary eye movements in salamandersNaturwissenschaften 64 533ndash534 [PubMed]
Marchetti E Gauthier G M amp Pellet J (1983)Cerebellar control of eye movements studied withinjection of harmaline in the trained baboon ArchivesItaliennes de Biologies 121 1ndash17 [PubMed]
Martinez-Conde S (2006) Fixational eye movements innormal and pathological vision Progress in BrainResearch 154 151ndash176 [PubMed]
Martinez-Conde S amp Macknik S L (2007a) Bobbingand perceiving Scientific American 297 16
Martinez-Conde S amp Macknik S L (2007b) Windowson the mind Scientific American 297 56ndash63[PubMed]
Martinez-Conde S Macknik S L amp Hubel D H(2000) Microsaccadic eye movements and firing ofsingle cells in the striate cortex of macaque monkeysNature Neuroscience 3 251ndash258 [PubMed]
Martinez-Conde S Macknik S L amp Hubel D H(2002) The function of bursts of spikes during visualfixation in the awake primate lateral geniculate nucleusand primary visual cortex Proceedings of theNational Academy of Sciences of the United Statesof America 99 13920ndash13925 [PubMed] [Article]
Martinez-Conde S Macknik S L amp Hubel D H(2004) The role of fixational eye movements invisual perception Nature Reviews Neuroscience 5229ndash240 [PubMed]
Martinez-Conde S Macknik S L Troncoso X G ampDyar T A (2006) Microsaccades counteract visualfading during fixation Neuron 49 297ndash305[PubMed] [Article]
Mensh B D Aksay E Lee D D Seung H S ampTank D W (2004) Spontaneous eye movements ingoldfish Oculomotor integrator performance plasti-
city and dependence on visual feedback VisionResearch 44 711ndash726 [PubMed]
Miller J A Denning K S George J S MarshakD W amp Kenyon G T (2006) A high frequencyresonance in the responses of retinal ganglion cells torapidly modulated stimuli A computer model VisualNeuroscience 23 779ndash794 [PubMed]
Montgomery J C McVean A R amp McCarthy D(1983) The effects of lowered temperature onspontaneous eye movements in a teleost fish Com-parative Biochemistry and Physiology A Compara-tive Physiology 75 363ndash368 [PubMed]
Motter B C amp Poggio G F (1984) Binocularfixation in the rhesus monkey Spatial and temporalcharacteristics Experimental Brain Research 54304ndash314 [PubMed]
Murakami I amp Cavanagh P (1998) A jitter after-effectreveals motion-based stabilization of vision Nature395 798ndash801 [PubMed]
Nye P W (1969) The monocular eye movements of thepigeon Vision Research 9 133ndash144 [PubMed]
Olveczky B P Baccus S A amp Meister M (2003)Segregation of object and background motion in theretina Nature 423 401ndash408 [PubMed]
Olveczky B P Baccus S A amp Meister M (2007)Retinal adaptation to object motion Neuron 56689ndash700 [PubMed] [Article]
Otero-Millan J Troncoso X Macknik S L Serrano-Pedraza I amp Martinez-Conde S (2008) Saccadesand microsaccades during visual fixation explorationand search Foundations for a common saccadicgenerator Journal of Vision 8(14)21 1ndash18 httpjournalofvisionorg81421 doi10116781421[PubMed] [Article]
Pastor A M Torres B Delgado-Garcia J M ampBaker R (1991) Discharge characteristics of medialrectus and abducens motoneurons in the goldfishJournal of Neurophysiology 66 2125ndash2140 [PubMed]
Pettigrew J D Wallman J amp Wildsoet C F (1990)Saccadic oscillations facilitate ocular perfusion fromthe avian pecten Nature 343 362ndash363 [PubMed]
Pritchard R M amp Heron W (1960) Small eye move-ments of the cat Canadian Journal of Psychology14 131ndash137 [PubMed]
Ratliff F amp Riggs L A (1950) Involuntary motions ofthe eye during monocular fixation Journal of Exper-imental Psychology 40 687ndash701 [PubMed]
Reppas J B UsreyWM amp Reid R C (2002) Saccadiceye movements modulate visual responses in thelateral geniculate nucleus Neuron 35 961ndash974[PubMed] [Article]
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 14
Riggs L A amp Ratliff F (1952) The effects of counter-acting the normal movements of the eye Journal ofthe Optical Society of America 42 872ndash873
Rolfs M Kliegl R amp Engbert R (2008) Toward amodel of microsaccade generation The case ofmicrosaccadic inhibition Journal of Vision 8(11)51ndash23 httpjournalofvisionorg8115 doi1011678115 [PubMed] [Article]
Rolfs M Laubrock J amp Kliegl R (2006) Shorteningand prolongation of saccade latencies followingmicrosaccades Experimental Brain Research 169369ndash376 [PubMed]
Rucci M Iovin R Poletti M amp Santini F (2007)Miniature eye movements enhance fine spatial detailNature 447 851ndash854 [PubMed]
Salas C Navarro F Torres B amp Delgado-Garcia J M(1992) Effects of diazepam and D-amphetamine onrhythmic pattern of eye movements in goldfishNeuroreport 3 131ndash134 [PubMed]
Schipperheyn J J (1963) Respiratory eye movementsand perception of stationary objects in the frog ActaPhysiologica et Pharmacologica Neerlandica 12157ndash159 [PubMed]
Schipperheyn J J (1965) Contrast detection in frogrsquosretina Acta Physiologica et Pharmacologica Neer-landica 13 231ndash277 [PubMed]
Schipperheyn J J (1967) Influence of eye movements onretinal ganglion cell activity in frogs Acta Physi-ologica et Pharmacologica Neerlandica 14 506[PubMed]
Segev R Schneidman E Goodhouse J amp Berry M J2nd (2007) Role of eye movements in the retinalcode for a size discrimination task Journal ofNeurophysiology 98 1380ndash1391 [PubMed] [Article]
Sharpe C R (1972) The visibility and fading of thinlines visualized by their controlled movement acrossthe retina The Journal of Physiology 222 113ndash134[PubMed] [Article]
Shelley M McLaughlin D Shapley R amp Wielaard J(2002) States of high conductance in a large-scalemodel of the visual cortex Journal of ComputationalNeuroscience 13 93ndash109 [PubMed]
Skavenski A A Robinson D A Steinman R M ampTimberlake G T (1975) Miniature eye movementsof fixation in rhesus monkey Vision Research 151269ndash1273 [PubMed]
Snodderly D M (1987) Effects of light and darkenvironments on macaque and human fixational eyemovements Vision Research 27 401ndash415 [PubMed]
Snodderly D M Kagan I amp Gur M (2001) Selectiveactivation of visual cortex neurons by fixational eyemovements Implications for neural coding VisualNeuroscience 18 259ndash277 [PubMed]
Snodderly D M amp Kurtz D (1985) Eye position duringfixation tasks Comparison of macaque and humanVision Research 25 83ndash98 [PubMed]
Steinbach M J amp Money K E (1973) Eye movementsof the owl Vision Research 13 889ndash891 [PubMed]
Steinman R M Haddad G M Skavenski A A ampWyman D (1973) Miniature eye movement Science181 810ndash819 [PubMed]
Torres B Pastor A M Cabrera B Salas C ampDelgado-Garcia J M (1992) Afferents to theoculomotor nucleus in the goldfish (Carassius aur-atus) as revealed by retrograde labeling with horse-radish peroxidase Journal of Comparative Neurology324 449ndash461 [PubMed]
Troncoso X G Macknik S L amp Martinez-Conde S(2008) Microsaccades counteract perceptual filling-inJournal of Vision 8(14)15 1ndash9 httpjournalofvisionorg81415 doi10116781415 [Article]
Troncoso X G Macknik S L Otero-Millan J ampMartinez-Conde S (2008) Microsaccades driveillusory motion in the Enigma illusion Proceedingsof the National Academy of Sciences of the UnitedStates of America 105 16033ndash16038 [PubMed]
Tse P U Sheinberg D S amp Logothetis N K (2004)The distribution of microsaccade directions need notreveal the location of attention Psychological Science15 708ndash710
Turatto M Valsecchi M Tame L amp Betta E (2007)Microsaccades distinguish between global and localvisual processing Neuroreport 18 1015ndash1018[PubMed]
Uchida N Kepecs A amp Mainen Z F (2006) Seeing ata glance smelling in a whiff Rapid forms ofperceptual decision making Nature Reviews Neuro-science 7 485ndash491 [PubMed]
Valsecchi M Betta E amp Turatto M (2007) Visualoddballs induce prolonged microsaccadic inhibitionExperimental Brain Research 177 196ndash208[PubMed]
van Dam L C amp van Ee R (2006) Retinal image shiftsbut not eye movements per se cause alternations inawareness during binocular rivalry Journal of Vision6(11)3 1172ndash1179 httpjournalofvisionorg6113doi1011676113 [PubMed] [Article]
Van der Steen J amp Collewijn H (1984) Ocular stabilityin the horizontal frontal and sagittal planes in therabbit Experimental Brain Research 56 263ndash274[PubMed]
Van Gisbergen J A Robinson D A amp Gielen S(1981) A quantitative analysis of generation ofsaccadic eye movements by burst neurons Journalof Neurophysiology 45 417ndash442 [PubMed]
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 15
Van Gisbergen J A M amp Robinson D A (1977)Generation of micro and macrosaccades by burstneurons in the monkey In R Baker amp A Berthoz(Eds) Control of gaze by brain stem neurons (vol 1pp 301ndash308) New York ElsevierNorth-Holland
Wallman J Pettigrew J D amp Letelier J C (1994)The oscillatory saccades of birds Motoneural activ-ity and possible functions In B T Fuchs A FV Buttner amp D Zee (Eds) Contemporary ocularmotor and vestibular research A tribute to David ARobinson (pp 480ndash487) Stuttgart Germany ThiemePublishing
Williams P E amp Shapley R M (2007) A dynamicnonlinearity and spatial phase specificity in macaque
V1 neurons Journal of Neuroscience 27 5706ndash5718[PubMed] [Article]
Winterson B J amp Robinson D A (1975) Fixationby the alert but solitary cat Vision Research 151349ndash1352 [PubMed]
Yarbus A L (1967) Eye movements and vision(B Haigh Trans) New York Plenum Press
Zuber B L amp Stark L (1965) Microsaccades and thevelocityltamplitude relationship for saccadic eyemovements Science 150 1459ndash1460 [PubMed]
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 16

(1968) stated that a frog ldquowill starve to death surroundedby food if it is not movingrdquo (but see Dieringer amp Daunicht1986 for a counterpoint) Small head motions or eyemovements correlated with respiratory motions maydiminish fading to some extent (Dieringer amp Precht1982 Schipperheyn 1963 1965 1967)As a side note the frogrsquos respiratory eye movements
play no role in the respiratory function ldquoin the course ofeach respiratory cycle increase of pressure in the frogrsquosbuccal cavity protrudes the eye and causes the opticalaxis to deviate upwards over 25ndash35 minutes of arcrdquo(Schipperheyn 1963) That is respiratory eye movementsare merely synchronized with respirationAutrum (1959) found no evidence of respiratory eye
movements in the frog Ewert andBorchers (1974) observedthat stationary stimuli elicit no responses from visualneurons in the common toad (Bufo Bufo L) regardless ofrespiratory pressure changes in the buccal cavity
Fixational eye movements in the salamander
Manteuffel et al (1977) identified two types of fixa-tional eye movements in salamanders tremor-like andrespiratory eye movements Tremor-like is a rapid move-ment of small amplitude Torsional eye movements resultfrom the animalrsquos breathing (also found in the frog bySchipperheyn 1963) Additionally flicks are occasionallyobserved which rapidly change the position of gaze It isunclear whether such flicks may generally correspond towhat we now call microsaccades andor to voluntarysaccades Based on these observations the authors pro-posed that unlike frogs and toads (Autrum 1959 Ewert ampBorchers 1974 Lettvin et al 1968) salamanders cancontinuously perceive stationary objects (Manteuffel et al1977) Fixational eye movements in the salamander (Tigersalamander Ambystoma tigrinum) may also help segre-gate object and background motion in the retina(Olveczky et al 2003 see also Fixational eye movementsin the rabbit section) and even work to emphasize novelvisual features (Olveczky et al 2007 see Baccus et al2008 for details on the retinal neurons and synapses thatmay underlie the computation of differential motionbetween an object and its background)
Fixational eye movements in fish
Spontaneous eye movements in the goldfish
Eye movements during fixation have been studied inboth afoveate (goldfish Carassius auratus) and foveate(archer fish Toxotes chatareus) fish species The goldfisha long-standing model organism in vision and oculomotor
research (Aksay et al 2003 2007 Cabrera Torres PasaroPastor amp Delgado-Garcia 1992 Pastor Torres Delgado-Garcia amp Baker 1991 Salas Navarro Torres ampDelgado-Garcia 1992 Torres Pastor Cabrera Salas ampDelgado-Garcia 1992) produces three types of sponta-neous eye movements saccades stretch and drift (Easter1971 Mensh Aksay Lee Seung amp Tank 2004) Weshould note that in the motionless goldfish eye move-ments are very infrequent Those that do occur consistmainly of saccades slow drifts are very rare (Easter Johnsamp Heckenlively 1974) During saccades (identified bythresholding the eye velocity at 5 degsec) both eyes moverapidly simultaneously and usually in the same direction(Mensh et al 2004) Stretches occur every 2ndash3 min and arecharacterized by a brief simultaneous large excursion ofboth eyes to extreme temporal positions (Easter 1971Mensh et al 2004) The function of stretches is not known(Mensh et al 2004) Because fixations were defined asthose periods that were free of saccadic and stretchmovements (Easter 1971) only drifts (from the threetypes of spontaneous movements) can technically beconsidered ldquofixational eye movementsrdquo However thegoldfish saccadic main sequence includes saccades withequivalent amplitudes and velocities to those found inhuman microsaccades (Mensh et al 2004 Zuber amp Stark1965) Indeed microsaccades have been identified in other(foveate) fish species although they are rarer than inhumans (Segev et al 2007) Small saccades in thegoldfish tend to be divergent whereas large saccadestend to be convergent (Mensh et al 2004) Drift occurssimultaneously in both eyes usually in the samedirection Drift velocities are almost exclusively e1 degsec(Mensh et al 2004) No tremor has been identified in thegoldfish
Spontaneous eye movements in teleost fish
Spontaneous eye movements have been studied in theNew Zealand parore (Girella tricuspidata MontgomeryMcVean amp McCarthy 1983) The pattern of eye move-ments was similar to that previously described in goldfish(Easter 1971 Hermann amp Constantine 1971)
Fixational eye movements in the archer fish
A recent study in the archer fish (Segev et al 2007) hasprovided the first perceptual and physiological correlatesof fixational eye movements in any fish species Archerfish have remarkable visual abilities they shoot downinsects resting on foliageVas well as stationary targetscovering a single retinal photoreceptorVwith squirts ofwater from their mouths they predict the location atwhich the insect will splash down in G100 msec and theydeduce the absolute size of an object presented at differentviewing distances (Supplementary Videos 2 and 3) Segev
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 9
et al (2007) identified three types of fixational eyemovements in the archer fish microsaccades drift andtremor Microsaccades were rare and were not analyzed indetail Drift velocities were e040 degsec Tremor was ahigh-frequency oscillatory movement (the primary com-ponent was 5 Hz) with peak-to-peak amplitude of 01ndash02 deg (corresponding to an image displacement of 1ndash2photoreceptors) and velocity of about 1 degsec (Figure 4)Whereas the amplitude and velocity of tremor in thearcher fish is comparable to those in humans its frequencyis much lower (5 Hz vs 940 Hz) Interestingly archer fishtremor is very similar both in amplitude (photoreceptorwidths covered) as well as in frequency (5 Hz) to thetremor-like periodic motion in the turtle (discussedearlier Greschner et al 2002)Segev et al (2007) trained archer fish to perform a size
discrimination task the fish had to distinguish a medium-sized target from larger and smaller distracters (theyindicated their choice by shooting a jet of water) To relatethis behavior to responses from retinal neurons theauthors recorded from populations of retinal ganglioncells while presenting objects of different sizes moved tosimulate saccades drifts and tremor Drifts and tremorelicited weak neuronal responses that were not veryinformative about the spatial structure of the stimulus(although they did provide some information about target
size in agreement with the results of Greschner et al2002 in the turtle) Moreover performance was similar fortremor alone and for tremor plus drift indicating that driftplays a negligible role in revealing spatial structure (alsoin agreement with Greschner et al 2002)When present the firing rate was phase-locked to
tremor Saccades were more effective than drifts or tremorin driving informative responses from the ganglion cells(consistent with theoretical and physiological studies ofmicrosaccade-driven neuronal activity in the primateretina and LGN Donner amp Hemila 2007 Martinez-Conde et al 2002) Segev et al (2007) proposed that inprimates both saccades and microsaccades (which arerare in the archer fish) may contribute to size discrim-ination tasks as well as enhance spatial detail whereasdrifts and tremor by themselves may not be sufficient toimprove visual perception However it is important toremember that central vision in humans can be maintainedby tremor andor drift even when microsaccades aresuppressed during strict fixation (Martinez-Conde 2006Martinez-Conde et al 2004 2006) On the other handone cannot rule out the possibility that if one couldcompletely eliminate drifts and tremor microsaccadesmight then suffice to sustain both central and peripheralvision in primates and humans Future experiments areneeded to further explore these ideas
Figure 4 Archer fish eye movements (A) Example trace of eye position as a function of time as measured by search coil (B) Example ofa single 15-s fixation superimposed on a grid representing the photoreceptor mosaic The back and forth movement is caused by tremor(C) Distribution of fixation durations (taken as time interval between saccades) (D) Power spectrum of horizontal fixational eyemovements (E) Distribution of tremor amplitude (data taken from 1 animal in a 15-min session) Modified from Segev et al (2007)
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 10
Discussion
The vast majority of eye movement studies conductedin non-human species to date have not analyzed eyeposition data in the range of fixational eye movementsThus eye movements during fixation have been reportedin only a handful of vertebrates Most of these studieshave concentrated solely on the physical parameters ofthe fixational eye movements without addressing theirphysiological and perceptual consequences Fortunatelythis trend has started to reverse in recent years with agrowing list of papers addressing the effects of non-human fixational eye movements in physiology andperception This is an important development A largeamount of all psychophysical and physiological visualresearch has been conducted during conditions of visualfixation Therefore understanding the precise physiolog-ical and perceptual contributions of fixational eye move-ments may be critical to the interpretation of previous andfuture vision researchThere are some interesting differences and similarities
in the types of fixational eye movements produced bydifferent species For instance ocular tremor is possiblyubiquitous to all vertebrates The fact that tremor has notbeen observed in all species studied could be due to thetechnical difficulty of recording tremor accurately (ietremor amplitudes and frequencies are usually in the rangeof the recording systemrsquos noise) Drift has also beenrecorded in most vertebrates and it may be as omnipre-sent as tremor Conversely microsaccades seem mostimportant in species with foveal vision (ie primates andhumans) whereas they are scarce or absent in afoveatespecies (ie rabbits frogs goldfish) A link betweenmicrosaccades and foveal vision may reflect the two-pronged functional significance of microsaccades inhumans and primates first microsaccades may benecessary to correct the slow drift of the eye and bringback the objectregion of interest to the retinal area wherephotoreceptor density is highest (Cornsweet 1956) Thecorrective role of human microsaccades has been demon-strated in recent studies (Engbert amp Kliegl 2004 Engbertamp Mergenthaler 2006) Second the dynamics of micro-saccades themselves may improve spatial resolution(Donner amp Hemila 2005)In reviewing the patterns of fixational eye movements
across vertebrates it is tempting to draw parallels withother sensory modalities Fixational eye movementsrefresh the retinal images during fixation and thus preventneural adaptation and visual fading But this mechanism isnot circumscribed to the visual system Sniffs in rodentolfaction discretely sample sensory information every200ndash300 msec and thus are similar in their temporaldynamics to primate saccades (Otero-Millan et al 2008Uchida Kepecs amp Mainen 2006) and microsaccades(Otero-Millan et al 2008) A similar mode of discretesampling may also be at play when objects are recognized
through tactile information for instance if we sweep ourfingertips over an objectrsquos surface with our eyes closed orwhen blind individuals read Braille scriptOne intriguing possibility that merits further experi-
ments and analyses is the potential correlation between thespatiotemporal parameters of fixational eye movements(such as those summarized in Table 1) and the adaptationproperties of visual neurons across species For instancethe greater ocular stability of some species (rabbitgoldfish frog etc) versus others (primates) could beassociated with some particular adaptive properties oftheir visual cells Thus future comparative studies shouldpay attention to the response properties of visual cellstriggered by fixational eye movements in the presence ofstationary stimuli Other potential factors underlying thevariation in fixational eye movements across species couldbe differences in oculomotor control at the level of the eyemuscles or perhaps differences in retinal organizationFuture research will further improve our understanding
of fixational eye movements and the neural activity thatmaintains the visibility of stationary visual scenes inhuman and non-human species One interesting questionthat remains to be addressed concerns the role of tremor invision and visual physiology Because tremor frequenciesare well over the flicker fusion frequencies in humans ithas been argued that tremor of the visual image would beineffective as a stimulus (Ditchburn 1955 Gerrits ampVendrik 1970 Sharpe 1972) However recent studieshave found simulated tremor to enhance visual processingand improve visual acuity in a model retina (Hennig et al2002 Miller et al 2006) Follow-up research should beaimed to replicate these findings in actual neurons withcorrelated perceptual reports One should also note thatearly visual neurons can follow high-frequency flickeringthat surpasses the perceptual threshold for flicker fusion(Martinez-Conde et al 2002) Thus high-frequencytremor could be adequate to maintain activity in the earlyvisual system which then may lead to visual perception inhigher visual areas Another significant question thatremains unanswered concerns the oculomotor bases ofthe various types of fixational eye movements acrossspecies To date there is too little information on thisimportant topic to draw a conclusion
Conclusions
We have reviewed the parameters of fixational eyemovements in non-human mammals birds reptilesamphibians and fish as well as their consequences forphysiology and perception Fixational eye movementsand microsaccades in particular appear to be mostimportant in foveate vs afoveate species Animals lackingfixational eye movements such as afoveate frogs andtoads may be incapable of seeing stationary objects
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 11
during visual fixation Future research should furtherexplore the perceptual effects of fixational eye movementsand determine the oculomotor mechanisms responsible fortheir generation across species
Acknowledgments
We thank Mona Stewart and Isabel Gomez-Caraballo fortechnical assistance Jorge Otero-Millan for programmingassistance and Dr Xoana G Troncoso for comments on themanuscript This study was funded through grants fromthe Barrow Neurological Foundation (to SMC and SLM)the National Science Foundation (Award 0643306 toSMC) the Arizona Biomedical Research Commission(Award 07-102 to SMC) and Science Foundation Arizona(SLM)
Commercial relationships noneCorresponding author Susana Martinez-Conde PhDEmail smartneuralcorrelatecomAddress Barrow Neurological Institute 350 W ThomasRd Phoenix AZ 85013 USA
References
Aksay E Major G Goldman M S Baker R SeungH S amp Tank D W (2003) History dependence ofrate covariation between neurons during persistentactivity in an oculomotor integrator Cerebral Cortex13 1173ndash1184 [PubMed] [Article]
Aksay E Olasagasti I Mensh B D Baker RGoldman M S amp Tank D W (2007) Functionaldissection of circuitry in a neural integrator NatureNeuroscience 10 494ndash504 [PubMed]
Autrum H (1959) Das Fehlen unwillkurlicher Augenbe-wegungen beim Frosch Naturwissenschaften 46435
Baccus S A Olveczky B P Manu M amp Meister M(2008) A retinal circuit that computes object motionJournal of Neuroscience 28 6807ndash6817 [PubMed]
Bair W amp OrsquoKeefe L P (1998) The influence offixational eye movements on the response of neuronsin area MT of the macaque Visual Neuroscience 15779ndash786 [PubMed]
Betta E amp Turatto M (2006) Are you ready I can tellby looking at your microsaccades Neuroreport 171001ndash1004 [PubMed]
Bloch S Rivaud S amp Martinoya C (1984) Compar-ing frontal and lateral viewing in the pigeon IIIDifferent patterns of eye movements for binocularand monocular fixation Behavioural Brain Research13 173ndash182 [PubMed]
Cabrera B Torres B Pasaro R Pastor A M ampDelgado-Garcia J M (1992) A morphological studyof abducens nucleus motoneurons and internuclearneurons in the goldfish (Carassius auratus) BrainResearch Bulletin 28 137ndash144 [PubMed]
Carpenter R H S (1988) Movements of the eyes(2nd ed) Pion London
Collewijn H (1977) Eye- and head movements infreely moving rabbits The Journal of Physiology266 471ndash498 [PubMed] [Article]
Collewijn H amp van der Mark F (1972) Ocular stabilityin variable visual feedback conditions in the rabbitBrain Research 36 47ndash57 [PubMed]
Conway J L Timberlake G T amp Skavenski A A(1981) Oculomotor changes in cats reared withoutexperiencing continuous retinal image motion Exper-imental Brain Research 43 229ndash232 [PubMed]
Cornsweet T N (1956) Determination of the stimuli forinvoluntary drifts and saccadic eye movements Jour-nal of the Optical Society of America 46 987ndash993[PubMed]
Delgado-Garcia J M del Pozo F amp Baker R (1986)Behavior of neurons in the abducens nucleus of thealert catVII Internuclear neurons Neuroscience 17953ndash973 [PubMed]
Dieringer N amp Daunicht W J (1986) Image fadingVAproblem for frogs Naturwissenschaften 73 330ndash332[PubMed]
Dieringer N amp Precht W (1982) Compensatory headand eye movements in the frog and their contributionto stabilization of gaze Experimental BrainResearch 47 394ndash406 [PubMed]
Dieringer N Precht W amp Blight A R (1982)Resetting fast phases of head and eye and theirlinkage in the frog Experimental Brain Research 47407ndash416 [PubMed]
Ditchburn R W (1955) Eye-movements in relation toretinal action Optica Acta 1 171ndash176
Ditchburn R W amp Ginsborg B L (1952) Vision with astabilized retinal image Nature 170 36ndash37[PubMed]
Ditchburn R W amp Ginsborg B L (1953) Involuntaryeye movements during fixation The Journal ofPhysiology 119 1ndash17 [PubMed] [Article]
Donner K amp Hemila S (2007) Modelling the effect ofmicrosaccades on retinal responses to stationarycontrast patterns Vision Research 47 1166ndash1177[PubMed]
Donner K K amp Hemila S (2005) The possible purposeof microsaccades Paper presented at the EuropeanConference on Visual Perception A Coruna Spain
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 12
Easter S S Jr (1971) Spontaneous eye movements inrestrained goldfish Vision Research 11 333ndash342[PubMed]
Easter S S Jr Johns P R amp Heckenlively D R(1974) Horizontal compensatory eye movements ingoldfish (Carassius auratus) I The normal animalJournal of Comparative Physiology 92 23ndash35
Engbert R (2006) Microsaccades A microcosm forresearch on oculomotor control attention and visualperception Progress Brain Research 154 177ndash192[PubMed]
Engbert R amp Kliegl R (2003a) Binocular coordinationin microsaccades In R R J Hyona amp H Deubel(Eds) The mindrsquos eyes Cognitive and appliedaspects of eye movements (pp 103ndash117) OxfordUK Elsevier
Engbert R amp Kliegl R (2003b) Microsaccades uncoverthe orientation of covert attention Vision Research43 1035ndash1045 [PubMed]
Engbert R amp Kliegl R (2004) Microsaccades keep theeyesrsquo balance during fixation Psychological Science15 431ndash436 [PubMed]
Engbert R amp Mergenthaler K (2006) Microsaccadesare triggered by low retinal image slip Proceedingsof the National Academy of Sciences of the UnitedStates of America 103 7192ndash7197 [PubMed][Article]
Ewert J P amp Borchers H W (1974) Antwort vonretinalen Ganglienzellen bei freibeweglichen Kroten(Bufo Bufo L) Journal of Comparative and Physi-ology 92 117ndash130
Fuller J H (1980) Linkage of eye and head movementsin the alert rabbit Brain Research 194 219ndash222[PubMed]
Fuller J H (1981) Eye and head movements duringvestibular stimulation in the alert rabbit BrainResearch 205 363ndash381 [PubMed]
Galfano G Betta E amp Turatto M (2004) Inhibition ofreturn in microsaccades Experimental BrainResearch 159 400ndash404 [PubMed]
Gerrits H J amp Vendrik A J (1970) Artificial move-ments of a stabilized image Vision Research 101443ndash1456 [PubMed]
Gioanni H Bennis M amp Sansonetti A (1993) Visualand vestibular reflexes that stabilize gaze in thechameleon Visual Neuroscience 10 947ndash956[PubMed]
Goffart L Quinet J Chavane F amp Masson G S(2006) Influence of background illumination onfixation and visually guided saccades in the rhesusmonkey Vision Research 46 149ndash162 [PubMed]
Greschner M Bongard M Rujan P amp Ammermuller J(2002) Retinal ganglion cell synchronization by fixa-tional eye movements improves feature estimationNature Neuroscience 5 341ndash347 [PubMed]
Gur M Beylin A amp Snodderly D M (1997)Response variability of neurons in primary visualcortex (V1) of alert monkeys Journal of Neuro-science 17 2914ndash2920 [PubMed] [Article]
Hafed Z M amp Clark J J (2002) Microsaccades as anovert measure of covert attention shifts VisionResearch 42 2533ndash2545 [PubMed]
Hebbard F W amp Marg E (1960) Physiologicalnystagmus in the cat Journal of the Optical Societyof America 50 151ndash155 [PubMed]
Hennig M A Kerscher N J Funke K amp Worgotter F(2002) Stochastic resonance in visual cortical neuronsDoes the eye-tremor actually improve visual acuityNeurocomputing 44ndash46 115ndash120
Hennig M H amp Worgotter F (2004) Eye micro-movements improve stimulus detection beyond theNyquist limit in the peripheral retina In Advances inNeural Information Processing Systems (NIPS2003)(vol 16) Denver CO MIT Press
Hennig M H amp Worgotter F (2007) Effects offixational eye movements on retinal ganglion cellresponses A modelling study Frontiers in Computa-tional Neuroscience 1 2 [PubMed] [Article]
Hermann H T amp Constantine M M (1971) Eyemovements in the goldfish Vision Research 11313ndash331 [PubMed]
Horowitz T S Fine E M Fencsik D E Yurgenson Samp Wolfe J M (2007) Fixational eye movements arenot an index of covert attention PsychologicalScience 18 356ndash363 [PubMed]
Horwitz G D amp Albright T D (2003) Short-latencyfixational saccades induced by luminance incrementsJournal of Neurophysiology 90 1333ndash1339[PubMed] [Article]
Laubrock J Engbert R Rolfs M amp Kliegl R(2007) Microsaccades are an index of covertattention Commentary on Horowitz Fine FencsikYurgenson and Wolfe (2007) Psychological Sci-ence 18 364ndash366 discussion 367ndash368 [PubMed]
Leopold D A amp Logothetis N K (1998) Micro-saccades differentially modulate neural activity in thestriate and extrastriate visual cortex ExperimentalBrain Research 123 341ndash345 [PubMed]
Lettvin J Y Maturana H R MsCulloch W S amp PittsW H (1968) What the frogrsquos eye tells the frogrsquosbrain In W C C a M Balaban (Ed) The mind
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 13
Biological approaches to its functions (pp 233ndash258)John Wiley and Sons London
Macknik S L amp Livingstone M S (1998) Neuronalcorrelates of visibility and invisibility in the primatevisual system Nature Neuroscience 1 144ndash149[PubMed]
Macknik S L amp Martinez-Conde S (2004) The spatialand temporal effects of lateral inhibitory networksand their relevance to the visibility of spatiotemporaledges Neurocomputing 58ndash60 775ndash782
Macknik S L Martinez-Conde S amp Haglund M M(2000) The role of spatiotemporal edges in visibilityand visual masking Proceedings of the NationalAcademy of Sciences of the United States of America97 7556ndash7560 [PubMed] [Article]
Manteuffel G Plasa L Sommer T J amp Wess O(1977) Involuntary eye movements in salamandersNaturwissenschaften 64 533ndash534 [PubMed]
Marchetti E Gauthier G M amp Pellet J (1983)Cerebellar control of eye movements studied withinjection of harmaline in the trained baboon ArchivesItaliennes de Biologies 121 1ndash17 [PubMed]
Martinez-Conde S (2006) Fixational eye movements innormal and pathological vision Progress in BrainResearch 154 151ndash176 [PubMed]
Martinez-Conde S amp Macknik S L (2007a) Bobbingand perceiving Scientific American 297 16
Martinez-Conde S amp Macknik S L (2007b) Windowson the mind Scientific American 297 56ndash63[PubMed]
Martinez-Conde S Macknik S L amp Hubel D H(2000) Microsaccadic eye movements and firing ofsingle cells in the striate cortex of macaque monkeysNature Neuroscience 3 251ndash258 [PubMed]
Martinez-Conde S Macknik S L amp Hubel D H(2002) The function of bursts of spikes during visualfixation in the awake primate lateral geniculate nucleusand primary visual cortex Proceedings of theNational Academy of Sciences of the United Statesof America 99 13920ndash13925 [PubMed] [Article]
Martinez-Conde S Macknik S L amp Hubel D H(2004) The role of fixational eye movements invisual perception Nature Reviews Neuroscience 5229ndash240 [PubMed]
Martinez-Conde S Macknik S L Troncoso X G ampDyar T A (2006) Microsaccades counteract visualfading during fixation Neuron 49 297ndash305[PubMed] [Article]
Mensh B D Aksay E Lee D D Seung H S ampTank D W (2004) Spontaneous eye movements ingoldfish Oculomotor integrator performance plasti-
city and dependence on visual feedback VisionResearch 44 711ndash726 [PubMed]
Miller J A Denning K S George J S MarshakD W amp Kenyon G T (2006) A high frequencyresonance in the responses of retinal ganglion cells torapidly modulated stimuli A computer model VisualNeuroscience 23 779ndash794 [PubMed]
Montgomery J C McVean A R amp McCarthy D(1983) The effects of lowered temperature onspontaneous eye movements in a teleost fish Com-parative Biochemistry and Physiology A Compara-tive Physiology 75 363ndash368 [PubMed]
Motter B C amp Poggio G F (1984) Binocularfixation in the rhesus monkey Spatial and temporalcharacteristics Experimental Brain Research 54304ndash314 [PubMed]
Murakami I amp Cavanagh P (1998) A jitter after-effectreveals motion-based stabilization of vision Nature395 798ndash801 [PubMed]
Nye P W (1969) The monocular eye movements of thepigeon Vision Research 9 133ndash144 [PubMed]
Olveczky B P Baccus S A amp Meister M (2003)Segregation of object and background motion in theretina Nature 423 401ndash408 [PubMed]
Olveczky B P Baccus S A amp Meister M (2007)Retinal adaptation to object motion Neuron 56689ndash700 [PubMed] [Article]
Otero-Millan J Troncoso X Macknik S L Serrano-Pedraza I amp Martinez-Conde S (2008) Saccadesand microsaccades during visual fixation explorationand search Foundations for a common saccadicgenerator Journal of Vision 8(14)21 1ndash18 httpjournalofvisionorg81421 doi10116781421[PubMed] [Article]
Pastor A M Torres B Delgado-Garcia J M ampBaker R (1991) Discharge characteristics of medialrectus and abducens motoneurons in the goldfishJournal of Neurophysiology 66 2125ndash2140 [PubMed]
Pettigrew J D Wallman J amp Wildsoet C F (1990)Saccadic oscillations facilitate ocular perfusion fromthe avian pecten Nature 343 362ndash363 [PubMed]
Pritchard R M amp Heron W (1960) Small eye move-ments of the cat Canadian Journal of Psychology14 131ndash137 [PubMed]
Ratliff F amp Riggs L A (1950) Involuntary motions ofthe eye during monocular fixation Journal of Exper-imental Psychology 40 687ndash701 [PubMed]
Reppas J B UsreyWM amp Reid R C (2002) Saccadiceye movements modulate visual responses in thelateral geniculate nucleus Neuron 35 961ndash974[PubMed] [Article]
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 14
Riggs L A amp Ratliff F (1952) The effects of counter-acting the normal movements of the eye Journal ofthe Optical Society of America 42 872ndash873
Rolfs M Kliegl R amp Engbert R (2008) Toward amodel of microsaccade generation The case ofmicrosaccadic inhibition Journal of Vision 8(11)51ndash23 httpjournalofvisionorg8115 doi1011678115 [PubMed] [Article]
Rolfs M Laubrock J amp Kliegl R (2006) Shorteningand prolongation of saccade latencies followingmicrosaccades Experimental Brain Research 169369ndash376 [PubMed]
Rucci M Iovin R Poletti M amp Santini F (2007)Miniature eye movements enhance fine spatial detailNature 447 851ndash854 [PubMed]
Salas C Navarro F Torres B amp Delgado-Garcia J M(1992) Effects of diazepam and D-amphetamine onrhythmic pattern of eye movements in goldfishNeuroreport 3 131ndash134 [PubMed]
Schipperheyn J J (1963) Respiratory eye movementsand perception of stationary objects in the frog ActaPhysiologica et Pharmacologica Neerlandica 12157ndash159 [PubMed]
Schipperheyn J J (1965) Contrast detection in frogrsquosretina Acta Physiologica et Pharmacologica Neer-landica 13 231ndash277 [PubMed]
Schipperheyn J J (1967) Influence of eye movements onretinal ganglion cell activity in frogs Acta Physi-ologica et Pharmacologica Neerlandica 14 506[PubMed]
Segev R Schneidman E Goodhouse J amp Berry M J2nd (2007) Role of eye movements in the retinalcode for a size discrimination task Journal ofNeurophysiology 98 1380ndash1391 [PubMed] [Article]
Sharpe C R (1972) The visibility and fading of thinlines visualized by their controlled movement acrossthe retina The Journal of Physiology 222 113ndash134[PubMed] [Article]
Shelley M McLaughlin D Shapley R amp Wielaard J(2002) States of high conductance in a large-scalemodel of the visual cortex Journal of ComputationalNeuroscience 13 93ndash109 [PubMed]
Skavenski A A Robinson D A Steinman R M ampTimberlake G T (1975) Miniature eye movementsof fixation in rhesus monkey Vision Research 151269ndash1273 [PubMed]
Snodderly D M (1987) Effects of light and darkenvironments on macaque and human fixational eyemovements Vision Research 27 401ndash415 [PubMed]
Snodderly D M Kagan I amp Gur M (2001) Selectiveactivation of visual cortex neurons by fixational eyemovements Implications for neural coding VisualNeuroscience 18 259ndash277 [PubMed]
Snodderly D M amp Kurtz D (1985) Eye position duringfixation tasks Comparison of macaque and humanVision Research 25 83ndash98 [PubMed]
Steinbach M J amp Money K E (1973) Eye movementsof the owl Vision Research 13 889ndash891 [PubMed]
Steinman R M Haddad G M Skavenski A A ampWyman D (1973) Miniature eye movement Science181 810ndash819 [PubMed]
Torres B Pastor A M Cabrera B Salas C ampDelgado-Garcia J M (1992) Afferents to theoculomotor nucleus in the goldfish (Carassius aur-atus) as revealed by retrograde labeling with horse-radish peroxidase Journal of Comparative Neurology324 449ndash461 [PubMed]
Troncoso X G Macknik S L amp Martinez-Conde S(2008) Microsaccades counteract perceptual filling-inJournal of Vision 8(14)15 1ndash9 httpjournalofvisionorg81415 doi10116781415 [Article]
Troncoso X G Macknik S L Otero-Millan J ampMartinez-Conde S (2008) Microsaccades driveillusory motion in the Enigma illusion Proceedingsof the National Academy of Sciences of the UnitedStates of America 105 16033ndash16038 [PubMed]
Tse P U Sheinberg D S amp Logothetis N K (2004)The distribution of microsaccade directions need notreveal the location of attention Psychological Science15 708ndash710
Turatto M Valsecchi M Tame L amp Betta E (2007)Microsaccades distinguish between global and localvisual processing Neuroreport 18 1015ndash1018[PubMed]
Uchida N Kepecs A amp Mainen Z F (2006) Seeing ata glance smelling in a whiff Rapid forms ofperceptual decision making Nature Reviews Neuro-science 7 485ndash491 [PubMed]
Valsecchi M Betta E amp Turatto M (2007) Visualoddballs induce prolonged microsaccadic inhibitionExperimental Brain Research 177 196ndash208[PubMed]
van Dam L C amp van Ee R (2006) Retinal image shiftsbut not eye movements per se cause alternations inawareness during binocular rivalry Journal of Vision6(11)3 1172ndash1179 httpjournalofvisionorg6113doi1011676113 [PubMed] [Article]
Van der Steen J amp Collewijn H (1984) Ocular stabilityin the horizontal frontal and sagittal planes in therabbit Experimental Brain Research 56 263ndash274[PubMed]
Van Gisbergen J A Robinson D A amp Gielen S(1981) A quantitative analysis of generation ofsaccadic eye movements by burst neurons Journalof Neurophysiology 45 417ndash442 [PubMed]
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 15
Van Gisbergen J A M amp Robinson D A (1977)Generation of micro and macrosaccades by burstneurons in the monkey In R Baker amp A Berthoz(Eds) Control of gaze by brain stem neurons (vol 1pp 301ndash308) New York ElsevierNorth-Holland
Wallman J Pettigrew J D amp Letelier J C (1994)The oscillatory saccades of birds Motoneural activ-ity and possible functions In B T Fuchs A FV Buttner amp D Zee (Eds) Contemporary ocularmotor and vestibular research A tribute to David ARobinson (pp 480ndash487) Stuttgart Germany ThiemePublishing
Williams P E amp Shapley R M (2007) A dynamicnonlinearity and spatial phase specificity in macaque
V1 neurons Journal of Neuroscience 27 5706ndash5718[PubMed] [Article]
Winterson B J amp Robinson D A (1975) Fixationby the alert but solitary cat Vision Research 151349ndash1352 [PubMed]
Yarbus A L (1967) Eye movements and vision(B Haigh Trans) New York Plenum Press
Zuber B L amp Stark L (1965) Microsaccades and thevelocityltamplitude relationship for saccadic eyemovements Science 150 1459ndash1460 [PubMed]
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 16

et al (2007) identified three types of fixational eyemovements in the archer fish microsaccades drift andtremor Microsaccades were rare and were not analyzed indetail Drift velocities were e040 degsec Tremor was ahigh-frequency oscillatory movement (the primary com-ponent was 5 Hz) with peak-to-peak amplitude of 01ndash02 deg (corresponding to an image displacement of 1ndash2photoreceptors) and velocity of about 1 degsec (Figure 4)Whereas the amplitude and velocity of tremor in thearcher fish is comparable to those in humans its frequencyis much lower (5 Hz vs 940 Hz) Interestingly archer fishtremor is very similar both in amplitude (photoreceptorwidths covered) as well as in frequency (5 Hz) to thetremor-like periodic motion in the turtle (discussedearlier Greschner et al 2002)Segev et al (2007) trained archer fish to perform a size
discrimination task the fish had to distinguish a medium-sized target from larger and smaller distracters (theyindicated their choice by shooting a jet of water) To relatethis behavior to responses from retinal neurons theauthors recorded from populations of retinal ganglioncells while presenting objects of different sizes moved tosimulate saccades drifts and tremor Drifts and tremorelicited weak neuronal responses that were not veryinformative about the spatial structure of the stimulus(although they did provide some information about target
size in agreement with the results of Greschner et al2002 in the turtle) Moreover performance was similar fortremor alone and for tremor plus drift indicating that driftplays a negligible role in revealing spatial structure (alsoin agreement with Greschner et al 2002)When present the firing rate was phase-locked to
tremor Saccades were more effective than drifts or tremorin driving informative responses from the ganglion cells(consistent with theoretical and physiological studies ofmicrosaccade-driven neuronal activity in the primateretina and LGN Donner amp Hemila 2007 Martinez-Conde et al 2002) Segev et al (2007) proposed that inprimates both saccades and microsaccades (which arerare in the archer fish) may contribute to size discrim-ination tasks as well as enhance spatial detail whereasdrifts and tremor by themselves may not be sufficient toimprove visual perception However it is important toremember that central vision in humans can be maintainedby tremor andor drift even when microsaccades aresuppressed during strict fixation (Martinez-Conde 2006Martinez-Conde et al 2004 2006) On the other handone cannot rule out the possibility that if one couldcompletely eliminate drifts and tremor microsaccadesmight then suffice to sustain both central and peripheralvision in primates and humans Future experiments areneeded to further explore these ideas
Figure 4 Archer fish eye movements (A) Example trace of eye position as a function of time as measured by search coil (B) Example ofa single 15-s fixation superimposed on a grid representing the photoreceptor mosaic The back and forth movement is caused by tremor(C) Distribution of fixation durations (taken as time interval between saccades) (D) Power spectrum of horizontal fixational eyemovements (E) Distribution of tremor amplitude (data taken from 1 animal in a 15-min session) Modified from Segev et al (2007)
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 10
Discussion
The vast majority of eye movement studies conductedin non-human species to date have not analyzed eyeposition data in the range of fixational eye movementsThus eye movements during fixation have been reportedin only a handful of vertebrates Most of these studieshave concentrated solely on the physical parameters ofthe fixational eye movements without addressing theirphysiological and perceptual consequences Fortunatelythis trend has started to reverse in recent years with agrowing list of papers addressing the effects of non-human fixational eye movements in physiology andperception This is an important development A largeamount of all psychophysical and physiological visualresearch has been conducted during conditions of visualfixation Therefore understanding the precise physiolog-ical and perceptual contributions of fixational eye move-ments may be critical to the interpretation of previous andfuture vision researchThere are some interesting differences and similarities
in the types of fixational eye movements produced bydifferent species For instance ocular tremor is possiblyubiquitous to all vertebrates The fact that tremor has notbeen observed in all species studied could be due to thetechnical difficulty of recording tremor accurately (ietremor amplitudes and frequencies are usually in the rangeof the recording systemrsquos noise) Drift has also beenrecorded in most vertebrates and it may be as omnipre-sent as tremor Conversely microsaccades seem mostimportant in species with foveal vision (ie primates andhumans) whereas they are scarce or absent in afoveatespecies (ie rabbits frogs goldfish) A link betweenmicrosaccades and foveal vision may reflect the two-pronged functional significance of microsaccades inhumans and primates first microsaccades may benecessary to correct the slow drift of the eye and bringback the objectregion of interest to the retinal area wherephotoreceptor density is highest (Cornsweet 1956) Thecorrective role of human microsaccades has been demon-strated in recent studies (Engbert amp Kliegl 2004 Engbertamp Mergenthaler 2006) Second the dynamics of micro-saccades themselves may improve spatial resolution(Donner amp Hemila 2005)In reviewing the patterns of fixational eye movements
across vertebrates it is tempting to draw parallels withother sensory modalities Fixational eye movementsrefresh the retinal images during fixation and thus preventneural adaptation and visual fading But this mechanism isnot circumscribed to the visual system Sniffs in rodentolfaction discretely sample sensory information every200ndash300 msec and thus are similar in their temporaldynamics to primate saccades (Otero-Millan et al 2008Uchida Kepecs amp Mainen 2006) and microsaccades(Otero-Millan et al 2008) A similar mode of discretesampling may also be at play when objects are recognized
through tactile information for instance if we sweep ourfingertips over an objectrsquos surface with our eyes closed orwhen blind individuals read Braille scriptOne intriguing possibility that merits further experi-
ments and analyses is the potential correlation between thespatiotemporal parameters of fixational eye movements(such as those summarized in Table 1) and the adaptationproperties of visual neurons across species For instancethe greater ocular stability of some species (rabbitgoldfish frog etc) versus others (primates) could beassociated with some particular adaptive properties oftheir visual cells Thus future comparative studies shouldpay attention to the response properties of visual cellstriggered by fixational eye movements in the presence ofstationary stimuli Other potential factors underlying thevariation in fixational eye movements across species couldbe differences in oculomotor control at the level of the eyemuscles or perhaps differences in retinal organizationFuture research will further improve our understanding
of fixational eye movements and the neural activity thatmaintains the visibility of stationary visual scenes inhuman and non-human species One interesting questionthat remains to be addressed concerns the role of tremor invision and visual physiology Because tremor frequenciesare well over the flicker fusion frequencies in humans ithas been argued that tremor of the visual image would beineffective as a stimulus (Ditchburn 1955 Gerrits ampVendrik 1970 Sharpe 1972) However recent studieshave found simulated tremor to enhance visual processingand improve visual acuity in a model retina (Hennig et al2002 Miller et al 2006) Follow-up research should beaimed to replicate these findings in actual neurons withcorrelated perceptual reports One should also note thatearly visual neurons can follow high-frequency flickeringthat surpasses the perceptual threshold for flicker fusion(Martinez-Conde et al 2002) Thus high-frequencytremor could be adequate to maintain activity in the earlyvisual system which then may lead to visual perception inhigher visual areas Another significant question thatremains unanswered concerns the oculomotor bases ofthe various types of fixational eye movements acrossspecies To date there is too little information on thisimportant topic to draw a conclusion
Conclusions
We have reviewed the parameters of fixational eyemovements in non-human mammals birds reptilesamphibians and fish as well as their consequences forphysiology and perception Fixational eye movementsand microsaccades in particular appear to be mostimportant in foveate vs afoveate species Animals lackingfixational eye movements such as afoveate frogs andtoads may be incapable of seeing stationary objects
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 11
during visual fixation Future research should furtherexplore the perceptual effects of fixational eye movementsand determine the oculomotor mechanisms responsible fortheir generation across species
Acknowledgments
We thank Mona Stewart and Isabel Gomez-Caraballo fortechnical assistance Jorge Otero-Millan for programmingassistance and Dr Xoana G Troncoso for comments on themanuscript This study was funded through grants fromthe Barrow Neurological Foundation (to SMC and SLM)the National Science Foundation (Award 0643306 toSMC) the Arizona Biomedical Research Commission(Award 07-102 to SMC) and Science Foundation Arizona(SLM)
Commercial relationships noneCorresponding author Susana Martinez-Conde PhDEmail smartneuralcorrelatecomAddress Barrow Neurological Institute 350 W ThomasRd Phoenix AZ 85013 USA
References
Aksay E Major G Goldman M S Baker R SeungH S amp Tank D W (2003) History dependence ofrate covariation between neurons during persistentactivity in an oculomotor integrator Cerebral Cortex13 1173ndash1184 [PubMed] [Article]
Aksay E Olasagasti I Mensh B D Baker RGoldman M S amp Tank D W (2007) Functionaldissection of circuitry in a neural integrator NatureNeuroscience 10 494ndash504 [PubMed]
Autrum H (1959) Das Fehlen unwillkurlicher Augenbe-wegungen beim Frosch Naturwissenschaften 46435
Baccus S A Olveczky B P Manu M amp Meister M(2008) A retinal circuit that computes object motionJournal of Neuroscience 28 6807ndash6817 [PubMed]
Bair W amp OrsquoKeefe L P (1998) The influence offixational eye movements on the response of neuronsin area MT of the macaque Visual Neuroscience 15779ndash786 [PubMed]
Betta E amp Turatto M (2006) Are you ready I can tellby looking at your microsaccades Neuroreport 171001ndash1004 [PubMed]
Bloch S Rivaud S amp Martinoya C (1984) Compar-ing frontal and lateral viewing in the pigeon IIIDifferent patterns of eye movements for binocularand monocular fixation Behavioural Brain Research13 173ndash182 [PubMed]
Cabrera B Torres B Pasaro R Pastor A M ampDelgado-Garcia J M (1992) A morphological studyof abducens nucleus motoneurons and internuclearneurons in the goldfish (Carassius auratus) BrainResearch Bulletin 28 137ndash144 [PubMed]
Carpenter R H S (1988) Movements of the eyes(2nd ed) Pion London
Collewijn H (1977) Eye- and head movements infreely moving rabbits The Journal of Physiology266 471ndash498 [PubMed] [Article]
Collewijn H amp van der Mark F (1972) Ocular stabilityin variable visual feedback conditions in the rabbitBrain Research 36 47ndash57 [PubMed]
Conway J L Timberlake G T amp Skavenski A A(1981) Oculomotor changes in cats reared withoutexperiencing continuous retinal image motion Exper-imental Brain Research 43 229ndash232 [PubMed]
Cornsweet T N (1956) Determination of the stimuli forinvoluntary drifts and saccadic eye movements Jour-nal of the Optical Society of America 46 987ndash993[PubMed]
Delgado-Garcia J M del Pozo F amp Baker R (1986)Behavior of neurons in the abducens nucleus of thealert catVII Internuclear neurons Neuroscience 17953ndash973 [PubMed]
Dieringer N amp Daunicht W J (1986) Image fadingVAproblem for frogs Naturwissenschaften 73 330ndash332[PubMed]
Dieringer N amp Precht W (1982) Compensatory headand eye movements in the frog and their contributionto stabilization of gaze Experimental BrainResearch 47 394ndash406 [PubMed]
Dieringer N Precht W amp Blight A R (1982)Resetting fast phases of head and eye and theirlinkage in the frog Experimental Brain Research 47407ndash416 [PubMed]
Ditchburn R W (1955) Eye-movements in relation toretinal action Optica Acta 1 171ndash176
Ditchburn R W amp Ginsborg B L (1952) Vision with astabilized retinal image Nature 170 36ndash37[PubMed]
Ditchburn R W amp Ginsborg B L (1953) Involuntaryeye movements during fixation The Journal ofPhysiology 119 1ndash17 [PubMed] [Article]
Donner K amp Hemila S (2007) Modelling the effect ofmicrosaccades on retinal responses to stationarycontrast patterns Vision Research 47 1166ndash1177[PubMed]
Donner K K amp Hemila S (2005) The possible purposeof microsaccades Paper presented at the EuropeanConference on Visual Perception A Coruna Spain
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 12
Easter S S Jr (1971) Spontaneous eye movements inrestrained goldfish Vision Research 11 333ndash342[PubMed]
Easter S S Jr Johns P R amp Heckenlively D R(1974) Horizontal compensatory eye movements ingoldfish (Carassius auratus) I The normal animalJournal of Comparative Physiology 92 23ndash35
Engbert R (2006) Microsaccades A microcosm forresearch on oculomotor control attention and visualperception Progress Brain Research 154 177ndash192[PubMed]
Engbert R amp Kliegl R (2003a) Binocular coordinationin microsaccades In R R J Hyona amp H Deubel(Eds) The mindrsquos eyes Cognitive and appliedaspects of eye movements (pp 103ndash117) OxfordUK Elsevier
Engbert R amp Kliegl R (2003b) Microsaccades uncoverthe orientation of covert attention Vision Research43 1035ndash1045 [PubMed]
Engbert R amp Kliegl R (2004) Microsaccades keep theeyesrsquo balance during fixation Psychological Science15 431ndash436 [PubMed]
Engbert R amp Mergenthaler K (2006) Microsaccadesare triggered by low retinal image slip Proceedingsof the National Academy of Sciences of the UnitedStates of America 103 7192ndash7197 [PubMed][Article]
Ewert J P amp Borchers H W (1974) Antwort vonretinalen Ganglienzellen bei freibeweglichen Kroten(Bufo Bufo L) Journal of Comparative and Physi-ology 92 117ndash130
Fuller J H (1980) Linkage of eye and head movementsin the alert rabbit Brain Research 194 219ndash222[PubMed]
Fuller J H (1981) Eye and head movements duringvestibular stimulation in the alert rabbit BrainResearch 205 363ndash381 [PubMed]
Galfano G Betta E amp Turatto M (2004) Inhibition ofreturn in microsaccades Experimental BrainResearch 159 400ndash404 [PubMed]
Gerrits H J amp Vendrik A J (1970) Artificial move-ments of a stabilized image Vision Research 101443ndash1456 [PubMed]
Gioanni H Bennis M amp Sansonetti A (1993) Visualand vestibular reflexes that stabilize gaze in thechameleon Visual Neuroscience 10 947ndash956[PubMed]
Goffart L Quinet J Chavane F amp Masson G S(2006) Influence of background illumination onfixation and visually guided saccades in the rhesusmonkey Vision Research 46 149ndash162 [PubMed]
Greschner M Bongard M Rujan P amp Ammermuller J(2002) Retinal ganglion cell synchronization by fixa-tional eye movements improves feature estimationNature Neuroscience 5 341ndash347 [PubMed]
Gur M Beylin A amp Snodderly D M (1997)Response variability of neurons in primary visualcortex (V1) of alert monkeys Journal of Neuro-science 17 2914ndash2920 [PubMed] [Article]
Hafed Z M amp Clark J J (2002) Microsaccades as anovert measure of covert attention shifts VisionResearch 42 2533ndash2545 [PubMed]
Hebbard F W amp Marg E (1960) Physiologicalnystagmus in the cat Journal of the Optical Societyof America 50 151ndash155 [PubMed]
Hennig M A Kerscher N J Funke K amp Worgotter F(2002) Stochastic resonance in visual cortical neuronsDoes the eye-tremor actually improve visual acuityNeurocomputing 44ndash46 115ndash120
Hennig M H amp Worgotter F (2004) Eye micro-movements improve stimulus detection beyond theNyquist limit in the peripheral retina In Advances inNeural Information Processing Systems (NIPS2003)(vol 16) Denver CO MIT Press
Hennig M H amp Worgotter F (2007) Effects offixational eye movements on retinal ganglion cellresponses A modelling study Frontiers in Computa-tional Neuroscience 1 2 [PubMed] [Article]
Hermann H T amp Constantine M M (1971) Eyemovements in the goldfish Vision Research 11313ndash331 [PubMed]
Horowitz T S Fine E M Fencsik D E Yurgenson Samp Wolfe J M (2007) Fixational eye movements arenot an index of covert attention PsychologicalScience 18 356ndash363 [PubMed]
Horwitz G D amp Albright T D (2003) Short-latencyfixational saccades induced by luminance incrementsJournal of Neurophysiology 90 1333ndash1339[PubMed] [Article]
Laubrock J Engbert R Rolfs M amp Kliegl R(2007) Microsaccades are an index of covertattention Commentary on Horowitz Fine FencsikYurgenson and Wolfe (2007) Psychological Sci-ence 18 364ndash366 discussion 367ndash368 [PubMed]
Leopold D A amp Logothetis N K (1998) Micro-saccades differentially modulate neural activity in thestriate and extrastriate visual cortex ExperimentalBrain Research 123 341ndash345 [PubMed]
Lettvin J Y Maturana H R MsCulloch W S amp PittsW H (1968) What the frogrsquos eye tells the frogrsquosbrain In W C C a M Balaban (Ed) The mind
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 13
Biological approaches to its functions (pp 233ndash258)John Wiley and Sons London
Macknik S L amp Livingstone M S (1998) Neuronalcorrelates of visibility and invisibility in the primatevisual system Nature Neuroscience 1 144ndash149[PubMed]
Macknik S L amp Martinez-Conde S (2004) The spatialand temporal effects of lateral inhibitory networksand their relevance to the visibility of spatiotemporaledges Neurocomputing 58ndash60 775ndash782
Macknik S L Martinez-Conde S amp Haglund M M(2000) The role of spatiotemporal edges in visibilityand visual masking Proceedings of the NationalAcademy of Sciences of the United States of America97 7556ndash7560 [PubMed] [Article]
Manteuffel G Plasa L Sommer T J amp Wess O(1977) Involuntary eye movements in salamandersNaturwissenschaften 64 533ndash534 [PubMed]
Marchetti E Gauthier G M amp Pellet J (1983)Cerebellar control of eye movements studied withinjection of harmaline in the trained baboon ArchivesItaliennes de Biologies 121 1ndash17 [PubMed]
Martinez-Conde S (2006) Fixational eye movements innormal and pathological vision Progress in BrainResearch 154 151ndash176 [PubMed]
Martinez-Conde S amp Macknik S L (2007a) Bobbingand perceiving Scientific American 297 16
Martinez-Conde S amp Macknik S L (2007b) Windowson the mind Scientific American 297 56ndash63[PubMed]
Martinez-Conde S Macknik S L amp Hubel D H(2000) Microsaccadic eye movements and firing ofsingle cells in the striate cortex of macaque monkeysNature Neuroscience 3 251ndash258 [PubMed]
Martinez-Conde S Macknik S L amp Hubel D H(2002) The function of bursts of spikes during visualfixation in the awake primate lateral geniculate nucleusand primary visual cortex Proceedings of theNational Academy of Sciences of the United Statesof America 99 13920ndash13925 [PubMed] [Article]
Martinez-Conde S Macknik S L amp Hubel D H(2004) The role of fixational eye movements invisual perception Nature Reviews Neuroscience 5229ndash240 [PubMed]
Martinez-Conde S Macknik S L Troncoso X G ampDyar T A (2006) Microsaccades counteract visualfading during fixation Neuron 49 297ndash305[PubMed] [Article]
Mensh B D Aksay E Lee D D Seung H S ampTank D W (2004) Spontaneous eye movements ingoldfish Oculomotor integrator performance plasti-
city and dependence on visual feedback VisionResearch 44 711ndash726 [PubMed]
Miller J A Denning K S George J S MarshakD W amp Kenyon G T (2006) A high frequencyresonance in the responses of retinal ganglion cells torapidly modulated stimuli A computer model VisualNeuroscience 23 779ndash794 [PubMed]
Montgomery J C McVean A R amp McCarthy D(1983) The effects of lowered temperature onspontaneous eye movements in a teleost fish Com-parative Biochemistry and Physiology A Compara-tive Physiology 75 363ndash368 [PubMed]
Motter B C amp Poggio G F (1984) Binocularfixation in the rhesus monkey Spatial and temporalcharacteristics Experimental Brain Research 54304ndash314 [PubMed]
Murakami I amp Cavanagh P (1998) A jitter after-effectreveals motion-based stabilization of vision Nature395 798ndash801 [PubMed]
Nye P W (1969) The monocular eye movements of thepigeon Vision Research 9 133ndash144 [PubMed]
Olveczky B P Baccus S A amp Meister M (2003)Segregation of object and background motion in theretina Nature 423 401ndash408 [PubMed]
Olveczky B P Baccus S A amp Meister M (2007)Retinal adaptation to object motion Neuron 56689ndash700 [PubMed] [Article]
Otero-Millan J Troncoso X Macknik S L Serrano-Pedraza I amp Martinez-Conde S (2008) Saccadesand microsaccades during visual fixation explorationand search Foundations for a common saccadicgenerator Journal of Vision 8(14)21 1ndash18 httpjournalofvisionorg81421 doi10116781421[PubMed] [Article]
Pastor A M Torres B Delgado-Garcia J M ampBaker R (1991) Discharge characteristics of medialrectus and abducens motoneurons in the goldfishJournal of Neurophysiology 66 2125ndash2140 [PubMed]
Pettigrew J D Wallman J amp Wildsoet C F (1990)Saccadic oscillations facilitate ocular perfusion fromthe avian pecten Nature 343 362ndash363 [PubMed]
Pritchard R M amp Heron W (1960) Small eye move-ments of the cat Canadian Journal of Psychology14 131ndash137 [PubMed]
Ratliff F amp Riggs L A (1950) Involuntary motions ofthe eye during monocular fixation Journal of Exper-imental Psychology 40 687ndash701 [PubMed]
Reppas J B UsreyWM amp Reid R C (2002) Saccadiceye movements modulate visual responses in thelateral geniculate nucleus Neuron 35 961ndash974[PubMed] [Article]
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 14
Riggs L A amp Ratliff F (1952) The effects of counter-acting the normal movements of the eye Journal ofthe Optical Society of America 42 872ndash873
Rolfs M Kliegl R amp Engbert R (2008) Toward amodel of microsaccade generation The case ofmicrosaccadic inhibition Journal of Vision 8(11)51ndash23 httpjournalofvisionorg8115 doi1011678115 [PubMed] [Article]
Rolfs M Laubrock J amp Kliegl R (2006) Shorteningand prolongation of saccade latencies followingmicrosaccades Experimental Brain Research 169369ndash376 [PubMed]
Rucci M Iovin R Poletti M amp Santini F (2007)Miniature eye movements enhance fine spatial detailNature 447 851ndash854 [PubMed]
Salas C Navarro F Torres B amp Delgado-Garcia J M(1992) Effects of diazepam and D-amphetamine onrhythmic pattern of eye movements in goldfishNeuroreport 3 131ndash134 [PubMed]
Schipperheyn J J (1963) Respiratory eye movementsand perception of stationary objects in the frog ActaPhysiologica et Pharmacologica Neerlandica 12157ndash159 [PubMed]
Schipperheyn J J (1965) Contrast detection in frogrsquosretina Acta Physiologica et Pharmacologica Neer-landica 13 231ndash277 [PubMed]
Schipperheyn J J (1967) Influence of eye movements onretinal ganglion cell activity in frogs Acta Physi-ologica et Pharmacologica Neerlandica 14 506[PubMed]
Segev R Schneidman E Goodhouse J amp Berry M J2nd (2007) Role of eye movements in the retinalcode for a size discrimination task Journal ofNeurophysiology 98 1380ndash1391 [PubMed] [Article]
Sharpe C R (1972) The visibility and fading of thinlines visualized by their controlled movement acrossthe retina The Journal of Physiology 222 113ndash134[PubMed] [Article]
Shelley M McLaughlin D Shapley R amp Wielaard J(2002) States of high conductance in a large-scalemodel of the visual cortex Journal of ComputationalNeuroscience 13 93ndash109 [PubMed]
Skavenski A A Robinson D A Steinman R M ampTimberlake G T (1975) Miniature eye movementsof fixation in rhesus monkey Vision Research 151269ndash1273 [PubMed]
Snodderly D M (1987) Effects of light and darkenvironments on macaque and human fixational eyemovements Vision Research 27 401ndash415 [PubMed]
Snodderly D M Kagan I amp Gur M (2001) Selectiveactivation of visual cortex neurons by fixational eyemovements Implications for neural coding VisualNeuroscience 18 259ndash277 [PubMed]
Snodderly D M amp Kurtz D (1985) Eye position duringfixation tasks Comparison of macaque and humanVision Research 25 83ndash98 [PubMed]
Steinbach M J amp Money K E (1973) Eye movementsof the owl Vision Research 13 889ndash891 [PubMed]
Steinman R M Haddad G M Skavenski A A ampWyman D (1973) Miniature eye movement Science181 810ndash819 [PubMed]
Torres B Pastor A M Cabrera B Salas C ampDelgado-Garcia J M (1992) Afferents to theoculomotor nucleus in the goldfish (Carassius aur-atus) as revealed by retrograde labeling with horse-radish peroxidase Journal of Comparative Neurology324 449ndash461 [PubMed]
Troncoso X G Macknik S L amp Martinez-Conde S(2008) Microsaccades counteract perceptual filling-inJournal of Vision 8(14)15 1ndash9 httpjournalofvisionorg81415 doi10116781415 [Article]
Troncoso X G Macknik S L Otero-Millan J ampMartinez-Conde S (2008) Microsaccades driveillusory motion in the Enigma illusion Proceedingsof the National Academy of Sciences of the UnitedStates of America 105 16033ndash16038 [PubMed]
Tse P U Sheinberg D S amp Logothetis N K (2004)The distribution of microsaccade directions need notreveal the location of attention Psychological Science15 708ndash710
Turatto M Valsecchi M Tame L amp Betta E (2007)Microsaccades distinguish between global and localvisual processing Neuroreport 18 1015ndash1018[PubMed]
Uchida N Kepecs A amp Mainen Z F (2006) Seeing ata glance smelling in a whiff Rapid forms ofperceptual decision making Nature Reviews Neuro-science 7 485ndash491 [PubMed]
Valsecchi M Betta E amp Turatto M (2007) Visualoddballs induce prolonged microsaccadic inhibitionExperimental Brain Research 177 196ndash208[PubMed]
van Dam L C amp van Ee R (2006) Retinal image shiftsbut not eye movements per se cause alternations inawareness during binocular rivalry Journal of Vision6(11)3 1172ndash1179 httpjournalofvisionorg6113doi1011676113 [PubMed] [Article]
Van der Steen J amp Collewijn H (1984) Ocular stabilityin the horizontal frontal and sagittal planes in therabbit Experimental Brain Research 56 263ndash274[PubMed]
Van Gisbergen J A Robinson D A amp Gielen S(1981) A quantitative analysis of generation ofsaccadic eye movements by burst neurons Journalof Neurophysiology 45 417ndash442 [PubMed]
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 15
Van Gisbergen J A M amp Robinson D A (1977)Generation of micro and macrosaccades by burstneurons in the monkey In R Baker amp A Berthoz(Eds) Control of gaze by brain stem neurons (vol 1pp 301ndash308) New York ElsevierNorth-Holland
Wallman J Pettigrew J D amp Letelier J C (1994)The oscillatory saccades of birds Motoneural activ-ity and possible functions In B T Fuchs A FV Buttner amp D Zee (Eds) Contemporary ocularmotor and vestibular research A tribute to David ARobinson (pp 480ndash487) Stuttgart Germany ThiemePublishing
Williams P E amp Shapley R M (2007) A dynamicnonlinearity and spatial phase specificity in macaque
V1 neurons Journal of Neuroscience 27 5706ndash5718[PubMed] [Article]
Winterson B J amp Robinson D A (1975) Fixationby the alert but solitary cat Vision Research 151349ndash1352 [PubMed]
Yarbus A L (1967) Eye movements and vision(B Haigh Trans) New York Plenum Press
Zuber B L amp Stark L (1965) Microsaccades and thevelocityltamplitude relationship for saccadic eyemovements Science 150 1459ndash1460 [PubMed]
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 16

Discussion
The vast majority of eye movement studies conductedin non-human species to date have not analyzed eyeposition data in the range of fixational eye movementsThus eye movements during fixation have been reportedin only a handful of vertebrates Most of these studieshave concentrated solely on the physical parameters ofthe fixational eye movements without addressing theirphysiological and perceptual consequences Fortunatelythis trend has started to reverse in recent years with agrowing list of papers addressing the effects of non-human fixational eye movements in physiology andperception This is an important development A largeamount of all psychophysical and physiological visualresearch has been conducted during conditions of visualfixation Therefore understanding the precise physiolog-ical and perceptual contributions of fixational eye move-ments may be critical to the interpretation of previous andfuture vision researchThere are some interesting differences and similarities
in the types of fixational eye movements produced bydifferent species For instance ocular tremor is possiblyubiquitous to all vertebrates The fact that tremor has notbeen observed in all species studied could be due to thetechnical difficulty of recording tremor accurately (ietremor amplitudes and frequencies are usually in the rangeof the recording systemrsquos noise) Drift has also beenrecorded in most vertebrates and it may be as omnipre-sent as tremor Conversely microsaccades seem mostimportant in species with foveal vision (ie primates andhumans) whereas they are scarce or absent in afoveatespecies (ie rabbits frogs goldfish) A link betweenmicrosaccades and foveal vision may reflect the two-pronged functional significance of microsaccades inhumans and primates first microsaccades may benecessary to correct the slow drift of the eye and bringback the objectregion of interest to the retinal area wherephotoreceptor density is highest (Cornsweet 1956) Thecorrective role of human microsaccades has been demon-strated in recent studies (Engbert amp Kliegl 2004 Engbertamp Mergenthaler 2006) Second the dynamics of micro-saccades themselves may improve spatial resolution(Donner amp Hemila 2005)In reviewing the patterns of fixational eye movements
across vertebrates it is tempting to draw parallels withother sensory modalities Fixational eye movementsrefresh the retinal images during fixation and thus preventneural adaptation and visual fading But this mechanism isnot circumscribed to the visual system Sniffs in rodentolfaction discretely sample sensory information every200ndash300 msec and thus are similar in their temporaldynamics to primate saccades (Otero-Millan et al 2008Uchida Kepecs amp Mainen 2006) and microsaccades(Otero-Millan et al 2008) A similar mode of discretesampling may also be at play when objects are recognized
through tactile information for instance if we sweep ourfingertips over an objectrsquos surface with our eyes closed orwhen blind individuals read Braille scriptOne intriguing possibility that merits further experi-
ments and analyses is the potential correlation between thespatiotemporal parameters of fixational eye movements(such as those summarized in Table 1) and the adaptationproperties of visual neurons across species For instancethe greater ocular stability of some species (rabbitgoldfish frog etc) versus others (primates) could beassociated with some particular adaptive properties oftheir visual cells Thus future comparative studies shouldpay attention to the response properties of visual cellstriggered by fixational eye movements in the presence ofstationary stimuli Other potential factors underlying thevariation in fixational eye movements across species couldbe differences in oculomotor control at the level of the eyemuscles or perhaps differences in retinal organizationFuture research will further improve our understanding
of fixational eye movements and the neural activity thatmaintains the visibility of stationary visual scenes inhuman and non-human species One interesting questionthat remains to be addressed concerns the role of tremor invision and visual physiology Because tremor frequenciesare well over the flicker fusion frequencies in humans ithas been argued that tremor of the visual image would beineffective as a stimulus (Ditchburn 1955 Gerrits ampVendrik 1970 Sharpe 1972) However recent studieshave found simulated tremor to enhance visual processingand improve visual acuity in a model retina (Hennig et al2002 Miller et al 2006) Follow-up research should beaimed to replicate these findings in actual neurons withcorrelated perceptual reports One should also note thatearly visual neurons can follow high-frequency flickeringthat surpasses the perceptual threshold for flicker fusion(Martinez-Conde et al 2002) Thus high-frequencytremor could be adequate to maintain activity in the earlyvisual system which then may lead to visual perception inhigher visual areas Another significant question thatremains unanswered concerns the oculomotor bases ofthe various types of fixational eye movements acrossspecies To date there is too little information on thisimportant topic to draw a conclusion
Conclusions
We have reviewed the parameters of fixational eyemovements in non-human mammals birds reptilesamphibians and fish as well as their consequences forphysiology and perception Fixational eye movementsand microsaccades in particular appear to be mostimportant in foveate vs afoveate species Animals lackingfixational eye movements such as afoveate frogs andtoads may be incapable of seeing stationary objects
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 11
during visual fixation Future research should furtherexplore the perceptual effects of fixational eye movementsand determine the oculomotor mechanisms responsible fortheir generation across species
Acknowledgments
We thank Mona Stewart and Isabel Gomez-Caraballo fortechnical assistance Jorge Otero-Millan for programmingassistance and Dr Xoana G Troncoso for comments on themanuscript This study was funded through grants fromthe Barrow Neurological Foundation (to SMC and SLM)the National Science Foundation (Award 0643306 toSMC) the Arizona Biomedical Research Commission(Award 07-102 to SMC) and Science Foundation Arizona(SLM)
Commercial relationships noneCorresponding author Susana Martinez-Conde PhDEmail smartneuralcorrelatecomAddress Barrow Neurological Institute 350 W ThomasRd Phoenix AZ 85013 USA
References
Aksay E Major G Goldman M S Baker R SeungH S amp Tank D W (2003) History dependence ofrate covariation between neurons during persistentactivity in an oculomotor integrator Cerebral Cortex13 1173ndash1184 [PubMed] [Article]
Aksay E Olasagasti I Mensh B D Baker RGoldman M S amp Tank D W (2007) Functionaldissection of circuitry in a neural integrator NatureNeuroscience 10 494ndash504 [PubMed]
Autrum H (1959) Das Fehlen unwillkurlicher Augenbe-wegungen beim Frosch Naturwissenschaften 46435
Baccus S A Olveczky B P Manu M amp Meister M(2008) A retinal circuit that computes object motionJournal of Neuroscience 28 6807ndash6817 [PubMed]
Bair W amp OrsquoKeefe L P (1998) The influence offixational eye movements on the response of neuronsin area MT of the macaque Visual Neuroscience 15779ndash786 [PubMed]
Betta E amp Turatto M (2006) Are you ready I can tellby looking at your microsaccades Neuroreport 171001ndash1004 [PubMed]
Bloch S Rivaud S amp Martinoya C (1984) Compar-ing frontal and lateral viewing in the pigeon IIIDifferent patterns of eye movements for binocularand monocular fixation Behavioural Brain Research13 173ndash182 [PubMed]
Cabrera B Torres B Pasaro R Pastor A M ampDelgado-Garcia J M (1992) A morphological studyof abducens nucleus motoneurons and internuclearneurons in the goldfish (Carassius auratus) BrainResearch Bulletin 28 137ndash144 [PubMed]
Carpenter R H S (1988) Movements of the eyes(2nd ed) Pion London
Collewijn H (1977) Eye- and head movements infreely moving rabbits The Journal of Physiology266 471ndash498 [PubMed] [Article]
Collewijn H amp van der Mark F (1972) Ocular stabilityin variable visual feedback conditions in the rabbitBrain Research 36 47ndash57 [PubMed]
Conway J L Timberlake G T amp Skavenski A A(1981) Oculomotor changes in cats reared withoutexperiencing continuous retinal image motion Exper-imental Brain Research 43 229ndash232 [PubMed]
Cornsweet T N (1956) Determination of the stimuli forinvoluntary drifts and saccadic eye movements Jour-nal of the Optical Society of America 46 987ndash993[PubMed]
Delgado-Garcia J M del Pozo F amp Baker R (1986)Behavior of neurons in the abducens nucleus of thealert catVII Internuclear neurons Neuroscience 17953ndash973 [PubMed]
Dieringer N amp Daunicht W J (1986) Image fadingVAproblem for frogs Naturwissenschaften 73 330ndash332[PubMed]
Dieringer N amp Precht W (1982) Compensatory headand eye movements in the frog and their contributionto stabilization of gaze Experimental BrainResearch 47 394ndash406 [PubMed]
Dieringer N Precht W amp Blight A R (1982)Resetting fast phases of head and eye and theirlinkage in the frog Experimental Brain Research 47407ndash416 [PubMed]
Ditchburn R W (1955) Eye-movements in relation toretinal action Optica Acta 1 171ndash176
Ditchburn R W amp Ginsborg B L (1952) Vision with astabilized retinal image Nature 170 36ndash37[PubMed]
Ditchburn R W amp Ginsborg B L (1953) Involuntaryeye movements during fixation The Journal ofPhysiology 119 1ndash17 [PubMed] [Article]
Donner K amp Hemila S (2007) Modelling the effect ofmicrosaccades on retinal responses to stationarycontrast patterns Vision Research 47 1166ndash1177[PubMed]
Donner K K amp Hemila S (2005) The possible purposeof microsaccades Paper presented at the EuropeanConference on Visual Perception A Coruna Spain
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 12
Easter S S Jr (1971) Spontaneous eye movements inrestrained goldfish Vision Research 11 333ndash342[PubMed]
Easter S S Jr Johns P R amp Heckenlively D R(1974) Horizontal compensatory eye movements ingoldfish (Carassius auratus) I The normal animalJournal of Comparative Physiology 92 23ndash35
Engbert R (2006) Microsaccades A microcosm forresearch on oculomotor control attention and visualperception Progress Brain Research 154 177ndash192[PubMed]
Engbert R amp Kliegl R (2003a) Binocular coordinationin microsaccades In R R J Hyona amp H Deubel(Eds) The mindrsquos eyes Cognitive and appliedaspects of eye movements (pp 103ndash117) OxfordUK Elsevier
Engbert R amp Kliegl R (2003b) Microsaccades uncoverthe orientation of covert attention Vision Research43 1035ndash1045 [PubMed]
Engbert R amp Kliegl R (2004) Microsaccades keep theeyesrsquo balance during fixation Psychological Science15 431ndash436 [PubMed]
Engbert R amp Mergenthaler K (2006) Microsaccadesare triggered by low retinal image slip Proceedingsof the National Academy of Sciences of the UnitedStates of America 103 7192ndash7197 [PubMed][Article]
Ewert J P amp Borchers H W (1974) Antwort vonretinalen Ganglienzellen bei freibeweglichen Kroten(Bufo Bufo L) Journal of Comparative and Physi-ology 92 117ndash130
Fuller J H (1980) Linkage of eye and head movementsin the alert rabbit Brain Research 194 219ndash222[PubMed]
Fuller J H (1981) Eye and head movements duringvestibular stimulation in the alert rabbit BrainResearch 205 363ndash381 [PubMed]
Galfano G Betta E amp Turatto M (2004) Inhibition ofreturn in microsaccades Experimental BrainResearch 159 400ndash404 [PubMed]
Gerrits H J amp Vendrik A J (1970) Artificial move-ments of a stabilized image Vision Research 101443ndash1456 [PubMed]
Gioanni H Bennis M amp Sansonetti A (1993) Visualand vestibular reflexes that stabilize gaze in thechameleon Visual Neuroscience 10 947ndash956[PubMed]
Goffart L Quinet J Chavane F amp Masson G S(2006) Influence of background illumination onfixation and visually guided saccades in the rhesusmonkey Vision Research 46 149ndash162 [PubMed]
Greschner M Bongard M Rujan P amp Ammermuller J(2002) Retinal ganglion cell synchronization by fixa-tional eye movements improves feature estimationNature Neuroscience 5 341ndash347 [PubMed]
Gur M Beylin A amp Snodderly D M (1997)Response variability of neurons in primary visualcortex (V1) of alert monkeys Journal of Neuro-science 17 2914ndash2920 [PubMed] [Article]
Hafed Z M amp Clark J J (2002) Microsaccades as anovert measure of covert attention shifts VisionResearch 42 2533ndash2545 [PubMed]
Hebbard F W amp Marg E (1960) Physiologicalnystagmus in the cat Journal of the Optical Societyof America 50 151ndash155 [PubMed]
Hennig M A Kerscher N J Funke K amp Worgotter F(2002) Stochastic resonance in visual cortical neuronsDoes the eye-tremor actually improve visual acuityNeurocomputing 44ndash46 115ndash120
Hennig M H amp Worgotter F (2004) Eye micro-movements improve stimulus detection beyond theNyquist limit in the peripheral retina In Advances inNeural Information Processing Systems (NIPS2003)(vol 16) Denver CO MIT Press
Hennig M H amp Worgotter F (2007) Effects offixational eye movements on retinal ganglion cellresponses A modelling study Frontiers in Computa-tional Neuroscience 1 2 [PubMed] [Article]
Hermann H T amp Constantine M M (1971) Eyemovements in the goldfish Vision Research 11313ndash331 [PubMed]
Horowitz T S Fine E M Fencsik D E Yurgenson Samp Wolfe J M (2007) Fixational eye movements arenot an index of covert attention PsychologicalScience 18 356ndash363 [PubMed]
Horwitz G D amp Albright T D (2003) Short-latencyfixational saccades induced by luminance incrementsJournal of Neurophysiology 90 1333ndash1339[PubMed] [Article]
Laubrock J Engbert R Rolfs M amp Kliegl R(2007) Microsaccades are an index of covertattention Commentary on Horowitz Fine FencsikYurgenson and Wolfe (2007) Psychological Sci-ence 18 364ndash366 discussion 367ndash368 [PubMed]
Leopold D A amp Logothetis N K (1998) Micro-saccades differentially modulate neural activity in thestriate and extrastriate visual cortex ExperimentalBrain Research 123 341ndash345 [PubMed]
Lettvin J Y Maturana H R MsCulloch W S amp PittsW H (1968) What the frogrsquos eye tells the frogrsquosbrain In W C C a M Balaban (Ed) The mind
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 13
Biological approaches to its functions (pp 233ndash258)John Wiley and Sons London
Macknik S L amp Livingstone M S (1998) Neuronalcorrelates of visibility and invisibility in the primatevisual system Nature Neuroscience 1 144ndash149[PubMed]
Macknik S L amp Martinez-Conde S (2004) The spatialand temporal effects of lateral inhibitory networksand their relevance to the visibility of spatiotemporaledges Neurocomputing 58ndash60 775ndash782
Macknik S L Martinez-Conde S amp Haglund M M(2000) The role of spatiotemporal edges in visibilityand visual masking Proceedings of the NationalAcademy of Sciences of the United States of America97 7556ndash7560 [PubMed] [Article]
Manteuffel G Plasa L Sommer T J amp Wess O(1977) Involuntary eye movements in salamandersNaturwissenschaften 64 533ndash534 [PubMed]
Marchetti E Gauthier G M amp Pellet J (1983)Cerebellar control of eye movements studied withinjection of harmaline in the trained baboon ArchivesItaliennes de Biologies 121 1ndash17 [PubMed]
Martinez-Conde S (2006) Fixational eye movements innormal and pathological vision Progress in BrainResearch 154 151ndash176 [PubMed]
Martinez-Conde S amp Macknik S L (2007a) Bobbingand perceiving Scientific American 297 16
Martinez-Conde S amp Macknik S L (2007b) Windowson the mind Scientific American 297 56ndash63[PubMed]
Martinez-Conde S Macknik S L amp Hubel D H(2000) Microsaccadic eye movements and firing ofsingle cells in the striate cortex of macaque monkeysNature Neuroscience 3 251ndash258 [PubMed]
Martinez-Conde S Macknik S L amp Hubel D H(2002) The function of bursts of spikes during visualfixation in the awake primate lateral geniculate nucleusand primary visual cortex Proceedings of theNational Academy of Sciences of the United Statesof America 99 13920ndash13925 [PubMed] [Article]
Martinez-Conde S Macknik S L amp Hubel D H(2004) The role of fixational eye movements invisual perception Nature Reviews Neuroscience 5229ndash240 [PubMed]
Martinez-Conde S Macknik S L Troncoso X G ampDyar T A (2006) Microsaccades counteract visualfading during fixation Neuron 49 297ndash305[PubMed] [Article]
Mensh B D Aksay E Lee D D Seung H S ampTank D W (2004) Spontaneous eye movements ingoldfish Oculomotor integrator performance plasti-
city and dependence on visual feedback VisionResearch 44 711ndash726 [PubMed]
Miller J A Denning K S George J S MarshakD W amp Kenyon G T (2006) A high frequencyresonance in the responses of retinal ganglion cells torapidly modulated stimuli A computer model VisualNeuroscience 23 779ndash794 [PubMed]
Montgomery J C McVean A R amp McCarthy D(1983) The effects of lowered temperature onspontaneous eye movements in a teleost fish Com-parative Biochemistry and Physiology A Compara-tive Physiology 75 363ndash368 [PubMed]
Motter B C amp Poggio G F (1984) Binocularfixation in the rhesus monkey Spatial and temporalcharacteristics Experimental Brain Research 54304ndash314 [PubMed]
Murakami I amp Cavanagh P (1998) A jitter after-effectreveals motion-based stabilization of vision Nature395 798ndash801 [PubMed]
Nye P W (1969) The monocular eye movements of thepigeon Vision Research 9 133ndash144 [PubMed]
Olveczky B P Baccus S A amp Meister M (2003)Segregation of object and background motion in theretina Nature 423 401ndash408 [PubMed]
Olveczky B P Baccus S A amp Meister M (2007)Retinal adaptation to object motion Neuron 56689ndash700 [PubMed] [Article]
Otero-Millan J Troncoso X Macknik S L Serrano-Pedraza I amp Martinez-Conde S (2008) Saccadesand microsaccades during visual fixation explorationand search Foundations for a common saccadicgenerator Journal of Vision 8(14)21 1ndash18 httpjournalofvisionorg81421 doi10116781421[PubMed] [Article]
Pastor A M Torres B Delgado-Garcia J M ampBaker R (1991) Discharge characteristics of medialrectus and abducens motoneurons in the goldfishJournal of Neurophysiology 66 2125ndash2140 [PubMed]
Pettigrew J D Wallman J amp Wildsoet C F (1990)Saccadic oscillations facilitate ocular perfusion fromthe avian pecten Nature 343 362ndash363 [PubMed]
Pritchard R M amp Heron W (1960) Small eye move-ments of the cat Canadian Journal of Psychology14 131ndash137 [PubMed]
Ratliff F amp Riggs L A (1950) Involuntary motions ofthe eye during monocular fixation Journal of Exper-imental Psychology 40 687ndash701 [PubMed]
Reppas J B UsreyWM amp Reid R C (2002) Saccadiceye movements modulate visual responses in thelateral geniculate nucleus Neuron 35 961ndash974[PubMed] [Article]
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 14
Riggs L A amp Ratliff F (1952) The effects of counter-acting the normal movements of the eye Journal ofthe Optical Society of America 42 872ndash873
Rolfs M Kliegl R amp Engbert R (2008) Toward amodel of microsaccade generation The case ofmicrosaccadic inhibition Journal of Vision 8(11)51ndash23 httpjournalofvisionorg8115 doi1011678115 [PubMed] [Article]
Rolfs M Laubrock J amp Kliegl R (2006) Shorteningand prolongation of saccade latencies followingmicrosaccades Experimental Brain Research 169369ndash376 [PubMed]
Rucci M Iovin R Poletti M amp Santini F (2007)Miniature eye movements enhance fine spatial detailNature 447 851ndash854 [PubMed]
Salas C Navarro F Torres B amp Delgado-Garcia J M(1992) Effects of diazepam and D-amphetamine onrhythmic pattern of eye movements in goldfishNeuroreport 3 131ndash134 [PubMed]
Schipperheyn J J (1963) Respiratory eye movementsand perception of stationary objects in the frog ActaPhysiologica et Pharmacologica Neerlandica 12157ndash159 [PubMed]
Schipperheyn J J (1965) Contrast detection in frogrsquosretina Acta Physiologica et Pharmacologica Neer-landica 13 231ndash277 [PubMed]
Schipperheyn J J (1967) Influence of eye movements onretinal ganglion cell activity in frogs Acta Physi-ologica et Pharmacologica Neerlandica 14 506[PubMed]
Segev R Schneidman E Goodhouse J amp Berry M J2nd (2007) Role of eye movements in the retinalcode for a size discrimination task Journal ofNeurophysiology 98 1380ndash1391 [PubMed] [Article]
Sharpe C R (1972) The visibility and fading of thinlines visualized by their controlled movement acrossthe retina The Journal of Physiology 222 113ndash134[PubMed] [Article]
Shelley M McLaughlin D Shapley R amp Wielaard J(2002) States of high conductance in a large-scalemodel of the visual cortex Journal of ComputationalNeuroscience 13 93ndash109 [PubMed]
Skavenski A A Robinson D A Steinman R M ampTimberlake G T (1975) Miniature eye movementsof fixation in rhesus monkey Vision Research 151269ndash1273 [PubMed]
Snodderly D M (1987) Effects of light and darkenvironments on macaque and human fixational eyemovements Vision Research 27 401ndash415 [PubMed]
Snodderly D M Kagan I amp Gur M (2001) Selectiveactivation of visual cortex neurons by fixational eyemovements Implications for neural coding VisualNeuroscience 18 259ndash277 [PubMed]
Snodderly D M amp Kurtz D (1985) Eye position duringfixation tasks Comparison of macaque and humanVision Research 25 83ndash98 [PubMed]
Steinbach M J amp Money K E (1973) Eye movementsof the owl Vision Research 13 889ndash891 [PubMed]
Steinman R M Haddad G M Skavenski A A ampWyman D (1973) Miniature eye movement Science181 810ndash819 [PubMed]
Torres B Pastor A M Cabrera B Salas C ampDelgado-Garcia J M (1992) Afferents to theoculomotor nucleus in the goldfish (Carassius aur-atus) as revealed by retrograde labeling with horse-radish peroxidase Journal of Comparative Neurology324 449ndash461 [PubMed]
Troncoso X G Macknik S L amp Martinez-Conde S(2008) Microsaccades counteract perceptual filling-inJournal of Vision 8(14)15 1ndash9 httpjournalofvisionorg81415 doi10116781415 [Article]
Troncoso X G Macknik S L Otero-Millan J ampMartinez-Conde S (2008) Microsaccades driveillusory motion in the Enigma illusion Proceedingsof the National Academy of Sciences of the UnitedStates of America 105 16033ndash16038 [PubMed]
Tse P U Sheinberg D S amp Logothetis N K (2004)The distribution of microsaccade directions need notreveal the location of attention Psychological Science15 708ndash710
Turatto M Valsecchi M Tame L amp Betta E (2007)Microsaccades distinguish between global and localvisual processing Neuroreport 18 1015ndash1018[PubMed]
Uchida N Kepecs A amp Mainen Z F (2006) Seeing ata glance smelling in a whiff Rapid forms ofperceptual decision making Nature Reviews Neuro-science 7 485ndash491 [PubMed]
Valsecchi M Betta E amp Turatto M (2007) Visualoddballs induce prolonged microsaccadic inhibitionExperimental Brain Research 177 196ndash208[PubMed]
van Dam L C amp van Ee R (2006) Retinal image shiftsbut not eye movements per se cause alternations inawareness during binocular rivalry Journal of Vision6(11)3 1172ndash1179 httpjournalofvisionorg6113doi1011676113 [PubMed] [Article]
Van der Steen J amp Collewijn H (1984) Ocular stabilityin the horizontal frontal and sagittal planes in therabbit Experimental Brain Research 56 263ndash274[PubMed]
Van Gisbergen J A Robinson D A amp Gielen S(1981) A quantitative analysis of generation ofsaccadic eye movements by burst neurons Journalof Neurophysiology 45 417ndash442 [PubMed]
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 15
Van Gisbergen J A M amp Robinson D A (1977)Generation of micro and macrosaccades by burstneurons in the monkey In R Baker amp A Berthoz(Eds) Control of gaze by brain stem neurons (vol 1pp 301ndash308) New York ElsevierNorth-Holland
Wallman J Pettigrew J D amp Letelier J C (1994)The oscillatory saccades of birds Motoneural activ-ity and possible functions In B T Fuchs A FV Buttner amp D Zee (Eds) Contemporary ocularmotor and vestibular research A tribute to David ARobinson (pp 480ndash487) Stuttgart Germany ThiemePublishing
Williams P E amp Shapley R M (2007) A dynamicnonlinearity and spatial phase specificity in macaque
V1 neurons Journal of Neuroscience 27 5706ndash5718[PubMed] [Article]
Winterson B J amp Robinson D A (1975) Fixationby the alert but solitary cat Vision Research 151349ndash1352 [PubMed]
Yarbus A L (1967) Eye movements and vision(B Haigh Trans) New York Plenum Press
Zuber B L amp Stark L (1965) Microsaccades and thevelocityltamplitude relationship for saccadic eyemovements Science 150 1459ndash1460 [PubMed]
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 16

during visual fixation Future research should furtherexplore the perceptual effects of fixational eye movementsand determine the oculomotor mechanisms responsible fortheir generation across species
Acknowledgments
We thank Mona Stewart and Isabel Gomez-Caraballo fortechnical assistance Jorge Otero-Millan for programmingassistance and Dr Xoana G Troncoso for comments on themanuscript This study was funded through grants fromthe Barrow Neurological Foundation (to SMC and SLM)the National Science Foundation (Award 0643306 toSMC) the Arizona Biomedical Research Commission(Award 07-102 to SMC) and Science Foundation Arizona(SLM)
Commercial relationships noneCorresponding author Susana Martinez-Conde PhDEmail smartneuralcorrelatecomAddress Barrow Neurological Institute 350 W ThomasRd Phoenix AZ 85013 USA
References
Aksay E Major G Goldman M S Baker R SeungH S amp Tank D W (2003) History dependence ofrate covariation between neurons during persistentactivity in an oculomotor integrator Cerebral Cortex13 1173ndash1184 [PubMed] [Article]
Aksay E Olasagasti I Mensh B D Baker RGoldman M S amp Tank D W (2007) Functionaldissection of circuitry in a neural integrator NatureNeuroscience 10 494ndash504 [PubMed]
Autrum H (1959) Das Fehlen unwillkurlicher Augenbe-wegungen beim Frosch Naturwissenschaften 46435
Baccus S A Olveczky B P Manu M amp Meister M(2008) A retinal circuit that computes object motionJournal of Neuroscience 28 6807ndash6817 [PubMed]
Bair W amp OrsquoKeefe L P (1998) The influence offixational eye movements on the response of neuronsin area MT of the macaque Visual Neuroscience 15779ndash786 [PubMed]
Betta E amp Turatto M (2006) Are you ready I can tellby looking at your microsaccades Neuroreport 171001ndash1004 [PubMed]
Bloch S Rivaud S amp Martinoya C (1984) Compar-ing frontal and lateral viewing in the pigeon IIIDifferent patterns of eye movements for binocularand monocular fixation Behavioural Brain Research13 173ndash182 [PubMed]
Cabrera B Torres B Pasaro R Pastor A M ampDelgado-Garcia J M (1992) A morphological studyof abducens nucleus motoneurons and internuclearneurons in the goldfish (Carassius auratus) BrainResearch Bulletin 28 137ndash144 [PubMed]
Carpenter R H S (1988) Movements of the eyes(2nd ed) Pion London
Collewijn H (1977) Eye- and head movements infreely moving rabbits The Journal of Physiology266 471ndash498 [PubMed] [Article]
Collewijn H amp van der Mark F (1972) Ocular stabilityin variable visual feedback conditions in the rabbitBrain Research 36 47ndash57 [PubMed]
Conway J L Timberlake G T amp Skavenski A A(1981) Oculomotor changes in cats reared withoutexperiencing continuous retinal image motion Exper-imental Brain Research 43 229ndash232 [PubMed]
Cornsweet T N (1956) Determination of the stimuli forinvoluntary drifts and saccadic eye movements Jour-nal of the Optical Society of America 46 987ndash993[PubMed]
Delgado-Garcia J M del Pozo F amp Baker R (1986)Behavior of neurons in the abducens nucleus of thealert catVII Internuclear neurons Neuroscience 17953ndash973 [PubMed]
Dieringer N amp Daunicht W J (1986) Image fadingVAproblem for frogs Naturwissenschaften 73 330ndash332[PubMed]
Dieringer N amp Precht W (1982) Compensatory headand eye movements in the frog and their contributionto stabilization of gaze Experimental BrainResearch 47 394ndash406 [PubMed]
Dieringer N Precht W amp Blight A R (1982)Resetting fast phases of head and eye and theirlinkage in the frog Experimental Brain Research 47407ndash416 [PubMed]
Ditchburn R W (1955) Eye-movements in relation toretinal action Optica Acta 1 171ndash176
Ditchburn R W amp Ginsborg B L (1952) Vision with astabilized retinal image Nature 170 36ndash37[PubMed]
Ditchburn R W amp Ginsborg B L (1953) Involuntaryeye movements during fixation The Journal ofPhysiology 119 1ndash17 [PubMed] [Article]
Donner K amp Hemila S (2007) Modelling the effect ofmicrosaccades on retinal responses to stationarycontrast patterns Vision Research 47 1166ndash1177[PubMed]
Donner K K amp Hemila S (2005) The possible purposeof microsaccades Paper presented at the EuropeanConference on Visual Perception A Coruna Spain
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 12
Easter S S Jr (1971) Spontaneous eye movements inrestrained goldfish Vision Research 11 333ndash342[PubMed]
Easter S S Jr Johns P R amp Heckenlively D R(1974) Horizontal compensatory eye movements ingoldfish (Carassius auratus) I The normal animalJournal of Comparative Physiology 92 23ndash35
Engbert R (2006) Microsaccades A microcosm forresearch on oculomotor control attention and visualperception Progress Brain Research 154 177ndash192[PubMed]
Engbert R amp Kliegl R (2003a) Binocular coordinationin microsaccades In R R J Hyona amp H Deubel(Eds) The mindrsquos eyes Cognitive and appliedaspects of eye movements (pp 103ndash117) OxfordUK Elsevier
Engbert R amp Kliegl R (2003b) Microsaccades uncoverthe orientation of covert attention Vision Research43 1035ndash1045 [PubMed]
Engbert R amp Kliegl R (2004) Microsaccades keep theeyesrsquo balance during fixation Psychological Science15 431ndash436 [PubMed]
Engbert R amp Mergenthaler K (2006) Microsaccadesare triggered by low retinal image slip Proceedingsof the National Academy of Sciences of the UnitedStates of America 103 7192ndash7197 [PubMed][Article]
Ewert J P amp Borchers H W (1974) Antwort vonretinalen Ganglienzellen bei freibeweglichen Kroten(Bufo Bufo L) Journal of Comparative and Physi-ology 92 117ndash130
Fuller J H (1980) Linkage of eye and head movementsin the alert rabbit Brain Research 194 219ndash222[PubMed]
Fuller J H (1981) Eye and head movements duringvestibular stimulation in the alert rabbit BrainResearch 205 363ndash381 [PubMed]
Galfano G Betta E amp Turatto M (2004) Inhibition ofreturn in microsaccades Experimental BrainResearch 159 400ndash404 [PubMed]
Gerrits H J amp Vendrik A J (1970) Artificial move-ments of a stabilized image Vision Research 101443ndash1456 [PubMed]
Gioanni H Bennis M amp Sansonetti A (1993) Visualand vestibular reflexes that stabilize gaze in thechameleon Visual Neuroscience 10 947ndash956[PubMed]
Goffart L Quinet J Chavane F amp Masson G S(2006) Influence of background illumination onfixation and visually guided saccades in the rhesusmonkey Vision Research 46 149ndash162 [PubMed]
Greschner M Bongard M Rujan P amp Ammermuller J(2002) Retinal ganglion cell synchronization by fixa-tional eye movements improves feature estimationNature Neuroscience 5 341ndash347 [PubMed]
Gur M Beylin A amp Snodderly D M (1997)Response variability of neurons in primary visualcortex (V1) of alert monkeys Journal of Neuro-science 17 2914ndash2920 [PubMed] [Article]
Hafed Z M amp Clark J J (2002) Microsaccades as anovert measure of covert attention shifts VisionResearch 42 2533ndash2545 [PubMed]
Hebbard F W amp Marg E (1960) Physiologicalnystagmus in the cat Journal of the Optical Societyof America 50 151ndash155 [PubMed]
Hennig M A Kerscher N J Funke K amp Worgotter F(2002) Stochastic resonance in visual cortical neuronsDoes the eye-tremor actually improve visual acuityNeurocomputing 44ndash46 115ndash120
Hennig M H amp Worgotter F (2004) Eye micro-movements improve stimulus detection beyond theNyquist limit in the peripheral retina In Advances inNeural Information Processing Systems (NIPS2003)(vol 16) Denver CO MIT Press
Hennig M H amp Worgotter F (2007) Effects offixational eye movements on retinal ganglion cellresponses A modelling study Frontiers in Computa-tional Neuroscience 1 2 [PubMed] [Article]
Hermann H T amp Constantine M M (1971) Eyemovements in the goldfish Vision Research 11313ndash331 [PubMed]
Horowitz T S Fine E M Fencsik D E Yurgenson Samp Wolfe J M (2007) Fixational eye movements arenot an index of covert attention PsychologicalScience 18 356ndash363 [PubMed]
Horwitz G D amp Albright T D (2003) Short-latencyfixational saccades induced by luminance incrementsJournal of Neurophysiology 90 1333ndash1339[PubMed] [Article]
Laubrock J Engbert R Rolfs M amp Kliegl R(2007) Microsaccades are an index of covertattention Commentary on Horowitz Fine FencsikYurgenson and Wolfe (2007) Psychological Sci-ence 18 364ndash366 discussion 367ndash368 [PubMed]
Leopold D A amp Logothetis N K (1998) Micro-saccades differentially modulate neural activity in thestriate and extrastriate visual cortex ExperimentalBrain Research 123 341ndash345 [PubMed]
Lettvin J Y Maturana H R MsCulloch W S amp PittsW H (1968) What the frogrsquos eye tells the frogrsquosbrain In W C C a M Balaban (Ed) The mind
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 13
Biological approaches to its functions (pp 233ndash258)John Wiley and Sons London
Macknik S L amp Livingstone M S (1998) Neuronalcorrelates of visibility and invisibility in the primatevisual system Nature Neuroscience 1 144ndash149[PubMed]
Macknik S L amp Martinez-Conde S (2004) The spatialand temporal effects of lateral inhibitory networksand their relevance to the visibility of spatiotemporaledges Neurocomputing 58ndash60 775ndash782
Macknik S L Martinez-Conde S amp Haglund M M(2000) The role of spatiotemporal edges in visibilityand visual masking Proceedings of the NationalAcademy of Sciences of the United States of America97 7556ndash7560 [PubMed] [Article]
Manteuffel G Plasa L Sommer T J amp Wess O(1977) Involuntary eye movements in salamandersNaturwissenschaften 64 533ndash534 [PubMed]
Marchetti E Gauthier G M amp Pellet J (1983)Cerebellar control of eye movements studied withinjection of harmaline in the trained baboon ArchivesItaliennes de Biologies 121 1ndash17 [PubMed]
Martinez-Conde S (2006) Fixational eye movements innormal and pathological vision Progress in BrainResearch 154 151ndash176 [PubMed]
Martinez-Conde S amp Macknik S L (2007a) Bobbingand perceiving Scientific American 297 16
Martinez-Conde S amp Macknik S L (2007b) Windowson the mind Scientific American 297 56ndash63[PubMed]
Martinez-Conde S Macknik S L amp Hubel D H(2000) Microsaccadic eye movements and firing ofsingle cells in the striate cortex of macaque monkeysNature Neuroscience 3 251ndash258 [PubMed]
Martinez-Conde S Macknik S L amp Hubel D H(2002) The function of bursts of spikes during visualfixation in the awake primate lateral geniculate nucleusand primary visual cortex Proceedings of theNational Academy of Sciences of the United Statesof America 99 13920ndash13925 [PubMed] [Article]
Martinez-Conde S Macknik S L amp Hubel D H(2004) The role of fixational eye movements invisual perception Nature Reviews Neuroscience 5229ndash240 [PubMed]
Martinez-Conde S Macknik S L Troncoso X G ampDyar T A (2006) Microsaccades counteract visualfading during fixation Neuron 49 297ndash305[PubMed] [Article]
Mensh B D Aksay E Lee D D Seung H S ampTank D W (2004) Spontaneous eye movements ingoldfish Oculomotor integrator performance plasti-
city and dependence on visual feedback VisionResearch 44 711ndash726 [PubMed]
Miller J A Denning K S George J S MarshakD W amp Kenyon G T (2006) A high frequencyresonance in the responses of retinal ganglion cells torapidly modulated stimuli A computer model VisualNeuroscience 23 779ndash794 [PubMed]
Montgomery J C McVean A R amp McCarthy D(1983) The effects of lowered temperature onspontaneous eye movements in a teleost fish Com-parative Biochemistry and Physiology A Compara-tive Physiology 75 363ndash368 [PubMed]
Motter B C amp Poggio G F (1984) Binocularfixation in the rhesus monkey Spatial and temporalcharacteristics Experimental Brain Research 54304ndash314 [PubMed]
Murakami I amp Cavanagh P (1998) A jitter after-effectreveals motion-based stabilization of vision Nature395 798ndash801 [PubMed]
Nye P W (1969) The monocular eye movements of thepigeon Vision Research 9 133ndash144 [PubMed]
Olveczky B P Baccus S A amp Meister M (2003)Segregation of object and background motion in theretina Nature 423 401ndash408 [PubMed]
Olveczky B P Baccus S A amp Meister M (2007)Retinal adaptation to object motion Neuron 56689ndash700 [PubMed] [Article]
Otero-Millan J Troncoso X Macknik S L Serrano-Pedraza I amp Martinez-Conde S (2008) Saccadesand microsaccades during visual fixation explorationand search Foundations for a common saccadicgenerator Journal of Vision 8(14)21 1ndash18 httpjournalofvisionorg81421 doi10116781421[PubMed] [Article]
Pastor A M Torres B Delgado-Garcia J M ampBaker R (1991) Discharge characteristics of medialrectus and abducens motoneurons in the goldfishJournal of Neurophysiology 66 2125ndash2140 [PubMed]
Pettigrew J D Wallman J amp Wildsoet C F (1990)Saccadic oscillations facilitate ocular perfusion fromthe avian pecten Nature 343 362ndash363 [PubMed]
Pritchard R M amp Heron W (1960) Small eye move-ments of the cat Canadian Journal of Psychology14 131ndash137 [PubMed]
Ratliff F amp Riggs L A (1950) Involuntary motions ofthe eye during monocular fixation Journal of Exper-imental Psychology 40 687ndash701 [PubMed]
Reppas J B UsreyWM amp Reid R C (2002) Saccadiceye movements modulate visual responses in thelateral geniculate nucleus Neuron 35 961ndash974[PubMed] [Article]
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 14
Riggs L A amp Ratliff F (1952) The effects of counter-acting the normal movements of the eye Journal ofthe Optical Society of America 42 872ndash873
Rolfs M Kliegl R amp Engbert R (2008) Toward amodel of microsaccade generation The case ofmicrosaccadic inhibition Journal of Vision 8(11)51ndash23 httpjournalofvisionorg8115 doi1011678115 [PubMed] [Article]
Rolfs M Laubrock J amp Kliegl R (2006) Shorteningand prolongation of saccade latencies followingmicrosaccades Experimental Brain Research 169369ndash376 [PubMed]
Rucci M Iovin R Poletti M amp Santini F (2007)Miniature eye movements enhance fine spatial detailNature 447 851ndash854 [PubMed]
Salas C Navarro F Torres B amp Delgado-Garcia J M(1992) Effects of diazepam and D-amphetamine onrhythmic pattern of eye movements in goldfishNeuroreport 3 131ndash134 [PubMed]
Schipperheyn J J (1963) Respiratory eye movementsand perception of stationary objects in the frog ActaPhysiologica et Pharmacologica Neerlandica 12157ndash159 [PubMed]
Schipperheyn J J (1965) Contrast detection in frogrsquosretina Acta Physiologica et Pharmacologica Neer-landica 13 231ndash277 [PubMed]
Schipperheyn J J (1967) Influence of eye movements onretinal ganglion cell activity in frogs Acta Physi-ologica et Pharmacologica Neerlandica 14 506[PubMed]
Segev R Schneidman E Goodhouse J amp Berry M J2nd (2007) Role of eye movements in the retinalcode for a size discrimination task Journal ofNeurophysiology 98 1380ndash1391 [PubMed] [Article]
Sharpe C R (1972) The visibility and fading of thinlines visualized by their controlled movement acrossthe retina The Journal of Physiology 222 113ndash134[PubMed] [Article]
Shelley M McLaughlin D Shapley R amp Wielaard J(2002) States of high conductance in a large-scalemodel of the visual cortex Journal of ComputationalNeuroscience 13 93ndash109 [PubMed]
Skavenski A A Robinson D A Steinman R M ampTimberlake G T (1975) Miniature eye movementsof fixation in rhesus monkey Vision Research 151269ndash1273 [PubMed]
Snodderly D M (1987) Effects of light and darkenvironments on macaque and human fixational eyemovements Vision Research 27 401ndash415 [PubMed]
Snodderly D M Kagan I amp Gur M (2001) Selectiveactivation of visual cortex neurons by fixational eyemovements Implications for neural coding VisualNeuroscience 18 259ndash277 [PubMed]
Snodderly D M amp Kurtz D (1985) Eye position duringfixation tasks Comparison of macaque and humanVision Research 25 83ndash98 [PubMed]
Steinbach M J amp Money K E (1973) Eye movementsof the owl Vision Research 13 889ndash891 [PubMed]
Steinman R M Haddad G M Skavenski A A ampWyman D (1973) Miniature eye movement Science181 810ndash819 [PubMed]
Torres B Pastor A M Cabrera B Salas C ampDelgado-Garcia J M (1992) Afferents to theoculomotor nucleus in the goldfish (Carassius aur-atus) as revealed by retrograde labeling with horse-radish peroxidase Journal of Comparative Neurology324 449ndash461 [PubMed]
Troncoso X G Macknik S L amp Martinez-Conde S(2008) Microsaccades counteract perceptual filling-inJournal of Vision 8(14)15 1ndash9 httpjournalofvisionorg81415 doi10116781415 [Article]
Troncoso X G Macknik S L Otero-Millan J ampMartinez-Conde S (2008) Microsaccades driveillusory motion in the Enigma illusion Proceedingsof the National Academy of Sciences of the UnitedStates of America 105 16033ndash16038 [PubMed]
Tse P U Sheinberg D S amp Logothetis N K (2004)The distribution of microsaccade directions need notreveal the location of attention Psychological Science15 708ndash710
Turatto M Valsecchi M Tame L amp Betta E (2007)Microsaccades distinguish between global and localvisual processing Neuroreport 18 1015ndash1018[PubMed]
Uchida N Kepecs A amp Mainen Z F (2006) Seeing ata glance smelling in a whiff Rapid forms ofperceptual decision making Nature Reviews Neuro-science 7 485ndash491 [PubMed]
Valsecchi M Betta E amp Turatto M (2007) Visualoddballs induce prolonged microsaccadic inhibitionExperimental Brain Research 177 196ndash208[PubMed]
van Dam L C amp van Ee R (2006) Retinal image shiftsbut not eye movements per se cause alternations inawareness during binocular rivalry Journal of Vision6(11)3 1172ndash1179 httpjournalofvisionorg6113doi1011676113 [PubMed] [Article]
Van der Steen J amp Collewijn H (1984) Ocular stabilityin the horizontal frontal and sagittal planes in therabbit Experimental Brain Research 56 263ndash274[PubMed]
Van Gisbergen J A Robinson D A amp Gielen S(1981) A quantitative analysis of generation ofsaccadic eye movements by burst neurons Journalof Neurophysiology 45 417ndash442 [PubMed]
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 15
Van Gisbergen J A M amp Robinson D A (1977)Generation of micro and macrosaccades by burstneurons in the monkey In R Baker amp A Berthoz(Eds) Control of gaze by brain stem neurons (vol 1pp 301ndash308) New York ElsevierNorth-Holland
Wallman J Pettigrew J D amp Letelier J C (1994)The oscillatory saccades of birds Motoneural activ-ity and possible functions In B T Fuchs A FV Buttner amp D Zee (Eds) Contemporary ocularmotor and vestibular research A tribute to David ARobinson (pp 480ndash487) Stuttgart Germany ThiemePublishing
Williams P E amp Shapley R M (2007) A dynamicnonlinearity and spatial phase specificity in macaque
V1 neurons Journal of Neuroscience 27 5706ndash5718[PubMed] [Article]
Winterson B J amp Robinson D A (1975) Fixationby the alert but solitary cat Vision Research 151349ndash1352 [PubMed]
Yarbus A L (1967) Eye movements and vision(B Haigh Trans) New York Plenum Press
Zuber B L amp Stark L (1965) Microsaccades and thevelocityltamplitude relationship for saccadic eyemovements Science 150 1459ndash1460 [PubMed]
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 16

Easter S S Jr (1971) Spontaneous eye movements inrestrained goldfish Vision Research 11 333ndash342[PubMed]
Easter S S Jr Johns P R amp Heckenlively D R(1974) Horizontal compensatory eye movements ingoldfish (Carassius auratus) I The normal animalJournal of Comparative Physiology 92 23ndash35
Engbert R (2006) Microsaccades A microcosm forresearch on oculomotor control attention and visualperception Progress Brain Research 154 177ndash192[PubMed]
Engbert R amp Kliegl R (2003a) Binocular coordinationin microsaccades In R R J Hyona amp H Deubel(Eds) The mindrsquos eyes Cognitive and appliedaspects of eye movements (pp 103ndash117) OxfordUK Elsevier
Engbert R amp Kliegl R (2003b) Microsaccades uncoverthe orientation of covert attention Vision Research43 1035ndash1045 [PubMed]
Engbert R amp Kliegl R (2004) Microsaccades keep theeyesrsquo balance during fixation Psychological Science15 431ndash436 [PubMed]
Engbert R amp Mergenthaler K (2006) Microsaccadesare triggered by low retinal image slip Proceedingsof the National Academy of Sciences of the UnitedStates of America 103 7192ndash7197 [PubMed][Article]
Ewert J P amp Borchers H W (1974) Antwort vonretinalen Ganglienzellen bei freibeweglichen Kroten(Bufo Bufo L) Journal of Comparative and Physi-ology 92 117ndash130
Fuller J H (1980) Linkage of eye and head movementsin the alert rabbit Brain Research 194 219ndash222[PubMed]
Fuller J H (1981) Eye and head movements duringvestibular stimulation in the alert rabbit BrainResearch 205 363ndash381 [PubMed]
Galfano G Betta E amp Turatto M (2004) Inhibition ofreturn in microsaccades Experimental BrainResearch 159 400ndash404 [PubMed]
Gerrits H J amp Vendrik A J (1970) Artificial move-ments of a stabilized image Vision Research 101443ndash1456 [PubMed]
Gioanni H Bennis M amp Sansonetti A (1993) Visualand vestibular reflexes that stabilize gaze in thechameleon Visual Neuroscience 10 947ndash956[PubMed]
Goffart L Quinet J Chavane F amp Masson G S(2006) Influence of background illumination onfixation and visually guided saccades in the rhesusmonkey Vision Research 46 149ndash162 [PubMed]
Greschner M Bongard M Rujan P amp Ammermuller J(2002) Retinal ganglion cell synchronization by fixa-tional eye movements improves feature estimationNature Neuroscience 5 341ndash347 [PubMed]
Gur M Beylin A amp Snodderly D M (1997)Response variability of neurons in primary visualcortex (V1) of alert monkeys Journal of Neuro-science 17 2914ndash2920 [PubMed] [Article]
Hafed Z M amp Clark J J (2002) Microsaccades as anovert measure of covert attention shifts VisionResearch 42 2533ndash2545 [PubMed]
Hebbard F W amp Marg E (1960) Physiologicalnystagmus in the cat Journal of the Optical Societyof America 50 151ndash155 [PubMed]
Hennig M A Kerscher N J Funke K amp Worgotter F(2002) Stochastic resonance in visual cortical neuronsDoes the eye-tremor actually improve visual acuityNeurocomputing 44ndash46 115ndash120
Hennig M H amp Worgotter F (2004) Eye micro-movements improve stimulus detection beyond theNyquist limit in the peripheral retina In Advances inNeural Information Processing Systems (NIPS2003)(vol 16) Denver CO MIT Press
Hennig M H amp Worgotter F (2007) Effects offixational eye movements on retinal ganglion cellresponses A modelling study Frontiers in Computa-tional Neuroscience 1 2 [PubMed] [Article]
Hermann H T amp Constantine M M (1971) Eyemovements in the goldfish Vision Research 11313ndash331 [PubMed]
Horowitz T S Fine E M Fencsik D E Yurgenson Samp Wolfe J M (2007) Fixational eye movements arenot an index of covert attention PsychologicalScience 18 356ndash363 [PubMed]
Horwitz G D amp Albright T D (2003) Short-latencyfixational saccades induced by luminance incrementsJournal of Neurophysiology 90 1333ndash1339[PubMed] [Article]
Laubrock J Engbert R Rolfs M amp Kliegl R(2007) Microsaccades are an index of covertattention Commentary on Horowitz Fine FencsikYurgenson and Wolfe (2007) Psychological Sci-ence 18 364ndash366 discussion 367ndash368 [PubMed]
Leopold D A amp Logothetis N K (1998) Micro-saccades differentially modulate neural activity in thestriate and extrastriate visual cortex ExperimentalBrain Research 123 341ndash345 [PubMed]
Lettvin J Y Maturana H R MsCulloch W S amp PittsW H (1968) What the frogrsquos eye tells the frogrsquosbrain In W C C a M Balaban (Ed) The mind
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 13
Biological approaches to its functions (pp 233ndash258)John Wiley and Sons London
Macknik S L amp Livingstone M S (1998) Neuronalcorrelates of visibility and invisibility in the primatevisual system Nature Neuroscience 1 144ndash149[PubMed]
Macknik S L amp Martinez-Conde S (2004) The spatialand temporal effects of lateral inhibitory networksand their relevance to the visibility of spatiotemporaledges Neurocomputing 58ndash60 775ndash782
Macknik S L Martinez-Conde S amp Haglund M M(2000) The role of spatiotemporal edges in visibilityand visual masking Proceedings of the NationalAcademy of Sciences of the United States of America97 7556ndash7560 [PubMed] [Article]
Manteuffel G Plasa L Sommer T J amp Wess O(1977) Involuntary eye movements in salamandersNaturwissenschaften 64 533ndash534 [PubMed]
Marchetti E Gauthier G M amp Pellet J (1983)Cerebellar control of eye movements studied withinjection of harmaline in the trained baboon ArchivesItaliennes de Biologies 121 1ndash17 [PubMed]
Martinez-Conde S (2006) Fixational eye movements innormal and pathological vision Progress in BrainResearch 154 151ndash176 [PubMed]
Martinez-Conde S amp Macknik S L (2007a) Bobbingand perceiving Scientific American 297 16
Martinez-Conde S amp Macknik S L (2007b) Windowson the mind Scientific American 297 56ndash63[PubMed]
Martinez-Conde S Macknik S L amp Hubel D H(2000) Microsaccadic eye movements and firing ofsingle cells in the striate cortex of macaque monkeysNature Neuroscience 3 251ndash258 [PubMed]
Martinez-Conde S Macknik S L amp Hubel D H(2002) The function of bursts of spikes during visualfixation in the awake primate lateral geniculate nucleusand primary visual cortex Proceedings of theNational Academy of Sciences of the United Statesof America 99 13920ndash13925 [PubMed] [Article]
Martinez-Conde S Macknik S L amp Hubel D H(2004) The role of fixational eye movements invisual perception Nature Reviews Neuroscience 5229ndash240 [PubMed]
Martinez-Conde S Macknik S L Troncoso X G ampDyar T A (2006) Microsaccades counteract visualfading during fixation Neuron 49 297ndash305[PubMed] [Article]
Mensh B D Aksay E Lee D D Seung H S ampTank D W (2004) Spontaneous eye movements ingoldfish Oculomotor integrator performance plasti-
city and dependence on visual feedback VisionResearch 44 711ndash726 [PubMed]
Miller J A Denning K S George J S MarshakD W amp Kenyon G T (2006) A high frequencyresonance in the responses of retinal ganglion cells torapidly modulated stimuli A computer model VisualNeuroscience 23 779ndash794 [PubMed]
Montgomery J C McVean A R amp McCarthy D(1983) The effects of lowered temperature onspontaneous eye movements in a teleost fish Com-parative Biochemistry and Physiology A Compara-tive Physiology 75 363ndash368 [PubMed]
Motter B C amp Poggio G F (1984) Binocularfixation in the rhesus monkey Spatial and temporalcharacteristics Experimental Brain Research 54304ndash314 [PubMed]
Murakami I amp Cavanagh P (1998) A jitter after-effectreveals motion-based stabilization of vision Nature395 798ndash801 [PubMed]
Nye P W (1969) The monocular eye movements of thepigeon Vision Research 9 133ndash144 [PubMed]
Olveczky B P Baccus S A amp Meister M (2003)Segregation of object and background motion in theretina Nature 423 401ndash408 [PubMed]
Olveczky B P Baccus S A amp Meister M (2007)Retinal adaptation to object motion Neuron 56689ndash700 [PubMed] [Article]
Otero-Millan J Troncoso X Macknik S L Serrano-Pedraza I amp Martinez-Conde S (2008) Saccadesand microsaccades during visual fixation explorationand search Foundations for a common saccadicgenerator Journal of Vision 8(14)21 1ndash18 httpjournalofvisionorg81421 doi10116781421[PubMed] [Article]
Pastor A M Torres B Delgado-Garcia J M ampBaker R (1991) Discharge characteristics of medialrectus and abducens motoneurons in the goldfishJournal of Neurophysiology 66 2125ndash2140 [PubMed]
Pettigrew J D Wallman J amp Wildsoet C F (1990)Saccadic oscillations facilitate ocular perfusion fromthe avian pecten Nature 343 362ndash363 [PubMed]
Pritchard R M amp Heron W (1960) Small eye move-ments of the cat Canadian Journal of Psychology14 131ndash137 [PubMed]
Ratliff F amp Riggs L A (1950) Involuntary motions ofthe eye during monocular fixation Journal of Exper-imental Psychology 40 687ndash701 [PubMed]
Reppas J B UsreyWM amp Reid R C (2002) Saccadiceye movements modulate visual responses in thelateral geniculate nucleus Neuron 35 961ndash974[PubMed] [Article]
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 14
Riggs L A amp Ratliff F (1952) The effects of counter-acting the normal movements of the eye Journal ofthe Optical Society of America 42 872ndash873
Rolfs M Kliegl R amp Engbert R (2008) Toward amodel of microsaccade generation The case ofmicrosaccadic inhibition Journal of Vision 8(11)51ndash23 httpjournalofvisionorg8115 doi1011678115 [PubMed] [Article]
Rolfs M Laubrock J amp Kliegl R (2006) Shorteningand prolongation of saccade latencies followingmicrosaccades Experimental Brain Research 169369ndash376 [PubMed]
Rucci M Iovin R Poletti M amp Santini F (2007)Miniature eye movements enhance fine spatial detailNature 447 851ndash854 [PubMed]
Salas C Navarro F Torres B amp Delgado-Garcia J M(1992) Effects of diazepam and D-amphetamine onrhythmic pattern of eye movements in goldfishNeuroreport 3 131ndash134 [PubMed]
Schipperheyn J J (1963) Respiratory eye movementsand perception of stationary objects in the frog ActaPhysiologica et Pharmacologica Neerlandica 12157ndash159 [PubMed]
Schipperheyn J J (1965) Contrast detection in frogrsquosretina Acta Physiologica et Pharmacologica Neer-landica 13 231ndash277 [PubMed]
Schipperheyn J J (1967) Influence of eye movements onretinal ganglion cell activity in frogs Acta Physi-ologica et Pharmacologica Neerlandica 14 506[PubMed]
Segev R Schneidman E Goodhouse J amp Berry M J2nd (2007) Role of eye movements in the retinalcode for a size discrimination task Journal ofNeurophysiology 98 1380ndash1391 [PubMed] [Article]
Sharpe C R (1972) The visibility and fading of thinlines visualized by their controlled movement acrossthe retina The Journal of Physiology 222 113ndash134[PubMed] [Article]
Shelley M McLaughlin D Shapley R amp Wielaard J(2002) States of high conductance in a large-scalemodel of the visual cortex Journal of ComputationalNeuroscience 13 93ndash109 [PubMed]
Skavenski A A Robinson D A Steinman R M ampTimberlake G T (1975) Miniature eye movementsof fixation in rhesus monkey Vision Research 151269ndash1273 [PubMed]
Snodderly D M (1987) Effects of light and darkenvironments on macaque and human fixational eyemovements Vision Research 27 401ndash415 [PubMed]
Snodderly D M Kagan I amp Gur M (2001) Selectiveactivation of visual cortex neurons by fixational eyemovements Implications for neural coding VisualNeuroscience 18 259ndash277 [PubMed]
Snodderly D M amp Kurtz D (1985) Eye position duringfixation tasks Comparison of macaque and humanVision Research 25 83ndash98 [PubMed]
Steinbach M J amp Money K E (1973) Eye movementsof the owl Vision Research 13 889ndash891 [PubMed]
Steinman R M Haddad G M Skavenski A A ampWyman D (1973) Miniature eye movement Science181 810ndash819 [PubMed]
Torres B Pastor A M Cabrera B Salas C ampDelgado-Garcia J M (1992) Afferents to theoculomotor nucleus in the goldfish (Carassius aur-atus) as revealed by retrograde labeling with horse-radish peroxidase Journal of Comparative Neurology324 449ndash461 [PubMed]
Troncoso X G Macknik S L amp Martinez-Conde S(2008) Microsaccades counteract perceptual filling-inJournal of Vision 8(14)15 1ndash9 httpjournalofvisionorg81415 doi10116781415 [Article]
Troncoso X G Macknik S L Otero-Millan J ampMartinez-Conde S (2008) Microsaccades driveillusory motion in the Enigma illusion Proceedingsof the National Academy of Sciences of the UnitedStates of America 105 16033ndash16038 [PubMed]
Tse P U Sheinberg D S amp Logothetis N K (2004)The distribution of microsaccade directions need notreveal the location of attention Psychological Science15 708ndash710
Turatto M Valsecchi M Tame L amp Betta E (2007)Microsaccades distinguish between global and localvisual processing Neuroreport 18 1015ndash1018[PubMed]
Uchida N Kepecs A amp Mainen Z F (2006) Seeing ata glance smelling in a whiff Rapid forms ofperceptual decision making Nature Reviews Neuro-science 7 485ndash491 [PubMed]
Valsecchi M Betta E amp Turatto M (2007) Visualoddballs induce prolonged microsaccadic inhibitionExperimental Brain Research 177 196ndash208[PubMed]
van Dam L C amp van Ee R (2006) Retinal image shiftsbut not eye movements per se cause alternations inawareness during binocular rivalry Journal of Vision6(11)3 1172ndash1179 httpjournalofvisionorg6113doi1011676113 [PubMed] [Article]
Van der Steen J amp Collewijn H (1984) Ocular stabilityin the horizontal frontal and sagittal planes in therabbit Experimental Brain Research 56 263ndash274[PubMed]
Van Gisbergen J A Robinson D A amp Gielen S(1981) A quantitative analysis of generation ofsaccadic eye movements by burst neurons Journalof Neurophysiology 45 417ndash442 [PubMed]
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 15
Van Gisbergen J A M amp Robinson D A (1977)Generation of micro and macrosaccades by burstneurons in the monkey In R Baker amp A Berthoz(Eds) Control of gaze by brain stem neurons (vol 1pp 301ndash308) New York ElsevierNorth-Holland
Wallman J Pettigrew J D amp Letelier J C (1994)The oscillatory saccades of birds Motoneural activ-ity and possible functions In B T Fuchs A FV Buttner amp D Zee (Eds) Contemporary ocularmotor and vestibular research A tribute to David ARobinson (pp 480ndash487) Stuttgart Germany ThiemePublishing
Williams P E amp Shapley R M (2007) A dynamicnonlinearity and spatial phase specificity in macaque
V1 neurons Journal of Neuroscience 27 5706ndash5718[PubMed] [Article]
Winterson B J amp Robinson D A (1975) Fixationby the alert but solitary cat Vision Research 151349ndash1352 [PubMed]
Yarbus A L (1967) Eye movements and vision(B Haigh Trans) New York Plenum Press
Zuber B L amp Stark L (1965) Microsaccades and thevelocityltamplitude relationship for saccadic eyemovements Science 150 1459ndash1460 [PubMed]
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 16

Biological approaches to its functions (pp 233ndash258)John Wiley and Sons London
Macknik S L amp Livingstone M S (1998) Neuronalcorrelates of visibility and invisibility in the primatevisual system Nature Neuroscience 1 144ndash149[PubMed]
Macknik S L amp Martinez-Conde S (2004) The spatialand temporal effects of lateral inhibitory networksand their relevance to the visibility of spatiotemporaledges Neurocomputing 58ndash60 775ndash782
Macknik S L Martinez-Conde S amp Haglund M M(2000) The role of spatiotemporal edges in visibilityand visual masking Proceedings of the NationalAcademy of Sciences of the United States of America97 7556ndash7560 [PubMed] [Article]
Manteuffel G Plasa L Sommer T J amp Wess O(1977) Involuntary eye movements in salamandersNaturwissenschaften 64 533ndash534 [PubMed]
Marchetti E Gauthier G M amp Pellet J (1983)Cerebellar control of eye movements studied withinjection of harmaline in the trained baboon ArchivesItaliennes de Biologies 121 1ndash17 [PubMed]
Martinez-Conde S (2006) Fixational eye movements innormal and pathological vision Progress in BrainResearch 154 151ndash176 [PubMed]
Martinez-Conde S amp Macknik S L (2007a) Bobbingand perceiving Scientific American 297 16
Martinez-Conde S amp Macknik S L (2007b) Windowson the mind Scientific American 297 56ndash63[PubMed]
Martinez-Conde S Macknik S L amp Hubel D H(2000) Microsaccadic eye movements and firing ofsingle cells in the striate cortex of macaque monkeysNature Neuroscience 3 251ndash258 [PubMed]
Martinez-Conde S Macknik S L amp Hubel D H(2002) The function of bursts of spikes during visualfixation in the awake primate lateral geniculate nucleusand primary visual cortex Proceedings of theNational Academy of Sciences of the United Statesof America 99 13920ndash13925 [PubMed] [Article]
Martinez-Conde S Macknik S L amp Hubel D H(2004) The role of fixational eye movements invisual perception Nature Reviews Neuroscience 5229ndash240 [PubMed]
Martinez-Conde S Macknik S L Troncoso X G ampDyar T A (2006) Microsaccades counteract visualfading during fixation Neuron 49 297ndash305[PubMed] [Article]
Mensh B D Aksay E Lee D D Seung H S ampTank D W (2004) Spontaneous eye movements ingoldfish Oculomotor integrator performance plasti-
city and dependence on visual feedback VisionResearch 44 711ndash726 [PubMed]
Miller J A Denning K S George J S MarshakD W amp Kenyon G T (2006) A high frequencyresonance in the responses of retinal ganglion cells torapidly modulated stimuli A computer model VisualNeuroscience 23 779ndash794 [PubMed]
Montgomery J C McVean A R amp McCarthy D(1983) The effects of lowered temperature onspontaneous eye movements in a teleost fish Com-parative Biochemistry and Physiology A Compara-tive Physiology 75 363ndash368 [PubMed]
Motter B C amp Poggio G F (1984) Binocularfixation in the rhesus monkey Spatial and temporalcharacteristics Experimental Brain Research 54304ndash314 [PubMed]
Murakami I amp Cavanagh P (1998) A jitter after-effectreveals motion-based stabilization of vision Nature395 798ndash801 [PubMed]
Nye P W (1969) The monocular eye movements of thepigeon Vision Research 9 133ndash144 [PubMed]
Olveczky B P Baccus S A amp Meister M (2003)Segregation of object and background motion in theretina Nature 423 401ndash408 [PubMed]
Olveczky B P Baccus S A amp Meister M (2007)Retinal adaptation to object motion Neuron 56689ndash700 [PubMed] [Article]
Otero-Millan J Troncoso X Macknik S L Serrano-Pedraza I amp Martinez-Conde S (2008) Saccadesand microsaccades during visual fixation explorationand search Foundations for a common saccadicgenerator Journal of Vision 8(14)21 1ndash18 httpjournalofvisionorg81421 doi10116781421[PubMed] [Article]
Pastor A M Torres B Delgado-Garcia J M ampBaker R (1991) Discharge characteristics of medialrectus and abducens motoneurons in the goldfishJournal of Neurophysiology 66 2125ndash2140 [PubMed]
Pettigrew J D Wallman J amp Wildsoet C F (1990)Saccadic oscillations facilitate ocular perfusion fromthe avian pecten Nature 343 362ndash363 [PubMed]
Pritchard R M amp Heron W (1960) Small eye move-ments of the cat Canadian Journal of Psychology14 131ndash137 [PubMed]
Ratliff F amp Riggs L A (1950) Involuntary motions ofthe eye during monocular fixation Journal of Exper-imental Psychology 40 687ndash701 [PubMed]
Reppas J B UsreyWM amp Reid R C (2002) Saccadiceye movements modulate visual responses in thelateral geniculate nucleus Neuron 35 961ndash974[PubMed] [Article]
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 14
Riggs L A amp Ratliff F (1952) The effects of counter-acting the normal movements of the eye Journal ofthe Optical Society of America 42 872ndash873
Rolfs M Kliegl R amp Engbert R (2008) Toward amodel of microsaccade generation The case ofmicrosaccadic inhibition Journal of Vision 8(11)51ndash23 httpjournalofvisionorg8115 doi1011678115 [PubMed] [Article]
Rolfs M Laubrock J amp Kliegl R (2006) Shorteningand prolongation of saccade latencies followingmicrosaccades Experimental Brain Research 169369ndash376 [PubMed]
Rucci M Iovin R Poletti M amp Santini F (2007)Miniature eye movements enhance fine spatial detailNature 447 851ndash854 [PubMed]
Salas C Navarro F Torres B amp Delgado-Garcia J M(1992) Effects of diazepam and D-amphetamine onrhythmic pattern of eye movements in goldfishNeuroreport 3 131ndash134 [PubMed]
Schipperheyn J J (1963) Respiratory eye movementsand perception of stationary objects in the frog ActaPhysiologica et Pharmacologica Neerlandica 12157ndash159 [PubMed]
Schipperheyn J J (1965) Contrast detection in frogrsquosretina Acta Physiologica et Pharmacologica Neer-landica 13 231ndash277 [PubMed]
Schipperheyn J J (1967) Influence of eye movements onretinal ganglion cell activity in frogs Acta Physi-ologica et Pharmacologica Neerlandica 14 506[PubMed]
Segev R Schneidman E Goodhouse J amp Berry M J2nd (2007) Role of eye movements in the retinalcode for a size discrimination task Journal ofNeurophysiology 98 1380ndash1391 [PubMed] [Article]
Sharpe C R (1972) The visibility and fading of thinlines visualized by their controlled movement acrossthe retina The Journal of Physiology 222 113ndash134[PubMed] [Article]
Shelley M McLaughlin D Shapley R amp Wielaard J(2002) States of high conductance in a large-scalemodel of the visual cortex Journal of ComputationalNeuroscience 13 93ndash109 [PubMed]
Skavenski A A Robinson D A Steinman R M ampTimberlake G T (1975) Miniature eye movementsof fixation in rhesus monkey Vision Research 151269ndash1273 [PubMed]
Snodderly D M (1987) Effects of light and darkenvironments on macaque and human fixational eyemovements Vision Research 27 401ndash415 [PubMed]
Snodderly D M Kagan I amp Gur M (2001) Selectiveactivation of visual cortex neurons by fixational eyemovements Implications for neural coding VisualNeuroscience 18 259ndash277 [PubMed]
Snodderly D M amp Kurtz D (1985) Eye position duringfixation tasks Comparison of macaque and humanVision Research 25 83ndash98 [PubMed]
Steinbach M J amp Money K E (1973) Eye movementsof the owl Vision Research 13 889ndash891 [PubMed]
Steinman R M Haddad G M Skavenski A A ampWyman D (1973) Miniature eye movement Science181 810ndash819 [PubMed]
Torres B Pastor A M Cabrera B Salas C ampDelgado-Garcia J M (1992) Afferents to theoculomotor nucleus in the goldfish (Carassius aur-atus) as revealed by retrograde labeling with horse-radish peroxidase Journal of Comparative Neurology324 449ndash461 [PubMed]
Troncoso X G Macknik S L amp Martinez-Conde S(2008) Microsaccades counteract perceptual filling-inJournal of Vision 8(14)15 1ndash9 httpjournalofvisionorg81415 doi10116781415 [Article]
Troncoso X G Macknik S L Otero-Millan J ampMartinez-Conde S (2008) Microsaccades driveillusory motion in the Enigma illusion Proceedingsof the National Academy of Sciences of the UnitedStates of America 105 16033ndash16038 [PubMed]
Tse P U Sheinberg D S amp Logothetis N K (2004)The distribution of microsaccade directions need notreveal the location of attention Psychological Science15 708ndash710
Turatto M Valsecchi M Tame L amp Betta E (2007)Microsaccades distinguish between global and localvisual processing Neuroreport 18 1015ndash1018[PubMed]
Uchida N Kepecs A amp Mainen Z F (2006) Seeing ata glance smelling in a whiff Rapid forms ofperceptual decision making Nature Reviews Neuro-science 7 485ndash491 [PubMed]
Valsecchi M Betta E amp Turatto M (2007) Visualoddballs induce prolonged microsaccadic inhibitionExperimental Brain Research 177 196ndash208[PubMed]
van Dam L C amp van Ee R (2006) Retinal image shiftsbut not eye movements per se cause alternations inawareness during binocular rivalry Journal of Vision6(11)3 1172ndash1179 httpjournalofvisionorg6113doi1011676113 [PubMed] [Article]
Van der Steen J amp Collewijn H (1984) Ocular stabilityin the horizontal frontal and sagittal planes in therabbit Experimental Brain Research 56 263ndash274[PubMed]
Van Gisbergen J A Robinson D A amp Gielen S(1981) A quantitative analysis of generation ofsaccadic eye movements by burst neurons Journalof Neurophysiology 45 417ndash442 [PubMed]
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 15
Van Gisbergen J A M amp Robinson D A (1977)Generation of micro and macrosaccades by burstneurons in the monkey In R Baker amp A Berthoz(Eds) Control of gaze by brain stem neurons (vol 1pp 301ndash308) New York ElsevierNorth-Holland
Wallman J Pettigrew J D amp Letelier J C (1994)The oscillatory saccades of birds Motoneural activ-ity and possible functions In B T Fuchs A FV Buttner amp D Zee (Eds) Contemporary ocularmotor and vestibular research A tribute to David ARobinson (pp 480ndash487) Stuttgart Germany ThiemePublishing
Williams P E amp Shapley R M (2007) A dynamicnonlinearity and spatial phase specificity in macaque
V1 neurons Journal of Neuroscience 27 5706ndash5718[PubMed] [Article]
Winterson B J amp Robinson D A (1975) Fixationby the alert but solitary cat Vision Research 151349ndash1352 [PubMed]
Yarbus A L (1967) Eye movements and vision(B Haigh Trans) New York Plenum Press
Zuber B L amp Stark L (1965) Microsaccades and thevelocityltamplitude relationship for saccadic eyemovements Science 150 1459ndash1460 [PubMed]
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 16

Riggs L A amp Ratliff F (1952) The effects of counter-acting the normal movements of the eye Journal ofthe Optical Society of America 42 872ndash873
Rolfs M Kliegl R amp Engbert R (2008) Toward amodel of microsaccade generation The case ofmicrosaccadic inhibition Journal of Vision 8(11)51ndash23 httpjournalofvisionorg8115 doi1011678115 [PubMed] [Article]
Rolfs M Laubrock J amp Kliegl R (2006) Shorteningand prolongation of saccade latencies followingmicrosaccades Experimental Brain Research 169369ndash376 [PubMed]
Rucci M Iovin R Poletti M amp Santini F (2007)Miniature eye movements enhance fine spatial detailNature 447 851ndash854 [PubMed]
Salas C Navarro F Torres B amp Delgado-Garcia J M(1992) Effects of diazepam and D-amphetamine onrhythmic pattern of eye movements in goldfishNeuroreport 3 131ndash134 [PubMed]
Schipperheyn J J (1963) Respiratory eye movementsand perception of stationary objects in the frog ActaPhysiologica et Pharmacologica Neerlandica 12157ndash159 [PubMed]
Schipperheyn J J (1965) Contrast detection in frogrsquosretina Acta Physiologica et Pharmacologica Neer-landica 13 231ndash277 [PubMed]
Schipperheyn J J (1967) Influence of eye movements onretinal ganglion cell activity in frogs Acta Physi-ologica et Pharmacologica Neerlandica 14 506[PubMed]
Segev R Schneidman E Goodhouse J amp Berry M J2nd (2007) Role of eye movements in the retinalcode for a size discrimination task Journal ofNeurophysiology 98 1380ndash1391 [PubMed] [Article]
Sharpe C R (1972) The visibility and fading of thinlines visualized by their controlled movement acrossthe retina The Journal of Physiology 222 113ndash134[PubMed] [Article]
Shelley M McLaughlin D Shapley R amp Wielaard J(2002) States of high conductance in a large-scalemodel of the visual cortex Journal of ComputationalNeuroscience 13 93ndash109 [PubMed]
Skavenski A A Robinson D A Steinman R M ampTimberlake G T (1975) Miniature eye movementsof fixation in rhesus monkey Vision Research 151269ndash1273 [PubMed]
Snodderly D M (1987) Effects of light and darkenvironments on macaque and human fixational eyemovements Vision Research 27 401ndash415 [PubMed]
Snodderly D M Kagan I amp Gur M (2001) Selectiveactivation of visual cortex neurons by fixational eyemovements Implications for neural coding VisualNeuroscience 18 259ndash277 [PubMed]
Snodderly D M amp Kurtz D (1985) Eye position duringfixation tasks Comparison of macaque and humanVision Research 25 83ndash98 [PubMed]
Steinbach M J amp Money K E (1973) Eye movementsof the owl Vision Research 13 889ndash891 [PubMed]
Steinman R M Haddad G M Skavenski A A ampWyman D (1973) Miniature eye movement Science181 810ndash819 [PubMed]
Torres B Pastor A M Cabrera B Salas C ampDelgado-Garcia J M (1992) Afferents to theoculomotor nucleus in the goldfish (Carassius aur-atus) as revealed by retrograde labeling with horse-radish peroxidase Journal of Comparative Neurology324 449ndash461 [PubMed]
Troncoso X G Macknik S L amp Martinez-Conde S(2008) Microsaccades counteract perceptual filling-inJournal of Vision 8(14)15 1ndash9 httpjournalofvisionorg81415 doi10116781415 [Article]
Troncoso X G Macknik S L Otero-Millan J ampMartinez-Conde S (2008) Microsaccades driveillusory motion in the Enigma illusion Proceedingsof the National Academy of Sciences of the UnitedStates of America 105 16033ndash16038 [PubMed]
Tse P U Sheinberg D S amp Logothetis N K (2004)The distribution of microsaccade directions need notreveal the location of attention Psychological Science15 708ndash710
Turatto M Valsecchi M Tame L amp Betta E (2007)Microsaccades distinguish between global and localvisual processing Neuroreport 18 1015ndash1018[PubMed]
Uchida N Kepecs A amp Mainen Z F (2006) Seeing ata glance smelling in a whiff Rapid forms ofperceptual decision making Nature Reviews Neuro-science 7 485ndash491 [PubMed]
Valsecchi M Betta E amp Turatto M (2007) Visualoddballs induce prolonged microsaccadic inhibitionExperimental Brain Research 177 196ndash208[PubMed]
van Dam L C amp van Ee R (2006) Retinal image shiftsbut not eye movements per se cause alternations inawareness during binocular rivalry Journal of Vision6(11)3 1172ndash1179 httpjournalofvisionorg6113doi1011676113 [PubMed] [Article]
Van der Steen J amp Collewijn H (1984) Ocular stabilityin the horizontal frontal and sagittal planes in therabbit Experimental Brain Research 56 263ndash274[PubMed]
Van Gisbergen J A Robinson D A amp Gielen S(1981) A quantitative analysis of generation ofsaccadic eye movements by burst neurons Journalof Neurophysiology 45 417ndash442 [PubMed]
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 15
Van Gisbergen J A M amp Robinson D A (1977)Generation of micro and macrosaccades by burstneurons in the monkey In R Baker amp A Berthoz(Eds) Control of gaze by brain stem neurons (vol 1pp 301ndash308) New York ElsevierNorth-Holland
Wallman J Pettigrew J D amp Letelier J C (1994)The oscillatory saccades of birds Motoneural activ-ity and possible functions In B T Fuchs A FV Buttner amp D Zee (Eds) Contemporary ocularmotor and vestibular research A tribute to David ARobinson (pp 480ndash487) Stuttgart Germany ThiemePublishing
Williams P E amp Shapley R M (2007) A dynamicnonlinearity and spatial phase specificity in macaque
V1 neurons Journal of Neuroscience 27 5706ndash5718[PubMed] [Article]
Winterson B J amp Robinson D A (1975) Fixationby the alert but solitary cat Vision Research 151349ndash1352 [PubMed]
Yarbus A L (1967) Eye movements and vision(B Haigh Trans) New York Plenum Press
Zuber B L amp Stark L (1965) Microsaccades and thevelocityltamplitude relationship for saccadic eyemovements Science 150 1459ndash1460 [PubMed]
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 16

Van Gisbergen J A M amp Robinson D A (1977)Generation of micro and macrosaccades by burstneurons in the monkey In R Baker amp A Berthoz(Eds) Control of gaze by brain stem neurons (vol 1pp 301ndash308) New York ElsevierNorth-Holland
Wallman J Pettigrew J D amp Letelier J C (1994)The oscillatory saccades of birds Motoneural activ-ity and possible functions In B T Fuchs A FV Buttner amp D Zee (Eds) Contemporary ocularmotor and vestibular research A tribute to David ARobinson (pp 480ndash487) Stuttgart Germany ThiemePublishing
Williams P E amp Shapley R M (2007) A dynamicnonlinearity and spatial phase specificity in macaque
V1 neurons Journal of Neuroscience 27 5706ndash5718[PubMed] [Article]
Winterson B J amp Robinson D A (1975) Fixationby the alert but solitary cat Vision Research 151349ndash1352 [PubMed]
Yarbus A L (1967) Eye movements and vision(B Haigh Trans) New York Plenum Press
Zuber B L amp Stark L (1965) Microsaccades and thevelocityltamplitude relationship for saccadic eyemovements Science 150 1459ndash1460 [PubMed]
Journal of Vision (2008) 8(14)28 1ndash16 Martinez-Conde amp Macknik 16